Строение. Поверхностный аппарат митохондрий состоит из двух мембран - наружной и внутренней. Внешняя мембрана гладкая, она отделяет митохондрию от гиалоплазмы. Под ней находится складчатая внутренняя мембрана, которая образует кристи (гребни). С обеих сторон крист обнаружены мелкие грибовидные тельца, называемые оксисомамы, или АТФ-сомами. Они содержат ферменты, участвующие в окислительном фосфорилирования (присоединении фосфатных остатков к АДФ с образованием АТФ). Количество крист в митохондриях связана с энергетическими потребностями клетки, в частности в мышечных клетках митохондрии содержат очень большое количество крист. При повышенной функции клетки митохондрии приобретают более овальной или удлиненной формы, и количество крист у них растет.
Митохондрии имеют собственный геном, их рибосомы типа 70S отличаются от рибосом цитоплазмы. ДНК митохондрий преимущественно имеет циклическую форму (плазмиды), кодирует все три вида собственных РНК и поставляет информацию для синтеза части митохондриальных белков (около 9%). Итак, митохондрии можно считать полуавтономными органелл. Митохондрии относятся к саморепликуючих (способных к размножению) органелл. Обновления митохондрий происходит в течение всего клеточного цикла. Например, в клетках печени они заменяются новыми спустя почти 10 дней. Наиболее вероятным путем воспроизведения митохондрий считают их разделение: посередине митохондрии появляется перетяжка или возникает перегородка, после чего органеллы распадаются на две новые митохондрии. Образуются митохондрии с промитохондрий - округлых телец диаметром до 50 нм с двойной мембраной.
Функции . Митохондрии участвуют в энергетических процессах клетки, они содержат ферменты, связанные с образованием энергии и клеточным дыханием. Иными словами, митохондрия является своеобразной биохимической мини-фабрикой, которая превращает энергию органических соединений на прикладное энергию АТФ. В митохондриях энергетический процесс начинается в матриксе, где происходит расщепление пировиноградной кислоты в цикле Кребса. Во время этого процесса освобождаются атомы водорода, которые транспортируются дыхательным цепью. Энергия, которая при этом высвобождается, используется в нескольких участках дыхательной цепи для осуществления реакции фосфорилирования - синтеза АТФ, то есть присоединения фосфатной группы к АДФ. Это происходит на внутренней мембране митохондрий. Итак, энергетическая функция митохондрий интегрируется с: а) окисления органических соединений, что происходит в матриксе, благодаря чему митохондрии называют дыхательным центром клеток, б) синтеза АТФ, осуществляется на кристах, благодаря чему митохондрии называют энергетическими станциями клеток. Кроме того, митохондрии берут участие в регуляции обмена воды, депонировании ионов кальция, продукции предшественников стероидных гормонов, в обмене веществ (например, митохондрии в клетках печени содержат ферменты, которые позволяют им обезвреживать аммиак) и другие.
БИОЛОГИЯ + Митохондриальные болезни - группа наследственных заболеваний, связанных с дефектами митохондрий, которые приводят к нарушению клеточного дыхания. Они передаются по женской линии детям обоего пола, поскольку яйцеклетка имеет больший объем цитоплазмы и, соответственно, передает потомкам и большее количество митохондрий. Митохондриальная ДНК, в отличие от ядерной, не защищена белками-гистонами, а механизмы репарации, которые достались от бактерий-предков, несовершенны. Поэтому в митохондриальной ДНК мутации накапливаются у 10-20 раз быстрее, чем в ядерной, что и приводит к митохондриальных болезней. В современной медицине их сейчас известно уже около 50. Например, синдром хронической усталости, мигрень, синдром Барта, синдром Пирсона и многие другие.
Строение и функции ядра растительной клетки.
Ядро – обязательная часть эукариотической клетки. Это место хранения и воспроизведения наследственной информации. Ядро также служит центром управления обменом веществ и почти всех процессов, происходящих в клетке. Чаще всего в клетках имеется лишь одно ядро, редко - два или несколько. Форма его чаще всего шаровидная или эллипсоидальная. В молодых, особенно меристематических, клетках оно занимает центральное положение, но позднее обычно смещается к оболочке, оттесняемое растущей вакуолью. Снаружи ядро покрыто двойной мембраной – ядерной оболочкой, пронизанной порами (поры ядра - динамичные образования, они могут открываться и закрываться; таким путем может осуществляться регуляция обмена между ядром и цитоплазмой) на краях которых наружная мембрана переходит во внутреннюю. Наружная ядерная мембрана соединяется с мембранными каналами ЭПС. На ней располагаются рибосомы. Внутренняя мембрана может давать впячивания.
Внутреннее содержимое ядра – кариоплазма с погруженными в нее хроматином и ядрышками, и рибосомами. Кариоплазма (нуклеоплазма) представляет собой желеобразный раствор, который заполняет пространство между структурами ядра (хроматином и ядрышками). Она содержит ионы, нуклеотиды, ферменты.
Хроматин – это деспирализованная форма существования хромосом. В деспирализованном состоянии хроматин находится в ядре неделящейся клетке. Хроматин и хромосомы взаимно переходят друг в друга. По химической организации как хроматин, так и хромосомы не отличаются. Химическую основу составляет дезоксирибонуклеопротеин – комплекс ДНК с белками. С помощью белков происходит многоуровневая упаковка молекул ДНК, при этом хроматин приобретает компактную форму.
Ядрышко, обычно шаровидной формы (одно или несколько), не окружено мембраной, содержит фибриллярные белковые нити и РНК. Ядрышки – не постоянные образования, они исчезают в начале деления клетки и восстанавливаются после его окончания. Ядрышки имеются только в неделящихся клетках. В ядрышках происходит формирование рибосом, синтез ядерных белков. Сами же ядрышки образуются на участках вторичных перетяжек хромосом (ядрышковых организаторах).
Ядро – обязательная часть эукариотической клетки. Диаметр ядра колеблется от 5 до 20 мкм. Главная функция ядра – хранение генетического материала в форме ДНК и передача ее дочерним клеткам при клеточном делении. Кроме того, ядро управляет белковыми синтезами, контролирует все процессы жизнедеятельности клетки. (в растительной клетке ядро описал Р.Броун в 1831г., в животной – Т.Шванн в 1838г.).
Химический состав ядра представлен, главным образом, нуклеиновыми кислотами и белками.
Строение и функции митохондрий.
Митохондрии или хондриосомы - «силовые» станции клетки, в них локализована большая часть реакций дыхания (аэробная фаза). В митохондриях происходит аккумуляция энергии дыхания в аденозинтрифосфате (АТФ). Энергия, запасаемая в АТФ, служит основным источником для физиологической деятельности клетки. Митохондрии обычно имеют удлиненную палочковидную форму длиной 4-7 мкм и диаметром 0,5-2 мкм. Число митохондрий в клетке может быть различным от 500 до 1000 и зависит от роли данного органа в процессах энергетического обмена.
Химический состав митохондрий несколько колеблется. В основном это белково-липидные органеллы. Содержание белка в них составляет 60-65%, причем структурные и ферментативные белки содержатся примерно в равной пропорции, а также около 30% липидов. Очень важно, что митохондрии содержат нуклеиновые кислоты: РНК - 1 % и ДНК -0,5%. В митохондриях имеется не только ДНК, но и вся система синтеза белка, в том числе рибосомы.
Митохондрии окружены двойной мембраной. Толщина мембран составляет 6-10 нм. Мембраны митохондрий на 70% состоят из белка. Фосфолипиды мембран представлены фосфатидтилхолином, фосфатидилэтаноламином, а также специфическими фосфолипидами, например, кардиолипином. Мембраны митохондрий не пропускают Н+ и служат барьером для их транспорта.
Между мембранами находится заполненное жидкостью перимитохондриальное пространство. Внутреннее пространство митохондрий заполняет матрикс в виде студнеобразной полужидкой массы. В матриксе сосредоточены ферменты цикла Кребса. Внутренняя мембрана дает выросты - кристы в виде пластин и трубочек, они разделяют внутреннее пространство митохондрий на отдельные отсеки. Во внутренней мембране локализована дыхательная цепь (цепь переноса электронов).
Основной функцией митохондрий является синтез АТФ - универсальной формы химической энергии в любой живой клетке. Как и у прокариот, данная молекула может образовываться двумя путями: в результате субстратного фосфорилирования в жидкой фазе (например, при гликолизе) или в процессе мембранного фосфорилирования, связанного с использованием энергии трансмембранного электрохимического градиента (англ.)русск. протонов (ионов водорода). Митохондрии реализуют оба эти пути, первый из которых характерен для начальных процессов окисления субстрата и происходит в матриксе, а второй завершает процессы энергообразования и связан с кристами митохондрий. При этом своеобразие митохондрий как энергообразующих органелл эукариотической клетки определяет именно второй путь генерации АТФ, получивший название «хемиосмотического сопряжения». По сути это последовательное превращение химической энергии восстанавливающих эквивалентов НАДН в электрохимический протонный градиент ΔμН + по обе стороны внутренней мембраны митохондрии, что приводит в действие мембранно-связанную АТФ-синтетазу и завершается образованием макроэргической связи в молекуле АТФ.
В целом весь процесс энергообразования в митохондриях может быть разбит на четыре основные стадии, первые две из которых протекают в матриксе, а две последние - на кристах митохондрий:
- Превращение поступивших из цитоплазмы в митохондрию пирувата и жирных кислот в ацетил-СоА;
- Окисление ацетил-СоА в цикле Кребса, ведущее к образованию НАДН;
- Перенос электронов с НАДН на кислород по дыхательной цепи;
- Образование АТФ в результате деятельности мембранного АТФ-синтетазного комплекса.
Ещё в цитоплазме в серии из 10 отдельных ферментативных реакций шестиуглеродная молекула глюкозы частично окисляется до двух трёхуглеродных молекул пирувата с образованием двух молекул АТФ. Затем пируват переносится из цитозоля через наружную и внутреннюю мембраны в матрикс, где первоначально превращается в ацетил-СоА. Этот процесс катализируется крупным пируватдегидрогеназным комплексом, имеющим размер, сопоставимый с размером рибосомы, и состоящим из трёх ферментов, пяти коферментов и двух регуляторных белков. Точно также жирные кислоты, полученные при расщеплении нерастворимых триглицеридов в цитоплазме, переносятся в митохондриальный матрикс в виде ацетил-СоА-производных.
На следующем этапе, также протекающем в матриксе митохондрии, ацетил-СоА полностью окисляется в цикле Кребса. В его работе задействованы четыре отдельных фермента, за каждый цикл обеспечивающие укорочение углеводородной цепи на два атома углерода, которые в дальнейшем превращаются в СО 2 . Этот процесс обеспечивает образование одной молекулы АТФ, а также НАДН - высокоэнергетического промежуточного соединения, которое легко отдаёт электроны в цепь переноса электронов на кристах митохондрий.
Дальнейшие процессы энергообразования в митохондрии происходят на её кристах и связаны с переносом электронов от НАДН к кислороду. В соответствии с тем, что потребление кислорода в качестве окислителя обычно называют «внутриклеточным дыханием», электронно-транспортную цепь ферментов, осуществляющих последовательный перенос электронов от НАДН к кислороду, часто называют «дыхательной цепью». При этом трансформация энергии окисления осуществляется ферментами, расположенными на кристах митохондрий и осуществляющими векторный (направленный по отношению к сторонам мембраны) перенос протонов водорода из матрикса митохондрии в межмембранное пространство. В этом состоит принципиальное отличие работы оксидоредуктаз дыхательной цепи от функционирования ферментов, катализирующих реакции в гомогенном (изотропном) растворе, где вопрос о направлении реакции в пространстве не имеет смысла.
Весь процесс переноса электрона по дыхательной цепи может быть разбит на три стадии, каждая из которых катализируется отдельным трансмембранным липопротеидным комплексом (I, III и IV), встроенным в мембрану кристы митохондрии. В состав каждого из названных комплексов входят следующие компоненты:
- Большой олигомерный фермент, катализирующий перенос электронов;
- Небелковые органические (простетические) группы, принимающие и высвобождающие электроны;
- Белки, обеспечивающие движение электронов.
Каждый из этих комплексов осуществляет перенос электронов от донора к акцептору по градиенту редокс-потенциала через ряд последовательно функционирующих переносчиков. В качестве последних в дыхательной цепи митохондрий функционируют мигрирующие в плоскости мембраны жирорастворимые молекулы убихинона, а также небольшие (молекулярная масса 13кДа) водорастворимые белки, содержащие ковалентно связанный гем и называемые «цитохромами с ». При этом три из пяти компонентов, составляющих дыхательную цепь, работают так, что перенос электронов сопровождается переносом протонов через мембрану крист митохондрий в направлении из матрикса в межмембранное пространство.
Дыхательная цепь начинается с комплекса I (НАДН-убихинон-оксидоредуктаза), состоящего из 16-26 полипептидных цепей и имеющего молекулярную массу около 850 кДа. Функциональная активность этого комплекса определяется тем, что он содержит в своём составе более 20 атомов железа, упакованных в ячейки из атомов серы, а также флавин (Фл - производное витамина рибофлавина). Комплекс I катализирует окисление НАДН, отщепляя от него два электрона, которые после «путешествия» по окислительно-восстановительным компонентам комплекса I попадают на молекулу-переносчик, в качестве которой выступает убихинон (Q). Последний способен ступенчато восстанавливаться, принимая на себя по два электрона и протона и, таким образом, превращаясь в восстановленную форму - убихинол (QH 2).
Энергетический потенциал (запас энергии) в молекуле убихинола существенно ниже, чем в молекуле НАДН, а разница в подобной энергии временно запасается в виде особого вида - электрохимического протонного градиента. Последний возникает в результате того, что перенос электронов по простетическим группам комплекса I, ведущий к снижению энергетического потенциала электронов, сопровождается трансмембранным переносом двух протонов из матрикса в межмембранное пространство митохондрии.
Восстановленный убихинол мигрирует в плоскости мембраны, где достигает второго фермента дыхательной цепи - комплекса III (bc 1 ). Последний представляет собой димер из субъединиц b и c 1 с молекулярной массой более 300 кДа, сформированный из восьми полипептидных цепей и содержащий атомы железа как в серных ячейках, так и в виде комплексов с гемами b (I), b (II) и c 1 - сложными гетероциклическими молекулами с четырьмя атомами азота, расположенными по углам металлосвязывающего квадрата. Комплекс III катализирует реакцию восстановления убихинола до убихинона с передачей электронов на атом железа второй молекулы переносчика (находящегося в межмембранном пространстве цитохрома c ). Отщепляющиеся при этом от убихинола два протона водорода освобождаются в межмембранное пространство, продолжая формирование электрохимического градиента. Наконец, ещё два протона водорода переносятся в межмембранное пространство митохондрии за счёт энергии электронов, проходящих по простетических группам комплекса III.
Последняя стадия катализируется комплексом IV (цитохром c -оксидаза) с молекулярной массой около 200 кДа, состоящим из 10-13 полипептидных цепей и, помимо двух различных гемов, включающим также несколько атомов меди, прочно связанных с белками. При этом электроны, отбираемые у восстановленного цитохрома c , пройдя по атомам железа и меди в составе комплекса IV, попадают на связанный в активном центре этого фермента кислород, что приводит к образованию воды.
Таким образом, суммарная реакция, катализируемая ферментами дыхательной цепи, состоит в окислении НАДН кислородом с образованием воды. По сути этот процесс заключается в ступенчатом переносе электронов между атомами металлов, присутствующих в простетических группах белковых комплексов дыхательной цепи, где каждый последующий комплекс обладает более высоким сродством к электрону, чем предыдущий. При этом сами электроны передаются по цепи до тех пор, пока не соединятся с молекулярным кислородом, обладающим наибольшим сродством к электронам. Освобождаемая же при этом энергия запасается в виде электрохимического (протонного) градиента по обе стороны внутренней мембраны митохондрий. При этом считается, что в процессе транспорта по дыхательной цепи пары электронов перекачивается от трёх до шести протонов.
Завершающим этапом функционирования митохондрии является генерация АТФ, осуществляемая встроенным во внутреннюю мембрану специальным макромолекулярным комплексом с молекулярной массой 500 кДа. Этот комплекс, называемый АТФ-синтетазой, как раз и катализирует синтез АТФ путём конверсии энергии трансмембранного электрохимического градиента протонов водорода в энергию макроэргической связи молекулы АТФ.
АТФ-синтаза
В структурно-функциональном плане АТФ-синтаза состоит из двух крупных фрагментов, обозначаемых символами F 1 и F 0 . Первый из них (фактор сопряжения F 1) обращён в сторону матрикса митохондрии и заметно выступает из мембраны в виде сферического образования высотой 8 нм и шириной 10 нм. Он состоит из девяти субъединиц, представленных пятью типами белков. Полипептидные цепи трёх субъединиц α и стольких же субъединиц β уложены в похожие по строению белковые глобулы, которые вместе образуют гексамер (αβ) 3 , имеющий вид слегка приплюснутого шара. Подобно плотно уложенным долькам апельсина, последовательно расположенные субъединицы α и β образуют структуру, характеризующуюся осью симметрии третьего порядка с углом поворота 120°. В центре этого гексамера находится субъединица γ, которая образована двумя протяжёнными полипептидными цепями и напоминает слегка деформированный изогнутый стержень длиной около 9 нм. При этом нижняя часть субъединицы γ выступает из шара на 3 нм в сторону мембранного комплекса F 0 . Также внутри гексамера находится минорная субъединица ε, связанная с γ. Последняя (девятая) субъединица обозначается символом δ и расположена на внешней стороне F 1 .
Мембранная часть АТФ-синтазы, называемая фактором сопряжения F 0 , представляет собой гидрофобный белковый комплекс, пронизывающий мембрану насквозь и имеющий внутри себя два полуканала для прохождения протонов водорода. Всего в состав комплекса F 0 входит одна белковая субъединица типа а , две копии субъединицы b , а также от 9 до 12 копий мелкой субъединицы c . Субъединица а (молекулярная масса 20 кДа) полностью погружена в мембрану, где образует шесть пересекающих её α-спиральных участков. Субъединица b (молекулярная масса 30 кДа) содержит лишь один сравнительно короткий погружённый в мембрану α-спиральный участок, а остальная её часть заметно выступает из мембраны в сторону F 1 и закрепляется за расположенную на её поверхности субъединицу δ. Каждая из 9-12 копий субъединицы c (молекулярная масса 6-11 кДа) представляет собой сравнительно небольшой белок из двух гидрофобных α-спиралей, соединённых друг с другом короткой гидрофильной петлёй, ориентированной в сторону F 1 , а все вместе образуют единый ансамбль, имеющий форму погружённого в мембрану цилиндра. Выступающая из комплекса F 1 в сторону F 0 субъединица γ как раз и погружена внутрь этого цилиндра и достаточно прочно зацеплена за него.
Таким образом, в молекуле АТФ-синтазы можно выделить две группы белковых субъединиц, которые могут быть уподоблены двум деталям мотора: ротору и статору. «Статор» неподвижен относительно мембраны и включает в себя шарообразный гексамер (αβ) 3 , находящуюся на его поверхности и субъединицу δ, а также субъединицы a и b мембранного комплекса F 0 . Подвижный относительно этой конструкции «ротор» состоит из субъединиц γ и ε, которые, заметно выступая из комплекса (αβ) 3 , соединяются с погружённым в мембрану кольцом из субъединиц c .
Способность синтезировать АТФ - свойство единого комплекса F 0 F 1 , сопряжённого с переносом протонов водорода через F 0 к F 1 , в последнем из которых как раз и расположены каталитические центры, осуществляющие преобразование АДФ и фосфата в молекулу АТФ. Движущей же силой для работы АТФ-синтазы является протонный потенциал, создаваемый на внутренней мембране митохондрий в результате работы цепи электронного транспорта.
Сила, приводящая в движение «ротор» АТФ-синтазы, возникает при достижении разности потенциалов между наружной и внутренней сторонами мембраны > 220 мВ и обеспечивается потоком протонов, протекающих через специальный канал в F 0 , расположенный на границе между субъединицами a и c . При этом путь переноса протонов включает в себя следующие структурные элементы:
- Два расположенных несоосно «полуканала», первый из которых обеспечивает поступление протонов из межмембранного пространства к существенно важным функциональным группам F 0 , а другой обеспечивает их выход в матрикс митохондрии;
- Кольцо из субъединиц c , каждая из которых в своей центральной части содержит протонируемую карбоксильную группу, способную присоединять H + из межмембранного пространства и отдавать их через соответствующие протонные каналы. В результате периодических смещений субъединиц с , обусловленных потоком протонов через протонный канал происходит поворот субъединицы γ, погружённой в кольцо из субъединиц с .
Таким образом, каталитическая активность АТФ-синтазы непосредственно связана с вращением её «ротора», при котором поворот субъединицы γ вызывает одновременное изменение конформации всех трёх каталитических субъединиц β, что в конечном счёте и обеспечивает работу фермента. При этом в случае образования АТФ «ротор» крутится по часовой стрелке со скоростью четыре оборота в секунду, а само подобное вращение происходит дискретными скачками по 120°, каждый из которых сопровождается образованием одной молекулы АТФ.
Непосредственная функция синтеза АТФ локализована на β-субъединицах сопрягающего комплекса F 1 . При этом самым первым актом в цепи событий, приводящих к образованию АТФ, является связывание АДФ и фосфата с активным центром свободной β-субъединицы, находящейся в состоянии 1. За счёт энергии внешнего источника (тока протонов) в комплексе F 1 происходят конформационные изменения, в результате которых АДФ и фосфат становятся прочно связанными с каталитическим центром (состояние 2), где становится возможным образование ковалентной связи между ними, ведущей к образованию АТФ. На данной стадии АТФ-синтазы ферменту практически не требуется энергии, которая будет необходима на следующем этапе для освобождения прочно связанной молекулы АТФ из ферментативного центра. Поэтому следующий этап работы фермента заключается в том, чтобы в результате энергозависимого структурного изменения комплекса F 1 каталитическая β-субъединица, содержащая прочно связанную молекулу АТФ, перешла в состояние 3, в котором связь АТФ с каталитическим центром ослаблена. В результате этого молекула АТФ покидает фермент, а β-субъединица возвращается в исходное состояние 1, благодаря чему обеспечивается цикличность работы фермента.
Работа АТФ-синтазы связана с механическими движениями её отдельных частей, что позволило отнести этот процесс к особому типу явлений, названных «вращательным катализом». Подобно тому, как электрический ток в обмотке электродвигателя приводит в движение ротор относительно статора, направленный перенос протонов через АТФ-синтетазу вызывает вращение отдельных субъединиц фактора сопряжения F 1 относительно других субъединиц ферментного комплекса, в результате чего это уникальное энергообразующее устройство совершает химическую работу - синтезирует молекулы АТФ. В дальнейшем АТФ поступает в цитоплазму клетки, где расходуется на самые разнообразные энергозависимые процессы. Подобный перенос осуществляется специальным встроенным в мембрану митохондрий ферментом АТФ/АДФ-транслоказой, который обменивает вновь синтезированную АТФ на цитоплазматическую АДФ, что гарантирует сохранность фонда адениловых нуклеотидов внутри митохондрий.
От д-ра Меркола
Митохондрии: вы можете не знать, что это такое, но они жизненно важны для вашего здоровья. Доктор наук Ронда Патрик – биомедик, которая изучила взаимодействие митохондриального метаболизма, аномального метаболизма и рака.
Часть ее работы предполагает выявление ранних биомаркеров заболевания. Например, повреждение ДНК – это ранний биомаркер рака. Затем она пытается определить, какие питательные микроэлементы помогают восстановить это повреждение ДНК.
Она также исследовала митохондриальную функцию и метаболизм, которыми и я увлекаюсь с недавних пор. Если, прослушав это интервью, вы захотите узнать об этом побольше, рекомендую начать с книги д-ра Ли Ноу «Жизнь - эпическая история наших митохондрий».
Митохондрии обладают огромным влиянием на здоровье, особенно на рак, и я начинаю верить, что оптимизация митохондриального метаболизма может лежать в основе эффективного лечения рака.
Важность оптимизации митохондриального метаболизма
Митохондрии представляют собой крошечные органеллы, которые, как первоначально считалось, мы унаследовали от бактерий. В красных кровяных тельцах и клетках кожи их почти нет, зато в зародышевых клетках их по 100 000, но в большинстве клеток их от одной до 2 000. Они – главный источник энергии для вашего организма.
Чтобы органы могли функционировать должным образом, им нужна энергия, и эта энергия вырабатывается митохондриями.
Поскольку митохондриальная функция лежит в основе всего, что происходит в организме, то оптимизация митохондриальной функции, и предотвращение нарушения функции митохондрий путем получения всех необходимых питательных веществ и прекурсоров, необходимых митохондриям, чрезвычайно важна для здоровья и профилактики заболеваний.
Так, одной из универсальных характеристик раковых клеток является серьезное нарушение функции митохондрий, при котором радикально снижено количество функциональных митохондрий.
Д-р Отто Варбург был врачом с научной степенью по химии и тесно дружил с Альбертом Эйнштейном. Большинство экспертов признают Варбурга величайшим биохимиком 20-го века.
В 1931 году он получил Нобелевскую премию – он открыл, что в качестве источника производства энергии раковые клетки используют глюкозу. Это назвали «эффектом Варбурга» но, к сожалению, это явление и по сей день игнорируется почти всеми.
Я убежден, что кетогенная диета, которая радикально улучшает здоровье митохондрий, может помочь при большинстве видов рака, особенно в сочетании с поглотителем продуктов брожения глюкозы, таким как 3-бромопируват.
Как митохондрии вырабатывают энергию
Чтобы производить энергию, митохондриям нужен кислород из воздуха, которым вы дышите, и жира и глюкоза из пищи, которую вы едите.
Эти два процесса - дыхания и приема пищи – соединяются друг с другом в процессе, который называется окислительное фосфорилирование. Именно он используется митохондриями для производства энергии в виде АТФ.
Митохондрии обладают рядом электронных транспортных цепочек, по которым они передают электроны из восстановленной формы съедаемой вами пищи, чтобы объединить их с кислородом из воздуха, которым вы дышите, и в конечном счете, образовать воду.
Этот процесс приводит протоны через митохондриальную мембрану, подзаряжая АТФ (аденозинтрифосфат) из АДФ (аденозина дифосфат). АТФ переносит энергию по всему организму
Но в ходе этого процесса образуются побочные продукты, такие как активные формы кислорода (АФК), которые повреждают клетки и митохондриальную ДНК, перенося их затем в ДНК ядра.
Таким образом, происходит компромисс. Вырабатывая энергию, организм стареет из-за возникающих в процессе разрушительных аспектов АФК. Скорость старения организма в значительной степени зависит от того, насколько хорошо функционируют митохондрии, и объема повреждений, который можно компенсировать с помощью оптимизации диеты.
Роль митохондрий при раковых заболеваниях
Когда появляются раковые клетки, активные формы кислорода, полученные в качестве побочного продукта производства АТФ, посылают сигнал, запускающий процесс клеточного самоубийства, также известный как апоптоз.
Поскольку клетки рака образуются каждый день, это хорошо. Убивая поврежденные клетки, организм избавляется от них и заменяет их здоровыми.
Раковые клетки, однако, устойчивы к этому протоколу самоубийства – у них против него встроена защита, как объяснил д-р Варбург и, впоследствии, Томас Сейфрид, который глубоко исследовал рак как заболевание обмена веществ.
Как поясняет Патрик:
«Одним из механизмов действия химиотерапевтических препаратов является образование активных форм кислорода. Они создают повреждения, и этого достаточно, чтобы подтолкнуть раковую клетку к смерти.
Думаю, причина этого в том, что раковая клетка, которая не использует свои митохондрии, то есть, больше не производит активные формы кислорода, и вдруг вы ее заставляете пользоваться митохондриями, и получается всплеск активных форм кислорода (ведь именно это делают митохондрии), и - бум, смерть, потому что раковая клетка уже готова к этой смерти. Она готова умереть».
Почему полезно не есть по вечерам
Уже довольно долгое время я – поклонник чередующегося голодания по целому ряду причин, разумеется, из соображений долголетия и здоровья, а также потому, что оно, как представляется, обеспечивает мощную профилактику рака и благотворное влияние, как от лечения. А механизм этого связан с эффектом, который голодание оказывает на митохондрии.
Как уже упоминалось, основной побочный эффект переноса электронов, в котором участвуют митохондрии, состоит в том, что некоторые утекают из цепи переноса электронов и вступают в реакцию с кислородом, образуя свободные радикалы супероксида.
Анион супероксида (результат уменьшения кислорода на один электрон), является предшественником большинства активных форм кислорода и медиатором окислительных цепных реакций. Свободные радикалы кислорода атакуют липиды клеточных мембран, белковых рецепторов, ферментов и ДНК, что может преждевременно убивать митохондрии.
Некоторые свободные радикалы, вообще-то, даже полезные, необходимые организму для регулирования клеточных функций, но при избыточном образовании свободных радикалов возникают проблемы. К сожалению, именно поэтому у большинства населения развивается большинство заболеваний, особенно рак. Решить эту проблему можно двумя способами:
- Увеличить антиоксиданты
- Уменьшить выработку митохондриальных свободных радикалов
По моему мнению, одной из наиболее эффективных стратегий снижения митохондриальных свободных радикалов является ограничение количества топлива, которым вы заправляете организм. Это совсем непротиворечащее положение, ведь ограничение калорий последовательно демонстрирует много терапевтических преимуществ. Это одна из причин эффективности чередующегося голодания, поскольку оно ограничивает период времени, в который принимается пища, что автоматически уменьшает количество калорий.
Это особенно эффективно, если не есть за несколько часов до сна, потому что это – самое метаболически низкое состояние.
Возможно, неспециалистам все это покажется слишком сложным, но следует понять одно: поскольку во время сна организм использует наименьшее количество калорий, то следует избегать еды перед сном, ведь избыточное количество топлива в это время приведет к образованию избыточного количества свободных радикалов, которые разрушают ткани, ускоряют старение и способствуют возникновению хронических заболеваний.
Как еще голодание помогает здоровью функции митохондрий
Патрик также отмечает, что частично механизм эффективности голодания объясняется тем, что энергию организм вынужден получать из липидов и запасов жира, а это означает, что клетки вынуждены использовать свои митохондрии.
Митохондрии – это единственный механизм, с помощью которых организм может создавать энергию из жира. Таким образом, голодание помогает активировать митохондрии.
Она также считает, что это играет огромную роль в механизме, с помощью которого чередующееся голодание и кетогенная диета убивают раковые клетки, и объясняет, почему некоторые препараты, активирующие митохондрии, способны убивать раковые клетки. Опять же, это потому, что образуется всплеск активных форм кислорода, ущерб от которых и решает исход дела, вызывая гибель раковых клеток.
Питание митохондрий
С точки зрения питания, Патрик подчеркивает значение следующих питательных веществ и важных сопутствующих факторов, необходимых для правильного функционирования митохондриальных ферментов:
- Коэнзим Q10 или убихинол (восстановленная форма)
- L-карнитин, который переносит жирные кислоты в митохондрии
- D-рибоза, которая является сырьем для молекул АТФ
- Магний
- Все витамины группы В, в том числе рибофлавин, тиамин и B6
- Альфа-липоевая кислота (АЛК)
Как замечает Патрик:
«Я предпочитаю получать как можно больше питательных микроэлементов из цельных продуктов по целому ряду причин. Во-первых, они образуют между собой комплекс с волокнами, благодаря которому облегчается их всасывание.
Кроме того, в этом случае обеспечивается их правильное соотношение. Получить их с избытком не удастся. Соотношение именно такое, как нужно. Есть и другие компоненты, которые, вероятно, еще предстоит определить.
Нужно быть очень бдительными, следя за тем, чтобы есть широкий спектр [продуктов] и получать правильные питательные микроэлементы. Я думаю, по этой причине полезно принимать добавки с комплексом витаминов В.
По этой причине их принимаю я. Другая причина заключается в том, что с возрастом мы перестаем так же легко усваивать витамины группы В, в основном, из-за увеличивающейся жесткости клеточных мембран. Это изменяет способ, которым витамины группы В транспортируются в клетку. Они водорастворимые, поэтому не хранятся в жире. Ими невозможно отравиться. В крайнем случае, будете мочиться чуть больше. Но я уверена в том, что они очень полезны».
Сохранить молодость митохондрий помогут физические упражнения
Физические упражнения тоже способствуют митохондриальному здоровью, поскольку они заставляют митохондрии трудиться. Как упоминалось ранее, одним из побочных эффектов усиленной работы митохондрий является создание активных форм кислорода, выступающих в качестве сигнальных молекул.
Одна из функций, сигнализируемых ими, является образование большего количества митохондрий. Поэтому, когда вы тренируетесь, организм реагирует, создавая больше митохондрий, чтобы удовлетворять повышенные запросы в энергии.
Старение неизбежно. Но ваш биологический возраст может сильно отличаться от хронологического, причем митохондрии имеют много общего с биологическим старением. Патрик цитирует недавнее исследование, которое показывает, как люди могут биологически стареть очень разными темпами.
Исследователи измерили более десятка различных биомаркеров, таких как длина теломера, повреждение ДНК, холестерин ЛПНП, метаболизм глюкозы и чувствительность к инсулину, в трех точках жизни людей: в возрасте 22, 32 и 38 лет.
«Мы обнаружили, что кто-то в возрасте 38 лет биологически мог выглядеть на 10 лет моложе или старше, судя по биологическим маркерам. Несмотря на одинаковый возраст, биологическое старение происходит совершенно разными темпами.
Интересно, что когда этих людей сфотографировали и показали их фотографии прохожим с просьбой угадать хронологический возраст изображенных людей, то люди угадывали биологический, а не хронологической возраст».
Таким образом, независимо от фактического возраста, на сколько лет вы выглядите, соответствует вашим биологическим биомаркерам, которые в значительной степени обусловлены здоровьем митохондрий. Поэтому, хотя старения и не избежать, вы в значительной степени можете управлять тем, как вы стареете, а это, согласитесь, дает очень много возможностей. И одним из ключевых факторов является поддержание митохондрий в хорошем рабочем состоянии.
Как считает Патрик, «молодость» - это не столько хронологический возраст, сколько то, на какой возраст вы себя чувствуете, и насколько хорошо работает ваш организм:
«Я хочу знать, как оптимизировать свою мыслительную деятельность и свои спортивные результаты. Я хочу продлить молодость. Я хочу дожить до 90. И когда я доживу, хочу заниматься серфингом в Сан-Диего точно так же, как и в свои 20. Я хотела бы угасать не так быстро, как некоторые люди. Мне нравится оттягивать это угасание и продлевать молодость столько, сколько получится, чтобы я как можно дальше радовалась жизни».
Митохондрии (МТ) – одно из самых интересных мне направлений исследований. Объединение митохондрий с другой клеткой в ходе эндосимбиоза около 1,6 млрд лет назад стало основной всех многоклеточных эукариотов со сложной структурой. Предположительно митохондрии произошли от клеток, напоминающих α-протеобактерии.
Лучшее обзорное исследование последнего времени по митохондриях – работа Вернера Кулбрандта «Структура и функция митохондриальных белковых комплексов мембраны ». Если вы знаете английский язык и интересуетесь устройством этих органелл, то настоятельно рекомендую к прочтению. Эта статья так хороша, что может быть смело главой хорошего учебника по молекулярной биологии. Сначала я хотел перевести всю статью, но это бы заняло непростительно много времени и оторвало бы от других дел. Поэтому ограничусь тезисами и картинками. Периодически разбавляя все своими мыслями.
Митохондрия кодирует сама только 13 белков, не смотря на наличие отдельной от клетки ДНК (мтДНК) и всего «производственного» цикла по транскрипции белков. Изолированная митохондрия какое-то время может сохранять композицию и функционировать.
Рисунок 1. Компоненты мембраны митохондриона. Внешняя мембрана отделяет митохондрию от цитоплазмы. Она окружает внутреннюю мембрану, которая отделяет межмембранное пространство от богатого белками центрального матрикса. Внутреннюю мембрану разделяют на внутреннюю пограничную мембрану и кристы. Две эти части непрерывны в местах крепления крист (cristae junction ). Кристы простираются более или менее глубоко в матрикс и являются основным место митохондриального преобразования энергии. Небольшой протоновый градиент в межмембранном пространстве (pH 7,2-7,4) и матрикс (pH 7,9-8,0) приводят к образованию АТФ АТФ-синтазой в мембранах крист.
Внешняя мембрана пористая и позволяет веществам из цитоплазмы проходить через нее. Внутренняя мембрана плотная, для ее пересечения нужны транспортные белки [Гилберт Линг обоснованно не согласен], непрерывность барьера позволяет иметь внутренней мембране электрохимический потенциал в -180 mV. У матрикса довольно большой pH (7,9-8). Еще раз углублюсь в Линга. Щелочной (выше 7) pH способствует более развернутой конформации белков . Высокий pH нарушает водородные и солевые связи, делая поляризованные CO и NH доступными молекулам воды, там самым усиливая дипольный момент всей внутриклеточной воды и связывая ее. В этом ключе наличие мембраны нужно не для «удержания» протоплазмы внутри клетки (это делают сами белки при высоком pH), а для наличия потенциала.
мтДНК находится в нуклеотидах, которых примерно 1000 на клетку. Белковая плотность матрикса довольна высокая (до 500 мг/мл), что близко к кристаллизованным белкам.
Внутренняя мембрана образует инвагинации, называемые кристами, которые глубоко проникают в матрикс. Кристы определяют третий «отсек» митохондрий – просвет крист (cristae lumen). Мембраны кристы содержат большинство, если не все, полностью «собранные» комплексы цепи переноса электронов и АТФ-синтазы. Просвет кристы содержит большое количество маленького растворимого белкового переносчика электронов (цитохром с). Митохондриальные кристы, таким образом, основное место биологической конверсии энергии во всех не фотосинтетических эукариотах.
С кристами тоже много всего интересного. Оптические свойства кристы влияют на распространение и генерацию света в тканях. Я даже встречал идеи о том, что поверхность крист подобна (предположение) поверхностям топологических изоляторов (подразумевалась суперпроводимость без диссипации заряда).

Рисунок 2. Мембранные белковые комплексы дыхательной цепи. Комплекс I (NADH / убихинон оксидоредуктаза, синий), Комплекс II (сукцинат дегидрогеназа, розовы), Комплекс III (цитохром С редуктаза, оранжевый), Комплекс IV (цитохром С оксидаза, зеленый) и митохондриальная АТФ синтаза (известная как комплекс V , бежевая) работают вместе во время окислительного фосфорилирования, чтобы клетки могли использовать энергию. Комплексы I , III , IV выкачивают протоны вдоль мембраны кристы, создавая протоновый градиент, стимулирующий синтез АТФ.
Теперь немного внимания на комплекс II. Вы помните, что жир (кето) делает упор в метаболизме на FADH2 и комплекс II. Они восстанавливают пару CoQ, в какой-то момент окисленного CoQ не хватает для транспортировки электронов на комплекс III и образует обратный поток электронов на комплекс I с образованием супероксида. При долгом HFLC-питании комплекс I будет обратимо разрушен, при этом это нормальная физиологическая оптимизация.
Еще прошу вас заметить, что комплекс II не выкачивает протоны. Что у нас рассеивает протоновый градиент, нарушает фосфорилирование и стимулирует сжигание жиров на тепло? Правильно, стресс холода. Термогенез связан с метаболизмом через комплекс, который не выкачивает протоны, тем самым не давая дополнительных протонов для АТФ-синтазы. Можно только удивляться как замечательно у нас продуман организм.
Крепления крист и MICOS
Места крепления крист (cristae junctions) – маленькие круглы отверстия примерно 25 нм диаметром. В митохондриях всех организмов есть система MICOS (mitochondria contact site and cristae to outer membrane), сборка из пяти мембранных и одного растворимого белков, прикрепляющих кристы к наружной мембране.
В клетках с повышенной потребностью в энергии, такие как скелетные и сердечные мышцы, кристы плотно заполняют большую часть объема митохондрии. В тканях с меньшими потребностями в энергии, таких как печень и почки, кристы находятся не так плотно по отношению друг к другу. Остается больше места в матрице для биосинтетических ферментов.
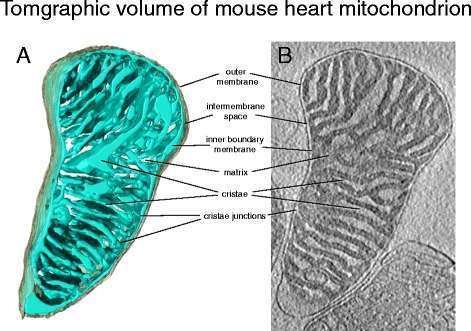
Рисунок 3. Томографический объем митохондрии сердца мыши. А) Трехмерный объем митохондрии сердца мыши, снятый cryo-ET. Наружная мембрана (серая) окутывает внутреннюю мембрану (светло-синяя). Внутренняя мембрана плотно наполнена кристами б) Томографический срез объема. Плотно заполненный матрикс, содержащий большую часть митохондриальных белков, выглядит темным на электронном микроскопе. В то время как межмембранное пространство и просветы крист выглядят светлыми из-за низкой концентрации белков.
Димеры АТФ синтазы
Митохондриальная F1-F0 АТФ синтаза является самым заметным белковым комплексом кристы. АТФ синтаза – это древняя наномашина, которая использует электрохимический протновых градиент вокруг внутренней мембраны для создания АФТ посредством вращательного катализа . Протоны, двигающиеся через F0 комплекс мембраны, вращают ротор из 8 (у млекопитающих) или 10 (у дрожжей) с-узлов. Центральный стебель передает крутящий момент c-ротора каталитической головке F1, где АТФ образуется из АДФ и фосфата через последовательность конформационных изменения. Периферийный стебель предотвращает непродуктивное вращение Головы F1 против комплекса F0.
Многие годы считалось, что АТФ синтаза случайным образом располагается на внутренней мембране. Но оказалось, что АТФ синтаза располагается двойными рядами . Причем линейные ряды АТФ синтазы – фундаментальный атрибут всех живых митохондрий.

Рисунок 4. Двойные ряды АТФ синтазы у семи разных видов.
Ряды АТФ синтазы располагаются в основном вдоль хребтов крист. Димеры изгибают липидный бислой и как следствие само-организуются в ряды. Когда у митохондрий дрожжей выбивали узлы e и g АФТ синтазы, то штамм рос на 60% медленней диких собратьев, и потенциал мембран их митохондрий был снижен вдвое. У АФТ синтазы прокариотов недостает нескольких узлов, связанных с димерами, ряды димеров не были найдены у бактерий и архей. Кристы и ряды димеров АФТ синтазы, таким образом, являются адаптацией к большим энергетическим потребностям организма.

Рисунок 5. Структура димера АТФ синтазы митохондрии polymella sp. Вид сбоку на V-образный димер АТФ синтазы.
Комплексы и суперкомплексы дыхательной цепи
Протоновый градиент вокруг внутренней мембраны создается тремя крупными мембранными комплексами, известными как комплекс I, комплекс III и комплекс IV (см. рисунок 2). Комплекс I кормится электронами из NADH, высвобождаемая при передаче электрона энергия выкачивает четыре протона. Комплекс III получает электрон от восстановленного хинола и передает его носителю электронов (цитохрому с), выкачивая в процессе один протон. Комплекс IV получает электрон из цитохрома с и передает его молекулярному кислороду, выкачивая 4 протона за каждую молекулу кислорода, превращенную в воду. Комплекс II не выкачивает протоны, напрямую передавая электроны хинолу. Как перенос электронов из NADH в хинол связан с транслокацией протонов пока не ясно. Комплекс I – крупнее III и IV вместе взятых.

Рисунок 6. Комплекс I митохондрии коровьего сердца. Матриксная часть содержит ряд из восьми железно-серных (Fe-S) кластеров, которые направляют электроны из NADH в хинол на пересечении матрикса и мембраны. Мембранная часть состоит из 78 лопастей, включая выкачивающие протоны молекулы.
Комплексы I, III и IV соединяются в суперкомплексы или респирасомы. У пекарских дрожжей (saccharomyces cerevisiae ) нет комплекса I, их суперкомплексы состоят из III и IV. Роль суперкомплексов пока еще не ясна. Предполагают, что это делает транспорт электронов более эффективным, но прямых доказательств этому пока нет.

Рисунок 7. Суперкомплекс митохондрии коровьего сердца. Обратите внимание на дистанцию между комплексами I и III, который надо проделать хинолу. Стрелки – движения электрона в суперкомплексе.
Основным белком просвета кристы является цитохром с, который переносит электрон из комплекса III в комплекс IV. Если цитохром с высвобождается в цитоплазму клетки, то вызывает апоптоз .

Рисунок 8. Ряды димеров АТФ синтазы задают форму кристам. У хребта кристы АФТ синтаза (желтый) образует слив для протонов (красный), протоновые насосы электронной цепи (зеленый) находятся по обоим сторона рядов димеров. Направляя протоны от источника к АТФ синтазе, кристы работают как протоновые направляющие, позволяющие эффективное производство АТФ. Красные стрелки показывают направление потока протонов.
Реорганизация мембраны во время старения
Старение – фундаментальный и плохо понимаемый процесс всех эукариотов. Исследовали старение митохондрий на грибах Podospora anserina , которые живут всего 18 дней. В нормальной митохондрии кристы проникают глубоко в матрикс. Для этого нужны ряды димеров АТФ синтазы и MICOS комплекс у мест крепления крист. С возрастом кристы начинают все ближе подходить к поверхности мембраны, димеры АФТ синтазы превщаются в мономеры, и все заканчивается высвобождением цитохрома с и клеточной смертью.
Транспорт электронов создает супероксид в комплексах I и III. Это побочный продукт метаболизма. Одновременно необходимый и смертельно опасный. Во время старения деление (fission) начинает превалировать над сращением (fussion). Это не дает поврежденным митохондриям «спастись» путем сращения и ускоряет неизбежное.

Рисунок 9. Изменения морфологии внутренней мембраны и димеров АТФ синтазы во время старения митохондрии.
Как видите, полей для будущих исследований очень много. Я предполагаю стык физики и биологии, где физики будут пытаются объяснить почему такая структура более энергетически эффективна. Тем более публикации по квантовой биологии
