§ 2. Основные компоненты эукариотной клетки
Эукариотные клетки (рис. 8 и 9) организованы значительно сложнее прокариотных. Весьма разнообразны они и по своим размерам (от нескольких микрометров до нескольких сантиметров), и по форме, и по структурным особенностям (рис. 10).
Рис. 8. Строение клетки эукариот. Обобщенная схема
Рис. 9. Строение клетки по данным электронной микроскопии
Рис. 10. Разные эукариотные клетки: 1 – эпителиальная; 2 – крови (е – эритроиит, / – лейкоиит); 3 – хряша; 4 – кости; 5 – гладкая мышечная; 6 – соединительной ткани; 7 – нервные клетки; 8 – поперечно-полосатое мышечное волокно
Однако общая организация и наличие основополагающих компонентов у всех эукариотных клеток одинаковы (рис. 11).
Рис. 11. Эукариотная клетка (схема)
Плазмалемма (наружная клеточная мембрана). Основу плазмалеммы, как и других мембран в клетках (например, митохондрий, пластид и т. д.), составляет слой липидов, имеющий два ряда молекул (рис. 12). Поскольку молекулы липидов полярны (один полюс у них гидрофилен, т. е. притягивается водой, а другой гидрофобен, т. е. отталкивается от воды), то и располагаются они в определенном порядке. Гидрофильные концы молекул одного слоя направлены в сторону водной среды – в цитоплазму клетки, а другого слоя – наружу от клетки – в сторону межклеточного вещества (у многоклеточных) или водной среды (у одноклеточных).
Рис. 12. Строение клеточноймембраны согласно жидкостно-мозаичной модели. Белкии гликопротеины погружены в двойной слой липидных молекул, обращенных своими гидрофильными концами (кружки) наружу, а гидрофобными (волнистые линии) – в глубь мембраны
Молекулы белков мозаично встроены в бимолекулярный слой липидов. С внешней стороны животной клетки к липидам и молекулам белков плазмалеммы присоединяются молекулы полисахаридов, образуя гликолипиды и гликопротеины.
Эта совокупность формирует слой гликокаликса. С ним связана рецепторная функция плазмалеммы (см. ниже); также в нем могут накапливаться различные вещества, используемые клеткой. Кроме того, гликокаликс усиливает механическую устойчивость плазмалеммы.
В клетках растений и грибов есть еще клеточная стенка, играющая опорную и защитную роль. У растений она состоит из целлюлозы, а у грибов – из хитина.
Наружная клеточная мембрана выполняет ряд функций, среди которых:
♦ механическая (опорная, формообразующая);
♦ барьерно-транспортная (избирательная проницаемость в отношении разных веществ: поступление в клетку необходимых и выведение ненужных и вредных);
♦ рецепторная (определение различных химических веществ, оказавшихся в непосредственной близости от клетки; восприятие сигналов в виде гормонов; узнавание «чужого» белка клетками иммунной системы и т. д.).
Обмен веществ между клеткой и окружающей средой осуществляется разными способами – пассивными и активными.
Молекулы воды и различных ионов пассивно (за счет диффузии, осмоса), без затраты клеткой энергии, поступают через особые поры – это пассивный транспорт. Макромолекулы, такие как белки, полисахариды, даже целые клетки, поступают путем фагоцитоза и пиноцитоза с затратой энергии – активный транспорт.
Путем фагоцитоза поглощаются целые клетки или крупные частицы (например, вспомните питание у амеб или фагоцитоз защитными клетками крови бактерий). При пиноцитозе происходит поглощение мелких частиц или капелек жидкого вещества. Общим для обоих процессов является то, что поглощаемые вещества окружаются впячивающейся наружной мембраной с образованием вакуоли, которая затем перемещается в глубь цитоплазмы клетки.
Экзоцитоз представляет собой процесс (будучи также активным транспортом), противоположный по направлению фагоцитозу и пиноцитозу (рис. 13). С его помощью могут выводиться непереваренные остатки пищи у простейших либо образованные в секреторной клетке биологически активные вещества.
Цитоплазма. Цитоплазма – это содержимое клетки, ограниченное плазмалеммой, за исключением ядра. В ее составе выделяют основное вещество (гиалоплазму), органоиды и включения.
Гиалоплазма – вязкая жидкость, способная находиться в состоянии либо золя (жидком), либо геля (студнеобразном).
При необходимости цитоплазма способна обратимо переходить из одного состояния в другое. Например, при амебоидном движении (вспомните раздел «Простейшие» из курса зоологии) в ходе образования ложноножки происходят быстрые переходы цитоплазмы из геля в золь и наоборот. Это обусловлено наличием в цитоплазме большого количества нитевидных молекул из белка актина. Когда они, соединяясь друг с другом, образуют трехмерную сеть, цитоплазма находится в состоянии геля, а когда сеть распадается – в состоянии золя.
В гиалоплазме содержатся различные вещества – ферменты, белки, углеводы, жиры и другие, органические и минеральные. Здесь осуществляются различные химические процессы – расщепление веществ, их синтез и модификации (изменения).
Органоиды. Это постоянные компоненты клетки с определенным строением и функциями, находящиеся в ее цитоплазме. В дальнейшем речь будет идти об органоидах общего назначения, присущих любым типам клеток всех эукариот. С ними связано обеспечение жизнедеятельности последних. Органоиды специального назначения встречаются только в клетках определенного (узкоспециализированного) типа – например, миофибриллы в мышечных клетках.
Органоиды общего назначения имеют одинаковое строение независимо от того, каким клеткам и каких организмов они принадлежат. Но среди них выделяют группы с мембранным (эндоплазматическая сеть, аппарат Гольджи, митохондрии, пластиды, лизосомы, вакуоли), а также немембранным (рибосомы, клеточный центр) строением.
Эндоплазматическая сеть (ЭПС). ЭПС состоит из мембран и представляет собой сложно разветвленную систему канальцев и цистерн, пронизывающую всю цитоплазму клетки (рис. 14). Различают два вида ЭПС – шероховатую и гладкую. К мембранам шероховатой (со стороны цитоплазмы) прикрепляются рибосомы, а на гладкой их нет.
Рис. 14. Эндоплазматическаясеть
Эндоплазматическая сеть выполняет в эукариотной клетке ряд важнейших функций:
♦ разграничивающую (разделение внутреннего объема клетки на различные реакционные пространства);
♦ участие в синтезе органических веществ (на мембранах шероховатой ЭПС располагаются рибосомы, а на гладкой – ферментные комплексы, обеспечивающие синтез липидов, углеводов и т. д.);
♦ участие в формировании элементов аппарата Гольджи, лизосом;
♦ транспорт веществ.
Аппарат Гольджи. Аппарат Гольджи (АГ) представляет собой систему цистерн (плоских вакуолей) и пузырьков (везикул), расположенную в непосредственной близости от ядра клетки, которые образуются за счет ЭПС в результате отделения небольших ее фрагментов (рис. 15). При слиянии этих фрагментов и возникают новые цистерны аппарата Гольджи, при этом из ЭПС транспортируются различные вещества, которые участвуют в сборке сложных органических соединений (белки + углеводы, белки + липиды и т. д.), выводимых с помощью АГ за пределы клетки. Эти биологически активные вещества либо выводятся из клетки (с помощью секреторных вакуолей путем экзоцитоза), либо входят в состав лизосом (см. ниже), образующихся за счет АГ.
Рис. 15. Аппарат Гольджи:
Аппарат Гольджи выполняет следующие функции:
♦ синтез биологически активных веществ, вырабатываемых клеткой;
♦ секрецию (выведение из клетки) различных веществ (гормонов, ферментов, веществ, из которых строится клеточная стенка, и т. п.);
♦ участие в образовании лизосом.
Митохондрии. Митохондрии есть у всех типов эукариотных клеток (рис. 16). Они имеют вид либо округлых телец, либо палочек, реже – нитей. Их размеры колеблются от 1 до 7 мкм. Число митохондрий в клетке составляет от нескольких сотен до десятков тысяч (у крупных простейших).
Рис. 16. Митохондрии. Вверху – митохондрии (a) в мочевых канальиах, видимые в световом микроскопе. Внизу – трехмерная модель организации митохондрии: 1 – кристы; 2 – внешняя мембрана; 3 – внутренняя мембрана; 4 – матрикс
Митохондрия образована двумя мембранами – внешней и внутренней, между которыми расположено межмембранное пространство. Внутренняя мембрана образует множество впячиваний – крист, представляющих собой либо пластины, либо трубочки. Такая ее организация обеспечивает огромную площадь внутренней мембраны. На ней располагаются ферменты, обеспечивающие преобразование энергии, заключенной в органических веществах (углеводах, липидах), в энергию АТФ, необходимую для жизнедеятельности клетки. Следовательно, функция митохондрий – участие в энергетических клеточных процессах. Именно поэтому большое количество митохондрий присуще, например, мышечным клеткам, выполняющим большую работу.
Пластиды. В растительных клетках обнаруживаются особые органоиды – пластиды, имеющие чаще веретеновидную или округлую форму, иногда более сложную. Различают три вида пластид – хлоропласты (рис. 17), хромопласты и лейкопласты.
Хлоропласты отличаются зеленым цветом, который обусловлен пигментом – хлорофиллом, обеспечивающим процесс фотосинтеза, т. е. синтеза органических веществ из воды (Н 2 О) и углекислого газа (СО 2) с использованием энергии солнечного света. Хлоропласты содержатся преимущественно в клетках листьев (у высших растений). Они сформированы двумя параллельно расположенными друг другу мембранами, окружающими содержимое хлоропластов – строму. Внутренняя мембрана образует многочисленные уплощенные мешочки – тилакоиды, которые сложены в стопки (наподобие стопки монет) – граны – и лежат в строме. Именно в тилакоидах и содержится хлорофилл.
Хромопласты определяют желтый, оранжевый и красный цвет многих цветков и плодов, в клетках которых присутствуют в большом количестве. Основными пигментами в их составе являются каротины. Функциональное назначение хромопластов состоит в цветовом привлечении животных, обеспечивающих опыление цветков и распространение семян.
Рис. 17. Пластиды: а – хлоропласты в клетках листа элодеи, видимые в световом микроскопе; б – схема внутреннего строения хлоропласта с гранами, представляющими собой стопки плоских мешочков, расположенных перпендикулярно поверхности хлоропласта; в – более подробная схема, на которой видны анастомозируюшие трубочки, соединяющие отдельные камеры фан
Лейкопласты – это бесцветные пластиды, содержащиеся в клетках подземных частей растений (например, в клубнях картофеля), семян и сердцевины стеблей. В лейкопластах, главным образом, происходит образование из глюкозы крахмала и накапливание его в запасающих органах растений.
Пластиды одного вида могут превращаться в другой. Например, при осеннем изменении цвета листьев хлоропласты превращаются в хромопласты.
Лизосомы. Эти органоиды имеют вид пузырьков, окруженных мембраной, диаметром до 2 мкм. Они содержат несколько десятков ферментов, расщепляющих белки, нуклеиновые кислоты, полисахариды и липиды. Функция лизосом – участие в процессах внутриклеточного расщепления сложных органических соединений (например, пищевых веществ или веществ «отработавших» клеточных компонентов). Лизосомы сливаются с фагоцитарными (или пиноцитарными) вакуолями, образуя пищеварительную вакуоль.
Образование лизосом происходит за счет отпочковывания от цистерн аппарата Гольджи.
Рибосомы. Рибосомы (рис. 18) присутствуют в клетках как эукариот, так и прокариот, поскольку выполняют важную функцию в биосинтезе белков (см. главу 5). В каждой клетке имеются десятки, сотни тысяч (до нескольких миллионов) этих мелких округлых органоидов.
Рис. 18. Схема строения рибосомы, сидяшей на мембране эндоплазматической сети: 1 – малая субьединица; 2 – тРНК; 3 – аминоацил-тРНК; 4 – аминокислота; 5 – большая субъединица; 6 – мембрана эндоплазматической сети; 7 – синтезируемая полипептидная цепь
Рибосома состоит из двух неравных субъединиц (частей). Они образуются отдельно и объединяются, «охватывая» информационную РНК, в процессе синтеза белковой молекулы. В состав рибосом входят различные белки и рибосомальные РНК.
Клеточные включения. Так называются непостоянные компоненты в клетке, присутствующие в основном веществе цитоплазмы в виде зерен, гранул или капелек. Включения могут быть окружены мембраной или же не окружаются ею.
В функциональном отношении выделяют три вида включений: запасные питательные вещества (крахмал, гликоген, жиры, белки), секреторные включения (вещества, характерные для железистых клеток, продуцируемые ими, – гормоны желез внутренней секреции и т. п.) и включения специального назначения (в узкоспециализированных клетках, например гемоглобин в эритроцитах).
§ 3. Организация клеточного ядра. Хромосомы
Клеточное ядро (см. рис. 8и9) имеет важнейшее значение в жизнедеятельности клетки, поскольку служит хранилищем наследственной информации, содержащейся в хромосомах (см. ниже).
Ядро ограничено ядерной оболочкой, отделяющей его содержимое (кариоплазму) от цитоплазмы. Оболочка состоит из двух мембран, разделенных промежутком. Обе они пронизаны многочисленными порами, благодаря которым возможен обмен веществами между ядром и цитоплазмой. В ядре клетки у большинства эукариот находится от 1 до 7 ядрышек. С ними связаны процессы синтеза РНК и тРНК.
Основные компоненты ядра – хромосомы, образованные из молекулы ДНК и различных белков. В световом микроскопе они хорошо различимы лишь в период клеточного деления (митоза, мейоза). В неделящейся клетке хромосомы имеют вид длинных тонких нитей, распределенных по всему объему ядра.
Во время деления клеток хромосомные нити образуют плотные спирали, вследствие чего становятся видимыми (с помощью обычного микроскопа) в форме палочек, «шпилек». Весь объем генетической информации распределен между хромосомами ядра. В процессе их изучения были выявлены следующие закономерности:
♦ в ядрах соматических клеток (т. е. клеток тела, неполовых) у всех особей одного вида содержится одинаковое количество хромосом, составляющих набор хромосом (рис. 19);
Рис. 19. Хромосомы разных видов растений и животных, изображенные в одном масштабе: 1,2 – амеба; 3,4 – диатомовые водоросли; 5–8, 18,19 – зеленые водоросли; 9 – мухомор; 10 – липа; 11–12 – дрозофила; 13 – семга; 14 – скерда (семейство сложноцветных); 15 – растение из семейства ароидных; 16 – бабочка-хохлатка; 17 – насекомое из семейства саранчовых; 20 – клоп-водомерка; 21 – цветочный клоп; 22 – земноводное амбистома; 23 – алоэ (семейство лилейных)
♦ для каждого вида характерен свой хромосомный набор по их количеству (например, у человека 46 хромосом, у мушки дрозофилы – 8, у аскариды – 4, у речного рака – 196, у лошади – 66, у кукурузы – 104);
♦ хромосомы в ядрах соматических клеток могут быть сгруппированы парами, получившими название гомологичных хромосом на основании их сходства (по строению и функциям);
♦ в ядрах половых клеток (гамет) из каждой пары гомологичных хромосом содержится только одна, т. е. общий набор хромосом вдвое меньше, чем в соматических клетках;
♦ одинарный набор хромосом в половых клетках называется гаплоидным и обозначается буквой n, а в соматических – диплоидным (2n).
Из изложенного ясно, что каждая пара гомологичных хромосом образована объединением отцовских и материнских хромосом при оплодотворении, т. е. слиянии половых клеток (гамет). И наоборот, при образовании половых клеток из каждой пары гомологичных хромосом в гамету попадает только одна.
Хромосомы разных гомологичных пар отличаются по размерам и форме (рис. 20 и 21).
Рис. 20. Строение и типы хромосом: а – внешний вид 1 – цетромера; 2 – короткое плечо; 3 – длинное плечо); внутренняя структура той же хромосомы (1 – центромера; – молекулы ДНК); в – типы хромосом (1 – одноплечая; разноплечая; 3 – равноплечая: X – плечо, V – центромера)
Рис. 21. Хромосома состоит из ДНК и белков. Молекула ДНК реплицируется. Две идентичные двойные спирали ДНК остаются соединенными в области иентромеры. Эти копии превращаются в отдельные хромосомы позднее, во время деления клетки
В теле хромосом выделяют первичную перетяжку (называемую центромерой), к которой прикрепляются нити веретена деления. Она делит хромосому на два плеча. Хромосомы могут быть равноплечими, разноплечими и одноплечими.
Глава 5. Обмен веществ
§ 1. Метаболизм как единство ассимиляции и диссимиляции
Все клетки и живые организмы – это открытые системы, т. е. они пребывают в состоянии постоянного обмена энергий и веществ с окружающей средой. Имеются открытые системы и в неживой природе, но их существование качественно отличается от живых организмов. Рассмотрим такой пример: горящий кусок самородной серы находится в состоянии обмена с окружающей средой. При его горении поглощается О 2 , а выделяются SO 2 и энергия (в виде тепла). Однако при этом кусок серы как физическое тело разрушается, утрачивает свою первичную структуру.
Для живых же организмов обмен с окружающей средой оказывается условием сохранения, поддержания их структурной организации путем самообновления всех веществ и компонентов, из которых они состоят.
Обмен веществ (метаболизм) – совокупность протекающих в живых организмах процессов (потребления, превращения, накопления и выделения веществ и энергии), обеспечивающих ихжизнедеятельность, развитие, рост, воспроизведение. В процессе обмена веществ происходит расщепление и синтез молекул, входящих в состав клеток; обновление клеточных структур и межклеточного вещества.
В основе метаболизма лежат взаимосвязанные процессы ассимиляции (анаболизм) и диссимиляции (катаболизм). При ассимиляции (пластический обмен) происходит синтез сложных веществ из простых. Именно благодаря этому создаются все органические вещества в клетке, необходимые для построения ее структурных компонентов, ферментных систем и т. д. Ассимиляция всегда осуществляется с затратой энергии.
В ходе диссимиляции (энергетический обмен) сложные органические вещества расщепляются до более простых или до неорганических. При этом выделяется энергия, которая расходуется клеткой на выполнение различных процессов, обеспечивающих ее жизнедеятельность (синтез и транспорт веществ, механическую работу и т. д.).
Все живые организмы могут быть разделены на две группы: автотрофы и гетеротрофы, которые отличаются источниками энергии и необходимых веществ для обеспечения своей жизнедеятельности.
Автотрофы – организмы, синтезирующие из неорганических веществ органические соединения с использованием энергии солнечного света (как фототрофы – растения, цианобактерии) или энергии, получаемой при окислении минеральных (неорганических) веществ (таких, как хемотрофы – серобактерии, железобактерии и др.). Следовательно, они способны самостоятельно создавать требуемые для своей жизнедеятельности вещества.
§ 2. Диссимиляция у анаэробных и аэробных организмов
Организмы могут быть разделены на две группы и по характеру диссимиляции – аэробы и анаэробы. Аэробы (от греч. бес – воздух) нуждаются в свободном кислороде для жизнедеятельности. У анаэробов (греч. ац – отрицательная частица) в нем нет необходимости. У них диссимиляция осуществляется путем брожения – бескислородного, ферментативного расщепления органического вещества с образованием более простых органических же веществ и выделением энергии. Например:
♦ молочнокислое брожение:
C 6 H 12 O 6 + 2H 3 PO 4 + 2АДФ → 2Ф Н + 2АТФ + 2H 2 O;
♦ спиртовое брожение:
C 6 H 12 O 6 + 2Ф Н + 2АДФ → 2C 2 H 5 OH + 2CO 2 + 2АТФ + 2H 2 O.
Образующиеся при брожении вещества являются органическими и, следовательно, содержат еще много энергии.
Рис. 22. Взаимоотношения ассимиляиии и диссимиляции у автотрофных и гетеротрофных организмов
У аэробных организмов в процессе дыхания в митохондриях происходит полное расщепление органических веществ (при использовании О 2) до бедных энергией конечных продуктов СО 2 иН 2 О и высвобождается значительно большее количество энергии:
С 6 Н 12 0 6 (глюкоза) + 0 2 > 6С0 2 + 6Н 2 0 + энергия (за счет которой синтезируются 38 молекул АТФ).
Рассмотрим в виде обобщенных схем метаболизм у автотрофных и гетеротрофных аэробных организмов (рис. 22).
Ассимиляция. Ее важнейшие процессы – фотосинтез и биосинтез белков.
Связь организма с окружающей средой, с физико-химической точки зрения, представляет собой открытую систему, т. е. систему, где биохимические процессы идут постоянно. Исходные вещества поступают из окружающей среды, а вещества, образующиеся также непрерывно, выносятся вовне. Равновесие между скоростью и концентрацией продуктов разнонаправленных реакций в организме является условным, мнимым, т. к. поступление и вынос веществ не прекращаются. Непрерывная связь с окружающей средой и позволяет рассматривать живой организм как открытую систему.
Для всех живых клеток источником энергии является Солнце. Растительные клетки улавливают энергию солнечного света с помощью хлорофилла, используя ее для реакций ассимиляции в процессе фотосинтеза. Клетки животных, грибов, бактерий используют солнечную энергию косвенно, при расщеплении органических веществ, синтезированных земным растением.
Часть питательных веществ клетки расщепляется в процессе клеточного дыхания, поставляя таким образом энергию, необходимую для разного рода клеточной активности. Протекает этот процесс в органеллах, называемых митохондриями. Митохондрия состоит из двух мембран: наружной, отделяющей органеллу от цитоплазмы, и внутренней, образующей многочисленные складики. Главным продуктом дыхания является АТФ. Он покидает митохондрии и используется в качестве источника энергии для многих химических реакций в цитоплазме и клеточной мембране. Если для осуществления клеточного дыхания требуется кислород, то дыхание называют аэробным, если же реакции идут в отсутствие кислорода, то говорят об анаэробном дыхании.
Для любого вида работы, выполняемой в клетке, используется энергия в одной-единственной форме - в форме энергии фосфатных связей АТФ. АТФ - легко подвижное соединение. Образование АТФ происходит на внутренней мембране митохондрий. АТФ синтезируется во всех клетках при дыхании за счет энергии окисления углеводов, жиров и других органических веществ. В клетках зеленых растений основное количество АТФ синтезируется в хлоропластах за счет солнечной энергии. В них при фотосинтезе образуется во много раз больше АТФ, чем в митохондриях. Разлагается АТФ с разрывом фосфорно-кислородных связей и выделением энергии. Это происходит под действием фермента АТФазы в процессе гидролиза АТФ - присоединения воды с отщеплением молекулы фосфорной кислоты. В результате АТФ превращается в АДФ, а если отщепляются две молекулы фосфорной кислоты, то в АМФ. Реакция отщепления каждой грамм-молекулы кислоты сопровождается освобождением 40 кДж. Это очень большой выход энергии, поэтому фосфорно-кислородные связи АТФ принято называть макроэргитическими (высокоэнергетическими).
Использование АТФ в реакциях пластического обмена осуществляется путем их сопряжения с гидролизом АТФ. Молекулы разных веществ заряжаются энергией путем присоединения освобожденной при гидролизе фосфорной группы от молекулы АТФ, т. е. путем фосфорилирования.
Особенность фосфатных производных состоит в том, что они не могут покинуть клетку, хотя их «разряженные» формы свободно проходят через мембрану. Благодаря этому фосфорилированные молекулы остаются в клетке до тех пор, пока они не будут использованы в соответствующих реакциях.
Обратный процесс превращения АДФ в АТФ происходит путем присоединения молекулы фосфорной кислоты к АДФ с выделением воды и поглощением большого количества энергии.
Таким образом АТФ - универсальный и непосредственный источник энергии для деятельности клетки. Это создает единый клеточный фонд энергии и делает возможным ее перераспределение и транспортировку из одних участков клетки в другие.
Перенос фосфатной группы играет важную роль в химических реакциях типа сборки макромолекул из мономеров. Например, аминокислоты могут соединяться в пептиды, лишь будучи предварительно профосфорилированными. Механические процессы сокращения или движения, перенос растворенного вещества против градиента концентрации и другие процессы сопряжены с расходованием энергии, запасенной в АТФ.
Процесс энергетического обмена может быть представлен следующим образом. Высокомолекулярные органические вещества в цитоплазме ферментативно, путем гидролиза превращаются в более простые, из которых они состоят: белки - в аминокислоты, поли- и дисахариды - в моносахариды (+ глюкозу), жиры в глицерин и жирные кислоты. Окислительные процессы отсутствуют, освобождается мало энергии, которая не используется и переходит в тепловую форму. Большинство клеток в первую очередь тратят углеводы. Полисахариды (крахмал у растений и гликоген у животных) гидрализуются до глюкозы. Окисление глюкозы происходит в три фазы: гликолиз, окислительное декарбоксилирование (цикл Кребса - цикл лимонной кислоты) и окислительное фосфорилирование (дыхательная цепь). Гликолиз, в результате которого одна молекула глюкозы расщепляется на две молекулы пировиноградной кислоты с выделением двух молекул АТФ, протекает в цитоплазме. При отсутствии кислорода пировиноградная кислота превращается либо в этанол (брожение), либо молочную кислоту (анаэробное дыхание).
Если гликолиз осуществляется в клетках животных, шестиуглеродная молекула глюкозы распадается на две молекулы молочной кислоты. Процесс этот многоступенчатый. Его последовательно осуществляют 13 ферментов. При спиртовом брожении из молекулы глюкозы образуется две молекулы этанола и две молекулы СО2.
Гликолиз - фаза, общая для анаэробного и аэробного дыхания, две остальные осуществляются лишь в аэробных условиях. Процесс бескислородного окисления, при котором выделяется и используется лишь часть энергии метаболитов, для анаэробных организмов является конечным. В присутствии же кислорода пировиноградная кислота переходит в митохондрии, где в результате целого ряда последовательных реакций она полностью окисляется аэробным путем до Н2O и СО2 с одновременным фосфорилированием АДФ до АТФ. При этом две молекулы АТФ дает гликолиз, две - цикл Кребса, 34 - дыхательная цепь. Чистый выход при полном окислении одной молекулы глюкозы до Н2O и СО2 составляет 38 молекул.
Таким образом, у аэробных организмов окончательный распад органических веществ осуществляется путем окисления их кислородом воздуха до простых неорганических: СО2 и Н2O. Процесс этот протекает на кристах митохондрий. При этом выделяется максимальное количество свободной энергии, значительная часть которой резервируется в молекулах АТФ. Легко видеть, что аэробное окисление в наибольшей степени обеспечивает клетку свободной энергией.
В результате катаболизма в клетке накапливаются богатые энергией молекулы АТФ, а во внешнюю среду выделяется СО2 и избыточное количество воды.
Молекулы сахаров, не требующиеся для дыхания, могут запасаться в клетке. Избыточные липиды либо расщепляются, после чего продукты их расщепления поступают в митохондрии в качестве субстрата для дыхания, либо отлагаются про запас в цитоплазме в виде жировых капель. Из поступающих в клетку аминокислот строятся белки. Синтез белка происходит в органеллах, называемых рибосомами. Каждая рибосома состоит из двух субчастиц - большой и малой: в состав обеих субчастиц входят белковые молекулы и молекулы РНК.
Рибосомы часто бывают прикреплены к особой системе мембран, состоящей из цистерн и пузырьков, - к так называемому эндоплазматическому ретикулуму (ЭР); в клетках, вырабатывающих много белка, эндоплазматический ретикулум часто развит очень хорошо и весь усеян рибосомами. Некоторые ферменты эффективны лишь в том случае, если они прикреплены к мембране. Здесь находится большая часть ферментов, участвующих в синтезе липидов. Таким образом, эндоплазматический ретикулум - это как бы своеобразный рабочий стол клетки.
Кроме того, ЭР делит цитоплазму на отдельные отсеки, или компартменты, т. е. разобщает различные химические процессы, одновременно протекающие в цитоплазме, и тем самым снижает вероятность того, что эти процессы будут мешать друг другу.
Часто образуемые данной клеткой продукты используются вне этой клетки. В подобных случаях синтезированные на рибосомах белки проходят через мембраны эндоплазматического ретикулума и упаковываются в формирующиеся вокруг них мембранные пузырьки, которые затем отшнуровываются от ЭР. Эти пузырьки, уплощаясь и укладываясь друг на друга, как блины в стопке, образуют характерную структуру, называемую комплексом Гольджи, или аппаратом Гольджи. Во время своего пребывания в аппарате Гольджи, белки претерпевают определенные изменения. Когда для них наступает время покинуть клетку, мембранные пузырьки сливаются с клеточной мембраной и опорожняются, изливая свое содержимое наружу, т. е. секреция происходит путем экзоцитоза.
В аппарате Гольджи образуются также лизосомы - мембранные мешочки, содержащие пищеварительные ферменты. Выяснение того, как клетка производит, упаковывает и экспортирует некоторые белки, а также как она «узнает», какие белки ей следует сохранять для себя, составляет один из увлекательнейших разделов современной цитологии.
Мембраны любой клетки непрерывно перемещаются и видоизменяются. Мембраны ЭР медленно перемещаются по клетке. Отдельные участки этих мембран отделяются и образуют пузырьки, которые на время становятся частью аппарата Гольджи, а затем, в процессе экзоцитоза, сливаются с клеточной мембраной.
Позже мембранный материал возвращается в цитоплазму, где он используется вновь.
Обмен веществ, поступающих внутрь клетки или выделяющихся ею наружу, а также обмен разными сигналами с микро- и макроокружением происходит через наружную мембрану клетки. Как известно, клеточная мембрана является липидным бислоем, в который встроены различные белковые молекулы, выполняющие роль специализированных рецепторов, ионных каналов, устройств, которые активно переносят или удаляют различные химические вещества, межклеточных контактов и т. п. В здоровых клетках эукариотов фосфолипиды в мембране распределены асимметрично: наружная поверхность состоит из сфингомиелина и фосфатидилхолина, внутренняя - из фосфатидилсерина и фосфатидилэтаноламина. Поддержание такой асимметрии нуждается в затратах энергии. Поэтому в случае повреждения клетки, ее инфицирования, энергетического голодания происходит обогащение наружной поверхности мембраны несвойственными ей фосфолипидами, что становится сигналом для других клеток и ферментов о повреждении клетки с соответствующей реакцией на это. Наибольшую роль играет растворимая форма фосфолипазы A2, которая расщепляет арахидоновую кислоту и создает лизоформы из вышеназванных фосфолипидов. Арахидоновая кислота является лимитирующим звеном для создания таких медиаторов воспаления, как эйкозаноиды, а к лизоформам в мембране присоединяются защитные молекулы - пентраксины (С-реактивный протеин (СРП), предшественники амилоидных белков) - с последующей активацией системы комплемента по классическому пути и разрушением клетки.
Строение мембраны способствует сохранению особенностей внутренней среды клетки, ее отличий от внешней среды. Это обеспечивается выборочной проницаемостью клеточной мембраны, существованием в ней механизмов активного транспорта. Их нарушение в результате прямого повреждения, например, тетродотоксином, уабаином, тетраэтиламмонием, или в случае недостаточного энергетического обеспечения соответствующих “насосов” приводит к нарушению электролитного состава клетки, изменению в ней метаболизма, нарушению специфических функций - сокращения, проведения импульса возбуждения и т. п. Нарушение клеточных ионных каналов (кальциевых, натриевых, калиевых и хлорных) у человека также может быть генетически обусловлено мутацией генов, отвечающих за структуру этих каналов. Tак называемые каналопатии служат причиной наследственных болезней нервной, мышечной, пищеварительной систем. Избыточное поступление внутрь клетки воды может привести к ее разрыву - цитолизу - вследствие перфорации мембраны при активации комплемента или атаки цитотоксических лимфоцитов и натуральных киллеров.
В клеточную мембрану встроено множество рецепторов - структур, которые в случае объединения с соответствующими специфическими сигнальными молекулами (лигандами) передают сигнал внутрь клетки. Это происходит с помощью различных регуляторных каскадов, состоящих из ферментативно активных молекул, которые последовательно активируются и в конечном итоге способствуют реализации различных клеточных программ, таких как рост и пролиферация, дифференцировка, подвижность, старение, гибель клетки. Регуляторные каскады достаточно многочисленны, но их количество до конца еще не определено. Система рецепторов и объединенных с ними регуляторных каскадов существует и внутри клетки; они создают определенную регуляторную сеть с пунктами концентрации, распределения и выбора дальнейшего пути сигнала в зависимости от функционального состояния клетки, этапа ее развития, одновременного действия сигналов из других рецепторов. Результатом этого может быть торможение или усиление сигнала, направление его по другому регуляторному пути. Как рецепторный аппарат, так и пути передачи сигнала через регуляторные каскады, например к ядру, могут нарушаться в результате генетического дефекта, который возникает как врожденный порок на уровне организма или вследствие соматической мутации в определенном типе клеток. Указанные механизмы могут повреждаться инфекционными агентами, токсинами, а также изменяться в процессе старения. Завершающим этапом этого может быть нарушение функций клетки, процессов ее пролиферации и дифференцировки.
На поверхности клеток также расположены молекулы, играющие важную роль в процессах межклеточного взаимодействия. К ним могут относиться белки клеточной адгезии, антигены тканевой совместимости, тканевоспецифические, дифференцирующие антигены и т. п. Изменения в составе этих молекул обусловливают нарушение межклеточных взаимодействий и могут стать причиной включения соответствующих механизмов элиминации таких клеток, ведь они представляют определенную опасность для целостности организма как резервуар инфекции, особенно вирусной, или как потенциальные инициаторы опухолевого роста.
Нарушение энергетического обеспечения клетки
Источником энергии в клетке служат продукты питания, после расщепления которых до конечных веществ выделяется энергия. Главным местом образования энергии являются митохондрии, в которых вещества окисляются с помощью ферментов дыхательной цепи. Окисление - основной поставщик энергии, поскольку в результате гликолиза из такого же количества субстратов окисления (глюкозы) высвобождается, по сравнению с окислением, не больше 5 % энергии. Около 60 % энергии, высвобождаемой при окислении, аккумулируется путем окислительного фосфорилирования в макроэргических фосфатах (АТФ, креатинфосфат), остальное ее количество рассеивается в виде тепла. В дальнейшем макроэргические фосфаты используются клеткой для таких процессов, как работа насосов, синтез, деление, движение, секреция и др. Выделяют три механизма, повреждение которых может вызвать нарушение обеспечения клетки энергией: первый - механизм синтеза ферментов энергетического обмена, второй - механизм окислительного фосфорилирования, третий - механизм использования энергии.
Нарушение электронного транспорта в дыхательной цепи митохондрий или разобщение окисления и фосфорилирования АДФ при потере протонного потенциала - движущей силы генерации АТФ, приводит к ослаблению окислительного фосфорилирования таким образом, что большая часть энергии рассеивается в виде тепла и уменьшается количество макроэргических соединений. Разобщение окисления и фосфорилирования под влиянием адреналина используется клетками гомойотермных организмов для повышения теплопродукции при поддержании постоянной температуры тела во время охлаждения или ее повышении при лихорадке. Значительные изменения в структуре митохондрий и энергетическом метаболизме наблюдаются при тиреотоксикозе. Эти изменения вначале обратимы, но после определенной черты становятся необратимыми: митохондрии фрагментируются, распадаются или набухают, теряют кристы, превращаясь в вакуоли, и в конце концов накапливают такие вещества, как гиалин, ферритин, кальций, липофусцин. У больных цингой происходит слияние митохондрий с образованием хондриосфер, возможно, вследствие повреждения мембран пероксидными соединениями. Значительные повреждения митохондрий возникают под действием ионизирующего излучения, в ходе преобразования нормальной клетки в злокачественную.
Митохондрии являются мощным депо ионов кальция, где его концентрация на несколько порядков превышает таковую в цитоплазме. При повреждении митохондрий кальций выходит в цитоплазму, обусловливая активизацию протеиназ с повреждением внутриклеточных структур и нарушением функций соответствующей клетки, например, кальциевые контрактуры или даже “кальциевую смерть” в нейронах. В результате нарушения функциональной способности митохондрий резко увеличивается образование свободнорадикальных пероксидных соединений, которые имеют очень высокую реакционную способность и поэтому повреждают важные компоненты клетки - нуклеиновые кислоты, белки и липиды. Это явление наблюдается при так называемом оксидационном стрессе и может иметь отрицательные последствия для существования клетки. Так, повреждение наружной мембраны митохондрии сопровождается выходом в цитоплазму веществ, содержащихся в межмембранном пространстве, прежде всего цитохрома С и некоторых других БАВ, которые запускают цепные реакции, являющиеся причиной запрограммированной гибели клетки - апоптоза. Повреждая ДНК митохондрий, свободнорадикальные реакции искажают генетическую информацию, необходимую для образования некоторых ферментов дыхательной цепи, которые продуцируются именно в митохондриях. Это приводит к еще большему нарушению окислительных процессов. В целом собственный генетический аппарат митохондрий по сравнению с генетическим аппаратом ядра хуже защищен от вредных влияний, способных изменять закодированную в нем генетическую информацию. Как результат - возникает нарушение функций митохондрий на протяжении жизни, например, в процессе старения, при злокачественной трансформации клетки, а также на фоне наследственных митохондриальных болезней, связанных с мутацией ДНК митохондрий в яйцеклетке. В настоящее время описаны свыше 50 митохондриальных мутаций, вызывающих наследственные дегенеративные болезни нервной и мышечной систем. Они передаются ребенку исключительно от матери, так как митохондрии сперматозоида не входят в состав зиготы и, соответственно, нового организма.
Нарушение сохранения и передачи генетической информации
Ядро клетки содержит большую часть генетической информации и тем самым обеспечивает ее нормальное функционирование. С помощью выборочной экспрессии генов оно координирует работу клетки в интерфазе, сохраняет генетическую информацию, воссоздает и передает генетический материал в процессе деления клеток. В ядре происходят репликация ДНК и транскрипция РНК. Различные патогенные факторы, такие как ультрафиолетовое и ионизирующее излучение, свободнорадикальное окисление, химические вещества, вирусы, способны повреждать ДНК. Подсчитано, что каждая клетка теплокровного животного за 1 сут. теряет более 10 000 оснований. Сюда следует добавить нарушения при копировании во время деления. При сохранении этих повреждений клетка была бы не способна выжить. Защита заключается в существовании мощных систем репарации, таких как ультрафиолетовая эндонуклеаза, система репаративной репликации и рекомбинационного восстановления, которые замещают нарушения ДНК. Генетические дефекты в репаративных системах вызывают развитие болезней, обусловленных повышенной чувствительностью к факторам, повреждающим ДНК. Это пигментная ксеродерма, а также некоторые синдромы ускоренного старения, сопровождающиеся повышенной склонностью к возникновению злокачественных опухолей.
Система регуляции процессов репликации ДНК, транскрипции информационой РНК (иРНК), трансляции генетической информации из нуклеиновых кислот в структуру белков довольно сложная и многоуровневая. Кроме регуляторных каскадов, запускающих действие факгоров транскрипции общим количеством свыше 3000, которые активируют определенные гены, существует еще многоуровневая регуляторная система, опосредованная малыми молекулами РНК (интерферирующие РНК; РНКи). Геном человека, который состоит приблизительно из 3 млрд пуриновых и пиримидиновых оснований, содержит лишь 2 % структурных генов, отвечающих за синтез белков. Остальные обеспечивают синтез регуляторных РНК, которые одновременно с факторами транскрипции активируют или блокируют работу структурных генов на уровне ДНК в хромосомах или влияют на процессы трансляции матричной РНК (мРНК) при образовании молекулы полипептида в цитоплазме. Нарушение генетической информации может происходить как на уровне структурных генов, так и регуляторной части ДНК с соответствующими проявлениями в виде разнообразных наследственных заболеваний.
В последнее время большое внимание привлекают изменения в генетическом материале, которые происходят в процессе индивидуального развития организма и связаны с торможением или активацией определенных участков ДНК и хромосом вследствие их метилирования, ацетилирования и фосфорилирования. Эти изменения сохраняются длительно, иногда - на протяжении всей жизни организма от эмбриогенеза до старости, и получили название эпигеномной наследственности.
Размножению клеток с измененной генетической информацией препятствуют также системы (факторы) контроля митотического цикла. Они взаимодействуют с циклинзависимыми протеинкиназами и их каталитическими субъединицами - циклинами - и блокируют прохождение клеткой полного митотического цикла, останавливая деление на границе между пресинтетической и синтетической фазами (блок G1/S) вплоть до завершения репарации ДНК, а в случае ее невозможности - инициируют запрограммированную смерть клетки. К таким факторам относится ген р53, мутация которого служит причиной потери контроля над пролиферацией трансформированных клеток; она наблюдается почти в 50 % случаев рака у человека. Второй контрольный пункт прохождения митотического цикла находится на границе G2/M. Здесь контролируется правильность распределения хромосомного материала между дочерними клетками в митозе или мейозе с помощью комплекса механизмов, контролирующих клеточное веретено, центр и центромеры (кинетохоры). Неэффективность этих механизмов приводит к нарушению распределения хромосом или их частей, что проявляется отсутствием какой-либо хромосомы в одной из дочерних клеток (анеуплоидия), наличием лишней хромосомы (полиплоидия), отрывом части хромосомы (делеция) и переносом ее на другую хромосому (транслокация). Такие процессы очень часто наблюдаются при размножении злокачественно перерожденных и трансформированных клеток. Если же это происходит во время мейоза с половыми клетками, то приводит или к гибели плода на раннем этапе эмбрионального развития, или к рождению организма с хромосомной болезнью.
Неконтролируемое размножение клеток во время роста опухолей возникает как результат мутации в генах, которые контролируют пролиферацию клеток и получили название онкогенов. Среди более 70 известных в настоящее время онкогенов большая часть относится к компонентам регуляции роста клетки, часть представлена факторами транскрипции, регулирующими активность генов, а также факторами, которые тормозят деление и рост клеток. Еще одним фактором, ограничивающим чрезмерную экспансию (распространение) пролиферирующих клеток, является укорочение концов хромосом - теломер, которые не способны в результате сугубо стерического взаимодействия полностью реплицироваться, поэтому после каждого деления клетки теломеры укорачиваются на определенную часть оснований. Таким образом, пролиферирующие клетки взрослого организма после определенного количества делений (обычно от 20 до 100 в зависимости от вида организма и его возраста) исчерпывают длину теломеры и дальнейшая репликация хромосом прекращается. Это явление не возникает в сперматозогенном эпителии, энтероцитах и эмбриональных клетках благодаря наличию фермента теломеразы, восстанавливающей длину теломер после каждого деления. В большинстве клеток взрослых организмов теломераза заблокирована, но, к сожалению, она активирована в клетках опухолей.
Связь между ядром и цитоплазмой, транспорт веществ в обоих направлениях осуществляются через поры в ядерной мембране при участии специальных транспортных систем с потреблением энергии. Таким образом транспортируются к ядру энергетические и пластические вещества, сигнальные молекулы (факторы транскрипции). Обратный поток выносит в цитоплазму молекулы иРНК и транспортной РНК (тРНК), рибосомы, необходимые для синтеза белка в клетке. Этот же путь транспорта веществ присущ и вирусам, в частности таким, как ВИЧ. Они переносят свой генетический материал в ядро клетки-хозяина с дальнейшим включением его в геном хозяина и переносом новообразованной вирусной РНК в цитоплазму для дальнейшего синтеза белков новых вирусных частичек.
Нарушение процессов синтеза
Процессы синтеза белков происходят в цистернах эндоплазматической сети, тесно связанных с порами в ядерной мембране, через которые в эндоплазматическую сеть поступают рибосомы, тРНК и иРНК. Здесь осуществляется синтез полипептидных цепочек, которые в дальнейшем приобретают свой окончательный вид в агранулярной эндоплазматической сети и пластинчатом комплексе (комплексе Гольджи), где подвергаются пос трансляционной модификации и соединению с молекулами углеводов и липидов. Новообразованные белковые молекулы не остаются на месте синтеза, а с помощью сложного регулируемого процесса, который носит название протеинкинезиса , активно переносятся к той изолированной части клетки, где они будут выполнять предназначенную им функцию. При этом очень важным этапом является структурирование перенесенной молекулы в соответствующую пространственную конфигурацию, способную выполнять присущую ей функцию. Такое структурирование происходит с помощью специальных ферментов или на матрице специализированных белковых молекул - шаперонов, которые помогают белковой молекуле, новообразованной или измененной вследствие внешнего влияния, приобрести правильную трехмерную структуру. В случае неблагоприятного влияния на клетку, когда возникает вероятность нарушения структуры белковых молекул (например, при повышении температуры тела, инфекционном процессе, интоксикации) концентрация шаперонов в клетке резко увеличивается. Поэтому такие молекулы получили еще название стресс-белков , или белков теплового шока . Нарушение структурирования белковой молекулы приводит к образованию химически инертных конгломератов, которые откладываются в клетке или вне ее при амилоидозе, болезни Альцгеймера и др. Иногда матрицей может служить предварительно структурированная аналогичная молекула, и в данном случае, если первичное структурирование произошло неправильно, все последующие молекулы также будут дефектными. Эта ситуация возникает при так называемых прионовых болезнях (скрепи у овец, бешенство коров, куру, болезнь Крейтцфельдта-Якоба у человека), когда дефект одного из мембранных белков нервной клетки обусловливает последующее скопление инертных масс внутри клетки и нарушение ее жизнедеятельности.
Нарушение процессов синтеза в клетке может происходить на различных его этапах: транскрипция РНК в ядре, трансляция полипептидов в рибосомах, посттрансляционная модификация, гиперметилирование и гликозилирование бежовой молекулы, транспорт и распределение белков в клетке и выведение их наружу. При этом можно наблюдать увеличение или уменьшение количества рибосом, распад полирибосом, расширение цистерн гранулярной эндоплазматической сети, потерю ею рибосом, образование везикул и вакуолей. Так, при отравлении бледной поганкой повреждается фермент РНК-полимераза, что нарушает транскрипцию. Дифтерийный токсин, инактивируя фактор элонгации, нарушает процессы трансляции, обусловливая повреждение миокарда. Причиной нарушения синтеза некоторых специфических белковых молекул могут служить инфекционные агенты. Например, герпесвирусы тормозят синтез и экспрессию молекул антигенов ГКГС, что позволяет им частично избежать иммунного контроля, бациллы чумы - синтез медиаторов острого воспаления. Появление необычных белков может приостанавливать их дальнейший распад и приводить к накоплению инертного или даже токсичного материала. Этому в определенной мере может способствовать и нарушение процессов распада.
Нарушение процессов распада
Одновременно с синтезом белка в клетке непрерывно происходит его распад. В нормальных условиях это имеет важное регуляторное и формообразующее значение, например, во время активации неактивных форм ферментов, белковых гормонов, белков митотического цикла. Нормальные рост и развитие клетки нуждаются в тонко контролируемом балансе между синтезом и деградацией белков и органелл. Однако в процессе синтеза белков вследствие ошибок в работе синтезирующего аппарата, аномального структурирования белковой молекулы, ее повреждения химическими и бактериальными агентами постоянно образуется довольно большое количество дефектных молекул. По некоторым оценкам, их доля составляет около трети всех синтезированных белков.
Клетки млекопитающих имеют несколько главных путей разрушения белков: через лизосомальные протеазы (пентидгидролазы), кальцийзависимые протеиназы (эндопептидазы) и систему протеасом. Кроме того, есть еще и специализированные протеиназы, например каспазы. Основной органеллой, в которой происходит деградация веществ в эукариотических клетках, является лизосома, содержащая многочисленные гидролитические ферменты. Вследствие процессов эндоцитоза и различных типов аутофагии в лизосомах и фаголизосомах разрушаются как дефектные белковые молекулы, так и целые органеллы: поврежденные митохондрии, участки плазматической мембраны, некоторые экстрацеллюлярные белки, содержимое секреторных гранул.
Важным механизмом деградации белков является протеасома - мультиката-литическая протеиназная структура сложного строения, локализирующаяся в цитозоле, ядре, эндоплазматической сети и на мембране клетки. Эта ферментная система отвечает за разрушение поврежденных белков, а также здоровых белков, которые должны быть удалены для нормального функционирования клетки. При этом белки, подлежащие разрушению, предварительно соединяются со специфическим полипептидом убиквитином. Однако частично в протеасомах могуг разрушаться и неубиквитированные белки. Распад белковой молекулы в протеасомах до коротких полипептидов (процессинг) с последующей их презентацией совместно с молекулами ГКГС I типа является важным звеном в осуществлении иммунного контроля антигенного гомеостаза организма. При ослаблении функции протеасом происходит аккумуляция поврежденных и ненужных белков, сопровождающая старение клетки. Нарушение деградации циклинзависимых белков приводит к нарушению клеточного деления, деградации секреторных белков - к развитию цистофиброза. И наоборот, повышение функции протеасом сопровождает истощение организма (СПИД, рак).
При генетически обусловленных нарушениях деградации белков организм нежизнеспособен и гибнет на ранних стадиях эмбриогенеза. Если же нарушается распад жиров или углеводов, то возникают болезни накопления (тезаурисмозы). При этом внутри клетки накапливается избыточное количество определенных веществ или продуктов их неполного распада - липидов, полисахаридов, что существенно повреждает функцию клетки. Чаще всего это наблюдается в эпите-лиоцитах печени (гепатоцитах), нейронах, фибробластах и макрофагоцитах.
Приобретенные нарушения процессов распада веществ могут возникать как результат патологических процессов (например, белковой, жировой, углеводной и пигментной дистрофии) и сопровождаться образованием непривычных веществ. Нарушения в системе лизосомного протеолиза приводят к снижению адаптации при голодании или повышенной нагрузке, к возникновению некоторых эндокринных дисфункций - снижению уровня инсулина, тиреоглобулина, цитокинов и их рецепторов. Нарушения деградации белков замедляют скорость заживления ран, обусловливают развитие атеросклероза, влияют на иммунный ответ. При гипоксии, изменении внутриклеточного pH, лучевом поражении, характеризующихся усиленной пероксидацией мембранных липидов, а также под влиянием лизосомотропных веществ - эндотоксинов бактерий, метаболитов токсических грибов (спорофузарин), кристаллов оксида кремния - изменяется стабильность мембраны лизосом, в цитоплазму высвобождаются акгивированные лизосомаль-ные ферменты, что вызывает разрушение структур клетки и ее гибель.
Глава 1
ОСНОВЫ КЛЕТОЧНОЙ ФИЗИОЛОГИИ
И. Дудель
Плазматическая мембрана . Животные клетки ограничены плазматической мембраной (рис. 1.1). На ее строении, очень сходном со строением многих внутриклеточных мембран, мы остановимся несколько подробнее. Основной матрикс мембраны состоит из липидов, главным образом фосфатидил–холина. Эти липиды состоят из головной гидрофильной группы, к которой присоединены длинные гидрофобные углеводородные цепи. В воде такие липиды спонтанно формируют двуслойную пленку толщиной 4–5 нм, в которой гидрофильные группы обращены к водной среде, а гидрофобные углеводородные цепи располагаются в два ряда, образуя безводную липидную фазу. Клеточные мембраны представляют собой липидные бислои именно такого типа и содержат гликолипиды, холестерол и фосфолипиды (рис. 1.2). Гидрофильная часть гликолипидов образована олигосахаридами. Гликолипиды всегда располагаются на наружной поверхности плазматической мембраны, причем олигосахаридная часть молекулы ориентирована подобно волоску, погруженному в окружающую среду. Разбросанные среди фосфолипидов в почти равном с ними количестве молекулы холестерола стабилизируют мембрану. Распределение различных липидов во внутреннем и наружном слоях мембраны неодинаково, и даже в пределах одного слоя имеются участки, в которых концентрируются отдельные виды липидов. Такое неравномерное распределение,
Рис. 1.1 . Схематический рисунок клетки, на котором изображены наиболее важные органеллы
вероятно, имеет какое–то, пока еще неясное, функциональное значение.
Главными функциональными элементами, погруженными в сравнительно инертный липидный матрикс мембраны, являются белки (рис. 1.2). Белок по массе составляет от 25 до 75% в различных мембранах, но, поскольку белковые молекулы намного крупнее, чем липидные, 50% по массе эквивалентны соотношению: 1 молекула белка на 50 молекул липида. Одни белки пронизывают мембрану от ее наружной до внутренней поверхности, другие же закреплены в каком–то одном слое. Белковые молекулы обычно ориентированы так, что их гидрофобные группы погружены в липидную мембрану, а полярные гидрофильные группы на поверхности мембраны погружены в водную фазу. Многие белки наружной поверхности мембраны представляют собой гликопротеины; их гидрофильные сахаридные группы обращены во внеклеточную среду.
Мембранные системы внутриклеточных органелл .
Примерно половина объема клетки занята органеллами, изолированными от цитозоля мембранами. Общая поверхность мембран внутриклеточных органелл по крайней мере в 10 раз превышает поверхность плазматической мембраны. Наиболее широко распространенная мембранная система – эндоплазматический ретикулум, представляющий собой сеть

Рис. 1.2. Схематическое изображение плазматической мембраны. Белки погружены в фосфолипидный бислой, причем некоторые из них пронизывают бислой, тогда как другие только заякорены на наружном или внутреннем слое
сильно извитых трубочек или мешковидных вытянутых структур; большие участки эндоплазматического ретикулума усеяны рибосомами; такой ретикулум получил название гранулярного, или шероховатого (рис. 1.1). Аппарат Гольджи также состоит из мембрансвязных ламелл, от которых отрываются пузырьки, или везикулы (рис. 1.1). Лизосомы и пероксисомы – это небольшие специализированные везикулы. Во всех этих разнообразных органеллах мембрана и охватываемое ею пространство содержат специфические наборы ферментов; внутри органелл накапливаются особые продукты обмена, используемые для осуществления различных функций органелл .
Ядро и митохондрии отличаются тем, что каждая из этих органелл окружена двумя мембранами. Ядро ответственно за кинетический контроль метаболизма; складчатая внутренняя мембрана митохондрии – место окислительного метаболизма; здесь за счет окисления пирувата или жирных кислот синтезируется высокоэнергетическое соединение аденозинтрифосфат (АТФ, или АТР).
Цитоскелет . Цитоплазму, окружающую органеллы, никоим образом нельзя считать аморфной; ее пронизывает сеть цитоскелета. Цитоскелет состоит из микротрубочек, актиновых филаментов и промежуточных филаментов (рис. 1.1). Микротрубочки имеют наружный диаметр около 25 нм; они образуются, как и обычный полимер, в результате сборки молекул белка тубулина. Актиновые филаменты– сократительные волокна, располагающиеся в примембранном слое и во всей клетке– в основном принимают участие в процессах, связанных с движением. Промежуточные филаменты состоят из блоков различного химического состава в разных типах клеток; они образуют разнообразные связи между упомянутыми выше двумя другими элементами цитоскелета. Органеллы и плазматическая мембрана также связаны с цитоскелетом, который не только обеспечивает поддержание формы клетки и положение органелл в ней, но и обусловливает изменение формы клетки и ее подвижность.
Цитозоль . Около половины объема клетки занято цитозолем. Поскольку он приблизительно на 20% (по массе) состоит из белка, это скорее гель, чем водный раствор. Малые молекулы, в том числе органические и неорганические ионы, растворены в водной фазе. Между клеткой и окружающей средой (внеклеточным пространством) происходит обмен ионов; эти обменные процессы будут рассмотрены в следующем разделе. Концентрация ионов во внеклеточном пространстве поддерживается со значительной точностью на постоянном уровне; внутриклеточная концентрация каждого из ионов также имеет специфический уровень, отличающийся от такового вне клетки (табл. 1.1). Наиболее распространенным катионом во внеклеточной среде является Na + в клетке его концентрация более чем в 10 раз ниже. Напротив, внутри клетки наиболее высока концентрация К + , вне клетки она ниже более чем на порядок. Наибольший градиент между внеклеточной и внутриклеточной концентрациями существует для Са 2+ концентрация свободных ионов которого внутри клетки по крайней мере в 10000 раз ниже, чем вне ее. Не все ионы растворены в цитозоле, часть их адсорбирована на белках или депонирована в органеллах. Например, в случае Са 2+ связанные ионы намного более многочисленны, чем свободные. Большая часть белков цитозоля – ферменты, при участии которых осуществляется множество процессов промежуточного метаболизма: гликолиз и глюконеогенез, синтез или разрушение аминокислот, синтез белка на рибосомах (рис. 1.1). В цитозоле содержатся также капли липидов и гранулы гликогена, служащие резервами важных молекул.
Таблица 1.1. Внутри– и внеклеточные концентрации ионов в мышечных клетках гомойотермных животных. А – –«высокомолекулярные клеточные анионы»
|
Внутриклеточная концентрация |
Внеклеточная концентрация |
||
|
Na + |
12 ммоль/л |
Na + |
145 ммоль/л |
|
155 ммоль/л |
К + |
4 ммоль/л |
|
|
Са 2+ |
10 –7 10 –8 ммоль/л |
Са 2+ |
2 ммоль/л |
|
С l – |
4 ммоль/л |
С1 – |
120 ммоль/л |
|
HCO 3 – |
8 ммоль/л |
HCO 3 – |
27 ммоль/л |
|
A – |
155 ммоль/л |
Прочие катионы |
5 ммоль/л |
|
Потенциал покоя –90 мВ |
|||
1.2. Обмен веществами между клеткой и окружающей средой
Мы кратко описали строение клетки, чтобы использовать это описание для рассмотрения основ клеточной физиологии. Клетку ни в коем случае нельзя считать статичным образованием, поскольку здесь происходит постоянный обмен веществами между различными внутриклеточными компартментами, а также между компартментами и окружающей средой. Структуры клетки находятся в динамическом равновесии, и взаимодействия клеток между собой и с внешней средой являются необходимым условием для поддержания жизни функционирующего организма. В данной главе мы рассмотрим фундаментальные механизмы такого обмена. В последующих главах эти механизмы будут рассмотрены в приложении к нервной клетке и ее функциям;
однако те же самые механизмы лежат в основе функционирования всех других органов.
Диффузия. Простейшим процессом перемещения вещества является диффузия. В растворах (или газах) атомы и молекулы перемещаются свободно, а разность концентраций уравновешивается благодаря диффузии. Рассмотрим два объема, заполненные жидкостью или газом (рис. 1.3), в которых вещества имеют концентрации c 1 и c 2 и разделены слоем с площадью поверхности А и толщиной d . Поток вещества m за время t описывается первым законом диффузии Фика:
dm / dt = D A / d ( C 1 –С 2)= D A / d D C (1)
где D–коэффициент диффузии, постоянный для данного вещества, растворителя и температуры. В более общем виде для разности концентраций dc на расстоянии dx
dm/dt= –D A dc/dx,(2)
поток через сечение А пропорционален градиенту концентрации dc / dx . Знак «минус» возникает в уравнении потому, что изменение концентрации по направлению х–отрицательно.
Диффузия–это наиболее важный процесс, благодаря которому большая часть молекул в водных растворах перемещается на небольшие расстояния. Это относится и к их движению в клетке постольку, поскольку диффузии не препятствуют мембраны. Множество веществ могут свободно диффундировать через липидные мембраны, в особенности вода и растворенные газы, такие, как О 2 и СО 2 . Жирорастворимые

Рис. 1.3. Количественная схема диффузии. Два пространства разделены слоем толщиной d и площадью А. С;–высокая концентрация частиц в левой части объема, С:,–низкая концентрация частиц в правой части, розовая поверхность –концентрационный градиент в диффузионном слое. Диффузионный поток dm/dt–см. уравнение (1)
вещества также хорошо диффундируют через мембраны; это касается и полярных молекул достаточно малого размера, таких, как этанол и мочевина, тогда как сахара проходят через липидный слой с трудом. Вместе с тем липидные слои практически непроницаемы для заряженных молекул, в том числе даже для неорганических ионов. Для неэлектролитов уравнение диффузии (1) обычно преобразуют, соединяя характеристки мембраны и диффундирующего вещества в один параметр–проницаемость (Р):
dm/dt=P A D c. (3)
На рис. 1.4 сравниваются проницаемости (Р) липидной мембраны для различных молекул.
Диффузия через мембранные поры . Плазматическая мембрана (и другие клеточные мембраны) проницаемы не только для веществ, диффундирующих через липидный слой, но и для многих ионов, сахаров, аминокислот и нуклеотидов. Эти вещества преодолевают мембрану через поры, образованные транспортными белками, погруженными в мембрану. Внутри таких белков имеется заполненный водой канал диаметром менее 1 нм, через который могут диффундировать малые молекулы. Они движутся по градиенту концентрации, и если они несут заряд, то их движение по каналам регулируется также мембранным потенциалом. Мембранные каналы обладают относительной избирательностью

Рис. 1.4. Проницаемость искусственных липидных бислоев для различных веществ
по отношению к типу молекул, которые могут через них проходить, Существуют, например, калиевые, натриевые и кальциевые каналы, каждый из которых непроницаем практически для любого иона, кроме специфического. Такая избирательность обусловлена зарядом или структурой мест связывания в стенках канала, что облегчает транспорт специфической молекулы и предотвращает проникновение через канал других веществ (рис. 1,5, А) .
За поведением мембранных ионных каналов легко наблюдать, поскольку возникающий при движении ионов ток можно измерить, причем даже для одиночного канала. Показано, что каналы спонтанно и с высокой частотой меняют свое состояние от открытого к закрытому. Для калиевого канала характерны импульсы тока амплитудой около 2 пА (2 10 –12 А) и длительностью в несколько миллисекунд (см. рис. 2.12, стр. 37) [З]. За этот период через него проходят десятки тысяч ионов. Переход белков из одной конформации в другую изучают методами рентгеновской дифракции, мессбауэровской спектроскопии и ядерно–магнитного резонанса (ЯМР). Белки, таким образом, являются очень динамичными подвижными структурами, а канал, проходящий через белок, не просто жесткая, наполненная водой трубка (рис. 1.5, А), но лабиринт быстро двигающихся молекулярных групп и зарядов. Эта динамическая характеристика канала отражается в энергетическом профиле канала, показанном на рис. 1.5, Б. Здесь по оси абсцисс представлена длина канала от внешнего раствора с концентрацией ионов С 0 и потенциалом 0 до внутреннего раствора с концентрацией С 1 и потенциалом Е. По оси ординат

Рис. 1.5. А. Схема белка, образующего калиевый канал, погруженный в липидный бислой плазматической мембраны. Четыре отрицательных заряда зафиксированы на «стенке» канала. Б. Схематический энергетический профиль канала, показанного на рис. А. По оси ординат отложены величины кинетической энергии, необходимые для прохождения канала; по оси абсцисс –расстояния между внутренней и наружной поверхностями мембраны. Энергетические минимумы соответствуют местам связывания положительно заряженных ионов с фиксированными отрицательными зарядами в стенке канала. Энергетические максимумы соответствуют препятствиям диффузии в канале. Предполагается, что конформация канального белка спонтанно осциллирует; варианты энергетических профилей изображены сплошными и штриховыми линиями; эти осцилляции в значительной степени облегчают связывание ионов при преодолении энергетического барьера (по с изменениями)
представлены уровни энергии иона в местах связывания канала; пик на графике представляет барьер проницаемости, который энергия иона должна превосходить для проникновения через канал, а «провал» графика–сравнительно стабильное состояние (связывание). Несмотря на препятствие в виде энергетического пика, ион может проникнуть через канал, если энергетический профиль спонтанно циклически меняется; ион, таким образом, может внезапно оказаться «по ту сторону» энергетического пика и может продолжить движение в клетку. В зависимости от заряда, размеров и степени гидратации иона и его способности связываться со структурами стенок канала энергетический профиль канала варьирует для различных ионов, чем может объясняться избирательность отдельных типов каналов.
Диффузионное равновесие ионов . Диффузия различных ионов через мембранные каналы должна была бы привести к устранению различий в концентрациях между вне– и внутриклеточной средами. Как, однако, видно из табл. 1.1, такие различия сохраняются, следовательно, должно существовать некое равновесие между диффузией и другими процессами транспорта через мембрану. Следующие два раздела касаются тех путей, с помощью которых такое равновесие устанавливается. В случае ионов на диффузионное равновесие влияет их заряд. Диффузия незаряженных молекул обеспечивается разностью концентраций dc , и, когда концентрации уравниваются, собственно транспорт прекращается. На заряженные частицы дополнительно влияет электрическое поле. Например, когда ион калия выходит из клетки по градиенту своей концентрации, он несет один положительный заряд. Таким образом, внутриклеточная среда становится более отрицательно заряженной, в результате чего на мембране возникает разность потенциалов. Внутриклеточный отрицательный заряд препятствует выходу из клетки новых ионов калия, а те ионы, которые все же выйдут из клетки, еще больше увеличат заряд на мембране. Поток ионов калия прекращается, когда действие электрического поля компенсирует диффузионное давление вследствие разности концентраций. Ионы продолжают проходить через мембрану, но в равных количествах в обоих направлениях. Следовательно, для данной разности концентраций ионов на мембране существует равновесный потенциал Е ion при котором поток ионов через мембрану прекращается. Равновесный потенциал может быть легко определен с помощью уравнения Нернста:
Е ion = R T / z F * ln C out / C in (4)
где R –газовая постоянная, Т–абсолютная температура, z – валентность иона (отрицательная для анионов) C out – внеклеточная концентрация иона, C in –внутриклеточная концентрация иона, F число Фарадея. Если подставить в уравнение константы, то при температуре тела (Т = 310 К) равновесный потенциал для ионов калия Е K равен:
Ек = –61 м B log /(5)
Если [ K + out ]/[ K + in ] = 39, как это следует из табл. 1.1, тогда
Ек= –61 м B log 39= –97 мВ.
Действительно, было обнаружено, что все клетки имеют мембранный потенциал; в мышечных клетках млекопитающих его уровень составляет около -90 мВ. В зависимости от условий и относительных концентраций ионов клетки могут иметь мембранный потенциал в пределах от -40 до -120мВ. Для клетки в приведенном выше примере (табл. 1.1) потенциал покоя, равный примерно -90 мВ, показывает, что потоки ионов калия через мембранные каналы находятся приблизительно в равновесии. Это неудивительно, поскольку в покоящейся мембране открытое состояние калиевых каналов наиболее вероятно, т.е. мембрана наиболее проницаема для ионов калия. Мембранный потенциал, однако, определяется потоками и других ионов.
Легкость, с которой незаряженные частицы могут диффундировать через мембрану, количественно представлена в уравнении (3). Проницаемость для заряженных частиц описывается несколько более сложным уравнением:
P = m R T / dF (6)
где m –подвижность иона в мембране, d –толщина мембраны, a R , Т и F –известные термодинамические постоянные. Определенные таким образом значения проницаемости для различных ионов могут быть использованы для расчета мембранного потенциала Em , когда ионы калия, натрия и хлора проходят через мембрану одновременно (с проницаемостью Р K , Р Na и P Cl соответственно). При этом предполагается, что потенциал падает в мембране равномерно, так что напряженность поля постоянна. В этом случае применяется уравнение Голдмана, или уравнение постоянного поля :
Е m= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl )(7)
Для большинства клеточных мембран Р K приблизительно в 30 раз выше, чем Р Na (см. также разд. 1.3). Относительная величина P Cl сильно варьирует; для многих мембран P Cl мала по сравнению с Р K , однако для других (например, в скелетных мышцах) P Cl , значительно выше, чем Р K .
Активный транспорт, натриевый насос . В предыдущем разделе описаны пассивная диффузия ионов и образующийся вследствие этого мембранный потенциал при заданных внутри– и внеклеточных концентрациях ионов. Однако в результате этого процесса концентрация ионов внутри клетки стабилизируется не автоматически, поскольку мембранный
потенциал несколько более электроотрицателен, чем E K , и намного–по сравнению с Е Na (около +60 мВ). Благодаря диффузии внутриклеточные концентрации ионов, по крайней мере калия и натрия, должны уравниваться с внеклеточными. Стабильность градиента ионов достигается посредством активного транспорта: мембранные белки переносят ионы через мембрану против электрического и (или) концентрационного градиентов, потребляя для этого метаболическую энергию. Наиболее важный процесс активного транспорта – это работа Na / K –насоса, существующего практически во всех клетках;
насос выкачивает ионы натрия из клетки, одновременно накачивая ионы калия внутрь клетки. Таким образом обеспечивается низкая внутриклеточная концентрация ионов натрия и высокая–калия (табл. 1.1). Градиент концентрации ионов натрия на мембране имеет специфические функции, связанные с передачей информации в виде электрических импульсов (см. разд. 2.2), а также с поддержанием других активных транспортных механизмов и регулирования объема клетки (см. ниже). Поэтому неудивительно, что более 1/3 энергии, потребляемой клеткой, расходуется на Na/K–насос, а в некоторых наиболее активных клетках на его работу расходуется до 70% энергии .
Na/K–транспортный белок представляет собой АТФазу. На внутренней поверхности мембраны она расщепляет АТФ на АДФ и фосфат (рис. 1.6). На транспортировку трех ионов натрия из клетки и одновременно двух ионов калия в клетку используется энергия одной молекулы АТФ, т. е. суммарно за один цикл из клетки удаляется один положительный заряд. Таким образом, Na/K–насос является электрогенным (создает электрический ток через мембрану), что приводит к увеличению электроотрицательности мембранного потенциала приблизительно на 10 мВ. Транспортный белок выполняет эту операцию с высокой скоростью: от 150 до 600 ионов натрия в секунду. Аминокислотная последовательность транспортного белка известна, однако еще не ясен механизм этого сложного обменного транспорта. Данный процесс описывают с использованием энергетических профилей переноса белками ионов натрия или калия (рис. 1.5,5). По характеру изменения этих профилей, связанных с постоянными изменениями конформации транспортного белка (процесс, требующий затраты энергии), можно судить о стехиометрии обмена: два иона калия обмениваются на три иона натрия.
Na/K–насос, как и изолированная Na + /К + –зависимая мембранная АТФаза, специфически ингибируется сердечным гликозидом уабаином (строфантином). Поскольку работа Na/K–насоса представляет собой многоступенчатую химическую реакцию, она, подобно всем химическим реакциям, в значительной степени зависит от температуры, что

Рис. 1.6. Схема Na/K–насоса–АТФазы (погруженной в липидный бислой плазматической мембраны), которая за один цикл выносит из клетки три иона Na + против градиентов потенциала и концентрации и приносит в клетку два иона К + . В ходе этого процесса одна молекула АТФ расщепляется на АДФ и фосфат. На схеме АТФаза показана как димер, состоящий из большой (функциональной) и малой субъединиц; в мембране она существует как тетрамер, образованный двумя большими и двумя малыми субъединицами
продемонстрировано на рис. 1.7. Здесь поток ионов натрия из мышечных клеток показан относительно времени; практически это эквивалентно потоку ионов натрия, опосредованному работой Na/K–насоса, потому что пассивный поток ионов натрия против градиентов концентрации и потенциала крайне мал. Если препарат охладить примерно на 18°С, то поток ионов натрия из клетки быстро уменьшится в 15 раз, а сразу после нагревания восстановится до исходного уровня. Такое уменьшение потока ионов натрия из клетки в несколько раз больше, чем то, которое бы соответствовало температурной зависимости процесса диффузии или простой химической реакции. Сходный эффект наблюдается, когда запас метаболической энергии истощается в результате отравления динитрофенолом (ДНФ) (рис. 1.7,5). Следовательно, поток ионов натрия из клетки обеспечивается энергозависимой реакцией – активным насосом. Другой характеристикой насоса наряду со значительной температурной и энергетической зависимостью является наличие уровня насыщения (как и у всех других химических реакций); это означает, что скорость работы насоса не может возрастать бесконечно при повышении концентрации транспортируемых ионов (рис. 1.8). В отличие от этого поток пассивно диффундирующего вещества растет пропорционально разности концентраций в соответствии с законом диффузии (уравнения 1 и 2).

Рис. 1.7. А, Б. Активный транспорт Na + . Ось ординат: поток радиоактивного 24 Na + из клетки (имп./мин). Ось абсцисс: время с начала эксперимента. А. Клетка охлаждена с 18,3 °С до 0,5 °С; поток Na + из клетки в этот период заторможен. Б. Подавление потока Na + из клетки динитрофенолом (ДНФ) в концентрации 0,2 ммоль/л (по с изменениями)
Помимо Na/K–насоса плазматическая мембрана содержит по крайней мере еще один насос – кальциевый; это насос откачивает ионы кальция (Са 2+) из клетки и участвует в поддержании их внутриклеточной концентрации на крайне низком уровне (табл. 1.1). Кальциевый насос присутствует с очень высокой плотностью в саркоплазматическом ретикулуме мышечных клеток, которые накапливают ионы кальция в результате расщепления молекул АТФ (см. гл. 4).
Воздействие Na/K–насоса на мембранный потенциал и объем клетки . На рис. 1.9 показаны различные компоненты мембранного тока и приведены внутриклеточные концентрации ионов, которые

Рис. 1.8. Соотношение между скоростью транспорта молекул и их концентрацией (в месте входа в канал или в месте связывания насоса) при диффузии через канал или при насосном транспорте. Последний при высоких концентрациях насыщается (максимальная скорость, Vmax ) значение по оси абсцисс, соответствующее половине максимальной скорости насоса (Vmax /2), является равновесной концентрацией К m

Рис. 1.9. Схема, показывающая концентрации Na + , K+ и Cl – внутри и вне клетки и пути проникновения этих ионов через клеточную мембрану (через специфические ионные каналы или с помощью Na/K–насоса. При данных концентрационных градиентах равновесные потенциалы E Na , E K и E С l – равны указанным, мембранный потенциал Em = – 90 мВ
обеспечивают их существование. Через калиевые каналы наблюдается выходящий ток ионов калия, так как мембранный потенциал несколько более электроположителен, чем равновесный потенциал для ионов калия. Общая проводимость натриевых каналов намного ниже, чем калиевых, т.е. натриевые каналы открыты намного реже, чем калиевые при потенциале покоя; однако в клетку входит примерно столько же ионов натрия, сколько выходит из нее ионов калия, потому что для диффузии ионов натрия в клетку необходимы большие градиенты концентрации и потенциала. Na/K–насос обеспечивает идеальную компенсацию пассивных диффузионных токов, так как переносит ионы натрия из клетки а ионы калия–в нее. Таким образом, насос является электрогенным за счет разницы в числе перенесенных в клетку и из клетки зарядов, что при нормальной скорости его работы создает мембранный потенциал, примерно на 10мВ более электроотрицательный, чем если бы он образовывался только за счет пассивных потоков ионов (см. уравнение 7). В результате мембранный потенциал приближается к калиевому равновесному потенциалу, что уменьшает утечку ионов калия. Активность Na /K–насоса регулируется внутриклеточной концентрацией ионов натрия. Скорость работы насоса замедляется при снижении концентрации ионов натрия, подлежащих выводу из клетки (рис. 1.8), так что работа насоса и поток ионов натрия внутрь клетки уравновешивают друг друга, поддерживая внутриклеточную концентрацию ионов натрия на уровне примерно 10 ммоль/л.
Чтобы поддерживать равновесие между насосным и пассивными мембранными токами, необходимо намного больше молекул Na/K–насоса, чем канальных белков для ионов калия и натрия. При открытом состоянии канала через него проходят десятки тысяч ионов за несколько миллисекунд (см. выше), а поскольку канал обычно открывается несколько раз в секунду, всего за это время через него проходит более 10 5 ионов. Одиночный насосный белок перемещает несколько сотен ионов натрия в секунду, следовательно, плазматическая мембрана должна содержать примерно в 1000 раз больше насосных молекул, чем канальных. Измерения канальных токов в покое показало наличие в среднем одного калиевого и одного натриевого открытого канала на 1 мкм 2 мембраны; из этого следует, что на том же пространстве должно присутствовать около 1000 молекул Na/K–насоса, т.е. расстояние между ними составляет в среднем 34 нм; диаметр насосного белка, как канального, составляет 8–10 нм. Таким образом, мембрана достаточно плотно насыщена насосными молекулами
Тот факт, что поток ионов натрия внутрь клетки, а ионов калия из клетки компенсируется работой насоса, имеется и другое следствие, заключающееся в сохранении стабильного осмотического давления и постоянного объема. Внутри клетки существует высокая концентрация крупных анионов, главным образом белков (А – в табл. 1.1), которые не способны проникать через мембрану (или проникают через нее очень медленно) и поэтому являются фиксированным компонентом внутри клетки. Чтобы уравновесить заряд этих анионов, необходимо равное количество катионов. Благодаря действию Na/K–насоса этими катионами в основном оказываются ионы калия. Существенное повышение внутриклеточной концентрации ионов могло бы происходить только при возрастании концентрации анионов вследствие потока С1 – по градиенту концентрации в клетку (табл. 1.1), но мембранный потенциал противодействует этому. Входящий ток Cl – наблюдается только до тех пор, пока не будет достигнут равновесный потенциал для ионов хлора; это наблюдается, когда градиент ионов хлора практически противоположен градиенту ионов калия, так как ионы хлора заряжены отрицательно (уравнение 4). Таким образом, устанавливается низкая внутриклеточная концентрация ионов хлора, соответствующая низкой внеклеточной концентрации ионов калия. Результатом является ограничение общего количества ионов в клетке. Если мембранный потенциал падает при блокаде Na/K–насоса, например при аноксии, то равновесный потенциал для ионов хлора снижается, а внутриклеточная концентрация ионов хлора соответственно повышается. Восстанавливая равновесие зарядов, ионы калия также входят в клетку; суммарная концентрация ионов в клетке возрастает, что повышает осмотическое давление; это заставляет воду поступать в клетку. Клетка набухает. Такое набухание наблюдается in vivo в условиях недостатка энергии.
Концентрационный градиент Na + как движущая сила мембранного транспорта . Значение Na/K–насоса для клетки не ограничивается стабилизацией нормальных градиентов К + и Na + на мембране. Энергия, запасенная в мембранном градиенте Na + , часто используется для обеспечения мембранного транспорта других веществ. Например, на рис. 1.10 показан «симпорт» Na + и молекулы сахара в клетку. Мембранный транспортный белок переносит молекулу сахара в клетку даже против градиента концентрации, в то же время Na + движется по градиенту концентрации и потенциала, обеспечивая энергию для транспорта сахаров. Такой транспорт сахаров полностью зависит от существования высокого градиента Na + ; если внутриклеточная концентрация Na + существенно возрастает, то транспорт сахаров прекращается. Для различных c ахаров существуют разные симпортные системы. Транспорт аминокислот в клетку сходен с транспортом c ахаров, показанным на рис. 1.10; он также обеспечивается градиентом Na + , существует по крайней мере пять различных систем симпорта, каждая из которых специализирована для какой–либо одной группы родственных аминокислот.
Помимо симпортных систем существуют также «антипортные». Одна из них, например, за один цикл переносит один ион кальция из клетки в обмен на три входящих иона натрия (рис. 1.10). Энергия для транспорта Са 2+ образуется за счет входа трех ионов натрия по градиенту концентрации и потенциала. Этой энергии достаточно (при потенциале покоя) для поддержания высокого градиента ионов кальция (от менее 10 –7 моль/л внутри клетки до приблизительно 2 ммоль/л вне клетки).
Эндо– и экзоцитоз . Для некоторых веществ, которые поступают в клетку или должны быть выведены

Рис. 1.10. Белки, погруженные в липидный бислой мембраны, опосредуют симпорт глюкозы и Na + в клетку, а также Ca 2+ / Na + –антипорт, в котором движущей силой является градиент Na + на клеточной мембране
из нее, транспортные каналы отсутствуют; к таким веществам относятся, например, белки и холестерол. Они могут проходить через плазматическую мембрану в везикулах, или пузырьках, с помощью эндо– и экзоцитоза. На рис. 1.11 показаны основные механизмы этих процессов. При экзоцитозе определенные органеллы (см. ниже) формируют везикулы, заполненные веществом, которое необходимо вывести из клетки, например гормонами или ферментами внеклеточного действия. Когда такие везикулы достигают плазматической мембраны, их липидная мембрана сливается с ней, давая таким образом возможность содержимому выйти во внешнюю среду. При противоположном процессе–эндоцитозе– плазматическая мембрана инвагинирует, образуя ямку, которая затем углубляется и замыкается, формируя внутриклеточную везикулу, заполненную внеклеточной жидкостью и некоторыми макромолекулами. Чтобы обеспечить это слияние мембран и замыкание везикулы, сократительные элементы цитоскелета действуют совместно с самими мембранами (см. ниже). При эндоцитозе не всегда происходит просто захват внеклеточной среды в клетку. В клеточной мембране содержатся часто организованные в специализированные группы специфические рецепторы к макромолекулам, таким, как инсулин или антигены. После того как эти макромолекулы свяжутся со своими рецепторами, в окружающем рецептор участке мембраны происходит эндоцитоз, и макромолекула избирательно транспортируется в клетку (рис. 1.12,Б).
Эндо– и экзоцитоз происходят в клетках непрерывно. Количество мембранного материала, совершающего оборот, значительно; в течение 1 ч макрофаг поглощает в виде везикул двойную площадь поверхности своей цитоплазматической мембраны. В большинстве клеток оборот мембранного материала происходит не столь интенсивно, но все же должен быть значительным.

Рис. 1.11. Экзоцитоз и эндоцитоз. Вверху: внутриклеточная везикула сливается с липидным бислоем плазматической мембраны и открывается во внеклеточное пространство. Этот процесс называется экзоцитозом. Внизу: плазматическая мембрана инвагинирует на небольшом участке и отшнуровывает везикулу, заполненную внеклеточным материалом. Этот процесс называют эндоцитозом
1.3. Перенос веществ внутри клетки
Эндо– и экзоцитоз–это не только процессы транспорта веществ через клеточную мембрану, но и процессы обмена мембран – структурных компонентов самой клетки. Предметом рассмотрения в данном разделе и являются другие сходные транспортные процессы в клетке и ее органеллах.
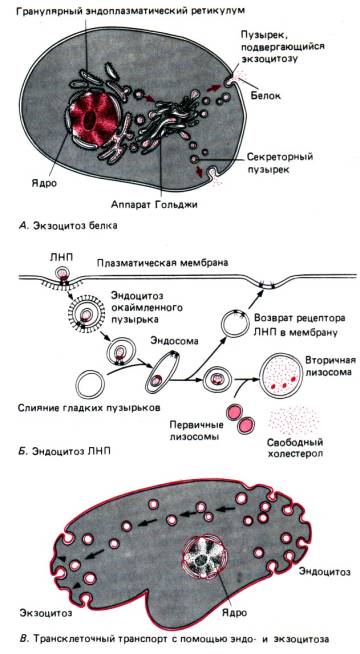
Рис. 1.12. А–В. Схема процессов, включающих экзо–и эндоцитоз. А. Белок, синтезированный в гранулярном эндоплазматическом ретикулуме, транспортируется посредством аппарата Гольджи к плазматической мембране, где секретируется путем экзоцитоза. Б. Холестерол, связанный с частицами ЛНП (липопротеина низкой плотности), присоединяется к плазматической мембране, индуцирует образование эндоцитозного пузырька в этом участке мембраны и транспортируется к лизосомам, где высвобождается. В. Внеклеточный материал, захваченный в процессе эндоцитоза (на рисунке справа), транспортируется через клетку в везикулах, или пузырьках, и выделяется посредством экзоцитоза (на рисунке слева)
Диффузия . Естественно, что в цитозоле разность концентраций устраняется за счет диффузии; то же самое справедливо и для жидкостей, заключенных в органеллах. Из–за высокой концентрации растворенного белка диффузия здесь протекает намного медленнее, чем в воде. Липидные мембраны–вокруг клетки и в составе органелл–это двумерные жидкости, в которых происходит диффузия. Липиды в мембранном бислое диффундируют в пределах собственного слоя, редко переходя из одного в другой. Погруженные в них белки также достаточно подвижны; они вращаются вокруг оси, перпендикулярной к мембране, или диффундируют латерально с весьма различными константами диффузии, в 2–10000 раз медленнее, чем фосфолипиды. Так, если некоторые белки перемещаются в липидном слое свободно и с такой же скоростью, как сами липидные молекулы, то другие заякорены, т.е. довольно прочно связаны с цитоскелетом. Существуют «постоянные» агрегаты специфических белков в мембране, например пре– и постсинаптические структуры нервных клеток. Свободно перемещающиеся белки можно продемонстрировать путем связывания их с флуоресцентными красителями, свечение которых индуцируют, освещая кратковременными вспышками маленький участок мембраны. Такие эксперименты показывают, что менее чем за 1 мин связавшиеся с красителем белки равномерно распределяются по мембране на расстояния до 10 мкм.
Активный транспорт в мембранах органелл .
Процессы активного транспорта, которые играют жизненно важную роль в функционировании плазматической мембраны, протекают также и внутри клетки–в мембранах органелл. Специфическое содержимое различных органелл создается отчасти благодаря внутреннему синтезу, а отчасти–за счет активного транспорта из цитозоля. Одним из примеров последнего является упоминавшийся выше Са 2+ –насос в саркоплазматическом ретикулуме мышечных клеток. Особенно интересно, что в случае синтеза АТФ в митохондриях действует принцип, обратный тому, что имеет место в АТФазных насосах плазматической мембраны (рис. 1.6). При синтезе АТФ окислительный метаболизм приводит к образованию крутого градиента H + на внутренних мембранах. Этот градиент является движущей силой для процесса, обратного насосному циклу активного транспорта молекул: ионы Н + движутся через мембрану по градиенту, а высвобождающаяся в результате этого энергия обеспечивает синтез АТФ из АДФ и фосфата. Образовавшаяся АТФ в свою очередь обеспечивает энергией клетку, в том числе и для активного транспорта.
Транспорт в везикулах . В клетке имеется большое число органелл и связанных с ними везикул (рис. 1.1). Эти органеллы, а везикулы особенно, находятся в постоянном движении, транспортируя свое содержимое к другим органеллам или к плазматической мембране. Везикулы могут также мигрировать из клеточной мембраны к органеллам, как при эндоцитозе.
Процесс секреции белка представлен на рис. 1.12, А. Белок синтезируется вблизи ядра клетки на рибосомах, связанных с эндоплазматическим ретикулумом (так называемый гранулярный, или шероховатый, эндоплазматический ретикулум); попав в эндоплазматический ретикулум, белок упаковывается в транспортные везикулы, которые отделяются от органеллы и мигрируют к аппарату Гольджи. Здесь они сливаются с цистернами аппарата Гольджи, где белок модифицируется (т.е. превращается в гликопротеин). На концах цистерн везикулы снова отделяются. Несущие модифицированный белок секреторные везикулы движутся к плазматической мембране и выделяют содержимое путем экзоцитоза.
Другой пример транспортного пути в клетке показан на рис. 1.12, Б; это–поглощение холестерола клеткой. Транспортируемый в крови холестерол связан в основном с белками, например частицами «липопротеина низкой плотности» (ЛНП). Такие частицы присоединяются к специфическим, содержащим рецепторы к ЛНП участкам мембраны, где происходит эндоцитоз и ЛНП переносится внутрь клетки в «окаймленных» везикулах. Эти везикулы сливаются, образуя эндосомы и утрачивая в ходе этого процесса «окаймление». Эндосомы в свою очередь сливаются с первичными лизосомами, содержащими преимущественно гидролитические ферменты, и образуют вторичные, более крупные лизосомы. В них холестерол высвобождается из частиц ЛНП и диффундирует в цитозоль, где становится доступным, например, для синтеза липидных мембран. От эндосом также отделяются везикулы, не содержащие ЛНП, которые особым путем движутся к плазматической мембране и сливаются с ней, возвращая мембранный материал и, вероятно, рецепторы к ЛНП. С момента связывания частицы ЛНП с мембраной до высвобождения холестерола из вторичной лизосомы проходит 10-15 мин. Нарушения в связывании и поглощении ЛНП, т. е– в снабжении клетки холестеролом, играют решающую роль в развитии серьезного и широко распространенного заболевания–атеросклероза («отвердение» артерий).
Существует множество других транспортных путей, сходных с показанными на рис. 1.11 и 1.12,А, с помощью которых движутся в клетке специфические везикулы. Неизвестно, как именно они передвигаются, но в этот процесс, вероятно, вовлечены элементы цитоскелета. Везикулы могут скользить по микротрубочкам, в этом случае энергия для движения, по–видимому, обеспечивается связанным с везикулами белком – АТФазой (см. ниже). Остается совершенно непонятным, как множество различных везикул, двигаясь одна за другой во всех направлениях, попадают по назначению. Они, очевидно, должны быть «помечены» таким образом, чтобы это распознавалось транспортной системой и преобразовывалось в целенаправленное движение.
Транспорт путем образования и разрушения органелл . До сих пор мы рассматривали эндо– и экзоцитоз как процессы транспортировки содержимого везикул. Существует и другой аспект этих процессов, заключающийся в том, что направленное удаление плазматической мембраны на одном участке клеточной поверхности путем эндоцитоза и, напротив, добавление ее на другом путем экзоцитоза перемещает значительные участки мембраны (рис. 1.12.Д), давая клетке возможность, например, сформировать вырост или двигаться.
Сходные перестройки типичны также и для цитоскелета, особенно для микрофиламентов и микротрубочек (рис. 1.1). Микрофиламенты состоят в первую очередь из белка F–актина, который способен к сборке в волокнистые пучки в результате полимеризации мономера из цитозоля. Пучки поляризованы, т. е. они часто нарастают только с одного конца, аккумулируя новые молекулы актина, тогда как другой конец инертен или здесь происходит разборка. За счет такого поляризованного роста микрофиламенты эффективно перемещаются и может изменяться структура их сети. Переход актина из деполимеризованного состояния (золя) в организованное (гель) может происходить очень быстро под действием других белков или изменений концентрации ионов (см. ниже). Существуют также белки, которые вызывают разрушение актиновых филаментов с образованием коротких фрагментов. Тонкие выросты многих клеток – филоподии – содержат центральный пучок актина (рис. 1.1), и различные движения филоподии, вероятно, обусловлены переходами актина: полимеризация – деполимеризация.
Микротрубочки также часто претерпевают подобные перемещения. Механизм этих перемещений сходен – полимеризация тубулина из цитозоля таким образом, что один из концов микротрубочки растет, тогда как другой либо не изменяется, либо там происходит разборка. Так микротрубочка путем соответствующего добавления или устранения материала может перемещаться по цитозолю.
Активные движения цитоскелета . Изменения цитосклетных структур могут происходить в результате как активных движений, так и перестроек, описанных выше. Во многих случаях движение микротрубочек и актиновых филаментов обусловлено сократительными белками, которые связывают филаменты или трубочки и могут перемещать их относительно друг друга. Белки миозин и динеин присутствуют в цитозоле всех клеток в сравнительно высоких концентрациях; они являются теми элементами, которые преобразуют энергию в движение в специализированных клетках (мышечных) и органеллах (ресничках). В мышечных клетках миозин образует толстые филаменты, ориентированные параллельно актиновым филаментам. Молекула миозина своей «головкой» присоединяется к актиновому филаменту и, используя энергию АТФ, смещает миозин вдоль молекулы актина. Затем миозин отсоединяется от актина. Совокупность множества таких циклов соединения–разъединения приводит к макроскопическому сокращению мышечных волокон (гл. 4). Динеин играет аналогичную роль в перемещении микротрубочек при работе ресничек (рис. 1.1). В цитоплазме неспециализированных клеток миозин и динеин образуют не правильные волокна, а в большинстве случаев маленькие группы молекул. Даже в виде таких малых агрегатов они способны перемещать актиновые филаменты или микротрубочки. Рис. 1.13 иллюстрирует этот процесс, когда к двум актиновым филаментам, поляризованным в разных направлениях, присоединены также противоположно поляризованные молекулы миозина. Головные группы миозина изгибаются к хвосту молекулы, расходуя при этом АТФ, а два актиновых филамента смещаются в противоположном направлении, после чего миозин отсоединяется от них. Перемещения такого рода, в ходе которых энергия АТФ преобразуется в механическую работу, могут изменять форму цитоскелета и, следовательно, клетки, а также обеспечивать транспорт связанных с цитоскелетом органелл.
Процессы внутриклеточного транспорта наиболее ярко могут быть продемонстрированы на аксоне нервной клетки. Аксонный транспорт рассматривается здесь подробно, чтобы проиллюстрировать события, которые, вероятно, сходным образом происходят в большинстве клеток. Аксон, диаметр которого составляет всего несколько микронов, может достигать длины одного метра и более, и движение белков путем диффузии от ядра к дистальному концу аксона заняло бы годы. Давно известно, что когда какой–либо из участков аксона подвергается констрикции, часть аксона, расположенная проксимальнее, расширяется. Это выглядит так, как будто в аксоне блокирован центробежный поток. Такой поток–быстрый аксонный транспорт может быть продемонстрирован движением радиоактивных маркеров, как в эксперименте, показанном на рис. 1.14. Лейцин, меченный радиоактивной меткой, инъецировали в ганглий дорсального корешка, и затем со 2–го по 10–й час измеряли радиоактивность в седалищном нерве на расстоянии 166 мм от тел нейронов. За 10 часов пик радиоактивности в месте инъекции менялся незначительно. Но волна радиоактивности распространялась по аксону с постоянной скоростью около 34 мм за 2 ч, или 410 мм/ сут. Показано, что во всех нейронах гомойотермных животных быстрый аксонный транспорт осуществляется с такой же скоростью, причем ощутимых различий между тонкими, безмиелиновыми волокнами и наиболее толстыми аксонами, а также между моторными и сенсорными волокнами не наблюдается. Тип радиоактивного маркера также не влияет на скорость быстрого аксонного транспорта; маркерами могут служить разнообразные радиоактивные

Рис. 1.13. Немышечный миозиновый комплекс при определенной ориентации может связываться с актиновыми филаментами различной полярности и, используя энергию АТФ, смещать их относительно друг друга
молекулы, такие, как различные аминокислоты, включающиеся в белки тела нейрона. Если проанализировать периферическую часть нерва, чтобы определить природу переносчиков транспортированной сюда радиоактивности, то такие переносчики обнаруживаются главным образом во фракции белков, но также в составе медиаторов и свободных аминокислот. Зная, что свойства этих веществ различны и особенно различны размеры их молекул, постоянную скорость транспорта мы можем объяснять только общим для всех них транспортным механизмом.
Описанный выше быстрый аксонный транспорт является антероградным, т. е. направленным от тела клетки. Показано, что некоторые вещества движутся от периферии к телу клетки с помощью ретроградного транспорта. Например, ацетилхолинэстераза транспортируется в этом направлении со скоростью в 2 раза меньшей, чем скорость быстрого аксонного транспорта. Маркер, часто используемый в нейроанатомии пероксидаза хрена–также перемещается ретроградным транспортом. Ретроградный транспорт, вероятно, играет важную роль в регуляции белкового синтеза в теле клетки. Через несколько дней после перерезки аксона в теле клетки наблюдается хроматолиз, что свидетельствует о нарушении белкового синтеза. Время, требующееся для хроматолиза, коррелирует с длительностью ретроградного транспорта от места перерезки аксона до тела клетки. Такой результат предполагает и объяснение этого нарушения – нарушается передача с периферии «сигнального вещества», регулирующего белковый синтез. Очевидно, что основными «средствами передвижения», используемыми для быстрого аксонного
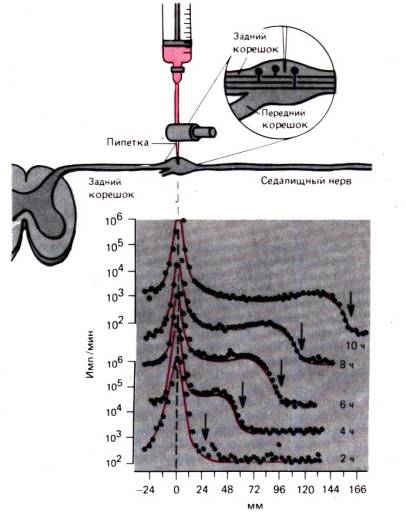
Рис. 1.14. Опыт, демонстрирующий быстрый аксонный транспорт в сенсорных волокнах седалищного нерва кошки. Меченный тритием лейцин вводят в ганглий дорсального корешка и измеряют радиоактивность в ганглии и сенсорных волокнах через 2, 4, 6, 8 и 10 ч после введения (нижняя часть рисунка). По оси абсцисс отложено расстояние от ганглия до участков седалищного нерва, где производят измерение. По оси ординат только для верхней и нижней кривой в логарифмическом масштабе отложена радиоактивность (имп./мин). «Волна» повышенной радиоактивности (стрелки) движется со скоростью 410мм/сут (по )
транспорта, являются везикулы (пузырьки) и органеллы, такие, как митохондрии, содержащие вещества, которые нужно транспортировать. Перемещение наиболее крупных везикул или митохондрии можно наблюдать с помощью микроскопа in vivo . Такие частицы совершают короткие быстрые движения в одном из направлений, останавливаются, часто двигаются немного назад или в сторону, снова останавливаются, а затем совершают рывок в основном направлении. 410 мм/сут соответствуют средней скорости антероградного движения приблизительно 5 мкм/с; скорость же каждого отдельного движения должна быть, следовательно, значительно выше, а если учесть размеры органелл, филаментов и микротрубочек, то эти движения действительно очень быстры. Быстрый аксонный транспорт требует значительной концентрации АТФ. Такие яды, как колхицин, разрушающий микротрубочки, также блокируют быстрый аксонный транспорт. Из этого следует, что в рассматриваемом нами транспортном процессе везикулы и органеллы движутся вдоль микротрубочек и актиновых филаментов; это движение обеспечивается малыми агрегатами молекул динеина и миозина, действующих, как показано на рис. 1.13, с использованием энергии АТФ.
Быстрый аксонный транспорт может участвовать и в патологических процессах. Некоторые нейротропные вирусы (например, вирусы герпеса или полиомиелита) проникают в аксон на периферии и движутся с помощью ретроградного транспорта к телу нейрона, где размножаются и оказывают свое токсическое действие. Токсин столбняка–белок, который продуцируется бактериями, попадающими в организм при повреждениях кожи, захватывается нервными окончаниями и транспортируется к телу нейрона, где он вызывает характерные мышечные спазмы. Известны случаи токсического воздействия на сам аксонный транспорт, например воздействие промышленным растворителем акриламидом. Кроме того, полагают, что патогенез авитаминоза «бери–бери» и алкогольной полинейропатии включает нарушение быстрого аксонного транспорта.
Помимо быстрого аксонного транспорта в клетке существует и довольно интенсивный медленный аксонный транспорт. Тубулин движется по аксону со скоростью около 1 мм/сут, а актин быстрее–до 5мм/сут. С этими компонентами цитоскелета мигрируют и другие белки; например, ферменты, по–видимому, связаны с актином или тубулином. Скорости перемещения тубулина и актина примерно согласуются со скоростью роста, обнаруженной для механизма, описанного ранее, когда молекулы включаются в активный конец микротрубочки или микрофиламента. Следовательно, этот механизм может лежать в основе медленного аксонного транспорта. Скорость медленного аксонного транспорта примерно соответствует также скорости роста аксона, что, по-видимому, указывает на ограничения, накладываемые структурой цитоскелета на второй процесс.
Завершая данный раздел, следует подчеркнуть, что клетки ни в коем случае не являются статичными структурами, каковыми они кажутся, например, на электронно–микроскопических фотографиях. Плазматическая мембрана и особенно органеллы находятся в постоянном быстром движении и постоянной перестройке; только поэтому они способны функционировать. Далее, это не простые камеры, в которых протекают химические реакции, а высокоорганизованные конгломераты мембран и волокон, в которых реакции протекают в оптимально организованной последовательности.
1.4. Регуляция клеточных функций
Поддержание индивидуальной клетки как функциональной единицы в основном регулируется ядром; изучение таких регуляторных механизмов –предмет клеточной биологии и биохимии. Вместе с тем клетки должны модифицировать свои функции в соответствии с условиями окружающей среды и нуждами других клеток организма, т. е. они служат объектами функциональной регуляции. Ниже мы кратко рассмотрим, каким образом эти регуляторные влияния действуют на плазматическую мембрану и как они доходят до внутриклеточных органелл.
Регуляторные воздействия на клеточную мембрану
Мембранный потенциал . Во многих случаях регуляция клеточных функций осуществляется путем изменения мембранного потенциала. Возможны локальные изменения потенциала, когда: 1) ток из соседнего участка клетки или генерированный другой клеткой протекает через мембрану; 2) изменяется внеклеточная концентрация ионов (часто [К + ] out ); 3) открываются мембранные ионные каналы. Изменения мембранного потенциала могут влиять на конформацию мембранных белков, заставляя, в частности, открываться или закрываться каналы. Как было описано выше, функционирование некоторых мембранных насосов зависит от мембранного потенциала. Нервные клетки специализированы воспринимать изменения мембранного потенциала как информацию, которая должна перерабатываться и передаваться (см. гл. 2).
Внеклеточные регуляторные вещества . Наиболее важным регуляторным механизмом с участием внеклеточных веществ является их взаимодействие со специфическими рецепторами на плазматической мембране или внутри клетки. К таким веществам относятся синаптические медиаторы, которые передают информацию между нервными клетками, локальные агенты и вещества, циркулирующие в крови и достигающие всех клеток организма, например гормоны и антигены. Синаптические медиаторы представляют собой небольшие молекулы, выделяющиеся из нервных окончаний в области синапса;
когда они достигают плазматической мембраны соседней, постсинаптической клетки, они запускают электрические сигналы или другие регуляторные механизмы. Этот вопрос подробно рассмотрен в гл. 3.
Локальные химические агенты часто выделяются специализированными клетками. Они свободно диффундируют во внеклеточном пространстве, однако их действие ограничено небольшой группой клеток вследствие быстрого разрушения этих веществ, либо спонтанного, либо под действием ферментов. Одним из примеров выделения таких агентов является высвобождение гистамина тучными клетками при повреждении или иммунной реакции. Гистамин вызывает расслабление гладкомышечных клеток сосудов, увеличивает проницаемость сосудистого эндотелия и стимулирует сенсорные нервные окончания, опосредующие ощущение зуда. Другие локальные химические агенты выделяются многими другими клетками. Типичными локальными агентами являются простагландины, составляющие группу приблизительно из 20 производных жирных кислот. Они выделяются непрерывно из широко распространенных клеток, но действуют только локально, так как быстро разрушаются фосфолипазами мембраны. Различные простагландины имеют широкий спектр действия: они могут запускать сокращение гладкомышечных клеток, вызывать агрегацию кровяных пластинок (тромбоцитов) или подавлять развитие желтого тела в яичниках.
Другие локальные агенты служат факторами роста. Наиболее известен фактор роста нервов (ФРН) для симпатических нейронов, который необходим для роста и выживания этих нейронов в ходе развития in vivo или в клеточной культуре. Очевидно, клетки–мишени для этого класса нейронов выделяют ФРН и тем самым обеспечивают правильную иннервацию. При формировании органов клеткам часто необходимо «находить путь» к клеткам–мишеням, которые могут располагаться на значительных расстояниях. Соответственно, должно существовать множество специализированных факторов роста, подобных ФРН.
Гормоны и антигены переносятся кровью ко всем клеткам. Антигены вызывают иммунный ответ клеток, несущих специфические антитела. Однако антигены, как правило, являются чужеродными веществами, не образуемыми в реагирующем организме (более подробно см. гл. 18). Некоторые гормоны, такие, как инсулин или тироксин, влияют на клетки, относящиеся к самым разнообразным типам, тогда как другие, например половые гормоны, – только на клетки определенного типа. Гормоны это либо пептиды, действие которых запускается связыванием их с рецептором на клеточной мембране, либо стероиды и тироксин, которые диффундируют через липидную мембрану и связываются с внутриклеточными рецепторами. Стероидные гормоны связываются с хроматином ядер, в результате чего запускается транскрипция определенных генов. Продуцирующиеся в результате белки вызывают изменение клеточных функций, в чем и заключается специфическое действие гормонов. Вопросы, связанные с выделением и действием гормонов, подробно рассмотрены в гл. 17.
Внутриклеточная коммуникация с участием вторых посредников
Регуляторные функции, описанные выше, включают воздействия на клеточную мембрану. Информация, полученная мембраной клетки, часто должна вызывать реакцию органелл и переносится к ним различными веществами, известными как вторые посредники (в отличие от первых, поступающих к клетке от внешних источников). Изучение вторых посредников развивается быстро, и нет гарантии, что нынешний уровень понимания проблемы окажется достаточно полным. Здесь мы коснемся трех хорошо изученных посредников: Са 2+ , цАМФ и инозитолтрифосфата.
Кальций. Простейший внутриклеточный посредник–это ион Са 2+ . Его свободная концентрация в покоящейся клетке очень низка и составляет 10 _ –8 –10 –7 моль/л. Он может проникать в клетку через специфические мембранные каналы, когда они находятся в открытом состоянии, например при изменениях мембранного потенциала (см. гл. 2). Возникающее в результате повышение концентрации Са 2+ запускает важные реакции в клетке, такие, как сокращение миофибрилл, которое является основой мышечного сокращения (см. гл. 4), или выделение везикул, содержащих медиаторы, из нервных окончаний (см. гл. 3). Обе реакции требуют концентрации Са 2+ , равной приблизительно 10 –5 моль/л. Са 2+ , оказывающий регуляторное действие, может высвобождаться также и из внутриклеточных депо, таких, как эндоплазматический ретикулум. Высвобождение Са 2+ из депо требует участия других посредников (см., например, рис. 1.16).
Циклический аденозинмонофосфат, цАМФ. В последнее время доказано, что циклический аденозинмонофосфат (цАМФ), производное основного источника энергии в организме–АТФ, является важным вторым посредником. Сложная цепь реакций, показанная на рис. 1.15, начинается с рецептора R s на наружной поверхности плазматической мембраны, который может служить местом специфического связывания для различных медиаторов и гормонов. После связывания со специфической «стимулирующей» молекулой R s изменяет свою конформацию; эти изменения влияют на белок G s на внутренней поверхности мембраны таким образом, что становится возможной активация последнего внутриклеточным гуанозинтрифосфатом (ГТФ). Активированный белок G s , в свою очередь, стимулирует фермент на внутренней поверхности мембраны–аденилатциклазу (АЦ), которая катализирует образование цАМФ из АТФ. Водорастворимый цАМФ и является посредником, передающим эффект

Рис. 1.15. Цепь реакций с участием внутриклеточного посредника цАМФ (циклического аденозинмонофосфата). Возбуждающий или тормозный внешние сигналы активируют мембранные рецепторы R s или Ri . Эти рецепторы регулируют процесс связывания G –белков с внутриклеточным ГТФ (гуанозинтрифосфатом), стимулируя или ингибируя тем самым внутриклеточную аденилатциклазу (АЦ). Усиливающий фермент АЦ превращает аденозинтрифосфат (АТФ) в цАМФ, который затем при участии фосфодиэстераэы расщепляется до АМФ. Свободный цАМФ диффундирует в клетку и активирует аденилаткиназу (А–киназу), высвобождая ее каталитическую субъединицу С, которая катализирует фосфорилирование внутриклеточных белков, т.е. формирует конечный эффект внеклеточного стимула. На схеме показаны также фармакологические препараты и токсины, которые запускают (+) или ингибируют (-) некоторые реакции (по с изменениями)
стимуляции внеклеточного рецептора R s к внутренним структурам клетки.
Параллельно со стимуляторной цепью реакций с участием R s возможно связывание тормозных медиаторов и гормонов с соответствующим рецептором R i который опять–таки через ГТФ–активируемый белок G , ингибирует АЦ и, таким образом, продукцию цАМФ. Диффундируя в клетку, цАМФ реагирует с аденилаткиназой (А–киназа); при этом высвобождается субъединица С, которая катализирует фосфорилирование белка Р. Это фосфорилирование переводит белки в активную форму, и теперь они могут Проявить свое специфическое регулирующее действие (например, вызвать деградацию гликогена). Эта сложная регуляторная система чрезвычайно эффективна, так как конечным результатом является фосфорилирование множества белков, т. е. регуляторный сигнал проходит цепь с большим коэффициентом усиления. Наружные медиаторы, которые связываются с рецепторами R s и R i специфическими для каждого из них, чрезвычайно разнообразны. Адреналин, связываясь с R s или R i участвует в регуляции метаболизма липидов и гликогена, а также в усилении сокращения сердечной мышцы и в других реакциях (см. гл. 19). Тиреотропный гормон, активируя R s , стимулирует выделение щитовидной железой гормона тироксина, а простагландин I тормозит агрегацию кровяных пластинок. Ингибиторные эффекты, в том числе адреналина, опосредованные через R i выражаются в замедлении липолиза. Таким образом, система цАМФ–это многофункциональная внутриклеточная регуляторная система, которая может точно контролироваться внеклеточными стимуляторными и ингибиторными сигнальными веществами.
Инозитолфосфат «ИФ з ». Внутриклеточная система второго посредника – инозитолфосфата – была открыта лишь недавно (рис. 1.16). В данном случае ингибиторный путь отсутствует, однако имеется сходство с системой цАМФ, в которой эффект стимуляции рецептора R переносится на ГТФ–активируемый G–белок на внутренней поверхности мембраны. На следующем этапе обычный мембранный липид фосфатидилинозитол (ФИ), предварительно получив две дополнительные фосфатные группы, превращается в ФИ–дифосфат (ФИФ 2), который расщепляется активированной фосфодиэстеразой (ФДЭ) на инозитолтрифосфат (ИФз) и липид диацилглицерол (ДАГ). Инозитолтрифосфат–это водорастворимый второй посредник, диффундирующий в цитозоль. Он действует в первую очередь путем высвобождения Ca 2+ из эндоплазматического ретикулума. Ca 2+ в свою очередь действует в качестве посредника, как было описано выше; например, он активирует Ca 2+ –зависимую фосфокиназу, фосфорилирующую ферменты. Липидная субъединица ДАГ (рис. 1.16) также переносит сигнал, диффундируя в липидной фазе плазматической мембраны к расположенной на ее внутренней поверхности С–киназе, которая и активируется с участием фосфатидилсерина в качестве кофактора. Затем С–киназа запускает фосфорилирование белков, переводя их в активную форму.
Внутриклеточная система второго посредника ИФ з также может управляться множеством внешних медиаторов и гормонов, в том числе ацетилхолином, серотонином, вазопрессином и тиреотропным гормоном; как и система цАМФ, она характеризуется разнообразными внутриклеточными эффектами. Возможно, эта система активируется и светом в зрительном рецепторе глаза, и играет центральную роль в фототрансдукции (см. гл. 11). Впервые в индивидуальном развитии организма рецептор системы ИФз активируется спермием, вследствие чего ИФз принимает участие в регуляторных реакциях, сопровождающих оплодотворение яйцеклетки.
Системы цАМФ и ИФз–ДАГ являются высокоэффективными биологическими усилителями. Они

Рис. 1.16. Цепь реакций с участием внутриклеточного посредника ИФз (инозитолтрифосфата). Как и в системе цАМФ, внеклеточный сигнал опосредован через белок G , который в данном случае активирует фосфодиэстеразу (ФДЭ). Этот фермент расщепляет фосфатидилинозиндифосфат (ФИФ 2 ) в плазматической мембране до ИФ з и диацилглицерола (ДАГ); ИФ з диффундирует в цитоплазму. Здесь он вызывает выброс Са 2+ из эндоплазматического ретикулума; повышение концентрации Са 2+ в цитоплазме ([Са 2+ ] i ) активирует протеинкиназу, которая фосфорилирует и, следовательно, активирует ферменты. Другой продукт, ДАГ, остается в мембране и активирует протеинкиназу С (кофактор–фосфатидилсерин, ФС). Протеинкиназа С также фосфорилирует ферменты, опосредующие специфическое действие, связанное со стимуляцией внешнего рецептора R . Ветви цепи реакций с участием ИФ з и ДАГ могут быть активированы независимо иономицином и форболовым эфиром соответственно (по с изменениями)
преобразуют реакцию между медиатором и наружным мембранным рецептором в фосфорилирование множества внутриклеточных белков, которые затем могут влиять на различные функции клетки. Один из существенных аспектов проблемы состоит в том, что, насколько известно на сегодняшний день, существуют только эти две тесно связанные регуляторные системы такого типа, используемые многочисленными внешними посредниками для регуляции разнообразных внутриклеточных процессов. Вместе с тем, эти регуляторные системы, в том числе и Са 2+ , тесно взаимодействуют друг с другом, что позволяет им осуществлять тонкую регуляцию клеточных функций.
Учебники и руководства
1. Alberts В ., Bray D., Lewis J., Raff M., Roberts К ., Watson J.D. Molecular Biology of the Cell, New York and London, Garland Publishing Inc., 1983.
2. Czihak G., Longer H., Ziegler H. (eds.). Biologie. Berlin, Heidelberg, New York, Springer, 1983.
3. Hille В . Ionic channels of excitable membranes. Sunderland, Mass., Sinauer Assoc., 1984.
4. Hoppe W., Lohmann W.. Marki H., Ziegler H. (eds.). Biophysik. Berlin, Heidelberg, New York, Springer, 1984.
5. Jungermann К ., Mahler H. Biochemie. Berlin, Heidelberg, New York, Springer, 1980.
6. Kandel E. R., Schwartz– J. H., (eds.). Principles of neural science, New York, Amsterdam, Oxford, Elsevier, 1985.
7. Schiebler Т . H., Schmidt W. Anatomic des Menschen. Berlin, Heidelberg, New York, Tokyo, Springer, 1983.
Оригинальные статьи и обзоры
8. Berridge M. J. The molecular basis of communication within the cell, Sci. Amer, 253, 124 134 (1985).
9. Berridge M. J., Irvine R. F. Inositol triphosphate, a novel second messenger in cellular signal transduction. Nature, 312, 315 321 (1984).
10. Bretscher M.S. The molecules of the cell membrane, Sci. Amer., 253, 124–134 (1985).
11. Daut J. The living cell as an energy–transducing machine. A minimal model of myocardial metabolism, Biochem. et Biophys. Acta, 895, 41–62 (1987).
12. Hodgkin A.L., Katz В . The effect of sodium ions on the electrical activity of the giant axon of the squid. J. Physiol. (Lond.), 108, 37–77 (1949).
13. Hodgkin A.L., Keynes R.D. Active transport of cations in giant axons from Sepia and Loligo, J. Physiol. (Lond.), 128, 28–42 (1955).
14. Longer P. Ionic channels with conformational substates, Biophys. J., 47, 581–590 (1985).
15. Ochs S., Worth P.M. Exoplasmic transport in normal and pathologic systems. In: Physiology and Pathology of Axons, S.G. Waxmann, Ed. New York, Raven Press, 1978.
краткое содержание других презентаций«Методика обучения биологии» - Школьная зоология. Знакомство учащихся с применением научных зоологических данных. Нравственное воспитание. Дополнительное освящение курятника. Выбор методов. Процессы жизнедеятельности. Аквариумные рыбки. Питание. Экологическое воспитание. Материальность жизненных процессов. Отрицательные результаты. Внимание учащихся. Обязательная форма. Рассматривание мелких животных. Цели и задачи биологии. Рассказ.
«Проблемное обучение на уроках биологии» - Знания. Новые учебники. Путь к решению. Проблема. Семинары. Что такое задача. Альбрехт Дюрер. Проблемное обучение на уроках биологии. Нестандартные уроки. Что подразумевается под проблемным обучением. Качество жизни. Биология как учебный предмет. Вопрос. Урок решения задач. Снижение интереса к предмету. Проблемно- лабораторные занятия.
«Критическое мышление на уроках биологии» - Технология «критического мышления». Использование технологии «развития критического мышления». Таблица к уроку. Мотивация к учению. Экосистемы. Значение «развития критического мышления». Признаки технологии. Технология «РКМ». Структура урока. Основные направления. История технологии. Педагогические технологии. Правила технологии. Задания по биологии. Фотосинтез. Приемы, используемые на разных стадиях урока.
«Уроки биологии с интерактивной доской» - Электронные учебники. Преимущества для учащихся. Интерактивная доска помогает донести информацию до каждого ученика. Дидактические задачи. Решение биологических задач. Преимущества работы с интерактивными досками. Работа с презентациями. Работа на сравнение объектов. Перемещение объектов. Использование электронных таблиц. Использование интерактивной доски в процессе обучения школьников. Преимущества для преподавателей.
«Системно-деятельностный подход в биологии» - Вопросы семинара. Деятельностный метод. Дриопитеки. Внеземной путь происхождения человека. Лизосомы. Химическая организация. Голосеменные растения. Метаболизм. Анализаторы. Системно-деятельностный подход в обучении биологии. Хромосомы. Цитоплазма. Слепота. Длина ушей. Классификация человека. Скелет млекопитающего. Пути эволюции человека. Митоз. Поверхностный комплекс. Проблемный вопрос. Ядрышко. Ядерная оболочка.
«Компьютер на биологии» - Совместная деятельность учащихся. Семейства покрытосеменных растений. Интерактивное обучение. Модели обучения. Пример системы оценивания. Вопросы инструктивной карточки. Пример инструктивной карточки. Исследователи. Микрогруппы. Технологии интерактивного обучения. Карусель. Интерактивные технологии обучения. Интерактивнные подходы на уроках биологии. Групповая форма работы. Задания для групп «исследователей».
