О СЛОЖНОМ ПРОСТЫМ ЯЗЫКОМ.
Тема эта сложная и комплексная, затрагивающая сразу же огромное количество биохимических процессов происходящих в нашем организме. Но давайте все таки попробуем разобраться, что же такое митохондрии и как они работают.
И так, митохондрии это одна из самых важных составляющих живой клетки. Если говорить простым языком то можно сказать, что это энергетическая станция клетки . Их деятельность основана на окисление органических соединений и генерации электрического потенциала (энергии освободившейся при распаде молекулы АТФ) для осуществления мышечного сокращения.
Все мы знаем, что работа нашего организма происходит в строгом соответствии с первым законом термодинамики. Энергия не создается в нашем организме, а лишь превращается. Организм только выбирает форму трансформации энергии, не производя ее, от химической к механической и тепловой. Основным источником всей энергии на планете Земля является Солнце. Приходя к нам в форме света, энергия поглощается хлорофиллом растений, там она возбуждает электрон атома водорода и таким образом дает энергию живой материи.
Своей жизнью мы обязаны энергии маленького электрона.
Работа митохондрии заключается в ступенчатом переносе энергии электрона водорода между атомами металлов, присутствующих в группах белковых комплексов дыхательной цепи (электронно-транспортной цепи белков), где каждый последующий комплекс обладает более высоким сродством к электрону притягивая его, чем предыдущий, до тех пор, пока электрон не соединиться с молекулярным кислородом, обладающим наибольшим сродством к электрону.
Каждый раз при передачи электрона по цепи высвобождается энергия которая аккумулируется в виде электрохимического градиента и затем реализовывается в виде мышечного сокращения и выделения тепла.
Серия окислительных процессов в митохондрии позволяющая перенести энергетический потенциал электрона называется «внутриклеточным дыханием» или часто «дыхательной цепью» , так как электрон по цепочки передается от атома к атому до тех пор пока не достигнет своей конечной цели атома кислорода.
Митохондриям нужен кислород для переноса энергии в процессе окисления.
Митохондрии потребляют до 80% кислорода который мы вдыхаем.
Митохондрия представляет из себя постоянную структуру клетки, расположенную в ее цитоплазме. Размер митохондрии обычно составляет от 0,5 до 1 мкм в диаметре. По форме она имеет зернистую структуру и может занимать до 20% объема клетки. Такая постоянная органическая структура клетки называется органелла . К органеллам относятся и миофибриллы – сократительные единицы мышечной клетки; и ядро клетки это тоже органелла. Вообще, любая постоянная структура клетки является органоидом-органеллой.
Открыл митохондрии и впервые описал немецкий анатом и гистолог Рихард Альтман в 1894 году, а название этой органелле дал другой немецкий гистолог К. Бенд в 1897 году. Но только в 1920 году, опять же немецкий биохимик Отто Вагбург, доказал, что с митохондриями связаны процессы клеточного дыхания.
Существует теория, согласно которой митохондрии появились в результате захвата примитивными клетками, клетками которые сами не могли использовать кислород для генерации энергии, бактерий протогенотов, которые могли это делать. Именно потому, что митохондрия ранее представляла из себя отдельный живой организм она и по сей день обладает собственным ДНК.
Митохондрии ранее представляли из себя самостоятельный живой организм.
В ходе эволюции прогеноты предали множество своих генов сформировавшемуся, благодаря повысившейся энергоэффективности, ядру и перестали быть самостоятельными организмами. Митохондрии присутствуют во всех клетках. Даже в сперматозоиде есть митохондрии. Именно благодаря им приводится в движение хвостик сперматозоида осуществляющий его движение. Но особенно много митахондрий в тех местах, где необходима энергия для любых жизненных процессов. И это конечно прежде всего мышечные клетки.
В мышечных клетках митохондрии могут объединяться в группы гигантских разветвленных митохондрий, связанных друг с другом с помощью межмитохондриальных контактов, в которых они создают согласованную работающую кооперативную систему . Пространство в такой зоне имеет повышенную электронную плотность. Новые митохондрии образуются путем простого деления предыдущих органелл. Наиболее «простой» и доступный всем клеткам механизм энергетического обеспечения чаще всего называют общим понятием гликолиз .
Это процесс последовательного разложения глюкозы до пировиноградной кислоты. Если этот процесс происходит без участия молекулярного кислорода или с недостаточным его присутствием, то он называется анаэробный гликолиз . При этом глюкоза расщепляется не до конечных продуктов, а до молочной и пировиноградной кислоты которая далее претерпевает дальнейшие превращения в ходе брожения. Поэтому высвобождающейся энергии бывает меньше, но и скорость получения энергии быстрее. В результате анаэробного гликолиза из одной молекулы глюкозы клетка получает 2 молекулы АТФ и 2 молекулы молочной кислоты. Такой «базовый» энергетический процесс может протекать внутри любой клетки без участия митохондрий .
В присутствии молекулярного кислорода внутри митохондрий осуществляется аэробный гликолиз в рамках «дыхательной цепи». Пировиноградная кислота в аэробных условиях вовлекается в цикл трикарбоновых кислот или цикл Кребса . В результате этого многостадийного процесса из одной молекулы глюкозы образуется 36 молекул АТФ. Сравнение энергетического баланса клетки, имеющей развитые митохондрии и клетки, где они не развиты показывает (при достаточном количестве кислорода) различие в полноте использования энергии глюкозы внутри клетки почти в 20 раз!
У человека, волокна скелетных мышц можно условно разделить на три типа исходя из механических и метаболических свойств: - медленные окислительные; - быстрые гликолитические; - быстрые окислительно-гликолитические.

Быстрые мышечные волокна предназначены для выполнения быстрой и тяжелой работы. Для своего сокращения они используют в основном быстрые источники энергии, а именно криатинфосфот и анаэробный гликолиз. Содержание митохондрий в таких типах волокон значительно меньше чем в медленных мышечных волокнах.
Медленные мышечные волокна выполняют медленные сокращения, но способны работать длительное время. В качестве энергии они используют аэробный гликолиз и синтез энергии из жиров. Это дает гораздо больше энергии чем анаэробный гликолиз, но требует в замен больше времени, так как цепочка деградации глюкозы более сложная и требует присутствия кислорода, транспортировка которого к месту преобразования энергии тоже занимает время. Медленные мышечные волокна называют красными из-за миоглобина – белка, ответственный за доставку кислорода внутрь волокна. Медленные мышечные волокна содержат значительное количество митохондрий.
Возникает вопрос, каким образом и с помощью каких упражнений можно развить в мышечных клетках разветвленную сеть митохондрий? Существуют различные теории и методики тренировок и о них в материале по .
Характерные для подавляющего большинства клеток. Основная функция – это окисление органических соединений и производство АТФ-молекул из освободившейся энергии. Маленькая митохондрия является главной энергетической станцией всего организма.
Происхождение митохондрий
Среди ученых на сегодняшний день весьма популярно мнение, что митохондрия появилась в клетке не самостоятельно в ходе эволюции. Скорее всего, это произошло из-за захвата примитивной клеткой, не способной на тот момент самостоятельно использовать кислород, бактерии, которая умела это и, соответственно, была превосходным источником энергии. Подобный симбиоз оказался успешным и закрепился в последующих поколениях. В пользу этой теории указывает наличие в митохондриях собственной ДНК.
Как устроены митохондрии
Митохондрия обладает двумя мембранами: наружной и внутренней. Главная функция наружной мембраны – это отделение органоида от цитоплазмы клетки. Она состоит из билипидного слоя и белков, пронизывающих его, через которые и осуществляется транспорт молекул и ионов, необходимых для работы. В то время как гладкая, внутренняя образует многочисленные складки – кристы, которые существенно увеличивают ее площадь. Внутренняя мембрана по большей части состоит из белков, среди которых присутствуют ферменты дыхательной цепи, транспортные белки и крупные АТФ-синтетазные комплексы. Именно в этом месте происходит синтез АТФ. Между наружной и внутренней мембраной находится межмембранное пространство с присущими ему ферментами.
Внутреннее пространство митохондрий называется матрикс. Здесь расположены ферментные системы окисления жирных кислот и пирувата, ферменты цикла Кребса, а также наследственный материал митохондрий – ДНК, РНК и белоксинтезирующий аппарат.
Для чего нужны митохондрии
Главной функцией митохондрий является синтез универсальной формы химической энергии – АТФ. Также они принимают участие в цикле трикарбоновых кислот, превращая пируват и жирные кислоты в ацетил-CoA, а затем окисляя его. В этом органоиде хранится и передается по наследству митохондриальная ДНК, кодирующая воспроизводство тРНК, рРНК и некоторых белков, необходимых для нормального функционирования митохондрий.
Митохондрия - это спиральная, округлая, вытянутая или разветвленная органелла.
Впервые понятие митохондрия было предложено Бенда в 1897 г. Митохондрии можно обнаружить в живых клетках с помощью фазово-контрастной и интерференционной микроскопии в виде зерен, гранул или нитей. Это довольно подвижные структуры, которые могут перемещаться, сливаться друг с другом, делиться. При окраске специальными методами в погибших клетках при световой микроскопии митохондрии имеют вид мелких зерен (гранул), диффузно распределенных в цитоплазме или концентрирующихся в каких-то определенных ее зонах.
В результате разрушения глюкозы и жиров в присутствии кислорода в митохондриях образуется энергия, а органические вещества превращаются в воду и диоксид углерода. Именно таким образом получают основную энергию, необходимую для жизнедеятельности, животные организмы. Энергия накапливается в аденозинтрифосфате (АТФ), а точнее, в его макроэргических связях. Функция митохондрий тесно связана с окислением органических соединений и использованием освобождающейся при их распаде энергии для синтеза молекул АТФ. Поэтому митохондрии часто называют энергетическими станциями клетки, или органеллами клеточного дыхания. АТФ выполняет функцию поставщика энергии, перенося одну из своих богатых энергией концевых фосфатных групп на другую молекулу, и превращается при этом в АДФ.
Предполагают, что в эволюции митохондрии были прокариотическими микроорганизмами, которые стали симбиотами в организме древней клетки. В последующем они стали жизненно необходимы, что было связано с увеличением содержания кислорода в атмосфере Земли. С одной стороны, митохондрии удаляли избыток токсичного для клетки кислорода, а с другой - обеспечивали энергией.
Без митохондрий клетка практически не в состоянии использовать кислород как вещество, обеспечивающее поставку энергии, и может восполнять свои энергетические потребности лишь путем анаэробных процессов. Таким образом, кислород - это яд, но яд жизненно важный для клетки, причем избыток кислорода так же вреден, как и его недостаток.
Митохондрии могут изменять свою форму и перемещаться в те области клетки, где потребность в них наиболее высока. Так, в кардиомиоцитах митохондрии находятся вблизи миофибрилл, в клетках почечных канальцев вблизи базальных впячиваний и т. д. В клетке содержится до тысячи митохондрий, и их количество зависит от активности клетки.
Митохондрии имеют средние поперечные размеры 0,5…3 мкм. В зависимости от размеров выделяют мелкие, средние, крупные и гигантские митохондрии (формируют разветвленную сеть - митохондриальный ретикулум). Размеры и число митохондрий тесно связаны с активностью клетки и ее энергопотреблением. Они крайне изменчивы и в зависимости от активности клетки, содержания кислорода, гормональных влияний могут набухать, изменять число и структуру крист, варьировать в числе, форме и размерах, а также ферментативной активности.
Объемная плотность митохондрий, степень развития их внутренней поверхности и другие показатели зависят от энергетических потребностей клетки. В лимфоцитах имеется всего по несколько митохондрий, а в печеночных клетках их 2…3 тыс.
Митохондрии состоят из матрикса, внутренней мембраны, перимитохондриального пространства и наружной мембраны. Наружная митохондриальная мембрана отделяет органеллу от гиалоплазмы. Обычно она имеет ровные контуры и замкнута так, что представляет собой мембранный мешок.
Внешнюю мембрану от внутренней отделяет перимитохондриальное пространство шириной около 10…20 нм. Внутренняя митохондриальная мембрана ограничивает собственно внутреннее содержимое митохондрии - матрикс. Внутренняя мембрана образует многочисленные выпячивания внутрь митохондрий, которые имеют вид плоских гребней, или крист.
По форме кристы могут иметь вид пластинок (трабекулярные) и трубочек (мультивезикулярные на срезе), а направлены они продольно или поперечно по отношению к митохондрии.
Каждая митохондрия заполнена матриксом, который на электронных микрофотографиях выглядит плотнее, чем окружающая цитоплазма. Матрикс митохондрии однородный (гомогенный), иногда мелкозернистый, различной электронной плотности. В нем выявляют тонкие нити толщиной около 2…3 нм и гранулы размером около 15…20 нм. Нити матрикса представляют собой молекулы ДНК, а мелкие гранулы - митохондриальные рибосомы. В матриксе содержатся ферменты, одна одноцепочечная, циклическая ДНК, митохондриальные рибосомы, много ионов Са 2+ .
Автономная система белкового синтеза митохондрий представлена молекулами ДНК, свободными от гистонов. ДНК короткая, имеет форму кольца (циклическая) и содержит 37 генов. В отличие от ядерной ДНК в ней практически нет некодирующих последовательностей нуклеотидов. Особенности строения и организации сближают ДНК митохондрий с ДНК бактериальных клеток. На ДНК митохондрий происходит синтез молекул РНК разных типов: информационных, трансфертных (транспортных) и рибосомальных. Информационная РНК митохондрий не подвергается сплайсингу (вырезанию участков, не несущих информационной нагрузки). Малые размеры молекул митохондриальных ДНК не могут определить синтез всех белков митохондрий. Большинство белков митохондрий находится под генетическим контролем клеточного ядра и синтезируется в цитоплазме, так как ДНК митохондрий слабо выражена и может обеспечить образование лишь части ферментов цепи окислительного фосфорилирования. Митохондриальная ДНК кодирует не более десяти белков, которые локализованы в мембранах и представляют собой структурные белки, ответственные за правильную интеграцию отдельных функциональных белковых комплексов митохондриальных мембран. Синтезируются также белки, осуществляющие транспортные функции. Такая система белкового синтеза не обеспечивает всех функций митохондрии, поэтому автономия митохондрий ограниченная и относительная.
У млекопитающих митохондрии при оплодотворении передаются лишь через яйцеклетку, а спермий привносит в новый организм ДНК ядра.
В матриксе митохондрий образуются рибосомы, отличающиеся от рибосом цитоплазмы. Они участвуют в синтезе ряда митохондриальных белков, не кодируемых ядром. Митохондриальные рибосомы имеют число седиментации 60 (в отличие от цитоплазматических с числом седиментации 80). Число седиментации - это скорость осаждения при центрифугировании и ультрацентрифугировании. По строению митохондриальные рибосомы близки к рибосомам прокариотических организмов, но меньшего размера и отличаются чувствительностью к определенным антибиотикам (левомицетину, тетрациклину и др.).
Внутренняя мембрана митохондрии обладает высокой степенью избирательности при транспорте веществ. К ее внутренней поверхности прикрепляются тесно прилежащие друг к другу ферменты цепи окислительного фосфорилирования, белки-переносчики электронов, транспортные системы АТФ, АДФ, пируват и др. В результате тесного расположения ферментов на внутренней мембране обеспечивается высокая сопряженность (взаимосвязанность) биохимических процессов, повышающая скорость и эффективность каталитических процессов.
При электронной микроскопии выявляют грибовидные частицы, выступающие в просвет матрикса. Они обладают АТФ-синтетичной (образует АТФ из АДФ) активностью. Транспорт электронов идет по дыхательной цепи, локализованной во внутренней мембране, которая содержит четыре крупных ферментных комплекса (цитохромы). При прохождении электронов по дыхательной цепи ионы водорода откачиваются из матрикса в перимитохондриальное пространство, что обеспечивает формирование протонного градиента (помпы). Энергия этого градиента (различия в концентрации веществ и формирование мембранного потенциала) используется для синтеза АТФ и транспорта метаболитов и неорганических ионов. Содержащиеся на внутренней мембране белки-переносчики транспортируют через нее органические фосфаты, АТФ, АДФ, аминокислоты, жирные кислоты, три — и дикарбоновые кислоты.
Наружная мембрана митохондрии более проницаема для низкомолекулярных веществ, так как в ней много гидрофильных белковых каналов. На наружной мембране располагаются специфические рецепторные комплексы, через которые белки из матрикса транспортируются в перимитохондриальное пространство.
По своему химическому составу и свойствам наружная мембрана близка к другим внутриклеточным мембранам и плазмолемме. В ней содержатся ферменты, метаболизирующие жиры, активирующие (катализирующие) превращения аминов, аминооксидаза. Если ферменты наружной мембраны сохраняют активность, то это показатель функциональной сохранности митохондрий.
В митохондриях имеются два автономных субкомпартмента. Вели перимитохондриальное пространство, или наружная камера митохондрии (внешний субкомпартмент), формируется за счет проникновения белковых комплексов гиалоплазмы, то внутренний субкомпартмент (матрикс митохондрии) частично образован за счет синтетической активности митохондриальной ДНК. Во внутреннем субкомпартменте (матриксе) содержатся ДНК, РНК и рибосомы. Он отличается высоким уровнем ионов Са 2+ в сравнении с гиалоплазмой. Во внешнем субкомпартменте накапливаются ионы водорода. Ферментативная активность внешнего и внутреннего субкомпартментов, состав белков сильно различаются. Внутренний субкомпартмент имеет более высокую электронную плотность, чем внешний.
Специфические маркеры митохондрий - ферменты цитохром-оксидаза и сукцинатдегидрогеназа, выявление которых позволяет количественно охарактеризовать энергетические процессы в митохондриях.
Основная функция митохондрий - синтез АТФ. Вначале в гиалоплазме разрушаются сахара (глюкоза) до молочной и пировиноградной кислот (пирувата) с одновременным синтезом небольшого количества АТФ. В результате гликолиза одной молекулы глюкозы используется две молекулы АТФ, а образуется четыре. Таким образом, положительный баланс составляют всего две молекулы АТФ. Эти процессы совершаются без кислорода (анаэробный гликолиз).
Все последующие этапы выработки энергии происходят в процессе аэробного окисления, который обеспечивает синтез большого количества АТФ. При этом органические вещества разрушаются до СO 2 и воды. Окисление сопровождается переносом протонов на их акцепторы. Эти реакции осуществляются с помощью ряда ферментов цикла трикарбоновых кислот, которые находятся в матриксе митохондрии.
В мембраны крист встроены системы переноса электронов и сопряженного с ним фосфорилирования АДФ (окислительное фосфорилирование). При этом происходит перенос электронов от одного белка-акцептора электронов к другому и, наконец, связывание их с кислородом, вследствие чего образуется вода. Одновременно с этим часть энергии, выделяемой при таком окислении в цепи переноса электронов, запасается в виде макроэргической связи при фосфорилировании АДФ, что приводит к образованию большого числа молекул АТФ - основного внутриклеточного энергетического эквивалента. На мембранах крист митохондрий происходит процесс окислительного фосфорилирования с помощью расположенных здесь белков цепи окисления и фермента фосфорилирования АДФ АТФ-синтетазы. В результате окислительного фосфорилирования из одной молекулы глюкозы образуется 36 молекул АТФ.
К некоторым гормонам и веществам на митохондриях имеются специализированные (аффинные) рецепторы. Трийодтиронин в норме ускоряет синтетическую активность митохондрий. Интерлейкин-1 и высокие концентрации трийодтиронина разобщают цепи окислительного фосфорилирования, вызывают набухание митохондрий, что сопровождается увеличением образования тепловой энергии.
Новые митохондрии образуются путем деления, перетяжкой или почкованием. В последнем случае образуется протомитохондрия, постепенно увеличивающаяся в размерах.
Протомитохондрия - мелкая органелла с наружной и внутренней мембранами. Внутренняя мембрана не имеет или содержит слаборазвитые кристы. Органелла характеризуется низким уровнем аэробного фосфорилирования. При образовании перетяжки содержимое митохондрии распределяется между двумя новыми довольно крупными органеллами. При любом способе размножения в каждой из вновь образующихся митохондрий имеется собственный геном.
Старые митохондрии разрушаются путем аутолиза (самопереваривания клеткой с помощью лизосом) с образованием аутолизосом. Из аутолизосомы образуется остаточное тельце. При полном переваривании содержимое остаточного тельца, состоящее из низкомолекулярных органических веществ, выводится путем экзоцитоза. При неполном переваривании остатки митохондрий могут накапливаться в клетке в виде слоистых телец или гранул с нипофусцином. В части митохондрий накапливаются нерастворимые соли кальция с образованием кристаллов - кальцинатов. Накопление продуктов дегенерации митохондрий может привести к дистрофии клетки.
Митохондрии (МТ) – одно из самых интересных мне направлений исследований. Объединение митохондрий с другой клеткой в ходе эндосимбиоза около 1,6 млрд лет назад стало основной всех многоклеточных эукариотов со сложной структурой. Предположительно митохондрии произошли от клеток, напоминающих α-протеобактерии.
Лучшее обзорное исследование последнего времени по митохондриях – работа Вернера Кулбрандта «Структура и функция митохондриальных белковых комплексов мембраны ». Если вы знаете английский язык и интересуетесь устройством этих органелл, то настоятельно рекомендую к прочтению. Эта статья так хороша, что может быть смело главой хорошего учебника по молекулярной биологии. Сначала я хотел перевести всю статью, но это бы заняло непростительно много времени и оторвало бы от других дел. Поэтому ограничусь тезисами и картинками. Периодически разбавляя все своими мыслями.
Митохондрия кодирует сама только 13 белков, не смотря на наличие отдельной от клетки ДНК (мтДНК) и всего «производственного» цикла по транскрипции белков. Изолированная митохондрия какое-то время может сохранять композицию и функционировать.
Рисунок 1. Компоненты мембраны митохондриона. Внешняя мембрана отделяет митохондрию от цитоплазмы. Она окружает внутреннюю мембрану, которая отделяет межмембранное пространство от богатого белками центрального матрикса. Внутреннюю мембрану разделяют на внутреннюю пограничную мембрану и кристы. Две эти части непрерывны в местах крепления крист (cristae junction ). Кристы простираются более или менее глубоко в матрикс и являются основным место митохондриального преобразования энергии. Небольшой протоновый градиент в межмембранном пространстве (pH 7,2-7,4) и матрикс (pH 7,9-8,0) приводят к образованию АТФ АТФ-синтазой в мембранах крист.
Внешняя мембрана пористая и позволяет веществам из цитоплазмы проходить через нее. Внутренняя мембрана плотная, для ее пересечения нужны транспортные белки [Гилберт Линг обоснованно не согласен], непрерывность барьера позволяет иметь внутренней мембране электрохимический потенциал в -180 mV. У матрикса довольно большой pH (7,9-8). Еще раз углублюсь в Линга. Щелочной (выше 7) pH способствует более развернутой конформации белков . Высокий pH нарушает водородные и солевые связи, делая поляризованные CO и NH доступными молекулам воды, там самым усиливая дипольный момент всей внутриклеточной воды и связывая ее. В этом ключе наличие мембраны нужно не для «удержания» протоплазмы внутри клетки (это делают сами белки при высоком pH), а для наличия потенциала.
мтДНК находится в нуклеотидах, которых примерно 1000 на клетку. Белковая плотность матрикса довольна высокая (до 500 мг/мл), что близко к кристаллизованным белкам.
Внутренняя мембрана образует инвагинации, называемые кристами, которые глубоко проникают в матрикс. Кристы определяют третий «отсек» митохондрий – просвет крист (cristae lumen). Мембраны кристы содержат большинство, если не все, полностью «собранные» комплексы цепи переноса электронов и АТФ-синтазы. Просвет кристы содержит большое количество маленького растворимого белкового переносчика электронов (цитохром с). Митохондриальные кристы, таким образом, основное место биологической конверсии энергии во всех не фотосинтетических эукариотах.
С кристами тоже много всего интересного. Оптические свойства кристы влияют на распространение и генерацию света в тканях. Я даже встречал идеи о том, что поверхность крист подобна (предположение) поверхностям топологических изоляторов (подразумевалась суперпроводимость без диссипации заряда).

Рисунок 2. Мембранные белковые комплексы дыхательной цепи. Комплекс I (NADH / убихинон оксидоредуктаза, синий), Комплекс II (сукцинат дегидрогеназа, розовы), Комплекс III (цитохром С редуктаза, оранжевый), Комплекс IV (цитохром С оксидаза, зеленый) и митохондриальная АТФ синтаза (известная как комплекс V , бежевая) работают вместе во время окислительного фосфорилирования, чтобы клетки могли использовать энергию. Комплексы I , III , IV выкачивают протоны вдоль мембраны кристы, создавая протоновый градиент, стимулирующий синтез АТФ.
Теперь немного внимания на комплекс II. Вы помните, что жир (кето) делает упор в метаболизме на FADH2 и комплекс II. Они восстанавливают пару CoQ, в какой-то момент окисленного CoQ не хватает для транспортировки электронов на комплекс III и образует обратный поток электронов на комплекс I с образованием супероксида. При долгом HFLC-питании комплекс I будет обратимо разрушен, при этом это нормальная физиологическая оптимизация.
Еще прошу вас заметить, что комплекс II не выкачивает протоны. Что у нас рассеивает протоновый градиент, нарушает фосфорилирование и стимулирует сжигание жиров на тепло? Правильно, стресс холода. Термогенез связан с метаболизмом через комплекс, который не выкачивает протоны, тем самым не давая дополнительных протонов для АТФ-синтазы. Можно только удивляться как замечательно у нас продуман организм.
Крепления крист и MICOS
Места крепления крист (cristae junctions) – маленькие круглы отверстия примерно 25 нм диаметром. В митохондриях всех организмов есть система MICOS (mitochondria contact site and cristae to outer membrane), сборка из пяти мембранных и одного растворимого белков, прикрепляющих кристы к наружной мембране.
В клетках с повышенной потребностью в энергии, такие как скелетные и сердечные мышцы, кристы плотно заполняют большую часть объема митохондрии. В тканях с меньшими потребностями в энергии, таких как печень и почки, кристы находятся не так плотно по отношению друг к другу. Остается больше места в матрице для биосинтетических ферментов.
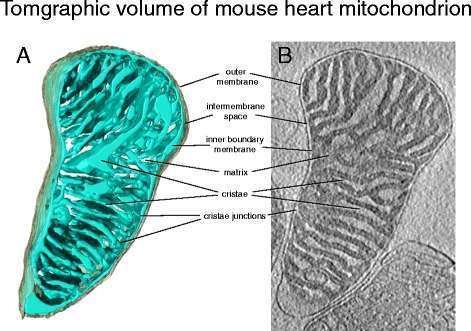
Рисунок 3. Томографический объем митохондрии сердца мыши. А) Трехмерный объем митохондрии сердца мыши, снятый cryo-ET. Наружная мембрана (серая) окутывает внутреннюю мембрану (светло-синяя). Внутренняя мембрана плотно наполнена кристами б) Томографический срез объема. Плотно заполненный матрикс, содержащий большую часть митохондриальных белков, выглядит темным на электронном микроскопе. В то время как межмембранное пространство и просветы крист выглядят светлыми из-за низкой концентрации белков.
Димеры АТФ синтазы
Митохондриальная F1-F0 АТФ синтаза является самым заметным белковым комплексом кристы. АТФ синтаза – это древняя наномашина, которая использует электрохимический протновых градиент вокруг внутренней мембраны для создания АФТ посредством вращательного катализа . Протоны, двигающиеся через F0 комплекс мембраны, вращают ротор из 8 (у млекопитающих) или 10 (у дрожжей) с-узлов. Центральный стебель передает крутящий момент c-ротора каталитической головке F1, где АТФ образуется из АДФ и фосфата через последовательность конформационных изменения. Периферийный стебель предотвращает непродуктивное вращение Головы F1 против комплекса F0.
Многие годы считалось, что АТФ синтаза случайным образом располагается на внутренней мембране. Но оказалось, что АТФ синтаза располагается двойными рядами . Причем линейные ряды АТФ синтазы – фундаментальный атрибут всех живых митохондрий.

Рисунок 4. Двойные ряды АТФ синтазы у семи разных видов.
Ряды АТФ синтазы располагаются в основном вдоль хребтов крист. Димеры изгибают липидный бислой и как следствие само-организуются в ряды. Когда у митохондрий дрожжей выбивали узлы e и g АФТ синтазы, то штамм рос на 60% медленней диких собратьев, и потенциал мембран их митохондрий был снижен вдвое. У АФТ синтазы прокариотов недостает нескольких узлов, связанных с димерами, ряды димеров не были найдены у бактерий и архей. Кристы и ряды димеров АФТ синтазы, таким образом, являются адаптацией к большим энергетическим потребностям организма.

Рисунок 5. Структура димера АТФ синтазы митохондрии polymella sp. Вид сбоку на V-образный димер АТФ синтазы.
Комплексы и суперкомплексы дыхательной цепи
Протоновый градиент вокруг внутренней мембраны создается тремя крупными мембранными комплексами, известными как комплекс I, комплекс III и комплекс IV (см. рисунок 2). Комплекс I кормится электронами из NADH, высвобождаемая при передаче электрона энергия выкачивает четыре протона. Комплекс III получает электрон от восстановленного хинола и передает его носителю электронов (цитохрому с), выкачивая в процессе один протон. Комплекс IV получает электрон из цитохрома с и передает его молекулярному кислороду, выкачивая 4 протона за каждую молекулу кислорода, превращенную в воду. Комплекс II не выкачивает протоны, напрямую передавая электроны хинолу. Как перенос электронов из NADH в хинол связан с транслокацией протонов пока не ясно. Комплекс I – крупнее III и IV вместе взятых.

Рисунок 6. Комплекс I митохондрии коровьего сердца. Матриксная часть содержит ряд из восьми железно-серных (Fe-S) кластеров, которые направляют электроны из NADH в хинол на пересечении матрикса и мембраны. Мембранная часть состоит из 78 лопастей, включая выкачивающие протоны молекулы.
Комплексы I, III и IV соединяются в суперкомплексы или респирасомы. У пекарских дрожжей (saccharomyces cerevisiae ) нет комплекса I, их суперкомплексы состоят из III и IV. Роль суперкомплексов пока еще не ясна. Предполагают, что это делает транспорт электронов более эффективным, но прямых доказательств этому пока нет.

Рисунок 7. Суперкомплекс митохондрии коровьего сердца. Обратите внимание на дистанцию между комплексами I и III, который надо проделать хинолу. Стрелки – движения электрона в суперкомплексе.
Основным белком просвета кристы является цитохром с, который переносит электрон из комплекса III в комплекс IV. Если цитохром с высвобождается в цитоплазму клетки, то вызывает апоптоз .

Рисунок 8. Ряды димеров АТФ синтазы задают форму кристам. У хребта кристы АФТ синтаза (желтый) образует слив для протонов (красный), протоновые насосы электронной цепи (зеленый) находятся по обоим сторона рядов димеров. Направляя протоны от источника к АТФ синтазе, кристы работают как протоновые направляющие, позволяющие эффективное производство АТФ. Красные стрелки показывают направление потока протонов.
Реорганизация мембраны во время старения
Старение – фундаментальный и плохо понимаемый процесс всех эукариотов. Исследовали старение митохондрий на грибах Podospora anserina , которые живут всего 18 дней. В нормальной митохондрии кристы проникают глубоко в матрикс. Для этого нужны ряды димеров АТФ синтазы и MICOS комплекс у мест крепления крист. С возрастом кристы начинают все ближе подходить к поверхности мембраны, димеры АФТ синтазы превщаются в мономеры, и все заканчивается высвобождением цитохрома с и клеточной смертью.
Транспорт электронов создает супероксид в комплексах I и III. Это побочный продукт метаболизма. Одновременно необходимый и смертельно опасный. Во время старения деление (fission) начинает превалировать над сращением (fussion). Это не дает поврежденным митохондриям «спастись» путем сращения и ускоряет неизбежное.

Рисунок 9. Изменения морфологии внутренней мембраны и димеров АТФ синтазы во время старения митохондрии.
Как видите, полей для будущих исследований очень много. Я предполагаю стык физики и биологии, где физики будут пытаются объяснить почему такая структура более энергетически эффективна. Тем более публикации по квантовой биологии
Митохондрии
Митохондрии - это структуры палочковидной или овальной формы (греч. mitos - нить, chondros - гранула). Они обнаружены во всех животных клетках (исключая зрелые эритроциты): у высших растений, у водорослей и простейших. Отсутствуют они только у прокариот бактерий.
Эти органеллы впервые были обнаружены и описаны в конце прошлого столетия Альтманом. Несколько позже эти структуры были названы митохондриями. В 1948 г. Хогебум указал на значение митохондрий как центра клеточного дыхания, а в 1949 г. Кеннеди и Ленинджер установили, что в митохондриях протекает цикл окислительного фосфорилирования. Так было доказано, что митохондрии служат местом генерирования энергии.
Митохондрии видны в обычном световом микроскопе при специальных методах окраски. В фазово - контрастном микроскопе и в «темном поле» их можно наблюдать в живых клетках.
Строение, размеры, форма митохондрий очень вариабельны. Это зависит в первую очередь от функционального состояния клеток. Например, установлено, что в мотонейронах мух, летающих непрерывно 2 часа, проявляется огромное количество шаровидных митохондрий, а у мух со склеенными крыльями число митохондрий значительно меньше и они имеют палочковидную форму (Л. Б. Левинсон). По форме они могут быть нитевидными, палочковидными, округлыми и гантелеобразными даже в пределах одной клетки.
Митохондрии локализованы в клетке, как правило, либо в тех участках, где расходуется энергия, либо около скоплений субстрата (например, липидных капель), если таковые имеются.
Строгая ориентация митохондрий обнаруживается вдоль жгутиков сперматозоидов, в поперечно-полосатой мышечной ткани, где они располагаются вдоль миофибрилл, в эпителии почечных канальцев локализуются во впячиваниях базальной мембраны и т.д.
Количество митохондрий в клетках имеет органные особенности, например, в клетках печени крыс содержится от 100 до 2500 митохондрий, а в клетках собирательных канальцев почки - 300, в сперматозоидах различных видов животных от 20 до 72, у гигантской амебы Chaos chaos их число достигает 500 000. Размеры митохондрий колеблются от 1 до 10 мкм.
Ультрамикроскопическое строение митохондрий однотипно, независимо от их формы и размера. Они покрыты двумя липопротеидными мембранами: наружной и внутренней. Между ними располагается межмембранное пространство.
Впячивания внутренней мембраны, которые вдаются в тело митохондрий, называются кристами . Расположение крист в митохондриях может быть поперечным и продольным. По форме кристы могут быть простыми и разветвленными. Иногда они образует сложную сеть. В некоторых клетках, например, в клетках клубочковой зоны надпочечника кристы имеют вид трубочек. Количество крист прямо пропорционально интенсивности окислительных процессов, протекающих в митохондриях. Например, в митохондриях кардиомиоцитов их в несколько раз больше, чем в митохондриях гепацитов. Пространство, ограниченное внутренней мембраной, составляет внутреннюю камеру митохондрий. В нем между кристами находится митохондриальный матрикс - относительно электронно плотное вещество.
Белки внутренней мембраны синтезируются миторибосомами, а белки внешней мембраны - циторибосомами.
"Наружная мембрана митохондрий по многим показателям сходна с мембранами ЭПС. Она бедна окислительными ферментами. Немного их и в мембранном пространстве. Зато внутренняя мембрана и митохондриальный матрикс буквально насыщены ими. Так, в матриксе митохондрий сосредоточены ферменты цикла Кребса и окисления жирных кислот. Во внутренней мембране локализована цепь переноса электронов, ферменты фосфорилирования (образования АТФ из АДФ), многочисленные транспортные системы.
Кроме белка и липидов, в состав мембран митохондрий входит РНК, ДНК, последняя обладает генетической специфичностью, и по своим физико-химическим свойствам отличается от ядерной ДНК.
При электронно-микроскопических исследованиях обнаружено, что поверхность наружной мембраны покрыта мелкими шаровидными элементарными частицами. Внутренняя мембрана и кристы содержат подобные элементарные частицы на «ножках», так называемые грибовидные тельца. Они -состоят из трех частей: головки сферической формы (диаметр 90-100 А°), ножки цилиндрической формы, длиной 5 нм и шириной 3-4 нм, основания, имеющего размеры 4 на 11 нм. Головки грибовидных телец связаны с фосфорилированием, затем обнаружено, что головки содержат фермент, обладающий АТФ-идной активностью.
В межмембранном пространстве находится вещество, обладающее более низкой электронной плотностью, чем матрикс. Оно обеспечивает сообщение между мембранами и поставляет для ферментов, находящихся в обеих мембранах, вспомогательные катализаторы-коферменты.
В настоящее время известно, что наружная мембрана митохондрий хорошо проницаема для веществ, имеющих низкий молекулярный вес, в частности, белковых соединений. Внутренняя мембрана митохондрий обладает избирательной проницаемостью. Она практически непроницаема для анионов (Cl -1 , Br -1 , SO 4 -2 , HCO 3 -1 , катионов Sn +2 , Mg +2 , ряда cахаров и большинства аминокислот, тогда как Са 2+ , Мп 2+ , фосфат, многокарбоновые кислоты легко проникают через нее. Имеются данные о наличии во внутренней мембране нескольких переносчиков, специфических к отдельным группам проникающих анионов и катионов. Активный транспорт веществ через мембраны осуществляется благодаря использованию энергии АТФ-азной системы или электрического потенциала, генерируемого на мембране в результате работы дыхательной цепи. Даже АТФ, синтезированная в митохондриях, может выйти с помощью переносчика (сопряженный транспорт).
Матрикс митохондрий представлен мелкозернистым электронно-плотным веществом. В нем располагаются миторибосомы, фибриллярные структуры, состоящие из молекул ДНК и гранул, имеющих диаметр более 200А ◦ образованные солями: Ca 3 (PO 4) , Ba 3 (PO 4) 2 , Mg 3 (PO 4) . Полагают, что гранулы служат резервуаром ионов Са +2 и Мg +2 . Их количество увеличивается при изменении проницаемости митохондриальных мембран.
Присутствие в митохондриях ДНК обеспечивает участие митохондрий в синтезе РНК и специфических белков, а также указывает на существование цитоплазматической наследственности. Каждая митохондрия содержит в зависимости от размера одну или несколько молекул ДНК (от 2 до 10). Молекулярный вес митохондриальной ДНК около (30-40)*10 6 у простейших, дрожжей, грибов. У высших животных около (9–10) *10 6.
Длина ее у дрожжей примерно равна 5 мкм, у растений - 30 мкм. Объем генетической информации, заключенный в митохондриальной ДНК, невелик: он состоит из 15-75 тыс. пар оснований, которые могут кодировать в среднем 25-125 белковых цепей с молекулярным весом около 40000.
Митохондриальная ДНК отличается от ядерной ДНК рядом особенностей: более высокой скоростью синтеза (в 5-7 раз), она более устойчива к действию ДНК-азы, представляет собой двухкольцевую молекулу, содержит больше гуанина и цитозина, денатурируется при более высокой температуре и легче восстанавливается. Однако не все митохондриальные белки синтезируются митохондриальной системой. Так, синтез цитохрома С и других ферментов обеспечивается информацией, содержащейся в ядре. В матриксе митохондрий локализованы, витамины А, В 2 , В 12 , К, Е, а также гликоген.
Функция митохондрий заключается в образовании энергии, необходимой для жизнедеятельности клеток. Источником энергии в клетке могут служить различные соединения: белки, жиры, углеводы. Однако единственным субстратом, который немедленно включается в энергетические процессы, является глюкоза.
Биологические процессы, в результате которых в митохондриях образуется энергия, можно подразделить на 3 группы: I группа - окислительные реакции, включающие две фазы: анаэробную (гликолиз) и аэробную. II группа - дефосфорилирование, расщепление АТФ и высвобождение энергии. III группа - фосфорилирование, сопряженное с процессом окисления.
Процесс окисления глюкозы вначале происходит без участия кислорода (анаэробным или гликолитическим путем) до пировиноградной или молочной кислоты.
Однако при этом энергии выделяется лишь небольшое количество. В дальнейшем эти кислоты вовлекаются в процессы окисления, которые протекают с участием кислорода, т. е. являются аэробными. В результате процесса окисления пировиноградной и молочной кислоты, названной циклом Кребса, образуется углекислый газ, вода и большое количество энергии.
Образующаяся энергия не выделяется в виде тепла, что привело бы к перегреванию клеток и гибели всего организма, а аккумулируется в удобной для хранения и транспорта форме в виде аденозинтрифосфорной кислоты (АТФ). Синтез АТФ происходит из АДФ и фосфорной кислоты и вследствие этого называется фосфорилированием .
В здоровых клетках фосфорилирование сопряжено с окислением. При заболеваниях сопряженность может разобщаться, поэтому субстрат окисляется, а фосфорилирование не происходит, и окисление переходит в тепло, а содержание АТФ в клетках снижается. В результате повышается температура и падает функциональная активность клеток.
Итак, основная функция митохондрий заключается в выработке практически всей энергии клетки и происходит синтез компонентов, необходимых для деятельности самого органоида, ферментов «дыхательного ансамбля», фосфолипидов и белков.
Еще одной стороной деятельности митохондрий является их участие в специфических синтезах, например, в синтезе стероидных гормонов и отдельных липидов. В ооцитах разных животных образуются скопления желтка в митохондриях, при этом они утрачивают свою основную систему. Отработавшие митохондрии могут накапливать также продукты экскреции.
В некоторых случаях (печень, почки) митохондрии способны аккумулировать вредные вещества и яды, попадающие в клетку, изолируя их от основной цитоплазмы и частично блокируя вредное действие этих веществ. Таким образом, митохондрии способны брать на себя функции других органоидов клетки, когда это требуется для полноценного обеспечения того или иного процесса в норме или в экстремальных условиях.
Биогенез митохондрий. Митохондрии представляют собой обновляющиеся структуры с довольно кратким жизненным циклом (в клетках печени крысы, например, период полужизни митохондрий охватывает около 10 дней). Митохондрии образуются в результате роста и деления предшествующих митохондрий. Деление их может происходить тремя способами: перетяжкой, отпочковыванием небольших участков и возникновением дочерних митохондрий внутри материнской. Делению (репродукции) митохондрий предшествует репродукция собственной генетической системы - митохондриальной ДНК.
Итак, согласно взглядам большинства исследователей, образование митохондрий происходит преимущественно путем саморепродукции их de novo.
