§ 2. यूकेरियोटिक कोशिका के मुख्य घटक
यूकेरियोटिक कोशिकाएं (चित्र 8 और 9) प्रोकैरियोटिक कोशिकाओं की तुलना में बहुत अधिक जटिल रूप से व्यवस्थित होती हैं। वे आकार में (कई माइक्रोमीटर से लेकर कई सेंटीमीटर तक), आकार में और संरचनात्मक विशेषताओं में बहुत विविध हैं (चित्र 10)।
चावल। 8. यूकेरियोटिक कोशिका की संरचना। सामान्यीकृत योजना
चावल। 9. इलेक्ट्रॉन माइक्रोस्कोपी के अनुसार कोशिका संरचना
चावल। 10. विभिन्न यूकेरियोटिक कोशिकाएँ: 1 - उपकला; 2 - रक्त (ई - एरिथ्रोइटिस, / - ल्यूकाइटिस); 3 - उपास्थि; 4 - हड्डियाँ; 5 - चिकनी मांसपेशी; 6 - संयोजी ऊतक; 7 - तंत्रिका कोशिकाएँ; 8 - धारीदार मांसपेशी फाइबर
हालाँकि, सामान्य संगठन और मूलभूत घटकों की उपस्थिति सभी यूकेरियोटिक कोशिकाओं में समान है (चित्र 11)।
चावल। 11. यूकेरियोटिक कोशिका (आरेख)
प्लाज़्मालेम्मा (बाहरी कोशिका झिल्ली)।प्लाज़्मालेम्मा का आधार, कोशिकाओं में अन्य झिल्लियों (उदाहरण के लिए, माइटोकॉन्ड्रिया, प्लास्टिड, आदि) की तरह, लिपिड की एक परत होती है जिसमें अणुओं की दो पंक्तियाँ होती हैं (चित्र 12)। चूँकि लिपिड अणु ध्रुवीय होते हैं (एक ध्रुव हाइड्रोफिलिक होता है, यानी, पानी से आकर्षित होता है, और दूसरा हाइड्रोफोबिक होता है, यानी, पानी से विकर्षित होता है), वे एक निश्चित क्रम में व्यवस्थित होते हैं। एक परत के अणुओं के हाइड्रोफिलिक सिरे जलीय वातावरण की ओर निर्देशित होते हैं - कोशिका के साइटोप्लाज्म में, और दूसरी परत - कोशिका से बाहर की ओर - अंतरकोशिकीय पदार्थ (बहुकोशिकीय जीवों में) या जलीय वातावरण (एककोशिकीय जीवों में) की ओर निर्देशित होती है। ).
चावल। 12. द्रव मोज़ेक मॉडल के अनुसार कोशिका झिल्ली की संरचना। प्रोटीन और ग्लाइकोप्रोटीन लिपिड अणुओं की दोहरी परत में डूबे होते हैं, उनके हाइड्रोफिलिक सिरे (वृत्त) बाहर की ओर होते हैं, और उनके हाइड्रोफोबिक सिरे (लहरदार रेखाएं) झिल्ली में गहराई की ओर होते हैं।
प्रोटीन अणु लिपिड की द्विआण्विक परत में मोज़ेक रूप से अंतर्निहित होते हैं। पशु कोशिका के बाहर, पॉलीसेकेराइड अणु प्लाज़्मालेम्मा के लिपिड और प्रोटीन अणुओं से जुड़े होते हैं, जिससे ग्लाइकोलिपिड्स और ग्लाइकोप्रोटीन बनते हैं।
यह समुच्चय एक परत बनाता है ग्लाइकोकैलिक्स।उसके साथ जुड़े रिसेप्टर कार्यप्लाज्मा झिल्ली (नीचे देखें); यह कोशिका द्वारा उपयोग किए जाने वाले विभिन्न पदार्थों को भी जमा कर सकता है। इसके अलावा, ग्लाइकोकैलिक्स प्लाज़्मालेम्मा की यांत्रिक स्थिरता को बढ़ाता है।
पौधों और कवक की कोशिकाओं में भी एक कोशिका भित्ति होती है जो सहायक और सुरक्षात्मक भूमिका निभाती है। पौधों में यह सेलूलोज़ से बना होता है, और कवक में यह काइटिन से बना होता है।
बाहरी कोशिका झिल्ली कई कार्य करती है, जिनमें शामिल हैं:
♦ यांत्रिक(सहायक, रूप-निर्माण);
♦ बाधा-परिवहन(विभिन्न पदार्थों के लिए चयनात्मक पारगम्यता: आवश्यक पदार्थों की कोशिका में प्रवेश और अनावश्यक और हानिकारक पदार्थों को हटाना);
♦ रिसेप्टर(कोशिका के निकट पाए जाने वाले विभिन्न रसायनों का निर्धारण; हार्मोन के रूप में संकेतों की धारणा; प्रतिरक्षा प्रणाली की कोशिकाओं द्वारा "विदेशी" प्रोटीन की पहचान, आदि)।
कोशिका और पर्यावरण के बीच पदार्थों का आदान-प्रदान विभिन्न तरीकों से होता है - निष्क्रिय और सक्रिय।
पानी के अणु और विभिन्न आयन निष्क्रिय रूप से (प्रसार, परासरण के कारण), कोशिका द्वारा ऊर्जा खर्च किए बिना, विशेष छिद्रों के माध्यम से प्रवेश करते हैं - ये हैं नकारात्मक परिवहन।प्रोटीन, पॉलीसेकेराइड, यहां तक कि संपूर्ण कोशिकाओं जैसे मैक्रोमोलेक्यूल्स की आपूर्ति की जाती है phagocytosisऔर पिनोसाइटोसिसऊर्जा की खपत के साथ- सक्रिय ट्रांसपोर्ट।
फागोसाइटोसिस द्वारा, संपूर्ण कोशिकाएं या बड़े कण शामिल हो जाते हैं (उदाहरण के लिए, बैक्टीरिया की सुरक्षात्मक रक्त कोशिकाओं द्वारा अमीबा या फागोसाइटोसिस में पोषण के बारे में सोचें)। पिनोसाइटोसिस के दौरान, तरल पदार्थ के छोटे कण या बूंदें अवशोषित हो जाती हैं। दोनों प्रक्रियाओं में सामान्य बात यह है कि अवशोषित पदार्थ रिक्तिका बनाने के लिए एक बाहरी बाहरी झिल्ली से घिरे होते हैं, जो फिर कोशिका के साइटोप्लाज्म में गहराई तक चला जाता है।
एक्सोसाइटोसिस फागोसाइटोसिस और पिनोसाइटोसिस (चित्र 13) की दिशा में विपरीत एक प्रक्रिया है (सक्रिय परिवहन भी)। इसकी सहायता से प्रोटोजोआ में बचे हुए अपच भोजन अथवा स्रावी कोशिका में बने जैविक रूप से सक्रिय पदार्थों को हटाया जा सकता है।
कोशिकाद्रव्य।साइटोप्लाज्म कोशिका की सामग्री है, जो नाभिक के अपवाद के साथ, प्लाज़्मालेम्मा द्वारा सीमित होती है। इसमें है जमीनी पदार्थ (हाइलोप्लाज्म), अंगकऔर समावेशन
हायलोप्लाज्मा- एक चिपचिपा तरल पदार्थ जो किसी भी अवस्था में हो सकता है ज़ोला(तरल), या जेल(जैली समान)।
यदि आवश्यक हो, तो साइटोप्लाज्म एक अवस्था से दूसरी अवस्था में उलटा संक्रमण करने में सक्षम है। उदाहरण के लिए, अमीबॉइड गति के दौरान (जूलॉजी पाठ्यक्रम से अनुभाग "प्रोटोज़ोआ" याद रखें), स्यूडोपोड के निर्माण के दौरान, जेल से सोल तक साइटोप्लाज्म का तेजी से संक्रमण होता है और इसके विपरीत। यह कोशिकाद्रव्य में बड़ी संख्या में फिलामेंटस प्रोटीन अणुओं की उपस्थिति के कारण होता है एक्टिना.जब वे त्रि-आयामी नेटवर्क बनाने के लिए एक-दूसरे से जुड़ते हैं, तो साइटोप्लाज्म जेल अवस्था में होता है, और जब नेटवर्क विघटित होता है, तो यह सोल अवस्था में होता है।
हाइलोप्लाज्म में विभिन्न पदार्थ होते हैं - एंजाइम, प्रोटीन, कार्बोहाइड्रेट, वसा और अन्य, कार्बनिक और खनिज। यहां विभिन्न रासायनिक प्रक्रियाएं होती हैं - पदार्थों का टूटना, उनका संश्लेषण और संशोधन (परिवर्तन)।
ऑर्गेनोइड्स।ये एक निश्चित संरचना और कार्यों के साथ कोशिका के स्थायी घटक होते हैं, जो उसके साइटोप्लाज्म में स्थित होते हैं। आगे हम किस बारे में बात करेंगे सामान्य प्रयोजन ऑर्गेनॉइड,सभी यूकेरियोट्स की सभी प्रकार की कोशिकाओं में निहित। वे बाद के महत्वपूर्ण कार्यों को सुनिश्चित करने से जुड़े हैं। विशेष प्रयोजन ऑर्गेनॉइडकेवल एक निश्चित (अत्यधिक विशिष्ट) प्रकार की कोशिकाओं में पाया जाता है - उदाहरण के लिए, मांसपेशी कोशिकाओं में मायोफिब्रिल्स।
सामान्य प्रयोजन के अंगों की संरचना एक समान होती है, भले ही वे किस कोशिका और किस जीव से संबंधित हों। लेकिन उनमें से झिल्ली वाले समूह भी हैं (एंडोप्लाज्मिक रेटिकुलम, गोल्गी उपकरण, माइटोकॉन्ड्रिया, प्लास्टिड्स, लाइसोसोम, रिक्तिकाएं),साथ ही गैर-झिल्ली ( राइबोसोम, कोशिका केंद्र)संरचना।
एंडोप्लाज्मिक रेटिकुलम (ईआर)।ईपीएस में झिल्लियां होती हैं और यह नलिकाओं और कुंडों की एक जटिल शाखित प्रणाली है जो कोशिका के संपूर्ण साइटोप्लाज्म में प्रवेश करती है (चित्र 14)। ईपीएस दो प्रकार के होते हैं - किसी न किसीऔर चिकना।राइबोसोम खुरदरी झिल्लियों (साइटोप्लाज्मिक पक्ष से) से जुड़े होते हैं, लेकिन वे चिकनी झिल्लियों पर मौजूद नहीं होते हैं।
चावल। 14. एंडोप्लाज्मिक रेटिकुलम
एंडोप्लाज्मिक रेटिकुलम यूकेरियोटिक कोशिका में कई महत्वपूर्ण कार्य करता है:
♦ परिसीमन(कोशिका के आंतरिक आयतन को विभिन्न प्रतिक्रिया स्थानों में विभाजित करना);
♦ कार्बनिक पदार्थों के संश्लेषण में भागीदारी(राइबोसोम खुरदुरे ईआर की झिल्लियों पर स्थित होते हैं, और एंजाइम कॉम्प्लेक्स चिकनी झिल्लियों पर स्थित होते हैं, जो लिपिड, कार्बोहाइड्रेट आदि के संश्लेषण को सुनिश्चित करते हैं);
♦ गोल्गी तंत्र, लाइसोसोम के तत्वों के निर्माण में भागीदारी;
♦ पदार्थों का परिवहन.
गॉल्जीकाय।गोल्गी उपकरण (एजी) एक प्रणाली है टैंक(फ्लैट रिक्तिकाएँ) और बबल(वेसिकल्स), कोशिका नाभिक के निकट स्थित होते हैं, जो इसके छोटे टुकड़ों के अलग होने के परिणामस्वरूप ईपीएस के कारण बनते हैं (चित्र 15)। जब ये टुकड़े विलीन हो जाते हैं, तो गोल्गी तंत्र के नए कुंड उत्पन्न होते हैं, जबकि ईपीएस से विभिन्न पदार्थों का परिवहन होता है, जो जटिल कार्बनिक यौगिकों (प्रोटीन + कार्बोहाइड्रेट, प्रोटीन + लिपिड, आदि) के संयोजन में शामिल होते हैं, जिन्हें हटा दिया जाता है। सेल के बाहर एजी की मदद. ये जैविक रूप से सक्रिय पदार्थ या तो कोशिका से हटा दिए जाते हैं (एक्सोसाइटोसिस द्वारा स्रावी रिक्तिका का उपयोग करके) या एजी द्वारा निर्मित लाइसोसोम (नीचे देखें) का हिस्सा होते हैं।
चावल। 15. गॉल्जी उपकरण:
गोल्गी तंत्र निम्नलिखित कार्य करता है:
♦ संश्लेषणकोशिका द्वारा उत्पादित जैविक रूप से सक्रिय पदार्थ;
♦ विभिन्न पदार्थों का स्राव (कोशिका से निकालना)।(हार्मोन, एंजाइम, पदार्थ जिनसे कोशिका भित्ति बनती है, आदि);
♦ लाइसोसोम के निर्माण में भागीदारी।
माइटोकॉन्ड्रिया।सभी प्रकार की यूकेरियोटिक कोशिकाओं में माइटोकॉन्ड्रिया होता है (चित्र 16)। वे या तो गोल शरीर या छड़ की तरह दिखते हैं, कम अक्सर - धागे की तरह। इनका आकार 1 से 7 माइक्रोन तक होता है। एक कोशिका में माइटोकॉन्ड्रिया की संख्या कई सौ से लेकर हजारों (बड़े प्रोटोजोआ में) तक होती है।
चावल। 16. माइटोकॉन्ड्रिया. ऊपर - मूत्र नलिकाओं में माइटोकॉन्ड्रिया (ए), एक प्रकाश सूक्ष्मदर्शी में दिखाई देता है। नीचे माइटोकॉन्ड्रियल संगठन का त्रि-आयामी मॉडल है: 1 - क्राइस्टे; 2 - बाहरी झिल्ली; 3 - आंतरिक झिल्ली; 4 - मैट्रिक्स
माइटोकॉन्ड्रिया दो झिल्लियों से बनता है - बाहरीऔर आंतरिक,जिसके बीच स्थित है इनतेरमेम्ब्रेन स्पेस।आंतरिक झिल्ली कई आक्रमण-क्रिस्टे बनाती है, जो या तो प्लेट या ट्यूब होती हैं। यह संगठन आंतरिक झिल्ली का एक विशाल क्षेत्र प्रदान करता है। इसमें एंजाइम होते हैं जो कोशिका के जीवन के लिए आवश्यक कार्बनिक पदार्थों (कार्बोहाइड्रेट, लिपिड) में निहित ऊर्जा को एटीपी ऊर्जा में परिवर्तित करना सुनिश्चित करते हैं। इसलिए, माइटोकॉन्ड्रिया का कार्य भाग लेना है ऊर्जासेलुलर प्रक्रियाएं. इसीलिए बड़ी संख्या में माइटोकॉन्ड्रिया अंतर्निहित होते हैं, उदाहरण के लिए, मांसपेशियों की कोशिकाओं में जो बहुत सारा काम करती हैं।
प्लास्टिड्स।पौधों की कोशिकाओं में, विशेष अंग पाए जाते हैं - प्लास्टिड, जो अक्सर धुरी के आकार या गोल आकार के होते हैं, कभी-कभी अधिक जटिल होते हैं। प्लास्टिड तीन प्रकार के होते हैं - क्लोरोप्लास्ट (चित्र 17), क्रोमोप्लास्ट और ल्यूकोप्लास्ट।
क्लोरोप्लास्टहरे रंग में भिन्नता, जो वर्णक के कारण होती है - क्लोरोफिल,प्रक्रिया सुनिश्चित करना प्रकाश संश्लेषण,यानी, सूर्य के प्रकाश की ऊर्जा का उपयोग करके पानी (एच 2 ओ) और कार्बन डाइऑक्साइड (सीओ 2) से कार्बनिक पदार्थों का संश्लेषण। क्लोरोप्लास्ट मुख्यतः पत्ती कोशिकाओं (उच्च पौधों में) में पाए जाते हैं। वे क्लोरोप्लास्ट की सामग्री के चारों ओर एक दूसरे के समानांतर स्थित दो झिल्लियों से बनते हैं - स्ट्रोमा.भीतरी झिल्ली असंख्य चपटी थैली बनाती है - थायलाकोइड्स,जो ढेर लगे हुए हैं (सिक्के के ढेर की तरह) - अनाज -और स्ट्रोमा में लेट जाओ। यह थायलाकोइड्स है जिसमें क्लोरोफिल होता है।
क्रोमोप्लास्टकई फूलों और फलों का पीला, नारंगी और लाल रंग निर्धारित करना, जिनकी कोशिकाओं में वे बड़ी मात्रा में मौजूद होते हैं। उनकी संरचना में मुख्य रंगद्रव्य हैं कैरोटीन.क्रोमोप्लास्ट का कार्यात्मक उद्देश्य जानवरों को रंग से आकर्षित करना, फूलों का परागण और बीजों का फैलाव सुनिश्चित करना है।
चावल। 17. प्लास्टिड्स: ए - एलोडिया पत्ती की कोशिकाओं में क्लोरोप्लास्ट, एक प्रकाश माइक्रोस्कोप में दिखाई देता है; बी - ग्रेना के साथ क्लोरोप्लास्ट की आंतरिक संरचना का आरेख, जो क्लोरोप्लास्ट की सतह पर लंबवत स्थित फ्लैट थैलियों के ढेर हैं; सी - व्यक्तिगत प्रशंसक कक्षों को जोड़ने वाले एनास्टोमोज़िंग ट्यूब दिखाने वाला एक अधिक विस्तृत आरेख
ल्यूकोप्लास्टपौधों के भूमिगत भागों (उदाहरण के लिए, आलू के कंदों में), बीज और तनों के मूल की कोशिकाओं में निहित रंगहीन प्लास्टिड हैं। ल्यूकोप्लास्ट में, स्टार्च मुख्य रूप से ग्लूकोज से बनता है और पौधों के भंडारण अंगों में जमा होता है।
एक प्रकार के प्लास्टिड दूसरे प्रकार में परिवर्तित हो सकते हैं। उदाहरण के लिए, जब पतझड़ में पत्तियाँ रंग बदलती हैं, तो क्लोरोप्लास्ट क्रोमोप्लास्ट में बदल जाते हैं।
लाइसोसोम।ये अंगक 2 माइक्रोन तक के व्यास वाली झिल्ली से घिरे पुटिकाओं की तरह दिखते हैं। उनमें कई दर्जन एंजाइम होते हैं जो प्रोटीन, न्यूक्लिक एसिड, पॉलीसेकेराइड और लिपिड को तोड़ते हैं। लाइसोसोम का कार्य जटिल कार्बनिक यौगिकों (उदाहरण के लिए, "खर्च किए गए" सेलुलर घटकों के पोषक तत्व या पदार्थ) के इंट्रासेल्युलर टूटने की प्रक्रियाओं में भाग लेना है। लाइसोसोम फागोसाइटिक (या पिनोसाइटिक) रिक्तिका के साथ मिलकर एक पाचन रसधानी बनाते हैं।
लाइसोसोम का निर्माण गोल्गी तंत्र के सिस्टर्न से निकलने के कारण होता है।
राइबोसोम.राइबोसोम (चित्र 18) यूकेरियोट्स और प्रोकैरियोट्स दोनों की कोशिकाओं में मौजूद होते हैं, क्योंकि वे एक महत्वपूर्ण कार्य करते हैं प्रोटीन जैवसंश्लेषण(अध्याय 5 देखें)। प्रत्येक कोशिका में दसियों, सैकड़ों हजारों (कई लाखों तक) ये छोटे गोल अंग होते हैं।
चावल। 18. एंडोप्लाज्मिक रेटिकुलम की झिल्ली पर बैठे राइबोसोम की संरचना की योजना: 1 - छोटी सबयूनिट; 2 - टीआरएनए; 3 - एमिनोएसिल-टीआरएनए; 4 - अमीनो एसिड; 5 - बड़ी सबयूनिट; 6 - एंडोप्लाज्मिक रेटिकुलम झिल्ली; 7 - संश्लेषित पॉलीपेप्टाइड श्रृंखला
राइबोसोम में दो असमान उपइकाइयाँ (भाग) होते हैं। वे अलग-अलग बनते हैं और एक प्रोटीन अणु के संश्लेषण के दौरान संदेशवाहक आरएनए को "घेरते हुए" संयोजित होते हैं। राइबोसोम में विभिन्न प्रोटीन और राइबोसोमल आरएनए होते हैं।
सेलुलर समावेशन.यह कोशिका में गैर-स्थायी घटकों को दिया गया नाम है, जो अनाज, कणिकाओं या बूंदों के रूप में साइटोप्लाज्म के मुख्य पदार्थ में मौजूद होते हैं। समावेशन एक झिल्ली से घिरा हो भी सकता है और नहीं भी।
कार्यात्मक रूप से, तीन प्रकार के समावेशन हैं: पोषक तत्वों को सुरक्षित रखें(स्टार्च, ग्लाइकोजन, वसा, प्रोटीन), स्रावी समावेशन(ग्रंथि कोशिकाओं की विशेषता वाले पदार्थ, उनके द्वारा उत्पादित - अंतःस्रावी ग्रंथियों के हार्मोन, आदि) और विशेष प्रयोजन का समावेश(अत्यधिक विशिष्ट कोशिकाओं में, उदाहरण के लिए लाल रक्त कोशिकाओं में हीमोग्लोबिन)।
§ 3. कोशिका केन्द्रक का संगठन। गुणसूत्रों
कोशिका केन्द्रक (चित्र 8 और 9 देखें) कोशिका के जीवन में अत्यंत महत्वपूर्ण है, क्योंकि यह गुणसूत्रों में निहित वंशानुगत जानकारी के भंडार के रूप में कार्य करता है (नीचे देखें)।
नाभिक एक परमाणु आवरण से घिरा होता है जो इसकी सामग्री को अलग करता है (कैरियोप्लाज्म)साइटोप्लाज्म से. खोल में दो झिल्लियाँ होती हैं जो एक अंतराल से अलग होती हैं। ये दोनों असंख्य छिद्रों से व्याप्त हैं, जिनकी बदौलत केन्द्रक और साइटोप्लाज्म के बीच पदार्थों का आदान-प्रदान संभव है। अधिकांश यूकेरियोट्स के कोशिका केन्द्रक में 1 से 7 तक होते हैं न्यूक्लियोली.आरएनए और टीआरएनए संश्लेषण की प्रक्रियाएं इनके साथ जुड़ी हुई हैं।
मुख्य कर्नेल घटक - गुणसूत्र,एक डीएनए अणु और विभिन्न प्रोटीन से निर्मित। प्रकाश सूक्ष्मदर्शी में वे केवल कोशिका विभाजन की अवधि के दौरान ही स्पष्ट रूप से दिखाई देते हैं (माइटोसिस, अर्धसूत्रीविभाजन)।एक गैर-विभाजित कोशिका में, गुणसूत्र नाभिक के पूरे आयतन में वितरित लंबे पतले धागों की तरह दिखते हैं।
कोशिका विभाजन के दौरान, क्रोमोसोमल स्ट्रैंड घने सर्पिल बनाते हैं, जिसके परिणामस्वरूप वे छड़, "हेयरपिन" के रूप में (पारंपरिक माइक्रोस्कोप का उपयोग करके) दिखाई देने लगते हैं। आनुवंशिक जानकारी की संपूर्ण मात्रा नाभिक के गुणसूत्रों के बीच वितरित होती है। उनके अध्ययन की प्रक्रिया में, निम्नलिखित पैटर्न की पहचान की गई:
♦ दैहिक कोशिकाओं (अर्थात, शरीर की कोशिकाएं, गैर-यौन) के नाभिक में एक ही प्रजाति के सभी व्यक्तियों में समान संख्या में गुणसूत्र होते हैं जो बनाते हैं गुणसूत्रों का सेट(चित्र 19);
चावल। 19. पौधों और जानवरों की विभिन्न प्रजातियों के गुणसूत्र, एक ही पैमाने पर दर्शाए गए: 1,2 - अमीबा; 3,4 - डायटम; 5-8, 18,19 - हरा शैवाल; 9 - फ्लाई एगारिक; 10 - लिंडेन; 11-12 - ड्रोसोफिला; 13 - सामन; 14 - स्केरडा (एस्टेरेसी परिवार); 15 - थायरॉइड परिवार का पौधा; 16 - कोरीडेलिस तितली; 17 - टिड्डी परिवार से कीट; 20 - वॉटर स्ट्राइडर बग; 21 - फूल बग; 22 - उभयचर एम्बिस्टोमा; 23 - एलो (लिली परिवार)
♦ प्रत्येक प्रजाति को उनकी संख्या के अनुसार अपने स्वयं के गुणसूत्र सेट की विशेषता होती है (उदाहरण के लिए, एक व्यक्ति में 46 गुणसूत्र होते हैं, एक ड्रोसोफिला मक्खी में 8, एक राउंडवॉर्म में 4, एक क्रेफ़िश में 196, एक घोड़े में 66 और मकई में 104 होते हैं);
♦ दैहिक कोशिकाओं के नाभिक में गुणसूत्रों को जोड़ियों में समूहित किया जा सकता है मुताबिक़ गुणसूत्रोंउनकी समानता के आधार पर (संरचना और कार्य में);
♦ रोगाणु कोशिकाओं (युग्मक) के नाभिक में, समजात गुणसूत्रों की प्रत्येक जोड़ी में केवल एक होता है, अर्थात, गुणसूत्रों का कुल सेट दैहिक कोशिकाओं का आधा होता है;
♦ जनन कोशिकाओं में गुणसूत्रों के एकल सेट को कहा जाता है अगुणितऔर अक्षर n द्वारा निरूपित किया जाता है, और दैहिक में - द्विगुणित(2एन).
उपरोक्त से यह स्पष्ट है कि समजात गुणसूत्रों का प्रत्येक जोड़ा निषेचन के दौरान पैतृक और मातृ गुणसूत्रों के मिलन से बनता है, अर्थात, सेक्स कोशिकाओं (युग्मक) के संलयन से। इसके विपरीत, जनन कोशिकाओं के निर्माण के दौरान, समजात गुणसूत्रों के प्रत्येक जोड़े से केवल एक ही युग्मक में प्रवेश करता है।
गुणसूत्रों अलगसजातीय जोड़े आकार और आकार में भिन्न होते हैं (चित्र 20 और 21)।
चावल। 20. गुणसूत्रों की संरचना और प्रकार: ए - उपस्थिति 1 - सेंट्रोमियर; 2 - छोटा कंधा; 3 - लंबा कंधा); एक ही गुणसूत्र की आंतरिक संरचना (1 - सेंट्रोमियर; - डीएनए अणु); सी - गुणसूत्रों के प्रकार (1 - एकल-सशस्त्र; भिन्न-सशस्त्र; 3 - समान-सशस्त्र: एक्स - भुजा, वी - सेंट्रोमियर)
चावल। 21. एक गुणसूत्र में DNA और प्रोटीन होते हैं। डीएनए अणु की प्रतिकृति बनाई जाती है। सेंट्रोमियर क्षेत्र में दो समान डीएनए डबल हेलिकॉप्टर जुड़े रहते हैं। ये प्रतियां बाद में कोशिका विभाजन के दौरान अलग-अलग गुणसूत्र बन जाती हैं
शरीर में गुणसूत्र होते हैं प्राथमिक संकुचन (जिसे सेंट्रोमियर कहा जाता है),जिससे धागे जुड़े हुए हैं विखंडन धुरी.यह गुणसूत्र को दो भागों में विभाजित करता है कंधागुणसूत्र समान-सशस्त्र, असमान-सशस्त्र या एकल-सशस्त्र हो सकते हैं।
अध्याय 5. चयापचय
§ 1. आत्मसात और प्रसार की एकता के रूप में चयापचय
सभी कोशिकाएँ और जीवित जीव खुली प्रणालियाँ हैं, अर्थात वे पर्यावरण के साथ ऊर्जा और पदार्थों के निरंतर आदान-प्रदान की स्थिति में हैं। निर्जीव प्रकृति में खुली प्रणालियाँ हैं, लेकिन उनका अस्तित्व जीवित जीवों से गुणात्मक रूप से भिन्न है। इस उदाहरण पर विचार करें: देशी सल्फर का एक जलता हुआ टुकड़ा पर्यावरण के साथ आदान-प्रदान की स्थिति में है। जब यह जलता है, तो O 2 अवशोषित हो जाता है, और SO 2 और ऊर्जा (गर्मी के रूप में) निकल जाती है। हालाँकि, इस मामले में, भौतिक शरीर के रूप में सल्फर का एक टुकड़ा नष्ट हो जाता है और अपनी प्राथमिक संरचना खो देता है।
जीवित जीवों के लिए, पर्यावरण के साथ आदान-प्रदान उन सभी पदार्थों और घटकों के आत्म-नवीकरण के माध्यम से उनके संरचनात्मक संगठन को संरक्षित करने और बनाए रखने के लिए एक शर्त बन जाता है, जिनसे वे बने होते हैं।
चयापचय (चयापचय) जीवित जीवों (उपभोग, परिवर्तन, संचय और पदार्थों और ऊर्जा की रिहाई) में होने वाली प्रक्रियाओं का एक सेट है जो उनकी महत्वपूर्ण गतिविधि, विकास, विकास, प्रजनन सुनिश्चित करता है। चयापचय प्रक्रिया के दौरान, कोशिकाओं को बनाने वाले अणु टूट जाते हैं और संश्लेषित होते हैं; सेलुलर संरचनाओं और अंतरकोशिकीय पदार्थ का नवीनीकरण।
मेटाबॉलिज्म परस्पर जुड़ी प्रक्रियाओं पर आधारित है मिलाना(उपचय) और भेद(अपचय)। आत्मसात (प्लास्टिक विनिमय) के दौरान, जटिल पदार्थों को सरल पदार्थों से संश्लेषित किया जाता है। यह इसके लिए धन्यवाद है कि कोशिका में इसके संरचनात्मक घटकों, एंजाइम सिस्टम आदि के निर्माण के लिए आवश्यक सभी कार्बनिक पदार्थ बनते हैं। आत्मसात्करण हमेशा ऊर्जा व्यय के साथ किया जाता है।
प्रसार (ऊर्जा चयापचय) के दौरान, जटिल कार्बनिक पदार्थ सरल या अकार्बनिक में टूट जाते हैं। इस मामले में, ऊर्जा जारी की जाती है, जिसे कोशिका द्वारा विभिन्न प्रक्रियाओं को करने के लिए खर्च किया जाता है जो इसकी महत्वपूर्ण गतिविधि (पदार्थों का संश्लेषण और परिवहन, यांत्रिक कार्य, आदि) सुनिश्चित करती हैं।
सभी जीवित जीवों को दो समूहों में विभाजित किया जा सकता है: स्वपोषकऔर विषमपोषी,जो अपने महत्वपूर्ण कार्यों को सुनिश्चित करने के लिए ऊर्जा के स्रोतों और आवश्यक पदार्थों में भिन्न होते हैं।
स्वपोषक- जीव जो सूर्य के प्रकाश की ऊर्जा का उपयोग करके अकार्बनिक पदार्थों से कार्बनिक यौगिकों का संश्लेषण करते हैं (जैसे)। फोटोट्रॉफ़्स- पौधे, साइनोबैक्टीरिया) या खनिज (अकार्बनिक) पदार्थों (जैसे) के ऑक्सीकरण से प्राप्त ऊर्जा रसोपोषी- सल्फर बैक्टीरिया, आयरन बैक्टीरिया, आदि)। नतीजतन, वे अपनी जीवन गतिविधि के लिए आवश्यक पदार्थों को स्वतंत्र रूप से बनाने में सक्षम हैं।
§ 2. अवायवीय और एरोबिक जीवों में विसंकरण
जीवों को विसंकरण की प्रकृति के अनुसार दो समूहों में विभाजित किया जा सकता है - एरोबिक्सऔर अवायवीय.एरोबेस (ग्रीक से। राक्षस– वायु) को जीवन के लिए निःशुल्क ऑक्सीजन की आवश्यकता होती है। अवायवीय जीवों में (ग्रीक। एसी– ऋणात्मक कण) यह आवश्यक नहीं है। उनमें, किण्वन द्वारा विघटन किया जाता है - सरल कार्बनिक पदार्थों के निर्माण और ऊर्जा की रिहाई के साथ कार्बनिक पदार्थों का ऑक्सीजन मुक्त, एंजाइमेटिक टूटना। उदाहरण के लिए:
♦ लैक्टिक एसिड किण्वन:
सी 6 एच 12 ओ 6 + 2एच 3 पीओ 4 + 2एडीपी → 2पीएच + 2एटीपी + 2एच 2 ओ;
♦ अल्कोहलिक किण्वन:
सी 6 एच 12 ओ 6 + 2पीएच + 2एडीपी → 2सी 2 एच 5 ओएच + 2सीओ 2 + 2एटीपी + 2एच 2 ओ।
किण्वन के दौरान बनने वाले पदार्थ कार्बनिक होते हैं और इसलिए उनमें अभी भी बहुत अधिक ऊर्जा होती है।
चावल। 22. स्वपोषी और विषमपोषी जीवों में स्वांगीकरण और असमीकरण के बीच संबंध
एरोबिक जीवों में, माइटोकॉन्ड्रिया में श्वसन की प्रक्रिया के दौरान, कार्बनिक पदार्थों का पूर्ण विघटन (ओ 2 का उपयोग करके) ऊर्जा-खराब अंतिम उत्पादों सीओ 2 और एच 2 ओ में होता है और काफी बड़ी मात्रा में ऊर्जा निकलती है:
C 6 H 12 0 6 (ग्लूकोज) + 0 2 > 6C0 2 + 6H 2 0 + ऊर्जा (जिससे 38 एटीपी अणु संश्लेषित होते हैं)।
आइए सामान्यीकृत योजनाओं के रूप में ऑटोट्रॉफ़िक और हेटरोट्रॉफ़िक एरोबिक जीवों के चयापचय पर विचार करें (चित्र 22)।
मिलाना।इसकी सबसे महत्वपूर्ण प्रक्रियाएं हैं प्रकाश संश्लेषणऔर प्रोटीन जैवसंश्लेषण.
भौतिक-रासायनिक दृष्टिकोण से पर्यावरण के साथ जीव का संबंध एक खुली प्रणाली है, यानी एक ऐसी प्रणाली जहां जैव रासायनिक प्रक्रियाएं चल रही हैं। प्रारंभिक पदार्थ पर्यावरण से आते हैं, और जो पदार्थ लगातार बनते रहते हैं वे भी बाहर ले जाये जाते हैं। शरीर में बहुदिशात्मक प्रतिक्रियाओं के उत्पादों की गति और एकाग्रता के बीच संतुलन सशर्त, काल्पनिक है, क्योंकि पदार्थों का सेवन और निष्कासन बंद नहीं होता है। पर्यावरण के साथ निरंतर संबंध हमें एक जीवित जीव को एक खुली प्रणाली के रूप में मानने की अनुमति देता है।
सभी जीवित कोशिकाओं के लिए ऊर्जा का स्रोत सूर्य है। पादप कोशिकाएँ क्लोरोफिल की मदद से सूर्य के प्रकाश से ऊर्जा ग्रहण करती हैं, और इसका उपयोग प्रकाश संश्लेषण की प्रक्रिया के दौरान आत्मसात प्रतिक्रियाओं के लिए करती हैं। जानवरों, कवक और बैक्टीरिया की कोशिकाएं, सांसारिक पौधों द्वारा संश्लेषित कार्बनिक पदार्थों के टूटने के दौरान, अप्रत्यक्ष रूप से सौर ऊर्जा का उपयोग करती हैं।
कोशिकीय श्वसन के दौरान कोशिका के कुछ पोषक तत्व टूट जाते हैं, जिससे विभिन्न प्रकार की कोशिकीय गतिविधियों के लिए आवश्यक ऊर्जा की आपूर्ति होती है। यह प्रक्रिया माइटोकॉन्ड्रिया नामक अंगकों में होती है। माइटोकॉन्ड्रिया में दो झिल्लियाँ होती हैं: बाहरी एक, कोशिकांग को साइटोप्लाज्म से अलग करती है, और भीतरी एक, जो कई तह बनाती है। श्वसन का मुख्य उत्पाद एटीपी है। यह माइटोकॉन्ड्रिया को छोड़ देता है और साइटोप्लाज्म और कोशिका झिल्ली में कई रासायनिक प्रतिक्रियाओं के लिए ऊर्जा स्रोत के रूप में उपयोग किया जाता है। यदि कोशिकीय श्वसन के लिए ऑक्सीजन की आवश्यकता होती है, तो श्वसन को एरोबिक श्वसन कहा जाता है, लेकिन यदि प्रतिक्रियाएं ऑक्सीजन की अनुपस्थिति में होती हैं, तो हम अवायवीय श्वसन कहते हैं।
किसी कोशिका में किए जाने वाले किसी भी प्रकार के कार्य के लिए, ऊर्जा का उपयोग केवल एक ही रूप में किया जाता है - एटीपी के फॉस्फेट बांड से ऊर्जा के रूप में। एटीपी एक आसानी से गतिशील यौगिक है। एटीपी का निर्माण माइटोकॉन्ड्रिया की आंतरिक झिल्ली पर होता है। कार्बोहाइड्रेट, वसा और अन्य कार्बनिक पदार्थों के ऑक्सीकरण की ऊर्जा के कारण श्वसन के दौरान सभी कोशिकाओं में एटीपी का संश्लेषण होता है। हरे पौधों की कोशिकाओं में, एटीपी की मुख्य मात्रा सौर ऊर्जा के कारण क्लोरोप्लास्ट में संश्लेषित होती है। प्रकाश संश्लेषण के दौरान, वे माइटोकॉन्ड्रिया की तुलना में कई गुना अधिक एटीपी का उत्पादन करते हैं। फॉस्फोरस-ऑक्सीजन बांड के टूटने और ऊर्जा की रिहाई के साथ एटीपी विघटित हो जाता है। यह एटीपी के हाइड्रोलिसिस के दौरान एंजाइम एटीपीस की कार्रवाई के तहत होता है - फॉस्फोरिक एसिड अणु के उन्मूलन के साथ पानी जोड़ना। परिणामस्वरूप, एटीपी को एडीपी में बदल दिया जाता है, और यदि फॉस्फोरिक एसिड के दो अणुओं को विभाजित किया जाता है, तो एएमपी में बदल दिया जाता है। एसिड के प्रत्येक ग्राम-अणु के उन्मूलन की प्रतिक्रिया 40 kJ की रिहाई के साथ होती है। यह एक बहुत बड़ा ऊर्जा उत्पादन है, यही कारण है कि एटीपी के फॉस्फोरस-ऑक्सीजन बांड को आमतौर पर मैक्रोर्जिस्टिक (उच्च-ऊर्जा) कहा जाता है।
प्लास्टिक विनिमय प्रतिक्रियाओं में एटीपी का उपयोग एटीपी हाइड्रोलिसिस के साथ जोड़कर किया जाता है। एटीपी अणु से हाइड्रोलिसिस के दौरान निकलने वाले फॉस्फोरस समूह को जोड़कर, यानी फॉस्फोराइलेशन द्वारा विभिन्न पदार्थों के अणुओं को ऊर्जा से चार्ज किया जाता है।
फॉस्फेट डेरिवेटिव की ख़ासियत यह है कि वे कोशिका को नहीं छोड़ सकते हैं, हालांकि उनके "डिस्चार्ज" रूप झिल्ली से स्वतंत्र रूप से गुजरते हैं। इसके कारण, फॉस्फोराइलेटेड अणु कोशिका में तब तक बने रहते हैं जब तक कि उनका उचित प्रतिक्रियाओं में उपयोग नहीं किया जाता है।
एडीपी को एटीपी में परिवर्तित करने की विपरीत प्रक्रिया एडीपी में फॉस्फोरिक एसिड अणु जोड़ने, पानी छोड़ने और बड़ी मात्रा में ऊर्जा को अवशोषित करने से होती है।
इस प्रकार, एटीपी कोशिका गतिविधि के लिए ऊर्जा का एक सार्वभौमिक और प्रत्यक्ष स्रोत है। यह ऊर्जा का एक एकल कोशिकीय कोष बनाता है और इसे कोशिका के एक क्षेत्र से दूसरे क्षेत्र में पुनर्वितरित और परिवहन करना संभव बनाता है।
फॉस्फेट समूह का स्थानांतरण रासायनिक प्रतिक्रियाओं जैसे मोनोमर्स से मैक्रोमोलेक्यूल्स की असेंबली में महत्वपूर्ण भूमिका निभाता है। उदाहरण के लिए, पहले फॉस्फोराइलेटेड होने के बाद ही अमीनो एसिड को पेप्टाइड्स में जोड़ा जा सकता है। संकुचन या गति की यांत्रिक प्रक्रियाओं, एक सांद्रता प्रवणता के विरुद्ध एक विघटित पदार्थ का परिवहन और अन्य प्रक्रियाओं में एटीपी में संग्रहीत ऊर्जा की खपत शामिल होती है।
ऊर्जा चयापचय की प्रक्रिया को निम्नानुसार दर्शाया जा सकता है। साइटोप्लाज्म में उच्च-आणविक कार्बनिक पदार्थ एंजाइमेटिक रूप से, हाइड्रोलिसिस द्वारा, सरल पदार्थों में परिवर्तित हो जाते हैं, जिनसे वे बनते हैं: प्रोटीन - अमीनो एसिड में, पॉली- और डिसैकराइड - मोनोसेकेराइड (+ ग्लूकोज) में, वसा ग्लिसरॉल और फैटी एसिड में। कोई ऑक्सीडेटिव प्रक्रियाएं नहीं होती हैं, थोड़ी ऊर्जा निकलती है, जिसका उपयोग नहीं किया जाता है और थर्मल रूप में चला जाता है। अधिकांश कोशिकाएँ पहले कार्बोहाइड्रेट का उपयोग करती हैं। पॉलीसेकेराइड (पौधों में स्टार्च और जानवरों में ग्लाइकोजन) ग्लूकोज में हाइड्रोलाइज्ड होते हैं। ग्लूकोज ऑक्सीकरण तीन चरणों में होता है: ग्लाइकोलाइसिस, ऑक्सीडेटिव डीकार्बाक्सिलेशन (क्रेब्स चक्र - साइट्रिक एसिड चक्र) और ऑक्सीडेटिव फॉस्फोराइलेशन (श्वसन श्रृंखला)। ग्लाइकोलाइसिस, जिसके परिणामस्वरूप ग्लूकोज का एक अणु एटीपी के दो अणुओं की रिहाई के साथ पाइरुविक एसिड के दो अणुओं में विभाजित हो जाता है, साइटोप्लाज्म में होता है। ऑक्सीजन की अनुपस्थिति में, पाइरुविक एसिड या तो इथेनॉल (किण्वन) या लैक्टिक एसिड (एनारोबिक श्वसन) में परिवर्तित हो जाता है।
जब पशु कोशिकाओं में ग्लाइकोलाइसिस होता है, तो ग्लूकोज का छह-कार्बन अणु लैक्टिक एसिड के दो अणुओं में टूट जाता है। यह प्रक्रिया बहु-चरणीय है. यह 13 एंजाइमों द्वारा क्रमिक रूप से संचालित होता है। अल्कोहलिक किण्वन के दौरान, ग्लूकोज अणु से इथेनॉल के दो अणु और CO2 के दो अणु बनते हैं।
ग्लाइकोलाइसिस अवायवीय और एरोबिक श्वसन के लिए सामान्य चरण है; अन्य दो केवल एरोबिक स्थितियों में होते हैं। ऑक्सीजन मुक्त ऑक्सीकरण की प्रक्रिया, जिसमें चयापचयों की ऊर्जा का केवल एक हिस्सा जारी और उपयोग किया जाता है, अवायवीय जीवों के लिए अंतिम है। ऑक्सीजन की उपस्थिति में, पाइरुविक एसिड माइटोकॉन्ड्रिया में चला जाता है, जहां, कई अनुक्रमिक प्रतिक्रियाओं के परिणामस्वरूप, यह एडीपी से एटीपी के फॉस्फोराइलेशन के साथ-साथ एच 2 ओ और सीओ 2 में एरोबिक रूप से पूरी तरह से ऑक्सीकृत हो जाता है। इस मामले में, दो एटीपी अणु ग्लाइकोलाइसिस द्वारा, दो क्रेब्स चक्र द्वारा और 34 श्वसन श्रृंखला द्वारा निर्मित होते हैं। एक ग्लूकोज अणु के H2O और CO2 में पूर्ण ऑक्सीकरण के लिए शुद्ध उपज 38 अणु है।
इस प्रकार, एरोबिक जीवों में, कार्बनिक पदार्थों का अंतिम अपघटन उन्हें वायुमंडलीय ऑक्सीजन के साथ सरल अकार्बनिक पदार्थों में ऑक्सीकरण करके किया जाता है: CO2 और H2O। यह प्रक्रिया माइटोकॉन्ड्रिया के क्राइस्टे पर होती है। इस मामले में, मुक्त ऊर्जा की अधिकतम मात्रा जारी होती है, जिसका एक महत्वपूर्ण हिस्सा एटीपी अणुओं में आरक्षित होता है। यह देखना आसान है कि एरोबिक ऑक्सीकरण कोशिका को अधिकतम सीमा तक मुक्त ऊर्जा प्रदान करता है।
अपचय के परिणामस्वरूप, ऊर्जा से भरपूर एटीपी अणु कोशिका में जमा हो जाते हैं, और CO2 और अतिरिक्त पानी बाहरी वातावरण में छोड़ दिया जाता है।
श्वसन के लिए आवश्यक चीनी अणुओं को कोशिका में संग्रहित किया जा सकता है। अतिरिक्त लिपिड या तो टूट जाते हैं, जिसके बाद उनके टूटने के उत्पाद श्वसन के लिए सब्सट्रेट के रूप में माइटोकॉन्ड्रिया में प्रवेश करते हैं, या वसा की बूंदों के रूप में साइटोप्लाज्म में भंडार के रूप में जमा होते हैं। प्रोटीन कोशिका में प्रवेश करने वाले अमीनो एसिड से निर्मित होते हैं। प्रोटीन संश्लेषण राइबोसोम नामक अंगकों में होता है। प्रत्येक राइबोसोम में दो उपकण होते हैं - बड़े और छोटे: दोनों उपकणों में प्रोटीन अणु और आरएनए अणु शामिल होते हैं।
राइबोसोम अक्सर सिस्टर्न और वेसिकल्स से युक्त एक विशेष झिल्ली प्रणाली से जुड़े होते हैं - तथाकथित एंडोप्लाज्मिक रेटिकुलम (ईआर); उन कोशिकाओं में जो बहुत अधिक प्रोटीन का उत्पादन करती हैं, एंडोप्लाज्मिक रेटिकुलम अक्सर बहुत अच्छी तरह से विकसित होता है और राइबोसोम से ढका होता है। कुछ एंजाइम केवल तभी प्रभावी होते हैं जब वे किसी झिल्ली से जुड़े होते हैं। लिपिड संश्लेषण में शामिल अधिकांश एंजाइम यहीं स्थित हैं। इस प्रकार, एंडोप्लाज्मिक रेटिकुलम एक प्रकार की कोशिका कार्यक्षेत्र की तरह है।
इसके अलावा, ईआर साइटोप्लाज्म को अलग-अलग हिस्सों में विभाजित करता है, यानी, यह साइटोप्लाज्म में एक साथ होने वाली विभिन्न रासायनिक प्रक्रियाओं को अलग करता है, और इस तरह यह संभावना कम हो जाती है कि ये प्रक्रियाएं एक-दूसरे के साथ हस्तक्षेप करेंगी।
अक्सर किसी सेल द्वारा उत्पादित उत्पादों का उपयोग उस सेल के बाहर किया जाता है। ऐसे मामलों में, राइबोसोम पर संश्लेषित प्रोटीन एंडोप्लाज्मिक रेटिकुलम की झिल्लियों से होकर गुजरते हैं और उनके चारों ओर बनने वाली झिल्ली पुटिकाओं में पैक हो जाते हैं, जो फिर ईआर से अलग हो जाते हैं। ये पुटिकाएँ, चपटी और एक-दूसरे के ऊपर खड़ी, पैनकेक की तरह, एक विशिष्ट संरचना बनाती हैं जिसे गोल्गी कॉम्प्लेक्स या गोल्गी तंत्र कहा जाता है। गोल्गी तंत्र में रहने के दौरान, प्रोटीन में कुछ परिवर्तन होते हैं। जब उनके कोशिका छोड़ने का समय आता है, तो झिल्ली पुटिकाएं कोशिका झिल्ली में विलीन हो जाती हैं और खाली हो जाती हैं, जिससे उनकी सामग्री बाहर निकल जाती है, यानी एक्सोसाइटोसिस द्वारा स्राव होता है।
गोल्गी तंत्र लाइसोसोम का भी उत्पादन करता है - झिल्ली थैली जिसमें पाचन एंजाइम होते हैं। यह पता लगाना कि कोई कोशिका कुछ प्रोटीन कैसे बनाती है, पैकेज करती है और निर्यात करती है, और यह कैसे "जानती है" कि उसे कौन से प्रोटीन अपने पास रखना चाहिए, आधुनिक कोशिका विज्ञान की सबसे आकर्षक शाखाओं में से एक है।
किसी भी कोशिका की झिल्लियाँ लगातार गतिशील और परिवर्तित होती रहती हैं। ईआर झिल्ली पूरे कोशिका में धीरे-धीरे चलती है। इन झिल्लियों के अलग-अलग खंड अलग हो जाते हैं और पुटिकाओं का निर्माण करते हैं, जो अस्थायी रूप से गोल्गी तंत्र का हिस्सा बन जाते हैं, और फिर, एक्सोसाइटोसिस की प्रक्रिया के माध्यम से, कोशिका झिल्ली के साथ विलय हो जाते हैं।
बाद में, झिल्ली सामग्री को साइटोप्लाज्म में वापस कर दिया जाता है, जहां इसका फिर से उपयोग किया जाता है।
कोशिका में प्रवेश करने वाले या उसके द्वारा बाहर छोड़े गए पदार्थों का आदान-प्रदान, साथ ही सूक्ष्म और स्थूल वातावरण के साथ विभिन्न संकेतों का आदान-प्रदान, कोशिका की बाहरी झिल्ली के माध्यम से होता है। जैसा कि ज्ञात है, कोशिका झिल्ली एक लिपिड बाईलेयर है जिसमें विभिन्न प्रोटीन अणु अंतर्निहित होते हैं जो विशेष रिसेप्टर्स, आयन चैनल, उपकरणों के रूप में कार्य करते हैं जो विभिन्न रसायनों, अंतरकोशिकीय संपर्कों आदि को सक्रिय रूप से परिवहन या हटाते हैं। स्वस्थ यूकेरियोटिक कोशिकाओं में, फॉस्फोलिपिड वितरित होते हैं झिल्ली असममित रूप से: बाहरी सतह में स्फिंगोमाइलिन और फॉस्फेटिडिलकोलाइन होते हैं, आंतरिक सतह - फॉस्फेटिडिलसेरिन और फॉस्फेटिडाइलथेनॉलमाइन से बनी होती है। ऐसी विषमता बनाए रखने के लिए ऊर्जा व्यय की आवश्यकता होती है। इसलिए, कोशिका क्षति, संक्रमण या ऊर्जा भुखमरी की स्थिति में, झिल्ली की बाहरी सतह फॉस्फोलिपिड्स से समृद्ध होती है जो इसके लिए असामान्य हैं, जो इसी प्रतिक्रिया के साथ कोशिका क्षति के बारे में अन्य कोशिकाओं और एंजाइमों के लिए एक संकेत बन जाती है। सबसे महत्वपूर्ण भूमिका फॉस्फोलिपेज़ ए2 के घुलनशील रूप द्वारा निभाई जाती है, जो एराकिडोनिक एसिड को तोड़ता है और उपर्युक्त फॉस्फोलिपिड्स से लाइसोफॉर्म बनाता है। एराकिडोनिक एसिड ईकोसैनोइड्स जैसे सूजन मध्यस्थों के निर्माण के लिए सीमित लिंक है, और सुरक्षात्मक अणु - पेंट्राक्सिन (सी-रिएक्टिव प्रोटीन (सीआरपी), अमाइलॉइड प्रोटीन के अग्रदूत) - झिल्ली में लाइसोफॉर्म से जुड़े होते हैं, जिसके बाद पूरक सक्रिय होता है शास्त्रीय मार्ग और कोशिका विनाश के साथ प्रणाली।
झिल्ली की संरचना कोशिका के आंतरिक वातावरण की विशेषताओं, बाहरी वातावरण से इसके अंतर को संरक्षित करने में मदद करती है। यह कोशिका झिल्ली की चयनात्मक पारगम्यता और उसमें सक्रिय परिवहन तंत्र के अस्तित्व द्वारा सुनिश्चित किया जाता है। प्रत्यक्ष क्षति के परिणामस्वरूप उनका विघटन, उदाहरण के लिए, टेट्रोडोटॉक्सिन, ओबैन, टेट्राएथिलमोनियम द्वारा, या संबंधित "पंप" को अपर्याप्त ऊर्जा आपूर्ति के मामले में सेल की इलेक्ट्रोलाइट संरचना में व्यवधान होता है, इसके चयापचय में परिवर्तन होता है, व्यवधान होता है विशिष्ट कार्यों का - संकुचन, उत्तेजना आवेगों का संचालन, आदि। मनुष्यों में सेलुलर आयन चैनलों (कैल्शियम, सोडियम, पोटेशियम और क्लोराइड) का विघटन आनुवंशिक रूप से इन चैनलों की संरचना के लिए जिम्मेदार जीन में उत्परिवर्तन द्वारा निर्धारित किया जा सकता है। तथाकथित चैनलोपैथी तंत्रिका, मांसपेशियों और पाचन तंत्र के वंशानुगत रोगों का कारण बनती है। कोशिका में पानी के अत्यधिक प्रवेश से इसका टूटना हो सकता है - साइटोलिसिस - पूरक सक्रिय होने पर झिल्ली के छिद्र के कारण या साइटोटॉक्सिक लिम्फोसाइट्स और प्राकृतिक हत्यारा कोशिकाओं के हमले के कारण।
कोशिका झिल्ली में कई रिसेप्टर्स निर्मित होते हैं - संरचनाएं, जो संबंधित विशिष्ट सिग्नलिंग अणुओं (लिगैंड्स) के साथ मिलकर कोशिका के अंदर एक सिग्नल संचारित करती हैं। यह विभिन्न नियामक कैस्केड के माध्यम से होता है जिसमें एंजाइमेटिक रूप से सक्रिय अणु होते हैं जो क्रमिक रूप से सक्रिय होते हैं और अंततः विभिन्न सेलुलर कार्यक्रमों के कार्यान्वयन में योगदान करते हैं, जैसे कि विकास और प्रसार, भेदभाव, गतिशीलता, उम्र बढ़ने और कोशिका मृत्यु। विनियामक कैस्केड काफी संख्या में हैं, लेकिन उनकी संख्या अभी तक पूरी तरह से निर्धारित नहीं की गई है। रिसेप्टर्स और उनसे जुड़े नियामक कैस्केड की प्रणाली भी कोशिका के अंदर मौजूद होती है; वे सेल की कार्यात्मक स्थिति, इसके विकास के चरण और अन्य रिसेप्टर्स से सिग्नल की एक साथ कार्रवाई के आधार पर आगे के सिग्नल पथ के एकाग्रता, वितरण और चयन के बिंदुओं के साथ एक विशिष्ट नियामक नेटवर्क बनाते हैं। इसका परिणाम सिग्नल का अवरोध या सुदृढ़ीकरण हो सकता है, इसे एक अलग नियामक मार्ग पर निर्देशित किया जा सकता है। नियामक कैस्केड के माध्यम से रिसेप्टर तंत्र और सिग्नल ट्रांसडक्शन मार्ग दोनों, उदाहरण के लिए नाभिक, एक आनुवंशिक दोष के परिणामस्वरूप बाधित हो सकते हैं जो जीव स्तर पर जन्मजात दोष के रूप में या एक विशिष्ट कोशिका प्रकार में दैहिक उत्परिवर्तन के कारण होता है। ये तंत्र संक्रामक एजेंटों, विषाक्त पदार्थों से क्षतिग्रस्त हो सकते हैं और उम्र बढ़ने की प्रक्रिया के दौरान भी बदल सकते हैं। इसका अंतिम चरण कोशिका के कार्यों, उसके प्रसार और विभेदन की प्रक्रियाओं में व्यवधान हो सकता है।
कोशिकाओं की सतह पर ऐसे अणु भी होते हैं जो अंतरकोशिकीय संपर्क की प्रक्रियाओं में महत्वपूर्ण भूमिका निभाते हैं। इनमें कोशिका आसंजन प्रोटीन, हिस्टोकम्पैटिबिलिटी एंटीजन, ऊतक-विशिष्ट, विभेदक एंटीजन आदि शामिल हो सकते हैं। इन अणुओं की संरचना में परिवर्तन से अंतरकोशिकीय अंतःक्रिया में व्यवधान होता है और ऐसी कोशिकाओं के उन्मूलन के लिए उपयुक्त तंत्र के सक्रियण का कारण बन सकता है, क्योंकि वे एक उत्पन्न करते हैं। संक्रमण के भंडार के रूप में, विशेष रूप से वायरल, या ट्यूमर के विकास के संभावित आरंभकर्ताओं के रूप में शरीर की अखंडता के लिए कुछ खतरा।
सेल की ऊर्जा आपूर्ति का उल्लंघन
कोशिका में ऊर्जा का स्रोत भोजन है, जिसके टूटने के बाद ऊर्जा अंतिम पदार्थों में मुक्त हो जाती है। ऊर्जा उत्पादन का मुख्य स्थान माइटोकॉन्ड्रिया है, जिसमें श्वसन श्रृंखला के एंजाइमों की सहायता से पदार्थों का ऑक्सीकरण होता है। ऑक्सीकरण ऊर्जा का मुख्य आपूर्तिकर्ता है, क्योंकि ग्लाइकोलाइसिस के परिणामस्वरूप, ऑक्सीकरण की तुलना में ऑक्सीकरण सब्सट्रेट (ग्लूकोज) की समान मात्रा से 5% से अधिक ऊर्जा जारी नहीं होती है। ऑक्सीकरण के दौरान निकलने वाली ऊर्जा का लगभग 60% उच्च-ऊर्जा फॉस्फेट (एटीपी, क्रिएटिन फॉस्फेट) में ऑक्सीडेटिव फॉस्फोराइलेशन द्वारा जमा होता है, बाकी गर्मी के रूप में नष्ट हो जाता है। इसके बाद, पंप संचालन, संश्लेषण, विभाजन, गति, स्राव आदि जैसी प्रक्रियाओं के लिए कोशिका द्वारा उच्च-ऊर्जा फॉस्फेट का उपयोग किया जाता है। तीन तंत्र हैं, जिनके क्षतिग्रस्त होने से कोशिका की ऊर्जा आपूर्ति में व्यवधान हो सकता है: पहला है ऊर्जा चयापचय एंजाइमों के संश्लेषण का तंत्र, दूसरा ऑक्सीडेटिव फास्फारिलीकरण का तंत्र है, तीसरा ऊर्जा उपयोग का तंत्र है।
माइटोकॉन्ड्रियल श्वसन श्रृंखला में इलेक्ट्रॉन परिवहन में व्यवधान या प्रोटॉन क्षमता के नुकसान के साथ एडीपी ऑक्सीकरण और फॉस्फोराइलेशन का विघटन, एटीपी उत्पादन के लिए प्रेरक शक्ति, ऑक्सीडेटिव फॉस्फोराइलेशन को इस तरह से कमजोर कर देती है कि अधिकांश ऊर्जा गर्मी के रूप में नष्ट हो जाती है और उच्च-ऊर्जा यौगिकों की संख्या घट जाती है। एड्रेनालाईन के प्रभाव में ऑक्सीकरण और फॉस्फोराइलेशन के अनयुग्मन का उपयोग होमोथर्मिक जीवों की कोशिकाओं द्वारा गर्मी के उत्पादन को बढ़ाने के लिए किया जाता है, जबकि ठंडक के दौरान शरीर के तापमान को स्थिर बनाए रखा जाता है या बुखार के दौरान इसे बढ़ाया जाता है। थायरोटॉक्सिकोसिस में माइटोकॉन्ड्रियल संरचना और ऊर्जा चयापचय में महत्वपूर्ण परिवर्तन देखे जाते हैं। ये परिवर्तन शुरू में प्रतिवर्ती होते हैं, लेकिन एक निश्चित बिंदु के बाद वे अपरिवर्तनीय हो जाते हैं: माइटोकॉन्ड्रिया टुकड़े हो जाते हैं, विघटित हो जाते हैं या सूज जाते हैं, क्रिस्टा खो देते हैं, रिक्तिका में बदल जाते हैं, और अंततः हाइलिन, फेरिटिन, कैल्शियम, लिपोफसिन जैसे पदार्थ जमा हो जाते हैं। स्कर्वी के रोगियों में, माइटोकॉन्ड्रिया आपस में जुड़कर चोंड्रियोस्फेयर बनाते हैं, संभवतः पेरोक्साइड यौगिकों द्वारा झिल्ली क्षति के कारण। माइटोकॉन्ड्रिया को महत्वपूर्ण क्षति एक सामान्य कोशिका के घातक कोशिका में परिवर्तन के दौरान आयनीकृत विकिरण के प्रभाव में होती है।
माइटोकॉन्ड्रिया कैल्शियम आयनों का एक शक्तिशाली डिपो है, जहां इसकी सांद्रता साइटोप्लाज्म की तुलना में कई गुना अधिक होती है। जब माइटोकॉन्ड्रिया क्षतिग्रस्त हो जाते हैं, तो कैल्शियम साइटोप्लाज्म में प्रवेश कर जाता है, जिससे इंट्रासेल्युलर संरचनाओं को नुकसान होने और संबंधित कोशिका के कार्यों में व्यवधान के साथ प्रोटीनेस सक्रिय हो जाता है, उदाहरण के लिए, कैल्शियम संकुचन या यहां तक कि न्यूरॉन्स में "कैल्शियम मृत्यु"। माइटोकॉन्ड्रिया की कार्यात्मक क्षमता में व्यवधान के परिणामस्वरूप, मुक्त कण पेरोक्साइड यौगिकों का निर्माण तेजी से बढ़ जाता है, जिनमें बहुत अधिक प्रतिक्रिया होती है और इसलिए कोशिका के महत्वपूर्ण घटकों - न्यूक्लिक एसिड, प्रोटीन और लिपिड को नुकसान होता है। यह घटना तथाकथित ऑक्सीडेटिव तनाव के दौरान देखी जाती है और कोशिका के अस्तित्व पर इसके नकारात्मक परिणाम हो सकते हैं। इस प्रकार, माइटोकॉन्ड्रिया की बाहरी झिल्ली को नुकसान इंटरमेम्ब्रेन स्पेस में निहित पदार्थों के साइटोप्लाज्म में रिलीज के साथ होता है, मुख्य रूप से साइटोक्रोम सी और कुछ अन्य जैविक रूप से सक्रिय पदार्थ, जो श्रृंखला प्रतिक्रियाओं को ट्रिगर करते हैं जो क्रमादेशित कोशिका मृत्यु का कारण बनते हैं - एपोप्टोसिस। माइटोकॉन्ड्रियल डीएनए को नुकसान पहुंचाकर, मुक्त कट्टरपंथी प्रतिक्रियाएं कुछ श्वसन श्रृंखला एंजाइमों के निर्माण के लिए आवश्यक आनुवंशिक जानकारी को विकृत कर देती हैं, जो विशेष रूप से माइटोकॉन्ड्रिया में उत्पन्न होती हैं। इससे ऑक्सीडेटिव प्रक्रियाओं में और भी अधिक व्यवधान होता है। सामान्य तौर पर, माइटोकॉन्ड्रिया का अपना आनुवंशिक तंत्र, नाभिक के आनुवंशिक तंत्र की तुलना में, हानिकारक प्रभावों से कम सुरक्षित होता है जो इसमें एन्कोड की गई आनुवंशिक जानकारी को बदल सकता है। नतीजतन, माइटोकॉन्ड्रिया की शिथिलता जीवन भर होती है, उदाहरण के लिए, उम्र बढ़ने की प्रक्रिया के दौरान, कोशिका के घातक परिवर्तन के दौरान, साथ ही अंडे में माइटोकॉन्ड्रियल डीएनए के उत्परिवर्तन से जुड़े वंशानुगत माइटोकॉन्ड्रियल रोगों की पृष्ठभूमि के खिलाफ। वर्तमान में, 50 से अधिक माइटोकॉन्ड्रियल उत्परिवर्तन का वर्णन किया गया है जो तंत्रिका और मांसपेशी प्रणालियों के वंशानुगत अपक्षयी रोगों का कारण बनते हैं। वे विशेष रूप से मां से बच्चे में संचरित होते हैं, क्योंकि शुक्राणु के माइटोकॉन्ड्रिया युग्मनज का हिस्सा नहीं होते हैं और तदनुसार, नए जीव का हिस्सा नहीं होते हैं।
आनुवंशिक जानकारी के संरक्षण और प्रसारण का उल्लंघन
कोशिका केन्द्रक में अधिकांश आनुवंशिक जानकारी होती है और इस तरह यह इसकी सामान्य कार्यप्रणाली सुनिश्चित करती है। चयनात्मक जीन अभिव्यक्ति के माध्यम से, यह इंटरफ़ेज़ के दौरान कोशिका गतिविधि का समन्वय करता है, आनुवंशिक जानकारी संग्रहीत करता है, और कोशिका विभाजन के दौरान आनुवंशिक सामग्री को पुन: बनाता और प्रसारित करता है। डीएनए प्रतिकृति और आरएनए प्रतिलेखन नाभिक में होता है। विभिन्न रोगजनक कारक, जैसे पराबैंगनी और आयनकारी विकिरण, मुक्त कण ऑक्सीकरण, रसायन, वायरस, डीएनए को नुकसान पहुंचा सकते हैं। यह अनुमान लगाया गया है कि गर्म रक्त वाले जानवर की प्रत्येक कोशिका को 1 दिन लगता है। 10,000 से अधिक आधार खो देता है। यहां हमें विभाजन के दौरान नकल करते समय उल्लंघनों को जोड़ना चाहिए। यदि ये क्षतियाँ बनी रहीं, तो कोशिका जीवित नहीं रह पाएगी। सुरक्षा शक्तिशाली मरम्मत प्रणालियों के अस्तित्व में निहित है, जैसे कि पराबैंगनी एंडोन्यूक्लिज़, मरम्मत प्रतिकृति और पुनर्संयोजन मरम्मत प्रणाली, जो डीएनए क्षति की भरपाई करती हैं। मरम्मत प्रणालियों में आनुवंशिक दोष डीएनए को नुकसान पहुंचाने वाले कारकों के प्रति बढ़ती संवेदनशीलता के कारण होने वाली बीमारियों के विकास का कारण बनते हैं। यह ज़ेरोडर्मा पिगमेंटोसम है, साथ ही कुछ त्वरित उम्र बढ़ने वाले सिंड्रोम भी हैं, जिनमें घातक ट्यूमर विकसित होने की प्रवृत्ति बढ़ जाती है।
डीएनए प्रतिकृति, मैसेंजर आरएनए (एमआरएनए) के प्रतिलेखन और न्यूक्लिक एसिड से प्रोटीन की संरचना में आनुवंशिक जानकारी के अनुवाद की प्रक्रियाओं को विनियमित करने की प्रणाली काफी जटिल और बहु-स्तरीय है। 3000 से अधिक की कुल संख्या के साथ प्रतिलेखन कारकों की कार्रवाई को ट्रिगर करने वाले नियामक कैस्केड के अलावा, जो कुछ जीनों को सक्रिय करते हैं, छोटे आरएनए अणुओं (आरएनए; आरएनएआई में हस्तक्षेप) द्वारा मध्यस्थता वाली एक बहु-स्तरीय नियामक प्रणाली भी है। मानव जीनोम, जिसमें लगभग 3 बिलियन प्यूरीन और पाइरीमिडीन आधार होते हैं, में प्रोटीन संश्लेषण के लिए जिम्मेदार संरचनात्मक जीन का केवल 2% होता है। बाकी नियामक आरएनए का संश्लेषण प्रदान करते हैं, जो प्रतिलेखन कारकों के साथ-साथ गुणसूत्रों में डीएनए स्तर पर संरचनात्मक जीन के काम को सक्रिय या अवरुद्ध करते हैं या पॉलीपेप्टाइड अणु के निर्माण के दौरान मैसेंजर आरएनए (एमआरएनए) के अनुवाद की प्रक्रियाओं को प्रभावित करते हैं। साइटोप्लाज्म. आनुवंशिक जानकारी का उल्लंघन संरचनात्मक जीन और डीएनए के नियामक भाग दोनों के स्तर पर विभिन्न वंशानुगत रोगों के रूप में संबंधित अभिव्यक्तियों के साथ हो सकता है।
हाल ही में, किसी जीव के व्यक्तिगत विकास के दौरान होने वाले आनुवंशिक सामग्री में परिवर्तनों पर बहुत अधिक ध्यान आकर्षित किया गया है और उनके मिथाइलेशन, एसिटिलेशन और फॉस्फोराइलेशन के कारण डीएनए और गुणसूत्रों के कुछ वर्गों के अवरोध या सक्रियण से जुड़े हैं। ये परिवर्तन लंबे समय तक बने रहते हैं, कभी-कभी भ्रूणजनन से लेकर बुढ़ापे तक जीव के पूरे जीवन भर, और एपिजेनोमिक आनुवंशिकता कहलाते हैं।
परिवर्तित आनुवंशिक जानकारी वाली कोशिकाओं के प्रसार को उन प्रणालियों (कारकों) द्वारा भी रोका जाता है जो माइटोटिक चक्र को नियंत्रित करते हैं। वे साइक्लिन-आश्रित प्रोटीन किनेसेस और उनके उत्प्रेरक सबयूनिट्स - साइक्लिन - के साथ बातचीत करते हैं और कोशिका को पूर्ण माइटोटिक चक्र से गुजरने से रोकते हैं, डीएनए की मरम्मत पूरी होने तक प्रीसिंथेटिक और सिंथेटिक चरणों (जी 1/एस ब्लॉक) के बीच की सीमा पर विभाजन को रोकते हैं। और यदि यह असंभव है, तो वे क्रमादेशित मृत्यु कोशिकाएँ आरंभ करते हैं। इन कारकों में p53 जीन शामिल है, जिसके उत्परिवर्तन के कारण रूपांतरित कोशिकाओं के प्रसार पर नियंत्रण खत्म हो जाता है; यह लगभग 50% मानव कैंसर में देखा जाता है। माइटोटिक चक्र की दूसरी जांच चौकी G2/M सीमा पर है। यहां, माइटोसिस या अर्धसूत्रीविभाजन में बेटी कोशिकाओं के बीच क्रोमोसोमल सामग्री का सही वितरण तंत्र के एक सेट का उपयोग करके नियंत्रित किया जाता है जो सेल स्पिंडल, केंद्र और सेंट्रोमियर (कीनेटोकोर्स) को नियंत्रित करता है। इन तंत्रों की अप्रभावीता से गुणसूत्रों या उनके भागों के वितरण में व्यवधान होता है, जो बेटी कोशिकाओं (एन्यूप्लोइडी) में से किसी एक में किसी गुणसूत्र की अनुपस्थिति, एक अतिरिक्त गुणसूत्र (पॉलीप्लोइडी) की उपस्थिति, एक के अलग होने से प्रकट होता है। एक गुणसूत्र का भाग (विलोपन) और उसका दूसरे गुणसूत्र में स्थानांतरण (स्थानांतरण)। ऐसी प्रक्रियाएं अक्सर घातक रूप से विकृत और रूपांतरित कोशिकाओं के प्रसार के दौरान देखी जाती हैं। यदि यह रोगाणु कोशिकाओं के साथ अर्धसूत्रीविभाजन के दौरान होता है, तो इससे या तो भ्रूण के विकास के प्रारंभिक चरण में भ्रूण की मृत्यु हो जाती है, या क्रोमोसोमल बीमारी वाले जीव का जन्म होता है।
ट्यूमर के विकास के दौरान अनियंत्रित कोशिका प्रसार उन जीनों में उत्परिवर्तन के परिणामस्वरूप होता है जो कोशिका प्रसार को नियंत्रित करते हैं और इन्हें ऑन्कोजीन कहा जाता है। वर्तमान में ज्ञात 70 से अधिक ऑन्कोजीन में से अधिकांश कोशिका वृद्धि विनियमन के घटकों से संबंधित हैं, कुछ प्रतिलेखन कारकों द्वारा दर्शाए जाते हैं जो जीन गतिविधि को नियंत्रित करते हैं, साथ ही ऐसे कारक जो कोशिका विभाजन और वृद्धि को रोकते हैं। प्रसार करने वाली कोशिकाओं के अत्यधिक विस्तार (प्रसार) को सीमित करने वाला एक अन्य कारक गुणसूत्रों - टेलोमेरेस के सिरों का छोटा होना है, जो पूरी तरह से स्थैतिक संपर्क के परिणामस्वरूप पूरी तरह से दोहराने में सक्षम नहीं हैं, इसलिए, प्रत्येक कोशिका विभाजन के बाद, टेलोमेरेस को छोटा कर दिया जाता है। आधारों का एक निश्चित भाग। इस प्रकार, एक निश्चित संख्या में विभाजन (आमतौर पर जीव के प्रकार और उसकी उम्र के आधार पर 20 से 100 तक) के बाद एक वयस्क जीव की कोशिकाओं का प्रसार टेलोमेयर की लंबाई को समाप्त कर देता है और आगे गुणसूत्र प्रतिकृति रुक जाती है। यह घटना शुक्राणु उपकला, एंटरोसाइट्स और भ्रूण कोशिकाओं में एंजाइम टेलोमेरेज़ की उपस्थिति के कारण नहीं होती है, जो प्रत्येक विभाजन के बाद टेलोमेर की लंबाई को बहाल करता है। वयस्क जीवों की अधिकांश कोशिकाओं में, टेलोमेरेज़ अवरुद्ध होता है, लेकिन, दुर्भाग्य से, यह ट्यूमर कोशिकाओं में सक्रिय होता है।
नाभिक और साइटोप्लाज्म के बीच संबंध और दोनों दिशाओं में पदार्थों का परिवहन ऊर्जा की खपत करने वाली विशेष परिवहन प्रणालियों की भागीदारी के साथ परमाणु झिल्ली में छिद्रों के माध्यम से किया जाता है। इस प्रकार, ऊर्जा और प्लास्टिक पदार्थ, सिग्नलिंग अणु (प्रतिलेखन कारक) को नाभिक तक पहुंचाया जाता है। रिवर्स प्रवाह एमआरएनए के साइटोप्लाज्म अणुओं में ले जाता है और कोशिका में प्रोटीन संश्लेषण के लिए आवश्यक राइबोसोम, आरएनए (टीआरएनए) को स्थानांतरित करता है। पदार्थों के परिवहन का यही मार्ग विषाणुओं में भी अंतर्निहित है, विशेष रूप से एचआईवी जैसे विषाणुओं में। वे अपनी आनुवंशिक सामग्री को मेजबान कोशिका के केंद्रक में स्थानांतरित करते हैं, इसके बाद इसे मेजबान जीनोम में शामिल करते हैं और नए वायरल कणों के प्रोटीन के आगे संश्लेषण के लिए नवगठित वायरल आरएनए को साइटोप्लाज्म में स्थानांतरित करते हैं।
संश्लेषण प्रक्रियाओं का उल्लंघन
प्रोटीन संश्लेषण प्रक्रियाएं एंडोप्लाज्मिक रेटिकुलम के सिस्टर्न में होती हैं, जो परमाणु झिल्ली में छिद्रों से निकटता से जुड़ी होती हैं, जिसके माध्यम से राइबोसोम, टीआरएनए और एमआरएनए एंडोप्लाज्मिक रेटिकुलम में प्रवेश करते हैं। यहां, पॉलीपेप्टाइड श्रृंखलाओं का संश्लेषण किया जाता है, जो बाद में एग्रानुलर एंडोप्लाज्मिक रेटिकुलम और लैमेलर कॉम्प्लेक्स (गोल्गी कॉम्प्लेक्स) में अपना अंतिम रूप प्राप्त करते हैं, जहां वे पोस्ट-ट्रांसलेशनल संशोधन से गुजरते हैं और कार्बोहाइड्रेट और लिपिड अणुओं के साथ संयोजन करते हैं। नवगठित प्रोटीन अणु संश्लेषण स्थल पर नहीं रहते, बल्कि एक जटिल विनियमित प्रक्रिया के माध्यम से बने रहते हैं जिसे कहा जाता है प्रोटीनकिनेसिस, सक्रिय रूप से कोशिका के उस अलग हिस्से में स्थानांतरित हो जाते हैं जहां वे अपना इच्छित कार्य करेंगे। इस मामले में, एक बहुत ही महत्वपूर्ण कदम स्थानांतरित अणु को उसके अंतर्निहित कार्य को करने में सक्षम उचित स्थानिक विन्यास में संरचना करना है। यह संरचना विशेष एंजाइमों की मदद से या विशेष प्रोटीन अणुओं - चैपरोन के मैट्रिक्स पर होती है, जो बाहरी प्रभाव के कारण नवगठित या परिवर्तित प्रोटीन अणु को सही त्रि-आयामी संरचना प्राप्त करने में मदद करती है। कोशिका पर प्रतिकूल प्रभाव की स्थिति में, जब प्रोटीन अणुओं की संरचना में व्यवधान की संभावना होती है (उदाहरण के लिए, शरीर के तापमान में वृद्धि, एक संक्रामक प्रक्रिया, नशा के साथ), कोशिका में चैपरोन की सांद्रता बढ़ जाती है तेजी से. इसलिए, ऐसे अणुओं को भी कहा जाता है तनाव प्रोटीन, या हीट शॉक प्रोटीन. प्रोटीन अणु की संरचना के उल्लंघन से रासायनिक रूप से निष्क्रिय समूहों का निर्माण होता है, जो अमाइलॉइडोसिस, अल्जाइमर रोग आदि के दौरान कोशिका में या उसके बाहर जमा हो जाते हैं। कभी-कभी एक पूर्व-संरचित समान अणु एक मैट्रिक्स के रूप में काम कर सकता है, और इसमें मामले में, यदि प्राथमिक संरचना गलत तरीके से होती है, तो बाद के सभी अणु भी दोषपूर्ण होंगे। यह स्थिति तथाकथित प्रियन रोगों (भेड़ में स्क्रैपी, पागल गायों, कुरु, मनुष्यों में क्रुट्ज़फेल्ड-जैकब रोग) में होती है, जब तंत्रिका कोशिका के झिल्ली प्रोटीन में से एक में दोष कोशिका के अंदर अक्रिय द्रव्यमान के संचय का कारण बनता है। और इसके महत्वपूर्ण कार्यों में व्यवधान।
कोशिका में संश्लेषण प्रक्रियाओं का विघटन इसके विभिन्न चरणों में हो सकता है: नाभिक में आरएनए प्रतिलेखन, राइबोसोम में पॉलीपेप्टाइड्स का अनुवाद, पोस्ट-ट्रांसलेशनल संशोधन, बेज अणु का हाइपरमेथिलेशन और ग्लाइकोसिलेशन, कोशिका में प्रोटीन का परिवहन और वितरण और उनका निष्कासन। बाहर की ओर. इस मामले में, कोई राइबोसोम की संख्या में वृद्धि या कमी, पॉलीराइबोसोम का टूटना, दानेदार एंडोप्लाज्मिक रेटिकुलम के कुंडों का विस्तार, राइबोसोम की हानि और पुटिकाओं और रिक्तिका के गठन का निरीक्षण कर सकता है। इस प्रकार, जब पीले ग्रीब द्वारा जहर दिया जाता है, तो आरएनए पोलीमरेज़ एंजाइम क्षतिग्रस्त हो जाता है, जो प्रतिलेखन को बाधित करता है। डिप्थीरिया विष, बढ़ाव कारक को निष्क्रिय करके, अनुवाद प्रक्रियाओं को बाधित करता है, जिससे मायोकार्डियल क्षति होती है। कुछ विशिष्ट प्रोटीन अणुओं के संश्लेषण में व्यवधान का कारण संक्रामक एजेंट हो सकते हैं। उदाहरण के लिए, हर्पीस वायरस एमएचसी एंटीजन अणुओं के संश्लेषण और अभिव्यक्ति को रोकते हैं, जो उन्हें प्रतिरक्षा नियंत्रण प्लेग बेसिली - तीव्र सूजन के मध्यस्थों के संश्लेषण से आंशिक रूप से बचने की अनुमति देता है; असामान्य प्रोटीन की उपस्थिति उनके आगे टूटने को रोक सकती है और अक्रिय या यहां तक कि विषाक्त पदार्थों के संचय का कारण बन सकती है। यह, कुछ हद तक, क्षय प्रक्रियाओं के विघटन से सुगम हो सकता है।
क्षय प्रक्रियाओं का विघटन
कोशिका में प्रोटीन के संश्लेषण के साथ-साथ उसका टूटना भी लगातार होता रहता है। सामान्य परिस्थितियों में, इसका महत्वपूर्ण विनियामक और रचनात्मक महत्व है, उदाहरण के लिए, एंजाइमों, प्रोटीन हार्मोन और माइटोटिक चक्र प्रोटीन के निष्क्रिय रूपों के सक्रियण के दौरान। सामान्य कोशिका वृद्धि और विकास के लिए प्रोटीन और ऑर्गेनेल के संश्लेषण और क्षरण के बीच एक सूक्ष्म नियंत्रित संतुलन की आवश्यकता होती है। हालाँकि, प्रोटीन संश्लेषण की प्रक्रिया में, संश्लेषण उपकरण के संचालन में त्रुटियों, प्रोटीन अणु की असामान्य संरचना और रासायनिक और जीवाणु एजेंटों द्वारा इसकी क्षति के कारण, काफी बड़ी संख्या में दोषपूर्ण अणु लगातार बनते रहते हैं। कुछ अनुमानों के अनुसार, उनका हिस्सा सभी संश्लेषित प्रोटीन का लगभग एक तिहाई है।
स्तनधारी कोशिकाओं में कई मुख्य होते हैं प्रोटीन विनाश के तरीके:लाइसोसोमल प्रोटीज़ (पेंटाइड हाइड्रॉलेज़), कैल्शियम-निर्भर प्रोटीनेज़ (एंडोपेप्टिडेज़) और प्रोटीसोम प्रणाली के माध्यम से। इसके अलावा, कैसपेज़ जैसे विशिष्ट प्रोटीनेस भी होते हैं। मुख्य अंग जिसमें यूकेरियोटिक कोशिकाओं में पदार्थों का क्षरण होता है वह लाइसोसोम है, जिसमें कई हाइड्रोलाइटिक एंजाइम होते हैं। लाइसोसोम और फागोलिसोसोम में एंडोसाइटोसिस और विभिन्न प्रकार की ऑटोफैगी की प्रक्रियाओं के कारण, दोषपूर्ण प्रोटीन अणु और संपूर्ण अंग नष्ट हो जाते हैं: क्षतिग्रस्त माइटोकॉन्ड्रिया, प्लाज्मा झिल्ली के खंड, कुछ बाह्य प्रोटीन और स्रावी कणिकाओं की सामग्री।
प्रोटीन क्षरण के लिए एक महत्वपूर्ण तंत्र प्रोटीसोम है, जो साइटोसोल, न्यूक्लियस, एंडोप्लाज्मिक रेटिकुलम और कोशिका झिल्ली पर स्थानीयकृत जटिल संरचना की एक बहु-उत्प्रेरक प्रोटीनेज़ संरचना है। यह एंजाइम प्रणाली क्षतिग्रस्त प्रोटीन के साथ-साथ स्वस्थ प्रोटीन को तोड़ने के लिए जिम्मेदार है जिन्हें सामान्य कोशिका कार्य के लिए हटाया जाना चाहिए। इस मामले में, नष्ट किए जाने वाले प्रोटीन को प्रारंभिक रूप से एक विशिष्ट पॉलीपेप्टाइड, यूबिकिटिन के साथ जोड़ा जाता है। हालाँकि, गैर-सर्वव्यापी प्रोटीन को प्रोटीसोम में आंशिक रूप से नष्ट भी किया जा सकता है। प्रोटीसोम्स में प्रोटीन अणुओं का छोटे पॉलीपेप्टाइड्स (प्रसंस्करण) में टूटना और बाद में टाइप I एमएचसी अणुओं के साथ उनकी प्रस्तुति शरीर में एंटीजेनिक होमियोस्टेसिस के प्रतिरक्षा नियंत्रण में एक महत्वपूर्ण कड़ी है। जब प्रोटीसोम फ़ंक्शन कमजोर हो जाता है, क्षतिग्रस्त हो जाता है और अनावश्यक प्रोटीन जमा हो जाता है, जो कोशिका की उम्र बढ़ने के साथ होता है। साइक्लिन-निर्भर प्रोटीन के क्षरण के उल्लंघन से कोशिका विभाजन में व्यवधान होता है, स्रावी प्रोटीन का क्षरण होता है - सिस्टोफिब्रोसिस का विकास होता है। इसके विपरीत, प्रोटीसोम फ़ंक्शन में वृद्धि शरीर की कमी (एड्स, कैंसर) के साथ होती है।
प्रोटीन क्षरण के आनुवंशिक रूप से निर्धारित विकारों के साथ, जीव व्यवहार्य नहीं होता है और भ्रूणजनन के प्रारंभिक चरण में ही मर जाता है। यदि वसा या कार्बोहाइड्रेट का टूटना बाधित हो जाता है, तो भंडारण रोग (थिसॉरिज्मोसिस) उत्पन्न हो जाते हैं। इस मामले में, कुछ पदार्थों या उनके अधूरे टूटने के उत्पादों - लिपिड, पॉलीसेकेराइड - की अत्यधिक मात्रा कोशिका के अंदर जमा हो जाती है, जो कोशिका के कार्य को काफी नुकसान पहुंचाती है। अधिकतर यह यकृत उपकला कोशिकाओं (हेपेटोसाइट्स), न्यूरॉन्स, फ़ाइब्रोब्लास्ट और मैक्रोफैगोसाइट्स में देखा जाता है।
पदार्थों के टूटने की प्रक्रियाओं में अर्जित विकार रोग प्रक्रियाओं (उदाहरण के लिए, प्रोटीन, वसा, कार्बोहाइड्रेट और वर्णक अध: पतन) के परिणामस्वरूप उत्पन्न हो सकते हैं और असामान्य पदार्थों के निर्माण के साथ होते हैं। लाइसोसोमल प्रोटियोलिसिस प्रणाली में गड़बड़ी के कारण उपवास के दौरान अनुकूलन में कमी आती है या तनाव बढ़ता है, और कुछ अंतःस्रावी विकारों की घटना होती है - इंसुलिन, थायरोग्लोबुलिन, साइटोकिन्स और उनके रिसेप्टर्स के स्तर में कमी आती है। बिगड़ा हुआ प्रोटीन क्षरण घाव भरने की दर को धीमा कर देता है, एथेरोस्क्लेरोसिस के विकास का कारण बनता है, और प्रतिरक्षा प्रतिक्रिया को प्रभावित करता है। हाइपोक्सिया के साथ, इंट्रासेल्युलर पीएच में परिवर्तन, विकिरण की चोट, झिल्ली लिपिड के बढ़े हुए पेरोक्सीडेशन की विशेषता, साथ ही लाइसोसोमोट्रोपिक पदार्थों के प्रभाव में - बैक्टीरियल एंडोटॉक्सिन, विषाक्त कवक (स्पोरोफ्यूसरिन) के मेटाबोलाइट्स, सिलिकॉन ऑक्साइड क्रिस्टल - लाइसोसोम झिल्ली की स्थिरता परिवर्तन, सक्रिय लाइसोसोमल एंजाइम साइटोप्लाज्म में छोड़े जाते हैं, जो कोशिका संरचनाओं के विनाश और उसकी मृत्यु का कारण बनता है।
अध्याय 1
कोशिका शरीर क्रिया विज्ञान की मूल बातें
मैं. डुडेल
प्लाज्मा झिल्ली . जंतु कोशिकाएं प्लाज्मा झिल्ली से घिरी होती हैं (चित्र 1.1)। हम इसकी संरचना पर थोड़ा और विस्तार से ध्यान देंगे, जो कई अंतःकोशिकीय झिल्लियों की संरचना के समान है। झिल्ली के मुख्य मैट्रिक्स में शामिल हैं लिपिड,मुख्य रूप से फॉस्फेटिडिलकोलाइन। इन लिपिडों में एक हाइड्रोफिलिक हेड समूह होता है जिससे लंबी हाइड्रोफोबिक हाइड्रोकार्बन श्रृंखलाएं जुड़ी होती हैं। पानी में, ऐसे लिपिड अनायास 4-5 एनएम मोटी एक बाइलेयर फिल्म बनाते हैं, जिसमें हाइड्रोफिलिक समूह जलीय माध्यम का सामना करते हैं, और हाइड्रोफोबिक हाइड्रोकार्बन श्रृंखलाएं दो पंक्तियों में व्यवस्थित होती हैं, जिससे एक निर्जल लिपिड चरण बनता है। कोशिका झिल्ली इस प्रकार की लिपिड बाईलेयर होती है और इसमें ग्लाइकोलिपिड्स, कोलेस्ट्रॉल और फॉस्फोलिपिड्स होते हैं (चित्र 1.2)। ग्लाइकोलिपिड्स का हाइड्रोफिलिक भाग ऑलिगोसेकेराइड्स द्वारा बनता है। ग्लाइकोलिपिड्स हमेशा प्लाज्मा झिल्ली की बाहरी सतह पर स्थित होते हैं, अणु का ऑलिगोसेकेराइड भाग पर्यावरण में डूबे हुए बाल की तरह उन्मुख होता है। फॉस्फोलिपिड्स के बीच लगभग समान मात्रा में बिखरे हुए, कोलेस्ट्रॉल अणु झिल्ली को स्थिर करते हैं। झिल्ली की आंतरिक और बाहरी परतों में विभिन्न लिपिड का वितरण समान नहीं होता है, और यहां तक कि एक परत के भीतर भी ऐसे क्षेत्र होते हैं जिनमें कुछ प्रकार के लिपिड केंद्रित होते हैं। यह असमान वितरण
चावल। 1.1. सबसे महत्वपूर्ण अंगकों को दर्शाने वाली कोशिका का योजनाबद्ध चित्रण
संभवतः इसका कुछ, अभी तक अस्पष्ट, कार्यात्मक महत्व है।
झिल्ली के अपेक्षाकृत निष्क्रिय लिपिड मैट्रिक्स में अंतर्निहित मुख्य कार्यात्मक तत्व हैं गिलहरी(चित्र 1.2)। वजन के हिसाब से प्रोटीन विभिन्न झिल्लियों में 25 से 75% तक होता है, लेकिन चूंकि प्रोटीन अणु लिपिड अणुओं की तुलना में बहुत बड़े होते हैं, वजन के हिसाब से 50% 1 प्रोटीन अणु और 50 लिपिड अणुओं के अनुपात के बराबर होता है। कुछ प्रोटीन झिल्ली की बाहरी से भीतरी सतह तक प्रवेश करते हैं, जबकि अन्य एक परत में स्थिर रहते हैं। प्रोटीन अणु आमतौर पर उन्मुख होते हैं ताकि उनके हाइड्रोफोबिक समूह लिपिड झिल्ली में एम्बेडेड हों और झिल्ली की सतह पर ध्रुवीय हाइड्रोफिलिक समूह जलीय चरण में एम्बेडेड हों। झिल्ली की बाहरी सतह पर मौजूद कई प्रोटीन ग्लाइकोप्रोटीन होते हैं; उनके हाइड्रोफिलिक सैकेराइड समूह बाह्य कोशिकीय वातावरण का सामना करते हैं।
इंट्रासेल्युलर ऑर्गेनेल की झिल्ली प्रणाली .
कोशिका आयतन का लगभग आधा भाग झिल्लियों द्वारा साइटोसोल से पृथक अंगकों द्वारा व्याप्त होता है। इंट्रासेल्युलर ऑर्गेनेल की झिल्लियों का कुल सतह क्षेत्र प्लाज्मा झिल्ली की सतह से कम से कम 10 गुना अधिक है। सबसे व्यापक रूप से प्रयुक्त झिल्ली प्रणाली है अन्तः प्रदव्ययी जलिका,एक नेटवर्क का प्रतिनिधित्व करना

चावल। 1.2.प्लाज्मा झिल्ली का योजनाबद्ध प्रतिनिधित्व. प्रोटीन फॉस्फोलिपिड बाइलेयर में अंतर्निहित होते हैं, जिनमें से कुछ बाइलेयर में प्रवेश करते हैं, जबकि अन्य केवल बाहरी या आंतरिक परत से जुड़े होते हैं।
अत्यधिक जटिल नलिकाएं या थैली जैसी लम्बी संरचनाएं; एंडोप्लाज्मिक रेटिकुलम के बड़े क्षेत्र राइबोसोम से भरे हुए हैं; ऐसे जालिका को दानेदार या खुरदरा कहा जाता है (चित्र 1.1)। गॉल्जीकायइसमें झिल्ली से बंधी लैमेला भी होती है, जिसमें से पुटिकाएं या पुटिकाएं टूट जाती हैं (चित्र 1.1)। लाइसोसोम और पेरोक्सीसोम- ये छोटे विशेष पुटिकाएं हैं। इन सभी विभिन्न अंगों, झिल्ली और इसके द्वारा घेरे गए स्थान में एंजाइमों के विशिष्ट सेट होते हैं; ऑर्गेनेल के अंदर, विशेष चयापचय उत्पाद जमा होते हैं, जिनका उपयोग ऑर्गेनेल के विभिन्न कार्यों को पूरा करने के लिए किया जाता है।
मुख्यऔर माइटोकॉन्ड्रियाइनमें अंतर यह है कि इनमें से प्रत्येक अंगक दो झिल्लियों से घिरा होता है। नाभिक चयापचय के गतिज नियंत्रण के लिए जिम्मेदार है; माइटोकॉन्ड्रिया की मुड़ी हुई आंतरिक झिल्ली ऑक्सीडेटिव चयापचय का स्थल है; यहां, पाइरूवेट या फैटी एसिड के ऑक्सीकरण के कारण, उच्च-ऊर्जा यौगिक एडेनोसिन ट्राइफॉस्फेट (एटीपी, या एटीपी) को संश्लेषित किया जाता है।
cytoskeleton . अंगकों के आसपास के साइटोप्लाज्म को किसी भी तरह से अनाकार नहीं माना जा सकता है; यह साइटोस्केलेटल नेटवर्क द्वारा व्याप्त है। साइटोस्केलेटन में सूक्ष्मनलिकाएं, एक्टिन फिलामेंट्स और मध्यवर्ती फिलामेंट्स होते हैं (चित्र 1.1)। सूक्ष्मनलिकाएंइनका बाहरी व्यास लगभग 25 एनएम है; वे ट्यूबुलिन प्रोटीन अणुओं के संयोजन के परिणामस्वरूप, एक नियमित बहुलक की तरह बनते हैं। एक्टिन फिलामेंट-सिकुड़ा हुआ तंतु, निकट-झिल्ली परत में और पूरे कोशिका में स्थित होते हैं, मुख्य रूप से गति से जुड़ी प्रक्रियाओं में भाग लेते हैं। माध्यमिक रेशेविभिन्न प्रकार की कोशिकाओं में विभिन्न रासायनिक संरचना के ब्लॉक से मिलकर बनता है; वे ऊपर उल्लिखित दो अन्य साइटोस्केलेटल तत्वों के बीच विभिन्न प्रकार के संबंध बनाते हैं। ऑर्गेनेल और प्लाज़्मा झिल्ली भी साइटोस्केलेटन से जुड़े होते हैं, जो न केवल कोशिका के आकार और उसमें ऑर्गेनेल की स्थिति को बनाए रखता है, बल्कि कोशिका के आकार और उसकी गतिशीलता में परिवर्तन भी निर्धारित करता है।
साइटोसोल . कोशिका आयतन का लगभग आधा भाग साइटोसोल द्वारा व्याप्त होता है। चूंकि इसमें वजन के हिसाब से लगभग 20% प्रोटीन होता है, इसलिए यह जलीय घोल की तुलना में अधिक जेल जैसा होता है। कार्बनिक और अकार्बनिक सहित छोटे अणु आयन,जलीय चरण में घुल गया. कोशिका और पर्यावरण (बाह्यकोशिकीय स्थान) के बीच आयनों का आदान-प्रदान होता है; इन चयापचय प्रक्रियाओं पर अगले भाग में चर्चा की जाएगी। बाह्यकोशिकीय स्थान में आयनों की सांद्रता को एक स्थिर स्तर पर काफी सटीकता के साथ बनाए रखा जाता है; प्रत्येक आयन की अंतःकोशिकीय सांद्रता का भी एक विशिष्ट स्तर होता है, जो कोशिका के बाहर से भिन्न होता है (तालिका 1.1)। बाह्य कोशिकीय वातावरण में धनायन सर्वाधिक प्रचुर मात्रा में होता हैना+ कोशिका में इसकी सांद्रता 10 गुना से भी कम है। इसके विपरीत, K+ की सांद्रता कोशिका के अंदर सबसे अधिक होती है; कोशिका के बाहर यह परिमाण के एक क्रम से कम होती है। बाह्यकोशिकीय और अंतःकोशिकीय सांद्रता के बीच सबसे बड़ी प्रवणता Ca 2+ के लिए मौजूद है, कोशिका के अंदर मुक्त आयनों की सांद्रता इसके बाहर की तुलना में कम से कम 10,000 गुना कम है। सभी आयन साइटोसोल में नहीं घुलते हैं; उनमें से कुछ प्रोटीन पर अवशोषित होते हैं या ऑर्गेनेल में जमा होते हैं। उदाहरण के लिए, Ca 2+ के मामले में, बाध्य आयन मुक्त आयनों की तुलना में बहुत अधिक संख्या में होते हैं। अधिकांश साइटोसोलिक प्रोटीन एंजाइम होते हैं, जिनकी भागीदारी से मध्यवर्ती चयापचय की कई प्रक्रियाएं होती हैं: ग्लाइकोलाइसिस और ग्लूकोनियोजेनेसिस, अमीनो एसिड का संश्लेषण या विनाश, राइबोसोम पर प्रोटीन संश्लेषण (चित्र 1.1)। साइटोसोल में लिपिड बूंदें और ग्लाइकोजन कणिकाएं भी होती हैं, जो महत्वपूर्ण अणुओं के भंडार के रूप में काम करती हैं।
तालिका 1.1.होमोथर्मिक जानवरों की मांसपेशियों की कोशिकाओं में इंट्रा- और बाह्य कोशिकीय आयन सांद्रता। ए – - "उच्च आणविक भार सेलुलर आयन"
|
अंतःकोशिकीय सांद्रता |
बाह्यकोशिकीय सांद्रता |
||
|
ना+ |
12 एमएमओएल/एल |
ना+ |
145 एमएमओएल/ली |
|
155 एमएमओएल/ली |
के+ |
4 एमएमओएल/एल |
|
|
सीए 2+ |
10 -7 10 -8 mmol/ली |
सीए 2+ |
2 एमएमओएल/एल |
|
सी एल - |
4 एमएमओएल/एल |
सी1- |
120 एमएमओएल/एल |
|
एचसीओ 3 - |
8 एमएमओएल/एल |
एचसीओ 3 - |
27 एमएमओएल/एल |
|
ए - |
155 एमएमओएल/ली |
अन्य फैटायनों |
5 एमएमओएल/एल |
|
विश्राम क्षमता-90 एमवी |
|||
1.2. कोशिका और पर्यावरण के बीच पदार्थों का आदान-प्रदान
हमने कोशिका की संरचना का संक्षेप में वर्णन किया है ताकि हम इस विवरण का उपयोग सेलुलर शरीर क्रिया विज्ञान की बुनियादी बातों की समीक्षा करने के लिए कर सकें। किसी भी स्थिति में किसी कोशिका को स्थिर इकाई नहीं माना जा सकता, क्योंकि विभिन्न अंतःकोशिकीय डिब्बों के साथ-साथ डिब्बों और पर्यावरण के बीच पदार्थों का निरंतर आदान-प्रदान होता रहता है। कोशिका संरचनाएं गतिशील संतुलन में हैं, और एक दूसरे के साथ और बाहरी वातावरण के साथ कोशिकाओं की बातचीत एक कार्यशील जीव के जीवन को बनाए रखने के लिए एक आवश्यक शर्त है। इस अध्याय में हम ऐसे आदान-प्रदान के मूलभूत तंत्रों को देखेंगे। अगले अध्यायों में इन तंत्रों पर विचार किया जाएगा क्योंकि वे तंत्रिका कोशिका और उसके कार्यों पर लागू होते हैं;
हालाँकि, वही तंत्र अन्य सभी अंगों के कामकाज का आधार हैं।
प्रसार.गतिमान पदार्थ की सबसे सरल प्रक्रिया प्रसार है। समाधानों (या गैसों) में, परमाणु और अणु स्वतंत्र रूप से चलते हैं, और सांद्रता में अंतर प्रसार के कारण संतुलित होता है। आइए तरल या गैस से भरे दो आयतनों पर विचार करें (चित्र 1.3), जिनमें पदार्थों की सांद्रता होती हैसी 1 और सी 2 और सतह क्षेत्र ए और मोटाई के साथ एक परत द्वारा अलग किया गयाडी। समय t के दौरान पदार्थ का प्रवाह m बताया गया है फ़िक का प्रसार का पहला नियम:
डी.एम/ डीटी= डीए/ डी ( सी 1 –सी 2)=डीए/ डीडी सी(1)
जहां डी किसी दिए गए पदार्थ, विलायक और तापमान के लिए प्रसार गुणांक, स्थिरांक है। एकाग्रता अंतर के लिए अधिक सामान्य रूप में dx दूरी पर dc
डीएम/डीटी= –डी ए डीसी/डीएक्स,(2)
खंड ए के माध्यम से प्रवाह एकाग्रता ढाल के समानुपाती होता हैडीसी/डीएक्स . समीकरण में ऋण चिह्न दिखाई देता है क्योंकि x दिशा में सांद्रता में परिवर्तन ऋणात्मक है।
प्रसार सबसे महत्वपूर्ण प्रक्रिया है जिसके कारण जलीय घोल में अधिकांश अणु कम दूरी तक चलते हैं। यह बात कोशिका में उनकी गति पर भी लागू होती है, क्योंकि झिल्लियों द्वारा प्रसार बाधित नहीं होता है। कई पदार्थ लिपिड झिल्लियों में स्वतंत्र रूप से फैल सकते हैं, विशेष रूप से पानी और ओ 2 और सीओ 2 जैसी घुली हुई गैसें। वसा में घुलनशील

चावल। 1.3.मात्रात्मक प्रसार योजना. दोनों स्थानों को मोटाई की एक परत द्वारा अलग किया जाता हैडीऔर क्षेत्र एक।सी; - आयतन के बाएँ भाग में कणों की उच्च सांद्रता, सी: - दाएँ भाग में कणों की कम सांद्रता भाग, गुलाबी सतह- प्रसार परत में सांद्रता प्रवणता। प्रसार प्रवाह डीएम/डीटी - देखें समीकरण (1)
पदार्थ झिल्लियों के माध्यम से भी अच्छी तरह फैलते हैं; यह इथेनॉल और यूरिया जैसे काफी छोटे आकार के ध्रुवीय अणुओं पर भी लागू होता है, जबकि शर्करा कठिनाई के साथ लिपिड परत से गुजरती है। साथ ही, लिपिड परतें आवेशित अणुओं के लिए व्यावहारिक रूप से अभेद्य होती हैं, जिनमें अकार्बनिक आयन भी शामिल हैं। गैर-इलेक्ट्रोलाइट्स के लिए, प्रसार समीकरण (1) आमतौर पर झिल्ली और फैलाने वाले पदार्थ की विशेषताओं को एक में जोड़कर रूपांतरित किया जाता है पैरामीटर-पारगम्यता (पी):
डीएम/डीटी=पी एडी सी।(3)
चित्र में. 1.4 की तुलना की गई है विभिन्न अणुओं के लिए लिपिड झिल्ली की पारगम्यता (पी)।
झिल्ली छिद्रों के माध्यम से प्रसार . प्लाज्मा झिल्ली (और अन्य कोशिका झिल्ली) न केवल लिपिड परत के माध्यम से फैलने वाले पदार्थों के लिए पारगम्य है, बल्कि कई आयनों, शर्करा, अमीनो एसिड और न्यूक्लियोटाइड के लिए भी पारगम्य है। ये पदार्थ बने छिद्रों के माध्यम से झिल्ली को पार करते हैं परिवहन प्रोटीन,झिल्ली में डूबा हुआ। ऐसे प्रोटीन के अंदर 1 एनएम से कम व्यास वाला एक पानी से भरा चैनल होता है, जिसके माध्यम से छोटे अणु फैल सकते हैं। वे एक सांद्रण प्रवणता के साथ चलते हैं, और यदि वे चार्ज ले जाते हैं, तो चैनलों के माध्यम से उनका आंदोलन भी झिल्ली क्षमता द्वारा नियंत्रित होता है। झिल्ली चैनल अपेक्षाकृत चयनात्मक होते हैं

चावल। 1.4.विभिन्न पदार्थों के लिए कृत्रिम लिपिड बाईलेयर्स की पारगम्यता
अणुओं के प्रकार के संबंध में जो उनके माध्यम से गुजर सकते हैं, उदाहरण के लिए, पोटेशियम, सोडियम और कैल्शियम चैनल हैं, जिनमें से प्रत्येक एक विशिष्ट को छोड़कर लगभग किसी भी आयन के लिए अभेद्य है। ऐसा चयनात्मकताचैनल की दीवारों में बाइंडिंग साइटों के चार्ज या संरचना द्वारा निर्धारित किया जाता है, जो एक विशिष्ट अणु के परिवहन की सुविधा प्रदान करता है और चैनल के माध्यम से अन्य पदार्थों के प्रवेश को रोकता है (चित्र)। 1.5, ए) .
व्यवहार के पीछे झिल्ली आयन चैनलनिरीक्षण करना आसान है, क्योंकि आयनों की गति से उत्पन्न होने वाली धारा को एक चैनल के लिए भी मापा जा सकता है। यह दिखाया गया है कि चैनल अनायास और उच्च आवृत्ति के साथ अपनी स्थिति को खुले से बंद में बदलते हैं। पोटेशियम चैनल को लगभग 2 पीए (2 · 10 -12 ए) के आयाम और कई मिलीसेकंड की अवधि के साथ वर्तमान दालों की विशेषता है (चित्र 2.12, पृष्ठ 37 देखें) [3]। इस अवधि के दौरान, दसियों हज़ार आयन इससे होकर गुजरते हैं। एक्स-रे विवर्तन, मोसबाउर स्पेक्ट्रोस्कोपी और परमाणु चुंबकीय अनुनाद (एनएमआर) का उपयोग करके प्रोटीन के एक संरचना से दूसरे में संक्रमण का अध्ययन किया जाता है। इस प्रकार प्रोटीन बहुत गतिशील, मोबाइल संरचनाएं हैं, और प्रोटीन से गुजरने वाला चैनल सिर्फ एक कठोर, पानी से भरी ट्यूब नहीं है (चित्र 1.5, ए), बल्कि तेजी से बढ़ने वाले आणविक समूहों और आवेशों की एक भूलभुलैया है। चैनल की यह गतिशील विशेषता परिलक्षित होती है चैनल की ऊर्जा प्रोफ़ाइल,चित्र में दिखाया गया है 1.5, बी। यहां, एब्सिस्सा अक्ष आयन सांद्रता सी 0 और संभावित 0 के साथ बाहरी समाधान से एकाग्रता सी 1 और संभावित ई के साथ आंतरिक समाधान तक चैनल की लंबाई का प्रतिनिधित्व करता है। कोटि अक्ष

चावल। 1.5.ए. प्लाज्मा झिल्ली के लिपिड बाईलेयर में एम्बेडेड पोटेशियम चैनल बनाने वाले प्रोटीन का योजनाबद्ध। चैनल की "दीवार" पर चार नकारात्मक चार्ज लगाए गए हैं। बी. चित्र में दिखाए गए चैनल की योजनाबद्ध ऊर्जा प्रोफ़ाइल। ए. कोटि अक्ष चैनल के पारित होने के लिए आवश्यक गतिज ऊर्जा के मूल्यों को दर्शाता है; एब्सिस्सा अक्ष के साथ - झिल्ली की आंतरिक और बाहरी सतहों के बीच की दूरी। ऊर्जा मिनिमा चैनल दीवार में निश्चित नकारात्मक चार्ज के साथ सकारात्मक रूप से चार्ज किए गए आयनों के बंधन स्थलों से मेल खाती है। ऊर्जा मैक्सिमा चैनल में प्रसार बाधाओं के अनुरूप है। यह माना जाता है कि चैनल प्रोटीन की संरचना स्वतःस्फूर्त रूप से दोलन करती है; ऊर्जा प्रोफ़ाइल विकल्प ठोस और धराशायी रेखाओं के साथ दिखाए गए हैं; ये दोलन ऊर्जा अवरोध (संशोधित) पर काबू पाने पर आयनों के बंधन को बहुत सुविधाजनक बनाते हैं
चैनल बाइंडिंग स्थलों पर आयन का ऊर्जा स्तर प्रस्तुत किया गया है; ग्राफ़ में शिखर पारगम्यता बाधा का प्रतिनिधित्व करता है जिसे आयन ऊर्जा को चैनल में प्रवेश करने के लिए पार करना होगा, और ग्राफ़ में "डिप्स" अपेक्षाकृत स्थिर स्थिति (बाइंडिंग) का प्रतिनिधित्व करता है। ऊर्जा शिखर की बाधा के बावजूद, यदि ऊर्जा प्रोफ़ाइल अनायास बदल जाती है, तो आयन चैनल में प्रवेश कर सकता है; इस प्रकार आयन अचानक खुद को ऊर्जा शिखर के "दूसरी तरफ" पा सकता है और कोशिका में जाना जारी रख सकता है। आयन के चार्ज, आकार और जलयोजन की डिग्री और चैनल दीवार संरचनाओं से जुड़ने की क्षमता के आधार पर, चैनल की ऊर्जा प्रोफ़ाइल विभिन्न आयनों के लिए भिन्न होती है, जो व्यक्तिगत चैनल प्रकारों की चयनात्मकता को समझा सकती है।
आयनों का प्रसार संतुलन . झिल्ली चैनलों के माध्यम से विभिन्न आयनों के प्रसार से बाह्य और अंतःकोशिकीय वातावरण के बीच सांद्रता में अंतर को समाप्त किया जाना चाहिए। हालाँकि, जैसा कि तालिका से देखा जा सकता है। 1.1, ऐसे मतभेद बने हुए हैं, इसलिए, कुछ तो होंगे ही संतुलनझिल्ली में प्रसार और अन्य परिवहन प्रक्रियाओं के बीच। अगले दो खंड उन तरीकों से निपटते हैं जिनसे यह संतुलन हासिल किया जाता है। आयनों के मामले में, प्रसार संतुलन उनके आवेश से प्रभावित होता है। अनावेशित अणुओं का प्रसार सांद्रता में अंतर से सुनिश्चित होता हैडीसी , और जब सांद्रता बराबर हो जाती है, तो परिवहन स्वयं बंद हो जाता है। आवेशित कण विद्युत क्षेत्र से अतिरिक्त रूप से प्रभावित होते हैं। उदाहरण के लिए, जब एक पोटेशियम आयन अपनी सांद्रता प्रवणता के साथ एक कोशिका को छोड़ता है, तो यह एक सकारात्मक चार्ज वहन करता है। इस प्रकार, इंट्रासेल्युलर वातावरण अधिक नकारात्मक रूप से चार्ज हो जाता है, जिसके परिणामस्वरूप झिल्ली में संभावित अंतर होता है। इंट्रासेल्युलर नकारात्मक चार्ज नए पोटेशियम आयनों को कोशिका छोड़ने से रोकता है, और जो आयन कोशिका छोड़ते हैं वे झिल्ली पर चार्ज को और बढ़ा देंगे। पोटेशियम आयनों का प्रवाह तब रुक जाता है जब विद्युत क्षेत्र की क्रिया सांद्रता में अंतर के कारण प्रसार दबाव की भरपाई कर देती है। आयन झिल्ली से गुजरते रहते हैं, लेकिन दोनों दिशाओं में समान मात्रा में। इसलिए, झिल्ली पर आयन सांद्रता में दिए गए अंतर के लिए, वहाँ है संतुलन क्षमताइआयन जिस पर झिल्ली से आयनों का प्रवाह रुक जाता है। संतुलन क्षमता को आसानी से उपयोग करके निर्धारित किया जा सकता है नर्नस्ट समीकरण:
इआयन= आरटी/ जेडएफ* एल.एनसी बाहर/ सी में(4)
जहां आर - गैस स्थिरांक, टी - पूर्ण तापमान, जेड - आयन वैलेंस (आयनों के लिए नकारात्मक)सी बाहर – बाह्यकोशिकीय आयन सांद्रता,सी में – इंट्रासेल्युलर आयन सांद्रता,एफ फैराडे संख्या. यदि हम समीकरण में स्थिरांकों को प्रतिस्थापित करते हैं, तो शरीर के तापमान (T = 310 K) पर पोटेशियम आयन E के लिए संतुलन क्षमता होती है K इसके बराबर है:
इक= –61 एमबी लॉग /(5)
यदि [K + बाहर ]/[ K + अंदर ] = 39, तालिका से निम्नानुसार है। 1.1 फिर
एक = -61 एम बी लॉग 39 = -97 एमवी।
वास्तव में, यह पाया गया कि सभी कोशिकाओं में है झिल्ली क्षमता;स्तनधारी मांसपेशी कोशिकाओं में इसका स्तर लगभग -90 mV होता है। स्थितियों और सापेक्ष आयन सांद्रता के आधार पर, कोशिकाओं की झिल्ली क्षमता -40 से -120 mV तक हो सकती है। उपरोक्त उदाहरण में सेल के लिए (तालिका 1.1) विराम विभव,लगभग -90 एमवी के बराबर, इंगित करता है कि झिल्ली चैनलों के माध्यम से पोटेशियम आयनों का प्रवाह लगभग संतुलन में है। यह आश्चर्य की बात नहीं है, क्योंकि विश्राम झिल्ली में पोटेशियम चैनलों की खुली अवस्था सबसे अधिक संभावना है, यानी। झिल्ली पोटेशियम आयनों के लिए सबसे अधिक पारगम्य है। हालाँकि, झिल्ली क्षमता अन्य आयनों के प्रवाह से निर्धारित होती है।
जिस आसानी से अनावेशित कण झिल्ली के माध्यम से फैल सकते हैं उसे समीकरण (3) में निर्धारित किया गया है। आवेशित कणों के प्रति पारगम्यता इसे थोड़े अधिक जटिल समीकरण द्वारा वर्णित किया गया है:
पी= एम आरटी/ डीएफ(6)
कहाँ एम- झिल्ली में आयन गतिशीलता,डी - झिल्ली की मोटाई,एक आर, टी और एफ -ज्ञात थर्मोडायनामिक स्थिरांक। इस तरह से निर्धारित विभिन्न आयनों के लिए पारगम्यता मूल्यों का उपयोग झिल्ली क्षमता की गणना के लिए किया जा सकता हैएम , जब पोटेशियम, सोडियम और क्लोरीन आयन एक साथ झिल्ली से गुजरते हैं (पारगम्यता पी के साथ)।के, पी ना और पी सीएल क्रमश)। यह माना जाता है कि क्षमता झिल्ली में समान रूप से गिरती है, ताकि क्षेत्र की ताकत स्थिर रहे। इस मामले में यह लागू होता है गोल्डमैन समीकरण, या स्थिर क्षेत्र समीकरण :
इएम= आर टी/ एफ * एलएन(पी के + पी ना + पी सीएल )/ (पी के + पी ना + पी सीएल )(7)
अधिकांश कोशिका झिल्लियों के लिए पीक पी से लगभग 30 गुना अधिकना (अनुभाग 1.3 भी देखें)। सापेक्ष मूल्यपी सीएल बहुत भिन्न होता है; कई झिल्लियों के लिएपी सीएल आर की तुलना में छोटाक , हालाँकि दूसरों के लिए (उदाहरण के लिए, कंकाल की मांसपेशियों में)पी सीएल , पी से काफी अधिकक।
सक्रिय परिवहन, सोडियम पंप . पिछला अनुभाग आयनों के निष्क्रिय प्रसार और दिए गए इंट्रा- और बाह्य कोशिकीय आयन सांद्रता पर परिणामी झिल्ली क्षमता का वर्णन करता है। हालाँकि, इस प्रक्रिया के परिणामस्वरूप, झिल्ली के बाद से कोशिका के अंदर आयनों की सांद्रता स्वचालित रूप से स्थिर नहीं होती है
क्षमता की तुलना में थोड़ा अधिक विद्युत ऋणात्मक हैईके, और ई की तुलना में बहुत अधिकना (लगभग +60 एमवी)। प्रसार के कारण, इंट्रासेल्युलर आयन सांद्रता, कम से कम पोटेशियम और सोडियम, को बाह्य कोशिकीय के साथ बराबर किया जाना चाहिए। आयन प्रवणता की स्थिरता सक्रिय परिवहन के माध्यम से प्राप्त की जाती है: झिल्ली प्रोटीन विद्युत और/या एकाग्रता प्रवणताओं के विरुद्ध झिल्ली में आयनों का परिवहन करते हैं, इसके लिए चयापचय ऊर्जा का उपभोग करते हैं। सक्रिय परिवहन की सबसे महत्वपूर्ण प्रक्रिया कार्य हैना/क - एक पंप जो लगभग सभी कोशिकाओं में मौजूद होता है;
पंप सोडियम आयनों को कोशिका से बाहर पंप करता है और साथ ही पोटेशियम आयनों को कोशिका में पंप करता है। यह सोडियम आयनों की कम इंट्रासेल्युलर सांद्रता और पोटेशियम आयनों की उच्च सांद्रता सुनिश्चित करता है (तालिका 1.1)। झिल्ली पर सोडियम आयनों की सांद्रता प्रवणता में विद्युत आवेगों के रूप में सूचना के प्रसारण (धारा 2.2 देखें) के साथ-साथ अन्य सक्रिय परिवहन तंत्रों के रखरखाव और सेल वॉल्यूम के विनियमन (नीचे देखें) से संबंधित विशिष्ट कार्य होते हैं। इसलिए, यह आश्चर्य की बात नहीं है कि एक सेल द्वारा खपत की गई ऊर्जा का 1/3 से अधिक Na/K पंप पर खर्च किया जाता है, और कुछ सबसे सक्रिय कोशिकाओं में 70% तक ऊर्जा इसके संचालन पर खर्च की जाती है।
Na/K ट्रांसपोर्ट प्रोटीन एक ATPase है। झिल्ली की आंतरिक सतह पर, यह एटीपी को एडीपी और फॉस्फेट में तोड़ देता है (चित्र 1.6)। एक एटीपी अणु की ऊर्जा का उपयोग कोशिका से तीन सोडियम आयनों और एक साथ दो पोटेशियम आयनों को कोशिका में ले जाने के लिए किया जाता है, यानी कुल मिलाकर, एक चक्र में कोशिका से एक सकारात्मक चार्ज हटा दिया जाता है। इस प्रकार, Na/K पंप है इलेक्ट्रोजेनिक(झिल्ली में एक विद्युत प्रवाह बनाता है), जिससे झिल्ली क्षमता की इलेक्ट्रोनगेटिविटी में लगभग 10 mV की वृद्धि होती है। ट्रांसपोर्ट प्रोटीन इस ऑपरेशन को उच्च गति से करता है: प्रति सेकंड 150 से 600 सोडियम आयन तक। परिवहन प्रोटीन का अमीनो एसिड अनुक्रम ज्ञात है, लेकिन इस जटिल चयापचय परिवहन का तंत्र अभी तक स्पष्ट नहीं है। इस प्रक्रिया को प्रोटीन द्वारा सोडियम या पोटेशियम आयनों के स्थानांतरण की ऊर्जा प्रोफाइल का उपयोग करके वर्णित किया गया है (चित्र 1.5,5)। परिवहन प्रोटीन (एक प्रक्रिया जिसमें ऊर्जा की आवश्यकता होती है) की संरचना में निरंतर परिवर्तन से जुड़े इन प्रोफाइलों में परिवर्तनों की प्रकृति से, कोई विनिमय की स्टोइकोमेट्री का न्याय कर सकता है: तीन सोडियम आयनों के लिए दो पोटेशियम आयनों का आदान-प्रदान किया जाता है।
Na/K पंप, पृथक के समानना+ /K + -निर्भर झिल्ली ATPase, विशेष रूप से कार्डियक ग्लाइकोसाइड ouabain (स्ट्रॉफ़ैन्थिन) द्वारा बाधित। चूँकि Na/K पंप का संचालन एक बहु-चरण रासायनिक प्रतिक्रिया है, यह, सभी रासायनिक प्रतिक्रियाओं की तरह, काफी हद तक तापमान पर निर्भर है, जो

चावल। 1.6.Na/K पंप-ATPase का आरेख (प्लाज्मा झिल्ली के लिपिड बाईलेयर में डूबा हुआ), जो एक चक्र में संभावित और एकाग्रता ग्रेडिएंट के विरुद्ध सेल से तीन Na + आयनों को निकालता है और दो K आयनों को सेल में लाता है + . इस प्रक्रिया के दौरान, एक एटीपी अणु एडीपी और फॉस्फेट में टूट जाता है। आरेख ATPase को एक बड़े (कार्यात्मक) और छोटे सबयूनिट से युक्त डिमर के रूप में दिखाता है; झिल्ली में यह दो बड़ी और दो छोटी उपइकाइयों द्वारा निर्मित टेट्रामर के रूप में मौजूद होता है
चित्र में दिखाया गया है 1.7. यहां, मांसपेशियों की कोशिकाओं से सोडियम आयनों का प्रवाह समय के सापेक्ष दिखाया गया है; यह व्यावहारिक रूप से Na/K पंप के संचालन द्वारा मध्यस्थ सोडियम आयनों के प्रवाह के बराबर है, क्योंकि एकाग्रता और संभावित ग्रेडिएंट के विरुद्ध सोडियम आयनों का निष्क्रिय प्रवाह बेहद छोटा है। यदि दवा को लगभग 18°C तक ठंडा किया जाता है, तो कोशिका से सोडियम आयनों का प्रवाह तेजी से 15 गुना कम हो जाएगा, और गर्म करने के तुरंत बाद यह अपने मूल स्तर पर बहाल हो जाएगा। कोशिका से सोडियम आयनों के प्रवाह में यह कमी उस कमी से कई गुना अधिक है जो प्रसार प्रक्रिया या एक साधारण रासायनिक प्रतिक्रिया की तापमान निर्भरता के अनुरूप होगी। इसी तरह का प्रभाव तब देखा जाता है जब डाइनिट्रोफेनोल (डीएनपी) विषाक्तता के परिणामस्वरूप चयापचय ऊर्जा समाप्त हो जाती है (चित्र 1.7,5)। नतीजतन, कोशिका से सोडियम आयनों का प्रवाह एक ऊर्जा-निर्भर प्रतिक्रिया - एक सक्रिय पंप द्वारा सुनिश्चित किया जाता है। पंप की एक अन्य विशेषता, महत्वपूर्ण तापमान और ऊर्जा निर्भरता के साथ, संतृप्ति स्तर की उपस्थिति है (अन्य सभी रासायनिक प्रतिक्रियाओं की तरह); इसका मतलब यह है कि परिवहन किए गए आयनों की सांद्रता बढ़ने पर पंप की गति अनिश्चित काल तक नहीं बढ़ सकती है (चित्र 1.8)। इसके विपरीत, निष्क्रिय रूप से फैलने वाले पदार्थ का प्रवाह प्रसार के नियम (समीकरण 1 और 2) के अनुसार एकाग्रता अंतर के अनुपात में बढ़ता है।

चावल। 1.7. ए, बी.सक्रिय ना परिवहन + . वाई अक्ष:सेल से रेडियोधर्मी 24 Na + का प्रवाह (imp./min)। भुजिका अक्ष:प्रयोग शुरू होने के बाद से समय. एक।सेल को 18.3°C से 0.5°C तक ठंडा किया जाता है; प्रवाहना+ इस अवधि के दौरान कोशिका से अवरोध उत्पन्न होता है। बी। 0.2 mmol/l (संशोधित) की सांद्रता पर डाइनिट्रोफेनोल (DNP) द्वारा कोशिका से Na + फ्लक्स का दमन
Na/K पंप के अलावा, प्लाज्मा झिल्ली में कम से कम एक और पंप होता है - कैल्शियम;यह पंप कैल्शियम आयनों (Ca 2+) को कोशिका से बाहर पंप करता है और उनकी अंतःकोशिकीय सांद्रता को अत्यंत निम्न स्तर पर बनाए रखने में शामिल होता है (तालिका 1.1)। कैल्शियम पंप मांसपेशियों की कोशिकाओं के सार्कोप्लाज्मिक रेटिकुलम में बहुत उच्च घनत्व में मौजूद होता है, जो एटीपी अणुओं के टूटने के परिणामस्वरूप कैल्शियम आयन जमा करता है (अध्याय 4 देखें)।
झिल्ली क्षमता और कोशिका आयतन पर Na/K पंप का प्रभाव . चित्र में. चित्र 1.9 झिल्ली धारा के विभिन्न घटकों को दर्शाता है और आयनों की अंतःकोशिकीय सांद्रता को दर्शाता है

चावल। 1.8.चैनल के माध्यम से प्रसार के दौरान या पंपिंग परिवहन के दौरान अणुओं के परिवहन की दर और उनकी एकाग्रता (चैनल के प्रवेश द्वार पर या पंप के बंधन स्थल पर) के बीच संबंध। उत्तरार्द्ध उच्च सांद्रता (अधिकतम गति) पर संतृप्त हो जाता हैवीमैक्स ) अधिकतम पंप गति के आधे के अनुरूप x-अक्ष पर मान (वीमैक्स /2), संतुलन सांद्रता है को एम

चावल। 1.9.Na+ सांद्रता दर्शाने वाला आरेख , के+ औरसीएल - कोशिका के अंदर और बाहर और कोशिका झिल्ली के माध्यम से इन आयनों के प्रवेश के रास्ते (विशिष्ट आयन चैनलों के माध्यम से या Na/K पंप का उपयोग करके। दिए गए एकाग्रता ग्रेडियेंट पर, संतुलन क्षमताएंई ना, ई के और ई सी एल - संकेतित, झिल्ली क्षमता के बराबरएम = – 90 एमवी
उनका अस्तित्व सुनिश्चित करें. पोटेशियम आयनों का एक बाहरी प्रवाह पोटेशियम चैनलों के माध्यम से देखा जाता है, क्योंकि झिल्ली क्षमता पोटेशियम आयनों के लिए संतुलन क्षमता की तुलना में थोड़ी अधिक विद्युत धनात्मक होती है। सोडियम चैनलों का समग्र संचालन पोटेशियम चैनलों की तुलना में बहुत कम है, अर्थात। विश्राम क्षमता पर सोडियम चैनल पोटेशियम चैनलों की तुलना में बहुत कम बार खुले होते हैं; हालाँकि, लगभग उतनी ही संख्या में सोडियम आयन कोशिका में प्रवेश करते हैं, जब पोटेशियम आयन कोशिका से बाहर निकलते हैं, क्योंकि सोडियम आयनों को कोशिका में फैलने के लिए बड़ी सांद्रता और संभावित ग्रेडिएंट की आवश्यकता होती है। Na/K पंप निष्क्रिय प्रसार धाराओं के लिए आदर्श मुआवजा प्रदान करता है, क्योंकि यह सोडियम आयनों को कोशिका से बाहर और पोटेशियम आयनों को उसमें स्थानांतरित करता है। इस प्रकार, सेल के अंदर और बाहर स्थानांतरित होने वाले आवेशों की संख्या में अंतर के कारण पंप इलेक्ट्रोजेनिक है, जो अपने संचालन की सामान्य गति पर लगभग 10 की झिल्ली क्षमता बनाता है। एमवी अकेले निष्क्रिय आयन फ्लक्स द्वारा निर्मित होने की तुलना में अधिक विद्युत ऋणात्मक है (समीकरण 7 देखें)। परिणामस्वरूप, झिल्ली क्षमता पोटेशियम संतुलन क्षमता के करीब पहुंच जाती है, जिससे पोटेशियम आयनों का रिसाव कम हो जाता है। ना गतिविधि/K पंप को सोडियम आयनों की इंट्रासेल्युलर सांद्रता द्वारा नियंत्रित किया जाता है। पंप की गति धीमी हो जाती है क्योंकि सेल से निकाले जाने वाले सोडियम आयनों की सांद्रता कम हो जाती है (चित्र 1.8), जिससे पंप संचालन और सेल में सोडियम आयनों का प्रवाह एक दूसरे को संतुलित करता है, जिससे सोडियम की इंट्रासेल्युलर सांद्रता बनी रहती है। आयन लगभग 10 mmol/L के स्तर पर।
पंपिंग और निष्क्रिय झिल्ली धाराओं के बीच संतुलन बनाए रखने के लिए, पोटेशियम और सोडियम आयनों के लिए चैनल प्रोटीन की तुलना में कई अधिक Na/K पंप अणुओं की आवश्यकता होती है। जब चैनल खुला होता है, तो कुछ मिलीसेकंड में हजारों आयन इससे गुजरते हैं (ऊपर देखें), और चूंकि चैनल आमतौर पर प्रति सेकंड कई बार खुलता है, इस दौरान कुल मिलाकर 10 5 से अधिक आयन इससे गुजरते हैं। एक एकल पंप प्रोटीन प्रति सेकंड कई सौ सोडियम आयनों को स्थानांतरित करता है, इसलिए प्लाज्मा झिल्ली में चैनल अणुओं की तुलना में लगभग 1000 गुना अधिक पंप अणु होने चाहिए। आराम के समय चैनल धाराओं के मापन से प्रति 1 माइक्रोन 2 झिल्ली में औसतन एक पोटेशियम और एक सोडियम खुले चैनल की उपस्थिति देखी गई; इससे यह निष्कर्ष निकलता है कि Na/K पंप के लगभग 1000 अणु एक ही स्थान में मौजूद होने चाहिए, यानी। उनके बीच की दूरी औसतन 34 एनएम है; चैनल प्रोटीन के रूप में पंप प्रोटीन का व्यास 8-10 एनएम है। इस प्रकार, झिल्ली पंपिंग अणुओं से काफी सघन रूप से संतृप्त होती है
तथ्य यह है कि कोशिका में सोडियम आयनों और कोशिका से बाहर पोटेशियम आयनों के प्रवाह की भरपाई पंप के संचालन से होती है, इसका एक और परिणाम है, जिसमें एक स्थिर आसमाटिक दबाव बनाए रखना शामिल है और स्थिर मात्रा.कोशिका के अंदर बड़े आयनों, मुख्य रूप से प्रोटीन (ए - तालिका 1.1 में) की उच्च सांद्रता होती है, जो झिल्ली में प्रवेश करने में सक्षम नहीं होते हैं (या बहुत धीरे-धीरे इसके माध्यम से प्रवेश करते हैं) और इसलिए कोशिका के अंदर एक निश्चित घटक होते हैं। इन आयनों के आवेश को संतुलित करने के लिए समान संख्या में धनायनों की आवश्यकता होती है। Na/K पंप की क्रिया के कारण, ये धनायन मुख्य रूप से पोटेशियम आयन होते हैं। इंट्रासेल्युलर आयन सांद्रता में उल्लेखनीय वृद्धि केवल C1 के प्रवाह के कारण आयनों की सांद्रता में वृद्धि के साथ हो सकती है - कोशिका में सांद्रता प्रवणता के साथ (तालिका 1.1), लेकिन झिल्ली क्षमता इसका प्रतिकार करती है। आने वाली धारासीएल - केवल तब तक मनाया जाता है जब तक क्लोरीन आयनों के लिए संतुलन क्षमता तक नहीं पहुंच जाता; यह तब देखा जाता है जब क्लोरीन आयन ग्रेडिएंट पोटेशियम आयन ग्रेडिएंट के लगभग विपरीत होता है, क्योंकि क्लोरीन आयन नकारात्मक रूप से चार्ज होते हैं (समीकरण 4)। इस प्रकार, क्लोरीन आयनों की एक कम इंट्रासेल्युलर सांद्रता स्थापित की जाती है, जो पोटेशियम आयनों की कम बाह्यकोशिकीय सांद्रता के अनुरूप होती है। परिणामस्वरुप कोशिका में आयनों की कुल संख्या सीमित हो जाती है। यदि Na/K पंप अवरुद्ध होने पर झिल्ली क्षमता कम हो जाती है, उदाहरण के लिए एनोक्सिया के दौरान, तो क्लोरीन आयनों के लिए संतुलन क्षमता कम हो जाती है, और क्लोराइड आयनों की इंट्रासेल्युलर सांद्रता तदनुसार बढ़ जाती है। आवेशों के संतुलन को बहाल करते हुए, पोटेशियम आयन भी कोशिका में प्रवेश करते हैं; कोशिका में आयनों की कुल सांद्रता बढ़ जाती है, जिससे आसमाटिक दबाव बढ़ जाता है; यह पानी को कोशिका में जाने के लिए बाध्य करता है। कोशिका सूज जाती है। यह सूजन देखी जाती हैविवो में ऊर्जा की कमी की स्थिति में.
कम और अधिक घनत्व के बीच में एक घुले हुए पदार्थ का जमाव ना + झिल्ली परिवहन के लिए एक प्रेरक शक्ति के रूप में . कोशिका के लिए Na/K पंप का महत्व झिल्ली पर सामान्य K+ और Na+ ग्रेडिएंट के स्थिरीकरण तक सीमित नहीं है। झिल्ली प्रवणता में संग्रहित ऊर्जाना+ , अक्सर अन्य पदार्थों के झिल्ली परिवहन को सुविधाजनक बनाने के लिए उपयोग किया जाता है। उदाहरण के लिए, चित्र में. 1.10 "महत्व" दिखाता हैना+ और कोशिका में शर्करा के अणु। झिल्ली परिवहन प्रोटीन एक सांद्रता प्रवणता के विरुद्ध भी चीनी अणु को कोशिका में पहुँचाता है, जबकि उसी समय Na + सांद्रता और संभावित प्रवणता के साथ चलता है, जिससे ऊर्जा मिलती है शर्करा का परिवहन.शर्करा का यह परिवहन पूरी तरह से उच्च ढाल के अस्तित्व पर निर्भर हैना+ ; अगर इंट्रासेल्युलर एकाग्रताना+ काफी बढ़ जाता है, शर्करा का परिवहन रुक जाता है। विभिन्न के लिएसी अखारोव, विभिन्न आयात प्रणालियाँ हैं। अमीनो एसिड का परिवहनपरिवहन के समान एक कोशिका मेंसी अखारोव को चित्र में दिखाया गया है। 1.10; यह ग्रेडिएंट द्वारा भी प्रदान किया जाता हैना+ , कम से कम पांच अलग-अलग सिंपोर्ट सिस्टम हैं, जिनमें से प्रत्येक संबंधित अमीनो एसिड के एक समूह के लिए विशिष्ट है।
आयात प्रणालियों के अलावा, वहाँ भी हैं "पोर्टर्स विरोधी"।उनमें से एक, उदाहरण के लिए, एक चक्र में तीन आने वाले सोडियम आयनों के बदले में कोशिका से एक कैल्शियम आयन स्थानांतरित करता है (चित्र 1.10)। के लिए ऊर्जा सीए 2+ परिवहनएक सांद्रता और संभावित ढाल के साथ तीन सोडियम आयनों के प्रवेश के कारण बनता है। यह ऊर्जा उच्च कैल्शियम आयन ग्रेडिएंट (कोशिका के अंदर 10-7 mol/L से कम से लेकर कोशिका के बाहर लगभग 2 mmol/L तक) बनाए रखने के लिए पर्याप्त है (विश्राम क्षमता पर)।
एंडो- और एक्सोसाइटोसिस . कुछ पदार्थों के लिए जो कोशिका में प्रवेश करते हैं या उत्सर्जित होने चाहिए

चावल। 1.10.झिल्ली के लिपिड बाईलेयर में एम्बेडेड प्रोटीन कोशिका में ग्लूकोज और Na + के सहसंबंध में मध्यस्थता करते हैं, साथ ही Ca2+/Na+ -एंटीपोर्ट, जिसमें प्रेरक शक्ति कोशिका झिल्ली पर Na + ग्रेडिएंट है
इससे कोई परिवहन चैनल नहीं हैं; इन पदार्थों में, उदाहरण के लिए, प्रोटीन और कोलेस्ट्रॉल शामिल हैं। वे प्लाज्मा झिल्ली से होकर गुजर सकते हैं पुटिका,या बुलबुले,एंडो- और एक्सोसाइटोसिस के माध्यम से। चित्र में. चित्र 1.11 इन प्रक्रियाओं के मुख्य तंत्र को दर्शाता है। एक्सोसाइटोसिस में, कुछ अंगक (नीचे देखें) ऐसे पदार्थ से भरे पुटिकाओं का निर्माण करते हैं जिन्हें कोशिका से निकालने की आवश्यकता होती है, जैसे हार्मोन या बाह्य कोशिकीय एंजाइम। जब ऐसे पुटिकाएं प्लाज्मा झिल्ली तक पहुंचती हैं, तो उनकी लिपिड झिल्ली इसके साथ विलीन हो जाती है, जिससे सामग्री बाहरी वातावरण में बच जाती है। विपरीत प्रक्रिया, एन्डोसाइटोसिस में, प्लाज्मा झिल्ली आक्रमण करती है, जिससे एक गड्ढा बनता है, जो फिर गहरा होता है और बंद हो जाता है, जिससे बाह्य कोशिकीय द्रव और कुछ मैक्रोमोलेक्यूल्स से भरा एक इंट्रासेल्युलर पुटिका बनता है। इस झिल्ली संलयन और पुटिका के बंद होने को सुनिश्चित करने के लिए, साइटोस्केलेटन के संकुचनशील तत्व स्वयं झिल्ली के साथ मिलकर कार्य करते हैं (नीचे देखें)। एन्डोसाइटोसिस में हमेशा कोशिका में बाह्य कोशिकीय वातावरण का कब्ज़ा शामिल नहीं होता है। कोशिका झिल्ली में अक्सर मैक्रोमोलेक्यूल्स के लिए विशिष्ट रिसेप्टर्स होते हैं, जैसे इंसुलिन या एंटीजन, जो अक्सर विशेष समूहों में व्यवस्थित होते हैं। इन मैक्रोमोलेक्यूल्स के अपने रिसेप्टर्स से बंधने के बाद, रिसेप्टर के आसपास के झिल्ली क्षेत्र में एंडोसाइटोसिस होता है, और मैक्रोमोलेक्यूल्स को चुनिंदा रूप से कोशिका में ले जाया जाता है (चित्र 1.12, बी)।
एंडो- और एक्सोसाइटोसिस कोशिकाओं में लगातार होते रहते हैं। झिल्ली सामग्री कारोबार की मात्रा महत्वपूर्ण है; 1 घंटे के भीतर, मैक्रोफेज पुटिकाओं के रूप में अपने साइटोप्लाज्मिक झिल्ली के सतह क्षेत्र को दोगुना अवशोषित कर लेता है। अधिकांश कोशिकाओं में, झिल्ली सामग्री का कारोबार इतनी तीव्रता से नहीं होता है, लेकिन फिर भी यह महत्वपूर्ण होना चाहिए।

चावल। 1.11.एक्सोसाइटोसिस और एंडोसाइटोसिस। ऊपर:अंतःकोशिकीय पुटिका प्लाज्मा झिल्ली के लिपिड बाइलेयर के साथ विलीन हो जाती है और बाह्यकोशिकीय स्थान में खुलती है। इस प्रक्रिया को एक्सोसाइटोसिस कहा जाता है। तल पर:प्लाज़्मा झिल्ली एक छोटे से क्षेत्र में आक्रमण करती है और बाह्य कोशिकीय सामग्री से भरी एक पुटिका छोड़ती है। इस प्रक्रिया को एन्डोसाइटोसिस कहा जाता है
1.3. कोशिका के भीतर पदार्थों का परिवहन
एंडो- और एक्सोसाइटोसिस न केवल कोशिका झिल्ली में पदार्थों के परिवहन की प्रक्रियाएं हैं, बल्कि झिल्ली के आदान-प्रदान की प्रक्रियाएं भी हैं - कोशिका के संरचनात्मक घटक। इस खंड में विचार का विषय कोशिका और उसके अंगों में अन्य समान परिवहन प्रक्रियाएं हैं।
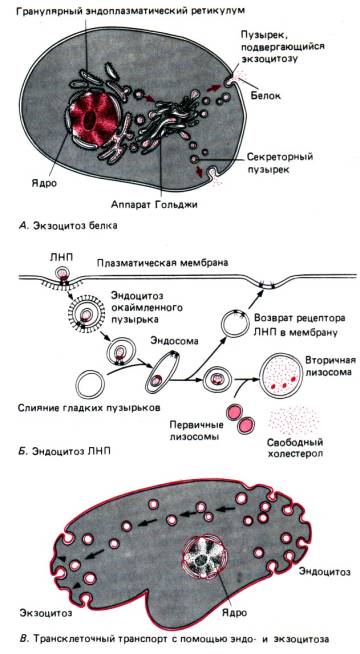
चावल। 1.12. ए-बी.एक्सो- और एंडोसाइटोसिस सहित प्रक्रियाओं की योजना। एक।दानेदार एंडोप्लाज्मिक रेटिकुलम में संश्लेषित प्रोटीन को गोल्गी तंत्र के माध्यम से प्लाज्मा झिल्ली में ले जाया जाता है, जहां इसे एक्सोसाइटोसिस द्वारा स्रावित किया जाता है। बी।एलडीएल (कम घनत्व वाले लिपोप्रोटीन) कणों से बंधा कोलेस्ट्रॉल प्लाज्मा झिल्ली से जुड़ जाता है, झिल्ली के इस क्षेत्र में एक एंडोसाइटिक पुटिका के गठन को प्रेरित करता है और लाइसोसोम में ले जाया जाता है, जहां इसे जारी किया जाता है। में।एंडोसाइटोसिस के दौरान कैप्चर की गई बाह्यकोशिकीय सामग्री (चित्र में)। दायी ओर),पुटिकाओं, या पुटिकाओं में कोशिका के माध्यम से परिवहन किया जाता है, और एक्सोसाइटोसिस के माध्यम से जारी किया जाता है (चित्र में) बाएं)
प्रसार . स्वाभाविक रूप से, साइटोसोल में प्रसार के कारण सांद्रता में अंतर समाप्त हो जाता है; यही बात अंगकों में मौजूद तरल पदार्थों के लिए भी सच है। घुले हुए प्रोटीन की उच्च सांद्रता के कारण, यहाँ प्रसार पानी की तुलना में बहुत धीमा है। लिपिड झिल्ली - कोशिकाओं के चारों ओर और अंगों के भीतर - द्वि-आयामी तरल पदार्थ हैं जिनमें प्रसार होता है। झिल्लीदार द्विपरत में लिपिड अपनी ही परत के भीतर फैलते हैं, शायद ही कभी एक से दूसरे में जाते हैं। उनमें डूबे प्रोटीन भी काफी गतिशील होते हैं; वे झिल्ली के लंबवत एक अक्ष के चारों ओर घूमते हैं या बहुत भिन्न प्रसार स्थिरांक के साथ पार्श्व में फैलते हैं, फॉस्फोलिपिड्स की तुलना में 2-10,000 गुना धीमी गति से। इसलिए, यदि कुछ प्रोटीन लिपिड परत में स्वतंत्र रूप से और लिपिड अणुओं के समान गति से चलते हैं, तो अन्य स्थिर हो जाते हैं, यानी। साइटोस्केलेटन से काफी मजबूती से जुड़ा हुआ है। झिल्ली में विशिष्ट प्रोटीन के "स्थायी" समुच्चय होते हैं, उदाहरण के लिए, तंत्रिका कोशिकाओं की पूर्व और पोस्टसिनेप्टिक संरचनाएं। स्वतंत्र रूप से घूमने वाले प्रोटीन को फ्लोरोसेंट रंगों से बांधकर प्रदर्शित किया जा सकता है, जिसकी चमक झिल्ली के एक छोटे से क्षेत्र को संक्षेप में रोशन करके प्रेरित होती है। ऐसे प्रयोगों से पता चलता है कि 1 मिनट से भी कम समय में, डाई से जुड़े प्रोटीन झिल्ली पर 10 माइक्रोन तक की दूरी पर समान रूप से वितरित हो जाते हैं।
ऑर्गेनेल झिल्लियों में सक्रिय परिवहन .
सक्रिय परिवहन प्रक्रियाएं, जो प्लाज्मा झिल्ली के कामकाज में महत्वपूर्ण भूमिका निभाती हैं, कोशिका के अंदर, ऑर्गेनेल की झिल्ली में भी होती हैं। विभिन्न अंगों की विशिष्ट सामग्री आंशिक रूप से आंतरिक संश्लेषण के कारण और आंशिक रूप से साइटोसोल से सक्रिय परिवहन के कारण बनती है। उत्तरार्द्ध का एक उदाहरण मांसपेशी कोशिकाओं के सार्कोप्लाज्मिक रेटिकुलम में उपर्युक्त सीए 2+ पंप है। यह विशेष रूप से दिलचस्प है कि माइटोकॉन्ड्रिया में एटीपी संश्लेषण के मामले में, प्लाज्मा झिल्ली के एटीपीस पंपों में जो होता है, उसके विपरीत सिद्धांत लागू होता है (चित्र 1.6)। एटीपी संश्लेषण के दौरान, ऑक्सीडेटिव चयापचय एक तीव्र ढाल के गठन की ओर जाता हैएच+ आंतरिक झिल्लियों पर. यह ढाल अणुओं के सक्रिय परिवहन के पंपिंग चक्र की रिवर्स प्रक्रिया के लिए प्रेरक शक्ति है: एच + आयन ढाल के साथ झिल्ली में चलते हैं, और परिणामस्वरूप जारी ऊर्जा एडीपी और फॉस्फेट से एटीपी का संश्लेषण प्रदान करती है। परिणामी एटीपी, बदले में, सक्रिय परिवहन सहित कोशिका को ऊर्जा प्रदान करता है।
पुटिकाओं में परिवहन . कोशिका में बड़ी संख्या में अंगक और संबद्ध पुटिकाएं होती हैं (चित्र 1.1)। ये अंगक, और विशेष रूप से पुटिकाएं, निरंतर गति में हैं, अपनी सामग्री को अन्य अंगक या प्लाज्मा झिल्ली तक पहुंचाते हैं। वेसिकल्स कोशिका झिल्ली से ऑर्गेनेल में भी स्थानांतरित हो सकते हैं, जैसे एन्डोसाइटोसिस में।
प्रक्रिया प्रोटीन स्रावचित्र में दिखाया गया है 1.12, एक।प्रोटीन को एंडोप्लाज्मिक रेटिकुलम (तथाकथित दानेदार, या खुरदरा, एंडोप्लाज्मिक रेटिकुलम) से जुड़े राइबोसोम पर कोशिका नाभिक के पास संश्लेषित किया जाता है; एक बार एंडोप्लाज्मिक रेटिकुलम में, प्रोटीन को परिवहन पुटिकाओं में पैक किया जाता है, जो ऑर्गेनेल से अलग हो जाते हैं और गोल्गी तंत्र में स्थानांतरित हो जाते हैं। यहां वे गोल्गी तंत्र के सिस्टर्न में विलीन हो जाते हैं, जहां प्रोटीन को संशोधित किया जाता है (यानी, ग्लाइकोप्रोटीन में परिवर्तित किया जाता है)। सिस्टर्न के सिरों पर, पुटिकाएं फिर से अलग हो जाती हैं। संशोधित प्रोटीन ले जाने वाले स्रावी पुटिकाएं प्लाज्मा झिल्ली में चली जाती हैं और एक्सोसाइटोसिस द्वारा अपनी सामग्री को छोड़ देती हैं।
कोशिका में परिवहन मार्ग का एक अन्य उदाहरण चित्र में दिखाया गया है। 1.12, बी; यह कोशिका द्वारा कोलेस्ट्रॉल का अवशोषण है। रक्त में पहुँचाया जाने वाला कोलेस्ट्रॉल मुख्य रूप से कणों जैसे प्रोटीन से बंधा होता है "निम्न घनत्व लिपोप्रोटीन"(एलडीएल)। ऐसे कण एलडीएल रिसेप्टर्स वाले विशिष्ट झिल्ली क्षेत्रों से जुड़ते हैं, जहां एंडोसाइटोसिस होता है और एलडीएल को "सीमाबद्ध" पुटिकाओं में कोशिका में ले जाया जाता है। ये पुटिकाएं मिलकर एंडोसोम बनाती हैं और इस प्रक्रिया के दौरान अपना "किनारा" खो देती हैं। एंडोसोम, बदले में, प्राथमिक लाइसोसोम के साथ विलय करते हैं, जिसमें मुख्य रूप से हाइड्रोलाइटिक एंजाइम होते हैं, और माध्यमिक, बड़े लाइसोसोम बनाते हैं। उनमें, कोलेस्ट्रॉल एलडीएल कणों से निकलता है और साइटोसोल में फैल जाता है, जहां यह उपलब्ध हो जाता है, उदाहरण के लिए, लिपिड झिल्ली के संश्लेषण के लिए। जिन पुटिकाओं में एलडीएल नहीं होता है, उन्हें एंडोसोम से भी अलग किया जाता है, जो एक विशेष तरीके से प्लाज्मा झिल्ली में चले जाते हैं और इसके साथ विलय करते हैं, झिल्ली सामग्री और संभवतः, एलडीएल के लिए रिसेप्टर्स लौटाते हैं। एलडीएल कण के झिल्ली से जुड़ने के क्षण से लेकर द्वितीयक लाइसोसोम से कोलेस्ट्रॉल निकलने तक 10-15 मिनट बीत जाते हैं। एलडीएल के बंधन और अवशोषण में कमी, यानी कोशिका को कोलेस्ट्रॉल की आपूर्ति में कमी, एक गंभीर और व्यापक बीमारी, एथेरोस्क्लेरोसिस (धमनियों का "सख्त होना") के विकास में निर्णायक भूमिका निभाती है।
चित्र में दिखाए गए मार्गों के समान कई अन्य परिवहन मार्ग हैं। 1.11 और 1.12, ए, जिसकी सहायता से कोशिका में विशिष्ट पुटिकाएँ गति करती हैं। यह ठीक से ज्ञात नहीं है कि वे कैसे चलते हैं, लेकिन संभवतः साइटोस्केलेटल तत्व इस प्रक्रिया में शामिल होते हैं। पुटिकाएं सूक्ष्मनलिकाएं के साथ सरक सकती हैं, ऐसी स्थिति में गति के लिए ऊर्जा स्पष्ट रूप से पुटिकाओं से जुड़े प्रोटीन, एटीपीस (नीचे देखें) द्वारा प्रदान की जाती है। यह पूरी तरह से अस्पष्ट है कि कितने अलग-अलग पुटिकाएँ, सभी दिशाओं में एक के बाद एक चलते हुए, अपने गंतव्य तक पहुँचती हैं। उन्हें स्पष्ट रूप से इस तरह से "चिह्नित" करने की आवश्यकता है कि परिवहन प्रणाली द्वारा पहचाना जाए और लक्षित आंदोलन में परिवर्तित किया जाए।
अंगकों के निर्माण और विनाश द्वारा परिवहन . अब तक, हमने एंडो- और एक्सोसाइटोसिस को पुटिकाओं की सामग्री के परिवहन की प्रक्रिया के रूप में माना है। इन प्रक्रियाओं का एक और पहलू है, जो इस तथ्य में निहित है कि एंडोसाइटोसिस द्वारा कोशिका की सतह के एक हिस्से पर प्लाज्मा झिल्ली को लक्षित हटाने और, इसके विपरीत, एक्सोसाइटोसिस द्वारा दूसरे पर इसका जोड़ झिल्ली के बड़े क्षेत्रों को स्थानांतरित करता है (चित्र) 1.12.ई), उदाहरण के लिए, कोशिका को एक वृद्धि या गति का अवसर देता है।
इसी तरह की पुनर्व्यवस्थाएं साइटोस्केलेटन के लिए भी विशिष्ट हैं, विशेष रूप से माइक्रोफिलामेंट्स और सूक्ष्मनलिकाएं (चित्र 1.1) के लिए। माइक्रोफिलामेंट्समुख्य रूप से शामिल हैं एफ-एक्टिन प्रोटीन,जो साइटोसोल से मोनोमर के पोलीमराइजेशन के परिणामस्वरूप रेशेदार बंडलों में संयोजन करने में सक्षम है। बंडल ध्रुवीकृत होते हैं, यानी, वे अक्सर केवल एक छोर पर बढ़ते हैं, नए एक्टिन अणुओं को जमा करते हैं, जबकि दूसरा छोर निष्क्रिय होता है या यहां विघटन होता है। इस ध्रुवीकृत वृद्धि के कारण, माइक्रोफ़िलामेंट कुशलता से चलते हैं और उनके नेटवर्क की संरचना बदल सकती है। एक्टिन का डिपॉलीमराइज़्ड अवस्था (सोल) से संगठित अवस्था (जेल) में संक्रमण अन्य प्रोटीन के प्रभाव या आयन सांद्रता में परिवर्तन (नीचे देखें) के तहत बहुत जल्दी हो सकता है। ऐसे प्रोटीन भी होते हैं जो एक्टिन फिलामेंट्स को तोड़कर छोटे टुकड़े बनाते हैं। कई कोशिकाओं के पतले प्रक्षेपण - फ़िलाओपोडिया - में एक्टिन का एक केंद्रीय बंडल होता है (चित्र 1.1), और फ़िलोपोडिया की विभिन्न गतिविधियाँ संभवतः एक्टिन संक्रमण के कारण होती हैं: पोलीमराइज़ेशन - डीपोलीमराइज़ेशन।
सूक्ष्मनलिकाएंभी अक्सर इसी तरह की हरकतों से गुजरते हैं। इन आंदोलनों का तंत्र समान है - साइटोसोल से ट्यूबुलिन का पोलीमराइजेशन इस तरह से होता है कि सूक्ष्मनलिका का एक सिरा बढ़ता है, जबकि दूसरा या तो नहीं बदलता है, या वहां डिस्सेप्लर होता है। इस प्रकार, एक सूक्ष्मनलिका, सामग्री को उचित रूप से जोड़ने या हटाने से, साइटोसोल में घूम सकती है।
साइटोस्केलेटन की सक्रिय गतिविधियां . साइटोस्केलेटल संरचनाओं में परिवर्तन ऊपर वर्णित सक्रिय आंदोलनों और पुनर्व्यवस्था दोनों के परिणामस्वरूप हो सकता है। कई मामलों में, सूक्ष्मनलिकाएं और एक्टिन फिलामेंट्स की गति संकुचनशील प्रोटीन द्वारा संचालित होती है जो फिलामेंट्स या नलिकाओं को बांधती है और उन्हें एक दूसरे के सापेक्ष स्थानांतरित कर सकती है। गिलहरी मायोसिन और डायनेइनसभी कोशिकाओं के साइटोसोल में अपेक्षाकृत उच्च सांद्रता में मौजूद; वे ऐसे तत्व हैं जो ऊर्जा को विशेष कोशिकाओं (मांसपेशियों) और ऑर्गेनेल (सिलिया) में गति में परिवर्तित करते हैं। मांसपेशी कोशिकाओं में, मायोसिन एक्टिन फिलामेंट्स के समानांतर उन्मुख मोटे फिलामेंट्स बनाता है। मायोसिन अणु अपने "सिर" के साथ एक्टिन फिलामेंट से जुड़ जाता है और, एटीपी की ऊर्जा का उपयोग करके, एक्टिन अणु के साथ मायोसिन को विस्थापित करता है।मायोसिन फिर एक्टिन से अलग हो जाता है। ऐसे कई कनेक्शन-डिस्कनेक्शन चक्रों का संयोजन स्थूलता की ओर ले जाता है मांसपेशीय तंतुओं का संकुचन(अध्याय 4)। सिलिया के दौरान सूक्ष्मनलिकाएं की गति में डायनेइन एक समान भूमिका निभाता है (चित्र 1.1)। अविशिष्ट कोशिकाओं के साइटोप्लाज्म में, मायोसिन और डायनेइन नियमित फाइबर नहीं बनाते हैं, लेकिन ज्यादातर मामलों में अणुओं के छोटे समूह बनाते हैं। ऐसे छोटे समुच्चय में भी, वे एक्टिन फिलामेंट्स या सूक्ष्मनलिकाएं को स्थानांतरित करने में सक्षम हैं। चावल। चित्र 1.13 इस प्रक्रिया को दर्शाता है जब विपरीत ध्रुवीकृत मायोसिन अणु भी दो एक्टिन फिलामेंट्स से जुड़े होते हैं, जो अलग-अलग दिशाओं में ध्रुवीकृत होते हैं। मायोसिन हेडग्रुप अणु की पूंछ की ओर झुकते हैं, इस प्रक्रिया में एटीपी का उपभोग करते हैं, और दो एक्टिन फिलामेंट विपरीत दिशा में चलते हैं, जिसके बाद मायोसिन उनसे अलग हो जाता है। इस प्रकार के आंदोलन, जिसके दौरान एटीपी ऊर्जा को यांत्रिक कार्य में परिवर्तित किया जाता है, साइटोस्केलेटन के आकार को बदल सकता है और, परिणामस्वरूप, कोशिका, और साइटोस्केलेटन से जुड़े ऑर्गेनेल का परिवहन भी प्रदान करता है।
इंट्रासेल्युलर परिवहन की प्रक्रियाओं को तंत्रिका कोशिका के अक्षतंतु पर सबसे स्पष्ट रूप से प्रदर्शित किया जा सकता है। एक्सोन परिवहनअधिकांश कोशिकाओं में समान तरीकों से घटित होने वाली घटनाओं को दर्शाने के लिए यहां विस्तार से चर्चा की गई है। एक अक्षतंतु जिसका व्यास केवल कुछ माइक्रोन है, एक मीटर या उससे अधिक की लंबाई तक पहुंच सकता है, और नाभिक से अक्षतंतु के दूरस्थ अंत तक प्रसार द्वारा प्रोटीन की गति में वर्षों लग जाएंगे। यह लंबे समय से ज्ञात है कि जब अक्षतंतु का कोई भी भाग संकुचन से गुजरता है, तो अक्षतंतु का अधिक समीप स्थित भाग फैलता है। ऐसा लगता है जैसे अक्षतंतु में केन्द्रापसारक प्रवाह अवरुद्ध हो गया है। ऐसा प्रवाह-तेज अक्षतंतु परिवहन कर सकते हैंरेडियोधर्मी मार्करों की गति द्वारा प्रदर्शित किया जा सकता है, जैसा कि चित्र में दिखाए गए प्रयोग में है। 1.14. रेडियोलेबल ल्यूसीन को पृष्ठीय जड़ नाड़ीग्रन्थि में इंजेक्ट किया गया था, और फिर दूसरे से 10 वें घंटे तक न्यूरोनल कोशिका निकायों से 166 मिमी की दूरी पर कटिस्नायुशूल तंत्रिका में रेडियोधर्मिता को मापा गया था। 10 घंटों में, इंजेक्शन स्थल पर रेडियोधर्मिता का चरम थोड़ा बदल गया। लेकिन रेडियोधर्मिता की लहर अक्षतंतु के साथ 2 घंटे में लगभग 34 मिमी, या 410 मिमी/दिन की निरंतर गति से फैलती है। यह दिखाया गया है कि होमोथर्मिक जानवरों के सभी न्यूरॉन्स में, तेज एक्सोनल परिवहन एक ही गति से होता है, और पतले, अनमाइलिनेटेड फाइबर और सबसे मोटे एक्सोन के साथ-साथ मोटर और संवेदी फाइबर के बीच कोई ध्यान देने योग्य अंतर नहीं देखा जाता है। रेडियोधर्मी मार्कर का प्रकार भी तेज़ एक्सोनल परिवहन की दर को प्रभावित नहीं करता है; मार्कर विभिन्न प्रकार के रेडियोधर्मी के रूप में काम कर सकते हैं

चावल। 1.13.गैर-मांसपेशी मायोसिन कॉम्प्लेक्स, एक निश्चित अभिविन्यास के साथ, विभिन्न ध्रुवता के एक्टिन फिलामेंट्स से बंध सकता है और एटीपी की ऊर्जा का उपयोग करके, उन्हें एक दूसरे के सापेक्ष विस्थापित कर सकता है।
अणु, जैसे विभिन्न अमीनो एसिड, जो न्यूरॉन के कोशिका शरीर के प्रोटीन में शामिल होते हैं। यदि हम यहां पहुंचाए गए रेडियोधर्मिता के वाहकों की प्रकृति निर्धारित करने के लिए तंत्रिका के परिधीय भाग का विश्लेषण करते हैं, तो ऐसे वाहक मुख्य रूप से प्रोटीन अंश में पाए जाते हैं, लेकिन मध्यस्थों और मुक्त अमीनो एसिड की संरचना में भी पाए जाते हैं। यह जानते हुए कि इन पदार्थों के गुण अलग-अलग हैं और उनके अणुओं के आकार विशेष रूप से भिन्न हैं, हम परिवहन की निरंतर गति को उन सभी के लिए सामान्य परिवहन तंत्र द्वारा ही समझा सकते हैं।
ऊपर वर्णित है तेज़ अक्षतंतु परिवहनहै अग्रगामी,यानी, कोशिका शरीर से दूर निर्देशित। यह दिखाया गया है कि कुछ पदार्थ परिधि से कोशिका शरीर की ओर बढ़ते हैं प्रतिगामी परिवहन.उदाहरण के लिए, एसिटाइलकोलिनेस्टरेज़ को इस दिशा में तेज़ एक्सोनल परिवहन की गति से 2 गुना धीमी गति से ले जाया जाता है। न्यूरोएनाटॉमी में अक्सर उपयोग किया जाने वाला मार्कर, हॉर्सरैडिश पेरोक्सीडेज, प्रतिगामी परिवहन द्वारा भी चलता है। प्रतिगामी परिवहन संभवतः कोशिका शरीर में प्रोटीन संश्लेषण के नियमन में महत्वपूर्ण भूमिका निभाता है। अक्षतंतु के कटने के कुछ दिनों बाद, कोशिका शरीर में क्रोमैटोलिसिस देखा जाता है, जो प्रोटीन संश्लेषण में व्यवधान का संकेत देता है। क्रोमैटोलिसिस के लिए आवश्यक समय अक्षतंतु संक्रमण के स्थल से कोशिका शरीर तक प्रतिगामी परिवहन की अवधि से संबंधित होता है। यह परिणाम इस विकार के लिए एक स्पष्टीकरण भी सुझाता है - प्रोटीन संश्लेषण को नियंत्रित करने वाले "सिग्नल पदार्थ" की परिधि से संचरण बाधित होता है। यह स्पष्ट है कि मुख्य "वाहन" का उपयोग तेजी से एक्सोनल के लिए किया जाता है
चावल। 1.14.बिल्ली कटिस्नायुशूल तंत्रिका के संवेदी तंतुओं में तेजी से एक्सोनल परिवहन का प्रदर्शन करने वाला प्रयोग। ट्रिटियम-लेबल ल्यूसीन को पृष्ठीय जड़ नाड़ीग्रन्थि में इंजेक्ट किया जाता है और नाड़ीग्रन्थि और संवेदी तंतुओं में रेडियोधर्मिता को इंजेक्शन के 2, 4, 6, 8 और 10 घंटे पर मापा जाता है। (तस्वीर के नीचे).द्वारा X- अक्षनाड़ीग्रन्थि से कटिस्नायुशूल तंत्रिका के क्षेत्रों तक की दूरी जहां माप किया जाता है, प्लॉट किया जाता है। केवल ऊपरी और निचले वक्रों के लिए कोर्डिनेट अक्ष पर, रेडियोधर्मिता (सीपीएस/मिनट) को लघुगणकीय पैमाने पर प्लॉट किया जाता है। बढ़ी हुई रेडियोधर्मिता की "लहर"। (तीर) 410 मिमी/दिन (साथ) की गति से चलती है
परिवहन हैं वेसिकल्स (वेसिकल्स) और ऑर्गेनेल,जैसे कि माइटोकॉन्ड्रिया, जिसमें ऐसे पदार्थ होते हैं जिन्हें परिवहन की आवश्यकता होती है। सबसे बड़े पुटिकाओं या माइटोकॉन्ड्रिया की गति को माइक्रोस्कोप का उपयोग करके देखा जा सकता हैविवो में . ऐसे कण एक दिशा में छोटी, तीव्र गति करते हैं, रुकते हैं, अक्सर थोड़ा पीछे या किनारे की ओर बढ़ते हैं, फिर रुकते हैं और फिर मुख्य दिशा में झटके मारते हैं। 410 मिमी/दिन लगभग 5 μm/s की पूर्ववर्ती गति की औसत गति से मेल खाता है; इसलिए, प्रत्येक व्यक्तिगत गति की गति बहुत अधिक होनी चाहिए, और यदि हम ऑर्गेनेल, फिलामेंट्स और सूक्ष्मनलिकाएं के आकार को ध्यान में रखते हैं, तो ये गति वास्तव में बहुत तेज होती हैं। तेज़ एक्सोनल परिवहन के लिए एटीपी की महत्वपूर्ण सांद्रता की आवश्यकता होती है। कोल्सीसिन जैसे जहर, जो सूक्ष्मनलिकाएं को बाधित करते हैं, तेजी से एक्सोनल परिवहन को भी रोकते हैं। इससे यह पता चलता है कि जिस परिवहन प्रक्रिया पर हम विचार कर रहे हैं, उसमें पुटिका और अंगक सूक्ष्मनलिकाएं और एक्टिन फिलामेंट्स के साथ चलते हैं; यह गति डायनेइन और मायोसिन अणुओं के छोटे समुच्चय द्वारा प्रदान की जाती है, जैसा कि चित्र में दिखाया गया है। 1.13, एटीपी ऊर्जा का उपयोग करते हुए।
तेज़ अक्षतंतु परिवहन भी इसमें शामिल हो सकता है पैथोलॉजिकल प्रक्रियाएं।कुछ न्यूरोट्रोपिक वायरस (उदाहरण के लिए, हर्पीस या पोलियो वायरस) परिधि पर अक्षतंतु में प्रवेश करते हैं और प्रतिगामी परिवहन के माध्यम से न्यूरॉन शरीर में चले जाते हैं, जहां वे गुणा करते हैं और अपने विषाक्त प्रभाव डालते हैं। टेटनस टॉक्सिन, बैक्टीरिया द्वारा निर्मित एक प्रोटीन है जो त्वचा के क्षतिग्रस्त होने पर शरीर में प्रवेश करता है, तंत्रिका अंत द्वारा पकड़ लिया जाता है और न्यूरॉन शरीर में ले जाया जाता है, जहां यह विशिष्ट मांसपेशियों में ऐंठन का कारण बनता है। एक्सॉन परिवहन पर विषाक्त प्रभाव के ज्ञात मामले हैं, उदाहरण के लिए, औद्योगिक विलायक एक्रिलामाइड के संपर्क में आना। इसके अलावा, यह माना जाता है कि विटामिन की कमी बेरीबेरी और अल्कोहल पोलीन्यूरोपैथी के रोगजनन में तेजी से एक्सोनल परिवहन का उल्लंघन शामिल है।
कोशिका में तेज एक्सोनल परिवहन के अलावा काफी तीव्र भी होता है धीमा अक्षतंतु परिवहन।ट्यूबुलिन अक्षतंतु के साथ लगभग 1 मिमी/दिन की गति से चलता है, और एक्टिन तेजी से चलता है - 5 मिमी/दिन तक। अन्य प्रोटीन इन साइटोस्केलेटल घटकों के साथ पलायन करते हैं; उदाहरण के लिए, एंजाइम एक्टिन या ट्यूबुलिन से जुड़े हुए प्रतीत होते हैं। ट्यूबुलिन और एक्टिन की गति की दरें मोटे तौर पर पहले वर्णित तंत्र के लिए पाई गई वृद्धि दर के अनुरूप हैं, जहां अणुओं को सूक्ष्मनलिका या माइक्रोफिलामेंट के सक्रिय अंत में शामिल किया जाता है। इसलिए, यह तंत्र धीमे अक्षीय परिवहन का आधार हो सकता है। धीमी अक्षीय परिवहन की दर भी लगभग अक्षतंतु वृद्धि की दर से मेल खाती है, जो स्पष्ट रूप से दूसरी प्रक्रिया पर साइटोस्केलेटन की संरचना द्वारा लगाए गए प्रतिबंधों को इंगित करती है।
इस खंड को समाप्त करते हुए, इस बात पर जोर दिया जाना चाहिए कि कोशिकाएं किसी भी तरह से स्थिर संरचनाएं नहीं हैं, जैसा कि वे दिखाई देते हैं, उदाहरण के लिए, इलेक्ट्रॉन सूक्ष्म तस्वीरों में। प्लाज्मा झिल्लीऔर विशेष रूप से अंगक निरंतर तीव्र गति और निरंतर पुनर्गठन में हैं;यही एकमात्र कारण है कि वे कार्य करने में सक्षम हैं। इसके अलावा, ये साधारण कक्ष नहीं हैं जिनमें रासायनिक प्रतिक्रियाएँ होती हैं, बल्कि झिल्लियों और तंतुओं के अत्यधिक संगठित समूह,जिसमें प्रतिक्रियाएँ एक सुव्यवस्थित क्रम में होती हैं।
1.4. सेलुलर कार्यों का विनियमन
एक कार्यात्मक इकाई के रूप में व्यक्तिगत कोशिका का रखरखाव मुख्य रूप से नाभिक द्वारा नियंत्रित होता है; ऐसे नियामक तंत्रों का अध्ययन कोशिका जीव विज्ञान और जैव रसायन का विषय है। साथ ही, कोशिकाओं को पर्यावरणीय परिस्थितियों और शरीर की अन्य कोशिकाओं की जरूरतों के अनुसार अपने कार्यों को संशोधित करना होगा, यानी वे कार्यात्मक विनियमन की वस्तुओं के रूप में कार्य करते हैं। नीचे हम संक्षेप में विचार करेंगे कि ये नियामक प्रभाव प्लाज्मा झिल्ली पर कैसे कार्य करते हैं और वे इंट्रासेल्युलर ऑर्गेनेल तक कैसे पहुंचते हैं।
कोशिका झिल्ली पर विनियामक प्रभाव
झिल्ली क्षमता . कई मामलों में, झिल्ली क्षमता को बदलकर सेलुलर कार्यों का विनियमन किया जाता है। क्षमता में स्थानीय परिवर्तन तब संभव होते हैं जब: 1) कोशिका के निकटवर्ती क्षेत्र से या किसी अन्य कोशिका द्वारा उत्पन्न धारा झिल्ली के माध्यम से प्रवाहित होती है; 2) आयनों की बाह्यकोशिकीय सांद्रता बदल जाती है (अक्सर [K + ]बाहर ); 3) झिल्ली आयन चैनल खुलते हैं। झिल्ली क्षमता में परिवर्तन झिल्ली प्रोटीन की संरचना को प्रभावित कर सकता है, जिससे, विशेष रूप से, चैनल खुल या बंद हो सकते हैं। जैसा कि ऊपर बताया गया है, कुछ झिल्ली पंपों की कार्यप्रणाली झिल्ली क्षमता पर निर्भर करती है। तंत्रिका कोशिकाओं को झिल्ली क्षमता में परिवर्तन को सूचना के रूप में समझने के लिए विशेषीकृत किया जाता है जिसे संसाधित और प्रसारित किया जाना चाहिए (अध्याय 2 देखें)।
बाह्यकोशिकीय नियामक पदार्थ . बाह्यकोशिकीय पदार्थों से जुड़ा सबसे महत्वपूर्ण नियामक तंत्र प्लाज्मा झिल्ली पर या कोशिका के अंदर विशिष्ट रिसेप्टर्स के साथ उनकी बातचीत है। इन पदार्थों में सिनैप्टिक ट्रांसमीटर शामिल हैं, जो तंत्रिका कोशिकाओं, स्थानीय एजेंटों और उन पदार्थों के बीच सूचना प्रसारित करते हैं जो रक्त में घूमते हैं और शरीर की सभी कोशिकाओं तक पहुंचते हैं, जैसे हार्मोन और एंटीजन। सिनैप्टिक ट्रांसमीटरसिनैप्स पर तंत्रिका अंत से निकलने वाले छोटे अणु होते हैं;
जब वे पड़ोसी, पोस्टसिनेप्टिक सेल के प्लाज्मा झिल्ली तक पहुंचते हैं, तो वे विद्युत संकेतों या अन्य नियामक तंत्रों को ट्रिगर करते हैं। इस मुद्दे पर अध्याय में विस्तार से चर्चा की गई है। 3.
स्थानीय रासायनिक एजेंट अक्सर विशिष्ट कोशिकाओं द्वारा स्रावित होता है। वे बाह्य कोशिकीय स्थान में स्वतंत्र रूप से फैलते हैं, लेकिन इन पदार्थों के तेजी से नष्ट होने के कारण या तो अनायास या एंजाइमों की कार्रवाई के कारण उनकी क्रिया कोशिकाओं के एक छोटे समूह तक ही सीमित होती है। ऐसे एजेंटों की रिहाई का एक उदाहरण रिहाई है हिस्टामिनक्षति या प्रतिरक्षा प्रतिक्रिया के कारण मस्तूल कोशिकाएं। हिस्टामाइन संवहनी चिकनी मांसपेशियों की कोशिकाओं को आराम देता है, संवहनी एंडोथेलियम की पारगम्यता को बढ़ाता है और संवेदी तंत्रिका अंत को उत्तेजित करता है जो खुजली की अनुभूति में मध्यस्थता करता है। अन्य स्थानीय रासायनिक एजेंट कई अन्य कोशिकाओं द्वारा जारी किए जाते हैं। विशिष्ट स्थानीय एजेंट हैं प्रोस्टाग्लैंडिंस,लगभग 20 फैटी एसिड डेरिवेटिव का एक समूह बनाना। वे व्यापक रूप से वितरित कोशिकाओं से लगातार जारी होते हैं, लेकिन केवल स्थानीय रूप से कार्य करते हैं, क्योंकि वे झिल्ली फॉस्फोलिपेज़ द्वारा जल्दी से नष्ट हो जाते हैं। विभिन्न प्रोस्टाग्लैंडिंस में कार्रवाई का एक व्यापक स्पेक्ट्रम होता है: वे चिकनी मांसपेशियों की कोशिकाओं के संकुचन को ट्रिगर कर सकते हैं, रक्त प्लेटलेट्स (प्लेटलेट्स) के एकत्रीकरण का कारण बन सकते हैं, या अंडाशय में कॉर्पस ल्यूटियम के विकास को रोक सकते हैं।
अन्य स्थानीय एजेंट सेवा देते हैं वृद्धि कारक।सहानुभूति न्यूरॉन्स के लिए सबसे प्रसिद्ध तंत्रिका विकास कारक (एनजीएफ) है, जो विकास के दौरान इन न्यूरॉन्स के विकास और अस्तित्व के लिए आवश्यक है।विवो में या सेल कल्चर में. जाहिरा तौर पर, न्यूरॉन्स के इस वर्ग के लिए लक्ष्य कोशिकाएं एनजीएफ का स्राव करती हैं और इस तरह उचित संरक्षण सुनिश्चित करती हैं। अंग बनाते समय, कोशिकाओं को अक्सर लक्षित कोशिकाओं तक "अपना रास्ता खोजने" की आवश्यकता होती है, जो काफी दूरी पर स्थित हो सकती हैं। तदनुसार, एनजीएफ के समान कई विशिष्ट विकास कारक होने चाहिए।
हार्मोन और एंटीजन रक्त द्वारा सभी कोशिकाओं तक पहुँचाया जाता है। एंटीजन विशिष्ट एंटीबॉडी वाले कोशिकाओं से प्रतिरक्षा प्रतिक्रिया को ट्रिगर करते हैं। हालाँकि, एंटीजन, एक नियम के रूप में, विदेशी पदार्थ हैं जो प्रतिक्रिया करने वाले जीव में उत्पन्न नहीं होते हैं (अधिक विवरण के लिए, अध्याय 18 देखें)। कुछ हार्मोन, जैसे इंसुलिन या थायरोक्सिन, विभिन्न प्रकार की कोशिका को प्रभावित करते हैं, जबकि अन्य, जैसे सेक्स हार्मोन, केवल एक विशिष्ट प्रकार की कोशिका को प्रभावित करते हैं। हार्मोन या तो पेप्टाइड होते हैं, जिनकी क्रिया कोशिका झिल्ली पर एक रिसेप्टर से जुड़ने से शुरू होती है, या स्टेरॉयड और थायरोक्सिन, जो लिपिड झिल्ली के माध्यम से फैलते हैं और इंट्रासेल्युलर रिसेप्टर्स से जुड़ते हैं। स्टेरॉयड हार्मोन परमाणु क्रोमेटिन से बंधते हैं, जिसके परिणामस्वरूप कुछ जीनों का प्रतिलेखन होता है। परिणामी प्रोटीन सेलुलर कार्यों में परिवर्तन का कारण बनता है, जो हार्मोन का विशिष्ट प्रभाव है। अध्याय में हार्मोन की रिहाई और क्रिया से संबंधित मुद्दों पर विस्तार से चर्चा की गई है। 17.
दूसरे दूतों को शामिल करते हुए अंतरकोशिकीय संचार
ऊपर वर्णित नियामक कार्यों में कोशिका झिल्ली पर प्रभाव शामिल है। कोशिका झिल्ली द्वारा प्राप्त जानकारी अक्सर ऑर्गेनेल को प्रतिक्रिया करने के लिए प्रेरित करती है और उन्हें दूसरे दूतों के रूप में जाने जाने वाले विभिन्न पदार्थों द्वारा स्थानांतरित किया जाता है (पहले दूतों के विपरीत, जो बाहरी स्रोतों से कोशिका में आते हैं)। दूसरे मध्यस्थों का अध्ययन तेजी से विकसित हो रहा है, और इस बात की कोई गारंटी नहीं है कि समस्या की समझ का वर्तमान स्तर पर्याप्त रूप से पूर्ण होगा। यहां हम तीन अच्छी तरह से अध्ययन किए गए मध्यस्थों पर बात करेंगे: सीए 2+, सीएमपी और इनोसिटोल ट्राइफॉस्फेट।
कैल्शियम.सबसे सरल अंतःकोशिकीय मध्यस्थ Ca 2+ आयन है। आराम करने वाली कोशिका में इसकी मुक्त सांद्रता बहुत कम है और इसकी मात्रा 10_–8–10–7 mol/l है। यह विशिष्ट झिल्ली चैनलों के माध्यम से कोशिका में प्रवेश कर सकता है जब वे खुली अवस्था में होते हैं, उदाहरण के लिए, जब झिल्ली क्षमता बदलती है (अध्याय 2 देखें)। सीए 2+ एकाग्रता में परिणामी वृद्धि कोशिका में महत्वपूर्ण प्रतिक्रियाओं को ट्रिगर करती है, जैसे मायोफिब्रिल्स का संकुचन, जो मांसपेशियों के संकुचन का आधार है (अध्याय 4 देखें), या तंत्रिका अंत से न्यूरोट्रांसमीटर युक्त पुटिकाओं की रिहाई (अध्याय 3 देखें) . दोनों प्रतिक्रियाओं के लिए लगभग 10 -5 mol/L की Ca 2+ सांद्रता की आवश्यकता होती है। सीए 2+, जिसका नियामक प्रभाव होता है, को एंडोप्लाज्मिक रेटिकुलम जैसे इंट्रासेल्युलर स्टोर्स से भी जारी किया जा सकता है। डिपो से Ca 2+ की रिहाई के लिए अन्य मध्यस्थों की भागीदारी की आवश्यकता होती है (उदाहरण के लिए, चित्र 1.16 देखें)।
चक्रीय एडेनोसिन मोनोफॉस्फेट, सीएमपी। हाल ही में, यह साबित हुआ है कि शरीर के मुख्य ऊर्जा स्रोत, एटीपी का व्युत्पन्न, चक्रीय एडेनोसिन मोनोफॉस्फेट (सीएमपी), एक महत्वपूर्ण दूसरा संदेशवाहक है। प्रतिक्रियाओं की जटिल श्रृंखला चित्र में दिखाई गई है। 1.15, रिसेप्टर से शुरू होता हैआर एस प्लाज्मा झिल्ली की बाहरी सतह पर, जो विभिन्न मध्यस्थों और हार्मोनों के लिए विशिष्ट बंधन स्थल के रूप में काम कर सकता है। एक विशिष्ट "उत्तेजक" अणु से बंधने के बादआर एस इसकी संरचना बदलता है; ये परिवर्तन प्रोटीन को प्रभावित करते हैंजी एस झिल्ली की आंतरिक सतह पर इस तरह से कि इंट्रासेल्युलर ग्वानोसिन ट्राइफॉस्फेट (जीटीपी) द्वारा उत्तरार्द्ध का सक्रियण संभव हो जाता है। सक्रिय प्रोटीनजी एस बदले में, झिल्ली की आंतरिक सतह पर एंजाइम, एडिनाइलेट साइक्लेज़ (एसी) को उत्तेजित करता है, जो एटीपी से सीएमपी के गठन को उत्प्रेरित करता है। पानी में घुलनशील सीएमपी मध्यस्थ है जो प्रभाव को प्रसारित करता है

चावल। 1.15.इंट्रासेल्युलर मैसेंजर सीएमपी (चक्रीय एडेनोसिन मोनोफॉस्फेट) से जुड़ी प्रतिक्रियाओं की एक श्रृंखला। उत्तेजक या निरोधात्मक बाहरी संकेत झिल्ली रिसेप्टर्स आर को सक्रिय करते हैंस या रि . ये रिसेप्टर्स बाइंडिंग प्रक्रिया को नियंत्रित करते हैंजी - इंट्रासेल्युलर जीटीपी (ग्वानोसिन ट्राइफॉस्फेट) वाले प्रोटीन, जिससे इंट्रासेल्युलर एडिनाइलेट साइक्लेज (एसी) उत्तेजित या बाधित होता है। प्रवर्धक एंजाइम एसी एडेनोसिन ट्राइफॉस्फेट (एटीपी) को सीएमपी में परिवर्तित करता है, जिसे बाद में फॉस्फोडिएस्टरेज़ द्वारा एएमपी में तोड़ दिया जाता है। मुक्त सीएमपी कोशिका में फैलता है और एडिनाइलेट किनेज़ (ए-किनेज़) को सक्रिय करता है, इसके उत्प्रेरक सबयूनिट सी को जारी करता है, जो इंट्रासेल्युलर प्रोटीन के फॉस्फोराइलेशन को उत्प्रेरित करता है, यानी। बाह्य कोशिकीय उत्तेजना का अंतिम प्रभाव बनता है। आरेख औषधीय दवाओं और विषाक्त पदार्थों को भी दिखाता है जो कुछ प्रतिक्रियाओं (संशोधित) को ट्रिगर (+) या रोकते हैं (-)
बाह्यकोशिकीय रिसेप्टर उत्तेजनाआर एस कोशिका की आंतरिक संरचनाओं के लिए.
प्रतिक्रियाओं की उत्तेजक श्रृंखला के समानांतरआर एस निरोधात्मक मध्यस्थों और हार्मोनों को संबंधित रिसेप्टर से बांधना संभव हैआर मैं जो फिर से जीटीपी-सक्रिय प्रोटीन के माध्यम से होता हैजी , एसी को रोकता है और इस प्रकार सीएमपी का उत्पादन होता है। कोशिका में फैलते हुए, सीएमपी एडिनाइलेट किनेज़ (ए-किनेज़) के साथ प्रतिक्रिया करता है; यह सबयूनिट सी जारी करता है, जो प्रोटीन पी के फास्फारिलीकरण को उत्प्रेरित करता है।यह फॉस्फोराइलेशन प्रोटीन को सक्रिय रूप में परिवर्तित करता है, और वे अब अपने विशिष्ट नियामक प्रभाव डाल सकते हैं (उदाहरण के लिए, ग्लाइकोजन क्षरण का कारण)। यह जटिल नियामक प्रणाली बेहद कुशल है क्योंकि अंतिम परिणाम कई प्रोटीनों का फॉस्फोराइलेशन होता है, यानी, नियामक संकेत उच्च लाभ के साथ सर्किट से गुजरता है। बाहरी मध्यस्थ जो रिसेप्टर्स से जुड़ते हैंआर एस और आर आई उनमें से प्रत्येक के लिए विशिष्ट, अत्यंत विविध। एड्रेनालाईन, के साथ जुड़ रहा हैआर एस या आर आई लिपिड और ग्लाइकोजन चयापचय के नियमन के साथ-साथ हृदय की मांसपेशियों के संकुचन को बढ़ाने और अन्य प्रतिक्रियाओं में भाग लेता है (अध्याय 19 देखें)। थायराइड उत्तेजक हार्मोन, सक्रिय आर एस , थायरॉयड ग्रंथि द्वारा थायरोक्सिन हार्मोन की रिहाई को उत्तेजित करता है, और प्रोस्टाग्लैंडीन I रक्त प्लेटलेट्स के एकत्रीकरण को रोकता है। एड्रेनालाईन सहित निरोधात्मक प्रभाव, के माध्यम से मध्यस्थआर मैं लिपोलिसिस को धीमा करने में व्यक्त किया जाता है। इस प्रकार, सीएमपी प्रणाली एक बहुक्रियाशील इंट्रासेल्युलर नियामक प्रणाली है,जिसे बाह्यकोशिकीय उत्तेजक और निरोधात्मक सिग्नलिंग पदार्थों द्वारा सटीक रूप से नियंत्रित किया जा सकता है।
इनोसिटोल फॉस्फेट "आईएफ" एच " दूसरे संदेशवाहक, इनोसिटोल फॉस्फेट की इंट्रासेल्युलर प्रणाली की खोज हाल ही में की गई थी (चित्र 1.16)। इस मामले में, कोई निरोधात्मक मार्ग नहीं है, लेकिन सीएमपी प्रणाली में समानता है, जिसमें आर रिसेप्टर की उत्तेजना का प्रभाव झिल्ली की आंतरिक सतह पर जीटीपी-सक्रिय जी प्रोटीन में स्थानांतरित हो जाता है। अगले चरण में, सामान्य झिल्ली लिपिड फॉस्फेटिडिलिनोसिटोल (पीआई), जिसे पहले दो अतिरिक्त फॉस्फेट समूह प्राप्त हुए थे, को पीआई-डिफॉस्फेट (पीआईपी 2) में परिवर्तित कर दिया जाता है, जिसे सक्रिय फॉस्फोडिएस्टरेज़ (पीडीई) द्वारा विभाजित किया जाता है। इनोसिटोल ट्राइफॉस्फेट(आईएफजेड) और लिपिड diacylglycerol(डीएजी)। इनोसिटोल ट्राइफॉस्फेट एक पानी में घुलनशील दूसरा संदेशवाहक है जो साइटोसोल में फैलता है। यह मुख्य रूप से एंडोप्लाज्मिक रेटिकुलम से Ca 2+ जारी करके कार्य करता है। Ca 2+ बदले में एक संदेशवाहक के रूप में कार्य करता है, जैसा कि ऊपर वर्णित है; उदाहरण के लिए, यह Ca 2+-निर्भर फॉस्फोकाइनेज को सक्रिय करता है, जो एंजाइमों को फॉस्फोराइलेट करता है। डीएजी की लिपिड सबयूनिट (चित्र 1.16) प्लाज्मा झिल्ली के लिपिड चरण में फैलते हुए सिग्नल को इसकी आंतरिक सतह पर स्थित सी-किनेज तक ले जाती है, जो सहकारक के रूप में फॉस्फेटिडिलसेरिन की भागीदारी से सक्रिय होता है। फिर सी-किनेज प्रोटीन के फॉस्फोराइलेशन को ट्रिगर करता है, उन्हें सक्रिय रूप में परिवर्तित करता है।
इंट्रासेल्युलर सेकंड मैसेंजर सिस्टम IF 3 को विभिन्न प्रकार के बाहरी मध्यस्थों और हार्मोनों द्वारा भी नियंत्रित किया जा सकता है, जिनमें एसिटाइलकोलाइन, सेरोटोनिन, वैसोप्रेसिन और थायरॉयड-उत्तेजक हार्मोन शामिल हैं; सीएमपी प्रणाली की तरह, यह विभिन्न प्रकार के इंट्रासेल्युलर प्रभावों की विशेषता है। यह संभव है कि यह प्रणाली आंख के दृश्य रिसेप्टर में प्रकाश द्वारा भी सक्रिय होती है और फोटोट्रांसडक्शन में केंद्रीय भूमिका निभाती है (अध्याय 11 देखें)। जीव के व्यक्तिगत विकास में पहली बार, आईजी प्रणाली का रिसेप्टर शुक्राणु द्वारा सक्रिय होता है, जिसके परिणामस्वरूप आईजी अंडे के निषेचन के साथ होने वाली नियामक प्रतिक्रियाओं में भाग लेता है।
सीएमपी और आईएफजेड-डीएजी सिस्टम अत्यधिक प्रभावी हैं जैविक वर्धक.वे

चावल। 1.16.इंट्रासेल्युलर मध्यस्थ आईपीजी (इनोसिटोल ट्राइफॉस्फेट) से जुड़ी प्रतिक्रियाओं की एक श्रृंखला। जैसा कि सीएमपी प्रणाली में होता है, बाह्यकोशिकीय संकेत एक प्रोटीन के माध्यम से मध्यस्थ होता हैजी, जो इस मामले में फॉस्फोडिएस्टरेज़ (पीडीई) को सक्रिय करता है। यह एंजाइम फॉस्फेटिडिलिनोसिन डाइफॉस्फेट (पीआईपी) को तोड़ता है 2 ) IF से पहले प्लाज्मा झिल्ली मेंएच और डायसाइलग्लिसरॉल (डीएजी); अगरएच साइटोप्लाज्म में फैल जाता है। यहां यह Ca के विमोचन का कारण बनता है 2+ एंडोप्लाज्मिक रेटिकुलम से; Ca सांद्रण में वृद्धि 2+ साइटोप्लाज्म में ([Ca 2+ ]मैं ) प्रोटीन काइनेज को सक्रिय करता है, जो फॉस्फोराइलेट करता है और इसलिए एंजाइम को सक्रिय करता है। एक अन्य उत्पाद, डीएजी, झिल्ली में रहता है और प्रोटीन काइनेज सी (फॉस्फेटिडिलसेरिन कॉफ़ेक्टर, पीएस) को सक्रिय करता है। प्रोटीन काइनेज सी उन एंजाइमों को फॉस्फोराइलेट भी करता है जो बाहरी रिसेप्टर उत्तेजना से जुड़ी विशिष्ट क्रियाओं में मध्यस्थता करते हैंआर . IF से जुड़ी प्रतिक्रियाओं की श्रृंखला की शाखाएँएच और डीएजी को क्रमशः आयनोमाइसिन और फोर्बोल एस्टर द्वारा स्वतंत्र रूप से सक्रिय किया जा सकता है (संशोधित)
एक ट्रांसमीटर और एक बाहरी झिल्ली रिसेप्टर के बीच प्रतिक्रिया को विभिन्न इंट्रासेल्युलर प्रोटीन के फॉस्फोराइलेशन में परिवर्तित करें, जो तब विभिन्न कोशिका कार्यों को प्रभावित कर सकता है। समस्या के महत्वपूर्ण पहलुओं में से एक यह है कि, जहां तक आज ज्ञात है, इस प्रकार की केवल ये दो निकट से संबंधित नियामक प्रणालियां हैं, जिनका उपयोग कई बाहरी मध्यस्थों द्वारा विभिन्न इंट्रासेल्युलर प्रक्रियाओं को विनियमित करने के लिए किया जाता है। साथ ही, सीए 2+ समेत ये नियामक प्रणालियां एक-दूसरे के साथ निकटता से बातचीत करती हैं, जो उन्हें सेलुलर कार्यों का अच्छा विनियमन करने की अनुमति देती है।
ट्यूटोरियल और मार्गदर्शिकाएँ
1. अल्बर्ट्स में., ब्रे डी., लुईस जे., रैफ एम., रॉबर्ट्स को., वॉटसन जे.डी.सेल की आणविक जीवविज्ञान, न्यूयॉर्क और लंदन, गारलैंड पब्लिशिंग इंक, 1983।
2. ज़िहाक जी., लॉन्गर एच., ज़िग्लर एच.(सं.). जीवविज्ञान। बर्लिन, हीडलबर्ग, न्यूयॉर्क, स्प्रिंगर, 1983।
3. हिले में. उत्तेजनीय झिल्लियों के आयनिक चैनल। सुंदरलैंड, मास., सिनाउर एसोसिएट., 1984।
4. होप्पे डब्ल्यू., लोहमान डब्ल्यू.. मार्की एच., ज़िग्लर एच.(सं.). बायोफिज़िक्स। बर्लिन, हीडलबर्ग, न्यूयॉर्क, स्प्रिंगर, 1984।
5. जुंगरमैन को., महलर एच.जैव रसायन। बर्लिन, हीडलबर्ग, न्यूयॉर्क, स्प्रिंगर, 1980।
6. कैंडेल ई.आर., श्वार्ट्ज- जे.एच.,(सं.). तंत्रिका विज्ञान के सिद्धांत, न्यूयॉर्क, एम्स्टर्डम, ऑक्सफोर्ड, एल्सेवियर, 1985।
7. शिब्लर टी. एच., श्मिट डब्ल्यू.एनाटॉमिक डेस मेन्सचेन। बर्लिन, हीडलबर्ग, न्यूयॉर्क, टोक्यो, स्प्रिंगर, 1983।
मूल लेख और समीक्षाएँ
8. बेरिज एम.जे.कोशिका के भीतर संचार का आणविक आधार, विज्ञान। आमेर, 253, 124-134 (1985)।
9. बेरिज एम.जे., इरविन आर.एफ.इनोसिटोल ट्राइफॉस्फेट, सेलुलर सिग्नल ट्रांसडक्शन में एक नया दूसरा संदेशवाहक। प्रकृति, 312, 315 321 (1984)।
10. ब्रेत्शर एम.एस.कोशिका झिल्ली के अणु, विज्ञान। आमेर., 253, 124-134 (1985)।
11. डौट जे.जीवित कोशिका एक ऊर्जा-संचारण मशीन के रूप में। मायोकार्डियल चयापचय का एक न्यूनतम मॉडल, बायोकेम। एट बायोफिज़। एक्टा, 895, 41-62 (1987)।
12. हॉजकिन ए.एल., काट्ज़ में. स्क्विड के विशाल अक्षतंतु की विद्युत गतिविधि पर सोडियम आयनों का प्रभाव। जे. फिजियोल. (लंदन), 108, 37-77 (1949)।
13. हॉजकिन ए.एल., कीन्स आर.डी.से विशाल अक्षतंतु में धनायनों का सक्रिय परिवहन एक प्रकार की मछलीऔर लोलिगो,जे. फिजियोल. (लंदन), 128, 28-42 (1955)।
14. लंबा पी.गठनात्मक उपअवस्थाओं के साथ आयनिक चैनल, बायोफिज़। जे., 47, 581-590 (1985)।
15. ओच्स एस., वर्थ पी.एम.सामान्य और पैथोलॉजिकल प्रणालियों में एक्सोप्लाज्मिक परिवहन। इन: फिजियोलॉजी एंड पैथोलॉजी ऑफ एक्सॉन्स, एस.जी. वैक्समैन,ईडी। न्यूयॉर्क, रेवेन प्रेस, 1978।
अन्य प्रस्तुतियों का सारांश"जीव विज्ञान पढ़ाने के तरीके" - स्कूल प्राणीशास्त्र। छात्रों को वैज्ञानिक प्राणीशास्त्रीय डेटा के उपयोग से परिचित कराना। नैतिक शिक्षा। चिकन कॉप का अतिरिक्त आशीर्वाद. तरीकों का चयन. जीवन का चक्र। एक्वेरियम मछली. पोषण। पर्यावरण शिक्षा। जीवन प्रक्रियाओं की भौतिकता. नकारात्मक परिणाम. छात्र ध्यान दें. अनिवार्य प्रपत्र. छोटे जानवरों को देख रहे हैं. जीव विज्ञान के लक्ष्य और उद्देश्य। कहानी।
"जीव विज्ञान पाठों में समस्या-आधारित शिक्षा" - ज्ञान। नई पाठ्यपुस्तकें. समाधान का मार्ग. संकट। सेमिनार. कार्य क्या है? अल्ब्रेक्ट ड्यूरर. जीवविज्ञान पाठों में समस्या-आधारित शिक्षा। गैर-मानक पाठ. समस्या आधारित शिक्षा से क्या तात्पर्य है? जीवन की गुणवत्ता। जीव विज्ञान एक शैक्षणिक विषय के रूप में। सवाल। समस्या समाधान पाठ. विषय में रुचि कम हो गई। समस्या-आधारित प्रयोगशाला कक्षाएं।
"जीवविज्ञान पाठों में आलोचनात्मक सोच" - "महत्वपूर्ण सोच" की तकनीक। "महत्वपूर्ण सोच के विकास" की तकनीक का उपयोग करना। पाठ के लिए तालिका. पढ़ाई के लिए प्रेरणा. पारिस्थितिकी तंत्र. "आलोचनात्मक सोच विकसित करना" का अर्थ। प्रौद्योगिकी के लक्षण. आरकेएम प्रौद्योगिकी। पाठ संरचना. मुख्य दिशाएँ. प्रौद्योगिकी का इतिहास. शैक्षणिक प्रौद्योगिकियाँ। प्रौद्योगिकी के नियम. जीव विज्ञान असाइनमेंट. प्रकाश संश्लेषण. पाठ के विभिन्न चरणों में प्रयुक्त तकनीकें।
"इंटरैक्टिव व्हाइटबोर्ड के साथ जीवविज्ञान पाठ" - इलेक्ट्रॉनिक पाठ्यपुस्तकें। विद्यार्थियों के लिए लाभ. एक इंटरैक्टिव व्हाइटबोर्ड प्रत्येक छात्र तक जानकारी पहुंचाने में मदद करता है। उपदेशात्मक कार्य. जैविक समस्याओं का समाधान. इंटरैक्टिव व्हाइटबोर्ड के साथ काम करने के लाभ। प्रस्तुतियों के साथ कार्य करना. वस्तुओं की तुलना करने पर काम करें. चलती वस्तुएं. स्प्रेडशीट का उपयोग करना. स्कूली बच्चों को पढ़ाने की प्रक्रिया में एक इंटरैक्टिव व्हाइटबोर्ड का उपयोग करना। शिक्षकों के लिए लाभ.
"जीव विज्ञान में सिस्टम-गतिविधि दृष्टिकोण" - संगोष्ठी प्रश्न। गतिविधि विधि. ड्रायोपिथेकस। मानव उत्पत्ति का अलौकिक मार्ग। लाइसोसोम। रासायनिक संगठन. जिम्नोस्पर्म। उपापचय। विश्लेषक। जीव विज्ञान पढ़ाने के लिए सिस्टम-गतिविधि दृष्टिकोण। गुणसूत्र. कोशिकाद्रव्य। अंधापन. कान की लंबाई. किसी व्यक्ति का वर्गीकरण. एक स्तनपायी का कंकाल. मानव विकास के पथ. माइटोसिस। सतही जटिल. समस्याग्रस्त प्रश्न. न्यूक्लियोलस। परमाणु लिफाफा।
"जीव विज्ञान में कंप्यूटर" - छात्रों की सहयोगात्मक गतिविधि। आवृतबीजी परिवार. इंटरैक्टिव प्रशिक्षण. सीखने के मॉडल. मूल्यांकन प्रणाली का एक उदाहरण. निर्देश कार्ड प्रश्न. अनुदेश कार्ड का उदाहरण. शोधकर्ताओं। सूक्ष्म समूह। इंटरैक्टिव शिक्षण प्रौद्योगिकियाँ। हिंडोला। इंटरैक्टिव शिक्षण प्रौद्योगिकियाँ। जीव विज्ञान पाठों में इंटरैक्टिव दृष्टिकोण। कार्य का समूह स्वरूप. "शोधकर्ताओं" के समूहों के लिए कार्य।
