Essee
aiheesta: "Jäännös"
Johdanto 3
1. Lisääntymistyypit 4
1.1 Sukupuolinen lisääntyminen 4
1.2 Sukupuolinen lisääntyminen 6
2. Organismien yksilöllinen kehitys 10
2.1 Alkion kehityskausi 10
2.2 Postembryonaalinen kehityskausi 13
2.3 Yleiset kehitysmallit. Biogeneettinen laki 15
Johtopäätös 18
Viitteet 18
Johdanto
Lisääntymiskyky, ts. saman lajin yksilöiden uuden sukupolven tuottaminen on yksi elävien organismien pääominaisuuksista. Lisääntymisprosessin aikana geneettistä materiaalia siirretään vanhemmilta sukupolvelta seuraavalle sukupolvelle, mikä varmistaa paitsi tietyn lajin myös tiettyjen vanhemman yksilöiden ominaisuuksien lisääntymisen. Lajille lisääntymisen tarkoitus on korvata sen kuolevat edustajat, mikä varmistaa lajin olemassaolon jatkuvuuden; lisäksi lisääntyminen mahdollistaa sopivissa olosuhteissa lajien kokonaismäärän lisäämisen.
Jokaisen uuden yksilön on käytävä läpi useita kasvu- ja kehitysvaiheita, ennen kuin se saavuttaa sen vaiheen, jossa se pystyy lisääntymään. Jotkut yksilöt kuolevat ennen lisääntymisvaiheen (tai sukukypsyyden) saavuttamista petoeläinten aiheuttaman tuhon, sairauksien ja erilaisten satunnaisten tapahtumien seurauksena; siksi laji voi selviytyä vain sillä ehdolla, että jokainen sukupolvi tuottaa enemmän jälkeläisiä kuin oli lisääntymiseen osallistuneita vanhempia yksilöitä. Populaatioiden koot vaihtelevat yksilöiden lisääntymisen ja sukupuuttoon kuolemisen välisen tasapainon mukaan. On olemassa useita erilaisia leviämisstrategioita, joista jokaisella on selkeät edut ja haitat; ne kaikki kuvataan tässä tiivistelmässä.
1. Lisääntymistyypit
Useita lisääntymismuotoja tunnetaan, mutta ne kaikki voidaan yhdistää kahteen tyyppiin: seksuaaliseen ja aseksuaaliseen.
Sukupuolisella lisääntymisellä tarkoitetaan sukupolvien vaihtoa ja organismien kehittymistä sukurauhasissa muodostuneista erikoistuneista sukusoluista. Tässä tapauksessa uusi organismi kehittyy kahden eri vanhempien muodostaman sukusolun fuusion seurauksena. Selkärangattomilla eläimillä siittiöitä ja munasoluja muodostuu kuitenkin usein yhden organismin kehossa. Tätä ilmiötä, biseksuaalisuutta, kutsutaan hermafroditismiksi. Kukkivat kasvit ovat myös biseksuaaleja. Useimmissa koppisiemenisten (kukkivien) kasvien lajeissa biseksuaalinen kukka sisältää sekä heteitä, jotka muodostavat miessukusoluja (siemensoluja), että emiä, jotka sisältävät munia. Noin neljänneksellä lajeista uros- ja naaraskukat kehittyvät itsenäisesti, ts. niiden kukat ovat yksipuolisia. Esimerkki on hamppu. Joissakin kasveissa (maissi, koivu) sekä uros- että naaraskukat näkyvät samalla yksilöllä.
Jotkut eläin- ja kasvilajit kehittyvät
hedelmöittämätön muna. Tämän tyyppistä lisääntymistä kutsutaan neitseelliseksi tai partenogeneettiseksi.
Aseksuaaliselle lisääntymiselle on ominaista se, että uusi yksilö kehittyy ei-seksuaalisista, somaattisista (kehon) soluista.
1.1 Aseksuaalinen lisääntyminen
Aseksuaalisessa lisääntymisessä uusi organismi voi syntyä yhdestä solusta tai useista äidin aseksuaalisista (somaattisista) soluista. Aseksuaaliseen lisääntymiseen osallistuu vain yksi vanhempi. Koska tytärorganismeja synnyttävät solut syntyvät mitoosin seurauksena, kaikki jälkeläiset ovat perinnöllisiltä ominaisuuksiltaan samanlaisia kuin äidin yksilö.
Riisi. 1. Vihreän euglenan lisääntyminen
Monet alkueläimet (ameba, vihreä euglena jne.), yksisoluiset levät (Chlamydomonas) lisääntyvät mitoottisen solujakautumisen kautta (kuva 1). Muille yksisoluisille eräille alemmille sienille, leville (chlorella), eläimille, esimerkiksi malarian malariaplasmodiumin aiheuttajalle, on ominaista itiöityminen. Tässä tapauksessa solu hajoaa suureen määrään yksilöitä, joka on yhtä suuri kuin ytimien lukumäärä, joka on aiemmin muodostunut emosolussa sen ytimen toistuvan jakautumisen seurauksena. Monisoluiset organismit pystyvät myös itiöimään: näitä ovat sammalet, korkeammat sienet, monisoluiset levät, pteridofyytit ja jotkut muut.
Sekä yksisoluisissa että monisoluisissa organismeissa orastuminen on myös suvuttoman lisääntymisen menetelmä. Esimerkiksi hiivasienissä ja joissakin väreissä (imeissä väreissä) muodostuu emosolulle silmuttaessa aluksi pieni ytimen sisältävä tuberkkeli, silmu. Se kasvaa, saavuttaa koon, joka on lähellä äidin ruumiin kokoa, ja sitten eroaa siirtyen itsenäiseen olemassaoloon. Monisoluisissa organismeissa (makean veden hydra) munuainen koostuu joukosta soluja kehon seinämän molemmista kerroksista. Silmu kasvaa, pitenee ja sen etupäähän ilmestyy suuaukko, jota ympäröivät lonkerot. Orastus päättyy pienen hydran muodostumiseen, joka sitten eroaa emoorganismista.
Monisoluisissa eläimissä suvuton lisääntyminen tapahtuu samalla tavalla (meduusat, annelidit, litamadot, piikkinahkaiset). Jokaisesta tällaisesta osasta kehittyy täysivaltainen yksilö.
Vegetatiivinen lisääntyminen on yleistä kasveissa, ts. ruumiinosat pistokkaat, lonkerot, mukulat. Siten perunat lisääntyvät muunnetuilla varren maanalaisilla osilla - mukuloilla. Jasmiinin ja pajun versot ja pistokkaat juurtuvat helposti. Rypäleitä, herukoita ja karviaisia lisätään pistokkailla.
Mansikoiden pitkät hiipivät varret muodostavat silmuja, jotka juurtuessaan synnyttävät uuden kasvin. Harvoja kasveja, kuten begoniaa, voidaan lisätä lehtipistokkailla (lehti ja varsi). Lehden alapuolella, paikoissa, joissa suuret suonet haarautuvat, ilmestyy juuria, yläpuolella on silmuja ja sitten versoja.
Juuresta käytetään myös kasvullista lisäämistä. Puutarhanhoidossa vadelmia, kirsikoita, luumuja ja ruusuja lisätään käyttämällä sivujuurista peräisin olevia pistokkaita. Daaliat lisääntyvät juurimukuloilla. Myös varren juurakon maanalaisen osan muuttaminen muodostaa uusia kasveja. Esimerkiksi kylvä ohdake juurakoiden avulla voi tuottaa yli tuhat uutta yksilöä 1 m2 maaperää kohti.
1.2 Sukupuolinen lisääntyminen
Sukupuolisella lisääntymisellä on erittäin suuria evoluutionaalisia etuja verrattuna aseksuaaliseen lisääntymiseen. Tämä johtuu siitä, että jälkeläisten genotyyppi syntyy yhdistämällä molemmille vanhemmille kuuluvia geenejä. Tämän seurauksena organismien kyky sopeutua ympäristöolosuhteisiin paranee. Koska jokaisessa sukupolvessa tehdään uusia yhdistelmiä, paljon suurempi määrä yksilöitä voi sopeutua uusiin olemassaolon olosuhteisiin kuin aseksuaalisen lisääntymisen aikana. Uusien geeniyhdistelmien syntyminen varmistaa lajin onnistuneemman ja nopeamman sopeutumisen muuttuviin ympäristöolosuhteisiin.
Siten seksuaalisen lisääntymisen ydin on kahdesta eri lähteestä - vanhemmista - peräisin olevan geneettisen tiedon jälkeläisen yhdistelmästä perinnöllisissä materiaaleissa.
Sukurauhasissa kehittyvät sukupuolisolut: miehen siittiöt, naisen munasolut (tai munat). Ensimmäisessä tapauksessa niiden kehitystä kutsutaan spermatogeneesiksi, toisessa - oogeneesiksi (latinasta ovo - muna).
Sukusolujen muodostumisprosessissa erotetaan useita vaiheita. Ensimmäinen vaihe on lisääntymisjakso, jolloin alkusukusolut jakautuvat mitoosin kautta, mikä johtaa niiden lukumäärän kasvuun.
Toinen vaihe on kasvun aika. Epäkypsissä urospuolisissa sukusoluissa sitä ei lausuta. Niiden koko kasvaa hieman. Päinvastoin, tulevaisuuden munasolujen koko kasvaa, joskus satoja ja useammin tuhansia ja jopa miljoonia kertoja. Munasolujen kasvu tapahtuu muiden kehon solujen muodostamien aineiden vuoksi. Siten kaloissa, sammakkoeläimissä ja suuremmassa määrin matelijoissa ja linnuissa suurin osa munasta on keltuaista. Se syntetisoituu maksassa, kuljetetaan erityisessä liukoisessa muodossa veren mukana munasarjoihin, tunkeutuu kasvaviin munasoluihin ja kerrostuu sinne keltuaislevyjen muodossa. Lisäksi itse tulevassa lisääntymissolussa syntetisoidaan lukuisia proteiineja ja suuri määrä erilaisia RNA:ita: kuljetus-, ribosomaalisia ja informaatiota. Keltuainen on kokoelma kehittyvän alkion ravitsemiseen välttämättömiä ravintoaineita (rasvat, proteiinit, hiilihydraatit, vitamiinit jne.), ja RNA varmistaa proteiinien synteesin varhaisessa kehitysvaiheessa, kun sen omaa tuhoisaa tietoa ei vielä hyödynnetä.
Seuraava vaihe, kypsymisjakso eli meioosi, on esitetty kuvassa 2. Kypsymisvaiheeseen tulevat solut sisältävät diploidisen kromosomijoukon ja jo kaksinkertaisen määrän DNA:ta.

Riisi. 2. Sukusolujen kypsyminen (meioosi)
Meioosin ydin on, että jokainen sukupuolisolu vastaanottaa yhden haploidisen joukon kromosomeja. Samanaikaisesti meioosi on kuitenkin vaihe, jossa syntyy uusia geeniyhdistelmiä yhdistämällä erilaisia äidin ja isän kromosomeja, perinnöllisten taipumusten rekombinaatiota tapahtuu myös risteytyksen seurauksena - homologisten kromosomien välisen osien vaihdon prosessin aikana. meioosista.
Meioosiin kuuluu kaksi peräkkäistä jakautumista. Kuten mitoosissa, jokaisessa meioottisessa jakautumisessa on neljä vaihetta: profaasi, metafaasi, anafaasi ja telofaasi.
Ensimmäinen (I) meioottinen jakautuminen. Profaasi I alkaa kromosomien spiralisoinnilla. Kuten muistat, jokainen kromosomi koostuu kahdesta kromatidista, jotka on yhdistetty sentromeeriin. Sitten homologiset kromosomit tulevat lähemmäksi toisiaan, yhden kromosomin kunkin kromatidin jokainen piste yhdistetään toisen, homologisen kromosomin kromatidin vastaavaan pisteeseen. Tätä homologisten kromosomien tarkkaa ja läheistä yhdistämisprosessia meioosissa kutsutaan konjugaatioksi. Tulevaisuudessa tällaisten kromosomien välillä voi tapahtua risteytys - identtisten tai homologisten eli samoja geenejä sisältävien alueiden vaihto. Profaasin loppua kohti homologisten kromosomien välille syntyy hylkiviä voimia. Ensin ne näkyvät sentromeerialueella ja sitten muilla alueilla.
Metafaasissa I kromosomien spiralisoituminen on suurinta. Konjugoidut kromosomit sijaitsevat päiväntasaajaa pitkin, jolloin homologisten kromosomien sentromeerit ovat päin solun eri napoja. Karan kierteet on kiinnitetty niihin.
Anafaasissa I homologisten kromosomien käsivarret lopulta erottuvat ja kromosomit siirtyvät eri napoihin. Näin ollen jokaisesta homologisesta kromosomiparista vain yksi pääsee tytärsoluun. Kromosomien määrä vähenee puoleen, kromosomijoukosta tulee haploidi. Jokainen kromosomi koostuu kuitenkin kahdesta kromatidista, eli se sisältää silti kaksinkertaisen määrän DNA:ta.
Telofaasissa I ydinvaippa muodostuu lyhyeksi ajaksi. Meioosin ensimmäisen ja toisen jakautumisen välisen vaiheen aikana DNA:n replikaatiota ei tapahdu. Kypsymisen ensimmäisen jakautumisen seurauksena muodostuneet solut eroavat isän ja äidin kromosomien koostumuksesta ja siten geenijoukosta.
Esimerkiksi kaikki ihmisen solut, mukaan lukien alkuperäiset sukusolut, sisältävät 46 kromosomia. Näistä 23 oli saatu isältä ja 23 äidiltä. Kun sukusolut muodostuvat ensimmäisen meioottisen jakautumisen jälkeen, spermatosyytit ja munasolut saavat myös 23 kromosomia. Kuitenkin johtuen isän ja äidin kromosomien eroavaisuuksien satunnaisuudesta anafaasissa I, tuloksena olevat solut saavat laajan valikoiman vanhempien kromosomien yhdistelmiä. Esimerkiksi yhdellä niistä voi olla 3 isän ja 20 äidin kromosomia, toisessa 10 isän ja 13 äidin kromosomia, kolmannessa 20 isän ja 3 äidin kromosomia jne. Mahdollisten yhdistelmien määrä on erittäin suuri. Jos otamme huomioon myös kromosomien homologisten osien vaihdon meioosin ensimmäisen jakautumisen profaasissa, on aivan ilmeistä, että jokainen tuloksena oleva sukusolu on geneettisesti ainutlaatuinen, koska sillä on oma ainutlaatuinen geenisarjansa.
Näin ollen meioosi on kombinatiivisen genotyyppisen vaihtelun perusta.
Toinen (II) meioottinen jakautuminen. Meioosin toinen jakautuminen etenee yleensä samalla tavalla kuin tavallinen mitoottinen jakautuminen, ainoana erona on, että jakautuva solu on haploidi. Anafaasissa II sentromeerit, jotka yhdistävät sisarkromatideja kussakin kromosomissa, jakautuvat, ja kromatideista, kuten mitoosissa, tulee tästä hetkestä itsenäisiä kromosomeja. Telofaasi II:n päätyttyä koko meioosiprosessi päättyy: alkuperäisestä primaarisesta sukusolusta muodostuu neljä haploidista solua.
Miehillä ne kaikki muunnetaan sukusoluiksi - siittiöiksi. Naarailla epätasaisen meioosin vuoksi vain yksi solu tuottaa elinkelpoisen munan. Muut kolme tytärsolua ovat paljon pienempiä, ne muuttuvat niin sanotuiksi ohjaaviksi eli vähentäviksi kappaleiksi, jotka pian kuolevat. Biologiselta kannalta katsottuna vain yhden munan muodostuminen ja kolmen geneettisesti täydellisen ohjauselimen kuolema johtuu tarpeesta säilyttää yhdessä solussa kaikki tulevan alkion kehittymiseen tarvittavat vararavinteet.
Muodostumisjakso koostuu soluista, jotka saavat tietyn muodon ja koon, joka vastaa niiden toimintaa.
Kypsymisprosessin aikana naisen sukusolut peittyvät kalvoilla ja ovat valmiita hedelmöittymiseen heti meioosin päätyttyä. Monissa tapauksissa, esimerkiksi matelijoilla, linnuilla ja nisäkkäillä, munaa ympäröivien solujen toiminnan vuoksi sen ympärille ilmestyy joukko lisäkalvoja. Niiden tehtävänä on suojata munaa ja kehittyvää alkiota ulkoisilta haitallisilta vaikutuksilta. Siittiöt voivat olla erikokoisia ja -muotoisia.
Siittiöiden tehtävänä on toimittaa geneettistä tietoa munasoluun ja stimuloida sen kehitystä. Muodostunut siittiö sisältää mitokondrioita, Golgi-laitteistoa, joka erittää entsyymejä, jotka liuottavat munasolun kalvoa hedelmöityksen aikana eli siittiön ja munasolun fuusion aikana. Tuloksena olevaa diploidista solua kutsutaan tsygootiksi.
2. Organismien yksilöllinen kehitys
Yksilökehitys eli ontogeneesi viittaa yksilön koko elämänvaiheeseen siitä hetkestä, kun siittiö sulautuu munasoluun ja tsygootin muodostuu organismin kuolemaan asti. Ontogeneesi on jaettu kahteen jaksoon: 1) alkion tsygootin muodostumisesta syntymään tai munakalvoista poistumiseen asti; 2) postembryonaalinen poistumisesta munakalvoista tai organismin syntymästä kuolemaan.
Tiedettä, joka tutkii organismien yksilöllisen kehityksen malleja alkiovaiheessa, kutsutaan embryologiaksi (kreikan sanasta embryo embryo).
2.1 Alkion kehityskausi
Useimmissa monisoluisissa eläimissä, riippumatta niiden organisaation monimutkaisuudesta, alkion kehitysvaiheet, jotka alkio käy läpi, ovat samat. Alkiossa on kolme päävaihetta: pilkkoutuminen, gastrulaatio ja primaarinen organogeneesi.
Erota. Organismin kehitys alkaa yksisoluvaiheessa. Hedelmöitetty munasolu on solu ja samalla varhaisimmassa kehitysvaiheessa oleva organismi. Toistuvien jakautumisten seurauksena yksisoluinen organismi muuttuu monisoluiseksi. Hedelmöityksen aikana siittiön ja munasolun fuusioitumisen kautta ilmaantunut diploidiydin alkaa jakautua muutamassa minuutissa ja myös sytoplasma jakautuu sen mukana. Tuloksena olevien solujen koko pienenee jokaisen jakautumisen myötä, joten jakautumisprosessia kutsutaan pilkkomiseksi. Sirpaloitumisjakson aikana solumateriaalia kertyy jatkokehitystä varten. Sirpaloituminen päättyy monisoluisen alkion, blastulan, muodostumiseen. Blastulassa on nesteellä täytetty ontelo, niin sanottu primaarinen ruumiinontelo.
Tapauksissa, joissa munan sytoplasmassa on vähän keltuaista (kuten lansetissa) tai suhteellisen vähän (kuten sammakossa), fragmentoituminen on täydellinen, eli muna jakautuu kokonaan.
Muuten linnuissa esiintyy sirpaloitumisjaksoa. Keltuaiseton sytoplasma muodostaa vain 1 % kananmunan kokonaistilavuudesta; munan koko jäljellä oleva sytoplasma ja siten tsygootti on täytetty keltuaismassalla. Jos katsot tarkkaan kananmunaa, yhdessä sen napoista suoraan keltuaisessa näet pienen täplän - blastulan eli itulevyn, joka muodostuu ytimen sisältävän sytoplasman keltuaisen osan murskaamisen seurauksena. Tällaisissa tapauksissa murskaamista kutsutaan epätäydelliseksi. Epätäydellinen pirstoutuminen on ominaista myös joillekin kaloille ja matelijoille.
Kaikissa tapauksissa, sekä lansetissa että sammakkoeläimissä ja linnuissa sekä muissa eläimissä, solujen kokonaistilavuus blastula-vaiheessa ei ylitä tsygootin tilavuutta. Toisin sanoen tsygootin mitoottiseen jakautumiseen ei liity tuloksena olevien tytärsolujen kasvua äidin tilavuuteen, ja niiden koot peräkkäisten jakautumisten seurauksena pienenevät asteittain. Tämä mitoottisen solun jakautumisen piirre pilkkomisen aikana havaitaan hedelmöittyneiden munien kehittymisen aikana kaikissa eläimissä.
Myös eräät muut murskauksen piirteet ovat tyypillisiä eri eläinlajeille. Esimerkiksi kaikilla blastulan soluilla on diploidinen kromosomisarja, ne ovat rakenteeltaan identtisiä ja eroavat toisistaan pääasiassa niiden sisältämän keltuaisen määrän suhteen. Tällaisia soluja, joista puuttuu merkkejä erikoistumisesta tiettyjen toimintojen suorittamiseen, kutsutaan erikoistumattomiksi (tai erilaistumattomiksi) soluiksi. Toinen pilkkomisen piirre on blastomeerien erittäin lyhyt mitoottinen sykli verrattuna aikuisen organismin soluihin. Hyvin lyhyen välivaiheen aikana tapahtuu vain DNA-kaksoistumista.
Gastrulaatio. Blastula, joka koostuu yleensä suuresta määrästä blastomeereja (esimerkiksi lansetissa 3000 solusta), siirtyy kehitysprosessin aikana uuteen vaiheeseen, jota kutsutaan gastrulaksi (kreikkalaisesta mahavatsasta). Tässä vaiheessa alkio koostuu selvästi erotettavissa olevista solukerroksista - ns. itukerroksista: ulompi eli ektodermi (kreikan sanasta ectos - sijaitsee ulkopuolella) ja sisäinen eli endodermi (kreikan sanasta entos - sijaitsee sisällä) . Gastrulan muodostumiseen johtavia prosesseja kutsutaan gastrulaatioksi.
Lansetissa gastrulaatio tapahtuu tunkeutumalla blastulan yksi napoista sisäänpäin, toista kohti; muissa eläimissä joko blastulan seinämän delaminaatiolla tai kasvattamalla massiivinen vegetatiivinen napa umpeen pienillä soluilla. eläinten napa.
Monisoluisissa eläimissä, paitsi coelenteraatissa, rinnakkain gastrulaation kanssa tai, kuten lansetissa, sen jälkeen, kolmas itukerroksen mesodermi (keskellä sijaitsevasta kreikkalaisesta mesoksista), joka on joukko soluelementtejä, jotka sijaitsevat ekto- ja endodermi primaarisen ruumiinontelon blastoselessa. Mesodermin ilmaantumisen myötä alkiosta tulee kolmikerroksinen.
Siten gastrulaatioprosessin ydin on solumassojen liikkuminen. Alkion solut käytännössä jakautuvat eivätkä kasva. Tässä vaiheessa alkiosolujen geneettisen tiedon käyttö kuitenkin alkaa ja ensimmäiset erilaistumisen merkit näkyvät.
Erilaistuminen tai erilaistuminen on sen esiintymisprosessi ja rakenteellisten ja toiminnallisten erojen lisääntyminen yksittäisten solujen ja alkion osien välillä. Morfologisesta näkökulmasta erilaistuminen ilmaistaan useiden satojen tietyn rakenteen omaavien solutyyppien muodostumisena, jotka eroavat toisistaan. Erikoistumattomista blastulasoluista nousee vähitellen esiin ihon epiteelisoluja, suoliston epiteeliä, keuhkoja, hermo- ja lihassoluja jne. Biokemiallisesta näkökulmasta solujen erikoistuminen on kykyä syntetisoida tiettyjä proteiineja, jotka ovat ominaisia vain tietylle solutyypille. Lymfosyytit syntetisoivat suojaavia proteiineja vasta-aineita, lihassolut supistavat proteiinia myosiinia. Jokainen solutyyppi tuottaa omia proteiinejaan, jotka ovat ainutlaatuisia. Solujen biokemiallinen erikoistuminen varmistetaan geenien selektiivisellä, differentiaalisella aktiivisuudella, eli eri itukerrosten soluissa, tiettyjen elinten ja järjestelmien alkeet, eri geeniryhmät alkavat toimia.
Eri eläinlajeissa samat itukerrokset synnyttävät samat elimet ja kudokset. Tämä tarkoittaa, että ne ovat homologisia. Siten niveljalkaisten ulomman itukerroksen - ektodermin - soluista muodostuvat sondit, mukaan lukien kalat, sammakkoeläimet, matelijat, linnut ja nisäkkäät, iho ja niiden johdannaiset sekä hermosto ja aistielimet. Suurimman osan eläimistä itukerrosten homologia on yksi todiste eläinmaailman yhtenäisyydestä.
Organogeneesi. Gastrulaation päätyttyä alkio muodostaa aksiaalisten elinten kompleksin: hermoputken, notochordin ja suolistoputken. Lansetissa aksiaaliset elimet muodostuvat seuraavasti: alkion selkäpuolen ektodermi taipuu keskiviivaa pitkin muuttuen uraksi, ja sen oikealla ja vasemmalla puolella oleva ektodermi alkaa kasvaa sen reunoilla. Ura, hermoston alkuaine, vajoaa ektodermin alle ja sen reunat sulkeutuvat. Hermoputki muodostuu. Loput ektodermista on ihon epiteelin alkuosa.
Endodermin dorsaalinen osa, joka sijaitsee suoraan hermorudimentin alla, on erotettu muusta endodermista ja taittuu tiheäksi naruksi, jota kutsutaan notochordiksi. Endodermin jäljellä olevasta osasta kehittyy mesodermi ja suoliston epiteeli. Alkion solujen erilaistuminen edelleen johtaa lukuisten itukerrosten - elimien ja kudosten - johdannaisten syntymiseen. Itukerrokset muodostavien solujen erikoistumisprosessissa hermosto, aistielimet, ihoepiteeli ja hammaskiille muodostuvat ektodermista; endodermista suoliston epiteeli, ruoansulatusrauhaset maksa ja haima, kidusten ja keuhkojen epiteeli; mesodermilihaskudoksesta, sidekudoksesta, mukaan lukien löysä sidekudos, rusto ja luukudos, veri ja imusolmukkeet, samoin kuin verenkiertoelimistö, munuaiset, sukurauhaset.
2.2 Postembryonaalinen kehityskausi
Syntymähetkellä tai organismin vapautuessa munankuorista alkiokausi päättyy ja alkion jälkeinen kehityskausi alkaa. Postembryonaalinen kehitys voi olla suoraa tai siihen voi liittyä transformaatio (metamorfoosi).
Suoran kehityksen aikana (matelijoilla, linnuilla, nisäkkäillä) munankuorista tai emon ruumiista nousee pienikokoinen organismi, jonka kaikki aikuiselle eläimelle ominaiset pääelimet ovat jo muodostuneet. Postembryonaalinen kehitys rajoittuu tässä tapauksessa pääasiassa kasvuun ja murrosikään.
Kehityksen aikana muodonmuutoksen yhteydessä munasta nousee toukka, joka on yleensä rakenteeltaan yksinkertaisempi kuin täysikasvuinen eläin, ja jolla on erityisiä toukkaelimiä, joita aikuisessa tilassa ei ole. Toukka ruokkii, kasvaa ja ajan myötä toukkaelimet korvataan aikuisille yksilöille ominaisilla elimilla. Tämän seurauksena muodonmuutoksen aikana toukkaelimet tuhoutuvat ja aikuisille eläimille ominaisia elimiä ilmaantuu.
Tarkastellaanpa useita esimerkkejä epäsuorasta postembryonisesta kehityksestä. Askidian toukalla (hylä Chordata, subphylum Larval-Chordata) on kaikki sointujen tärkeimmät ominaisuudet: notochord, hermoputki ja kidusraot nielussa. Se ui vapaasti ja kiinnittyy sitten johonkin kiinteään pintaan merenpohjassa, jossa tapahtuu muodonmuutos: sen häntä, notochord ja lihakset katoavat, ja hermoputki hajoaa yksittäisiksi soluiksi, joista suurin osa on fagosytoosia. Toukkien hermostosta on jäljellä vain joukko soluja, jotka synnyttävät hermosolmukkeen. Kiintynyttä elämäntapaa johtavan aikuisen askidialaisen kehon rakenne ei muistuta lainkaan sointujen järjestämisen tavanomaisia piirteitä. Vain tieto ontogeneesin piirteistä mahdollistaa askidien systemaattisen sijainnin määrittämisen: toukkien rakenne osoittaa niiden alkuperän vapaata elämäntapaa johtaneista chordaateista. Metamorfoosin aikana askidiat siirtyvät istuvaan elämäntapaan, ja siksi heidän organisaationsa yksinkertaistuu.
Sammakkoeläinten toukkamuoto on nuijapolka, jolle on ominaista kidusraot, sivuviiva, kaksikammioinen sydän ja yksi verenkierron ympyrä. Kilpirauhashormonin vaikutuksen alaisena tapahtuvan metamorfoosin aikana häntä rauhoittuu, raajat ilmestyvät, sivuviiva katoaa, keuhkot ja toinen verenkierron ympyrä kehittyvät. Huomionarvoista on useiden nuijapäiden ja kalojen rakenteellisten piirteiden samankaltaisuus (sivuviiva, sydämen ja verenkiertoelimen rakenne, kidusraot).
Hyönteisten kehitys voi toimia myös esimerkkinä muodonmuutoksesta. Perhosen toukat tai sudenkorennon toukat eroavat rakenteeltaan, elämäntavultaan ja elinympäristöltään jyrkästi aikuisista eläimistä ja muistuttavat esi-isiään, annelideja.
Postembryonisella kehitysjaksolla on erilaisia kestoja. Esimerkiksi toukkaperhoset elävät 2-3 vuotta toukkatilassa ja 2-3 tunnista 2-3 päivään kypsässä tilassa lajista riippuen. Useimmissa tapauksissa postembryonaalinen ajanjakso on pidempi. Ihmisillä se sisältää murrosiän, kypsyysvaiheen ja vanhuusvaiheen.
Nisäkkäillä ja ihmisillä tiedetään olevan elinajanodote riippuvainen murrosiän ja raskauden kestosta. Elinajanodote yleensä ylittää
lisääntymistä edeltävä ontogeneesijakso 5-8 kertaa.
Postembryoniseen kehitykseen liittyy kasvua. Erotetaan jatkuva kasvu, joka jatkuu läpi elämän, ja määrätty kasvu, joka on rajoitettu tiettyyn ajanjaksoon. Epämääräistä kasvua havaitaan puumaisilla kasveilla, joillakin nilviäisillä, selkärankaisilla, kaloilla ja rotilla.
Monilla eläimillä kasvu pysähtyy pian sukukypsyyden saavuttamisen jälkeen. Ihmisellä kasvu päättyy 20-25 vuoden iässä.
2.3 Yleiset kehitysmallit. Biogeneettinen laki
Kaikki monisoluiset organismit kehittyvät hedelmöitetystä munasolusta. Alkioiden kehitys samaan tyyppiin kuuluvissa eläimissä on pitkälti samanlaista. Kaikissa chordaateissa alkiokaudella muodostuu aksiaalinen luuston notochord, hermoputki ilmestyy ja nielun etuosaan muodostuu kidusrakoja. Myös sointujen rakennesuunnitelma on sama. Kehityksen alkuvaiheessa selkärankaisten alkiot ovat hyvin samankaltaisia (kuva 3). Nämä tosiasiat vahvistavat K. Baerin muotoileman alkion samankaltaisuuden lain pätevyyden: "Alkioilla on jo varhaisimmista vaiheista lähtien tietty yleinen samankaltaisuus tyypin sisällä." Eri systemaattisten ryhmien alkioiden samankaltaisuus osoittaa niiden yhteisen alkuperän. Tämän jälkeen alkioiden rakenne paljastaa luokan, suvun, lajin ominaisuudet ja lopuksi tietylle yksilölle ominaiset ominaisuudet. Alkioiden ominaisuuksien poikkeamista kehityksen aikana kutsutaan alkion poikkeavuudeksi, ja se heijastaa tietyn systemaattisen eläinryhmän kehitystä, tietyn lajin kehityshistoriaa.

Riisi. 3. Sukulinjan samankaltaisuus selkärankaisilla: 1 monotreme (echidna), 2 pussieläintä (kengurut), 3 artiodaktyyliä (peura), 4 lihansyöjää (kissa), 5 kädellistä (apina), 6 - ihmiset
Suuri samankaltaisuus alkioiden välillä varhaisessa kehitysvaiheessa ja
myöhempien vaiheiden erojen ilmiöllä on oma selityksensä.
Keho on vaihteleva koko kehityksen ajan.
Mutaatioprosessi vaikuttaa geeneihin, jotka määrittävät nuorimpien alkioiden rakenteelliset ja metaboliset ominaisuudet. Mutta niissä syntyvillä rakenteilla (kaukaisille esivanhemmille ominaiset muinaiset ominaisuudet) on erittäin tärkeä rooli jatkokehitysprosesseissa. Kuten on osoitettu, notochord primordium indusoi hermoputken muodostumista, ja sen menetys johtaa kehityksen pysähtymiseen. Siksi muutokset alkuvaiheessa johtavat yleensä yksilön alikehittymiseen ja kuolemaan. Päinvastoin, myöhemmissä vaiheissa tapahtuvat muutokset, jotka vaikuttavat vähemmän merkittäviin ominaisuuksiin, voivat olla hyödyllisiä eliölle, ja sellaisissa tapauksissa ne poimivat luonnollisen valinnan.
Nykyaikaisten eläinten kaukaisille esivanhemmille ominaisten ominaisuuksien esiintyminen alkion kehitysvaiheessa heijastaa elinten rakenteen evoluutiomuutoksia.
Kehitysessään organismi kulkee yksisoluisen vaiheen (tsygoottivaiheen) läpi, jota voidaan pitää primitiivisen ameeban fylogeneettisen vaiheen toistona. Kaikissa selkärankaisissa, mukaan lukien niiden korkeimmat edustajat, muodostuu notokordi, joka sitten korvataan selkärangalla, ja heidän esivanhemmissaan lansetin perusteella päätellen notochord säilyi koko elämän. Lintujen ja nisäkkäiden, mukaan lukien ihmiset, alkionkehityksen aikana nieluun ilmestyy kidusrakoja ja vastaavia väliseiniä. Kiduslaitteiston osien muodostuminen maaselkärankaisten alkioissa selittyy niiden alkuperällä kalamaisista esivanhemmista, jotka hengittivät kidusten kanssa. Ihmisalkion sydämen rakenne alkuvaiheessa muistuttaa tämän elimen rakennetta kalassa: siinä on yksi eteinen ja yksi kammio. Hampaattomat valaat kehittävät hampaat alkion aikana. Nämä hampaat eivät puhkea, ne tuhoutuvat ja hajoavat.
Tässä annetut esimerkit ja monet muut viittaavat syvään yhteyteen organismien yksilöllisen kehityksen ja niiden historiallisen kehityksen välillä. Tämä yhteys ilmaistaan F. Müllerin ja E. Haeckelin 1800-luvulla muotoilemassa biogeneettisessä laissa: jokaisen yksilön ontogeneesi (yksilökehitys) on lyhyt ja nopea toisto sen lajin fysikaalisesta kehityksestä (historiallinen kehitys), johon tämä yksilö kuuluu. .
Johtopäätös
Abstraktia käsittelevän työn valmistuttuamme voimme tulla siihen tulokseen, että lisääntymiskyky eli itsensä lisääntyminen on yksi orgaanisen luonnon tärkeimmistä ominaisuuksista. Lisääntyminen on ominaisuus, joka kuuluu poikkeuksetta kaikille eläville organismeille bakteereista nisäkkäisiin.
Kaikkien eläin- ja kasvilajien, bakteerien ja sienten olemassaolo, jatkuvuus vanhempien yksilöiden ja heidän jälkeläistensä välillä säilyy vain lisääntymisen kautta. Läheisesti itsensä lisääntymiseen liittyy toinen elävien olentojen ominaisuus.organismien kehitystä. SeSe kuuluu myös kaikkeen maapallon elämään: pienimmille yksisoluisille organismeille sekä monisoluisille kasveille ja eläimille.
Bibliografia
Bogen G. Moderni biologia. - M.: Mir, 1970.
Green N., Stout W., Taylor D. Biology: 3 osassa T. 3: käännös. englannista/toim. R. Soper. - M.: Mir, 1990.
Mamontov S.G. Biologia. Yleiset kuviot. M.: Bustard, 2002.
Eläimistä ihmisiin. M.: Nauka, 1971.
Slyusarev A.A. Biologia ja yleinen genetiikka. - M.: Lääketiede, 1978.
Luento nro 3 Ontogeneesi
1. Gametogeneesi
2. Alkion aika
3. Postembryonaalinen ajanjakso
Ontogeneesi– organismin yksilöllinen kehitys tsygootista biologiseen kuolemaan. Progeneesi– Ontogeneesiä edeltävä ajanjakso sisältää gametogeneesin ja hedelmöittymisen. Fylogeneesi– lajin evoluutionaalinen kehitys.
Ihmisille on ominaista seksuaalinen lisääntyminen; sille on ominaista: sukurauhasten, sukusolujen, hedelmöitysprosessin ja sukupuolisen dimorfian esiintyminen.
Gametogeneesi - sukusolujen muodostumisprosessi ovogeneesi - munasolujen kypsyminen, spermatogeneesi- siittiöt. Gametogeneesi tapahtuu kehon sukurauhasissa. Sukusolut muodostuvat alkion epiteelisoluista, jotka muodostuvat organismin alkion kehitysvaiheen aikana.
Muodostumisprosessissa sukusolut käyvät läpi kolme vaihetta:
1. Lisääntymisjakso (alkion epiteelin solut jakautuvat mitoosilla);
2. Kasvukausi;
3. Kypsymisaika, solut jakautuvat meioosin kautta, mikä johtaa sukusolujen muodostumiseen (kuva 5).
Riisi. 5. Sukusolujen jakautuminen
Siittiöt kypsyvät kivesten siementiehyissä. Sijaitsee siemenputkien välissä interstitiaalinen kudos tuottavat miessukupuolihormonia - testosteroni. Miesten sukupuolihormonit - androgeenit, säätelevät lisääntymistoimintoa, gametogeneesiä ja toissijaisten seksuaalisten ominaisuuksien muodostumista. Siittiöt kypsyvät 70 päivässä. Siemenmäiset tubulukset sisältävät sukusoluja eri kypsymisvaiheissa. 5 ml:ssa. Ihmisen siemenneste sisältää 12 miljoonaa siittiötä. Ne ovat negatiivisesti varautuneita, varaus estää niitä tarttumasta yhteen. Suuri määrä urospuolisia sukusoluja on biologisesti tarkoituksenmukaista, niiden määrän väheneminen 40 % tekee hedelmöitysprosessin mahdottomaksi. Siittiöt ovat pieniä, liikkuvia soluja, jotka koostuvat päästä, kaulasta ja hännästä. Päässä on acrosome, modifioitu lysosomi, se sisältää entsyymejä, jotka liuottavat munasolun kalvon hedelmöityksen aikana. Elinajanodote on 6-72 tuntia.
Oogeneesi alkaa naisvartalon alkion kehitysvaiheessa. Meioosin ensimmäisen jakautumisen profaasivaiheessa, kun konjugaatio ja risteytyminen tapahtuu, tapahtuu väliaikainen tauko. Munasolun kehitys jatkuu tytön murrosiän aikana. Naissukupuolihormonien - estrogeenien - vaikutuksesta ensimmäisen asteen munasolun ympärille muodostuu follikkeli, solua suojaava ja ravitseva rakkula.
Kun munarakkula kypsyy, se siirtyy munasarjan reunaan ja sitten puhkeaa; tätä munasolun kehitysvaihetta kutsutaan munasarjaksi tai follikkeliksi; se kestää 12 päivää. Follikkelin repeämistä ja munasolun vapautumista munasarjasta vatsaonteloon kutsutaan ovulaatioksi.
Ovulaation jälkeen hedelmöitys on mahdollista. Naispuolisen sukusolun vangitsevat munanjohtimen fimbriat. Ripsivärisen epiteelin avulla se siirtyy kohtuun, kohtuvaihe kestää 12-14 päivää. Tähän mennessä naissukupuolihormonien vaikutuksesta kohdun epiteeli on löystynyt. Jos hedelmöittymistä ei tapahdu, epiteeli ja munat erittyvät kehosta kohdun seinämien sileiden lihasten supistumisen seurauksena. Tätä prosessia kutsutaan kuukautiseksi ja kestää 3-4 päivää. Siten aika follikkelien muodostumisen alusta kuukautisten alkamiseen on 20-30 päivää ja sitä kutsutaan munasarja-kuukautiskierroksi (Kuva 6).

Riisi. 6. Ovulaatio ja munasolun istuttaminen kohtuun
Se sisältää munasarjavaihe, kohtu ja kuukautiset. Munasarja-kuukautiskierron aikana naisen koko lisääntymisjärjestelmä uusiutuu, hormonien laadullinen ja määrällinen koostumus muuttuu, suorituskyky, voima, joustavuus ja kestävyys muuttuvat. Naisen kehon fysiologiset muutokset otetaan huomioon harjoitteluprosessia organisoitaessa.
Lannoitus- Tämä on munasolun ja siittiön fuusio. Lannoitusvaihetta on kolme:
1) akrosomaalinen reaktio (20 sekuntia) - munankuoren liukeneminen akrosomientsyymien vaikutuksesta ja siittiöiden tunkeutuminen munaan. Uskotaan, että siittiö, joka luo suurimman tunkeutumispotentiaalin (jännityksen) munakalvolle, tunkeutuu munan läpi;
2) väliaikainen tauko – munassa ei tapahdu näkyviä muutoksia, aineenvaihdunta kiihtyy;
3) munasolun ja siittiön ytimien fuusio, diploidisen kromosomijoukon palauttaminen.
Hedelmöityminen tapahtuu munanjohtimen ylemmässä kolmanneksessa. Hedelmöitetty munasolu on ns tsygootti. Tsygootin muodostumisen myötä ihmisen ontogeneesi alkaa.
2. Ontogeneesi ihmisellä on kaksi vaihetta: embryonaalinen ja postembryonaalinen.
Alkion kehitysvaihe jatkuu tsygootista syntymään. Sisältää vaiheet: tsygootti, blastula, gastrula, histogeneesi, organogeneesi.
Tsygootti- jakautuu toistuvasti mitoosilla, jolloin muodostuu monisoluinen yksikerroksinen alkio blastula. Blastulan koko ei kasva, koska tällä hetkellä se on munanjohtimessa ja sen pitäisi liikkua vapaasti kohtuun. Kuudentena päivänä se menee kohtuun ja kiinnittyy sen seinämään syöksymällä löystyneeseen epiteelin sisään. Tätä prosessia kutsutaan implantaatio. Alkio jatkaa kehittymistä ja siitä tulee kaksi-, sitten kolmikerroksinen. Tässä kehitysvaiheessa sitä kutsutaan gastrula. Tämän seurauksena muodostuu kolme alkiokerrosta: ektodermi, endodermi, mesodermi. Kudokset ja elimet muodostuvat alkiojärjestelmistä.
Täten: blastula- yksikerroksinen alkio, gastrula– kaksi- ja kolmikerroksinen alkio. Histogeneesi– kudosten levittäminen alkiokerroksista. Organogeneesi- urkujen laskeminen. 8 viikon iässä ihmisalkion massa on 4 g, koko 5 mm. Tähän mennessä hän oli käynyt läpi muodostumisprosesseja ja saanut ihmiskehon muodot. Kahdeksan raskausviikon jälkeen ihmisalkiota kutsutaan sikiöksi.
Ihmisalkion kehityksen aikana väliaikaiset viranomaiset, jotka menettävät toimintansa syntymän jälkeen. Väliaikaisiin viranomaisiin kuuluvat: itiö kuoret, chorion, amnion ja istukka, napanuora. Istukka varmistaa läheisen kosketuksen lapsen ja äidin kapillaarien välillä. Kehittyvä organismi saa ravinteita, happea, vasta-aineita istukan kautta ja poistaa aineenvaihduntatuotteita. Istukka suorittaa estetoimintoa ja suojaa alkiota mikro-organismeilta, eräiltä lääkkeiltä ja myrkkyiltä (kuva 7).

Riisi. 7. Istukka
1 - kohtu; 2 – amnion; 3 - alkio; 4 – lapsivesi; 5 – istukka; 6 – istukan kapillaarit; 7 – suonet; 8 – valtimot
Lapsen elinten ja kudosten hienovaraiset muodostumismekanismit voivat häiriintyä joutuessaan alttiiksi myrkyllisille, huumausaineille, alkoholille ja äidin veressä olevalle nikotiinille. Ihmisalkion kehityshäiriöitä kutsutaan synnynnäisiksi epämuodostumiksi ja epämuodostumiksi. Näitä ovat: "suulakihalkio" (ylemmän kitalaen puuttuminen), "huulihalkio" (leukaluiden yhteensulautumattomuus), polydaktylia (lisäsormet), syndaktylia (sulautuneet sormet). Epäsuotuisia ympäristötekijöitä, jotka aiheuttavat synnynnäisiä epämuodostumia, kutsutaan teratogeeninen(aiheuttaa epämuodostumia) (Kuva 8).

Riisi. 8. Ihmiskehon kehitykseen vaikuttavat ympäristötekijät
Ihmisen alkion kehitysjakso kestää 36 viikkoa.
3. Syntymän jälkeistä ja biologista kuolemaa edeltävää ajanjaksoa kutsutaan sikiön jälkeinen. Se sisältää nuori, murrosikä kehitys- ja ikääntymiskausia.
Nuori - kausi ennen murrosikää. Murrosikä– murrosiän aika.
Urheiluharjoittelussa, fysiologiassa, pedagogiikassa, psykologiassa käytetään seuraavaa ikäjakoa ihmisen postembryonaalinen ontogeneesi.
1-10 päivän aikana lapsi kutsutaan vastasyntynyt;
10 päivästä 1 vuoteen - rintakehä;
yhdestä vuodesta kolmeen ajanjaksoon varhaislapsuus;
4-7 vuotta - ensimmäinen lapsuus;
8-12-vuotiaat tytöille ja 13-vuotiaille pojille toinen lapsuus;
tytöille 12-15 ja pojille 13-16 - teini-ikäiset;
17-21 (pojat), 16-20 vuotta (tytöt) – teini-iässä;
20-55 naisille ja 21-60 miehille murrosikä(murrosikä);
55 ja 60 - 70 - iäkkäät ihmiset;
70-90 vanhat miehet;
yli 90 satavuotiaat.
Ihmisen ontogeneesissä on herkkä ja kriittinen kausia.
Kriittiset jaksot niille on ominaista yksittäisten geenien ja niiden kompleksien lisääntynyt aktiivisuus, jotka ohjaavat kehon mahdollisten merkkien kehittymistä. Tänä aikana säätelyprosessit muuttuvat merkittävästi, yksittäisten elinten ja toiminnallisten järjestelmien kehityksessä tapahtuu laadullinen ja määrällinen harppaus. Tänä aikana keho on herkin ympäristötekijöiden vaikutuksille. Ontogeneesin kriittisinä jaksoina pidetään implantaatiota, istukan muodostumista, aksiaalisten elinten muodostumista, hermo- ja suolistoputkia, notokordeja, sydämen muodostumista ja muita.
Jos nainen sairastuu vihurirokkoon 3. ja 9. raskausviikon välillä, on olemassa riski, että sikiölle kehittyy vikoja, kuten sydänsairauksia, kaihia ja kuuroutta. Muina aikoina vihurirokko ei aiheuta sikiön epämuodostumia.
Herkät jaksot– Nämä ovat aikoja, jolloin geneettinen kontrolli on heikentynyt ja kehon yksilölliset ominaisuudet ovat lisääntyneet ympäristövaikutuksille, mukaan lukien pedagogiset ja valmennus. Niinpä herkkä ajanjakso erilaisten nopeuden laadun indikaattoreiden ilmentymiselle tapahtuu 11-14-vuotiaana ja 15-vuotiaana sen maksimitaso saavutetaan, kun korkeat urheilusaavutukset ovat mahdollisia. Samanlainen kuva on havaittavissa ontogeneesissä kätevyyden ja joustavuuden ominaisuuksien ilmentymisestä.
Liikuntakasvatuksen ja urheilun parissa työskenteleville valmentajille ja opettajille herkkien ajanjaksojen tuntemuksella on suuri käytännön merkitys, sillä herkkien ajanjaksojen aikana harjoitusvaikutus.
Ne voidaan jakaa sen mukaan, kuinka monta solua on mukana suvuttomassa lisääntymisprosessissa: suvuton lisääntyminen, jossa yhdestä solusta syntyy tytärsukupolvi: solun jakautuminen monisolujen jakautuminen skitsogonia itiöityminen itiöityminen orastavassa yksisoluisessa hiivassa...
Jaa työsi sosiaalisessa mediassa
Jos tämä työ ei sovi sinulle, sivun alalaidassa on luettelo vastaavista teoksista. Voit myös käyttää hakupainiketta
Eliöiden lisääntyminen ja kehitys
Suunnitelma
- Lisääntymisen käsite ja merkitys.
- Lisääntymisen muodot ja tyypit.
- Solusykli. Mitoosi. Meioosi.
- Sukusolujen rakenne. Gametogeneesi.
1. Lisääntymisen käsite ja merkitys
Yksi elävien olentojen ominaisuuksista on diskreetti, nuo. millä tahansa organisaatiotasolla elävää ainetta edustavat perusrakenneyksiköt. Jokainen tietyn lajin yksilö on kuolevainen, ja lajin olemassaolo säilyy jäljentäminen eliöt. Siten elämän diskreetti edellyttää sen lisääntymistä, ts. lisääntymisprosessi.
Jäljentäminen tämä on elävien olentojen kyky lisääntyä omaa lajiaan. Tämä takaa elämän jatkuvuuden ja jatkuvuuden. Jatkuvuuden määrää se, että lisääntymisprosessin aikana perintöainesta siirtyy vanhemmilta jälkeläisille, minkä seurauksena vanhempien ominaisuudet ilmenevät tavalla tai toisella tytärorganismeissa.
Jäljennösarvo:
- Lajien lukumäärän lisääminen tai säilyttäminen. Lisääntymisen seurauksena ei tapahdu vain lisääntymistä (eli omaa lisääntymistä), vaan myös elävien organismien lukumäärän lisääntyminen.
- Elämän jatkuvuus. Lisääntymisen ansiosta enemmän tai vähemmän suuria yksilöryhmiä (esimerkiksi populaatioita ja lajeja) voi olla olemassa melko pitkään, koska yksilöiden luonnollisesta kuolemasta johtuvaa lukumäärän vähenemistä kompensoi organismien jatkuva lisääntyminen ja lisääntyminen. kuolleiden korvaaminen vastasyntyneillä.
- Jatkuvuus sukupolvien välillä. Lisääntymisprosessin aikana geneettistä tietoa välitetään vanhempien sukupolven yksilöiltä, mikä varmistaa tiettyjen vanhempien ja koko lajin, johon organismi kuuluu, ominaisuuksien lisääntymisen.
- Sellaiset elävien asioiden ominaisuudet kuin perinnöllisyys ja vaihtelevuus toteutuvat.
2. Lisääntymismuodot ja -tyypit
Lisääntymistä on kaksi päämuotoa: seksuaalinen ja aseksuaalinen.
Suvuton lisääntyminen
Aseksuaalisen lisääntymisen aikana uusi yksilö ilmestyy kehon erikoistumattomista ei-reproduktiivisista somaattisista soluista. Siksi aseksuaalinen lisääntyminen koskee yhtä yksilöä. Aseksuaalinen lisääntyminen varmistaa suuren määrän identtisten yksilöiden lisääntymisen, mikä on hyödyllistä jatkuvissa olosuhteissa eläville lajeille. Äidin kanssa identtisten jälkeläisten nopea ja runsas lisääntyminen on suvuttoman lisääntymisen biologinen merkitys.
Luonnossa on olemassa erilaisia vaihtoehtoja aseksuaaliseen lisääntymiseen:jakautuminen, itiöinti, pirstoutuminen, orastuminen, vegetatiivinen lisääntyminen, kloonaus.
Ne voidaan jakaa sen mukaan, kuinka monta solua on mukana suvuttoman lisääntymisen prosessissa:
- aseksuaalinen lisääntyminen, jossa tytärsukupolvi syntyy yhdestä solusta:
- solujen jakautuminen
- useiden solujen jakautuminen (skitsogonia)
- itiöinti (sporulaatio)
- orastava yksisoluisissa organismeissa (hiiva)
2) suvuton lisääntyminen, joka perustuu soluryhmän jakautumiseen:
- kasvullinen
- pirstoutuminen
- orastava monisoluisissa organismeissa (hydra)
On olemassa toinen luokitus, jonka mukaan aseksuaalisen lisääntymisen vaihtoehdot ovat:
- vegetatiivinen lisääntyminen (eli jakautuminen äidin kehon osiin):
Division
Moninkertainen fissio (skitsogonia)
Orastava
Hajanaisuus
Kasvien lisääminen pistokkailla, sipulilla, lehdillä, juurakoilla
- lisääntyminen itiöillä, ts. itiöintiä
Katsotaanpa tarkemmin aseksuaalisen lisääntymisen tyyppejä.
Division. Yksinkertaisin aseksuaalisen lisääntymisen muoto. Jakovaihtoehtoja on useita:
- Jakautuminen yksinkertaisella supistuksella, jolloin yhdestä emoorganismista muodostuu kaksi tytärorganismia. Tyypillistä bakteereille ja sinileville.
- Jakautuminen ytimen mitoottisella jakautumisella, jota seuraa sytoplasman erottaminen. Yksisoluisille organismeille ominaista (monet alkueläimet - ameba, vihreä euglena jne.; yksisoluiset levät - chlamydomonas jne.).
Molemmissa tapauksissa se tapahtuu binääri divisioona, ts. kahteen soluun. Toinen vaihtoehto on kuitenkin mahdollinen:
- Moninkertainen fissio (skitsogonia). Ensin tuma jakautuu toistuvasti ja sitten sytoplasma jakautuu osiin. Esimerkiksi Plasmodium falciparumin (malarian aiheuttaja) lisääntyminen ihmisen punasoluissa. Tässä tapauksessa plasmodia läpikäy ytimen toistuvan jakautumisen useita kertoja, minkä jälkeen sytoplasma jakautuu. Tämän seurauksena 1 plasmodium synnyttää 12-24 tytärorganismia.
Itiöiden muodostuminen (itiöityminen). Erittäin laajalle levinnyt elävien organismien aseksuaalinen lisääntymismenetelmä, jota löytyy melkein kaikista kasveista, sienistä ja joistakin alkueläimistä (esimerkiksi itiötyyppi) sekä prokaryoottisista organismeista (monet bakteerit, sinilevät).
Spore Tämä on solu, joka on peitetty kehittyneellä suojapäällysteellä - itiökalvolla, jonka avulla se kestää erilaisten epäsuotuisten ympäristötekijöiden vaikutuksia.Pienen koonsa vuoksi itiö sisältää yleensä vain vähän ravintovarastoja. Monissa organismeissa se on leviämisen pääyksikkö, koska muodostuneet suuret määrät kevyitä itiöitä kulkeutuvat vapaasti merkittävien etäisyyksien päähän ilmamassojen ja vesivirtojen liikkeestä.Usein itiöitä muodostuu suuria määriä ja niiden paino on vähäinen, mikä helpottaa niiden leviämistä tuulen sekä eläinten, pääasiassa hyönteisten, välityksellä.
Lisäksi joissain muodoissa (alkueläimet, sienet) itiöt voivat toimia elinkaaren erityisenä vaiheena, jolloin ne voivat "selviytyä" epäsuotuisten ympäristötekijöiden vaikutuksista.Bakteeri-itiöt eivät tarkalleen ottaen palvele lisääntymistä, vaan selviytymistä epäsuotuisissa olosuhteissa, koska jokainen bakteeri tuottaa vain yhden itiön. Bakteeri-itiöt ovat vastustuskykyisimpiä: esimerkiksi ne kestävät usein käsittelyä vahvoilla desinfiointiaineilla ja vedessä keittämistä.
Monissa kasveissa itiöiden muodostumisprosessi(sporogeneesi) suoritetaan erityisissä pussimaisissa rakenteissa, joita kutsutaan sporangiumiksi. Itiöt voivat olla joko liikkuvia piippulaitteen läsnäolon vuoksi (tässä tapauksessa niitä kutsutaan ns. eläinporit), ja liikkumaton, vailla kykyä liikkua aktiivisesti.
Zoosporeja havaitaan joissakin viherlevissä. Eläimillä itiöitymistä havaitaan malariaplasmodiumissa ja koko ryhmässä itiöitä (yksisoluisia organismeja).
Orastava. Sitä löytyy sekä yksisoluisista organismeista, esimerkiksi hiivasta ja tietyntyyppisistä väreistä, että monisoluisista organismeista, esimerkiksi coelenterate-tyypin (hydra) edustajista, sekä vaippaeläimistä (luokan askidiaanit).
Yksisoluisille organismeille tämä menetelmä koostuu tuberkkelin (uloskasvun) muodostumisesta emosolun ytimellä, joka sitten erottuu ja siitä tulee itsenäinen organismi.
Monisoluisille organismeille tämä menetelmä koostuu siitä, että äidin yksilön kehoon ilmestyy ensin pieni tuberkuloosi, jonka koko kasvaa, sitten ilmestyvät kaikkien äidin organismille ominaisten rakenteiden ja elinten alkeet. Sitten tapahtuu tytäryksilön erottaminen (silmuminen). Tämän jälkeen nuori, juuri erotettu organismi kasvaa ja saavuttaa alkuperäisen näytteen koon.
Riisi. Orastava orastavassa (hydra), 1 aikuinen organismi, 2 orastavaa tytärorganismia.
Epätavallinen orastumisen muoto on kuvattu mehevässä kasvissa Bryophyllum - kserofyytti, jota usein kasvatetaan koristekasvina: pienillä juurilla varustetut miniatyyrikasvit kehittyvät sen lehtien reunoja pitkin; nämä "silmut" putoavat lopulta ja alkavat olla itsenäisinä kasveina.
Hajanaisuus yksilön jakautuminen kahteen tai useampaan osaan, joista jokaisesta kehittyy uusi organismi. Fragmentoitumista tapahtuu esimerkiksi rihmalevissä, kuten Spirogyrassa. Spirogyra-filamentti voi hajota kahteen osaan missä tahansa paikassa, joista muodostuu myöhemmin kaksi organismia.
Fragmentoitumista havaitaan myös joissakin alemmissa eläimissä, jotka, toisin kuin paremmin organisoituneet muodot, säilyttävät merkittävän kyvyn uusiutua suhteellisen heikosti erilaistuneista soluista. Esimerkiksi nemertealaisten (ryhmä primitiivisiä, pääasiassa merimatoja) ruumis repeytyy erityisen helposti moniin osiin, joista jokainen voi synnyttää uusiutumisen seurauksena uuden yksilön. Tässä tapauksessa regeneraatio on normaali ja säännelty prosessi; kuitenkin joissakin eläimissä (esimerkiksi meritähtissä) yksittäisistä osista palautuminen tapahtuu vasta vahingossa tapahtuvan pirstoutumisen jälkeen. Sirpaloitumista havaitaan sienissä, coelenteraateissa (hydroissa), meduusoissa, piikkinahkaisissa, annelideissa ja lattamatoissa. Joskus tämä kyky on niin hyvin kehittynyt, että koko yksilö palautuu erillisestä fragmentista.
Riisi. . Meritähdin uusiutuminen yhdestä säteestä. A, B, C peräkkäiset regeneraatiovaiheet
Vegetatiivinen lisääntyminentämä on lisääntyminen, jossa uusi tytärorganismi kehittyy emoorganismista erotetusta osasta. Tässä tapauksessa emänäytteestä erotetaan melko hyvin erottuva osa, joka kehittyy edelleen itsenäiseksi kasveksi. Tai kasvi muodostaa erityisiä rakenteita, jotka on suunniteltu erityisesti vegetatiiviseen lisääntymiseen. Esimerkiksi sipulit, mukulat, mukulat, juurakot, langat, silmut. Jotkut näistä rakenteista toimivat myös ravinteiden varastoinnissa, jolloin kasvi selviää epäsuotuisista olosuhteista, kuten kylmästä tai kuivuudesta. Vegetatiivinen lisääntyminen on ominaista monille kasviryhmille levistä kukkiviin kasveihin.
Periaatteessa kasvullinen lisääntyminen ei käytännössä eroa sirpaloitumisesta tai silmusta, mutta perinteisesti tätä termiä käytetään kasviorganismeihin ja vain toisinaan eläimiin, toisin kuin pirstoutumisesta ja silmutuksesta.
Kloonaus. Tämä on keinotekoinen lisääntymismenetelmä, jota ei esiinny luonnossa. Se on yleistynyt vasta viimeisten 30-40 vuoden aikana ja sitä käytetään yhä enemmän taloudellisiin tarkoituksiin. On olemassa useita erikoistekniikoita, joiden avulla voit kloonata joitain kasveja ja eläimiä. Klooni Termillä tarkoitetaan geneettisesti identtisiä jälkeläisiä, jotka on saatu yhdestä yksilöstä yhden tai toisen suvuttoman lisääntymismenetelmän seurauksena. Tästä tämän menetelmän nimi tulee.
Kloonauskokeita tehdään erityisen laajalti kasveilla, mikä johtuu niiden korkeasta uusiutumiskyvystä. Yksittäiset solut asetetaan ravintoalustaan, jossa ne jakautuvat ja erityistekniikoilla saadaan aikaan epäjärjestynyt solumassa tai kallus. Sitten ne aiheuttavat primääristen homogeenisten kallusten erilaistumisen ja erilaisten kudosten ja elinten muodostumisen ja lopulta koko kasviorganismin, jolla on kaikki samat ominaisuudet kuin alkuperäisellä, josta solut on otettu.
Kloonausmenetelmää käyttämällä voidaan saada erilaisia hybridimuotoja. Siten entsyymien tai ultraäänen avulla poistetaan kasvisolujen soluseinät, minkä jälkeen syntyneet "paljaat" protoplastit voivat sulautua yhteen, jolloin syntyy hybridisoluja (esimerkiksi tomaatti-peruna tai tupakka-petunian hybridit). Tämän jälkeen soluseinät palautetaan, muodostuu kallus ja sitten koko hybridikasvi.
Eläinten tapauksessa käytetään seuraavaa menetelmää: munan tuma poistetaan tai tuhotaan ja sen tilalle asetetaan somaattisen solun (esimerkiksi epiteelisolun) ydin. Tulevaisuudessa tällaisesta munasta voidaan saada organismi, joka on ominaisuuksiltaan identtinen eläimen ytimen luovuttajan kanssa. Tällä tavalla voit saada klooneja joistakin eläimistä kynsi sammakoita(Xenopus), newts (Triturus). Tällä hetkellä on saatu jopa nisäkkäiden klooneja, kuten tunnettu lammas Dolly. Kloonaustekniikoiden kehittämistä suorittaa erityinen biologian haara - biotekniikka, jonka tehtävät eivät rajoitu tähän.
Seksuaalinen lisääntyminen
Seksuaalinen lisääntyminenkutsutaan sukupolvien vaihdoksi ja organismien kehitykseksi, joka perustuu erikoistuneiden sukusolujen fuusioon ( sukusolut ) ja tsygootin muodostuminen. Sukusolut (naaras ja uros, muna ja siittiöt) muodostuvat sukurauhasissa.
Sukupuolinen lisääntyminen on tyypillistä suurimmalle osalle elävistä olentoista, se on edistyksellisempää kuin sukupuolinen lisääntyminen ja sillä on valtavia geneettisiä etuja. Sukupuolinen lisääntyminen varmistaa parhaiten jälkeläisten geneettisen monimuotoisuuden, koska on yhdistelmä geenejä, jotka aiemmin kuuluivat molemmille vanhemmille. Lajien muodostavien yksilöiden genotyyppien monimuotoisuus mahdollistaa lajin onnistuneemman ja nopeamman sopeutumisen muuttuviin ympäristöolosuhteisiin.
Hedelmöityksen aikana sukusolut sulautuvat yhteen muodostaen diploidin tsygootti josta kehitysprosessissa saadaan kypsä organismi. Sukusolut ovat haploideja - ne sisältävät yhden joukon kromosomeja, tsygootti on diploidi, joka sisältää kaksinkertaisen joukon kromosomeja. Tämä on tulevan organismin ensimmäinen solu.
Sukusoluja on yleensä kahta tyyppiä - miehiä ja naisia. Niitä tuottavat uros- ja naarasvanhemmat, jos laji on kaksikotinen; ( sellaisia ovat jotkin kukkivat kasvit, useimmat eläimet ja ihmiset) tai sama yksilö (harmafroditismi).
Tyypillinen uros- ja naaraseläimilleseksuaalinen dimorfismi- eli seksuaaliset erot rakenteessa, ulkonäössä (koko, väri ja muut ominaisuudet) sekä käyttäytymisessä. Eläimillä sitä esiintyy jo evolutionaarisen kehityksen alemmissa vaiheissa, esimerkiksi pyöreissä helminteissä ja niveljalkaisissa, ja saavuttaa suurimman ilmenemismuotonsa selkärankaisissa, joissa urosten ja naarasten väliset ulkoiset erot ovat erittäin ilmeisiä. Niiden lajien kasveissa, joille on ominaista uros- ja naarasyksilöiden läsnäolo, esiintyy myös seksuaalista dimorfismia, mutta se on hyvin vähän ilmaistua.
Sukupuolisen ja aseksuaalisen lisääntymisen vertailu on esitetty taulukossa. 5
Pöytä 5
Aseksuaalisen ja seksuaalisen lisääntymisen vertailu
|
Suvuton lisääntyminen |
Seksuaalinen lisääntyminen (pois lukien bakteerit) |
|
|
Yksi vanhempi |
Yleensä kaksi vanhempaa |
|
|
Sukusoluja ei muodostu |
Muodostuu haploidisia sukusoluja, joiden ytimet sulautuvat (hedelmöitys) muodostaen diploidisen tsygootin |
|
|
Ei meioosia |
Jossain elinkaaren vaiheessa esiintyy meioosia, joka estää kromosomien kaksinkertaistumisen jokaisessa sukupolvessa. |
|
|
Jälkeläiset ovat identtisiä vanhempiensa kanssa. Ainoa geneettisen variaation lähde on satunnaiset mutaatiot |
Jälkeläiset eivät ole identtisiä vanhempainhenkilöitä. Niillä on geneettistä vaihtelua, joka johtuu geneettisestä rekombinaatiosta. Tämä edistää vahvimpien ja vahvimpien yksilöiden luonnollista valintaa ja siten evoluutiota. |
|
|
Kasveille, joillekin alemmille eläimille ja mikro-organismeille ominaista. Ei löydy korkeammista eläimistä |
Ominaista useimmille kasveille ja eläimille |
|
|
Usein johtaa nopeaan suuren määrän jälkeläisiä syntymiseen |
Vähemmän nopeaa määrän kasvua |
Seksuaalinen lisääntyminen, riippuen sukusolujen koon ja toiminnan suhteesta, voi olla kolme vaihtoehtoa:
Isogamia (levät, alkueläimet). Miesten ja naisten sukusolut ovat kooltaan, rakenteeltaan ja liikkuvuudeltaan samanlaisia.
Heterogamia. Solut eroavat kooltaan ja rakenteeltaan.
Oogamy. Tämä on yksi heterogamian muunnelmista, kun muna on suuri ja liikkumaton ja siittiö on kooltaan pienempi, sillä on liikeelin ja liikkuvuus.
On olemassa erityisiä seksuaalisen lisääntymisen muotoja, kuten hermafroditismi ja partenogeneesi.
Hermafroditismi. Termi "hermafroditismi" on yhdistelmä kreikkalaisista nimistä Hermes (miesten kauneuden jumala) ja Aphrodite (naisen kauneuden jumalatar).
Useimmissa hermafrodiittilajeissa hedelmöityksessä on kuitenkin mukana eri yksilöistä peräisin olevia sukusoluja, ja niillä on lukuisia geneettisiä, morfologisia ja fysiologisia mukautuksia, jotka estävät itsensä hedelmöittymisen ja suosivat ristihedelmöitystä. Esimerkiksi monilla alkueläimillä itsensä hedelmöittymistä estää geneettinen yhteensopimattomuus, monilla kukkakasveilla androeciumin ja gynoeciumin rakenne ja monilla eläimillä se, että munasolut ja siittiöt muodostuvat samassa yksilössä eri aikoina.
Todellisen hermafroditismin ilmiö löytyy myös järjestäytyneemmistä olennoista. Erityisesti sitä löytyy nisäkkäistä. Esimerkiksi sioilla havaitaan toisinaan munasarjojen kehittymistä toisella kehon puolella ja kivesten kehittymistä toisella tai yhdistettyjen rakenteiden kehittymistä (ovotestis) ja molemmissa tapauksissa toiminnallista synteesiä. aktiivisia munasoluja ja siittiöitä tapahtuu. Tällaiset eläimet luokitellaan "keskikokoiseen" sukupuoleen, ja suurin osa keskitason sukupuolen yksilöistä on naaraita, joilla on kaksi XX-kromosomia. Samanlainen ilmiö on havaittu vuohilla.
Todellista hermafroditismia esiintyy myös ihmisillä, mikä johtuu kehityshäiriöistä. Hermafrodiittien genotyypit ovat 46XX tai 46 XY , useimmissa tapauksissa viitaten XX (noin 60 %). Väärä hermafroditismi tunnetaan myös, kun yksilöillä on molemmille sukupuolille tyypillisiä ulkoisia sukupuolielimiä ja toissijaisia sukupuoliominaisuuksia, mutta ne tuottavat vain yhden tyyppisiä sukusoluja - miehen tai naisen.
Useimmilla kukkivilla kasveilla on hermafrodiittikukkia, joita kutsutaan yleensä biseksuaaleiksi, koska jokainen kukka sisältää emen ja heteitä. Tästä syystä hedelmät kehittyvät kaikista kukista. Vehnä, kirsikka, omena ja monet muut kasvilajit ovat biseksuaaleja. Biseksuaalien lisäksi kehittyi evoluution aikana kasveja, joiden sukupuolet ovat eronneet saman lajin sisällä, eli kasveista syntyi yksikotisia ja kaksikotisia. Yksikotisiksi kutsutaan kasveja, jotka sisältävät sekä naaraskukkia että uroskukkia. Yksikotisissa kasveissa hedelmät kehittyvät vain emikukkaista. Maissi on yksikotinenkurkku, kurpitsa ja muut. Sitä vastoin kaksikotiset kasvit ovat kasveja, jotka sisältävät joko emi- tai tärinäkukkia (saman lajin sisällä). Kaksikotisissa kasveissa vain ne, joilla on emikukkaiset kukat (naaraat), kantavat hedelmää. Poppeli, mansikka ja muut puu- ja ruohomaiset kasvit ovat kaksikotisia.
Parthenogenesis (kreikaksi. parthenos neitsyt ja synty syntymä) on yksi sukupuolisen lisääntymisen muunnelmista, jossa naarassukusolusta kehittyy uusi yksilö ilman urospuolisen sukusolun hedelmöittymistä. Partenogeneesin etuna on, että joissakin tapauksissa se lisää lisääntymisnopeutta, ja sosiaalisissa hyönteisissä sen avulla voit säädellä kunkin tyypin jälkeläisten määrää. Partenogeneesi voi olla pakollinen (pakollinen) ja fakultatiivinen (valinnainen).
Esimerkiksi mehiläisessä(Apis mellifera) kuningatar munii hedelmöittyneitä munia(2п = 32), jotka kehittyessään synnyttävät naaraat (kuningattaret tai työläiset) ja hedelmöittämättömiä munia(P = 16), jotka tuottavat miehiä (droneja), jotka tuottavat spermiiniä mieluummin mitoosilla kuin meioosilla.
Riisi. Partenogeneesityyppien kaavio.
- Solusykli. Mitoosi. Meioosi
Solujen jakautuminen on organismien lisääntymisen ja yksilöllisen kehityksen perusta.
Kaikki elävät organismit koostuvat soluista. Kehon tyypillisen rakenteen kehitys, kasvu ja muodostuminen tapahtuu yhden tai ryhmän alkuperäisiä soluja lisääntymisen kautta. Elämän aikana osa kehon soluista kuluu, vanhenee ja kuolee. Säilyttääkseen rakenteen ja normaalin toiminnan kehon on tuotettava uusia soluja vanhojen tilalle. Ainoa tapa muodostaa soluja on jakaa edelliset.
Solunjako elintärkeä prosessi kaikille organismeille. Ihmiskehossa, joka koostuu noin 10 13 soluja, useiden miljoonien on jaettava joka sekunti.
Kolme eukaryoottisolujen jakautumismenetelmää on kuvattu: amitoosi (suora jako), mitoosi (epäsuora jako) ja meioosi (vähennysjako).
Amitoosi suhteellisen harvinainen ja vähän tutkittu solunjakomenetelmä. Se on kuvattu ikääntyville ja patologisesti muuttuneille soluille. Amitoosissa faasien välinen ydin jaetaan supistumisen avulla, eikä perinnöllisen materiaalin tasaista jakautumista taata. Usein tuma jakautuu ilman myöhempää sytoplasman erottamista ja muodostuu kaksitumaisia soluja. Amitoosin läpikäynyt solu ei pysty myöhemmin osallistumaan normaaliin mitoosisykliin. Siksi amitoosia esiintyy yleensä kuolemaan tuomituissa soluissa ja kudoksissa, esimerkiksi nisäkkäiden alkiokalvojen soluissa ja kasvainsoluissa.
Mitoosi universaali menetelmä eukaryoottisolujen jakamiseen. Sen kesto eläinsoluissa on noin 1 tunti Mitoosi on jatkuva prosessi, joka on perinteisesti jaettu neljään vaiheeseen: profaasi, metafaasi, anafaasi ja telofaasi.
Tapahtumasarjaa, joka tapahtuu tietyn solun muodostumisen ja sen tytärsoluiksi jakautumisen välillä, kutsutaansolusykli. Tämä sykli koostuu kolmesta päävaiheesta:
1. Välivaihe. Intensiivisen synteesin jakasvu. Solu syntetisoi monia aineita, joita tarvitaan sen kasvuun ja kaikkien sen luontaisten toimintojen toteuttamiseen. Interfaasin aikana tapahtuu DNA:n replikaatio.
2. Mitoosi. Tämä on tuman jakautumisprosessi (karyokineesi), jossa kromatidit erotetaan toisistaan ja jaetaan uudelleen kromosomeiksi tytärsolujen välillä.
3. Sytokineesi on sytoplasman jakautumisprosessi (sytokineesi) kahden tytärsolun välillä.
Solusyklin pituus riippuu solutyypistä ja ulkoisista tekijöistä, kuten lämpötilasta, ravintoaineista ja hapesta. Bakteerisolut voivat jakautua 20 minuutin välein, suolen epiteelisolut - 8-10 välein, sipulin juuren kärjessä olevat solut - 20 tunnin välein, ja monet hermoston solut eivät koskaan jakautu.
Prosessien välinen suhde ajan kuluessa on esitetty kohdassa riisi.

Riisi. Solusyklin vaiheet
Interfaasi koostuu useista jaksoista: G 1, S, G 2.
Jakso G 1 kutsutaan presynteettisiksi. Kestoltaan eniten vaihteleva. Tällä hetkellä solussa aktivoituvat biologiset synteesiprosessit, pääasiassa rakenteelliset ja toiminnalliset proteiinit. Solu kasvaa ja valmistautuu seuraavaan jaksoon. Tänä aikana tapahtuu intensiivisiä biosynteesiprosesseja. Mitokondrioiden, kloroplastien (kasveissa), endoplasmisen retikulumin, lysosomien, Golgi-laitteiston, tyhjiöiden ja rakkuloiden muodostuminen. Tuma tuottaa rRNA:ta, mRNA:ta ja tRNA:ta; muodostuu ribosomeja; solu syntetisoi rakenteellisia ja toiminnallisia proteiineja. Entsyymien hallitsema intensiivinen soluaineenvaihdunta. Solujen kasvu. Sellaisten aineiden muodostuminen, jotka estävät tai stimuloivat seuraavan vaiheen alkamista.
Kausi S kutsutaan synteettisiksi. Tämä on mitoosisyklin pääjakso. Jakautuvissa nisäkässoluissa se kestää noin 6 x 10 tuntia, missä tapahtuu DNA:n replikaatio. Synteesi proteiinimolekyylejä kutsutaan histoneiksi, jotka sitoutuvat jokaiseen DNA-juosteeseen. Jokainen kromosomi muuttuu kahdeksi kromatidiksi.
G2-jaksoa kutsutaan postsynteettinen.Se on suhteellisen lyhyt, nisäkässoluissa noin 2 x 5 tuntia.Tänä aikana sentriolien, mitokondrioiden ja plastidien määrä kaksinkertaistuu, tapahtuu aktiivisia aineenvaihduntaprosesseja, kerääntyy proteiineja ja energiaa tulevaa jakautumista varten. Solu alkaa jakautua. Tapahtuu intensiivisiä biosynteesiprosesseja. Mitokondrioiden ja kloroplastien jakautuminen. Lisääntynyt energiavarasto. Sentriolien replikaatio (soluissa, joissa niitä on) ja karan muodostumisen alku
Mitoosi ehdollisesti jaettu neljään vaiheeseen: profaasi, metafaasi, anafaasi ja telofaasi
Prophase . DNA:n spiralisoituminen alkaa ja lisääntyy vähitellen ytimessä. Kromosomit lyhenevät, paksuuntuvat, näkyvät ja saavat tyypillisen bikromatidisen rakenteen. Tuma häviää vähitellen. Sytoplasmassa mikrotubulukset ovat suunnattu jokaisen sentrioliparin ympärille muodostaen karakeskuksia. Sentriolit siirtyvät eri napoihin, mikrotubulukset ulottuvat soluakselia pitkin ja alkaa muodostua akromatiinikara. Ydinvaippa hajoaa erillisiksi pieniksi paloiksi. Kromosomit liikkuvat kohti solun keskustaa
Metafaasi . Kromosomit ovat maksimaalisesti spiraalimaisia ja järjestetty siten, että niiden sentromeerit sijaitsevat samassa tasossa - soluekvaattorin tasolla. Muodostuu metafaasilevy, mitoottisen karan muodostuminen on valmis. Sentriolit sijaitsevat pareittain vastakkaisissa navoissa, ja eri napojen karalangat on kiinnitetty kunkin kromosomin sentromeeriin.
Anafaasi . Tämä on mitoosin lyhin vaihe. Täällä tapahtuu kunkin kromosomin pituussuuntainen halkeaminen, lankojen väheneminenkarat ja kromatidien (tytärkromosomien) hajoaminen solun napoja kohti.
Telofaasi . Tytärkromosomit, jotka koostuvat yhdestä kromatidista, saavuttavat solun navat. DNA, joka muodostaa ne, alkaa hajota, ytime ilmestyy, ydinkalvo muodostuu kunkin tytärkromosomiryhmän ympärille ja akromatiinikaran langat hajoavat vähitellen. Ydinfissio on valmis.
Sytoplasman jakautuminen alkaa(sytotomia) ja väliseinän muodostuminen tytärsolujen välille. Eläinsolut suorittavat sytotomian supistamalla sytoplasmista kalvoa. Kasveissa solun ekvaattorin tasoon muodostuu kalvoseinä, joka kasvaa sivusuunnassa ja saavuttaa soluseinän. Tämän seurauksena muodostuu kaksi täysin erillistä tytärsolua.
Seurataan muutos perinnöllisissä aineissamitoosisyklin aikana. Mitoosisyklin päätapahtumat ovat DNA kopiointi esiintyy välivaiheessa ja johtaa perinnöllisen tiedon määrän kaksinkertaistumiseen, jakromatidisegregaatio,tapahtuu mitoosin anafaasissa ja varmistaa perinnöllisen tiedon tasaisen jakautumisen tytärsolujen välillä. Perinnöllinen materiaali suorittaa nämä prosessit eri rakenteellisissa muodoissa. Replikatiivinen synteesi käy läpiinterfaasikromatiini, injossa DNA-molekyyli on suhteellisen despiralisoituneessa tilassa. Geneettisen tiedon jakelu tapahtuumitoottiset kromosomit,jossa DNA on maksimaalisesti spiraalistunut.
Mitoosisyklissä myös perinnöllisen materiaalin määrä muuttuu. Jos kromosomien lukumäärä haploidissa joukossa on merkitty kirjaimella p (diploidisessa joukossa vastaavasti 2n), ja DNA-molekyylien lukumäärä merkitään kirjaimella Kanssa, silloin on mahdollista jäljittää somaattisen solun ytimen kaavan muutos mitoosisyklin eri vaiheissa. Ennen S -jakso, jolloin jokainen kromosomi koostuu yhdestä DNA-molekyylistä, DNA:n kokonaismäärä ytimessä vastaa siinä olevien kromosomien lukumäärää ja diploidisen solun kaava on muotoa 2p2s. Replikaation jälkeen, kun kunkin kromosomin DNA kaksinkertaistuu, DNA:n kokonaismäärä tumassa kaksinkertaistuu ja solukaava saa muotoa 2n4c. Kromatidierotuksen seurauksena mitoosin anafaasissa tytärytimet saavat diploidisen joukon yksikromatidisia kromosomeja. Tytärsolujen kaava muuttuu jälleen 2p2s.
Mitoosin biologinen merkityson, että tämän jakautumismenetelmän seurauksena soluihin muodostuu perinnöllistä tietoa, joka on laadullisesti ja määrällisesti identtinen emosolun tiedon kanssa. Perinnöllisen materiaalin tasainen jakautuminen varmistetaan DNA:n replikaatio- ja kromosomien kaksinkertaistumisprosesseilla mitoosisyklin välivaiheessa sekä kromatidien spiralisoitumisella ja tasaisella jakautumisella tytärsolujen välillä mitoosin aikana. Mitoosi varmistaa karyotyypin pysyvyyden ylläpidon useiden solusukupolvien ajan ja toimii solumekanismina kehon kasvu- ja kehitysprosesseille sekä regeneraatiolle ja aseksuaaliselle lisääntymiselle.
Useiden ympäristötekijöiden toiminta voi häiritä mitoosin normaalia kulkua ja johtaa kromosomien vaurioitumiseen sekä muutoksiin yksittäisten kromosomien tai kokonaisten kromosomiryhmien lukumäärässä kehon somaattisissa soluissa. Patologiset mitoosit voivat aiheuttaa useita kromosomisairauksia. Patologisia mitoosia havaitaan erityisen usein kasvainsoluissa.
Mitoosikaavio:
MITOOSI
|
1. Välivaihe Sitä kutsutaan usein väärin lepovaiheeksi. Interfaasin kesto vaihtelee ja riippuu tietyn solun toiminnasta. Tämä on ajanjakso, jonka aikana solu normaalisti syntetisoi organelleja ja kasvaa kokoa. Nukleolit ovat selvästi näkyvissä ja syntetisoivat aktiivisesti ribosomaalista materiaalia. Juuri ennen solujen jakautumista kunkin kromosomin DNA ja histonit replikoituvat. Jokaista kromosomia edustaa nyt kromatidipari, joka on liitetty toisiinsa sentromeerillä. Kromosomien aines värjäytyy ja sitä kutsutaan kromatiiniksi, mutta itse näitä rakenteita on vaikea nähdä. |
|
|
2. Prophase Yleensä solunjakautumisen pisin vaihe. Kromatidit lyhenevät (jopa 4 % alkuperäisestä pituudestaan) ja paksuuntuvat spiralisoitumisen ja kondensoitumisen seurauksena. Värjättäessä kromatidit ovat selvästi näkyvissä, mutta sentromeerit eivät ole näkyvissä. Eri kromatidipareissa sentromeeri sijaitsee eri tavalla. Eläinsoluissa ja alemmissa kasveissa sentriolit hajoavat solun vastakkaisiin napoihin. Jokaisesta sentriolista lähtee lyhyitä mikrotubuluksia säteiden muodossa, jotka muodostavat yhdessä tähden. Nukleolit pienenevät, koska niiden nukleiinihappo siirtyy osittain tiettyihin kromatidipareihin. Profaasin loppua kohti ydinkalvo hajoaa ja muodostuu fissiokara. |
|
|
3. Metafaasi Kromatidiparit kiinnittyvät sentromeerien avulla karafilamentteihin (mikrotubuluksiin) ja liikkuvat ylös ja alas karassa, kunnes niiden sentromeerit ovat linjassa karan ekvaattoria pitkin kohtisuoraan sen akseliin nähden. |
|
|
4. Anafaasi Tämä on hyvin lyhyt vaihe. Jokainen sentromeeri jakautuu kahteen osaan, ja karafilamentit vetävät tytärsentromeerit vastakkaisille napoille. Sentromeerit vetävät taakseen toisistaan eronneet kromatidit, joita nykyään kutsutaan kromosomeiksi. |
|
|
5. Telofaasi Kromosomit saavuttavat solun navat, venyvät, pidentyvät, eikä niitä enää voida erottaa selvästi. Karan filamentit tuhoutuvat ja sentriolit replikoituvat. Ydinkalvo muodostuu kromosomien ympärille jokaiseen napaan. Nucleolus ilmestyy uudelleen. Telofaasia voi välittömästi seurata sytokineesi (koko solun jakautuminen kahteen osaan). |
|





Meiosis (kreikan sanasta meiosis vähentäminen) on erikoinen tapa jakautua soluissa, mikä johtaa kromosomien lukumäärän puolittumiseen niissä. Meioosi on keskeinen linkki gametogeneesi eläimillä ja sporogeneesi kasveissa. Meioosi koostuu kahdesta peräkkäisestä jakautumisesta, joita edeltää yksittäinen DNA:n replikaatio. Kaikki molempien jakautumiseen tarvittavat aineet ja energia varastoituvat meioosia edeltävän interfaasin aikana minä Interfaasi II käytännössä poissa, ja jakautumiset seuraavat nopeasti yksi toisensa jälkeen. Jokaisessa meioottisessa jaossa erotetaan samat neljä vaihetta: profaasi, metafaasi, anafaasi ja telofaasi, jotka ovat ominaisia mitoosille, mutta eroavat useista piirteistä.
Ensimmäinen meioottinen jakautuminen (meioosi minä ) johtaa kromosomien lukumäärän puolittumiseen ja sitä kutsutaan pelkistymiseksi. Seurauksena yhdestä diploidisesta solusta(2s 4c) muodostuu kaksi haploidista solua(p 2c) solut.
Profaasi I meioosi on pisin ja monimutkaisin. Mitoosin profaasille tyypillisten DNA-heliksaatio- ja karan muodostumisprosessien lisäksi minä Kaksi erittäin tärkeää biologista tapahtumaa tapahtuu: konjugaatio tai synapsi homologiset kromosomit ja ylittämällä.
Konjugaatio on prosessi, jossa homologiset kromosomit ovat lähellä toisiaan. Nämä kromosomiparit muodostuvat bivalenttinen ja ne säilyvät koostumuksessaan erityisten proteiinien avulla. Koska jokainen kromosomi koostuu kahdesta kromatidista, kaksiarvoinen sisältää neljä kromatidia ja sitä kutsutaan myös muistikirja. Diploidisolu tuottaa P bivalentteja. Konjugaation jälkeen solukaava saa muodon kulkea.
Joissakin bivalenttien paikoissa konjugoitujen kromosomien kromatidit leikkaavat, rikkoutuvat ja vaihtavat vastaavia osia. Tätä homologisten kromosomien fragmenttien vaihtoprosessia kutsutaan crossing overiksi. Se varmistaa uusien isän ja äidin geenien yhdistelmien muodostumisen tulevien sukusolujen kromosomeihin. Crossing over voi tapahtua useilla alueilla (multiple crossing over), mikä tarjoaa suuremman perinnöllisen tiedon rekombinaatioasteen sukusoluissa. Profaasin loppuun mennessä minä kromosomien spiralisoitumisaste kasvaa, kromatidit tulevat selvästi erotettavissa, karalangat jokaisesta navasta kiinnittyvät yhden bivalentin kromosomin sentromeeriin. Ydinvaippa tuhoutuu ja kaksiarvoiset aineet suuntautuvat solun ekvatoriaalista tasoa kohti.
Metavaiheessa I Meioosi viimeistelee karan muodostumisen, kaksiarvoiset aineet asennetaan solun ekvatoriaaliseen tasoon. Yhden navan karasäikeet on kiinnitetty kunkin kromosomin sentromeeriin.
Anafaasissa I Meioosissa karan säikeiden vaikutuksesta homologiset kromosomit siirtyvät poispäin toisistaan ja suuntautuvat solun vastakkaisiin napoihin. Tämän seurauksena kennon jokaisessa navassa ahaploidi settikromosomit, jotka sisältävät yhden bikromatidikromosomin jokaisesta homologisesta kromosomiparista. Anafaasissa minä eri parien kromosomit, ts. ei-homologiset kromosomit käyttäytyvät täysin toisistaan riippumatta ja varmistavat monenlaisten kromosomien muodostumisen. yhdistelmiä isän ja äidin kromosomit tulevien sukusolujen haploidisessa sarjassa. Tällaisten yhdistelmien lukumäärä vastaa kaavaa 2 P, missä p homologisten kromosomien parien lukumäärä. Ihmisillä tämä arvo on 2, ts. 8.4 Ihmisen sukusoluissa on mahdollista 10 muunnelmaa isän ja äidin kromosomien yhdistelmistä.
Joten, homologisten kromosomien erot anafaasissa minä meioosi ei takaa vain kromosomien määrän vähenemistä tulevissa sukusoluissa, vaan myös jälkimmäisten valtavan monimuotoisuuden johtuen eri parien isän ja äidin kromosomien satunnaisesta yhdistelmästä.
Telofaasissa I Meioosi tapahtuu sellaisten solujen muodostumisessa, joiden ytimissä on haploidinen kromosomisarja ja kaksinkertainen määrä DNA:ta, koska jokainen kromosomi koostuu kahdesta kromatidista. Ensimmäisestä meioottisesta jakautumisesta syntyvillä soluilla on kaava p2s ja lyhyen välivaiheen jälkeen he aloittavat seuraavan jaon.
Toinen meioottinen jakautuminen (meioosi II ) etenee tyypillisenä mitoosina (kuva 5.3), mutta eroaa siinä, että siihen tulevat solut sisältävät haploidisen kromosomijoukon. Tämän jaon seurauksena P bikromatidiset kromosomit (r2c), halkeilu, muoto P yksikromatidiset kromosomit(ps). Tätä jakoa kutsutaan yhtälö (tai tasaus).
Siten kahden peräkkäisen meioottisen jakautumisen jälkeen yhdestä solusta, jossa on diploidisarja kaksikromatidisia kromosomeja (2x4c), muodostuu neljä solua, joissa on haploidinen yksikromatidisten kromosomien sarja(ps).
Meioosin biologinen merkityskoostuu sellaisten solujen muodostumisesta, joissa on vähentynyt kromosomisarja, ja karyotyypin pysyvyyden ylläpitämisestä useissa lisääntyvien organismien sukupolvissa. sukupuoliteitse tarttuvat. Meioosi toimii perustana kombinatiiviselle vaihtelulle tarjoamalla sukusolujen geneettistä monimuotoisuutta isän ja äidin kromosomien risteytys-, eroamis- ja kombinatoriikan kautta. Muutokset kromosomirakenteessa, jotka johtuvat epätasaisesta risteyksestä, kaikkien tai yksittäisten kromosomien hajoamisen häiriintymisestä anafaasissa I ja II meioottiset jakautumiset johtavat epänormaalien sukusolujen muodostumiseen ja voivat toimia perustana organismin kuolemalle tai useiden kromosomaalisten oireyhtymien kehittymiselle jälkeläisissä.
Kaavio ja lyhyet kuvaukset meioosin peräkkäisistä vaiheista eläinsolussa.
|
1. Välivaihe Kesto vaihtelee lajeittain. Organellien replikaatio tapahtuu ja solun koko kasvaa. DNA:n ja histonien replikaatio päättyy pääasiassa premeioottiseen interfaasiin, mutta ulottuu osittain myös profaasin alkuun. Jokaista kromosomia edustaa nyt kromatidipari, joka on yhdistetty sentromeerillä. Kromosomimateriaali värjäytyy, mutta kaikista rakenteista vain nukleolit ovat selvästi näkyvissä. |
A. Varhainen profaasi minä |
|
2. Prophase Pisin vaihe. Se on usein jaettu viiteen vaiheeseen (leptoteema, zygonema, pakyneema, diplonema ja diakineesi), mutta tässä sitä pidetään jatkuvana kromosomimuutosten sarjana. A . Kromosomit lyhenevät ja tulevat näkyviin erillisinä rakenteina. Joissakin organismeissa ne näyttävät helmijonoilta: voimakkaasti värjäytyneen materiaalin alueet - kromomeerit - vuorottelevat värjäytymättömien alueiden kanssa. Kromomeerit ovat paikkoja, joissa kromosomimateriaali on voimakkaasti kiertynyt. |
|
|
B. Homologiset kromosomit, jotka ovat peräisin äidin ja isän sukusolujen ytimistä, lähestyvät toisiaan ja konjugoituvat. Nämä kromosomit ovat saman pituisia, niiden sentromeerit ovat samassa paikassa ja ne sisältävät yleensä saman määrän geenejä samassa lineaarisessa sekvenssissä. Homologisten kromosomien kromomeerit sijaitsevat vierekkäin. Konjugaatioprosessi voi alkaa useista pisteistä kromosomeissa, jotka sitten yhdistetään koko pituudelta (ikään kuin vetoketjut yhteen). Konjugoitujen homologisten kromosomien pareja kutsutaan usein bivalentteiksi. Bivalentit lyhenevät ja paksuuntuvat. Tässä tapauksessa tapahtuu sekä tiheämpää pakkausta molekyylitasolla että ulospäin havaittavaa vääntymistä (spiralisoitumista). Nyt jokainen kromosomi sentromeeriineen on selvästi näkyvissä. |
B. Profaasi I |
|
SISÄÄN. Homologiset kromosomit, jotka muodostavat bivalentin, ovat osittain erotettuja, ikään kuin työntäisivät pois toisistaan. Nyt voit nähdä, että jokainen kromosomi koostuu kahdesta kromatidista. Kromosomit ovat edelleen yhteydessä toisiinsa useissa kohdissa. Näitä pisteitä kutsutaan chiasmataksi (kreikasta. chiasma - risti). Jokaisessa kiasmassa kromatidien osiot vaihtuvat katkosten ja jälleenyhtymien seurauksena, joissa kaksi kussakin kiasmassa olevista neljästä langasta on mukana. Tämän seurauksena yhden kromosomin geenit (esimerkiksi isän - A, B, C) liittyvät toisen kromosomin geeneihin (äidin a, b , c), mikä johtaa uusiin geeniyhdistelmiin tuloksena olevissa kromatideissa. Tätä prosessia kutsutaan ylittämiseksi. Homologiset kromosomit eivät erotu risteytyksen jälkeen, koska sisarkromatidit (molempien kromosomien) pysyvät tiukasti yhteydessä anafaasiin asti. |
B. Ylittäminen profaasin aikana minä |
|
G . Homologisten kromosomien kromatidit hylkivät edelleen toisiaan, ja bivalentit saavat tietyn konfiguraation chiasmaattien lukumäärästä riippuen. Bivalentit, joissa on yksi chiasmata, ovat ristin muotoisia, kahdella chiasmatalla ne ovat renkaan muotoisia ja kolmella tai useammalla ne muodostavat kohtisuorassa toisiinsa nähden olevia silmukoita. Profaasin loppuun mennessä kaikki kromosomit ovat täysin tiivistyneet ja värjäytyneet voimakkaasti. Solussa tapahtuu muitakin muutoksia: sentriolien (jos sellaisia on) siirtyminen napoihin, nukleolien ja ydinkalvon tuhoutuminen ja sitten karan filamenttien muodostuminen. |
|
|
2. Metafaasi Bivalentit asettuvat ekvatoriaaliseen tasoon muodostaen metafaasilevyn. Niiden sentromeerit käyttäytyvät yksittäisinä rakenteina (vaikka ne näyttävät usein kaksinkertaisilta) ja järjestävät niihin kiinnittyneitä karafilamentteja, joista jokainen on suunnattu vain yhtä napaa kohti. Näiden lankojen heikon vetovoiman seurauksena kukin bivalentti sijaitsee päiväntasaajan alueella ja sen molemmat sentromeerit ovat samalla etäisyydellä siitä, toinen alapuolella ja toinen yläpuolella. |
D. Myöhäinen metafaasi minä |
|
3. Anafaasi Kummassakin bivalentissa olevat kaksi sentromeeriä eivät ole vielä jakautuneet, mutta sisarkromatidit eivät ole enää vierekkäin. Karan filamentit vetävät sentromeerit, joista jokainen liittyy kahteen kromatidiin, karan vastakkaisia napoja kohti. Tämän seurauksena kromosomit jakautuvat kahteen haploidiseen joukkoon, jotka päätyvät tytärsoluihin. |
E. Anafaasi I |
|
4. Telofaasi Homologisten sentromeerien ja niihin liittyvien kromatidien hajoaminen vastakkaisiin napoihin tarkoittaa ensimmäisen meioottisen jakautumisen päättymistä. Yhden sarjan kromosomien määrä on kasvanut puoleen, mutta kunkin navan kromosomit koostuvat kahdesta kromatidista. Chiasmatan muodostumisen aikana tapahtuvan risteytymisen vuoksi nämä kromatidit ovat geneettisesti epäidenttisiä, ja toisen meioottisen jakautumisen aikana ne eroavat. Karat ja niiden kierteet yleensä katoavat. Eläimissä ja joissakin kasveissa kromatidit hajoavat, niiden ympärille muodostuu ydinkalvo jokaiseen napaan, ja tuloksena oleva ydin siirtyy interfaasiin. Sitten alkaa sytoplasman jakautuminen (eläimillä) tai jakautuvan soluseinän muodostuminen (kasveissa), kuten mitoosissa. Monissa kasveissa ei havaita telofaasia, soluseinän muodostumista eikä interfaasia, ja solu siirtyy suoraan anafaasista I profaasiin II. |
G. Telofaasi I eläimen solussa |







|
Interfaasi II Tämä vaihe havaitaan yleensä vain eläinsoluissa: sen kesto vaihtelee. Vaihe S puuttuu, eikä DNA-replikaatiota tapahdu enempää. Meioosin toiseen jakautumiseen liittyvät prosessit ovat mekanismiltaan samanlaisia kuin mitoosissa tapahtuvat prosessit. Ne sisältävät kromatidien erottamisen molemmissa tytärsoluissa ensimmäisestä meioottisesta jakautumisesta. Meioosin toinen jakautuminen eroaa mitoosista pääasiassa kahdella tavalla: 1) metafaasissa II meioosi, sisarkromatidit ovat usein voimakkaasti erotettu toisistaan; 2) kromosomien lukumäärä on haploidinen. |
|
|
Profaasi II Soluissa, jotka menettävät interfaasin II , myös tämä vaihe puuttuu. Profaasin kesto II kääntäen verrannollinen telofaasin kestoon minä . Nukleolit ja tumakalvot tuhoutuvat ja kromatidit lyhenevät ja paksuuntuvat. Centriolit, jos niitä on, siirtyvät solujen vastakkaisiin napoihin; karafilamentteja tulee näkyviin. Kromatidit on järjestetty siten, että niiden pitkät akselit ovat kohtisuorassa ensimmäisen meioottisen jaon karan akseliin nähden. |
Z. Prophase P |
|
Metafaasi II Toisen jaon aikana sentromeerit käyttäytyvät kaksoisrakenteiden tavoin. Ne järjestävät karan kierteet molempia napoja kohti ja kohdistavat siten karan päiväntasaajalle. |
Metafaasi II |
|
Anafaasi II Keskusta Omeeri jakautuu ja karan kierteet vetävät ne vastakkaisille napoille. Sentromeerit vetävät pitkin erotettuja kromatideja, joita nyt kutsutaan kromosomeiksi. Telofaasi II Tämä vaihe on hyvin samanlainen kuin mitoosin telofaasi. Kromosomit hajoavat, venyvät ja on sitten vaikea erottaa toisistaan. Karan filamentit katoavat ja sentriolit replikoituvat. Jokaisen ytimen ympärille, joka sisältää nyt puolet alkuperäisen emosolun (haploidi) kromosomien lukumäärästä, muodostuu jälleen tumakalvo. Sytoplasman myöhemmän jakautumisen (eläimissä) tai soluseinän muodostumisen (kasveissa) seurauksena yhdestä alkuperäisestä emosolusta saadaan neljä tytärsolua. |
|


Erot mitoosin ja meioosin vaiheiden välillä
|
Vaihe |
Mitoosi |
Meioosi |
|
Prophase |
Kromomeerejä ei näy Homologiset kromosomit erotetaan toisistaan Chiasmataa ei muodostu Risteyksiä ei tapahdu |
Kromimeerit näkyvissä Homologiset kromosomit konjugoituvat. Chiasmatat muodostuvat. Ylitys voi tapahtua |
|
Metafaasi |
Kromatidiparit sijaitsevat karaekvaattorilla Sentromeerit ovat samassa tasossa karaekvaattorilla |
Kromatidiparit sijaitsevat karan ekvaattorilla vain meioosin toisessa jaossa Sentromeerit meioosin ensimmäisessä jaossa sijaitsevat päiväntasaajan ylä- ja alapuolella yhtä etäisyydellä siitä |
|
Anafaasi |
Sentromeerit jakautuvat. Kromatidit erottuvat. Erilaiset kromatidit ovat identtisiä |
Sentromeerit jakautuvat vasta meioosin toisessa vaiheessa. Kromatidit eroavat meioosin toisen jakautumisen aikana. Ensimmäisessä jaossa kokonaiset kromosomit erottuvat. Erilaiset kromosomit eivät välttämättä ole identtisiä risteytymisen seurauksena |
|
Telolofaasi |
Kromosomien lukumäärä tytärsoluissa on sama kuin emosoluissa Tytärsolut sisältävät molemmat homologiset kromosomit (diploideissa) |
Tytärsolujen kromosomien määrä on puolet emosoluista Tytärsolut sisältävät vain yhden jokaisesta homologisesta kromosomiparista |
|
Missä tällainen jakautuminen tapahtuu? |
Mahdollista haploidisissa, diploidisissa ja polyploidisissa soluissa Esiintyy somaattisten solujen ja joidenkin itiöiden muodostumisen aikana sekä sukusolujen muodostumisen aikana kasveissa, joissa tapahtuu sukupolvien vuorottelua |
Vain diploidisissa ja polyploidisissa soluissa Gameto- tai sporogeneesin aikana |
- Sukusolujen rakenne. Gametogeneesi
Seksuaalinen lisääntyminen tapahtuu käyttämällä erityisiä sukusoluja, joita kutsutaan sukusoluiksi. Naisten sukusoluja kutsutaan munat, uros siittiöt.Sukusolut eroavat somaattisista soluista ensisijaisesti sillä, että niissä on puolet vähemmän kromosomeja ja vähäisiä aineenvaihduntaprosesseja.
Munasolut suhteellisen suuret ei-liikkuvat solut, tavallisesti muodoltaan pyöreät; Tyypillisten organellien lisäksi sytoplasmassa on vararavinteiden sulkeumia keltuaisen muodossa ( riisi .). Munien ytimissä muodostuu monia kopioita ribosomaalisista geeneistä ja mRNA:sta, mikä varmistaa tulevan alkion elintärkeiden proteiinien synteesin. Eri organismien munat eroavat toisistaan keltuaisen määrän ja jakautumisen luonteen suhteen. Munatyyppejä on useita.

Isolesitaalinenkutsutaan suhteellisen pieniksi muniksi, joissa on pieni määrä tasaisesti jakautunutta keltuaista. Niiden ydin sijaitsee lähempänä keskustaa. Tällaisia munia löytyy matoista, simpukoista ja kotiloista, piikkinahkaisista ja lanseteista.Kohtalaisen telolesitaalinenSamen ja sammakkoeläinten munien halkaisija on noin 1,5 × 2 mm ja ne sisältävät keskimäärin keltuaisen, josta suurin osa on keskittynyt johonkin navoista (kasvillinen). Vastakkaisella navalla (eläin), jossa on vähän keltuaista, on munan ydin.
Voimakkaasti telolesitaalinenJoidenkin kalojen, matelijoiden, lintujen ja munasolujen nisäkkäiden munat sisältävät paljon keltuaista, jotka vievät melkein koko munan sytoplasman tilavuuden. Eläimen navassa on itulevy, jossa on aktiivinen sytoplasma, jossa ei ole keltuaista. Näiden munien koot ovat suuria, 10 15 mm tai enemmän.
Alecital munissa ei käytännössä ole keltuaista, ne ovat mikroskooppisesti pieniä (0,1 × 0,3 mm) ja ovat tyypillisiä istukan nisäkkäille, mukaan lukien ihmiset.
Sperma tai siittiöt ovat hyvin pieniä liikkuvia urospuolisia sukusoluja (esimerkiksi ihmisen siittiöt ovat 50 × 70 µm pitkiä ja krokotiili 20 µm), jotka muodostuvat miehen sukurauhasista - kiveksistä; niiden määrä on miljoonia. Siittiöiden muoto vaihtelee eri eläimillä, mutta niiden rakenne on sama, useimmilla niistä on pää ja kaula. Siittiön päässä on ydin, joka sisältää haploidisen määrän kromosomeja ja peitettynä acrosome. Akrosomi on erityinen rakenne, modifioitu Golgi-kompleksi, joka sisältää entsyymejä munasolun kalvon liuottamiseksi hedelmöityksen aikana, kalvon rajoittamana. Kaula sisältää useita mitokondrioita ja kaksi sentriolia. Kaulasta kasvaa mikrotubulusten muodostama häntä, joka varmistaa siittiöiden liikkuvuuden. Keskiosa on laajentunut sen sisältämien lukuisten mitokondrioiden ansiosta, jotka on koottu spiraaliksi siimakalvon ympärille. Nämä mitokondriot tarjoavat energiaa supistumismekanismeille, jotka tarjoavat siimalihaksen liikkeen. Siittiöiden pää- ja häntäosilla on siimaleille ominaista rakenne.

Jos katsot ihmisen siittiön päätä ylhäältä, se näyttää pyöreältä, ja sivulta katsottuna se näyttää litistyneeltä. Lippuliike ei yksin riitä siihen, että siittiö kulkee etäisyyttä emättimestä hedelmöityskohtaan. Siittiöiden pääasiallinen liikkumistehtävä on parvella munasolun ympärillä ja orientoitua tietyllä tavalla ennen munasolun kalvojen tunkeutumista.
Sukusolujen muodostuminen (gametogeneesi).Miesten ituepiteelin solut Ja Naisten sukurauhaset läpikäyvät sarjan peräkkäisiä mitoottisia ja meioottisia jakautumisia, joita kutsutaan yhteisesti gametogeneesiksi, mikä johtaa kypsien miessukusolujen muodostumiseen (spermatogeneesi) ja naissukusolujen muodostumiseen (oogeneesi). Molemmissa tapauksissa prosessi on jaettu kolmeen vaiheeseen - lisääntymisvaiheeseen, kasvuvaiheeseen ja kypsymisvaiheeseen. Lisääntymisvaihe sisältää toistuvia mitoottisia jakautumisia, jotka johtavat monien siittiöiden tai oogonioiden muodostumiseen. Jokainen niistä käy läpi kasvujakson valmistautuessaan ensimmäiseen meioottiseen jakautumiseen ja sitä seuraavaan sytokineesiin. Sitten alkaa kypsymisvaihe, jonka aikana tapahtuu ensimmäinen ja toinen meioottinen jakautuminen Kanssa myöhempi haploidisten solujen erilaistuminen Ja kypsien sukusolujen muodostuminen.
Siittiöiden kehitys (spermatogeneesi).Siittiöt (spermiini) muodostuvat sarjan peräkkäisten solujen jakautumisen seurauksena, joita kutsutaan yhteisestispermatogeneesi,jota seuraa monimutkainen erilaistumisprosessi nimeltään spermiogeneesi (kuva 20.31 ). Siittiöiden muodostumisprosessi kestää noin 70 päivää; 1 g kivespainoa kohti muodostuu 10 7 spermaa päivässä. Siementiehyen epiteeli koostuu solujen ulkokerroksestaituepiteelija noin kuusi solukerrosta, jotka muodostuivat tämän kerroksen toistuvien solujakautumisten seurauksena (kuvat 20.32 ja 20.33); nämä kerrokset vastaavat peräkkäisiä siittiöiden kehitysvaiheita. Ensinnäkin ituepiteelisolujen jakautuminen aiheuttaa lukuisia spermatogonia jotka kasvavat ja muuttuvatensimmäisen asteen spermatosyytit.Ensimmäisen meioottisen jakautumisen seurauksena nämä spermatosyytit muodostavat haploidejatoisen asteen spermatosyytit,jonka jälkeen ne läpikäyvät toisen meioottisen jakautumisen ja muuttuvat siittiöiksi. Kehittyvien solujen "säikeiden" välissä on suuria Sertoli-solut tai troofiset solut,sijaitsee koko tilassa tubuluksen ulkokerroksesta sen luumeniin.

Spermatosyytit sijaitsevat lukuisissa invaginaatioissa Sertoli-solujen sivupinnoilla; täällä ne muuttuvat siittiöiksi ja siirtyvät sitten siihen Sertoli-solun reunaan, joka on siementiehyen luumenia päin, missä ne kypsyvät muodostaen spermiini (kuva 20.33 ). Ilmeisesti Sertoli-solut tarjoavat mekaanista tukea, suojaa ja ravintoa kypsyville siittiöille. Kaikki siementiehyitä ympäröivien verisuonten kautta kehittyville sukusoluille toimitetut ravintoaineet ja happi sekä vereen vapautuneet aineenvaihduntajätteet kulkevat Sertoli-solujen läpi. Nämä solut erittävät myös nestettä, jonka kanssa siittiöt kulkevat tubulusten läpi.
Munan kehitys ihmisillä (oogeneesi).Toisin kuin siittiöiden muodostuminen, joka alkaa miehillä vasta murrosiässä, munasolujen muodostuminen alkaa naisilla jo ennen heidän syntymäänsä ja päättyy kullekin munalle vasta sen hedelmöittymisen jälkeen. Oogeneesin vaiheet on esitetty kuvassa. 20.36. Sikiön kehityksen aikana alkuperäiset sukusolut jakautuvat toistuvasti mitoosin kautta tuottaen monia suuria soluja ns. oogonia. Oogonia käy taas läpi mitoosin ja muodostuuensimmäisen asteen munasolut,jotka pysyvät profaasivaiheessa melkein ovulaation asti. Ensimmäisen asteen munasoluja ympäröi yksi kerros soluja-granulosakalvo-ja muodostavat nsprimordiaaliset follikkelit.Naarassikiö juuri ennen syntymää sisältää noin 2 10 6 nämä munarakkulat, mutta vain noin 450 niistä saavuttaa toisen asteen munasolut ja poistuu munasarjasta (ovulaatio). Ennen ovulaatiota ensimmäisen asteen munasolu käy läpi ensimmäisen meioottisen jakautumisen muodostaen haploidintoisen asteen munasolu Ja ensimmäinen napakappale.Toinen meioottinen jakautuminen saavuttaa metafaasivaiheen, mutta ei jatku ennen kuin munasolu fuusioituu siittiöiden kanssa. Hedelmöityksen aikana toisen asteen munasolussa tapahtuu toinen meioottinen jakautuminen, jolloin muodostuu suuri solu - muna ja myös toinen napakappale.Kaikki napakappaleet ovat pieniä soluja; niillä ei ole merkitystä oogeneesissä ja ne tuhoutuvat lopulta.

partenogeneesi
velvoittaa
valinnainen
Organismien kehitys vain hedelmöittämättömästä munasta.
Vain naaraat muodostuvat.
Esimerkiksi valkoihoinen kivilisko
letlet voivat kehittyä sekä ilman lannoitusta että lannoituksen kanssa
Nainen tyyppi
Urokset kehittyvät hedelmöittämättömistä munista
Mies tyyppi
Joissakin isogamisissa levissä
Muita vastaavia teoksia, jotka saattavat kiinnostaa sinua.vshm> |
|||
| 6644. | Organismien yksilöllinen kehitys (ontogeneesi) | 78,69 kt | |
| Uskotaan, että vain yhden siittiön ydinsisältö tunkeutuu eläinten munaan. Ihmisten ja joskus korkeampien eläinten tapauksessa syntymää edeltävää kehitysjaksoa kutsutaan usein syntymää edeltäväksi syntymän jälkeen ja postnataaliseksi. Useimmissa monisoluisissa eläimissä, riippumatta niiden organisaation monimutkaisuudesta, alkion kehitysvaiheet, jotka alkio käy läpi, ovat samat. Murskaantumisen luonne ja blastulatyypit eri selkärankaisissa... | |||
| 13714. | Organismien yksilöllinen kehitys ja käyttäytyminen. Ontogeneesi. Kasvien ja eläinten elinkaari | 9,96 kt | |
| Ontogeneesi persoonallisuuden alkuperä on organismin yksilöllinen kehitys tsygootin muodostumishetkestä sen kuolemaan. Sukusolut ovat sukupuolisoluja, jotka kantavat perinnöllistä tietoa ja joilla on haploidi joukko. Hedelmöitys on kasvien tai eläinten uros- ja naaraspuolisten sukusolujen fuusioprosessi ja se on seksuaalisen prosessin perusta. Alkion itämisjakso on ontogeneesin ajanjakso tsygootin muodostumisesta syntymään tai munakalvoista ilmestymiseen tai itämiseen. | |||
| 10427. | Jäljentäminen | 6,75 kt | |
| Kun organismien lisääntymismuotoja on valtavasti, ne voidaan pelkistää kahteen päätyyppiin: aseksuaalisiin ja seksuaalisiin. Aseksuaalisessa lisääntymisessä jälkeläisten lisääntyminen tapahtuu toisesta vanhemmasta itiöiden muodostumisen kautta tai kasvullisesti. Kasvillisen lisääntymisen aikana jälkeläisiä syntyy emosta erotetuista ruumiinosista. Kasvillisen lisääntymisen aikana kasvit säilyttävät heterotsygoottisuuden useiden sukupolvien ajan. | |||
| 21332. | Rajoittavat tekijät. Organismien sopeutuminen tekijöihin | 303,8 kt | |
| Jotkut eläimet rakastavat voimakasta lämpöä, toiset sietävät paremmin maltillisia ympäristölämpötiloja jne. Lisäksi elävät organismit jaetaan sellaisiin, jotka voivat esiintyä laajalla tai kapealla vaihtelualueella missä tahansa ympäristötekijässä. Jos ympäristöolosuhteiden vaikutus ei saavuta ääriarvoja, elävät organismit reagoivat siihen tietyillä toimilla tai tilan muutoksilla, mikä lopulta johtaa lajin selviytymiseen. Tämän työn tutkimuskohteena ovat ympäristötekijät, aiheena rajoittavat tekijät ja organismien sopeutuminen... | |||
| 8875. | Organismien ja ympäristötekijöiden vuorovaikutuksen yleiset mallit | 193,58 kt | |
| Maaperän maa- ja vesiympäristön bioottiset tekijät Elävien organismien biologisesti aktiiviset aineet Antropogeeniset tekijät Organismien ja ympäristötekijöiden vuorovaikutuksen yleiset mallit Rajoittavan tekijän käsite. Liebigin minimilaki Shelfordin laki Antropogeenisten tekijöiden vaikutuksen erityispiirteet organismiin Organismien luokittelu ympäristötekijöiden suhteen 1. Höyhenruohoarojen olosuhteet edustavat täysin erilaisia abioottisten tekijöiden järjestelmiä. | |||
| 12700. | Tuholaisten biologiset ominaisuudet ja toimenpiteet niiden torjumiseksi | 62,79 kt | |
| Erityisen merkittäviä satohäviöitä aiheutuu rikkaruohojen esiintymisestä, jotka poistavat ravinteita ja kosteutta maaperästä, varjostavat viljelykasveja ja monissa tapauksissa saastuttavat tuotteita myrkyllisillä aineilla ja siemenillä, jotka aiheuttavat ihmisten ja eläinten myrkytyksen. Maatalouden kemialisoinnin pääsuunnat: lannoitteiden, kemikaalien käyttö, kasvinsuojelu tuholaisilta, taudeilta ja rikkaruohoilta, kemiallisten tuotteiden käyttö karjassa, maataloustuotteiden purku ja... | |||
| 13403. | Virukset soluttomina elämänmuotoina. Viruksen rakenne, luokitus, vuorovaikutus eri organismien solujen kanssa | 12,75 kt | |
| Nämä ovat biologisia esineitä, genomeja, jotka koostuvat nukleiinihapoista DNA:sta tai RNA:sta, joita tuotetaan elävissä soluissa niiden biosynteettisellä laitteistolla. Virusten ja muiden elämänmuotojen väliset erot: niillä ei ole solurakennetta; 1-tyyppisiä nukleiinihappoja; vain DNA tai RNA; niillä ei ole omaa aineenvaihduntaa. Hypoteesi virusten alkuperästä: virukset syntyivät normaalin solun komponenteista, jotka pakenivat säätelymekanismien hallinnasta ja muuttuivat itsenäiseksi yksiköksi; luultavasti DNA-osassa tapahtui sarja geneettisiä muutoksia, jotka... | |||
| 18798. | Maaperän mesofauna-organismien rakenteelliset ja toiminnalliset indikaattorit Pozim-joen laakson metsissä ja avoimissa biotoopeissa | 61,54 kt | |
| Nykyään tämän alueen kehitys on kuitenkin seurausta useista ympäristöongelmista. Varsinkin laaksossa tehdään laajasti töitä tulvaniityjen kuivattamiseksi, mikä vaikuttaa suoraan luonnollisiin biotooppeihin. | |||
| 19386. | Liberalismin kehitys IVY:ssä | 35,55 kt | |
| Ulkomaan taloudellisten suhteiden kehitysprosesseissa tarkasteltavien muutosten monimutkainen järjestelmäluonne määrää perinteisten kansantalouden identifiointia, rajoja ja suvereniteettia koskevien käsitysten koko aksiomatian tarkistamisen ja muuttamisen... | |||
| 8867. | KOULUTUS JA KEHITYS | 162,32 kt | |
| Koulutuksen ja kehityksen välisen suhteen ongelman ydin. Peruslähestymistapoja koulutuksen ja kehityksen välisen suhteen ongelman ratkaisemiseen. Proksimaalisen kehityksen vyöhykkeen käsite L. Harjoittelun ja kehityksen välisen suhteen ongelman ydin. | |||
Lisääntyminen on eliöiden kykyä tuottaa jälkeläisiä tai organismien kykyä lisääntyä. Elävien olentojen tärkein ominaisuus lisääntyminen varmistaa elämän jatkuvuuden ja lajien jatkuvuuden.
Lisääntymisprosessi on erittäin monimutkainen, ja se ei liity pelkästään geneettisen tiedon siirtämiseen vanhemmilta jälkeläisille, vaan myös organismien anatomisiin ja fysiologisiin ominaisuuksiin, niiden käyttäytymiseen ja hormonaaliseen hallintaan. Organismien lisääntymiseen liittyy niiden kasvu- ja kehitysprosessit.
Eläville olennoille on ominaista lisääntymismenetelmien äärimmäinen monimuotoisuus. Siitä huolimatta on olemassa kaksi pääasiallista lisääntymismenetelmää - aseksuaalinen ja seksuaalinen (kuva 16). Aseksuaalinen lisääntyminen tai apomixis (kreikasta. aro - ilman, mixis - sekoitus) on prosessi, jossa vain yksi vanhempi (solu tai monisoluinen organismi) osallistuu. Sitä vastoin seksuaalisessa lisääntymisessä on kaksi vanhempaa, joilla kummallakin on oma lisääntymisjärjestelmä ja jotka tuottavat sukusoluja (sukusoluja), jotka fuusion jälkeen muodostavat tsygootin (hedelmöitetty muna), joka sitten erilaistuu alkioksi. Näin ollen seksuaalisen lisääntymisen aikana on sekoitus perinnöllisiä tekijöitä, eli prosessi, jota kutsutaan amfimiksiksi (kreikasta. amfi - molemmin puolin, mixis - sekoitus).
Suvuton lisääntyminen
Aseksuaalinen lisääntyminen on ominaista monille lajeille, sekä kasveille että eläimille. Sitä löytyy viruksista, bakteereista, levistä, sienistä, vaskulaarisista kasveista, alkueläimistä, sienistä, coelenteraateista, sammaleläimistä ja vaippaeläimistä.
Yksinkertaisin suvuttoman lisääntymisen muoto on ominaista viruksille. Niiden lisääntymisprosessi liittyy nukleiinihappomolekyyleihin ja näiden molekyylien kykyyn kopioida itseään
ja perustuu suhteellisen heikkojen vetysidosten spesifisyyteen nukleotidien välillä.
Riisi. 16.Eliöiden lisääntymismenetelmät
Suhteessa muihin suvuttomasti lisääntyviin organismeihin tehdään ero vegetatiivisen lisääntymisen ja itiöinnin kautta lisääntymisen välillä.
Vegetatiivinen lisääntyminen on lisääntymistä, jossa uusi eliö kehittyy äidin organismista erotetusta osasta. Tämän tyyppinen lisääntyminen on ominaista sekä yksisoluisille että monisoluisille organismeille, mutta sillä on erilaisia ilmenemismuotoja.
Yksisoluisissa organismeissa vegetatiivista lisääntymistä edustavat sellaiset muodot kuin jakautuminen, moninkertainen fissio ja orastuminen. Jakautuminen yksinkertaisella supistumisella kahden tytärorganismin muodostumisella yhdestä emoorganismista on ominaista bakteereille ja sinileville (sinileville). Päinvastoin, lisääntyminen ruskeiden ja vihreiden levien sekä yksisoluisten eläinten (sarkoodien, siimalevien ja värpästen) jakautumisen kautta tapahtuu ytimen mitoottisen jakautumisen kautta, jota seuraa sytoplasman supistuminen.
Lisääntyminen moninkertaisella fissiolla (skitsogonia) sisältää ytimen jakautumisen, jota seuraa sytoplasman jakaminen osiin. Tämän jakautumisen seurauksena yhdestä solusta muodostuu useita tytärorganismeja. Esimerkki moninkertaisesta jakautumisesta on falciparum plasmodiumin lisääntyminen (P. vivax) ihmisen punasoluissa. Tässä tapauksessa plasmodiassa toistuva tuman jakautuminen tapahtuu monta kertaa ilman sytokineesia, jota seuraa sytokineesi. Tämän seurauksena yksi plasmodium synnyttää 12-24 tytärorganismia.
Monisoluisissa kasviorganismeissa vegetatiivinen lisääntyminen jakautumalla tapahtuu pistokkaiden, sipulien, lehtien ja juurakoiden avulla. Mutta tämä on pohjimmiltaan keinotekoista lisäystä, jota käytetään maatalouskäytännössä. Korkeampien kasvien lisääntyminen keinotekoisissa olosuhteissa on myös mahdollista yhdestä solusta. Organismeilla, jotka kehittyvät yhdestä solusta (kloonista), on kaikki alkuperäisen monisoluisen organismin ominaisuudet. Tätä lisääntymistä kutsutaan klonaaliseksi mikrolisäykseksi. Yksi vegetatiivisen lisäämisen muodoista voi olla useiden viljelykasvien varttaminen tai istuttaminen, joka koostuu silmun tai verson osan siirtämisestä kasvista toiseen. Tietenkin tämä on myös lisääntymismenetelmä, jota ei esiinny luonnossa, mutta jota käytetään hyvin laajasti maataloudessa.
Monisoluisissa eläimissä vegetatiivinen lisääntyminen tapahtuu hajottamalla niiden ruumis osiin, minkä jälkeen jokainen
osasta kehittyy uusi eläin. Tällainen lisääntyminen on tyypillistä sienille, coelenteraateille (hydraille), neerteaneille, lattamatoille, piikkinahkaisille (meritähtille) ja joillekin muille organismeille. Eläinten vegetatiiviselle lisääntymiselle läheinen pirstoutumisen muoto on eläinten polyembryonia, joka koostuu siitä, että tietyssä kehitysvaiheessa alkio on jaettu useisiin osiin, joista jokainen kehittyy itsenäiseksi organismiksi. Polyembryonia esiintyy armadilloissa. Jälkimmäiset kuitenkin lisääntyvät seksuaalisesti. Siksi polyembryonia on melko ainutlaatuinen vaihe seksuaalisessa lisääntymisessä, ja polyembryoniasta syntyneitä jälkeläisiä edustavat yksitsygoottiset kaksoset.
Orastuminen koostuu tuberkuloosin (uloskasvun) muodostumisesta emosolun ytimellä, joka sitten erottuu ja siitä tulee itsenäinen organismi. Orastumista esiintyy sekä yksisoluisissa kasveissa, esimerkiksi hiivassa, että yksisoluisissa eläimissä, esimerkiksi tietyissä väreissä.
Lisääntyminen itiöinnin avulla liittyy erikoistuneiden solujen muodostumiseen - itiöt, jotka sisältävät ytimen, sytoplasman, on peitetty tiheällä kalvolla ja kykenevät pitkäaikaiseen olemassaoloon epäsuotuisissa olosuhteissa, mikä lisäksi edistää niiden leviämistä. Useimmiten tällainen lisääntyminen tapahtuu bakteereissa, levissä, sienissä, sammalissa ja saniaisissa. Joissakin viherlevissä yksittäisistä soluista voi muodostua niin sanottuja zoosporeja.
Eläimillä lisääntymistä itiöinnin kautta havaitaan itiöeläimillä, erityisesti falciparum-plasmodiumilla.
Monissa lajeissa suvuton lisääntyminen voi vaihdella sukupuolisen lisääntymisen kanssa.
SUKUPUOLINEN LISÄÄNTYMINEN
Sukupuolista lisääntymistä tapahtuu sekä yksi- että monisoluisissa kasveissa ja eläimissä.
Kuten luvuissa V ja XIII mainitaan, sukupuolinen lisääntyminen bakteereissa tapahtuu konjugaatiolla, joka toimii sukupuoliprosessin analogina ja on näiden organismien rekombinaatiojärjestelmä, kun taas alkueläimissä sukupuolinen lisääntyminen tapahtuu myös konjugaation tai syngamian kautta. ja autogamiaa.
Monisoluisissa organismeissa (kasveissa ja eläimissä) sukupuolinen lisääntyminen liittyy suku- tai sukusolujen (sukusolujen) muodostumiseen, hedelmöittymiseen ja tsygoottien muodostumiseen.
Sukupuolinen lisääntyminen on merkittävä organismien evoluutiohankinta. Toisaalta se edistää geenien uudelleenlajittelua, organismien monimuotoisuuden syntymistä ja kilpailukyvyn lisäämistä jatkuvasti muuttuvissa ympäristöolosuhteissa.
Yksisoluisissa organismeissa sukupuolinen lisääntyminen tapahtuu useissa muodoissa. Bakteereissa sukupuolinen lisääntyminen voidaan analogistaa niissä tapahtuvan konjugaation kanssa, joka koostuu plasmidin tai kromosomaalisen DNA:n siirtämisestä luovuttajasoluista (sisältää plasmideja) vastaanottajasoluihin (jotka eivät sisällä plasmideja) sekä bakteerien transduktion kanssa. , joka koostuu geneettisen materiaalin siirrosta joistakin bakteerisoluista muihin faageihin. Konjugaatiota löytyy myös väreistä, joissa tämän prosessin aikana tapahtuu ytimien siirtymistä yksilöstä toiseen, minkä jälkeen jälkimmäinen jakautuu.
Monisoluisissa kasveissa ja eläimissä solujen sukupuolinen lisääntyminen tapahtuu naaras- ja miessukusolujen (munien ja siittiöiden) muodostumisen kautta, hedelmöittämällä munasolu siittiöllä ja muodostamalla tsygootti. Kasveissa sukupuolisoluja tuotetaan erityisissä lisääntymisrakenteissa; eläimissä niitä tuotetaan sukurauhasissa, joita kutsutaan sukurauhasiksi (kreikasta. mennyt - siemen).
Eläinten somaattisten ja lisääntymissolujen välillä on tärkeä ero. Se johtuu siitä, että somaattiset solut pystyvät jakautumaan, eli ne lisääntyvät itseään ja lisäksi niistä muodostuu sukusoluja. Päinvastoin, sukupuolisolut eivät jakautu, vaan ne "aloittaa" koko organismin lisääntymisen.
Diploidisia somaattisia soluja, joissa miehen sukusoluja muodostuu, kutsutaan spermatogoniaksi ja joissa naissukusoluja muodostuu - oogoniaksi. Miesten ja naisten sukusolujen muodostumisprosessia (kasvua ja erilaistumista) kutsutaan gametogeneesiksi.
Gametogeneesi perustuu meioosiin (kreikasta. meioosi - vähentää), joka on soluytimien pelkistysjakauma, johon liittyy kromosomien lukumäärän väheneminen tumaa kohti. Meioosia esiintyy seksuaalisesti lisääntyvien elävien olentojen sukuelinten erikoistuneissa soluissa (kuva 17). Esimerkiksi pteridofyyteissä meioosia esiintyy erikoistuneissa sporangisoluissa, jotka sijaitsevat näiden kasvien lehtien alapinnalla ja kehittyvät itiöiksi ja sitten gametofyyteiksi. Jälkimmäiset ovat olemassa erikseen ja tuottavat lopulta uros- ja naarassukusoluja. Kukkivissa kasveissa meioosia esiintyy munasolujen erikoistuneissa soluissa, jotka kehittyvät itiöiksi. Jälkimmäiset tuottavat gametofyytin yhdellä munalla. Lisäksi näissä kasveissa meioosia esiintyy myös erikoistuneissa ponnesoluissa, jotka myös kehittyvät itiöiksi, jotka lopulta tuottavat siitepölyä kahden urossukusolun kanssa. Kastematoissa, jotka ovat hermafrodiitteja ja sisältävät urospuolisia sukuelimiä yhdessä ja naaraspuolisia toisessa kehon osassa ja joille on ominaista
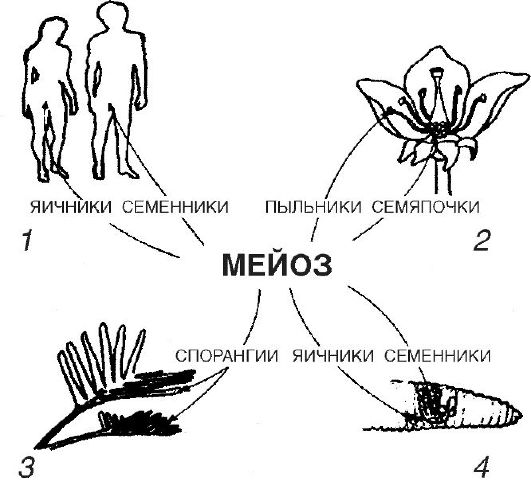
Riisi. 17. Meioosi eri organismeissa: 1 - Ihmisen; 2 - kukkivat kasvit; 3 - saniainen; 4 - kastemato
Heillä on kyky hedelmöittää eri yksilöiden välillä; heillä on kyky suorittaa samanaikaisesti spermatogeneesiä ja oogeneesia.
Meioosia esiintyy kivesten ja munasarjojen erikoistuneissa soluissa, jotka tuottavat vastaavasti uros- ja naarassukusoluja. Proteiineja, jotka ovat meioosin indusoijia, on tunnistettu.
Meioosin aikana somaattisille soluille (soluytimille) ja epäkypsille soluiduille tyypillinen kromosomien diploidiluku (2n) muuttuu kypsille sukusoluille ominaiseksi haploidilukumääräksi (n). Siten sukusolut saavat gametogeneesin seurauksena vain puolet somaattisten solujen kromosomeista (kuvio 18).

Riisi. 18.
Kromosomien käyttäytyminen gametogeneesin aikana eläimillä on sama sekä miehillä että naarailla. Sukupuolet eroavat kuitenkin meioosin eri vaiheiden syntyajasta, mikä on erityisen tärkeää
erityisesti ihmisillä havaittavissa. Postpubertaalisilla miehillä meioosin koko prosessi päättyy noin kahdessa kuukaudessa, kun taas naisilla ensimmäinen meioottinen jakautuminen alkaa sikiön munasarjassa ja päättyy vasta ovulaation alkaessa, joka tapahtuu noin 15 vuoden iässä.
Korkeammissa eläimissä, urosten tapauksessa, meioosiin liittyy neljän toiminnallisesti aktiivisen sukusolun muodostuminen.
Päinvastoin, naarailla jokainen toisen asteen munasolu tuottaa vain yhden munan. Naarasmeioosin muut ydintuotteet ovat kolme pelkistyskappaletta, jotka eivät osallistu lisääntymiseen ja rappeutuvat.
Meioosi koostuu kahdesta ydinjaosta. Ensimmäinen meioottinen ydinjako erottaa kunkin homologisen kromosomiparin jäsenet sen jälkeen, kun ne ovat pariutuneet keskenään (synapsis) ja vaihtaneet geneettistä materiaalia (risteytyminen). Tämän jakautumisen seurauksena muodostuu kaksi haploidista ydintä. Toinen meioottinen jako erottaa kromosomien (kromatidit) kaksi pitkittäistä puoliskoa kussakin näistä ytimistä ja tuottaa neljä haploidista ydintä.
Gametogeneesin aikana tapahtuu myös munien (ovogeneesi) ja siittiöiden (spermatogeneesi) erilaistumista, mikä on niiden toiminnan edellytys. Eläinten munat ovat paljon suurempia kuin siittiö, ovat yleensä liikkumattomia ja sisältävät ravintoaineita, jotka takaavat alkion kehityksen alkuvaiheessa hedelmöityksen jälkeen. Useimpien eläinten siittiöissä on siima, joka varmistaa niiden liikkumattomuuden.
Meioosilla on erinomainen biologinen merkitys. Meioosin ansiosta organismien soluissa säilyy jatkuva määrä kromosomeja sukupolvien lukumäärästä riippumatta. Siksi meioosi ylläpitää lajien pysyvyyttä. Lopuksi, meioosissa risteytyksen seurauksena tapahtuu geenien rekombinaatio, joka on yksi evoluution tekijöistä.
SPERMATOGENEESI JA OVOGENEESI
Spermatogeneesi on kypsien miehen sukusolujen muodostumisprosessi. Siittiöt kehittyvät miehen sukurauhasissa (kiveksissä tai kiveksissä) erikoistuneista somaattisista soluista (kuva 19). Tällaiset erikoistuneet solut toimivat seuraavasti:
joita kutsutaan primordiaalisiksi sukusoluiksi, jotka siirtyvät kiveksiin uroshenkilön alkion alkuvaiheessa. Tästä johtuen primordiaalisolut ovat kypsien sukusolujen esiasteita (prekursoreita).
Ihmisen kivekset koostuvat lukuisista tubuluksista, joiden seinämät muodostuvat siittiöiden eri vaiheissa olevista solukerroksista. Tubulusten ulkokerroksen muodostavat suuret solut, joita kutsutaan spermatogoniaksi. Nämä solut sisältävät diploidisen joukon kromosomeja ja ovat kiveksessä olevien alkuperäisten sukusolujen jälkeläisiä. Yksilön murrosiän aikana osa siittiösoluista siirtyy tubulusten sisäkerrokseen, jossa ne kehittyvät meioosin seurauksena soluiksi, joita kutsutaan ensimmäisen asteen spermatosyyteiksi (spermatosyytit I), sitten toisen asteen spermatosyyteiksi. (spermatosyytit II) ja lopuksi

Riisi. 19.Spermatogeneesi ja oogeneesi
spermatideiksi, jotka ovat haploideja sukusoluja, jotka lopulta erilaistuvat kypsiksi siittiöiksi. Yleisesti voidaan siis sanoa, että spermatogeneesi alkaa diploidisissa somaattisissa soluissa (spermatogonia), jota seuraa sukusolujen kypsymisjakso, jossa kaksi tuman jakautumista tapahtuu meioosin kautta, mikä johtaa siittiöiden muodostumiseen. Tämä on kuva henkilöstä.
Meioosi (kuva 20) spermatogeneesissä tapahtuu useissa vaiheissa (vaiheissa). Jakojen välillä on kaksi välivaihetta. Siten meioottinen jakautuminen voidaan esittää sarjana tapahtumia, jotka seuraavat peräkkäin, nimittäin: interfaasi I - ensimmäinen meioottinen jakautuminen (varhainen profaasi I, myöhäinen profaasi I, metafaasi I, anafaasi I, telofaasi I) - interfaasi II (interokinesis) - toinen meioottinen jakautuminen (profaasi I, metafaasi II, anafaasi II, telofaasi II). Meioosiprosessi on erittäin dynaaminen, joten mikroskooppiset erot eri vaiheiden välillä eivät heijasta itse vaiheiden luonnetta, vaan pikemminkin eri vaiheiden kromosomien ominaisuuksia. Interfaasi I

Riisi. 20.Meioosin vaiheet: 1 - ensimmäinen meioottinen jakautuminen; 2 - toinen meioottinen jakautuminen
jolle on tunnusomaista se, että siinä tapahtuu kromosomin replikaatio (DNA:n kaksinkertaistuminen), mikä on lähes täysin valmis varhaisen I-profaasin alkuun mennessä.
Ensimmäinen meioottinen jakautuminen alkaa primaarisesta siittiösolusta ja sille on tunnusomaista pitkä profaasi, joka koostuu profaasi I ja profaasi II siirtymisestä toisiinsa. Profaasissa I on viisi päävaihetta - leptonema, zygonema, pakyneema, diploneema ja diakineesi.
Leptoneema-vaiheessa ytimessä olevat kromosomit esitetään ohuina spiraalisäikeinä, jotka sisältävät lukuisia tummia rakeita (kromomeerejä). Kromomeerien ja lankojen halkeamista ei ole havaittu, mutta uskotaan, että kromosomit ovat tässä vaiheessa kaksinkertaisia eli diploideja. Kunkin kromosomiparin homologeja yhdistävät kromomeerit pituudeltaan kiinnitysperiaatteen mukaisesti.
Tsygonema-vaiheelle on ominaista synapsien muodostuminen homologisten kromosomien välille, mikä johtaa parillisten kromosomien (kaksiarvoisten) muodostumiseen. Kromosomit X ja Y käyttäytyvät hieman eri tavalla kuin autosomit. Ne tiivistyvät tummanvärisiksi heterokromaattisiksi kappaleiksi, jotka muodostavat parin päissä olevien homologisten alueiden seurauksena.
Pakyneemavaiheessa, joka on meioottisen profaasin pisin vaihe, tapahtuu bivalenttien kondensaatiota ja kunkin kromatidin jakautumista kahdeksi, minkä seurauksena jokainen bivalentti on monimutkainen kierukkarakenne, joka koostuu neljästä sisarkromatidista (tetradista). Tämän vaiheen lopussa parillisten kaksiarvoisten kromosomien erottaminen alkaa. Nyt homologisia kromosomeja voidaan havaita vierekkäin. Siksi joissakin valmisteissa voidaan nähdä neljä kromosomia, jotka muodostuvat kunkin homologin päällekkäisyyden seurauksena muodostaen sisarkromatideja. Tässä vaiheessa tapahtuu vaihtoa homologien välillä ja muodostuu chiasmata.
Diplonema-vaiheessa tapahtuu sisarkromatidien lyhenemistä, paksuuntumista ja keskinäistä hylkimistä, minkä seurauksena kaksiarvoisen kromatidit ovat melkein eronneet. Erottamista pidetään epätäydellisenä, koska sentromeeriä ei ole vielä jaettu jokaiseen kromosomipariin. Mitä tulee bivalentteihin, chiasmatat pitävät niitä eri paikoissa pitkin pituutta, jotka ovat homologisten kromatidien väliin muodostuneita rakenteita.
edellinen ylitys synaptisesti yhdistettyjen homologien välillä. Hyvissä preparaatioissa voidaan havaita yhdestä useaan chiasmataa bivalentin pituudesta riippuen. Jokainen tässä vaiheessa havaittu chiasma edustaa tulosta vaihdosta, joka tapahtui ei-sisarkromatidien välillä pakyneemavaiheen aikana. Kun bivalenttien puristus ja hylkiminen voimistuvat, chiasmata siirtyy kohti kromosomien päitä, eli tapahtuu kromosomien terminaalistumista. Diploneman lopussa tapahtuu kromosomien despiralisaatiota; homologit hylkivät edelleen toisiaan.
Diakineesin vaiheessa, joka on samanlainen kuin diplonema, bivalenttien lyheneminen jatkuu ja chiasmata heikkenee (vähenee), minkä seurauksena muodostuu erillisiä yksiköitä kromatidien muodossa (neljä). Välittömästi tämän vaiheen päätyttyä tapahtuu ydinkalvon liukeneminen.
Metafaasissa I kaksiarvoiset aineet saavuttavat suurimman pitoisuutensa. Ovaalin muotoisina ne sijaitsevat ytimen ekvatoriaalisessa osassa, jossa ne muodostavat meioottisen metafaasin I ekvatoriaaliset levyt. Jokaisen bivalentin muodon määrää chiasmatan lukumäärä ja sijainti. Miehillä chiasmaattien määrä bivalenttia kohden metafaasissa I on yleensä 1-5. XY-kaksiarvoisesta tulee sauvan muotoinen yhden terminaalissa olevan kiasmin seurauksena.
Anafaasissa I vastakkaisten sentromeerien liike alkaa solun vastakkaisille napoille. Tämän seurauksena homologiset kromosomit erotetaan. Jokainen kromosomi koostuu nyt kahdesta kromatidista, joita pitää yhdessä sentromeeri, joka ei jakautu ja pysyy ehjänä. Tämä erottaa meioosin anafaasin I mitoosin anafaasista, jossa sentromeeri jakautuu. On tärkeää huomata, että risteytys tekee jokaisesta kromatidista geneettisesti erilaisia.
Telofaasi I -vaiheessa kromosomit saavuttavat navat, mikä päättää ensimmäisen meioottisen jakautumisen. Telofaasin I jälkeen tapahtuu lyhyt välivaihe (interkineesi), jossa kromosomit hajoavat ja muuttuvat diffuusiksi, tai telofaasi I siirtyy suoraan toisen meioottisen jakautumisen profaasiin II. Kummassakaan tapauksessa DNA:n replikaatiota ei havaita. Ensimmäisen meioottisen jakautumisen jälkeen soluja kutsutaan toisen asteen spermatosyyteiksi. Kromosomien lukumäärä kussakin tällaisessa solussa vähenee 2n 1n, mutta DNA-sisältö ei vielä muutu.
Toinen meioottinen jakautuminen tapahtuu useissa vaiheissa (profaasi II, metafaasi II, anafaasi II, telofaasi II) ja on samanlainen kuin mitoottinen jakautuminen. Profaasissa II sekundaaristen spermatosyyttien kromosomit jäävät napoihin. Metafaasissa II kunkin kaksoiskromosomin sentromeeri jakautuu, jolloin jokaiselle uudelle kromosomille on oma sentromeeri. Alkaa karan muodostuminen, jonka napaa kohti uudet kromosomit liikkuvat. Telofaasissa II toinen meioottinen jakautuminen päättyy, minkä seurauksena jokainen toisen asteen siittiösolu tuottaa kaksi siittiötä, joista siittiöt sitten erilaistuvat. Kuten sekundaarisessa spermatosyytissä, kromosomien lukumäärä siittiössä on haploidinen (1n). Spermatidien kromosomit ovat kuitenkin yksittäisiä, kun taas sekundaaristen spermatosyyttien II kromosomit ovat kaksinkertaisia, ja ne on rakennettu kahdesta kromatidista. Näin ollen kunkin spermatidin ytimessä on yksi sarja ei-homologisia kromosomeja. Toissijainen meioottinen jakautuminen on mitoottista tyyppiä oleva jakautuminen (ekvatoriaalinen jakautuminen). Se erottaa kaksoissisarkromatidit ja eroaa pelkistysjaosta, jossa homologiset kromosomit erotetaan. Ainoa merkittävä ero klassiseen mitoosiin on se, että kromosomit ovat haploidisia.
Joten ensimmäisen asteen spermatosyyttien ensimmäinen meioottinen jakautuminen johtaa kahden sekundaarisen spermatosyytin muodostumiseen (toinen kerta). Pelkistysjakauman seurauksena muodostuneiden rakenteiden molemmat kromatidit ovat sisarkromatideja. Jälkimmäiset syntyvät ensimmäistä meioottista jakautumista edeltävän replikoinnin tuloksena. Kunkin sekundaarisen spermatosyytin toinen meioottinen jakautuminen johtaa neljän spermatidin muodostumiseen. Siten tyypillisessä meioosissa solut jakautuvat kahdesti, kun taas kromosomit vain kerran (kuva 21).
Spermatogeneesin viimeinen vaihe liittyy erilaistumiseen, joka päättyy siihen, että jokainen suhteellisen suuri, pallomaisesti liikkumaton spermatidi muuttuu pieneksi pitkänomaiseksi liikkuvaksi siittiöksi.
Useimmissa aikuisissa (seksuaalisesti kypsissä) uroseläimissä spermatogeneesiä tapahtuu kiveksissä jatkuvasti tai ajoittain (kausiluonteisesti). Esimerkiksi hyönteisillä spermatogeneesisyklin loppuun saattaminen kestää vain muutaman päivän, kun taas nisäkkäillä tämä sykli kestää viikkoja tai jopa

Riisi. 21.Kromosomien jakautuminen gametogeneesin aikana
kuukaudet. Aikuisella spermatogeneesiä tapahtuu ympäri vuoden. Primitiivisen spermatogonian kehittymisaika kypsiksi siittiöiksi on noin 74 päivää.
Eri lajien organismien tuottamille miesten lisääntymissoluille on ominaista liikkuvuus ja äärimmäinen koon ja rakenteen monimuotoisuus (kuva 22). Jokainen ihmisen siittiö koostuu kolmesta osasta - pää, keskiosa ja häntä (kuva 23). Siittiön pää sisältää ytimen, joka sisältää haploidisen joukon kromosomeja.
Pää on varustettu akrosomilla, joka sisältää lyyttisiä entsyymejä, jotka ovat välttämättömiä siittiön sisällön pääsylle munasoluun. Päässä on myös kaksi sentriolia - proc-

Riisi. 22. Siittiöiden muodot

Riisi. 23. Siittiöiden rakenne: A - valomikroskooppinen kuva eri tasoissa: 1 - pää, 2 - keskiosa, 3 - häntä; B - elektronimikroskooppisen kuvan kaavamainen rekonstruktio: 1 - ydin, 2 - akrosomi, 3 - sentrosomi (proksimaalinen sentrioli), 4 - keskusrengas, 5 - mitokondrioiden heliksi,
6 - aksiaalinen kierre
simaali, joka stimuloi siittiöiden hedelmöittämän munasolun jakautumista, ja distaalinen, joka synnyttää hännän aksimaalisen varren. Siittiön keskiosa sisältää hännän tyviosan ja mitokondriot. Siittiöiden häntä (prosessi) muodostuu sisäisestä aksimaalisesta sauvasta ja ulkoisesta vaipasta, joka on sytoplasmista alkuperää. Ihmisen siittiöille on ominaista merkittävä liikkuvuus.
OVOGENESIS- JA OGGLE-SOLUJAT
Munasolujen muodostumisprosessia kutsutaan oogeneesiksi. Sen tehtävänä on varmistaa haploidinen kromosomisarja munan ytimessä ja tsygootin ravitsemustarpeet. Oogeneesi ilmenemismuodossaan on periaatteessa verrattavissa spermatogeneesiin.
Nisäkkäillä ja ihmisillä oogeneesi alkaa synnytystä edeltävänä aikana (ennen syntymää). Oogoniat, jotka ovat pieniä soluja, joissa on melko suuri tuma ja jotka sijaitsevat munasarjan follikkeleissa, alkavat erilaistua primäärisiksi munasoluiksi. Jälkimmäiset muodostuvat jo kolmannessa kohdunsisäisen kehityksen kuukaudessa, minkä jälkeen ne siirtyvät ensimmäisen meioottisen jakautumisen profaasiin. Kun tyttö syntyy, kaikki primaariset munasolut ovat jo ensimmäisen meioottisen jakautumisen profaasissa. Primaariset munasolut pysyvät profaasissa naarasyksilön murrosiän alkamiseen asti. Yksi munasolu sisältää jopa 100 000 mitokondriota. Kun munasarjan follikkelit kypsyvät murrosiän alkaessa, meioottinen profaasi alkaa uudelleen primaarisissa munasoluissa. Kunkin kehittyvän munan ensimmäinen meioottinen jakautuminen valmistuu vähän ennen munan ovulaation alkamisajankohtaa. Ensimmäisen meioottisen jakautumisen ja sytoplasman epätasaisen jakautumisen seurauksena yhdestä tuloksena olevasta solusta tulee sekundaarinen munasolu, toisesta polaarinen (pelkistys)kappale.
Toissijainen meioottinen jakautuminen tapahtuu, kun sekundaarinen munasolu (kehittynyt muna) siirtyy munasarjasta munanjohtimeen. Tämä jakautuminen ei kuitenkaan valmistu ennen kuin siittiö tunkeutuu sekundaariseen munasoluun, joka yleensä tapahtuu munanjohtimessa. Kun siittiö tunkeutuu sekundaariseen munasoluun, jälkimmäinen jakautuu, jolloin muodostuu munasolu (kypsä munasolu), jossa pronukleus sisältää yhden sarjan
23 äidin kromosomista. Toinen tästä jakautumisesta syntynyt solu on toinen polaarinen kappale, joka ei pysty kehittymään edelleen. Tällä hetkellä myös polaarinen (pelkistys)kappale jakautuu kahteen osaan. Siten yhden ensimmäisen asteen munasolun kehittymiseen liittyy yhden ovotidin ja kolmen pelkistyskappaleen muodostuminen. Munasarjoissa yleensä 300-400 munasolua kypsyy tällä tavalla läpi elämän, mutta vain yksi munasolu kypsyy kuukaudessa. Munien erilaistumisen aikana muodostuu kalvoja ja niiden ytimen koko pienenee.
Joissakin eläinlajeissa oogeneesi tapahtuu nopeasti ja jatkuvasti ja johtaa suuren munamäärän tuotantoon.
Huolimatta yhtäläisyyksistä spermatogeneesin kanssa, oogeneesille on ominaista joitain erityispiirteitä. Primaarisen munasolun ravintoaine (keltuainen) ei jakautunut tasaisesti meioottisen jakautumisen seurauksena muodostuvien neljän solun kesken. Suurin osa keltuaisesta varastoituu yhteen suureen soluun, kun taas polaariset kappaleet sisältävät hyvin vähän tätä ainetta. Jakautumisten seurauksena ensimmäinen ja toinen polaarikappale saavat samat kromosomisarjat kuin sekundaariset munasolut, mutta niistä ei tule sukusoluja. Siksi munat ovat paljon rikkaampia ravintoaineista verrattuna siittiöihin. Tämä ero on erityisen selvä munivien eläinten tapauksessa.
Nisäkkään munien muoto on soikea tai hieman pitkänomainen (kuva 24), ja niille on tunnusomaista tyypilliset solurakenteen piirteet. Ne sisältävät kaikki somaattisille soluille tyypilliset rakenteet, mutta munan solunsisäinen organisaatio on hyvin spesifinen ja sen määrää se, että muna on myös ympäristö, joka varmistaa tsygootin kehittymisen. Yksi munien ominaispiirteistä on niiden kalvojen rakenteen monimutkaisuus. Monilla eläimillä erotetaan munien primaariset, sekundaariset ja tertiaariset kalvot. Ensisijainen kuori (sisäinen) muodostuu munasoluvaiheessa. Se edustaa munasolun pintakerrosta, ja sillä on monimutkainen rakenne, koska sen viereisten follikulaaristen solujen kasvut tunkeutuvat siihen. Toissijainen (keski) kuori muodostuu kokonaan follikulaarisista soluista, ja tertiäärinen (ulompi) kuori muodostuu aineista, jotka ovat munanjohtimien rauhasten eritystuotteita, joiden läpi munat kulkevat. Lintuissa esim.
Munien tertiäärisiä kalvoja ovat valkuaiskalvo, alakuori ja kuorikalvot. Nisäkkään munille on ominaista kahden kalvon läsnäolo. Munien solunsisäisten komponenttien rakenne on lajispesifinen ja joskus jopa yksilöllisiä.

Riisi. 24. Munan rakenne: 1 - follikulaariset solut; 2 - kuori; 3 - sytoplasma; 4- ydin
LANNOITUS
Hedelmöitys on miesten ja naisten sukusolujen yhdistämisprosessi, joka johtaa tsygootin muodostumiseen ja sitä seuraavan uuden organismin kehittymiseen. Hedelmöitysprosessin aikana tsygoottiin muodostuu diploidinen kromosomijoukko, joka määrittää tämän prosessin erinomaisen biologisen merkityksen.
Sukupuolisesti lisääntyvien organismien lajista riippuen erotetaan ulkoinen ja sisäinen hedelmöitys. Ulkoinen hedelmöitys tapahtuu ympäristössä, johon miehen ja naisen sukusolut tulevat. Esimerkiksi kalojen lannoitus on ulkoista. Niiden vapauttamat uros- (maito) ja naaraspuoliset (kaviaari) sukusolut tulevat veteen, jossa ne kohtaavat ja yhdistyvät.
Sisäinen hedelmöitys varmistetaan siittiöiden siirtymisellä miehen kehosta naisen kehoon. Tällaista hedelmöitystä tapahtuu nisäkkäillä, joiden keskeinen kohta on sukusolujen ytimien fuusio. Uskotaan, että yhden siittiön sisällön tunkeutuu munasoluun. Lannoitusmekanismeissa
valmistumisesta, paljon on vielä epäselvää. Merisiilien hedelmöityksestä saadut tiedot osoittavat, että jo 2 sekuntia siittiön ja munasolun kosketuksesta tapahtuu muutoksia jälkimmäisen plasmakalvon sähköisissä ominaisuuksissa. Sukusolujen fuusio tapahtuu 7 sekunnissa. Oletetaan, että vain yhden monista siittiöiden sisällön tunkeutuminen munasoluun selittyy sen plasmakalvon sähköisten ominaisuuksien muutoksilla. Hedelmöitetty munasolu synnyttää tsygootin. On olemassa kaksi mielipidettä siitä, miksi siittiöt aktivoivat munan aineenvaihduntaa. Jotkut uskovat, että siittiöiden sitoutuminen solujen pinnalla oleviin ulkoisiin reseptoreihin on signaali, joka tulee munasoluun kalvon kautta ja aktivoi siellä inositolitrifosfaattia ja kalsiumioneja. Toiset uskovat, että siittiöt sisältävät erityisen aloitustekijän.
Viime vuosien kokeellinen kehitys on osoittanut, että nisäkkäiden, myös ihmisten, munien hedelmöittäminen on mahdollista in vitro. Lisäksi in vitro -kehitetyt alkiot voidaan istuttaa naisen kohtuun, jossa ne käyvät läpi normaalin kehityksen. On olemassa lukuisia "koeputkilasten" syntymätapauksia.
Toisin kuin tsygogeneesin avulla lisääntyvät eläimet, monet organismit pystyvät lisääntymään partenogeneesin avulla (kreikasta. partenos - neitsyt ja genos - syntymä), joka viittaa organismien lisääntymiseen hedelmöittämättömästä munasta. On olemassa pakollista ja fakultatiivista partenogeneesiä. Pakollisesta partenogeneesistä on tullut pääasiallinen menetelmä tiettyjen lajien organismien, esimerkiksi valkoihoisen kiviliskon, lisääntymiselle. Tämän lajin eläimet ovat vain naaraita. Päinvastoin, fakultatiivinen partenogeneesi tarkoittaa, että munasolut pystyvät kehittymään sekä ilman hedelmöitystä että hedelmöityksen jälkeen. Fakultatiivinen partenogeneesi puolestaan on nainen ja mies. Naaraspartenogeneesiä havaitaan usein mehiläisissä, muurahaisissa ja rotifereissa, joissa urokset kehittyvät hedelmöittämättömistä munista. Urospartenogeneesiä tapahtuu joissakin isogamisissa levissä.
Partenogeneesi voi olla sekä luonnollista että keinotekoista (indusoitua). Keinotekoisen partenogeneesin mekanismi on munien ärsytys fysikaalisesti tai kemiallisesti
ikaalisia tekijöitä, mikä johtaa munien aktivoitumiseen ja sen seurauksena hedelmöittämättömien munien kehittymiseen. Keinotekoista partenogeneesiä on havaittu moniin systemaattisiin ryhmiin kuuluvilla eläimillä - piikkinahkaisilla, madoilla, nilviäisillä ja jopa nisäkkäillä.
Tunnettua partenogeneesin muotoa kutsutaan androgeneesiksi (kreikasta. andros - mies, synty - alkuperä). Jos munasolun ydin inaktivoituu ja siihen tunkeutuu useita siittiöitä, tällaisesta munasta kehittyy miespuolinen organismi urosytimien (siementen) fuusion seurauksena.
On tapauksia, joissa partenogeneesi tapahtuu syklisesti vuodenajasta riippuen. Esimerkiksi rotiferit, vesikirput ja kirvat lisääntyvät kesällä partenogeneesillä ja syksyllä munasolujen hedelmöityksellä ja tsygoottien muodostuksella eli tsygogeneesillä.
Partenogeneesin ja sen muotojen rooli luonnossa on pieni, koska se ei tarjoa organismien laajoja sopeutumiskykyjä.
Toisin kuin tsygogeneesi ja partenogeneesi, on olemassa gynogeneesiä, joka on pseudogamiaa, kun siittiö pääsee munasoluun ja aktivoi sen, mutta siittiön ydin ei fuusioidu munan ytimeen. Tässä tapauksessa tuloksena olevat jälkeläiset koostuvat vain naaraista. Gynogeneesiä esiintyy luontaisesti sukkulamadoissa ja kaloissa, mutta se voidaan indusoida myös keinotekoisesti. Erityisesti on kuvattu keinotekoisen gynogeneesin tapauksia silkkiäistoukeissa, kaloissa ja sammakkoeläimissä.
VUOROTTAVA HAPLOIDIA JA DIPLOIDIA.
VAIHTOLUVIA SUKUPOLVIA
Sukupuolisesti lisääntyville organismeille on ominaista vaihtelevat haploidi- ja diploidivaiheet niiden kehityksessä. Monissa organismeissa, mukaan lukien nisäkkäät, tämä vuorottelu on säännöllistä, ja organismien lajiominaisuuksien säilyminen perustuu siihen.
Monille organismeille on ominaista myös sukupolvien vaihtuminen, jolloin sukupuolisesti lisääntyvien yksilöiden sukupolvet korvataan sukupuolisesti lisääntyvien sukupolvien kanssa sukusolujen muodostuessa. Tätä kutsutaan ensisijaiseksi sukupolvenvaihdokseksi. Sitä esiintyy itiöeläimissä, siimakasveissa ja monissa kasveissa. Ensisijainen sukupolvien vaihto on säännöllistä, ja se on läsnä
Se, mitä luonnosta löytyy, osoittaa, että monien organismien fysiologiassa on säilynyt sekä aseksuaalinen että seksuaalinen lisääntyminen. Muissa organismeissa tapahtuu sukupuolisen lisääntymisen vuorottelua partenogeneesin kanssa. Tätä kutsutaan toissijaiseksi sukupolvenvaihdoksi. Esimerkiksi trematodeissa sukupuolinen lisääntyminen korvataan säännöllisesti partenogeneesillä. Tätä toissijaista sukupolvien vuorottelua kutsutaan heterogoniaksi. Koelenteraateissa joissakin kehitysvaiheissa tapahtuu siirtymä seksuaalisesta lisääntymisestä aseksuaaliseen (vegetatiiviseen). Tätä sukupolvien toissijaisen vuorottelun muotoa kutsutaan metageneesiksi.
SEKSUAALINEN DIMORFISMI. HERMAFRODITISMI
Miehillä ja naarailla on erityisiä fenotyyppisiä piirteitä. Naisten ja urosten välisiä eroja niiden ominaisuuksissa kutsutaan seksuaaliseksi dimorfismiksi. Eläimillä sitä esiintyy jo evoluution alemmissa vaiheissa, esimerkiksi pyöreissä helminteissä ja niveljalkaisissa, ja saavuttaa suurimman ilmenemismuotonsa selkärankaisissa.
Jos miehen ja naisen sukusoluja tuottaa sama yksilö, jolla on sekä miehen että naisen sukurauhaset, tätä ilmiötä kutsutaan todelliseksi hermafroditismiksi. Sitä löytyy lattamatoista, nilviäisistä ja nilviäisistä. Tasomadoilla uros- ja naaraspuoliset sukurauhaset toimivat koko yksilön elämän ajan. Sitä vastoin nilviäisten sukurauhaset tuottavat vuorotellen munia ja siittiöitä.
Todellista hermafroditismia esiintyy myös ihmisillä, mikä johtuu kehityshäiriöistä. Hermafrodiittien genotyypit ovat 46 XX tai 46 XY, ja suurin osa tapauksista on XX (noin 60 %). Genotyyppi XX löytyy useimmiten negroidiafrikkalaisten populaatioiden hermafrodiiteista, kun taas XY löytyy useimmiten japanilaisista. Molemmissa hermafrodiiteissa havaittiin taipumus molemminpuoliseen sukurauhasten epäsymmetriaan. Todellisten hermafrodiittien joukossa on myös kromosomaalisia mosaiikkeja, joissa joissakin somaattisissa soluissa on pari XX kromosomia, toisissa - pari XY.
Väärä hermafroditismi tunnetaan myös, kun yksilöillä on molemmille sukupuolille tyypillisiä ulkoisia sukupuolielimiä ja toissijaisia seksuaalisia ominaisuuksia, mutta ne tuottavat vain yhden tyyppisiä sukusoluja - uros- tai naarassoluja.
LISÄYSMENETELMIEN ALKUPERÄ
Uskotaan, että vanhin on suvuton lisääntyminen, erityisesti kasvullinen lisääntyminen. Jälkimmäisestä kehittyi itiöiden lisääntyminen, jonka kiistaton etu on, että se tarjoaa paremmat mahdollisuudet lajien säilymiseen ja erityisesti niiden leviämiseen.
Sukupuolinen lisääntyminen on tehokkain tapa organismien lisääntymiseen. Uskotaan, että se kehittyi aseksuaalista, syntyi noin 1 miljardi vuotta sitten, ja ensimmäiset vaiheet liittyivät sukusolujen kehityksen komplikaatioon. Primitiivisille sukusoluille oli ominaista riittämätön morfologinen erilaistuminen, minkä seurauksena isogamia (kreikasta. isos- yhtä suuri, gamos - avioliitto), kun sukupuolisolut olivat liikkuvia isogameetteja, joita ei erilaistu uros- ja naarasmuotoihin.
Myöhemmin kehittyi anisogamia (kreikasta. anisos- epätasa-arvoinen, gamos - avioliitto), jolle on ominaista erilaisten sukusolujen läsnäolo, jotka eroavat vain kooltaan. Myöhemmissä evoluution vaiheissa sukusolujen liikkuvuudessa, muodossa ja koossa syntyi jyrkkiä eroja. Evoluutioprosessissa selkärankaiset kehittivät myös useita lisälaitteita, jotka helpottavat miehen siittiöiden siirtymistä naisen lisääntymiselimiin ja luovat olosuhteet hedelmöittyneen munan kehittymiselle. Evoluution aikana nämä laitteet kehittyivät eritysjärjestelmästä, mikä johti virtsaelimen muodostumiseen.
Diploiditila tarjoaa äärimmäisiä etuja organismeille, koska tässä tilassa tapahtuu erilaisten alleelien kertymistä. Siksi seksuaalisella lisääntymisellä on myös se etu, että se tarjoaa organismeilleOsuurempi mahdollisuus vaihtelua verrattuna aseksuaaliin, ja tällä on ratkaiseva rooli evoluutiossa.
Parthenogeneesillä on selviä lisääntymisetuja, koska se tuottaa vain naarasjälkeläisiä. Se on kuitenkin harvinaista. Kahta hypoteesia käytetään selittämään sukupuolisen lisääntymisen vähäistä korvaamista partenogeneesillä luonnollisissa organismipopulaatioissa. Yhden niistä (mutaatiokertyvä) mukaan sukupuoli on mukautuva sopeutuminen, koska se "puhdistaa" genomin ajan myötä toistuvista mutaatioista, kun taas toisen hypoteesin (ekologinen) mukaan sukupuoli on
Ensimmäiset ajatukset kasvusta ja kehityksestä juontavat juurensa antiikin maailmaan. Jopa Hippokrates (460-377 eKr.) oletti, että munat sisältävät jo täysin muodostuneen organismin, mutta hyvin pelkistetyssä muodossa. Tämä ajatus kehitettiin sitten preformationismin opiksi (lat. preformatio - esimuodostus), joka osoittautui erityisen suosituksi 1600-1700-luvuilla. Preformationismin kannattajia olivat Harvey, Malpighi ja monet muut tuon ajan tunnetut biologit ja lääkärit. Preformationistien kannalta kiistanalainen kysymys oli vain, missä sukupuolisoluissa organismi esimuodostettiin - naaras- vai miespuolinen. Niitä, jotka suosivat munaa, kutsuttiin ovisteiksi, ja niitä, jotka pitivät enemmän miehen sukusoluja tärkeänä, kutsuttiin eläinkulisteiksi. Preformationismi on metafyysinen oppi alusta loppuun, sillä se kielsi kehityksen. Ratkaisevan iskun preformationismille antoi C. Bonnet (1720-1793), joka löysi partenogeneesin vuonna 1745 käyttämällä esimerkkiä kirvojen kehittymisestä hedelmöittämättömistä munista. Tämän jälkeen preformationismi ei enää toipunut ja alkoi menettää merkityksensä.
Muinaisessa maailmassa syntyi toinen oppi, joka oli vastakohta preformationismille ja sai myöhemmin nimen epigenesis (kreikasta. epi- jälkeen, synty - kehitys). Kuten preformationismi, myös epigeneesi yleistyi 1600-1700-luvuilla. Epigeneesin leviämisessä K.F:n näkemykset olivat erittäin tärkeitä. Wolf (1733-1794), tiivistetty kirjassaan "Kehityksen teoria" (1759). K.F. Susi uskoi, että muna ei sisällä esimuodostettua organismia eikä sen osia ja että se muodostuu alun perin homogeenisesta massasta. Toisin kuin esiformistit, K.F. Susi ja muut epigeneesin kannattajat olivat aikansa edistyksellisiä, koska he sisälsivät ajatuksen kehityksestä. Myöhemmin ilmaantui kuitenkin uusia hetkiä. Erityisesti vuonna 1828 K. Baer julkaisi teoksensa "The History of Animal Development", jossa hän osoitti, että munan sisältö on heterogeenista eli rakenteellista ja rakenteen aste kasvaa alkion kehittyessä. Siten K. Baer osoitti sekä preformationismin että epigeneesin epäjohdonmukaisuuden.
Kasvu ja kehitys ovat elävien olentojen tärkeimpiä ominaisuuksia. Kasvu on koko organismin massan kasvua solujen määrän lisääntymisen seurauksena, kun taas kehitys on laadullisia muutoksia elimistössä, jotka määräytyvät solujen erilaistumisesta ja morfogeneesistä, mikä varmistaa yksilöissä progressiiviset muutokset munasta alkaen. ja päättyen heidän aikuisuuteensa.
Ontogeneesi (kreikasta. päälle - olento, synty - kehitys) on yksilön kehityksen historia (sykli), joka alkaa sen synnyttäneiden sukusolujen muodostumisesta ja päättyy sen kuolemaan. Ajatukset ontogeniasta perustuvat tietoon kasvusta, kehityksestä ja erilaistumisesta. Ontogeneesin perusperiaatteiden tutkiminen on tärkeää ihmisen biologian ymmärtämisen kannalta.
Nykyaikaiset tiedot organismien kehityksestä torjuvat sekä preformationismin että epigeneesin. Nykyaikaisten käsitteiden puitteissa organismin kehittyminen ymmärretään prosessiksi, jossa aikaisemmin muodostuneet rakenteet stimuloivat myöhempien rakenteiden kehittymistä. Kehitysprosessi määräytyy geneettisesti ja liittyy läheisesti ympäristöön. Kehityksen määrää siis sisäisten ja ulkoisten tekijöiden yhtenäisyys.
KASVUUN JA KEHITTYMISEN YKSITYISYYS
Organismin kasvu on sen massan asteittaista kasvua ja muodon muutosta, joka johtuu solujen lukumäärän ja niiden erilaistumisen lisääntymisestä, kudosten ja elinten muodostumisesta sekä solujen ja kudosten biokemiallisista muutoksista. Siten kasvu on seurausta kvantitatiivisista muutoksista solujen lukumäärän (ruumiinpainon) kasvun muodossa ja laadullisista muutoksista solujen erilaistumisen ja morfogeneesin muodossa. Solujen erilaistuminen on prosessi, jossa jotkut solut muuttuvat morfologisesti, biokemiallisesti ja toiminnallisesti erilaisiksi muista soluista. Joidenkin solujen lisääntyminen ja erilaistuminen koordinoidaan aina toisten kasvun ja erilaistumisen kanssa. Molemmat prosessit tapahtuvat koko organismin elinkaaren ajan. Koska erilaistuvat solut muuttavat muotoaan ja soluryhmät osallistuvat muodonmuutoksiin, tähän liittyy morfogeneesi, joka on joukko prosesseja, jotka määräävät solujen ja kudosten rakenteellisen organisoinnin sekä organismien yleisen morfologian.
Kasvua voidaan mitata rakentamalla mittaustulosten perusteella käyriä kehon koosta, painosta, kuivamassasta, solujen lukumäärästä, typpipitoisuudesta ja muista indikaattoreista.
ONTOGENEESI JA SEN TYYPIT. ONTOGENEESIIN PERIODIOINTI
Ontogeneesi luokitellaan organismien kehityksen luonteesta riippuen suoraan ja epäsuoraan, ja siksi tehdään ero suoran ja epäsuoran kehityksen välillä. Organismien suora kehitys luonnossa tapahtuu ei-toukkien ja kohdunsisäisen kehityksen muodossa, kun taas epäsuora kehitys havaitaan toukkien kehityksen muodossa. Toisin kuin ontogenia, lajiluokka on fylogia.
Toukkien kehitys. Tämä kehitys ymmärretään epäsuoraksi kehitykseksi, koska organismien kehityksessä on yksi tai useampi toukkavaihe. Toukkien kehitys on tyypillistä hyönteisille, sammakkoeläimille ja piikkinahkaisille. Näiden eläinten toukat elävät itsenäistä elämäntapaa, minkä jälkeen ne muuttuvat. Siksi tätä kehitystä kutsutaan myös kehitykseksi metamorfoosilla (katso alla).
Ei-toukkien kehitys. Tämä kehitysmuoto on ominaista suoraan kehittyville organismeille, esimerkiksi kaloille, matelijoille ja linnuille, joiden munassa on runsaasti keltuaista (ravintoaine). Tästä johtuen merkittävä osa ontogeneesistä tapahtuu ulkoiseen ympäristöön munivissa munissa. Alkioiden aineenvaihdunta varmistetaan kehittämällä väliaikaisia elimiä, jotka ovat alkion kalvoja (keltuaispussi, amnion, allantois).
Kohdunsisäinen kehitys. Tämä kehitys on ominaista myös suoraan kehittyville organismeille, kuten nisäkkäille, mukaan lukien ihmiset. Koska näiden organismien munat ovat ravintoaineltaan erittäin köyhiä, äidin keho tarjoaa kaikki alkioiden elintärkeät toiminnot muodostamalla väliaikaisia elimiä äidin ja alkion kudoksista, joista tärkein on istukka. Evoluutioteoriassa kohdunsisäinen kehitys on viimeisin muoto, mutta se on kaikkein hyödyllisin alkioille, koska se varmistaa tehokkaasti niiden selviytymisen.
Ontogeneesi on jaettu alkusikiön, alkion ja postembryoniseen ajanjaksoon. Ihmisten tapauksessa kehitysjaksoa ennen syntymää kutsutaan prenataaliseksi tai synnytystä edeltäväksi, syntymän jälkeen - postnataaliseksi. Kehittyvää alkiota ennen elinrudimenttien muodostumista kutsutaan alkioksi, elinrudimenttien muodostumisen jälkeen sikiöksi.
ALKUALEN KEHITYS
Tämä organismien yksilöllisen kehityksen ajanjakso liittyy sukusolujen muodostumiseen gametogeneesin prosessissa. Miesten lisääntymissoluilla ei ole merkittäviä eroja muista soluista, kun taas munat eroavat siinä, että ne sisältävät paljon keltuaista. Ottaen huomioon keltuaisen määrän ja sen jakautumisen munissa, jälkimmäiset luokitellaan kolmeen tyyppiin:
1) isosetaaliset munat, jossa on vähän keltuaista, joka jakautuu tasaisesti koko soluun. Näitä munia tuottavat piikkinahkaiset (merisiilit), alemmat jänteet (lansetit) ja nisäkkäät;
2) telolesitaaliset munat sisältävät suuren määrän keltuaista, joka on keskittynyt yhteen navoista - vegetatiiviseen. Tällaisia munia tuottavat nilviäiset, sammakkoeläimet, matelijat ja linnut. Esimerkiksi sammakonmunat koostuvat 50% keltuaisesta, kananmunista (yleensä kananmunista) - 95%. Telolesitaalien munien toisessa navassa (eläin) sytoplasma ja tuma keskittyvät;
3) sentrolesitaaliset munasolut, jossa keltuaista on vähän ja se sijaitsee keskeisellä paikalla. Sytoplasma sijaitsee tällaisten munien reunalla. Sentrolesitaalimunia tuottavat niveljalkaiset.
Esialkion aikakaudelle on ominaista myös se, että tänä aikana sukusoluissa tapahtuu DNA-molekyylien kertymiseen liittyviä metabolisia prosesseja.
ALKIÖKEHITYS
Embryogenesis (kreikasta. etbryop - alkio) eli alkiokausi alkaa uros- ja naarassolujen fuusiolla, jotka edustavat munasolujen hedelmöitysprosessia.
Organismeissa, joille on ominaista kohdunsisäinen kehitys, alkiojakso päättyy syntymään, ja organismeissa, joille on tunnusomaista toukka- ja ei-toukkatyyppinen kehitys, alkiojakso päättyy organismin vapautumiseen munasta tai alkion kalvoista. Alkion aikana erotetaan tsygootin, pilkkoutumisen, blastulan, itukerrosten muodostumisen, histogeneesin ja organogeneesin vaiheet.
Tsygootti.Hedelmöitykseen liittyy sarja prosesseja, joissa miehen sukusolu aloittaa munasolun kehittymisen. Urospuolisen sukusolun aktivoimassa munassa tapahtuu useita fysikaalisia ja kemiallisia prosesseja, mukaan lukien lisääntynyt proteiinisynteesi. Protoplasman liike johtaa munan kahdenvälisen symmetrian muodostumiseen. Ytimet fuusioituvat ja kromosomien diploidisarja palautuu. Tämä luo yksisoluisen organismin.
Erota.Se edustaa tsygootin (hedelmöitetyn munan) kehitysvaihetta, joka koostuu tsygootin jakamisesta mitoosin kautta. Jakautuminen alkaa uurteen ilmestymisestä munan pinnalle. Ensimmäinen uurre johtaa kahden solun muodostumiseen - kaksi blastomeeriä, toinen - neljä blastomeeriä, kolmas - kahdeksan blastomeeriä (kuva 25). Peräkkäisten fragmenttien seurauksena muodostunutta soluryhmää kutsutaan morulaksi (latinasta. morum- mulperipuu).
Tämän vaiheen biologinen merkitys on siinä, että suuresta solusta, joka on muna, muodostuu pienempiä soluja, joissa sytoplasman suhde ytimeen vähenee.
Tsygootin pirstoutuminen päättyy monisoluisen rakenteen muodostumiseen, jota kutsutaan blastulaksi (kreikasta. blastot - versoa). Tämä rakenne on blastodermiksi kutsutun vesikkelin muotoinen, ja se koostuu yhdestä solukerroksesta. Nyt näitä soluja kutsutaan alkiosoluiksi. Blastula on kooltaan samanlainen kuin muna. Fragmentoitumisjakson aikana ytimien lukumäärä ja DNA:n kokonaismäärä kasvaa. Myös pieniä määriä mRNA:ta ja tRNA:ta syntetisoidaan, kun taas ribosomaalista RNA:ta ei ole vielä havaittavissa.
Kaikki eläimet käyvät läpi blastula-vaiheen, mutta jokaisella tapauksella on omat ominaisuutensa. Nisäkkäillä jakautuminen on epätasaista, joten morulat koostuvat eri määrästä soluja. Lisäksi osa soluista muodostaa rakenteen, jota kutsutaan trofoblastiksi,

Blastocoel Larva Nuijapää
muodossa
Riisi. 25.Tsygoottien fragmentoituminen ja blastulien muodostuminen eri organismeissa: 1 - alkuperäinen muna; 2 - kaksi blastomeeria; 3 - neljä blastomeeria; 4 - kahdeksan blastomeeria; 5 - blastula; 6 - aikuisten muoto
joiden solut ravitsevat alkiota ja entsyymien ansiosta varmistavat sen tunkeutumisen kohdun seinämään. Myöhemmin trofoblastisolut irtoavat alkiosta ja muodostavat rakkulan, joka täyttyy kohtukudoksesta tulevalla nesteellä.
Tämän vaiheen biologinen merkitys on siinä, että suuresta solusta, kuten munasolusta, muodostuu pienempiä soluja, joissa sytoplasman suhde ytimeen vähenee ja ytimessä on uusi sytoplasminen ympäristö.
Gastrulaatio(kreikasta vatsa - suonen ontelo). Tämä on alkiosolujen liikkumisprosessi blastulan muodostumisen jälkeen, johon liittyy kahden tai kolmen (eläimen tyypistä riippuen) alkion kerroksen tai ns. itukerrosten muodostuminen (kuva 26). ).
Isolekitaalisten munien kehittyminen (gastrulaatio) tapahtuu blastulan sisällä olevan vegetatiivisen navan invaginaatiolla (invaginaatiolla), jonka seurauksena vastakkaiset navat melkein sulautuvat ja blastocoel (blastulan ontelo) katoaa melkein tai kokonaan. Ulkoinen
Riisi. 26.Gastrulaatio eri organismeissa: 1 - blastocoel; 2 - primaarinen mesenkyymi; 3, 4, 5 - primaarinen suolisto
Alkioiden solukerrosta kutsutaan ektodermiksi (kreikasta. ectos - ulkopuolella, derma - iho) tai ulompi itukerros, kun taas sisempi on endodermi (kreikasta. entos - sisällä) tai sisäinen alkiokerros. Tässä tapauksessa muodostunutta onteloa kutsutaan gastrocoeliksi tai primaariseksi suoleksi, jonka sisäänkäyntiä kutsutaan blastoporeksi (primaarinen suu).
Kahden itukerroksen kehittyminen on ominaista sienille ja coelenteraateille. Kuitenkin chordaateille gastrulaatiojakson aikana on ominaista kolmannen itukerroksen - mesodermin - kehittyminen (kreikasta. mesos - keskellä), muodostuu ektodermin ja endodermin väliin.
Gastrulaatio on välttämätön edellytys tuleville kehitysvaiheille, koska se tuo solut asentoon, joka mahdollistaa elimien muodostumisen. Kolmeen alkioanlageen erilaistunut alkiomateriaali synnyttää kaikki kehittyvän alkion kudokset ja elimet.
Alkukerrosten kehittymiseen (erilaistumiseen) liittyy erilaisten kudosten ja elinten muodostuminen niistä. Erityisesti ihon orvaskesi, kynnet ja hiukset, tali- ja hikirauhaset, hermosto (aivot, selkäydin, hermosolmut, hermot), aistielinten reseptorisolut, silmän linssi, suun epiteeli , nenäontelo ja peräaukko, hammas -
ei emali. Endodermista kehittyy ruokatorven, mahan, suoliston, sappirakon, henkitorven, keuhkoputkien, keuhkojen, virtsaputken epiteeli sekä maksa, haima, kilpirauhanen, lisäkilpirauhanen ja kateenkorva. Mesodermista kehittyvät sileät lihakset, luusto- ja sydänlihakset, dermis, sidekudos, luut ja rustot, hammasdentiini, veri ja verisuonet, suoliliepe, munuaiset, kivekset ja munasarjat. Ihmisillä aivot ja selkäydin eroavat ensimmäisenä. 2 kuukauden kuluttua melkein kaikki kehon rakenteet ilmestyvät. Alkion yhteys ympäristöön tapahtuu väliaikaisten elinten kautta. Organogeneesi päättyy alkiokauden lopussa. Jos lopullinen suuaukko muodostuu ensisijaisen suun (blastopore) kohdalle, näitä eläimiä kutsutaan protostomeiksi (madot, nilviäiset, niveljalkaiset).
Jos lopullinen suu muodostuu vastakkaiseen paikkaan, näitä eläimiä kutsutaan deuterostomeiksi (piikkinahkaiset, chordates).
Alkion ja ympäristön välisen yhteyden varmistamiseksi käytetään ns. väliaikaisia elimiä, jotka ovat olemassa väliaikaisesti. Munatyypistä riippuen väliaikaiset elimet ovat erilaisia rakenteita. Kaloilla, matelijoilla ja linnuilla väliaikaisia elimiä ovat keltuainen pussi. Nisäkkäillä keltuainen pussi muodostuu alkion synnyn alussa, mutta ei kehity. Myöhemmin sitä vähennetään. Alkion ulkokalvoa kutsutaan chorioniksi. Se kasvaa kohtuun. Paikka, jossa kohtuun kasvaa eniten, on nimeltään istukka. Alkio on yhteydessä istukkaan napanuoran tai napanuoran kautta, joka sisältää verisuonia, jotka tarjoavat istukan verenkierron. Sikiön aineenvaihdunta varmistetaan istukan kautta.
Alkion osien muodostava vuorovaikutus perustuu tiettyihin koordinoituihin aineenvaihduntaprosesseihin. Kehitysmalli on heterokronia, jolla tarkoitetaan elinten kielien erilaista muodostumista ajan myötä ja niiden kehittymisen erilaista intensiteettiä. Ne elimet ja järjestelmät, joiden pitäisi alkaa toimia aikaisemmin, kehittyvät nopeammin. Esimerkiksi ihmisillä yläraajojen alkeet kehittyvät nopeammin kuin alaraajojen alkeet.
Alkiot ovat erittäin herkkiä erilaisille vaikutuksille. Siksi erotetaan kriittiset jaksot, eli jaksot, jolloin alkiot ovat herkimpiä vahingollisille tekijöille. Ihmisten tapauksessa alkion ontogeneesin kriittiset jaksot
ovat ensimmäiset päivät hedelmöityksen jälkeen, istukan muodostumisen ja synnytyksen aika.
Somaattisten solujen ytimet pystyvät varmistamaan munien normaalin kehityksen, mikä selvisi somaattisten solujen ytimien siirtokokeissa muniin, joissa ei ole ytimiä.
Kokeet osoittivat myös, että yhden rodun lampaiden 8- ja 16-soluisista alkioista peräisin olevan yksittäisen blastomeerin muuntumiseen toisen rodun munan tumapuoliskoksi (jälkimmäisen puolikkaan leikkaamisen jälkeen) seurasi elävien alkioiden kehittyminen. ja karitsojen syntymä.
Hippokrateen ajoista (5. vuosisata eKr.) lähtien on keskusteltu sikiön syntymän käynnistävistä syistä. Erityisesti Hippokrates itse ehdotti, että sikiön kehitys käynnistää oman syntymänsä. Englantilaisten tutkijoiden uusin lampailla tekemä kokeellinen työ on osoittanut, että lampaiden poikimisen alkamista ohjaa monimutkainen hypotalamus + aivolisäke + sikiön lisämunuaiset. Hypotalamuksen ytimien vaurioituminen, aivolisäkkeen tai lisämunuaisen etulohkon poisto pidentää lampaiden tiineyttä. Päinvastoin, adenokortikotrooppisen hormonin (erite aivolisäkkeen etuosasta) tai kortisolin (lisämunuaisen erite) antaminen lampaille lyhentää niiden tiineyden kestoa.
Joten korkeampien eukaryoottien kehitysprosessissa yksi hedelmöitetty tsygoottisolu synnyttää mitoosin seurauksena tapahtuvan jatkokehityksen aikana erityyppisiä soluja - epiteeli-, hermo-, luu-, verisoluja ja muita, joille on ominaista erilaisia morfologioita ja makromolekyylikoostumuksia. Erityyppisille soluille on kuitenkin myös ominaista, että ne sisältävät samat geenisarjat, mutta ovat erittäin erikoistuneita ja suorittavat vain yhtä tai useampaa erityistoimintoa, toisin sanoen jotkut geenit "toimivat" soluissa, kun taas toiset ovat inaktiivisia. Esimerkiksi,

Riisi. 27. Yksitsygoottisten kaksosten kehitys
vain punasolut ovat spesifisiä hemoglobiinin synteesissä ja varastoinnissa. Samalla tavalla vain epidermaaliset solut syntetisoivat keratiinia. Siksi somaattisten solujen tumien geneettisestä identiteetistä ja hedelmöittyneiden munasolujen kehityksen ohjausmekanismeista on jo pitkään herännyt kysymyksiä, jotka ovat edellytyksenä solujen erilaistumisen taustalla olevien mekanismien ymmärtämiselle. Kantasoluilla on suuri merkitys solujen erilaistumisessa (katso alla). Hiirillä tehdyt kokeet ovat osoittaneet, että yhden tyypin solut pystyvät muuntumaan toisen tyyppisiksi soluiksi. Erityisesti erilaistuvien tai erilaistuneiden maksasolujen on osoitettu muuttuvan haimasoluiksi.
50-luvulta lähtien. XX vuosisadalla Monissa laboratorioissa suoritettiin kokeita somaattisten solujen tumien onnistuneesta siirrosta muniin, joista oli keinotekoisesti poistettu omat ytimensä. Erilaistuneiden solujen ytimistä peräisin olevan DNA:n tutkimus osoitti, että lähes kaikissa tapauksissa genomit sisältävät samat nukleotidiparien sekvenssit. Tunnetaan poikkeuksia, joissa nisäkkään punasolut menettävät tumansa viimeisen erilaistumisvaiheen aikana. Mutta tähän mennessä altaat
Jotkut hemoglobiinin mRNA:t on jo syntetisoitu, joten punasolut eivät enää tarvitse ytimiä. Muita esimerkkejä ovat immunoglobuliini- ja T-solugeenit, joita modifioidaan kehityksen aikana.
Yksi tärkeimmistä vaiheista alkion ontogeneesin ohjausmekanismien ymmärtämisessä olivat 1960-1970-luvuilla tehtyjen kokeiden tulokset. Englantilainen tutkija D. Gurdon selvittääkseen, onko somaattisten solujen ytimillä kykyä varmistaa munien jatkokehitys, jos nämä ytimet viedään muniin, joista niiden omat ytimet on aiemmin poistettu. Kuvassa Kuvassa 28 on kaavio yhdestä näistä kokeista, jossa nuijapäiden somaattisten solujen tumat siirrettiin sammakon muniin, joista tumat oli aiemmin poistettu. Nämä kokeet osoittivat, että somaattisten solujen ytimet voivat todellakin varmistaa munien jatkokehityksen, koska ne pystyivät hedelmöittämään munia ja "pakottamaan" ne kehittymään edelleen. Tämä osoitti eläinten kloonauksen mahdollisuuden.

Riisi. 28.Kaavio kokeesta somaattisten solujen tumien siirtämiseksi tumallisiin muniin (D. Gurdon, 1968)
Myöhemmin muut tutkijat suorittivat kokeita, joissa he osoittivat, että yksittäisten blastomeerien siirtyminen yhden rodun lampaiden 8- ja 16-päiväisistä alkioista toisen rodun munan tumalliseen puolikkaaseen (sen jälkeen kun jälkimmäinen oli leikattu puoliksi) seurasi elinkelpoisten alkioiden muodostuminen ja sitä seuraava karitsojen syntymä.
Englantilaiset kirjoittajat totesivat vuoden 1997 alussa, että somaattisten solujen ytimien (aikuisten lampaiden alkioiden, sikiöiden tai utareiden solut) tuominen keinotekoisesti poistettuihin lampaanmuniin ja sitten näin hedelmöitettyjen munien istuttaminen lampaan kohtuun liittyy tiineyden esiintyminen ja sen jälkeen karitsojen syntymä (kuva 29). Yksi karitsoista oli nimeltään Dolly. Vuonna 2003 Dolly kuoli. Tänä aikana saatiin hiirten, lehmien, kanien, hevosten, rottien ja muiden eläinten alkioita.

Kulttuurin tuominen äidin kehoon
Riisi. 29.Somaattisten solujen ytimien siirto tumallisiin muniin
Näiden tulosten arviointi osoittaa, että nisäkkäät voivat lisääntyä aseksuaalisesti tuottaen jälkeläisiä eläimistä, joiden solut sisältävät isältä tai emältä peräisin olevaa tumamateriaalia luovuttajalampaiden sukupuolesta riippuen. Tällaisissa soluissa
vain sytoplasma ja mitokondriot ovat äidin alkuperää. Tällä päätelmällä on erittäin tärkeä yleinen biologinen merkitys ja se laajentaa näkemyksiämme eläinten lisääntymispotentiaalista. Mutta on myös tärkeää lisätä, että puhumme geenimanipulaatioista, joita luonnossa ei ole. Toisaalta käytännössä nämä geneettiset manipulaatiot edustavat suoraa tapaa kloonata hyvin organisoituneita eläimiä, joilla on halutut ominaisuudet, millä on suuri taloudellinen merkitys. Lääketieteellisesti tätä reittiä voidaan käyttää tulevaisuudessa miesten hedelmättömyyden voittamiseksi.
Alkion normaalille kehitykselle välttämätön geneettinen informaatio ei siis katoa solujen erilaistumisen aikana ns. kantasolujen takia, joilla on potentiaalia kehittyä erilaisiksi kehon soluiksi. Kun kantasolut jakautuvat, jokaisella uudella solulla on potentiaalia joko pysyä kantasoluna tai tulla soluksi, jolla on erityistoiminto (lihassolut, verisolut tai aivosolut). Hedelmöitetty munasolu on totipotentti, koska se synnyttää kehossa erilaisia soluja. Totipotentit kantasolut voivat synnyttää minkä tahansa tyyppisiä soluja paitsi ne, jotka ovat välttämättömiä sikiön kehitykselle. Kantasoluja, jotka voivat synnyttää monenlaisia solutyyppejä, kutsutaan yleisesti multitotipotentti soluja. On osoitettu, että aikuiset kantasolut voivat tuottaa erilaistuneita soluja alkion muista kudoksista. Somaattisilla soluilla on myös ominaisuus nimeltä totipotenssi, eli niiden genomi sisältää kaiken tiedon, jonka ne ovat saaneet erilaistumisen seurauksena hedelmöittyneestä munasolusta. Näiden tietojen läsnäolo tarkoittaa epäilemättä, että solujen erilaistuminen on geneettisen kontrollin alaista. Kantasolujen tutkimuksella on vaikutuksia lääketieteeseen.
On osoitettu, että hedelmöityksen jälkeiseen intensiiviseen proteiinisynteesiin useimmissa eukaryooteissa ei liity mRNA-synteesiä. Oogeneesin tutkimus selkärankaisilla, erityisesti sammakkoeläimillä, on osoittanut, että intensiivistä transkriptiota tapahtuu jopa meioosin I-vaiheen (erityisesti diploneman) aikana. Siksi geenitranskriptit mRNA:n tai pro-mRNA-molekyylien muodossa säilytetään munissa lepotilassa. On todettu, että alkiosoluissa on niin sanottu epäsymmetrinen
kolmiosainen jakautuminen, joka koostuu siitä, että alkiosolun jakautuminen synnyttää kaksi solua, joista vain yksi perii transkriptioon osallistuvat proteiinit. Siten transkriptiotekijöiden epätasainen jakautuminen tytärsolujen välillä johtaa erilaisten geenijoukkojen ilmentymiseen niissä jakautumisen jälkeen, eli solujen erilaistumiseen.
Sammakkoeläimissä ja ehkä useimmissa selkärankaisissa geneettiset ohjelmat, jotka ohjaavat varhaista kehitystä (ennen blastulavaihetta), vakiintuvat oogeneesin aikana. Myöhemmät kehitysvaiheet, kun solujen erilaistuminen alkaa (noin gastrula-vaiheesta lähtien), vaativat uusia ohjelmia geenien ilmentämiseen. Siten solujen erilaistuminen liittyy geneettisen tiedon uudelleenohjelmointiin suuntaan tai toiseen.
Solujen erilaistumisen tunnusomainen piirre on, että se johtaa peruuttamattomasti yhteen tai toiseen solutyyppiin. Tätä prosessia kutsutaan päättäväisyys ja on myös geneettisen kontrollin alainen, ja kuten nyt oletetaan, solujen erilaistumista ja määritystä säätelee solujen vuorovaikutus peptidikasvutekijöiden tyrosiinikinaasireseptorien kautta suorittamien signaalien perusteella. Tällaisia järjestelmiä on varmaan monia. Yksi niistä on se, että lihas- ja hermosolujen erilaistumista säätelevät neuroreguliinit, jotka ovat kalvoproteiineja, jotka toimivat yhden tai useamman tyrosiinikinaasireseptorin kautta.
Määrityksen geneettistä kontrollia osoittavat myös niin sanotut homeiotrooppiset tai homeoottiset mutaatiot, joiden on osoitettu hyönteisissä aiheuttavan muutoksia määrittelyssä tietyissä mielikuvituslevyissä. Tämän seurauksena jotkut kehon osat kehittyvät paikoilleen. Esimerkiksi Drosophilassa mutaatiot muuttavat antennilevyn määrityksen levyksi, joka kehittyy päästä ulottuvan raajan umpilisäkkeeksi. Suvun hyönteisissä Oftalmoptera silmälevystä voi kehittyä siipirakenteita. Hiirillä on osoitettu geeniklusterin (kompleksi) Hox olemassaolo, joka koostuu 38 geenistä ja ohjaa raajojen kehitystä.
Geenitoiminnan säätely alkion kehityksen aikana on itsenäinen merkitys. Uskotaan, että erilaistumisen aikana geenit toimivat eri aikoina, mikä ilmenee
erilaisten mRNA:iden transkriptiossa eri erilaistuneissa soluissa, ts. tapahtuu geenien repressiota ja derepressiota. Esimerkiksi merisiilin blastosyyteissä RNA:ksi transkriptoituneiden geenien määrä on 10 %, rotan maksasoluissa myös 10 % ja naudan kateenkorvan soluissa 15 %. Oletetaan, että ei-histoniproteiinit osallistuvat geenien transkription tilan säätelyyn. Seuraavat tiedot tukevat tätä oletusta. Kun solukromatiini on vaiheessa S kirjoitettu järjestelmään in vitro, sitten syntetisoidaan vain histoni-mRNA, jonka jälkeen histonit. Päinvastoin, kun käytetään β-vaiheen solujen kromatiinia, histoni-mRNA:ta ei syntetisoidu, kun ei-mäntäproteiinit poistetaan β-faasin kromatiinista ja korvataan ei-histonisilla kromosomiproteiineilla, jotka syntetisoidaan vaihe S, sitten sellaisen kromatiinin transkription jälkeen in vitro histoni-mRNA syntetisoidaan. Lisäksi kun ei-histoniproteiinit ovat peräisin 1-faasisista soluista ja DNA ja histonit tulevat S-vaiheen soluista, histoni-mRNA:ta ei syntetisoidu.Nämä tulokset osoittavat, että kromatiinin sisältämät ei-histoniproteiinit määräävät kyvyn transkriptoida histoneja koodaavia geenejä. Siksi uskomme, että ei-histonikromosomiproteiineilla voi olla tärkeä rooli geenien säätelyssä ja ilmentymisessä eukaryooteissa.
Saatavilla olevat tiedot viittaavat siihen, että proteiini- ja steroidihormonit osallistuvat transkription säätelyyn eläimillä. Proteiini (insuliini) ja steroidi (estrogon ja testosteroni) hormonit ovat kaksi signaalijärjestelmää, joita käytetään solujen välisessä viestinnässä. Korkeammissa eläimissä hormonit syntetisoidaan erikoistuneissa erityssoluissa. Vapautuessaan verenkiertoon ne pääsevät kudoksiin. Koska proteiinihormonien molekyylit ovat suhteellisen suuria, ne eivät tunkeudu soluihin, joten niiden vaikutukset saadaan aikaan kohdesolujen kalvoissa sijaitsevista reseptoriproteiineista ja syklisen AMP:n (cAMP) solunsisäisistä tasoista. Päinvastoin, steroidihormonit ovat pieniä molekyylejä, minkä seurauksena ne tunkeutuvat helposti soluihin kalvojen läpi. Kun ne ovat solujen sisällä, ne sitoutuvat spesifisiin reseptoriproteiineihin, joita löytyy vain kohdesolujen sytoplasmasta. Kompleksit otetaan huomioon hormoni-proteiinireseptori, keskittyessään kohdesolujen ytimiin, ne aktivoivat tiettyjen geenien transkription vuorovaikutuksessa tiettyjen
mi ei-histoniproteiineja, jotka sitoutuvat spesifisten geenien promoottorialueisiin. Näin ollen kompleksin sitoutuminen hormoni + proteiini (proteiinireseptori) ei-histoniproteiinien kanssa vapauttaa promoottorialueita RNA-polymeraasin liikkumista varten. Yhteenvetona saadut tiedot alkiojakson geneettisestä kontrollista organismien ontogeneesissä, voimme päätellä, että sen kulkua ohjaa geenien toiminnan erilainen päälle- ja poiskytkentä eri soluissa (kudoksissa) niiden derepression ja repression kautta.
POISTEMBRYONALINEN KEHITYS
Organismin syntymän jälkeen alkaa sen postembryonaalinen kehitys (ihmisillä postnataalinen), joka eri organismeissa kestää lajista riippuen useista päivistä satoihin vuosiin. Näin ollen elinajanodote on organismeille ominainen laji, joka ei riipu niiden organisaation tasosta (katso alla).
Postembryonisessa ontogeneesissä erotetaan nuoriso- ja murrosikä sekä kuolemaan päättyvä vanhuuden aika.
Nuorten aikakausi. Tämä ajanjakso (lat. juvenilis- nuori) määräytyy ajan organismin syntymästä murrosikään. Se tapahtuu eri tavoin ja riippuu organismien ontogeneesin tyypistä. Tälle ajanjaksolle on ominaista joko suora tai epäsuora kehitys.
Suoran kehityksen omaavien eliöiden (monet selkärangattomat, kalat, matelijat, linnut, nisäkkäät, ihmiset) tapauksessa munankuoresta kuoriutuneet tai vastasyntyneet ovat aikuisten muotojen kaltaisia ja eroavat jälkimmäisistä vain pienemmissä koossa. yksittäisten elinten alikehittyneisyys ja epätäydelliset ruumiinmittasuhteet (kuva 30).
Suoran kehityksen alaisten organismien nuoruuden kasvulle ominaista on, että solujen lukumäärä ja koko lisääntyvät ja kehon mittasuhteet muuttuvat. Ihmisen kasvu hänen ontogeneesinsä eri ajanjaksoina on esitetty kuvassa. 31. Ihmisen eri elinten kasvu on epätasaista. Esimerkiksi pään kasvu päättyy lapsuudessa, jalat saavuttavat suhteellisen koon noin 10 vuotta. Ulkoiset sukuelimet kasvavat erittäin nopeasti 12-14-vuotiaana. Tehdään ero selvän ja määrittelemättömän kasvun välillä. Tietty kasvu on ominaista organismeille, jotka lopettavat kasvun tietyssä iässä,
esimerkiksi hyönteiset, nisäkkäät, ihmiset. Epämääräinen kasvu on ominaista organismeille, jotka kasvavat koko elämänsä ajan, esimerkiksi nilviäisille, kaloille, sammakkoeläimille, matelijoille ja monille kasvilajeille.

Riisi. kolmekymmentä.Eri lajien organismien suora ja epäsuora kehitys
Epäsuoran kehityksen tapauksessa organismit käyvät läpi muunnoksia, joita kutsutaan metamorfooseiksi (lat. metamorfoosi - muunnos).

Riisi. 31.Kasvu ja kehitys ihmisen ontogeneesin eri aikoina
Ne edustavat organismien muutoksia kehityksen aikana. Metamorfoosit löytyvät laajalti coelenteraateista (hydra, meduusat, korallipolyypit), lattamatoista (fasciola), sukkulamadoista (syöksymadoista), nilviäisistä (osterit, simpukat, mustekalat), niveljalkaisista (ravut, jokiravut, hummerit, katkaravut, spiders, punkki , hyönteiset) ja jopa joissakin sointuissa (vaippaeläimet ja sammakkoeläimet). Tässä tapauksessa erotetaan täydelliset ja epätäydelliset metamorfoosit. Ilmeisimpiä metamorfoosin muotoja havaitaan hyönteisissä, jotka käyvät läpi sekä epätäydellisen että täydellisen metamorfoosin.
Epätäydellinen transformaatio on kehitys, jossa munankuorista tulee esiin organismi, jonka rakenne on samanlainen kuin aikuisen organismin rakenne, mutta sen koko on paljon pienempi. Tällaista organismia kutsutaan toukkaksi. Kasvu- ja kehitysprosessin aikana toukkien koko kasvaa, mutta olemassa oleva kitinoitu peite estää kehon koon lisääntymisen edelleen, mikä johtaa moltingiin eli kitinoidun peitteen irtoamiseen, jonka alla on pehmeä kynsinauho. Jälkimmäinen suoristuu, ja tähän liittyy eläimen koon kasvu. Useiden sulkien jälkeen eläin saavuttaa kypsyyden. Epätäydellinen transformaatio on tyypillistä esimerkiksi lutikoiden kehittymiselle.
Täydellinen metamorfoosi on kehitys, jossa munankuorista vapautuu toukka, joka on merkittävästi erilainen
rakenteessa aikuisilta yksilöiltä. Esimerkiksi perhosissa ja monissa hyönteisissä toukat ovat toukkia. Toukat ovat alttiina sulamiselle ja voivat sulaa useita kertoja ja muuttua sitten pupuiksi. Jälkimmäisistä kehittyy aikuisia muotoja (imago), jotka eivät eroa alkuperäisistä.
Selkärankaisilla metamorfoosia esiintyy sammakkoeläinten ja luisten kalojen keskuudessa. Toukkavaiheelle on ominaista väliaikaisten elinten läsnäolo, jotka joko toistavat esi-isien ominaisuudet tai joilla on selkeä adaptiivinen merkitys. Esimerkiksi nuijapäiselle, joka on sammakon toukkamuoto ja toistaa alkuperäisen muodon ominaisuuksia, on ominaista kalamainen muoto, kidusten hengittäminen ja yksi verenkiertokierros. Nuijapäiden mukautuvat ominaisuudet ovat niiden imevät ja pitkät suolet. Toukkamuodoille on myös ominaista, että ne ovat aikuisiin muotoihin verrattuna sopeutuneet elämään täysin erilaisissa olosuhteissa, eri ekologisesti ja eri paikassa ravintoketjussa. Esimerkiksi sammakon toukilla on kidusten hengitys, kun taas aikuisilla muodoilla on keuhkohengitys. Toisin kuin aikuiset muodot, jotka ovat lihansyöjiä, sammakon toukat syövät kasviperäisiä ruokia.
Organismien kehityksen tapahtumasarjaa kutsutaan usein elinkaareksi, jotka voivat olla yksinkertaisia tai monimutkaisia. Yksinkertaisimmat kehityssyklit ovat tyypillisiä esimerkiksi nisäkkäille, kun hedelmöittyneestä munasta kehittyy organismi, joka taas tuottaa munia jne. Monimutkaiset biologiset syklit ovat eläimissä kiertokulkuja, joille on ominaista kehitys ja muodonmuutos. Biologisten kiertokulkujen tuntemus on käytännönläheistä, erityisesti tapauksissa, joissa organismit ovat patogeenejä tai patogeenien kantajia eläimissä ja kasveissa.
Metamorfoosiin liittyvä kehitys ja erilaistuminen ovat seurausta luonnollisesta valinnasta, jonka ansiosta monet toukkamuodot, kuten hyönteistoukat ja sammakkopatikka, ovat paremmin sopeutuneet ympäristöön kuin aikuiset sukukypsät muodot.
Murrosikä. Tätä ajanjaksoa kutsutaan myös kypsäksi, ja se liittyy organismien sukukypsyyteen. Organismien kehitys tänä aikana saavuttaa maksiminsa.
Sikiönjälkeisen ajanjakson kasvuun ja kehitykseen vaikuttavat suuresti ympäristötekijät. Kasveille ratkaisevia tekijöitä ovat valo, kosteus, lämpötila, maaperän ravinteiden määrä ja laatu. Eläimille oikea ruokinta on ensiarvoisen tärkeää (proteiinien, hiilihydraattien, lipidien, mineraalisuolojen, vitamiinien, hivenaineiden läsnäolo rehussa). Myös happi, lämpötila, valo (D-vitamiinin synteesi) ovat tärkeitä.
Eläinorganismien kasvu ja yksilöllinen kehitys ovat neurohumoraalisen säätelyn alaisia humoraalisten ja hermostollisten säätelymekanismien avulla. Kasveista on löydetty hormonin kaltaisia vaikuttavia aineita, joita kutsutaan fytohormoneiksi. Jälkimmäiset vaikuttavat kasviorganismien elintärkeisiin toimintoihin.
Eläinsoluissa syntetisoituu niiden elinprosessien aikana kemiallisesti aktiivisia aineita, jotka vaikuttavat elämänprosesseihin. Selkärangattomien ja selkärankaisten hermosolut tuottavat aineita, joita kutsutaan neurosekreteiksi. Endokriiniset eli sisäiset eritysrauhaset erittävät myös hormoneiksi kutsuttuja aineita. Endokriiniset rauhaset, erityisesti kasvuun ja kehitykseen liittyvät rauhaset, säätelevät neuroerityksiä. Niveljalkaisten kasvun ja kehityksen säätelyä havainnollistaa hyvin hormonien vaikutus sulamiseen. Solujen toukkien eritteiden synteesiä säätelevät aivoihin kerääntyvät hormonit. Äyriäisten erityinen rauhanen tuottaa hormonia, joka estää moldingin. Näiden hormonien tasot määräävät sulamisen tiheyden. Hyönteisillä munasolujen kypsymisen ja diapausin hormonaalinen säätely on todettu.
Selkärankaisilla endokriiniset rauhaset ovat aivolisäke, käpylisäke, kilpirauhanen, lisäkilpirauhanen, haima, lisämunuaiset ja sukurauhaset, jotka ovat tiiviisti yhteydessä toisiinsa. Selkärankaisten aivolisäke tuottaa gonadotrooppista hormonia, joka stimuloi sukurauhasten toimintaa. Ihmisillä aivolisäkkeen hormoni vaikuttaa kasvuun. Puutteella kehittyy kääpiö, ylimäärällä gigantismi. Käpyrauhanen tuottaa hormonia, joka vaikuttaa eläinten seksuaalisen aktiivisuuden vuodenaikojen vaihteluihin. Kilpirauhashormoni vaikuttaa hyönteisten ja sammakkoeläinten metamorfoosiin. Nisäkkäillä kilpirauhasen vajaakehitys johtaa kasvun hidastumiseen ja sukuelinten alikehittymiseen. Ihmisillä kilpirauhasen vajaatoiminnan vuoksi luutuminen ja kasvu viivästyvät.
(kääpiö), murrosikää ei tapahdu, henkinen kehitys pysähtyy (kretinismi). Lisämunuaiset tuottavat hormoneja, jotka vaikuttavat aineenvaihduntaan, kasvuun ja solujen erilaistumiseen. Sukurauhaset tuottavat sukupuolihormoneja, jotka määrittävät toissijaiset seksuaaliset ominaisuudet. Sukurauhasten poistaminen johtaa peruuttamattomiin muutoksiin useissa ominaisuuksissa. Esimerkiksi kastroiduilla kukoilla kamman kasvu pysähtyy ja sukupuolivaisto menetetään. Kastroitu mies saa ulkoisen samankaltaisuuden naisen kanssa (parta ja karvat eivät kasva iholle, rasvaa kertyy rintakehälle ja lantion alueelle, äänen sointi säilyy jne.).
Kasvien fytohormonit ovat auksiinit, sytokiniinit ja gibberelliinit. Ne säätelevät solujen kasvua ja jakautumista, uusien juurien muodostumista, kukkien kehitystä ja muita kasvien ominaisuuksia.
Kaikilla ontogeneesin jaksoilla organismit pystyvät palauttamaan kadonneita tai vaurioituneita ruumiinosia. Tätä organismien ominaisuutta kutsutaan uudistuminen, jotka voivat olla fysiologisia ja korjaavia.
Fysiologinen regeneraatio - Tämä on kadonneiden ruumiinosien korvaaminen kehon elinkaaren aikana. Tämän tyyppiset regeneraatiot ovat hyvin yleisiä eläinmaailmassa. Esimerkiksi niveljalkaisissa sitä edustaa sulaminen, joka liittyy kasvuun. Matelijoilla uusiutuminen ilmaistaan hännän ja suomujen korvaamisena, linnuilla - höyhenillä, kynsillä ja kannuilla. Nisäkkäillä esimerkki fysiologisesta uusiutumisesta on hirven vuotuinen sarvien irtoaminen.
Korjaava regeneraatio - Tämä on väkivaltaisesti repeytyneen organismin ruumiinosan ennallistaminen. Tämän tyyppinen uusiutuminen on mahdollista monilla eläimillä, mutta sen ilmenemismuodot vaihtelevat. Se on yleinen esimerkiksi hydroissa ja liittyy jälkimmäisen lisääntymiseen, koska koko organismi uusiutuu osasta. Muissa organismeissa regeneraatio ilmenee yksittäisten elinten kyvynä toipua minkä tahansa osan menettämisen jälkeen. Ihmisellä epiteeli-, side-, lihas- ja luukudoksilla on melko korkea regeneraatiokyky.
Monien lajien kasvit pystyvät myös uusiutumaan.
Regeneraatiotiedot ovat erittäin tärkeitä paitsi biologiassa. Niitä käytetään laajalti maataloudessa, lääketieteessä, erityisesti kirurgiassa.
Vanhuus ontogeneesin vaiheena. Vanhuus on eläimen ontogeneesin toiseksi viimeinen vaihe, ja sen kesto määräytyy
kokonaiselinikä, joka toimii lajin ominaispiirteenä ja vaihtelee eri eläinten välillä. Vanhuutta on tutkittu tarkimmin ihmisillä.
Ihmisen vanhuudelle on olemassa erilaisia määritelmiä. Erityisesti yksi suosituimmista määritelmistä on, että vanhuus on peräkkäisten muutosten kertymistä, joka liittyy organismin iän nousuun ja lisää sen sairauden tai kuoleman todennäköisyyttä. Tiedettä ihmisen ikääntymisestä kutsutaan gerontologiaksi (kreikaksi. geron - vanha mies, vanha mies, logot - Tiede). Sen tehtävänä on tutkia iän siirtymisen malleja kypsyyden ja kuoleman välillä.
Gerontologian tieteellinen tutkimus ulottuu useille alueille, solujen entsyymien aktiivisuuden muutosten tutkimuksista ympäristöstressin psykologisten ja sosiologisten lievennysten vaikutuksen selvittämiseen vanhusten käyttäytymiseen.
Ihmisten tapauksessa erotetaan fysiologinen vanhuus; kalenteriikään liittyvä vanhuus; sosiaalisten tekijöiden ja sairauksien aiheuttama ennenaikainen ikääntyminen. WHO:n suositusten mukaan iäkkääksi tulee katsoa noin 60-75-vuotias ja vanhaksi 75-vuotias tai vanhempi.
Ihmisen vanhuudelle on ominaista useita ulkoisia ja sisäisiä merkkejä.
Vanhuuden ulkoisista merkeistä havaittavimmat ovat liikkeiden sujuvuuden heikkeneminen, asennon muutokset, ihon kimmoisuuden, kehon painon, lihasten kiinteyden ja kimmoisuuden heikkeneminen, ryppyjen ilmaantuminen kasvoille ja muille ihon osille. kehon ja hampaiden menetys. Joten esimerkiksi yleisten tietojen mukaan henkilö menettää 30-vuotiaana 2 hammasta (menetyksen seurauksena), 40-vuotiaana - 4 hammasta, 50-vuotiaana - 8 hammasta ja 60-vuotiaana - jo 11 hammasta. Ensimmäisessä signalointijärjestelmässä tapahtuu havaittavia muutoksia (aistielinten tarkkuus on tylsistynyt). Esimerkiksi suurin etäisyys, jolla terveet ihmiset erottavat tietyt identtiset äänet, on 12 m 20-30-vuotiaana, 10 m 50-vuotiaana, 7 m 60-vuotiaana ja vain 4 m 70-vuotiaana. toinen merkinantojärjestelmä (puheen intonaatio muuttuu, ääni muuttuu tylsäksi).
Sisäisistä merkeistä on ensinnäkin mainittava sellaiset merkit kuin elinten käänteinen kehitys (involuutio). Maksan ja munuaisten koko sekä lukumäärä pienenee
munuaisten nefronien määrä (80-vuotiaana lähes puolet), mikä heikentää munuaisten toimintaa ja vaikuttaa vesi-elektrolyyttiaineenvaihduntaan. Verisuonten elastisuus heikkenee, kudosten ja elinten verenkierto heikkenee ja perifeerinen verisuonivastus kasvaa. Epäorgaaniset suolat kerääntyvät luihin, rusto muuttuu (kalkkiutuu) ja elinten uusiutumiskyky heikkenee. Soluissa tapahtuu merkittäviä muutoksia, niiden jakautuminen ja toiminnallisen sävyn palautuminen hidastuu, vesipitoisuus laskee, soluentsyymien aktiivisuus vähenee, assimilaation ja dissimilaation välinen koordinaatio häiriintyy. Aivoissa proteiinisynteesi häiriintyy, mikä johtaa epänormaalien proteiinien muodostumiseen. Solukalvojen viskositeetti kasvaa, sukupuolihormonien synteesi ja hyötykäyttö häiriintyvät ja hermosolujen rakenteessa tapahtuu muutoksia. Sidekudoksen proteiineissa ja tämän kudoksen elastisuudessa tapahtuu rakenteellisia muutoksia. Immunologiset reaktiot heikkenevät ja autoimmuunireaktioiden mahdollisuus kasvaa. Endokriinisten järjestelmien, erityisesti sukurauhasten, toiminta heikkenee. Muiden merkkien käyttäytyminen vanhuudessa on esitetty kuvassa. 32.

Riisi. 32.Muutokset joissakin ihmisen ominaisuuksissa iän myötä: 1 - hermoimpulssien nopeus; 2 - perusaineenvaihdunnan taso; 3 - sydänindeksi; 4 - insuliinin munuaissuodatuksen taso; 5 - keuhkojen vuorovesitilavuudet; 6 - plasmavirtauksen taso munuaisissa
Halu ymmärtää kehon ikääntymisen luonne on ollut olemassa jo pitkään. Muinaisessa Kreikassa Hippokrates uskoi, että ikääntyminen liittyi ylimääräiseen ruokaan ja riittämättömään raittiiseen ilmaan. Aristoteleen mukaan ikääntyminen johtuu kehon lämpöenergian kulutuksesta. Ruoan merkityksen ikääntymiseen vaikuttavana tekijänä totesi myös Galen. Mutta pitkään aikaan ei ollut tarpeeksi tieteellistä tietoa tämän ongelman objektiiviseen ymmärtämiseen. Vasta 1800-luvulla. Ikääntymisen tutkimuksessa on edistytty jonkin verran, ja ikääntymisen teorioita on alkanut ilmaantua.
Yksi ensimmäisistä tunnetuimmista ihmiskehon ikääntymisen teorioista on saksalaisen lääkärin H. Hufelandin (1762-1836) teoria, joka piti pitkäikäisyydessä tärkeänä työvoimaa. Olemme kuulleet hänen lausuntonsa, ettei yksikään laiska ole elänyt vanhuuteen. Vielä kuuluisampi endokriininen ikääntymisen teoria, joka on peräisin Bertholdin (1849) vielä vuosisadan puolivälissä suorittamista kokeista, jotka osoittivat, että kivesten siirtämiseen eläimestä toiseen liittyy toissijaisten sukupuoliominaisuuksien kehittyminen. Myöhemmin ranskalainen fysiologi C. Brown-Séquard (1818-1894) väitti, että näillä injektioilla on hyödyllinen ja nuorentava vaikutus. 1900-luvun alussa. Jo nyt uskotaan, että vanhuuden alkaminen liittyy endokriinisten rauhasten, erityisesti sukurauhasten, toiminnan loppumiseen. 20-30 luvulla. XX vuosisadalla Tämän uskomuksen perusteella eri maissa on tehty monia leikkauksia vanhusten tai vanhusten nuorentamiseksi. Esimerkiksi G. Steinach Itävallassa sitoi miesten siittiönauhat, mikä johti sukurauhasten ulkoisen erittymisen lakkaamiseen ja oletettavasti nuorentumiseen. S.A. Voronov Ranskassa siirsi kiveksiä nuorista eläimistä vanhoille ja apinoista miehille, ja Tushnov Neuvostoliitossa nuorensi kukkoja ruiskuttamalla niihin sukurauhasten histolysaattia. Kaikki nämä toimet johtivat joihinkin vaikutuksiin, mutta vain väliaikaisiin. Näiden vaikutusten jälkeen ikääntymisprosessi jatkui ja vieläkin voimakkaammin.
1900-luvun alussa. nousi mikrobiologinen ikääntymisen teoria, jonka luoja oli I.I. Mechnikov, joka erotti fysiologisen ja patologisen vanhuuden. Hän uskoi, että ihmisen vanhuus on patologista, ts. ennenaikaista. I.I:n ajatusten perusta Mechnikovin oppi ortobioosista (ortos -
oikea, bios - elämä), jonka mukaan pääasiallinen ikääntymisen syy on paksusuolen mätänemisen seurauksena muodostuneiden myrkytystuotteiden vaurioituminen hermosoluille. Normaalin elämäntavan opin kehittäminen (hygieniasääntöjen noudattaminen, säännöllinen työ, pidättäytyminen huonoista tavoista), I.I. Mechnikov ehdotti myös tapaa tukahduttaa mädäntyneitä suoliston bakteereja kuluttamalla fermentoituja maitotuotteita.
30-luvulla XX vuosisadalla tuli laajalle levinneeksi teoria keskushermoston (CNS) roolista ikääntymisessä. Tämän teorian luoja on I.P. Pavlov, joka totesi keskushermoston integroivan roolin organismien normaalissa toiminnassa. I.P:n seuraajat Pavlovin eläinkokeet osoittivat, että ennenaikainen ikääntyminen johtuu hermoshokit ja pitkittynyt hermoston ylikuormitus.
Mainitsemisen arvoinen teoria ikään liittyvistä muutoksista sidekudoksessa, muotoiltu 30-luvulla. XX vuosisadalla A.A. Bogomolets (1881-1946). Hän uskoi, että sidekudos (luukudos, rusto, jänteet, nivelsiteet ja sidekudos) varmistaa kehon fysiologisen toiminnan ja että solujen kolloidisen tilan muutokset, niiden turgorin menetys jne. määräävät ikään liittyviä muutoksia. organismeissa. Nykyaikaiset tiedot osoittavat kalsiumin kertymisen tärkeyden sidekudoksiin, koska se myötävaikuttaa sen elastisuuden menettämiseen sekä verisuonten kovettumiseen.
Nykyaikaisille lähestymistavoille ikääntymisen olemuksen ja mekanismien ymmärtämiseksi on ominaista fysikaalis-kemiallisen biologian tiedon laaja käyttö ja erityisesti molekyyligenetiikan saavutukset. Yleisimmät nykyaikaiset ajatukset ikääntymisen mekanismista kiteytyvät siihen, että elämän aikana kehon soluihin kertyy somaattisia mutaatioita, joiden seurauksena tapahtuu viallisten proteiinien synteesi tai korjaamattomien DNA-ristisidosten proteiinien kanssa. Koska viallisilla proteiineilla on hajottava rooli solujen aineenvaihdunnassa, tämä johtaa ikääntymiseen. Viljeltyjen fibroblastien tapauksessa vanhoihin soluihin liittyvien proteiinien ja mRNA:iden on osoitettu suppressoivan DNA-synteesiä nuorissa fibroblasteissa.
On myös tunnettu hypoteesi, jonka mukaan ikääntymisen katsotaan johtuvan mitokondrioiden aineenvaihduntatuotteiden muutoksista, joita seuraa entsyymien toimintahäiriö.
Ihmisillä on osoitettu geenien olemassaolo, jotka määräävät ikääntymiseen liittyvien perinnöllisten rappeutumisprosessien kehittymisen ajoituksen. Useat tutkijat uskovat, että ikääntymisen syynä ovat muutokset kehon immunologisessa puolustusjärjestelmässä, erityisesti autoimmuunireaktioissa kehon rakenteisiin, jotka ovat elintärkeitä. Lopuksi, selittäessään ikääntymisen mekanismeja, asiantuntijat kiinnittävät suurta huomiota vapaiden radikaalien muodostumiseen liittyviin proteiinivaurioihin. Lopuksi joskus pidetään tärkeänä lysosomien hajoamisen jälkeen vapautuvia hydrolaaseja, jotka tuhoavat soluja.
Kattavaa teoriaa ikääntymisestä ei kuitenkaan ole vielä luotu, koska on selvää, että mikään näistä teorioista ei pysty itsenäisesti selittämään ikääntymisen mekanismeja.
Kuolema.Kuolema on ontogeneesin viimeinen vaihe. Kysymys kuolemasta on biologiassa erityisen tärkeällä sijalla, koska kuoleman tunne "...on täysin vaistomainen ihmisluonnossa ja on aina ollut yksi ihmisen suurimmista huolenaiheista" (I.I. Mechnikov, 1913). Lisäksi kysymys kuolemasta oli ja on kaikkien filosofisten ja uskonnollisten opetusten huomion keskipisteessä, vaikka kuolemanfilosofia esitettiin eri tavalla eri historiallisina aikoina. Muinaisessa maailmassa Sokrates ja Platon väittivät sielun kuolemattomuuden puolesta, kun taas Aristoteles kielsi Platonin käsityksen sielun kuolemattomuudesta ja uskoi ihmishengen kuolemattomuuteen, joka jatkaa elämäänsä ihmisen kuoleman jälkeen. Myös Cicero ja Seneca tunnustivat tulevan elämän, mutta Marcus Aurelius piti kuolemaa luonnonilmiönä, joka pitäisi hyväksyä valittamatta. 1700-luvulla Myös I. Kant ja I. Fichte (1762-1814) uskoivat tulevaan elämään, ja G. Hegel piti kiinni siitä uskomuksesta, että sielu imeytyy "absoluuttiseen olentoon", vaikka tämän "olennon" luonnetta ei paljastettu. .
Kaikkien tunnettujen uskonnollisten opetusten mukaisesti ihmisen maallinen elämä jatkuu hänen kuolemansa jälkeen, ja ihmisen on väsymättä valmistauduttava tähän tulevaan kuolemaan. Kuitenkin luonnontieteilijät ja filosofit, jotka eivät tunnusta kuolemattomuutta, uskoivat ja uskovat edelleen, että kuolema on, kuten I.I. on toistuvasti korostanut. Mechnikov, eliön elämän luonnollinen tulos. Kuvannollisempi määritelmä kuolemasta on, että se "...on selvä merkityksettömyyden voitto merkityksestä, kaaos avaruudesta" (V. Solovjov, 1894).
Tieteelliset todisteet viittaavat siihen, että yksisoluisissa organismeissa (kasveissa ja eläimissä) kuolema olisi erotettava lopettamisesta.
tietoa heidän olemassaolostaan. Kuolema on heidän tuhonsa, kun taas olemassaolon lakkaaminen liittyy heidän jakautumiseensa. Näin ollen yksisoluisten organismien hauraus kompensoituu niiden lisääntymisellä. Monisoluisissa kasveissa ja eläimissä kuolema on sanan täydessä merkityksessä eliön elämän loppua.
Ihmisillä kuoleman todennäköisyys kasvaa murrosiän aikana. Erityisesti kehittyneissä maissa kuoleman todennäköisyys kasvaa lähes eksponentiaalisesti 28 vuoden iän jälkeen.
On olemassa ihmisen kliininen ja biologinen kuolema. Kliininen kuolema ilmaistaan tajunnan menetyksenä, sydämen sykkeen ja hengityksen lakkaamisena, mutta useimmat solut ja elimet pysyvät edelleen hengissä. Solut uusiutuvat itsestään ja suoliston peristaltiikka jatkuu. Kliininen kuolema ei "päästä" biologista kuolemaa, koska se on palautuva, koska kliinisen kuoleman tilassa voi "palata" elämään. Esimerkiksi koirat "palautuvat" elämään 5-6 minuutin kuluttua, ihmiset - 6-7 minuutin kuluttua kliinisen kuoleman alkamisesta. Biologiselle kuolemalle on ominaista se, että se on peruuttamaton. Sydämen lyönnin ja hengityksen pysähtymiseen liittyy itseuudistumisprosessien pysähtyminen, solujen kuolema ja hajoaminen. Solukuolema ei kuitenkaan ala kaikissa elimissä yhtä aikaa. Ensin kuolee aivokuori, sitten suoliston, keuhkojen, maksan, lihassolujen ja sydämen epiteelisolut.
Toimenpiteet eliöiden elvyttämiseksi (elvytykseen) perustuvat ajatukseen kliinisestä kuolemasta, jolla on poikkeuksellisen tärkeä merkitys nykylääketieteessä.
ELÄMÄ
Kasviston ja eläimistön edustajien elinajanodotetietojen vertailu osoittaa, että kasveilla ja eläimillä eri organismit elävät eri aikoja. Esimerkiksi ruohokasvit (luonnonvaraiset ja viljellyt) elävät yhden kauden. Päinvastoin, niiden puumaisille kasveille on ominaista pidempi elinikä. Esimerkiksi kirsikka elää 100 vuotta, kuusi - 1000 vuotta, tammi -
2000 vuotta, mänty - jopa 2000-4000 vuotta.
Monien lajien kalat elävät 55-80 vuotta, sammakot - 16 vuotta, krokotiilit - 50-60 vuotta, joidenkin lajien linnut - jopa 100 vuotta. Nisäkkäiden elinikä on lyhyempi.
Esimerkiksi pienet karjat elävät 20-25 vuotta, karja - 30 vuotta tai enemmän, hevoset - jopa 30 vuotta, koirat - 20 vuotta tai enemmän, sudet - 15 vuotta, karhut - 50 vuotta, norsut - 100 vuotta. Nisäkkäistä ihmiset elävät pisimpään. Monet ihmiset elivät 115-120-vuotiaaksi tai enemmän.
Olemassa olevien käsitysten mukaisesti elinajanodote on lajikohtainen määrällinen ominaisuus, joka on genotyypin hallinnassa. Uskotaan, että geneettisesti määrätty elinajanodote korreloi ontogeneesijaksojen kanssa (taulukko 5).
Taulukko 5.Fibroblastien tason riippuvuus viljelyn aikana luovuttajan iästä
Oletetaan, että lajin elinikä on lajin evoluution mukainen hankinta. Mitä tulee yksilöiden pitkäikäisyyteen, se voidaan selittää olettamalla tiettyjen geenien yhdistelmien läsnäoloa heidän genotyypeissään tai pienellä määrällä tai täydellisellä mutaatioiden puuttumisella heidän soluissaan.
A.A. Bogomolets ja I.I. Schmalhausen laski, että ihmisen luonnollisen eliniän odotteen tulisi olla 120-150 vuotta. Kuitenkin vain harvat selviävät tähän ikään. Siksi todellinen elinajanodote ei ole sama kuin luonnollinen elinajanodote.
Keskimääräiseen eliniän odotteeseen vaikuttavat useat tekijät (lastekuolleisuuden väheneminen, infektioiden hallinnan tehokkuus, kirurgian edistyminen, ravitsemuksen ja yleisten elinolojen paraneminen). Tärkeimmät syyt eliniän lyhenemiseen ovat nälkäkuolleisuus, sairaudet ja riittämätön sairaanhoito. Venäjällä on viime vuosina havaittu negatiivinen väestönkasvu kaikissa Venäjän kaupungeissa ja maaseudulla johtuen syntyvyyden laskusta ja korkeasta kuolleisuudesta.
Lääketieteessä elinajanodote on kansakunnan terveyden indikaattori. Neuvostoliitto sijoittui ensimmäisellä sijalla maailmassa vanhusten lukumäärällä mitattuna. Esimerkiksi miljoonaa asukasta kohden on 104 yli 90-vuotiasta, kun taas Englannissa - 6, Ranskassa - 7
ja USA - 15 henkilöä. Tällä hetkellä työikäisen väestön rajoissa on tapahtunut muutos esimerkiksi 30-luvulle verrattuna. XX vuosisadalla Myös ero eläkeiän ja ihmisten aktiivisuustason välillä vaihtelee. Vuonna 1982 Wienissä pidettiin maailman väestön ikääntymisongelmia käsittelevä maailmankokous, jossa asiasta laadittiin ennusteita vuoteen 2025 asti. Oletetaan, että 60-vuotiaiden ja sitä vanhempien määrä kasvaa viisinkertaiseksi verrattuna vuoteen 1950, yli 80-vuotiaat - 7 kertaa. Tämän kansainvälisen foorumin mukaan väestön ikääntymisvauhdissa on eroja eri kansojen, maiden ja alueiden välillä.
Geriatria on tiede, jonka tehtävänä on kehittää tapoja normalisoida ikääntyvän kehon muuttuvia toimintoja. Nykylääketieteessä ei kuitenkaan vielä ole menetelmiä ja keinoja vaikuttaa normaaleihin fysiologisiin prosesseihin. Siksi geriatrian rooli rajoittuu vanhuudessa syntyvien sairauksien hoitoon ja ennenaikaista ikääntymistä aiheuttavien riskitekijöiden eliminoimiseen (jos mahdollista).
KESKUSTELUA KOSKEVAT KYSYMYKSET
1. Mitä ymmärrät organismien seksuaalisella lisääntymisellä ja mikä on sen biologinen rooli?
2. Kuvaile aseksuaalista lisääntymistä ja nimeä sen muodot.
3. Kuvaile sukupuoliprosessin piirteitä yksi- ja monisoluisissa organismeissa.
4. Mikä on gametogeneesi?
5. Mikä on kunkin sukusolutyypin tehtävä?
6. Mitä vaiheita sukusolujen kehityksessä tiedät?
7. Mitä yhtäläisyyksiä ja eroja spermatogeneesin ja oogeneesin välillä on?
8. Mitä meioosi on ja mikä on sen biologinen merkitys?
9. Kuvaile meioosin vaiheita.
10. Vaikuttaako ylitys meioosin tuloksiin ja miten?
11. Kuvaile lannoituksen olemusta.
12. Mitä eroa on tsigogeneesillä ja partenogeneesillä?
13. Mikä on vaihtelevan haploidisuuden ja diploidisuuden biologinen rooli?
14. Mikä on seksuaalinen dimorfismi? Mitä ymmärrät hermafroditismilla? Onko ihmisillä havaittu hermafroditismia ja kuinka usein?
15. Millaisena kuvittelet lisääntymismenetelmien kehityksen?
16. Mitä ymmärrät organismien kasvulla ja kehityksellä? Mikä on solujen kasvun ja erilaistumisen välinen suhde?
17. Mikä on solujen erilaistumisen molekyyliperusta? Mitä tiedät kantasoluista?
18. Muotoile ontogeneesin käsite ja nimeä ontogeneesin jaksot.
19. Mitä eroa on suoralla ja epäsuoralla kehittämisellä?
20. Mikä on hedelmöityksen vaikutus munasoluihin?
21. Missä geneettisen tiedon toteutusvaiheessa geenien toiminnan ohjaus tapahtuu?
22. Miten hedelmöittyneestä munasolusta kehittyy monisoluinen rakenne?
23. Miten kehittyvät solut ja kudokset eroavat toisistaan kehityksen aikana?
24. Voidaanko kadonneita tai vaurioituneita ruumiinosia palauttaa?
25. Mikä yhteys ontogeneesissä on vanhuuden ja elinajanodotteen välillä?
26. Muotoile ja määritä luonnollisen (todennäköisen) ja todellisen elinajanodotteen välinen ero.
27. Mitä teorioita organismien ikääntymisestä tiedät?
29. Mitkä tekijät vaikuttavat elinajanodotteeseen?
30. Mihin organismien kloonaus perustuu? Anna esimerkkejä kloonaavista organismeista.
31. Läpäiseekö siittiö tai sen sisältö solun läpi?
32. Mitä kloonaustiedot osoittavat?
33. Preformationismi ja epigeneesi biologian historiassa. Onko niillä tieteellistä merkitystä?
34. Mitä eroja on kliinisen ja biologisen kuoleman välillä?
Lisääntyminen on eliöiden kykyä muodostaa omaa lajiaan. Lisääntyminen on yksi elämän tärkeimmistä ominaisuuksista, ja se on mahdollista organismien yleisen kyvyn tuottaa jälkeläisiä ansiosta. Välittömät jälkeläiset eivät kuitenkaan aina ole vanhempiensa kaltaisia. Esimerkiksi saniaisten itiöistä kasvaa lukuisia jälkeläisiä, joita edustavat versot, jotka eivät ole samanlaisia kuin emo-itiöitä kantava kasvi. Uloskasvuun ilmestyy puolestaan itsestä poikkeava kasvi - sporofyytti. Tätä ilmiötä kutsutaan sukupolvien vuorotteluksi.
Erityinen seksuaalisen lisääntymisen muoto on partenogeneesi eli neitseellinen lisääntyminen eli organismin kehittyminen hedelmöittämättömästä munasolusta. Tämä lisääntymismuoto on tyypillistä pääasiassa lajeille, joiden elinkaari on lyhyt, ja vuodenaikojen vaihtelut ovat voimakkaat. Partenogeneesi voi olla haploidista tai diploidista. Haploidisessa (generatiivisessa) partenogeneesissä haploidisesta munasta kehittyy uusi organismi. Tuloksena olevat yksilöt voivat olla vain miehiä, vain naisia tai molempia. Tämä riippuu kromosomaalisen sukupuolen määrityksestä.
Organismin yksilöllinen kehitys eli ontogeneesi on joukko peräkkäisiä morfologisia, fysiologisia ja biokemiallisia muutoksia, jotka organismi käy läpi sen syntymähetkestä kuolemaan. Ontogeneesin aikana tapahtuu perinnöllisen tiedon täytäntöönpano, jonka keho on vastaanottanut vanhemmiltaan. Ontogeneesissä on kaksi pääjaksoa - alkio ja postembryonaalinen. Alkiovaiheessa eläimissä muodostuu alkio, jossa muodostuvat pääelinjärjestelmät. Postembryonaalisella kaudella kehitysprosessit päättyvät, murrosikää, lisääntymistä, ikääntymistä ja kuolemaa tapahtuu.
