§ 2. Componentele principale ale unei celule eucariote
Celulele eucariote (Fig. 8 și 9) sunt organizate mult mai complexe decât cele procariote. Ele sunt foarte diverse ca mărime (de la câțiva micrometri la câțiva centimetri), ca formă și ca caracteristici structurale (Fig. 10).
Orez. 8. Structura unei celule eucariote. Schema generalizata
Orez. 9. Structura celulei conform microscopiei electronice
Orez. 10. Diverse celule eucariote: 1 – epiteliale; 2 – sânge (e – eritroită, / – leukită); 3 – cartilaj; 4 – oase; 5 – musculatura neteda; 6 – țesut conjunctiv; 7 – celule nervoase; 8 – fibra musculara striata
Cu toate acestea, organizarea generală și prezența componentelor fundamentale sunt aceleași în toate celulele eucariote (Fig. 11).
Orez. 11. Celula eucariotă (diagrama)
Plasmalemma (membrana celulară exterioară). Baza membranei plasmatice, ca și alte membrane din celule (de exemplu, mitocondrii, plastide etc.), este un strat de lipide care are două rânduri de molecule (Fig. 12). Deoarece moleculele de lipide sunt polare (un pol este hidrofil, adică atras de apă, iar celălalt este hidrofob, adică respins de apă), ele sunt aranjate într-o anumită ordine. Capetele hidrofile ale moleculelor unui strat sunt îndreptate spre mediul apos - în citoplasma celulei, iar celălalt strat - spre exterior din celulă - către substanța intercelulară (la organismele pluricelulare) sau mediul apos (la organismele unicelulare). ).
Orez. 12. Structura membranei celulare după modelul mozaic fluid. Proteinele și glicoproteinele sunt scufundate într-un strat dublu de molecule de lipide, cu capetele lor hidrofile (cercurile) îndreptate spre exterior și capetele lor hidrofobe (linii ondulate) îndreptate adânc în membrană.
Moleculele de proteine sunt încorporate mozaic într-un strat bimolecular de lipide. La exteriorul celulei animale, moleculele de polizaharide sunt atașate de lipidele și moleculele proteice ale plasmalemei, formând glicolipide și glicoproteine.
Acest agregat formează un strat glicocalix. Asociat cu el funcția receptorului membrane plasmatice (vezi mai jos); poate acumula si diverse substante folosite de celula. În plus, glicocalixul sporește stabilitatea mecanică a plasmalemei.
În celulele plantelor și ciupercilor există și un perete celular care joacă un rol de susținere și de protecție. La plante este alcătuită din celuloză, iar la ciuperci este făcută din chitină.
Membrana celulară exterioară îndeplinește o serie de funcții, inclusiv:
♦ mecanic(sprijinirea, formarea);
♦ bariera-transport(permeabilitatea selectivă la diferite substanțe: intrarea în celulă a substanțelor necesare și îndepărtarea celor inutile și nocive);
♦ receptor(determinarea diferitelor substanțe chimice găsite în imediata apropiere a celulei; percepția semnalelor sub formă de hormoni; recunoașterea proteinei „străine” de către celulele sistemului imunitar etc.).
Schimbul de substanțe între celulă și mediu se realizează în moduri diferite - pasiv și activ.
Moleculele de apă și diverși ioni pasiv (datorită difuziei, osmozei), fără ca celula să consume energie, intră prin pori speciali - aceștia sunt transport pasiv. Macromolecule precum proteinele, polizaharidele, chiar și celulele întregi, sunt furnizate de fagocitozăȘi pinocitoza cu consum de energie - transport activ.
Prin fagocitoză, celulele întregi sau particulele mari sunt înghițite (de exemplu, gândiți-vă la nutriția în amibe sau la fagocitoza de către celulele sanguine protectoare ale bacteriilor). În timpul pinocitozei, particulele mici sau picăturile unei substanțe lichide sunt absorbite. Comun pentru ambele procese este faptul că substanțele absorbite sunt înconjurate de o membrană exterioară invaginată pentru a forma o vacuola, care apoi se deplasează adânc în citoplasma celulei.
Exocitoza este un proces (fiind și transport activ) opus în direcție fagocitozei și pinocitozei (Fig. 13). Cu ajutorul acestuia, alimentele nedigerate rămân în protozoare sau pot fi îndepărtate substanțele biologic active formate în celula secretorie.
Citoplasma. Citoplasma este conținutul celulei, limitat de plasmalemă, cu excepția nucleului. Contine substanță fundamentală (hialoplasmă), organeleȘi incluziuni.
Hialoplasma- un lichid vâscos care poate fi într-o stare de oricare Zola(lichid), sau gel(Ca un jeleu).
Dacă este necesar, citoplasma este capabilă să treacă reversibil de la o stare la alta. De exemplu, în timpul mișcării amoeboidului (amintiți-vă secțiunea „Protozoare” din cursul de zoologie), în timpul formării unui pseudopod, au loc tranziții rapide ale citoplasmei de la gel la sol și invers. Acest lucru se datorează prezenței în citoplasmă a unui număr mare de molecule de proteine filamentoase actina. Când se conectează între ele pentru a forma o rețea tridimensională, citoplasma este în stare de gel, iar când rețeaua se dezintegrează, este în stare de sol.
Hialoplasma conține diverse substanțe - enzime, proteine, carbohidrați, grăsimi și altele, organice și minerale. Aici au loc diferite procese chimice - descompunerea substanțelor, sinteza și modificările lor (modificări).
Organoizi. Acestea sunt componente permanente ale unei celule cu o anumită structură și funcții, localizate în citoplasma acesteia. În cele ce urmează vom vorbi despre organoizi de uz general, inerente tuturor tipurilor de celule ale tuturor eucariotelor. Ele sunt asociate cu asigurarea funcțiilor vitale ale acestora din urmă. Organoizi cu scop special găsit doar în celule de un anumit tip (foarte specializate) - de exemplu, miofibrile din celulele musculare.
Organelele de uz general au aceeași structură, indiferent de celule și organisme cărora le aparțin. Dar printre ele există grupuri cu membrană (reticul endoplasmatic, aparat Golgi, mitocondrii, plastide, lizozomi, vacuole), precum și non-membrană ( ribozomi, centru celular) structura.
Reticulul endoplasmatic (RE). EPS este format din membrane și este un sistem complex ramificat de tubuli și cisterne care pătrunde în întreaga citoplasmă a celulei (Fig. 14). Există două tipuri de EPS - stare brutăȘi neted. Ribozomii sunt atașați de membranele aspre (din partea citoplasmatică), dar nu sunt prezenți pe membranele netede.
Orez. 14. Reticulul endoplasmatic
Reticulul endoplasmatic îndeplinește o serie de funcții importante într-o celulă eucariotă:
♦ delimitând(diviziunea volumului intern al celulei în diferite spații de reacție);
♦ participarea la sinteza substanțelor organice(ribozomii sunt localizați pe membranele ER rugoase, iar complexele enzimatice sunt localizate pe cele netede, asigurând sinteza lipidelor, glucidelor etc.);
♦ participarea la formarea elementelor aparatului Golgi, lizozomi;
♦ transport de substante.
Aparate Golgi. Aparatul Golgi (AG) este un sistem tancuri(vacuole plate) și bule(veziculele), situate în imediata apropiere a nucleului celular, care se formează datorită EPS ca urmare a separării fragmentelor sale mici (Fig. 15). Atunci când aceste fragmente se îmbină, apar noi cisterne ale aparatului Golgi, în timp ce din EPS sunt transportate diverse substanțe, care sunt implicate în asamblarea compușilor organici complecși (proteine + glucide, proteine + lipide etc.), care sunt îndepărtate cu ajutorul ajutorul AG în afara celulei. Aceste substanțe biologic active sunt fie îndepărtate din celulă (folosind vacuole secretoare prin exocitoză), fie fac parte din lizozomi (vezi mai jos) formați de AG.
Orez. 15. Aparatul Golgi:
Aparatul Golgi îndeplinește următoarele funcții:
♦ sinteză substanțe biologic active produse de celulă;
♦ secretia (eliminarea din celula) a diverselor substante(hormoni, enzime, substanțe din care este construit peretele celular etc.);
♦ participarea la formarea lizozomilor.
Mitocondriile. Toate tipurile de celule eucariote au mitocondrii (Fig. 16). Arată fie corpuri rotunjite, fie tije, mai rar - fire. Dimensiunile lor variază de la 1 la 7 microni. Numărul de mitocondrii dintr-o celulă variază de la câteva sute la zeci de mii (în protozoarele mari).
Orez. 16. Mitocondriile. Deasupra - mitocondrii (a) în canalele urinare, vizibile la microscop cu lumină. Mai jos este un model tridimensional de organizare mitocondrială: 1 – cristae; 2 – membrana exterioară; 3 – membrană internă; 4 – matrice
Mitocondria este formată din două membrane - externȘi intern,între care se află spațiu intermembranar. Membrana interioară formează multe invaginări - cristae, care sunt fie plăci, fie tuburi. Această organizare oferă o zonă imensă a membranei interne. Contine enzime care asigura conversia energiei continute de substantele organice (glucide, lipide) in energie ATP, necesara vietii celulei. Prin urmare, funcția mitocondriilor este de a participa energie procesele celulare. De aceea, un număr mare de mitocondrii sunt inerente, de exemplu, în celulele musculare care efectuează multă muncă.
Plastide.În celulele vegetale se găsesc organele speciale - plastide, care au adesea o formă fusiformă sau rotunjită, uneori mai complexă. Există trei tipuri de plastide - cloroplaste (Fig. 17), cromoplaste și leucoplaste.
Cloroplaste diferă în culoarea verde, care se datorează pigmentului - clorofilă, asigurarea procesului fotosinteză, adică sinteza substanțelor organice din apă (H 2 O) și dioxid de carbon (CO 2) folosind energia luminii solare. Cloroplastele se găsesc în principal în celulele frunzelor (în plantele superioare). Sunt formate din două membrane situate paralele una cu cealaltă, care înconjoară conținutul cloroplastelor - stroma. Membrana interioară formează numeroși saci turtiți - tilacoizi, care sunt stivuite (ca un teanc de monede) – cereale -și se află în stromă. Tilacoizii sunt cei care conțin clorofilă.
Cromoplastele determina culoarea galbena, portocalie si rosu a multor flori si fructe, in celulele carora sunt prezente in cantitati mari. Principalii pigmenți din compoziția lor sunt carotenii. Scopul funcțional al cromoplastelor este de a atrage animalele cu culoare, asigurând polenizarea florilor și răspândirea semințelor.
Orez. 17. Plastide: a – cloroplaste din celulele frunzei Elodea, vizibile la microscop optic; b – schema structurii interne a cloroplastului cu grana, care sunt stive de saci plati situati perpendicular pe suprafata cloroplastului; c – o diagramă mai detaliată care arată tuburile de anastomoză care conectează camerele individuale ale ventilatorului
Leucoplaste sunt plastide incolore conținute în celulele părților subterane ale plantelor (de exemplu, în tuberculii de cartofi), semințe și miezul tulpinilor. În leucoplaste, amidonul se formează în principal din glucoză și se acumulează în organele de depozitare ale plantelor.
Plastidele de un tip se pot transforma în altul. De exemplu, când frunzele își schimbă culoarea toamna, cloroplastele se transformă în cromoplaste.
Lizozomi. Aceste organite arată ca niște vezicule înconjurate de o membrană cu un diametru de până la 2 microni. Conțin câteva zeci de enzime care descompun proteinele, acizii nucleici, polizaharidele și lipidele. Funcția lizozomilor este de a participa la procesele de descompunere intracelulară a compușilor organici complecși (de exemplu, nutrienți sau substanțe ale componentelor celulare „uzate”). Lizozomii fuzionează cu vacuolele fagocitare (sau pinocitare) pentru a forma o vacuola digestivă.
Formarea lizozomilor are loc datorită înmuguririi din cisternele aparatului Golgi.
Ribozomi. Ribozomii (Fig. 18) sunt prezenți atât în celulele eucariotelor cât și ale procariotelor, deoarece îndeplinesc o funcție importantă în biosinteza proteinelor(vezi capitolul 5). Fiecare celulă conține zeci, sute de mii (până la câteva milioane) din aceste mici organele rotunde.
Orez. 18. Schema structurii unui ribozom aşezat pe membrana reticulului endoplasmatic: 1 – subunitate mică; 2 – ARNt; 3 – aminoacil-ARNt; 4 – aminoacid; 5 – subunitate mare; 6 – membrana reticulului endoplasmatic; 7 – lanț polipeptidic sintetizat
Ribozomul este format din două subunități (părți) inegale. Ele se formează separat și se combină, „cuprinzând” ARN mesager, în timpul sintezei unei molecule de proteine. Ribozomii conțin diverse proteine și ARN ribozomal.
Incluziuni celulare. Acesta este numele dat componentelor nepermanente din celulă, prezente în substanța principală a citoplasmei sub formă de boabe, granule sau picături. Incluziunile pot fi sau nu înconjurate de o membrană.
Din punct de vedere funcțional, există trei tipuri de incluziuni: rezerve de nutrienți(amidon, glicogen, grăsimi, proteine), incluziuni secretoare(substanțe caracteristice celulelor glandulare, produse de acestea - hormoni ai glandelor endocrine etc.) și includerea scopului special(în celule înalt specializate, de exemplu hemoglobina din globulele roșii).
§ 3. Organizarea nucleului celular. Cromozomii
Nucleul celular (vezi Fig. 8 și 9) este de cea mai mare importanță în viața celulei, deoarece servește ca depozit al informațiilor ereditare conținute în cromozomi (vezi mai jos).
Nucleul este delimitat de o înveliș nuclear care separă conținutul său (carioplasma) din citoplasmă. Învelișul este format din două membrane separate printr-un gol. Ambele sunt pătrunse de numeroși pori, datorită cărora este posibil schimbul de substanțe între nucleu și citoplasmă. În nucleul celular al majorității eucariotelor există de la 1 la 7 nucleoli. Procesele de sinteză a ARN și ARNt sunt asociate cu acestea.
Componentele principale ale nucleului - cromozomi, format dintr-o moleculă de ADN și diverse proteine. Într-un microscop cu lumină, acestea sunt clar vizibile numai în perioada de diviziune celulară (mitoză, meioză).Într-o celulă care nu se divide, cromozomii arată ca fire lungi și subțiri distribuite pe întregul volum al nucleului.
În timpul diviziunii celulare, firele cromozomiale formează spirale dense, în urma cărora devin vizibile (folosind un microscop convențional) sub formă de tije, „agrafe de păr”. Întregul volum de informații genetice este distribuit între cromozomii nucleului. În procesul de studiere a acestora, au fost identificate următoarele modele:
♦ în nucleele celulelor somatice (adică celulele corpului nesexuale) toți indivizii aceleiași specii conțin același număr de cromozomi care alcătuiesc set de cromozomi(Fig. 19);
Orez. 19. Cromozomi de diferite specii de plante și animale, reprezentați la aceeași scară: 1,2 – amibe; 3,4 – diatomee; 5–8, 18,19 – alge verzi; 9 – agaric musca; 10 – tei; 11–12 – Drosophila; 13 – somon; 14 – skerda (familia Asteraceae); 15 – plantă din familia aroid; 16 – fluture corydalis; 17 – insectă din familia lăcustelor; 20 – insectă strider de apă; 21 – insectă de flori; 22 – amfibian Ambystoma; 23 – aloe (familia crinilor)
♦ fiecare specie este caracterizată de propriul set de cromozomi în funcție de numărul lor (de exemplu, o persoană are 46 de cromozomi, o muscă Drosophila are 8, un vierme rotund are 4, un rac are 196, un cal are 66 și porumbul are 104);
♦ cromozomii din nucleele celulelor somatice pot fi grupați în perechi numite cromozomi omologi pe baza asemănării lor (ca structură și funcție);
♦ în nucleele celulelor germinale (gameți), fiecare pereche de cromozomi omologi conține doar unul, adică setul total de cromozomi este jumătate din cel al celulelor somatice;
♦ se numește un singur set de cromozomi din celulele germinale haploidși este notat cu litera n, iar în somatic - diploid(2n).
Din cele de mai sus este clar că fiecare pereche de cromozomi omologi este formată prin unirea cromozomilor paterni și materni în timpul fecundației, adică fuziunea celulelor sexuale (gameți). În schimb, în timpul formării celulelor germinale, din fiecare pereche de cromozomi omologi intră doar unul în gamet.
Cromozomii diferit perechile omoloage diferă ca mărime și formă (Fig. 20 și 21).
Orez. 20. Structura și tipurile de cromozomi: a – aspect 1 – centromer; 2 – umăr scurt; 3 – umăr lung); structura internă a aceluiași cromozom (1 – centromer; – molecule de ADN); c – tipuri de cromozomi (1 – cu un singur braț; cu brațe diferite; 3 – cu brațe egale: X – braț, V – centromer)
Orez. 21. Un cromozom este format din ADN și proteine. Molecula de ADN este replicată. Două elice duble ADN identice rămân conectate la regiunea centromerului. Aceste copii devin cromozomi separați mai târziu în timpul diviziunii celulare
În corpul cromozomilor există constricția primară (numită centromer), de care sunt atașate firele fusuri de fisiune.Împarte cromozomul în două umăr Cromozomii pot fi cu brațe egale, cu brațe inegale sau cu un singur braț.
Capitolul 5. Metabolism
§ 1. Metabolismul ca unitate de asimilare si disimilare
Toate celulele și organismele vii sunt sisteme deschise, adică se află într-o stare de schimb constant de energii și substanțe cu mediul. Există sisteme deschise în natura neînsuflețită, dar existența lor este diferită calitativ de organismele vii. Luați în considerare acest exemplu: o bucată de sulf nativ care arde se află într-o stare de schimb cu mediul. Când arde, O 2 este absorbit și SO 2 și energie (sub formă de căldură) sunt eliberate. Cu toate acestea, în acest caz, o bucată de sulf ca corp fizic este distrusă și își pierde structura primară.
Pentru organismele vii, schimbul cu mediul se dovedește a fi o condiție pentru păstrarea și menținerea organizării lor structurale prin auto-înnoirea tuturor substanțelor și componentelor din care sunt compuse.
Metabolismul (metabolismul) este un ansamblu de procese care au loc în organismele vii (consum, transformare, acumulare și eliberare de substanțe și energie) care asigură activitatea lor vitală, dezvoltarea, creșterea, reproducerea. În timpul procesului metabolic, moleculele care alcătuiesc celulele sunt descompuse și sintetizate; reînnoirea structurilor celulare și a substanței intercelulare.
Metabolismul se bazează pe procese interconectate asimilare(anabolism) și disimilare(catabolism). În timpul asimilării (schimbului plastic), substanțele complexe sunt sintetizate din cele simple. Datorită acestui fapt, sunt create toate substanțele organice din celulă necesare pentru construirea componentelor sale structurale, a sistemelor enzimatice etc. Asimilarea are loc întotdeauna cu cheltuirea energiei.
În timpul disimilării (metabolismului energetic), substanțele organice complexe sunt descompuse în unele mai simple sau anorganice. În acest caz, se eliberează energie, care este cheltuită de celulă pentru a efectua diverse procese care îi asigură activitatea vitală (sinteza și transportul substanțelor, lucru mecanic etc.).
Toate organismele vii pot fi împărțite în două grupe: autotrofiȘi heterotrofi, care se deosebesc prin sursele de energie şi substanţele necesare pentru a-şi asigura funcţiile vitale.
Autotrofi– organisme care sintetizează compuși organici din substanțe anorganice folosind energia luminii solare (cum fototrofe– plante, cianobacterii) sau energie obținută din oxidarea substanțelor minerale (anorganice) (cum ar fi chimiotrofe– bacterii sulfuroase, bacterii fier etc.). În consecință, ei sunt capabili să creeze în mod independent substanțele necesare activității lor de viață.
§ 2. Disimilarea la organismele anaerobe si aerobe
Organismele pot fi împărțite în două grupe și în funcție de natura disimilației - aerobiȘi anaerobi. Aerobi (din greacă. demon– aer) necesită oxigen liber pentru viață. În anaerobi (greacă. ac– particulă negativă) nu este necesar. În ele, disimilarea se realizează prin fermentație - descompunerea enzimatică fără oxigen a materiei organice cu formarea de substanțe organice mai simple și eliberarea de energie. De exemplu:
♦ fermentarea acidului lactic:
C6H12O6 + 2H3PO4 + 2ADP → 2PH + 2ATP + 2H20;
♦ fermentație alcoolică:
C6H12O6 + 2PH + 2ADP → 2C2H5OH + 2CO2 + 2ATP + 2H2O.
Substanțele formate în timpul fermentației sunt organice și, prin urmare, conțin încă multă energie.
Orez. 22. Relații dintre asimilare și disimilare la organismele autotrofe și heterotrofe
La organismele aerobe, în timpul procesului de respirație în mitocondrii, are loc o descompunere completă a substanțelor organice (folosind O 2 ) la produsele finale sărace în energie CO 2 și H 2 O și este eliberată o cantitate semnificativ mai mare de energie:
C 6 H 12 0 6 (glucoză) + 0 2 > 6C0 2 + 6H 2 0 + energie (datorită căreia se sintetizează 38 de molecule de ATP).
Să considerăm sub forma unor scheme generalizate metabolismul organismelor aerobe autotrofe și heterotrofe (Fig. 22).
Asimilare. Cele mai importante procese ale sale sunt fotosintezăȘi biosinteza proteinelor.
Legătura organismului cu mediul, din punct de vedere fizico-chimic, este un sistem deschis, adică un sistem în care procesele biochimice sunt în desfășurare. Substanțele inițiale provin din mediul înconjurător, iar substanțele care se formează continuu sunt transportate în exterior. Echilibrul dintre viteza și concentrația produselor reacțiilor multidirecționale din organism este condiționat, imaginar, deoarece aportul și eliminarea substanțelor nu se oprește. Conexiunea continuă cu mediul ne permite să considerăm un organism viu ca un sistem deschis.
Pentru toate celulele vii, sursa de energie este Soarele. Celulele vegetale captează energia din lumina soarelui cu ajutorul clorofilei, folosind-o pentru reacții de asimilare în timpul procesului de fotosinteză. Celulele animalelor, ciupercilor și bacteriilor folosesc energia solară indirect, în timpul descompunerii substanțelor organice sintetizate de plantele pământești.
Unele dintre substanțele nutritive ale celulei sunt descompuse în timpul respirației celulare, furnizând astfel energia necesară pentru diferite tipuri de activitate celulară. Acest proces are loc în organele numite mitocondrii. Mitocondriile sunt formate din două membrane: cea exterioară, care separă organitele de citoplasmă, și cea interioară, formând numeroase pliuri. Produsul principal al respirației este ATP. Paraseste mitocondriile si este folosita ca sursa de energie pentru multe reactii chimice din citoplasma si membrana celulara. Dacă oxigenul este necesar pentru respirația celulară, atunci respirația se numește aerobă, dar dacă reacțiile apar în absența oxigenului, atunci vorbim de respirație anaerobă.
Pentru orice tip de muncă efectuată într-o celulă, energia este utilizată într-o singură formă - sub formă de energie din legăturile de fosfat ale ATP. ATP este un compus ușor mobil. Formarea ATP are loc pe membrana interioară a mitocondriilor. ATP este sintetizat în toate celulele în timpul respirației datorită energiei de oxidare a carbohidraților, grăsimilor și a altor substanțe organice. În celulele vegetale verzi, cantitatea principală de ATP este sintetizată în cloroplaste datorită energiei solare. În timpul fotosintezei, ele produc de multe ori mai mult ATP decât mitocondriile. ATP se descompune prin ruperea legăturilor fosfor-oxigen și eliberarea de energie. Acest lucru se întâmplă sub acțiunea enzimei ATPază în timpul hidrolizei ATP - adăugarea de apă cu eliminarea unei molecule de acid fosforic. Ca rezultat, ATP este transformat în ADP, iar dacă două molecule de acid fosforic sunt separate, atunci în AMP. Reacția de eliminare a fiecărei molecule gram de acid este însoțită de eliberarea a 40 kJ. Aceasta este o ieșire de energie foarte mare, motiv pentru care legăturile fosfor-oxigen ale ATP sunt de obicei numite macroergistice (de înaltă energie).
Utilizarea ATP în reacțiile de schimb plastic se realizează prin cuplarea lor cu hidroliza ATP. Moleculele diferitelor substanțe sunt încărcate cu energie prin atașarea grupării fosfor eliberate în timpul hidrolizei din molecula de ATP, adică prin fosforilare.
Particularitatea derivaților de fosfat este că nu pot părăsi celula, deși formele lor „descărcate” trec liber prin membrană. Datorită acestui fapt, moleculele fosforilate rămân în celulă până când sunt utilizate în reacții adecvate.
Procesul invers de conversie a ADP în ATP are loc prin adăugarea unei molecule de acid fosforic la ADP, eliberând apă și absorbind o cantitate mare de energie.
Astfel, ATP este o sursă universală și directă de energie pentru activitatea celulară. Acest lucru creează un singur fond celular de energie și face posibilă redistribuirea și transportul acestuia dintr-o zonă a celulei în alta.
Transferul grupării fosfat joacă un rol important în reacțiile chimice precum asamblarea macromoleculelor din monomeri. De exemplu, aminoacizii pot fi combinați în peptide numai după ce au fost fosforilați anterior. Procesele mecanice de contracție sau mișcare, transportul unei substanțe dizolvate împotriva unui gradient de concentrație și alte procese presupun consumul de energie stocată în ATP.
Procesul de metabolism energetic poate fi reprezentat astfel. Substanțele organice cu molecul mare din citoplasmă sunt transformate enzimatic, prin hidroliză, în altele mai simple din care constau: proteine - în aminoacizi, poli- și dizaharide - în monozaharide (+ glucoză), grăsimile în glicerol și acizi grași. Nu există procese oxidative, se eliberează puțină energie, care nu este folosită și intră în formă termică. Majoritatea celulelor folosesc carbohidrații mai întâi. Polizaharidele (amidonul la plante și glicogenul la animale) sunt hidrolizate la glucoză. Oxidarea glucozei are loc in trei faze: glicoliza, decarboxilarea oxidativa (ciclul Krebs - ciclul acidului citric) si fosforilarea oxidativa (lantul respirator). În citoplasmă are loc glicoliza, în urma căreia o moleculă de glucoză este împărțită în două molecule de acid piruvic cu eliberarea a două molecule de ATP. În absența oxigenului, acidul piruvic este transformat fie în etanol (fermentare), fie în acid lactic (respirație anaerobă).
Când glicoliza are loc în celulele animale, molecula de glucoză cu șase atomi de carbon se descompune în două molecule de acid lactic. Acest proces este în mai multe etape. Se efectuează secvenţial de către 13 enzime. În timpul fermentației alcoolice, dintr-o moleculă de glucoză se formează două molecule de etanol și două molecule de CO2.
Glicoliza este o fază comună respirației anaerobe și aerobe; celelalte două apar numai în condiții aerobe. Procesul de oxidare fără oxigen, în care doar o parte din energia metaboliților este eliberată și utilizată, este final pentru organismele anaerobe. În prezența oxigenului, acidul piruvic trece în mitocondrii, unde, ca urmare a unui număr de reacții secvențiale, este complet oxidat aerob la H2O și CO2 cu fosforilarea simultană a ADP la ATP. În acest caz, două molecule de ATP sunt produse prin glicoliză, două prin ciclul Krebs și 34 prin lanțul respirator. Randamentul net pentru oxidarea completă a unei molecule de glucoză la H2O și CO2 este de 38 de molecule.
Astfel, la organismele aerobe, descompunerea finală a substanțelor organice se realizează prin oxidarea lor cu oxigen atmosferic în substanțe anorganice simple: CO2 și H2O. Acest proces are loc pe cresta mitocondriilor. În acest caz, cantitatea maximă de energie liberă este eliberată, o parte semnificativă din care este rezervată în moleculele de ATP. Este ușor de observat că oxidarea aerobă oferă celulei energie liberă în cea mai mare măsură.
Ca urmare a catabolismului, moleculele de ATP bogate în energie se acumulează în celulă, iar CO2 și excesul de apă sunt eliberate în mediul extern.
Moleculele de zahăr care nu sunt necesare pentru respirație pot fi stocate în celulă. Lipidele în exces sunt fie descompuse, după care produsele defalcării lor intră în mitocondrii ca substrat pentru respirație, fie sunt depuse ca rezerve în citoplasmă sub formă de picături de grăsime. Proteinele sunt construite din aminoacizii care intră în celulă. Sinteza proteinelor are loc în organele numite ribozomi. Fiecare ribozom este format din două subparticule - mari și mici: ambele subparticule includ molecule de proteine și molecule de ARN.
Ribozomii sunt adesea atașați la un sistem membranar special format din cisterne și vezicule - așa-numitul reticul endoplasmatic (ER); în celulele care produc multe proteine, reticulul endoplasmatic este adesea foarte bine dezvoltat și acoperit cu ribozomi. Unele enzime sunt eficiente doar dacă sunt atașate de o membrană. Cele mai multe dintre enzimele implicate în sinteza lipidelor se află aici. Astfel, reticulul endoplasmatic este ca un fel de banc de lucru cu celule.
În plus, ER împarte citoplasma în compartimente separate, adică separă diferite procese chimice care au loc simultan în citoplasmă și, prin urmare, reduce probabilitatea ca aceste procese să interfereze între ele.
Produsele produse de o anumită celulă sunt adesea folosite în afara celulei. În astfel de cazuri, proteinele sintetizate pe ribozomi trec prin membranele reticulului endoplasmatic și sunt împachetate în vezicule membranare care se formează în jurul lor, care sunt apoi desprinse din RE. Aceste vezicule, aplatizate și stivuite una peste alta, precum clătitele stivuite, formează o structură caracteristică numită complexul Golgi sau aparatul Golgi. În timpul șederii lor în aparatul Golgi, proteinele suferă anumite modificări. Când vine momentul să părăsească celula, veziculele membranei se contopesc cu membrana celulară și sunt golite, revărsându-și conținutul, adică secreția are loc prin exocitoză.
Aparatul Golgi produce, de asemenea, lizozomi - saci membranari care conțin enzime digestive. A afla cum o celulă produce, împachetează și exportă anumite proteine și cum „știe” ce proteine ar trebui să păstreze pentru ea însăși, este una dintre cele mai fascinante ramuri ale citologiei moderne.
Membranele oricărei celule se mișcă și se schimbă în mod constant. Membranele ER se deplasează lent în întreaga celulă. Secțiuni individuale ale acestor membrane se separă și formează vezicule, care devin temporar parte a aparatului Golgi și apoi, prin procesul de exocitoză, fuzionează cu membrana celulară.
Mai târziu, materialul membranei este returnat în citoplasmă, unde este folosit din nou.
Schimbul de substanțe care intră în celulă sau eliberate de aceasta în exterior, precum și schimbul de diferite semnale cu micro- și macromediul, are loc prin membrana exterioară a celulei. După cum se știe, membrana celulară este un dublu strat lipidic în care sunt încorporate diverse molecule de proteine care acționează ca receptori specializați, canale ionice, dispozitive care transportă sau îndepărtează activ diferite substanțe chimice, contacte intercelulare etc. În celulele eucariote sănătoase, fosfolipidele sunt distribuite în membrana asimetric: suprafața exterioară este formată din sfingomielină și fosfatidilcolină, suprafața interioară - din fosfatidilserina și fosfatidiletanolamină. Menținerea unei astfel de asimetrii necesită cheltuieli de energie. Prin urmare, în cazul deteriorării celulelor, infecției sau lipsei de energie, suprafața exterioară a membranei este îmbogățită cu fosfolipide neobișnuite pentru aceasta, ceea ce devine un semnal pentru alte celule și enzime despre deteriorarea celulelor cu o reacție corespunzătoare la aceasta. Cel mai important rol îl joacă forma solubilă a fosfolipazei A2, care descompune acidul arahidonic și creează lizoforme din fosfolipidele menționate mai sus. Acidul arahidonic este veriga limitatoare pentru crearea de mediatori inflamatori precum eicosanoizii, iar moleculele protectoare - pentraxinele (proteina C reactiva (CRP), precursori ai proteinelor amiloide) - sunt atasate de lizoformele din membrana, urmate de activarea complementului. sistem de-a lungul căii clasice și distrugerea celulelor.
Structura membranei ajută la păstrarea caracteristicilor mediului intern al celulei, a diferențelor sale față de mediul extern. Acest lucru este asigurat de permeabilitatea selectivă a membranei celulare și de existența mecanismelor active de transport în aceasta. Perturbarea lor ca urmare a deteriorării directe, de exemplu, de către tetrodotoxină, ouabaină, tetraetilamoniu sau în cazul unei aprovizionări insuficiente cu energie a „pompelor” corespunzătoare duce la perturbarea compoziției electrolitice a celulei, modificări ale metabolismului acesteia, perturbări. a funcțiilor specifice – contracția, conducerea impulsurilor de excitație etc. Perturbarea canalelor ionice celulare (calciu, sodiu, potasiu și clorură) la om poate fi determinată și genetic de mutații ale genelor responsabile de structura acestor canale. Așa-numitele canalopatii provoacă boli ereditare ale sistemului nervos, muscular și digestiv. Intrarea excesiva a apei in celula poate duce la ruperea acesteia - citoliza - datorita perforarii membranei atunci cand complementul este activat sau un atac al limfocitelor citotoxice si al celulelor natural killer.
Membrana celulară are mulți receptori încorporați în ea - structuri care, atunci când sunt combinate cu moleculele de semnalizare specifice corespunzătoare (liganzi), transmit un semnal în interiorul celulei. Acest lucru are loc prin diverse cascade de reglare constând din molecule active enzimatic care sunt activate secvenţial şi care în cele din urmă contribuie la implementarea diferitelor programe celulare, cum ar fi creşterea şi proliferarea, diferenţierea, motilitatea, îmbătrânirea şi moartea celulară. Cascadele de reglementare sunt destul de numeroase, dar numărul lor nu a fost încă pe deplin determinat. Sistemul de receptori și cascadele de reglare asociate acestora există și în interiorul celulei; ele creează o rețea de reglementare specifică cu puncte de concentrare, distribuție și selecție a căii ulterioare a semnalului în funcție de starea funcțională a celulei, stadiul dezvoltării acesteia și acțiunea simultană a semnalelor de la alți receptori. Rezultatul acestui lucru poate fi inhibarea sau întărirea semnalului, direcționându-l de-a lungul unei căi de reglementare diferite. Atât aparatul receptor, cât și căile de transducție a semnalului prin cascade de reglare, de exemplu către nucleu, pot fi perturbate ca urmare a unui defect genetic care apare ca defect congenital la nivel de organism sau datorită unei mutații somatice într-un anumit tip de celulă. Aceste mecanisme pot fi deteriorate de agenți infecțioși, toxine și, de asemenea, se pot modifica în timpul procesului de îmbătrânire. Etapa finală a acesteia poate fi o perturbare a funcțiilor celulei, procesele de proliferare și diferențiere a acesteia.
Pe suprafața celulelor există și molecule care joacă un rol important în procesele de interacțiune intercelulară. Acestea pot include proteine de adeziune celulară, antigene de histocompatibilitate, antigeni specifici țesuturilor, de diferențiere etc. Modificările în compoziția acestor molecule determină perturbarea interacțiunilor intercelulare și pot determina activarea mecanismelor adecvate de eliminare a unor astfel de celule, deoarece acestea reprezintă o un anumit pericol pentru integritatea organismului ca rezervor de infecție, în special virală, sau ca potențiali inițiatori ai creșterii tumorii.
Încălcarea alimentării cu energie a celulei
Sursa de energie în celulă este hrana, după descompunerea căreia energia este eliberată în substanțe finale. Principalul loc de producere a energiei este mitocondriile, în care substanțele sunt oxidate cu ajutorul enzimelor din lanțul respirator. Oxidarea este principalul furnizor de energie, deoarece, ca urmare a glicolizei, nu mai mult de 5% din energie este eliberată din aceeași cantitate de substraturi de oxidare (glucoză), în comparație cu oxidarea. Aproximativ 60% din energia eliberată în timpul oxidării este acumulată prin fosforilarea oxidativă în fosfații de înaltă energie (ATP, creatină fosfat), restul este disipat sub formă de căldură. Ulterior, fosfații de înaltă energie sunt utilizați de către celulă pentru procese precum funcționarea pompei, sinteza, diviziunea, mișcarea, secreția etc. Există trei mecanisme, a căror deteriorare poate provoca o întrerupere a aprovizionării cu energie a celulei: primul este mecanism de sinteză a enzimelor de metabolism energetic, al doilea este mecanismul de fosforilare oxidativă, al treilea este mecanismul de utilizare a energiei.
Întreruperea transportului de electroni în lanțul respirator mitocondrial sau decuplarea oxidării și fosforilării ADP cu pierderea potențialului de protoni, forța motrice pentru generarea de ATP, duce la o slăbire a fosforilării oxidative în așa fel încât cea mai mare parte a energiei este disipată sub formă de căldură și numărul compușilor cu energie înaltă scade. Decuplarea oxidării și fosforilării sub influența adrenalinei este folosită de celulele organismelor homeoterme pentru a crește producția de căldură menținând în același timp o temperatură constantă a corpului în timpul răcirii sau creșterea acesteia în timpul febrei. În tireotoxicoză se observă modificări semnificative ale structurii mitocondriale și ale metabolismului energetic. Aceste modificări sunt inițial reversibile, dar după un anumit punct devin ireversibile: mitocondriile se fragmentează, se dezintegrează sau se umflă, pierd cresta, transformându-se în vacuole și în cele din urmă acumulează substanțe precum hialina, feritina, calciul, lipofuscina. La pacienții cu scorbut, mitocondriile fuzionează pentru a forma condriosfere, posibil din cauza deteriorării membranei de către compușii peroxid. Deteriorarea semnificativă a mitocondriilor are loc sub influența radiațiilor ionizante în timpul transformării unei celule normale într-una malignă.
Mitocondriile sunt un depozit puternic de ioni de calciu, unde concentrația sa este cu câteva ordine de mărime mai mare decât cea din citoplasmă. Când mitocondriile sunt deteriorate, calciul intră în citoplasmă, provocând activarea proteinazelor cu deteriorarea structurilor intracelulare și întreruperea funcțiilor celulei corespunzătoare, de exemplu, contracturi de calciu sau chiar „moartea calciului” în neuroni. Ca urmare a perturbării capacității funcționale a mitocondriilor, crește brusc formarea compușilor peroxid de radicali liberi, care au o reactivitate foarte mare și, prin urmare, dăunează componente importante ale celulei - acizi nucleici, proteine și lipide. Acest fenomen se observă în timpul așa-numitului stres oxidativ și poate avea consecințe negative asupra existenței celulei. Astfel, deteriorarea membranei exterioare a mitocondriilor este însoțită de eliberarea în citoplasmă a unor substanțe conținute în spațiul intermembranar, în primul rând citocromul C și alte substanțe biologic active, care declanșează reacții în lanț care provoacă moartea celulară programată - apoptoza. Prin deteriorarea ADN-ului mitocondrial, reacțiile radicalilor liberi distorsionează informațiile genetice necesare formării anumitor enzime ale lanțului respirator, care sunt produse în mod specific în mitocondrii. Acest lucru duce la o întrerupere și mai mare a proceselor oxidative. În general, aparatul genetic propriu al mitocondriilor, în comparație cu aparatul genetic al nucleului, este mai puțin protejat de influențele nocive care pot modifica informațiile genetice codificate în acesta. Ca urmare, disfuncția mitocondriilor apare pe tot parcursul vieții, de exemplu, în timpul procesului de îmbătrânire, în timpul transformării maligne a celulei, precum și pe fondul bolilor mitocondriale ereditare asociate cu mutația ADN-ului mitocondrial în ou. În prezent, au fost descrise peste 50 de mutații mitocondriale care cauzează boli degenerative ereditare ale sistemelor nervos și muscular. Ele sunt transmise copilului exclusiv de la mamă, deoarece mitocondriile spermatozoizilor nu fac parte din zigot și, în consecință, din noul organism.
Încălcarea conservării și transmiterii informațiilor genetice
Nucleul celular conține cea mai mare parte a informațiilor genetice și, prin urmare, asigură funcționarea sa normală. Prin expresia selectivă a genelor, coordonează activitatea celulară în timpul interfazei, stochează informații genetice și recreează și transmite materialul genetic în timpul diviziunii celulare. Replicarea ADN-ului și transcripția ARN au loc în nucleu. Diferiți factori patogeni, cum ar fi radiațiile ultraviolete și ionizante, oxidarea radicalilor liberi, substanțele chimice, virușii, pot deteriora ADN-ul. Se estimează că fiecare celulă a unui animal cu sânge cald durează 1 zi. pierde peste 10.000 de baze. Aici ar trebui să adăugăm încălcări la copiere în timpul divizării. Dacă aceste daune ar persista, celula nu ar putea supraviețui. Protecția constă în existența unor sisteme de reparare puternice, cum ar fi endonucleaza ultravioletă, sisteme de reparare de replicare și recombinare, care înlocuiesc deteriorarea ADN-ului. Defectele genetice ale sistemelor de reparare determină dezvoltarea bolilor cauzate de sensibilitatea crescută la factorii care dăunează ADN-ului. Aceasta este xeroderma pigmentosum, precum și unele sindroame de îmbătrânire accelerată, însoțite de o tendință crescută de a dezvolta tumori maligne.
Sistemul de reglare a proceselor de replicare a ADN-ului, transcrierea ARN-ului mesager (ARNm) și traducerea informațiilor genetice din acizii nucleici în structura proteinelor este destul de complex și cu mai multe niveluri. Pe lângă cascadele de reglare care declanșează acțiunea factorilor de transcripție cu un număr total de peste 3000, care activează anumite gene, există și un sistem de reglare pe mai multe niveluri mediat de molecule mici de ARN (ARN interferent; ARNi). Genomul uman, care constă din aproximativ 3 miliarde de baze purinice și pirimidinice, conține doar 2% din genele structurale responsabile de sinteza proteinelor. Restul asigură sinteza ARN-urilor reglatoare, care, simultan cu factorii de transcripție, activează sau blochează activitatea genelor structurale la nivel de ADN în cromozomi sau influențează procesele de translație a ARN-ului mesager (ARNm) în timpul formării unei molecule polipeptidice în citoplasma. Încălcarea informațiilor genetice poate apărea atât la nivelul genelor structurale, cât și la nivelul părții reglatoare a ADN-ului cu manifestări corespunzătoare sub formă de diferite boli ereditare.
Recent, a fost atrasă multă atenție modificările materialului genetic care apar în timpul dezvoltării individuale a unui organism și sunt asociate cu inhibarea sau activarea anumitor secțiuni ale ADN-ului și cromozomilor datorită metilării, acetilării și fosforilării acestora. Aceste modificări persistă mult timp, uneori de-a lungul întregii vieți a organismului, de la embriogeneză până la bătrânețe, și se numesc ereditate epigenomică.
Proliferarea celulelor cu informații genetice modificate este împiedicată și de sistemele (factorii) care controlează ciclul mitotic. Ele interacționează cu proteine kinaze dependente de ciclină și subunitățile lor catalitice - ciclinele - și împiedică celula să treacă prin ciclul mitotic complet, oprind diviziunea la granița dintre fazele presintetice și sintetice (blocul G1/S) până când repararea ADN-ului este finalizată, iar dacă acest lucru este imposibil, ei inițiază celulele morții programate. Acești factori includ gena p53, a cărei mutație provoacă pierderea controlului asupra proliferării celulelor transformate; se observă în aproape 50% dintre cancerele umane. Al doilea punct de control al ciclului mitotic este la granița G2/M. Aici, distribuția corectă a materialului cromozomial între celulele fiice în mitoză sau meioză este controlată folosind un set de mecanisme care controlează fusul celular, centrul și centromerii (kinetocori). Ineficacitatea acestor mecanisme duce la perturbarea distribuției cromozomilor sau a părților acestora, care se manifestă prin absența oricărui cromozom într-una dintre celulele fiice (aneuploidie), prezența unui cromozom suplimentar (poliploidie), separarea unui parte dintr-un cromozom (deleție) și transferul acestuia într-un alt cromozom (translocare). Astfel de procese sunt foarte des observate în timpul proliferării celulelor degenerate și transformate malign. Dacă acest lucru se întâmplă în timpul meiozei cu celule germinale, duce fie la moartea fătului într-un stadiu incipient al dezvoltării embrionare, fie la nașterea unui organism cu o boală cromozomială.
Proliferarea celulară necontrolată în timpul creșterii tumorii are loc ca urmare a mutațiilor în genele care controlează proliferarea celulară și sunt numite oncogene. Dintre cele peste 70 de oncogene cunoscute în prezent, majoritatea aparțin componentelor de reglare a creșterii celulare, unele sunt reprezentate de factori de transcripție care reglează activitatea genelor, precum și de factori care inhibă diviziunea și creșterea celulară. Un alt factor care limitează expansiunea (răspândirea) excesivă a celulelor în proliferare este scurtarea capetelor cromozomilor - telomeri, care nu se pot replica complet ca urmare a interacțiunii pur sterice, prin urmare, după fiecare diviziune celulară, telomerii sunt scurtați cu o anumită parte a bazelor. Astfel, celulele în proliferare ale unui organism adult după un anumit număr de diviziuni (de obicei de la 20 la 100 în funcție de tipul de organism și de vârsta acestuia) epuizează lungimea telomerilor și replicarea ulterioară a cromozomilor se oprește. Acest fenomen nu apare în epiteliul spermatozoizilor, enterocite și celule embrionare din cauza prezenței enzimei telomerazei, care restabilește lungimea telomerilor după fiecare diviziune. În majoritatea celulelor organismelor adulte, telomeraza este blocată, dar, din păcate, este activată în celulele tumorale.
Conexiunea dintre nucleu și citoplasmă și transportul substanțelor în ambele direcții se realizează prin porii din membrana nucleară cu participarea unor sisteme speciale de transport care consumă energie. În acest fel, energia și substanțele plastice, moleculele de semnalizare (factori de transcripție) sunt transportate către nucleu. Fluxul invers transportă în citoplasmă molecule de ARNm și ARN de transfer (ARNt), ribozomi necesari pentru sinteza proteinelor în celulă. Aceeași cale de transport al substanțelor este, de asemenea, inerentă virusurilor, în special precum HIV. Ei își transferă materialul genetic în nucleul celulei gazdă cu încorporarea lui în continuare în genomul gazdă și transferul de ARN viral nou format în citoplasmă pentru sinteza ulterioară a proteinelor de noi particule virale.
Încălcarea proceselor de sinteză
Procesele de sinteză a proteinelor au loc în cisterne ale reticulului endoplasmatic, strâns legate de porii din membrana nucleară, prin care ribozomii, ARNt și ARNm intră în reticulul endoplasmatic. Aici se realizează sinteza lanțurilor polipeptidice, care ulterior capătă forma lor finală în reticulul endoplasmatic agranular și complexul lamelar (complexul Golgi), unde suferă modificări post-translaționale și se combină cu moleculele de carbohidrați și lipide. Moleculele proteice nou formate nu rămân la locul sintezei, ci printr-un proces complex reglementat numit proteinkineza, sunt transferate activ în acea parte izolată a celulei unde își vor îndeplini funcția dorită. În acest caz, un pas foarte important este structurarea moleculei transferate într-o configurație spațială adecvată capabilă să își îndeplinească funcția inerentă. Această structurare are loc cu ajutorul unor enzime speciale sau pe o matrice de molecule proteice specializate - chaperone, care ajută molecula proteică, nou formată sau modificată ca urmare a influenței externe, să dobândească structura tridimensională corectă. În cazul unui efect advers asupra celulei, atunci când există o posibilitate de perturbare a structurii moleculelor de proteine (de exemplu, cu creșterea temperaturii corpului, un proces infecțios, intoxicație), concentrația de însoțitori în celulă crește brusc. Prin urmare, astfel de molecule sunt numite și proteine de stres, sau proteine de șoc termic. Încălcarea structurării unei molecule proteice duce la formarea de conglomerate inerte chimic, care se depun în celulă sau în afara acesteia în timpul amiloidozei, bolii Alzheimer etc. Uneori, o moleculă similară pre-structurată poate servi ca matrice, iar în aceasta În cazul în care structurarea primară are loc incorect, toate moleculele ulterioare vor fi, de asemenea, defecte. Această situație apare în așa-numitele boli prionice (scrapie la oi, vaci turbate, kuru, boala Creutzfeldt-Jakob la om), când un defect al uneia dintre proteinele membranei celulei nervoase determină acumularea ulterioară de mase inerte în interiorul celulei. și perturbarea funcțiilor sale vitale.
Perturbarea proceselor de sinteză într-o celulă poate apărea în diferitele sale etape: transcripția ARN-ului în nucleu, translația polipeptidelor în ribozomi, modificarea post-translațională, hipermetilarea și glicozilarea moleculei bej, transportul și distribuția proteinelor în celulă și îndepărtarea acestora. spre exterior. În acest caz, se poate observa o creștere sau scădere a numărului de ribozomi, descompunerea poliribozomilor, extinderea cisternelor reticulului endoplasmatic granular, pierderea ribozomilor și formarea veziculelor și vacuolelor. Astfel, atunci când este otrăvită de un greb palid, enzima ARN polimerază este deteriorată, ceea ce perturbă transcripția. Toxina difterică, prin inactivarea factorului de alungire, perturbă procesele de translație, provocând leziuni miocardice. Cauza perturbării sintezei unor molecule de proteine specifice poate fi agenți infecțioși. De exemplu, virusurile herpetice inhibă sinteza și exprimarea moleculelor de antigen MHC, ceea ce le permite să evite parțial controlul imun al bacililor ciumei - sinteza mediatorilor inflamației acute; Apariția unor proteine neobișnuite poate opri descompunerea lor ulterioară și poate duce la acumularea de material inert sau chiar toxic. Acest lucru poate fi, într-o anumită măsură, facilitat de întreruperea proceselor de dezintegrare.
Perturbarea proceselor de degradare
Concomitent cu sinteza proteinelor în celulă, are loc continuu descompunerea acesteia. În condiții normale, acest lucru are o semnificație de reglementare și formativă importantă, de exemplu, în timpul activării formelor inactive de enzime, hormoni proteici și proteine ciclului mitotic. Creșterea și dezvoltarea normală a celulelor necesită un echilibru fin controlat între sinteza și degradarea proteinelor și organelelor. Cu toate acestea, în procesul de sinteză a proteinelor, din cauza erorilor în funcționarea aparatului de sinteză, a structurii anormale a moleculei de proteine și a deteriorării acesteia de către agenți chimici și bacterieni, se formează în mod constant un număr destul de mare de molecule defecte. Potrivit unor estimări, ponderea lor este de aproximativ o treime din toate proteinele sintetizate.
Celulele de mamifere au mai multe principale Modalități de distrugere a proteinelor: prin proteaze lizozomale (pentide hidrolaze), proteinaze calciu dependente (endopeptidaze) și sistemul proteazomal. În plus, există și proteinaze specializate, precum caspazele. Principalul organel în care are loc degradarea substanțelor în celulele eucariote este lizozomul, care conține numeroase enzime hidrolitice. Datorită proceselor de endocitoză și diferitelor tipuri de autofagie în lizozomi și fagolizozomi, atât moleculele proteice defecte, cât și organele întregi sunt distruse: mitocondriile deteriorate, secțiunile membranei plasmatice, unele proteine extracelulare și conținutul granulelor secretoare.
Un mecanism important pentru degradarea proteinelor este proteazomul, o structură proteinază multicatalitică cu structură complexă localizată în citosol, nucleu, reticul endoplasmatic și pe membrana celulară. Acest sistem enzimatic este responsabil pentru descompunerea proteinelor deteriorate, precum și a proteinelor sănătoase care trebuie eliminate pentru funcționarea normală a celulelor. În acest caz, proteinele care urmează să fie distruse sunt combinate preliminar cu o polipeptidă specifică, ubiquitina. Cu toate acestea, proteinele non-ubiquitinate pot fi, de asemenea, parțial distruse în proteazomi. Defalcarea moleculelor proteice din proteazomi în polipeptide scurte (procesare) cu prezentarea lor ulterioară împreună cu moleculele MHC de tip I este o legătură importantă în controlul imun al homeostaziei antigenice în organism. Când funcția proteazomului este slăbită, se acumulează proteine deteriorate și inutile, care însoțesc îmbătrânirea celulară. Încălcarea degradării proteinelor dependente de ciclină duce la perturbarea diviziunii celulare, degradarea proteinelor secretoare - la dezvoltarea cistofibrozei. În schimb, o creștere a funcției proteazomului însoțește epuizarea organismului (SIDA, cancer).
Cu tulburări determinate genetic de degradare a proteinelor, organismul nu este viabil și moare în stadiile incipiente ale embriogenezei. Dacă descompunerea grăsimilor sau a carbohidraților este perturbată, apar boli de depozitare (tesaurioză). În acest caz, în interiorul celulei se acumulează o cantitate excesivă de anumite substanțe sau produse ale descompunerii lor incomplete - lipide, polizaharide, ceea ce dăunează semnificativ funcției celulei. Cel mai adesea acest lucru se observă în celulele epiteliale hepatice (hepatocite), neuroni, fibroblaste și macrofagocite.
Tulburările dobândite ale proceselor de descompunere a substanțelor pot apărea ca urmare a proceselor patologice (de exemplu, proteine, grăsimi, carbohidrați și degenerare pigmentară) și sunt însoțite de formarea de substanțe neobișnuite. Tulburările în sistemul de proteoliză lizozomală duc la scăderea adaptării în timpul postului sau la creșterea stresului și la apariția anumitor disfuncții endocrine - scăderea nivelului de insulină, tiroglobuline, citokine și receptorii acestora. Degradarea proteinelor încetinește rata de vindecare a rănilor, determină dezvoltarea aterosclerozei și afectează răspunsul imun. Cu hipoxie, modificări ale pH-ului intracelular, leziuni ale radiațiilor, caracterizate prin peroxidarea crescută a lipidelor membranare, precum și sub influența substanțelor lizozomotrope - endotoxine bacteriene, metaboliți ai ciupercilor toxice (sporofusarină), cristale de oxid de siliciu - stabilitatea membranei lizozomului modificări, enzimele lizozomale activate sunt eliberate în citoplasmă, ceea ce provoacă distrugerea structurilor celulare și moartea acesteia.
Capitolul 1
BAZELE FIZIOLOGIEI CELULARE
I. Dudel
Membrană plasmatică . Celulele animale sunt delimitate de o membrană plasmatică (Figura 1.1). Ne vom opri asupra structurii sale, care este foarte asemănătoare cu structura multor membrane intracelulare, mai detaliat. Matricea principală a membranei este formată din lipide,în principal fosfatidilcolină. Aceste lipide constau dintr-un grup de cap hidrofil de care sunt atașate lanțuri hidrocarburi hidrofobe lungi. În apă, astfel de lipide formează spontan o peliculă dublu strat de 4-5 nm grosime, în care grupările hidrofile se confruntă cu mediul apos, iar lanțurile de hidrocarburi hidrofobe sunt dispuse pe două rânduri, formând o fază lipidică anhidră. Membranele celulare sunt bistraturi lipidice de acest tip și conțin glicolipide, colesterol și fosfolipide (Fig. 1.2). Partea hidrofilă a glicolipidelor este formată din oligozaharide. Glicolipidele sunt întotdeauna situate pe suprafața exterioară a membranei plasmatice, cu partea oligozaharidă a moleculei orientată ca un fir de păr scufundat în mediu. Împrăștiate printre fosfolipide în cantități aproape egale, moleculele de colesterol stabilizează membrana. Distribuția diferitelor lipide în straturile interioare și exterioare ale membranei nu este aceeași și chiar și în cadrul unui singur strat există zone în care sunt concentrate anumite tipuri de lipide. Această distribuție neuniformă
Orez. 1.1. Desen schematic al unei celule care prezintă cele mai importante organite
are probabil o semnificație funcțională, încă neclară.
Principalele elemente funcționale înglobate în matricea lipidică relativ inertă a membranei sunt veverite(Fig. 1.2). Proteina în greutate reprezintă 25 până la 75% în diferite membrane, dar deoarece moleculele de proteine sunt mult mai mari decât moleculele de lipide, 50% în greutate este echivalent cu raportul dintre 1 moleculă de proteină la 50 de molecule de lipide. Unele proteine pătrund în membrana de la suprafața ei exterioară spre interioară, în timp ce altele sunt fixate într-un singur strat. Moleculele de proteine sunt de obicei orientate astfel încât grupările lor hidrofobe să fie înglobate în membrana lipidică, iar grupările hidrofile polare de pe suprafața membranei să fie încorporate în faza apoasă. Multe proteine de pe suprafața exterioară a membranei sunt glicoproteine; grupările lor zaharide hidrofile se confruntă cu mediul extracelular.
Sisteme membranare ale organitelor intracelulare .
Aproximativ jumătate din volumul celular este ocupat de organele izolate din citosol prin membrane. Suprafața totală a membranelor organelelor intracelulare este de cel puțin 10 ori mai mare decât suprafața membranei plasmatice. Cel mai utilizat sistem de membrană este reticul endoplasmatic, reprezentand o retea

Orez. 1.2.Reprezentarea schematică a membranei plasmatice. Proteinele sunt încorporate în stratul dublu fosfolipidic, unele dintre ele pătrund în stratul dublu, în timp ce altele sunt ancorate doar în stratul exterior sau interior.
tuburi foarte contorte sau structuri alungite asemănătoare sacului; zone mari ale reticulului endoplasmatic sunt împânzite cu ribozomi; un astfel de reticul se numește granular sau aspru (Fig. 1.1). aparate Golgi constă, de asemenea, din lamele legate de membrană, din care veziculele sau veziculele se desprind (Fig. 1.1). Lizozomi și peroxizomi- Acestea sunt mici vezicule specializate. În toate aceste diverse organite, membrana și spațiul pe care îl cuprinde conțin seturi specifice de enzime; În interiorul organelelor se acumulează produse metabolice speciale, utilizate pentru îndeplinirea diferitelor funcții ale organelelor.
MiezȘi mitocondriile diferă prin faptul că fiecare dintre aceste organite este înconjurată de două membrane. Nucleul este responsabil de controlul cinetic al metabolismului; membrana interioară pliată a mitocondriilor este locul metabolismului oxidativ; aici, datorită oxidării piruvatului sau acizilor grași, este sintetizat compusul de înaltă energie adenozin trifosfat (ATP, sau ATP).
Citoscheletul . Citoplasma care înconjoară organelele nu poate fi în niciun fel considerată amorfă; este pătruns de o reţea citoscheletică. Citoscheletul este format din microtubuli, filamente de actină și filamente intermediare (Fig. 1.1). Microtubuli au un diametru exterior de aproximativ 25 nm; se formează, ca un polimer obișnuit, ca rezultat al asamblarii moleculelor de proteină tubulină. filamente de actină - fibrele contractile, situate în stratul aproape de membrană și în întreaga celulă, participă în principal la procesele asociate cu mișcarea. Filamente intermediare constau din blocuri de compoziție chimică diferită în diferite tipuri de celule; ele formează o varietate de conexiuni între celelalte două elemente citoscheletice menționate mai sus. Organelele și membrana plasmatică sunt, de asemenea, asociate cu citoscheletul, care nu numai că menține forma celulei și poziția organelelor în ea, dar determină și modificări ale formei celulei și mobilității acesteia.
Citosol . Aproximativ jumătate din volumul celular este ocupat de citosol. Deoarece are aproximativ 20% proteine în greutate, este mai mult un gel decât o soluție apoasă. Molecule mici, inclusiv organice și anorganice ioni, dizolvat în faza apoasă. Se produce un schimb de ioni între celulă și mediu (spațiul extracelular); aceste procese metabolice vor fi discutate în secțiunea următoare. Concentrația ionilor în spațiul extracelular se menține cu o acuratețe considerabilă la un nivel constant; concentrația intracelulară a fiecărui ion are și un nivel specific, diferit de cel din exteriorul celulei (Tabelul 1.1). Cel mai abundent cation din mediul extracelular este Na+ în celulă concentrația sa este de peste 10 ori mai mică. Dimpotrivă, concentrația de K + este cea mai mare în interiorul celulei, în afara celulei, este mai mică cu mai mult de un ordin de mărime. Cel mai mare gradient între concentrațiile extracelulare și intracelulare există pentru Ca 2+, concentrația de ioni liberi în interiorul celulei este de cel puțin 10.000 de ori mai mică decât în afara acesteia. Nu toți ionii sunt dizolvați în citosol; unii dintre ei sunt adsorbiți pe proteine sau depozitați în organele. De exemplu, în cazul Ca 2+, ionii legați sunt mult mai numeroși decât cei liberi. Majoritatea proteinelor citosolice sunt enzime, cu participarea cărora se desfășoară multe procese de metabolism intermediar: glicoliză și gluconeogeneză, sinteza sau distrugerea aminoacizilor, sinteza proteinelor pe ribozomi (Fig. 1.1). Citosolul conține, de asemenea, picături de lipide și granule de glicogen, care servesc drept rezerve de molecule importante.
Tabelul 1.1.Concentrațiile ionilor intra și extracelulari în celulele musculare ale animalelor homeoterme. A – – „anioni celulari cu greutate moleculară mare”
|
Concentrația intracelulară |
Concentrația extracelulară |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca 2+ |
10 –7 10 –8 mmol/l |
Ca 2+ |
2 mmol/l |
|
C l – |
4 mmol/l |
C1 – |
120 mmol/l |
|
HCO 3 – |
8 mmol/l |
HCO 3 – |
27 mmol/l |
|
A - |
155 mmol/l |
Alții cationi |
5 mmol/l |
|
Potențial de repaus –90 mV |
|||
1.2. Schimb de substanțe între celulă și mediu
Am descris pe scurt structura celulei, astfel încât să putem folosi această descriere pentru a trece în revistă elementele de bază ale fiziologiei celulare. În niciun caz o celulă nu poate fi considerată o entitate statică, deoarece există un schimb constant de substanțe între diferitele compartimente intracelulare, precum și între compartimente și mediu. Structurile celulare sunt în echilibru dinamic, iar interacțiunile celulelor între ele și cu mediul extern sunt o condiție necesară pentru menținerea vieții unui organism funcțional. În acest capitol ne vom uita la mecanismele fundamentale ale unui astfel de schimb. În capitolele următoare vor fi luate în considerare aceste mecanisme în măsura în care se aplică celulei nervoase și funcțiilor acesteia;
totuși, aceleași mecanisme stau la baza funcționării tuturor celorlalte organe.
Difuzie.Cel mai simplu proces de mișcare a materiei este difuzia. În soluții (sau gaze), atomii și moleculele se mișcă liber, iar diferențele de concentrație sunt echilibrate datorită difuziei. Să considerăm două volume pline cu lichid sau gaz (Fig. 1.3), în care substanțele au concentrații c 1 și c 2 și separate printr-un strat cu suprafața A și grosimea d. Curgerea materiei m în timpul t descris Prima lege a difuziei a lui Fick:
dm/ dt= DA/ d ( C 1 –С 2)=DA/ dD C(1)
unde D este coeficientul de difuzie, constant pentru o anumită substanță, solvent și temperatură. Într-o formă mai generală pentru diferența de concentrație dc la distanta dx
dm/dt= –D A dc/dx,(2)
debitul prin secţiunea A este proporţional cu gradientul de concentraţie dc/dx . Semnul minus apare în ecuație deoarece modificarea concentrației în direcția x este negativă.
Difuzia este cel mai important proces datorită căruia majoritatea moleculelor din soluțiile apoase se deplasează pe distanțe scurte. Acest lucru se aplică și mișcării lor în celulă, în măsura în care difuzia nu este împiedicată de membrane. Multe substanțe pot difuza liber prin membranele lipidice, în special apa și gazele dizolvate precum O 2 și CO 2 . Solubil în grăsime

Orez. 1.3.Schema de difuzie cantitativă. Cele două spații sunt separate printr-un strat de grosimedsi zona A. C – concentrație mare de particule în partea stângă a volumului, C: – concentrație scăzută de particule în partea dreaptă piese, suprafata roz– gradient de concentraţie în stratul de difuzie. Debit de difuzie dm/dt – vezi ecuația (1)
substanțele difuzează bine și prin membrane; Acest lucru se aplică și moleculelor polare de dimensiuni destul de mici, cum ar fi etanolul și ureea, în timp ce zaharurile trec cu dificultate prin stratul lipidic. În același timp, straturile lipidice sunt practic impermeabile la moleculele încărcate, inclusiv la ionii anorganici. Pentru non-electroliți, ecuația de difuzie (1) este de obicei transformată prin combinarea caracteristicilor membranei și a substanței care difuzează într-una singură. permeabilitatea parametrului (P):
dm/dt=P AD c.(3)
În fig. 1.4 sunt comparate permeabilitatea (P) a membranei lipidice pentru diferite molecule.
Difuzia prin porii membranei . Membrana plasmatică (și alte membrane celulare) este permeabilă nu numai la substanțele care difuzează prin stratul lipidic, ci și la mulți ioni, zaharuri, aminoacizi și nucleotide. Aceste substanțe traversează membrana prin porii formați proteine de transport, scufundat în membrană. În interiorul unor astfel de proteine există un canal umplut cu apă cu un diametru mai mic de 1 nm, prin care moleculele mici pot difuza. Se mișcă de-a lungul unui gradient de concentrație, iar dacă poartă o sarcină, atunci mișcarea lor prin canale este, de asemenea, reglată de potențialul membranei. Canalele de membrană sunt relativ selective

Orez. 1.4.Permeabilitatea straturilor duble lipidice artificiale la diferite substanțe
în raport cu tipul de molecule care pot trece prin ele, există, de exemplu, canale de potasiu, sodiu și calciu, fiecare dintre acestea fiind impermeabil la aproape orice ion, cu excepția unuia specific. Astfel de selectivitatea este determinată de încărcătura sau structura locurilor de legare din pereții canalului, care facilitează transportul unei anumite molecule și împiedică pătrunderea altor substanțe prin canal (Fig. 1.5, A) .
În spatele comportamentului canale ionice membranare ușor de observat, deoarece curentul rezultat din mișcarea ionilor poate fi măsurat, chiar și pentru un singur canal. Se arată că canalele spontan și cu frecvență înaltă își schimbă starea de la deschis la închis. Canalul de potasiu se caracterizează prin impulsuri de curent cu o amplitudine de aproximativ 2 pA (2 10 –12 A) și o durată de câteva milisecunde (vezi Fig. 2.12, p. 37) [3]. În această perioadă trec prin ea zeci de mii de ioni. Tranziția proteinelor de la o conformație la alta este studiată folosind difracția de raze X, spectroscopie Mössbauer și rezonanța magnetică nucleară (RMN). Proteinele sunt astfel structuri foarte dinamice, mobile, iar canalul care trece prin proteină nu este doar un tub rigid, umplut cu apă (Fig. 1.5, A), ci un labirint de grupuri moleculare și sarcini care se mișcă rapid. Această caracteristică dinamică a canalului se reflectă în profilul energetic al canalului, prezentată în fig. 1.5, B. Aici, axa absciselor reprezintă lungimea canalului de la soluția externă cu concentrație ionică C 0 și potențial 0 până la soluția internă cu concentrație C 1 și potențial E. Axa ordonatelor

Orez. 1.5.A. Schema unei proteine care formează un canal de potasiu încorporat în stratul dublu lipidic al membranei plasmatice. Patru sarcini negative sunt fixate pe „peretele” canalului. B. Profil energetic schematic al canalului prezentat în Fig. A. Axa ordonatelor arată valorile energiei cinetice necesare trecerii canalului; de-a lungul axei absciselor – distanța dintre suprafețele interioare și exterioare ale membranei. Minimele energetice corespund locurilor de legare ale ionilor încărcați pozitiv cu sarcini negative fixe în peretele canalului. Maximele energetice corespund obstacolelor de difuzie din canal. Se presupune că conformația proteinei canalului oscilează spontan; opțiunile profilului energetic sunt afișate cu linii continue și întrerupte; aceste oscilații facilitează foarte mult legarea ionilor la depășirea barierei energetice (modificat)
sunt prezentate nivelurile de energie ionică la locurile de legare a canalelor; vârful din grafic reprezintă bariera de permeabilitate pe care energia ionică trebuie să o depășească pentru a pătrunde în canal, iar „cufundarea” din grafic reprezintă o stare relativ stabilă (legare). În ciuda obstacolului vârfului de energie, ionul poate pătrunde în canal dacă profilul energetic se modifică spontan; ionul se poate găsi brusc „de cealaltă parte” a vârfului de energie și poate continua să se miște în celulă. În funcție de sarcina, dimensiunea și gradul de hidratare a ionului și capacitatea acestuia de a se lega de structurile peretelui canalului, profilul energetic al canalului variază pentru diferiți ioni, ceea ce poate explica selectivitatea tipurilor individuale de canale.
Echilibrul de difuzie al ionilor . Difuzia diferiților ioni prin canalele membranare ar trebui să conducă la eliminarea diferențelor de concentrație dintre mediile extracelulare și intracelulare. După cum, totuși, se poate observa din tabel. 1.1, astfel de diferențe rămân, prin urmare, trebuie să existe unele echilibruîntre difuzie și alte procese de transport prin membrană. Următoarele două secțiuni tratează modalitățile prin care se realizează acest echilibru. În cazul ionilor, echilibrul de difuzie este afectat de sarcina lor. Difuzia moleculelor neîncărcate este asigurată de diferența de concentrații DC , iar când concentrațiile sunt egalizate, transportul în sine se oprește. Particulele încărcate sunt afectate suplimentar de câmpul electric. De exemplu, atunci când un ion de potasiu părăsește o celulă de-a lungul gradientului său de concentrație, acesta poartă o singură sarcină pozitivă. Astfel, mediul intracelular devine mai încărcat negativ, rezultând o diferență de potențial de-a lungul membranei. Sarcina negativă intracelulară împiedică noi ioni de potasiu să părăsească celula, iar acei ioni care părăsesc celula vor crește și mai mult sarcina pe membrană. Fluxul ionilor de potasiu se oprește atunci când acțiunea câmpului electric compensează presiunea de difuzie din cauza diferenței de concentrație. Ionii continuă să treacă prin membrană, dar în cantități egale în ambele direcții. Prin urmare, pentru o diferență dată în concentrațiile ionilor de pe membrană, există potenţial de echilibru E ion la care se oprește fluxul ionilor prin membrană. Potențialul de echilibru poate fi ușor determinat folosind Ecuații Nernst:
Eion= RT/ zF* lnC afară/ C în(4)
unde R – constanta gazului, T – temperatura absolută, z – valența ionilor (negativ pentru anioni) C afară - concentrația ionilor extracelulari, C în – concentrația ionilor intracelulari, F Numărul Faraday. Dacă înlocuim constante în ecuație, atunci la temperatura corpului (T = 310 K) potențialul de echilibru pentru ionii de potasiu E K este egal cu:
Ek= –61 mjurnal B /(5)
Dacă [ K + out ]/[ K + in ] = 39, după cum urmează din tabel. 1.1 atunci
Ek= –61 m B log 39= –97 mV.
Într-adevăr, s-a constatat că toate celulele au potențial de membrană;în celulele musculare de mamifere nivelul său este de aproximativ -90 mV. În funcție de condiții și concentrațiile relative de ioni, celulele pot avea un potențial de membrană cuprins între -40 și -120 mV. Pentru celula din exemplul de mai sus (Tabelul 1.1) potential de odihna, egal cu aproximativ -90 mV, indică faptul că fluxurile ionilor de potasiu prin canalele membranare sunt aproximativ în echilibru. Acest lucru nu este surprinzător, deoarece într-o membrană de repaus starea deschisă a canalelor de potasiu este cea mai probabilă, adică. membrana este cea mai permeabilă la ionii de potasiu. Potențialul membranei este însă determinat de fluxurile altor ioni.
Ușurința cu care particulele neîncărcate pot difuza prin membrană este cuantificată în ecuația (3). Permeabilitatea la particulele încărcate este descrisă de o ecuație ceva mai complexă:
P= m RT/ dF(6)
Unde m- mobilitate ionică în membrană, d - grosimea membranei, a R, T și F – constante termodinamice cunoscute. Valorile permeabilității pentru diverși ioni determinate în acest fel pot fi utilizate pentru a calcula potențialul membranei Em , când ionii de potasiu, sodiu și clor trec prin membrană simultan (cu permeabilitate P K, PNa și PCI respectiv). Se presupune că potențialul scade uniform în membrană, astfel încât intensitatea câmpului este constantă. În acest caz se aplică Ecuația Goldman sau ecuația câmpului constant :
Em= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl )(7)
Pentru majoritatea membranelor celulare P K de aproximativ 30 de ori mai mare decât P N / A (vezi și secțiunea 1.3). Valoare relativă P Cl diferă în mare măsură; pentru multe membrane P Cl mic în comparație cu R K , cu toate acestea pentru alții (de exemplu, în mușchii scheletici) P Cl , semnificativ mai mare decât P K.
Transport activ, pompa de sodiu . Secțiunea anterioară descrie difuzia pasivă a ionilor și potențialul de membrană rezultat la concentrații date de ioni intra și extracelulari. Cu toate acestea, ca urmare a acestui proces, concentrația de ioni în interiorul celulei nu este stabilizată automat, deoarece membrana
potențialul este puțin mai electronegativ decât EK, și mult în comparație cu E N / A (aproximativ +60 mV). Datorită difuziei, concentrațiile ionilor intracelulari, cel puțin potasiu și sodiu, trebuie egalate cu cele extracelulare. Stabilitatea gradientului ionic se realizează prin transport activ: proteinele membranei transportă ionii prin membrană împotriva gradienților electrici și/sau de concentrație, consumând energie metabolică pentru aceasta. Cel mai important proces de transport activ este munca Na/K – o pompă care există în aproape toate celulele;
pompa pompează ioni de sodiu din celulă, pompând simultan ioni de potasiu în celulă. Aceasta asigură o concentrație intracelulară scăzută de ioni de sodiu și o concentrație mare de ioni de potasiu (Tabelul 1.1). Gradientul de concentrație al ionilor de sodiu pe membrană are funcții specifice legate de transmiterea informațiilor sub formă de impulsuri electrice (vezi Secțiunea 2.2), precum și menținerea altor mecanisme active de transport și reglarea volumului celular (vezi mai jos). Prin urmare, nu este de mirare că mai mult de 1/3 din energia consumată de o celulă este cheltuită pe pompa Na/K, iar în unele dintre cele mai active celule până la 70% din energie este cheltuită pentru funcționarea acesteia.
Proteina de transport Na/K este o ATPază. Pe suprafața interioară a membranei, acesta descompune ATP în ADP și fosfat (Fig. 1.6). Energia unei molecule de ATP este utilizată pentru a transporta trei ioni de sodiu din celulă și simultan doi ioni de potasiu în celulă, adică, în total, o sarcină pozitivă este îndepărtată din celulă într-un ciclu. Astfel, pompa Na/K este electrogen(creează un curent electric prin membrană), ceea ce duce la o creștere a electronegativității potențialului membranei cu aproximativ 10 mV. Proteina de transport efectuează această operație la viteză mare: de la 150 la 600 de ioni de sodiu pe secundă. Secvența de aminoacizi a proteinei de transport este cunoscută, dar mecanismul acestui transport metabolic complex nu este încă clar. Acest proces este descris folosind profile energetice ale transferului ionilor de sodiu sau potasiu de către proteine (Fig. 1.5,5). După natura modificărilor acestor profiluri, asociate cu schimbări constante în conformația proteinei de transport (un proces care necesită energie), se poate judeca stoichiometria schimbului: doi ioni de potasiu sunt schimbați cu trei ioni de sodiu.
Pompă Na/K, la fel ca izolată Na+ ATPaza membranară dependentă de /K+, inhibată în mod specific de glicozidul cardiac ouabain (strofantina). Deoarece funcționarea pompei Na/K este o reacție chimică în mai multe etape, aceasta, ca toate reacțiile chimice, depinde în mare măsură de temperatură, care

Orez. 1.6.Diagrama unei pompe-ATPaze Na/K (cufundată în stratul dublu lipidic al membranei plasmatice), care într-un ciclu elimină trei ioni Na + din celulă împotriva gradienților de potențial și concentrație și aduce doi ioni de K în celulă + . În timpul acestui proces, o moleculă de ATP este descompusă în ADP și fosfat. Diagrama prezintă ATPaza ca un dimer constând dintr-o subunități mari (funcționale) și mici; în membrană există ca un tetramer format din două subunităţi mari şi două mici
prezentată în fig. 1.7. Aici, fluxul ionilor de sodiu din celulele musculare este prezentat în raport cu timpul; aceasta este practic echivalentă cu fluxul de ioni de sodiu mediat de funcționarea pompei Na/K, deoarece fluxul pasiv de ioni de sodiu față de gradienții de concentrație și potențial este extrem de mic. Dacă medicamentul este răcit cu aproximativ 18 ° C, fluxul de ioni de sodiu din celulă va scădea rapid de 15 ori și imediat după încălzire va fi restabilit la nivelul inițial. Această scădere a fluxului de ioni de sodiu din celulă este de câteva ori mai mare decât cea care ar corespunde dependenței de temperatură a procesului de difuzie sau unei simple reacții chimice. Un efect similar este observat atunci când energia metabolică este epuizată ca urmare a intoxicației cu dinitrofenol (DNP) (Fig. 1.7,5). În consecință, fluxul ionilor de sodiu din celulă este asigurat de o reacție dependentă de energie - o pompă activă. O altă caracteristică a pompei, împreună cu dependența semnificativă de temperatură și energie, este prezența unui nivel de saturație (ca toate celelalte reacții chimice); aceasta înseamnă că viteza pompei nu poate crește la infinit pe măsură ce crește concentrația ionilor transportați (Fig. 1.8). În schimb, fluxul unei substanțe care difuzează pasiv crește proporțional cu diferența de concentrație în conformitate cu legea difuziei (ecuațiile 1 și 2).

Orez. 1.7. A, B. Transport Na activ + . Axa Y: flux de 24 Na + radioactiv din celulă (imp./min). Axa absciselor: timp de la începutul experimentului. A. Celula este răcită de la 18,3 °C la 0,5 °C; curgere Na+ din celulă în această perioadă este inhibată. B. Suprimarea fluxului de Na + din celulă cu dinitrofenol (DNP) la o concentrație de 0,2 mmol/l (modificat)
Pe lângă pompa Na/K, membrana plasmatică conține cel puțin încă o pompă - calciu; această pompă pompează ionii de calciu (Ca 2+) din celulă și este implicată în menținerea concentrației lor intracelulare la un nivel extrem de scăzut (Tabelul 1.1). Pompa de calciu este prezentă în densitate foarte mare în reticulul sarcoplasmatic al celulelor musculare, care acumulează ioni de calciu ca urmare a defalcării moleculelor de ATP (vezi capitolul 4).
Efectul pompei Na/K asupra potențialului membranei și volumului celulei . În fig. Figura 1.9 prezintă diferitele componente ale curentului membranar şi prezintă concentraţiile intracelulare ale ionilor care

Orez. 1.8.Relația dintre viteza de transport a moleculelor și concentrația lor (la intrarea în canal sau la locul de legare al pompei) în timpul difuziei prin canal sau în timpul transportului prin pompare. Acesta din urmă devine saturat la concentrații mari (viteza maximă, Vmax ) valoarea pe axa x corespunzătoare jumătate din viteza maximă a pompei ( Vmax /2), este concentrația de echilibru LA m

Orez. 1.9.Diagrama care arată concentrațiile de Na+ , K+ și Cl – în interiorul și în exteriorul celulei și căile de penetrare a acestor ioni prin membrana celulară (prin canale ionice specifice sau folosind o pompă Na/K. La gradienți de concentrație dat, potențialele de echilibru E Na, E K și E С l – egale cu cele indicate, potenţial de membrană Em = – 90 mV
asigura existenta acestora. Un curent exterior de ioni de potasiu este observat prin canalele de potasiu, deoarece potențialul de membrană este puțin mai electropozitiv decât potențialul de echilibru pentru ionii de potasiu. Conductanța generală a canalelor de sodiu este mult mai mică decât cea a canalelor de potasiu, adică canalele de sodiu sunt deschise mult mai rar decât canalele de potasiu la potenţial de repaus; cu toate acestea, aproximativ același număr de ioni de sodiu intră în celulă pe măsură ce ionii de potasiu ies din celulă, deoarece sunt necesare concentrații mari și gradienți potențiali pentru ca ionii de sodiu să difuzeze în celulă. Pompa Na/K oferă compensarea ideală pentru curenții de difuzie pasivă, deoarece transportă ionii de sodiu din celulă și ionii de potasiu în ea. Astfel, pompa este electrogenă datorită diferenței dintre numărul de sarcini transferate în și din celulă, care la viteza normală de funcționare creează un potențial de membrană de aproximativ 10. mV este mai electronegativ decât dacă ar fi produs numai de fluxuri de ioni pasivi (vezi ecuația 7). Ca urmare, potențialul de membrană se apropie de potențialul de echilibru al potasiului, ceea ce reduce scurgerea ionilor de potasiu. Na activitate Pompa /K este reglată de concentrația intracelulară a ionilor de sodiu. Viteza pompei scade pe măsură ce concentrația ionilor de sodiu care urmează să fie îndepărtați din celulă scade (Fig. 1.8), astfel încât funcționarea pompei și fluxul de ioni de sodiu în celulă se echilibrează reciproc, menținând concentrația intracelulară de sodiu. ioni la un nivel de aproximativ 10 mmol/L.
Pentru a menține echilibrul între curenții de pompare și pasivi de membrană, sunt necesare mult mai multe molecule de pompă Na/K decât proteinele canal pentru ionii de potasiu și sodiu. Când canalul este deschis, zeci de mii de ioni trec prin el în câteva milisecunde (vezi mai sus), iar din moment ce canalul se deschide de obicei de mai multe ori pe secundă, în total trec prin el mai mult de 10 5 ioni în acest timp. O singură proteină cu pompă mișcă câteva sute de ioni de sodiu pe secundă, astfel încât membrana plasmatică trebuie să conțină de aproximativ 1000 de ori mai multe molecule de pompă decât moleculele canal. Măsurătorile curenților de canal în repaus au arătat prezența în medie a unui canal deschis de potasiu și de sodiu pe membrană de 1 μm 2; De aici rezultă că aproximativ 1000 de molecule ale pompei Na/K ar trebui să fie prezente în același spațiu, adică. distanța dintre ele este în medie de 34 nm; Diametrul proteinei pompei, ca proteină canal, este de 8-10 nm. Astfel, membrana este destul de dens saturată cu molecule de pompare
Faptul că fluxul de ioni de sodiu în celulă și ionii de potasiu din celulă este compensat de funcționarea pompei, există o altă consecință, care constă în menținerea unei presiuni osmotice stabile și volum constant.În interiorul celulei există o concentrație mare de anioni mari, în principal proteine (A - în tabelul 1.1), care nu sunt capabili să pătrundă în membrană (sau să pătrundă prin ea foarte lent) și, prin urmare, sunt o componentă fixă în interiorul celulei. Pentru a echilibra sarcina acestor anioni, este nevoie de un număr egal de cationi. Datorită acțiunii pompei Na/K, acești cationi sunt în principal ioni de potasiu. O creștere semnificativă a concentrației ionilor intracelulari ar putea avea loc numai cu o creștere a concentrației de anioni datorită fluxului de C1 - de-a lungul gradientului de concentrație în celulă (Tabelul 1.1), dar potențialul de membrană contracarează acest lucru. Curent de intrare Cl – observat numai până la atingerea potențialului de echilibru pentru ionii de clor; acest lucru se observă atunci când gradientul ionic de clor este aproape opus gradientului ionic de potasiu, deoarece ionii de clor sunt încărcați negativ (Ecuația 4). Astfel, se stabilește o concentrație intracelulară scăzută a ionilor de clor, corespunzătoare unei concentrații extracelulare scăzute a ionilor de potasiu. Rezultatul este o limitare a numărului total de ioni din celulă. Dacă potențialul membranei scade atunci când pompa Na/K este blocată, de exemplu în timpul anoxiei, atunci potențialul de echilibru pentru ionii de clor scade, iar concentrația intracelulară a ionilor de clor crește în consecință. Restabilind echilibrul sarcinilor, ionii de potasiu intră și ei în celulă; concentrația totală de ioni din celulă crește, ceea ce crește presiunea osmotică; aceasta forțează apa să intre în celulă. Celula se umfla. Se observă această umflare in vivo în condiţii de lipsă de energie.
Gradient de concentrație N / A + ca forță motrice pentru transportul membranei . Importanța pompei Na/K pentru celulă nu se limitează la stabilizarea gradienților normali de K+ și Na+ pe membrană. Energia stocată în gradientul membranei Na+ , este adesea folosit pentru a facilita transportul membranar al altor substanțe. De exemplu, în Fig. 1.10 arată „simport” Na+ și molecule de zahăr în celulă. Proteina de transport membranar transportă molecula de zahăr în celulă chiar și împotriva unui gradient de concentrație, în timp ce, în același timp, Na + se mișcă de-a lungul gradientului de concentrație și potențial, furnizând energie pentru transportul zaharurilor. Acest transport de zaharuri este in intregime dependent de existenta unui gradient mare Na+ ; dacă concentraţia intracelulară Na+ creste semnificativ, transportul zaharurilor se opreste. Pentru diverse c akharov, există diferite sisteme de simport. Transportul aminoacizilorîntr-o celulă asemănătoare transportului c akharov prezentat în fig. 1,10; este asigurat si de gradient Na+ , Există cel puțin cinci sisteme de transport diferite, fiecare specializat pentru un grup de aminoacizi înrudiți.
Pe lângă sistemele de simport, există și „anti-portari”. Unul dintre ei, de exemplu, transferă un ion de calciu din celulă într-un ciclu, în schimbul a trei ioni de sodiu (Fig. 1.10). Energie pentru Ca 2+ transport se formează ca urmare a pătrunderii a trei ioni de sodiu de-a lungul unui gradient de concentrație și potențial. Această energie este suficientă (la potențialul de repaus) pentru a menține un gradient ridicat de ioni de calciu (de la mai puțin de 10–7 mol/L în interiorul celulei la aproximativ 2 mmol/L în afara celulei).
Endo- și exocitoză . Pentru anumite substanțe care intră în celulă sau trebuie excretate

Orez. 1.10.Proteinele încorporate în stratul dublu lipidic al membranei mediază simportul glucozei și Na+ în celulă, precum și Ca2+/Na+ –antiport, în care forța motrice este gradientul Na + pe membrana celulară
din el, nu există canale de transport; Aceste substanțe includ, de exemplu, proteine și colesterol. Ele pot trece prin membrana plasmatică în vezicule, sau bule, prin endo- și exocitoză. În fig. Figura 1.11 prezintă principalele mecanisme ale acestor procese. În exocitoză, anumite organele (vezi mai jos) formează vezicule pline cu o substanță care trebuie îndepărtată din celulă, cum ar fi hormonii sau enzimele extracelulare. Când astfel de vezicule ajung la membrana plasmatică, membrana lor lipidică fuzionează cu aceasta, permițând astfel conținutului să scape în mediul extern. În procesul opus, endocitoza, membrana plasmatică se invaginează, formând o groapă, care apoi se adâncește și se închide, formând o veziculă intracelulară umplută cu lichid extracelular și câteva macromolecule. Pentru a asigura această fuziune a membranei și închiderea veziculei, elementele contractile ale citoscheletului acționează împreună cu membranele în sine (vezi mai jos). Endocitoza nu implică întotdeauna pur și simplu capturarea mediului extracelular în celulă. Membrana celulară conține adesea receptori specifici pentru macromolecule, cum ar fi insulina sau antigenele, adesea organizate în grupuri specializate. După ce aceste macromolecule se leagă de receptorii lor, apare endocitoza în zona membranei din jurul receptorului, iar macromolecula este transportată selectiv în celulă (Fig. 1.12, B).
Endo- și exocitoza apar continuu în celule. Cantitatea de rotație a materialului membranar este semnificativă; în decurs de 1 oră, macrofagul absoarbe suprafața dublă a membranei sale citoplasmatice sub formă de vezicule. În majoritatea celulelor, rotația materialului membranei nu are loc la fel de intens, dar ar trebui să fie totuși semnificativă.

Orez. 1.11.Exocitoză și endocitoză. Sus: vezicula intracelulară fuzionează cu stratul dublu lipidic al membranei plasmatice și se deschide în spațiul extracelular. Acest proces se numește exocitoză. În partea de jos: membrana plasmatică se invaginează într-o zonă mică și eliberează o veziculă plină cu material extracelular. Acest proces se numește endocitoză
1.3. Transportul substanțelor în interiorul celulei
Endo- și exocitoza nu sunt numai procese de transport de substanțe prin membrana celulară, ci și procese de schimb de membrane - componentele structurale ale celulei în sine. Subiectul de luat în considerare în această secțiune sunt alte procese de transport similare în celulă și organelele sale.
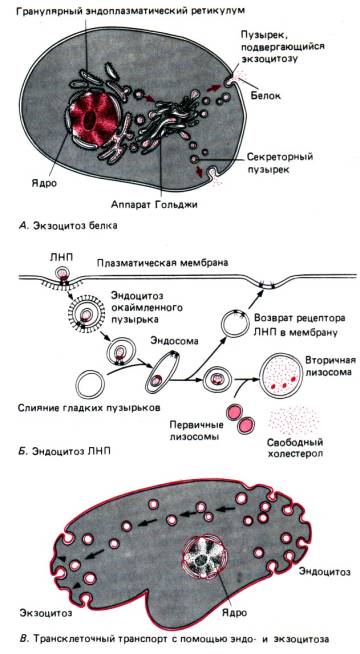
Orez. 1.12. A–B. Schema proceselor inclusiv exo- și endocitoza. A. Proteina sintetizată în reticulul endoplasmatic granular este transportată prin aparatul Golgi către membrana plasmatică, unde este secretată prin exocitoză. B. Colesterolul legat de particulele de LDL (lipoproteine cu densitate joasă) se atașează de membrana plasmatică, induce formarea unei vezicule endocitare în această regiune a membranei și este transportat la lizozomi, unde este eliberat. ÎN. Material extracelular capturat în timpul endocitozei (în figură pe dreapta), transportat prin celulă în vezicule sau vezicule și eliberat prin exocitoză (în figură stânga)
Difuzia . În mod firesc, în citosol diferența de concentrații este eliminată datorită difuziei; același lucru este valabil și pentru fluidele conținute în organele. Datorită concentrației mari de proteine dizolvate, difuzia aici este mult mai lentă decât în apă. Membranele lipidice - în jurul celulelor și în interiorul organelelor - sunt fluide bidimensionale în care are loc difuzia. Lipidele dintr-un strat dublu de membrană difuzează în propriul lor strat, mișcându-se rar de la unul la altul. Proteinele scufundate în ele sunt și ele destul de mobile; se rotesc în jurul unei axe perpendiculare pe membrană sau difuzează lateral cu constante de difuzie foarte diferite, de 2–10.000 de ori mai lente decât fosfolipidele. Deci, dacă unele proteine se mișcă liber în stratul lipidic și cu aceeași viteză cu moleculele lipidice în sine, atunci altele sunt ancorate, adică. destul de ferm legat de citoschelet. Există agregate „permanente” de proteine specifice în membrană, de exemplu, structuri pre- și postsinaptice ale celulelor nervoase. Proteinele care se mișcă liber pot fi demonstrate prin legarea lor de coloranți fluorescenți, a căror luminescență este indusă prin iluminarea scurtă a unei zone mici a membranei. Astfel de experimente arată că, în mai puțin de 1 minut, proteinele legate de colorant sunt distribuite uniform pe membrană pe distanțe de până la 10 μm.
Transport activ în membranele organitelor .
Procesele de transport activ, care joacă un rol vital în funcționarea membranei plasmatice, au loc și în interiorul celulei, în membranele organitelor. Conținutul specific al diferitelor organite este creat parțial datorită sintezei interne și parțial datorită transportului activ din citosol. Un exemplu al acestuia din urmă este pompa de Ca2+ menționată mai sus din reticulul sarcoplasmatic al celulelor musculare. Este deosebit de interesant că, în cazul sintezei ATP în mitocondrii, principiul opus se aplică la ceea ce se întâmplă în pompele ATPazei ale membranei plasmatice (Fig. 1.6). În timpul sintezei ATP, metabolismul oxidativ duce la formarea unui gradient abrupt H+ pe membranele interne. Acest gradient este forța motrice pentru procesul invers al ciclului de pompare al transportului activ al moleculelor: ionii H + se deplasează prin membrană de-a lungul gradientului, iar energia eliberată ca rezultat asigură sinteza ATP din ADP și fosfat. ATP-ul rezultat, la rândul său, furnizează energie celulei, inclusiv pentru transportul activ.
Transport în vezicule . Celula are un număr mare de organite și vezicule asociate (Fig. 1.1). Aceste organite, si mai ales veziculele, sunt in continua miscare, transportand continutul lor catre alte organite sau catre membrana plasmatica. Veziculele pot migra, de asemenea, de la membrana celulară la organele, ca în endocitoză.
Proces secretia de proteine prezentată în fig. 1.12, A. Proteina este sintetizată în apropierea nucleului celular pe ribozomii asociați cu reticulul endoplasmatic (așa-numitul reticul endoplasmatic granular sau rugos); Odată ajunsă în reticulul endoplasmatic, proteina este ambalată în vezicule de transport, care se separă de organele și migrează către aparatul Golgi. Aici se contopesc cu cisternele aparatului Golgi, unde proteina este modificată (adică, transformată într-o glicoproteină). La capetele cisternelor, veziculele se separă din nou. Veziculele secretoare care transportă proteina modificată se deplasează către membrana plasmatică și își eliberează conținutul prin exocitoză.
Un alt exemplu de cale de transport într-o celulă este prezentat în Fig. 1,12, B; Aceasta este absorbția colesterolului de către celulă. Colesterolul transportat în sânge este legat în principal de proteine, cum ar fi particulele "lipoproteine cu densitate joasa"(LDL). Astfel de particule se atașează de zone specifice membranei care conțin receptori LDL, unde are loc endocitoza și LDL este transportat în celulă în vezicule „mărginite”. Aceste vezicule fuzionează pentru a forma endozomi și își pierd „marginile” în timpul acestui proces. Endozomii, la rândul lor, fuzionează cu lizozomii primari, conținând predominant enzime hidrolitice, și formează lizozomi secundari, mai mari. În ele, colesterolul este eliberat din particulele LDL și difuzează în citosol, unde devine disponibil, de exemplu, pentru sinteza membranelor lipidice. Veziculele care nu conțin LDL sunt, de asemenea, separate de endozomi, care se deplasează într-un mod special către membrana plasmatică și se contopesc cu aceasta, returnând materialul membranar și, probabil, receptorii pentru LDL. Din momentul în care particula LDL se leagă de membrană până când colesterolul este eliberat din lizozomul secundar, trec 10-15 minute. Deficiențele în legarea și absorbția LDL, adică în furnizarea de colesterol către celulă, joacă un rol decisiv în dezvoltarea unei boli grave și răspândite, ateroscleroza („întărirea” arterelor).
Există multe alte rute de transport similare cu cele prezentate în Fig. 1.11 și 1.12, A, cu ajutorul cărora vezicule specifice se mișcă în celulă. Nu se știe exact cum se mișcă, dar elementele citoscheletice sunt probabil implicate în acest proces. Veziculele pot aluneca de-a lungul microtubulilor, caz în care energia pentru mișcare este aparent furnizată de proteina asociată veziculelor, ATPaza (vezi mai jos). Rămâne complet neclar câte vezicule diferite, mișcându-se una după alta în toate direcțiile, ajung la destinație. Evident, trebuie să fie „marcate” într-un mod care să fie recunoscut de sistemul de transport și transformat în mișcare țintită.
Transport prin formarea și distrugerea organelelor . Până acum, am considerat endo- și exocitoza drept procese de transport a conținutului veziculelor. Există un alt aspect al acestor procese, care constă în faptul că îndepărtarea țintită a membranei plasmatice pe o parte a suprafeței celulare prin endocitoză și, dimpotrivă, adăugarea acesteia pe alta prin exocitoză, deplasează suprafețe mari ale membranei (Fig. 1.12.E), oferind celulei posibilitatea, de exemplu, de a forma o excrescere sau de a muta.
Rearanjamente similare sunt, de asemenea, tipice pentru citoschelet, în special pentru microfilamente și microtubuli (Fig. 1.1). Microfilamente constau în primul rând din proteina F-actină, care este capabil de asamblare în fascicule fibroase ca urmare a polimerizării monomerului din citosol. Fasciculele sunt polarizate, adică cresc adesea doar la un capăt, acumulând noi molecule de actină, în timp ce celălalt capăt este inert sau aici are loc dezasamblarea. Datorită acestei creșteri polarizate, microfilamentele se mișcă eficient, iar structura rețelei lor se poate modifica. Trecerea actinei dintr-o stare depolimerizată (sol) la o stare organizată (gel) poate avea loc foarte rapid sub influența altor proteine sau modificări ale concentrației ionilor (vezi mai jos). Există, de asemenea, proteine care determină descompunerea filamentelor de actină pentru a forma fragmente scurte. Proiecțiile subțiri ale multor celule - filopodii - conțin un fascicul central de actină (Fig. 1.1), iar diferitele mișcări ale filopodiilor se datorează probabil tranzițiilor actinei: polimerizare - depolimerizare.
Microtubulide asemenea, suferă adesea mișcări similare. Mecanismul acestor mișcări este similar - polimerizarea tubulinei din citosol în așa fel încât un capăt al microtubulului să crească, în timp ce celălalt fie nu se schimbă, fie dezasamblarea are loc acolo. Astfel, un microtubul, prin adăugarea sau îndepărtarea corespunzătoare a materialului, se poate deplasa prin citosol.
Mișcări active ale citoscheletului . Modificări ale structurilor citoscheletice pot apărea ca rezultat atât al mișcărilor active, cât și al rearanjamentelor descrise mai sus. În multe cazuri, mișcarea microtubulilor și a filamentelor de actină este condusă de proteinele contractile care leagă filamentele sau tubulii și le pot muta unul față de celălalt. Veverițe miozina si dineina prezent în citosolul tuturor celulelor în concentrații relativ mari; sunt elementele care transformă energia în mișcare în celulele specializate (mușchi) și organele (cilii). În celulele musculare, miozina formează filamente groase orientate paralel cu filamentele de actină. Molecula de miozină cu „capul” său se atașează de filamentul de actină și, folosind energia ATP, deplasează miozina de-a lungul moleculei de actină. Miozina se desprinde apoi de actină. Combinația multor astfel de cicluri de conectare-deconectare duce la macroscopic contracția fibrelor musculare(capitolul 4). Dineina joacă un rol similar în mișcarea microtubulilor în timpul cililor (Fig. 1.1). În citoplasma celulelor nespecializate, miozina și dineina nu formează fibre regulate, ci în cele mai multe cazuri grupuri mici de molecule. Chiar și în astfel de agregate mici, ele sunt capabile să miște filamente de actină sau microtubuli. Orez. Figura 1.13 ilustrează acest proces atunci când moleculele de miozină cu polarizare opusă sunt atașate de asemenea la două filamente de actină, polarizate în direcții diferite. Grupurile de cap de miozină se îndoaie spre coada moleculei, consumând ATP în acest proces, iar cele două filamente de actină se deplasează în direcția opusă, după care miozina se desprinde de ele. Mișcările de acest fel, în timpul cărora energia ATP este convertită în lucru mecanic, pot schimba forma citoscheletului și, în consecință, a celulei și, de asemenea, asigură transportul organelelor asociate cu citoscheletul.
Procesele de transport intracelular pot fi demonstrate cel mai clar pe axonul unei celule nervoase. Transportul axonilor este discutat aici în detaliu pentru a ilustra evenimentele care pot avea loc în moduri similare în majoritatea celulelor. Un axon care are doar câțiva microni în diametru poate atinge o lungime de un metru sau mai mult, iar mișcarea proteinelor prin difuzie de la nucleu la capătul distal al axonului ar dura ani. Se știe de mult timp că atunci când orice parte a axonului suferă constricție, partea axonului situată mai proximal se extinde. Se pare că fluxul centrifugal este blocat în axon. Astfel de flux-transport rapid axonilor poate fi demonstrat prin mișcarea markerilor radioactivi, ca în experimentul prezentat în Fig. 1.14. Leucina marcată radioactiv a fost injectată în ganglionul rădăcinii dorsale, iar apoi radioactivitatea a fost măsurată în nervul sciatic la o distanță de 166 mm de corpurile celulare neuronale din a 2-a până la a 10-a oră. Peste 10 ore, vârful radioactivității la locul injectării s-a modificat ușor. Dar valul de radioactivitate s-a răspândit de-a lungul axonului cu o viteză constantă de aproximativ 34 mm în 2 ore, sau 410 mm/zi. S-a demonstrat că la toți neuronii animalelor homeoterme, transportul axonal rapid are loc cu aceeași viteză și nu se observă diferențe vizibile între fibrele subțiri, nemielinice și cei mai groși axoni, precum și între fibrele motorii și senzoriale. De asemenea, tipul de marker radioactiv nu afectează viteza de transport axonal rapid; markerii pot servi ca o varietate de radioactivi

Orez. 1.13.Complexul de miozină non-musculară, cu o anumită orientare, se poate lega de filamente de actină de polaritate diferită și, folosind energia ATP, le poate deplasa unul față de celălalt.
molecule, cum ar fi diverși aminoacizi, care sunt incluse în proteinele corpului celular al neuronului. Dacă analizăm partea periferică a nervului pentru a determina natura purtătorilor de radioactivitate transportați aici, atunci astfel de purtători se găsesc mai ales în fracția proteică, dar și în compoziția mediatorilor și a aminoacizilor liberi. Știind că proprietățile acestor substanțe sunt diferite și dimensiunile moleculelor lor sunt deosebit de diferite, putem explica viteza constantă de transport doar printr-un mecanism de transport comun tuturor acestora.
Descris mai sus transport rapid axonilor este anterograd, adică direcționat departe de corpul celular. S-a demonstrat că unele substanțe se deplasează de la periferie către corpul celular folosind transport retrograd. De exemplu, acetilcolinesteraza este transportată în această direcție cu o viteză de 2 ori mai mică decât viteza de transport axonal rapid. Un marker folosit adesea în neuroanatomie, peroxidaza de hrean, se mișcă și prin transport retrograd. Transportul retrograd joacă probabil un rol important în reglarea sintezei proteinelor în corpul celular. La câteva zile după tăierea axonului, se observă cromatoliza în corpul celular, ceea ce indică o întrerupere a sintezei proteinelor. Timpul necesar cromatolizei se corelează cu durata transportului retrograd de la locul transecției axonilor la corpul celular. Acest rezultat sugerează și o explicație pentru această tulburare - transmiterea de la periferie a „substanței semnal” care reglează sinteza proteinelor este întreruptă. Este evident că principalele „vehicule” utilizate pentru axonal rapid
Orez. 1.14.Experiment care demonstrează transportul axonal rapid în fibrele senzoriale ale nervului sciatic al pisicii. Leucina marcată cu tritiu este injectată în ganglionul rădăcinii dorsale și radioactivitatea în ganglion și fibrele senzoriale este măsurată la 2, 4, 6, 8 și 10 ore după injectare. (partea de jos a imaginii). De axa x Este trasată distanța de la ganglion până la zonele nervului sciatic unde se face măsurarea. Pe axa ordonatelor numai pentru curbele superioare și inferioare, radioactivitatea (pulsuri/min) este reprezentată pe o scară logaritmică. „Val” de radioactivitate crescută (săgeți) se deplasează cu o viteză de 410 mm/zi (de-a lungul)
transportul sunt vezicule (vezicule) și organele, precum mitocondriile, care conțin substanțe care trebuie transportate. Mișcarea celor mai mari vezicule sau mitocondrii poate fi observată cu ajutorul unui microscop in vivo . Astfel de particule fac mișcări scurte și rapide într-o direcție, se opresc, adesea se mișcă ușor înapoi sau în lateral, se opresc din nou și apoi se smuciază în direcția principală. 410 mm/zi corespunde unei viteze medii de mișcare anterogradă de aproximativ 5 μm/s; viteza fiecărei mișcări individuale trebuie, așadar, să fie mult mai mare, iar dacă luăm în considerare dimensiunile organelelor, filamentelor și microtubulilor, atunci aceste mișcări sunt într-adevăr foarte rapide. Transportul axonal rapid necesită o concentrație semnificativă de ATP. Otrăvurile precum colchicina, care perturbă microtubulii, blochează și transportul axonal rapid. De aici rezultă că în procesul de transport pe care îl luăm în considerare, veziculele și organelele se deplasează de-a lungul microtubulilor și filamentelor de actină; această mișcare este asigurată de mici agregate de molecule de dineină și miozină, acționând așa cum se arată în Fig. 1.13, folosind energia ATP.
Transportul rapid axonilor poate fi, de asemenea, implicat procese patologice. Unii virusuri neurotropice (de exemplu, virusurile herpes sau polio) intră în axon la periferie și se deplasează prin transport retrograd către corpul neuronului, unde se înmulțesc și își exercită efectele toxice. Toxina tetanica, o proteina produsa de bacteriile care patrund in organism atunci cand pielea este deteriorata, este captata de terminatiile nervoase si transportata in corpul neuronului, unde provoaca spasme musculare caracteristice. Există cazuri cunoscute de efecte toxice asupra transportului axonilor în sine, de exemplu, expunerea la solventul industrial acrilamidă. În plus, se crede că patogeneza deficienței de vitamine beriberi și polineuropatia alcoolică implică o încălcare a transportului axonal rapid.
Pe lângă transportul axonal rapid în celulă, există și destul de intens transport lent axonilor. Tubulina se deplasează de-a lungul axonului cu o viteză de aproximativ 1 mm/zi, iar actina se mișcă mai repede – până la 5 mm/zi. Alte proteine migrează cu aceste componente citoscheletice; de exemplu, enzimele par a fi asociate cu actina sau tubulina. Ratele de mișcare a tubulinei și actinei sunt aproximativ în concordanță cu ratele de creștere găsite pentru mecanismul descris anterior, în care moleculele sunt încorporate în capătul activ al unui microtubul sau microfilament. Prin urmare, acest mecanism poate sta la baza transportului axonal lent. Viteza de transport axonal lent corespunde, de asemenea, aproximativ cu viteza de creștere a axonilor, ceea ce aparent indică restricții impuse de structura citoscheletului asupra celui de-al doilea proces.
În încheierea acestei secțiuni, trebuie subliniat faptul că celulele nu sunt deloc structuri statice, așa cum apar, de exemplu, în fotografiile microscopice electronice. Membrană plasmatică si in special organitele sunt într-o mișcare rapidă constantă și o restructurare constantă; de aceea sunt capabili să funcţioneze. Mai mult, acestea nu sunt simple camere în care au loc reacții chimice, ci conglomerate foarte organizate de membrane și fibre,în care reacţiile apar într-o succesiune organizată optim.
1.4. Reglarea funcțiilor celulare
Menținerea celulei individuale ca unitate funcțională este reglementată în primul rând de nucleu; studiul unor astfel de mecanisme de reglare face obiectul biologiei celulare și al biochimiei. În același timp, celulele trebuie să își modifice funcțiile în conformitate cu condițiile de mediu și cu nevoile altor celule ale corpului, adică servesc ca obiecte de reglare funcțională. Mai jos vom analiza pe scurt modul în care acţionează aceste influenţe reglatoare asupra membranei plasmatice şi cum ajung în organele intracelulare.
Efecte de reglare asupra membranei celulare
Potențial de membrană . În multe cazuri, reglarea funcțiilor celulare se realizează prin modificarea potențialului membranei. Modificările locale ale potențialului sunt posibile atunci când: 1) curentul dintr-o zonă adiacentă a celulei sau generat de o altă celulă trece prin membrană; 2) se modifică concentrația extracelulară a ionilor (deseori [K + ] afară ); 3) canalele ionice membranare se deschid. Modificările potenţialului de membrană pot afecta conformaţia proteinelor membranei, determinând, în special, deschiderea sau închiderea canalelor. După cum este descris mai sus, funcționarea unor pompe cu membrană depinde de potențialul membranei. Celulele nervoase sunt specializate să perceapă modificările potențialului membranei ca informații care trebuie procesate și transmise (vezi capitolul 2).
Substanțe reglatoare extracelulare . Cel mai important mecanism de reglare care implică substanțele extracelulare este interacțiunea acestora cu receptorii specifici de pe membrana plasmatică sau din interiorul celulei. Aceste substanțe includ transmițători sinaptici, care transmit informații între celulele nervoase, agenți locali și substanțe care circulă în sânge și ajung la toate celulele corpului, cum ar fi hormonii și antigenii. Transmițători sinaptici sunt molecule mici eliberate din terminațiile nervoase de la sinapsă;
când ajung la membrana plasmatică a unei celule vecine, postsinaptice, declanșează semnale electrice sau alte mecanisme de reglare. Această problemă este discutată în detaliu în Cap. 3.
Agenți chimici locali secretat adesea de celule specializate. Acestea difuzează liber în spațiul extracelular, dar acțiunea lor este limitată la un grup restrâns de celule din cauza distrugerii rapide a acestor substanțe, fie spontan, fie sub acțiunea enzimelor. Un exemplu de eliberare a unor astfel de agenți este eliberarea histamina mastocite din cauza leziunilor sau a răspunsului imun. Histamina determină relaxarea celulelor musculare netede vasculare, crește permeabilitatea endoteliului vascular și stimulează terminațiile nervoase senzoriale care mediază senzația de mâncărime. Alți agenți chimici locali sunt eliberați de multe alte celule. Agenții locali tipici sunt prostaglandine, alcătuind un grup de aproximativ 20 de derivaţi ai acizilor graşi. Ele sunt eliberate continuu din celulele larg distribuite, dar acționează doar local, deoarece sunt distruse rapid de fosfolipazele membranare. Diverse prostaglandine au o gamă largă de acțiuni: pot declanșa contracția celulelor musculare netede, pot provoca agregarea trombocitelor din sânge (trombocite) sau pot suprima dezvoltarea corpului galben în ovare.
Alți agenți locali servesc factori de crestere. Cel mai cunoscut este factorul de creștere a nervilor (NGF) pentru neuronii simpatici, care este necesar pentru creșterea și supraviețuirea acestor neuroni în timpul dezvoltării. in vivo sau în cultura celulară. Aparent, celulele țintă pentru această clasă de neuroni secretă NGF și, prin urmare, asigură o inervație adecvată. Atunci când formează organe, celulele trebuie adesea să „găsească drumul” spre celulele țintite, care pot fi situate la distanțe considerabile. În consecință, trebuie să existe mulți factori de creștere specializați similari cu NGF.
Hormoni și antigeni transportat de sânge către toate celulele. Antigenele declanșează un răspuns imun din partea celulelor care poartă anticorpi specifici. Cu toate acestea, antigenele, de regulă, sunt substanțe străine care nu sunt produse în organismul care reacţionează (pentru mai multe detalii, vezi capitolul 18). Unii hormoni, cum ar fi insulina sau tiroxina, afectează o mare varietate de tipuri de celule, în timp ce alții, cum ar fi hormonii sexuali, afectează doar un anumit tip de celulă. Hormonii sunt fie peptide, a căror acțiune este declanșată prin legarea la un receptor de pe membrana celulară, fie steroizi și tiroxina, care difuzează prin membrana lipidică și se leagă de receptorii intracelulari. Hormonii steroizi se leagă de cromatina nucleară, ducând la transcrierea anumitor gene. Proteinele rezultate provoacă modificări ale funcțiilor celulare, care este efectul specific al hormonilor. Problemele legate de eliberarea și acțiunea hormonilor sunt discutate în detaliu în capitolul. 17.
Comunicarea intracelulară care implică mesageri secundi
Funcțiile de reglare descrise mai sus includ efecte asupra membranei celulare. Informațiile primite de membrana celulară trebuie adesea să determine reacția organelelor și sunt transferate acestora prin diferite substanțe cunoscute sub denumirea de mesageri secundi (spre deosebire de primii mesageri, care vin în celulă din surse externe). Studiul mediatorilor secund evoluează rapid și nu există nicio garanție că nivelul actual de înțelegere a problemei va fi suficient de complet. Aici vom atinge trei mediatori bine studiați: Ca 2+ , cAMP și inozitol trifosfat.
Calciu.Cel mai simplu mediator intracelular este ionul Ca 2+. Concentrația sa liberă într-o celulă de repaus este foarte scăzută și se ridică la 10_–8–10–7 mol/l. Poate pătrunde în celulă prin anumite canale membranare atunci când sunt în stare deschisă, de exemplu, când se modifică potențialul membranei (vezi capitolul 2). Creșterea rezultată a concentrației de Ca 2+ declanșează reacții importante în celulă, cum ar fi contracția miofibrilelor, care este baza contracției musculare (vezi capitolul 4), sau eliberarea veziculelor care conțin neurotransmițători de la terminațiile nervoase (vezi capitolul 3) . Ambele reacții necesită o concentrație de Ca 2+ de aproximativ 10 –5 mol/L. Ca 2+, care are un efect de reglare, poate fi eliberat și din depozitele intracelulare, cum ar fi reticulul endoplasmatic. Eliberarea Ca 2+ din depozit necesită participarea altor intermediari (vezi, de exemplu, Fig. 1.16).
Adenozin monofosfat ciclic, cAMP. Recent, s-a dovedit că adenozin monofosfatul ciclic (cAMP), un derivat al sursei principale de energie a organismului, ATP, este un al doilea mesager important. Lanțul complex de reacții prezentat în Fig. 1.15, începe cu receptor R s pe suprafața exterioară a membranei plasmatice, care poate servi ca loc de legare specifică pentru diferiți mediatori și hormoni. După legarea de o anumită moleculă „stimulatoare”. R s își schimbă conformația; aceste modificări afectează proteina G s pe suprafața interioară a membranei în așa fel încât să devină posibilă activarea acesteia din urmă de către guanozin trifosfat intracelular (GTP). Proteine activate G s , la rândul său, stimulează enzima de pe suprafața interioară a membranei, adenilat ciclaza (AC), care catalizează formarea cAMP din ATP. cAMP solubil în apă este mediatorul care transmite efectul

Orez. 1.15.Un lanț de reacții care implică mesagerul intracelular cAMP (adenozin monofosfat ciclic). Semnalele externe excitatorii sau inhibitorii activează receptorii membranari R s sau Ri . Acești receptori reglează procesul de legare G – proteine cu GTP intracelular (guanozin trifosfat), stimulând sau inhibând astfel adenilat ciclaza (AC) intracelular. Enzima de amplificare AC transformă adenozin trifosfatul (ATP) în cAMP, care este apoi descompus în AMP de către fosfodiesterază. AMPc liber difuzează în celulă și activează adenilat kinaza (A-kinaza), eliberând subunitatea sa catalitică C, care catalizează fosforilarea proteinelor intracelulare, adică. formează efectul final al unui stimul extracelular. Diagrama prezintă și medicamentele farmacologice și toxine care declanșează (+) sau inhibă (-) anumite reacții (modificate)
stimularea receptorilor extracelulari R s la structurile interne ale celulei.
În paralel cu lanțul stimulator de reacții care implică R s legarea mediatorilor inhibitori și a hormonilor de receptorul corespunzător este posibilă R i care din nou prin proteina activată de GTP G , inhibă AC și astfel producția de cAMP. Difuzând în celulă, cAMP reacţionează cu adenilat kinaza (A-kinaza); aceasta eliberează subunitatea C, care catalizează fosforilarea proteinei P. Această fosforilare transformă proteinele într-o formă activă, iar ele își pot exercita acum efectele lor specifice de reglare (de exemplu, provoacă degradarea glicogenului). Acest sistem de reglare complex este extrem de eficient deoarece rezultatul final este fosforilarea multor proteine, adică semnalul de reglare călătorește prin circuit cu un câștig mare. Mediatori externi care se leagă de receptori Rs și Ri specifice fiecăruia dintre ele, extrem de diverse. Adrenalina, conectarea cu R s sau Ri participă la reglarea metabolismului lipidelor și glicogenului, precum și la îmbunătățirea contracției mușchiului inimii și la alte reacții (vezi capitolul 19). Hormon de stimulare a tiroidei, activând R s , stimulează eliberarea hormonului tiroxină de către glanda tiroidă, iar prostaglandina I inhibă agregarea trombocitelor din sânge. Efecte inhibitoare, inclusiv adrenalina, mediate prin R i sunt exprimate în încetinirea lipolizei. Prin urmare, Sistemul cAMP este un sistem de reglare intracelular multifuncțional, care poate fi controlat cu precizie de substanțe de semnalizare extracelulare stimulatoare și inhibitorii.
Inozitol fosfat „IF” h " Sistemul intracelular al celui de-al doilea mesager, fosfatul de inozitol, a fost descoperit abia recent (Fig. 1.16). În acest caz, nu există o cale inhibitorie, dar există o similitudine cu sistemul cAMP, în care efectul de stimulare a receptorului R este transferat proteinei G activate de GTP pe suprafața interioară a membranei. În etapa următoare, fosfatidilinozitol (PI) lipidic membranar obișnuit, după ce a primit anterior două grupe fosfat suplimentare, este transformat în PI-difosfat (PIP 2), care este scindat de fosfodiesteraza activată (PDE) în inozitol trifosfat(IFz) și lipide diacilglicerol(DAG). Inozitol trifosfat este un al doilea mesager solubil în apă care difuzează în citosol. Acţionează în primul rând prin eliberarea de Ca 2+ din reticulul endoplasmatic. Ca 2+ la rândul său acționează ca un mesager, așa cum este descris mai sus; de exemplu, activează fosfokinaza dependentă de Ca 2+, care fosforilează enzimele. Subunitatea lipidică a DAG (Fig. 1.16) poartă, de asemenea, semnalul, difuzând în faza lipidică a membranei plasmatice la C-kinaza situată pe suprafața sa interioară, care este activată cu participarea fosfatidilserinei ca cofactor. Apoi C-kinaza declanșează fosforilarea proteinelor, transformându-le într-o formă activă.
Sistemul de mesager secundar intracelular IF3 poate fi, de asemenea, controlat de o varietate de mediatori externi și hormoni, incluzând acetilcolina, serotonina, vasopresina și hormonul de stimulare a tiroidei; ca și sistemul cAMP, este caracterizat printr-o varietate de efecte intracelulare. Este posibil ca acest sistem să fie activat și de lumină în receptorul vizual al ochiului și să joace un rol central în fototransducție (vezi capitolul 11). Pentru prima dată în dezvoltarea individuală a organismului, receptorul sistemului IG este activat de spermatozoizi, drept urmare IG participă la reacțiile de reglare care însoțesc fertilizarea ovulului.
Sistemele cAMP și IFz-DAG sunt foarte eficiente amplificatori biologici. ei

Orez. 1.16.Un lanț de reacții care implică mediatorul intracelular IPG (inozitol trifosfat). Ca și în sistemul cAMP, semnalul extracelular este mediat printr-o proteinăG, care în acest caz activează fosfodiesteraza (PDE). Această enzimă descompune fosfatidilinozină difosfat (PIP 2 ) în membrana plasmatică înainte de IF h şi diacilglicerol (DAG); DACĂ h difuzează în citoplasmă. Aici provoacă eliberarea de Ca 2+ din reticulul endoplasmatic; creșterea concentrației de Ca 2+ în citoplasmă ([Ca 2+ ] i ) activează protein kinaza, care fosforilează și, prin urmare, activează enzimele. Un alt produs, DAG, rămâne în membrană și activează protein kinaza C (cofactorul fosfatidilserinei, PS). Protein kinaza C fosforilează, de asemenea, enzimele care mediază acțiuni specifice asociate cu stimularea receptorilor extrinseci R . Ramuri ale lanțului de reacții care implică IF h și DAG pot fi activate independent de ionomicină și, respectiv, esterul de forbol (modificat)
transformă reacția dintre un transmițător și un receptor membranar extern în fosforilarea unei varietăți de proteine intracelulare, care pot influența apoi diferite funcții celulare. Unul dintre aspectele semnificative ale problemei este că, din câte se știe astăzi, există doar aceste două sisteme de reglare strâns legate de acest tip, utilizate de numeroși mediatori externi pentru a regla o varietate de procese intracelulare. În același timp, aceste sisteme de reglare, inclusiv Ca 2+, interacționează strâns între ele, ceea ce le permite să efectueze o reglare fină a funcțiilor celulare.
Tutoriale și ghiduri
1. Alberts ÎN., Bray D., Lewis J., Raff M., Roberts LA., Watson J.D.Molecular Biology of the Cell, New York și Londra, Garland Publishing Inc., 1983.
2. Czihak G., Longer H., Ziegler H.(eds.). Biologie. Berlin, Heidelberg, New York, Springer, 1983.
3. Hille ÎN. Canalele ionice ale membranelor excitabile. Sunderland, Mass., Sinauer Assoc., 1984.
4. Hoppe W., Lohmann W.. Marki H., Ziegler H.(eds.). Biofizică. Berlin, Heidelberg, New York, Springer, 1984.
5. Jungermann LA., Mahler H.Biochimie. Berlin, Heidelberg, New York, Springer, 1980.
6. Kandel E. R., Schwartz– J. H.,(eds.). Principles of Neural Science, New York, Amsterdam, Oxford, Elsevier, 1985.
7. Schiebler T. H., Schmidt W.Anatomic des Menschen. Berlin, Heidelberg, New York, Tokyo, Springer, 1983.
Articole și recenzii originale
8. Berridge M.J. Baza moleculară a comunicării în interiorul celulei, Sci. Amer, 253, 124-134 (1985).
9. Berridge M.J., Irvine R.F. Inozitol trifosfat, un nou mesager secundar în transducția semnalului celular. Nature, 312, 315 321 (1984).
10. Bretscher M.S. Moleculele membranei celulare, Sci. Amer., 253, 124–134 (1985).
11. Daut J. Celula vie ca mașină de transducere a energiei. Un model minim de metabolism miocardic, Biochem. et Biophys. Acta, 895, 41–62 (1987).
12. Hodgkin A.L., Katz ÎN. Efectul ionilor de sodiu asupra activității electrice a axonului gigant al calmarului. J. Physiol. (Land.), 108, 37–77 (1949).
13. Hodgkin A.L., Keynes R.D. Transportul activ al cationilor în axonii giganți din Sepiași Loligo, J. Physiol. (Land.), 128, 28–42 (1955).
14. Mai lung P. Canale ionice cu substari conformaționale, Biophys. J., 47, 581–590 (1985).
15. Ochs S., Worth P.M. Transportul exoplasmatic în sistemele normale și patologice. În: Fiziologia și patologia axonilor, S.G. Waxmann, Ed. New York, Raven Press, 1978.
rezumatul altor prezentări„Metode de predare a biologiei” - Zoologie școlară. Introducerea elevilor în utilizarea datelor zoologice științifice. Educatie morala. Binecuvântare suplimentară a cotetului de găini. Selectarea metodelor. Procesele vieții. Pești de acvariu. Nutriție. Educația pentru mediu. Materialitatea proceselor vieții. Rezultate negative. Atenție studenți. Formular obligatoriu. Privind animalele mici. Scopurile și obiectivele biologiei. Poveste.
„Învățare bazată pe probleme în lecțiile de biologie” - Cunoștințe. Manuale noi. Calea către soluție. Problemă. Seminarii. Ce este o sarcină? Albrecht Durer. Învățare bazată pe probleme în lecțiile de biologie. Lecții non-standard. Ce se înțelege prin învățarea bazată pe probleme. Calitatea vieții. Biologia ca disciplină academică. Întrebare. Lecție de rezolvare a problemelor. Scăderea interesului pentru subiect. Clase de laborator bazate pe probleme.
„Gândirea critică în lecțiile de biologie” - Tehnologia „gândirii critice”. Utilizarea tehnologiei de „dezvoltare a gândirii critice”. Tabel pentru lecție. Motivația de a studia. Ecosisteme. Sensul „dezvoltare a gândirii critice”. Semne ale tehnologiei. Tehnologia RKM. Structura lecției. Direcții principale. Istoria tehnologiei. Tehnologii pedagogice. Regulile tehnologiei. Sarcini de biologie. Fotosinteză. Tehnici utilizate în diferite etape ale lecției.
„Lecții de biologie cu o tablă interactivă” - Manuale electronice. Beneficii pentru elevi. O tablă interactivă ajută la transmiterea informațiilor fiecărui elev. Sarcini didactice. Rezolvarea problemelor biologice. Avantajele lucrului cu table interactive. Lucrul cu prezentări. Lucrați la compararea obiectelor. Obiecte în mișcare. Utilizarea foilor de calcul. Utilizarea unei table interactive în procesul de predare a școlarilor. Beneficii pentru profesori.
„Abordare sistem-activitate în biologie” - Întrebări seminar. Metoda de activitate. Dryopithecus. Cale extraterestră de origine umană. Lizozomi. Organizarea chimică. Gimnosperme. Metabolism. Analizoare. Abordarea sistem-activitate a predării biologiei. Cromozomii. Citoplasma. Orbire. Lungimea urechii. Clasificarea unei persoane. Scheletul unui mamifer. Căile evoluției umane. Mitoză. Complex superficial. Întrebare problematică. Nucleol. Plic nuclear.
„Computer în Biologie” - Activitate de colaborare a studenților. Familiile de angiosperme. Antrenament interactiv. Modele de învățare. Un exemplu de sistem de evaluare. Întrebări pe cardul de instrucțiuni. Exemplu de card de instrucțiuni. Cercetători. Microgrupuri. Tehnologii de învățare interactivă. Carusel. Tehnologii de învățare interactivă. Abordări interactive în lecțiile de biologie. Forma de lucru în grup. Sarcini pentru grupuri de „cercetători”.
