5. Fungsi regulasi. Protein melakukan fungsi zat pemberi sinyal - beberapa hormon, histohormon dan neurotransmiter, merupakan reseptor untuk memberi sinyal zat dari struktur apa pun, dan memastikan transmisi sinyal lebih lanjut dalam rantai sinyal biokimia sel. Contohnya termasuk hormon pertumbuhan somatotropin, hormon insulin, reseptor H- dan M-kolinergik.
6. Fungsi motorik. Dengan bantuan protein, proses kontraksi dan pergerakan biologis lainnya dilakukan. Contohnya termasuk tubulin, aktin, dan miosin.
7. Fungsi cadangan. Tumbuhan mengandung protein cadangan, yang merupakan nutrisi berharga; pada tubuh hewan, protein otot berfungsi sebagai nutrisi cadangan yang dimobilisasi ketika benar-benar diperlukan.
Protein dicirikan oleh adanya beberapa tingkat organisasi struktural.
Struktur primer Protein adalah rangkaian residu asam amino dalam rantai polipeptida. Ikatan peptida adalah ikatan karboksamida antara gugus α-karboksil dari satu asam amino dan gugus α-amino dari asam amino lain.
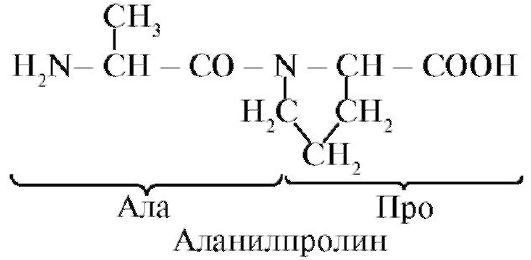
alanylphenylalanylcysteylproline
Ke atas ikatan eptida ada beberapa fitur:
a) ia distabilkan secara resonansi dan oleh karena itu terletak secara praktis pada bidang yang sama - bidang; rotasi di sekitar ikatan C-N memerlukan banyak energi dan sulit;
b) ikatan -CO-NH- mempunyai sifat khusus, lebih kecil dari ikatan biasa, tetapi lebih besar dari ikatan rangkap, yaitu terdapat tautomerisme keto-enol:

c) substituen yang berhubungan dengan ikatan peptida ada di kesurupan-posisi;
d) tulang punggung peptida dikelilingi oleh rantai samping yang sifatnya berbeda, berinteraksi dengan molekul pelarut di sekitarnya, gugus karboksil dan amino bebas terionisasi, membentuk pusat kationik dan anionik dari molekul protein. Tergantung pada rasionya, molekul protein menerima muatan positif atau negatif total, dan juga dicirikan oleh nilai pH tertentu dari medium ketika mencapai titik isoelektrik protein. Radikal membentuk jembatan garam, eter, dan disulfida di dalam molekul protein, dan juga menentukan rentang reaksi yang merupakan karakteristik protein.
Saat ini sepakat untuk mempertimbangkan polimer yang terdiri dari 100 atau lebih residu asam amino sebagai protein, polipeptida - polimer yang terdiri dari 50-100 residu asam amino, peptida dengan berat molekul rendah - polimer yang terdiri dari kurang dari 50 residu asam amino.
Beberapa berat molekul rendah peptida memainkan peran biologis independen. Contoh beberapa peptida ini:
Glutathione - γ-glu-cis-gly - satu salah satu peptida intraseluler yang paling tersebar luas, mengambil bagian dalam proses redoks dalam sel dan transfer asam amino melintasi membran biologis.
Karnosin - β-ala-his - peptida, terkandung dalam otot hewan, menghilangkan produk pemecahan lipid peroksida, mempercepat proses pemecahan karbohidrat di otot dan terlibat dalam metabolisme energi di otot dalam bentuk fosfat.
Vasopresin adalah hormon lobus posterior kelenjar pituitari, yang terlibat dalam pengaturan metabolisme air dalam tubuh:

phalloidin- polipeptida agaric lalat beracun, dalam konsentrasi yang dapat diabaikan menyebabkan kematian tubuh karena pelepasan enzim dan ion kalium dari sel:

Gramisidin - antibiotika, bekerja pada banyak bakteri gram positif, mengubah permeabilitas membran biologis untuk senyawa dengan berat molekul rendah dan menyebabkan kematian sel:

Met-enkephalin - tyr-gly-gly-phen-met - peptida yang disintesis dalam neuron dan mengurangi rasa sakit.
Struktur sekunder protein adalah struktur spasial yang terbentuk sebagai hasil interaksi antar gugus fungsi tulang punggung peptida.
Rantai peptida mengandung banyak gugus ikatan peptida CO dan NH, yang masing-masing berpotensi mampu berpartisipasi dalam pembentukan ikatan hidrogen. Ada dua jenis struktur utama yang memungkinkan hal ini terjadi: heliks α, yang rantainya digulung seperti kabel telepon, dan struktur β terlipat, yang bagian memanjang dari satu atau lebih rantai diletakkan berdampingan. samping. Kedua struktur ini sangat stabil.
α-heliks dicirikan oleh pengepakan rantai polipeptida bengkok yang sangat padat; untuk setiap putaran heliks kanan terdapat 3,6 residu asam amino, yang radikalnya selalu mengarah ke luar dan sedikit ke belakang, yaitu ke awal rantai polipeptida.
Karakteristik utama dari α-helix:
1) α-heliks distabilkan oleh ikatan hidrogen antara atom hidrogen pada nitrogen dari gugus peptida dan oksigen karbonil dari residu yang terletak empat posisi di sepanjang rantai;
2) semua kelompok peptida berpartisipasi dalam pembentukan ikatan hidrogen, ini memastikan stabilitas maksimum α-heliks;
3) semua atom nitrogen dan oksigen dari gugus peptida terlibat dalam pembentukan ikatan hidrogen, yang secara signifikan mengurangi hidrofilisitas daerah heliks dan meningkatkan hidrofobisitasnya;
4) α-heliks terbentuk secara spontan dan merupakan konformasi rantai polipeptida yang paling stabil, sesuai dengan energi bebas minimum;
5) dalam rantai polipeptida asam L-amino, heliks sisi kanan, biasanya ditemukan dalam protein, jauh lebih stabil daripada heliks sisi kiri.
Kemungkinan pembentukan α-helix ditentukan oleh struktur primer protein. Beberapa asam amino mencegah tulang punggung peptida terpelintir. Misalnya, gugus karboksil glutamat dan aspartat yang berdekatan saling tolak menolak, sehingga mencegah pembentukan ikatan hidrogen pada α-heliks. Untuk alasan yang sama, helikalisasi rantai sulit dilakukan di tempat di mana residu lisin dan arginin bermuatan positif terletak berdekatan satu sama lain. Namun, prolin memainkan peran terbesar dalam gangguan α-helix. Pertama, dalam prolin, atom nitrogen merupakan bagian dari cincin kaku, yang mencegah rotasi di sekitar ikatan N-C, dan kedua, prolin tidak membentuk ikatan hidrogen karena tidak adanya hidrogen pada atom nitrogen.
Terpal β adalah struktur berlapis, dibentuk oleh ikatan hidrogen antara fragmen peptida yang tersusun linier. Kedua rantai bisa independen atau termasuk dalam molekul polipeptida yang sama. Jika rantai berorientasi pada arah yang sama, maka struktur β seperti itu disebut paralel. Dalam kasus arah rantai yang berlawanan, yaitu ketika ujung-N dari satu rantai bertepatan dengan ujung-C dari rantai lainnya, struktur β disebut antiparalel. Lembar β antiparalel dengan jembatan hidrogen yang hampir linier lebih disukai secara energetik.


lembaran β paralel lembaran β antiparalel
Berbeda dengan α-heliks jenuh dengan ikatan hidrogen, setiap bagian rantai lembaran terbuka untuk pembentukan ikatan hidrogen tambahan. Radikal samping asam amino berorientasi hampir tegak lurus terhadap bidang lembaran, bergantian ke atas dan ke bawah.
Di daerah-daerah di mana rantai peptida membengkok cukup tajam, seringkali mengandung β-loop. Ini adalah fragmen pendek di mana 4 residu asam amino dibengkokkan 180° dan distabilkan oleh satu jembatan hidrogen antara residu pertama dan keempat. Radikal asam amino besar mengganggu pembentukan β-loop, sehingga paling sering mengandung asam amino terkecil, glisin.
Struktur suprasekunder protein- ini adalah urutan spesifik pergantian struktur sekunder. Domain dipahami sebagai bagian terpisah dari molekul protein yang memiliki tingkat otonomi struktural dan fungsional tertentu. Domain sekarang dianggap sebagai elemen fundamental dari struktur molekul protein, dan hubungan serta sifat susunan heliks α dan lembaran β memberikan lebih banyak pemahaman tentang evolusi molekul protein dan hubungan filogenetik daripada perbandingan struktur primer.
Tugas utama evolusi adalah merancang lebih banyak protein baru. Ada kemungkinan yang sangat kecil untuk secara tidak sengaja mensintesis rangkaian asam amino yang akan memenuhi kondisi pengemasan dan memastikan pemenuhan tugas fungsional. Oleh karena itu, merupakan hal yang umum untuk menemukan protein dengan fungsi berbeda namun strukturnya sangat mirip sehingga tampaknya memiliki nenek moyang yang sama atau berevolusi satu sama lain. Tampaknya evolusi, ketika dihadapkan pada kebutuhan untuk memecahkan masalah tertentu, memilih untuk tidak merancang protein untuk tujuan ini sejak awal, tetapi mengadaptasi struktur yang sudah ada untuk tujuan ini, mengadaptasinya untuk tujuan baru.
Beberapa contoh struktur suprasekunder yang sering diulang:
1) αα’ - protein yang hanya mengandung heliks α (mioglobin, hemoglobin);
2) ββ' - protein yang hanya mengandung struktur β (imunoglobulin, superoksida dismutase);
3) βαβ' - struktur β-barel, setiap lapisan β terletak di dalam laras dan terhubung ke α-heliks yang terletak di permukaan molekul (triose fosfoisomerase, laktat dehidrogenase);
4) "jari seng" - sebuah fragmen protein yang terdiri dari 20 residu asam amino, atom seng terikat pada dua residu sistein dan dua residu histidin, menghasilkan pembentukan "jari" dari sekitar 12 residu asam amino, yang dapat berikatan dengan daerah pengatur molekul DNA;
5) "ritsleting leusin" - protein yang berinteraksi memiliki daerah heliks yang mengandung setidaknya 4 residu leusin, letaknya terpisah 6 asam amino, yaitu berada di permukaan setiap putaran kedua dan dapat membentuk ikatan hidrofobik dengan residu leusin protein lain. Dengan bantuan ritsleting leusin, misalnya, molekul protein histon yang sangat basa dapat dikomplekskan, mengatasi muatan positif.
Struktur tersier protein- ini adalah susunan spasial molekul protein, yang distabilkan oleh ikatan antara radikal samping asam amino.
Jenis ikatan yang menstabilkan struktur tersier suatu protein:

ikatan interaksi disulfida hidrogen hidrofobik elektrostatik ikatan interaksi
Tergantung lipatnya Struktur tersier protein dapat diklasifikasikan menjadi dua jenis utama - fibrilar dan globular.
Protein fibrilar- molekul panjang seperti benang yang tidak larut dalam air, rantai polipeptidanya memanjang sepanjang satu sumbu. Ini terutama protein struktural dan kontraktil. Beberapa contoh protein fibrilar yang paling umum:
1. α- Keratin. Disintesis oleh sel epidermis. Mereka menyumbang hampir seluruh berat kering rambut, bulu, bulu, tanduk, kuku, cakar, duri, sisik, kuku dan cangkang penyu, serta sebagian besar berat lapisan luar kulit. Ini adalah seluruh keluarga protein; komposisi asam aminonya serupa, mengandung banyak residu sistein dan memiliki susunan spasial rantai polipeptida yang sama.
Dalam sel rambut, rantai polipeptida keratin mula-mula diorganisasikan menjadi serat, yang kemudian dibentuk struktur seperti tali atau kabel yang dipilin, yang akhirnya memenuhi seluruh ruang sel. Sel-sel rambut menjadi pipih dan akhirnya mati, dan dinding sel membentuk selubung berbentuk tabung yang disebut kutikula di sekeliling setiap rambut. Dalam α-keratin, rantai polipeptida berbentuk α-heliks, dipelintir satu sama lain menjadi kabel tiga inti dengan pembentukan ikatan silang disulfida.
Residu N-terminal berada pada satu sisi (paralel). Keratin tidak larut dalam air karena dominasi asam amino dalam komposisinya dengan radikal samping non-polar yang bertabrakan dengan fase air. Selama pengeritingan, proses berikut terjadi: pertama, jembatan disulfida dihancurkan dengan reduksi dengan tiol, dan kemudian, ketika rambut diberi bentuk yang diperlukan, dikeringkan dengan pemanasan, sedangkan karena oksidasi dengan oksigen atmosfer, jembatan disulfida baru terbentuk. , yang mempertahankan bentuk gaya rambut.
2. β-Keratin. Ini termasuk serat sutra dan jaring laba-laba. Mereka adalah lapisan lipit β antiparalel dengan dominasi glisin, alanin dan serin dalam komposisi.
3. Kolagen. Protein paling umum pada hewan tingkat tinggi dan protein fibrilar utama jaringan ikat. Kolagen disintesis dalam fibroblas dan kondrosit - sel jaringan ikat khusus, yang kemudian dikeluarkan. Serat kolagen ditemukan di kulit, tendon, tulang rawan, dan tulang. Mereka tidak meregang, lebih kuat dari kawat baja, dan fibril kolagen ditandai dengan lurik melintang.
Saat direbus dalam air, berserat, kolagen yang tidak larut dan tidak dapat dicerna diubah menjadi gelatin melalui hidrolisis beberapa ikatan kovalen. Kolagen mengandung 35% glisin, 11% alanin, 21% prolin, dan 4-hidroksiprolin (asam amino unik untuk kolagen dan elastin). Komposisi ini menentukan relatif rendahnya nilai gizi gelatin sebagai protein pangan. Fibril kolagen terdiri dari subunit polipeptida berulang yang disebut tropokolagen. Subunit ini disusun sepanjang fibril dalam bentuk berkas paralel secara head-to-tail. Perpindahan kepala memberikan ciri khas lurik melintang. Rongga pada struktur ini bila diperlukan dapat berfungsi sebagai tempat pengendapan kristal hidroksiapatit Ca 5 (OH)(PO 4) 3 yang berperan penting dalam mineralisasi tulang.


Subunit tropocollagen terdiri dari dari tiga rantai polipeptida yang digulung erat menjadi tali tiga untai, berbeda dari keratin α dan β. Pada beberapa kolagen, ketiga rantai memiliki urutan asam amino yang sama, sedangkan pada kolagen lain hanya dua rantai yang identik, dan pada rantai ketiga berbeda. Rantai polipeptida tropokolagen membentuk heliks kidal, dengan hanya tiga residu asam amino per putaran karena lengkungan rantai yang disebabkan oleh prolin dan hidroksiprolin. Ketiga rantai tersebut dihubungkan satu sama lain, selain ikatan hidrogen, melalui ikatan tipe kovalen yang terbentuk antara dua residu lisin yang terletak di rantai yang berdekatan:

Seiring bertambahnya usia, semakin banyak ikatan silang yang terbentuk di dalam dan di antara subunit tropokolagen, yang membuat fibril kolagen lebih kaku dan rapuh, dan ini mengubah sifat mekanik tulang rawan dan tendon, membuat tulang lebih rapuh dan mengurangi transparansi kornea.
4. elastin. Pada jaringan elastis kuning terdapat ligamen dan lapisan elastis jaringan ikat pada dinding arteri besar. Subunit utama fibril elastin adalah tropoelastin. Elastin kaya akan glisin dan alanin, banyak mengandung lisin dan sedikit prolin. Bagian spiral elastin meregang ketika tegangan diterapkan, tetapi kembali ke panjang aslinya ketika beban dihilangkan. Residu lisin dari empat rantai berbeda membentuk ikatan kovalen satu sama lain dan memungkinkan elastin meregang secara reversibel ke segala arah.
Protein globular- protein, yang rantai polipeptidanya terlipat menjadi globul kompak, mampu melakukan berbagai macam fungsi.
Struktur tersier protein globular Cara paling mudah untuk mempertimbangkannya adalah dengan menggunakan contoh mioglobin. Mioglobin adalah protein pengikat oksigen yang relatif kecil yang ditemukan di sel otot. Ini menyimpan oksigen terikat dan mendorong transfernya ke mitokondria. Molekul mioglobin mengandung satu rantai polipeptida dan satu hemogroup (heme) - kompleks protoporfirin dengan besi.
Properti dasar mioglobin:
a) molekul mioglobin sangat kompak sehingga hanya 4 molekul air yang dapat masuk ke dalamnya;
b) semua residu asam amino polar, kecuali dua, terletak di permukaan luar molekul, dan semuanya dalam keadaan terhidrasi;
c) sebagian besar residu asam amino hidrofobik terletak di dalam molekul mioglobin dan, dengan demikian, terlindung dari kontak dengan air;
d) masing-masing dari empat residu prolin dalam molekul mioglobin terletak di lokasi pembengkokan rantai polipeptida; residu serin, treonin, dan asparagin terletak di lokasi pembengkokan lainnya, karena asam amino tersebut mencegah pembentukan α-heliks jika mereka terletak bersebelahan;
e) gugus heme datar terletak pada rongga (kantong) dekat permukaan molekul, atom besi mempunyai dua ikatan koordinasi yang diarahkan tegak lurus bidang heme, salah satunya berikatan dengan residu histidin 93, dan yang lainnya berfungsi untuk mengikat sebuah molekul oksigen.
Dimulai dengan struktur tersier protein menjadi mampu melakukan fungsi biologis yang melekat padanya. Dasar dari fungsi protein adalah ketika struktur tersier diletakkan pada permukaan protein, terbentuk area yang dapat mengikat molekul lain, yang disebut ligan. Spesifisitas yang tinggi dari interaksi protein dengan ligan dijamin oleh struktur pusat aktif yang saling melengkapi dengan struktur ligan. Komplementaritas adalah korespondensi spasial dan kimiawi dari permukaan yang berinteraksi. Bagi sebagian besar protein, struktur tersier adalah tingkat lipatan maksimum.
Struktur protein kuarter- karakteristik protein yang terdiri dari dua atau lebih rantai polipeptida yang dihubungkan satu sama lain secara eksklusif melalui ikatan non-kovalen, terutama elektrostatik dan hidrogen. Paling sering, protein mengandung dua atau empat subunit; lebih dari empat subunit biasanya mengandung protein pengatur.
Protein dengan struktur kuaterner, sering disebut oligomer. Ada protein homomer dan heteromer. Protein homomer termasuk protein yang semua subunitnya memiliki struktur yang sama, misalnya enzim katalase terdiri dari empat subunit yang benar-benar identik. Protein heteromerik memiliki subunit yang berbeda; misalnya, enzim RNA polimerase terdiri dari lima subunit berbeda secara struktural yang menjalankan fungsi berbeda.
Interaksi subunit tunggal dengan ligan tertentu menyebabkan perubahan konformasi pada seluruh protein oligomer dan mengubah afinitas subunit lain terhadap ligan; sifat ini mendasari kemampuan protein oligomer untuk regulasi alosterik.
Struktur kuaterner suatu protein dapat diperiksa menggunakan contoh hemoglobin. Berisi empat rantai polipeptida dan empat gugus prostetik heme, di mana atom besinya berbentuk besi Fe 2+. Bagian protein dari molekul - globin - terdiri dari dua rantai α dan dua rantai β yang mengandung hingga 70% heliks α. Masing-masing dari empat rantai memiliki struktur tersier yang khas, dan satu hemogroup dikaitkan dengan setiap rantai. Heme dari rantai yang berbeda letaknya relatif berjauhan dan memiliki sudut kemiringan yang berbeda. Beberapa kontak langsung terbentuk antara dua rantai α dan dua rantai β, sementara banyak kontak tipe α 1 β 1 dan α 2 β 2 yang dibentuk oleh radikal hidrofobik muncul antara rantai α dan β. Antara α 1 β 1 dan α 2 β 2 masih ada saluran.
Berbeda dengan mioglobin hemoglobin dicirikan afinitasnya terhadap oksigen secara signifikan lebih rendah, yang memungkinkannya, pada tekanan parsial rendah oksigen yang ada di jaringan, memberi mereka sebagian besar oksigen yang terikat. Oksigen lebih mudah diikat oleh besi hemoglobin pada nilai pH yang lebih tinggi dan konsentrasi CO 2 yang rendah, karakteristik alveoli paru-paru; pelepasan oksigen dari hemoglobin dipengaruhi oleh nilai pH yang lebih rendah dan konsentrasi CO 2 yang tinggi, karakteristik jaringan.
Selain oksigen, hemoglobin membawa ion hidrogen, yang mengikat residu histidin dalam rantai. Hemoglobin juga membawa karbon dioksida, yang menempel pada gugus amino terminal dari masing-masing empat rantai polipeptida, menghasilkan pembentukan karbaminohemoglobin:

DI DALAM sel darah merah dalam konsentrasi yang cukup tinggi terdapat zat 2,3-difosfogliserat (DPG), kandungannya meningkat seiring dengan naiknya ketinggian dan selama hipoksia, memfasilitasi pelepasan oksigen dari hemoglobin dalam jaringan. DPG terletak di saluran antara α 1 β 1 dan α 2 β 2, berinteraksi dengan kelompok rantai β yang terkontaminasi secara positif. Ketika hemoglobin mengikat oksigen, DPG dipaksa keluar dari rongga. Sel darah merah beberapa burung tidak mengandung DPG, tetapi inositol heksa-fosfat, yang selanjutnya mengurangi afinitas hemoglobin terhadap oksigen.
 2,3-Difosfogliserat (DPG)
2,3-Difosfogliserat (DPG)
HbA - hemoglobin dewasa normal, HbF - hemoglobin janin, memiliki afinitas lebih besar terhadap O 2, HbS - hemoglobin pada anemia sel sabit. Anemia sel sabit adalah penyakit bawaan serius yang disebabkan oleh kelainan genetik hemoglobin. Di dalam darah orang sakit terdapat sejumlah besar sel darah merah tipis berbentuk sabit, yang pertama, mudah pecah, dan kedua, menyumbat kapiler darah.
Pada tingkat molekuler, hemoglobin S berbeda dari hemoglobin A terdapat satu residu asam amino pada posisi 6 rantai β, sedangkan residu asam glutamat terdapat valin. Jadi, hemoglobin S mengandung dua muatan negatif yang lebih sedikit; munculnya valin menyebabkan munculnya kontak hidrofobik yang “lengket” pada permukaan molekul; sebagai akibatnya, selama deoksigenasi, molekul deoksihemoglobin S saling menempel dan membentuk tidak larut, panjangnya tidak normal agregat seperti benang, menyebabkan deformasi sel darah merah.
Tidak ada alasan untuk berpikir bahwa terdapat kendali genetik yang independen atas pembentukan tingkat organisasi struktural protein di atas primer, karena struktur primer menentukan sekunder, tersier, dan kuaterner (jika ada). Konformasi asli suatu protein adalah struktur yang paling stabil secara termodinamika dalam kondisi tertentu.
KULIAH 6
Ada sifat fisik, kimia dan biologi protein.
Sifat fisik protein adalah adanya berat molekul, birefringence (perubahan karakteristik optik larutan protein yang bergerak dibandingkan dengan larutan diam), karena bentuk protein yang tidak bulat, mobilitas dalam medan listrik, karena muatan molekul protein . Selain itu, protein dicirikan oleh sifat optiknya, yaitu kemampuannya memutar bidang polarisasi cahaya, menghamburkan sinar cahaya karena ukuran partikel protein yang besar, dan menyerap sinar ultraviolet.
Salah satu ciri sifat fisik protein adalah kemampuan untuk menyerap di permukaan, dan terkadang untuk menangkap molekul, senyawa organik dengan berat molekul rendah, dan ion di dalamnya.
Sifat kimia protein berbeda-beda keanekaragaman yang luar biasa, karena protein dicirikan oleh semua reaksi radikal asam amino dan dicirikan oleh reaksi hidrolisis ikatan peptida.
Memiliki sejumlah besar gugus asam dan basa, protein menunjukkan sifat amfoter. Berbeda dengan asam amino bebas, sifat asam-basa protein tidak ditentukan oleh gugus α-amino dan α-karboksi yang terlibat dalam pembentukan ikatan peptida, tetapi oleh radikal bermuatan dari residu asam amino. Sifat utama protein ditentukan oleh residu arginin, lisin dan histidin. Sifat asam disebabkan oleh residu asam aspartat dan glutamat.
Kurva titrasi protein sudah cukup sulit untuk ditafsirkan, karena protein apa pun memiliki terlalu banyak gugus yang dapat dititrasi, terdapat interaksi elektrostatik antara gugus protein yang terionisasi, dan pK setiap gugus yang dapat dititrasi dipengaruhi oleh residu hidrofobik di dekatnya dan ikatan hidrogen. Penerapan praktis terbesarnya adalah titik isoelektrik suatu protein - nilai pH di mana muatan total protein adalah nol. Pada titik isoelektrik, protein bersifat inert maksimal, tidak bergerak dalam medan listrik, dan mempunyai cangkang hidrasi tertipis.
Protein menunjukkan sifat buffering, namun kapasitas buffernya tidak signifikan. Pengecualiannya adalah protein yang mengandung residu histidin dalam jumlah besar. Misalnya hemoglobin yang terkandung dalam eritrosit, karena kandungan residu histidin yang sangat tinggi, memiliki kapasitas buffer yang signifikan pada pH sekitar 7, yang sangat penting untuk peran eritrosit dalam pengangkutan oksigen dan karbon dioksida di darah.
Protein dicirikan oleh kelarutannya dalam air, dan dari sudut pandang fisik mereka membentuk solusi molekuler yang sebenarnya. Namun, larutan protein dicirikan oleh beberapa sifat koloid: efek Tendahl (fenomena hamburan cahaya), ketidakmampuan melewati membran semipermeabel, viskositas tinggi, dan pembentukan gel.
Kelarutan protein sangat bergantung pada konsentrasi garam, yaitu pada kekuatan ionik larutan. Dalam air suling, protein seringkali sulit larut, namun kelarutannya meningkat seiring dengan meningkatnya kekuatan ionik. Pada saat yang sama, semakin banyak ion anorganik terhidrasi yang berikatan dengan permukaan protein dan dengan demikian tingkat agregasinya menurun. Pada kekuatan ionik yang tinggi, ion garam menghilangkan lapisan hidrasi dari molekul protein, yang menyebabkan agregasi dan pengendapan protein (fenomena salting out). Dengan menggunakan perbedaan kelarutan, campuran protein dapat dipisahkan dengan menggunakan garam biasa.
Diantara sifat biologis protein terutama mencakup aktivitas katalitiknya. Sifat biologis penting lainnya dari protein adalah aktivitas hormonalnya, yaitu kemampuannya untuk mempengaruhi seluruh kelompok reaksi dalam tubuh. Beberapa protein memiliki sifat toksik, aktivitas patogen, fungsi pelindung dan reseptor, dan bertanggung jawab atas fenomena adhesi sel.
Sifat biologis unik lainnya dari protein- denaturasi. Protein dalam keadaan alaminya disebut asli. Denaturasi adalah penghancuran struktur spasial protein di bawah pengaruh agen denaturasi. Struktur utama protein tidak rusak selama denaturasi, namun aktivitas biologisnya hilang, begitu pula kelarutan, mobilitas elektroforesis, dan beberapa reaksi lainnya. Ketika didenaturasi, radikal asam amino yang membentuk pusat aktif protein secara spasial berjauhan satu sama lain, yaitu pusat pengikatan spesifik protein dengan ligan hancur. Radikal hidrofobik, biasanya terletak di inti hidrofobik protein globular, ketika didenaturasi berakhir di permukaan molekul, sehingga menciptakan kondisi agregasi protein yang mengendap.
Reagen dan kondisi yang menyebabkan denaturasi protein:
Suhu di atas 60 o C - rusaknya ikatan lemah pada protein,
Asam dan basa - perubahan ionisasi gugus ionogenik, pemutusan ikatan ionik dan hidrogen,
Urea - penghancuran ikatan hidrogen intramolekul sebagai akibat dari pembentukan ikatan hidrogen dengan urea,
Alkohol, fenol, kloramin - penghancuran ikatan hidrofobik dan hidrogen,
Garam logam berat - pembentukan garam protein yang tidak larut dengan ion logam berat.
Ketika zat pendenaturasi dihilangkan, renativasi dapat dilakukan, karena rantai peptida cenderung mengadopsi konformasi dengan energi bebas terendah dalam larutan.
Dalam kondisi seluler, protein bisa mengalami denaturasi secara spontan, meskipun lajunya lebih rendah dibandingkan pada suhu tinggi. Renativasi protein secara spontan di dalam sel sulit dilakukan, karena konsentrasinya yang tinggi ada kemungkinan besar agregasi molekul yang terdenaturasi sebagian.
Sel mengandung protein- pendamping molekuler yang memiliki kemampuan untuk mengikat protein terdenaturasi sebagian yang berada dalam keadaan tidak stabil, rentan terhadap agregasi, dan mengembalikan konformasi aslinya. Awalnya, protein ini ditemukan sebagai protein kejutan panas, karena sintesisnya meningkat ketika sel terkena stres, misalnya ketika suhu meningkat. Pendamping diklasifikasikan menurut massa subunitnya: hsp-60, hsp-70 dan hsp-90. Setiap kelas mencakup keluarga protein terkait.
Pendamping molekuler ( hsp-70) kelas protein yang sangat terkonservasi yang ditemukan di semua bagian sel: sitoplasma, nukleus, retikulum endoplasma, mitokondria. Di ujung C rantai polipeptida tunggal, hsp-70 memiliki wilayah alur yang mampu berinteraksi dengan peptida sepanjang 7-9 residu asam amino, diperkaya dengan radikal hidrofobik. Daerah seperti itu pada protein globular terdapat kira-kira setiap 16 asam amino. Hsp-70 mampu melindungi protein dari inaktivasi suhu dan mengembalikan konformasi serta aktivitas protein yang terdenaturasi sebagian.
Pendamping-60 (hsp-60) berpartisipasi dalam pembentukan struktur tersier protein. Hsp-60 berfungsi sebagai protein oligomer yang terdiri dari 14 subunit. Hsp-60 membentuk dua cincin, masing-masing cincin terdiri dari 7 subunit yang terhubung satu sama lain.
Setiap subunit terdiri dari tiga domain:
Domain apikal memiliki sejumlah residu asam amino hidrofobik yang menghadap ke dalam rongga yang dibentuk oleh subunit;
Domain ekuator memiliki aktivitas ATPase dan diperlukan untuk pelepasan protein dari kompleks pendamping;
Domain perantara menghubungkan domain apikal dan khatulistiwa.
Protein yang memiliki fragmen pada permukaannya, diperkaya dengan asam amino hidrofobik, memasuki rongga kompleks pendamping. Dalam lingkungan spesifik rongga ini, dalam kondisi isolasi dari molekul lain dari sitosol sel, pemilihan konformasi protein yang mungkin terjadi sampai ditemukan konformasi yang lebih menguntungkan secara energi. Pembentukan konformasi asli yang bergantung pada pendamping dikaitkan dengan pengeluaran sejumlah besar energi, yang sumbernya adalah ATP.
MODUL 1 STRUKTUR, SIFAT DAN FUNGSI PROTEIN
MODUL 1 STRUKTUR, SIFAT DAN FUNGSI PROTEIN
Struktur modul | Tema |
Unit modular 1 | 1.1. Organisasi struktural protein. Tahapan pembentukan konformasi protein asli 1.2. Dasar-dasar fungsi protein. Obat sebagai ligan mempengaruhi fungsi protein 1.3. Denaturasi protein dan kemungkinan renativasi spontannya |
Unit modular 2 | 1.4. Ciri-ciri struktur dan fungsi protein oligomer pada contoh hemoglobin 1.5. Pemeliharaan konformasi protein asli dalam kondisi seluler 1.6. Berbagai protein. Keluarga protein menggunakan contoh imunoglobulin 1.7. Sifat fisikokimia protein dan metode pemisahannya |
Unit modular 1 ORGANISASI STRUKTUR PROTEIN MONOMER DAN DASAR FUNGSINYA
Tujuan pembelajaran Mampu:
1. Menggunakan pengetahuan tentang ciri-ciri struktural protein dan ketergantungan fungsi protein pada strukturnya untuk memahami mekanisme perkembangan proteinopati herediter dan didapat.
2. Menjelaskan mekanisme kerja terapeutik beberapa obat sebagai ligan yang berinteraksi dengan protein dan mengubah aktivitasnya.
3. Menggunakan pengetahuan tentang struktur dan labilitas konformasi protein untuk memahami ketidakstabilan struktural dan fungsionalnya serta kecenderungan denaturasinya dalam kondisi yang berubah.
4. Menjelaskan kegunaan bahan denaturasi sebagai alat untuk mensterilkan bahan dan instrumen kesehatan, serta sebagai antiseptik.
Tahu:
1. Tingkat organisasi struktural protein.
2. Pentingnya struktur primer protein, yang menentukan keanekaragaman struktural dan fungsionalnya.
3. Mekanisme pembentukan pusat aktif pada protein dan interaksi spesifiknya dengan ligan yang mendasari berfungsinya protein.
4. Contoh pengaruh ligan eksogen (obat, toksin, racun) terhadap konformasi dan aktivitas fungsional protein.
5. Penyebab dan akibat denaturasi protein, faktor penyebab denaturasi.
6. Contoh penggunaan faktor denaturasi dalam pengobatan sebagai antiseptik dan alat sterilisasi alat kesehatan.
TOPIK 1.1. ORGANISASI STRUKTUR PROTEIN. TAHAP PEMBENTUKAN ASLI
KONFORMASI PROTEIN
Protein adalah molekul polimer yang monomernya hanya 20 asam α-amino. Himpunan dan urutan kombinasi asam amino dalam suatu protein ditentukan oleh struktur gen dalam DNA individu. Setiap protein, sesuai dengan struktur spesifiknya, menjalankan fungsi karakteristiknya. Kumpulan protein suatu organisme menentukan karakteristik fenotipiknya, serta adanya penyakit keturunan atau kecenderungan perkembangannya.
1. Asam amino penyusun protein. Ikatan peptida. Protein adalah polimer yang dibangun dari monomer - 20 asam α-amino, rumus umumnya adalah
Asam amino berbeda dalam struktur, ukuran, dan sifat fisikokimia radikal yang terikat pada atom karbon α. Gugus fungsi asam amino menentukan karakteristik sifat-sifat asam α-amino yang berbeda. Radikal yang terdapat pada asam α-amino dapat dibagi menjadi beberapa kelompok:
prolin, Berbeda dengan 19 monomer protein lainnya, ini bukanlah asam amino, melainkan asam imino; radikal dalam prolin dikaitkan dengan atom karbon α dan gugus imino
 Asam amino bervariasi dalam kelarutan dalam air. Hal ini disebabkan kemampuan radikal berinteraksi dengan air (hidrat).
Asam amino bervariasi dalam kelarutan dalam air. Hal ini disebabkan kemampuan radikal berinteraksi dengan air (hidrat).
KE hidrofilik termasuk radikal yang mengandung gugus fungsi tak bermuatan anionik, kationik, dan polar.
KE hidrofobik termasuk radikal yang mengandung gugus metil, rantai atau cincin alifatik.
2. Ikatan peptida menghubungkan asam amino membentuk peptida. Selama sintesis peptida, gugus α-karboksil dari satu asam amino berinteraksi dengan gugus α-amino dari asam amino lain untuk membentuk ikatan peptida:
Protein adalah polipeptida, mis. polimer linier asam α-amino dihubungkan oleh ikatan peptida (Gbr. 1.1.)
 Beras. 1.1. Istilah yang digunakan untuk menggambarkan struktur peptida
Beras. 1.1. Istilah yang digunakan untuk menggambarkan struktur peptida
Monomer asam amino yang menyusun polipeptida disebut residu asam amino. Rantai kelompok yang berulang - NH-CH-CO- formulir tulang punggung peptida. Residu asam amino yang mempunyai gugus α-amino bebas disebut terminal-N, dan yang mempunyai gugus α-karboksil bebas disebut terminal-C. Peptida ditulis dan dibaca dari N-terminus ke C-terminus.
Ikatan peptida yang dibentuk oleh gugus imino prolin berbeda dari ikatan peptida lainnya: atom nitrogen pada gugus peptida kekurangan hidrogen,
sebaliknya, terdapat ikatan dengan radikal, akibatnya salah satu sisi cincin termasuk dalam tulang punggung peptida:
 Peptida berbeda dalam komposisi asam amino, jumlah asam amino dan urutan ikatan asam amino, misalnya Ser-Ala-Glu-Gis dan His-Glu-Ala-Ser adalah dua peptida yang berbeda.
Peptida berbeda dalam komposisi asam amino, jumlah asam amino dan urutan ikatan asam amino, misalnya Ser-Ala-Glu-Gis dan His-Glu-Ala-Ser adalah dua peptida yang berbeda.
Ikatan peptida sangat kuat, dan hidrolisis kimia non-enzimatiknya memerlukan kondisi yang keras: protein yang dianalisis dihidrolisis dalam asam klorida pekat pada suhu sekitar 110° selama 24 jam. Dalam sel hidup, ikatan peptida dapat diputus oleh enzim proteolitik, ditelepon protease atau hidrolase peptida.
3. Struktur primer protein. Residu asam amino dalam rantai peptida protein yang berbeda tidak bergantian secara acak, tetapi tersusun dalam urutan tertentu. Urutan linier atau urutan pergantian residu asam amino dalam rantai polipeptida disebut struktur utama protein.
Struktur utama setiap protein individu dikodekan dalam molekul DNA (di wilayah yang disebut gen) dan diwujudkan selama transkripsi (menyalin informasi ke mRNA) dan translasi (sintesis struktur primer protein). Akibatnya, struktur utama protein seseorang adalah informasi yang diturunkan secara turun-temurun dari orang tua ke anak-anak, yang menentukan ciri-ciri struktural protein suatu organisme tertentu, yang menjadi sandaran fungsi protein yang ada (Gbr. 1.2.).
 Beras. 1.2. Hubungan antara genotipe dan konformasi protein yang disintesis dalam tubuh individu
Beras. 1.2. Hubungan antara genotipe dan konformasi protein yang disintesis dalam tubuh individu
Masing-masing dari sekitar 100.000 protein individu dalam tubuh manusia miliki unik struktur primer. Molekul dari jenis protein yang sama (misalnya albumin) memiliki pergantian residu asam amino yang sama, yang membedakan albumin dari protein individu lainnya.
Urutan residu asam amino dalam rantai peptida dapat dianggap sebagai bentuk pencatatan informasi. Informasi ini menentukan susunan spasial rantai peptida linier menjadi struktur tiga dimensi yang lebih kompak yang disebut konformasi tupai. Proses pembentukan konformasi protein yang aktif secara fungsional disebut Melipat
4. Konformasi protein. Rotasi bebas pada tulang punggung peptida dimungkinkan antara atom nitrogen dari gugus peptida dan atom karbon α yang berdekatan, serta antara atom karbon α dan karbon dari gugus karbonil. Karena interaksi gugus fungsi residu asam amino, struktur primer protein dapat memperoleh struktur spasial yang lebih kompleks. Dalam protein globular, ada dua tingkat utama pelipatan konformasi rantai peptida: sekunder Dan struktur tersier.
Struktur sekunder protein adalah struktur spasial yang terbentuk sebagai hasil pembentukan ikatan hidrogen antara gugus fungsi -C=O dan -NH- dari tulang punggung peptida. Dalam hal ini, rantai peptida dapat memperoleh dua jenis struktur teratur: α-heliks Dan struktur β.
DI DALAM α-heliks ikatan hidrogen terbentuk antara atom oksigen dari gugus karbonil dan hidrogen dari nitrogen amino dari asam amino ke-4 darinya; rantai samping residu asam amino
terletak di sepanjang pinggiran spiral, tanpa berpartisipasi dalam pembentukan struktur sekunder (Gbr. 1.3.).
Radikal curah, atau radikal yang membawa muatan yang sama, mencegah pembentukan α-helix. Residu prolin, yang memiliki struktur cincin, memutus α-heliks, karena kurangnya hidrogen pada atom nitrogen dalam rantai peptida, tidak mungkin terbentuk ikatan hidrogen. Ikatan antara nitrogen dan atom karbon α merupakan bagian dari cincin prolin, sehingga tulang punggung peptida menjadi bengkok pada titik ini.
β-Struktur terbentuk antara daerah linier tulang punggung peptida dari satu rantai polipeptida, sehingga membentuk struktur terlipat. Rantai polipeptida atau bagiannya dapat terbentuk paralel atau struktur β antiparalel. Dalam kasus pertama, ujung N dan C dari rantai peptida yang berinteraksi bertepatan, dan pada kasus kedua, keduanya memiliki arah yang berlawanan (Gbr. 1.4).
Beras. 1.3. Struktur sekunder protein - α-helix
 Beras. 1.4. Struktur lembaran β paralel dan antiparalel
Beras. 1.4. Struktur lembaran β paralel dan antiparalel
Struktur β ditunjukkan dengan panah lebar: A - Struktur β Antiparalel. B - Struktur lembaran β paralel
Pada beberapa protein, struktur β dapat terbentuk karena pembentukan ikatan hidrogen antara atom-atom tulang punggung peptida dari rantai polipeptida yang berbeda.
Juga ditemukan dalam protein daerah dengan sekunder tidak beraturan struktur, yang meliputi tikungan, putaran, dan putaran tulang punggung polipeptida. Mereka sering terletak di tempat di mana arah rantai peptida berubah, misalnya, ketika struktur lembaran β paralel terbentuk.
Berdasarkan keberadaan heliks α dan struktur β, protein globular dapat dibagi menjadi empat kategori.
Beras. 1.5. Struktur sekunder mioglobin (A) dan rantai β hemoglobin (B), mengandung delapan heliks α

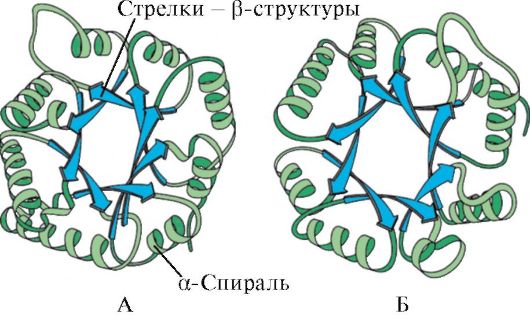 Beras. 1.6. Struktur sekunder domain triosefosfat isomerase dan piruvat kinase
Beras. 1.6. Struktur sekunder domain triosefosfat isomerase dan piruvat kinase
 Beras. 1.7. Struktur sekunder dari domain konstan imunoglobulin (A) dan enzim superoksida dismutase (B)
Beras. 1.7. Struktur sekunder dari domain konstan imunoglobulin (A) dan enzim superoksida dismutase (B)
DI DALAM kategori keempat termasuk protein yang mengandung sejumlah kecil struktur sekunder biasa. Protein ini termasuk protein kecil yang kaya sistein atau metalloprotein.
Struktur tersier protein- jenis konformasi yang terbentuk karena interaksi antara radikal asam amino, yang dapat ditempatkan pada jarak yang cukup jauh satu sama lain dalam rantai peptida. Kebanyakan protein membentuk struktur spasial menyerupai globule (protein globular).
Karena radikal asam amino hidrofobik cenderung bergabung melalui apa yang disebut interaksi hidrofobik dan gaya van der Waals antarmolekul, inti hidrofobik padat terbentuk di dalam globul protein. Radikal hidrofilik terionisasi dan non-terionisasi terutama terletak di permukaan protein dan menentukan kelarutannya dalam air.
 Beras. 1.8. Jenis ikatan yang timbul antara radikal asam amino selama pembentukan struktur tersier suatu protein
Beras. 1.8. Jenis ikatan yang timbul antara radikal asam amino selama pembentukan struktur tersier suatu protein
1 - ikatan ionik- terjadi antara gugus fungsi yang bermuatan positif dan negatif;
2 - ikatan hidrogen- terjadi antara gugus hidrofilik tak bermuatan dan gugus hidrofilik lainnya;
3 - interaksi hidrofobik- timbul di antara radikal hidrofobik;
4 - ikatan disulfida- terbentuk karena oksidasi gugus SH residu sistein dan interaksinya satu sama lain
Residu asam amino hidrofilik yang terdapat di dalam inti hidrofobik dapat berinteraksi satu sama lain menggunakan ionik Dan ikatan hidrogen(Gbr. 1.8).
Ikatan ionik dan hidrogen, serta interaksi hidrofobik, lemah: energinya tidak jauh lebih tinggi daripada energi gerak termal molekul pada suhu kamar. Konformasi protein dipertahankan dengan pembentukan banyak ikatan lemah. Karena atom-atom yang membentuk protein terus bergerak, beberapa ikatan lemah dapat diputus dan membentuk ikatan lain, yang menyebabkan sedikit pergerakan pada masing-masing bagian rantai polipeptida. Sifat protein untuk mengubah konformasi sebagai akibat putusnya sebagian dan pembentukan ikatan lemah lainnya disebut labilitas konformasi.
Tubuh manusia mempunyai sistem yang mendukung homeostatis- keteguhan lingkungan internal dalam batas-batas tertentu yang dapat diterima untuk kesehatan tubuh. Dalam kondisi homeostatis, perubahan kecil pada konformasi tidak mengganggu keseluruhan struktur dan fungsi protein. Konformasi protein yang aktif secara fungsional disebut konformasi asli. Perubahan lingkungan internal (misalnya konsentrasi glukosa, ion Ca, proton, dll) menyebabkan perubahan konformasi dan terganggunya fungsi protein.
Struktur tersier beberapa protein menjadi stabil ikatan disulfida, terbentuk karena interaksi gugus -SH dari dua residu
 Beras. 1.9. Pembentukan ikatan disulfida dalam molekul protein
Beras. 1.9. Pembentukan ikatan disulfida dalam molekul protein
sistein (Gbr. 1.9). Kebanyakan protein intraseluler tidak memiliki ikatan kovalen disulfida pada struktur tersiernya. Kehadiran mereka merupakan karakteristik protein yang disekresikan oleh sel, yang menjamin stabilitasnya yang lebih besar dalam kondisi ekstraseluler. Dengan demikian, ikatan disulfida terdapat dalam molekul insulin dan imunoglobulin.
Insulin- hormon protein yang disintesis di sel pankreas dan disekresikan ke dalam darah sebagai respons terhadap peningkatan konsentrasi glukosa dalam darah. Dalam struktur insulin, terdapat dua ikatan disulfida yang menghubungkan rantai polipeptida A dan B, dan satu ikatan disulfida dalam rantai A (Gbr. 1.10).
 Beras. 1.10. Ikatan disulfida dalam struktur insulin
Beras. 1.10. Ikatan disulfida dalam struktur insulin
5. Struktur protein supersekunder. Pada protein dengan struktur dan fungsi primer yang berbeda, kadang-kadang terdeteksi kombinasi serupa dan posisi relatif struktur sekunder, yang disebut struktur supersekunder. Ia menempati posisi perantara antara struktur sekunder dan tersier, karena merupakan kombinasi spesifik elemen struktur sekunder dalam pembentukan struktur tersier protein. Struktur supersekunder memiliki nama spesifik, seperti “α-helix-turn-a-helix”, “leucine ritsleting”, “jari seng”, dll. Struktur supersekunder tersebut merupakan karakteristik protein pengikat DNA.
"Ritsleting leusin." Jenis struktur supersekunder ini digunakan untuk menggabungkan dua protein menjadi satu. Pada permukaan protein yang berinteraksi terdapat daerah heliks α yang mengandung setidaknya empat residu leusin. Residu leusin dalam α-heliks terletak terpisah enam asam amino. Karena setiap putaran α-heliks mengandung 3,6 residu asam amino, radikal leusin terletak di permukaan setiap putaran kedua. Residu leusin dari α-heliks suatu protein dapat berinteraksi dengan residu leusin dari protein lain (interaksi hidrofobik), menghubungkan keduanya (Gbr. 1.11.). Banyak protein pengikat DNA berfungsi dalam kompleks oligomer di mana masing-masing subunit dihubungkan satu sama lain melalui “ritsleting leusin”.
 Beras. 1.11. "Ritsleting leusin" antara daerah heliks α dari dua protein
Beras. 1.11. "Ritsleting leusin" antara daerah heliks α dari dua protein
Contoh protein tersebut adalah histon. sejarah- protein inti, yang mengandung sejumlah besar asam amino bermuatan positif - arginin dan lisin (hingga 80%). Molekul histon digabungkan menjadi kompleks oligomer yang mengandung delapan monomer menggunakan “ritsleting leusin”, meskipun molekul-molekul ini memiliki muatan homonim yang signifikan.
"jari seng"- varian struktur supersekunder, karakteristik protein pengikat DNA, berbentuk fragmen memanjang pada permukaan protein dan mengandung sekitar 20 residu asam amino (Gbr. 1.12). Bentuk “jari terentang” didukung oleh atom seng yang terikat pada empat radikal asam amino - dua residu sistein dan dua residu histidin. Dalam beberapa kasus, selain residu histidin, terdapat residu sistein. Dua residu sistein yang letaknya berdekatan dipisahkan dari dua residu Gisili lainnya melalui rangkaian Cys yang terdiri dari sekitar 12 residu asam amino. Wilayah protein ini membentuk α-heliks, yang radikalnya dapat secara spesifik berikatan dengan wilayah pengatur alur utama DNA. Kekhususan pengikatan individu
 Beras. 1.12. Struktur utama wilayah protein pengikat DNA yang membentuk struktur “jari seng” (huruf menunjukkan asam amino yang menyusun struktur ini)
Beras. 1.12. Struktur utama wilayah protein pengikat DNA yang membentuk struktur “jari seng” (huruf menunjukkan asam amino yang menyusun struktur ini)
Protein pengikat DNA pengatur bergantung pada urutan residu asam amino yang terletak di daerah jari seng. Struktur tersebut mengandung, khususnya, reseptor hormon steroid yang terlibat dalam regulasi transkripsi (membaca informasi dari DNA ke RNA).
TOPIK 1.2. DASAR FUNGSI PROTEIN. OBAT SEBAGAI LIGAND YANG MEMPENGARUHI FUNGSI PROTEIN
1. Pusat aktif protein dan interaksinya dengan ligan. Selama pembentukan struktur tersier, suatu daerah terbentuk pada permukaan protein yang aktif secara fungsional, biasanya dalam suatu ceruk, dibentuk oleh radikal asam amino yang berjauhan satu sama lain dalam struktur primer. Wilayah ini, yang memiliki struktur unik untuk protein tertentu dan mampu berinteraksi secara spesifik dengan molekul tertentu atau kelompok molekul serupa, disebut situs pengikatan protein-ligan atau situs aktif. Ligan adalah molekul yang berinteraksi dengan protein.
Spesifisitas tinggi Interaksi protein dengan ligan dijamin oleh struktur pusat aktif yang saling melengkapi dengan struktur ligan.
Komplementaritas- ini adalah korespondensi spasial dan kimia dari permukaan yang berinteraksi. Pusat aktif tidak hanya harus sesuai secara spasial dengan ligan yang termasuk di dalamnya, tetapi juga ikatan (interaksi ionik, hidrogen, dan hidrofobik) harus terbentuk antara gugus fungsi radikal yang termasuk dalam pusat aktif dan ligan yang menahan ligan. di pusat aktif (Gbr. 1.13 ).
 Beras. 1.13. Interaksi komplementer protein dengan ligan
Beras. 1.13. Interaksi komplementer protein dengan ligan
Beberapa ligan, ketika melekat pada pusat aktif suatu protein, memainkan peran tambahan dalam fungsi protein. Ligan seperti itu disebut kofaktor, dan protein yang mengandung bagian non-protein disebut protein kompleks(berlawanan dengan protein sederhana, hanya terdiri dari bagian protein). Bagian bukan protein yang terikat kuat pada protein disebut kelompok prostetik. Misalnya, mioglobin, hemoglobin, dan sitokrom mengandung gugus prostetik, heme, yang mengandung ion besi, terikat kuat pada pusat aktif. Protein kompleks yang mengandung heme disebut hemoprotein.
Ketika ligan tertentu melekat pada protein, fungsi protein ini terwujud. Jadi, albumin, protein terpenting dalam plasma darah, menunjukkan fungsi transpornya dengan menempelkan ligan hidrofobik, seperti asam lemak, bilirubin, obat-obatan tertentu, dll. ke pusat aktif (Gbr. 1.14)
Ligan yang berinteraksi dengan struktur tiga dimensi rantai peptida tidak hanya berupa molekul organik dan anorganik dengan berat molekul rendah, tetapi juga makromolekul:
DNA (contoh protein pengikat DNA dibahas di atas);
Polisakarida;
 Beras. 1.14. Hubungan antara genotipe dan fenotipe
Beras. 1.14. Hubungan antara genotipe dan fenotipe
Struktur primer unik protein manusia, yang dikodekan dalam molekul DNA, diwujudkan dalam sel dalam bentuk konformasi unik, struktur pusat aktif, dan fungsi protein.
Dalam kasus ini, protein mengenali wilayah tertentu dari ligan yang sepadan dan saling melengkapi dengan tempat pengikatan. Jadi, pada permukaan hepatosit terdapat protein reseptor hormon insulin, yang juga memiliki struktur protein. Interaksi insulin dengan reseptor menyebabkan perubahan konformasi dan aktivasi sistem sinyal, yang menyebabkan penyimpanan nutrisi di hepatosit setelah makan.
Dengan demikian, Fungsi protein didasarkan pada interaksi spesifik pusat aktif protein dengan ligan.
2. Struktur domain dan perannya dalam fungsi protein. Rantai polipeptida panjang dari protein globular sering kali terlipat menjadi beberapa daerah yang kompak dan relatif independen. Mereka memiliki struktur tersier independen, mengingatkan pada struktur protein globular, dan disebut domain. Karena struktur domain protein, struktur tersiernya lebih mudah dibentuk.
Dalam protein domain, situs pengikatan ligan sering kali terletak di antara domain. Jadi, trypsin adalah enzim proteolitik yang diproduksi oleh bagian eksokrin pankreas dan diperlukan untuk pencernaan protein makanan. Ia memiliki struktur dua domain, dan pusat pengikatan trypsin dengan ligannya - protein makanan - terletak di alur antara dua domain. Di pusat aktif, kondisi yang diperlukan untuk pengikatan efektif bagian tertentu dari protein makanan dan hidrolisis ikatan peptidanya tercipta.
Domain yang berbeda dalam suatu protein dapat berpindah relatif satu sama lain ketika pusat aktif berinteraksi dengan ligan (Gbr. 1.15).
Heksokinase- enzim yang mengkatalisis fosforilasi glukosa menggunakan ATP. Situs aktif enzim terletak pada celah antara dua domain. Ketika heksokinase berikatan dengan glukosa, domain di sekitarnya menutup dan substrat menjadi terperangkap, tempat terjadinya fosforilasi (lihat Gambar 1.15).
 Beras. 1.15. Pengikatan domain heksokinase ke glukosa
Beras. 1.15. Pengikatan domain heksokinase ke glukosa
Pada beberapa protein, domain menjalankan fungsi independen dengan mengikat berbagai ligan. Protein semacam itu disebut multifungsi.
3. Obat adalah ligan yang mempengaruhi fungsi protein. Interaksi protein dengan ligan bersifat spesifik. Namun, karena labilitas konformasi protein dan pusat aktifnya, zat lain dapat dipilih yang juga dapat berinteraksi dengan protein di pusat aktif atau bagian lain dari molekul.
Suatu zat yang strukturnya mirip dengan ligan alami disebut analog struktural ligan atau ligan non-alami. Ia juga berinteraksi dengan protein di situs aktif. Analog struktural ligan dapat meningkatkan fungsi protein (agonis), dan menguranginya (antagonis). Ligan dan analog strukturalnya bersaing satu sama lain untuk berikatan dengan protein di tempat yang sama. Zat yang demikian disebut modulator kompetitif(pengatur) fungsi protein. Banyak obat bertindak sebagai penghambat protein. Beberapa di antaranya diperoleh dengan modifikasi kimia ligan alami. Penghambat fungsi protein dapat berupa obat dan racun.
Atropin adalah penghambat kompetitif reseptor M-kolinergik. Asetilkolin adalah neurotransmitter untuk transmisi impuls saraf melalui sinapsis kolinergik. Untuk melakukan eksitasi, asetilkolin yang dilepaskan ke celah sinaptik harus berinteraksi dengan protein reseptor membran postsinaptik. Ditemukan dua jenis reseptor kolinergik:
reseptor M selain asetilkolin, ia secara selektif berinteraksi dengan muscarine (toksin lalat agaric). M - reseptor kolinergik terdapat pada otot polos dan, ketika berinteraksi dengan asetilkolin, menyebabkan kontraksinya;
reseptor H secara khusus mengikat nikotin. Reseptor N-kolinergik ditemukan di sinapsis otot rangka lurik.
Penghambat spesifik Reseptor M-kolinergik adalah atropin. Hal ini ditemukan di tanaman belladonna dan henbane.
 Atropin memiliki gugus fungsi yang mirip strukturnya dengan asetilkolin dan susunan spasialnya, oleh karena itu atropin merupakan penghambat kompetitif reseptor M-kolinergik. Mengingat pengikatan asetilkolin dengan reseptor M-kolinergik menyebabkan kontraksi otot polos, maka atropin digunakan sebagai obat yang meredakan kejangnya. (antispasmodik). Oleh karena itu, diketahui penggunaan atropin untuk mengendurkan otot mata saat melihat fundus, serta untuk meredakan kejang pada kolik gastrointestinal. Reseptor M-kolinergik juga terdapat di sistem saraf pusat (SSP), sehingga atropin dosis besar dapat menyebabkan reaksi yang tidak diinginkan dari sistem saraf pusat: agitasi motorik dan mental, halusinasi, kejang.
Atropin memiliki gugus fungsi yang mirip strukturnya dengan asetilkolin dan susunan spasialnya, oleh karena itu atropin merupakan penghambat kompetitif reseptor M-kolinergik. Mengingat pengikatan asetilkolin dengan reseptor M-kolinergik menyebabkan kontraksi otot polos, maka atropin digunakan sebagai obat yang meredakan kejangnya. (antispasmodik). Oleh karena itu, diketahui penggunaan atropin untuk mengendurkan otot mata saat melihat fundus, serta untuk meredakan kejang pada kolik gastrointestinal. Reseptor M-kolinergik juga terdapat di sistem saraf pusat (SSP), sehingga atropin dosis besar dapat menyebabkan reaksi yang tidak diinginkan dari sistem saraf pusat: agitasi motorik dan mental, halusinasi, kejang.
Ditilin adalah agonis kompetitif reseptor H-kolinergik, menghambat fungsi sinapsis neuromuskular.
 Sinapsis neuromuskular otot rangka mengandung reseptor H-kolinergik. Interaksi mereka dengan asetilkolin menyebabkan kontraksi otot. Selama beberapa operasi bedah, serta dalam pemeriksaan endoskopi, obat-obatan digunakan yang menyebabkan relaksasi otot rangka (relaksan otot). Ini termasuk dithiline, yang merupakan analog struktural asetilkolin. Ia menempel pada reseptor H-kolinergik, tetapi tidak seperti asetilkolin, ia dihancurkan dengan sangat lambat oleh enzim asetilkolinesterase. Akibat pembukaan saluran ion yang berkepanjangan dan depolarisasi membran yang terus-menerus, konduksi impuls saraf terganggu dan terjadi relaksasi otot. Awalnya, sifat-sifat ini ditemukan dalam racun curare, itulah sebabnya obat tersebut disebut seperti curare.
Sinapsis neuromuskular otot rangka mengandung reseptor H-kolinergik. Interaksi mereka dengan asetilkolin menyebabkan kontraksi otot. Selama beberapa operasi bedah, serta dalam pemeriksaan endoskopi, obat-obatan digunakan yang menyebabkan relaksasi otot rangka (relaksan otot). Ini termasuk dithiline, yang merupakan analog struktural asetilkolin. Ia menempel pada reseptor H-kolinergik, tetapi tidak seperti asetilkolin, ia dihancurkan dengan sangat lambat oleh enzim asetilkolinesterase. Akibat pembukaan saluran ion yang berkepanjangan dan depolarisasi membran yang terus-menerus, konduksi impuls saraf terganggu dan terjadi relaksasi otot. Awalnya, sifat-sifat ini ditemukan dalam racun curare, itulah sebabnya obat tersebut disebut seperti curare.
TOPIK 1.3. DENATURASI PROTEIN DAN KEMUNGKINAN RENATIVASI SPONTANNYA
1. Karena konformasi asli protein dipertahankan karena interaksi yang lemah, perubahan komposisi dan sifat lingkungan sekitar protein, paparan reagen kimia dan faktor fisik menyebabkan perubahan konformasi (sifat labilitas konformasi). Pemutusan sejumlah besar ikatan menyebabkan rusaknya konformasi asli dan denaturasi protein.
Denaturasi protein- ini adalah penghancuran konformasi aslinya di bawah pengaruh zat denaturasi, yang disebabkan oleh putusnya ikatan lemah yang menstabilkan struktur spasial protein. Denaturasi disertai dengan rusaknya struktur tiga dimensi unik dan pusat aktif protein serta hilangnya aktivitas biologisnya (Gbr. 1.16).
Semua molekul terdenaturasi dari satu protein memperoleh konformasi acak yang berbeda dari molekul lain dari protein yang sama. Radikal asam amino yang membentuk pusat aktif ternyata berjauhan secara spasial satu sama lain, yaitu. situs pengikatan spesifik protein dengan ligan dihancurkan. Selama denaturasi, struktur utama protein tetap tidak berubah.
Penerapan agen denaturasi dalam penelitian biologi dan kedokteran. Dalam studi biokimia, sebelum menentukan senyawa dengan berat molekul rendah dalam bahan biologis, protein biasanya dihilangkan terlebih dahulu dari larutan. Asam trikloroasetat (TCA) paling sering digunakan untuk tujuan ini. Setelah menambahkan TCA ke dalam larutan, protein yang terdenaturasi mengendap dan mudah dihilangkan dengan penyaringan (Tabel 1.1.)
Dalam pengobatan, bahan pendenaturasi sering digunakan untuk mensterilkan peralatan dan bahan medis dalam autoklaf (bahan pendenaturasi adalah suhu tinggi) dan sebagai antiseptik (alkohol, fenol, kloramin) untuk merawat permukaan terkontaminasi yang mengandung mikroflora patogen.
2. Reaktivasi protein secara spontan- bukti determinisme struktur primer, konformasi dan fungsi protein. Protein individu adalah produk dari satu gen yang memiliki urutan asam amino yang identik dan memperoleh konformasi yang sama di dalam sel. Kesimpulan mendasar bahwa struktur primer suatu protein sudah berisi informasi tentang konformasi dan fungsinya dibuat berdasarkan kemampuan beberapa protein (khususnya, ribonuklease dan mioglobin) untuk melakukan reaktifasi secara spontan - mengembalikan konformasi aslinya setelah denaturasi.
Pembentukan struktur protein spasial dilakukan dengan metode self-assembly - suatu proses spontan di mana rantai polipeptida, yang memiliki struktur primer unik, cenderung mengadopsi konformasi dengan energi bebas terendah dalam larutan. Kemampuan untuk mengaktifkan kembali protein yang mempertahankan struktur primernya setelah denaturasi dijelaskan dalam percobaan dengan enzim ribonuklease.
Ribonuklease adalah enzim yang memecah ikatan antara nukleotida individu dalam molekul RNA. Protein globular ini mempunyai satu rantai polipeptida, struktur tersiernya distabilkan oleh banyak ikatan lemah dan empat ikatan disulfida.
Perlakuan ribonuklease dengan urea, yang memutus ikatan hidrogen dalam molekul, dan zat pereduksi, yang memutus ikatan disulfida, menyebabkan denaturasi enzim dan hilangnya aktivitasnya.
Penghapusan agen denaturasi melalui dialisis mengarah pada pemulihan konformasi dan fungsi protein, yaitu untuk kelahiran kembali. (Gbr. 1.17).
 Beras. 1.17. Denaturasi dan renativasi ribonuklease
Beras. 1.17. Denaturasi dan renativasi ribonuklease
A - konformasi asli ribonuklease, dalam struktur tersiernya terdapat empat ikatan disulfida; B - molekul ribonuklease terdenaturasi;
B - molekul ribonuklease yang diaktifkan kembali dengan struktur dan fungsi yang dipulihkan
1. Isi tabel 1.2.
Tabel 1.2. Klasifikasi asam amino menurut polaritas radikalnya
2. Tuliskan rumus tetrapeptida:
Asp - Pro - Fen - Liz
a) menyoroti gugus berulang dalam peptida yang membentuk tulang punggung peptida dan gugus variabel yang diwakili oleh radikal asam amino;
b) memberi label pada ujung N dan C;
c) menyorot ikatan peptida;
d) tulis peptida lain yang terdiri dari asam amino yang sama;
e) menghitung jumlah kemungkinan varian tetrapeptida dengan komposisi asam amino yang serupa.
3. Jelaskan peran struktur primer protein dengan menggunakan contoh analisis komparatif dua hormon peptida neurohipofisis mamalia yang serupa secara struktural dan dekat secara evolusioner - oksitosin dan vasopresin (Tabel 1.3).
Tabel 1.3. Struktur dan fungsi oksitosin dan vasopresin
 Untuk ini:
Untuk ini:
a) membandingkan komposisi dan urutan asam amino dari dua peptida;
b) menemukan kesamaan struktur utama kedua peptida dan kesamaan tindakan biologisnya;
c) menemukan perbedaan struktur dua peptida dan perbedaan fungsinya;
d) menarik kesimpulan tentang pengaruh struktur primer peptida terhadap fungsinya.
4. Jelaskan tahapan utama pembentukan konformasi protein globular (sekunder, struktur tersier, konsep struktur supersekunder). Tunjukkan jenis ikatan yang terlibat dalam pembentukan struktur protein. Radikal asam amino manakah yang dapat berpartisipasi dalam pembentukan interaksi hidrofobik, ionik, ikatan hidrogen.
Berikan contoh.
5. Definisikan konsep "labilitas konformasi protein", tunjukkan alasan keberadaan dan signifikansinya.
6. Perluas arti dari frasa berikut: “Fungsi protein didasarkan pada interaksi spesifiknya dengan ligan”, dengan menggunakan istilah dan penjelasan maknanya: konformasi protein, pusat aktif, ligan, saling melengkapi, fungsi protein.
7. Dengan menggunakan satu contoh, jelaskan apa itu domain dan apa perannya dalam fungsi protein.
TUGAS PENGENDALIAN DIRI
1. Cocok.
Gugus fungsi dalam radikal asam amino:
A. Gugus karboksil B. Gugus hidroksil C Gugus guanidin D. Gugus tiol E. Gugus amino
2. Pilihlah jawaban yang benar.
Asam amino dengan radikal tak bermuatan polar adalah:
A. Cis B. Asn
B. Glu G. Tiga
3. Pilihlah jawaban yang benar.
Radikal asam amino:
A. Memberikan kekhususan struktur primer B. Berpartisipasi dalam pembentukan struktur tersier
B. Terletak di permukaan protein, mempengaruhi kelarutannya D. Membentuk pusat aktif
D. Berpartisipasi dalam pembentukan ikatan peptida
4. Pilihlah jawaban yang benar.
Interaksi hidrofobik dapat terbentuk antara radikal asam amino:
A. Tre Lay B. Pro Tiga
B. Bertemu Ile G. Tir Ala D. Val Fen
5. Pilihlah jawaban yang benar.
Ikatan ionik dapat terbentuk antara radikal asam amino:
A. Gln Asp B. Apr Liz
B.Liz Glu G.Gis Asp D.Asn Apr
6. Pilihlah jawaban yang benar.
Ikatan hidrogen dapat terbentuk antara radikal asam amino:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Cocok.
Jenis ikatan yang terlibat dalam pembentukan struktur protein:
A. Struktur primer B. Struktur sekunder
B. Struktur tersier
D. Struktur Supersekunder E. Konformasi.
1. Ikatan hidrogen antar atom tulang punggung peptida
2. Lemahnya ikatan antar gugus fungsi radikal asam amino
3. Ikatan antara gugus α-amino dan α-karboksil asam amino
8. Pilihlah jawaban yang benar. Tripsin:
A. Enzim proteolitik B. Berisi dua domain
B. Menghidrolisis pati
D. Situs aktif terletak di antara domain. D. Terdiri dari dua rantai polipeptida.
9. Pilihlah jawaban yang benar. Atropin:
A.Neurotransmiter
B. Analog struktural asetilkolin
B. Berinteraksi dengan reseptor H-kolinergik
D. Memperkuat konduksi impuls saraf melalui sinapsis kolinergik
D. Penghambat kompetitif reseptor M-kolinergik
10. Pilihlah pernyataan yang benar. Dalam protein:
A. Struktur primer berisi informasi tentang struktur situs aktifnya
B. Pusat aktif terbentuk pada tingkat struktur primer
B. Konformasinya ditetapkan secara kaku oleh ikatan kovalen
D. Situs aktif dapat berinteraksi dengan sekelompok ligan serupa
karena labilitas konformasi protein D. Perubahan lingkungan dapat mempengaruhi afinitas zat aktif
pusat ke ligan
1.1-B, 2-G, 3-B.
3.A,B,C,D.
7. 1-B, 2-D, 3-A.
8.A,B,C,D.
SYARAT DAN KONSEP DASAR
1. Protein, polipeptida, asam amino
2. Struktur protein primer, sekunder, tersier
3. Konformasi, konformasi protein asli
4. Ikatan kovalen dan lemah pada protein
5. Labilitas konformasi
6. Situs aktif protein
7. Ligan
8. Pelipatan protein
9. Analog struktur ligan
10. Protein domain
11. Protein sederhana dan kompleks
12. Denaturasi protein, zat pendenaturasi
13. Reaktivasi protein
Menyelesaikan masalah
“Organisasi struktural protein dan dasar fungsinya”
1. Fungsi utama protein - hemoglobin A (HbA) adalah pengangkutan oksigen ke jaringan. Dalam populasi manusia, berbagai bentuk protein ini diketahui mengalami perubahan sifat dan fungsi - yang disebut hemoglobin abnormal. Misalnya, hemoglobin S, yang ditemukan dalam sel darah merah pasien dengan penyakit sel sabit (HbS), diketahui memiliki kelarutan yang rendah dalam kondisi tekanan parsial oksigen yang rendah (seperti halnya pada darah vena). Hal ini mengarah pada pembentukan agregat protein ini. Protein kehilangan fungsinya, mengendap, dan sel darah merah menjadi tidak beraturan (beberapa di antaranya berbentuk sabit) dan dihancurkan lebih cepat dari biasanya di limpa. Akibatnya, anemia sel sabit berkembang.
Satu-satunya perbedaan dalam struktur primer HbA ditemukan di wilayah N-terminal rantai β hemoglobin. Bandingkan daerah N-terminal dari untai β dan tunjukkan bagaimana perubahan struktur primer protein mempengaruhi sifat dan fungsinya.
 Untuk ini:
Untuk ini:
a) tuliskan rumus asam amino yang membedakan HbA dan bandingkan sifat asam amino tersebut (polaritas, muatan).
b) menarik kesimpulan tentang penyebab penurunan kelarutan dan terganggunya pengangkutan oksigen ke jaringan.
2. Gambar tersebut menunjukkan diagram struktur protein yang mempunyai pusat pengikatan dengan ligan (pusat aktif). Jelaskan mengapa protein bersifat selektif dalam pemilihan ligannya. Untuk ini:
a) ingat apa itu pusat aktif suatu protein dan perhatikan struktur pusat aktif protein yang ditunjukkan pada gambar;
b) tuliskan rumus radikal asam amino yang menyusun pusat aktif;
c) menggambar ligan yang secara spesifik dapat berinteraksi dengan situs aktif protein. Tunjukkan gugus fungsi yang dapat membentuk ikatan dengan radikal asam amino yang membentuk pusat aktif;
d) menunjukkan jenis ikatan yang timbul antara ligan dan radikal asam amino pusat aktif;
e) menjelaskan kekhususan interaksi protein-ligan berdasarkan apa.
 3.
Gambar tersebut menunjukkan situs aktif protein dan beberapa ligan.
3.
Gambar tersebut menunjukkan situs aktif protein dan beberapa ligan.
Tentukan ligan mana yang paling mungkin berinteraksi dengan situs aktif protein dan alasannya.
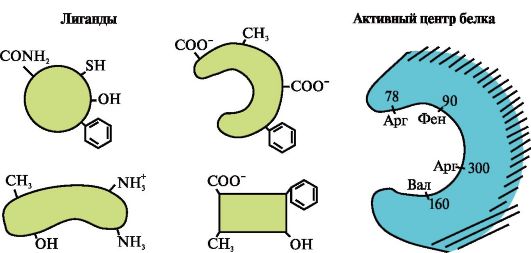 Jenis ikatan apa yang timbul selama pembentukan kompleks protein-ligan?
Jenis ikatan apa yang timbul selama pembentukan kompleks protein-ligan?
4. Analog struktural ligan protein alami dapat digunakan sebagai obat untuk memodifikasi aktivitas protein.
Asetilkolin adalah mediator transmisi eksitasi pada sinapsis neuromuskular. Ketika asetilkolin berinteraksi dengan protein - reseptor membran postsinaptik otot rangka, saluran ion terbuka dan terjadi kontraksi otot. Ditilin adalah obat yang digunakan dalam beberapa operasi untuk mengendurkan otot, karena mengganggu transmisi impuls saraf melalui sinapsis neuromuskular. Jelaskan mekanisme kerja ditilin sebagai pelemas otot. Untuk ini:
a) tuliskan rumus asetilkolin dan ditilin serta bandingkan strukturnya;
b) mendeskripsikan mekanisme efek relaksasi ditilin.
5. Pada beberapa penyakit, suhu tubuh penderita meningkat, yang dianggap sebagai reaksi perlindungan tubuh. Namun suhu tinggi merusak protein tubuh. Jelaskan mengapa pada suhu di atas 40 °C fungsi protein terganggu dan timbul ancaman bagi kehidupan manusia. Untuk melakukan ini, ingatlah:
1) Struktur protein dan ikatan yang menjaga strukturnya dalam konformasi asli;
2) Bagaimana struktur dan fungsi protein berubah seiring dengan meningkatnya suhu?;
3) Apa itu homeostatis dan mengapa penting untuk menjaga kesehatan manusia.
Unit modular 2 PROTEIN OLIGOMERICK SEBAGAI TARGET PENGARUH REGULASI. KEANEKARAGAMAN STRUKTURAL DAN FUNGSIONAL PROTEIN. METODE PEMISAHAN DAN PEMURNIKAN PROTEIN
Tujuan pembelajaran Mampu:
1. Menggunakan pengetahuan tentang ciri-ciri struktur dan fungsi protein oligomer untuk memahami mekanisme adaptif pengaturan fungsinya.
2. Jelaskan peran pendamping dalam sintesis dan pemeliharaan konformasi protein dalam kondisi seluler.
3. Menjelaskan keragaman manifestasi kehidupan melalui keragaman struktur dan fungsi protein yang disintesis dalam tubuh.
4. Menganalisis hubungan antara struktur protein dan fungsinya menggunakan contoh perbandingan hemoprotein terkait - mioglobin dan hemoglobin, serta perwakilan dari lima kelas protein dari keluarga imunoglobulin.
5. Menerapkan pengetahuan tentang kekhasan sifat fisikokimia protein untuk memilih metode pemurniannya dari protein dan pengotor lain.
6. Menafsirkan hasil komposisi kuantitatif dan kualitatif protein plasma darah untuk memastikan atau memperjelas diagnosis klinis.
Tahu:
1. Ciri-ciri struktur protein oligomer dan mekanisme adaptif untuk mengatur fungsinya menggunakan contoh hemoglobin.
2. Struktur dan fungsi pendamping dan pentingnya menjaga konformasi asli protein dalam kondisi seluler.
3. Prinsip penggabungan protein ke dalam famili berdasarkan kesamaan konformasi dan fungsinya dengan menggunakan contoh imunoglobulin.
4. Cara pemisahan protein berdasarkan ciri-ciri sifat fisikokimianya.
5. Elektroforesis plasma darah sebagai metode untuk menilai komposisi protein secara kualitatif dan kuantitatif.
TOPIK 1.4. FITUR STRUKTUR DAN FUNGSI PROTEIN OLIGOMER MENGGUNAKAN CONTOH HEMOGLOBIN
1. Banyak protein mengandung beberapa rantai polipeptida. Protein seperti ini disebut oligomer, dan rantai individu - protomer. Protomer dalam protein oligomer dihubungkan oleh banyak ikatan non-kovalen lemah (hidrofobik, ionik, hidrogen). Interaksi
protomer dilakukan berkat komplementaritas permukaan kontak mereka.
Jumlah protomer dalam protein oligomer dapat sangat bervariasi: hemoglobin mengandung 4 protomer, enzim aspartat aminotransferase memiliki 12 protomer, dan protein virus mosaik tembakau mengandung 2.120 protomer yang dihubungkan melalui ikatan non-kovalen. Akibatnya, protein oligomer dapat memiliki berat molekul yang sangat tinggi.
Interaksi satu protomer dengan protomer lainnya dapat dianggap sebagai kasus khusus interaksi protein-ligan, karena setiap protomer berfungsi sebagai ligan bagi protomer lainnya. Jumlah dan cara bergabungnya protomer dalam suatu protein disebut struktur protein kuaterner.
Protein dapat mengandung protomer dengan struktur yang sama atau berbeda, misalnya homodimer adalah protein yang mengandung dua protomer identik, dan heterodimer adalah protein yang mengandung dua protomer berbeda.
Jika protein mengandung protomer yang berbeda, maka pusat pengikatan dengan ligan berbeda yang strukturnya berbeda dapat terbentuk pada protein tersebut. Ketika ligan berikatan dengan situs aktif, fungsi protein ini terwujud. Pusat yang terletak pada protomer berbeda disebut alosterik (berbeda dengan protomer aktif). Menghubungi ligan atau efektor alosterik, ia melakukan fungsi pengaturan (Gbr. 1.18). Interaksi pusat alosterik dengan efektor menyebabkan perubahan konformasi pada struktur seluruh protein oligomer karena labilitas konformasinya. Hal ini mempengaruhi afinitas situs aktif untuk ligan tertentu dan mengatur fungsi protein tersebut. Perubahan konformasi dan fungsi semua protomer selama interaksi protein oligomer dengan setidaknya satu ligan disebut perubahan konformasi kooperatif. Efektor yang meningkatkan fungsi protein disebut aktivator, dan efektor yang menghambat fungsinya - penghambat.
Jadi, protein oligomer, serta protein dengan struktur domain, memiliki sifat baru dibandingkan dengan protein monomer - kemampuan untuk mengatur fungsi secara alosterik (regulasi dengan menempelkan ligan berbeda ke protein). Hal ini dapat dilihat dengan membandingkan struktur dan fungsi dua protein kompleks yang berkaitan erat, mioglobin dan hemoglobin.
 Beras. 1.18. Skema struktur protein dimer
Beras. 1.18. Skema struktur protein dimer
2. Pembentukan struktur spasial dan fungsi mioglobin.
Mioglobin (Mb) adalah protein yang terdapat pada otot merah, yang fungsi utamanya adalah menciptakan cadangan O2 yang diperlukan untuk kerja otot yang intens. Mb adalah protein kompleks yang mengandung bagian protein - apoMb dan bagian non-protein - heme. Struktur utama apoMB menentukan konformasi globular kompaknya dan struktur pusat aktif, tempat melekatnya bagian non-protein mioglobin, heme. Oksigen yang berasal dari darah ke otot berikatan dengan heme Fe+2 di mioglobin. Mb merupakan protein monomer yang memiliki afinitas sangat tinggi terhadap O 2, sehingga pelepasan oksigen oleh mioglobin hanya terjadi selama kerja otot yang intens, ketika tekanan parsial O 2 menurun tajam.
Pembentukan konformasi Mv. Di otot merah, pada ribosom, selama translasi, struktur utama MB disintesis, diwakili oleh urutan spesifik 153 residu asam amino. Struktur sekunder Mb mengandung delapan heliks α, yang disebut dengan huruf Latin dari A sampai H, di antaranya terdapat daerah non-heliks. Struktur tersier Mb berbentuk globul padat, di dalam ceruknya terdapat pusat aktif antara heliks F dan E (Gbr. 1.19).
 Beras. 1.19. Struktur mioglobin
Beras. 1.19. Struktur mioglobin
3. Ciri-ciri struktur dan fungsi pusat aktif MV. Pusat aktif Mb dibentuk terutama oleh radikal asam amino hidrofobik, yang berjarak jauh satu sama lain dalam struktur primer (misalnya, Tri 3 9 dan Fen 138) Ligan yang sukar larut dalam air - heme dan O 2 - menempel pada pusat aktif. Heme adalah ligan spesifik apoMB (Gbr. 1.20), yang dasarnya terdiri dari empat cincin pirol yang dihubungkan oleh jembatan metenil; di tengahnya terdapat atom Fe+ 2 yang terhubung ke atom nitrogen pada cincin pirol melalui empat ikatan koordinasi. Di pusat aktif Mb, selain radikal asam amino hidrofobik, juga terdapat residu dua asam amino dengan radikal hidrofilik - Gis E 7(Gis 64) dan GIS F 8(93 miliknya) (Gbr. 1.21).
 Beras. 1.20. Struktur heme - bagian non-protein dari mioglobin dan hemoglobin
Beras. 1.20. Struktur heme - bagian non-protein dari mioglobin dan hemoglobin
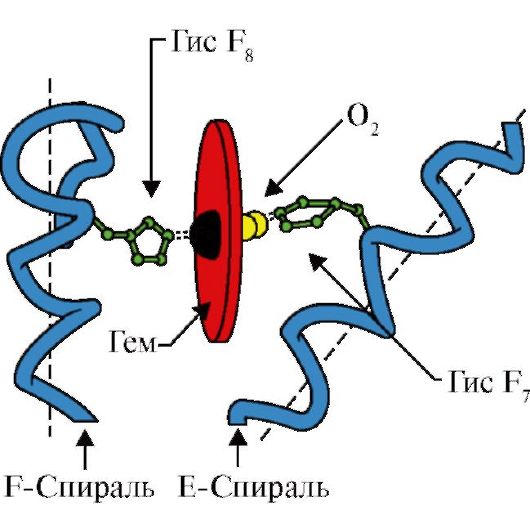 Beras. 1.21. Lokasi heme dan O2 di situs aktif apomioglobin dan protomer hemoglobin
Beras. 1.21. Lokasi heme dan O2 di situs aktif apomioglobin dan protomer hemoglobin
Heme terikat secara kovalen ke F8-Nya melalui atom besi. O 2 menempel pada besi di sisi lain bidang heme. E 7-nya diperlukan untuk orientasi O 2 yang benar dan memfasilitasi penambahan oksigen ke Fe + 2 heme
GIS F 8 membentuk ikatan koordinasi dengan Fe+ 2 dan mengikat heme dengan kuat di pusat aktif. Gis E 7 diperlukan untuk orientasi yang benar di pusat aktif ligan lain - O 2 selama interaksinya dengan Fe + 2 heme. Lingkungan mikro heme menciptakan kondisi pengikatan O2 ke Fe+2 yang kuat namun reversibel dan mencegah air memasuki situs aktif hidrofobik, yang dapat menyebabkan oksidasi menjadi Fe+3.
Struktur monomer Mb dan pusat aktifnya menentukan tingginya afinitas protein terhadap O2.
4. Struktur oligomer Hb dan pengaturan afinitas Hb terhadap ligan O2. Hemoglobin manusia- keluarga protein, seperti mioglobin, yang berhubungan dengan protein kompleks (hemoprotein). Mereka memiliki struktur tetramerik dan mengandung dua rantai α, tetapi berbeda dalam struktur dua rantai polipeptida lainnya (rantai 2α-, 2x). Struktur rantai polipeptida kedua menentukan ciri-ciri fungsi bentuk Hb ini. Sekitar 98% hemoglobin dalam sel darah merah orang dewasa adalah hemoglobin A(rantai 2α-, 2p).
Selama perkembangan janin, ada dua jenis utama hemoglobin yang berfungsi: Hb embrionik(2α, 2ε), yang ditemukan pada tahap awal perkembangan janin, dan hemoglobin F (janin)- (2α, 2γ), yang menggantikan hemoglobin janin awal pada bulan keenam perkembangan intrauterin dan hanya setelah lahir digantikan oleh Hb A.
HB A adalah protein terkait mioglobin (MB) yang ditemukan pada sel darah merah manusia dewasa. Struktur masing-masing protomernya mirip dengan mioglobin. Struktur sekunder dan tersier dari protomer mioglobin dan hemoglobin sangat mirip, meskipun faktanya dalam struktur primer rantai polipeptidanya hanya 24 residu asam amino yang identik (struktur sekunder protomer hemoglobin, seperti mioglobin, mengandung delapan heliks α, ditunjuk dengan huruf latin dari A sampai H , dan struktur tersiernya berbentuk bola kompak). Namun berbeda dengan mioglobin, hemoglobin memiliki struktur oligomer, terdiri dari empat rantai polipeptida yang dihubungkan melalui ikatan non-kovalen (Gambar 1.22).
Setiap protomer Hb dikaitkan dengan bagian non-protein - heme dan protomer tetangganya. Hubungan bagian protein Hb dengan heme mirip dengan mioglobin: pada pusat aktif protein, bagian hidrofobik heme dikelilingi oleh radikal asam amino hidrofobik kecuali His F 8 dan His E 7, yang terletak di kedua sisi bidang heme dan memainkan peran serupa dalam fungsi protein dan pengikatannya dengan oksigen (lihat struktur mioglobin).
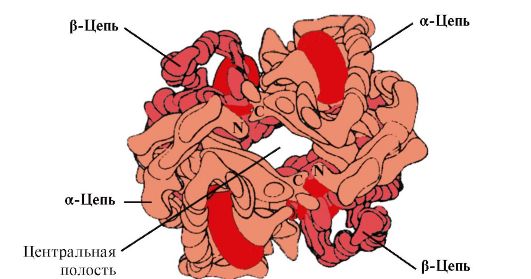 Beras. 1.22. Struktur oligomer hemoglobin
Beras. 1.22. Struktur oligomer hemoglobin
Di samping itu, Gis E 7 melakukan hal yang penting peran tambahan dalam berfungsinya Nv. Heme bebas memiliki afinitas 25.000 kali lebih tinggi terhadap CO dibandingkan O2. CO terbentuk dalam jumlah kecil di dalam tubuh dan, mengingat afinitasnya yang tinggi terhadap heme, CO dapat mengganggu pengangkutan O2 yang diperlukan untuk kehidupan sel. Namun, dalam komposisi hemoglobin, afinitas heme terhadap karbon monoksida melebihi afinitas terhadap O 2 hanya 200 kali lipat karena adanya His E 7 di pusat aktif. Sisa asam amino ini menciptakan kondisi optimal untuk pengikatan heme ke O2 dan melemahkan interaksi heme dengan CO.
5. Fungsi utama HB adalah mengangkut O2 dari paru-paru ke jaringan. Berbeda dengan mioglobin monomer, yang memiliki afinitas sangat tinggi terhadap O2 dan berfungsi menyimpan oksigen di otot merah, struktur oligomer hemoglobin menyediakan:
1) cepat saturasi HB dengan oksigen di paru-paru;
2) kemampuan HB melepaskan oksigen dalam jaringan pada tekanan parsial O 2 yang relatif tinggi (20-40 mm Hg);
3) kemungkinan mengatur afinitas Hb terhadap O2.
6. Perubahan kooperatif dalam konformasi protomer hemoglobin mempercepat pengikatan O2 di paru-paru dan pelepasannya ke jaringan. Di paru-paru, tekanan parsial O2 yang tinggi mendorong pengikatannya dengan Hb di situs aktif empat protomer (2α dan 2β). Pusat aktif setiap protomer, seperti pada mioglobin, terletak di antara dua heliks α (F dan E) dalam kantong hidrofobik. Ini mengandung bagian non-protein - heme, melekat pada bagian protein melalui banyak interaksi hidrofobik lemah dan satu ikatan kuat antara Fe 2 + heme dan His F 8 (lihat Gambar 1.21).
Dalam deoksihemoglobin, karena ikatan dengan F 8 His, atom Fe 2+ menonjol dari bidang heme menuju histidin. Pengikatan O 2 ke Fe 2 + terjadi di sisi lain heme di wilayah His E 7 menggunakan ikatan koordinasi bebas tunggal. E 7 miliknya memberikan kondisi optimal untuk pengikatan O 2 ke besi heme.
Penambahan O2 pada atom Fe+2 dari salah satu protomer menyebabkan pergerakannya ke bidang heme, diikuti oleh residu histidin yang terkait dengannya.
 Beras. 1.23. Perubahan konformasi protomer hemoglobin bila dikombinasikan dengan O2
Beras. 1.23. Perubahan konformasi protomer hemoglobin bila dikombinasikan dengan O2
Hal ini menyebabkan perubahan konformasi semua rantai polipeptida karena labilitas konformasinya. Mengubah konformasi rantai lain memfasilitasi interaksinya dengan molekul O2 berikutnya.
Molekul O2 keempat menempel pada hemoglobin 300 kali lebih mudah dibandingkan molekul pertama (Gbr. 1.24).
 Beras. 1.24. Perubahan kooperatif dalam konformasi protomer hemoglobin selama interaksinya dengan O2
Beras. 1.24. Perubahan kooperatif dalam konformasi protomer hemoglobin selama interaksinya dengan O2
Dalam jaringan, setiap molekul O2 berikutnya lebih mudah dibelah daripada molekul sebelumnya, juga karena perubahan kooperatif dalam konformasi protomer.
7. CO 2 dan H+, yang terbentuk selama katabolisme zat organik, mengurangi afinitas hemoglobin terhadap O 2 sebanding dengan konsentrasinya. Energi yang dibutuhkan untuk fungsi sel diproduksi terutama di mitokondria selama oksidasi zat organik menggunakan O2 yang dikirim dari paru-paru melalui hemoglobin. Sebagai hasil oksidasi zat organik, produk akhir penguraiannya terbentuk: CO 2 dan K 2 O, yang jumlahnya sebanding dengan intensitas proses oksidasi yang sedang berlangsung.
CO 2 berdifusi dari sel ke dalam darah dan menembus ke dalam sel darah merah, di mana, di bawah aksi enzim karbanhidrase, CO2 diubah menjadi asam karbonat. Asam lemah ini berdisosiasi menjadi proton dan ion bikarbonat.
H+ mampu bergabung dengan radikal-radikalnya 14 6 dalam rantai α- dan β hemoglobin, mis. di daerah yang jauh dari heme. Protonasi hemoglobin mengurangi afinitasnya terhadap O2, mendorong pembuangan O2 dari oksiHb, pembentukan deoksiHb, dan meningkatkan suplai oksigen ke jaringan sebanding dengan jumlah proton yang terbentuk (Gbr. 1.25).
Peningkatan jumlah oksigen yang dilepaskan tergantung pada peningkatan konsentrasi H+ dalam sel darah merah disebut efek Bohr (dinamai menurut ahli fisiologi Denmark Christian Bohr, yang pertama kali menemukan efek ini).
Di paru-paru, tekanan parsial oksigen yang tinggi mendorong pengikatannya dengan deoksiHb, yang mengurangi afinitas protein terhadap H+. Proton yang dilepaskan di bawah pengaruh asam karbonat bereaksi dengan bikarbonat membentuk CO 2 dan H 2 O
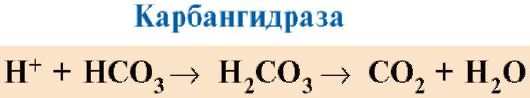
 Beras. 1.25. Ketergantungan afinitas Hb terhadap O 2 pada konsentrasi CO 2 dan proton (efek Bohr):
Beras. 1.25. Ketergantungan afinitas Hb terhadap O 2 pada konsentrasi CO 2 dan proton (efek Bohr):
A- pengaruh konsentrasi CO 2 dan H+ terhadap pelepasan O 2 dari kompleks dengan HB (efek Bohr); B- oksigenasi deoksihemoglobin di paru-paru, pembentukan dan pelepasan CO2.
CO 2 yang dihasilkan memasuki ruang alveolar dan dikeluarkan bersama udara yang dihembuskan. Dengan demikian, jumlah oksigen yang dilepaskan oleh hemoglobin dalam jaringan diatur oleh produk katabolisme zat organik: semakin intens pemecahan zat, misalnya selama latihan fisik, semakin tinggi konsentrasi CO 2 dan H + serta semakin banyak oksigen. jaringan menerima sebagai akibat dari penurunan afinitas Hb terhadap O2.
8. Regulasi alosterik afinitas Hb terhadap O2 oleh ligan - 2,3-bifosfogliserat. Dalam eritrosit, ligan alosterik hemoglobin, 2,3-bifosfogliserat (2,3-BPG), disintesis dari produk oksidasi glukosa - 1,3-bifosfogliserat. Dalam kondisi normal, konsentrasi 2,3-BPG tinggi dan sebanding dengan konsentrasi Hb. 2,3-BPG memiliki muatan negatif kuat -5.
 Bifosfogliserat di kapiler jaringan, berikatan dengan deoksihemoglobin, meningkatkan pelepasan oksigen ke jaringan, mengurangi afinitas Hb terhadap O2.
Bifosfogliserat di kapiler jaringan, berikatan dengan deoksihemoglobin, meningkatkan pelepasan oksigen ke jaringan, mengurangi afinitas Hb terhadap O2.
Di tengah molekul hemoglobin tetramerik terdapat rongga. Ini dibentuk oleh residu asam amino dari keempat protomer (lihat Gambar 1.22). Di kapiler jaringan, protonasi Hb (efek Bohr) menyebabkan putusnya ikatan antara besi heme dan O2. Dalam sebuah molekul
deoksihemoglobin, dibandingkan dengan oksihemoglobin, muncul ikatan ion tambahan yang menghubungkan protomer, akibatnya dimensi rongga pusat meningkat dibandingkan dengan oksihemoglobin. Rongga sentral merupakan tempat melekatnya 2,3-BPG pada hemoglobin. Karena perbedaan ukuran rongga tengah, 2,3-BPG hanya dapat menempel pada deoksihemoglobin.
2,3-BPG berinteraksi dengan hemoglobin di tempat yang jauh dari pusat aktif protein dan termasuk dalam alosterik ligan (pengaturan), dan rongga tengah Hb adalah pusat alosterik. 2,3-BPG memiliki muatan negatif yang kuat dan berinteraksi dengan lima gugus bermuatan positif dari dua rantai β Hb: gugus N-terminal α-amino dari Val dan radikal Lys 82 His 143 (Gbr. 1.26).
 Beras. 1.26. BPG di rongga tengah deoksihemoglobin
Beras. 1.26. BPG di rongga tengah deoksihemoglobin
BPG berikatan dengan tiga gugus bermuatan positif pada setiap untai β.
Dalam kapiler jaringan, deoksihemoglobin yang dihasilkan berinteraksi dengan 2,3-BPG dan ikatan ionik terbentuk antara radikal rantai β bermuatan positif dan ligan bermuatan negatif, yang mengubah konformasi protein dan mengurangi afinitas Hb terhadap O2. . Penurunan afinitas Hb terhadap O 2 mendorong pelepasan O 2 yang lebih efisien ke dalam jaringan.
Di paru-paru, pada tekanan parsial tinggi, oksigen berinteraksi dengan Hb, bergabung dengan besi heme; dalam hal ini, konformasi protein berubah, rongga pusat berkurang dan 2,3-BPG dipindahkan dari pusat alosterik
Dengan demikian, protein oligomer memiliki sifat baru dibandingkan dengan protein monomer. Perlekatan ligan di situs
berjauhan secara spasial satu sama lain (alosterik), dapat menyebabkan perubahan konformasi pada seluruh molekul protein. Akibat interaksi dengan ligan pengatur, terjadi perubahan konformasi dan adaptasi fungsi molekul protein terhadap perubahan lingkungan.
TOPIK 1.5. PEMELIHARAAN KONFORMASI ASLI PROTEIN DALAM KONDISI SEL
Dalam sel, selama sintesis rantai polipeptida, pengangkutannya melalui membran ke bagian sel yang sesuai, selama proses pelipatan (pembentukan konformasi asli) dan selama perakitan protein oligomer, serta selama fungsinya, zat antara , konformasi yang rawan agregasi dan tidak stabil muncul dalam struktur protein. Radikal hidrofobik, biasanya tersembunyi di dalam molekul protein dalam konformasi asli, muncul di permukaan dalam konformasi tidak stabil dan cenderung bergabung dengan kelompok protein lain yang sulit larut dalam air. Protein khusus telah ditemukan dalam sel semua organisme yang diketahui yang memastikan pelipatan protein sel yang optimal, menstabilkan konformasi aslinya selama berfungsi dan, yang paling penting, mempertahankan struktur dan fungsi protein intraseluler ketika homeostasis terganggu. Protein-protein ini disebut "pendamping" yang berarti "pengasuh" dalam bahasa Perancis.
1. Pendamping molekuler dan perannya dalam mencegah denaturasi protein.
Pendamping (CH) diklasifikasikan menurut massa subunitnya. Pendamping dengan berat molekul tinggi memiliki massa 60 hingga 110 kDa. Diantaranya, tiga kelas yang paling banyak dipelajari: Sh-60, Sh-70 dan Sh-90. Setiap kelas mencakup keluarga protein terkait. Jadi, Sh-70 mencakup protein dengan berat molekul 66 hingga 78 kDa. Pendamping dengan berat molekul rendah memiliki berat molekul 40 hingga 15 kDa.
Di antara para pendamping ada pokok protein, sintesis basalnya yang tinggi tidak bergantung pada efek stres pada sel-sel tubuh, dan dapat diinduksi, sintesisnya lemah dalam kondisi normal, tetapi meningkat tajam di bawah tekanan. Pendamping yang dapat diinduksi juga disebut “protein kejutan panas” karena pertama kali ditemukan dalam sel yang terkena suhu tinggi. Dalam sel, karena tingginya konsentrasi protein, reaktivasi spontan protein yang terdenaturasi sebagian sulit dilakukan. Sh-70 dapat mencegah timbulnya denaturasi dan membantu mengembalikan konformasi asli protein. Pendamping molekuler-70- kelas protein yang sangat terkonservasi yang ditemukan di semua bagian sel: sitoplasma, nukleus, retikulum endoplasma, mitokondria. Pada ujung karboksil rantai polipeptida tunggal Ш-70 terdapat daerah berupa alur yang mampu berinteraksi dengan peptida yang panjangnya
dari 7 hingga 9 residu asam amino yang diperkaya dengan radikal hidrofobik. Daerah seperti itu pada protein globular terdapat kira-kira setiap 16 asam amino. Sh-70 mampu melindungi protein dari inaktivasi suhu dan memulihkan konformasi serta aktivitas protein yang terdenaturasi sebagian.
2. Peran pendamping dalam pelipatan protein. Selama sintesis protein di ribosom, daerah terminal-N dari polipeptida disintesis sebelum terminal-C. Untuk membentuk konformasi asli, diperlukan rangkaian asam amino lengkap dari protein. Dalam proses sintesis protein, pendamping-70 karena struktur pusat aktifnya mampu menutup area polipeptida yang rentan agregasi, diperkaya dengan radikal asam amino hidrofobik hingga sintesis selesai (Gambar 1.27, A ).
 Beras. 1.27. Partisipasi pendamping dalam pelipatan protein
Beras. 1.27. Partisipasi pendamping dalam pelipatan protein
A - partisipasi pendamping-70 dalam pencegahan interaksi hidrofobik antara bagian polipeptida yang disintesis; B - pembentukan konformasi asli protein di kompleks pendamping
Banyak protein bermolekul tinggi yang memiliki konformasi kompleks, seperti struktur domain, terlipat dalam ruang khusus yang dibentuk oleh Sh-60. Sh-60 berfungsi sebagai kompleks oligomer yang terdiri dari 14 subunit. Mereka membentuk dua cincin berongga, yang masing-masing terdiri dari tujuh subunit, cincin ini terhubung satu sama lain. Setiap subunit Sh-60 terdiri dari tiga domain: apikal (apikal), diperkaya dengan radikal hidrofobik yang menghadap rongga cincin, perantara dan khatulistiwa (Gbr. 1.28).
 Beras. 1.28. Struktur kompleks pendamping terdiri dari 14 Ш-60
Beras. 1.28. Struktur kompleks pendamping terdiri dari 14 Ш-60
A - tampak samping; B - tampilan atas
Protein yang disintesis, yang memiliki unsur-unsur pada permukaan yang merupakan karakteristik molekul yang tidak terlipat, khususnya radikal hidrofobik, memasuki rongga cincin pendamping. Dalam lingkungan spesifik rongga-rongga ini, kemungkinan konformasi dicari sampai satu-satunya konformasi yang paling disukai secara energetik ditemukan (Gbr. 1.27, B). Pembentukan konformasi dan pelepasan protein disertai dengan hidrolisis ATP di daerah ekuator. Biasanya, pelipatan yang bergantung pada pendamping membutuhkan banyak energi.
Selain berpartisipasi dalam pembentukan struktur tiga dimensi protein dan renativasi protein yang terdenaturasi sebagian, pendamping juga diperlukan untuk terjadinya proses mendasar seperti perakitan protein oligomer, pengenalan dan pengangkutan protein terdenaturasi ke dalam lisosom, pengangkutan protein melintasi membran, dan partisipasi dalam pengaturan aktivitas kompleks protein.
TOPIK 1.6. VARIETAS PROTEIN. KELUARGA PROTEIN: CONTOH IMUNOGLOBULIN
1. Protein memainkan peran penting dalam kehidupan sel individu dan seluruh organisme multiseluler, dan fungsinya sangat beragam. Hal ini ditentukan oleh karakteristik struktur primer dan konformasi protein, struktur unik pusat aktif dan kemampuan mengikat ligan tertentu.
Hanya sebagian kecil dari semua kemungkinan varian rantai peptida yang dapat mengadopsi struktur spasial yang stabil; mayoritas
diantaranya dapat mengambil banyak konformasi dengan energi Gibbs yang kira-kira sama, tetapi dengan sifat yang berbeda. Struktur utama dari sebagian besar protein yang diketahui, dipilih melalui evolusi biologis, memastikan stabilitas luar biasa dari salah satu konformasi, yang menentukan karakteristik fungsi protein ini.
2. Keluarga protein. Dalam spesies biologis yang sama, substitusi residu asam amino dapat menyebabkan munculnya protein berbeda yang menjalankan fungsi terkait dan memiliki rangkaian asam amino homolog. Protein terkait tersebut memiliki konformasi yang sangat mirip: jumlah dan posisi relatif heliks α dan/atau struktur β, sebagian besar putaran dan lengkungan rantai polipeptida serupa atau identik. Protein dengan daerah rantai polipeptida homolog, konformasi serupa dan fungsi terkait diklasifikasikan ke dalam keluarga protein. Contoh keluarga protein: proteinase serin, keluarga imunoglobulin, keluarga mioglobin.
Proteinase serin- keluarga protein yang menjalankan fungsi enzim proteolitik. Ini termasuk enzim pencernaan - chymotrypsin, trypsin, elastase dan banyak faktor pembekuan darah. Protein ini memiliki asam amino yang identik pada 40% posisinya dan konformasi yang sangat mirip (Gbr. 1.29).

Beras. 1.29. Struktur spasial elastase (A) dan kimotripsin (B)
Beberapa substitusi asam amino telah menyebabkan perubahan spesifisitas substrat protein ini dan munculnya keragaman fungsional dalam famili tersebut.
3. Keluarga imunoglobulin. Dalam berfungsinya sistem kekebalan tubuh, protein dari superfamili imunoglobulin memainkan peran besar, yang mencakup tiga keluarga protein:
Antibodi (imunoglobulin);
Reseptor limfosit T;
Protein dari kompleks histokompatibilitas utama - MHC kelas 1 dan 2 (Kompleks Histokompatibilitas Utama).
Semua protein ini memiliki struktur domain, terdiri dari domain mirip imun yang homolog dan melakukan fungsi serupa: mereka berinteraksi dengan struktur asing, baik yang terlarut dalam darah, getah bening atau cairan antar sel (antibodi), atau terletak di permukaan sel (milik atau luar negeri).
4. Antibodi- protein spesifik yang diproduksi oleh limfosit B sebagai respons terhadap masuknya struktur asing ke dalam tubuh, disebut antigen.
Fitur struktur antibodi
Molekul antibodi paling sederhana terdiri dari empat rantai polipeptida: dua rantai ringan identik - L, mengandung sekitar 220 asam amino, dan dua rantai berat identik - H, terdiri dari 440-700 asam amino. Keempat rantai dalam molekul antibodi dihubungkan oleh banyak ikatan non-kovalen dan empat ikatan disulfida (Gbr. 1.30).
Rantai ringan antibodi terdiri dari dua domain: domain variabel (VL), yang terletak di wilayah terminal-N rantai polipeptida, dan domain konstan (CL), yang terletak di terminal-C. Rantai berat biasanya memiliki empat domain: satu variabel (VH), terletak di ujung N, dan tiga domain konstan (CH1, CH2, CH3) (lihat Gambar 1.30). Setiap domain imunoglobulin memiliki superstruktur lembaran β di mana dua residu sistein dihubungkan oleh ikatan disulfida.
Di antara dua domain konstan CH1 dan CH2 terdapat wilayah yang mengandung sejumlah besar residu prolin, yang mencegah pembentukan struktur sekunder dan interaksi rantai H yang berdekatan di segmen ini. Daerah engsel ini memberikan fleksibilitas molekul antibodi. Di antara domain variabel rantai berat dan ringan terdapat dua situs pengikatan antigen yang identik (situs aktif untuk mengikat antigen), oleh karena itu antibodi semacam itu sering disebut bivalen. Tidak seluruh rangkaian asam amino dari daerah variabel kedua rantai terlibat dalam pengikatan antigen ke antibodi, tetapi hanya 20-30 asam amino yang terletak di daerah hipervariabel dari setiap rantai. Wilayah inilah yang menentukan kemampuan unik setiap jenis antibodi untuk berinteraksi dengan antigen komplementer terkait.
Antibodi adalah salah satu garis pertahanan tubuh terhadap serangan organisme asing. Fungsinya dapat dibagi menjadi dua tahap: tahap pertama adalah pengenalan dan pengikatan antigen pada permukaan organisme asing, yang dimungkinkan karena adanya situs pengikatan antigen dalam struktur antibodi; tahap kedua adalah permulaan proses inaktivasi dan penghancuran antigen. Spesifisitas tahap kedua bergantung pada kelas antibodi. Ada lima kelas rantai berat yang berbeda satu sama lain dalam struktur domain konstan: α, δ, ε, γ dan μ, yang menurutnya lima kelas imunoglobulin dibedakan: A, D, E, G dan M.
Ciri-ciri struktural rantai berat memberikan daerah engsel dan daerah terminal-C rantai berat suatu karakteristik konformasi dari setiap kelas. Setelah antigen berikatan dengan antibodi, perubahan konformasi dalam domain konstan menentukan jalur penghilangan antigen.
 Beras. 1. 30. Struktur domain IgG
Beras. 1. 30. Struktur domain IgG
Imunoglobulin M
Imunoglobulin M memiliki dua bentuk.
Bentuk monomer- Antibodi kelas 1 yang diproduksi oleh pengembangan limfosit B. Selanjutnya, banyak sel B beralih untuk memproduksi antibodi kelas lain, tetapi dengan tempat pengikatan antigen yang sama. IgM tertanam dalam membran dan bertindak sebagai reseptor pengenalan antigen. Integrasi IgM ke dalam membran sel dimungkinkan karena adanya 25 residu asam amino hidrofobik di bagian ekor.
Bentuk sekretori IgM mengandung lima subunit monomer yang dihubungkan satu sama lain melalui ikatan disulfida dan rantai J polipeptida tambahan (Gbr. 1.31). Rantai berat monomer bentuk ini tidak mengandung ekor hidrofobik. Pentamer memiliki 10 situs pengikatan antigen sehingga efektif dalam mengenali dan menghilangkan antigen yang pertama kali masuk ke dalam tubuh. Bentuk sekretori IgM adalah kelas antibodi utama yang disekresikan ke dalam darah selama respon imun primer. Pengikatan IgM ke antigen mengubah konformasi IgM dan menginduksi pengikatannya pada komponen protein pertama dari sistem komplemen (sistem komplemen adalah sekumpulan protein yang terlibat dalam penghancuran antigen) dan aktivasi sistem ini. Jika antigen terletak pada permukaan suatu mikroorganisme, maka sistem komplemen menyebabkan terganggunya integritas membran sel dan kematian sel bakteri.
Imunoglobulin G
Secara kuantitatif, kelas imunoglobulin ini mendominasi dalam darah (75% dari seluruh Ig). IgG - monomer, kelas utama antibodi yang disekresikan ke dalam darah selama respon imun sekunder. Setelah interaksi IgG dengan antigen permukaan mikroorganisme, kompleks antigen-antibodi mampu mengikat dan mengaktifkan protein sistem komplemen atau dapat berinteraksi dengan reseptor spesifik makrofag dan neutrofil. Interaksi dengan fagosit mengarah
 Beras. 1.31. Struktur bentuk sekretori IgM
Beras. 1.31. Struktur bentuk sekretori IgM
untuk penyerapan kompleks antigen-antibodi dan penghancurannya dalam fagosom sel. IgG merupakan satu-satunya golongan antibodi yang mampu menembus penghalang plasenta dan memberikan perlindungan intrauterin pada janin dari infeksi.
Imunoglobulin A
Kelas utama antibodi terdapat dalam sekret (susu, air liur, sekret saluran pernafasan dan saluran usus). IgA disekresi terutama dalam bentuk dimer, di mana monomer dihubungkan satu sama lain melalui rantai J tambahan (Gbr. 1.32).
IgA tidak berinteraksi dengan sistem komplemen dan sel fagositik, tetapi dengan mengikat mikroorganisme, antibodi mencegah perlekatan mereka pada sel epitel dan penetrasi ke dalam tubuh.
Imunoglobulin E
Imunoglobulin E diwakili oleh monomer yang mengandung rantai ε berat, seperti rantai μ imunoglobulin M, satu variabel dan empat domain konstan. Setelah disekresi, IgE berikatan dengannya
 Beras. 1.32. Struktur IgA
Beras. 1.32. Struktur IgA
Daerah terminal-C dengan reseptor yang sesuai pada permukaan sel mast dan basofil. Akibatnya, mereka menjadi reseptor antigen pada permukaan sel tersebut (Gbr. 1.33).
 Beras. 1.33. Interaksi IgE dengan antigen pada permukaan sel mast
Beras. 1.33. Interaksi IgE dengan antigen pada permukaan sel mast
Setelah antigen menempel pada situs pengikatan antigen IgE yang sesuai, sel menerima sinyal untuk mengeluarkan zat aktif biologis (histamin, serotonin), yang sebagian besar bertanggung jawab untuk perkembangan reaksi inflamasi dan manifestasi reaksi alergi seperti sebagai asma, urtikaria, demam.
Imunoglobulin D
Imunoglobulin D ditemukan dalam jumlah yang sangat kecil dalam serum; Rantai δ berat memiliki satu domain variabel dan tiga domain konstan. IgD bertindak sebagai reseptor untuk limfosit B; fungsi lainnya masih belum diketahui. Interaksi antigen spesifik dengan reseptor pada permukaan limfosit B (IgD) menyebabkan transmisi sinyal-sinyal ini ke dalam sel dan aktivasi mekanisme yang memastikan proliferasi klon limfosit tertentu.
TOPIK 1.7. SIFAT FISIK DAN KIMIA PROTEIN SERTA METODE PEMISAHANNYA
1. Protein individu berbeda dalam sifat fisik dan kimia:
Bentuk molekul;
Berat molekul;
Muatan total, yang besarnya bergantung pada perbandingan gugus asam amino anionik dan kationik;
Rasio radikal asam amino polar dan nonpolar pada permukaan molekul;
Tingkat resistensi terhadap berbagai agen denaturasi.
2. Kelarutan protein tergantung tentang sifat-sifat protein yang tercantum di atas, serta tentang komposisi media di mana protein dilarutkan (nilai pH, komposisi garam, suhu, keberadaan zat organik lain yang dapat berinteraksi dengan protein). Besarnya muatan molekul protein merupakan salah satu faktor yang mempengaruhi kelarutannya. Ketika muatan pada titik isoelektrik hilang, protein lebih mudah berkumpul dan mengendap. Hal ini terutama terjadi pada protein terdenaturasi, di mana radikal asam amino hidrofobik muncul di permukaan.
Pada permukaan molekul protein terdapat radikal asam amino bermuatan positif dan negatif. Jumlah gugus ini, dan muatan total protein, bergantung pada pH medium, yaitu. perbandingan konsentrasi gugus H+ - dan OH -. Dalam lingkungan asam Peningkatan konsentrasi H+ menyebabkan penekanan disosiasi gugus karboksil -COO - + H+ > - COOH dan penurunan muatan negatif protein. Dalam lingkungan basa, pengikatan kelebihan OH - oleh proton yang terbentuk selama disosiasi gugus amino -NH 3 + + OH - - NH 2 + H 2 O dengan pembentukan air, menyebabkan penurunan muatan positif protein . Nilai pH dimana suatu protein mempunyai muatan nol disebut titik isoelektrik (IEP). Dalam IET, jumlah gugus bermuatan positif dan negatif adalah sama, yaitu. protein berada dalam keadaan isoelektrik.
3. Pemisahan protein individu. Ciri-ciri struktur dan fungsi tubuh bergantung pada kumpulan protein yang disintesis di dalamnya. Mempelajari struktur dan sifat protein tidak mungkin dilakukan tanpa mengisolasinya dari sel dan memurnikannya dari protein lain dan molekul organik. Tahapan isolasi dan pemurnian protein individu:
Penghancuran sel jaringan yang sedang dipelajari dan memperoleh homogenat.
Pemisahan homogenat menjadi pecahan dengan sentrifugasi, memperoleh fraksi nuklir, mitokondria, sitosol atau lainnya yang mengandung protein yang diinginkan.
Denaturasi termal selektif- pemanasan jangka pendek larutan protein, di mana beberapa pengotor protein yang terdenaturasi dapat dihilangkan (jika protein relatif stabil terhadap panas).
Mengasinkan. Protein yang berbeda mengendap pada konsentrasi garam yang berbeda dalam larutan. Dengan meningkatkan konsentrasi garam secara bertahap, dimungkinkan untuk memperoleh sejumlah fraksi terpisah dengan kandungan dominan protein terisolasi di salah satunya. Amonium sulfat paling sering digunakan untuk fraksinasi protein. Protein dengan kelarutan paling kecil akan mengendap pada konsentrasi garam rendah.
Filtrasi gel- metode menyaring molekul melalui butiran Sephadex yang membengkak (rantai polisakarida tiga dimensi dari dekstran yang memiliki pori-pori). Kecepatan protein melewati kolom yang diisi dengan Sephadex akan bergantung pada berat molekulnya: semakin kecil massa molekul protein, semakin mudah mereka menembus ke dalam butiran dan bertahan di sana lebih lama; semakin besar massanya, semakin cepat mereka terelusi dari butiran kolom.
Ultrasentrifugasi- metode yang melibatkan penempatan protein dalam tabung sentrifus ke dalam rotor ultrasentrifugasi. Ketika rotor berputar, laju sedimentasi protein sebanding dengan berat molekulnya: fraksi protein yang lebih berat terletak lebih dekat ke dasar tabung reaksi, fraksi protein yang lebih ringan terletak lebih dekat ke permukaan.
Elektroforesis- metode yang didasarkan pada perbedaan kecepatan pergerakan protein dalam medan listrik. Nilai ini sebanding dengan muatan protein. Elektroforesis protein dilakukan di atas kertas (dalam hal ini kecepatan pergerakan protein hanya sebanding dengan muatannya) atau dalam gel poliakrilamida dengan ukuran pori tertentu (kecepatan pergerakan protein sebanding dengan muatan dan berat molekulnya) .
Kromatografi pertukaran ion- metode fraksinasi berdasarkan pengikatan gugus protein terionisasi dengan gugus resin penukar ion yang bermuatan berlawanan (bahan polimer tidak larut). Kekuatan pengikatan protein pada resin sebanding dengan muatan protein. Protein yang teradsorpsi pada polimer penukar ion dapat tersapu dengan meningkatnya konsentrasi larutan NaCl; semakin rendah muatan protein, semakin rendah konsentrasi NaCl yang diperlukan untuk menghilangkan protein yang terikat pada gugus ionik resin.
Kromatografi afinitas- metode paling spesifik untuk mengisolasi protein individu. Ligan suatu protein terikat secara kovalen pada polimer inert. Ketika larutan protein dilewatkan melalui kolom dengan polimer, hanya protein spesifik untuk ligan tertentu yang teradsorpsi pada kolom karena pengikatan komplementer protein ke ligan.
Dialisis- metode yang digunakan untuk menghilangkan senyawa dengan berat molekul rendah dari larutan protein yang diisolasi. Metode ini didasarkan pada ketidakmampuan protein untuk melewati membran semipermeabel, tidak seperti zat dengan berat molekul rendah. Ini digunakan untuk memurnikan protein dari pengotor dengan berat molekul rendah, misalnya garam setelah penggaraman.
TUGAS KERJA EKSTRAKURIKULER
1. Isi tabelnya. 1.4.
Tabel 1.4. Analisis perbandingan struktur dan fungsi protein terkait - mioglobin dan hemoglobin
a) mengingat struktur pusat aktif Mb dan Hb. Apa peran radikal asam amino hidrofobik dalam pembentukan pusat aktif protein ini? Jelaskan struktur pusat aktif Mb dan Hb serta mekanisme perlekatan ligan padanya. Apa peran residu His F 8 dan His E 7 dalam berfungsinya pusat aktif Mv iHv?
b) sifat baru apa yang dimiliki protein oligomer, hemoglobin, yang berkerabat dekat dengan mioglobin monomer? Jelaskan peran perubahan kooperatif dalam konformasi protomer dalam molekul hemoglobin, pengaruh CO2 dan konsentrasi proton terhadap afinitas hemoglobin terhadap oksigen, serta peran 2,3-BPG dalam regulasi alosterik fungsi Hb .
2. Ciri-ciri pendamping molekuler, dengan memperhatikan hubungan antara struktur dan fungsinya.
3. Protein apa yang dikelompokkan ke dalam keluarga? Dengan menggunakan contoh keluarga imunoglobulin, identifikasi ciri-ciri struktural serupa dan fungsi terkait dari protein keluarga ini.
4. Protein individu yang dimurnikan seringkali diperlukan untuk tujuan biokimia dan pengobatan. Jelaskan sifat fisikokimia protein yang menjadi dasar metode pemisahan dan pemurniannya.
TUGAS PENGENDALIAN DIRI
1. Pilihlah jawaban yang benar.
Fungsi hemoglobin:
A. Pengangkutan O2 dari paru ke jaringan B. Pengangkutan H+ dari jaringan ke paru
B. Mempertahankan pH darah yang konstan D. Transportasi CO 2 dari paru-paru ke jaringan
D. Transportasi CO 2 dari jaringan ke paru-paru
2. Pilihlah jawaban yang benar. Liganα -protomer Hb adalah: A.Heme
B.Oksigen
B.CO G.2,3-BPG
D. β-Protomer
3. Pilihlah jawaban yang benar.
Hemoglobin berbeda dengan mioglobin:
A. Memiliki struktur kuaterner
B. Struktur sekunder hanya diwakili oleh heliks α
B. Milik protein kompleks
D. Berinteraksi dengan ligan alosterik D. Terikat secara kovalen dengan heme
4. Pilihlah jawaban yang benar.
Afinitas Hb terhadap O2 menurun:
A. Ketika satu molekul O 2 ditambahkan B. Ketika satu molekul O 2 dihilangkan
B. Saat berinteraksi dengan 2,3-BPG
D. Bila dilekatkan pada protomer H + D. Bila konsentrasi 2,3-BPG menurun
5. Cocok.
Tipe HB mempunyai ciri-ciri :
A. Dalam bentuk deoksi membentuk agregat fibrilar B. Berisi dua rantai α- dan dua rantai δ
B. Bentuk Hb yang dominan pada eritrosit dewasa D. Mengandung heme dengan Fe+3 pada pusat aktifnya
D. Berisi dua rantai α- dan dua γ 1. HbA 2.
6. Cocok.
Ligan Hb:
A. Berikatan dengan Hb di pusat alosterik
B. Memiliki afinitas yang sangat tinggi terhadap situs aktif Hb
B. Dengan bergabung, meningkatkan afinitas Hb terhadap O 2 G. Mengoksidasi Fe+ 2 menjadi Fe+ 3
D. Membentuk ikatan kovalen dengan F8 miliknya
7. Pilihlah jawaban yang benar.
Pendamping:
A. Protein terdapat di seluruh bagian sel
B. Sintesis meningkat di bawah tekanan
B. Berpartisipasi dalam hidrolisis protein terdenaturasi
D. Berpartisipasi dalam mempertahankan konformasi asli protein
D. Mereka menciptakan organel di mana konformasi protein terbentuk.
8. Cocokkan. Imunoglobulin:
A. Bentuk sekretorinya adalah pentamerik.
B. Kelas Ig yang menembus sawar plasenta
B. Ig - reseptor sel mast
D. Kelas utama Ig hadir dalam sekresi sel epitel. D. Reseptor limfosit B, aktivasi yang memastikan proliferasi sel
9. Pilihlah jawaban yang benar.
Imunoglobulin E:
A. Diproduksi oleh makrofag B. Memiliki rantai ε yang berat.
B. Tertanam dalam membran limfosit T
D. Bertindak sebagai reseptor antigen membran pada sel mast dan basofil
D. Bertanggung jawab atas reaksi alergi
10. Pilihlah jawaban yang benar.
Metode pemisahan protein didasarkan pada perbedaan berat molekulnya:
A. Filtrasi gel
B. Ultrasentrifugasi
B. Elektroforesis gel poliakrilamida D. Kromatografi penukar ion
D. Kromatografi afinitas
11. Pilih jawaban yang benar.
Metode pemisahan protein didasarkan pada perbedaan kelarutannya dalam air:
A. Filtrasi gel B. Pengasinan
B. Kromatografi penukar ion D. Kromatografi afinitas
D. Elektroforesis dalam gel poliakrilamida
STANDAR JAWABAN TERHADAP “TUGAS PENGENDALIAN DIRI”
1.A,B,C,D
2.A,B,C,D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7.A,B,D,D
8. 1-G; 2-B, 3-B
SYARAT DAN KONSEP DASAR
1. Protein oligomer, protomer, struktur protein kuaterner
2. Perubahan kooperatif dalam konformasi protomer
3. Efek Bohr
4. Regulasi alosterik fungsi protein, pusat alosterik dan efektor alosterik
5. Pendamping molekuler, protein kejutan panas
6. Keluarga protein (serin protease, imunoglobulin)
7. Hubungan IgM-, G-, E-, A-struktur-fungsi
8. Muatan total protein, titik isoelektrik protein
9. Elektroforesis
10. Pengasinan
11. Filtrasi gel
12. Kromatografi pertukaran ion
13. Ultrasentrifugasi
14. Kromatografi afinitas
15. Elektroforesis protein plasma darah
TUGAS UNTUK PEKERJAAN KELAS
1. Bandingkan ketergantungan derajat kejenuhan hemoglobin (Hb) dan mioglobin (Mb) dengan oksigen terhadap tekanan parsialnya dalam jaringan
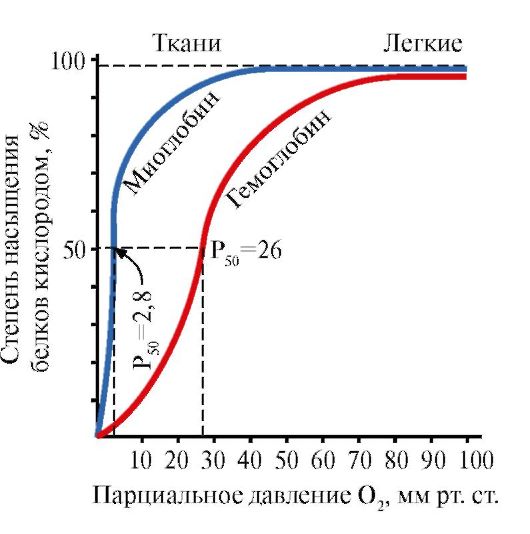 Beras. 1.34. Ketergantungan saturasi Mv danNHoksigen dari tekanan parsialnya
Beras. 1.34. Ketergantungan saturasi Mv danNHoksigen dari tekanan parsialnya
Harap dicatat bahwa bentuk kurva saturasi oksigen protein berbeda: untuk mioglobin - hiperbola, untuk hemoglobin - bentuk sigmoid.
1. Bandingkan nilai tekanan parsial oksigen dimana Mb dan Hb jenuh dengan O 2 sebesar 50%. Manakah dari protein berikut yang memiliki afinitas lebih tinggi terhadap O2?
2. Ciri-ciri struktural Mb apa yang menentukan afinitasnya yang tinggi terhadap O 2?
3. Ciri-ciri struktural HB apa yang memungkinkannya melepaskan O2 di kapiler jaringan istirahat (dengan tekanan parsial O2 yang relatif tinggi) dan secara tajam meningkatkan pelepasan ini pada otot yang bekerja? Sifat protein oligomer apa yang memberikan efek ini?
4. Hitung berapa jumlah O2 (dalam%) hemoglobin teroksigenasi yang diberikan kepada otot istirahat dan bekerja?
5. menarik kesimpulan tentang hubungan antara struktur suatu protein dan fungsinya.
2. Jumlah oksigen yang dilepaskan oleh hemoglobin di kapiler tergantung pada intensitas proses katabolik di jaringan (efek Bohr). Bagaimana perubahan metabolisme jaringan mengatur afinitas Hb terhadap O2? Pengaruh CO 2 dan H+ terhadap afinitas Hb terhadap O 2
 1. jelaskan efek Bohr.
1. jelaskan efek Bohr.
2. ke arah mana proses yang ditunjukkan pada diagram berlangsung:
a) di kapiler paru-paru;
b) di kapiler jaringan?
3. Apa arti fisiologis dari efek Bohr?
4. Mengapa interaksi Hb dengan H+ pada tempat yang jauh dari heme mengubah afinitas protein terhadap O 2?
3. Afinitas Hb terhadap O2 bergantung pada konsentrasi ligannya - 2,3-bifosfogliserat, yang merupakan pengatur alosterik afinitas Hb terhadap O2. Mengapa interaksi ligan pada lokasi yang jauh dari situs aktif mempengaruhi fungsi protein? Bagaimana 2,3-BPG mengatur afinitas Hb terhadap O2? Untuk mengatasi masalah tersebut, jawablah pertanyaan berikut:
1. di mana dan dari apa 2,3-bifosfogliserat (2,3-BPG) disintesis? Tulis rumusnya, tunjukkan muatan molekul ini.
2. Bentuk hemoglobin (oksi atau deoksi) manakah yang berinteraksi dengan BPG dan mengapa? Di bagian molekul Hb manakah interaksi terjadi?
3. ke arah manakah proses yang ditunjukkan pada diagram berlangsung?
a) di kapiler jaringan;
b) di kapiler paru-paru?
4. dimana konsentrasi kompleks harus lebih tinggi
Nv-2,3-BFG:
a) di kapiler otot saat istirahat,
b) di kapiler otot yang bekerja (asalkan konsentrasi BPG yang sama dalam eritrosit)?
5. Bagaimana afinitas HB terhadap oksigen berubah ketika seseorang beradaptasi dengan kondisi dataran tinggi, jika konsentrasi BPG dalam eritrosit meningkat? Apa signifikansi fisiologis dari fenomena ini?
4. Penghancuran 2,3-BPG selama penyimpanan darah yang diawetkan mengganggu fungsi HB. Bagaimana afinitas HB terhadap O 2 berubah dalam darah yang diawetkan jika konsentrasi 2,3-BPG dalam eritrosit dapat menurun dari 8 menjadi 0,5 mmol/l. Apakah mungkin untuk mentransfusikan darah tersebut ke pasien yang sakit parah jika konsentrasi 2,3-BPG pulih tidak lebih awal dari setelah tiga hari? Apakah mungkin mengembalikan fungsi sel darah merah dengan menambahkan 2,3-BPG ke dalam darah?
5. Ingat struktur molekul imunoglobulin paling sederhana. Apa peran imunoglobulin dalam sistem kekebalan? Mengapa Ig sering disebut bivalen? Bagaimana hubungan struktur Ig dengan fungsinya? (Jelaskan dengan menggunakan contoh kelas imunoglobulin.)
Sifat fisikokimia protein dan metode pemisahannya.
6. Bagaimana muatan bersih suatu protein mempengaruhi kelarutannya?
a) tentukan muatan total peptida pada pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) bagaimana muatan peptida ini berubah pada pH >7, pH<7, рН <<7?
c) berapa titik isoelektrik suatu protein (IEP) dan di lingkungan manakah letaknya?
IET dari peptida ini?
d) pada nilai pH berapa kelarutan peptida ini paling kecil akan diamati.
7. Mengapa susu asam, tidak seperti susu segar, “mengental” ketika direbus (yaitu, kasein protein susu mengendap)? Pada susu segar, molekul kasein mempunyai muatan negatif.
8. Filtrasi gel digunakan untuk memisahkan protein individu. Campuran yang mengandung protein A, B, C dengan berat molekul masing-masing sebesar 160.000, 80.000 dan 60.000, dianalisis dengan filtrasi gel (Gbr. 1.35). Butiran gel yang membengkak dapat ditembus oleh protein dengan berat molekul kurang dari 70.000. Prinsip apa yang mendasari metode pemisahan ini? Grafik manakah yang mencerminkan hasil fraksinasi dengan benar? Tunjukkan urutan pelepasan protein A, B, dan C dari kolom.
 Beras. 1.35. Menggunakan Filtrasi Gel untuk Pemisahan Protein
Beras. 1.35. Menggunakan Filtrasi Gel untuk Pemisahan Protein
9. Pada Gambar. 1.36, A menunjukkan diagram elektroforesis di atas kertas protein serum darah orang sehat. Jumlah relatif fraksi protein yang diperoleh dengan metode ini adalah: albumin 54-58%, α 1 -globulin 6-7%, α 2 -globulin 8-9%, β-globulin 13%, γ-globulin 11-12% .
 Beras. 1.36 Elektroforesis pada kertas protein plasma darah orang sehat (A) dan pasien (B)
Beras. 1.36 Elektroforesis pada kertas protein plasma darah orang sehat (A) dan pasien (B)
I - γ-globulin; II - β-globulin; AKU AKU AKU -α 2 -globulin; IV -α 2 -globulin; V - albumin
Banyak penyakit disertai dengan perubahan kuantitatif komposisi protein serum (disproteinemia). Sifat perubahan ini diperhitungkan saat membuat diagnosis dan menilai tingkat keparahan dan stadium penyakit.
Menggunakan data yang diberikan dalam tabel. 1.5, tebak penyakitnya, yang ditandai dengan profil elektroforesis yang ditunjukkan pada Gambar. 1.36.
Tabel 1.5. Perubahan konsentrasi protein serum pada patologi
Tupai- senyawa organik dengan berat molekul tinggi yang terdiri dari residu asam α-amino.
DI DALAM komposisi protein termasuk karbon, hidrogen, nitrogen, oksigen, belerang. Beberapa protein membentuk kompleks dengan molekul lain yang mengandung fosfor, besi, seng, dan tembaga.
Protein memiliki berat molekul besar: albumin telur - 36.000, hemoglobin - 152.000, miosin - 500.000. Sebagai perbandingan: berat molekul alkohol adalah 46, asam asetat - 60, benzena - 78.
Komposisi asam amino protein
Tupai- polimer non-periodik, yang monomernya adalah asam α-amino. Biasanya, 20 jenis asam α-amino disebut monomer protein, meskipun lebih dari 170 di antaranya ditemukan di sel dan jaringan.
Tergantung pada apakah asam amino dapat disintesis dalam tubuh manusia dan hewan lain, ada: asam amino nonesensial- dapat disintesis; asam amino esensial- tidak dapat disintesis. Asam amino esensial harus dipasok ke tubuh melalui makanan. Tumbuhan mensintesis semua jenis asam amino.
Tergantung pada komposisi asam aminonya, protein adalah: lengkap- mengandung seluruh rangkaian asam amino; cacat- beberapa asam amino hilang dalam komposisinya. Jika protein hanya terdiri dari asam amino, maka disebut sederhana. Jika protein mengandung, selain asam amino, komponen non-asam amino (gugus prostetik), maka disebut kompleks. Gugus prostetik dapat diwakili oleh logam (metaloprotein), karbohidrat (glikoprotein), lipid (lipoprotein), asam nukleat (nukleoprotein).
Semua mengandung asam amino: 1) gugus karboksil (-COOH), 2) gugus amino (-NH 2), 3) radikal atau gugus R (sisa molekul). Struktur radikal berbeda untuk berbagai jenis asam amino. Tergantung pada jumlah gugus amino dan gugus karboksil yang termasuk dalam asam amino, mereka dibedakan: asam amino netral memiliki satu gugus karboksil dan satu gugus amino; asam amino basa memiliki lebih dari satu gugus amino; asam amino yang bersifat asam mempunyai lebih dari satu gugus karboksil.
Asam amino adalah senyawa amfoter, karena dalam larutan mereka dapat bertindak sebagai asam dan basa. Dalam larutan air, asam amino ada dalam bentuk ionik yang berbeda.
Ikatan peptida
Peptida- zat organik yang terdiri dari residu asam amino yang dihubungkan oleh ikatan peptida.
Pembentukan peptida terjadi sebagai akibat dari reaksi kondensasi asam amino. Ketika gugus amino dari satu asam amino berinteraksi dengan gugus karboksil asam amino lainnya, terjadi ikatan kovalen nitrogen-karbon di antara keduanya, yang disebut peptida. Tergantung pada jumlah residu asam amino yang termasuk dalam peptida, ada dipeptida, tripeptida, tetrapeptida dll. Pembentukan ikatan peptida dapat terjadi berkali-kali. Hal ini mengarah pada pembentukan polipeptida. Di salah satu ujung peptida terdapat gugus amino bebas (disebut ujung-N), dan di ujung lainnya terdapat gugus karboksil bebas (disebut ujung-C).

Organisasi spasial molekul protein
Kinerja fungsi spesifik tertentu oleh protein bergantung pada konfigurasi spasial molekulnya; selain itu, secara energetik tidak menguntungkan bagi sel untuk menyimpan protein dalam bentuk tidak terlipat, dalam bentuk rantai, oleh karena itu rantai polipeptida mengalami pelipatan, memperoleh suatu rantai. struktur tiga dimensi tertentu, atau konformasi. Ada 4 level organisasi spasial protein.
Struktur protein primer- urutan susunan residu asam amino pada rantai polipeptida penyusun molekul protein. Ikatan antar asam amino merupakan ikatan peptida.

Jika suatu molekul protein hanya terdiri dari 10 residu asam amino, maka jumlah varian molekul protein yang mungkin secara teoritis berbeda dalam urutan pergantian asam amino adalah 10 20. Memiliki 20 asam amino, Anda dapat membuat kombinasi yang lebih beragam. Sekitar sepuluh ribu protein berbeda telah ditemukan di dalam tubuh manusia, yang berbeda satu sama lain dan dari protein organisme lain.
Struktur utama molekul proteinlah yang menentukan sifat molekul protein dan konfigurasi spasialnya. Penggantian hanya satu asam amino dengan asam amino lain dalam rantai polipeptida menyebabkan perubahan sifat dan fungsi protein. Misalnya, penggantian asam amino glutamat keenam dengan valin pada subunit β hemoglobin menyebabkan fakta bahwa molekul hemoglobin secara keseluruhan tidak dapat menjalankan fungsi utamanya - transportasi oksigen; Dalam kasus seperti itu, orang tersebut menderita penyakit yang disebut anemia sel sabit.
Struktur sekunder- pelipatan rantai polipeptida yang teratur menjadi spiral (terlihat seperti pegas yang memanjang). Putaran heliks tersebut diperkuat oleh ikatan hidrogen yang timbul antara gugus karboksil dan gugus amino. Hampir semua gugus CO dan NH berperan dalam pembentukan ikatan hidrogen. Mereka lebih lemah dari peptida, namun, diulang berkali-kali, memberikan stabilitas dan kekakuan pada konfigurasi ini. Pada tingkat struktur sekunder terdapat protein: fibroin (sutra, jaring laba-laba), keratin (rambut, kuku), kolagen (tendon).



Struktur tersier- pengepakan rantai polipeptida menjadi butiran-butiran yang dihasilkan dari pembentukan ikatan kimia (hidrogen, ionik, disulfida) dan pembentukan interaksi hidrofobik antara radikal residu asam amino. Peran utama dalam pembentukan struktur tersier dimainkan oleh interaksi hidrofilik-hidrofobik. Dalam larutan berair, radikal hidrofobik cenderung bersembunyi dari air, berkelompok di dalam globul, sedangkan radikal hidrofilik, akibat hidrasi (interaksi dengan dipol air), cenderung muncul di permukaan molekul. Pada beberapa protein, struktur tersier distabilkan oleh ikatan kovalen disulfida yang terbentuk antara atom belerang dari dua residu sistein. Pada tingkat struktur tersier terdapat enzim, antibodi, dan beberapa hormon.

Struktur Kuarter karakteristik protein kompleks yang molekulnya dibentuk oleh dua atau lebih butiran. Subunit ditahan dalam molekul melalui interaksi ionik, hidrofobik, dan elektrostatis. Kadang-kadang, selama pembentukan struktur kuaterner, ikatan disulfida terjadi antar subunit. Protein yang paling banyak dipelajari dengan struktur kuaterner adalah hemoglobin. Ini dibentuk oleh dua subunit α (141 residu asam amino) dan dua subunit β (146 residu asam amino). Terkait dengan setiap subunit adalah molekul heme yang mengandung besi.
Jika karena alasan tertentu konformasi spasial protein menyimpang dari normal, protein tersebut tidak dapat menjalankan fungsinya. Misalnya, penyebab “penyakit sapi gila” (ensefalopati spongiform) adalah konformasi abnormal prion, protein permukaan sel saraf.
Sifat-sifat protein
Komposisi asam amino dan struktur molekul protein menentukannya properti. Protein menggabungkan sifat basa dan asam, ditentukan oleh radikal asam amino: semakin asam asam amino dalam suatu protein, semakin besar sifat asamnya. Kemampuan menyumbang dan menambah H+ ditentukan sifat penyangga protein; Salah satu buffer yang paling kuat adalah hemoglobin dalam sel darah merah, yang menjaga pH darah pada tingkat yang konstan. Ada protein larut (fibrinogen), dan ada protein tidak larut yang menjalankan fungsi mekanis (fibroin, keratin, kolagen). Ada protein yang aktif secara kimia (enzim), ada protein yang tidak aktif secara kimia yang tahan terhadap berbagai kondisi lingkungan, dan ada pula yang sangat tidak stabil.
Faktor eksternal (panas, radiasi ultraviolet, logam berat dan garamnya, perubahan pH, radiasi, dehidrasi)
dapat menyebabkan terganggunya organisasi struktural molekul protein. Proses hilangnya konformasi tiga dimensi yang melekat pada molekul protein tertentu disebut denaturasi. Penyebab terjadinya denaturasi adalah putusnya ikatan yang menstabilkan struktur protein tertentu. Pada awalnya, ikatan yang paling lemah akan putus, dan ketika kondisi menjadi lebih ketat, ikatan yang lebih kuat pun akan putus. Oleh karena itu, pertama-tama struktur kuaterner, kemudian struktur tersier dan sekunder hilang. Perubahan konfigurasi spasial menyebabkan perubahan sifat protein dan, sebagai akibatnya, protein tidak dapat menjalankan fungsi biologis bawaannya. Jika denaturasi tidak disertai dengan kerusakan struktur primer, maka hal itu mungkin terjadi dapat dibalik, dalam hal ini, terjadi pemulihan sendiri karakteristik konformasi protein. Misalnya, protein reseptor membran mengalami denaturasi seperti itu. Proses pemulihan struktur protein setelah denaturasi disebut renaturasi. Jika pemulihan konfigurasi spasial protein tidak mungkin dilakukan, maka disebut denaturasi tidak dapat diubah.
Fungsi protein
| Fungsi | Contoh dan penjelasannya |
|---|---|
| Konstruksi | Protein terlibat dalam pembentukan struktur seluler dan ekstraseluler: mereka adalah bagian dari membran sel (lipoprotein, glikoprotein), rambut (keratin), tendon (kolagen), dll. |
| Mengangkut | Protein darah hemoglobin mengikat oksigen dan mengangkutnya dari paru-paru ke seluruh jaringan dan organ, dan darinya mentransfer karbon dioksida ke paru-paru; Komposisi membran sel mencakup protein khusus yang memastikan transfer zat dan ion tertentu secara aktif dan selektif dari sel ke lingkungan luar dan sebaliknya. |
| Peraturan | Hormon protein berperan dalam pengaturan proses metabolisme. Misalnya, hormon insulin mengatur kadar glukosa darah, meningkatkan sintesis glikogen, dan meningkatkan pembentukan lemak dari karbohidrat. |
| Protektif | Menanggapi penetrasi protein asing atau mikroorganisme (antigen) ke dalam tubuh, protein khusus terbentuk - antibodi yang mampu mengikat dan menetralisirnya. Fibrin, terbentuk dari fibrinogen, membantu menghentikan pendarahan. |
| Motor | Protein kontraktil aktin dan miosin memberikan kontraksi otot pada hewan multiseluler. |
| Sinyal | Di dalam membran permukaan sel terdapat molekul protein yang mampu mengubah struktur tersiernya sebagai respons terhadap faktor lingkungan, sehingga menerima sinyal dari lingkungan luar dan mengirimkan perintah ke sel. |
| Penyimpanan | Di dalam tubuh hewan, protein biasanya tidak disimpan, kecuali albumin telur dan kasein susu. Namun berkat protein, beberapa zat dapat disimpan di dalam tubuh; misalnya, selama pemecahan hemoglobin, zat besi tidak dikeluarkan dari tubuh, tetapi disimpan, membentuk kompleks dengan protein feritin. |
| Energi | Ketika 1 g protein dipecah menjadi produk akhir, 17,6 kJ dilepaskan. Pertama, protein terurai menjadi asam amino, dan kemudian menjadi produk akhir - air, karbon dioksida, dan amonia. Namun, protein digunakan sebagai sumber energi hanya jika sumber lain (karbohidrat dan lemak) telah habis. |
| Katalis | Salah satu fungsi terpenting protein. Disediakan oleh protein - enzim yang mempercepat reaksi biokimia yang terjadi di dalam sel. Misalnya, ribulosa bifosfat karboksilase mengkatalisis fiksasi CO2 selama fotosintesis. |
Enzim
Enzim, atau enzim, adalah kelas protein khusus yang merupakan katalis biologis. Berkat enzim, reaksi biokimia terjadi dengan kecepatan luar biasa. Laju reaksi enzimatik puluhan ribu kali (dan terkadang jutaan) lebih tinggi daripada laju reaksi yang melibatkan katalis anorganik. Zat yang menjadi tempat kerja enzim disebut substrat.
Enzim adalah protein globular, fitur struktural enzim dapat dibagi menjadi dua kelompok: sederhana dan kompleks. Enzim sederhana adalah protein sederhana, yaitu hanya terdiri dari asam amino. Enzim kompleks adalah protein kompleks, mis. Selain bagian protein, mereka mengandung kelompok yang bersifat non-protein - kofaktor. Beberapa enzim menggunakan vitamin sebagai kofaktor. Molekul enzim mengandung bagian khusus yang disebut pusat aktif. Pusat aktif- bagian kecil dari enzim (dari tiga hingga dua belas residu asam amino), di mana terjadi pengikatan substrat atau substrat untuk membentuk kompleks enzim-substrat. Setelah reaksi selesai, kompleks enzim-substrat terurai menjadi enzim dan produk reaksi. Beberapa enzim memiliki (kecuali aktif) pusat alosterik- area di mana pengatur kecepatan enzim dipasang ( enzim alosterik).

Reaksi katalisis enzimatik dicirikan oleh: 1) efisiensi tinggi, 2) selektivitas dan arah kerja yang ketat, 3) spesifisitas substrat, 4) regulasi yang halus dan tepat. Spesifisitas substrat dan reaksi katalisis enzimatik dijelaskan oleh hipotesis E. Fischer (1890) dan D. Koshland (1959).
E. Fisher (hipotesis kunci-kunci) menyarankan bahwa konfigurasi spasial pusat aktif enzim dan substrat harus sesuai satu sama lain. Substrat dibandingkan dengan “kunci”, enzim dengan “gembok”.
D. Koshland (hipotesis sarung tangan) menyarankan bahwa korespondensi spasial antara struktur substrat dan pusat aktif enzim tercipta hanya pada saat interaksinya satu sama lain. Hipotesis ini disebut juga hipotesis korespondensi yang diinduksi.
Laju reaksi enzimatik bergantung pada: 1) suhu, 2) konsentrasi enzim, 3) konsentrasi substrat, 4) pH. Perlu ditekankan bahwa karena enzim adalah protein, aktivitasnya paling tinggi dalam kondisi fisiologis normal.
Kebanyakan enzim hanya dapat bekerja pada suhu antara 0 dan 40°C. Dalam batas ini, laju reaksi meningkat kira-kira 2 kali lipat setiap kenaikan suhu 10 °C. Pada suhu di atas 40°C, protein mengalami denaturasi dan aktivitas enzim menurun. Pada suhu mendekati titik beku, enzim menjadi tidak aktif.
Dengan bertambahnya jumlah substrat, laju reaksi enzimatik meningkat hingga jumlah molekul substrat sama dengan jumlah molekul enzim. Dengan peningkatan lebih lanjut dalam jumlah substrat, kecepatannya tidak akan meningkat, karena pusat aktif enzim sudah jenuh. Peningkatan konsentrasi enzim menyebabkan peningkatan aktivitas katalitik, karena lebih banyak molekul substrat yang mengalami transformasi per satuan waktu.

Untuk setiap enzim, terdapat nilai pH optimal yang menunjukkan aktivitas maksimum (pepsin - 2.0, amilase ludah - 6.8, lipase pankreas - 9.0). Pada nilai pH yang lebih tinggi atau lebih rendah, aktivitas enzim menurun. Dengan perubahan pH yang tiba-tiba, enzim mengalami denaturasi.
Kecepatan kerja enzim alosterik diatur oleh zat yang menempel pada pusat alosterik. Jika zat tersebut mempercepat suatu reaksi maka disebut aktivator, jika mereka melambat - penghambat.
Klasifikasi enzim
Menurut jenis transformasi kimia yang dikatalisisnya, enzim dibagi menjadi 6 kelas:
- oksireduktase(transfer atom hidrogen, oksigen atau elektron dari satu zat ke zat lain - dehidrogenase),
- transferase(pemindahan gugus metil, asil, fosfat atau amino dari satu zat ke zat lain - transaminase),
- hidrolase(reaksi hidrolisis di mana dua produk terbentuk dari substrat - amilase, lipase),
- liase(penambahan non-hidrolitik pada substrat atau pelepasan sekelompok atom darinya, dalam hal ini ikatan C-C, C-N, C-O, C-S dapat diputus - dekarboksilase),
- isomerase(penataan ulang intramolekul - isomerase),
- ligase(hubungan dua molekul akibat pembentukan ikatan C-C, C-N, C-O, C-S - sintetase).
Kelas pada gilirannya dibagi lagi menjadi subkelas dan subsubkelas. Dalam klasifikasi internasional saat ini, setiap enzim memiliki kode tertentu, terdiri dari empat angka yang dipisahkan oleh titik. Angka pertama adalah kelas, yang kedua adalah subkelas, yang ketiga adalah subsubkelas, yang keempat adalah nomor urut enzim pada subkelas tersebut, misalnya kode arginase adalah 3.5.3.1.
Pergi ke kuliah nomor 2"Struktur dan fungsi karbohidrat dan lipid"
Pergi ke kuliah nomor 4"Struktur dan fungsi asam nukleat ATP"
Protein (protein) adalah senyawa polimer bermolekul tinggi yang bersifat peptida (asam poliheteroamino).
Struktur utama protein adalah urutan residu asam amino bergantian dalam rantai polipeptida (PPC).
Struktur utama protein adalah kovalen struktur, karena didasarkan pada peptida hubungan antara gugus a-amino dan a-karboksil asam amino. Akibatnya rantai polipeptida tidak bercabang.
Kerangka (tulang punggung, kerangka) rantai polipeptida terdiri dari elemen struktur yang berulang secara teratur
Rantai polipeptida mempunyai sifat vektorialitas, arah rantai adalah dari ujung N (awal rantai) sampai ujung C (ujung rantai), ujung N merupakan ujung tempat gugus a-amino bebas berada. terletak. Terminal-C adalah ujung yang mengandung gugus a-karboksil bebas. Urutan asam amino protein ditetapkan mulai dari ujung N, menggunakan nama asam amino yang disingkat tiga huruf, misalnya: gly-ala-cis-pro. Penunjukan satu huruf untuk residu asam amino dalam suatu protein juga dapat digunakan.
Ujung N dan C protein dapat dimodifikasi. Gugus amino di terminal-N dapat diasetilasi, diformilasi, atau dimetilasi. Pada sejumlah protein, residu N-terminal adalah pirolidon karbonat (piroglutamat), yang tidak mengandung gugus amino bebas. Terminal-C mungkin berada di tengah. Modifikasi terminal-C lebih jarang terjadi dibandingkan modifikasi terminal-N.
Koefisien polikondensasi protein bervariasi antara 50 hingga 2500. Biasanya, suatu protein mengandung 100-300 residu asam amino. Karena berat molekul rata-rata residu asam amino tunggal adalah sekitar 110 Da, berat molekul protein bervariasi dari 6000 hingga jutaan Da.
Setiap protein individu memiliki struktur primer yang unik. Protein pertama yang struktur utamanya terbentuk adalah insulin. Sanger berhasil melakukan ini. Strateginya adalah sebagai berikut. Dia pertama-tama memisahkan dua rantai polipeptida dan kemudian melakukan pembelahan enzimatik spesifiknya menjadi peptida kecil yang mengandung urutan yang tumpang tindih. Residu N-terminal kemudian diidentifikasi menggunakan 1-fluoro-2,4-dinitrobenzene. Selain itu, ia menentukan komposisi asam amino peptida dan akhirnya dapat menentukan strukturnya dengan membandingkan urutan peptida yang tumpang tindih. Secara umum, strategi Sanger masih tetap penting hingga saat ini. Namun, pendekatan lain juga telah diusulkan. Edman mengembangkan metode prosedur otomatis untuk pembelahan berurutan dan identifikasi residu asam amino terminal-N. Analisis difraksi sinar-X dapat digunakan untuk menguraikan struktur primer. Urutan residu asam amino dapat ditentukan dari urutan nukleotida RNA pembawa pesan.
Saat ini, struktur utama lebih dari 2000 protein telah terbentuk. Secara teoritis, jumlah varian struktur primer protein tidak terbatas. Bahkan untuk polipeptida yang terdiri dari 20 asam amino berbeda, jumlah urutan yang mungkin adalah 20´10 18 . Di alam yang hidup, hanya sebagian kecil dari kemungkinan urutan yang terwujud, yang jumlah totalnya pada semua jenis organisme hidup diperkirakan 10 10 -10 12.
Struktur utama protein ditentukan secara genetik, yaitu. Urutan asam amino dalam suatu protein ditentukan oleh urutan nukleotida di dalamnya DNA. Distorsi urutan nukleotida DNA menyebabkan munculnya protein abnormal dengan perubahan sifat biologis, yang merupakan penyebab patologi molekuler. Secara khusus, anemia sel sabit disebabkan oleh mutasi titik pada gen yang mengontrol rantai hemoglobin b. Konsekuensinya adalah tergantinya residu glutamat pada posisi ke-6 rantai b dengan valin. Substitusi ini mengakibatkan hilangnya satu muatan negatif pada masing-masing dua rantai b, yang menyebabkan perubahan konformasi hemoglobin dan hilangnya fungsi biologisnya.
Protein homolog adalah protein yang melakukan fungsi yang sama pada spesies berbeda. Contohnya adalah hemoglobin: pada semua vertebrata ia melakukan fungsi yang sama terkait dengan transportasi oksigen. Protein homolog dicirikan oleh adanya asam amino yang sama di banyak posisi. Ternyata, jumlah residu asam amino yang membedakan protein homolog sebanding dengan perbedaan filogenetik antara spesies ini. Misalnya, molekul sitokrom C kuda dan ragi berbeda dalam 48 residu asam amino, sedangkan molekul yang sama pada ayam dan bebek hanya berbeda dalam 2 residu. Adapun sitokrom C dari ayam dan kalkun memiliki urutan asam amino yang identik. Informasi tentang jumlah perbedaan urutan asam amino protein homolog dari spesies yang berbeda digunakan untuk menyusun peta evolusi yang mencerminkan tahapan berturut-turut kemunculan dan perkembangan berbagai spesies hewan dan tumbuhan dalam proses evolusi.
Kuliah 3. Struktur protein
Definisi:
Protein adalah polimer tidak beraturan yang monomernya L- asam amino.
Asam amino
Ada dua bentuk stereoisomer di alam: L (kidal) dan D (dekstrorotatori). Di samping itu L -asam amino yang membentuk protein ada di dalam tubuh D -asam amino yang tidak termasuk dalam protein.
Rumus umum asam amino ditunjukkan pada gambar.
Hal ini berlaku untuk 19 dari 20 asam amino yang ditemukan dalam protein. Selain 19 asam amino ini, protein mengandung satu asam amino asam imino - prolin
Semua asam amino mengandung α -kelompok amino. Oleh karena itu namanya – “asam α-amino”. Dalam prolin – α- imino kelompok.
Klasifikasi asam amino penyusun protein menurut prinsip polaritas (non polaritas) radikal.
1. Radikal non-polar atau hidrofobik.
Alifatik - alanin, valin, leusin, isoleusin. Mengandung belerang metionin Aromatik - fenilalanin, triptofan. asam imino prolin
2. Radikal yang bersifat polar namun tidak bermuatan. Glisin.
Asam hidroksiamino - serin, treonin, tirosin. Mengandung gugus sulfhidril sistein. Mengandung gugus Amida: asparagin, glutamin.
3. Radikal bermuatan negatif. Asam aspartat, asam glutamat.
4. Radikal bermuatan positif. Lisin, arginin, histidin.
Struktur protein primer
Definisi:
Struktur utama suatu protein adalah urutan residu asam amino dalam rantai polipeptida.
Asam amino digabungkan untuk membentuk polipeptida menggunakan ikatan kovalen (amida).
Tripeptida yang terdiri dari tiga asam amino berbeda memiliki kemungkinan 3! = 6 struktur primer yang berbeda.
Oligopeptida yang terdiri dari dua puluh asam amino berbeda memiliki keragaman struktur primer 20!, yang berarti 2x10 18.
Keragaman struktur primer suatu protein berukuran rata-rata (kira-kira 500 asam amino) sudah berjumlah sekitar 20.500 varian (jika semua asam amino disajikan dalam rasio ekuimolar).
PadaBelum ada, dan tidak akan ada, dua orang di Bumi dengan kumpulan protein yang persis sama.
Struktur sekunder protein
Definisi:
Struktur sekunder suatu protein adalah struktur rantai polipeptida yang teratur, yang disebabkan oleh ikatan hidrogen antara C=O dan N- Hasam amino yang berbeda.
Struktur sekunder dapat teratur (α-helix) atau tidak teratur (struktur β-sheet). Dalam α-heliks kelompok NH n Residu asam amino ke-th berinteraksi dengan gugus C=O dari residu asam amino (n-4). Terdapat 3,6 residu asam amino per putaran β-heliks dengan diameter 10,1 Å. Periode identitas α-heliks beraturan adalah 18 asam amino (5 putaran). Pengganggu α-helix biasa adalah yang utama prolin. Efek terpenting kedua diberikan oleh radikal bermuatan sama yang terletak di dekatnya.
Lipatan β dapat dibentuk tidak hanya oleh polipeptida tunggal, tetapi juga oleh polipeptida tetangga yang merupakan bagian dari satu protein.
Alfa atau beta alami murni - protein tidak ada.
Struktur tersier protein
Definisi
Struktur tersier suatu protein adalah konformasi spasial suatu polipeptida yang mempunyai struktur sekunder dan ditentukan oleh interaksi antar radikal.
Ada empat jenis interaksi antar radikal.
Jenis interaksi antar radikal
1 . Kovalen Komunikasi antar sisa dua sistein (disulfida jembatan).
2. Interaksi ionik (elektrostatik) antara residu asam amino yang bermuatan berlawanan (tiga radikal dengan tanda “+” dan dua dengan tanda “-”).
Misalnya, gugus ε-amino yang bermuatan positif dari lisin (- NH3 +) tertarik oleh gugus karboksil bermuatan negatif – (COO-) glutamat atau asam aspartat.
3. Ikatan hidrogen.
Semua asam amino yang mempunyai gugus hidroksil,amida atau karboksil terlibat.
4. Interaksi hidrofobik . Terbentuk antara radikal non-polar dalam lingkungan berair. 8 asam amino terlibat (kelas satu).
Struktur tersier suatu protein sepenuhnya ditentukan oleh struktur primernya, yaitu urutan asam amino, yang pada gilirannya ditentukan oleh kode genetik.
Interaksi hidrofobik sangat menentukan karena non-selektivitas (non-spesifisitas) dan kelimpahannya.
Kebanyakan protein memiliki inti hidrofobik.
Struktur protein kuarter
Definisi: Struktur kuaterner suatu protein adalah agregasi dua atau lebih rantai polipeptida yang memiliki struktur tersier menjadi komposisi oligomer yang signifikan secara fungsional.
Ikatan yang membentuk dan memelihara struktur kuaterner sama dengan ikatan yang membentuk struktur tersier, kecuali ikatan hidrofobik.
N Ujung - dari rantai beta mengandung asam glutamat polar ("-" bermuatan). Pada pasien dengan anemia sel sabit, valin nonpolar digunakan sebagai gantinya.Dari 574 asam amino, 2 diganti.
Hemoglobin tersebut kehilangan kelarutannya dan terbentuk endapan berserat, sehingga merusak sel darah merah.
Anemia sel sabit adalah penyakit genetik. Penyebabnya adalah penggantian satu nukleotida saja pada gen yang mengkode rantai hemoglobin B. Anak-anak yang homozigot resesif untuk alel ini tidak dapat hidup sampai usia dua tahun. Heterozigot memiliki 85% sel darah merah normal dan 15% sel darah merah rusak. Homozigot dominan menderita malaria, sedangkan heterozigot tidak menderita malaria.
Protein globular dan fibrilar
95% protein memiliki inti hidrofobik. 5% protein fibrilar.
Sebagian besar protein globular dapat larut. Kebanyakan fibrillar tidak larut (α-keratin - mereka menyumbang hampir seluruh berat kering rambut, wol, tanduk, kuku, kuku, sisik, bulu; kolagen - protein tendon, tulang rawan; fibroin - protein sutra).
Protein fibrilar mengandung lebih banyak asam amino bermuatan daripada asam amino globular - rantai individu dapat larut, dan kompleksnya bersifat non-polar dan tidak larut.
