1. განმარტეთ გენეტიკური ინფორმაციის გადაცემის თანმიმდევრობა: გენი – ცილა – თვისება.
2. დაიმახსოვრეთ რომელი ცილის სტრუქტურა განაპირობებს მის აგებულებას და თვისებებს. როგორ არის ეს სტრუქტურა კოდირებული დნმ-ის მოლეკულაში?
3. რა არის გენეტიკური კოდი?
4. აღწერეთ გენეტიკური კოდის თვისებები.
7. მატრიცის სინთეზის რეაქციები. ტრანსკრიფცია
ცილის შესახებ ინფორმაცია ჩაწერილია როგორც ნუკლეოტიდური თანმიმდევრობა დნმ-ში და მდებარეობს ბირთვში. სინამდვილეში ციტოსინთეზი ხდება ციტოპლაზმაში რიბოზომებზე. აქედან გამომდინარე, ცილის სინთეზს სჭირდება სტრუქტურა, რომელიც გადაიტანს ინფორმაციას დნმ-დან ცილის სინთეზის ადგილზე. ასეთი შუამავალი არის ინფორმაცია, ანუ მატრიცა, რნმ, რომელიც გადასცემს ინფორმაციას დნმ-ის მოლეკულის კონკრეტული გენიდან რიბოზომებზე ცილის სინთეზის ადგილზე.
ინფორმაციის მატარებლის გარდა, საჭიროა ნივთიერებები, რომლებიც უზრუნველყოფენ ამინომჟავების მიწოდებას სინთეზის ადგილზე და მათი ადგილის დადგენას პოლიპეპტიდურ ჯაჭვში. ასეთი ნივთიერებებია გადაცემის რნმ, რომლებიც უზრუნველყოფენ ამინომჟავების კოდირებას და მიწოდებას სინთეზის ადგილზე. ცილის სინთეზი მიმდინარეობს რიბოსომებზე, რომელთა სხეული აგებულია რიბოსომური რნმ-ისგან. ეს ნიშნავს, რომ საჭიროა სხვა ტიპის რნმ - რიბოსომული.
გენეტიკური ინფორმაცია რეალიზდება სამი სახის რეაქციაში: რნმ სინთეზი, ცილის სინთეზი, დნმ-ის რეპლიკაცია. თითოეულ მათგანში ნუკლეოტიდების ხაზოვანი თანმიმდევრობის შემცველი ინფორმაცია გამოიყენება სხვა წრფივი თანმიმდევრობის შესაქმნელად: ნუკლეოტიდები (რნმ ან დნმ-ის მოლეკულებში) ან ამინომჟავები (ცილის მოლეკულებში). ექსპერიმენტულად დადასტურდა, რომ ეს არის დნმ, რომელიც ემსახურება როგორც შაბლონს ყველა ნუკლეინის მჟავის სინთეზისთვის. ამ ბიოსინთეზურ რეაქციებს ე.წ მატრიცის სინთეზი.მატრიცული რეაქციების საკმარისმა სიმარტივემ და მათმა ერთგანზომილებიანობამ შესაძლებელი გახადა მათი მექანიზმის დეტალური შესწავლა და გაგება, უჯრედში მიმდინარე სხვა პროცესებისგან განსხვავებით.
ტრანსკრიფცია
დნმ-დან რნმ-ის ბიოსინთეზის პროცესს ე.წ ტრანსკრიფცია.ეს პროცესი ხდება ბირთვში. დნმ-ის მატრიცაზე სინთეზირებულია რნმ-ის ყველა ტიპი - ინფორმაციული, სატრანსპორტო და რიბოსომული, რომლებიც შემდგომში მონაწილეობენ ცილის სინთეზში. დნმ-ის გენეტიკური კოდი ტრანსკრიფციის დროს გადაიწერება მესენჯერ რნმ-ში. რეაქცია ეფუძნება კომპლემენტარობის პრინციპს.
რნმ-ს სინთეზს აქვს მთელი რიგი მახასიათებლები. რნმ-ის მოლეკულა გაცილებით მოკლეა და დნმ-ის მხოლოდ მცირე ნაწილის ასლია. ამიტომ, დნმ-ის მხოლოდ გარკვეული ნაწილი, სადაც მოცემული ნუკლეინის მჟავის შესახებ ინფორმაციაა განთავსებული, მატრიცას ემსახურება. ახლად სინთეზირებული რნმ არასოდეს რჩება შეკრული დნმ-ის თავდაპირველ შაბლონთან, მაგრამ გამოიყოფა რეაქციის დასრულების შემდეგ. ტრანსკრიფციის პროცესი სამ ეტაპად მიმდინარეობს.
პირველი ეტაპი - ინიცირება- პროცესის დასაწყისი. რნმ-ის ასლების სინთეზი იწყება დნმ-ზე კონკრეტული უბნით, რომელსაც ე.წ პრომოუტერი.ეს ზონა შეიცავს ნუკლეოტიდების სპეციფიკურ კომპლექტს, რომლებიც არიან დაწყების სიგნალები.პროცესი კატალიზებულია ფერმენტების მიერ რნმ პოლიმერაზები.რნმ პოლიმერაზას ფერმენტი აკავშირებს პრომოტორს, ხსნის ორმაგ სპირალს და არღვევს წყალბადურ კავშირებს დნმ-ის ორ ჯაჭვს შორის. მაგრამ მხოლოდ ერთი მათგანი ემსახურება რნმ-ის სინთეზის შაბლონს.
მეორე ეტაპი - დრეკადობა.ამ ეტაპზე ხდება ძირითადი პროცესი. დნმ-ის ერთ ჯაჭვზე, ისევე როგორც მატრიცაზე, ნუკლეოტიდები რიგდებიან კომპლემენტარობის პრინციპის მიხედვით (სურ. 19). რნმ პოლიმერაზას ფერმენტი, რომელიც მოძრაობს ნაბიჯ-ნაბიჯ დნმ-ის ჯაჭვის გასწვრივ, აკავშირებს ნუკლეოტიდებს ერთმანეთთან და მუდმივად ხსნის დნმ-ის ორმაგ სპირალს შემდგომში. ამ მოძრაობის შედეგად სინთეზირდება რნმ-ის ასლი.
მესამე ეტაპი - შეწყვეტა.ეს ბოლო ეტაპია. რნმ-ის სინთეზი გრძელდება მანამ გაჩერების სიგნალი- ნუკლეოტიდების გარკვეული თანმიმდევრობა, რომელიც აჩერებს ფერმენტისა და რნმ-ის სინთეზის მოძრაობას. პოლიმერაზა გამოყოფილია დნმ-დან და სინთეზირებული რნმ-ის ასლიდან. ამავდროულად, რნმ-ის მოლეკულა ასევე ამოღებულია მატრიციდან. დნმ აღადგენს ორმაგ სპირალს. სინთეზი დასრულდა. დნმ-ის რეგიონიდან გამომდინარე, ამ გზით სინთეზირდება რიბოსომული, სატრანსპორტო და მესინჯერი რნმ.
რნმ-ის მოლეკულის ტრანსკრიფციის შაბლონი დნმ-ის მხოლოდ ერთ-ერთი ჯაჭვია. თუმცა, დნმ-ის სხვადასხვა ჯაჭვები შეიძლება იყოს შაბლონი ორი მიმდებარე გენისთვის. ორი ჯაჭვიდან რომელი იქნება გამოყენებული სინთეზისთვის, განსაზღვრავს პრომოტორი, რომელიც მიმართავს რნმ პოლიმერაზას ფერმენტს ამა თუ იმ მიმართულებით.
ტრანსკრიფციის შემდეგ, ევკარიოტული უჯრედების მესინჯერი რნმ-ის მოლეკულა განიცდის გადაწყობას. მასში ამოჭრილია ნუკლეოტიდური თანმიმდევრობები, რომლებიც არ ატარებენ ინფორმაციას ამ ცილის შესახებ. ამ პროცესს ე.წ შერწყმა.უჯრედის ტიპისა და განვითარების სტადიიდან გამომდინარე, შესაძლებელია რნმ-ის მოლეკულის სხვადასხვა ნაწილის ამოღება. შესაბამისად, დნმ-ის ერთ მონაკვეთში სინთეზირდება სხვადასხვა რნმ, რომლებიც ატარებენ ინფორმაციას სხვადასხვა ცილების შესახებ. ეს უზრუნველყოფს მნიშვნელოვანი გენეტიკური ინფორმაციის გადაცემას ერთი გენიდან და ასევე ხელს უწყობს გენეტიკურ რეკომბინაციას.
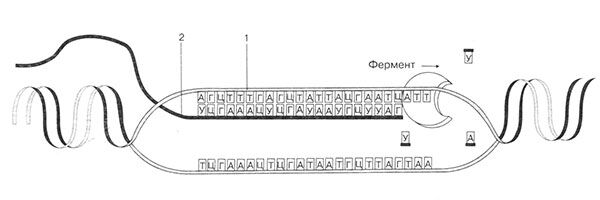
ბრინჯი. 19. მესინჯერი რნმ-ის სინთეზი. 1 - დნმ ჯაჭვი; 2 - სინთეზირებული რნმ
კითხვები და ამოცანები თვითკონტროლისთვის
1. რა რეაქციები უკავშირდება მატრიცის სინთეზის რეაქციებს?
2. რა არის საწყისი მატრიცა მატრიცის სინთეზის ყველა რეაქციისთვის?
3. რა ჰქვია mRNA ბიოსინთეზის პროცესს?
4. რა ტიპის რნმ არის სინთეზირებული დნმ-ზე?
5. დააყენეთ mRNA ფრაგმენტის თანმიმდევრობა, თუ დნმ-ის შესაბამის ფრაგმენტს აქვს თანმიმდევრობა: AAGCTCTGATTCTGATCGGACCTAATGA.
8. ცილის ბიოსინთეზი
ცილები ყველა უჯრედის აუცილებელი კომპონენტია, ამიტომ პლასტიკური მეტაბოლიზმის ყველაზე მნიშვნელოვანი პროცესი ცილის ბიოსინთეზია. ის გვხვდება ორგანიზმის ყველა უჯრედში. ეს არის უჯრედის ერთადერთი კომპონენტები (გარდა ნუკლეინის მჟავებისა), რომელთა სინთეზი ხდება უჯრედის გენეტიკური მასალის უშუალო კონტროლის ქვეშ. უჯრედის მთელი გენეტიკური აპარატი - დნმ და სხვადასხვა ტიპის რნმ - მორგებულია ცილების სინთეზისთვის.
გენი- ეს არის დნმ-ის მოლეკულის განყოფილება, რომელიც პასუხისმგებელია ერთი ცილის მოლეკულის სინთეზზე. ცილის სინთეზისთვის აუცილებელია დნმ-თან დაკავშირებული გარკვეული გენის კოპირება მესინჯერ რნმ-ის მოლეკულის სახით. ეს პროცესი ადრე იყო განხილული. ცილის სინთეზი რთული მრავალსაფეხურიანი პროცესია და დამოკიდებულია სხვადასხვა ტიპის რნმ-ის აქტივობაზე. ცილების პირდაპირი ბიოსინთეზისთვის საჭიროა შემდეგი კომპონენტები:
1. მესინჯერი რნმ - ინფორმაციის გადამზიდავი დნმ-დან სინთეზის ადგილამდე. mRNA მოლეკულები სინთეზირდება ტრანსკრიფციის დროს.
2. რიბოსომები – ორგანელები, სადაც ხდება ცილის სინთეზი.
3. არსებითი ამინომჟავების ნაკრები ციტოპლაზმაში.
4. ამინომჟავების მაკოდირებელი რნმ-ების გადატანა და მათი გადატანა რიბოზომებზე სინთეზის ადგილზე.
5. ATP - ნივთიერება, რომელიც უზრუნველყოფს ენერგიას ამინომჟავების კოდირების პროცესებისა და პოლიპეპტიდური ჯაჭვის სინთეზისთვის.
გადაცემის რნმ სტრუქტურა და ამინომჟავის კოდირება
გადაცემის რნმ (tRNA) არის პატარა მოლეკულები 70-დან 90 ნუკლეოტიდთან ერთად. tRNAs შეადგენს მთელი უჯრედის რნმ-ის დაახლოებით 15%-ს. tRNA-ს ფუნქცია დამოკიდებულია მის სტრუქტურაზე. tRNA მოლეკულების სტრუქტურის შესწავლამ აჩვენა, რომ ისინი გარკვეულწილად იკეცება და ჰგავს სამყურა ფოთოლი(სურ. 20). მოლეკულაში გამოიყოფა მარყუჟები და ორმაგი სექციები, რომლებიც დაკავშირებულია დამატებითი ბაზების ურთიერთქმედების გამო. ყველაზე მნიშვნელოვანია ცენტრალური მარყუჟი, რომელიც შეიცავს ანტიკოდონი -ნუკლეოტიდის ტრიპლეტი, რომელიც შეესაბამება კონკრეტული ამინომჟავის კოდს. თავისი ანტიკოდონით, tRNA-ს შეუძლია გაერთიანდეს mRNA-ზე შესაბამის კოდონთან კომპლემენტარობის პრინციპის მიხედვით.
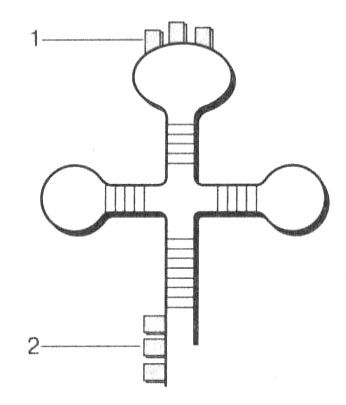
ბრინჯი. 20. tRNA მოლეკულის სტრუქტურა: 1 - ანტიკოდონი; 2 - ამინომჟავის მიმაგრების ადგილი
თითოეულ tRNA-ს შეუძლია 20 ამინომჟავიდან მხოლოდ ერთის გადატანა. ეს ნიშნავს, რომ თითოეული ამინომჟავისთვის არის მინიმუმ ერთი tRNA. ვინაიდან ამინომჟავას შეიძლება ჰქონდეს რამდენიმე სამეული, tRNA სახეობების რაოდენობა უდრის ამინომჟავის სამეულების რაოდენობას. ამრიგად, tRNA სახეობების საერთო რაოდენობა შეესაბამება კოდონების რაოდენობას და უდრის 61-ს. არცერთი tRNA არ შეესაბამება სამ გაჩერების კოდს.
tRNA მოლეკულის ერთ ბოლოში ყოველთვის არის გუანინის ნუკლეოტიდი (5'-ბოლო), ხოლო მეორეზე (3'-ბოლო) ყოველთვის არის სამი CCA ნუკლეოტიდი. სწორედ ამ მიზნით არის მიმაგრებული ამინომჟავა (სურ. 21). თითოეული ამინომჟავა ერთვის თავის სპეციფიკურ tRNA-ს შესაბამისი ანტიკოდონით. ამ მიმაგრების მექანიზმი დაკავშირებულია სპეციფიკური ფერმენტების - ამინოაცილ-tRNA სინთეზების მუშაობასთან, რომლებიც თითოეულ ამინომჟავას ამაგრებენ შესაბამის tRNA-ს. თითოეულ ამინომჟავას აქვს საკუთარი სინთეტაზა. ამინომჟავის შეერთება tRNA-სთან ხორციელდება ATP-ის ენერგიის გამო, ხოლო მაკროერგიული ბმა გადადის ბმაში tRNA-სა და ამინომჟავას შორის. ასე ხდება ამინომჟავების გააქტიურება და კოდირება.
ცილის ბიოსინთეზის ეტაპები. რიბოსომაზე განხორციელებული პოლიპეპტიდური ჯაჭვის სინთეზის პროცესს ე.წ გადაცემა.მესინჯერი რნმ (მრნმ) არის შუამავალი ცილის პირველადი სტრუქტურის შესახებ ინფორმაციის გადაცემისას, tRNA გადასცემს კოდირებულ ამინომჟავებს სინთეზის ადგილზე და უზრუნველყოფს მათი ნაერთების თანმიმდევრობას. რიბოსომები აგროვებენ პოლიპეპტიდურ ჯაჭვს.
მემკვიდრეობითი ინფორმაციის გადაცემა და განხორციელება ეფუძნება მატრიცის სინთეზის რეაქციებს. მათგან მხოლოდ სამია: დნმ-ის რეპლიკაცია, ტრანსკრიფცია და ტრანსლაცია. ყველა ეს რეაქცია დაკავშირებულია პლასტიკური გაცვლის რეაქციებთან და მოითხოვს ენერგიის ხარჯვას და ფერმენტების მონაწილეობას.
რეპლიკაცია.
რეპლიკაცია- დნმ-ის მოლეკულების თვითგაორმაგება - საფუძვლად უდევს მემკვიდრეობითი ინფორმაციის გადაცემას თაობიდან თაობაში. ერთი მშობლის დნმ-ის მოლეკულის რეპლიკაციის შედეგად წარმოიქმნება ორი ქალიშვილი, რომელთაგან თითოეული არის ორმაგი სპირალი, რომელშიც დნმ-ის ერთი ჯაჭვი არის მშობელი, ხოლო მეორე ახლად სინთეზირებული. რეპლიკაცია მოითხოვს სხვადასხვა ფერმენტებს, ნუკლეოტიდებს და ენერგიას.
სპეციალური ფერმენტების დახმარებით წყდება დედის დნმ-ის ორი ჯაჭვის დამატებითი ფუძის დამაკავშირებელი წყალბადის ბმები. დნმ-ის ძაფები განსხვავდება. დნმ პოლიმერაზას ფერმენტის მოლეკულები მოძრაობენ დნმ-ის მშობელი ჯაჭვების გასწვრივ და თანმიმდევრულად აკავშირებენ ნუკლეოტიდებს შვილობილი დნმ-ის ძაფების წარმოქმნით. ნუკლეოტიდების დამატების პროცესი მიჰყვება კომპლემენტარობის პრინციპს. შედეგად, ორი დნმ-ის მოლეკულა იქმნება მშობლისა და ერთმანეთის იდენტური.
ცილის ბიოსინთეზი.
ცილის ბიოსინთეზი, ე.ი. მემკვიდრეობითი ინფორმაციის რეალიზაციის პროცესი ორ ეტაპად მიმდინარეობს. პირველ ეტაპზე ცილის პირველადი სტრუქტურის შესახებ ინფორმაცია კოპირდება დნმ-დან mRNA-მდე. ამ პროცესს ტრანსკრიფცია ეწოდება. მეორე ეტაპი - ტრანსლაცია - ხდება რიბოზომებზე. ტრანსლაციის დროს ამინომჟავებიდან ცილები სინთეზირდება mRNA-ში ჩაწერილი თანმიმდევრობის შესაბამისად, ე.ი. ნუკლეოტიდური თანმიმდევრობა ითარგმნება ამინომჟავების თანმიმდევრობაში. ამრიგად, მემკვიდრეობითი ინფორმაციის რეალიზაციის პროცესი შეიძლება გამოიხატოს სქემით:
დნმ → mRNA → ცილა → თვისება, ნიშანი
ტრანსკრიფცია- მესინჯერი რნმ-ის სინთეზი დნმ-ის შაბლონზე. ეს პროცესი ხდება იქ, სადაც არის დნმ. ევკარიოტებში ტრანსკრიფცია ხდება ბირთვში, მიტოქონდრიაში და ქლოროპლასტებში (მცენარეებში), პროკარიოტებში კი უშუალოდ ციტოპლაზმაში. ტრანსკრიფციის დროს დნმ-ის მოლეკულა არის შაბლონი, ხოლო mRNA არის რეაქციის პროდუქტი.
ტრანსკრიფცია იწყება დნმ-ის ჯაჭვების გამოყოფით, რაც ხდება ისევე, როგორც რეპლიკაციის დროს (წყალბადის ბმები იშლება ფერმენტების დახმარებით). შემდეგ რნმ პოლიმერაზას ფერმენტი თანმიმდევრულად აერთიანებს ნუკლეოტიდებს ჯაჭვში კომპლემენტარობის პრინციპის მიხედვით, სინთეზირებს mRNA მოლეკულას. შედეგად მიღებული mRNA მოლეკულა გამოყოფილია და იგზავნება ციტოპლაზმაში რიბოსომის „საძიებლად“.
რიბოზომებზე ცილების სინთეზს ე.წ გადაცემა. ევკარიოტებში ტრანსლაცია ხდება რიბოსომებზე, რომლებიც მდებარეობს ციტოპლაზმაში, EPS-ის ზედაპირზე, მიტოქონდრიებში და ქლოროპლასტებში (მცენარეებში) და პროკარიოტებში ციტოპლაზმის რიბოზომებზე. თარგმანი მოიცავს mRNA, tRNA, რიბოზომებს, ამინომჟავებს, ATP მოლეკულებს და ფერმენტებს.
· Ამინომჟავებისემსახურება როგორც მასალას ცილის მოლეკულის სინთეზისთვის.
· ATPარის ენერგიის წყარო ამინომჟავების ერთმანეთთან დასაკავშირებლად.
· ფერმენტებიმონაწილეობენ ამინომჟავების tRNA-სთან მიმაგრებაში და ამინომჟავების ერთმანეთთან შეერთებაში.
· რიბოზომებიისინი შედგება rRNA და ცილის მოლეკულებისგან, რომლებიც ქმნიან აქტიურ ცენტრს, რომელშიც მიმდინარეობს ტრანსლაციის ძირითადი მოვლენები.
· მესინჯერი რნმამ შემთხვევაში, ეს არის ცილის მოლეკულის სინთეზის შაბლონი. mRNA ტრიპლეტებს, რომელთაგან თითოეული ამინომჟავას კოდირებს, ეწოდება კოდონები.
· გადაცემის რნმმოაქვს ამინომჟავები რიბოსომებში და მონაწილეობს ნუკლეოტიდური თანმიმდევრობის ამინომჟავების თანმიმდევრობაში ტრანსლაციაში. გადაცემის რნმ, ისევე როგორც რნმ-ის სხვა ტიპები, სინთეზირდება დნმ-ის შაბლონზე. ისინი ჰგავს სამყურას ფოთოლს (სურ. 28.3). სამი ნუკლეოტიდი, რომელიც მდებარეობს tRNA მოლეკულის ცენტრალური მარყუჟის ზედა ნაწილში, იქმნება ანტიკოდონი.
თარგმანის პროგრესი.
 თარგმანი იწყება mRNA-ს რიბოსომასთან შეერთებით. რიბოსომა მოძრაობს mRNA-ს გასწვრივ, ყოველ ჯერზე მოძრაობს ერთი სამეული. mRNA-ს ორი სამეული (კოდონი) შეიძლება ერთდროულად იყოს რიბოსომის აქტიურ ცენტრში. თითოეულ ამ კოდონს ემთხვევა tRNA, რომელსაც აქვს დამატებითი ანტიკოდონი და ატარებს სპეციფიკურ ამინომჟავას. წყალბადის ბმები იქმნება კოდონებსა და ანტიკოდონებს შორის, რომლებიც ინარჩუნებენ tRNA-ს აქტიურ ადგილზე. ამ დროს ამინომჟავებს შორის წარმოიქმნება პეპტიდური ბმა. მზარდი პოლიპეპტიდური ჯაჭვი „შეჩერებულია“ tRNA-ზე, რომელიც ამ უკანასკნელის აქტიურ ცენტრში შევიდა. რიბოსომა წინ მიიწევს ერთი ტრიპლეტით, რის შედეგადაც წარმოიქმნება ახალი კოდონი და შესაბამისი tRNA აქტიურ ადგილზე. გამოთავისუფლებული tRNA გამოყოფილია mRNA-სგან და იგზავნება ახალი ამინომჟავისთვის.
თარგმანი იწყება mRNA-ს რიბოსომასთან შეერთებით. რიბოსომა მოძრაობს mRNA-ს გასწვრივ, ყოველ ჯერზე მოძრაობს ერთი სამეული. mRNA-ს ორი სამეული (კოდონი) შეიძლება ერთდროულად იყოს რიბოსომის აქტიურ ცენტრში. თითოეულ ამ კოდონს ემთხვევა tRNA, რომელსაც აქვს დამატებითი ანტიკოდონი და ატარებს სპეციფიკურ ამინომჟავას. წყალბადის ბმები იქმნება კოდონებსა და ანტიკოდონებს შორის, რომლებიც ინარჩუნებენ tRNA-ს აქტიურ ადგილზე. ამ დროს ამინომჟავებს შორის წარმოიქმნება პეპტიდური ბმა. მზარდი პოლიპეპტიდური ჯაჭვი „შეჩერებულია“ tRNA-ზე, რომელიც ამ უკანასკნელის აქტიურ ცენტრში შევიდა. რიბოსომა წინ მიიწევს ერთი ტრიპლეტით, რის შედეგადაც წარმოიქმნება ახალი კოდონი და შესაბამისი tRNA აქტიურ ადგილზე. გამოთავისუფლებული tRNA გამოყოფილია mRNA-სგან და იგზავნება ახალი ამინომჟავისთვის.
ბიოლოგიის ოლიმპიადა. სკოლის ეტაპი. 2016-2017 სასწავლო წელი.
10-11 კლასი
1. უჯრედისა და ქსოვილის არასწორი კორელაციაა
ა) ფესვის თმა – მთლიანი ქსოვილი
ბ) პოლისადური პარენქიმის უჯრედი – ძირითადი ქსოვილი
გ) მიმავალი უჯრედი – მთლიანი ქსოვილი
დ) კომპანიონური უჯრედი – ექსკრეტორული ქსოვილი
2. ღონისძიებისთვის, რომელიც ჩატარდება სამ დღეში, საჭიროა მწიფე მსხალი. თუმცა ის მსხალი, რომელიც ამ მიზნით იყიდა, ჯერ არ იყო მომწიფებული. მომწიფების პროცესი შეიძლება დაჩქარდეს მათი დაყენებით
ა) ბნელ ადგილას
ბ) მაცივარში
ბ) ფანჯრის რაფაზე
დ) სქელი ქაღალდის ტომარაში მწიფე ვაშლებთან ერთად
3. ბრიოფიტებმა მოახერხეს ხმელეთზე გადარჩენა, რადგან
ა) ისინი იყვნენ პირველი მცენარეები, რომლებმაც განავითარეს სტომატები
ბ) მათ არ სჭირდებათ ტენიანი გარემო რეპროდუქციული ციკლისთვის
გ) იზრდებიან ნიადაგის ზემოთ შედარებით ნოტიო რაიონებში
დ) სპოროფიტი დამოუკიდებელი გახდა გამეტოფიტისგან
4 ძუძუმწოვარი ლოყები ჩამოყალიბებულია მსგავსად
ა) მოწყობილობა დიდი რაოდენობით საკვების შესაგროვებლად
ბ) თავის ქალას და, კერძოდ, ყბების სტრუქტურული თავისებურებების შედეგი
ბ) წოვის მოწყობილობა
დ) სუნთქვის დამხმარე საშუალება
5. ნიანგის გული თავისი აგებულებით
ა) სამკამერიანი პარკუჭში არასრული ძგიდით
ბ) სამკამერიანი
ბ) ოთხკამერიანი
დ) ოთხკამერიანი ნახვრეტით ძგიდეში პარკუჭებს შორის
6. ფიბრინოგენი, რომელიც წარმოადგენს ცილას, მონაწილეობს სისხლის შედედებაში
ა) სისხლის პლაზმა
ბ) ლეიკოციტების ციტოპლაზმა
ბ) თრომბოციტების ნაწილი
დ) წარმოიქმნება სისხლის წითელი უჯრედების განადგურების დროს
7. აბიოტური ფაქტორები მოიცავს ისეთ ეკოლოგიურ ერთეულს, როგორიცაა
ა) ბიოცენოზი
ბ) ეკოსისტემა
ბ) მოსახლეობა
8. ფორმირებისას ხდება რედუქციური გაყოფა (მეიოზი).
ა) ბაქტერიული სპორები
ბ) ულოტრიქსის ზოოსპორები
ბ) მარჩანტიას სპორები
დ) ზოოსპორები ფიტოფტორა
9. ჩამოთვლილი ბიოპოლიმერებიდან განშტოებული სტრუქტურა აქვს
დ) პოლისაქარიდები
10. ფენილკეტონურია არის გენეტიკური დაავადება, რომელიც გამოწვეულია რეცესიული მუტაციით. ავადმყოფი ბავშვის გაჩენის ალბათობა, თუ ორივე მშობელი ამ მახასიათებლის მიმართ ჰეტეროზიგოტურია
11. ახსნილია მხედველობის ორგანოების აგებულების მსგავსება კეფალოპოდებსა და ხერხემლიანებში.
ა) კონვერგენცია
ბ) პარალელიზმი
ბ) ადაპტაცია
დ) დამთხვევა
12. თავისუფლად მოცურავე ასციდის ლარვას აქვს აკორდი და ნერვული მილი. ზრდასრული ასციდიის დროს, რომელიც უვლის ცხოვრების წესს, ისინი ქრება. ეს არის მაგალითი
ა) ადაპტაციები
ბ) დეგენერაცია
ბ) ცენოგენეზი
13. ფიჭვის წყლის მატარებელი ელემენტებია
ა) რგოლოვანი და სპირალური ჭურჭელი
ბ) მხოლოდ რგოლოვანი ჭურჭელი
ბ) ტრაქეიდები
დ) სპირალური და ფოროვანი ჭურჭელი
14. ნაყოფიერება დამახასიათებელია
ბ) ანანასი
ბ) ბანანი
15. მცენარეთა უჯრედების ქლოროპლასტებში განლაგებულია სინათლის მოსავლის კომპლექსები
ა) გარე მემბრანაზე
ბ) შიდა მემბრანაზე
ბ) თილაკოიდურ მემბრანაზე
დ) სტრომაში
Მე -2 ნაწილი.
მატჩი (6 ქულა).
2.1. დაადგინეთ შესაბამისობა ნაცრისფერი ვირთხის ნიშანსა და იმ სახეობის კრიტერიუმს შორის, რომლისთვისაც ის დამახასიათებელია.
2.2. ფუნქციების რეგულირების მახასიათებლებსა და მის მეთოდს შორის შესაბამისობის დადგენა.
დააყენეთ სწორი თანმიმდევრობა (6 ქულა).
2.3. დაადგინეთ გეოგრაფიული სახეობების ეტაპების სწორი თანმიმდევრობა.
1) ტერიტორიული იზოლაციის გაჩენა იმავე სახეობის პოპულაციებს შორის
2) სახეობების დიაპაზონის გაფართოება ან დაყოფა
3) მუტაციების გამოჩენა იზოლირებულ პოპულაციებში
4) ბუნებრივი გადარჩევით ინდივიდების შენარჩუნება იმ თვისებებით, რომლებიც სასარგებლოა სპეციფიკურ გარემო პირობებში
5) სხვადასხვა პოპულაციის ინდივიდების მიერ შეჯვარების უნარის დაკარგვა
2.4. დაადგინეთ თანმიმდევრობა, რომელშიც ეს პროცესები ხდება მიტოზური უჯრედების გაყოფის დროს.
1) ქრომოსომა განლაგებულია უჯრედის ეკვატორის გასწვრივ
2) ქრომატიდები განსხვავდებიან უჯრედის პოლუსებისკენ
3) წარმოიქმნება ორი ქალიშვილი უჯრედი
4) ქრომოსომა სპირალიზებულია, თითოეული შედგება ორი ქრომატიდისგან
5) ქრომოსომა დესპირალიზებულია
2.5. თქვენ გთავაზობენ ტესტის დავალებებს განსჯის სახით, რომელთაგან თითოეულს ან უნდა დაეთანხმოთ ან უარყოთ. პასუხის მატრიცაში მიუთითეთ პასუხის ვარიანტი „დიახ“ ან „არა“: (10 ქულა).
1. Nightshade ყვავილები შეგროვებული ქოლგის inflorescence.
2. კილიარულ ჭიებს ანუსი არ აქვთ.
3. პეროქსისომა არის ევკარიოტული უჯრედის სავალდებულო ორგანელა.
4. პეპტიდური ბმა არ არის მაკროერგიული.
5. ღვიძლის უჯრედებში გლუკაგონის დამატება იწვევს გლიკოგენის დაშლას.
6. აბიოტური ფაქტორები არ მოქმედებს ორი მონათესავე სახეობის კონკურენტულ ურთიერთობებზე.
7. ფოთოლზე გაზის გაცვლის ფუნქციები შესაძლებელია ლანციელის და ჰიდათოდების გამო.
8. ძუძუმწოვრების ერთკამერიანი კუჭის შესაბამისი მონაკვეთი მომცრო ცხოველების კუჭის ნაწიბურია.
9. კვებითი ჯაჭვების სიგრძე შემოიფარგლება ენერგიის დაკარგვით.
10. რაც უფრო მცირეა სხეულში სისხლძარღვების დიამეტრი, მით მეტია მათში სისხლის ნაკადის წრფივი სიჩქარე.
ნაწილი 3
3.1. იპოვეთ სამი შეცდომა მოცემულ ტექსტში. მიუთითეთ წინადადებების ნომრები, რომლებშიც ისინი შედგენილია, შეასწორეთ (6 ქულა).
1. მატრიცის სინთეზის რეაქციები მოიცავს სახამებლის წარმოქმნას, mRNA სინთეზს, ცილების შეკრებას რიბოზომებში. 2. მატრიცის სინთეზი წააგავს მონეტების ჩამოსხმას მატრიცაზე: ახალი მოლეკულების სინთეზირება ხდება არსებული მოლეკულების სტრუქტურაში დასახული „გეგმის“ შესაბამისად. 3. მატრიქსის როლს უჯრედში ქლოროფილის მოლეკულები, ნუკლეინის მჟავები (დნმ და რნმ) ასრულებენ. 4. მონომერები ფიქსირდება მატრიცებზე, შემდეგ ისინი გაერთიანებულია პოლიმერულ ჯაჭვებში. 5. მზა პოლიმერები გამოდიან მატრიცებიდან. 6. ძველი მატრიცები მაშინვე ნადგურდება, რის შემდეგაც ყალიბდება ახალი.
ადამიანს აქვს ოთხი ფენოტიპი სისხლის ჯგუფების მიხედვით: I (0), II (A), III (B), IV (AB). გენს, რომელიც განსაზღვრავს სისხლის ჯგუფს, აქვს სამი ალელი: IA, IB, i0; უფრო მეტიც, i0 ალელი რეცესიულია IA და IB ალელებთან მიმართებაში. მშობლებს აქვთ II (ჰეტეროზიგოტური) და III (ჰომოზიგოტური) სისხლის ჯგუფი. განსაზღვრეთ მშობლების სისხლის ჯგუფების გენოტიპები. მიუთითეთ ბავშვების სისხლის ჯგუფის შესაძლო გენოტიპები და ფენოტიპები (რიცხვი). შეადგინეთ პრობლემის გადაჭრის სქემა. განსაზღვრეთ მემკვიდრეობის ალბათობა II სისხლის ჯგუფის ბავშვებში.
პასუხები 10-11 კლასი
ნაწილი 1. აირჩიეთ ერთი სწორი პასუხი. (15 ქულა)
2.2. მაქსიმალური - 3 ქულა, ერთი შეცდომა - 2 ქულა, ორი შეცდომა - 1ბ, სამი ან მეტი შეცდომა - 0 ქულა
2.4. მაქსიმალური - 3 ქულა, ერთი შეცდომა - 2 ქულა, ორი შეცდომა - 1ბ, სამი ან მეტი შეცდომა - 0 ქულა
ნაწილი 3
3.1. იპოვეთ სამი შეცდომა მოცემულ ტექსტში. მიუთითეთ წინადადებების რაოდენობა, რომლებშიც ისინი შესრულდა, შეასწორეთ ისინი (3b შეცდომების მქონე წინადადებების სწორად ამოცნობისთვის და 3b შეცდომების გამოსწორებისთვის).
1. - მატრიცის სინთეზის რეაქციები არ მოიცავს სახამებლის წარმოქმნას, ამისთვის მატრიცა არ არის საჭირო;
3. - ქლოროფილის მოლეკულებს არ შეუძლიათ მატრიცის სახით მოქმედება, არ გააჩნიათ კომპლემენტარობის თვისება;
6. - მატრიცები გამოიყენება არაერთხელ.
3.2. ამოცანის ამოხსნა (3 ქულა).
პრობლემის გადაჭრის სქემა მოიცავს:
1) მშობლებს აქვთ სისხლის ჯგუფები: II ჯგუფი - IAi0 (გამეტები IA, i0), III ჯგუფი - IB IB (გამეტები IB);
2) ბავშვთა სისხლის ჯგუფების შესაძლო ფენოტიპები და გენოტიპები: IV ჯგუფი (IAIB) და III ჯგუფი (IBi0);
3) II ჯგუფის სისხლის მემკვიდრეობის ალბათობა არის 0%.
პასუხის ფორმა
ბიოლოგიის რუსულენოვანი ოლიმპიადის სასკოლო ეტაპი
მონაწილის კოდი _____________
ნაწილი 1. აირჩიეთ ერთი სწორი პასუხი. (15 ქულა)
Მე -2 ნაწილი.
ნაწილი 3
3.1._______________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________________
3.2. პრობლემის გადაწყვეტა
1869 წელს შვეიცარიელმა ბიოქიმიკოსმა იოჰან ფრიდრიხ მიშერმა პირველად აღმოაჩინა, გამოყო უჯრედის ბირთვებიდან და აღწერა დნმ. მაგრამ მხოლოდ 1944 წელს O. Avery, S. McLeod და M. McCarthy დაამტკიცეს დნმ-ის გენეტიკური როლი, ანუ საიმედოდ დადგინდა, რომ მემკვიდრეობითი ინფორმაციის გადაცემა დაკავშირებულია დეზოქსირიბონუკლეინის მჟავასთან. ეს აღმოჩენა იყო ძლიერი ფაქტორი, რომელიც ასტიმულირებდა მემკვიდრეობის შესწავლას მოლეკულურ დონეზე. მას შემდეგ დაიწყო მოლეკულური ბიოლოგიისა და გენეტიკის სწრაფი განვითარება.
Ნუკლეინის მჟავა (ლათ. ბირთვი - ბირთვი) არის ბუნებრივი მაღალმოლეკულური ორგანული ნაერთები, რომლებიც უზრუნველყოფენ მემკვიდრეობითი (გენეტიკური) ინფორმაციის შენახვას და გადაცემას ცოცხალ ორგანიზმებში. მათ შორისაა: ნახშირბადი (C), წყალბადი (H), ჟანგბადი (O), ფოსფორი (P). ნუკლეინის მჟავები არის არარეგულარული ბიოპოლიმერები, რომლებიც შედგება მონომერებისგან - ნუკლეოტიდებისგან. თითოეული ნუკლეოტიდი შეიცავს:
· აზოტის ბაზა,
· მარტივი ნახშირბადი - 5-ნახშირბადოვანი შაქრის პენტოზა (რიბოზა ან დეზოქსირიბოზა),
· ფოსფორის მჟავის ნარჩენი.
არსებობს ნუკლეინის მჟავების ორი ტიპი: დეზოქსირიბონუკლეინის მჟავა - დნმ, რომელიც შეიცავს დეზოქსირიბოზას და რიბონუკლეინის მჟავა - რნმ, რომელიც შეიცავს რიბოზას.
განვიხილოთ ნუკლეინის მჟავების თითოეული ტიპი.
დნმ გვხვდება თითქმის ექსკლუზიურად უჯრედის ბირთვში, ზოგჯერ ორგანელებში: მიტოქონდრიებში, პლასტიდებში. დნმ არის პოლიმერული ნაერთი უჯრედში მუდმივი (სტაბილური) შემცველობით.
დნმ-ის სტრუქტურა.თავისი სტრუქტურით, დნმ-ის მოლეკულა შედგება ორი პოლიმერული ჯაჭვისგან, რომლებიც ერთმანეთთან არის დაკავშირებული და გრეხილი ორმაგი სპირალის სახით (ნახ. 1).
დ. უოტსონმა და ფ. კრიკმა 1953 წელს შექმნეს დნმ-ის სტრუქტურის მოდელი, რისთვისაც ორივეს მიენიჭა ნობელის პრემია. ორმაგი სპირალის სიგანე არის მხოლოდ დაახლოებით 0,002 მიკრონი (20 ანგსტრომი), მაგრამ მისი სიგრძე განსაკუთრებით დიდია - რამდენიმე ათეულამდე და ასობით მიკრომეტრამდეც კი (შედარებისთვის: უმსხვილესი ცილის მოლეკულის სიგრძე გაშლილ ფორმაშია. არაუმეტეს 0,1 მიკრონი).
ნუკლეოტიდები განლაგებულია ერთმანეთისგან დაშორებით - 0,34 ნმ და არის 10 ნუკლეოტიდი სპირალის ყოველ შემობრუნებაზე. დნმ-ის მოლეკულური წონა დიდია: ის ათობით და ასობით მილიონიც კი არის. მაგალითად, მოლეკულური წონა (მ რ) დროზოფილას ყველაზე დიდი ქრომოსომა არის 7.9 10 10.
ერთი ჯაჭვის ძირითადი სტრუქტურული ერთეულია ნუკლეოტიდი, რომელიც შედგება აზოტოვანი ფუძისგან, დეზოქსირიბოზისა და ფოსფატის ჯგუფისგან. დნმ შეიცავს 4 ტიპის აზოტოვან ფუძეს:
· პურინი - ადენინი (A) და გუანინი (G),
· პირიმიდინი - ციტოზინი (C) და თიმინი (T).
პურინის ფუძეების საერთო რაოდენობა უდრის პირიმიდინის ფუძეების ჯამს.
დნმ ნუკლეოტიდები ასევე იქნება 4 ტიპის, შესაბამისად: ადენილი (A), გუანილი (G), ციტიდილი (C) და თიმიდილ (T) დნმ-ის ყველა ნუკლეოტიდი დაკავშირებულია პოლინუკლეოტიდურ ჯაჭვში დეოქსირიბოზებს შორის მდებარე ფოსფორმჟავას ნარჩენების გამო. პოლინუკლეოტიდურ ჯაჭვში შეიძლება იყოს 300000-მდე ან მეტი ნუკლეოტიდი.
ამრიგად, დნმ-ის თითოეული ჯაჭვი არის პოლინუკლეოტიდი, რომელშიც ნუკლეოტიდები განლაგებულია მკაცრად განსაზღვრული თანმიმდევრობით. აზოტოვანი ფუძეები ისე უახლოვდება ერთმანეთს, რომ წყალბადის ბმები იქმნება მათ შორის. მნიშვნელოვანი ნიმუში აშკარად ვლინდება მათ განლაგებაში: ერთი ჯაჭვის ადენინი (A) უკავშირდება მეორე ჯაჭვის თიმინს (T) ორი წყალბადის ბმით, ხოლო ერთი ჯაჭვის გუანინი (G) სამი წყალბადური ბმით უკავშირდება ციტოზინს.
 (გ) სხვა ჯაჭვი, რის შედეგადაც წარმოიქმნება A-T და G-C წყვილები. ნუკლეოტიდების შერჩევითი შეერთების ამ უნარს ეწოდება კომპლემენტარულობა, ანუ სივრცითი და ქიმიური კორესპონდენცია ნუკლეოტიდების წყვილებს შორის (იხ. სურ. 2).
(გ) სხვა ჯაჭვი, რის შედეგადაც წარმოიქმნება A-T და G-C წყვილები. ნუკლეოტიდების შერჩევითი შეერთების ამ უნარს ეწოდება კომპლემენტარულობა, ანუ სივრცითი და ქიმიური კორესპონდენცია ნუკლეოტიდების წყვილებს შორის (იხ. სურ. 2).
ერთი ჯაჭვის ნუკლეოტიდების შეერთების თანმიმდევრობა საპირისპიროა (შემავსებელი) მეორე ჯაჭვისა, ანუ ჯაჭვები, რომლებიც ქმნიან დნმ-ის ერთ მოლეკულას მრავალ მიმართულების, ანუ ანტიპარალელურია. ჯაჭვები ტრიალებს ერთმანეთზე და ქმნიან ორმაგ სპირალს. წყალბადის ბმების დიდი რაოდენობა უზრუნველყოფს დნმ-ის ჯაჭვების ძლიერ კავშირს და აძლევს მოლეკულას სტაბილურობას, ხოლო მობილურობას ინარჩუნებს - ფერმენტების გავლენის ქვეშ ადვილად იხსნება (დესპირალიზდება).
დნმ-ის რეპლიკაცია (დნმ-ის რედუპლიკაცია) - ნუკლეინის მჟავების მაკრომოლეკულების თვითრეპროდუქციის (თვით გაორმაგების) პროცესი, რომელიც უზრუნველყოფს გენეტიკური ინფორმაციის ზუსტ კოპირებას და მის გადაცემას თაობიდან თაობას.
დნმ-ის რეპლიკაცია ხდება უჯრედების გაყოფამდე ინტერფაზის დროს. დნმ-ის მთავარი მოლეკულა (უჯრედში დნმ-ის ჯაჭვების რაოდენობა არის 2n) ფერმენტების მოქმედებით იხსნება ერთი ბოლოდან, შემდეგ კი შვილობილი პოლინუკლეოტიდური ჯაჭვები სრულდება თავისუფალი ნუკლეოტიდებიდან ორივე ჯაჭვის კომპლემენტარობის პრინციპის მიხედვით. მატრიქსის რეაქციების შედეგად წარმოიქმნება ერთი და იგივე ნუკლეოტიდური შემადგენლობის ორი ქალიშვილი დნმ-ის მოლეკულა, რომლებშიც ერთ-ერთი ჯაჭვი არის ძველი მშობელი, ხოლო მეორე არის ახალი, ახლად სინთეზირებული (უჯრედში დნმ-ის რაოდენობა ხდება 4n. = 2 X 2n).
დნმ-ის ფუნქციები.
1. ცილების სტრუქტურის ან მისი ცალკეული ორგანელების შესახებ მემკვიდრეობითი ინფორმაციის შენახვა. გენეტიკური ინფორმაციის უმცირესი ერთეული ნუკლეოტიდის შემდეგ არის სამი თანმიმდევრული ნუკლეოტიდი - სამეული. პოლინუკლეოტიდურ ჯაჭვში ტრიპლეტების თანმიმდევრობა განსაზღვრავს ერთი ცილის მოლეკულის ამინომჟავების თანმიმდევრობას (ცილის პირველადი სტრუქტურა) და წარმოადგენს გენს. ცილებთან ერთად, დნმ არის ქრომატინის ნაწილი, ნივთიერება, რომელიც ქმნის უჯრედის ბირთვის ქრომოსომებს.
2. მემკვიდრეობითი ინფორმაციის გადაცემა უჯრედების გაყოფის დროს რეპლიკაციის შედეგად დედა უჯრედიდან ქალიშვილზე.
3. მემკვიდრეობითი ინფორმაციის დანერგვა (გენების სახით შენახული) ბიოსინთეზის მატრიქსული რეაქციების შედეგად უჯრედისა და ორგანიზმისთვის სპეციფიკური ცილების წარმოქმნით. ამავდროულად, მის ერთ-ერთ ჯაჭვზე, კომპლემენტარობის პრინციპის მიხედვით, სინთეზირებულია მესინჯერი რნმ-ის მოლეკულები მოლეკულის მიმდებარე გარემოს ნუკლეოტიდებიდან.
რნმ არის უჯრედში მერყევი (ლაბილური) შემცველობის ნაერთი.
რნმ-ის სტრუქტურა.მათი სტრუქტურის მიხედვით, რნმ-ის მოლეკულები უფრო მცირეა ვიდრე დნმ-ის მოლეკულები, რომელთა მოლეკულური წონაა 20-30 ათასი (tRNA) 1 მილიონამდე (rRNA), რნმ არის ერთჯაჭვიანი მოლეკულა, რომელიც აგებულია დნმ-ის ერთ-ერთი ჯაჭვის მსგავსად. . რნმ მონომერები - ნუკლეოტიდები შედგება აზოტოვანი ფუძისგან, რიბოზის (პენტოზა) და ფოსფატის ჯგუფისგან. რნმ შეიცავს 4 აზოტოვან ბაზას:
· პურინი - ადენინი (A);
· პირიმიდინი - გუანინი (G), ციტოზინი (C), ურაცილი (U).
რნმ-ში თიმინს ცვლის ურაცილი, რომელიც აგებულებით ახლოსაა მასთან (ნუკლეოტიდი – ურიდილ. ნუკლეოტიდები პოლინუკლეოტიდურ ჯაჭვში ისევე არიან დაკავშირებული, როგორც დნმ-ში, რიბოზებს შორის მდებარე ფოსფორმჟავას ნარჩენების გამო.
მდებარეობა საკანში რნმ-ს შორის გამოირჩევა: ბირთვული, ციტოპლაზმური, მიტოქონდრიული, პლასტიდური.
ფუნქციის მიხედვით რნმ-ებს შორის გამოირჩევა: ტრანსპორტი, ინფორმაცია და რიბოსომული.
 გადაცემის რნმ (tRNAs)
- ერთჯაჭვიანი, მაგრამ სამგანზომილებიანი "სამყურა" სტრუქტურის მქონე, რომელიც შექმნილია ინტრამოლეკულური წყალბადის ბმებით (ნახ. 3). tRNA მოლეკულები ყველაზე მოკლეა. შედგება 80-100 ნუკლეოტიდისგან. ისინი შეადგენენ უჯრედში რნმ-ის მთლიანი შემცველობის დაახლოებით 10%-ს. ისინი უჯრედში ცილების ბიოსინთეზის დროს რიბოსომებს გადასცემენ გააქტიურებულ ამინომჟავებს (თითოეულ tRNA-ს აქვს საკუთარი ამინომჟავა, საერთო ჯამში ცნობილია 61 tRNA).
გადაცემის რნმ (tRNAs)
- ერთჯაჭვიანი, მაგრამ სამგანზომილებიანი "სამყურა" სტრუქტურის მქონე, რომელიც შექმნილია ინტრამოლეკულური წყალბადის ბმებით (ნახ. 3). tRNA მოლეკულები ყველაზე მოკლეა. შედგება 80-100 ნუკლეოტიდისგან. ისინი შეადგენენ უჯრედში რნმ-ის მთლიანი შემცველობის დაახლოებით 10%-ს. ისინი უჯრედში ცილების ბიოსინთეზის დროს რიბოსომებს გადასცემენ გააქტიურებულ ამინომჟავებს (თითოეულ tRNA-ს აქვს საკუთარი ამინომჟავა, საერთო ჯამში ცნობილია 61 tRNA).
საინფორმაციო (მატრიცული) რნმ (mRNA, mRNA) - ერთჯაჭვიანი მოლეკულა, რომელიც წარმოიქმნება ბირთვში დნმ-ის მოლეკულაზე ტრანსკრიფციის შედეგად (აკოპირებს გენებს) და აწვდის ინფორმაციას ერთი ცილის მოლეკულის პირველადი სტრუქტურის შესახებ რიბოსომებში ცილის სინთეზის ადგილზე. mRNA მოლეკულა შეიძლება შედგებოდეს 300-3000 ნუკლეოტიდისგან. mRNA-ის წილი უჯრედში მთლიანი რნმ-ის შემცველობის 0,5-1%-ს შეადგენს.
რიბოსომული რნმ (rRNA) - ყველაზე დიდი ერთჯაჭვიანი მოლეკულები, რომლებიც ქმნიან კომპლექსურ კომპლექსებს ცილებთან, რომლებიც მხარს უჭერენ რიბოზომების სტრუქტურას, რომლებზეც ხდება ცილის სინთეზი.
rRNA შეადგენს უჯრედში რნმ-ის მთლიანი შემცველობის დაახლოებით 90%-ს.
ორგანიზმის მთელი გენეტიკური ინფორმაცია (მისი ცილების სტრუქტურა) შეიცავს მის დნმ-ში, რომელიც შედგება გენებში გაერთიანებული ნუკლეოტიდებისგან. შეგახსენებთ, რომ გენი არის მემკვიდრეობითი ინფორმაციის ერთეული (დნმ-ის მოლეკულის მონაკვეთი), რომელიც შეიცავს ინფორმაციას ერთი ცილის - ფერმენტის სტრუქტურის შესახებ. გენები, რომლებიც განსაზღვრავენ ორგანიზმების თვისებებს, ე.წ სტრუქტურული.და გენები, რომლებიც არეგულირებენ სტრუქტურული გენების გამოხატვას, ე.წ მარეგულირებელი.გენის გამოვლინება (გამოხატვა) (მემკვიდრეობითი ინფორმაციის რეალიზაცია) ხდება შემდეგნაირად:

გენის ექსპრესიის განსახორციელებლად, არსებობს გენეტიკური კოდი - მკაცრად მოწესრიგებული ურთიერთობა ნუკლეოტიდურ ფუძეებსა და ამინომჟავებს შორის (ცხრილი 12).
ცხრილი 12 გენეტიკური კოდი

გენეტიკური კოდის ძირითადი თვისებები.
სამმაგი- ამინომჟავების კოდირება ხორციელდება ნუკლეოტიდების ფუძეების ტრიპლეტებით (სამებით). კოდირების ტრიპლეტების რაოდენობაა 64 (ნუკლეოტიდების 4 ტიპი: A, T, C, G, 4 3 = 64).
გაურკვევლობათითოეული ტრიპლეტი აკოდირებს მხოლოდ ერთ ამინომჟავას.
დეგენერაცია- კოდირების სამეულების რაოდენობა აღემატება ამინომჟავების რაოდენობას (64 > 20). არსებობს ამინომჟავები, რომლებიც კოდირებულია ერთზე მეტი სამეულით (ასეთი ამინომჟავები უფრო ხშირია ცილებში). არსებობს სამი ტრიპლეტი, რომლებიც არ კოდირებენ არცერთ ამინომჟავას (UAA, UAG, UGA). მათ უწოდებენ "უაზრო კოდონებს" და ასრულებენ "გაჩერების სიგნალების" როლს, რაც ნიშნავს გენის ჩაწერის დასასრულს (კოდონების საერთო რაოდენობა 61-ია).
გადახურვის გარეშე (განგრძობადობა) - დნმ-დან ტრიპლეტის კითხვა mRNA სინთეზის დროს მიმდინარეობს მკაცრად სამი თანმიმდევრული ნუკლეოტიდის გასწვრივ, მეზობელი კოდონების გადახურვის გარეშე. გენში არ არის „სასვენი ნიშნები“.
მრავალმხრივობა - იგივე ტრიპლეტები ერთნაირი ამინომჟავების კოდირებას ახდენენ დედამიწაზე მცხოვრებ ყველა ორგანიზმში.
ამინომჟავების სახელების საერთო აბრევიატურები:
FEN - ფენილალანინი; GIS - ჰისტიდინი;
LEI - ლეიცინი; GLN - გლუტამინი;
ILE - იზოლეიცინი; GLU - გლუტამინის მჟავა;
MET - მეთიონინი; LYS - ლიზინი;
VAL - ვალინი; ASN - ასპარაგინი;
SER - სერია; ASP - ასპარტინის მჟავა;
PRO - პროლინი; დსთ - ცისტეინი;
TPE - თრეონინი; THREE - ტრიპტოფანი;
ALA - ალანინი; ARG - არგინინი;
TIR - ტიროზინი; GLI - გლიცინი.
ამრიგად, უჯრედში არსებული ყველა გენეტიკური ინფორმაციის დნმ-ის მატარებელი პირდაპირ მონაწილეობას არ იღებს ცილის სინთეზში (ანუ ამ მემკვიდრეობითი ინფორმაციის განხორციელებაში). ცხოველურ და მცენარეულ უჯრედებში დნმ-ის მოლეკულები გამოყოფილია ბირთვული მემბრანით ციტოპლაზმისგან.პლაზმა, სადაც ხდება ცილების სინთეზი. შუამავალი იგზავნება ბირთვიდან რიბოზომებში - ცილების შეკრების ადგილებამდე - რომელიც ატარებს კოპირებულ ინფორმაციას და შეუძლია გაიაროს ბირთვული მემბრანის ფორებში. მესინჯერი რნმ, რომელიც ჩართულია მატრიცულ რეაქციებში, ასეთი შუამავალია.
მატრიცული რეაქციები
- ეს არის რეაქციები "ძველ" მაკრომოლეკულებზე დაფუძნებული ახალი ნაერთების სინთეზისთვის, რომლებიც მოქმედებენ როგორც მატრიცა, ანუ ფორმა, მოდელი ახალი მოლეკულების კოპირებისთვის. მემკვიდრეობითი ინფორმაციის რეალიზაციის მატრიცული რეაქციები, რომელშიც დნმ და რნმ მონაწილეობენ, არის: 
1. დნმ-ის რეპლიკაცია- დნმ-ის მოლეკულების გაორმაგება, რის გამოც ხდება გენეტიკური ინფორმაციის გადაცემა თაობიდან თაობაში. მატრიცა არის დედის დნმ და ამ მატრიცის მიხედვით წარმოქმნილი ახლები არის ქალიშვილი, ახლად სინთეზირებული 2 დნმ მოლეკულა (ნახ. 4).
2.
ტრანსკრიფცია(ლათ. ტრანსკრიფცია - გადაწერა) არის რნმ-ის მოლეკულების სინთეზი კომპლემენტარობის პრინციპის მიხედვით დნმ-ის ერთ-ერთი ჯაჭვის შაბლონზე. წარმოიქმნება ბირთვში დნმ-დამოკიდებული ფერმენტის - რნმ პოლიმერაზას მოქმედებით. მესინჯერი რნმ არის ერთი არაჯაჭვიანი მოლეკულა და გენის კოდირება მოდის ორჯაჭვიანი დნმ-ის მოლეკულის ერთი ჯაჭვიდან. თუ ნუკლეოტიდი G არის ტრანსკრიბირებულ დნმ-ის ჯაჭვში, მაშინ დნმ პოლიმერაზა მოიცავს C-ს mRNA-ში; თუ ეს არის T, მაშინ ის მოიცავს A-ს mRNA-ში; თუ ის T არის, მოიცავს Y (თიმინი T არ შედის რნმ-ში; სურ. 5). დნმ-ის ტრიპლეტების ენა ითარგმნება mRNA კოდონების ენაზე (მრნმ-ის სამეულს კოდონებს უწოდებენ).
არაჯაჭვიანი მოლეკულა და გენის კოდირება მოდის ორჯაჭვიანი დნმ-ის მოლეკულის ერთი ჯაჭვიდან. თუ ნუკლეოტიდი G არის ტრანსკრიბირებულ დნმ-ის ჯაჭვში, მაშინ დნმ პოლიმერაზა მოიცავს C-ს mRNA-ში; თუ ეს არის T, მაშინ ის მოიცავს A-ს mRNA-ში; თუ ის T არის, მოიცავს Y (თიმინი T არ შედის რნმ-ში; სურ. 5). დნმ-ის ტრიპლეტების ენა ითარგმნება mRNA კოდონების ენაზე (მრნმ-ის სამეულს კოდონებს უწოდებენ).
სხვადასხვა გენის ტრანსკრიფციის შედეგად სინთეზირდება ყველა სახის რნმ. შემდეგ, mRNA, tRNA, rRNA ბირთვის ფორების მეშვეობით შედიან უჯრედის ციტოპლაზმაში თავიანთი ფუნქციების შესასრულებლად.
3. მაუწყებლობა(ლათ. translatio - გადაცემა, ტრანსლაცია) არის ცილების პოლიპეპტიდური ჯაჭვების სინთეზი მომწიფებულ mRNA მატრიცაზე, რომელსაც ახორციელებს რიბოსომები. ამ პროცესში რამდენიმე ეტაპია:
პირველი ეტაპი არის ინიცირება (სინთეზის დასაწყისი - ჯაჭვები). ციტოპლაზმაში რიბოსომა შედის mRNA-ს ერთ-ერთ ბოლოში (ზუსტად ის, საიდანაც დაიწყო ბირთვში მოლეკულის სინთეზი) და იწყებს პოლიპეპტიდის სინთეზს. tRNA მოლეკულა, რომელიც ახორციელებს ამინომჟავას მეთიონინის (tRNA meth) ტრანსპორტირებას, უკავშირდება რიბოსომას და მიმაგრებულია mRNA ჯაჭვის საწყისთან (ყოველთვის AUG კოდით). პირველი ტრნმ-ის გვერდით (რომელსაც საერთო არაფერი აქვს სინთეზირებელ ცილასთან), მიმაგრებულია მეორე tRNA ამინომჟავით. თუ ანტიკოდონი არის tRNA, მაშინ ამინომჟავებს შორის წარმოიქმნება პეპტიდური ბმა, რომელიც წარმოიქმნება გარკვეული ფერმენტის მიერ. ამის შემდეგ, tRNA ტოვებს რიბოსომას (მიდის ციტოპლაზმაში ახალი ამინომჟავისთვის), ხოლო mRNA მოძრაობს ერთ კოდონს.
მეორე ეტაპი არის დრეკადობა (ჯაჭვის დრეკადობა). რიბოსომა მოძრაობს mRNA მოლეკულის გასწვრივ არა შეუფერხებლად, არამედ პერიოდულად, სამჯერ სამჯერ.  მესამე tRNA ამინომჟავასთან ერთად აკავშირებს თავის ანტიკოდონთან mRNA-ს კოდონს. როდესაც კავშირის კომპლემენტარულობა დამყარდება, რიბოსომა გადადგამს კიდევ ერთ საფეხურს, ერთი „კოდონი“ და სპეციფიური ფერმენტი „ჯვარედინად აკავშირებს“ მეორე და მესამე ამინომჟავებს პეპტიდურ ბმასთან - წარმოიქმნება პეპტიდური ჯაჭვი. მზარდი პოლიპეპტიდური ჯაჭვის ამინომჟავები დაკავშირებულია იმ თანმიმდევრობით, რომელშიც განლაგებულია მათ მაკოდირებელი mRNA კოდონები (ნახ. 6).
მესამე tRNA ამინომჟავასთან ერთად აკავშირებს თავის ანტიკოდონთან mRNA-ს კოდონს. როდესაც კავშირის კომპლემენტარულობა დამყარდება, რიბოსომა გადადგამს კიდევ ერთ საფეხურს, ერთი „კოდონი“ და სპეციფიური ფერმენტი „ჯვარედინად აკავშირებს“ მეორე და მესამე ამინომჟავებს პეპტიდურ ბმასთან - წარმოიქმნება პეპტიდური ჯაჭვი. მზარდი პოლიპეპტიდური ჯაჭვის ამინომჟავები დაკავშირებულია იმ თანმიმდევრობით, რომელშიც განლაგებულია მათ მაკოდირებელი mRNA კოდონები (ნახ. 6).
მესამე ეტაპი არის ჯაჭვის შეწყვეტა (სინთეზის დასასრული). ხდება მაშინ, როდესაც რიბოსომა თარგმნის სამი „უაზრო კოდონიდან“ ერთ-ერთს (UAA, UAG, UGA). რიბოსომები იხტებიან mRNA-დან, ცილის სინთეზი დასრულებულია.
ამრიგად, ცილის მოლეკულაში ამინომჟავების განლაგების თანმიმდევრობის ცოდნით, შესაძლებელია mRNA ჯაჭვში ნუკლეოტიდების (სამების) რიგის დადგენა და მისი მიხედვით, ნუკლეოტიდების წყვილების რიგი დნმ-ის განყოფილებაში და პირიქით, ნუკლეოტიდების კომპლემენტარობის პრინციპის გათვალისწინებით.
ბუნებრივია, მატრიცული რეაქციების პროცესში, ნებისმიერი მიზეზის გამო (ბუნებრივი თუ ხელოვნური) შეიძლება მოხდეს ცვლილებები - მუტაციები. ეს არის გენის მუტაციები მოლეკულურ დონეზე - დნმ-ის მოლეკულების სხვადასხვა დაზიანების შედეგი. მოლეკულურ დონეზე წარმოქმნილი გენის მუტაციები ჩვეულებრივ გავლენას ახდენს ერთ ან მეტ ნუკლეოტიდზე. გენის მუტაციების ყველა ფორმა შეიძლება დაიყოს ორ დიდ ჯგუფად.
პირველი ჯგუფი- ჩარჩოს ცვლა - არის ნუკლეოტიდის ერთი ან მეტი წყვილის ჩასმა ან წაშლა. დარღვევის ადგილიდან გამომდინარე, იცვლება კოდონების ერთი ან სხვა რაოდენობა. ეს არის ყველაზე მძიმე გენის დაზიანება, ვინაიდან ცილაში სრულიად განსხვავებული ამინომჟავები შევა.
ასეთი წაშლა და ჩასმა წარმოადგენს ყველა სპონტანური გენის მუტაციის 80%-ს.
ყველაზე დამაზიანებელ ეფექტს გააჩნია ეგრეთ წოდებული უაზრო მუტაციები, რომლებიც დაკავშირებულია ტერმინატორის კოდონების გამოჩენასთან, რომლებიც იწვევენ გაჩერებას.რომ ცილის სინთეზი. ამან შეიძლება გამოიწვიოს ცილის სინთეზის ნაადრევი შეწყვეტა, რომელიც სწრაფად იშლება. შედეგი არის უჯრედის სიკვდილი ან ინდივიდუალური განვითარების ბუნების ცვლილება.
მუტაციები, რომლებიც დაკავშირებულია გენის კოდირების ნაწილში ჩანაცვლებასთან, წაშლასთან ან ჩასმასთან, ფენოტიპურად ვლინდება როგორც ამინომჟავების ჩანაცვლება ცილაში. ამინომჟავების ბუნებიდან და დაზიანებული უბნის ფუნქციური მნიშვნელობიდან გამომდინარე, ხდება ცილის ფუნქციური აქტივობის სრული ან ნაწილობრივი დაკარგვა. როგორც წესი, ეს გამოიხატება სიცოცხლისუნარიანობის დაქვეითებით, ორგანიზმების მახასიათებლების ცვლილებით და ა.შ.
მეორე ჯგუფიარის გენური მუტაციები ნუკლეოტიდების ბაზის წყვილის ჩანაცვლებით. ბაზის ჩანაცვლების ორი ტიპი არსებობს:
1. გარდამავალი- ერთი პურინის ჩანაცვლება პურინული ფუძისთვის (A G-სთვის ან G-სთვის A) ან ერთი პირიმიდინით პირიმიდინისთვის (C T-სთვის ან T, C-სთვის).
2. ტრანსვერსია- ერთი პურინის ბაზის შეცვლა პირიმიდინის ბაზით ან პირიქით (A C-სთვის, ან G T-სთვის, ან A-სთვის Y).
ტრანსვერსიის თვალსაჩინო მაგალითია ნამგლისებრუჯრედოვანი ანემია, რომელიც ხდება ჰემოგლობინის სტრუქტურაში მემკვიდრეობითი დარღვევის გამო. მუტანტის გენში, რომელიც აკოდირებს ჰემოგლობინის ერთ-ერთ ჯაჭვს, დარღვეულია მხოლოდ ერთი ნუკლეოტიდი, ხოლო ადენინი ჩანაცვლებულია ურაცილით (GAA by GUA) mRNA-ში.
შედეგად ხდება ბიოქიმიური ფენოტიპის ცვლილება, ჰემოგლობინის ჯაჭვში გლუტამინის მჟავა იცვლება ვალინით. ეს ჩანაცვლება ცვლის ჰემოგლობინის მოლეკულის ზედაპირს: ორმხრივ ჩაზნექილი დისკის ნაცვლად, ერითროციტების უჯრედები ნამგლისებრი ხდება და ან ბლოკავს პატარა გემებს ან სწრაფად იხსნება ცირკულაციისგან, რაც სწრაფად იწვევს ანემია.
ამრიგად, გენის მუტაციების მნიშვნელობა ორგანიზმის სიცოცხლისთვის არ არის იგივე:
· ზოგიერთი „ჩუმი მუტაცია“ არ ახდენს გავლენას ცილის სტრუქტურასა და ფუნქციაზე (მაგალითად, ნუკლეოტიდის ჩანაცვლება, რომელიც არ იწვევს ამინომჟავის ჩანაცვლებას);
· ზოგიერთი მუტაცია იწვევს ცილის ფუნქციის სრულ დაკარგვას და უჯრედების სიკვდილს (მაგალითად, უაზრო მუტაციები);
· სხვა მუტაციები - mRNA და ამინომჟავების თვისებრივი ცვლილებით, იწვევს ორგანიზმის მახასიათებლების ცვლილებას;
· და ბოლოს, ზოგიერთი მუტაცია, რომელიც ცვლის ცილის მოლეკულების თვისებებს, საზიანო გავლენას ახდენს უჯრედების სასიცოცხლო აქტივობაზე - ასეთი მუტაციები იწვევს დაავადების მძიმე მიმდინარეობას (მაგალითად, ტრანსვერსიები).
დნმ- ხაზოვანი პოლიმერი, რომელსაც აქვს ორმაგი სპირალის ფორმა, რომელიც წარმოიქმნება წყვილი ანტიპარალელური დამატებითი ჯაჭვებით. დნმ-ის მონომერები არის ნუკლეოტიდები.
თითოეული დნმ ნუკლეოტიდი შედგება პურინის (A - ადენინი ან G - გუანინი) ან პირიმიდინის (T - თიმინი ან C - ციტოზინი) აზოტოვანი ბაზისგან, ხუთნახშირბადოვანი შაქრისგან - დეოქსირიბოზასა და ფოსფატის ჯგუფისგან.
დნმ-ის მოლეკულას აქვს შემდეგი პარამეტრები: სპირალის სიგანე არის დაახლოებით 2 ნმ, სიმაღლე ან სპირალის სრული შემობრუნება არის 3,4 ნმ. ერთი ნაბიჯი შეიცავს 10 დამატებით ბაზის წყვილს.
დნმ-ის მოლეკულაში ნუკლეოტიდები ერთმანეთს უპირისპირდება აზოტოვანი ფუძეებით და წყვილად გაერთიანებულია კომპლემენტარობის წესების შესაბამისად: თიმინი მდებარეობს ადენინის საპირისპიროდ, ხოლო ციტოზინი გუანინის საპირისპიროდ. A-T წყვილი დაკავშირებულია ორი წყალბადის ბმით, ხოლო G-C წყვილი სამით.
დნმ-ის ჯაჭვების ხერხემალი წარმოიქმნება შაქრის ფოსფატის ნარჩენებით.
დნმ-ის რეპლიკაცია არის დნმ-ის მოლეკულის თვითგაორმაგების პროცესი, რომელიც ხორციელდება ფერმენტების კონტროლის ქვეშ.
წყალბადის ბმების გაწყვეტის შემდეგ წარმოქმნილ თითოეულ ჯაჭვზე, დნმ პოლიმერაზას ფერმენტის მონაწილეობით, სინთეზირებულია ქალიშვილი დნმ-ის ჯაჭვი. სინთეზის მასალაა უჯრედების ციტოპლაზმაში არსებული თავისუფალი ნუკლეოტიდები.
მეზობელ ჯაჭვებზე ქალიშვილის მოლეკულების სინთეზი სხვადასხვა სიჩქარით მიმდინარეობს. ერთ ჯაჭვზე უწყვეტად იკრიბება ახალი მოლეკულა, მეორეზე - გარკვეული დაგვიანებით და ფრაგმენტულად. პროცესის დასრულების შემდეგ, ახალი დნმ-ის მოლეკულების ფრაგმენტები უკავშირდება დნმ ლიგაზას ფერმენტს. ასე რომ, ერთი დნმ-ის მოლეკულიდან წარმოიქმნება ორი, რომლებიც ერთმანეთისა და მთავარი მოლეკულის ზუსტი ასლია. ამ ტიპის რეპლიკაციას ეწოდება ნახევრად კონსერვატიული.
რეპლიკაციის ბიოლოგიური მნიშვნელობა მდგომარეობს მემკვიდრეობითი ინფორმაციის ზუსტ გადაცემაში მშობლის მოლეკულიდან ქალიშვილზე, რაც ხდება სომატური უჯრედების დაყოფის დროს.
დნმ-ის შეკეთება- მექანიზმი, რომელიც უზრუნველყოფს დნმ-ის მოლეკულაში ნუილეოტიდების დარღვეული თანმიმდევრობის გამოსწორების უნარს.
თუ დნმ-ის რეპლიკაციის დროს რაიმე მიზეზით ირღვევა ნუკლეოტიდების თანმიმდევრობა მის მოლეკულაში, მაშინ უმეტეს შემთხვევაში ეს დაზიანებები აღმოიფხვრება თავად უჯრედის მიერ. ცვლილება ჩვეულებრივ ხდება დნმ-ის ერთ-ერთ ჯაჭვში. მეორე ჯაჭვი უცვლელი რჩება. პირველი ჯაჭვის დაზიანებული მონაკვეთის „გამოჭრა“ შესაძლებელია ფერმენტების – დნმ-ის აღმდგენი ნუკლეაზების დახმარებით. კიდევ ერთი ფერმენტი, დნმ პოლიმერაზა, აკოპირებს ინფორმაციას ხელუხლებელი ჯაჭვიდან, ათავსებს საჭირო ნუკლეოტიდებს დაზიანებულ ჯაჭვში. ამის შემდეგ დნმ ლიგაზა „ჯვარედინად აკავშირებს“ დნმ-ის მოლეკულას და დაზიანებული მოლეკულა აღდგება.
რნმ - ხაზოვანი პოლიმერი, რომელიც შედგება, როგორც წესი, ნუკლეოტიდების ერთი ჯაჭვისგან. რნმ-ში თიმინის ნუკლეოტიდი ჩანაცვლებულია ურაცილის ნუკლეოტიდით (U). თითოეული რნმ ნუკლეოტიდი შეიცავს ხუთნახშირბადოვან შაქარს - რიბოზას, ოთხი აზოტოვანი ბაზიდან ერთ-ერთს და ფოსფორის მჟავას ნარჩენს.
მატრიქსი ანუ ინფორმაციული რნმ სინთეზირდება ბირთვში ფერმენტ რნმ პოლიმერაზას მონაწილეობით, რომელიც ავსებს დნმ-ის ადგილს, სადაც ხდება სინთეზი და შეადგენს უჯრედის რნმ-ის 5%-ს. რიბოსომური რნმ სინთეზირდება ბირთვში და წარმოადგენს რიბოსომების ნაწილს, რომელიც შეადგენს უჯრედის რნმ-ის 85%-ს. გადამტანი რნმ (40-ზე მეტი ტიპი) ატარებს ამინომჟავებს ცილების სინთეზის ადგილზე, აქვს სამყურას ფორმა და შედგება 70-90 ნუკლეოტიდისგან.

მატრიქსის სინთეზის რეაქციები მოიცავს დნმ-ის რეპლიკაციას, რნმ-ის სინთეზს დნმ-ზე (ტრანსკრიფცია), ცილის სინთეზს mRNA-ზე (თარგმანი) და რნმ ან დნმ-ის სინთეზს ვირუსულ რნმ-ზე.
ტრანსკრიფციის დროს რნმ პოლიმერაზას ფერმენტი ერთვის დნმ ნუკლეოტიდების ჯგუფს – პრომოტორს. პრომოტორი განსაზღვრავს ადგილს, საიდანაც უნდა დაიწყოს mRNA სინთეზი. იგი აგებულია თავისუფალი ნუკლეოტიდებისგან, რომლებიც ავსებენ დნმ-ის მოლეკულას. ფერმენტი მუშაობს მანამ, სანამ არ შეხვდება დნმ-ის ნუკლეოტიდების სხვა ჯგუფს - გაჩერების სიგნალს, რომელიც აცხადებს mRNA სინთეზის დასრულებას.
mRNA მოლეკულა შედის ციტოპლაზმაში რიბოზომებზე, სადაც ხდება პოლიპეპტიდური ჯაჭვების სინთეზი. mRNA ნუკლეოტიდის თანმიმდევრობაში შემავალი ინფორმაციის პოლიპეპტიდში ამინომჟავების თანმიმდევრობით გადაყვანის პროცესს ტრანსლაცია ეწოდება.
გარკვეული ამინომჟავა მიეწოდება რიბოსომებს გარკვეული ტიპის tRNA.
