ساختار اولیه - دنباله ای از اسیدهای آمینه در یک زنجیره پلی پپتیدی در یک مولکول پروتئین، وقتی بخشهای سفت و سخت (پیوند پپتیدی) و انعطافپذیر (اتم α-کربن) متناوب میشوند، آرایش فشردهای از زنجیره در فضا شکل میگیرد.
روش آکوبوریاستفاده است فنیل هیدرازین. فنیل هیدرازین پیوندهای پپتیدی پروتئین را می شکند و به تمام آمینو اسیدها به جز آمینو ترمینال می چسبد. تجزیه و تحلیل کروماتوگرافی بعدی تشخیص آمینو اسید C ترمینال در پروتئین را ممکن می سازد.
مطالعه ساختار اولیه مهم است اهمیت بیولوژیکی و پزشکی عمومی:
- ساختار اولیه ساختارهای پروتئینی بعدی را تعیین می کند.
- دانش ساختار اولیه پروتئین برای سنتز مصنوعی پروتئین ها ضروری است.
- ساختار اولیه ویژگی گونه را تعیین می کند، به عنوان مثال، در پروتئین انسولین، معمولا در وسط مولکول در گونه های مختلف حیوانی و انسان، به عنوان یک قاعده، جایگزینی 3 اسید آمینه معادل رخ می دهد.
- تغییرات در ساختار اولیه می تواند منجر به بسیاری از بیماری ها شود، به عنوان مثال، کم خونی داسی شکل، که در آن در هموگلوبین در زنجیره β در موقعیت 6، اسید گلوتامیک با والین جایگزین می شود. این جایگزینی با یک اسید آمینه نابرابر منجر به اختلال در عملکرد هموگلوبین و ظهور گلبول های قرمز داسی شکل می شود.
ساختار ثانویه -یک الگوی مرتب تکراری از آرایش زنجیره پلی پپتیدی در فضا. اغلب، 2 نوع ساختار ثانویه در پروتئین ها یافت می شود: α - مارپیچ و ساختار β.
α - مارپیچدر سال 1951 توسط L. Pauling با استفاده از روش پراش اشعه ایکس مورد مطالعه قرار گرفت. این یک ساختار مارپیچ راست دست است که در یک دور آن 3.6 اسید آمینه جای می گیرد. گام مارپیچ (فاصله بین پیچ های مجاور) 0.54 نانومتر است. مارپیچ α توسط پیوندهای هیدروژنی ثابت می شود که بین پیوندهای پپتیدی تشکیل شده توسط هر چهارمین اسید آمینه بسته می شود. ساختار ثانویه α - به طور خود به خود تا می شود و توسط ساختار اولیه پروتئین تعیین می شود. نسبت نواحی چیده شده در یک ساختار مارپیچ در پروتئین های مختلف متفاوت است. به عنوان مثال، در هموگلوبین و میوگلوبین، چین ساختاری α غالب است، که اندازه مولکول پروتئین را تا 4 برابر کاهش می دهد.
ساختار βشکل "آکاردئون" دارد و توسط پیوندهای هیدروژنی بین بخشهای دور از یک زنجیره پلی پپتیدی یا بین چندین مولکول پروتئین تثبیت می شود. ساختارهای β موازی که در آن انتهای N و C با یکدیگر مطابقت دارند و ساختارهای ضد موازی وجود دارد. نمونه ای از پروتئین هایی که عمدتاً حاوی ساختارهای β هستند، ایمونوگلوبولین ها هستند.
ساختار ثانویه با تجزیه و تحلیل پراش اشعه ایکس و با مطالعه جذب پرتوهای فرابنفش توسط پروتئین مورد مطالعه قرار می گیرد (هرچه نسبت ساختارهای α بیشتر باشد، جذب بیشتر است).
ساختار ثانویه در حین دناتوراسیون از بین می رود.
ساختار سوم - باشکلی از آرایش فضایی زنجیره پلی پپتیدی خاص برای هر پروتئین. این ساختار به صورت خود به خود شکل می گیرد و توسط ساختار اولیه تعیین می شود. ساختار سوم به طور قابل توجهی فشردگی پروتئین را ده ها افزایش می دهد. پیوندهای غیر کووالانسی (آبگریز، یونی) و پیوندهای کووالانسی (دی سولفیدی) در تشکیل ساختار سوم نقش دارند.
ساختار سوم فعالیت بیولوژیکی و خواص فیزیکوشیمیایی پروتئین ها را تعیین می کند. اگر ساختار سوم مختل شود، پروتئین فعالیت بیولوژیکی خود را از دست می دهد.
روشهای مطالعه ساختار سوم، آنالیز پراش اشعه ایکس و تعیین فعالیت شیمیایی رادیکالهای اسید آمینه منفرد در یک پروتئین است. ساختار سوم پروتئین میوگلوبین برای اولین بار توسط J. Kendrew (1957) مورد مطالعه قرار گرفت. M. Perutz (1959) ساختار هموگلوبین را مطالعه کرد.
ساختار سوم پروتئین ها شامل ساختارهای α - مارپیچ، β - تا شده، حلقه های β (که در آن زنجیره پلی پپتیدی 180 0 خم می شود) و به اصطلاح سیم پیچ بی نظم است. به عنوان مثال، پروتئین انسولین شامل 57٪ α - مناطق مارپیچ، 6٪ - ساختارهای چین خورده β، 10٪ از مولکول به شکل حلقه های β مرتب شده است و 27٪ از مولکول یک سیم پیچ بی نظم است.
مجموع اولیه، ثانویه، ثالثی است ساختار مولکول پروتئین ترکیب مادام العمر (بومی) به طور خود به خود تشکیل می شود و تشکیل آن را تاشو می گویند. ترکیب پروتئین ها بسیار ناپایدار است و با مشارکت پروتئین های خاص تشکیل می شود - همراهان(اصحاب). چپرون ها می توانند به پروتئین های نیمه دناتوره شده ای که در حالت ناپایدار هستند متصل شوند و ترکیب اصلی خود را بازیابی کنند. چاپرون ها بر اساس وزن مولکولی (60 تا 100 سی دی) طبقه بندی می شوند. بیشترین مطالعه شده عبارتند از Sh-60، Sh-70 و Sh-90. به عنوان مثال، Sh-70 با پروتئین های غنی از رادیکال های آبگریز تعامل می کند و آنها را از دناتوره شدن در دمای بالا محافظت می کند. به طور کلی، چاپرون ها از پروتئین های اصلی بدن محافظت می کنند، از دناتوره شدن جلوگیری می کنند و به شکل گیری ترکیب کمک می کنند، انتقال پروتئین های دناتوره شده به لیزوزوم ها را تسهیل می کنند و در فرآیند سنتز پروتئین شرکت می کنند.
بر اساس ترکیب، تمام پروتئین ها به سه گروه تقسیم می شوند:
- پروتئین های فیبریل: کلاژن، الاستین، فیبروئین.
- پروتئین های کروی: هموگلوبین، آلبومین، گلوبولین.
- پروتئین های مخلوط: میوزین.
ساختار سوم در تمام پروتئین ها ذاتی است. فقط پروتئین های الیگومری که حاوی چندین زیر واحد، پروتومرها هستند، ساختار چهارتایی دارند. یک پروتومر یک زنجیره پلی پپتیدی جداگانه در نظر گرفته می شود، یک زیرواحد جزء فعال عملکردی یک پروتئین الیگومری است. یک زیرواحد ممکن است شامل یک پروتومر یا چندین باشد.
ساختار کواترنری -تعداد و آرایش نسبی زیر واحدها در پروتئین های الیگومری فقط پروتئین های الیگومری که حاوی چندین زیر واحد، پروتومرها هستند، ساختار چهارتایی دارند. یک پروتومر یک زنجیره پلی پپتیدی جداگانه در نظر گرفته می شود، یک زیرواحد جزء فعال عملکردی یک پروتئین الیگومری است. یک زیر واحد ممکن است حاوی یک پروتومر یا چند پروتومر باشد.
تشکیل ساختار چهارتایی شامل پیوندهای غیر کووالانسی ضعیف (آبگریز، یونی، هیدروژن) است. ساختار چهارتایی پروتئین ها خود به خود شکل می گیرد و در صورت دناتوره شدن به راحتی شکسته می شود. زیر واحدهای منفرد در یک پروتئین الیگومری با یکدیگر تعامل دارند که منجر به تغییرات در ساختار سوم پروتومرهای منفرد می شود. این پدیده تغییرات مشارکتی در ساختار پروتومر نامیده می شود و معمولاً با افزایش فعالیت پروتئین همراه است.
پروتئین های الیگومر در مقایسه با پروتئین های مونومر دارای تعدادی ویژگی هستند.
- آنها بسته بندی بسیار فشرده و سطح رابط نسبتاً کوچکی دارند، بنابراین با قرار گرفتن درون سلولی، آب کمتری را به هم متصل می کنند.
- فعالیت آنها در بدن تنظیم می شود. پروتومرها معمولاً غیر فعال هستند، اما پروتئین های الیگومری بسیار فعال تر هستند.
- اگر همان نوع پروتومرها در سنتز یک پروتئین الیگومری نقش داشته باشند، باعث صرفه جویی در مواد ژنتیکی می شود (چندین پروتومر یکسان روی بخش کوتاهی از DNA "مهر" می شوند).
- آنها از نظر عملکردی بیشتر با شرایط بدن سازگار هستند.
عملکرد پروتئین های الیگومری با مقایسه پروتئین های هموگلوبین و میوگلوبین که در انتقال اکسیژن به بافت ها نقش دارند نشان داده شده است. هموگلوبین گلبول های قرمز یک پروتئین الیگومری است که شامل 4 زنجیره پلی پپتیدی است. میوگلوبین عضلانی یک پروتئین مونومر است که شامل 1 زنجیره پلی پپتیدی است. منحنی اشباع اکسیژن میوگلوبین نشان دهنده وابستگی مستقیم آن به غلظت اکسیژن است. برای هموگلوبین، منحنی اشباع اکسیژن S شکل است. این به دلیل تغییر تدریجی متوالی در ساختار (ترکیب) هر یک از 4 پروتومر در ترکیب هموگلوبین است که در نتیجه میل هموگلوبین برای اکسیژن به شدت افزایش می یابد. این ماهیت اشباع هموگلوبین با اکسیژن به شدت ظرفیت اکسیژن آن را در مقایسه با میوگلوبین افزایش می دهد.
جایگاه ویژه ای در بین پروتئین ها اشغال شده است پروتئین های دامنه .
دامنه ها از نظر ساختاری و عملکردی بخش های مجزا از یک زنجیره پلی پپتیدی هستند. دامنه ها می توانند مسئول برهمکنش یک پروتئین با مواد مختلف - لیگاندها (مواد کم مولکولی، DNA، RNA، پلی ساکاریدها، و غیره) باشند.
به دلیل گزینش پذیری بالای پروتئین ها، می توان آنها را به مجتمع هایی ترکیب کرد که اغلب آنها را مجتمع های چند آنزیمی می نامند - اینها انجمن های ساختاری چندین آنزیم هستند که مراحل جداگانه یک فرآیند شیمیایی پیچیده را کاتالیز می کنند. به عنوان مثال: کمپلکس پیروات دهیدروژناز (PDC)، مجموعه ای از سه نوع آنزیم که اکسیداسیون اسید پیروویک (PVA) را کاتالیز می کند.
می توان به طور خاص نه تنها پروتئین های فردی، بلکه پروتئین ها را با لیپیدها (چربی ها) در طول تشکیل غشای سلولی و پروتئین ها را با اسیدهای نوکلئیک در طول تشکیل کروماتین ترکیب کرد.
خواص فیزیکوشیمیایی پروتئین ها.
آنها تا حد زیادی توسط ترکیب مولکول پروتئین (ساختار اولیه - سوم پروتئین) تعیین می شوند. خواص فیزیکوشیمیایی پروتئین ها در محلول ها ظاهر می شود.
انحلال پذیریپروتئین ها از پروتئینی به پروتئین دیگر متفاوت است.
به طور کلی، حلالیت پروتئین ها زیاد است، اما در بین انواع مختلف پروتئین ها متفاوت است. تحت تأثیر عوامل زیر است:
- شکل مولکول پروتئین (پروتئین های کروی محلول تر از پروتئین های فیبریلار هستند)
- ماهیت رادیکال اسید آمینه پروتئین، نسبت رادیکال های قطبی غیر قطبی (هرچه رادیکال های آبدوست قطبی در پروتئین بیشتر باشد، حلالیت آن بهتر است)
- خواص حلال، وجود نمک. غلظت کم نمک ها (KCL، NaCl) گاهی حلالیت پروتئین ها را افزایش می دهد. به عنوان مثال، آلبومین ها در آب مقطر خالص بهتر محلول هستند، گلوبولین ها فقط در حضور 10٪ نمک (KCL، NaCl) حل می شوند. پروتئین های بافت همبند کلاژن و الاستین در آب یا محلول های نمکی نامحلول هستند.
جرم مولکولیپروتئین ها بسیار بزرگ هستند، از 6000 تا 1000000 متغیر است، برای مثال، وزن مولکولی هموگلوبین 68000، آلبومین 100000، ریبونوکلئاز حدود 14000، میوزین 500000 است.
روش های تعیین جرم مولی پروتئین هاباید ملایم باشد و مولکول های پروتئین را از بین نبرد. به عنوان مثال، روش بولیوسکوپی که بر اساس اندازه گیری نقطه جوش محلول ها است، برای پروتئین ها قابل اجرا نیست. دقیق ترین روش ها برای تعیین وزن مولکولی پروتئین ها روش اولتراسانتریفیوژ و روش پراش اشعه ایکس است.
روش اولتراسانتریفیوژ(رسوب) بر اساس تغییر در سرعت ته نشینی پروتئین هایی با وزن های مولکولی مختلف هنگام چرخش محلول های پروتئینی با سرعت بالا است. وزن مولکولی پروتئین های یافت شده توسط این روش توسط واحد Svedberg تعیین می شود (S = 10-13 c.)
روش پراش اشعه ایکسبه شما امکان می دهد وزن مولکولی را با تجزیه و تحلیل تصاویر اشعه ایکس متعدد از یک مولکول پروتئین محاسبه کنید.
روش الکتروفورتیکبر اساس وابستگی سرعت حرکت پروتئین ها در یک میدان الکتریکی ثابت به وزن مولکولی پروتئین است (تحرک الکتروفورتیک برای پروتئین هایی با وزن مولکولی پایین تر است)
روش کروماتوگرافیبر اساس سرعت های مختلف عبور پروتئین های مختلف از طریق "الک" ژل مولکولی است.
مولکولهای بزرگتر از اندازه منافذ ژل سریعتر از مولکولهای پروتئینی کوچکتر که در دانههای ژل باقی میمانند از ژل عبور میکنند.
روش میکروسکوپی الکترونیبا مقایسه اندازه یک مولکول پروتئین با نمونه های مرجع با جرم شناخته شده انجام شد.
روش های شیمیاییمرتبط با ویژگی های ترکیب شیمیایی پروتئین ها
شکل مولکول های پروتئینناهمسان. مولکول های پروتئین می توانند فیبریلار یا کروی شکل باشند. پروتئین های فیبریلار شکل مولکولی نخ مانندی دارند. آنها معمولاً در آب و محلول های نمکی رقیق نامحلول هستند. پروتئین های فیبریلار شامل پروتئین های ساختاری اصلی بافت همبند هستند: کلاژن، کراتین، الاستین. در پروتئینهای کروی، زنجیرههای پلی پپتیدی بهطور محکم در ساختارهای کروی فشرده پیچیده میشوند. بیشتر پروتئین های کروی در آب و محلول های نمکی ضعیف بسیار محلول هستند. پروتئین های کروی شامل آنزیم ها، آنتی بادی ها، آلبومین ها و هموگلوبین هستند. برخی از پروتئین ها دارای یک نوع مولکول میانی هستند که شامل هر دو ناحیه نخ مانند و کروی است. نمونه ای از این پروتئین ها پروتئین ماهیچه ای میوزین است که در محلول های نمکی محلول است.
اندازه مولکول های پروتئیندر محدوده 1 تا 100 نانومتر، نزدیک به اندازه ذرات کلوئیدی هستند. به همین دلیل، محلول های پروتئینی دارای خواص محلول های واقعی و محلول های کلوئیدی هستند.
بسیاری از خواص جنبشی مولکولی محلول های پروتئین مشابه هستند خواص محلول های کلوئیدی .
- سرعت آهسته انتشار پروتئین های لازم برای تبادل آنها.
- عدم توانایی پروتئین ها در عبور از غشاهای نیمه تراوا. در محفظه هایی با غلظت پروتئین بالا، فشار هیدرواستاتیک اضافی به دلیل حرکت یک طرفه مولکول های آب از طریق غشای نیمه تراوا به سمت غلظت بالای پروتئین ایجاد می شود. فشار اضافی ایجاد شده توسط پروتئین ها فشار انکوتیک نامیده می شود. این یک عامل مهم تعیین کننده حرکت آب بین بافت ها، خون و روده ها است.
- ویسکوزیته بالای پروتئین ها به دلیل برهمکنش های بین مولکولی مختلف مولکول های پروتئینی بزرگ است. افزایش ویسکوزیته خون، به ویژه، بار روی عضله قلب را افزایش می دهد.
- برخی از پروتئین ها قادر به تشکیل ژل هستند که باعث افزایش قدرت پروتئین ها می شود (به عنوان مثال کلاژن).
خواص نوری پروتئین ها با اندازه مولکول های پروتئین، ساختار رادیکال های اسید آمینه در پروتئین ها، وجود پیوندهای پپتیدی و مناطق آلفا-مارپیچ در پروتئین ها تعیین می شود.
- محلول های پروتئینی دارند اثر شکست نور (انکسار) و پراکندگی نور.این ویژگی ها به دلیل اندازه بزرگ مولکول های پروتئین، متناسب با طول موج قسمت مرئی طیف است. درجه شکست متناسب با غلظت محلول پروتئین است.
- محلول های پروتئینی جذب اشعه ماوراء بنفشدر محدوده 190-230 نانومتر به دلیل وجود پیوندهای پپتیدی و در محدوده 260-280 نانومتر به دلیل وجود اسیدهای آمینه حلقوی در پروتئین ها. میزان جذب UV متناسب با غلظت پروتئین در محلول است.
- محلول های پروتئینی می توانند صفحه نور قطبی شده را بچرخانید،که به دلیل فعالیت نوری اسیدهای آمینه موجود در پروتئین و وجود مقاطع آلفا-مارپیچ در آن است. بین قطبش نور و غلظت پروتئین ها در محلول رابطه مستقیم وجود دارد.
پروتئین ها به عنوان محلول های مولکولی دارای خواص راه حل های واقعی . محلول های پروتئینی به عنوان محلول های واقعی، بسیار پایدار هستند.
ماژول 1 ساختار، خواص و عملکرد پروتئین ها
ماژول 1 ساختار، خواص و عملکرد پروتئین ها
ساختار ماژول | تم ها |
واحد مدولار 1 | 1.1. سازماندهی ساختاری پروتئین ها مراحل تشکیل ترکیب پروتئین بومی 1.2. اصول عملکرد پروتئین داروها به عنوان لیگاندهای موثر بر عملکرد پروتئین 1.3. دناتوره شدن پروتئین ها و امکان احیای خود به خودی آنها |
واحد ماژولار 2 | 1.4. ویژگی های ساختار و عملکرد پروتئین های الیگومری با استفاده از مثال هموگلوبین 1.5. حفظ ساختار پروتئین بومی تحت شرایط سلولی 1.6. انواع پروتئین ها. خانواده های پروتئینی با استفاده از مثال ایمونوگلوبولین ها 1.7. خواص فیزیکوشیمیایی پروتئین ها و روش های جداسازی آنها |
واحد مدولار 1 سازمان ساختاری پروتئین های مونومر و مبانی عملکرد آنها
اهداف آموزشی قادر باشند:
1. استفاده از دانش در مورد ویژگی های ساختاری پروتئین ها و وابستگی عملکرد پروتئین به ساختار آنها برای درک مکانیسم های ایجاد پروتئینوپاتی های ارثی و اکتسابی.
2. مکانیسم های عمل درمانی برخی داروها را به عنوان لیگاندهایی که با پروتئین ها برهمکنش می کنند و فعالیت آنها را تغییر می دهند، توضیح دهید.
3. استفاده از دانش در مورد ساختار و ثبات ساختاری پروتئین ها برای درک ناپایداری ساختاری و عملکردی و تمایل به دناتوره شدن آنها در شرایط متغیر.
4. استفاده از عوامل دناتوره کننده به عنوان وسیله ای برای استریل کردن مواد و ابزار پزشکی و همچنین ضد عفونی کننده ها را توضیح دهید.
بدانید:
1. سطوح سازماندهی ساختاری پروتئین ها.
2. اهمیت ساختار اولیه پروتئین ها که تنوع ساختاری و عملکردی آنها را تعیین می کند.
3. مکانیسم تشکیل مرکز فعال در پروتئین ها و برهمکنش خاص آن با لیگاند که زمینه ساز عملکرد پروتئین ها است.
4. نمونه هایی از تأثیر لیگاندهای خارجی (داروها، سموم، سموم) بر روی ترکیب و فعالیت عملکردی پروتئین ها.
5. علل و پیامدهای دناتوره شدن پروتئین، عوامل ایجاد دناتوره.
6. نمونه هایی از استفاده از عوامل دناتوره کننده در پزشکی به عنوان ضد عفونی کننده و وسیله ای برای استریل کردن وسایل پزشکی.
موضوع 1.1. سازمان ساختاری پروتئین ها. مراحل تشکیل بومی
ترکیبات پروتئینی
پروتئینها مولکولهای پلیمری هستند که مونومرهای آنها فقط 20 اسیدآمینه است. مجموعه و ترتیب ترکیب اسیدهای آمینه در یک پروتئین توسط ساختار ژن ها در DNA افراد تعیین می شود. هر پروتئین، مطابق با ساختار خاص خود، عملکرد خاص خود را انجام می دهد. مجموعه پروتئین های یک ارگانیسم مشخص ویژگی های فنوتیپی آن و همچنین وجود بیماری های ارثی یا مستعد رشد آنها را تعیین می کند.
1. آمینو اسیدهایی که پروتئین ها را می سازند. پیوند پپتیدیپروتئین ها پلیمرهایی هستند که از مونومرها - 20 اسیدآمینه آلفا ساخته شده اند که فرمول کلی آنها عبارت است از
اسیدهای آمینه در ساختار، اندازه و خواص فیزیکوشیمیایی رادیکال های متصل به اتم آلفا کربن متفاوت است. گروه های عاملی اسیدهای آمینه ویژگی های خواص اسیدهای آمینه مختلف را تعیین می کنند. رادیکال های موجود در اسیدهای آمینه α را می توان به چند گروه تقسیم کرد:
پرولین،بر خلاف 19 مونومر پروتئین دیگر، این یک اسید آمینه نیست، بلکه یک رادیکال در پرولین با اتم α-کربن و گروه ایمنو مرتبط است
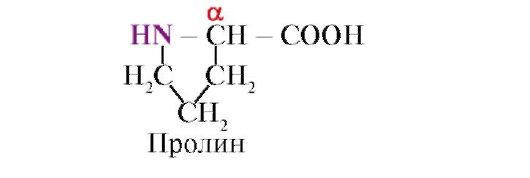 آمینو اسیدها از نظر حلالیت در آب متفاوت هستند.این به دلیل توانایی رادیکال ها در تعامل با آب (هیدرات) است.
آمینو اسیدها از نظر حلالیت در آب متفاوت هستند.این به دلیل توانایی رادیکال ها در تعامل با آب (هیدرات) است.
به آب دوستشامل رادیکال های حاوی گروه های عاملی آنیونی، کاتیونی و بدون بار قطبی است.
به آبگریزشامل رادیکال های حاوی گروه های متیل، زنجیره های آلیفاتیک یا حلقه ها می شود.
2. پیوندهای پپتیدی اسیدهای آمینه را به هم متصل می کنند تا پپتیدها را تشکیل دهند.در طول سنتز پپتید، گروه α-کربوکسیل یک اسید آمینه با گروه آلفا-آمینه یک اسید آمینه دیگر برهمکنش می کند و تشکیل می شود. پیوند پپتیدی:
پروتئین ها پلی پپتید هستند، به عنوان مثال. پلیمرهای خطی اسیدهای آمینه α که توسط یک پیوند پپتیدی متصل شده اند (شکل 1.1.)
 برنج. 1.1. اصطلاحات مورد استفاده برای توصیف ساختار پپتیدها
برنج. 1.1. اصطلاحات مورد استفاده برای توصیف ساختار پپتیدها
مونومرهای اسیدهای آمینه که پلی پپتیدها را می سازند نامیده می شوند باقی مانده های اسید آمینهزنجیره ای از گروه های تکراری - NH-CH-CO- تشکیل می دهد ستون فقرات پپتیدیبقایای اسید آمینه ای که دارای گروه آلفا-آمینه آزاد باشد N- ترمینال و آن که دارای گروه آلفا-کربوکسیل آزاد باشد C-terminal نامیده می شود. پپتیدها از انتهای N تا C پایانه نوشته و خوانده می شوند.
پیوند پپتیدی تشکیل شده توسط گروه ایمنو پرولین با سایر پیوندهای پپتیدی متفاوت است: اتم نیتروژن گروه پپتید فاقد هیدروژن است.
در عوض، یک پیوند با یک رادیکال وجود دارد که در نتیجه یک طرف حلقه در ستون فقرات پپتیدی قرار می گیرد:
 پپتیدها از نظر ترکیب اسید آمینه، تعداد اسیدهای آمینه و ترتیب اتصال اسیدهای آمینه متفاوت هستند، برای مثال Ser-Ala-Glu-Gis و His-Glu-Ala-Ser دو پپتید متفاوت هستند.
پپتیدها از نظر ترکیب اسید آمینه، تعداد اسیدهای آمینه و ترتیب اتصال اسیدهای آمینه متفاوت هستند، برای مثال Ser-Ala-Glu-Gis و His-Glu-Ala-Ser دو پپتید متفاوت هستند.
پیوندهای پپتیدی بسیار قوی هستند و هیدرولیز شیمیایی غیر آنزیمی آنها به شرایط سخت نیاز دارد: پروتئین مورد تجزیه و تحلیل در اسید هیدروکلریک غلیظ در دمای حدود 110 درجه به مدت 24 ساعت هیدرولیز می شود. در یک سلول زنده، پیوندهای پپتیدی را می توان شکست آنزیم های پروتئولیتیک،تماس گرفت پروتئازهایا هیدرولازهای پپتیدی
3. ساختار اولیه پروتئین ها.باقی مانده های اسید آمینه در زنجیره های پپتیدی پروتئین های مختلف به طور تصادفی متناوب نمی شوند، بلکه به ترتیب خاصی مرتب می شوند. توالی خطی یا ترتیب تناوب باقی مانده اسیدهای آمینه در یک زنجیره پلی پپتیدی نامیده می شود. ساختار اولیه پروتئین
ساختار اولیه هر پروتئین منفرد در مولکول DNA (در ناحیه ای به نام ژن) کدگذاری می شود و در طول رونویسی (کپی اطلاعات روی mRNA) و ترجمه (سنتز ساختار اولیه پروتئین) تحقق می یابد. در نتیجه، ساختار اولیه پروتئینهای یک فرد، اطلاعاتی است که به طور ارثی از والدین به فرزندان منتقل میشود، که ویژگیهای ساختاری پروتئینهای یک موجود زنده را تعیین میکند، که عملکرد پروتئینهای موجود به آن بستگی دارد (شکل 1.2).
 برنج. 1.2. رابطه بین ژنوتیپ و ترکیب پروتئین های سنتز شده در بدن فرد
برنج. 1.2. رابطه بین ژنوتیپ و ترکیب پروتئین های سنتز شده در بدن فرد
هر یک از حدود 100000 پروتئین فردی در بدن انسان وجود دارد منحصر بفردساختار اولیه مولکول های همان نوع پروتئین (به عنوان مثال، آلبومین) دارای تناوب یکسانی از باقی مانده اسیدهای آمینه هستند که آلبومین را از هر پروتئین فردی دیگر متمایز می کند.
توالی باقی مانده اسیدهای آمینه در یک زنجیره پپتیدی را می توان نوعی ثبت اطلاعات در نظر گرفت. این اطلاعات آرایش فضایی زنجیره پپتیدی خطی را به یک ساختار سه بعدی فشرده تر به نام تعیین می کند. ساختارسنجاب فرآیند تشکیل یک ترکیب پروتئین فعال عملکردی نامیده می شود تاشو
4. ترکیب پروتئین.چرخش آزاد در ستون فقرات پپتید بین اتم نیتروژن گروه پپتید و اتم آلفا کربن همسایه و همچنین بین اتم آلفا کربن و کربن گروه کربونیل امکان پذیر است. با توجه به برهمکنش گروه های عاملی بقایای اسید آمینه، ساختار اولیه پروتئین ها می تواند ساختارهای فضایی پیچیده تری به دست آورد. در پروتئین های کروی، دو سطح اصلی چین خوردگی ترکیب زنجیره های پپتیدی وجود دارد: ثانویو ساختار سوم
ساختار ثانویه پروتئین هایک ساختار فضایی است که در نتیجه تشکیل پیوندهای هیدروژنی بین گروه های عاملی -C=O و -NH- ستون فقرات پپتیدی ایجاد شده است. در این مورد، زنجیره پپتیدی می تواند ساختارهای منظمی از دو نوع به دست آورد: مارپیچ αو ساختارهای β
که در مارپیچ αپیوندهای هیدروژنی بین اتم اکسیژن گروه کربونیل و هیدروژن نیتروژن آمید اسید آمینه چهارم از آن تشکیل می شود. زنجیره های جانبی باقی مانده اسید آمینه
در امتداد حاشیه مارپیچ قرار دارند، بدون شرکت در تشکیل ساختار ثانویه (شکل 1.3.).
رادیکال های حجیم یا رادیکال های حامل بارهای مساوی از تشکیل مارپیچ α جلوگیری می کنند. باقیمانده پرولین، که دارای ساختار حلقه ای است، مارپیچ α را قطع می کند، زیرا به دلیل کمبود هیدروژن در اتم نیتروژن در زنجیره پپتیدی، تشکیل پیوند هیدروژنی غیرممکن است. پیوند بین نیتروژن و اتم آلفا کربن بخشی از حلقه پرولین است، بنابراین ستون فقرات پپتیدی در این نقطه خم می شود.
ب-ساختاربین نواحی خطی ستون فقرات پپتیدی یک زنجیره پلی پپتیدی تشکیل می شود و در نتیجه ساختارهای چین خورده تشکیل می شود. زنجیره های پلی پپتیدی یا قسمت هایی از آن می توانند تشکیل شوند موازییا ساختارهای β ضد موازیدر حالت اول، انتهای N و C زنجیرههای پپتیدی متقابل بر هم منطبق هستند و در حالت دوم جهت مخالف دارند (شکل 1.4).
برنج. 1.3. ساختار ثانویه پروتئین - α-مارپیچ
 برنج. 1.4. ساختارهای موازی و ضد موازی ورق β
برنج. 1.4. ساختارهای موازی و ضد موازی ورق β
ساختارهای β با فلش های پهن نشان داده می شوند: A - ساختار β ضد موازی. ب - سازه های موازی ورق بتا
در برخی پروتئین ها، ساختارهای β می توانند به دلیل تشکیل پیوندهای هیدروژنی بین اتم های ستون فقرات پپتیدی زنجیره های پلی پپتیدی مختلف تشکیل شوند.
همچنین در پروتئین ها یافت می شود مناطق با ثانویه نامنظمساختاری که شامل خم ها، حلقه ها و چرخش های ستون فقرات پلی پپتیدی است. آنها اغلب در مکان هایی قرار دارند که جهت زنجیره پپتیدی تغییر می کند، به عنوان مثال زمانی که یک ساختار ورقه β موازی تشکیل می شود.
بر اساس وجود مارپیچ های α و ساختارهای β، پروتئین های کروی را می توان به چهار دسته تقسیم کرد.
برنج. 1.5. ساختار ثانویه میوگلوبین (A) و زنجیره β هموگلوبین (B)، حاوی هشت مارپیچ α

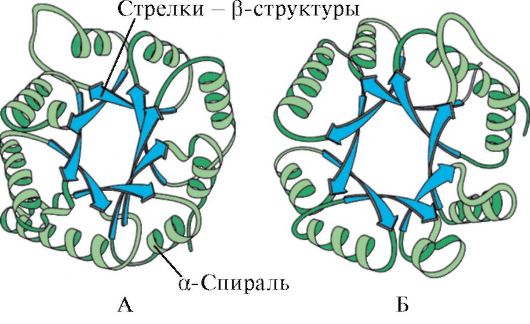 برنج. 1.6. ساختار ثانویه تریوسفسفات ایزومراز و دامنه پیروات کیناز
برنج. 1.6. ساختار ثانویه تریوسفسفات ایزومراز و دامنه پیروات کیناز
 برنج. 1.7. ساختار ثانویه دامنه ثابت ایمونوگلوبولین (A) و آنزیم سوپراکسید دیسموتاز (B)
برنج. 1.7. ساختار ثانویه دامنه ثابت ایمونوگلوبولین (A) و آنزیم سوپراکسید دیسموتاز (B)
که در دسته چهارمشامل پروتئین هایی است که حاوی مقدار کمی از ساختارهای ثانویه منظم است. این پروتئین ها شامل پروتئین های کوچک غنی از سیستئین یا متالوپروتئین ها هستند.
ساختار سوم پروتئین- نوعی ترکیب تشکیل شده به دلیل برهمکنش بین رادیکال های اسید آمینه، که می توانند در فاصله قابل توجهی از یکدیگر در زنجیره پپتیدی قرار گیرند. بیشتر پروتئین ها یک ساختار فضایی شبیه یک کروی (پروتئین های کروی) را تشکیل می دهند.
از آنجایی که رادیکال های اسید آمینه آبگریز تمایل به ترکیب از طریق به اصطلاح فعل و انفعالات آبگریزو نیروهای بین مولکولی واندروالس، یک هسته آبگریز متراکم در داخل گلبول پروتئین تشکیل می شود. رادیکال های یونیزه و غیریونیزه آبدوست عمدتاً روی سطح پروتئین قرار دارند و حلالیت آن را در آب تعیین می کنند.
 برنج. 1.8. انواع پیوندهایی که بین رادیکال های اسید آمینه در طی تشکیل ساختار سوم پروتئین ایجاد می شود.
برنج. 1.8. انواع پیوندهایی که بین رادیکال های اسید آمینه در طی تشکیل ساختار سوم پروتئین ایجاد می شود.
1 - پیوند یونی- بین گروه های عملکردی دارای بار مثبت و منفی رخ می دهد.
2 - پیوند هیدروژنی- بین یک گروه بدون بار آبدوست و هر گروه آبدوست دیگر رخ می دهد.
3 - فعل و انفعالات آبگریز- بین رادیکال های آبگریز ایجاد می شود.
4 - پیوند دی سولفیدی- به دلیل اکسیداسیون گروه های SH باقی مانده های سیستئین و تعامل آنها با یکدیگر ایجاد می شود.
بقایای اسید آمینه آبدوست واقع در داخل هسته آبگریز می توانند با استفاده از یکدیگر با یکدیگر تعامل کنند یونیو پیوند های هیدروژنی(شکل 1.8).
پیوندهای یونی و هیدروژنی و همچنین برهمکنش های آبگریز ضعیف هستند: انرژی آنها خیلی بیشتر از انرژی حرکت حرارتی مولکول ها در دمای اتاق نیست. ساختار پروتئین با تشکیل بسیاری از پیوندهای ضعیف حفظ می شود. از آنجایی که اتمهایی که یک پروتئین را میسازند در حرکت دائمی هستند، ممکن است برخی از پیوندهای ضعیف شکسته شوند و برخی دیگر تشکیل شوند که منجر به حرکتهای جزئی بخشهای جداگانه زنجیره پلی پپتیدی میشود. این خاصیت پروتئین ها برای تغییر ساختار در نتیجه شکستن برخی و تشکیل پیوندهای ضعیف دیگر نامیده می شود پایداری ساختاری
بدن انسان دارای سیستم هایی است که پشتیبانی می کنند هموستاز- پایداری محیط داخلی در حدود معین قابل قبول برای یک بدن سالم. در شرایط هموستاز، تغییرات کوچک در ساختار ساختار و عملکرد کلی پروتئین ها را مختل نمی کند. ترکیب فعال عملکردی یک پروتئین نامیده می شود ترکیب بومیتغییرات در محیط داخلی (به عنوان مثال، غلظت گلوکز، یون های کلسیم، پروتون ها و غیره) منجر به تغییر در ترکیب و اختلال در عملکرد پروتئین می شود.
ساختار سوم برخی از پروتئین ها تثبیت شده است پیوندهای دی سولفیدیبه دلیل تعامل گروه های -SH از دو باقی مانده تشکیل شده است
 برنج. 1.9. تشکیل پیوند دی سولفید در یک مولکول پروتئین
برنج. 1.9. تشکیل پیوند دی سولفید در یک مولکول پروتئین
سیستئین (شکل 1.9). اکثر پروتئین های درون سلولی در ساختار سوم خود پیوندهای دی سولفید کووالانسی ندارند. وجود آنها مشخصه پروتئین های ترشح شده توسط سلول است که پایداری بیشتر آنها را در شرایط خارج سلولی تضمین می کند. بنابراین، پیوندهای دی سولفید در مولکول های انسولین و ایمونوگلوبولین ها وجود دارد.
انسولین- یک هورمون پروتئینی که در سلولهای β پانکراس سنتز می شود و در پاسخ به افزایش غلظت گلوکز در خون به خون ترشح می شود. در ساختار انسولین، دو پیوند دی سولفیدی وجود دارد که زنجیره های پلی پپتیدی A و B را به هم متصل می کند و یک پیوند دی سولفیدی در زنجیره A وجود دارد (شکل 1.10).
 برنج. 1.10. پیوندهای دی سولفید در ساختار انسولین
برنج. 1.10. پیوندهای دی سولفید در ساختار انسولین
5. ساختار فوق ثانویه پروتئین ها.در پروتئین هایی با ساختار و عملکرد اولیه متفاوت، گاهی اوقات آنها را شناسایی می کنند ترکیبات مشابه و موقعیت های نسبی سازه های ثانویه،که ساختار فوق ثانویه نامیده می شوند. از آنجایی که ترکیب خاصی از عناصر ساختار ثانویه در تشکیل ساختار سوم پروتئین است، موقعیت میانی را بین ساختارهای ثانویه و سوم اشغال می کند. ساختارهای فوق ثانویه دارای اسامی خاص هستند، مانند "α-helix-turn-a-helix"، "زیپ لوسین"، "انگشتان روی"، و غیره. چنین ساختارهای فوق ثانویه مشخصه پروتئین های متصل به DNA هستند.
"زیپ لوسین."این نوع ساختار فوق ثانویه برای اتصال دو پروتئین به یکدیگر استفاده می شود. روی سطح پروتئینهای در حال تعامل، نواحی مارپیچ α حاوی حداقل چهار باقیمانده لوسین وجود دارد. بقایای لوسین در مارپیچ α با شش آمینو اسید فاصله دارند. از آنجایی که هر چرخش مارپیچ α حاوی 3.6 باقیمانده اسید آمینه است، رادیکالهای لوسین بر روی سطح هر چرخش دوم قرار دارند. باقیماندههای لوسین از مارپیچ α یک پروتئین میتوانند با باقیماندههای لوسین پروتئین دیگر (برهمکنشهای آبگریز) تعامل داشته باشند و آنها را به هم متصل کنند (شکل 1.11). بسیاری از پروتئینهای اتصالدهنده DNA در کمپلکسهای الیگومری عمل میکنند که در آن واحدهای فرعی توسط «زیپهای لوسینی» به یکدیگر مرتبط هستند.
 برنج. 1.11. "زیپ لوسین" بین مناطق α-مارپیچ دو پروتئین
برنج. 1.11. "زیپ لوسین" بین مناطق α-مارپیچ دو پروتئین
نمونه ای از این پروتئین ها هیستون ها هستند. هیستون ها- پروتئین های هسته ای، که حاوی تعداد زیادی اسید آمینه با بار مثبت - آرژنین و لیزین (تا 80٪). مولکولهای هیستون با استفاده از زیپهای لوسین، با وجود بار همنام قابل توجه این مولکولها، به کمپلکسهای الیگومری حاوی هشت مونومر ترکیب میشوند.
"انگشت روی"- گونه ای از ساختار فوق ثانویه، مشخصه پروتئین های متصل به DNA، به شکل یک قطعه دراز روی سطح پروتئین است و حاوی حدود 20 باقی مانده اسید آمینه است (شکل 1.12). شکل "انگشت کشیده" توسط یک اتم روی متصل به چهار رادیکال اسید آمینه - دو باقی مانده سیستئین و دو باقی مانده هیستیدین پشتیبانی می شود. در برخی موارد، به جای باقی مانده هیستیدین، باقی مانده سیستئین وجود دارد. دو باقیمانده سیستئین نزدیک به هم با یک توالی Cys متشکل از تقریباً 12 باقیمانده اسید آمینه از دو باقی مانده Gisili دیگر جدا می شوند. این ناحیه از پروتئین یک مارپیچ α را تشکیل می دهد که رادیکال های آن می توانند به طور خاص به مناطق تنظیم کننده شیار اصلی DNA متصل شوند. ویژگی اتصال فردی
 برنج. 1.12. ساختار اولیه ناحیه پروتئین های متصل به DNA که ساختار "انگشت روی" را تشکیل می دهند (حروف نشان دهنده آمینو اسیدهای تشکیل دهنده این ساختار هستند)
برنج. 1.12. ساختار اولیه ناحیه پروتئین های متصل به DNA که ساختار "انگشت روی" را تشکیل می دهند (حروف نشان دهنده آمینو اسیدهای تشکیل دهنده این ساختار هستند)
پروتئین تنظیم کننده اتصال به DNA به دنباله ای از باقی مانده های اسید آمینه واقع در ناحیه انگشت روی بستگی دارد. چنین ساختارهایی به ویژه حاوی گیرنده های هورمون های استروئیدی هستند که در تنظیم رونویسی (خواندن اطلاعات از DNA به RNA) دخیل هستند.
موضوع 1.2. مبانی عملکرد پروتئین. داروها به عنوان لیگاندهای مؤثر بر عملکرد پروتئین
1. مرکز فعال پروتئین و برهمکنش آن با لیگاند.در طول تشکیل ساختار سوم، ناحیه ای بر روی سطح یک پروتئین فعال از نظر عملکردی، معمولاً در یک فرورفتگی، تشکیل می شود که توسط رادیکال های اسید آمینه که در ساختار اولیه از یکدیگر دور هستند، تشکیل می شود. این ناحیه که ساختار منحصر به فردی برای یک پروتئین معین دارد و قادر است به طور خاص با یک مولکول خاص یا گروهی از مولکول های مشابه برهم کنش داشته باشد، محل اتصال پروتئین-لیگاند یا محل فعال نامیده می شود. لیگاندها مولکول هایی هستند که با پروتئین ها تعامل دارند.
ویژگی بالاتعامل پروتئین با لیگاند با مکمل بودن ساختار مرکز فعال با ساختار لیگاند تضمین می شود.
مکمل بودن- این مطابقت فضایی و شیمیایی سطوح در حال تعامل است. مرکز فعال نه تنها باید از نظر فضایی با لیگاند موجود در آن مطابقت داشته باشد، بلکه پیوندهایی (برهمکنش های یونی، هیدروژنی و آبگریز) نیز باید بین گروه های عاملی رادیکال های موجود در مرکز فعال و لیگاند که لیگاند را نگه می دارد تشکیل شود. در مرکز فعال (شکل 1.13).
 برنج. 1.13. برهمکنش مکمل پروتئین با لیگاند
برنج. 1.13. برهمکنش مکمل پروتئین با لیگاند
برخی لیگاندها وقتی به مرکز فعال پروتئین متصل می شوند، نقش کمکی در عملکرد پروتئین ها دارند. این لیگاندها کوفاکتور نامیده می شوند و پروتئین های حاوی بخش غیر پروتئینی نامیده می شوند پروتئین های پیچیده(بر خلاف پروتئین های ساده که فقط از قسمت پروتئینی تشکیل شده اند). قسمت غیر پروتئینی که محکم به پروتئین متصل است نامیده می شود گروه پروتزبه عنوان مثال، میوگلوبین، هموگلوبین و سیتوکروم ها حاوی یک گروه مصنوعی به نام هم هستند که حاوی یون آهن است و محکم به مرکز فعال متصل است. پروتئین های پیچیده حاوی هم، هموپروتئین نامیده می شوند.
وقتی لیگاندهای خاصی به پروتئین ها متصل می شوند، عملکرد این پروتئین ها آشکار می شود. بنابراین، آلبومین، مهمترین پروتئین در پلاسمای خون، عملکرد انتقال خود را با اتصال لیگاندهای آبگریز، مانند اسیدهای چرب، بیلی روبین، برخی داروها و غیره به مرکز فعال نشان می دهد (شکل 1.14).
لیگاندهایی که با ساختار سه بعدی زنجیره پپتیدی تعامل دارند نه تنها می توانند مولکول های آلی و معدنی کم مولکولی، بلکه درشت مولکول ها نیز باشند:
DNA (نمونه هایی با پروتئین های متصل شونده به DNA که در بالا مورد بحث قرار گرفت).
پلی ساکاریدها؛
 برنج. 1.14. رابطه بین ژنوتیپ و فنوتیپ
برنج. 1.14. رابطه بین ژنوتیپ و فنوتیپ
ساختار اولیه منحصر به فرد پروتئین های انسانی، رمزگذاری شده در مولکول DNA، در سلول ها به شکل یک ترکیب منحصر به فرد، ساختار مرکز فعال و عملکردهای پروتئین تحقق می یابد.
در این موارد، پروتئین ناحیه خاصی از لیگاند را که متناسب و مکمل محل اتصال است، تشخیص می دهد. بنابراین، در سطح سلولهای کبدی پروتئینهای گیرنده هورمون انسولین وجود دارد که ساختار پروتئینی نیز دارد. تعامل انسولین با گیرنده باعث تغییر در ساختار آن و فعال شدن سیستم های سیگنالینگ می شود که منجر به ذخیره مواد مغذی در سلول های کبدی بعد از غذا می شود.
بدین ترتیب، عملکرد پروتئین ها بر اساس تعامل خاص مرکز فعال پروتئین با لیگاند است.
2. ساختار دامنه و نقش آن در عملکرد پروتئین ها.زنجیره های پلی پپتیدی طولانی پروتئین های کروی اغلب به چندین ناحیه فشرده و نسبتا مستقل جمع می شوند. آنها ساختار سوم مستقلی دارند که یادآور پروتئین های کروی است و نامیده می شوند دامنه هابه لطف ساختار دامنه پروتئین ها، ساختار سوم آنها آسان تر است.
در پروتئین های دامنه، محل های اتصال لیگاند اغلب بین دامنه ها قرار دارند. بنابراین، تریپسین یک آنزیم پروتئولیتیک است که توسط بخش برونریز پانکراس تولید میشود و برای هضم پروتئینهای غذا ضروری است. این ساختار دو دامنه ای دارد و مرکز اتصال تریپسین با لیگاند آن - پروتئین غذا - در شیار بین دو دامنه قرار دارد. در مرکز فعال، شرایط لازم برای اتصال موثر یک محل خاص پروتئین غذایی و هیدرولیز پیوندهای پپتیدی آن ایجاد می شود.
هنگامی که مرکز فعال با لیگاند تعامل می کند، دامنه های مختلف در یک پروتئین می توانند نسبت به یکدیگر حرکت کنند (شکل 1.15).
هگزوکیناز- آنزیمی که فسفوریلاسیون گلوکز را با استفاده از ATP کاتالیز می کند. محل فعال آنزیم در شکاف بین دو حوزه قرار دارد. هنگامی که هگزوکیناز به گلوکز متصل می شود، حوزه های اطراف آن بسته می شود و بستر به دام می افتد، جایی که فسفوریلاسیون رخ می دهد (شکل 1.15 را ببینید).
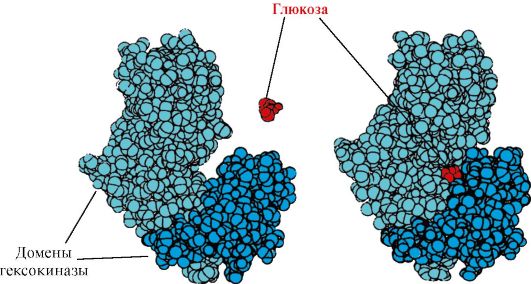 برنج. 1.15. اتصال دامنه های هگزوکیناز به گلوکز
برنج. 1.15. اتصال دامنه های هگزوکیناز به گلوکز
در برخی پروتئین ها، دامنه ها با اتصال به لیگاندهای مختلف، عملکردهای مستقلی را انجام می دهند. چنین پروتئین هایی چند منظوره نامیده می شوند.
3. داروها لیگاندهایی هستند که بر عملکرد پروتئین ها تأثیر می گذارند.برهمکنش پروتئین ها با لیگاندها خاص است. با این حال، به دلیل ناپایداری ساختاری پروتئین و مرکز فعال آن، می توان ماده دیگری را انتخاب کرد که بتواند با پروتئین موجود در مرکز فعال یا قسمت دیگری از مولکول نیز تعامل داشته باشد.
ماده ای مشابه ساختار لیگاند طبیعی نامیده می شود آنالوگ ساختاری لیگاندیا یک لیگاند غیر طبیعی. همچنین با پروتئین در محل فعال تعامل دارد. یک آنالوگ ساختاری لیگاند می تواند عملکرد پروتئین را افزایش دهد (آگونیست)،و آن را کاهش دهید (آنتاگونیست).لیگاند و آنالوگ های ساختاری آن برای اتصال به پروتئین در همان محل با یکدیگر رقابت می کنند. چنین موادی نامیده می شوند تعدیل کننده های رقابتی(تنظیم کننده) عملکرد پروتئین. بسیاری از داروها به عنوان مهارکننده های پروتئین عمل می کنند. برخی از آنها با اصلاح شیمیایی لیگاندهای طبیعی به دست می آیند. مهارکننده های عملکرد پروتئین می توانند داروها و سموم باشند.
آتروپین یک مهارکننده رقابتی گیرنده های M-کولینرژیک است.استیل کولین یک انتقال دهنده عصبی برای انتقال تکانه های عصبی از طریق سیناپس های کولینرژیک است. برای انجام تحریک، استیل کولین آزاد شده در شکاف سیناپسی باید با پروتئین گیرنده غشای پس سیناپسی تعامل داشته باشد. دو نوع پیدا شد گیرنده های کولینرژیک:
گیرنده Mعلاوه بر استیل کولین، به طور انتخابی با موسکارین (سم آگاریک مگس) تعامل دارد. M - گیرنده های کولینرژیک در عضلات صاف وجود دارند و هنگام تعامل با استیل کولین باعث انقباض آنها می شوند.
گیرنده Hبه طور خاص به نیکوتین متصل می شود. گیرنده های N-کولینرژیک در سیناپس های ماهیچه های اسکلتی مخطط یافت می شوند.
بازدارنده اختصاصی گیرنده های ام کولینرژیکآتروپین است. در گیاهان بلادونا و حنبان یافت می شود.
 آتروپین دارای گروههای عملکردی شبیه به استیل کولین و آرایش فضایی آنها است، بنابراین یک مهارکننده رقابتی گیرندههای M-کولینرژیک است. با توجه به اینکه اتصال استیل کولین به گیرنده های M کولینرژیک باعث انقباض عضلات صاف می شود، از آتروپین به عنوان دارویی برای رفع اسپاسم آنها استفاده می شود. (ضد اسپاسم).بنابراین، استفاده از آتروپین برای شل کردن عضلات چشم هنگام مشاهده فوندوس و همچنین برای تسکین اسپاسم در هنگام قولنج گوارشی شناخته شده است. گیرنده های M-کولینرژیک نیز در سیستم عصبی مرکزی (CNS) وجود دارند، بنابراین دوزهای زیاد آتروپین می تواند باعث واکنش نامطلوب از سیستم عصبی مرکزی شود: تحریک حرکتی و ذهنی، توهم، تشنج.
آتروپین دارای گروههای عملکردی شبیه به استیل کولین و آرایش فضایی آنها است، بنابراین یک مهارکننده رقابتی گیرندههای M-کولینرژیک است. با توجه به اینکه اتصال استیل کولین به گیرنده های M کولینرژیک باعث انقباض عضلات صاف می شود، از آتروپین به عنوان دارویی برای رفع اسپاسم آنها استفاده می شود. (ضد اسپاسم).بنابراین، استفاده از آتروپین برای شل کردن عضلات چشم هنگام مشاهده فوندوس و همچنین برای تسکین اسپاسم در هنگام قولنج گوارشی شناخته شده است. گیرنده های M-کولینرژیک نیز در سیستم عصبی مرکزی (CNS) وجود دارند، بنابراین دوزهای زیاد آتروپین می تواند باعث واکنش نامطلوب از سیستم عصبی مرکزی شود: تحریک حرکتی و ذهنی، توهم، تشنج.
دیتیلین آگونیست رقابتی گیرنده های کولینرژیک H است که عملکرد سیناپس های عصبی عضلانی را مهار می کند.
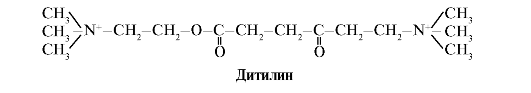 سیناپس های عصبی عضلانی ماهیچه های اسکلتی حاوی گیرنده های کولینرژیک H هستند. تعامل آنها با استیل کولین منجر به انقباض عضلانی می شود. در برخی از اعمال جراحی و همچنین در مطالعات آندوسکوپی از داروهایی استفاده می شود که باعث شل شدن عضلات اسکلتی می شود. (شل کننده های عضلانی).اینها شامل دیتیلین است که آنالوگ ساختاری استیل کولین است. به گیرنده های کولینرژیک H می چسبد، اما برخلاف استیل کولین، به آرامی توسط آنزیم استیل کولین استراز از بین می رود. در نتیجه باز شدن طولانی مدت کانال های یونی و دپلاریزاسیون مداوم غشاء، هدایت تکانه های عصبی مختل شده و شل شدن عضلات رخ می دهد. در ابتدا، این خواص در سم کورار کشف شد، به همین دلیل است که چنین داروهایی نامیده می شود کورار مانند
سیناپس های عصبی عضلانی ماهیچه های اسکلتی حاوی گیرنده های کولینرژیک H هستند. تعامل آنها با استیل کولین منجر به انقباض عضلانی می شود. در برخی از اعمال جراحی و همچنین در مطالعات آندوسکوپی از داروهایی استفاده می شود که باعث شل شدن عضلات اسکلتی می شود. (شل کننده های عضلانی).اینها شامل دیتیلین است که آنالوگ ساختاری استیل کولین است. به گیرنده های کولینرژیک H می چسبد، اما برخلاف استیل کولین، به آرامی توسط آنزیم استیل کولین استراز از بین می رود. در نتیجه باز شدن طولانی مدت کانال های یونی و دپلاریزاسیون مداوم غشاء، هدایت تکانه های عصبی مختل شده و شل شدن عضلات رخ می دهد. در ابتدا، این خواص در سم کورار کشف شد، به همین دلیل است که چنین داروهایی نامیده می شود کورار مانند
موضوع 1.3. دناتوره کردن پروتئین ها و امکان تجدید خودبه خودی آنها
1. از آنجایی که ترکیب بومی پروتئین ها به دلیل برهمکنش های ضعیف حفظ می شود، تغییر در ترکیب و خواص محیط اطراف پروتئین، قرار گرفتن در معرض معرف های شیمیایی و عوامل فیزیکی باعث تغییر در ترکیب آنها می شود (خاصیت ثبات ساختاری). شکستن تعداد زیادی از پیوندها منجر به تخریب ساختار بومی و دناتوره شدن پروتئین ها می شود.
دناتوره شدن پروتئین ها- این تخریب ساختار بومی آنها تحت تأثیر عوامل دناتوره کننده است که ناشی از پارگی پیوندهای ضعیف است که ساختار فضایی پروتئین را تثبیت می کند. دناتوره شدن با تخریب ساختار سه بعدی منحصر به فرد و مرکز فعال پروتئین و از دست دادن فعالیت بیولوژیکی آن همراه است (شکل 1.16).
تمام مولکولهای دناتورهشده یک پروتئین ترکیبی تصادفی پیدا میکنند که با سایر مولکولهای همان پروتئین متفاوت است. رادیکالهای اسید آمینهای که مرکز فعال را تشکیل میدهند، از نظر مکانی از یکدیگر دور هستند، یعنی. محل اتصال خاص پروتئین با لیگاند از بین می رود. در طول دناتوره شدن، ساختار اولیه پروتئین ها بدون تغییر باقی می ماند.
کاربرد عوامل دناتوره کننده در تحقیقات بیولوژیکی و پزشکی.در مطالعات بیوشیمیایی، قبل از تعیین ترکیبات با وزن مولکولی کم در مواد بیولوژیکی، معمولاً ابتدا پروتئین ها از محلول حذف می شوند. اسید تری کلرواستیک (TCA) اغلب برای این منظور استفاده می شود. پس از افزودن TCA به محلول، پروتئین های دناتوره شده رسوب می کنند و به راحتی با فیلتراسیون حذف می شوند (جدول 1.1).
در پزشکی، عوامل دناتوره کننده اغلب برای استریل کردن ابزار و مواد پزشکی در اتوکلاوها (عامل دناتوره کننده دمای بالا است) و به عنوان ضد عفونی کننده (الکل، فنل، کلرامین) برای درمان سطوح آلوده حاوی میکرو فلور بیماری زا استفاده می شود.
2. فعال شدن مجدد پروتئین خود به خود- اثبات قطعیت ساختار اولیه، ترکیب و عملکرد پروتئین ها. پروتئین های منفرد محصولات یک ژن هستند که دارای یک توالی اسید آمینه یکسان هستند و ترکیب مشابهی را در سلول به دست می آورند. نتیجه گیری اساسی که ساختار اولیه یک پروتئین از قبل حاوی اطلاعاتی در مورد ترکیب و عملکرد آن است، بر اساس توانایی برخی از پروتئین ها (به ویژه ریبونوکلئاز و میوگلوبین) برای بازسازی خود به خود - بازیابی ساختار اصلی آنها پس از دناتوره سازی ساخته شده است.
تشکیل ساختارهای پروتئینی فضایی با روش خودآرایی انجام می شود - یک فرآیند خود به خودی که در آن یک زنجیره پلی پپتیدی که دارای ساختار اولیه منحصر به فرد است، تمایل به اتخاذ یک ترکیب با کمترین انرژی آزاد در محلول دارد. توانایی احیای پروتئین هایی که ساختار اولیه خود را پس از دناتوره شدن حفظ می کنند، در آزمایشی با آنزیم ریبونوکلئاز توصیف شد.
ریبونوکلئاز آنزیمی است که پیوندهای بین نوکلئوتیدهای منفرد را در یک مولکول RNA تجزیه می کند. این پروتئین کروی دارای یک زنجیره پلی پپتیدی است که ساختار سوم آن توسط بسیاری از پیوندهای ضعیف و چهار دی سولفیدی تثبیت شده است.
درمان ریبونوکلئاز با اوره که پیوندهای هیدروژنی موجود در مولکول را می شکند و یک عامل احیا کننده که پیوندهای دی سولفیدی را می شکند، منجر به دناتوره شدن آنزیم و از دست دادن فعالیت آن می شود.
حذف عوامل دناتوره کننده با دیالیز منجر به ترمیم ساختار و عملکرد پروتئین می شود، به عنوان مثال. به تولد دوباره (شکل 1.17).
 برنج. 1.17. دناتوره سازی و احیای ریبونوکلئاز
برنج. 1.17. دناتوره سازی و احیای ریبونوکلئاز
الف - ترکیب بومی ریبونوکلئاز که در ساختار سوم آن چهار پیوند دی سولفیدی وجود دارد. ب - مولکول ریبونوکلئاز دناتوره شده.
ب - مولکول ریبونوکلئاز فعال شده با ساختار و عملکرد بازسازی شده
1. جدول 1.2 را پر کنید.
جدول 1.2. طبقه بندی اسیدهای آمینه بر اساس قطبیت رادیکال ها
2. فرمول تتراپپتید را بنویسید:
Asp - Pro - Fen - Liz
الف) گروههای تکرار شونده در پپتید که ستون فقرات پپتید را تشکیل میدهند و گروههای متغیری که توسط رادیکالهای اسید آمینه نشان داده میشوند را برجسته کنید.
ب) انتهای N و C را برچسب بزنید.
ج) پیوندهای پپتیدی را برجسته کنید.
د) پپتید دیگری متشکل از همان اسیدهای آمینه بنویسید.
ه) تعداد انواع احتمالی یک تتراپپتید با ترکیب اسید آمینه مشابه را بشمارید.
3. نقش ساختار اولیه پروتئین ها را با استفاده از مثالی از تجزیه و تحلیل مقایسه ای دو هورمون پپتیدی از نظر ساختاری مشابه و از نظر تکاملی نزدیک نوروهیپوفیز پستانداران - اکسی توسین و وازوپرسین توضیح دهید (جدول 1.3).
جدول 1.3. ساختار و عملکرد اکسی توسین و وازوپرسین
 برای این:
برای این:
الف) ترکیب و توالی اسید آمینه دو پپتید را مقایسه کنید.
ب) تشابه ساختار اولیه دو پپتید و شباهت عمل بیولوژیکی آنها را بیابید.
ج) تفاوت در ساختار دو پپتید و تفاوت در عملکرد آنها را بیابید.
د) در مورد تأثیر ساختار اولیه پپتیدها بر عملکرد آنها نتیجه گیری کنید.
4. مراحل اصلی تشکیل ترکیب پروتئین های کروی (ساختارهای ثانویه، سوم، مفهوم ساختار فوق ثانویه) را شرح دهید. انواع پیوندهای دخیل در تشکیل ساختارهای پروتئینی را مشخص کنید. کدام رادیکال های اسید آمینه می توانند در تشکیل فعل و انفعالات آبگریز، پیوندهای یونی و هیدروژنی شرکت کنند.
مثال بزن.
5. مفهوم "پایداری ساختاری پروتئین ها" را تعریف کنید، دلایل وجود و اهمیت آن را نشان دهید.
6. معنی عبارت زیر را بسط دهید: "عملکرد پروتئین ها بر اساس برهمکنش خاص آنها با لیگاند است" با استفاده از اصطلاحات و توضیح معنای آنها: ترکیب پروتئین، مرکز فعال، لیگاند، مکمل، عملکرد پروتئین.
7. با استفاده از یک مثال، توضیح دهید که دامنه ها چیست و نقش آنها در عملکرد پروتئین ها چیست.
وظایف خود کنترلی
1. همخوانی داشتن.
گروه عملکردی در رادیکال اسید آمینه:
A. گروه کربوکسیل B. گروه هیدروکسیل C گروه گوانیدین D. گروه تیول E. گروه آمینو
2. پاسخ های درست را انتخاب کنید.
اسیدهای آمینه با رادیکال های بدون بار قطبی عبارتند از:
الف سیس ب اصن
ب گلو جی سه
3. پاسخ های درست را انتخاب کنید.
رادیکال های اسید آمینه:
الف. ارائه ویژگی ساختار اولیه ب. مشارکت در شکل گیری ساختار سوم
ب- روی سطح پروتئین قرار دارند و بر حلالیت آن تأثیر می گذارند D. مرکز فعال را تشکیل می دهند
د. در تشکیل پیوندهای پپتیدی شرکت کنید
4. پاسخ های درست را انتخاب کنید.
فعل و انفعالات هیدروفوبیک می تواند بین رادیکال های اسید آمینه ایجاد شود:
A. Tre Lay B. Pro Three
بی مت ایل جی تیر آلا د وال فن
5. پاسخ های درست را انتخاب کنید.
پیوندهای یونی می توانند بین رادیکال های اسید آمینه ایجاد شوند:
آ.گلن آسپ ب.آپر لیز
بی لیز گلو جی گیس آسپ د اصن آوریل
6. پاسخ های درست را انتخاب کنید.
پیوندهای هیدروژنی می توانند بین رادیکال های اسید آمینه ایجاد شوند:
A. Ser Gln B. Cis Tre
ب آسپ لیز جی گلو آسپ د اسن تره
7. همخوانی داشتن.
نوع پیوند دخیل در تشکیل ساختار پروتئین:
الف- ساختار اولیه ب- ساختار ثانویه
ب- ساختار سوم
د. ساختار فوق ثانویه E. انطباق.
1. پیوندهای هیدروژنی بین اتم های ستون فقرات پپتیدی
2. پیوندهای ضعیف بین گروه های عاملی رادیکال های اسید آمینه
3. پیوند بین گروه های α-آمینو و α-کربوکسیل اسیدهای آمینه
8. پاسخ های درست را انتخاب کنید. تریپسین:
الف- آنزیم پروتئولیتیک ب- شامل دو حوزه است
ب- نشاسته را هیدرولیز می کند
د- سایت فعال بین دامنه ها قرار دارد. د- از دو زنجیره پلی پپتیدی تشکیل شده است.
9. پاسخ های درست را انتخاب کنید. آتروپین:
الف. انتقال دهنده عصبی
ب. آنالوگ ساختاری استیل کولین
ب- با گیرنده های H-cholinergic تعامل دارد
د- هدایت تکانه های عصبی را از طریق سیناپس های کولینرژیک تقویت می کند
د. بازدارنده رقابتی گیرنده های ام کولینرژیک
10. عبارات صحیح را انتخاب کنید. در پروتئین ها:
الف- ساختار اولیه حاوی اطلاعاتی در مورد ساختار سایت فعال آن است
ب- مرکز فعال در سطح ساختار اولیه تشکیل می شود
ب. ساختار به طور صلب توسط پیوندهای کووالانسی ثابت شده است
د- سایت فعال می تواند با گروهی از لیگاندهای مشابه تعامل داشته باشد
به دلیل ثبات ساختاری پروتئین ها D. تغییرات در محیط می تواند بر میل ترکیبی فعال تأثیر بگذارد.
مرکز به لیگاند
1. 1-B، 2-G، 3-B.
3. الف، ب، ج، د.
7. 1-B، 2-D، 3-A.
8. الف، ب، ج، د.
اصطلاحات و مفاهیم اساسی
1. پروتئین، پلی پپتید، اسیدهای آمینه
2. ساختارهای پروتئینی اولیه، ثانویه، سوم
3. ترکیب، ترکیب پروتئین بومی
4. پیوندهای کووالانسی و ضعیف در پروتئین
5. ثبات ساختاری
6. سایت فعال پروتئین
7. لیگاندها
8. تا زدن پروتئین
9. آنالوگ های ساختاری لیگاندها
10. پروتئین های دامنه
11. پروتئین های ساده و پیچیده
12. دناتوره شدن پروتئین، عوامل دناتوره کننده
13. فعال شدن مجدد پروتئین ها
حل مشکلات
"سازمان ساختاری پروتئین ها و اساس عملکرد آنها"
1. عملکرد اصلی پروتئین - هموگلوبین A (HbA) انتقال اکسیژن به بافت ها است. در جمعیت انسانی، اشکال متعددی از این پروتئین با خواص و عملکرد تغییر یافته شناخته شده است - به اصطلاح هموگلوبین های غیر طبیعی. به عنوان مثال، هموگلوبین S که در گلبول های قرمز بیماران مبتلا به بیماری سلول داسی شکل (HbS) یافت می شود، در شرایط فشار جزئی کم اکسیژن (همانطور که در خون وریدی وجود دارد) حلالیت کمی دارد. این منجر به تشکیل توده های این پروتئین می شود. پروتئین عملکرد خود را از دست می دهد، رسوب می کند و گلبول های قرمز به شکل نامنظمی در می آیند (برخی از آنها به شکل داسی شکل می گیرند) و سریعتر از حد معمول در طحال از بین می روند. در نتیجه کم خونی داسی شکل ایجاد می شود.
تنها تفاوت در ساختار اولیه HbA در ناحیه N ترمینال زنجیره β هموگلوبین مشاهده شد. مناطق N ترمینال زنجیره β را مقایسه کنید و نشان دهید که چگونه تغییرات در ساختار اولیه پروتئین بر خواص و عملکردهای آن تأثیر می گذارد.
 برای این:
برای این:
الف) فرمول اسیدهای آمینه را بنویسید که HbA با آنها تفاوت دارد و خواص این اسیدهای آمینه (قطبی، بار) را با هم مقایسه کنید.
ب) در مورد دلیل کاهش حلالیت و اختلال در انتقال اکسیژن به بافت ها نتیجه گیری کنید.
2. شکل، نموداری از ساختار پروتئینی را نشان می دهد که دارای مرکز اتصال با لیگاند (مرکز فعال) است. توضیح دهید که چرا پروتئین در انتخاب لیگاند انتخابی است. برای این:
الف) به یاد داشته باشید که مرکز فعال یک پروتئین چیست و ساختار مرکز فعال پروتئین نشان داده شده در شکل را در نظر بگیرید.
ب) فرمول رادیکال های اسید آمینه ای که مرکز فعال را تشکیل می دهند بنویسید.
ج) لیگاندی را رسم کنید که به طور خاص با محل فعال پروتئین تعامل داشته باشد. روی آن گروههای عاملی را که میتوانند با رادیکالهای اسید آمینهای که مرکز فعال را تشکیل میدهند پیوند ایجاد کنند، نشان دهید.
د) انواع پیوندهایی را که بین لیگاند و رادیکالهای اسید آمینه محل فعال ایجاد میشود، نشان دهد.
ه) توضیح دهید که ویژگی تعامل پروتئین-لیگاند بر چه اساس است.
 3.
شکل مکان فعال پروتئین و چندین لیگاند را نشان می دهد.
3.
شکل مکان فعال پروتئین و چندین لیگاند را نشان می دهد.
تعیین کنید کدام لیگاند به احتمال زیاد با محل فعال پروتئین تعامل دارد و چرا.
 چه نوع پیوندهایی در طی تشکیل کمپلکس پروتئین-لیگاند ایجاد می شوند؟
چه نوع پیوندهایی در طی تشکیل کمپلکس پروتئین-لیگاند ایجاد می شوند؟
4. آنالوگ های ساختاری لیگاندهای پروتئین طبیعی را می توان به عنوان دارو برای اصلاح فعالیت پروتئین ها استفاده کرد.
استیل کولین واسطه انتقال تحریک در سیناپس های عصبی عضلانی است. هنگامی که استیل کولین با پروتئین ها - گیرنده های غشای پس سیناپسی عضلات اسکلتی تعامل می کند، کانال های یونی باز می شوند و انقباض عضلانی رخ می دهد. دیتیلین دارویی است که در برخی از عملیات ها برای شل کردن عضلات استفاده می شود، زیرا انتقال تکانه های عصبی از طریق سیناپس های عصبی عضلانی را مختل می کند. مکانیسم اثر دیتیلین را به عنوان یک داروی شل کننده عضلانی توضیح دهید. برای این:
الف) فرمول های استیل کولین و دی تیلین را بنویسید و ساختار آنها را مقایسه کنید.
ب) مکانیسم اثر آرامش بخش دیتیلین را شرح دهد.
5. در برخی از بیماری ها دمای بدن بیمار بالا می رود که به عنوان واکنش محافظتی بدن محسوب می شود. با این حال، دمای بالا برای پروتئین های بدن مضر است. توضیح دهید که چرا در دمای بالای 40 درجه سانتیگراد عملکرد پروتئین ها مختل می شود و تهدیدی برای زندگی انسان ایجاد می شود. برای انجام این کار، به یاد داشته باشید:
1) ساختار پروتئین ها و پیوندهایی که ساختار آن را در ساختار بومی نگه می دارند.
2) ساختار و عملکرد پروتئین ها با افزایش دما چگونه تغییر می کند؟
3) هموستاز چیست و چرا برای حفظ سلامت انسان مهم است.
واحد مدولار 2 پروتئین های الیگومریک به عنوان اهداف تأثیرات تنظیمی. تنوع ساختاری و عملکردی پروتئین ها. روشهای جداسازی و خالصسازی پروتئینها
اهداف آموزشی قادر باشند:
1. از دانش در مورد ویژگی های ساختار و عملکرد پروتئین های الیگومری برای درک مکانیسم های تطبیقی تنظیم عملکرد آنها استفاده کنید.
2. نقش چپرون ها را در سنتز و حفظ ترکیب پروتئین در شرایط سلولی توضیح دهید.
3. انواع تظاهرات حیات را با تنوع ساختارها و عملکردهای پروتئین های سنتز شده در بدن توضیح دهید.
4. تجزیه و تحلیل رابطه بین ساختار پروتئین ها و عملکرد آنها با استفاده از مثال هایی از مقایسه هموپروتئین های مرتبط - میوگلوبین و هموگلوبین، و همچنین نمایندگان پنج کلاس پروتئین های خانواده ایمونوگلوبولین.
5. استفاده از دانش در مورد ویژگی های فیزیکی و شیمیایی پروتئین ها برای انتخاب روش های خالص سازی آنها از سایر پروتئین ها و ناخالصی ها.
6. نتایج ترکیب کمی و کیفی پروتئین های پلاسمای خون را برای تایید یا روشن شدن تشخیص بالینی تفسیر کنید.
بدانید:
1. ویژگی های ساختار پروتئین های الیگومری و مکانیسم های تطبیقی برای تنظیم عملکرد آنها با استفاده از مثال هموگلوبین.
2. ساختار و عملکرد چپرون ها و اهمیت آنها برای حفظ ترکیب بومی پروتئین ها در شرایط سلولی.
3. اصول ترکیب پروتئین ها در خانواده ها بر اساس شباهت ساختار و عملکرد آنها با استفاده از مثال ایمونوگلوبولین ها.
4. روش های جداسازی پروتئین ها بر اساس ویژگی های خواص فیزیکوشیمیایی آنها.
5. الکتروفورز پلاسمای خون به عنوان روشی برای ارزیابی ترکیب کیفی و کمی پروتئین ها.
موضوع 1.4. ویژگی های ساختار و عملکرد پروتئین های الیگومر با استفاده از نمونه هموگلوبین
1. بسیاری از پروتئین ها حاوی چندین زنجیره پلی پپتیدی هستند. چنین پروتئین هایی نامیده می شوند الیگومریک،و زنجیرهای فردی - پروتومرهاپروتومرها در پروتئین های الیگومری با بسیاری از پیوندهای غیر کووالانسی ضعیف (آب گریز، یونی، هیدروژن) به هم متصل می شوند. اثر متقابل
پروتومرها به لطف انجام می شوند مکمل بودنسطوح تماس آنها
تعداد پروتومرها در پروتئین های الیگومری می تواند بسیار متفاوت باشد: هموگلوبین حاوی 4 پروتومر، آنزیم آسپارتات آمینوترانسفراز دارای 12 پروتومر، و پروتئین ویروس موزاییک تنباکو حاوی 2120 پروتومر است که با پیوندهای غیر کووالانسی به هم متصل شده اند. در نتیجه، پروتئین های الیگومری می توانند وزن مولکولی بسیار بالایی داشته باشند.
برهمکنش یک پروتومر با دیگران را می توان مورد خاصی از برهمکنش پروتئین-لیگاند در نظر گرفت، زیرا هر پروتومر به عنوان لیگاند برای پروتومرهای دیگر عمل می کند. تعداد و روش اتصال پروتومرها در یک پروتئین نامیده می شود ساختار پروتئین چهارتایی
پروتئین ها می توانند حاوی پروتومرهایی با ساختارهای یکسان یا متفاوت باشند، به عنوان مثال، همودایمرها پروتئین هایی هستند که حاوی دو پروتومر یکسان هستند و هترودایمرها پروتئین هایی هستند که حاوی دو پروتومر متفاوت هستند.
اگر پروتئین ها حاوی پروتومرهای مختلفی باشند، می توان مراکز اتصال با لیگاندهای مختلف که از نظر ساختار متفاوت هستند روی آنها تشکیل شود. هنگامی که یک لیگاند به محل فعال متصل می شود، عملکرد این پروتئین آشکار می شود. مرکزی که روی یک پروتومر متفاوت قرار دارد آلوستریک (متفاوت از مرکز فعال) نامیده می شود. تماس گرفتن لیگاند یا عامل آلوستریک،یک عملکرد تنظیمی را انجام می دهد (شکل 1.18). برهمکنش مرکز آلوستریک با عامل باعث تغییرات ساختاری در ساختار کل پروتئین الیگومری به دلیل ثبات ساختاری آن می شود. این بر میل ترکیبی محل فعال برای یک لیگاند خاص تأثیر می گذارد و عملکرد آن پروتئین را تنظیم می کند. تغییر در ساختار و عملکرد همه پروتومرها در طی برهمکنش یک پروتئین الیگومری با حداقل یک لیگاند، تغییرات ساختاری مشارکتی نامیده می شود. عواملی که عملکرد پروتئین را تقویت می کنند نامیده می شوند فعال کننده ها،و عواملی که عملکرد آن را مهار می کنند هستند مهار کننده ها
بنابراین، پروتئین های الیگومری، و همچنین پروتئین هایی با ساختار دامنه، دارای ویژگی جدیدی در مقایسه با پروتئین های مونومر هستند - توانایی تنظیم آلوستریک عملکردها (تنظیم با اتصال لیگاندهای مختلف به پروتئین). این را می توان با مقایسه ساختار و عملکرد دو پروتئین پیچیده نزدیک به هم، میوگلوبین و هموگلوبین مشاهده کرد.
 برنج. 1.18. طرح ساختار یک پروتئین دیمری
برنج. 1.18. طرح ساختار یک پروتئین دیمری
2. تشکیل ساختارهای فضایی و عملکرد میوگلوبین.
میوگلوبین (Mb) پروتئینی است که در ماهیچه های قرمز یافت می شود و وظیفه اصلی آن ایجاد ذخایر O 2 لازم برای کار شدید عضلات است. Mb یک پروتئین پیچیده حاوی یک بخش پروتئینی - apoMb و یک بخش غیر پروتئینی - هم است. ساختار اولیه apoMB ساختار کروی فشرده آن و ساختار مرکز فعال را تعیین می کند که بخش غیر پروتئینی میوگلوبین، هِم، به آن متصل است. اکسیژنی که از خون به ماهیچه ها می رسد به هم های Fe+2 در میوگلوبین متصل می شود. Mb یک پروتئین مونومر است که میل ترکیبی بسیار بالایی برای O 2 دارد، بنابراین آزاد شدن اکسیژن توسط میوگلوبین تنها در حین کار شدید عضلانی رخ می دهد، زمانی که فشار جزئی O 2 به شدت کاهش می یابد.
تشکیل ترکیب Mv.در ماهیچه های قرمز، روی ریبوزوم ها در حین ترجمه، ساختار اولیه MB سنتز می شود که با دنباله خاصی از 153 باقی مانده اسید آمینه نشان داده می شود. ساختار ثانویه Mb شامل هشت مارپیچ α است که با حروف لاتین از A تا H نامیده می شوند و بین آنها مناطق غیر مارپیچ وجود دارد. ساختار سوم Mb به شکل یک گلبول فشرده است که در شکاف آن مرکز فعال بین مارپیچ های F و E قرار دارد (شکل 1.19).
 برنج. 1.19. ساختار میوگلوبین
برنج. 1.19. ساختار میوگلوبین
3. ویژگی های ساختار و عملکرد مرکز فعال MV.مرکز فعال Mb عمدتاً توسط رادیکال های اسید آمینه آبگریز تشکیل شده است که در ساختار اولیه به طور گسترده از یکدیگر فاصله دارند (به عنوان مثال Tri 3 9 و Fen 138) در لیگاندهای آب محلول ضعیف - هم و O 2 - به مرکز فعال متصل می شوند. Heme یک لیگاند خاص apoMB است (شکل 1.20)، که اساس آن از چهار حلقه پیرول تشکیل شده است که توسط پل های متنیل به هم متصل شده اند. در مرکز یک اتم Fe + 2 وجود دارد که با چهار پیوند هماهنگی به اتم های نیتروژن حلقه های پیرول متصل است. در مرکز فعال Mb، علاوه بر رادیکال های اسید آمینه آبگریز، بقایای دو اسید آمینه با رادیکال های آبدوست نیز وجود دارد - Gis E 7(Gis 64) و GIS F 8(93 او) (شکل 1.21).
 برنج. 1.20. ساختار هم - بخش غیر پروتئینی میوگلوبین و هموگلوبین
برنج. 1.20. ساختار هم - بخش غیر پروتئینی میوگلوبین و هموگلوبین
 برنج. 1.21. محل هم و O2 در محل فعال آپومیوگلوبین و پروتومرهای هموگلوبین
برنج. 1.21. محل هم و O2 در محل فعال آپومیوگلوبین و پروتومرهای هموگلوبین
هِم به صورت کووالانسی به F8 از طریق یک اتم آهن متصل می شود. O 2 به آهن در طرف دیگر صفحه هم می چسبد. E 7 او برای جهت گیری صحیح O 2 ضروری است و افزودن اکسیژن به هم Fe + 2 را تسهیل می کند.
GIS F 8یک پیوند هماهنگی با Fe+2 ایجاد می کند و هم را در مرکز فعال محکم می کند. Gis E 7برای جهت گیری صحیح در مرکز فعال لیگاند دیگر - O 2 در طول برهمکنش آن با Fe + 2 Heme ضروری است. ریزمحیط هِم شرایطی را برای اتصال قوی اما برگشت پذیر O 2 به Fe + 2 ایجاد می کند و از ورود آب به محل فعال آبگریز جلوگیری می کند که می تواند منجر به اکسیداسیون آن به Fe + 3 شود.
ساختار مونومر Mb و مرکز فعال آن میل ترکیبی بالای پروتئین را برای O 2 تعیین می کند.
4. ساختار الیگومری Hb و تنظیم میل ترکیبی Hb به لیگاندهای O 2. هموگلوبین های انسانی- خانواده ای از پروتئین ها، مانند میوگلوبین، مربوط به پروتئین های پیچیده (هموپروتئین ها). آنها ساختار تترامری دارند و شامل دو زنجیره α هستند، اما در ساختار دو زنجیره پلی پپتیدی دیگر (زنجیره 2α-، 2x) متفاوت هستند. ساختار دومین زنجیره پلی پپتیدی ویژگی های عملکرد این اشکال Hb را تعیین می کند. حدود 98 درصد از هموگلوبین موجود در گلبول های قرمز خون بزرگسالان است هموگلوبین A(2α-، 2p-زنجیره).
در طول رشد جنین، دو نوع اصلی هموگلوبین عمل می کنند: Hb جنینی(2α, 2ε) که در مراحل اولیه رشد جنین یافت می شود و هموگلوبین F (جنین)- (2α, 2γ) که جایگزین هموگلوبین اولیه جنین در ماه ششم رشد داخل رحمی می شود و تنها پس از تولد با Hb A جایگزین می شود.
HB A یک پروتئین مربوط به میوگلوبین (MB) است که در گلبول های قرمز خون انسان بالغ یافت می شود. ساختار تک تک پروتومرهای آن شبیه به میوگلوبین است. ساختارهای ثانویه و ثانویه پروتومرهای میوگلوبین و هموگلوبین بسیار شبیه هستند، علیرغم این واقعیت که در ساختار اولیه زنجیره های پلی پپتیدی آنها تنها 24 باقیمانده اسید آمینه یکسان هستند (ساختار ثانویه پروتومرهای هموگلوبین، مانند میوگلوبین، حاوی هشت مارپیچ α است. با حروف لاتین از A تا H مشخص می شود و ساختار سوم به شکل یک کروی فشرده است). اما بر خلاف میوگلوبین، هموگلوبین دارای ساختار الیگومری است که شامل چهار زنجیره پلی پپتیدی است که با پیوندهای غیر کووالانسی به هم متصل شده اند (شکل 1.22).
هر پروتومر Hb با یک بخش غیر پروتئینی مرتبط است - هم و پروتومرهای مجاور. ارتباط بخش پروتئینی Hb با هم شبیه به میوگلوبین است: در مرکز فعال پروتئین، قسمت های آبگریز هم توسط رادیکال های اسید آمینه آبگریز احاطه شده است به استثنای His F 8 و His E 7. که در دو طرف صفحه هم قرار دارند و نقش مشابهی در عملکرد پروتئین و اتصال آن با اکسیژن دارند (به ساختار میوگلوبین مراجعه کنید).
 برنج. 1.22. ساختار الیگومری هموگلوبین
برنج. 1.22. ساختار الیگومری هموگلوبین
بعلاوه، Gis E 7مهمی را انجام می دهد نقش اضافیدر عملکرد Nv. هِم آزاد 25000 برابر بیشتر میل ترکیبی برای CO نسبت به O2 دارد. CO در مقادیر کمی در بدن تشکیل می شود و با توجه به تمایل زیاد آن به هم، می تواند انتقال O 2 لازم برای زندگی سلولی را مختل کند. با این حال، در ترکیب هموگلوبین، میل ترکیبی هم برای مونوکسید کربن تنها 200 برابر بیشتر از میل ترکیبی برای O 2 به دلیل وجود His E 7 در مرکز فعال است. باقیمانده این اسید آمینه شرایط بهینه را برای اتصال هم به O 2 ایجاد می کند و برهمکنش هم با CO را ضعیف می کند.
5. وظیفه اصلی HB انتقال O2 از ریه ها به بافت ها است.بر خلاف میوگلوبین مونومر که میل ترکیبی بسیار بالایی با O2 دارد و وظیفه ذخیره اکسیژن را در عضلات قرمز انجام می دهد، ساختار الیگومری هموگلوبین فراهم می کند:
1) اشباع سریع HB با اکسیژن در ریه ها.
2) توانایی HB برای آزادسازی اکسیژن در بافت ها با فشار جزئی نسبتاً بالای O 2 (20-40 میلی متر جیوه).
3) امکان تنظیم میل Hb به O 2.
6. تغییرات مشارکتی در ترکیب پروتومرهای هموگلوبین باعث تسریع اتصال O 2 در ریه ها و انتشار آن در بافت ها می شود. در ریه ها، فشار جزئی بالای O 2 باعث اتصال آن به Hb در محل فعال چهار پروتومر (2α و 2β) می شود. مرکز فعال هر پروتومر، مانند میوگلوبین، بین دو مارپیچ α (F و E) در یک پاکت آبگریز قرار دارد. این شامل یک بخش غیر پروتئینی - هِم است که با بسیاری از فعل و انفعالات آبگریز ضعیف و یک پیوند قوی بین Fe 2 + هِم و His F 8 به بخش پروتئینی متصل شده است (شکل 1.21 را ببینید).
در دئوکسی هموگلوبین، به دلیل این پیوند با His F 8، اتم Fe 2 + از صفحه هم به سمت هیستیدین بیرون زده است. اتصال O 2 به Fe 2 + در طرف دیگر هم در ناحیه His E 7 با استفاده از یک پیوند هماهنگی آزاد منفرد اتفاق می افتد. E 7 او شرایط بهینه را برای اتصال O 2 به آهن هم فراهم می کند.
افزودن O 2 به اتم Fe + 2 یک پروتومر باعث حرکت آن به صفحه هم می شود و به دنبال آن باقی مانده هیستیدین مرتبط با آن است.
 برنج. 1.23. تغییر در ساختار پروتومر هموگلوبین هنگام ترکیب با O2
برنج. 1.23. تغییر در ساختار پروتومر هموگلوبین هنگام ترکیب با O2
این منجر به تغییر در ساختار تمام زنجیره های پلی پپتیدی به دلیل ثبات ساختاری آنها می شود. تغییر ساختار زنجیره های دیگر تعامل آنها با مولکول های بعدی O 2 را تسهیل می کند.
چهارمین مولکول O 2 300 برابر راحت تر از مولکول اول به هموگلوبین می چسبد (شکل 1.24).
 برنج. 1.24. تغییرات مشارکتی در ترکیب پروتومرهای هموگلوبین در طول تعامل آن با O2
برنج. 1.24. تغییرات مشارکتی در ترکیب پروتومرهای هموگلوبین در طول تعامل آن با O2
در بافتها، هر مولکول O2 بعدی راحتتر از مولکول قبلی جدا میشود، همچنین به دلیل تغییرات مشترک در ساختار پروتومرها.
7. CO 2 و H + که در طی کاتابولیسم مواد آلی تشکیل می شوند، میل هموگلوبین به O 2 را به نسبت غلظت آنها کاهش می دهند. انرژی مورد نیاز برای عملکرد سلول عمدتاً در میتوکندری در طی اکسیداسیون مواد آلی با استفاده از O 2 که توسط هموگلوبین از ریه ها تحویل می شود تولید می شود. در نتیجه اکسیداسیون مواد آلی، محصولات نهایی تجزیه آنها تشکیل می شود: CO 2 و K 2 O که مقدار آنها متناسب با شدت فرآیندهای اکسیداسیون در حال انجام است.
CO 2 از سلول ها به خون منتشر می شود و به گلبول های قرمز نفوذ می کند و در آنجا تحت تأثیر آنزیم کربان هیدراز به اسید کربنیک تبدیل می شود. این اسید ضعیف به یک پروتون و یک یون بی کربنات تجزیه می شود.
H+ قادر به پیوستن به رادیکال های او هستند 14 6 در زنجیره های α و β هموگلوبین، به عنوان مثال. در مناطق دور از هم پروتوناسیون هموگلوبین میل آن را برای O 2 کاهش می دهد، باعث حذف O 2 از oxyHb، تشکیل deoxyHb می شود و عرضه اکسیژن به بافت ها را متناسب با تعداد پروتون های تشکیل شده افزایش می دهد (شکل 1.25).
افزایش در میزان اکسیژن آزاد شده بسته به افزایش غلظت H+ در گلبول های قرمز، اثر بور نامیده می شود (به نام فیزیولوژیست دانمارکی کریستین بور، که اولین بار این اثر را کشف کرد).
در ریه ها، فشار جزئی بالای اکسیژن باعث اتصال آن به deoxyHb می شود که میل پروتئین را به H + کاهش می دهد. پروتون های آزاد شده تحت اثر اسید کربنیک با بی کربنات ها واکنش داده و CO 2 و H 2 O را تشکیل می دهند.
 برنج. 1.25. وابستگی تمایل Hb به O 2 به غلظت CO 2 و پروتون ها (اثر Bohr):
برنج. 1.25. وابستگی تمایل Hb به O 2 به غلظت CO 2 و پروتون ها (اثر Bohr):
آ- تأثیر غلظت CO 2 و H + در آزادسازی O 2 از مجتمع با HB (اثر Bohr). ب- اکسیژن رسانی دی اکسی هموگلوبین در ریه ها، تشکیل و آزادسازی CO 2.
CO 2 حاصل وارد فضای آلوئولی شده و با هوای بازدمی خارج می شود. بنابراین، مقدار اکسیژن آزاد شده توسط هموگلوبین در بافت ها توسط محصولات کاتابولیسم مواد آلی تنظیم می شود: هرچه تجزیه مواد شدیدتر باشد، به عنوان مثال در حین تمرین بدنی، غلظت CO 2 و H + بیشتر و اکسیژن بیشتر می شود. بافت ها در نتیجه کاهش میل ترکیبی Hb برای O 2 دریافت می کنند.
8. تنظیم آلوستریک میل Hb برای O2 توسط لیگاند - 2،3-bisphosphoglycerate.در گلبول های قرمز، لیگاند آلوستریک هموگلوبین، 2،3-بیس فسفوگلیسرات (2،3-BPG)، از محصول اکسیداسیون گلوکز - 1،3-بیس فسفوگلیسرات سنتز می شود. در شرایط عادی، غلظت 2،3-BPG بالا و قابل مقایسه با غلظت Hb است. 2,3-BPG دارای بار منفی قوی 5- است.
 بیس فسفوگلیسرات در مویرگ های بافتی که به دی اکسی هموگلوبین متصل می شود، آزادسازی اکسیژن را در بافت ها افزایش می دهد و میل هموگلوبین به O 2 را کاهش می دهد.
بیس فسفوگلیسرات در مویرگ های بافتی که به دی اکسی هموگلوبین متصل می شود، آزادسازی اکسیژن را در بافت ها افزایش می دهد و میل هموگلوبین به O 2 را کاهش می دهد.
در مرکز مولکول هموگلوبین تترامریک یک حفره وجود دارد. این توسط بقایای اسید آمینه هر چهار پروتومر تشکیل می شود (شکل 1.22 را ببینید). در مویرگ های بافتی، پروتونه شدن Hb (اثر Bohr) منجر به گسیختگی پیوند بین آهن هِم و O2 می شود. در یک مولکول
دئوکسی هموگلوبین، در مقایسه با اکسی هموگلوبین، پیوندهای یونی اضافی ظاهر می شود که پروتومرها را به هم متصل می کند، در نتیجه ابعاد حفره مرکزی در مقایسه با اکسی هموگلوبین افزایش می یابد. حفره مرکزی محل اتصال 2،3-BPG به هموگلوبین است. به دلیل تفاوت در اندازه حفره مرکزی، 2،3-BPG فقط می تواند به دئوکسی هموگلوبین متصل شود.
2،3-BPG با هموگلوبین در محلی دور از مراکز فعال پروتئین تعامل دارد و متعلق به آلوستریکلیگاندهای (تنظیمی) و حفره مرکزی Hb است مرکز آلوستریک 2،3-BPG دارای بار منفی قوی است و با پنج گروه با بار مثبت از دو زنجیره β Hb تعامل می کند: گروه N ترمینال α-آمینه Val و رادیکال Lys 82 His 143 (شکل 1.26).
 برنج. 1.26. BPG در حفره مرکزی دئوکسی هموگلوبین
برنج. 1.26. BPG در حفره مرکزی دئوکسی هموگلوبین
BPG به سه گروه با بار مثبت در هر رشته β متصل می شود.
در مویرگ های بافتی، دئوکسی هموگلوبین حاصل با 2،3-BPG برهمکنش می کند و پیوندهای یونی بین رادیکال های با بار مثبت زنجیره های β و لیگاند با بار منفی تشکیل می شود که ترکیب پروتئین را تغییر می دهد و میل هموگلوبین به O2 را کاهش می دهد. . کاهش میل ترکیبی هموگلوبین برای O 2 باعث آزادسازی موثرتر O 2 در بافت می شود.
در ریه ها، در فشار جزئی بالا، اکسیژن با هموگلوبین برهمکنش می کند و به آهن هِم می پیوندد. در این حالت، ترکیب پروتئین تغییر می کند، حفره مرکزی کاهش می یابد و 2،3-BPG از مرکز آلوستریک جابجا می شود.
بنابراین پروتئین های الیگومری در مقایسه با پروتئین های مونومری خواص جدیدی دارند. چسباندن لیگاندها در سایت ها
از نظر مکانی از یکدیگر دور هستند (آلوستریک)، می توانند باعث تغییرات ساختاری در کل مولکول پروتئین شوند. به دلیل تعامل با لیگاندهای تنظیمی، تغییر در ترکیب و انطباق عملکرد مولکول پروتئین با تغییرات محیطی رخ می دهد.
موضوع 1.5. حفظ ساختار بومی پروتئین ها تحت شرایط سلولی
در سلول ها، در طول سنتز زنجیره های پلی پپتیدی، حمل و نقل آنها از طریق غشاء به قسمت های مربوطه سلول، در طول فرآیند تاخوردگی (تشکیل ترکیب بومی) و در طول مونتاژ پروتئین های الیگومری، و همچنین در طول عملکرد آنها، میانجی است. ، ترکیبات مستعد تجمع و ناپایدار در ساختار پروتئین بوجود می آیند. رادیکالهای آبگریز که معمولاً در داخل مولکول پروتئین در ترکیب اصلی پنهان میشوند، در یک ترکیب ناپایدار روی سطح ظاهر میشوند و تمایل دارند با گروههایی از پروتئینهای دیگر که در آب محلول نیستند ترکیب شوند. در سلولهای همه موجودات شناخته شده، پروتئینهای خاصی یافت شدهاند که چینخوردگی بهینه پروتئینهای سلولی را تضمین میکنند، ساختار بومی آنها را در طول عملکرد تثبیت میکنند و مهمتر از همه، ساختار و عملکرد پروتئینهای درون سلولی را در هنگام اختلال هموستاز حفظ میکنند. این پروتئین ها نامیده می شوند "همراهان"که در فرانسوی به معنی پرستار بچه است.
1. چاپرون های مولکولی و نقش آنها در جلوگیری از دناتوره شدن پروتئین.
چاپرون ها (CH) بر اساس جرم زیر واحدهایشان طبقه بندی می شوند. شاپرون های با وزن مولکولی بالا دارای جرمی بین 60 تا 110 کیلو دالتون هستند. در میان آنها، سه کلاس بیشتر مورد مطالعه قرار گرفته است: Sh-60، Sh-70 و Sh-90. هر کلاس شامل یک خانواده از پروتئین های مرتبط است. بنابراین، Sh-70 حاوی پروتئین هایی با وزن مولکولی از 66 تا 78 کیلو دالتون است. شاپرون های با وزن مولکولی پایین دارای وزن مولکولی بین 40 تا 15 کیلو دالتون هستند.
در میان همراهان وجود دارد سازندهپروتئین هایی که سنتز پایه بالایی آنها به اثرات استرس بر سلول های بدن بستگی ندارد و القایی،سنتز آن در شرایط عادی ضعیف است، اما تحت استرس به شدت افزایش می یابد. چاپرون های القایی "پروتئین های شوک حرارتی" نیز نامیده می شوند زیرا برای اولین بار در سلول هایی که در معرض دمای بالا قرار داشتند کشف شدند. در سلول ها به دلیل غلظت بالای پروتئین ها، فعال سازی مجدد خود به خودی پروتئین های نیمه دناتوره شده مشکل است. Sh-70 می تواند از شروع دناتوره شدن جلوگیری کند و به بازیابی ساختار بومی پروتئین ها کمک کند. چاپرون های مولکولی-70- یک کلاس بسیار حفاظت شده از پروتئین ها که در تمام قسمت های سلول یافت می شود: سیتوپلاسم، هسته، شبکه آندوپلاسمی، میتوکندری. در انتهای کربوکسیل زنجیره پلی پپتیدی منفرد Ш-70 ناحیه ای وجود دارد که یک شیار است که قادر به تعامل با پپتیدهای طولی است.
از 7 تا 9 باقی مانده اسید آمینه غنی شده با رادیکال های آبگریز. چنین مناطقی در پروتئین های کروی تقریباً در هر 16 اسید آمینه وجود دارد. Sh-70 قادر به محافظت از پروتئین ها در برابر غیرفعال شدن دما و بازیابی ساختار و فعالیت پروتئین های نیمه دناتوره شده است.
2. نقش چاپرون ها در تاخوردگی پروتئین.در طول سنتز پروتئین روی ریبوزوم، ناحیه N ترمینال پلی پپتید قبل از C ترمینال سنتز می شود. برای تشکیل ترکیب بومی، توالی اسید آمینه کامل پروتئین مورد نیاز است. در فرآیند سنتز پروتئین، چاپرون-70، به دلیل ساختار مرکز فعال خود، قادرند مناطقی از پلی پپتید را که مستعد تجمع هستند، تا زمانی که سنتز کامل شود، غنی شده با رادیکال های اسید آمینه آبگریز ببندند (شکل 1.27، A. ).
 برنج. 1.27. مشارکت چپرون ها در تاخوردگی پروتئین
برنج. 1.27. مشارکت چپرون ها در تاخوردگی پروتئین
الف - مشارکت chaperones-70 در جلوگیری از فعل و انفعالات آبگریز بین بخشهای پلی پپتید سنتز شده. ب - تشکیل ترکیب بومی پروتئین در کمپلکس چاپرون
بسیاری از پروتئین های مولکولی بالا که ترکیب پیچیده ای دارند، مانند ساختار دامنه، در فضای خاصی که توسط Sh-60 تشکیل شده است، جمع می شوند. Ш-60به عنوان یک کمپلکس الیگومری متشکل از 14 زیر واحد عمل می کند. آنها دو حلقه توخالی را تشکیل می دهند که هر یک از هفت زیر واحد تشکیل شده است که این حلقه ها به یکدیگر متصل هستند. هر زیر واحد Sh-60 از سه حوزه تشکیل شده است: آپیکال (آپیکال)، غنی شده با رادیکال های آبگریز رو به حفره حلقه، میانی و استوایی (شکل 1.28).
 برنج. 1.28. ساختار کمپلکس چاپرونین متشکل از 14 Ш-60
برنج. 1.28. ساختار کمپلکس چاپرونین متشکل از 14 Ш-60
الف - نمای جانبی؛ ب - نمای بالا
پروتئین های سنتز شده، که دارای عناصری بر روی سطح مشخصه مولکول های باز شده، به ویژه رادیکال های آبگریز هستند، وارد حفره حلقه های چاپرون می شوند. در محیط خاص این حفره ها، انطباق های ممکن جستجو می شوند تا زمانی که تنها موردی که از نظر انرژی مطلوب تر است پیدا شود (شکل 1.27، B). تشکیل ترکیبات و آزادسازی پروتئین با هیدرولیز ATP در ناحیه استوایی همراه است. به طور معمول، چنین تا کردن وابسته به چاپرون به مقدار قابل توجهی انرژی نیاز دارد.
چپرون ها علاوه بر مشارکت در تشکیل ساختار سه بعدی پروتئین ها و احیای پروتئین های نیمه دناتوره شده، برای وقوع فرآیندهای اساسی مانند تجمع پروتئین های الیگومری، شناسایی و انتقال پروتئین های دناتوره شده به لیزوزوم ها نیز ضروری هستند. انتقال پروتئین ها از طریق غشاها و مشارکت در تنظیم فعالیت کمپلکس های پروتئینی.
موضوع 1.6. انواع پروتئین ها. خانواده های پروتئینی: نمونه ای از ایمونوگلوبولین ها
1. پروتئین ها نقش تعیین کننده ای در زندگی تک تک سلول ها و کل ارگانیسم چند سلولی دارند و عملکرد آنها به طرز شگفت انگیزی متنوع است. این با ویژگی های ساختار اولیه و ترکیبات پروتئین ها، ساختار منحصر به فرد مرکز فعال و توانایی اتصال لیگاندهای خاص تعیین می شود.
تنها بخش بسیار کوچکی از تمام انواع احتمالی زنجیرههای پپتیدی میتوانند یک ساختار فضایی پایدار اتخاذ کنند. اکثریت
از آنها می توانند بسیاری از ترکیبات را با انرژی تقریباً یکسان گیبس، اما با خواص متفاوت، بگیرند. ساختار اولیه اکثر پروتئین های شناخته شده انتخاب شده توسط تکامل بیولوژیکی، ثبات استثنایی یکی از ترکیبات را تضمین می کند، که ویژگی های عملکرد این پروتئین را تعیین می کند.
2. خانواده های پروتئینی.در یک گونه بیولوژیکی، جایگزینی بقایای اسید آمینه می تواند منجر به ظهور پروتئین های مختلف شود که عملکردهای مرتبط را انجام می دهند و دارای توالی اسید آمینه همولوگ هستند. چنین پروتئینهای مرتبط دارای ترکیبهای بسیار مشابهی هستند: تعداد و موقعیتهای نسبی مارپیچهای α و/یا ساختارهای β، و بیشتر چرخشها و خمهای زنجیرههای پلی پپتیدی مشابه یا یکسان هستند. پروتئین هایی با مناطق همولوگ زنجیره پلی پپتیدی، ترکیب مشابه و عملکردهای مرتبط در خانواده های پروتئینی طبقه بندی می شوند. نمونه هایی از خانواده های پروتئینی: سرین پروتئینازها، خانواده ایمونوگلوبولین ها، خانواده میوگلوبین ها.
پروتئینازهای سرین- خانواده ای از پروتئین ها که عملکرد آنزیم های پروتئولیتیک را انجام می دهند. اینها شامل آنزیم های گوارشی - کیموتریپسین، تریپسین، الاستاز و بسیاری از فاکتورهای لخته شدن خون است. این پروتئین ها دارای اسیدهای آمینه یکسان در 40 درصد موقعیت های خود و ترکیب بسیار مشابهی هستند (شکل 1.29).

برنج. 1.29. ساختارهای فضایی الاستاز (A) و کیموتریپسین (B)
برخی از جایگزینی اسیدهای آمینه منجر به تغییر در ویژگی سوبسترای این پروتئین ها و ظهور تنوع عملکردی در خانواده شده است.
3. خانواده ایمونوگلوبولین.در عملکرد سیستم ایمنی، پروتئین های ابرخانواده ایمونوگلوبولین ها نقش بزرگی ایفا می کنند که شامل سه خانواده پروتئین است:
آنتی بادی ها (ایمونوگلوبولین ها)؛
گیرنده های لنفوسیت T؛
پروتئین های کمپلکس اصلی سازگاری بافتی - کلاس های MHC 1 و 2 (Major Histocompatibility Complex).
همه این پروتئینها ساختار دامنه دارند، از حوزههای همولوگ شبیه ایمنی تشکیل شدهاند و عملکردهای مشابهی را انجام میدهند: آنها با ساختارهای خارجی، یا محلول در خون، لنف یا مایع بین سلولی (آنتی بادیها)، یا در سطح سلولها (خود یا قرار دارند) تعامل دارند. خارجی).
4. آنتی بادی ها- پروتئین های خاصی که توسط لنفوسیت های B در پاسخ به ورود یک ساختار خارجی به بدن تولید می شوند، نامیده می شوند آنتی ژن
ویژگی های ساختار آنتی بادی ها
ساده ترین مولکول های آنتی بادی از چهار زنجیره پلی پپتیدی تشکیل شده است: دو زنجیره سبک یکسان - L، حاوی حدود 220 اسید آمینه، و دو نوع سنگین یکسان - H، متشکل از 440-700 اسید آمینه. هر چهار زنجیره در مولکول آنتی بادی توسط بسیاری از پیوندهای غیر کووالانسی و چهار پیوند دی سولفیدی به هم متصل هستند (شکل 1.30).
زنجیره های سبک آنتی بادی از دو حوزه تشکیل شده اند: یک دامنه متغیر (VL) که در ناحیه N ترمینال زنجیره پلی پپتیدی قرار دارد و یک دامنه ثابت (CL) که در انتهای C قرار دارد. زنجیرههای سنگین معمولاً دارای چهار حوزه هستند: یک متغیر (VH) که در انتهای N قرار دارد و سه حوزه ثابت (CH1، CH2، CH3) (شکل 1.30 را ببینید). هر دامنه ایمونوگلوبولین دارای یک روبنای ورقه β است که در آن دو باقیمانده سیستئین توسط یک پیوند دی سولفیدی به هم متصل شده اند.
بین دو حوزه ثابت CH1 و CH2 یک ناحیه حاوی تعداد زیادی باقیمانده پرولین وجود دارد که از تشکیل ساختار ثانویه و تعامل زنجیرههای H همسایه در این بخش جلوگیری میکند. این ناحیه لولا به مولکول آنتی بادی انعطاف پذیری می دهد. بین حوزه های متغیر زنجیره های سنگین و سبک دو محل اتصال آنتی ژن یکسان (محل های فعال برای اتصال آنتی ژن ها) وجود دارد، بنابراین چنین آنتی بادی هایی اغلب نامیده می شوند. دو ظرفیتینه کل توالی اسید آمینه نواحی متغیر هر دو زنجیره در اتصال آنتی ژن به آنتی بادی دخالت دارد، بلکه تنها 20-30 اسید آمینه در نواحی بیش متغیر هر زنجیره قرار دارند. این مناطق هستند که توانایی منحصر به فرد هر نوع آنتی بادی را برای برهمکنش با آنتی ژن مکمل مربوطه تعیین می کنند.
آنتی بادی ها یکی از خطوط دفاعی بدن در برابر موجودات خارجی مهاجم هستند. عملکرد آنها را می توان به دو مرحله تقسیم کرد: مرحله اول شناسایی و اتصال آنتی ژن بر روی سطح ارگانیسم های خارجی است که به دلیل وجود محل های اتصال آنتی ژن در ساختار آنتی بادی امکان پذیر است. مرحله دوم شروع فرآیند غیرفعال سازی و تخریب آنتی ژن است. ویژگی مرحله دوم به کلاس آنتی بادی ها بستگی دارد. پنج کلاس از زنجیره های سنگین وجود دارد که در ساختار حوزه های ثابت با یکدیگر متفاوت هستند: α، δ، ε، γ و μ، که بر اساس آنها پنج کلاس ایمونوگلوبولین ها متمایز می شوند: A، D، E، G و M.
ویژگیهای ساختاری زنجیرههای سنگین به نواحی لولا و نواحی C ترمینال زنجیرههای سنگین یک ویژگی ترکیبی از هر کلاس میدهد. پس از اتصال آنتی ژن به آنتی بادی، تغییرات ساختاری در حوزه های ثابت مسیر حذف آنتی ژن را تعیین می کند.
 برنج. 1. 30. ساختار دامنه IgG
برنج. 1. 30. ساختار دامنه IgG
ایمونوگلوبولین M
ایمونوگلوبولین های M دو شکل دارند.
فرم مونومر- کلاس 1 آنتی بادی های تولید شده توسط لنفوسیت های B. متعاقبا، بسیاری از سلولهای B به تولید کلاسهای دیگر آنتیبادیها، اما با همان محل اتصال آنتیژن، روی میآورند. IgM در غشاء تعبیه شده است و به عنوان یک گیرنده تشخیص آنتی ژن عمل می کند. ادغام IgM در غشای سلولی به دلیل وجود 25 باقی مانده اسید آمینه آبگریز در ناحیه دم امکان پذیر است.
فرم ترشحی IgMشامل پنج زیر واحد مونومر است که توسط پیوندهای دی سولفیدی و یک زنجیره پلی پپتیدی J اضافی به یکدیگر متصل شده اند (شکل 1.31). زنجیره های سنگین مونومرهای این شکل حاوی دم آبگریز نیستند. پنتامر دارای 10 محل اتصال آنتی ژن است و بنابراین در شناسایی و حذف آنتی ژنی که ابتدا وارد بدن می شود موثر است. شکل ترشحی IgM کلاس اصلی آنتی بادی هایی است که در طی پاسخ ایمنی اولیه به خون ترشح می شود. اتصال IgM به یک آنتی ژن، ترکیب IgM را تغییر می دهد و باعث اتصال آن به اولین جزء پروتئینی سیستم کمپلمان (سیستم کمپلمان مجموعه ای از پروتئین های دخیل در تخریب آنتی ژن است) و فعال شدن این سیستم می شود. اگر آنتی ژن روی سطح میکروارگانیسم قرار گیرد، سیستم کمپلمان باعث اختلال در یکپارچگی غشای سلولی و مرگ سلول باکتری می شود.
ایمونوگلوبولین G
از نظر کمی، این دسته از ایمونوگلوبولین ها در خون غالب هستند (75٪ از کل Igs). IgG - مونومرها، دسته اصلی آنتی بادی هایی هستند که در طی یک پاسخ ایمنی ثانویه در خون ترشح می شوند. پس از برهمکنش IgG با آنتی ژن های سطحی میکروارگانیسم ها، کمپلکس آنتی ژن-آنتی بادی قادر است پروتئین های سیستم کمپلمان را متصل و فعال کند یا می تواند با گیرنده های خاص ماکروفاژها و نوتروفیل ها تعامل داشته باشد. تعامل با فاگوسیت ها منجر می شود
 برنج. 1.31. ساختار فرم ترشحی IgM
برنج. 1.31. ساختار فرم ترشحی IgM
به جذب کمپلکس های آنتی ژن-آنتی بادی و تخریب آنها در فاگوزوم های سلولی. IgG تنها کلاس آنتی بادی هایی است که قادر به نفوذ به سد جفت و محافظت داخل رحمی از جنین در برابر عفونت ها هستند.
ایمونوگلوبولین های A
کلاس اصلی آنتی بادی های موجود در ترشحات (شیر، بزاق، ترشحات مجاری تنفسی و روده). IgA عمدتاً به شکل دایمر ترشح می شود، جایی که مونومرها از طریق یک زنجیره J اضافی به یکدیگر متصل می شوند (شکل 1.32).
IgA با سیستم کمپلمان و سلول های فاگوسیت کننده تعامل ندارد، اما با اتصال به میکروارگانیسم ها، آنتی بادی ها از اتصال آنها به سلول های اپیتلیال و نفوذ به بدن جلوگیری می کنند.
ایمونوگلوبولین E
ایمونوگلوبولینهای E توسط مونومرهایی نشان داده میشوند که در آنها زنجیرههای ε سنگین، مانند زنجیرههای μ ایمونوگلوبولینهای M، یک متغیر و چهار حوزه ثابت دارند. IgE پس از ترشح با آن متصل می شود
 برنج. 1.32. ساختار IgA
برنج. 1.32. ساختار IgA
مناطق C ترمینال با گیرنده های مربوطه در سطح ماست سل ها و بازوفیل ها. در نتیجه، آنها به گیرنده های آنتی ژن روی سطح این سلول ها تبدیل می شوند (شکل 1.33).
 برنج. 1.33. برهمکنش IgE با آنتی ژن در سطح ماست سل
برنج. 1.33. برهمکنش IgE با آنتی ژن در سطح ماست سل
پس از اتصال آنتی ژن به محل های اتصال آنتی ژن مربوطه IgE، سلول ها سیگنالی برای ترشح مواد فعال بیولوژیکی (هیستامین، سروتونین) دریافت می کنند که تا حد زیادی مسئول ایجاد واکنش التهابی و بروز واکنش های آلرژیک مانند آسم، کهیر، تب یونجه.
ایمونوگلوبولین D
ایمونوگلوبولین های D در مقادیر بسیار کمی در سرم یافت می شوند. زنجیره δ سنگین دارای یک متغیر و سه حوزه ثابت است. IgD ها به عنوان گیرنده های لنفوسیت های B عمل می کنند. تعامل آنتی ژن های خاص با گیرنده های روی سطح لنفوسیت های B (IgD) منجر به انتقال این سیگنال ها به سلول و فعال شدن مکانیسم هایی می شود که تکثیر یک کلون لنفوسیتی معین را تضمین می کند.
موضوع 1.7. خواص فیزیکی و شیمیایی پروتئین ها و روش های جداسازی آنها
1. پروتئین های منفرد از نظر خواص فیزیکی و شیمیایی متفاوت هستند:
شکل مولکول ها؛
وزن مولکولی؛
بار کل، که مقدار آن به نسبت گروه های آنیونی و کاتیونی اسیدهای آمینه بستگی دارد.
نسبت رادیکال های اسید آمینه قطبی و غیر قطبی روی سطح مولکول ها.
درجات مقاومت در برابر عوامل دناتوره کننده مختلف.
2. حلالیت پروتئین بستگی دارددر مورد خواص پروتئین های ذکر شده در بالا و همچنین در مورد ترکیب محیطی که پروتئین در آن حل می شود (مقادیر pH، ترکیب نمک، دما، وجود سایر مواد آلی که می توانند با پروتئین تعامل داشته باشند). میزان بار روی مولکول های پروتئین یکی از عوامل موثر بر حلالیت آنهاست. هنگامی که بار در نقطه ایزوالکتریک از بین می رود، پروتئین ها راحت تر جمع می شوند و رسوب می کنند. این امر به ویژه برای پروتئین های دناتوره شده، که در آن رادیکال های اسید آمینه آبگریز روی سطح ظاهر می شوند، معمول است.
در سطح یک مولکول پروتئین، رادیکال های اسید آمینه با بار مثبت و منفی وجود دارد. تعداد این گروه ها و در نتیجه بار کل پروتئین ها به pH محیط بستگی دارد. نسبت غلظت گروه های H+ - و OH -. در محیط اسیدیافزایش غلظت H+ منجر به سرکوب تفکیک گروه های کربوکسیل -COO - + H + > - COOH و کاهش بار منفی پروتئین ها می شود. در یک محیط قلیایی، اتصال OH اضافی - توسط پروتون های تشکیل شده در هنگام تفکیک گروه های آمینه -NH 3 + + OH - - NH 2 + H 2 O با تشکیل آب، منجر به کاهش بار مثبت پروتئین ها می شود. . مقدار pH که در آن یک پروتئین دارای بار خالص صفر است نامیده می شود نقطه ایزوالکتریک (IEP).در IET، تعداد گروه های دارای بار مثبت و منفی یکسان است، یعنی. پروتئین در حالت ایزوالکتریک است.
3. جداسازی پروتئین های فردیویژگی های ساختار و عملکرد بدن به مجموعه پروتئین های سنتز شده در آن بستگی دارد. مطالعه ساختار و خواص پروتئین ها بدون جداسازی آنها از سلول و خالص سازی آنها از سایر پروتئین ها و مولکول های آلی غیرممکن است. مراحل جداسازی و خالص سازی پروتئین های فردی:
تخریب سلولیبافت مورد مطالعه و به دست آوردن یک هموژن.
جداسازی هموژنه به کسریبا سانتریفیوژ کردن، به دست آوردن یک بخش هسته ای، میتوکندری، سیتوزولی یا سایر بخش های حاوی پروتئین مورد نظر.
دناتوراسیون حرارتی انتخابی- حرارت دادن کوتاه مدت محلول پروتئین، که طی آن می توان برخی از ناخالصی های پروتئین دناتوره شده را حذف کرد (اگر پروتئین نسبتاً در برابر حرارت پایدار باشد).
نمک زدنپروتئین های مختلف در غلظت های مختلف نمک در محلول رسوب می کنند. با افزایش تدریجی غلظت نمک، می توان تعدادی فراکسیون جداگانه با محتوای غالب پروتئین جدا شده در یکی از آنها به دست آورد. سولفات آمونیوم اغلب برای شکنش پروتئین استفاده می شود. پروتئین هایی با کمترین حلالیت در غلظت های کم نمک رسوب می کنند.
فیلتراسیون ژل- روشی برای غربال کردن مولکول ها از طریق گرانول های متورم سفادکس (زنجیره های پلی ساکارید سه بعدی دکستران دارای منافذ). سرعت عبور پروتئین ها از یک ستون پر از Sephadex به وزن مولکولی آنها بستگی دارد: هر چه جرم مولکول های پروتئین کمتر باشد، راحت تر به درون گرانول ها نفوذ می کنند و هر چه جرم بیشتر باشد، سریعتر از گرانول ها خارج می شوند ستون
اولتراسانتریفیوژ- روشی که شامل قرار دادن پروتئین ها در یک لوله سانتریفیوژ در روتور یک اولتراسانتریفیوژ است. هنگامی که روتور می چرخد، سرعت ته نشینی پروتئین ها متناسب با وزن مولکولی آنها است: بخش هایی از پروتئین های سنگین تر نزدیک به انتهای لوله آزمایش قرار دارند، آنهایی که سبک تر هستند - نزدیک به سطح.
الکتروفورز- روشی مبتنی بر تفاوت در سرعت حرکت پروتئین ها در میدان الکتریکی. این مقدار با بار پروتئین ها متناسب است. الکتروفورز پروتئین ها بر روی کاغذ (در این مورد، سرعت حرکت پروتئین تنها با بار آنها متناسب است) یا در ژل پلی آکریل آمید با اندازه منافذ مشخص (سرعت حرکت پروتئین متناسب با بار و وزن مولکولی آنها است) انجام می شود. .
کروماتوگرافی تبادل یونی- یک روش شکنش مبتنی بر اتصال گروه های یونیزه شده پروتئین ها با گروه های بار مخالف رزین های تبادل یونی (مواد پلیمری نامحلول). قدرت اتصال پروتئین به رزین متناسب با بار پروتئین است. پروتئین های جذب شده به پلیمر تبادل یونی می توانند با افزایش غلظت محلول های NaCl شسته شوند. هرچه بار پروتئین کمتر باشد، غلظت NaCl کمتری برای شستن پروتئین متصل به گروه های یونی رزین لازم است.
کروماتوگرافی میل ترکیبی- خاص ترین روش برای جداسازی تک تک پروتئین ها، لیگاند یک پروتئین به صورت کووالانسی به یک پلیمر بی اثر متصل می شود. هنگامی که یک محلول پروتئین از طریق یک ستون با یک پلیمر عبور داده می شود، به دلیل اتصال مکمل پروتئین به لیگاند، فقط پروتئین مخصوص یک لیگاند معین روی ستون جذب می شود.
دیالیز- روشی که برای حذف ترکیبات با وزن مولکولی کم از محلول پروتئین ایزوله استفاده می شود. این روش بر خلاف مواد با وزن مولکولی کم بر اساس ناتوانی پروتئین ها در عبور از یک غشای نیمه تراوا است. برای تصفیه پروتئین ها از ناخالصی های کم مولکولی، به عنوان مثال، نمک ها پس از نمک استفاده می شود.
تکالیف برای کارهای فوق برنامه
1. جدول را پر کنید. 1.4.
جدول 1.4. تجزیه و تحلیل مقایسه ای ساختار و عملکرد پروتئین های مرتبط - میوگلوبین و هموگلوبین
الف) ساختار مرکز فعال Mb و Hb را به خاطر بسپارید. رادیکال های اسید آمینه آبگریز چه نقشی در تشکیل مراکز فعال این پروتئین ها دارند؟ ساختار مرکز فعال Mb و Hb و مکانیسم های اتصال لیگاندها به آن را شرح دهید. باقیمانده های His F 8 و His E 7 چه نقشی در عملکرد مرکز فعال Mv iHv ایفا می کنند؟
ب) پروتئین الیگومری نزدیک به هموگلوبین در مقایسه با میوگلوبین مونومری چه خواص جدیدی دارد؟ نقش تغییرات مشارکتی در ترکیب پروتومرها در مولکول هموگلوبین، تأثیر غلظت CO 2 و پروتون بر تمایل هموگلوبین به اکسیژن و همچنین نقش 2،3-BPG در تنظیم آلوستریک عملکرد Hb را توضیح دهید. .
2. همراهان مولکولی را با توجه به رابطه بین ساختار و عملکرد آنها مشخص کنید.
3. چه پروتئین هایی به خانواده ها تقسیم می شوند؟ با استفاده از مثال خانواده ایمونوگلوبولین ها، ویژگی های ساختاری مشابه و عملکردهای مرتبط پروتئین های این خانواده را شناسایی کنید.
4. پروتئین های منفرد خالص اغلب برای اهداف بیوشیمیایی و دارویی مورد نیاز هستند. روش های مورد استفاده برای جداسازی و خالص سازی پروتئین ها بر اساس چه ویژگی های فیزیکوشیمیایی پروتئین ها است.
وظایف خود کنترلی
1. پاسخ های درست را انتخاب کنید.
وظایف هموگلوبین:
الف. انتقال O 2 از ریه ها به بافت ب. انتقال H + از بافت ها به ریه ها
ب- حفظ pH خون ثابت D. انتقال CO 2 از ریه ها به بافت ها
د. انتقال CO 2 از بافت ها به ریه ها
2. پاسخ های درست را انتخاب کنید. لیگاندα پروتومر Hb عبارت است از: A. Heme
ب. اکسیژن
B. CO G. 2,3-BPG
D. β-پروتومر
3. پاسخ های درست را انتخاب کنید.
هموگلوبین در مقابل میوگلوبین:
الف- ساختار چهارتایی دارد
ب- ساختار ثانویه فقط با مارپیچ α نشان داده می شود
ب- متعلق به پروتئین های پیچیده است
د. با یک لیگاند آلوستریک تعامل دارد
4. پاسخ های درست را انتخاب کنید.
تمایل Hb به O2 کاهش می یابد:
الف. وقتی یک مولکول O 2 اضافه می شود ب. وقتی یک مولکول O 2 حذف می شود
ب. هنگام تعامل با 2،3-BPG
د. هنگامی که به پروتومرهای H + D متصل می شود. هنگامی که غلظت 2،3-BPG کاهش می یابد
5. همخوانی داشتن.
انواع HB با موارد زیر مشخص می شود:
الف. به شکل دئوکسی، سنگدانه های فیبریلار تشکیل می دهد. ب. شامل دو زنجیره α و دو زنجیره δ است.
ب- شکل غالب هموگلوبین در گلبول های قرمز بالغ D. حاوی هم با Fe+3 در مرکز فعال است.
د- شامل دو زنجیره α و دو γ 1. HbA 2.
6. همخوانی داشتن.
لیگاندهای Hb:
الف- در مرکز آلوستریک به Hb متصل می شود
ب- میل ترکیبی بسیار بالایی با محل فعال Hb دارد
ب- با پیوستن، میل ترکیبی Hb را برای O 2 G افزایش می دهد. Fe+2 را به Fe+3 اکسید می کند.
د- با hisF8 پیوند کووالانسی تشکیل می دهد
7. پاسخ های درست را انتخاب کنید.
همراهان:
الف- پروتئین های موجود در تمام قسمت های سلول
ب. سنتز تحت استرس افزایش می یابد
ب. در هیدرولیز پروتئین های دناتوره شده شرکت کنید
د. مشارکت در حفظ ترکیب بومی پروتئین ها
د- اندامک هایی ایجاد کنید که در آن ترکیب پروتئین ها تشکیل شود
8. مطابقت دادن. ایمونوگلوبولین ها:
الف- فرم ترشحی پنتامری است.
ب- کلاس Ig که به سد جفت نفوذ می کند
ب. Ig - گیرنده ماست سل
د- کلاس اصلی Ig موجود در ترشحات سلول های اپیتلیال. D. گیرنده لنفوسیت B که فعال شدن آن تکثیر سلولی را تضمین می کند
9. پاسخ های درست را انتخاب کنید.
ایمونوگلوبولین E:
الف. تولید شده توسط ماکروفاژها ب. آنها دارای زنجیره ε سنگین هستند.
ب. در غشای لنفوسیت های T تعبیه شده است
د. به عنوان گیرنده های آنتی ژن غشایی روی ماست سل ها و بازوفیل ها عمل می کنند
د. مسئول واکنش های آلرژیک
10. پاسخ های درست را انتخاب کنید.
روش جداسازی پروتئین ها بر اساس تفاوت در وزن مولکولی آنها است:
الف. فیلتراسیون ژل
ب. اولتراسانتریفیوژ
ب. الکتروفورز ژل پلی آکریل آمید D. کروماتوگرافی تبادل یونی
د. کروماتوگرافی میل ترکیبی
11. پاسخ صحیح را انتخاب کنید.
روش جداسازی پروتئین ها بر اساس تفاوت در حلالیت آنها در آب است:
الف. فیلتراسیون ژل ب. نمک زدایی
ب. کروماتوگرافی تبادل یونی D. کروماتوگرافی میل ترکیبی
د. الکتروفورز در ژل پلی آکریل آمید
استانداردهای پاسخ به "وظایف خودکنترلی"
1. الف، ب، ج، د
2. الف، ب، ج، د
5. 1-B، 2-A، 3-G
6. 1-B، 2-B، 3-A
7. الف، ب، د، د
8. 1-G; 2-B، 3-B
اصطلاحات و مفاهیم اساسی
1. پروتئین های الیگومر، پروتومر، ساختار چهارتایی پروتئین ها
2. تغییرات مشارکتی در ساختار پروتومر
3. اثر بور
4. تنظیم آلوستریک توابع پروتئین، مرکز آلوستریک و عامل آلوستریک
5. چاپرون های مولکولی، پروتئین های شوک حرارتی
6. خانواده های پروتئینی (سرین پروتئازها، ایمونوگلوبولین ها)
7. رابطه IgM-، G-، E-، A-ساختار-تابع
8. بار کل پروتئین ها، نقطه ایزوالکتریک پروتئین ها
9. الکتروفورز
10. نمک زدن
11. فیلتراسیون ژل
12. کروماتوگرافی تبادل یونی
13. اولتراسانتریفیوژ
14. کروماتوگرافی میل ترکیبی
15. الکتروفورز پروتئین های پلاسمای خون
وظایف برای کار در کلاس
1. مقایسه وابستگی درجه اشباع هموگلوبین (Hb) و میوگلوبین (Mb) با اکسیژن بر روی فشار جزئی آن در بافت ها
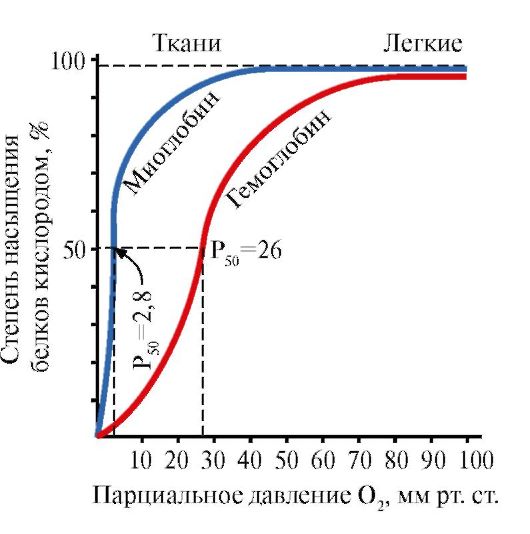 برنج. 1.34. وابستگی اشباع Mv وNHاکسیژن ناشی از فشار جزئی آن
برنج. 1.34. وابستگی اشباع Mv وNHاکسیژن ناشی از فشار جزئی آن
لطفا توجه داشته باشید که شکل منحنی های اشباع اکسیژن پروتئین متفاوت است: برای میوگلوبین - یک هذلولی، برای هموگلوبین - یک شکل سیگموئید.
1. مقادیر فشار جزئی اکسیژن را که در آن Mb و Hb 50٪ با O 2 اشباع شده است مقایسه کنید. کدام یک از این پروتئین ها تمایل بیشتری به O 2 دارد؟
2. چه ویژگی های ساختاری Mb میل ترکیبی بالای آن را برای O 2 تعیین می کند؟
3. چه ویژگی های ساختاری HB به آن اجازه می دهد O2 را در مویرگ های بافت های در حال استراحت (در فشار جزئی نسبتاً بالای O2) آزاد کند و این آزاد شدن را در عضلات در حال کار به شدت افزایش دهد؟ کدام ویژگی پروتئین های الیگومری این اثر را فراهم می کند؟
4. محاسبه کنید چه مقدار هموگلوبین اکسیژن دار O 2 (بر حسب درصد) به عضلات در حال استراحت و کار می دهد؟
5. در مورد رابطه بین ساختار پروتئین و عملکرد آن نتیجه گیری کنید.
2. میزان اکسیژن آزاد شده توسط هموگلوبین در مویرگ ها به شدت فرآیندهای کاتابولیک در بافت ها بستگی دارد (اثر بور). چگونه تغییرات در متابولیسم بافت میل هموگلوبین به O2 را تنظیم می کند؟ اثر CO 2 و H + بر میل ترکیبی Hb برای O 2
 1. اثر بور را توصیف کنید.
1. اثر بور را توصیف کنید.
2. فرآیند نشان داده شده در نمودار در چه جهتی پیش می رود:
الف) در مویرگ های ریه؛
ب) در مویرگ های بافتی؟
3. اهمیت فیزیولوژیکی اثر بور چیست؟
4. چرا برهمکنش Hb با H+ در مکان های دور از هم، میل پروتئین را برای O 2 تغییر می دهد؟
3. تمایل Hb به O2 به غلظت لیگاند آن بستگی دارد - 2،3-bisphosphoglycerate، که یک تنظیم کننده آلوستریک میل Hb برای O2 است. چرا برهمکنش لیگاند در محلی دور از محل فعال بر عملکرد پروتئین تأثیر می گذارد؟ چگونه 2،3-BPG میل هموگلوبین را برای O2 تنظیم می کند؟ برای حل مشکل به سوالات زیر پاسخ دهید:
1. 2.3-bisphosphoglycerate (2,3-BPG) از کجا و از چه چیزی سنتز می شود؟ فرمول آن را بنویسید، بار این مولکول را نشان دهید.
2. BPG با کدام شکل از هموگلوبین (اکسی یا دئوکسی) تعامل دارد و چرا؟ برهمکنش در کدام بخش از مولکول Hb رخ می دهد؟
3. فرآیند نشان داده شده در نمودار در چه جهتی پیش می رود؟
الف) در مویرگ های بافتی؛
ب) در مویرگ های ریه؟
4. جایی که غلظت کمپلکس باید بیشتر باشد
Nv-2،3-BFG:
الف) در مویرگ های عضلات در حالت استراحت،
ب) در مویرگ های ماهیچه های در حال کار (با وجود همان غلظت BPG در گلبول های قرمز)؟
5. در صورتی که غلظت BPG در گلبول های قرمز افزایش یابد، میل HB برای اکسیژن چگونه تغییر می کند زمانی که فرد با شرایط ارتفاع بالا سازگار شود؟ اهمیت فیزیولوژیکی این پدیده چیست؟
4. تخریب 2،3-BPG در طول ذخیره سازی خون حفظ شده، عملکرد HB را مختل می کند. اگر غلظت 2،3-BPG در گلبول های قرمز از 8 به 0.5 میلی مول در لیتر کاهش یابد، میل HB برای O 2 در خون حفظ شده چگونه تغییر می کند. اگر غلظت 2،3-BPG زودتر از سه روز بازیابی نشود، آیا می توان چنین خونی را به بیمارانی که به شدت بیمار هستند، تزریق کرد؟ آیا می توان عملکرد گلبول های قرمز را با افزودن 2،3-BPG به خون بازیابی کرد؟
5. ساختار ساده ترین مولکول های ایمونوگلوبولین را به خاطر بسپارید. ایمونوگلوبولین ها چه نقشی در سیستم ایمنی دارند؟ چرا Igs اغلب دو ظرفیتی نامیده می شود؟ ساختار Igs چگونه با عملکرد آنها ارتباط دارد؟ (با استفاده از نمونه ای از کلاس ایمونوگلوبولین ها توضیح دهید.)
خواص فیزیکوشیمیایی پروتئین ها و روش های جداسازی آنها
6. بار خالص پروتئین چگونه بر حلالیت آن تأثیر می گذارد؟
الف) بار کل پپتید را در pH 7 تعیین کنید
Ala-Glu-Tre-Pro-Asp-Liz-Cis
ب) بار این پپتید در pH> 7، pH چگونه تغییر می کند<7, рН <<7?
ج) نقطه ایزوالکتریک یک پروتئین (IEP) چیست و در چه محیطی قرار دارد؟
IET این پپتید؟
د) کمترین انحلال پذیری این پپتید در چه مقدار pH مشاهده خواهد شد.
7. چرا شیر ترش، بر خلاف شیر تازه، در زمان جوشاندن «منعک» می شود (یعنی پروتئین شیر کازئین رسوب می کند)؟ در شیر تازه، مولکول های کازئین دارای بار منفی هستند.
8. فیلتراسیون ژل برای جداسازی پروتئین های فردی استفاده می شود. مخلوطی حاوی پروتئین های A، B، C با وزن مولکولی به ترتیب برابر با 160000، 80000 و 60000 با فیلتراسیون ژل مورد تجزیه و تحلیل قرار گرفت (شکل 1.35). گرانول های ژل متورم به پروتئین هایی با وزن مولکولی کمتر از 70000 نفوذپذیر هستند. کدام نمودار نتایج حاصل از تقسیم بندی را به درستی منعکس می کند؟ ترتیب آزاد شدن پروتئین های A، B و C از ستون را مشخص کنید.
 برنج. 1.35. استفاده از فیلتراسیون ژل برای جداسازی پروتئین ها
برنج. 1.35. استفاده از فیلتراسیون ژل برای جداسازی پروتئین ها
9. در شکل 1.36، A نمودار الکتروفورز روی کاغذ پروتئین های سرم خون یک فرد سالم را نشان می دهد. مقادیر نسبی فراکسیون های پروتئینی به دست آمده با استفاده از این روش عبارتند از: آلبومین ها 54-58٪، α1-گلوبولین ها 6-7٪، α2-گلوبولین ها 8-9٪، β-گلوبولین ها 13٪، γ-گلوبولین ها 11-12٪.
 برنج. 1.36 الکتروفورز روی کاغذ پروتئین های پلاسمای خون یک فرد سالم (A) و یک بیمار (B)
برنج. 1.36 الکتروفورز روی کاغذ پروتئین های پلاسمای خون یک فرد سالم (A) و یک بیمار (B)
I - γ-گلوبولین ها؛ II - β-گلوبولین ها؛ III -α 2 -گلوبولین؛ IV -α 2 -گلوبولین؛ V - آلبومین ها
بسیاری از بیماری ها با تغییرات کمی در ترکیب پروتئین های سرم (دیسپروتئینمی) همراه هستند. ماهیت این تغییرات در هنگام تشخیص و ارزیابی شدت و مرحله بیماری در نظر گرفته می شود.
با استفاده از داده های ارائه شده در جدول 1.5، در مورد بیماری حدس بزنید، که با مشخصات الکتروفورتیک ارائه شده در شکل مشخص می شود. 1.36.
جدول 1.5. تغییرات در غلظت پروتئین های سرم در پاتولوژی
پروتئین ها مواد آلی هستند. این ترکیبات مولکولی بالا با ترکیب خاصی مشخص می شوند و پس از هیدرولیز به اسیدهای آمینه تجزیه می شوند. مولکولهای پروتئین میتوانند به اشکال مختلفی وجود داشته باشند که بسیاری از آنها از چندین زنجیره پلی پپتیدی تشکیل شدهاند. اطلاعات مربوط به ساختار یک پروتئین در DNA رمزگذاری شده است و فرآیند سنتز مولکول های پروتئین را ترجمه می نامند.
ترکیب شیمیایی پروتئین ها
پروتئین متوسط شامل:
- 52٪ کربن؛
- 7٪ هیدروژن؛
- 12٪ نیتروژن؛
- 21٪ اکسیژن؛
- 3 درصد گوگرد
مولکول های پروتئین پلیمر هستند. برای درک ساختار آنها، لازم است بدانیم مونومرهای آنها - اسیدهای آمینه - چیست.
آمینو اسید
آنها معمولاً به دو دسته تقسیم می شوند: دائماً و گاه به گاه رخ می دهند. اولی شامل 18 و 2 آمید دیگر است: آسپارتیک و اسید گلوتامیک. گاهی اوقات فقط سه اسید یافت می شود.
این اسیدها را می توان به روش های مختلفی طبقه بندی کرد: با توجه به ماهیت زنجیره های جانبی یا بار رادیکال های آنها، می توان آنها را بر تعداد گروه های CN و COOH نیز تقسیم کرد.
ساختار پروتئین اولیه
ترتیب تناوب اسیدهای آمینه در یک زنجیره پروتئینی سطوح بعدی سازمان، خواص و عملکرد آن را تعیین می کند. اصلی ترین مورد بین مونومرها پپتید است. از انتزاع هیدروژن از یک اسید آمینه و گروه OH از دیگری تشکیل می شود.
اولین سطح سازماندهی یک مولکول پروتئین، توالی اسیدهای آمینه در آن است، به سادگی زنجیره ای که ساختار مولکول های پروتئین را تعیین می کند. این شامل یک "اسکلت" است که ساختار منظمی دارد. این دنباله تکراری -NH-CH-CO- است. زنجیره های جانبی منفرد توسط رادیکال های اسید آمینه (R) نشان داده می شوند، خواص آنها ترکیب ساختار پروتئین را تعیین می کند.
حتی اگر ساختار مولکول های پروتئین یکسان باشد، تنها به این دلیل که مونومرهای آنها توالی متفاوتی در زنجیره دارند، می توانند از نظر خواص متفاوت باشند. ترتیب اسیدهای آمینه در یک پروتئین توسط ژن ها تعیین می شود و عملکردهای بیولوژیکی خاصی را به پروتئین دیکته می کند. توالی مونومرها در مولکول های مسئول عملکرد یکسان اغلب در گونه های مختلف مشابه است. چنین مولکول هایی از نظر سازماندهی یکسان یا مشابه هستند و عملکردهای یکسانی را در انواع مختلف موجودات - پروتئین های همولوگ انجام می دهند. ساختار، خواص و عملکرد مولکول های آینده در حال حاضر در مرحله سنتز زنجیره ای از اسیدهای آمینه ایجاد شده است.
برخی از ویژگی های مشترک
ساختار پروتئین ها برای مدت طولانی مورد مطالعه قرار گرفته است و تجزیه و تحلیل ساختار اولیه آنها امکان تعمیم برخی از آنها را فراهم کرده است. تعداد بیشتری از پروتئینها با حضور تمام بیست اسید آمینه مشخص میشوند که در میان آنها گلیسین، آلانین، گلوتامین و مقدار کمی تریپتوفان، آرژنین، متیونین و هیستیدین وجود دارد. تنها استثنا برخی از گروه های پروتئین، به عنوان مثال، هیستون ها هستند. آنها برای بسته بندی DNA مورد نیاز هستند و حاوی مقدار زیادی هیستیدین هستند.
هر نوع حرکت موجودات (عضله، حرکت پروتوپلاسم در سلول، سوسو زدن مژک ها در تک یاخته ها و غیره) توسط پروتئین ها انجام می شود. ساختار پروتئین ها به آنها اجازه می دهد تا حرکت کنند و رشته ها و حلقه ها را تشکیل دهند.

عملکرد انتقال این است که بسیاری از مواد در غشای سلولی توسط پروتئین های حامل خاص منتقل می شوند.
نقش هورمونی این پلیمرها بلافاصله مشخص می شود: تعدادی از هورمون ها در ساختار پروتئین هستند، به عنوان مثال انسولین، اکسی توسین.
عملکرد ذخیره با این واقعیت تعیین می شود که پروتئین ها قادر به تشکیل رسوب هستند. به عنوان مثال، والگومین تخم مرغ، کازئین شیر، پروتئین دانه های گیاهی - آنها مقدار زیادی مواد مغذی را ذخیره می کنند.
همه تاندونها، مفاصل مفصلی، استخوانهای اسکلتی و سمها توسط پروتئینها تشکیل میشوند که ما را به عملکرد بعدی آنها میرساند - پشتیبانی.
مولکول های پروتئین گیرنده هایی هستند که تشخیص انتخابی مواد خاصی را انجام می دهند. گلیکوپروتئین ها و لکتین ها به ویژه برای این نقش شناخته شده اند.
مهمترین عوامل ایمنی آنتی بادی ها و منشا پروتئینی هستند. به عنوان مثال، فرآیند لخته شدن خون بر اساس تغییرات در پروتئین فیبرینوژن است. دیواره های داخلی مری و معده با یک لایه محافظ از پروتئین های مخاطی - لیسین ها پوشیده شده اند. سموم نیز منشا پروتئینی دارند. اساس پوستی که از بدن حیوانات محافظت می کند کلاژن است. همه این عملکردهای پروتئینی محافظتی هستند.

خوب، آخرین عملکرد تنظیمی است. پروتئین هایی وجود دارند که عملکرد ژنوم را کنترل می کنند. یعنی رونویسی و ترجمه را تنظیم می کنند.
مهم نیست که نقش پروتئین ها چقدر مهم است، ساختار پروتئین ها مدت ها پیش توسط دانشمندان کشف شد. و اکنون آنها در حال کشف راه های جدیدی برای استفاده از این دانش هستند.
بلیط 2. 1. عوامل تغذیه ای ضروری طبیعت چربی.برخی از لیپیدها در بدن انسان سنتز نمی شوند و بنابراین از عوامل تغذیه ای ضروری هستند. اینها شامل اسیدهای چرب با دو یا چند پیوند دوگانه (پلین) است. اسیدهای چرب ضروری.برخی از این اسیدها بسترهایی برای سنتز هورمون های محلی - ایکوزانوئیدها هستند (موضوع 8.10).
ویتامین های محلول در چربیانجام عملکردهای مختلف: ویتامین Aدر روند بینایی و همچنین رشد و تمایز سلولی شرکت می کند. توانایی آن در مهار رشد انواع خاصی از تومورها ثابت شده است. ویتامین Kدر لخته شدن خون شرکت می کند؛ ویتامین دیدر تنظیم متابولیسم کلسیم شرکت می کند. ویتامین E- آنتی اکسیدان، از تشکیل رادیکال های آزاد جلوگیری می کند و در نتیجه با آسیب سلولی در نتیجه پراکسیداسیون لیپیدی مقابله می کند.
سند
2. ساختار و سطوح سازماندهی ساختاری پروتئین ها
چهار سطح از سازماندهی ساختاری پروتئین ها وجود دارد: اولیه، ثانویه، سوم و چهارم. هر سطح ویژگی های خاص خود را دارد.
ساختار پروتئین اولیه
ساختار اولیه پروتئین ها یک زنجیره پلی پپتیدی خطی از اسیدهای آمینه است که توسط پیوندهای پپتیدی به هم متصل شده اند. ساختار اولیه ساده ترین سطح سازماندهی ساختاری یک مولکول پروتئین است. پایداری بالایی توسط پیوندهای پپتیدی کووالانسی بین گروه α-آمینه یک اسید آمینه و گروه α-کربوکسیل یک اسید آمینه دیگر به آن داده می شود. [نمایش].
اگر گروه ایمنو پرولین یا هیدروکسی پرولین در تشکیل پیوند پپتیدی دخیل باشد، شکل متفاوتی دارد. [نمایش].
هنگامی که پیوندهای پپتیدی در سلول ها تشکیل می شود، ابتدا گروه کربوکسیل یک اسید آمینه فعال می شود و سپس با گروه آمینه دیگری ترکیب می شود. سنتز آزمایشگاهی پلی پپتیدها تقریباً به همین روش انجام می شود.
پیوند پپتیدی یک قطعه تکرار شونده از یک زنجیره پلی پپتیدی است. دارای تعدادی ویژگی است که نه تنها بر شکل ساختار اولیه، بلکه بر سطوح بالاتر سازماندهی زنجیره پلی پپتیدی نیز تأثیر می گذارد:

همسطحی - همه اتم های موجود در گروه پپتید در یک صفحه هستند.
توانایی وجود در دو شکل تشدید (کتو یا انول)؛
موقعیت ترانس جایگزین ها نسبت به پیوند C-N.
توانایی تشکیل پیوندهای هیدروژنی و هر یک از گروه های پپتیدی می توانند دو پیوند هیدروژنی با گروه های دیگر از جمله گروه های پپتیدی تشکیل دهند.
استثنا گروه های پپتیدی شامل گروه آمینه پرولین یا هیدروکسی پرولین است. آنها قادر به تشکیل تنها یک پیوند هیدروژنی هستند (به بالا مراجعه کنید). این بر تشکیل ساختار ثانویه پروتئین تأثیر می گذارد. زنجیره پلی پپتیدی در ناحیه ای که پرولین یا هیدروکسی پرولین در آن قرار دارد به راحتی خم می شود، زیرا طبق معمول توسط یک پیوند هیدروژنی دوم حفظ نمی شود.
نامگذاری پپتیدها و پلی پپتیدها. نام پپتیدها از نام اسیدهای آمینه تشکیل دهنده آنها ساخته شده است. دو اسید آمینه یک دی پپتید، سه اسید آمینه یک تری پپتید، چهار اسید یک تتراپپتید و غیره. هر پپتید یا زنجیره پلی پپتیدی با هر طولی دارای یک اسید آمینه N ترمینال حاوی یک گروه آمینه آزاد و یک اسید آمینه ترمینال C حاوی یک کربوکسیل آزاد است. گروه هنگام نامگذاری پلی پپتیدها، تمام آمینو اسیدها به ترتیب فهرست می شوند، که با N ترمینال شروع می شود و در نام آنها به جز C ترمینال، پسوند -in با -yl جایگزین می شود (زیرا اسیدهای آمینه در پپتیدها دیگر یک گروه کربوکسیل، اما یک کربونیل). به عنوان مثال، نام نشان داده شده در شکل. 1 تری پپتید - لوک سیلفنیل آلان سیلترئون که در.
ویژگی های ساختار اولیه پروتئین. در ستون فقرات زنجیره پلی پپتیدی، ساختارهای سفت و سخت (گروههای پپتیدی مسطح) با نواحی نسبتاً متحرک (-CHR)، که قادر به چرخش در اطراف پیوندها هستند، متناوب میشوند. چنین ویژگی های ساختاری زنجیره پلی پپتیدی بر آرایش فضایی آن تأثیر می گذارد.
ساختار ثانویه پروتئین
ساختار ثانویه راهی برای تا کردن یک زنجیره پلی پپتیدی به یک ساختار منظم به دلیل تشکیل پیوندهای هیدروژنی بین گروه های پپتیدی یک زنجیره یا زنجیره های پلی پپتیدی مجاور است. بر اساس پیکربندی، ساختارهای ثانویه به دو دسته مارپیچ (α-مارپیچ) و لایه چین خورده (ساختار β و فرم متقاطع β) تقسیم می شوند.


α-مارپیچ. این یک نوع ساختار پروتئین ثانویه است که شبیه یک مارپیچ منظم است که به دلیل پیوندهای هیدروژنی بین پپتیدی در یک زنجیره پلی پپتیدی ایجاد شده است. مدل ساختار مارپیچ α (شکل 2)، که تمام خواص پیوند پپتیدی را در نظر می گیرد، توسط پاولینگ و کوری پیشنهاد شد. ویژگی های اصلی مارپیچ α:
پیکربندی مارپیچ زنجیره پلی پپتیدی دارای تقارن مارپیچ.
تشکیل پیوندهای هیدروژنی بین گروه های پپتیدی هر اولین و چهارمین باقی مانده اسید آمینه.
منظم بودن چرخش های مارپیچی؛
هم ارزی تمام باقی مانده های اسید آمینه در مارپیچ α، صرف نظر از ساختار رادیکال های جانبی آنها.
رادیکال های جانبی اسیدهای آمینه در تشکیل مارپیچ α شرکت نمی کنند.
از نظر خارجی، مارپیچ α شبیه یک مارپیچ کمی کشیده از یک اجاق گاز الکتریکی است. منظم بودن پیوندهای هیدروژنی بین گروه های پپتیدی اول و چهارم، منظم بودن چرخش های زنجیره پلی پپتیدی را تعیین می کند. ارتفاع یک پیچ یا گام مارپیچ α 0.54 نانومتر است. این شامل 3.6 باقی مانده اسید آمینه است، به عنوان مثال، هر باقی مانده اسید آمینه در امتداد محور (ارتفاع یک باقی مانده اسید آمینه) به طول 0.15 نانومتر (0.54:3.6 = 0.15 نانومتر) حرکت می کند، که به ما امکان می دهد در مورد هم ارزی تمام باقی مانده های اسید آمینه صحبت کنیم. در مارپیچ α دوره منظم یک مارپیچ α 5 چرخش یا 18 باقیمانده اسید آمینه است. طول یک دوره 2.7 نانومتر است. برنج. 3. مدل a-helix Pauling-Corey
ب-ساختار. این یک نوع ساختار ثانویه است که دارای یک پیکربندی کمی خمیده از زنجیره پلی پپتیدی است و توسط پیوندهای هیدروژنی بین پپتیدی در بخش های جداگانه یک زنجیره پلی پپتیدی یا زنجیره های پلی پپتیدی مجاور تشکیل می شود. به آن ساختار تاشوی لایه ای نیز می گویند. انواعی از ساختارهای β وجود دارد. نواحی لایه ای محدودی که توسط یک زنجیره پلی پپتیدی از یک پروتئین تشکیل شده اند، فرم متقاطع β (ساختار β کوتاه) نامیده می شوند. پیوندهای هیدروژنی به شکل متقاطع β بین گروه های پپتیدی حلقه های زنجیره پلی پپتیدی تشکیل می شود. نوع دیگر - ساختار β کامل - مشخصه کل زنجیره پلی پپتیدی است که شکلی کشیده دارد و توسط پیوندهای هیدروژنی بین پپتیدی بین زنجیره های پلی پپتیدی موازی مجاور نگه داشته می شود (شکل 3). این سازه شبیه دم یک آکاردئون است. علاوه بر این، انواع ساختارهای β امکان پذیر است: آنها می توانند توسط زنجیره های موازی تشکیل شوند (انتهای پایانه N زنجیره های پلی پپتیدی در یک جهت هدایت می شوند) و ضد موازی (انتهای ترمینال N در جهات مختلف هدایت می شوند). رادیکال های جانبی یک لایه بین رادیکال های جانبی یک لایه دیگر قرار می گیرند.
در پروتئین ها، به دلیل بازآرایی پیوندهای هیدروژنی، انتقال از ساختار α به ساختار β و پشت امکان پذیر است. به جای پیوندهای هیدروژنی بین پپتیدی منظم در طول زنجیره (که به لطف آن زنجیره پلی پپتیدی به صورت مارپیچی پیچ خورده است)، بخش های مارپیچ باز می شوند و پیوندهای هیدروژنی بین قطعات دراز زنجیره های پلی پپتیدی بسته می شوند. این انتقال در کراتین، پروتئین مو یافت می شود. هنگام شستن مو با مواد شوینده قلیایی، ساختار مارپیچ بتا کراتین به راحتی از بین می رود و به α-کراتین تبدیل می شود (موهای مجعد صاف می شود).
تخریب ساختارهای ثانویه منظم پروتئین ها (آ-مارپیچ ها و ساختارهای β) بر اساس قیاس با ذوب یک کریستال، "ذوب" پلی پپتیدها نامیده می شود. در این حالت پیوندهای هیدروژنی شکسته می شود و زنجیره های پلی پپتیدی به شکل یک درهم تنیده تصادفی به خود می گیرند. در نتیجه، پایداری ساختارهای ثانویه توسط پیوندهای هیدروژنی بین پپتیدی تعیین می شود. سایر انواع پیوندها تقریباً هیچ نقشی در این امر ندارند، به استثنای پیوندهای دی سولفیدی در امتداد زنجیره پلی پپتیدی در محلهای باقیمانده سیستئین. پپتیدهای کوتاه به دلیل پیوندهای دی سولفید به چرخه بسته می شوند. بسیاری از پروتئین ها دارای هر دو ناحیه مارپیچ α و ساختار β هستند. تقریباً هیچ پروتئین طبیعی متشکل از 100٪ α-مارپیچ وجود ندارد (به استثنای پارامیوزین، پروتئین ماهیچه ای که 96-100٪ آلفا-مارپیچ است)، در حالی که پلی پپتیدهای مصنوعی دارای 100٪ مارپیچ هستند.
سایر پروتئین ها درجات مختلفی از پیچ خوردگی دارند. فرکانس بالایی از ساختارهای α-مارپیچ در پارامیوزین، میوگلوبین و هموگلوبین مشاهده می شود. در مقابل، در تریپسین، یک ریبونوکلئاز، بخش قابل توجهی از زنجیره پلی پپتیدی به ساختارهای بتا لایه ای تا می شود. پروتئین های بافت های حمایت کننده: کراتین (پروتئین مو، پشم)، کلاژن (پروتئین تاندون ها، پوست)، فیبروئین (پروتئین ابریشم طبیعی) دارای پیکربندی بتا از زنجیره های پلی پپتیدی هستند. درجات مختلف مارپیچ بودن زنجیره های پلی پپتیدی پروتئین ها نشان می دهد که بدیهی است که نیروهایی وجود دارند که تا حدی مارپیچ را مختل می کنند یا چین خوردگی منظم زنجیره پلی پپتیدی را "شکست می دهند". دلیل این امر تاخوردگی فشرده تر زنجیره پلی پپتیدی پروتئین در یک حجم معین، به عنوان مثال، به یک ساختار سوم است.
ساختار سوم پروتئین
ساختار سوم یک پروتئین روشی است که زنجیره پلی پپتیدی در فضا مرتب می شود. بر اساس شکل ساختار سوم، پروتئین ها عمدتا به کروی و فیبریلار تقسیم می شوند. پروتئین های کروی اغلب شکل بیضی دارند و پروتئین های فیبریلار (نخ مانند) شکل کشیده (میله ای یا دوکی شکل) دارند.
با این حال، پیکربندی ساختار سوم پروتئینها هنوز دلیلی برای این فکر نمیکند که پروتئینهای فیبریلار فقط ساختار β و پروتئینهای کروی ساختار α-مارپیچ دارند. پروتئین های فیبریلاری وجود دارند که ساختار ثانویه چین خورده ای دارند و نه لایه ای. به عنوان مثال، α-کراتین و پارامیوزین (پروتئین ماهیچه پرکننده نرم تنان)، تروپومیوزین ها (پروتئین های ماهیچه های اسکلتی) متعلق به پروتئین های فیبریلار (دارای شکل میله ای) هستند و ساختار ثانویه آنها α-مارپیچ است. در مقابل، پروتئین های کروی ممکن است حاوی تعداد زیادی ساختار β باشند.
مارپیچ شدن یک زنجیره پلی پپتیدی خطی اندازه آن را تقریباً 4 برابر کاهش می دهد. و بسته بندی در ساختار سوم آن را ده ها برابر فشرده تر از زنجیره اصلی می کند.
پیوندهایی که ساختار سوم پروتئین را تثبیت می کنند. پیوندهای بین رادیکال های جانبی اسیدهای آمینه در تثبیت ساختار سوم نقش دارند. این اتصالات را می توان به موارد زیر تقسیم کرد:
قوی (کووالانسی) [نمایش].
ضعیف (قطبی و واندروالس) [نمایش].
پیوندهای متعدد بین رادیکال های جانبی اسیدهای آمینه، پیکربندی فضایی مولکول پروتئین را تعیین می کند.
ویژگی های سازماندهی ساختار سوم پروتئین. انطباق ساختار سوم زنجیره پلی پپتیدی توسط خواص رادیکال های جانبی اسیدهای آمینه موجود در آن (که تأثیر قابل توجهی در تشکیل ساختارهای اولیه و ثانویه ندارند) و ریزمحیط، به عنوان مثال، تعیین می شود. محیط. هنگامی که تا می شود، زنجیره پلی پپتیدی یک پروتئین تمایل دارد شکلی از نظر انرژی مطلوب به خود بگیرد که با حداقل انرژی آزاد مشخص می شود. بنابراین، گروههای R غیرقطبی، با "اجتناب" از آب، بخش داخلی ساختار سوم پروتئین را تشکیل میدهند، جایی که بخش اصلی باقی ماندههای آبگریز زنجیره پلی پپتیدی در آن قرار دارد. تقریباً هیچ مولکولی آب در مرکز گلبول پروتئین وجود ندارد. گروه های قطبی (آب دوست) R اسید آمینه خارج از این هسته آبگریز قرار دارند و توسط مولکول های آب احاطه شده اند. زنجیره پلی پپتیدی به طور پیچیده در فضای سه بعدی خم شده است. هنگامی که خم می شود، ترکیب مارپیچ ثانویه مختل می شود. این زنجیره در نقاط ضعیفی که پرولین یا هیدروکسی پرولین در آن قرار دارند، می شکند، زیرا این اسیدهای آمینه در زنجیره متحرک تر هستند و تنها یک پیوند هیدروژنی را با سایر گروه های پپتیدی تشکیل می دهند. محل خم دیگر گلیسین است که دارای یک گروه R کوچک (هیدروژن) است. بنابراین، گروههای R از سایر اسیدهای آمینه، وقتی روی هم قرار میگیرند، تمایل دارند فضای آزاد را در محل گلیسین اشغال کنند. تعدادی از اسیدهای آمینه - آلانین، لوسین، گلوتامات، هیستیدین - به حفظ ساختارهای مارپیچ پایدار در پروتئین کمک می کنند و مانند متیونین، والین، ایزولوسین، اسید آسپارتیک به تشکیل ساختارهای β کمک می کنند. در یک مولکول پروتئینی با پیکربندی سوم، مناطقی به شکل مارپیچ α (مارپیچ)، ساختارهای β (لایه ای) و یک سیم پیچ تصادفی وجود دارد. فقط تا زدن فضایی صحیح پروتئین باعث فعال شدن آن می شود. نقض آن منجر به تغییر در خواص پروتئین و از دست دادن فعالیت بیولوژیکی می شود.
ساختار پروتئین کواترنر
پروتئین های متشکل از یک زنجیره پلی پپتیدی فقط ساختار سوم دارند. اینها شامل میوگلوبین - یک پروتئین بافت عضلانی است که در اتصال اکسیژن، تعدادی آنزیم (لیزوزیم، پپسین، تریپسین و غیره) نقش دارد. با این حال، برخی از پروتئین ها از چندین زنجیره پلی پپتیدی ساخته می شوند که هر کدام دارای ساختار سوم هستند. برای چنین پروتئین هایی، مفهوم ساختار چهارتایی معرفی شده است، که سازماندهی چندین زنجیره پلی پپتیدی با ساختار سوم در یک مولکول پروتئین عملکردی واحد است. چنین پروتئینی با ساختار چهارتایی، الیگومر و زنجیره های پلی پپتیدی آن با ساختار سوم، پروتومرها یا زیر واحدها نامیده می شوند (شکل 4).

در سطح چهارتایی سازمان، پروتئین ها پیکربندی اولیه ساختار سوم (کروبی یا فیبریلار) را حفظ می کنند. به عنوان مثال، هموگلوبین پروتئینی با ساختار چهارتایی است و از چهار زیر واحد تشکیل شده است. هر یک از زیر واحدها یک پروتئین کروی هستند و به طور کلی هموگلوبین دارای پیکربندی کروی است. پروتئینهای مو و پشم - کراتینها که از نظر ساختار سوم به پروتئینهای فیبریلار مربوط میشوند، دارای ساختار فیبریلار و ساختار چهارتایی هستند.
تثبیت ساختار چهارتایی پروتئین. تمام پروتئین هایی که ساختار چهارتایی دارند به شکل ماکرومولکول های مجزا جدا می شوند که به زیر واحدها تجزیه نمی شوند. تماس بین سطوح زیر واحدها تنها به دلیل گروه های قطبی باقی مانده های اسید آمینه امکان پذیر است، زیرا در طول تشکیل ساختار سوم هر یک از زنجیره های پلی پپتیدی، رادیکال های جانبی اسیدهای آمینه غیر قطبی (که اکثریت را تشکیل می دهند. تمام اسیدهای آمینه پروتئین زا) در داخل زیر واحد پنهان هستند. پیوندهای متعدد یونی (نمک)، هیدروژنی و در برخی موارد دی سولفیدی بین گروه های قطبی آنها تشکیل می شود که زیرواحدها را به شکل یک مجتمع سازمان یافته محکم نگه می دارند. استفاده از موادی که پیوندهای هیدروژنی را می شکند یا موادی که پل های دی سولفیدی را کاهش می دهند، باعث تجزیه پروتومرها و تخریب ساختار چهارتایی پروتئین می شود. روی میز 1 داده های مربوط به پیوندهایی را که سطوح مختلف سازماندهی مولکول پروتئین را تثبیت می کنند، خلاصه می کند [نمایش].
ویژگی های سازماندهی ساختاری برخی از پروتئین های فیبریلار
سازماندهی ساختاری پروتئین های فیبریلار در مقایسه با پروتئین های کروی دارای تعدادی ویژگی است. این ویژگی ها را می توان در مثال کراتین، فیبروئین و کلاژن مشاهده کرد. کراتین ها در ترکیبات α و β وجود دارند. α-کراتین ها و فیبروئین دارای ساختار ثانویه چین دار هستند، با این حال، در کراتین زنجیره ها موازی هستند و در فیبروئین آنها ضد موازی هستند (شکل 3 را ببینید). علاوه بر این، کراتین حاوی پیوندهای دی سولفید بین زنجیره ای است، در حالی که فیبروئین آنها را ندارد. شکستن پیوندهای دی سولفیدی منجر به جدا شدن زنجیره های پلی پپتیدی در کراتین ها می شود. در مقابل، تشکیل حداکثر تعداد پیوندهای دی سولفیدی در کراتین ها از طریق قرار گرفتن در معرض عوامل اکسید کننده یک ساختار فضایی قوی ایجاد می کند. به طور کلی، در پروتئین های فیبریلار، بر خلاف پروتئین های کروی، گاهی اوقات تشخیص دقیق بین سطوح مختلف سازمان دشوار است. اگر بپذیریم (در مورد یک پروتئین کروی) که ساختار سوم باید با قرار دادن یک زنجیره پلی پپتیدی در فضا و ساختار چهارتایی توسط چندین زنجیره تشکیل شود، در پروتئین های فیبریلار چندین زنجیره پلی پپتیدی از قبل در طول تشکیل ساختار ثانویه درگیر هستند. . یک مثال معمولی از پروتئین فیبریلار، کلاژن است که یکی از فراوان ترین پروتئین ها در بدن انسان است (حدود 1/3 جرم کل پروتئین ها). در بافت هایی که دارای استحکام بالا و کشش کم هستند (استخوان ها، تاندون ها، پوست، دندان ها و غیره) یافت می شود. در کلاژن، یک سوم از باقی مانده اسیدهای آمینه گلیسین است و حدود یک چهارم یا کمی بیشتر پرولین یا هیدروکسی پرولین است.

زنجیره پلی پپتیدی جدا شده از کلاژن (ساختار اولیه) مانند یک خط شکسته به نظر می رسد. حاوی حدود 1000 اسید آمینه و وزن مولکولی آن در حدود 10 5 است (شکل 5، a، b). زنجیره پلی پپتیدی از یک اسید آمینه تکراری (سه گانه) با ترکیب زیر ساخته شده است: gly-A-B، که در آن A و B هر اسید آمینه ای غیر از گلیسین (اغلب پرولین و هیدروکسی پرولین) هستند. زنجیره های پلی پپتیدی کلاژن (یا زنجیره های α) در طول تشکیل ساختارهای ثانویه و سوم (شکل 5، c و d) نمی توانند α-مارپیچ های معمولی با تقارن مارپیچ تولید کنند. پرولین، هیدروکسی پرولین و گلیسین (اسیدهای آمینه ضد هلیك) با این امر تداخل دارند. بنابراین، سه زنجیره α، همانطور که بود، مارپیچ های پیچ خورده را تشکیل می دهند، مانند سه رشته که دور یک استوانه می پیچند. سه زنجیره α مارپیچ یک ساختار کلاژن تکرار شونده به نام تروپوکلاژن را تشکیل می دهند (شکل 5d). تروپوکلاژن در سازمان خود ساختار سوم کلاژن است. حلقه های مسطح پرولین و هیدروکسی پرولین که به طور منظم در امتداد زنجیره قرار می گیرند، به آن استحکام می بخشد، همانطور که پیوندهای زنجیره ای بین زنجیره های α تروپوکلاژن (به همین دلیل است که کلاژن در برابر کشش مقاوم است). تروپوکلاژن اساسا زیر واحدی از فیبرهای کلاژن است. قرار دادن زیرواحدهای تروپوکلاژن در ساختار چهارتایی کلاژن در مراحل انجام می شود (شکل 5e).
تثبیت ساختارهای کلاژن به دلیل پیوندهای بین زنجیره ای هیدروژن، یونی و واندروالس و تعداد کمی پیوند کووالانسی رخ می دهد.
زنجیره آلفا کلاژن ساختار شیمیایی متفاوتی دارد. انواع مختلفی از زنجیره α 1 (I، II، III، IV) و زنجیره α 2 وجود دارد. بسته به اینکه کدام زنجیره α 1 و α 2 در تشکیل مارپیچ سه رشته ای تروپوکلاژن دخیل هستند، چهار نوع کلاژن متمایز می شوند:
نوع اول - دو زنجیره α 1 (I) و یک زنجیره α2.
نوع دوم - سه زنجیره α 1 (II).
نوع سوم - سه زنجیره α 1 (III).
نوع چهارم - سه زنجیره α 1 (IV).
رایج ترین کلاژن نوع اول است: در بافت استخوان، پوست، تاندون ها یافت می شود. کلاژن نوع دوم در بافت غضروف و غیره یافت می شود. در یک نوع بافت انواع مختلفی از کلاژن وجود دارد.
تجمع منظم ساختارهای کلاژن، سفتی و بی اثر بودن آنها استحکام بالای الیاف کلاژن را تضمین می کند. پروتئین های کلاژن همچنین حاوی اجزای کربوهیدراتی هستند، یعنی مجتمع های پروتئین-کربوهیدرات هستند.
کلاژن یک پروتئین خارج سلولی است که توسط سلول های بافت همبند موجود در همه اندام ها تشکیل می شود. بنابراین، با آسیب به کلاژن (یا اختلال در شکل گیری آن)، چندین نقض عملکردهای حمایتی بافت همبند اندام ها رخ می دهد.
زنجیره پلی پپتیدی آلفا با ترکیب اسید آمینه والین-لوسین و زنجیره پلی پپتیدی بتا با ترکیب والین-هیستیدین-لوسین به پایان می رسد. زنجیره های پلی پپتیدی آلفا و بتا در مولکول هموگلوبین به صورت خطی چیده نشده اند، این ساختار اولیه است. به دلیل وجود نیروهای درون مولکولی، زنجیره های پلی پپتیدی به شکل یک مارپیچ آلفا-مارپیچ (ساختار ثانویه) معمولی پروتئین ها پیچ خورده است. مارپیچ آلفا-مارپیچ خود به صورت مکانی برای هر زنجیره پلی پپتیدی آلفا و بتا خم می شود و شبکه هایی به شکل تخم مرغی (ساختار سوم) تشکیل می دهد. بخشهای جداگانه مارپیچهای آلفا-مارپیچ زنجیرههای پلی پپتیدی با حروف لاتین از A تا H مشخص میشوند. هر چهار زنجیره پلیپپتیدی منحنی آلفا و بتا از نظر فضایی در یک رابطه مشخص قرار دارند - یک ساختار چهارتایی. آنها نه با پیوندهای شیمیایی واقعی، بلکه توسط نیروهای بین مولکولی به هم متصل می شوند.
معلوم شد که انسان دارای سه نوع اصلی هموگلوبین طبیعی است: جنینی - U، جنینی - F و بالغ - A. HbU (نامگذاری شده از حرف اولیه کلمه رحم) در جنین بین 7 تا 12 هفته زندگی رخ می دهد. سپس ناپدید می شود و هموگلوبین جنینی ظاهر می شود که بعد از ماه سوم هموگلوبین اصلی جنین است. به دنبال این، هموگلوبین معمولی بزرگسالان به تدریج ظاهر می شود که HbA نامیده می شود، پس از حرف اولیه کلمه انگلیسی "adult". مقدار هموگلوبین جنین به تدریج کاهش می یابد به طوری که در لحظه تولد 80 درصد هموگلوبین HbA و تنها 20 درصد آن HbF است. پس از تولد، هموگلوبین جنین کاهش می یابد و در 2-3 سال زندگی فقط 1-2٪ است. همین مقدار هموگلوبین جنین در بزرگسالان وجود دارد. مقدار HbF بیش از 2% برای بزرگسالان و کودکان بالای 3 سال پاتولوژیک در نظر گرفته می شود.
علاوه بر انواع طبیعی هموگلوبین، بیش از 50 نوع پاتولوژیک در حال حاضر شناخته شده است. آنها ابتدا با حروف لاتین نامگذاری شدند. حرف B در تعیین انواع هموگلوبین وجود ندارد، زیرا در ابتدا HbS نام داشت.
هموگلوبین (Hb)- یک کروموپروتئین موجود در گلبول های قرمز خون و در انتقال اکسیژن به بافت ها نقش دارد. هموگلوبین در بزرگسالان هموگلوبین A (Hb A) نامیده می شود. وزن مولکولی آن حدود 65000 دالتون است. مولکول Hb A ساختار چهارتایی دارد و شامل چهار زیر واحد - زنجیره های پلی پپتیدی (با نامگذاری α1، α2، β1 و β2 است که هر کدام با هم مرتبط هستند.
به یاد داشته باشید که هموگلوبین یک پروتئین آلوستریک است. این امر میل پروتئین را برای لیگاندها تغییر می دهد. ترکیبی با کمترین میل ترکیبی برای لیگاند تنش یا T-conformation نامیده می شود. ترکیبی که بیشترین میل ترکیبی را برای لیگاند دارد، ترکیب آرام یا R-conformation نامیده می شود.
ترکیبات R و T مولکول هموگلوبین در حالت تعادل دینامیکی هستند:
عوامل محیطی مختلف می توانند این تعادل را در یک جهت یا جهت دیگر تغییر دهند. تنظیم کننده های آلوستریک موثر بر میل هموگلوبین برای O2 عبارتند از: 1) اکسیژن. 2) غلظت H+ (pH متوسط); 3) دی اکسید کربن (CO2)؛ 4) 2،3-دی فسفوگلیسرات (DPG). اتصال یک مولکول اکسیژن به یکی از زیرواحدهای هموگلوبین باعث انتقال یک ترکیب تنش به یک ساختار آرام می شود و میل ترکیبی برای اکسیژن سایر زیر واحدهای مولکول هموگلوبین مشابه را افزایش می دهد. این پدیده اثر تعاونی نامیده می شود. ماهیت پیچیده اتصال هموگلوبین به اکسیژن توسط منحنی اشباع هموگلوبین O2 منعکس می شود که دارای شکل S است (شکل 3.1).

شکل 3.1.منحنی اشباع اکسیژن میوگلوبین (1) و هموگلوبین (2).
اشکال مولکولیهموگلوبین در ساختار زنجیره های پلی پپتیدی با یکدیگر متفاوت است. نمونه ای از این تنوع هموگلوبین که در شرایط فیزیولوژیکی وجود دارد می باشد هموگلوبین جنینی (HbF)، در مرحله جنینی رشد انسان در خون وجود دارد. برخلاف HbA، مولکول آن شامل 2 زنجیره α و 2 γ است (یعنی زنجیره های β با زنجیره γ جایگزین می شوند). چنین هموگلوبین تمایل بیشتری به اکسیژن دارد. این چیزی است که به جنین اجازه می دهد تا از طریق جفت اکسیژن را از خون مادر دریافت کند. بلافاصله پس از تولد، HbF در خون نوزاد با HbA جایگزین می شود.
نمونه ای از هموگلوبین غیرطبیعی یا پاتولوژیک، هموگلوبین S است که قبلاً ذکر شد (نگاه کنید به 2.4.) که در بیماران مبتلا به کم خونی داسی شکل یافت می شود. همانطور که می دانید، با جایگزینی گلوتامات با والین در زنجیره β، با هموگلوبین A متفاوت است. این جایگزینی اسید آمینه باعث کاهش حلالیت HbS در آب و کاهش میل ترکیبی آن برای O2 می شود.
ساختار شیمیایی پروتئین ها توسط اسیدهای آمینه آلفا که در یک زنجیره از طریق یک پیوند پپتیدی متصل هستند نشان داده می شود. در موجودات زنده، ترکیب توسط کد ژنتیکی تعیین می شود. در فرآیند سنتز در اکثر موارد از 20 اسید آمینه از نوع استاندارد استفاده می شود. ترکیبات متعدد آنها مولکول های پروتئینی را با خواص بسیار متنوعی تشکیل می دهند. باقی مانده های اسید آمینه اغلب در معرض تغییرات پس از ترجمه قرار می گیرند. آنها می توانند قبل از شروع پروتئین به انجام وظایف خود و در طول فعالیت خود در سلول ایجاد شوند. در موجودات زنده، چندین مولکول اغلب مجتمع های پیچیده ای را تشکیل می دهند. یک مثال ارتباط فتوسنتزی است.
هدف از اتصالات
پروتئین ها به دلیل اینکه بدن آنها نمی تواند تمام اسیدهای آمینه لازم را سنتز کند، جزء مهمی از تغذیه انسان و حیوان محسوب می شود. برخی از آنها باید با غذاهای پروتئینی همراه باشند. منابع اصلی ترکیبات گوشت، آجیل، شیر، ماهی و غلات هستند. پروتئین ها به میزان کمتری در سبزیجات، قارچ ها و انواع توت ها وجود دارند. در طی هضم از طریق آنزیم ها، پروتئین های مصرف شده به اسیدهای آمینه تجزیه می شوند. آنها قبلاً در بیوسنتز پروتئین های خود در بدن استفاده می شوند یا برای به دست آوردن انرژی بیشتر تجزیه می شوند.

مرجع تاریخی
توالی ساختار پروتئین انسولین ابتدا توسط فردریج سنگر تعیین شد. او برای کارهایش در سال 1958 جایزه نوبل را دریافت کرد. سنگر از روش توالی یابی استفاده کرد. با استفاده از پراش اشعه ایکس، ساختارهای سه بعدی میوگلوبین و هموگلوبین متعاقبا (در اواخر دهه 1950) به دست آمد. این کار توسط جان کندرو و مکس پروتز انجام شد.
ساختار مولکول پروتئین
این شامل پلیمرهای خطی است. آنها به نوبه خود از باقی مانده های اسید آمینه آلفا تشکیل شده اند که مونومر هستند. علاوه بر این، ساختار پروتئین ممکن است شامل اجزایی با ماهیت غیر اسید آمینه و بقایای اسید آمینه اصلاح شده باشد. هنگام تعیین اجزاء، از اختصارات 1 یا 3 حرفی استفاده می شود. ترکیبی که حاوی دو تا چند ده باقیمانده است اغلب به عنوان "پلی پپتید" نامیده می شود. در نتیجه برهمکنش گروه آلفا کربوکسیل یک اسید آمینه با گروه آلفا آمینه دیگری، پیوندهایی ظاهر می شود (در طول تشکیل ساختار پروتئین). بسته به اینکه کدام گروه از باقی مانده اسید آمینه آزاد باشد، انتهای C و N ترمینال ترکیب از هم متمایز می شود: -COOH یا -NH2. در فرآیند سنتز پروتئین روی ریبوزوم، اولین باقیمانده انتهایی معمولاً باقی مانده متیونین است. موارد بعدی به C-پایانه های قبلی متصل می شوند.

سطوح سازمان
آنها توسط Lindrem-Lang پیشنهاد شدند. علیرغم اینکه این تقسیم بندی تا حدودی منسوخ تلقی می شود، هنوز از آن استفاده می شود. پیشنهاد شد که چهار سطح سازمان اتصال را متمایز کند. ساختار اولیه یک مولکول پروتئین توسط کد ژنتیکی و ویژگی های ژن تعیین می شود. سطوح بالاتر با تشکیل در طول تاخوردگی پروتئین مشخص می شود. ساختار فضایی یک پروتئین به عنوان یک کل توسط زنجیره اسید آمینه تعیین می شود. با این وجود، کاملاً ناپایدار است. ممکن است تحت تأثیر عوامل خارجی قرار گیرد. در این رابطه، صحیح تر است که در مورد ترکیب ترکیبی که مطلوب ترین و از نظر انرژی ارجح است صحبت کنیم.
سطح 1
آن را با دنباله ای از بقایای اسید آمینه یک زنجیره پلی پپتیدی نشان می دهد. به عنوان یک قاعده، با استفاده از نمادهای یک یا سه حرفی توصیف می شود. ساختار اولیه پروتئین ها با ترکیبات پایدار باقی مانده اسیدهای آمینه مشخص می شود. آنها وظایف خاصی را انجام می دهند. چنین "نقوش محافظه کارانه" در طول تکامل گونه ها حفظ می شود. آنها اغلب می توانند برای پیش بینی مشکل یک پروتئین ناشناخته استفاده شوند. با ارزیابی درجه تشابه (همسانی) در زنجیره های اسید آمینه از موجودات مختلف، می توان فاصله تکاملی تشکیل شده بین گونه های تشکیل دهنده این موجودات را تعیین کرد. ساختار اولیه پروتئین ها با تعیین توالی یا مجموعه اصلی mRNA آن با استفاده از جدول کد ژنتیکی تعیین می شود.

سفارش محلی یک بخش زنجیره ای
این سطح بعدی سازمان است - ساختار ثانویه پروتئین ها. انواع مختلفی از آن وجود دارد. ترتیب محلی بخشی از یک زنجیره پلی پپتیدی توسط پیوندهای هیدروژنی تثبیت می شود. محبوب ترین انواع عبارتند از:

ساختار فضایی
ساختار سوم پروتئین ها شامل عناصر سطح قبلی است. آنها با انواع مختلف تعاملات تثبیت می شوند. پیوندهای آبگریز از اهمیت بالایی برخوردار هستند. تثبیت شامل موارد زیر است:
- برهمکنش های کووالانسی
- پیوندهای یونی بین گروه های جانبی اسید آمینه که بارهای مخالف دارند تشکیل شده است.
- فعل و انفعالات هیدروژن
- پیوندهای آبگریز در فرآیند برهمکنش با عناصر اطراف H 2 O ، پروتئین تا می شود به طوری که گروه های اسید آمینه غیر قطبی جانبی از محلول آبی جدا می شوند. گروه های آب دوست (قطبی) روی سطح مولکول ظاهر می شوند.
ساختار سوم پروتئین ها با روش های تشدید مغناطیسی (هسته ای)، انواع خاصی از میکروسکوپ و روش های دیگر تعیین می شود.
اصل تخمگذار
تحقیقات نشان داده است که شناسایی یک سطح دیگر بین سطوح 2 و 3 راحت است. به آن "معماری"، "موتیف تخمگذار" می گویند. با موقعیت نسبی اجزای ساختار ثانویه (رشته های بتا و مارپیچ های آلفا) در داخل مرزهای یک گلبول فشرده - حوزه پروتئین تعیین می شود. این می تواند به طور مستقل وجود داشته باشد یا در یک پروتئین بزرگتر همراه با سایر پروتئین های مشابه گنجانده شود. مشخص شده است که انگیزه های یک ظاهر طراحی کاملا محافظه کارانه است. آنها در پروتئین هایی یافت می شوند که نه روابط تکاملی و نه عملکردی دارند. تعریف معماری اساس طبقه بندی عقلانی (فیزیکی) است.

سازمان دامنه
با آرایش متقابل چندین زنجیره پلی پپتیدی در یک مجتمع پروتئینی، ساختار چهارتایی پروتئین ها تشکیل می شود. عناصر تشکیل دهنده آن به طور جداگانه روی ریبوزوم ها تشکیل می شوند. تنها پس از اتمام سنتز، این ساختار پروتئین شروع به شکل گیری می کند. این می تواند شامل زنجیره های پلی پپتیدی متفاوت و یکسان باشد. ساختار چهارتایی پروتئین ها به دلیل برهمکنش های مشابه در سطح قبلی تثبیت می شود. برخی از کمپلکس ها ممکن است شامل چندین ده پروتئین باشند.
ساختار پروتئین: وظایف حفاظتی
پلی پپتیدهای اسکلت سلولی که به نوعی به عنوان تقویت کننده عمل می کنند، شکل بسیاری از اندامک ها را می دهند و در تغییر آن شرکت می کنند. پروتئین های ساختاری از بدن محافظت می کنند. به عنوان مثال، کلاژن چنین پروتئینی است. این پایه را در ماده بین سلولی بافت های همبند تشکیل می دهد. کراتین همچنین عملکرد محافظتی دارد. اساس شاخ، پر، مو و سایر مشتقات اپیدرم را تشکیل می دهد. هنگامی که پروتئین ها به سموم متصل می شوند، در بسیاری از موارد سم زدایی رخ می دهد. به این ترتیب وظیفه حفاظت شیمیایی بدن انجام می شود. آنزیم های کبدی نقش مهمی در فرآیند خنثی سازی سموم در بدن انسان دارند. آنها قادر به تجزیه سموم یا تبدیل آنها به شکل محلول هستند. این امر انتقال سریعتر آنها را از بدن تسهیل می کند. پروتئین های موجود در خون و سایر مایعات بدن با تحریک پاسخ به حمله و آسیب پاتوژن، دفاع ایمنی را ایجاد می کنند. ایمونوگلوبولین ها (آنتی بادی ها و اجزای سیستم کمپلمان) قادر به خنثی کردن باکتری ها، پروتئین های خارجی و ویروس ها هستند.
مکانیسم تنظیمی
مولکول های پروتئین که نه به عنوان منبع انرژی و نه به عنوان ماده ساختمانی عمل می کنند، بسیاری از فرآیندهای درون سلولی را کنترل می کنند. بنابراین، به واسطه آنها، ترجمه، رونویسی، برش و فعالیت سایر پلی پپتیدها تنظیم می شود. مکانیسم تنظیمی مبتنی بر فعالیت آنزیمی است یا به دلیل اتصال خاص به مولکول های دیگر خود را نشان می دهد. به عنوان مثال، فاکتورهای رونویسی، پلی پپتیدهای فعال کننده و پروتئین های سرکوب کننده قادر به کنترل شدت رونویسی ژن هستند. در انجام این کار، آنها با توالی های تنظیم کننده ژن تعامل دارند. مهمترین نقش در کنترل سیر فرآیندهای درون سلولی به پروتئین فسفاتازها و پروتئین کینازها اختصاص دارد. این آنزیم ها با افزودن یا حذف گروه های فسفات از پروتئین های دیگر باعث تحریک یا مهار فعالیت آن ها می شوند.

وظیفه سیگنال
اغلب با عملکرد تنظیمی ترکیب می شود. این به دلیل این واقعیت است که بسیاری از پلی پپتیدهای درون سلولی و همچنین خارج سلولی می توانند سیگنال ها را منتقل کنند. فاکتورهای رشد، سیتوکین ها، هورمون ها و سایر ترکیبات این توانایی را دارند. استروئیدها از طریق خون منتقل می شوند. تعامل هورمون با گیرنده به عنوان سیگنالی عمل می کند که پاسخ سلول را تحریک می کند. استروئیدها محتوای ترکیبات موجود در خون و سلول ها، تولید مثل، رشد و سایر فرآیندها را کنترل می کنند. یک مثال انسولین است. سطح گلوکز را تنظیم می کند. تعامل سلول ها از طریق ترکیبات پروتئینی سیگنالی که از طریق ماده بین سلولی منتقل می شود انجام می شود.
حمل و نقل عناصر
پروتئین های محلول درگیر در حرکت مولکول های کوچک میل ترکیبی بالایی با سوبسترا دارند که در غلظت افزایش یافته وجود دارد. همچنین این قابلیت را دارند که در مناطقی که محتوای آن کم است به راحتی آن را آزاد کنند. به عنوان مثال پروتئین انتقال هموگلوبین است. اکسیژن را از ریه ها به بافت های دیگر منتقل می کند و از آنها دی اکسید کربن را منتقل می کند. برخی از پروتئین های غشایی نیز در انتقال مولکول های کوچک از طریق دیواره های سلولی نقش دارند و آنها را تغییر می دهند. لایه لیپیدی سیتوپلاسم ضد آب است. این مانع از انتشار مولکول های باردار یا قطبی می شود. اتصالات حمل و نقل غشایی معمولاً به حامل ها و کانال ها تقسیم می شوند.
اتصالات پشتیبان
این پروتئین ها به اصطلاح ذخایر را تشکیل می دهند. آنها به عنوان مثال در دانه های گیاهی و تخم های حیوانی تجمع می یابند. چنین پروتئین هایی به عنوان منبع ذخیره ماده و انرژی عمل می کنند. برخی از ترکیبات توسط بدن به عنوان یک مخزن اسید آمینه استفاده می شود. آنها به نوبه خود پیش سازهای مواد فعالی هستند که در تنظیم متابولیسم نقش دارند.
گیرنده های سلولی
چنین پروتئین هایی می توانند مستقیماً در سیتوپلاسم قرار گیرند یا در دیواره جاسازی شوند. یک قسمت از اتصال سیگنال را دریافت می کند. به عنوان یک قاعده، این یک ماده شیمیایی و در برخی موارد یک اثر مکانیکی (مثلاً کشش)، نور و سایر محرک ها است. در فرآیند قرار گرفتن در معرض یک سیگنال به قطعه خاصی از مولکول - گیرنده پلی پپتیدی - تغییرات ساختاری آن شروع می شود. آنها باعث ایجاد تغییر در ساختار بقیه قسمتی می شوند که محرک را به سایر اجزای سلول منتقل می کند. ارسال سیگنال می تواند به روش های مختلفی انجام شود. برخی از گیرنده ها قادر به کاتالیز کردن یک واکنش شیمیایی هستند، در حالی که برخی دیگر به عنوان کانال های یونی عمل می کنند که تحت تأثیر یک محرک بسته یا باز می شوند. برخی از ترکیبات به طور خاص مولکول های پیام رسان را در داخل سلول متصل می کنند.
پلی پپتیدهای موتوری
یک کلاس کامل از پروتئین ها وجود دارد که حرکت را برای بدن فراهم می کند. پروتئین های حرکتی در انقباض عضلات، حرکت سلولی و فعالیت تاژک ها و مژک ها نقش دارند. آنها همچنین حمل و نقل جهت دار و فعال را ارائه می دهند. کینزین ها و داینین ها با استفاده از هیدرولیز ATP به عنوان منبع انرژی، مولکول ها را در طول میکروتوبول ها منتقل می کنند. دومی اندامک ها و سایر عناصر را از نواحی سلولی محیطی به سمت سانتروزوم حرکت می دهد. کینزین ها در جهت مخالف حرکت می کنند. Dyneins همچنین مسئول فعالیت تاژک ها و مژک ها هستند.
