Реферат
на тему: «Размножение»
Введение 3
1. Типы размножения 4
1.1 Бесполое размножение 4
1.2 Половое размножение 6
2. Индивидуальное развитие организмов 10
2.1 Эмбриональный период развития 10
2.2 Постэмбриональный период развития 13
2.3 Общие закономерности развития. Биогенетический закон 15
Заключение 18
Список литературы 18
Введение
Способность размножаться, т.е. производить новое поколение особей того же вида, - одна из основных особенностей живых организмов. В процессе размножения происходит передача генетического материала от родительского поколения следующему поколению, что обеспечивает воспроизведение признаков не только данного вида, но конкретных родительских особей. Для вида смысл размножения состоит в замещении тех его представителей, которые гибнут, что обеспечивает непрерывность существования вида; кроме того, при подходящих условиях размножение позволяет увеличить общую численность вида.
Каждая новая особь, прежде чем достигнуть стадии, на которой она будет способна к размножению, должна пройти ряд стадий роста и развития. Некоторые особи погибают, не достигнув репродуктивной стадии (или половозрелости) в результате уничтожения хищниками, болезней и разного рода случайных событий; поэтому вид может сохраниться лишь при условии, что каждое поколение будет производить больше потомков, чем было родительских особей, принимавших участие в размножении. Численность популяций колеблется в зависимости от баланса между размножением и вымиранием особей. Существует ряд различных стратегий размножения, каждая из которых имеет определенные преимущества и недостатки; все они будут описаны в этом реферате.
1. Типы размножения
Известны различные формы размножения, но все они могут быть объединены в два типа: половое и бесполое.
Половым размножением называют смену поколений и развитие организмов из специализированных половых клеток, образуются в половых железах. При этом новый организм развивается в результате слияния двух половых клеток, образованных разными родителями. Однако у беспозвоночных животных нередко сперматозоиды и яйцеклетки формируются в теле одного организма. Такое явление обоеполость называют гермафродитизмом. Цветковые растения также бывают обоеполыми. У большинства видов покрытосеменных (цветковых) растений обоеполый цветок включает и тычинки, образующие мужские половые клетки спермин, и пестики, содержащие яйцеклетки. Примерно у четвертой части видов мужскиe (тычиночные) и женские (пестичные) цветки развиваются независимо, т.е. у них цветки однополые. Примером может служить конопля. У некоторых растений кукурузы, березы и мужские и женские цветки возникают на одной особи.
У некоторых видов животных и растений наблюдается развитие
неоплодотворенной яйцеклетки. Такое размножение называют девственным или партеногенетическим.
Бесполое размножение характеризуется тем, что новая особь развивается из неполовых, соматических (телесных) клеток.
1.1 Бесполое размножение
При бесполом размножении новый организм может возникнуть из одной клетки или из нескольких неполовых (соматических) клеток материнской особи. В бесполом размножении участвует только одна родительская особь. Поскольку клетки, дающие начало дочерним организмам, возникают в результате митоза, то все потомки окажутся сходными по наследственным признакам с материнской особью.
Рис. 1. Размножение эвглены зеленой
Многие простейшие (амебы, эвглена зеленая и др.), одноклеточные водоросли (хламидомонада) размножаются путем митотического деления клетки (рис. 1). Другим одноклеточным некоторым низшим грибам, водорослям (хлорелла), животным, например возбудителю малярии малярийному плазмодию, свойственно спорообразование. При этом клетка распадается на большое число особей, равное количеству ядер, заранее образованных в родительской клетке в результате многократного деления ее ядра. Многоклеточные организмы также способны к спорообразованию: это мхи, высшие грибы, многоклеточные водоросли, папоротникообразные и некоторые другие.
Как у одноклеточных, так и у многоклеточных организмов способом бесполого размножения служит также почкование. Например, у дрожжевых грибов и некоторых инфузорий (сосущие инфузории) при почковании на материнской клетке первоначально образуется небольшой бугорок, содержащий ядро, почка. Она растет, достигает размеров, близких к размерам материнского организма, и затем отделяется, переходя к самостоятельному существованию. У многоклеточных (пресноводная гидра) почка состоит из группы клеток обоих слоев стенки тела. Почка растет, удлиняется, на переднем ее конце появляется ротовое отверстие, окруженное щупальцами. Почкование завершается образованием маленькой гидры, которая затем отделяется от материнского организма.
У многоклеточных животных бесполое размножение осуществляется таким же путем (медузы, кольчатые черви, плоские черви, иглокожие). Из каждой такой части развивается полноценная особь.
У растений широко распространено вегетативное размножение, т.е. частями тела черенками, усами, клубнями. Так, картофель размножается видоизмененными подземными частями стебля клубнями. У жасмина, ивы легко укореняются побеги черенки. С помощью черенков размножают виноград, смородину, крыжовник.
Длинные ползучие стебли земляники усы образуют почки, которые, укореняясь, дают начало новому растению. Немногие растения, например бегония, могут размножаться листовыми черенками (листовая пластинка и черешок). На нижней стороне листа, в местах разветвления крупных жилок, возникают корни, на верхней почки, а затем побеги.
Для вегетативного размножения используют также корень. В садоводстве с помощью черенков из боковых корней размножают малину, вишню, сливу, розу. С помощью корневых клубней размножаются георгины. Видоизменение подземной части стебля корневище также образует новые растения. Например, осот с помощью корневища может дать более тысячи новых особей на 1 м2 почвы.
1.2 Половое размножение
Половое размножение имеет очень большие эволюционные преимущества по сравнению с бесполым. Это обусловлено тем, что генотип потомков возникает путем объединения генов, принадлежащих обоим родителям. В результате повышаются возможности организмов в приспособлении к условиям окружающей среды. Так как новые комбинации осуществляются в каждом поколении, то приспособленными к новым условиям существования может оказаться гораздо большее количество особей, чем при бесполом размножении. Появление новых комбинаций генов обеспечивает более успешное и быстрое приспособление вида к меняющимся условиям обитания.
Таким образом, сущность полового размножения заключается в объединении в наследственном материале потомка генетической информации из двух разных источников родителей.
В половых железах развиваются половые клетки: мужские сперматозоиды, женские яйцеклетки (или яйца). В первом случае их развитие называют сперматогенезом, во втором овогенезом (от лат. ово яйцо).
В процессе образования половых клеток выделяют ряд стадий. Первая стадия период размножения, в котором первичные половые клетки делятся путем митоза, в результате чего увеличивается их количество.
Вторая стадия период роста. У незрелых мужских гамет он выражен не резко. Их размеры увеличиваются незначительно. Напротив, будущие яйцеклетки овоциты увеличиваются в размерах иногда в сотни, а чаще в тысячи и даже миллионы раз. Рост овоцитов осуществляется за счет веществ, образуемых другими клетками организма. Так, у рыб, амфибий и в большей степени у рептилий и птиц основную массу яйца составляет желток. Он синтезируется в печени, в особой растворимой форме переносится кровью в яичник, проникает в растущие овоциты и откладывается там в виде желточных пластинок. Кроме того, в самой будущей половой клетке синтезируются многочисленные белки и большое количество разнообразных РНК: транспортных, рибосомных и информационных. Желток совокупность питательных веществ (жиров, белков, углеводов, витаминов и др.), необходимых для питания развивающегося зародыша, а РНК обеспечивает синтез белков на ранней стадии развития, когда собственная бедственная информация еще не используется.
Следующая стадия период созревания, или мейоз, представлена на рисунке 2. Клетки, вступающие в период созревания, содержит диплоидный набор хромосом и уже удвоенное количество ДНК.

Рис. 2. Созревание половых клеток (мейоз)
Сущность мейоза состоит в том, что каждая половая клетка получает одинарный, гаплоидный, набор хромосом. Однако вместе с тем мейоз это стадия, во время которой создаются новые комбинации генов путем сочетания разных материнских и отцовских хромосом, рекомбинирование наследственных задатков возникает, кроме того в результате кроссинговера обмена участками между гомологичными хромосомами в процессе мейоза.
Мейоз включает два последовательных деления. Как и в митозе, в каждом мейотическом делении выделяют четыре стадии: профазу, метафазу, анафазу и телофазу.
Первое (I) мейотическое деление. Профаза I начинается спирализацией хромосом. Как вы помните, каждая хромосома состоит из двух хроматид, соединенных в области центромеры. Затем гомологические хромосомы сближаются, каждая точка каждой хроматиды одной хромосомы совмещается с соответствующей точкой хроматиды другой, гомологичной хромосомы. Этот процесс точного и тесного сближения гомологичных хромосом в мейозе называют конъюгацией. В дальнейшем между такими хромосомами может произойти кроссинговер обмен одинаковыми, или гомологичными, т. е. содержащими одни и те же гены, участками. К концу профазы между гомологичными хромосомами возникают силы отталкивания. Вначале они проявляются в области центромер, а затем в других участках.
В метафазе I спирализация хромосом максимальна. Конъюгированные хромосомы располагаются по экватору, причем центромеры гомологичных хромосом обращены к разным полюсам клетки. К ним прикрепляются нити веретена деления.
В анафазе I плечи гомологичных хромосом окончательно разделяются, и хромосомы расходятся к различным полюсам. Следовательно, из каждой пары гомологичных хромосом в дочернюю клетку попадает только одна. Число хромосом уменьшается в два раза, хромосомный набор становится гаплоидным. Однако каждая хромосома состоит из двух хроматид, т. е. по-прежнему содержит удвоенное количество ДНК.
В телофазе I на непродолжительное время образуется ядерная оболочка. Во время интерфазы между первым и вторым делениями мейоза редупликации ДНК не происходит. Клетки, образовавшиеся в результате первого деления созревания, различаются по составу отцовских и материнских хромосом и, следовательно, по набору генов.
Например, все клетки человека, в том числе первичные половые клетки, содержат 46 хромосом. Из них 23 получены от отца и 23 от матери. При образовании половых клеток после первого мейотического деления в сперматоциты и овоциты также попадает по 23 хромосомы. Однако вследствие случайности расхождения отцовских и материнских хромосом в анафазе I образующиеся клетки получают самые разнообразные комбинации родительских хромосом. Например, в одной из них может оказаться 3 отцовских и 20 материнских хромосом, в другой 10 отцовских и 13 материнских, в третьей 20 отцовских и 3 материнских и т. д. Число возможных комбинаций очень велико. Если учесть еще обмен гомологичными участками хромосом в профазе первого деления мейоза, то вполне очевидно, что каждая образующаяся половая клетка генетически уникальна, так как несет свой неповторимый набор генов.
Следовательно, мейоз основа комбинативной генотипической изменчивости.
Второе (II) мейотическое деление. Второе деление мейоза в общем протекает так же, как обычное митотическое деление, с той лишь разницей, что делящаяся клетка гаплоидна. В анафазе II центромеры, соединяющие сестринские хроматиды в каждой хромосоме, делятся, и хроматиды, как и в митозе, с этого момента становятся самостоятельными хромосомами. С завершением телофазы II заканчивается и весь процесс мейоза: из исходной первичной половой клетки образовались четыре гаплоидные клетки.
У особей мужского пола все они преобразуются в гаметы сперматозоиды. У особей женского пола вследствие неравномерного мейоза лишь из одной клетки получается жизнеспособное яйцо. Три другие дочерние клетки гораздо мельче, они превращаются в так называемые направительные, или редукционные, тельца, вскоре погибающие. Образование только одной яйцеклетки и гибель трех генетически полноценных направительных телец с биологической точки зрения обусловлено необходимостью сохранения в одной клетке всех запасных питательных веществ, которые понадобятся для развития будущего зародыша.
Период формирования состоит в приобретении клетками определенной формы и размеров, соответствующих их функции.
Женские половые клетки в процессе созревания покрываются оболочками и готовы к оплодотворению непосредственно после завершения мейоза. Во многих случаях, например у пресмыкающихся, птиц и млекопитающих, за счет деятельности клеток, окружающих яйцеклетку, вокруг нее возникает ряд дополнительных оболочек. Их функция заключается в защите яйцеклетки и развивающегося зародыша от внешних неблагоприятных воздействий. Сперматозоиды могут иметь различные размеры и форму.
Функция сперматозоидов состоит в доставке в яйцеклетку генетической информации и стимуляции ее развития. Сформированный сперматозоид содержит митохондрии, аппарат Гольджи, выделяющий ферменты, растворяющие мембрану яйца при оплодотворении, т. е. при слиянии сперматозоида и яйцеклетки. Возникающая при этом диплоидная клетка носит название зиготы.
2. Индивидуальное развитие организмов
Индивидуальным развитием, или онтогенезом, называют весь период жизни особи с момента слияния сперматозоида с яйцом и образования зиготы до гибели организма. Онтогенез делится на два периода: 1) эмбриональный от образования зиготы до рождения или выхода из яйцевых оболочек; 2) постэмбриональный от выхода из яйцевых оболочек или рождения до смерти организма.
Наука, изучающая закономерности индивидуального развития организмов на стадии зародыша, называется эмбриологией (от греч. эмбрион зародыш).
2.1 Эмбриональный период развития
У большинства многоклеточных животных, независимо от сложности их организации, стадии эмбрионального развития, которые проходит зародыш, едины. В эмбриональном периоде выделяют три основных этапа: дробление, гаструляцию и первичный органогенез.
Дробление. Развитие организма начинается со стадии одной клетки. Оплодотворенное яйцо это клетка и одновременно уже организм на самой ранней стадии его развития. В результате многократных делений одноклеточный организм превращается в многоклеточный. Возникшее при оплодотворении путем слияния сперматозоида и яйцеклетки диплоидное ядро через несколько минут начинает делиться, вместе с ним делится и цитоплазма. Образующиеся клетки с каждым делением уменьшаются в размерах, поэтому процесс деления носит название дробления. В период дробления накапливается клеточный материал для дальнейшего развития. Завершается дробление образованием многоклеточного зародыша бластулы. Бластула имеет полость, наполненную жидкостью, так называемую первичную полость тела.
В тех случаях, когда в цитоплазме яйцеклетки желтка мало (как у ланцетника) или относительно немного (как у лягушки), дробление бывает полным, т. е. яйцеклетка делится целиком.
Иначе протекает период дробления у птиц. Свободная от желтка цитоплазма составляет всего 1% от общего объема яйцеклетки курицы; вся остальная цитоплазма яйцеклетки, а следовательно и зигота, заполнена массивом желтка. Если присмотреться к куриному яйцу, на одном из его полюсов непосредственно на желтке можно увидеть маленькое пятнышко бластулу, или зародышевый диск, образовавшийся в результате дробления свободного от желтка участка цитоплазмы, содержащего ядро. В таких случаях дробление называют неполным. Неполное дробление свойственно и некоторым рыбам и рептилиям.
Во всех случаях и у ланцетника, и у амфибий, и у птиц, а также у других животных общий объем клеток на стадии бластулы не превышает объема зиготы. Другими словами, митотическое деление зиготы не сопровождается ростом образовавшихся дочерних клеток до объема материнской, и размеры их в результате ряда последовательных делений прогрессивно уменьшаются. Эта особенность митотического деления клеток в ходе дробления наблюдается при развитии оплодотворенных яиц у всех животных.
Некоторые другие черты дробления также свойственны различным видам животных. Например, все клетки в бластуле имеют диплоидный набор хромосом, одинаковы по строению и отличаются друг от друга главным образом количеством содержащегося в них желтка. Такие клетки, лишенные признаков специализации для выполнения определенных функций, называют неспециализированными (или недифференцированными) клетками. Другая особенность дробления чрезвычайно короткий митотический цикл бластомеров по сравнению с клетками взрослого организма. Во время очень короткой интерфазы происходит только удвоение ДНК.
Гаструляция. Бластула, как правило, состоящая из большого числа бластомеров (например, у ланцетника из 3000 клеток), в процессе развития переходит в новую стадию, которую называют гаструлой (от греч. гастер желудок). Зародыш на этой стадии состоит из отчетливо различимых пластов клеток так называемых зародышевых листков: наружного, или эктодермы (от греч. эктос находящийся снаружи), и внутреннего, или энтодермы (от греч. энтос находящийся внутри). Совокупность процессов, приводящих к образованию гаструлы, называют гаструляцией.
У ланцетника гаструляция осуществляется путем впячивания одного из полюсов бластулы внутрь, по направлению к другому, у других животных либо путем расслоения стенки бластулы, либо путем обрастания массивного вегетативного полюса мелкими клетками анимального полюса.
У многоклеточных животных, кроме кишечнополостных, параллельно с гаструляцией или, как у ланцетника, вслед за ней возникает и третий зародышевый листок мезодерма (от греч. мезос находящийся посередине), которая представляет собой совокупность клеточных элементов, расположенных между экто- и энтодермой в первичной полости тела бластоцеле. С появлением мезодермы зародыш становится трехслойным.
Таким образом, сущность процесса гаструляции заключается перемещении клеточных масс. Клетки зародыша практически делятся и не растут. Однако на этой стадии начинается использование генетической информации клеток зародыша, появляются первые признаки дифференцировки.
Дифференцировка, или дифференцирование, это процесс ее возникновения и нарастания структурных и функциональных различий между отдельными клетками и частями зародыша. С морфологической точки зрения дифференцирование выражается в том, что образуются несколько сотен типов клеток специфического строения, отличающихся друг от друга. Из неспециализированных клеток бластулы постепенно возникают клетки эпителия кожи, эпителия кишечника, легких, появляются нервные, мышечные клетки и т.д. С биохимической точки зрения специализация клеток заключается в способности синтезировать определенные белки, свойственные только данному типу клеток. Лимфоциты синтезируют защитные белки антитела, мышечные клетки сократительный белок миозин. Каждый тип клеток образует «свои», свойственные только ему белки. Биохимическая специализация клеток обеспечивается избирательной дифференциальной активностью генов, т. е. в клетках разных зародышевых листков зачатков определенных органов и систем начинают функционировать разные группы генов.
У разных видов животных одни и те же зародышевые листки дают начало одним и тем же органам и тканям. Это значит, что они гомологичны. Так, из клеток наружного зародышевого листка эктодермы у членистоногих, хордовых, в том числе у рыб, амфибий, рептилий, птиц и млекопитающих, формируются кожные покровы и их производные, а также нервная система и органы чувств. Гомология зародышевых листков подавляющего большинства животных одно из доказательств единства животного мира.
Органогенез. После завершения гаструляции у зародыша образуется комплекс осевых органов: нервная трубка, хорда, кишечная трубка. У ланцетника осевые органы формируются следующим образом: эктодерма на спинной стороне зародыша прогибается по средней линии, превращаясь в желобок, а эктодерма, расположенная справа и слева от него, начинает нарастать на его края. Желобок зачаток нервной системы погружается под эктодерму, и края его смыкаются. Образуется нервная трубка. Вся остальная эктодерма зачаток кожного эпителия.
Спинная часть энтодермы, располагающаяся непосредственно под нервным зачатком, обособляется от остальной энтодермы и сворачивается в плотный тяж хорду. Из оставшейся части энтодермы развиваются мезодерма и эпителий кишечника. Дальнейшая дифференцировка клеток зародыша приводит к возникновению многочисленных производных зародышевых листков органов и тканей. В процессе специализации клеток, входящих в состав зародышевых листков, из эктодермы образуются нервная система, органы чувств, эпителий кожи, эмаль зубов; из энтодермы эпителий кишки, пищеварительные железы печень и поджелудочная железа, эпителий жабр и легких; из мезодермы мышечная ткань, соединительная ткань, в том числе рыхлая соединительная ткань, хрящевая и костная ткани, кровь и лимфа, а также кровеносная система, почки, половые железы.
2.2 Постэмбриональный период развития
В момент рождения или выхода организма из яйцевых оболочек заканчивается эмбриональный и начинается постэмбриональный период развития. Постэмбриональное развитие может быть прямым или сопровождается превращением (метаморфозом).
При прямом развитии (у пресмыкающихся, птиц, млекопитающих) из яйцевых оболочек или из тела матери выходит организм небольших размеров, но с уже заложенными всеми основными органами, свойственными взрослому животному. Постэмбриональное развитие в этом случае сводится в основном к росту и половому созреванию.
При развитии с метаморфозом из яйца выходит личинка, обычно устроенная проще взрослого животного, со специальными личиночными органами, во взрослом состоянии отсутствующими. Личинка питается, растет, и со временем личиночные органы заменяются органами, свойственными взрослым особям. Следовательно, при метаморфозе разрушаются личиночные органы и возникают органы, присущие взрослым животным.
Разберем несколько примеров непрямого постэмбрионального развития. Личинка асцидий (тип Хордовые, подтип Личиночно-хордовые) обладает всеми основными признаками хордовых животных: хордой, нервной трубкой, жаберными щелями в глотке. Она свободно плавает, затем прикрепляется к какой-либо твердой поверхности на дне моря, где и совершается метаморфоз: у нее исчезают хвост, хорда, мышцы, а нервная трубка распадается на отдельные клетки, большая часть которых фагоцитируется. От нервной системы личинки остается лишь группа клеток, дающая начало нервному узлу. Строение тела взрослой асцидий, ведущей прикрепленный образ жизни, нисколько не напоминает обычные черты организации хордовых животных. Только знание особенностей онтогенеза позволяет определить систематическое положение асцидий: строение личинки указывает на происхождение их от хордовых животных, которые вели свободный образ жизни. В процессе метаморфоза асцидий переходят к сидячему образу жизни, в связи с чем упрощается их организация.
Личиночная форма амфибий головастик, для которого характерны жаберные щели, боковая линия, двухкамерное сердце, один круг кровообращения. В процессе метаморфоза, происходящего под влиянием гормона щитовидной железы, рассасывается хвост, появляются конечности, исчезает боковая линия, развиваются легкие и второй круг кровообращения. Обращает на себя внимание сходство ряда черт строения головастиков и рыб (боковая линия, строение сердца и кровеносной системы, жаберные щели).
Примером метаморфоза может служить также развитие насекомых. Гусеницы бабочек или личинки стрекоз резко отличаются по строению, образу жизни и среде обитания от взрослых животных и напоминают своих предков кольчатых червей.
Постэмбриональный период развития имеет разную продолжительность. Например, поденки в личиночном состоянии живут 2-3 года, а в половозрелом от 2-3 часов до 2-3 суток в зависимости от видовой принадлежности. В большинстве же случаев постэмбриональный период более продолжителен. У человека он включает стадию полового созревания, стадию зрелости и стадию старости.
У млекопитающих и человека наблюдается известная зависимость продолжительности жизни от длительности полового созревания и беременности. Обычно продолжительность жизни превышает
дорепродуктивный период онтогенеза в 5-8 раз.
Постэмбриональное развитие сопровождается ростом. Различают рост неопределенный, продолжающийся в течение всей жизни, и определенный, ограниченный каким-то сроком. Неопределенный рост наблюдается у древесных форм растений, некоторых моллюсков, из позвоночных у рыб, крыс.
У многих животных рост прекращается вскоре после достижения половой зрелости. У человека рост заканчивается к 20-25 годам.
2.3 Общие закономерности развития. Биогенетический закон
Все многоклеточные организмы развиваются из оплодотворенного яйца. Развитие зародышей у животных, относящихся к одному типу, во многом сходно. У всех хордовых животных в эмбриональном периоде закладывается осевой скелет хорда, возникает нервная трубка, в переднем отделе глотки образуются жаберные щели. План строения хордовых животных также одинаков. На ранних стадиях развития зародыши позвоночных очень похожи (рис. 3). Эти факты подтверждают справедливость сформулированного К. Бэром закона зародышевого сходства: «Эмбрионы обнаруживают, уже начиная с самых ранних стадий, известное общее сходство в пределах типа». Сходство зародышей разных систематических групп свидетельствует об общности их происхождения. В дальнейшем в строении зародышей проявляются признаки класса, рода, вида и, наконец, признаки, характерные для данной особи. Расхождение признаков зародышей в процессе развития называется эмбриональной дивергенцией и отражает эволюцию той или иной систематической группы животных, историю развития данного вида.

Рис. 3. Зародышевое сходство у позвоночных: 1 однопроходные (ехидна), 2 сумчатые (кенгуру), 3 парнокопытные (олень), 4 хищные (кошка), 5 приматы (мартышка), 6 - человек
Большое сходство зародышей на ранних стадиях развития и по
явление различий на более поздних стадиях имеют свое объяснение.
Организм подвержен изменчивости на протяжении всего развития.
Мутационный процесс затрагивает гены, обусловливающие особенности строения и обмена веществ у самых молодых эмбрионов. Но возникающие у них структуры (древние признаки, свойственные далеким предкам) играют весьма важную роль в процессах дальнейшего развития. Как указывалось, зачаток хорды индуцирует образование нервной трубки, а его утрата приводит к прекращению развития. Поэтому изменения на ранних стадиях обычно приводят к недоразвитию и гибели особи. Напротив, изменения на поздних стадиях, затрагивая менее значительные признаки, могут быть благоприятны для организма и в таких случаях подхватываются естественным отбором.
Появление в эмбриональном периоде развития современных животных признаков, свойственных их далеким предкам, отражает эволюционные преобразования в строении органов.
В своем развитии организм проходит одноклеточную стадию (стадия зиготы), что может рассматриваться как повторение филогенетической стадии первобытной амебы. У всех позвоночных, включая высших их представителей, закладывается хорда, которая далее замещается позвоночником, а у их предков, если судить по ланцетнику, хорда оставалась на всю жизнь. В ходе эмбрионального развития птиц и млекопитающих, включая человека, появляются жаберные щели в глотке и соответствующие им перегородки. Факт закладки частей жаберного аппарата у зародышей наземных позвоночных объясняется их происхождением от рыбообразных предков, дышащих жабрами. Строение сердца человеческого зародыша в ранний период формирования напоминает строение этого органа у рыб: оно с одним предсердием и одним желудочком. У беззубых китов в эмбриональном периоде появляются зубы. Зубы эти не прорезываются, они разрушаются и рассасываются.
Приведенные здесь и многие другие примеры указывают на глубокую связь между индивидуальным развитием организмов и их историческим развитием. Эта связь нашла свое выражение в биогенетическом законе, сформулированном Ф. Мюллером и Э. Геккелем в XIX в.: онтогенез (индивидуальное развитие) каждой особи есть краткое и быстрое повторение филогенеза (исторического развития) вида, к которому эта особь относится.
Заключение
Завершая работу над рефератом можно прийти к выводу, что способность к размножению, или самовоспроизведению, одна из важнейших характеристик органической природы. Размножение свойство, присущее всем без исключения живым организмам, от бактерий до млекопитающих.
Существование любого вида животных и растений, бактерий и грибов, преемственность между родительскими особями и их потомством поддерживаются только благодаря размножению. Тесно связано с самовоспроизведением и другое свойство живых организмов развитие. Оно также присуще всему живому на Земле: и мельчайшим одноклеточным организмам, и многоклеточным растениям и животным.
Список литературы
Боген Г. Современная биология. - М.: Мир, 1970.
Грин Н., Стаут У., Тейлор Д. Биология: в 3-х т. Т. 3: пер. с англ./под ред. Р. Сопера. - М.: Мир, 1990.
Мамонтов С.Г. Биология. Общие закономерности. М.: Дрофа, 2002.
От животных к человеку. М.: Наука, 1971.
Слюсарев А.А. Биология с общей генетикой. - М.: Медицина, 1978.
Лекция №3 Онтогенез
1. Гаметогенез
2. Эмбриональный период
3. Постэмбриональный период
Онтогенез – индивидуальное развитие организма от зиготы до биологической смерти. Прогенез – период до онтогенеза, включает гаметогенез и оплодотворение. Филогенез – эволюционное развитие вида.
Для человека характерно половое размножение, оно характеризуется: наличием половых желез, половых клеток, процессом оплодотворения, половым диморфизмом.
Гаметогенез – процесс образования половых клеток, овогенез – созревание яйцеклеток, сперматогенез – сперматозоидов. Гаметогенез происходит в половых железах организма. Гаметы образуются из клеток зачаточного эпителия, которые закладываются в эмбриональный период развития организма.
В процессе образования половые клетки – проходят три стадии:
1. Период размножения (клетки зачаточного эпителия делятся митозом);
2. Период роста;
3. Период созревания, клетки делятся мейозом, в результате, чего образуются гаметы (рис. 5).
Рис. 5. Деление половых клеток
Сперматозоиды созревают в семенных канальцев семенников. Между семенными канальцами расположена интерстициальная ткань , продуцирующая мужской половой гормон – тестостерон . Мужские половые гормоны – андрогены , регулируют детородную функцию, гаметогенез, формирование вторичных половых признаков. Сперматозоиды созревают 70 дней. В семенных канальцах находятся гаметы на разных стадиях созревания. В 5мл. семенной жидкости у человека содержится 12 млн. сперматозоидов. Они заряжены отрицательно, заряд препятствует их слипанию. Большое количество мужских гамет биологически целесообразно, уменьшение их численности на 40%, делает процесс оплодотворения не возможным. Сперматозоиды это мелкие подвижные клетки, состоят из головки, шейки, хвоста. На головке находится акросома, видоизмененная лизосома, в ней содержатся ферменты, растворяющие оболочку яйцеклетки при оплодотворении. Продолжительность жизни от 6 до 72 часов.
Овогенез начинается в эмбриональный период развития женского организма. На стадии профазы первого деления мейоза, когда происходит конъюгация и кроссинговер, наступает временная пауза. Дальнейшее развитие яйцеклетки возобновляется в период полового созревания девочки. Под действием женских половых гормонов – эстрогенов, вокруг ооцита первого порядка образуется фолликул, клеточный пузырек, защищающий и питающий клетку.
По мере созревания фолликул продвигается к краю яичника и затем лопается, эту стадию развития яйцеклетки называю яичниковой или фолликулярной, она продолжается 12 дней. Разрыв фолликула выход яйцеклетки из яичника в брюшную полость называют – овуляция.
После овуляции возможно оплодотворение. Женская гамета захватывается бахромками маточной трубы. С помощью реснитчатого эпителия она продвигается в матку, маточная стадии продолжается 12-14 дней. К этому времени под действием женских половых гормонов, эпителий матки разрыхляется. Если оплодотворение не произошло, эпителий и яйцеклетки выводятся из организма, в результате сокращения гладкой мускулатуры стенок матки. Этот процесс называется мензис, он длится 3-4 дня. Таким образом, время от начала образования фолликула и до наступления мензиса составляет 20-30 дней и называется овариально-менструальный цикл (рис. 6).

Рис. 6. Овуляция и имплантация яйцеклетки в матку
Он включает яичниковую стадию, маточную и мензис. В течение овариально-менструального цикла перестраивается вся репродуктивная система женщины, меняется качественный и количественный состав гормонов, изменяется работоспособность, сила гибкость, выносливость. Физиологические изменения женского организма учитываются при организации тренировочного процесса.
Оплодотворение – это слияние яйцеклетки и сперматозоида. Различают три этапа оплодотворения:
1) акросомальная реакция (20 секунд) – растворение оболочки яйцеклетки ферментами акросомы и проникновение сперматозоида в яйцеклетку. Считают, что в яйцеклетку проникает тот сперматозоид, который создает на мембране яйцеклетки наибольший потенциал проникновения (возбуждение);
2) временная пауза – в яйцеклетке не происходит видимых изменений, усиливается обмен веществ;
3) слияние ядер яйцеклетки и сперматозоида, восстановление диплоидного набора хромосом.
Оплодотворение происходит в верхней трети маточной трубы. Оплодотворенная яйцеклетка называется зиготой . С образованием зиготы начинается онтогенез человека.
2. Онтогенез человека включает два этапа: эмбриональный и постэмбриональный.
Эмбриональный этап развития продолжается от зиготы до рождения. Включает стадии: зиготы, бластулы, гаструлы, гистогенеза, органогенеза.
Зигота – многократно делится митозом, в результате чего образуется многоклеточный однослойный зародыш – бластула . Бластула не увеличивается в размерах, так как в это время она находится в маточной трубе и должна свободно продвигаться в матку. На шестой день она попадает в матку и прикрепляется к ее стенке, погружаясь в разрыхленный эпителий. Это процесс называется имплантация . Зародыш продолжает развиваться и становится двух, затем трехслойным. На этом этапе развития его называют гаструлой . В результате образуется три зародышевых листка: эктодерма, эндодерма, мезодерма. Из зародышевых систем образуются ткани и органы.
Таким образом: бластула – однослойный зародыш, гаструла – двух и трехслойный зародыш. Гистогенез – закладка тканей из зародышевых листков. Органогенез – закладка органов. В возрасте 8 недель масса зародыша человека составляет 4 г, размер 5мм. К этому времени у него произошли формообразовательные процессы, он приобрел контуры человеческого тела. После восьми недель беременности зародыш человек называют плод.
В процессе развития эмбриона человека формируются провизорные органы, которые после рождения утрачивают свои функции. К провизорным органам относятся: зародышевые оболочки, хорион, амнион и плацента, пупочный канатик . Плацента обеспечивает тесный контакт капилляров ребенка и матери. Развивающийся организм получает через плаценту питательные вещества, кислород, антитела, выводит продукты обмена. Плацента выполняет барьерную функцию, защищает эмбрион от микроорганизмов, некоторых лекарств и токсинов (рис. 7).

Рис. 7. Плацента
1 – матка; 2 – амнион; 3 – эмбрион; 4 – околоплодные воды; 5 – плацента; 6 – капилляры плаценты; 7 – вены; 8 – артерии
Тонкие механизмы формирования органов и тканей ребенка могут быть нарушены при воздействии, ядовитых, наркотических веществ, алкоголя, никотина, содержащихся в крови матери. Нарушения эмбрионального развития человека называются врожденными уродствами и пороками развития. К ним относятся: «волчья пасть» (отсутствие верхнего неба), «заячья губа» (несрастание верхнечелюстных костей), полидактилия (лишние пальцы), синдактилия (сросшиеся пальцы). Неблагоприятные факторы среды, вызывающие врожденные пороки развития называются тератогенными (вызывающие уродства) (рис. 8).

Рис. 8. Факторы среды влияющие на развитие организма человека
Эмбриональный период развития человека длится 36 недель.
3. Период после рождения и до биологической смерти называется постэмбриональным. Он включает ювенильный, пубертатный периоды развития и старение.
Ювенильный – период до полового созревания. Пубертатный – период половой зрелости.
В спортивной практике, физиологии, педагогике, психологии, используется следующая возрастная периодизация постэмбрионального онтогенеза человека.
В период от 1 до 10 дней ребенок называется новорожденный ;
С 10 дней до 1 года – грудной;
с 1 года до трех – период раннего детства ;
с 4 до 7 лет – первое детство ;
с 8 лет до 12 у девочек, и до 13 у мальчиков второе детство ;
с 12 лет до 15 у девочек и с 13 до 16 у мальчиков – подростки ;
17-21 (юноши), 16-20 лет (девушки) – юношеский возраст ;
20-55 у женщин и 21-60 у мужчин период половой зрелости (пубертат);
от 55 и 60 до 70 – пожилые люди ;
от 70 до 90 старики ;
более 90 долгожители .
В онтогенезе человека существуют сенситивные и критические периоды.
Критические периоды характеризуются повышенной активностью отдельных генов и их комплексов, контролирующих развитие, каких либо признаков организма. В эти периоды происходит значительная перестройка регуляторных процессов, качественный и количественный скачок в развитии отдельных органов и функциональных систем. В эти периоды организм наиболее чувствителен к воздействию факторов окружающей среды. Критическими периодами онтогенеза считают имплантацию, плацентацию, закладку осевых органов, нервной и кишечной трубки, хорды, формирование сердца и другие.
Если женщина заболеет краснухой между 3-й и 9-й неделями беременности, то возникает риск развития у плода таких пороков, как порок сердца, катаракта, глухота. В другие сроки краснуха не вызывает пороков развития плода.
Сенситивные периоды – это периоды снижения генетического контроля и повышенной чувствительности отдельных признаков организма к средовым влияниям, в том числе педагогическим и тренерским. Так сенситивный период проявления различных показателей качества быстроты приходится на возраст 11-14 лет и к 15-летнему возрасту достигается его максимальный уровень, когда возможны высокие спортивные достижения. Сходная картина наблюдается в онтогенезе и для проявления качества ловкости и гибкости.
Для тренеров и педагогов, работающих в области физического воспитания и спорта, знание сенситивных периодов имеет важное практическое значение, так как в сенситивные периоды обеспечивается наибольший тренировочный эффект .
Их можно разделить в зависимости от того сколько клеток участвуют в процессе бесполого размножения: бесполое размножение при котором дочернее поколение возникает из одной клетки: деление клетки множественное деление клетки шизогония спорообразование споруляция почкование у одноклеточных дрожжи...
Поделитесь работой в социальных сетях
Если эта работа Вам не подошла внизу страницы есть список похожих работ. Так же Вы можете воспользоваться кнопкой поиск
Размножение и развитие организмов
План
- Понятие и значение размножения.
- Формы и виды размножения.
- Клеточный цикл. Митоз. Мейоз.
- Строение гамет. Гаметогенез.
1. Понятие и значение размножения
Одним из свойств живого является дискретность, т.е. на любом уровне организации живая материя представлена элементарными структурными единицами. Каждая особь данного вида смертна, и существование вида поддерживается размножением организмов. Таким образом, дискретность жизни предполагает ее воспроизводство, т.е. процесс размножения.
Размножение это способность живых существ воспроизводить себе подобных. При этом обеспечивается непрерывность и преемственность жизни. Преемственность определяется тем, что в процессе размножения генетический материал передается от родителей к потомству, в результате родительские признаки в той или иной степени проявляются и у дочерних организмов.
Значение размножения:
- Увеличение или поддержание численности вида. За счет размножения происходит не только репродукция (т.е. воспроизведение себе подобных), но и увеличение численности живых организмов.
- Непрерывность жизни. Благодаря размножению более или менее крупные группы особей (например, популяции и виды) могут существовать на протяжении довольно длительного времени, так как снижение их численности вследствие естественной гибели особей компенсируется за счет постоянного воспроизведения организмов и замещения умерших вновь родившимися.
- Преемственность между поколениями. В процессе размножения передается генетическая информация от особей родительского поколения, обеспечивая воспроизведение признаков конкретных родителей и всего вида к которому принадлежит организм.
- Реализуются такие свойства живого как наследственность и изменчивость.
2. Формы и виды размножения
Известны две основные формы размножения: половое и бесполое.
Бесполое размножение
При бесполом размножении новая особь появляется из неспециализированных неполовых клеток тела соматических. Поэтому при бесполом размножении участвует одна особь. Бесполое размножение гарантирует воспроизведение большого количества идентичных особей, что выгодно для видов, обитающих в постоянных условиях. Быстрое и многочисленное воспроизведение потомства, идентичного материнскому является биологическим смыслом бесполого размножения.
В природе существуют разные варианты бесполого размножения: деление, споруляция, фрагментация, почкование, вегетативное размножение, клонирование.
Их можно разделить в зависимости от того, сколько клеток участвуют в процессе бесполого размножения:
- бесполое размножение, при котором дочернее поколение возникает из одной клетки:
- деление клетки
- множественное деление клетки (шизогония)
- спорообразование (споруляция)
- почкование у одноклеточных (дрожжи)
2) бесполое размножение, в основе которого лежит деление группы клеток:
- вегетативное
- фрагментация
- почкование у многоклеточных (гидра)
Существует ещё одна классификация, согласно которой вариантами бесполого размножения являются:
- вегетативное размножение (т.е. деление частями материнского организма):
Деление
Множественное деление (шизогония)
Почкование
Фрагментация
Размножение растений черенками, луковицами, листьями, корневищами
- размножение спорами, т.е. спорообразование
Рассмотрим типы бесполого размножения подробнее.
Деление . Наиболее простая форма бесполого размножения. Различают различные варианты деления:
- Деление путем простой перетяжки с образованием при этом из одного родительского организма двух дочерних. Характерно для бактерий и цианобактерий.
- Деление путем митотического деления ядра с последующим разделением цитоплазмы. Свойствено одноклеточным организмам (многим простейшим амеба, эвглена зеленая и др.; одноклеточным водорослям хламидомонада и др.).
И в том и в другом случае происходит бинарное деление, т.е. на две клетки. Однако возможен и другой вариант:
- Множественное деление (шизогония). Сначала многократно делится ядро и затем происходит деление цитоплазмы на части. Например, размножение малярийного плазмодия (возбудителя малярии) в эритроцитах человека. В этом случае у плазмодиев происходит много раз повторяющееся деление ядра, после чего делится цитоплазма. В результате 1 плазмодий дает начало 12-24 дочерним организмам.
Образование спор (споруляция) . Очень широко распространенный способ бесполого размножения среди живых организмов и встречается практически у всех растений, грибов и некоторых простейших (например, тип споровики), а также прокариотических организмов (у многих бактерий, синезеленых водорослей).
Спора это клетка, покрытая развитым защитным покровом споровой оболочкой, позволяющей переносить действие различных неблагоприятных факторов внешней среды. Вследствие малых размеров спора обычно содержит лишь минимальные запасы питательных веществ . У многих организмов она является основной единицей расселения, так как большие количества образующихся легких спор свободно переносятся на значительные расстояния движением воздушных масс и потоками воды. Нередко споры образуются в больших количествах и имеют ничтожный вес, что облегчает их распространение ветром, а также животными, главным образом насекомыми.
Кроме того, у некоторых форм (простейшие, грибы) споры могут выступать в качестве особой стадии жизненного цикла, позволяющей «переживать» действие неблагоприятных факторов среды. Споры бактерий служат, строго говоря, не для размножения, а для того, чтобы выжить при неблагоприятных условиях, поскольку каждая бактерия образует только одну спору. Бактериальные споры относятся к числу наиболее устойчивых: так, например, они нередко выдерживают обработку сильными дезинфицирующими веществами и кипячение в воде.
У многих растений процесс образования спор (спорогенез) осуществляется в особых мешковидных структурах спорангиях. Споры могут быть как подвижными, благодаря наличию жгутикового аппарата (в этом случае они называются зооспорами), так и неподвижными, лишенными способности активно передвигаться.
Зооспоры наблюдаются у некоторых зеленых водорослей. Среди животных спорообразование наблюдается у малярийного плазмодия и целой группы споровиков (одноклеточные).
Почкование . Встречается как у одноклеточных, например, у дрожжей и некоторых видов инфузорий, так и у многоклеточных, например, у представителей типа кишечнополостные (гидра), а также для оболочников (класс асцидии).
Для одноклеточных этот способ заключается в том, что на материнской клетке образуется бугорок (вырост) с ядром, который затем отделяется и становится самостоятельным организмом.
Для многоклеточных этот способ заключается в том, что вначале на теле материнской особи возникает небольшой бугорок, увеличивающийся в размерах, затем появляются зачатки всех структур и органов, характерных для материнского организма. Потом происходит отделение (отпочковывание) дочерней особи. После этого молодой, вновь отделившийся организм растет и достигает размеров исходного экземпляра.
Рис. Почкование у кишечнополостных (гидра), 1 взрослый организм, 2 дочерний почкующийся организм.
Необычная форма почкования описана у суккулентного растения Bryophyllum - ксерофита, часто выращиваемого в качестве декоративного комнатного растения: по краям его листьев развиваются миниатюрные растеньица, снабженные маленькими корешками, эти «почки» в конце концов отпадают и начинают существовать как самостоятельные растения.
Фрагментация разделение особи на две или несколько частей, каждая из которых развивается в новый организм. Фрагментация происходит, например, у нитчатых водорослей, таких как спирогира. Нить спирогиры может разорваться на две части в любом месте, из которых в дальнейшем образуются два организма.
Фрагментация наблюдается также у некоторых низших животных, которые в отличие от более высокоорганизованных форм сохраняют значительную способность к регенерации из относительно слабо дифференцированных клеток. Например, тело немертин (группа примитивных червей, главным образом морских) особенно легко разрывается на много частей, каждая из которых может дать в результате регенерации новую особь. В этом случае регенерация процесс нормальный и регулируемый; однако у некоторых животных (например, у морских звезд) восстановление из отдельных частей происходит только после случайной фрагментации. Фрагментация наблюдается у губок, кишечнополостных (гидр), медуз, иглокожих, кольчатых и плоских червей. Иногда такая способность развита настолько хорошо, что целая особь восстанавливается из отдельного фрагмента.
Рис. . Регенерация морской звезды из одного луча. А, Б, В последовательные стадии регенерации
Вегетативное размножение это размножение, при котором из части, отделившейся от материнского организма, развивается новый дочерний организм. При этом от материнского экземпляра отделяется достаточно хорошо дифференцированная часть, которая далее развивается в самостоятельное растение. Либо растение образует особые структуры, специально предназначенные для вегетативного размножения. Например, луковицы, клубни, клубнелуковицы, корневища, усы, почки. Некоторые из этих структур служат также для запасания питательных веществ, что позволяет растению пережить периоды неблагоприятных условий, таких как холода или засуха. Вегетативное размножение характерно для многих групп растений от водорослей до цветковых.
Принципиально вегетативное размножение практически не отличается от фрагментации или почкования, но традиционно этот термин применяется по отношению к растительным организмам и лишь иногда к животным, в противоположность фрагментации и почкованию.
Клонирование . Это искусственный способ размножения, не встречающийся в естественных условиях. Он получил распространение только в последние 30-40 лет и все чаще используется в хозяйственных целях. Существует ряд специальных методик, позволяющих клонировать некоторые растения и животных. Клоном называется совершенно одинаковое в генетическом отношении потомство, полученное от одной особи в результате того или иного способа бесполого размножения. Отсюда происходит и название данного способа.
Особенно широко эксперименты по клонированию проводятся на растениях, что связано с их высокой способностью к регенерации. Отдельные клетки помещают на питательные среды, где они делятся, и, используя особые методики, получают неорганизованную массу клеток, или каллус. Затем вызывают дифференциацию первично гомогенных каллусов и образование различных тканей и органов и, в конечном счете, целого растительного организма, обладающего всеми теми свойствами, что и исходный, от которого были взяты клетки.
Используя метод клонирования, можно получать различные гибридные формы. Так, с помощью ферментов или ультразвука удаляют клеточные стенки растительных клеток, после чего полученные «голые» протопласты могут сливаться, в результате возникают гибридные клетки (например, томатно-картофельный или табачно-петуниевый гибриды). После этого клеточные стенки восстанавливаются, образуется каллус, а затем и целое гибридное растение.
В случае с животными используется такой метод: ядро яйцеклетки удаляется или разрушается, а на его место помещается ядро какой-либо соматической клетки (например, клетки эпителия). В дальнейшем из такой яйцеклетки может быть получен организм, идентичный по признакам животному донору ядра. Таким способом можно получать клоны некоторых животных шпорцевых лягушек (Xenopus ), тритонов (Triturus ). В настоящее время получены даже клоны млекопитающих, как, например, всем известная овечка Долли. Разработкой методик клонирования занимается особая отрасль биологии биотехнология, задачи которой этим вовсе не ограничиваются.
Половое размножение
Половым размножением называется смена поколений и развитие организмов на основе слияния специализированных половых клеток (гамет ) и образования зиготы. Образование гамет (женской и мужской, яйцеклетки и сперматозоида), происходит в половых железах.
Половое размножение характерно для подавляющего большинства живых существ, является более прогрессивным по сравнению с бесполым и имеет колоссальные генетические преимущества. Половое размножение наилучшим образом обеспечивает генетическое разнообразие потомства, т.к. происходит комбинация генов, до этого принадлежавших обоим родителям. Разнообразие генотипов особей, составляющих вид, обеспечивает возможность более успешного и быстрого приспособления вида к меняющимся условиям обитания.
При оплодотворении гаметы сливаются, образуя диплоидную зиготу, из которой в процессе развития получается зрелый организм. Гаметы гаплоидны - они содержат один набор хромосом, зигота диплоидна, содержит двойной набор хромосом. Это первая клетка будущего организма.
Гаметы обычно бывают двух типов - мужские и женские. Они производятся мужскими и женскими родительскими особями, если вид является раздельнополым ; (таковы некоторые цветковые растения, большинство животных и человек) или одной и той же особью (гармафродитизм).
Для мужских и женских особей животных характерен половой диморфизм - т.е. половые различия в строении, внешнем облике (размеры, окраска и другие свойства), а также в поведении. У животных он встречается уже на низших ступенях эволюционного развития, например у круглых гельминтов, членистоногих, и достигает наибольшего выражения у позвоночных животных, у которых внешние различия между самцами и самками очень выразительны. У растений тех видов, для которых характерно наличие мужских и женских особей, также имеет место половой диморфизм, однако выражен он очень незначительно.
Сравнение полового и бесполого размножения приведено в табл. 5
Табл. 5
Сравнение бесполого и полового размножения
|
Бесполое размножение |
Половое размножение (исключая бактерий) |
|
|
Одна родительская особь |
Обычно две родительские особи |
|
|
Гаметы не образуются |
Образуются гаплоидные гаметы, ядра которых сливаются (оплодотворение), так что получается диплоидная зигота |
|
|
Мейоз отсутствует |
На какой-либо стадии жизненного цикла происходит мейоз, что препятствует удвоению хромосом в каждом поколении. |
|
|
Потомки идентичны родительским особям . Единственный источник генетической изменчивости случайные мутации |
Потомки не идентичны родительским особям. У них наблюдается генетическая изменчивость, возникающая в результате генетической рекомбинации . Это способствует естественному отбору наиболее сильных и приспособленных особей, и, следовательно, эволюции. |
|
|
Характерно для растений, некоторых низших животных и микроорганизмов. У высших животных не встречается |
Характерно для большинства растений и животных |
|
|
Часто приводит к быстрому созданию большого числа потомков |
Менее быстрое увеличение численности |
Половое размножение в зависимости от соотношения половых клеток по размерам и функциям может быть в трех вариантах:
Изогамия (водоросли, простейшие). Мужские и женские половые клетки одинаковы по размерам, строению и подвижности.
Гетерогамия. Клетки отличаются по размерам и строению.
Оогамия. Это один из вариантов гетерогамии, когда яйцеклетка крупная и неподвижная, а сперматозоид меньше по размерам, имеет орган передвижения и обладает подвижностью.
Существуют особые формы полового размножения, такие как гермафродитизм и партеногенез.
Гермафродитизм. Термин «гермафродитизм» является сочетанием греческих имен Гермеса (бог мужской красоты) и Афродиты (богини женской красоты).
Однако у большинства гермафродитных видов в оплодотворении участвуют гаметы, происходящие от разных особей, и у них имеются многочисленные генетические, морфологические и физиологические адаптации, препятствующие самооплодотворению и благоприятствующие перекрестному оплодотворению. Например, у многих простейших самооплодотворение предотвращается генетической несовместимостью, у многих цветковых растений - строением андроцея и гинецея, а у многих животных тем, что яйца и спермии образуются у одной и той же особи в разное время.
Явление истинного гермафродитизма встречается и у более организованных существ. В частности, оно встречается у млекопитающих. Например, у свиней иногда отмечается развитие яичников на одной стороне тела, а развитие тестисов (яичек) на другой, либо происходит развитие комбинированных структур (овотестисов), причем в обоих случаях имеет место синтеза функционально активных яйцеклеток и сперматозоидов. Таких животных относят к «промежуточному» полу, причем большинство особей промежуточного сексуального типа является особями женского пола с двумя ХХ-хромосомами. Аналогичное явление отмечено у коз.
Истинный гермафродитизм встречается и у человека, возникая в результате нарушений развития. Генотипами гермафродитов являются 46ХХ или 46 XY , причем большинство случаев относится к XX (около 60%). Известен и ложный гермафродитизм, когда индивидуумы обладают наружными половыми органами и вторичными половыми признаками, характерными для обоих полов, но продуцируют половые клетки лишь одного типа мужские или женские.
Для большинства цветковых растений характерны гермафродитные цветки, которые обычно называют обоеполыми, поскольку в каждом цветке имеется пестик и тычинки. По этой причине плоды развиваются со всех цветков. Обоеполыми являются пшеница, вишня, яблоня и многие другие виды растений. Помимо обоеполых в ходе эволюции развились растения с разделением полов в пределах одного и того же вида, т. е. возникли однодомность и двудомность растений. Растения, содержащие одновременно пестичные (женские) и тычиночные (мужские) цветки получили название однодомных. У однодомных растений плоды развиваются только из пестичных цветков. Однодомными являются кукуруза, огурец, тыква и другие. Напротив, двудомными являются растения, содержащие либо пестичные, либо тычиночные цветки (в пределах одного и того же вида). У двудомных растений плодоносящими бывают только те, которые имеют пестичные цветки (женские особи). Двудомными являются тополь, клубника и другие виды древесных и травянистых растений.
Партеногенез (от греч. parthenos девственница и genesis рождение) - одна из модификаций полового размножения, при которой женская гамета развивается в новую особь без оплодотворения мужской гаметой. Преимущество партеногенеза состоит в том, что в некоторых случаях он повышает скорость размножения, а у общественных насекомых позволяет регулировать численность потомков каждого типа. Партеногенез может быть облигатный (обязательный) и факультативный (необязательный).
Например, у медоносной пчелы (Apis mellifera ) матка откладывает оплодотворенные яйца (2п = 32), которые, развиваясь, дают самок (маток или рабочих особей), и неоплодотворенные яйца (п = 16), которые дают самцов (трутней), производящих спермин путем митоза, а не мейоза.
Рис. Схема типов партеногенеза.
- Клеточный цикл. Митоз. Мейоз
Деление клетки основа размножения и индивидуального развития организмов.
Все живые организмы состоят из клеток. Развитие, рост, становление типичной структуры организма осуществляются благодаря воспроизведению одной или группы исходных клеток. В процессе жизнедеятельности часть клеток организма изнашивается, стареет и погибает. Для поддержания структуры и нормального функционирования организм должен производить новые клетки на смену старым. Единственным способом образования клеток является деление предшествующих.
Деление клеток жизненно важный процесс для всех организмов. В человеческом организме, состоящем примерно из 10 13 клеток, каждую секунду должны делиться несколько миллионов из них.
Описано три способа деления эукариотических клеток: амитоз (прямое деление), митоз (непрямое деление) и мейоз (редукционное деление).
Амитоз относительно редкий и малоизученный способ деления клетки. Описан он для стареющих и патологически измененных клеток. При амитозе интерфазное ядро делится путем перетяжки, равномерное распределение наследственного материала не обеспечивается. Нередко ядро делится без последующего разделения цитоплазмы и образуются двухъядерные клетки. Клетка, претерпевшая амитоз, в дальнейшем не способна вступать в нормальный митотический цикл. Поэтому амитоз встречается, как правило, в клетках и тканях, обреченных на гибель, например в клетках зародышевых оболочек млекопитающих, в клетках опухолей.
Митоз универсальный способ деления эукариотических клеток. Его продолжительность в клетках животных составляет около 1 ч. Митоз представляет собой непрерывный процесс, который условно делят на четыре фазы: профазу, метафазу, анафазу и телофазу.
Последовательность событий, происходящих между образованием данной клетки и ее делением на дочерние клетки, называют клеточным циклом . Этот цикл состоит из трех главных стадий:
1. Интерфаза. Период интенсивного синтеза и роста. В клетке синтезируется много веществ, необходимых для ее роста и осуществления всех свойственных ей функций. Во время интерфазы происходит репликация ДНК.
2. Митоз. Это процесс деления ядра (кариокинез), при котором хроматиды отделяются одна от другой и перераспределяются в виде хромосом между дочерними клетками.
3. Цитокинез - процесс разделения цитоплазмы (цитокинез) между двумя дочерними клетками.
Продолжительность клеточного цикла зависит от типа клетки и от внешних факторов, таких как температура, питательные вещества и кислород. Бактериальные клетки могут делиться каждые 20 мин, клетки кишечного эпителия - каждые 8-10, клетки в кончике корня лука - каждые 20 ч, а многие клетки нервной системы не делятся никогда.
Соотношение процессов во времени показано на рис.

Рис . Фазы клеточного цикла
Интерфаза состоит из нескольких периодов: G 1, S , G 2.
Период G 1 называют пресинтетическим . Самый вариабельный по продолжительности. В это время в клетке активируются процессы биологического синтеза, в первую очередь структурных и функциональных белков. Клетка растет и готовится к следующему периоду. В данный период происходят интенсивные процессы биосинтеза. Образование митохондрий, хлоропластов (у растений), эндоплазматического ретикулума, лизосом, аппарата Гольджи, вакуолей и пузырьков. Ядрышко продуцирует рРНК, мРНК и тРНК; образуются рибосомы; клетка синтезирует структурные и функциональные белки. Интенсивный клеточный метаболизм, контролируемый ферментами. Рост клетки. Образование веществ, подавляющих или стимулирующих начало следующей фазы.
Период S называют синтетическим . Это главный период в митотическом цикле. В делящихся клетках млекопитающих он длится около 610 ч. Здесь происходит репликация ДНК. Синтез белковых молекул, называемых гистонами, с которыми связывается каждая нить ДНК. Каждая хромосома превращается в две хроматиды.
Период G 2 называют постсинтетическим. Он относительно короток, в клетках млекопитающих он составляет порядка 25 ч. В это время количество центриолей, митохондрий и пластид удваивается, идут активные метаболические процессы, накапливаются белки и энергия для предстоящего деления. Клетка приступает к делению. Происходят интенсивные процессы биосинтеза. Деление митохондрий и хлоропластов. Увеличение энергетических запасов. Репликация центриолей (в тех клетках, где они имеются) и начало образования веретена деления
Митоз условно делят на четыре фазы: профазу, метафазу, анафазу и телофазу
Профаза . В ядре начинается и постепенно нарастает спирализация ДНК. Хромосомы укорачиваются, утолщаются, становятся видимыми, приобретают типичную двухроматидную структуру. Ядрышко постепенно исчезает. В цитоплазме вокруг каждой пары центриолей ориентируются микротрубочки, образуя центры веретена деления. Центриоли движутся к разным полюсам, микротрубочки вытягиваются вдоль оси клетки начинается формирование ахроматинового веретена. Ядерная оболочка распадается на отдельные мелкие фрагменты. Хромосомы направляются к центру клетки,
Метафаза . Хромосомы максимально спирализуются и располагаются таким образом, что их центромеры лежат в одной плоскости плоскости экватора клетки. Образуется метафазная пластинка.Завершается формирование митотического веретена. Центриоли попарно располагаются на противоположных полюсах, а нити веретена от разных полюсов прикрепляются к центромере каждой хромосомы.
Анафаза . Это самая короткая фаза митоза. Здесь происходят продольное расщепление каждой хромосомы, сокращение нитей веретена и расхождение хроматид (дочерних хромосом) по направлению к полюсам клетки.
Телофаза . Дочерние хромосомы, состоящие из одной хро-матиды, достигают полюсов клетки. Составляющая их ДНК начинает деспирализоваться, появляется ядрышко, вокруг каждой группы дочерних хромосом образуется ядерная оболочка, нити ахроматинового веретена постепенно распадаются. Деление ядра завершается.
Начинается деление цитоплазмы (цитотомия) и образование перегородки между дочерними клетками. Животные клетки осуществляют цитотомию путем перетяжки цитоплазматической мембраны. У растений в плоскости экватора клетки образуется мембранная перегородка, которая растет в стороны, достигая клеточной стенки. В результате образуются две полностью разделенные дочерние клетки.
Проследим изменение наследственного материала в ходе митотического цикла. Главными событиями митотического цикла являются репликация ДНК, происходящая в интерфазе и приводящая к удвоению количества наследственной информации, и расхождение хроматид, происходящее в анафазе митоза и обеспечивающее равномерное распределение наследственной информации между дочерними клетками. Эти процессы наследственный материал осуществляет, находясь в разных структурных формах. Репликативный синтез претерпевает интерфазный хроматин, в составе которого молекула ДНК находится в относительно деспирализованном состоянии. Распределение генетической информации осуществляют митотические хромосомы, в составе которых ДНК максимально спирализована.
В митотическом цикле изменяется также количество наследственного материала. Если число хромосом в гаплоидном наборе обозначать буквой п (в диплоидном наборе соответственно 2п), а число молекул ДНК обозначить буквой с, то можно проследить изменение формулы ядра соматической клетки на разных стадиях митотического цикла. До S -периода, когда каждая хромосома состоит из одной молекулы ДНК, общее количество ДНК в ядре соответствует числу хромосом в нем, а формула диплоидной клетки имеет вид 2п2с. После репликации, когда ДНК каждой хромосомы себя удваивает, суммарное количество ДНК в ядре увеличивается вдвое и формула клетки приобретает вид 2п4с. В результате расхождения хроматид в анафазе митоза дочерние ядра получают диплоидный набор однохроматидных хромосом. Формула дочерних клеток вновь становится 2п2с.
Биологическое значение митоза заключается в том, что в результате этого способа деления образуются клетки с наследственной информацией, которая качественно и количественно идентична информации материнской клетки. Равномерное распределение наследственного материала обеспечивается процессами репликации ДНК и удвоения хромосом в интерфазе митотического цикла, а также спирализацией и равномерным распределением хроматид между дочерними клетками в процессе митоза. Митоз обеспечивает поддержание постоянства кариотипа в ряду поколений клеток и служит клеточным механизмом процессов роста и развития организма, а также регенерации и бесполого размножения.
Действие ряда факторор внешней среды может нарушать нормальное течение митоза и приводить к повреждениям хромосом, а также к изменению числа отдельных хромосом или целых хромосомных наборов в соматических клетках организма. Патологические митозы могут стать причиной ряда хромосомных болезней. Особенно часто патологические митозы наблюдаются в опухолевых клетках.
Схема митоза:
МИТОЗ
|
1. Интерфаза Ее часто неправильно называют стадией покоя. Продолжительность интерфазы различна и зависит от функции данной клетки. Это период, во время которого клетка обычно синтезирует органеллы и увеличивается в размерах. Ядрышки хорошо видны и активно синтезируют рибосомный материал. Непосредственно перед клеточным делением ДНК и гистоны каждой хромосомы реплицируются. Каждая хромосома представлена теперь парой хроматид, соединенных друг с другом центромерой. Вещество хромосом окрашивается и носит название хроматина, но сами эти структуры увидеть трудно. |
|
|
2. Профаза Обычно самая продолжительная фаза клеточного деления. Хроматиды укорачиваются (до 4% своей первоначальной длины) и утолщаются в результате их спирализации и конденсации. При окрашивании хроматиды четко видны, но центромеры не выявляются. В разных парах хроматид центромера располагается по-разному. В животных клетках и у низших растений центриоли расходятся к противоположным полюсам клетки. От каждой центриоли в виде лучей расходятся короткие микротрубочки, образующие в совокупности звезду. Ядрышки уменьшаются, так как их нуклеиновая кислота частично переходит в определенные пары хроматид. К концу профазы ядерная мембрана распадается и образуется веретено деления. |
|
|
3. Метафаза Пары хроматид прикрепляются своими центромерами к нитям веретена деления (микротрубочкам) и перемещаются вверх и вниз по веретену до тех пор, пока их центромеры не выстроятся по экватору веретена перпендикулярно его оси. |
|
|
4. Анафаза Это очень короткая стадия. Каждая центромера расщепляется на две, и нити веретена оттягивают дочерние центромеры к противоположным полюсам. Центромеры тянут за собой отделившиеся одна от другой хроматиды, которые теперь называются хромосомами. |
|
|
5. Телофаза Хромосомы достигают полюсов клетки, деспирализуются, удлиняются, и их уже нельзя четко различить. Нити веретена разрушаются, а центриоли реплицируются. Вокруг хромосом на каждом из полюсов образуется ядерная оболочка. Вновь появляется ядрышко. За телофазой может сразу следовать цитокинез (разделение всей клетки на две). |
|





Мейоз (от греч. мейозис уменьшение) своеобразный способ деления клеток, приводящий к уменьшению в них числа хромосом вдвое. Мейоз является центральным звеном гаметогенеза у животных и спорогенеза у растений. Мейоз состоит из двух последовательных делений, которым предшествует однократная редупликация ДНК. Все вещества и энергия, необходимые для осуществления обоих делений, запасаются в ходе предшествующей мейозу интерфазы I . Интерфаза II практически отсутствует, и деления быстро следуют одно за другим. В каждом из делений мейоза различают те же четыре стадии: профазу, метафазу, анафазу и телофазу, которые характерны для митоза, но отличаются рядом особенностей.
Первое мейотическое деление (мейоз I ) приводит к уменьшению вдвое числа хромосом и называется редукционным. В результате из одной диплоидной клетки (2п 4с) образуются две гаплоидные (п 2с) клетки.
Профаза I мейоза наиболее продолжительна и сложна. Помимо типичных для профазы митоза процессов спирализации ДНК и образования веретена деления в профазе I происходят два исключительно важных в биологическом отношении события: конъюгация, или синапсис, гомологичных хромосом и кроссинговер.
Конъюгация это процесс тесного сближения гомологичных хромосом. Такие спаренные хромосомы образуют бивалент и удерживаются в его составе с помощью специальных белков. Поскольку каждая из хромосом состоит из двух хроматид, бивалент включает четыре хроматиды и называется также тетрадой. В диплоидной клетке образуется п бивалентов. После конъюгации формула клетки приобретает вид пАс.
В некоторых местах бивалента хроматиды конъюгированных хромосом перекрещиваются, рвутся и обмениваются соответствующими участками. Такой процесс обмена фрагментами гомологичных хромосом называется кроссинговером. Он обеспечивает образование новых комбинаций отцовских и материнских генов в хромосомах будущих гамет. Кроссинговер может происходить в нескольких участках (множественный кроссинговер), обеспечивая более высокую степень рекомбинации наследственной информации в гаметах. К концу профазы I степень спирализации хромосом возрастает, хроматиды становятся хорошо различимыми, нити веретена деления от каждого полюса прикрепляются к центромере одной из хромосом бивалента. Ядерная оболочка разрушается, и биваленты направляются к плоскости экватора клетки.
В метафазе I мейоза завершается формирование веретена деления, биваленты устанавливаются в плоскости экватора клетки. Нити веретена с одного полюса прикрепляются к центромере каждой хромосомы.
В анафазе I мейоза под действием нитей веретена гомологичные хромосомы отходят друг от друга, направляясь к противоположным полюсам клетки. В результате у каждого из полюсов клетки формируется гаплоидный набор хромосом, содержащий по одной двухроматидной хромосоме из каждой пары гомологичных хромосом. В анафазе I хромосомы разных пар, т.е. негомологичные хромосомы, ведут себя совершенно независимо друг от друга, обеспечивая образование самых различных комбинаций отцовских и материнских хромосом в гаплоидном наборе будущих гамет. Число таких комбинаций соответствует формуле 2 П , где п число пар гомологичных хромосом. У человека эта величина равна 2 , т.е. 8,4 10 вариантов сочетаний отцовских и материнских хромосом возможно в гаметах человека.
Итак, расхождение гомологичных хромосом в анафазе I мейоза обеспечивает не только редукцию числа хромосом в будущих половых клетках, но и огромное разнообразие последних в силу случайного сочетания отцовских и материнских хромосом разных пар.
В телофазе I мейоза происходит формирование клеток, ядра которых имеют гаплоидный набор хромосом и удвоенное количество ДНК, поскольку каждая хромосома состоит из двух хроматид. Клетки, образующиеся в результате первого мейотического деления, имеют формулу п2с и после короткой интерфазы приступают к следующему делению.
Второе мейотическое деление (мейоз II ) протекает как типичный митоз (рис..5.3), но отличается тем, что вступающие в него клетки содержат гаплоидный набор хромосом. В результате такого деления п двухроматидных хромосом (я2с), расщепляясь, образуют п однохроматидных хромосом (пс). Такое деление называют эквационным (или уравнительным).
Таким образом, после двух последовательных мейотических делений из одной клетки с диплоидным набором двухроматидных хромосом (2я4с) образуются четыре клетки с гаплоидным набором однохроматидных хромосом (пс).
Биологическое значение мейоза заключается в образовании клеток с редуцированным набором хромосом и поддержании постоянства кариотипа в ряду поколений организмов, размножа-. ющихся половым путем. Мейоз служит основой комбинативной изменчивости, обеспечивая генетическое разнообразие гамет благодаря процессам кроссинговера, расхождения и комбинаторики отцовских и материнских хромосом. Изменения структуры хромосом вследствие неравного кроссинговера, нарушение расхождения всех или отдельных хромосом в анафазе I и II мейотических делений приводят к образованию аномальных гамет и могут служить основой гибели организма или развития у потомков ряда хромосомных синдромов.
Схема и краткие описания последовательных стадий мейоза в животной клетке.
|
1. Интерфаза Продолжительность различна у разных видов. Происходит репликация органелл, и клетка увеличивается в размерах. Репликация ДНК и гистонов в основном заканчивается в премейотической интерфазе, но частично захватывает и начало профазы. Каждая хромосома представлена теперь парой хроматид, соединенных центромерой. Хромосомный материал окрашивается, но из всех структур четко выявляются только ядрышки. |
А. Ранняя профаза I |
|
2. Профаза Самая продолжительная фаза. Ее часто делят на пять стадий (лептотема, зигонема, пахинема, диплонема и диакинез), но здесь оно будет рассматриваться как непрерывная последовательность изменений хромосом. А . Хромосомы укорачиваются и становятся видимыми как обособленные структуры. У некоторых организмов они выглядят как нитки бус: участки интенсивно окрашивающегося материала хромомеры - чередуются у них с неокрашивающимися участками. Хромомеры - это те места, где хромосомный материал сильно спирализован. |
|
|
Б. Гомологичные хромосомы, происходящие из ядер материнской и отцовской гамет, приближаются одна к другой и конъюгируют. Эти хромосомы одинаковой длины, их центромеры занимают одинаковое положение, и они обычно содержат одинаковое число генов, расположенных в одной и той же линейной последовательности. Хромомеры гомологичных хромосом лежат бок о бок. Процесс конъюгации может начинаться в нескольких точках хромосом, которые затем соединяются по всей длине (как бы застегиваясь на «молнию»). Пары конъюгировавших гомологичных хромосом часто называют бивалентами. Биваленты укорачиваются и утолщаются. При этом происходит как более плотная упаковка на молекулярном уровне, так и внешне заметное закручивание (спирализация). Теперь каждая хромосома с ее центромерой ясно видна. |
Б. Профаза I |
|
В. Гомологичные хромосомы, составляющие бивалент, частично разделяются, как будто, отталкиваясь, друг от друга. Теперь видно, что каждая хромосома состоит из двух хроматид. Хромосомы все еще соединены между собой в нескольких точках. Эти точки называют хиазмами (от греч. chiasma - перекрест). В каждой хиазме происходит обмен участками хроматид в результате разрывов и воссоединений, в которых участвуют две из четырех имеющихся в каждой хиазме нитей. В результате гены из одной хромосомы (например, отцовской - А, В, С) оказываются связанными с генами из другой хромосомы (материнской а, b , с), что приводит к новым генным комбинациям в образующихся хроматидах. Этот процесс называют кроссинговером. Гомологичные хромосомы после кроссинговера не расходятся, так как сестринские хроматиды (обеих хромосом) остаются прочно связанными вплоть до анафазы. |
В. Кроссинговер во время профазы I |
|
Г . Хроматиды гомологичных хромосом продолжают отталкиваться друг от друга, и биваленты приобретают определенную конфигурацию в зависимости от числа хиазм. Биваленты с одной хиазмой имеют крестообразную форму, с двумя хиазмами кольцевидную, а с тремя и более - образуют петли, лежащие перпендикулярно друг другу. К концу профазы все хромосомы полностью уплотнены и интенсивно окрашиваются. В клетке происходят и другие изменения: миграция центриолей (если они имеются) к полюсам, разрушение ядрышек и ядерной мембраны, а затем образование нитей веретена деления. |
|
|
2. Метафаза Биваленты выстраиваются в экваториальной плоскости, образуя метафазную пластинку. Их центромеры ведут себя как единые структуры (хотя часто выглядят двойными) и организуют прикрепленные к ним нити веретена, каждая из которых направлена только к одному из полюсов. В результате слабого тянущего усилия этих нитей каждый бивалент располагается в области экватора, причем обе его центромеры оказываются на одинаковом расстоянии от него одна снизу, а другая сверху. |
Д. Поздняя метафаза I |
|
3. Анафаза Имеющиеся у каждого бивалента две центромеры еще не делятся, но сестринские хроматиды уже не примыкают одна к другой. Нити веретена тянут центромеры, каждая из которых связана с двумя хроматидами, к противоположным полюсам веретена. В результате хромосомы разделяются на два гаплоидных набора, попадающих в дочерние клетки. |
Е . Анафаза I |
|
4. Телофаза Расхождение гомологичных центромер и связанных с ними хроматид к противоположным полюсам означает завершение первого деления мейоза. Число хромосом в одном наборе стало вдвое меньше, но находящиеся на каждом полюсе хромосомы состоят из двух хроматид. Вследствие кроссинговера при образовании хиазм эти хроматиды генетически неидентичны, и при втором делении мейоза им предстоит разойтись. Веретена и их нити обычно исчезают. У животных и у некоторых растений хроматиды деспирализуются, вокруг них на каждом полюсе формируется ядерная мембрана и образовавшееся ядро вступает в интерфазу. Затем начинается деление цитоплазмы (у животных) или формирование разделяющей клеточной стенки (у растений), как при митозе. У многих растений не наблюдается ни телофазы, ни образования клеточной стенки, ни интерфазы, и клетка прямо переходит из анафазы I в профазу II . |
Ж. Телофаза I в животной клетке |







|
Интерфаза II Эта стадия обычно наблюдается только в животных клетках: продолжительность ее варьирует. Фаза S отсутствует, и дальнейшей репликации ДНК не происходит. Процессы, участвующие во втором делении мейоза, по своему механизму сходны с происходящими в митозе. Они включают разделение хроматид в обеих дочерних клетках, получившихся в результате первого деления мейоза. Второе деление мейоза отличается от митоза главным образом двумя особенностями: 1) в метафазе II мейоза сестринские хроматиды часто сильно обособляются друг от друга; 2) число хромосом гаплоидное. |
|
|
Профаза II В клетках, у которых выпадает интерфаза II , эта стадия тоже отсутствует. Продолжительность профазы II обратно пропорциональна продолжительности телофазы I . Ядрышки и ядерные мембраны разрушаются, а хроматиды укорачиваются и утолщаются. Центриоли, если они есть, перемещаются к противоположным полюсам клеток; появляются нити веретена деления. Хроматиды располагаются таким образом, что их длинные оси перпендикулярны оси веретена первого деления мейоза. |
З. Профаза П |
|
Метафаза II При втором делении центромеры ведут себя кок двойные структуры. Они организуют нити веретена, направленные к обоим полюсам, и таким образом выстраиваются по экватору веретено. |
Метафаза II |
|
Анафаза II Центр омеры делятся, и нити веретена растаскивают их к противоположным полюсам. Центромеры тянут за собой отделившиеся друг от друга хроматиды, которые теперь называются хромосомами. Телофаза II Эта стадия очень сходна с телофазой митоза. Хромосомы деспирализуются, растягиваются и после этого плохо различимы. Нити веретена исчезают, а центриоли реплицируются. Вокруг каждого ядра, которое содержит теперь половинное (гаплоидное) число хромосом исходной родительской клетки, вновь образуется ядерная мембрана. В результате последующего деления цитоплазмы (у животных) или образования клеточной стенки (у растений) из одной исходной родительской клетки получается четыре дочерних клетки. |
|


Различия между стадиями митоза и мейоза
|
Стадия |
Митоз |
Мейоз |
|
Профаза |
Хромомеры не видны Гомологичные хромосомы обособлены Хиазмы не образуются Кроссинговсра не происходит |
Хромомеры видны Гомологичные хромосомы конъюгируют. Хиазмы образуются. Кроссинговер может иметь место |
|
Метафаза |
Пары хроматид располагаются на экваторе веретена Центромеры выстраиваются в одной плоскости на экваторе веретена |
Пары хроматид располагаются на экваторе веретена только во втором делении мейоза Центромеры в первом делении мейоза располагаются над и под экватором на одинаковых расстояниях от него |
|
Анафаза |
Центромеры делятся. Хроматиды расходятся. Расходящиеся хроматиды идентичны |
Центромеры делятся только во втором деянии мейоза. Хроматиды расходятся во втором делении мейоза. В первом делении расходятся целые хромосомы. Расходящиеся хромосомы могут оказаться неидентичными в результате кроссинговера |
|
Телолофаза |
Число хромосом в дочерних клетках то же, что и в родительских клетках Дочерние клетки содержат обе гомологичные хромосомы (у диплоидов) |
Число хромосом в дочерних клетках вдвое меньше, чем в родительских Дочерние клетки содержат только по одной из каждой пары гомологичных хромосом |
|
Где происходит деление данного типа |
Возможно в гаплоидных, диплоидных и полиплоидных клетках Происходит при образовании соматических клеток и некоторых спор, а также при образовании гамет у растений, у которых имеет место чередование поколений |
Только в диплоидных и полиплоидных клетках При гамето- или спорогенезе |
- Строение гамет. Гаметогенез
Половое размножение осуществляется с помощью специализированных половых клеток гамет. Женские гаметы называют яйцеклетками, мужские сперматозоидами. Гаметы отличаются от соматических клеток прежде всего вдвое меньшим числом хромосом, а также низким уровнем обменных процессов.
Яйцеклетки относительно крупные неподвижные клетки, обычно округлой формы; в цитоплазме помимо типичных органоидов содержатся включения запасных питательных веществ в виде желтка (рис .). В ядрах яйцеклеток образуется много копий рибосомальных генов и мРНК, обеспечивающих синтез жизненно важных белков будущего зародыша. Яйцеклетки разных организмов различаются количеством и характером распределения в них желтка. Различают несколько типов яйцеклеток.

Изолециталъными называют относительно мелкие яйцеклетки с небольшим количеством равномерно распределенного желтка. Ядро в них располагается ближе к центру. Такие яйцеклетки встречаются у червей, двустворчатых и брюхоногих моллюсков, иглокожих, ланцетника. Умеренно телолециталъные яйцеклетки осетровых рыб и земноводных имеют диаметр около 1,5 2 мм и содержат среднее количество желтка, основная масса которого сосредоточена на одном из полюсов (вегетативном). На противоположном полюсе (анимальном), где желтка мало, находится ядро яйцеклетки.
Резко телолециталъные яйцеклетки некоторых рыб, пресмыкающихся, птиц и яйцекладущих млекопитающих содержат очень много желтка, занимающего почти весь объем цитоплазмы яйцеклетки. На анимальном полюсе находится зародышевый диск с активной, лишенной желтка цитоплазмой. Размеры этих яиц крупные 10 15 мм и более.
Алециталъные яйцеклетки практически лишены желтка, имеют микроскопически малые размеры (0,1 0,3 мм) и характерны для плацентарных млекопитающих, в том числе и для человека.
Сперматозоиды или спермии - это очень мелкие подвижные мужские гаметы (например, спермий человека имеют длину 50 70 мкм, а крокодила 20 мкм), образуемые мужскими гонадами - семенниками; их количество исчисляется миллионами. Форма спермиев у разных животных различна, однако строение их однотипно, большинство из них имеет головку, шейку. В головке сперматозоида находится ядро, содержащее гаплоидное число хромосом и прикрытое акросомой . Акросома - особая структура, видоизмененный комплекс Гольджи, который содержит ферменты для растворения оболочки яйцеклетки при оплодотворении ограниченная мембраной. В шейке находятся многочисленные митохондрии и две центриоли. От шейки отрастает хвост, образованный микротрубочками и обеспечивающий подвижность сперматозоидов. Средняя часть расширена за счет содержащихся в ней многочисленных митохондрий, собранных в спираль вокруг жгутика. Эти митохондрии доставляют энергию для сократительных механизмов, обеспечивающих движения жгутика. Главная и хвостовая части сперматозоида имеют строение, характерное для жгутиков.

Если смотреть на головку человеческого спермия сверху, то она кажется округлой, а при взгляде сбоку - уплощенной. Одного только жгутикового движения недостаточно, чтобы сперматозоид мог пройти расстояние от влагалища до места, где происходит оплодотворение. Главная локомоторная задача спермиев состоит в том, чтобы скопиться вокруг ооцита и ориентироваться определенным образом, прежде чем проникнуть сквозь мембраны ооцита.
Образование гамет (гаметогенез). Клетки зачаткового эпителия в мужских и женских половых железах (гонадах) претерпевают ряд последовательных митотических и мейотических делений, называемых в совокупности гаметогенезом, в результате чего образуются зрелые мужские гаметы (сперматогенез) и женские гаметы (оогенез). В обоих случаях процесс делится на три фазы - фазу размножения, фазу роста и фазу созревания. Фаза размножения включает многократные митотические деления, приводящие к образованию множества сперматогоний или оогоний. Каждая из них проходит период роста в порядке подготовки к первому делению мейоза и последующему цитокинезу. Затем начинается фаза созревания, во время которой происходят первое и второе деления мейоза с последующей дифференцировкой гаплоидных клеток и формированием зрелых гамет.
Развитие сперматозоидов (сперматогенез). Сперматозоиды (спермин) образуются в результате ряда последовательных клеточных делений, называемых в совокупности сперматогенезом, за которыми следует сложный процесс дифференцировки, называемый спермиогенезом (рис. 20.31 ). Процесс образования спермия занимает примерно 70 дней; на 1 г веса яичка образуется 10 7 спермиев в сутки. Эпителий семенного канальца состоит из наружного слоя клеток зачаткового эпителия и примерно шести слоев клеток, образовавшихся в результате многократных делений клеток этого слоя (рис. 20.32 и 20.33); эти слои соответствуют последовательным стадиям развития сперматозоидов. Сначала деление клеток зачаткового эпителия дает начало многочисленным сперматогониям, которые увеличиваются в размерах и становятся сперматоцитами первого порядка. Эти сперматоциты в результате первого деления мейоза образуют гаплоидные сперматоциты второго порядка, после чего претерпевают второе деление мейоза и превращаются в сперматиды. Между «тяжами» из развивающихся клеток расположены крупные клетки Сертоли, или трофические клетки, расположенные во всем пространстве от наружного слоя канальца до его просвета.

Сперматоциты располагаются в многочисленных впячиваниях на боковых поверхностях клеток Сертоли; здесь они превращаются в сперматиды, а затем переходят на тот край клетки Сертоли, который обращен к просвету семенного канальца, где они созревают, образуя спермин (рис. 20.33 ). По-видимому, клетки Сертоли обеспечивают созревающим спермиям механическую опору, защиту и питание. Все питательные вещества и кислород, доставляемые развивающимся гаметам по окружающим семенные канальцы кровеносным сосудам, и выделяемые в кровь отходы метаболизма проходят через клетки Сертоли. Эти клетки секретируют и жидкость, с которой спермин проходят по канальцам.
Развитие яйца у человека (оогенез). В отличие от образования спермиев, которое начинается у мужчин только при половом созревании, образование яйцеклеток у женщин начинается еще до их рождения и завершается для каждой данной яйцеклетки только после ее оплодотворения. Стадии оогенеза показаны на рис. 20.36. Во время развития плода первичные половые клетки многократно делятся путем митоза, образуя много крупных клеток, называемых оогониями. Оогонии снова претерпевают митоз и образуют ооцигы первого порядка, которые остаются на стадии профазы почти до самой овуляции. Ооциты первого порядка окружены одним слоем клеток -гранулезной оболочкой -и образуют так называемые примордиальные фолликулы. Плод женского пола непосредственно перед рождением содержит около 2 10 6 этих фолликулов, но лишь примерно 450 из них достигают стадии ооцитов второго порядка и выходят из яичника (овуляция). Перед овуляцией ооцит первого порядка проделывает первое деление мейоза, образуя гаплоидный ооцит второго порядка и первое полярное тельце. Второе деление мейоза доходит до стадии метафазы, но не продолжается дальше до тех пор, пока ооцит не сольется со сперматозоидом. При оплодотворении ооцит второго порядка совершает второе деление мейоза, образуя крупную клетку - яйцо, а также второе полярное тельце. Все полярные тельца представляют собой мелкие клетки; они не играют никакой роли в оогенезе и в конечном счете разрушаются.

партеногенез
облигатный
факультативный
Развитие организмов только из неоплодотворенной яйцеклетки.
Формируются только женские особи.
Например, кавказская скальная ящерица
летки могут развиваться как без оплодотворения, так и с оплодотворением
Женский тип
Из неоплодотворенных яйцеклеток развиваются самцы
Мужской тип
У некоторых изогамных водорослей
Другие похожие работы, которые могут вас заинтересовать.вшм> |
|||
| 6644. | Индивидуальное развитие организмов (онтогенез) | 78.69 KB | |
| Считают что в яйцеклетку животных проникает ядерное содержимое лишь одного сперматозоида. В случае человека а иногда и высших животных период развития до рождения часто называют пренатальным после рождения постнатальным. У большинства многоклеточных животных независимо от сложности их организации стадии эмбрионального развития которые проходит зародыш едины. Характер дробления и типы бластул у разных позвоночных... | |||
| 13714. | Индивидуальное развитие организмов и их поведение. Онтогенез. Жизненный цикл у растений и животных | 9.96 KB | |
| Онтогенез личность происхождение это индивидуальное развитие организма с момента образования зиготы до его смерти. Гаметы это половые клетки которые несут наследственную информацию и имеют гаплоидный набор. Оплодотворение это процесс слияния мужской и женской половых клеток растений или животных является основой полового процесса. Эмбриональный зародышевый период это период онтогенез от образования зиготы до рождения или выхода из яичных оболочек или прорастания. | |||
| 10427. | Размножение | 6.75 KB | |
| При огромном разнообразии форм размножения организмов все они могут быть сведены к двум основным типам: бесполому и половому. При бесполом размножении воспроизведение потомства происходит от одной родительской особи путем образования спор или вегетативно. При вегетативном размножении потомство возникает от отделившихся от материнской особи участков тела. При вегетативном размножении у растений сохраняется гетерозиготность в течение многих поколений. | |||
| 21332. | Лимитирующие факторы. Адаптация организмов к факторам | 303.8 KB | |
| Некоторые животные любят сильную жару иные лучше переносят умеренную температуру среды и т. Кроме того живые организмы делят на способных существовать в широком или узком диапазонах изменения какого-либо фактора среды. Если влияние условий среды не достигает предельных значений живые организмы реагируют на него определенными действиями или изменениями своего состояния что в конечном итоге ведет к выживанию вида. Объектом исследования в данной работе являются факторы среды обитания предметом лимитирующие факторы и адаптация организмов к... | |||
| 8875. | Общие закономерности взаимодействия организмов и экологических факторов | 193.58 KB | |
| Биотические факторы наземной и водной среды почв Биологически активные вещества живых организмов Антропогенные факторы Общие закономерности взаимодействия организмов и экологических факторов Понятие лимитирующего фактора. Закон минимума Либиха закон Шелфорда Специфика воздействия антропогенных факторов на организм Классификация организмов по отношению к экологическим факторам 1. Условия ковыльных степей представляют совершенно иные режимы абиотических факторов. | |||
| 12700. | Биологические особенности вредных организмов и меры борьбы с ними | 62.79 KB | |
| Особенно значительные потери урожая происходят в результате присутствия сорных растений которые выносят питательные вещества и влагу из почвы затеняют культурные растения а во многих случаях и загрязняют продукцию ядовитыми веществами и семенами вызывающими отравление человека и животных. Основные направления химизации сельского хозяйства: применение удобрений химических средств защиты растений от вредителей болезней и сорняков использование химических продуктов в животноводстве консервировании сельскохозяйственных продуктов и... | |||
| 13403. | Вирусы как неклеточная форма жизни. Строение, классификация, взаимодействие вируса с кретками различных организмов | 12.75 KB | |
| Это биологические объекты геномы котх состт из нуклиновых кислот ДНК или РНК продуцируют в живых клетках используя их биосинтетический аппарат. Отличия вирусов от других форм жизни: не имеют клеточного строения 1 тип нуклеиновых кислот только ДНК или РНК нет собствго метаболизма. Гипотеза происхождения вирусов: вирусы возникли из компонентов нормальной клетки вышедших изпод контроля регулирующих механизмов и превратившихся в самостоятельную единицу вероятно на участке ДНК произошла серия генетических изменений которая... | |||
| 18798. | Структурно-функциональные показатели организмов почвенной мезофауны в лесных и открытых биотопах долины р.Позимь | 61.54 KB | |
| Однако на сегодняшний день освоение данной территории является следствием ряда экологических проблем. В частности в долине широко проводятся работы по осушению пойменных лугов, что непосредственно влияет на естественные биотопы. | |||
| 19386. | Развитие либерализма в СНГ | 35.55 KB | |
| Сложносистемный характер рассматриваемых изменений в процессах эволюции внешнеэкономических связей обусловливает пересмотр и изменение всей аксиоматики традиционных представлений об идентификации, границах и суверенности национального хозяйствования... | |||
| 8867. | ОБУЧЕНИЕ И РАЗВИТИЕ | 162.32 KB | |
| Суть проблемы соотношения обучения и развития. Основные подходы к решению проблемы соотношения обучения и развития. Концепция зоны ближайшего развития Л. Суть проблемы соотношения обучения и развития. | |||
Размножение - это свойство организмов производить потомство или способность организмов к самовоспроизведению. Являясь важнейшим свойством живого, размножение обеспечивает непрерывность жизни, продолжение видов.
Процесс размножения исключительно сложен и связан не только с передачей генетической информации от родителей к потомству, но и с анатомическими и физиологическими свойствами организмов, с их поведением, гормональным контролем. Размножение организмов сопровождается процессами их роста и развития.
Для живых существ характерно чрезвычайное разнообразие в способах размножения. Тем не менее различают два основных способа размножения - бесполое и половое (рис. 16). Бесполое размножение, или апомиксис (от греч. аро - без, mixis - смешение), представляет собой процесс, в котором участвует лишь один родитель (клетка или многоклеточный организм). Напротив, в половом размножении участвуют два родителя, каждый из которых имеет собственную репродуктивную систему и продуцирует половые клетки (гаметы), которые после слияния образуют зиготу (оплодотворенное яйцо), дифференцирующуюся затем в эмбрион. Следовательно, при половом размножении имеет место смешение наследственных факторов, т. е. процесс, называемый амфимиксисом (от греч. amphi - с обеих сторон, mixis - смешение).
БЕСПОЛОЕ РАЗМНОЖЕНИЕ
Бесполое размножение характерно для организмов многих видов как растений, так и животных. Оно встречается у вирусов, бактерий, водорослей, грибов, сосудистых растений, простейших, губок, кишечнополостных, мшанок и оболочников.
Наиболее простая форма бесполого размножения характерна для вирусов. Их репродуктивный процесс связан с молекулами нуклеиновых кислот, со способностью этих молекул к самоудвоению
и основан на специфичности относительно слабых водородных связей между нуклеотидами.
Рис. 16. Способы размножения организмов
Применительно к другим организмам, размножающимся бесполым путем, различают вегетативное размножение и размножение спорообразованием.
Вегетативное размножение - это размножение, при котором из части, отделившейся от материнского организма, развивается новый организм. Этот вид размножения характерен как для одноклеточных, так и многоклеточных организмов, но имеет у них разное проявление.
У одноклеточных организмов вегетативное размножение представлено такими формами, как деление, множественное деление и почкование. Деление путем простой перетяжки с образованием при этом из одного родительского организма двух дочерних присуще бактериям и синезеленым водорослям (цианобактериям). Напротив, размножение делением бурых и зеленых водорослей, а также одноклеточных животных (саркодовых, жгутиковых и инфузорий) происходит путем митотического деления ядра с последующей перетяжкой цитоплазмы.
Размножение путем множественного деления (шизогонии) заключается в делении ядра с последующим разделением цитоплазмы на части. В результате такого деления из одной клетки образуется несколько дочерних организмов. Примером множественного деления является размножение малярийного плазмодия (P. vivax) в эритроцитах человека. В этом случае у плазмодиев происходит повторяющееся много раз деление ядра без цитокинеза, после чего следует и цитокинез. В результате этого один плазмодий дает начало 12-24 дочерним организмам.
У многоклеточных растительных организмов вегетативное размножение путем деления осуществляется черенками, луковицами, листьями, корневищами. Но это, по существу, искусственное размножение, используемое в сельскохозяйственной практике. Размножение высших растений в искусственных условиях возможно и из одной клетки. Организмы, развивающиеся из одной клетки (клон), обладают всеми свойствами исходного многоклеточного организма. Это размножение получило название клонального микроразмножения. В качестве одной из форм вегетативного размножения могут служить прививки или трансплантации многих культурных растений, заключающиеся в пересадке почки или части побега от одного растения к другому. Конечно, это тоже способ размножения, который в природе не встречается, но в сельском хозяйстве используется очень широко.
У многоклеточных животных вегетативное размножение происходит путем фрагментации их тела на части, после чего каждая
часть развивается в новое животное. Такое размножение характерно для губок, кишечнополостных (гидр), немертин, плоских чер- вей, иглокожих (морских звезд) и некоторых других организмов. Близкой формой к вегетативному размножению животных фрагментацией является полиэмбриония животных, заключающаяся в том, что на определенной стадии развития эмбрион разделяется на несколько частей, каждая из которых развивается в самостоятель- ный организм. Полиэмбриония встречается у броненосцев. Однако последние размножаются половым путем. Поэтому полиэмбриония является скорее своеобразной стадией в половом размножении, а потомство, возникающее в результате полиэмбрионии, представлено монозиготными близнецами.
Почкование заключается в том, что на материнской клетке образуется бугорок (вырост) с ядром, который затем отделяется и становится самостоятельным организмом. Почкование встречается как у одноклеточных растений, например у дрожжей, так и у одноклеточных животных, например у инфузорий отдельных видов.
Размножение спорообразованием связано с образованием специализированных клеток - спор, которые содержат ядро, цитоплазму, покрыты плотной оболочкой и способны к длительному существованию в неблагоприятных условиях, что способствует, кроме того, и их расселению. Наиболее часто такое размножение встречается у бактерий, водорослей, грибов, мхов, папоротникообразных. У некоторых зеленых водорослей из отдельных клеток могут формироваться так называемые зооспоры.
Среди животных размножение спорообразованием отмечается у споровиков, в частности у малярийного плазмодия.
У организмов многих видов бесполое размножение может чередоваться с половым размножением.
ПОЛОВОЕ РАЗМНОЖЕНИЕ
Половое размножение встречается как у одноклеточных, так и у многоклеточных растений и животных.
Как отмечено в главах V и XIII, половое размножение у бактерий осуществляется путем конъюгации, служащей аналогом полового процесса и являющейся системой рекомбинации этих организмов, тогда как у простейших половое размножение происходит тоже путем конъюгации либо посредством сингамии и аутогамии.
У многоклеточных организмов (растений и животных) половое размножение связано с образованием зародышевых или половых клеток (гамет), оплодотворением и образованием зигот.
Половое размножение является значительным эволюционным приобретением организмов. С другой стороны, оно способствует пересортировке генов, появлению разнообразия организмов и повышению их конкурентоспособности в непрерывно меняющихся условиях окружающей среды.
У одноклеточных организмов половое размножение существует в нескольких формах. У бактерий половое размножение можно аналогизировать с имеющей место у них конъюгацией, заключающейся в передаче плазмидной или хромосомной ДНК от клеток-доноров (содержащих плазмиды) к клеткам-реципиентам (не содержащих плазмиды), а также с трансдукцией бактерий, заключающейся в передаче генетического материала от одних бактериальных клеток к другим фагам. Конъюгация встречается также у инфузорий, у которых во время этого процесса происходит переход ядер от одних особей к другим, после чего следует деление последних.
У многоклеточных растений и животных половое размножение клеток происходит через образование женских и мужских половых клеток (яйцеклеток и сперматозоидов), последующее оплодотворение яйцеклетки сперматозоидом и образование зиготы. У растений половые клетки продуцируются в специализированных репродуктивных структурах, у животных они продуцируются в половых железах, называемых гонадами (от греч. gone - семя).
Между соматическими и половыми клетками животных существует важное различие. Оно заключается в том, что соматические клетки способны к делению, т. е. репродуцируют себя, и, кроме того, из них образуются половые клетки. Напротив, половые клетки не делятся, но они «начинают» репродукцию целого организма.
Диплоидные соматические клетки, в которых образуются мужские половые клетки, называются сперматогониями, а в которых образуются женские половые клетки - овогониями. Процесс образования (роста и дифференциации) мужских и женских половых клеток носит название гаметогенеза.
Гаметогенез основан на мейозе (от греч. meiosis - понижать), который представляет собой редукционное деление ядер клеток, сопровождающееся понижением числа хромосом на ядро. Мейоз происходит в специализированных клетках репродуктивных органов живых существ, размножающихся половым путем (рис. 17). Например, у папоротникообразных мейоз встречается в специализированных клетках спорангий, располагающихся на нижней поверхности листьев этих растений и развивающихся в споры, а затем в гаметофиты. Последние существуют раздельно, продуцируя в конечном итоге мужские и женские гаметы. У цветковых растений мейоз осуществляется в специализированных клетках семяпочек, которые развиваются в споры. Последние продуцируют гаметофит с одной яйцеклеткой. Кроме того, у этих растений мейоз происходит и в специализированных клетках пыльников, которые также развиваются в споры, продуцирующие в конечном итоге пыльцу с двумя мужскими гаметами. У земляных червей, которые являются гермафродитами и содержат мужские половые органы в одном сегменте тела, а женские в другом и которые характеризу-
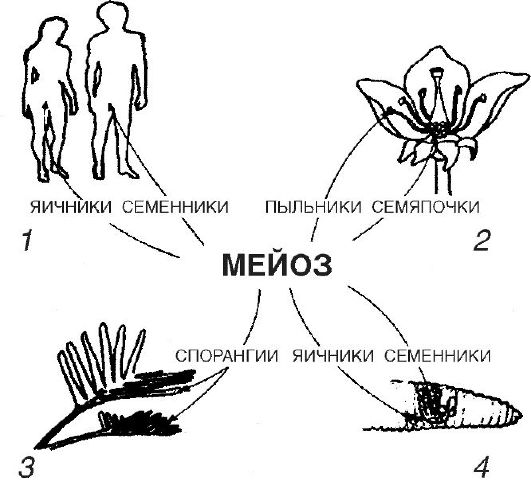
Рис. 17. Мейоз у разных организмов: 1 - человек; 2 - цветковые растения; 3 - папоротник; 4 - дождевой червь
ются способностью к перекрестному оплодотворению между разными особями, имеется способность одновременно к сперматогенезу и овогенезу.
Мейоз осуществляется в специализированных клетках семенников и яичников, в которых продуцируются мужские и женские гаметы соответственно. Выявлены белки - индукторы мейоза.
В процессе мейоза диплоидное число хромосом (2л), которое характерно для соматических клеток (клеточных ядер) и незрелых зародышей клеток, изменяется до гаплоидного числа (n), характерного для зрелых зародышевых клеток. Таким образом, в результате гаметогенеза половые клетки получают лишь половину хромосом соматических клеток (рис. 18).

Рис. 18.
Поведение хромосом в период гаметогенеза у животных является одинаковым как у мужских, так и у женских особей. Однако полы различаются по времени происхождения разных стадий мейоза, что осо-
бенно заметно у человека. У мужских особей в постпубертатный период полный процесс мейоза завершается примерно в течение двух месяцев, в то время как у женских особей первое мейотическое деление начинается в фетальном яичнике и не завершается, пока не начнется овуляция, которая наступает у них примерно в 15 лет.
У высших животных в случае мужских особей мейоз сопровождается образованием четырех функционально активных гамет.
Напротив, у женских особей каждый овоцит II порядка дает лишь одну яйцеклетку. Другие ядерные продукты женского мейоза представляют собой три редукционных тельца, которые не участвуют в размножении и дегенерируют.
Мейоз состоит из двух ядерных делений. Первое меиотическое деление ядра разделяет членов каждой пары гомологичных хромосом после того, как они спарились одна с другой (синапсис) и обменялись генетическим материалом (кроссинговер). В результате этого разделения образуется два гаплоидных ядра. Второе мейотическое деление разделяет две продольные половины хромосом (хроматиды) в каждом из этих ядер, продуцируя четыре гаплоидных ядра.
В процессе гаметогенеза происходит также дифференциация яйцеклеток (овогенез) и сперматозоидов (сперматогенез), являю- щаяся пререквизитом их функций. Яйцеклетки животных намного крупнее сперматозоидов, обычно неподвижны и содержат питательный материал, который обеспечивает развитие эмбриона в начальном периоде после оплодотворения. Сперматозоиды большинства животных обладают жгутиком, обеспечивающим независимость их движения.
Мейоз имеет выдающееся биологическое значение. Благодаря мейозу в клетках организмов поддерживается постоянное число хромосом независимо от количества поколений. Следовательно, мейоз поддерживает постоянство видов. Наконец, в мейозе в результате кроссинговера происходит рекомбинация генов, которая является одним из факторов эволюции.
СПЕРМАТОГЕНЕЗ И ОВОГЕНЕЗ
Сперматогенез - это процесс образования зрелых мужских половых клеток. Сперматозоиды развиваются в мужских половых железах (семенниках, или тестисах) из специализированных соматических клеток (рис. 19). Такими специализированными клетками служат так
называемые примордиальные зародышевые клетки, которые мигрируют к тестисам в раннем периоде эмбриогенеза мужского индивида. Следовательно, примордиальные клетки являются прародителями (предшественниками) зрелых половых клеток.
Семенники человека состоят из многочисленных канальцев, стенки которых сформированы слоями клеток, находящихся на разных стадиях развития сперматозоидов. Наружный слой канальцев образован крупными клетками, называемыми сперматогониями. Эти клетки содержат диплоидный набор хромосом и являются в тестисах потомками примордиальных зародышевых клеток. В период половой зрелости индивида часть сперматогоний перемещается во внутрен- ний слой канальцев, где в результате мейоза они развиваются в клетки, получившие название сперматоцитов I порядка (сперматоциты I), затем в сперматоциты II порядка (сперматоциты II) и, наконец,

Рис. 19. Сперматогенез и овогенез
в сперматиды, являющиеся гаплоидными зародышевыми клетками, дифференцирующимися в конечном итоге в зрелые сперматозоиды. Таким образом, в обобщенном виде можно сказать, что сперматоге- нез инициируется в диплоидных соматических клетках (сперматогониях), после чего следует период созревания зародышевых клеток, в котором происходит два деления ядер путем мейоза, приводящего к образованию сперматид. Такова картина у человека.
Мейоз (рис. 20) в сперматогенезе протекает в несколько стадий (фаз). Между делениями имеются две интерфазы. Таким образом, мейотическое деление можно представить в виде серии следующих одно за другим событий, а именно: интерфаза I -- первое мейотическое деление (ранняя профаза I, поздняя профаза I, метафаза I, анафаза I, телофаза I) - интерфаза II (интерокинез) - второе мейотическое деление (профаза I, метафаза II, анафаза II, телофаза II). Процесс мейоза очень динамичен, поэтому микроскопические различия между разными стадиями отражают скорее не характер самих стадий, а свойства хромосом на разных стадиях. Интерфаза I

Рис. 20. Фазы мейоза: 1 - первое мейотическое деление; 2 - второе мейотическое деление
характеризуется тем, что в ней происходит репликация хромосом (удвоение ДНК), которая к началу ранней профазы I почти полностью завершается.
Первое мейотическое деление начинается в первичном сперматоците и характеризуется длинной профазой, которая состоит из переходящих одна в другую профазы I и профазы II. В профазе I различают пять главных стадий - лептонему, зигонему, пахинему, диплонему и диакинез.
На стадии лептонемы хромосомы в ядре представлены в виде тонких спирализованных нитей, содержащих многочисленные темноокрашенные гранулы (хромомеры). Расщепления хромомер и нитей не отмечают, но считают, что хромосомы на уровне этой стадии являются двойными, т. е. диплоидными. Гомологи каждой хромосомной пары объединяются хромомерами вдоль их длины по принципу застежки.
На стадии зигонемы характерно установление синапсов между гомологичными хромосомами, в результате чего образуются спарен- ные хромосомы (биваленты). Хромосомы X и Y ведут себя по сравнению с аутосомами несколько по-другому. Они конденсируются в темноокрашиваемые гетерохроматиновые тела, спариваемые в результате гомологических районов на их концах.
На стадии пахинемы, которая является наиболее долгой во времени стадией в мейотической профазе, происходит конденсация бивалентов и разделение каждой хроматиды надвое, в результате чего каждый бивалент представляет собой сложную спиральную структуру, состоящую из четырех сестринских хроматид (тетрад). В конце этой стадии начинается разделение спаренных хромосом-бивалентов. Теперь гомологичные хромосомы могут наблюдаться рядом. Поэтому в некоторых препаратах можно видеть четыре хромосомы, которые образуются в результате дупликации каждого гомолога, формирующего сестринские хроматиды. На этой стадии осуществляются обмены между гомологами и формирование хиазм.
На стадии диплонемы происходит укорачивание, утолщение и взаимное отталкивание сестринских хроматид, в результате чего хроматиды в биваленте почти разъединены. Разделение считается неполным по той причине, что в каждой паре хромосом еще не расщеплена центромера. Что касается бивалентов, то они удерживаются на различных местах вдоль их длины с помощью хиазм, которые являются структурами, сформированными между гомологичными хроматидами в результате
предыдущего кроссинговера между синаптически связанными гомологами. В хороших препаратах можно наблюдать от одной до нескольких хиазм в зависимости от длины бивалента. Каждая наблюдаемая на этой стадии хиазма представляет собой результат обмена, который встречался между несестринскими хроматидами в течение стадии пахинемы. Поскольку сжатие и отталкивание бивалентов усиливается, хиазмы двигаются к концам хромосом, т. е. происходит терминализация хромосом. В конце диплонемы наступает деспирализация хромосом; гомологи продолжают отталкиваться друг от друга.
На стадии диакинеза, которая сходна с диплонемой, продолжается укорочение бивалентов и наступает ослабление (уменьшение) хиазм, вследствие чего формируются дискретные единицы в виде хроматид (четыре). Непосредственно после завершения этой стадии происходит растворение ядерной мембраны.
В метафазе I биваленты достигают наибольшей концентрации. Становясь овальными, они располагаются в экваториальной части ядра, где формируют экваториальные пластинки мейотической метафазы I. Форма каждого бивалента определяется числом и локализацией хиазм. У мужчин число хиазм на бивалент в метафазе I составляет обычно 1-5. Бивалент XY становится палочковидным в результате одиночной терминально расположенной хиазмы.
В анафазе I начинается движение противоположных центромер к противоположным полюсам клетки. В результате этого происходит разделение гомологичных хромосом. Каждая хромосома состоит теперь из двух хроматид, удерживаемых центромерой, которая не делится и остается интактной. Этим анафаза I мейоза отличается от анафазы митоза, при котором центромера подвергается разделению. Важно заметить, что благодаря кроссинговеру каждая хроматида является генетически различной.
В стадии телофазы I хромосомы достигают полюсов, чем заканчивается первое мейотическое деление. После телофазы I наступает короткая интерфаза (интеркинез), в которой хромосомы деспи- рализуются и становятся диффузными, или телофаза I переходит прямо в профазу II второго меиотического деления. Ни в одном, ни в другом случае репликации ДНК не отмечается. После первого мейотического деления клетки называют сперматоцитами II порядка. Количество хромосом в каждой такой клетке снижается от 2n до 1n, но содержание ДНК еще не изменяется.
Второе мейотическое деление осуществляется в течение нескольких фаз (профаза II, метафаза II, анафаза II, телофаза II) и сходно с митотическим делением. В профазе II хромосомы вторичных спер- матоцитов остаются у полюсов. В метафазе II центромера каждой из двойных хромосом делится, обеспечивая каждую новую хромосому собственной центромерой. Начинается формирование веретена, к полюсу которого двигаются новые хромосомы. В телофазе II второе мейотическое деление заканчивается, в результате чего каждый сперматоцит II порядка дает два сперматида, из которых дифферен- цируются затем сперматозоиды. Как и во вторичном сперматоците, число хромосом в сперматиде является гаплоидным (1n). Однако хромосомы сперматид являются одиночными, тогда как хромосомы вторичных сперматоцитов II - двойными, будучи построенными из двух хроматид. Следовательно, ядро каждого сперматида имеет одиночный набор негомологичных хромосом. Вторичное мейотическое деление - это деление митотического типа (экваториальным делением). Оно разделяет двойные сестринские хроматиды и отличается от редукционного деления, в котором гомологичные хромосомы разделены. Единственное существенное отличие от классического митоза заключается в том, что здесь имеется гаплоидный набор хромосом.
Итак, первое мейотическое деление сперматоцитов I порядка приводит к образованию двух вторичных сперматоцитов (II порядка). Обе хроматиды структур, образующихся в результате редукционного деления, являются сестринскими хроматидами. Последние возникают в результате репликации, предшествующей первому мейотическому делению. Второе мейотическое деление каждого вторичного сперматоцита приводит к образованию четырех сперматидов. Таким образом, в типичном мейозе клетки делятся дважды, тогда как хромосомы - только один раз (рис. 21).
Конечный этап в сперматогенезе связан с дифференциацией, которая заканчивается тем, что каждый из сравнительно больших, сферически неподвижных сперматидов превращается в небольшой вытянутый подвижный сперматозоид.
У большинства взрослых (сексуально зрелых) мужских особей животных сперматогенез происходит в семенниках постоянно или периодически (сезонно). Например, у насекомых для завершения цикла сперматогенеза требуется лишь несколько дней, тогда как у млекопитающих этот цикл затягивается на недели и даже

Рис. 21. Распределение хромосом при гаметогенезе
месяцы. У взрослого человека сперматогенез проходит в течение всего года. Время развития примитивных сперматогоний в зрелые сперматозоиды составляет около 74 дней.
Мужские половые клетки, продуцируемые организмами разной видовой принадлежности, характеризуются подвижностью и чрезвычайным разнообразием в размерах и по структуре (рис. 22). Каждый сперматозоид человека состоит из трех отделов - головки, средней части и хвоста (рис. 23). В головке сперматозоида располагается ядро, которое содержит гаплоидный набор хромосом.
Головка снабжена акросомой, которая содержит литические ферменты, необходимые содержимому сперматозоиду для вступления в яйцеклетку. В головке локализуются также две центриоли - прок-

Рис. 22. Формы сперматозоидов

Рис. 23. Строение сперматозоида: А - светомикроскопическое изображение в разных плоскостях: 1 - головка, 2 - средняя часть, 3 - хвост; Б - схематическая реконструкция электронномикроскопического изображения: 1 - ядро, 2 - акросома, 3 - центросома (проксимальная центриоль), 4 - центральное кольцо, 5 - митохондриальная спираль,
6 - осевая нить
симальная, которая побуждает деление оплодотворенной сперматозоидом яйцеклетки, и дистальная, которая дает начало аксимальному стержню хвоста. В средней части сперматозоида располагаются базальное тельце хвоста и митохондрии. Хвост (отросток) сперматозоида образован внутренним аксимальным стержнем и внешним футляром, имеющим цитоплазматическое происхождение. Сперматозоиды человека характеризуются значительной подвижностью.
ОВОГЕНЕЗ И ЯЙЦЕКЛЕТКИ
Процесс формирования яйцеклеток носит название овогенеза. Его функциями являются обеспечение гаплоидного набора хромосом в ядре яйцеклетки и питательных потребностей зиготы. Овогенез в своем проявлении в основном сопоставим со сперматогенезом.
У млекопитающих и человека овогенез начинается еще во внутриутробном периоде (до рождения). Овогонии, представляющие собой мелкие клетки с довольно крупным ядром и локализованные в фолликулах яичников, начинают дифференцироваться в первичные овоциты. Последние формируются уже на третьем месяце внутриутробного развития, после чего они вступают в профазу первого мейотического деления. Ко времени рождения девочки все первичные овоциты уже находятся в профазе первого мейотического деле- ния. Первичные овоциты остаются в профазе вплоть до наступления половой зрелости женского индивида. В одном овоците содержится до 100 000 митоходрий. Когда при наступлении половой зрелости фолликулы яичника созревают, мейотическая профаза в первичных овоцитах возобновляется. Первое мейотическое деление для каждой развивающейся яйцеклетки завершается вскоре перед временем овуляции этой яйцеклетки. В результате первого мейотического деления и неравномерного распределения цитоплазмы одна образовавшаяся клетка становится вторичным овоцитом, другая - полярным (редукционным) тельцем.
Вторичное мейотическое деление наступает, когда вторичный овоцит (развивающаяся яйцеклетка) проходит из яичника в фаллопиеву трубу. Однако это деление не завершается до тех пор, пока во вто- ричный овоцит не проникнет сперматозоид, что происходит обычно в фаллопиевой трубе. Когда сперматозоид проникает во вторичный овоцит, то последний делится, в результате чего образуется овотида (зрелая яйцеклетка) с пронуклеоусом, содержащим одиночный набор
из 23 материнских хромосом. Другая клетка, образующаяся в результате этого деления, является вторым полярным тельцем, не способным к дальнейшему развитию. В это время подвергается делению надвое и полярное (редукционное) тельце. Таким образом, развитие одного овоцита I порядка сопровождается образованием одной овотиды и трех редукционных телец. В яичниках таким путем на протяжении жизни обычно созревает 300-400 овоцитов, но в месяц созревает лишь один овоцит. В течение дифференциации яйцеклеток формируются мембраны, уменьшается в размере их ядро.
У некоторых видов животных овогенез протекает быстро и непрерывно и приводит к образованию большого количества яйцеклеток.
Вопреки сходству со сперматогенезом овогенез характеризуется некоторыми специфическими особенностями. Питательный материал (желток) первичного овоцита не распределяется поровну между четырьмя клетками, которые образуются в результате мейотических делений. Основное количество желтка сохраняется в одной большой клетке, тогда как полярные тельца содержат очень мало этого вещества. Первые и вторые полярные тельца получают в результате делений те же самые хромосомные наборы, что и вторичные овоциты, но они не становятся половыми клетками. Поэтому яйцеклетки намного богаче питательным материалом по сравнению со сперматозоидами. Особенно сильно это различие проявляется в случае яйцекладущих животных.
Яйцеклетки млекопитающих имеют овальную или несколько вытянутую форму (рис. 24) и характеризуются типичными чертами клеточного строения. Они содержат все структуры, характерные для соматических клеток, однако внутриклеточная организация яйцеклетки очень специфична и определяется тем, что яйцеклетка является также и средой, которая обеспечивает развитие зиготы. Одна из характерных особенностей яйцеклеток заключается в сложности строения их оболочек. У очень многих животных различают первичную, вторичную и третичную оболочки яйцеклеток. Первичная оболочка (внутренняя) формируется еще на стадии овоцита. Представляя собой поверхностный слой овоцита, она имеет сложную структуру, так как пронизана выростами прилегающих к ней фолликулярных клеток. Вторичная (средняя) оболочка полностью сформирована фолликулярными клетками, а третичная (наружная) образована веществами, представляющими собой продукты секреции желез яйцеводов, через которые проходят яйцеклетки. У птиц, например,
третичными оболочками яйцеклеток служат белковая, подскорлуповая и скорлуповая оболочки. Для яйцеклеток млекопитающих характерно наличие двух оболочек. Структура внутриклеточных компонентов яйцеклеток специфична в видовом отношении, а иногда имеет даже индивидуальные особенности.

Рис. 24. Строение яйцеклетки: 1 - фолликулярные клетки; 2 - оболочка; 3 - цитоплазма; 4 - ядро
ОПЛОДОТВОРЕНИЕ
Оплодотворение - это процесс объединения мужской и женской гамет, который приводит к формированию зиготы и последующему развитию нового организма. В процессе оплодотворения происходит установление диплоидного набора хромосом в зиготе, что определяет выдающееся биологическое значение этого процесса.
В зависимости от видовой принадлежности организмов, размножающихся половым путем, различают наружное и внутреннее оплодотворение. Наружное оплодотворение происходит в окружающей среде, в которую поступают мужские и женские половые клетки. Например, наружным является оплодотворение у рыб. Выделяемые ими мужские (молока) и женские (икра) половые клетки поступают в воду, где и происходит их встреча и объединение.
Внутреннее оплодотворение обеспечивается переносом сперматозоидов из мужского организма в женский. Такое оплодотворение встречается у млекопитающих, центральным моментом которого является слияние ядер половых клеток. Считают, что в яйцеклетку проникает содержимое одного сперматозоида. В механизмах опло-
дотворения многое еще остается неясным. Данные об оплодотворении у морских ежей свидетельствуют о том, что уже через 2 секунды после соприкосновения сперматозоидов и яйцеклетки наступают изменения в электрических свойствах плазменной мембраны последней. Слияние гамет наступает через 7 секунд. Предполагают, что проникновение в яйцеклетку содержимого лишь одного из множества сперматозоидов объясняется изменениями электрических свойств ее плазменной мембраны. Оплодотворенная яйцеклетка дает начало зиготе. По вопросу о причинах активации сперматозоидом метаболизма яйцеклетки существует два мнения. Одни считают, что связывание сперматозоида с внешними рецепторами на поверхности клеток представляет собой сигнап, который через мембрану поступает вовнутрь яйцеклетки и активизирует там инозитолтрифосфат и ионы кальция. Другие полагают, что сперматозоиды содержат специальный инициирующий фактор.
Экспериментальные разработки, выполненные в последние годы, показали, что оплодотворение яйцеклеток млекопитающих, включая человека, возможно и в пробирке. Болеее того, зародыши, развившиеся в пробирке, могут быть имплантированы в матку женщины, где они подвергаются дальнейшему нормальному развитию. Известны многочисленные случаи рождения «пробирочных» детей.
В отличие от животных, размножающихся путем зигогенеза, многие организмы способны к размножению путем партеногенеза (от греч. parthenos - девственница и genos - рождение), под которым понимают размножение организмов из неоплодотворенной яйцеклетки. Различают облигатный и факультативный партеногенез. Облигатный партеногенез стал главным способом размножения организмов отдельных видов, например кавказской скальной ящерицы. Животные этого вида являются только женскими особями. Напротив, факультативный партеногенез заключается в том, что яйцеклетки способны развиваться как без оплодотворения, так и после оплодотворения. Факультативный партеногенез, в свою очередь, бывает женским и мужским. Женский партеногенез часто наблюдается у пчел, муравьев, коловраток, у которых из неоплодотворенных яйцеклеток развиваются самцы. Мужской партеногенез встречается у некоторых изогамных водорослей.
Партеногенез бывает как естественным, так и искусственным (индуцированным). Механизм искусственного партеногенеза заключается в раздражении яйцеклеток с помощью физических или хими-
ческих факторов, что ведет к активации яйцеклеток и, как следствие, к развитию неоплодотворенных яиц. Искусственный партеногенез наблюдали в случае животных многих систематических групп - иглокожих, червей, моллюсков и даже млекопитающих.
Известна форма партеногенеза, получившая название андрогенеза (от греч. andros - мужчина, genesis - зарождение). Если в яйцеклетке инактивировать ядро и после этого в нее проникнет несколько сперматозоидов, то из такой яйцеклетки в результате слияния мужских (сперматозоидных) ядер развивается мужской организм.
Известны случаи, когда партеногенез проявляется циклически, в зависимости от сезона. Например, коловратки, дафнии и тли летом размножаются путем партеногенеза, осенью - оплодотворения яйце- клеток и образования зигот, т. е. путем зигогенеза.
Роль партеногенеза и его форм в природе невелика, так как он не обеспечивает широких адаптивных возможностей организмов.
В отличие от зигогенеза и партеногенеза существует гиногенез, представляющий собой псевдогамию, когда сперматозоид вступает в яйцеклетку и активизирует ее, но ядро сперматозоида не сливается с ядром яйцеклетки. В этом случае появляющееся потомство состоит только из женских особей. Гиногенез встречается в естественных условиях у нематод и рыб, но его можно вызвать и искусственно. В частности, описаны случаи искусственного гиногенеза у тутового шелкопряда, рыб и амфибий.
ЧЕРЕДОВАНИЕ ГАПЛОИДИИ И ДИПЛОИДИИ.
ЧЕРЕДОВАНИЕ ПОКОЛЕНИЙ
Организмам, размножающимся половым путем, свойственно чередование гаплоидной и диплоидной фаз в их развитии. У многих организмов, включая млекопитающих, это чередование имеет регулярный характер, и на нем основано сохранение видовых признаков организмов.
Для многих организмов характерна также смена поколений, когда поколения особей, размножающихся бесполым путем, сменяются поколениями особей, размножающихся половым путем с образованием гамет. Это получило название первичной смены поколений. Она встречается у споровиков, жгутиконосцев и многих растений. Первичная смена поколений имеет регулярный характер, и ее нали-
чие в природе свидетельствует о сохранении в филогенезе многих организмов как бесполого, так и полового размножения. У других организмов встречается чередование полового размножения с парте- ногенезом. Это называют вторичной сменой поколений. Например, у трематод половое размножение регулярно сменяется партеногенезом. Такое вторичное чередование поколений называют гетерогонией. У кишечнополостных на некоторых стадиях развития осуществляется переход от полового размножения к бесполому (вегетативному). Эту форму вторичного чередования поколений называют метагенезом.
ПОЛОВОЙ ДИМОРФИЗМ. ГЕРМАФРОДИТИЗМ
Для мужских и женских особей характерны специфические фенотипические черты. Различия между самками и самцами по их свойствам называют половым диморфизмом. У животных он встречается уже на низших ступенях эволюционного развития, например у круглых гельминтов, членистоногих, и достигает наибольшего выражения у позвоночных животных.
Если мужские и женские половые клетки продуцируются одной и той же особью, имеющей как мужские, так и женские половые железы, то это явление носит название истинного гермафродитизма. Он встречается у плоских червей, кольчатых червей, моллюсков. У плоских червей мужские и женские половые железы функционируют на протяжении всей жизни особей. Напротив, у моллюсков половые железы продуцируют яйцеклетки и сперматозоиды попеременно.
Истинный гермафродитизм встречается и у человека, являясь результатом нарушения развития. Генотипами гермафродитов являются 46 XX или 46 XY, причем большинство случаев относится к XX (около 60%). Генотип XX чаще всего встречается у гермафродитов негроидных афри- канских популяций, тогда как XY в большинстве случаев - среди японцев. У гермафродитов обоих типов отмечена тенденция в сторону билатеральной асимметрии гонад. Среди истинных гермафродитов встречаются также хромосомные мозаики, которые в одних соматических клетках имеют пару хромосом XX, в других - пару XY.
Известен и ложный гермафродитизм, когда индивиды обладают наружными половыми органами и вторичными половыми признака- ми, характерными для обоих полов, но продуцируют лишь половые клетки одного типа - мужские или женские.
ПРОИСХОЖДЕНИЕ СПОСОБОВ РАЗМНОЖЕНИЯ
Предполагают, что наиболее древним является бесполое размножение, в частности вегетативное размножение. Из последнего развилось размножение спорообразованием, несомненное преимущество которого заключается в том, что оно обеспечивает лучшие возможности в сохранении видов и особенно в их расселении.
Половое размножение - наиболее эффективный путь воспроизводства организмов. Предполагают, что оно развилось из бесполого, возникнув около 1 млрд лет назад, причем первые этапы были связаны с усложнением в развитии гамет. Примитивные гаметы характеризовались недостаточной морфологической дифференцировкой, в результате чего для многих организмов ведущей была изогамия (от греч. isos - равный, gamos - брак), когда половые клетки были подвижными изогаметами, не дифференцированными на мужские и женские формы.
В последующем получила развитие анизогамия (от греч. anisos - неравный, gamos - брак), характеризующаяся наличием дифференцированных гамет, различающихся лишь величиной. На более поздних этапах эволюции возникли резкие различия в подвижности, форме и размерах гамет. В процессе эволюции у позвоночных выработался также ряд добавочных приспособлений, облегчающих перенос спермы самца в половые пути самки и создающих условия для развития оплодотворенной яйцеклетки. Эти приспособления в ходе эволюции развились из выделительной системы, что привело к формированию мочеполовой системы.
Диплоидное состояние предоставляет чрезвычайные преимущества для организмов, поскольку в этом состоянии происходит накопление различных аллелей. Поэтому половое размножение имеет еще и то преимущество, что оно предоставляет организмам б о льшую возможность изменчивости по сравнению с бесполым, а это играет важнейшую роль в эволюции.
Явные репродуктивные преимущества имеет партеногенез, так как он продуцирует лишь потомство женского рода. Однако он редок. Для объяснения низкой частоты замещения партеногенезом полового размножения в естественных популяциях организмов используют две гипотезы. В соответствии с одной из них (мутационно-аккумулятивной) пол является адаптивным приспособлением, поскольку он «очищает» геном от повторяющихся во времени мутаций, тогда как по другой гипотезе (экологической) пол является
Первые представления о росте и развитии восходят к античному миру. Еще Гиппократ (460-377 гг. до н. э.) предполагал, что яйцеклетки уже содержат полностью сформированный организм, но в очень уменьшенном виде. Это представление затем было развито в учение о преформизме (от лат. preformatio - предобразование), которое особенно популярным оказалось в XVII-XVIII вв. Сторонниками преформизма были Гарвей, Мальпиги и многие другие видные биологи и медики того времени. Для преформистов спорный вопрос заключался лишь в том, в каких половых клетках преформирован орга- низм - женских или мужских. Тех, кто отдавал предпочтение яйцеклеткам, называли овистами, а тех, кто большее значение придавал мужским половым клеткам, - анималькулистами. Преформизм - это метафизическое учение от начала и до конца, ибо оно отрицало развитие. Решающий удар преформизму нанес Ш. Боннэ (1720-1793), который открыл в 1745 г. партеногенез на примере развития тлей из неоплодотворенных яиц. После этого преформизм уже не мог оправиться и стал терять свое значение.
В античном мире возникло и другое учение, противоположное преформизму и получившее впоследствии название эпигенеза (от греч. epi - после, genesis - развитие). Как и преформизм, эпи- генез большое распространение получил также в XVII-XVIII вв. В распространении эпигенеза большое значение имели взгляды К.Ф. Вольфа (1733-1794), обобщенные в его книге «Теория развития» (1759). К.Ф. Вольф считал, что в яйце нет ни преформированного организма, ни его частей и что оно состоит из первона- чально однородной массы. В отличие от преформистов взгляды К.Ф. Вольфа и других сторонников эпигенеза были прогрессивны применительно для своего времени, так как содержали мысль о развитии. Однако в дальнейшем появились новые моменты. В частности, в 1828 г. К. Бэр опубликовал свой труд «История развития животных», в котором показал, что содержимое яйца неоднородно, т. е. структурировано, причем степень структурированности возрастает по мере развития зародыша. Таким образом, К. Бэр показал несостоятельность как преформизма, так и эпигенеза.
Рост и развитие - важнейшие свойства живого. Рост - это повышение массы всего организма в результате увеличения количества клеток, тогда как развитие - это качественные изменения организма, которые определяются дифференциацией клеток и морфогенезом, обеспечивающими прогрессивные изменения индивидуумов, начиная с яйцеклеток и заканчивая их взрослым состоянием.
Онтогенез (от греч. ontos - существо, genesis - развитие) - это история (цикл) развития индивидуума, начинающаяся с обра- зования давших ему начало половых клеток и заканчивающаяся его смертью. Представления об онтогенезе основаны на данных о росте, развитии и дифференцировке. Изучение фундаментальных основ онтогенеза имеет важное значение для понимания биологии человека.
Современные данные о развитии организмов отвергают как преформизм, так и эпигенез. В рамках современных представлений развитие организма понимают в качестве процесса, при котором структуры, образовавшиеся ранее, побуждают развитие последующих структур. Процесс развития детерминирован генетически и теснейшим образом связан со средой. Следовательно, развитие определяется единством внутренних и внешних факторов.
ЕДИНСТВО РОСТА И РАЗВИТИЯ
Рост организма - это постепенное возрастание его массы и изменение формы в результате увеличения количества клеток и их дифференцировки, формирования тканей и органов, биохимических изменений в клетках и тканях. Таким образом, рост является результатом количественных изменений в виде увеличения количества клеток (массы тела) и качественных в виде дифференцировки клеток и морфогенеза. Дифференциация клеток - это процесс, благодаря которому одни клетки становятся морфологически, биохимически и функционально отличающимися от других клеток. Размножение и дифференцировка одних клеток всегда координированы с ростом и дифференцировкой других. Оба эти процесса происходят на протяжении всего жизненного цикла организма. Поскольку дифференцирующиеся клетки изменяют свою форму, а в изменения формы вовлекаются группы клеток, то это сопровождается морфогенезом, представляющим собой совокупность процессов, определяющих структурную организацию клеток и тканей, а также общую морфологию организмов.
Рост можно измерить, построив на основе результатов измерений кривые размеров организма, массы, сухой массы, количества клеток, содержания азота и других показателей.
ОНТОГЕНЕЗ И ЕГО ТИПЫ. ПЕРИОДИЗАЦИЯ ОНТОГЕНЕЗА
Онтогенез в зависимости от характера развития организмов типируют на прямой и непрямой, в связи с чем различают прямое и непрямое развитие. Прямое развитие организмов в природе встречается в виде неличиночного и внутриутробного развития, тогда как непрямое развитие наблюдается в форме личиночного развития. В противоположность онтогенезу видовой категорией является филогенез.
Личиночное развитие. Под этим развитием понимают непрямое развитие, поскольку организмы в своем развитии имеют одну или несколько личиночных стадий. Личиночное развитие характерно для насекомых, амфибий, иглокожих. Личинки этих животных ведут самостоятельный образ жизни, подвергаясь затем превращениям. Поэтому это развитие называют еще развитием с метаморфозами (см. ниже).
Неличиночное развитие. Такая форма развития характерна для организмов, развивающихся прямым образом, например для рыб, пресмыкающихся и птиц, яйца которых богаты желтком (питательным материалом). Благодаря этому в яйцах, откладываемых во внешнюю среду, проходит значительная часть онтогенеза. Метаболизм зародышей обеспечивается развивающимися провизорными органами, представляющими собой зародышевые оболочки (желточный мешок, амнион, аллантоис).
Внутриутробное развитие. Это развитие также характерно для организмов, развивающихся прямым путем, например для млекопитающих, включая человека. Поскольку яйцеклетки этих организмов очень бедны питательными веществами, то все жизненные функции зародышей обеспечиваются материнским организмом посредством образования из тканей матери и зародыша провизорных органов, среди которых главным является плацента. Эволюционно внутриутробное развитие - самая поздняя форма, однако оно наиболее выгодно для зародышей, так как эффективно обеспечивает их выживание.
Онтогенез подразделяют на проэмбриональный, эмбриональный и постэмбриональный периоды. В случае человека период развития до рождения называют пренатальным или антенатальным, после рождения - постнатальным. Развивающийся зародыш до образования зачатков органов называют эмбрионом, после образования зачатков органов - плодом.
ПРОЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
Этот период в индивидуальном развитии организмов связан с образованием гамет в процессе гаметогенеза. Мужские половые клетки не имеют существенных отличий от других клеток, тогда как яйцеклетки отличаются тем, что они содержат очень много желтка. Учитывая количество желтка и распространение его в яйцеклетках, последние классифицируют на три типа:
1) изолециталъные яйцеклетки, содержащие немного желтка, который локализован равномерно по всей клетке. Эти яйцеклетки продуцируются иглокожими (морскими ежами), низшими хордовыми (ланцетниками), млекопитающими;
2) телолецитальные яйцеклетки содержат большое количество желтка, который сосредоточен на одном из полюсов - вегетативном. Такие яйцеклетки продуцируются моллюсками, земноводными, реп- тилиями, птицами. Например, яйцеклетки лягушки состоят на 50% из желтка, яйцеклетки кур (в обиходе куриные яйца) - на 95%. На другом полюсе (анимальном) телолецитальных яйцеклеток сосредоточены цитоплазма и ядро;
3) центролецитальные яйцеклетки, в которых желтка немного и он занимает центральное положение. На периферии таких яйцеклеток располагается цитоплазма. Центролецитальные яйцеклетки проду- цируются членистоногими.
Для проэмбрионального периода характерно также то, что в этот период в гаметах происходят метаболические процессы, связанные с накоплением молекул ДНК.
ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
Эмбриогенез (от греч. етbryоп - зародыш), или эмбриональный период, начинается со слияния мужских и женских половых клеток, которые представляют собой процесс оплодотворения яйцеклеток.
У организмов, для которых характерно внутриутробное развитие, эмбриональный период заканчивается рождением, а у организмов, которым свойственны личиночный и неличиночный типы рзвития, эмбриональный период завершается выходом организма из яйцевых или зародышевых оболочек соответственно. В пределах эмбрионального периода различают стадии зиготы, дробления, бластулы, образования зародышевых листков, гистогенеза и органогенеза.
Зигота. Оплодотворение заключается в серии процессов, в которых мужская половая клетка инициирует развитие яйцеклетки. В активированной мужской гаметой яйцеклетке происходит ряд физических и химических процессов, включая повышенный синтез белков. Перемещение протоплазмы ведет к установлению билатериальной симметрии яйцеклетки. Ядра сливаются, восстанавливается диплоидный набор хромосом. Таким образом создается одноклеточный организм.
Дробление. Представляет собой начальный период развития зиготы (оплодотворенного яйца), которое заключается в делении зиготы путем митоза. Деление начинается с появления на поверхности яйцеклетки борозды. Первая борозда приводит к образованию двух клеток - двух бластомеров, вторая - четырех бластомеров, третья - восьми бластомеров (рис. 25). Группа клеток, образованная в результате последовательных дроблений, получила название морулы (от лат. morum - тутовая ягода).
Биологическое значение этой стадии заключается в том, что из крупной клетки, являющейся яйцеклеткой, образуются более мелкие клетки, в которых уменьшено отношение цитоплазмы к ядру.
Дробление зиготы завершается образованием многоклеточной структуры, получившей название бластулы (от греч. blastos - росток). Эта структура имеет форму пузырька, называемого бластодермой и состоящего из одного слоя клеток. Теперь эти клетки называют эмбриональными. По размерам бластула сходна с яйцеклеткой. В период дробления увеличивается количество ядер и общее число ДНК. Синтезируется также небольшое количество мРНК и тРНК, тогда как рибосомная РНК еще не обнаруживается.
Стадию бластулы проходят все животные, но в каждом случае есть особенности. У млекопитающих деление идет неравномерно, поэтому морулы состоят из разного количества клеток. Кроме того, из части клеток образуется структура, называемая трофобластом,

Бластоцель Личинка Головастик
форма
Рис. 25. Дробление зигот и образование бластул у разных организмов: 1 - исходная яйцеклетка; 2 - два бластомера; 3 - четыре бластомера; 4 - восемь бластомеров; 5 - бластула; 6 - взрослая форма
клетки которого питают зародыш и благодаря ферментам обеспечивают внедрение последнего в стенку матки. Позднее клетки трофобласта отслаиваются от зародыша и образуют пузырек, который заполняется жидкостью тканей матки.
Биологическое значение этой стадии заключается в том, что из крупной клетки, каковой является яйцеклетка, образуются более мелкие клетки, в которых уменьшено отношение цитоплазмы к ядру и ядро имеет новое цитоплазматическое окружение.
Гаструляция (от греч. gastre - полость сосуда). Это следующий за образованием бластулы процесс движения эмбриональных клеток, который сопровождается формированием двух или трех (в зависимости от вида животных) слоев зародыша, или так называемых зародышевых листков (рис. 26).
Развитие (гаструляция) изолецитальных яиц происходит путем инвагинации (впячивания) вегетативного полюса вовнутрь бластулы, в результате чего противоположные полюса почти сливаются, а бластоцель (полость бластулы) почти либо полностью исчезает. Внешний
Рис. 26. Гаструляция у различных организмов: 1 - бластоцель; 2 - первичная мезенхима; 3, 4, 5 - первичная кишка
слой клеток зародыша получил название эктодермы (от греч. ectos - снаружи, derma - кожа), или наружного зародышевого листка, тогда как внутренний - энтодермы (от греч. entos - внутри), или внутреннего зародышевого листка. Образующаяся при этом полость получила название гастроцеля, или первичной кишки, вход в которую называют бластопором (первичным ртом).
Развитие двух зародышевых листков характерно для губок и кишечнополостных. Однако хордовым в период гаструляции присуще развитие третьего зародышевого листка - мезодермы (от греч. mesos - средний), образующегося между эктодермой и энтодермой.
Гаструляция является необходимым пререквизитом для последующих стадий развития, поскольку она приводит клетки в положение, открывающее возможность формировать органы. Дифференцированный на три эмбриональных закладки зародышевый материал дает начало всем тканям и органам развивающегося зародыша.
Развитие (дифференцировка) зародышевых листков сопровождается тем, что из них формируются различные ткани и органы. В частности, из эктодермы развиваются эпидермис кожи, ногти и волосы, сальные и потовые железы, нервная система (головной мозг, спинной мозг, ганглии, нервы), рецепторные клетки органов чувств, хрусталик глаза, эпителий рта, носовой полости и анального отверстия, зуб-
ная эмаль. Из энтодермы развиваются эпителий пищевода, желудка, кишок, желчного пузыря, трахеи, бронхов, легких, мочеиспускательного канала, а также печень, поджелудочная железа, щитовидная, паращитовидные и зобная железы. Из мезодермы развиваются гладкая мускулатура, скелетные и сердечные мышцы, дерма, соединительная ткань, кости и хрящи, дентин зубов, кровь и кровеносные сосуды, брыжейка, почки, семенники и яичники. У человека первыми обособляются головной и спинной мозг. Через 2 месяца появляются почти все структуры тела. Связь зародыша со средой осуществляется через провизорные органы. Органогенез заканчивается к концу эмбрионального периода. Если дефинитивное ротовое отверстие образуется на месте первичного рта (бластопора), то этих животных называют первичноротыми (черви, моллюски, членистоногие).
Если же дефинитивный рот образуется в противоположном месте, то этих животных называют вторичноротыми (иглокожие, хордовые).
Для обеспечения связи зародыша со средой служат так называемые провизорные органы, которые существуют временно. В зависимости от типа яйцеклеток провизорными органами являются разные структуры. У рыб, рептилий и птиц к провизорным органам относится желточный мешок. У млекопитающих желточный мешок закладывается в начале эмбриогенеза, но не развивается. Позднее он редуцируется. Наружная оболочка эмбриона называется хорионом. Она врастает в матку. Место наибольшего врастания в матку называют плацентой. Зародыш с плацентой связан через пуповину, или пупочный кантик, в котором имеются кровеносные сосуды, обеспечивающие плацентарное кровообращение. Метаболизм плода обеспечивается через плаценту.
В основе формообразующего взаимодействия частей эмбриона лежат определенным образом скоординированные процессы обмена веществ. Закономерностью развития является гетерохронность, под которой понимают разное во времени образование закладок органов и различную интенсивность их развития. Быстрее развиваются те органы и системы, которые должны раньше начать функционировать. Например, у человека зачатки верхних конечностей развиваются быстрее, чем нижних.
Зародыши чрезвычайно чувствительны к разным воздействиям. Поэтому различают критические периоды, т. е. периоды, когда зародыши наиболее чувствительны к повреждающим факторам. В случае человека критическими периодами эмбрионального онтогенеза
являются первые дни после оплодотворения, время образования плаценты и роды.
Ядра соматических клеток способны обеспечивать нормальное развитие яйцеклеток, что было выяснено в экспериментах по пересадке ядер соматических клеток в яйцеклетки, лишенные ядер.
Эксперименты также показали, что трансформация отдельного бластомера из 8- и 16-клеточных эмбрионов овец одной породы в безъядерную половину яйцеклетки (после рассечения последней пополам) другой породы сопровождалась развитием жизнеспособных эмбрионов и рождением ягнят.
Со времен Гиппократа (V в. до н. э.) обсуждается вопрос о причинах, которые инициируют рождение плода. В частности, сам Гиппократ предполагал, что развитие плода инициирует собственное рождение. Новейшие экспериментальные работы английских исследователей, выполненные на овцах, показали, что у овец инициация окотов контролируется комплексом гипоталамус + гипофиз + надпочечники плодов. Повреждение ядер гипоталамуса, удаление передней доли гипофиза или надпочечников пролонгирует беременности овец. Напротив, введение овцам аденокортикотропного гормона (секрета передней доли гипофиза) или кортизола (секрета надпочечников) сокращает сроки их беременностей.
Итак, в процессе развития высших эукариотов одиночная оплодотворенная клетка-зигота в ходе дальнейшего развития в результате митоза дает начало клеткам разных типов - эпителиальным, нервным, костным, клеткам крови и другим, которые характеризуются разнообразием морфологии и макромолекулярного состава. Однако для клеток разных типов характерно и то, что они содержат одинаковые наборы генов, но являются высокоспециализированными, выполняя лишь одну или несколько специфических функций, т. е. одни гены «работают» в клетках, а другие неактивны. Например,

Рис. 27. Развитие монозиготных близнецов
только эритроциты специфичны в синтезе и хранении гемоглобина. Точно так лишь клетки эпидермиса синтезируют кератин. Поэтому давно возникли вопросы о генетической идентичности ядер соматических клеток и о контрольных механизмах развития оплодотворенных яйцеклеток как пререквизита в познании механизмов, лежащих в основе дифференцировки клеток. В дифференциации клеток имеют большое значение стволовые клетки (см. ниже). В экспериментах на мышах показано, что клетки одного типа способны к конверсии в клетки другого типа. В частности, показано, что дифференцирующиеся или дифференцированные клетки печени конвертируются в клетки поджелудочной железы.
Начиная с 50-х гг. XX в. во многих лабораториях были выполнены эксперименты по успешной пересадке ядер соматических клеток в яйцеклетки, искусственно лишенные собственных ядер. Исследование ДНК из ядер разных дифференцированных клеток показало, что почти во всех случаях геномы содержат одинаковые наборы последовательностей нуклеотидных пар. Известны исключения, когда эритроциты млекопитающих теряют свои ядра в течение последней стадии дифференцировки. Но к этому времени пулы стой-
ких гемоглобиновых мРНК уже синтезированы, так что ядра больше не нужны эритроцитам. Другими примерами служат гены иммуно- глобулинов и Т-клеток, модифицируемые в ходе развития.
Одним из крупных этапов в направлении познания контрольных механизмов эмбрионального онтогенеза стали результаты экспериментов, выполненных в 1960-1970 гг. английским исследователем Д. Гердоном с целью выяснить, обладают ли ядра соматических клеток способ- ностью обеспечивать дальнейшее развитие яйцеклеток, если эти ядра ввести в яйцеклетки, из которых предварительно удалены собственные ядра. На рис. 28 приведена схема одного из этих экспериментов, в котором ядра соматических клеток головастика пересаживали в яйцеклетки лягушки с предварительно удаленными ядрами. Эти эксперименты показали, что ядра соматических клеток действительно могут обеспечивать дальнейшее развитие яйцеклеток, так как они оказались способными оплодотворять яйцеклетки и «заставляли» их развиваться дальше. Этим была доказана возможность клонирования животных.

Рис. 28. Схема эксперимента по пересадке ядер соматических клеток в безъядерные яйцеклетки (Д. Гердон, 1968)
Позднее другими исследователями были выполнены эксперименты, в которых показано, что перенос отдельных бластомеров из 8- и 16-дневных эмбрионов овец одной породы в безъядерную половину яйцеклетки (после рассечения последней пополам) другой породы сопровождался формированием жизнеспособных эмбрионов с последующим рождением ягнят.
В начале 1997 г. английскими авторами было установлено, что введение в искусственно лишенные ядра яйцеклеток овец ядер соматических клеток (клеток эмбрионов, плодов или вымени взрослых овец), а затем имплантация оплодотворенных таким образом яйцеклеток в матку овец сопровождается возникновением беременности с последующим рождением ягнят (рис. 29). Один из ягнят был назван Долли. В 2003 г. Долли погибла. За это время были получены эмбрионы мышей, коров, кроликов, лошади, крысы и других животных.

Введение культуры в организм матери
Рис. 29. Пересадка ядер соматических клеток в безъядерные яйцеклетки
Оценка этих результатов показывает, что млекопитающих можно размножать асексуальным путем, получая потомство животных, клетки которых содержат ядерный материал отцовского или материнского происхождения в зависимости от пола овцы-донора. В таких клетках
лишь цитоплазма и митохондрии имеют материнское происхождение. Это заключение имеет чрезвычайно важное общебиологическое значение, расширяет наши взгляды на потенциал размножения животных. Но важно также добавить, что речь идет о генетических манипуляциях, которые в природе отсутствуют. С другой стороны, в практическом плане эти генетические манипуляции представляют собой прямой путь клонирования высокоорганизованных животных с заданными свойствами, что имеет важное экономическое значение. В медицинском плане этот путь, возможно, будет использован в будущем для преодоления мужского бесплодия.
Итак, генетическая информация, необходимая для нормального развития эмбриона, не теряется в течение дифференцировки клеток за счет так называемых стволовых клеток, которые обладают потенцией развиваться в различные типы клеток тела. Когда стволовые клетки делятся, каждая новая клетка имеет потенцию или оставаться стволовой клеткой, или стать клеткой с более специализированной функцией (мышечные клетки, клетки крови или клетки головного мозга). Оплодотворенная яйцеклетка является тотипотентной, так как дает начало разным типам клеток тела. Тотипотентные стволовые клетки могут дать начало любому типу клеток, кроме тех, которые необходимы для развития плода. Стволовые клетки, которые могут дать начало самым различным клеточным типам, обычно называют мультитотипотентными клетками. Установлено, что взрослые стволовые клетки могут продуцировать дифференцированные клетки из эмбриональных несвязанных тканей. Соматические клетки тоже обладают свойством, получившим название тотипотентности, т. е. в их геноме содержится вся информация, полученная ими от оплодотворенной яйцеклетки, давшей им начало в результате дифференциации. Наличие этих данных с несомненностью означает, что дифференциация клеток подвержена генетическому контролю. Изучение стволовых клеток имеет значение для медицины.
Установлено, что интенсивный белковый синтез, следующий за оплодотворением, у большинства эукариотов не сопровождается синтезом мРНК. Изучение овогенеза у позвоночных, в частности у амфибий, показало, что интенсивная транскрипция происходит еще в течение профазы I (особенно диплонемы) мейоза. Поэтому генные транскрипты в форме молекул мРНК или про-мРНК сохраняются в яйцеклетках в бездействующем состоянии. Установлено, что у эмбриональных клеток имеет место так называемое асимме-
тричное деление, заключающееся в том, что деление эмбриональной клетки дает начало двум клеткам, из которых лишь одна наследует белки, принимающие участие в транскрипции. Таким образом, неравное распределение транскрипционных факторов между дочерними клетками ведет к экспрессии в них разных наборов генов после деления, т. е. к дифференциации клеток.
У амфибий и, возможно, у большинства позвоночных генетические программы, контролирующие раннее развитие (до стадии бластулы), устанавливаются еще в течение овогенеза. Более поздние стадии развития, когда начинается клеточная дифференциация (примерно, со стадии гаструлы), нуждаются в новых программах для экспрессии генов. Таким образом, дифференцировка клеток связана с перепрограммированием генетической информации в том или ином направлении.
Характерная особенность дифференцировки клеток заключается в том, что она необратимо ведет к тому или иному типу клеток. Этот процесс носит название детерминации и также находится под генетическим контролем, а как сейчас предполагают, дифференциация и детерминация клеток регулируются взаимодействием клеток на основе сигналов, осуществляемых пептидными ростовыми факторами через тирозинкиназные рецепторы. Вероятно, существует много таких систем. Одна из них заключается в том, что дифференциация мышечных и нервных клеток регулируется нейрорегулина- ми, представляющими собой мембранные белки, действующие через один или более тирозинкиназный рецептор.
Генетический контроль детерминации демонстрируется также существованием так называемых гомейотропных или гомеозисных мутаций, которые, как показано у насекомых, вызывают изменения при детерминации в специфических имагинальных дисках. В результате некоторые части тела развиваются не на своих местах. Например, у дрозофил мутации трансформируют детерминацию антенного диска в диск, который развивается в аппендикс конечности, протянутой от головы. У насекомых из рода Ophthalmoptera структуры крыльев могут развиваться из диска для глаз. У мышей показано существование генного кластера (комплекса) Нох, который состоит из 38 генов и контролирует развитие конечностей.
Самостоятельное значение имеет вопрос о регуляции активности генов в период эмбрионального развития. Считают, что в ходе дифференцировки гены действуют в разное время, что выражается
в транскрипции в различных дифференцированных клетках разных мРНК, т. е. имеют место репрессия и дерепрессия генов. Например, количество генов, транскрибируемых в РНК в бластоцитах морского ежа, равно 10%, в клетках печени крыс - тоже 10%, а в клетках тимуса крупного рогатого скота - 15%. Предполагают, что в контроле транскрипционного статуса генов принимают участие негистоновые белки. В пользу этого предположения свидетельствуют следующие данные. Когда хроматин клеток в фазе S транскрибируется в системе in vitro, то синтезируется только гистоновая мРНК, а в след за нею и гистоны. Напротив, когда используют хроматин клеток из (^-фазы, то никакой гистоновой мРНК не синтезируется. Когда же непистоновые белки удаляются из хроматина (1 -фазы и замещаются негистоновыми хромосомными белками, синтезированными в фазе S, то после транскрипции такого хроматина in vitro синтезируется гистоновая мРНК. Более того, когда негистоновые белки происходят из (1 -фазы клеток, а ДНК и гистоны - из S-фазы клеток, никакой гистоновой мРНК не синтезируется. Эти результаты показывают, что содержащиеся в хроматине негистоновые белки определяют возможность транскрипции генов, кодирующих гистоны. Поэтому считают, что негистоновые хромосомные белки могут играть важную роль в регуляции и экспрессии генов у эукариот.
Имеющиеся данные позволяют считать, что в регуляции транскрипции у животных принимают участие белковые и стероидные гормоны. Белковый (инсулин) и стероидные (эстрогон и тестостерон) гормоны представляют собой две сигнальные системы, используемые в межклеточных коммуникациях. У высших животных гормоны синтезируются в специализированных секреторных клетках. Освобождаясь в кровяное русло, они поступают в ткани. Поскольку молекулы белковых гормонов имеют относительно крупные размеры, то они не проникают в клетки, поэтому их эффекты обеспечиваются белками-рецепторами, локализованными в мембранах клеток-мишеней, и внутриклеточными уровнями циклического АМФ (цАМФ). Напротив, стероидные гормоны являются малыми молекулами, вследствие чего легко проникают в клетки через мембраны. Оказавшись внутри клеток, они связываются со специфическими рецепторными белками, которые имеются в цитоплазме только клеток-мишеней. Как считают, комплексы гормон - белковый рецептор, концентрируясь в ядрах клеток-мишеней, активируют транскрипцию специфических генов через взаимодействие с определенны-
ми негистоновыми белками, которые связываются с промоторными районами специфических генов. Следовательно, связывание ком- плекса гормон + белок (белковый рецептор) с негистоновыми белками освобождает промоторные районы для движения РНК-полимеразы. Обобщая данные о генетическом контроле эмбрионального периода в онтогенезе организмов, можно заключить, что его ход контролируется дифференциальным включением и выключением действия генов в разных клетках (тканях) путем их дерепрессии и репрессии.
ПОСТЭМБРИОНАЛЬНОЕ РАЗВИТИЕ
После появления организма на свет начинается его постэмбриональное развитие (постнатальное для человека), которое у разных организмов протекает от нескольких дней до сотен лет в зависимости от их видовой принадлежности. Следовательно, продолжительность жизни - это видовой признак организмов, не зависящий от уровня их организации (см. ниже).
В постэмбриональном онтогенезе различают ювенильный и пубертатный периоды, а также период старости, заканчивающийся смертью.
Ювенильный период. Этот период (от лат. juvenilis - юный) опреде- ляется временем от рождения организма до полового созревания. Он протекает по-разному и зависит от типа онтогенеза организмов. Для этого периода характерно либо прямое, либо непрямое развитие.
В случае организмов, для которых характерно прямое развитие (многие беспозвоночные, рыбы, пресмыкающиеся, птицы, млекопи- тающие, человек), вылупившиеся из яйцевых оболочек или новорожденные сходны со взрослыми формами, отличаясь от последних лишь меньшими размерами, а также недоразвитием отдельных органов и несовершенными пропорциями тела (рис. 30).
Характерной особенностью роста в ювенильный период организмов, подверженных прямому развитию, является то, что проис- ходит увеличение количества и размеров клеток, изменяются пропорции тела. Рост человека в разные периоды его онтогенеза показан на рис. 31. Рост разных органов человека неравномерен. Например, рост головы заканчивается в детстве, ноги достигают пропорциональной величины примерно к 10 годам. Наружные половые органы очень быстро растут в возрасте 12-14 лет. Различают определенный и неопределенный рост. Определенный рост характерен для организмов, которые к определенному возрасту прекращают свой рост,
например насекомые, млекопитающие, человек. Неопределенный рост характерен для организмов, которые растут всю жизнь, напри- мер моллюски, рыбы, земноводные, рептилии, многие виды растений.

Рис. 30. Прямое и непрямое развитие организмов разных видов
В случае непрямого развития организмы претерпевают превращения, называемые метаморфозами (от лат. metamorphosis - превращение).

Рис. 31. Рост и развитие в разные периоды онтогенеза человека
Они представляют собой видоизменения организмов в процессе развития. Метаморфозы широко встречаются у кишечнополостных (гидры, медузы, коралловые полипы), плоских червей (фасциолы), круглых червей (аскариды), моллюсков (устрицы, мидии, осьминоги), членистоногих (раки, речные крабы, омары, креветки, скорпионы, пауки, клещи, насекомые) и даже у некоторых хордовых (оболочечники и земноводные). При этом различают полные и неполные метаморфозы. Наиболее выразительные формы метаморфоз наблюдаются у насекомых, которые подвергаются как неполным, так и полным метаморфозам.
Неполное превращение - это такое развитие, при котором из яйцевых оболочек выходит организм, строение которого сходно со строением взрослого организма, но размеры намного меньше. Такой организм называют личинкой. В процессе роста и развития размеры личинок увеличиваются, но имеющийся хитинизированный покров мешает дальнейшему увеличению размеров тела, что приводит к линьке, т. е. сбрасыванию хитинизированного покрова, под которым находится мягкая кутикула. Последняя расправляется, и это сопровождается увеличением размеров животного. После нескольких линек животное достигает зрелости. Неполное превращение характерно, например, в случае развития клопов.
Полное превращение - это такое развитие, при котором из яйцевых оболочек освобождается личинка, существенно отличающаяся
по строению от взрослых особей. Например, у бабочек и многих насекомых личинками являются гусеницы. Гусеницы подвержены линьке, причем могут линять по нескольку раз, превращаясь затем в куколки. Из последних развиваются взрослые формы (имаго), которые не отличаются от исходных.
У позвоночных метаморфозы встречаются среди земноводных и костных рыб. Для личиночной стадии характерно наличие про- визорных органов, которые либо повторяют признаки предков, либо имеют явно приспособительное значение. Например, для головастика, являющегося личиночной формой лягушки и повторяющего признаки исходной формы, характерны рыбообразная форма, наличие жаберного дыхания и одного круга кровообращения. Приспособительными признаками головастиков являются их присоски и длинный кишечник. Для личиночных форм характерно также и то, что по сравнению со взрослыми формами они оказываются приспособленными к жизни в совершенно иных условиях, занимая другую экологическую нишу и другое место в цепи питания. Например, личинки лягушек имеют жаберное дыхание, тогда как взрослые формы - легочное. В отличие от взрослых форм, которые являются плотоядными существами, личинки лягушек питаются растительной пищей.
Последовательность событий в развитии организмов часто называют жизненными циклами, которые могут быть простыми и сложными. Наиболее простые циклы развития характерны, например, для млекопитающих, когда из оплодотворенной яйцеклетки развивается организм, который снова продуцирует яйцеклетки, и т. д. Сложными биологическими циклами являются циклы у животных, для которых характерно развитие с метаморфозами. Знания о биологических циклах имеют практическое значение, особенно в случаях, когда организмы являются возбудителями или переносчиками воз- будителей болезни животных и растений.
Развитие и дифференциация, связанные с метаморфозами, являются результатом естественного отбора, благодаря которому многие личиночные формы, например гусеницы насекомых и головастики лягушек, адаптированы к среде лучше, чем взрослые половозрелые формы.
Пубертатный период. Этот период называют еще зрелым, и он связан с половой зрелостью организмов. Развитие организмов в этот период достигает максимума.
На рост и развитие в постэмбриональный период большое влияние оказывают факторы среды. Для растений решающими факторами являются свет, влажность, температура, количество и качество питательных веществ в почве. Для животных первостепенное значение имеет полноценное кормление (наличие в корме белков, углеводов, липидов, минеральных солей, витаминов, микроэлементов). Важны также кислород, температура, свет (синтез витамина D).
Рост и индивидуальное развитие животных организмов подвержены нейрогуморальной регуляции со стороны гуморальных и нервных механизмов регуляции. У растений обнаружены гормоноподобные активные вещества, получившие название фитогормонов. Последние влияют на жизненно важные отправления растительных организмов.
В клетках животных в процессе жизнедеятельности синтезируются химически активные вещества, влияющие на процессы жизнедеятельности. Нервные клетки беспозвоночных и позвоночных выра- батывают вещества, получившие название нейросекретов. Железы эндокринной, или внутренней, секреции также выделяют вещества, которые получили название гормонов. Эндокринные железы, в частности те, что имеют отношение к росту и развитию, регулируются нейросекретами. У членистоногих регуляция роста и развития очень хорошо показана на примере влияния гормонов на линьку. Синтез личиночного секрета клетками регулируются гормонами, накапливающимися в мозге. В особой железе у ракообразных вырабатывается гормон, тормозящий линьку. Уровни этих гормонов определяют периодичность линек. У насекомых установлены гормональная регуляция созревания яиц, протекающие диапаузы.
У позвоночных железами внутренней секреции являются гипофиз, эпифиз, щитовидная, паращитовидная, поджелудочная, над- почечники и половые железы, которые тесно связаны одна с другой. Гипофиз у позвоночных вырабатывает гонадотропный гормон, стимулирующий деятельность половых желез. У человека гормон гипофиза влияет на рост. При недостатке развивается карликовость, при избытке - гигантизм. Эпифиз продуцирует гормон, который влияет на сезонные колебания в половой активности животных. Гормон щитовидной железы влияет на метаморфоз насекомых и земноводных. У млекопитающих недоразвитие щитовидной железы ведет к задержке роста, недоразвитию половых органов. У человека из-за дефекта щитовидной железы задерживаются окостенение, рост
(карликовость), не наступает полового созревания, останавливается психическое развитие (кретинизм). Надпочечники продуцируют гормоны, оказывающие влияние на метаболизм, рост и дифференци- ровку клеток. Половые железы продуцируют половые гормоны, которые определяют вторичные половые признаки. Удаление половых желез ведет к необратимым изменениям ряда признаков. Например, у кастрированных петухов прекращается рост гребня, теряется половой инстинкт. Кастрированный мужчина приобретает внешнее сходство с женщиной (не растут борода и волосы на коже, отлагается жир на груди и в области таза, сохраняется тембр голоса и т. д.).
Фитогормонами растений являются ауксины, цитокинины и гиббереллины. Они регулирую рост и деление клеток, образование новых корней, развитие цветков и другие свойства растений.
На всех периодах онтогенеза организмы способны к восстановлению утраченных или поврежденных частей тела. Это свойство организмов носит название регенерации, которая бывает физиологической и репаративной.
Физиологическая регенерация - это замена утерянных частей тела в процессе жизнедеятельности организма. Регенерации этого типа очень распространены в животном мире. Например, у членистоногих она представлена линькой, которая связана с ростом. У рептилий регенерация выражается в замещении хвоста и чешуи, у птиц - перьев, когтей и шпор. У млекопитающих примером физиологической регенерации может быть ежегодное сбрасывание оленями рогов.
Репаративная регенерация - это восстановление части тела организма, отторгнутой насильственным путем. Регенерация этого типа возможна у многих животных, но ее проявления различны. Например, она часта у гидр и связана с размножением последних, поскольку из части регенерирует весь организм. У других организмов регенерации проявляются в виде способности отдельных органов к восста- новлению после утраты ими какой-либо части. У человека достаточно высокой регенеративной способностью обладают эпителиальная, соединительная, мышечная и костная ткани.
Растения многих видов также способны к регенерации.
Данные о регенерации имеют большое значение не только в биологии. Их широко используют в сельском хозяйстве, в медицине, в частности в хирургии.
Старость как этап онтогенеза. Старость является предпоследним этапом онтогенеза животных, причем ее длительность определяется
общей продолжительностью жизни, которая служит видовым признаком и у разных животных неодинаковая. Наиболее точно старость изучена у человека.
Известны самые различные определения старости человека. В частности, одно из наиболее популярных определений заключается в том, что старость есть накопление последовательных изменений, сопровождающих повышение возраста организма и увеличивающих вероятность его болезней или смерти. Науку о старости человека называют геронтологией (от греч. geron - старик, старец, logos - наука). Ее задачей является изучение закономерностей возрастного перехода между зрелостью и смертью.
Научные исследования в геронтологии распространяются на разные области, начиная с исследований изменений активности клеточных ферментов и заканчивая выяснением влияния психологических и социологических смягчений в стрессах среды на поведение старых людей.
В случае человека различают физиологическую старость; старость, связанную с календарным возрастом; и преждевременное старение, обусловленное социальными факторами и болезнями. В соответствии с рекомендациями ВОЗ пожилым возрастом человека следует считать возраст порядка 60-75 лет, а старым - 75 лет и более.
Старость человека характеризуется рядом внешних и внутренних признаков.
Среди внешних признаков старости наиболее заметными являются снижение плавности движений, изменение осанки, снижение эластичности кожи, массы тела, упругости и эластичности мышц, появление на лице и других участках тела морщин, выпадение зубов. Так, например, по обобщенным данным, человек в возрасте 30 лет теряет 2 зуба (в результате выпадения), в 40 лет - 4 зуба, в 50 лет - 8 зубов, а в 60 лет - уже 11 зубов. Заметным изменениям подвергается первая сигнальная система (притупляется острота органов чувств). Например, максимальное расстояние, при котором здоровые люди различают те или иные одинаковые звуки, в 20-30 лет составляет 12 м, в 50 лет - 10 м, в 60 лет - 7 м, а в 70 лет - только 4 м. Заметно изменяется также вторая сигнальная система (изменяется речевая интонация, голос становится глухим).
Среди внутренних признаков в первую очередь следует назвать такие признаки, как обратное развитие (инволюция) органов. Отмечается уменьшение размеров печени и почек, а также количе-
ства нефронов в почках (к 80 годам почти наполовину), что снижает функциональные возможности почек и отражается на водноэлектролитном обмене. Снижается эластичность кровеносных сосудов, уменьшается перфузия кровью тканей и органов, повышается периферическая сопротивляемость сосудов. В костях накапливаются неорганические соли, изменяются (обызвествляются) хрящи, снижается способность органов к регенерации. Происходят существенные изменения в клетках, замедляется деление и восстанов- ление их функционального тонуса, уменьшается содержание воды, снижается активность клеточных ферментов, нарушается координированность между ассимиляцией и диссимиляцией. В головном мозге нарушается синтез белков, в результате чего образуются аномальные белки. Повышается вязкость клеточных мембран, нарушаются синтез и утилизация половых гормонов, происходят изменения в структуре нейронов. Наступают структурные изменения белков соединительной ткани и эластичности этой ткани. Ослабляются иммунологические реакции, увеличивается возможность аутоиммунных реакций. Снижаются функции эндокринных систем, в частности половых желез. Поведение других признаков в старости показано на рис. 32.

Рис. 32. Изменение некоторых признаков человека с возрастом: 1 - скорость проведения нервных импульсов; 2 - уровень основного обмена; 3 - сердечный индекс; 4 - уровень почечной фильтрации по инсулину; 5 - дыхательные объемы легких; 6 - уровень плазмотока в почках
Стремления понять природу старения организма возникли давно. В Древней Греции Гиппократ считал, что старение связано с неумеренностью в пище, недостаточным пребыванием на свежем воздухе. По мнению Аристотеля, старение обусловлено расходом тепловой энергии организмом. Значение пищи как фактора старения отмечал также Гален. Но долгое время для объективного понимания этой проблемы не хватало научных данных. Лишь в XIX в. в изучении старения наметился некоторый прогресс, стали возникать теории старения.
Одной из первых наиболее известных теорий старения организма человека является теория немецкого врача X. Гуфеланда (1762-1836), который придавал в долголетии значение трудовой деятельности. До нас дошло его высказывание о том, что ни один лентяй не дожил до преклонного возраста. Еще более известна эндокринная теория старения, которая берет начало из опытов, выполненных еше в середине позапрошлого века Бертольдом (1849), который показал, что пересадка семенников от одних животных к другим сопровождается развитием вторичных половых признаков. Позднее французский физиолог Ш. Броун-Секар (1818-1894) на основе результатов впрыскиваний себе экстрактов из семенников утверждал, что эти инъекции производят благотворное и омолаживающее действие. В начале XX в. уже сложилось убеждение в том, что наступление старости связано с угасанием деятельности желез внутренней секреции, в частности половых желез. В 20-30-е гг. XX в. на основе этого убеждения в разных странах было проделано много операций по омолаживанию пожилых или старых людей. Например, Г. Штейнах в Австрии перевязывал у мужчин семенные канатики, что вело к прекращению внешней секреции половых желез и якобы к некоторому омоложению. С.А. Воронов во Франции пересаживал семенники от молодых животных к старым и от обезьян к мужчинам, а Тушнов в СССР омолаживал петухов, вводя им гистолизаты половых желез. Все эти операции приводили к некоторым эффектам, но лишь временным. После названных воздействий процессы старения продолжались, причем еще интенсивнее.
В начале XX в. возникла микробиологическая теория старения, творцом которой был И.И. Мечников, различавший физиологиче- скую старость и патологическую. Он считал, что старость человека является патологической, т.е. преждевременной. Основу представлений И.И. Мечникова составляло учение об ортобиозе (orthos -
правильный, bios - жизнь), в соответствии с которым основной причиной старения является повреждение нервных клеток продуктами интоксикации, образующимися в результате гниения в толстом кишечнике. Развивая учение о нормальном образе жизни (соблюдение правил гигиены, регулярный труд, воздержание от вредных привычек), И.И. Мечников предлагал также способ подавления гнилостных бактерий кишечника путем употребления кисломолочных продуктов.
В 30-е гг. XX в. широкое распространение получила теория о роли центральной нервной системы (ЦНС) в старении. Творцом этой теории является И.П. Павлов, который установил интегрирующую роль ЦНС в нормальном функционировании организмов. Последователи И.П. Павлова в экспериментах на животных показали, что преждевременное старение вызывается нервными потрясениями и продолжительным нервным перенапряжением.
Заслуживает упоминания теория возрастных изменений соединительной ткани, сформулированная в 30-е гг. XX в. А.А. Богомольцем (1881-1946). Он считал, что физиологическую активность организма обеспечивает соединительная ткань (костная ткань, хрящи, сухожилия, связки и волокнистая соединительная ткань) и что изменения коллоидного состояния клеток, потеря их тургора и т. д. определяют возрастные изменения организмов. Современные данные указывают на значение накопления кальция в соединительных тканях, так как он способствует потере ее упругости, а также уплотнению сосудов.
Для современных подходов к познанию сущности и механизмов старения характерно широкое использование данных физико- химической биологии, и в частности достижений молекулярной генетики. Наиболее распространенные современные представления о механизме старения сводятся к тому, что в процессе жизни в клетках организма накапливаются соматические мутации, в результате которых происходит синтез дефектных белков или нерепарируемые сшивки ДНК с белком. Поскольку дефектные белки играют дозинте- грирующую роль в клеточном метаболизме, то это ведет к старению. В случае культивируемых фибробластов показано, что связанные со старыми клетками белки и мРНК подавляют синтез ДНК в молодых фибробластах.
Известна также гипотеза, в соответствии с которой старение считается результатом изменения митохондриальных метаболитов с последующим нарушением функций ферментов.
У человека показано существование генов, определяющих сроки развития наследственных дегенеративных процессов, связанных со старением. Ряд исследователей считают, что причиной старения являются изменения в системе иммунологической защиты организма, в частности аутоиммунные реакции на структуры организма, имеющие жизненное значение. Наконец, в объяснениях механизмов старения специалисты большое внимание уделяют повреждениям белков, связанным с образованием свободных радикалов. Наконец, иногда придают значение освобождающимся после распада лизосом гидролазам, которые разрушают клетки.
Однако исчерпывающей теории старения все же не создано, поскольку ясно, что ни одна из этих теорий самостоятельно объяс- нить механизмы старения не может.
Смерть. Смерть является завершающим этапом онтогенеза. Вопрос о смерти в биологии занимает особое место, ибо чувство смерти «...совершенно инстинктивно присуще человеческой природе и всегда составляло одну из величайших забот человека» (И.И. Мечников, 1913). Более того, вопрос о смерти стоял и стоит в центре внимания всех философских и религиозных учений, хотя философия смерти в разные исторические времена представлялась неодинаково. В античном мире Сократ и Платон доказывали бессмертие души, тогда как Аристотель отрицал платоновскую идею бессмертия души и верил в бессмертность человеческого духа, продолжающего жить после смерти человека. Цицерон и Сенека также признавали будущую жизнь, но Марк Аврелий считал смерть естественным явлением, которое следует принимать безропотно. В XVIII в. И. Кант и И. Фихте (1762-1814) тоже верили в будущую жизнь, а Г. Гегель придерживался убеждений, по которым душа поглощается «абсолютным существом», хотя природа этого «существа» не раскрывалась.
В соответствии со всеми известными религиозными учениями земная жизнь человека продолжается и после его смерти, и человек должен неустанно готовиться к этой будущей смерти. Однако естество- испытатели и философы, не признающие бессмертия, считали и считают, что смерть представляет собой, как неоднократно подчеркивал И.И. Мечников, естественный исход жизни организма. Более образное определение смерти заключается в том, что она «...есть явная победа бессмыслия над смыслом, хаоса над космосом» (В. Соловьев, 1894).
Научные данные свидетельствуют о том, что у одноклеточных организмов (растений и животных) следует отличать смерть от прекраще-
ния их существования. Смертью является их гибель, тогда как прекращение существования связано с их делением. Следовательно, недолговечность одноклеточных организмов компенсируется их раз- множением. У многоклеточных растений и животных смерть является в полном смысле слова завершением жизни организма.
У человека вероятность смерти повышается в пубертатный период. В частности, в развитых странах вероятность смерти возрастает почти экспоненциально после 28 лет.
Различают клиническую и биологическую смерть человека. Клиническая смерть выражается в потере сознания, прекращении сердцебиения и дыхания, однако большинство клеток и органов все же остаются живыми. Происходит самообновление клеток, продолжается перистальтика кишечника. Клиническая смерть не «доходит» до биологической смерти, ибо она обратима, так как в состоянии клинической смерти можно «возвращать» к жизни. Например, собак «возвращают» к жизни через 5-6 мин, человека - через 6-7 мин от начала клинической смерти. Биологическая смерть характеризуется тем, что она необратима. Остановка сердцебиения и дыхания сопровождается прекращением процессов самообновления, гибелью и разложением клеток. Однако гибель клеток начинается не во всех органах одновременно. Вначале гибнет кора головного мозга, затем гибнут эпителиальные клетки кишечника, легких, печени, клетки мышц, сердца.
На представлениях о клинической смерти основаны мероприятия по реанимации (оживлении) организмов, что имеет исключительно важное значение в современной медицине.
ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
Сравнение данных о продолжительности жизни представителей флоры и фауны показывает, что среди растений и животных различ- ные организмы живут разное время. Например, травянистые растения (дикие и культурные) живут в течение одного сезона. Напротив, их древесные растения характеризуются большей продолжительностью жизни. Например, вишня живет 100 лет, ель - 1000 лет, дуб -
2000 лет, сосна - до 2000-4000 лет.
Рыбы многих видов живут 55-80 лет, лягушки - 16 лет, крокодилы - 50-60 лет, птицы некоторых видов - до 100 лет. Продолжительность жизни млекопитающих является меньшей.
Например, мелкий рогатый скот живет 20-25 лет, крупный рогатый скот - 30 лет и более, лошади - до 30 лет, собаки - 20 лет и более, волки - 15 лет, медведи - 50 лет, слоны - 100 лет. Среди млеко- питающих долгожителем является человек. Многие люди доживали до 115-120 лет и более.
В соответствии с существующими представлениями продолжительность жизни является видовым количественным признаком, подверженным контролю со стороны генотипа. Считают, что гене- тически детерминированная продолжительность жизни коррелирует с периодами онтогенеза (табл. 5).
Таблица 5. Зависимость уровня фибробластов при культивировании от возраста донора
Предполагают, что видовая продолжительность жизни является эволюционным приобретением вида. Что касается долгожительства отдельных индивидуумов, то его объяснение возможно допущением наличия в их генотипах сочетаний определенных генов, либо небольшого количества или полного отсутствия мутаций в их клетках.
А.А. Богомолец и И.И. Шмальгаузен подсчитали, что естественная продолжительность жизни человека должна составлять 120- 150 лет. Однако до этого возраста доживают лишь единицы. Поэтому фактическая продолжительность жизни не совпадает с естественной продолжительностью.
На среднюю продолжительность жизни оказывает влияние ряд факторов (снижение детской смертности, эффективность борьбы с инфекциями, успехи хирургии, улучшение питания и общих условий жизни). Главнейшие причины снижения продолжительности жизни заключаются в смертности от голода, болезней, недостаточной медицинской помощи. В России в последние годы отмечается отрицательная численность населения во всех русских городах и сельской местности за счет снижения рождаемости и высокой смертности.
В медицинском плане продолжительность жизни - это показатель здоровья нации. СССР по числу старых людей занимал первое место в мире. Например, на 1 млн жителей приходится 104 человека в возрасте свыше 90 лет, тогда как в Англии - 6, Франции - 7
и США - 15 человек. В настоящее время отмечается изменение границ трудоспособного населения по сравнению, например, с 30-ми гг. XX в. Отличается также разрыв между пенсионным возрастом и активно- стью людей. В 1982 г. в Вене состоялась Всемирная ассамблея по проблемам старения населения мира, на которой были сформулированы прогнозы по этой проблематике до 2025 г. Предполагается, что численность людей в возрасте 60 лет и старше по сравнению с 1950 г. возрастет в 5 раз, людей старше 80 лет - в 7 раз. По данным этого международного форума, имеются различия в скорости старения населения применительно к разным народам, странам и регионам.
Гериатрия - наука, задачей которой является разработка способов нормализации изменяющихся функций стареющего организма. Однако современная медицина еще не располагает методами и средствами воздействия на нормальные физиологические процессы. Поэтому роль гериатрии ограничивается лечением возникающих в пожилом и старческом возрасте заболеваний и исключением (при наличии возможностей) факторов риска, вызывающих прежде- временное старение.
ВОПРОСЫ ДЛЯ ОБСУЖДЕНИЯ
1. Что вы понимаете под половым размножением организмов и какова его биологическая роль?
2. Опишите бесполое размножение и назовите его формы.
3. Опишите особенности полового процесса у одноклеточных и многоклеточных организмов.
4. Что такое гаметогенез?
5. Какова функция гамет каждого типа?
6. Какие вы знаете стадии в развитии гамет?
7. В чем заключаются сходства и различия между сперматогенезом и овогенезом?
8. Что такое мейоз и каково его биологическое значение?
9. Опишите фазы мейоза.
10. Влияет ли кроссинговер на результаты мейоза и как?
11. Опишите сущность оплодотворения.
12. В чем заключается разница между зигогенезом и партеногенезом?
13. Какова биологическая роль чередования гаплоидии и диплоидии?
14. Что представляет собой половой диморфизм? Что вы понимаете под гермафродитизмом? Наблюдаются ли случаи гермафродитизма у человека и как часто?
15. Как вы представляете эволюцию способов размножения?
16. Что вы понимаете под ростом и развитием организмов? Какова связь между ростом и дифференциацией клеток?
17. В чем заключаются молекулярные основы дифференцировки клеток? Что вы знаете о стволовых клетках?
18. Сформулируйте понятие об онтогенезе и назовите периоды онтогенеза.
19. Каковы различия между прямым и непрямым развитием?
20. В чем заключается влияние оплодотворения на яйцеклетки?
21. На каком этапе реализации генетической информации осуществляется контроль действия генов?
22. Каким образом оплодотворенная яйцеклетка развивается в многоклеточную структуру?
23. Каким образом развивающиеся клетки и ткани становятся отличными одни от других в процессе развития?
24. Могут ли восстанавливаться утерянные или поврежденные части тела?
25. Какова связь в онтогенезе между старостью и продолжительностью жизни?
26. Сформулируйте и определите разницу между естественной (вероятной) и фактической продолжительностью жизни.
27. Какие вам известны теории старения организмов?
29. Какие факторы оказывают влияние на продолжительность жизни?
30. На чем основано клонирование организмов? Приведите примеры клонирования организмов.
31. В клетку проникает сперматозоид или его содержимое?
32. О чем свидетельствуют данные по клонированию?
33. Преформизм и эпигенез в истории биологии. Имеют ли они научное значение?
34. Каковы различия между клинической и биологической смертью?
Воспроизведение - это способность организмов образовывать себе подобных. Воспроизведение является одним из важнейших свойств жизни и возможно благодаря общей способности организмов производить потомство. Однако не всегда непосредственные потомки подобны родительским особям. Например, из спор папоротника вырастает многочисленное потомство, представленное заростками, не похожими на материнское спороносное растение. На заростке, в свою очередь, возникает непохожее на него растение - спорофит. Такое явление получило название чередование поколений.
Особую форму полового размножения представляет собой партеногенез, или девственное размножение, - развитие организма из неоплодотворенной яйцеклетки. Данная форма размножения характерна главным образом для видов, обладающих коротким жизненным циклом с выраженными сезонными изменениями. Партеногенез бывает гаплоидным и диплоидным. При гаплоидном (генеративном) партеногенезе новый организм развивается из гаплоидной яйцеклетки. Получаемые при этом особи могут быть только мужскими, только женскими или теми и другими. Это зависит от хромосомного определения пола.
Индивидуальное развитие организма, или онтогенез, - этосовокупность последовательных морфологических, физиологических и биохимических преобразований, претерпеваемых организмом от момента его зарождения до смерти. В онтогенезе происходит реализация наследственной информации, полученной организмом от родителей. В онтогенезе выделяют два основных периода - эмбриональный и постэмбриональный. В эмбриональном у животных формируется эмбрион, у которого закладываются основные системы органов. В постэмбриональном периоде завершаются формообразовательные процессы, происходит половое созревание, размножение, старение и смерть.
