5. Regulierungsfunktion. Proteine übernehmen die Funktion von Signalstoffen – einige Hormone, Histohormone und Neurotransmitter – sind Rezeptoren für Signalstoffe jeglicher Struktur und sorgen für die weitere Signalübertragung in den biochemischen Signalketten der Zelle. Beispiele hierfür sind das Wachstumshormon Somatotropin, das Hormon Insulin sowie H- und M-cholinerge Rezeptoren.
6. Motorische Funktion. Mit Hilfe von Proteinen werden Kontraktionsprozesse und andere biologische Bewegungen durchgeführt. Beispiele hierfür sind Tubulin, Aktin und Myosin.
7. Ersatzfunktion. Pflanzen enthalten Reserveproteine, die wertvolle Nährstoffe sind; Muskelproteine dienen als Reservenährstoffe, die bei Bedarf mobilisiert werden.
Proteine zeichnen sich durch das Vorhandensein mehrerer struktureller Organisationsebenen aus.
Primärstruktur Ein Protein ist eine Abfolge von Aminosäureresten in einer Polypeptidkette. Eine Peptidbindung ist eine Carbonsäureamidbindung zwischen der α-Carboxylgruppe einer Aminosäure und der α-Aminogruppe einer anderen Aminosäure.
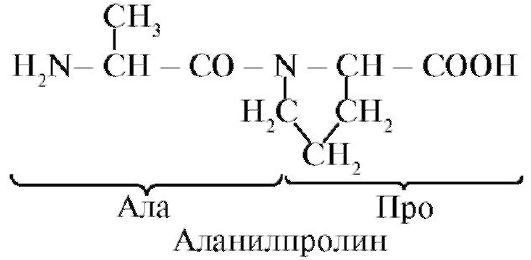
Alanylphenylalanylcysteylprolin
Hoch Peptidbindung Es gibt mehrere Funktionen:
a) es ist resonant stabilisiert und liegt daher praktisch in derselben Ebene – planar; die Rotation um die C-N-Bindung erfordert viel Energie und ist schwierig;
b) Die -CO-NH-Bindung hat einen besonderen Charakter, sie ist kleiner als eine reguläre, aber größer als eine doppelte, d. h. es liegt eine Keto-Enol-Tautomerie vor:

c) Substituenten in Bezug auf die Peptidbindung sind in Trance-Position;
d) Das Peptidrückgrat ist von Seitenketten verschiedener Art umgeben, die mit den umgebenden Lösungsmittelmolekülen interagieren, freie Carboxyl- und Aminogruppen ionisiert werden und kationische und anionische Zentren des Proteinmoleküls bilden. Abhängig von ihrem Verhältnis erhält das Proteinmolekül insgesamt eine positive oder negative Ladung und wird außerdem durch den einen oder anderen pH-Wert der Umgebung gekennzeichnet, wenn es den isoelektrischen Punkt des Proteins erreicht. Radikale bilden Salz-, Ether- und Disulfidbrücken innerhalb des Proteinmoleküls und bestimmen auch die Bandbreite der für Proteine charakteristischen Reaktionen.
Derzeit einigten sich darauf, Polymere, die aus 100 oder mehr Aminosäureresten bestehen, als Proteine, Polypeptide – Polymere, die aus 50–100 Aminosäureresten bestehen, und Peptide mit niedrigem Molekulargewicht – Polymere, die aus weniger als 50 Aminosäureresten bestehen – zu betrachten.
Manche niedriges molekulares Gewicht Peptide spielen eine eigenständige biologische Rolle. Beispiele für einige dieser Peptide:
Glutathion – γ-glu-cis-gly – eins eines der am weitesten verbreiteten intrazellulären Peptide, beteiligt sich an Redoxprozessen in Zellen und der Übertragung von Aminosäuren durch biologische Membranen.
Carnosin – β-ala-his – Peptid, kommt in den Muskeln von Tieren vor, eliminiert die Produkte des Lipidperoxidabbaus, beschleunigt den Prozess des Kohlenhydratabbaus in den Muskeln und ist in Form von Phosphat am Energiestoffwechsel in den Muskeln beteiligt.
Vasopressin ist ein Hormon des Hypophysenhinterlappens, das an der Regulierung des Wasserstoffwechsels im Körper beteiligt ist:

Phalloidin- giftiges Fliegenpilz-Polypeptid, das in vernachlässigbaren Konzentrationen den Tod des Körpers durch die Freisetzung von Enzymen und Kaliumionen aus Zellen verursacht:

Gramicidin - Antibiotikum wirkt auf viele grampositive Bakterien, verändert die Durchlässigkeit biologischer Membranen für Verbindungen mit niedrigem Molekulargewicht und führt zum Zelltod:

Meth-Enkephalin – Tyr-Gly-Gly-Phen-Met – ein Peptid, das in Neuronen synthetisiert wird und Schmerzen lindert.
Sekundärstruktur von Proteinen ist eine räumliche Struktur, die durch Wechselwirkungen zwischen funktionellen Gruppen des Peptidrückgrats entsteht.
Die Peptidkette enthält viele CO- und NH-Gruppen von Peptidbindungen, von denen jede potenziell in der Lage ist, an der Bildung von Wasserstoffbrückenbindungen teilzunehmen. Es gibt zwei Haupttypen von Strukturen, die dies ermöglichen: eine α-Helix, bei der die Kette wie ein Telefonkabel aufgerollt ist, und eine gefaltete β-Struktur, bei der längliche Abschnitte einer oder mehrerer Ketten nebeneinander gelegt sind Seite. Beide Strukturen sind sehr stabil.
Die α-Helix ist gekennzeichnet durch extrem dichte Packung einer verdrillten Polypeptidkette; auf jede Windung einer rechtsdrehenden Helix kommen 3,6 Aminosäurereste, deren Reste immer nach außen und leicht nach hinten gerichtet sind, also zum Anfang der Polypeptidkette.
Hauptmerkmale der α-Helix:
1) Die α-Helix wird durch Wasserstoffbrückenbindungen zwischen dem Wasserstoffatom am Stickstoff der Peptidgruppe und dem Carbonylsauerstoff des Rests an vier Positionen entlang der Kette stabilisiert;
2) alle Peptidgruppen sind an der Bildung einer Wasserstoffbindung beteiligt, dies gewährleistet maximale Stabilität der α-Helix;
3) alle Stickstoff- und Sauerstoffatome der Peptidgruppen sind an der Bildung von Wasserstoffbrückenbindungen beteiligt, was die Hydrophilie der α-helikalen Regionen deutlich verringert und ihre Hydrophobie erhöht;
4) Die α-Helix wird spontan gebildet und ist die stabilste Konformation der Polypeptidkette, entsprechend der minimalen freien Energie;
5) In einer Polypeptidkette aus L-Aminosäuren ist die rechtsgängige Helix, die normalerweise in Proteinen vorkommt, viel stabiler als die linksgängige.
Möglichkeit der Bildung einer α-Helix wird durch die Primärstruktur des Proteins bestimmt. Einige Aminosäuren verhindern die Verdrehung des Peptidrückgrats. Beispielsweise stoßen sich die benachbarten Carboxylgruppen von Glutamat und Aspartat gegenseitig ab, was die Bildung von Wasserstoffbrückenbindungen in der α-Helix verhindert. Aus dem gleichen Grund ist die Kettenhelikalisierung an Stellen schwierig, an denen positiv geladene Lysin- und Argininreste nahe beieinander liegen. Allerdings spielt Prolin die größte Rolle bei der Zerstörung der α-Helix. Erstens ist das Stickstoffatom in Prolin Teil eines starren Rings, der eine Rotation um die N-C-Bindung verhindert, und zweitens bildet Prolin aufgrund des Fehlens von Wasserstoff am Stickstoffatom keine Wasserstoffbindung.
β-Faltblattbildung ist eine Schichtstruktur, gebildet durch Wasserstoffbrückenbindungen zwischen linear angeordneten Peptidfragmenten. Beide Ketten können unabhängig sein oder zum selben Polypeptidmolekül gehören. Sind die Ketten in die gleiche Richtung ausgerichtet, so nennt man eine solche β-Struktur parallel. Bei entgegengesetzten Kettenrichtungen, also wenn der N-Terminus einer Kette mit dem C-Terminus einer anderen Kette zusammenfällt, spricht man von einer β-Struktur antiparallel. Antiparallele β-Faltblätter mit nahezu linearen Wasserstoffbrücken sind energetisch bevorzugter.


paralleles β-Faltblatt, antiparalleles β-Faltblatt
Im Gegensatz zur α-Helix Da jeder Abschnitt der β-Faltblattkette mit Wasserstoffbrückenbindungen gesättigt ist, ist er offen für die Bildung zusätzlicher Wasserstoffbrückenbindungen. Die Seitenreste der Aminosäuren sind nahezu senkrecht zur Blattebene abwechselnd nach oben und unten ausgerichtet.
In den Bereichen, in denen die Peptidkette biegt ziemlich stark ab und enthält oft eine β-Schleife. Hierbei handelt es sich um ein kurzes Fragment, in dem 4 Aminosäurereste um 180° gebogen sind und durch eine Wasserstoffbrücke zwischen dem ersten und vierten Rest stabilisiert werden. Große Aminosäurereste stören die Bildung der β-Schleife, daher umfasst sie am häufigsten die kleinste Aminosäure, Glycin.
Suprasekundäre Proteinstruktur- Dies ist eine bestimmte Reihenfolge des Wechsels von Sekundärstrukturen. Unter einer Domäne versteht man einen separaten Teil eines Proteinmoleküls, der über ein gewisses Maß an struktureller und funktioneller Autonomie verfügt. Domänen gelten heute als grundlegende Elemente der Struktur von Proteinmolekülen, und die Beziehung und Art der Anordnung von α-Helices und β-Faltblättern trägt eher zum Verständnis der Evolution von Proteinmolekülen und phylogenetischen Beziehungen bei als ein Vergleich von Primärstrukturen.
Die Hauptaufgabe der Evolution ist Entwicklung immer neuer Proteine. Die Chance, versehentlich eine Aminosäuresequenz zu synthetisieren, die die Verpackungsbedingungen erfüllt und die Erfüllung funktioneller Aufgaben gewährleistet, ist unendlich gering. Daher ist es üblich, Proteine mit unterschiedlichen Funktionen zu finden, die jedoch in ihrer Struktur so ähnlich sind, dass es den Anschein hat, als hätten sie einen gemeinsamen Vorfahren oder hätten sich voneinander entwickelt. Es scheint, dass die Evolution, wenn sie vor der Notwendigkeit steht, ein bestimmtes Problem zu lösen, es vorzieht, Proteine nicht von Anfang an für diesen Zweck zu entwerfen, sondern bereits etablierte Strukturen für diesen Zweck anzupassen und sie für neue Zwecke anzupassen.
Einige Beispiele für häufig wiederholte suprasekundäre Strukturen:
1) αα’ – Proteine, die nur α-Helices enthalten (Myoglobin, Hämoglobin);
2) ββ’ – Proteine, die nur β-Strukturen enthalten (Immunglobuline, Superoxiddismutase);
3) βαβ’ – β-Fass-Struktur, jede β-Schicht befindet sich im Inneren des Fasses und ist mit einer α-Helix verbunden, die sich auf der Oberfläche des Moleküls befindet (Triose-Phosphoisomerase, Lactat-Dehydrogenase);
4) „Zinkfinger“ – ein Proteinfragment bestehend aus 20 Aminosäureresten, das Zinkatom ist mit zwei Cysteinresten und zwei Histidinresten verknüpft, wodurch ein „Finger“ aus etwa 12 Aminosäureresten entsteht kann an die regulatorischen Regionen des DNA-Moleküls binden;
5) „Leucin-Reißverschluss“ – interagierende Proteine haben eine α-helikale Region, die mindestens 4 Leucinreste enthält, sie sind 6 Aminosäuren voneinander entfernt, das heißt, sie befinden sich auf der Oberfläche jeder zweiten Windung und können hydrophobe Bindungen mit Leucinresten bilden ein weiteres Protein. Mit Hilfe von Leucin-Reißverschlüssen können beispielsweise Moleküle stark basischer Histonproteine komplexiert und so eine positive Ladung überwunden werden.
Protein-Tertiärstruktur- Dies ist die räumliche Anordnung des Proteinmoleküls, stabilisiert durch die Bindungen zwischen den Nebenresten von Aminosäuren.
Arten von Bindungen, die die Tertiärstruktur eines Proteins stabilisieren:

elektrostatischer Wasserstoff, hydrophobe Disulfid-Wechselwirkung, Bindungen, Wechselwirkungen, Bindungen
Je nach Faltung Die Tertiärstruktur von Proteinen kann in zwei Haupttypen eingeteilt werden – fibrillär und globulär.
Fibrilläre Proteine- lange, fadenförmige, wasserunlösliche Moleküle, deren Polypeptidketten entlang einer Achse verlängert sind. Dabei handelt es sich vor allem um Struktur- und Kontraktilproteine. Einige Beispiele der häufigsten fibrillären Proteine:
1. α- Keratine. Von Epidermiszellen synthetisiert. Sie machen fast das gesamte Trockengewicht von Haaren, Fell, Federn, Hörnern, Nägeln, Krallen, Federkielen, Schuppen, Hufen und Schildkrötenpanzern sowie einen erheblichen Teil des Gewichts der äußeren Hautschicht aus. Dabei handelt es sich um eine ganze Familie von Proteinen; sie haben eine ähnliche Aminosäurezusammensetzung, enthalten viele Cysteinreste und weisen die gleiche räumliche Anordnung der Polypeptidketten auf.
In Haarzellen befinden sich Polypeptidketten von Keratin zunächst in Fasern organisiert, aus denen dann Strukturen wie ein Seil oder ein gedrehtes Kabel geformt werden, die schließlich den gesamten Raum der Zelle ausfüllen. Die Haarzellen werden abgeflacht und sterben schließlich ab, und die Zellwände bilden um jedes Haar eine röhrenförmige Hülle, die sogenannte Kutikula. Im α-Keratin haben die Polypeptidketten die Form einer α-Helix, die unter Bildung von Kreuzdisulfidbindungen zu einem dreiadrigen Kabel umeinander verdreht ist.
Die N-terminalen Reste sind lokalisiert auf einer Seite (parallel). Keratine sind in Wasser unlöslich, da in ihrer Zusammensetzung Aminosäuren mit unpolaren Seitenradikalen, die der wässrigen Phase zugewandt sind, überwiegen. Während der Dauerwelle laufen folgende Prozesse ab: Zunächst werden Disulfidbrücken durch Reduktion mit Thiolen zerstört, und wenn das Haar dann die gewünschte Form erhält, wird es durch Erhitzen getrocknet, während durch Oxidation mit Luftsauerstoff neue Disulfidbrücken gebildet werden , die die Form der Frisur beibehalten.
2. β-Keratine. Dazu gehören Seiden- und Spinnennetzfibroin. Es handelt sich um antiparallele β-gefaltete Schichten mit einem überwiegenden Anteil an Glycin, Alanin und Serin in der Zusammensetzung.
3. Kollagen. Das häufigste Protein bei höheren Tieren und das wichtigste fibrilläre Protein des Bindegewebes. Kollagen wird in Fibroblasten und Chondrozyten – spezialisierten Bindegewebszellen – synthetisiert und dann ausgestoßen. Kollagenfasern kommen in Haut, Sehnen, Knorpel und Knochen vor. Sie dehnen sich nicht, sind stärker als Stahldraht und Kollagenfibrillen zeichnen sich durch Querstreifen aus.
Beim Kochen in Wasser faserig, unlösliches und unverdauliches Kollagen wird durch Hydrolyse einiger kovalenter Bindungen in Gelatine umgewandelt. Kollagen enthält 35 % Glycin, 11 % Alanin, 21 % Prolin und 4-Hydroxyprolin (eine Aminosäure, die nur in Kollagen und Elastin vorkommt). Diese Zusammensetzung bestimmt den relativ geringen Nährwert von Gelatine als Nahrungsprotein. Kollagenfibrillen bestehen aus sich wiederholenden Polypeptiduntereinheiten, die Tropokollagen genannt werden. Diese Untereinheiten sind entlang der Fibrille in Form paralleler Bündel Kopf-an-Schwanz angeordnet. Durch die Verschiebung der Köpfe entstehen die charakteristischen Querstreifen. Die Hohlräume in dieser Struktur können bei Bedarf als Ort für die Ablagerung von Kristallen aus Hydroxylapatit Ca 5 (OH)(PO 4) 3 dienen, das eine wichtige Rolle bei der Knochenmineralisierung spielt.


Tropokollagen-Untereinheiten bestehen aus aus drei Polypeptidketten, die eng zu einem dreisträngigen Seil zusammengerollt sind und sich von α- und β-Keratinen unterscheiden. Bei manchen Kollagenen haben alle drei Ketten die gleiche Aminosäuresequenz, bei anderen sind nur zwei Ketten identisch und die dritte ist unterschiedlich. Die Polypeptidkette von Tropokollagen bildet aufgrund der durch Prolin und Hydroxyprolin verursachten Kettenbiegungen eine linksgängige Helix mit nur drei Aminosäureresten pro Windung. Die drei Ketten sind zusätzlich zu Wasserstoffbrücken durch eine kovalente Bindung miteinander verbunden, die zwischen zwei Lysinresten in benachbarten Ketten gebildet wird:

Wenn wir älter werden In und zwischen den Tropokollagen-Untereinheiten bilden sich immer mehr Vernetzungen, wodurch die Kollagenfibrillen steifer und spröder werden. Dadurch verändern sich die mechanischen Eigenschaften von Knorpel und Sehnen, die Knochen werden brüchiger und die Transparenz der Hornhaut verringert.
4. Elastin. Enthalten im gelben elastischen Gewebe der Bänder und der elastischen Bindegewebsschicht in den Wänden großer Arterien. Die Hauptuntereinheit der Elastinfibrillen ist Tropoelastin. Elastin ist reich an Glycin und Alanin, enthält viel Lysin und wenig Prolin. Die spiralförmigen Abschnitte aus Elastin dehnen sich bei Zugeinwirkung, kehren jedoch bei Entlastung wieder in ihre ursprüngliche Länge zurück. Die Lysinreste von vier verschiedenen Ketten bilden kovalente Bindungen miteinander und ermöglichen es Elastin, sich reversibel in alle Richtungen zu dehnen.
Kugelförmige Proteine- Proteine, deren Polypeptidkette zu einer kompakten Kügelchen gefaltet ist, können vielfältige Funktionen erfüllen.
Tertiärstruktur globulärer Proteine Am bequemsten ist es, dies am Beispiel von Myoglobin zu betrachten. Myoglobin ist ein relativ kleines sauerstoffbindendes Protein, das in Muskelzellen vorkommt. Es speichert gebundenen Sauerstoff und fördert dessen Übertragung in die Mitochondrien. Das Myoglobinmolekül enthält eine Polypeptidkette und eine Hämogruppe (Häm) – einen Komplex aus Protoporphyrin und Eisen.
Grundeigenschaften Myoglobin:
a) das Myoglobinmolekül ist so kompakt, dass nur 4 Wassermoleküle hineinpassen;
b) alle polaren Aminosäurereste, mit Ausnahme von zwei, befinden sich auf der äußeren Oberfläche des Moleküls und befinden sich alle in einem hydratisierten Zustand;
c) die meisten hydrophoben Aminosäurereste befinden sich innerhalb des Myoglobinmoleküls und sind somit vor dem Kontakt mit Wasser geschützt;
d) jeder der vier Prolinreste im Myoglobinmolekül befindet sich an der Biegestelle der Polypeptidkette, Serin- und Asparaginreste befinden sich an anderen Biegestellen, da solche Aminosäuren die Bildung einer α-Helix verhindern, wenn sie vorhanden sind liegen nebeneinander;
e) eine flache Hämgruppe liegt in einem Hohlraum (Tasche) nahe der Oberfläche des Moleküls, das Eisenatom verfügt über zwei senkrecht zur Hämebene gerichtete Koordinationsbindungen, eine davon ist mit dem Histidinrest 93 verbunden und die andere dient der Bindung ein Sauerstoffmolekül.
Beginnend mit der Protein-Tertiärstruktur wird in der Lage, seine inhärenten biologischen Funktionen zu erfüllen. Die Funktionsweise von Proteinen beruht darauf, dass beim Anlegen einer Tertiärstruktur auf der Oberfläche des Proteins Bereiche entstehen, die andere Moleküle, sogenannte Liganden, anlagern können. Die hohe Spezifität der Wechselwirkung des Proteins mit dem Liganden wird durch die Komplementarität der Struktur des aktiven Zentrums zur Struktur des Liganden gewährleistet. Komplementarität ist die räumliche und chemische Entsprechung interagierender Oberflächen. Bei den meisten Proteinen stellt die Tertiärstruktur den maximalen Faltungsgrad dar.
Quartäre Proteinstruktur- charakteristisch für Proteine, die aus zwei oder mehr Polypeptidketten bestehen, die ausschließlich durch nichtkovalente Bindungen, hauptsächlich elektrostatische und Wasserstoffbindungen, miteinander verbunden sind. Am häufigsten enthalten Proteine zwei oder vier Untereinheiten; mehr als vier Untereinheiten enthalten normalerweise regulatorische Proteine.
Proteine mit Quartärstruktur, werden oft als oligomer bezeichnet. Es gibt homomere und heteromere Proteine. Zu den homomeren Proteinen zählen Proteine, bei denen alle Untereinheiten die gleiche Struktur haben, beispielsweise besteht das Enzym Katalase aus vier absolut identischen Untereinheiten. Heteromere Proteine haben unterschiedliche Untereinheiten; das Enzym RNA-Polymerase besteht beispielsweise aus fünf strukturell unterschiedlichen Untereinheiten, die unterschiedliche Funktionen erfüllen.
Interaktion einzelner Untereinheiten mit einem bestimmten Liganden führt zu Konformationsänderungen im gesamten oligomeren Protein und verändert die Affinität anderer Untereinheiten für Liganden. Diese Eigenschaft liegt der Fähigkeit oligomerer Proteine zur allosterischen Regulierung zugrunde.
Die Quartärstruktur eines Proteins kann untersucht werden am Beispiel des Hämoglobins. Enthält vier Polypeptidketten und vier Häm-Prothesengruppen, in denen die Eisenatome in der Eisenform Fe 2+ vorliegen. Der Proteinteil des Moleküls – Globin – besteht aus zwei α-Ketten und zwei β-Ketten, die bis zu 70 % α-Helices enthalten. Jede der vier Ketten hat eine charakteristische Tertiärstruktur, und jeder Kette ist eine Hämogruppe zugeordnet. Die Häme verschiedener Ketten liegen relativ weit voneinander entfernt und haben unterschiedliche Neigungswinkel. Zwischen zwei α-Ketten und zwei β-Ketten werden nur wenige direkte Kontakte gebildet, während zwischen den α- und β-Ketten zahlreiche durch hydrophobe Radikale gebildete Kontakte vom Typ α 1 β 1 und α 2 β 2 entstehen. Zwischen α 1 β 1 und α 2 β 2 verbleibt ein Kanal.
Im Gegensatz zu Myoglobin Hämoglobin gekennzeichnet deutlich geringere Affinität zu Sauerstoff, was es ihm ermöglicht, bei den in den Geweben herrschenden niedrigen Sauerstoffpartialdrücken einen erheblichen Teil des gebundenen Sauerstoffs an diese abzugeben. Sauerstoff wird bei höheren pH-Werten und niedrigen CO 2 -Konzentrationen, die für die Alveolen der Lunge charakteristisch sind, leichter durch Hämoglobineisen gebunden; Die Freisetzung von Sauerstoff aus Hämoglobin wird durch für Gewebe charakteristische niedrigere pH-Werte und hohe CO 2 -Konzentrationen begünstigt.
Hämoglobin trägt neben Sauerstoff auch Wasserstoffionen, die an Histidinreste in den Ketten binden. Hämoglobin trägt auch Kohlendioxid, das sich an die terminale Aminogruppe jeder der vier Polypeptidketten bindet, was zur Bildung von Carbaminohämoglobin führt:

IN rote Blutkörperchen in relativ hohen Konzentrationen der Stoff 2,3-Diphosphoglycerat (DPG) ist vorhanden, dessen Gehalt mit dem Aufstieg in große Höhen und bei Hypoxie zunimmt und die Freisetzung von Sauerstoff aus dem Hämoglobin im Gewebe erleichtert. DPG befindet sich im Kanal zwischen α 1 β 1 und α 2 β 2 und interagiert mit positiv kontaminierten Gruppen von β-Ketten. Wenn Hämoglobin Sauerstoff bindet, wird DPG aus der Höhle gedrückt. Die roten Blutkörperchen einiger Vögel enthalten kein DPG, sondern Inositolhexaphosphat, was die Affinität von Hämoglobin zu Sauerstoff weiter verringert.
 2,3-Diphosphoglycerat (DPG)
2,3-Diphosphoglycerat (DPG)
HbA – normales Hämoglobin für Erwachsene, HbF – fetales Hämoglobin, hat eine größere Affinität zu O 2, HbS – Hämoglobin bei Sichelzellenanämie. Sichelzellenanämie ist eine schwere Erbkrankheit, die durch eine genetische Anomalie des Hämoglobins verursacht wird. Im Blut erkrankter Menschen befinden sich ungewöhnlich viele dünne, sichelförmige rote Blutkörperchen, die erstens leicht platzen und zweitens die Blutkapillaren verstopfen.
Auf molekularer Ebene ist Hämoglobin S anders Von Hämoglobin A gibt es einen Aminosäurerest an Position 6 der β-Ketten, wo anstelle eines Glutaminsäurerestes Valin vorhanden ist. Somit enthält Hämoglobin S zwei weniger negative Ladungen; das Auftreten von Valin führt zum Auftreten eines „klebrigen“ hydrophoben Kontakts auf der Oberfläche des Moleküls, wodurch Desoxyhämoglobin S-Moleküle während der Desoxygenierung zusammenkleben und ungewöhnlich lange unlösliche Formen bilden fadenförmige Aggregate, die zu einer Verformung der roten Blutkörperchen führen.
Es gibt keinen Grund zu der Annahme, dass es eine unabhängige genetische Kontrolle über die Bildung von Ebenen der Proteinstrukturorganisation oberhalb der Primärstruktur gibt, da die Primärstruktur die Sekundär-, Tertiär- und Quartärstruktur (falls vorhanden) bestimmt. Die native Konformation eines Proteins ist unter bestimmten Bedingungen die thermodynamisch stabilste Struktur.
VORTRAG 6
Es gibt physikalische, chemische und biologische Eigenschaften von Proteinen.
Physikalische Eigenschaften von Proteinen sind das Vorhandensein von Molekulargewicht, Doppelbrechung (eine Änderung der optischen Eigenschaften einer Proteinlösung in Bewegung im Vergleich zu einer ruhenden Lösung) aufgrund der nichtkugelförmigen Form von Proteinen, Mobilität in einem elektrischen Feld aufgrund der Ladung von Proteinmolekülen . Darüber hinaus zeichnen sich Proteine durch optische Eigenschaften aus, die in der Fähigkeit bestehen, die Polarisationsebene des Lichts zu drehen, Lichtstrahlen aufgrund der großen Größe der Proteinpartikel zu streuen und ultraviolette Strahlen zu absorbieren.
Eine der charakteristischen physikalischen Eigenschaften Proteine haben die Fähigkeit, an der Oberfläche zu adsorbieren und manchmal Moleküle, niedermolekulare organische Verbindungen und Ionen im Inneren einzufangen.
Die chemischen Eigenschaften von Proteinen sind unterschiedlich außergewöhnliche Vielfalt, da Proteine durch alle Reaktionen von Aminosäureresten und durch die Reaktion der Hydrolyse von Peptidbindungen gekennzeichnet sind.
Besitzt eine beträchtliche Anzahl saurer und basischer Gruppen Proteine weisen amphotere Eigenschaften auf. Im Gegensatz zu freien Aminosäuren werden die Säure-Base-Eigenschaften von Proteinen nicht durch α-Amino- und α-Carboxygruppen bestimmt, die an der Bildung von Peptidbindungen beteiligt sind, sondern durch geladene Radikale von Aminosäureresten. Die Haupteigenschaften von Proteinen werden durch Arginin-, Lysin- und Histidinreste bestimmt. Die sauren Eigenschaften sind auf Asparagin- und Glutaminsäurereste zurückzuführen.
Es genügen Proteintitrationskurven schwer zu interpretieren, da jedes Protein zu viele titrierbare Gruppen hat, es elektrostatische Wechselwirkungen zwischen den ionisierten Gruppen des Proteins gibt und der pK-Wert jeder titrierbaren Gruppe durch benachbarte hydrophobe Reste und Wasserstoffbrückenbindungen beeinflusst wird. Die größte praktische Anwendung ist der isoelektrische Punkt eines Proteins – der pH-Wert, bei dem die Gesamtladung des Proteins Null ist. Am isoelektrischen Punkt ist das Protein maximal inert, bewegt sich nicht in einem elektrischen Feld und hat die dünnste Hydratationshülle.
Proteine weisen Puffereigenschaften auf, aber ihre Pufferkapazität ist unbedeutend. Die Ausnahme bilden Proteine, die eine große Anzahl von Histidinresten enthalten. Beispielsweise verfügt das in Erythrozyten enthaltene Hämoglobin aufgrund des sehr hohen Gehalts an Histidinresten über eine erhebliche Pufferkapazität bei einem pH-Wert von etwa 7, was für die Rolle, die Erythrozyten beim Transport von Sauerstoff und Kohlendioxid spielen, sehr wichtig ist das Blut.
Proteine zeichnen sich durch ihre Löslichkeit in Wasser aus, und aus physikalischer Sicht bilden sie echte molekulare Lösungen. Allerdings zeichnen sich Proteinlösungen durch einige kolloidale Eigenschaften aus: den Tendahl-Effekt (Lichtstreuungsphänomen), die Unfähigkeit, semipermeable Membranen zu passieren, hohe Viskosität und die Bildung von Gelen.
Die Proteinlöslichkeit ist stark abhängig von der Konzentration der Salze, also von der Ionenstärke der Lösung. In destilliertem Wasser sind Proteine meist schlecht löslich, ihre Löslichkeit nimmt jedoch mit zunehmender Ionenstärke zu. Gleichzeitig binden immer mehr hydratisierte anorganische Ionen an die Oberfläche des Proteins und dadurch nimmt der Grad seiner Aggregation ab. Bei hoher Ionenstärke entziehen Salzionen den Proteinmolekülen die Hydratationshülle, was zur Aggregation und Ausfällung von Proteinen führt (Aussalzungsphänomen). Ausgehend von Löslichkeitsunterschieden ist es möglich, ein Proteingemisch mithilfe von Kochsalzen zu trennen.
Zu den biologischen Eigenschaften von Proteinen Dazu zählt vor allem ihre katalytische Aktivität. Eine weitere wichtige biologische Eigenschaft von Proteinen ist ihre hormonelle Aktivität, also die Fähigkeit, ganze Reaktionsgruppen im Körper zu beeinflussen. Einige Proteine haben toxische Eigenschaften, pathogene Aktivität, Schutz- und Rezeptorfunktionen und sind für Zelladhäsionsphänomene verantwortlich.
Eine weitere einzigartige biologische Eigenschaft von Proteinen- Denaturierung. Proteine in ihrem natürlichen Zustand werden als nativ bezeichnet. Unter Denaturierung versteht man die Zerstörung der räumlichen Struktur von Proteinen unter Einwirkung von Denaturierungsmitteln. Die Primärstruktur von Proteinen wird bei der Denaturierung nicht beschädigt, ihre biologische Aktivität geht jedoch verloren, ebenso wie Löslichkeit, elektrophoretische Mobilität und einige andere Reaktionen. Bei der Denaturierung sind die Aminosäurereste, die das aktive Zentrum des Proteins bilden, räumlich voneinander entfernt, d. h. das spezifische Bindungszentrum des Proteins mit dem Liganden wird zerstört. Hydrophobe Radikale, die sich normalerweise im hydrophoben Kern globulärer Proteine befinden, landen bei der Denaturierung auf der Oberfläche des Moleküls und schaffen so Bedingungen für die Aggregation der ausfallenden Proteine.
Reagenzien und Bedingungen, die eine Proteindenaturierung verursachen:
Temperatur über 60 °C – Zerstörung schwacher Bindungen im Protein,
Säuren und Laugen – Veränderung der Ionisierung ionogener Gruppen, Aufbrechen von Ionen- und Wasserstoffbindungen,
Harnstoff – Zerstörung intramolekularer Wasserstoffbrückenbindungen durch Bildung von Wasserstoffbrückenbindungen mit Harnstoff,
Alkohol, Phenol, Chloramin – Zerstörung hydrophober und Wasserstoffbrückenbindungen,
Salze von Schwermetallen – die Bildung unlöslicher Salze von Proteinen mit Schwermetallionen.
Wenn Denaturierungsmittel entfernt werden, ist eine Renativierung möglich, da die Peptidkette dazu neigt, in Lösung die Konformation mit der niedrigsten freien Energie anzunehmen.
Unter zellulären Bedingungen können Proteine spontan denaturieren, wenn auch mit geringerer Geschwindigkeit als bei hohen Temperaturen. Eine spontane Renativierung von Proteinen in der Zelle ist schwierig, da aufgrund der hohen Konzentration eine hohe Wahrscheinlichkeit der Aggregation teilweise denaturierter Moleküle besteht.
Zellen enthalten Proteine- molekulare Chaperone, die die Fähigkeit haben, an teilweise denaturierte Proteine zu binden, die sich in einem instabilen Zustand befinden und zur Aggregation neigen, und ihre native Konformation wiederherzustellen. Ursprünglich wurden diese Proteine als Hitzeschockproteine entdeckt, da ihre Synthese zunahm, wenn die Zelle Stress ausgesetzt war, beispielsweise wenn die Temperatur stieg. Chaperone werden nach der Masse ihrer Untereinheiten klassifiziert: hsp-60, hsp-70 und hsp-90. Jede Klasse umfasst eine Familie verwandter Proteine.
Molekulare Chaperone ( hsp-70) eine hochkonservierte Klasse von Proteinen, die in allen Teilen der Zelle vorkommt: Zytoplasma, Zellkern, endoplasmatisches Retikulum, Mitochondrien. Am C-Terminus der einzelnen Polypeptidkette verfügt Hsp-70 über eine Region, bei der es sich um eine Furche handelt, die mit 7–9 Aminosäureresten langen Peptiden interagieren kann, die mit hydrophoben Radikalen angereichert sind. Solche Regionen kommen in globulären Proteinen etwa alle 16 Aminosäuren vor. Hsp-70 ist in der Lage, Proteine vor Temperaturinaktivierung zu schützen und die Konformation und Aktivität teilweise denaturierter Proteine wiederherzustellen.
Chaperone-60 (hsp-60) sind an der Bildung der Tertiärstruktur von Proteinen beteiligt. Hsp-60 fungiert als oligomere Proteine, die aus 14 Untereinheiten bestehen. Hsp-60 bildet zwei Ringe, jeder Ring besteht aus 7 miteinander verbundenen Untereinheiten.
Jede Untereinheit besteht aus drei Domänen:
Die apikale Domäne weist eine Reihe hydrophober Aminosäurereste auf, die zum Inneren des von den Untereinheiten gebildeten Hohlraums weisen.
Die äquatoriale Domäne besitzt ATPase-Aktivität und ist für die Freisetzung des Proteins aus dem Chaperonin-Komplex notwendig;
Die Zwischendomäne verbindet die apikale und äquatoriale Domäne.
Ein Protein, dessen Oberfläche Fragmente aufweist, angereichert mit hydrophoben Aminosäuren, gelangt in den Hohlraum des Chaperonin-Komplexes. In der spezifischen Umgebung dieses Hohlraums erfolgt unter Bedingungen der Isolation von anderen Molekülen des Zellzytosols die Auswahl möglicher Proteinkonformationen, bis eine energetisch günstigere Konformation gefunden wird. Die Chaperon-abhängige Bildung der nativen Konformation ist mit dem Aufwand einer erheblichen Energiemenge verbunden, deren Quelle ATP ist.
MODUL 1 STRUKTUR, EIGENSCHAFTEN UND FUNKTIONEN VON PROTEINEN
MODUL 1 STRUKTUR, EIGENSCHAFTEN UND FUNKTIONEN VON PROTEINEN
Modulstruktur | Themen |
Moduleinheit 1 | 1.1. Strukturelle Organisation von Proteinen. Stadien der Bildung der nativen Proteinkonformation 1.2. Grundlagen der Proteinfunktion. Medikamente als Liganden, die die Proteinfunktion beeinflussen 1.3. Denaturierung von Proteinen und die Möglichkeit ihrer spontanen Renativierung |
Moduleinheit 2 | 1.4. Merkmale der Struktur und Funktionsweise oligomerer Proteine am Beispiel von Hämoglobin 1.5. Aufrechterhaltung der nativen Proteinkonformation unter zellulären Bedingungen 1.6. Vielzahl von Proteinen. Proteinfamilien am Beispiel der Immunglobuline 1.7. Physikochemische Eigenschaften von Proteinen und Methoden zu ihrer Trennung |
Moduleinheit 1 STRUKTURELLE ORGANISATION MONOMERER PROTEINE UND DIE GRUNDLAGEN IHRER FUNKTION
Lernziele: In der Lage sein:
1. Nutzen Sie Kenntnisse über die Strukturmerkmale von Proteinen und die Abhängigkeit von Proteinfunktionen von ihrer Struktur, um die Mechanismen der Entwicklung erblicher und erworbener Proteinopathien zu verstehen.
2. Erklären Sie die Mechanismen der therapeutischen Wirkung einiger Medikamente als Liganden, die mit Proteinen interagieren und deren Aktivität verändern.
3. Nutzen Sie Kenntnisse über die Struktur und Konformationslabilität von Proteinen, um deren strukturelle und funktionelle Instabilität und Tendenz zur Denaturierung unter sich ändernden Bedingungen zu verstehen.
4. Erklären Sie die Verwendung von Denaturierungsmitteln als Mittel zur Sterilisierung medizinischer Materialien und Instrumente sowie als Antiseptika.
Wissen:
1. Ebenen der strukturellen Organisation von Proteinen.
2. Die Bedeutung der Primärstruktur von Proteinen, die ihre strukturelle und funktionelle Vielfalt bestimmt.
3. Der Mechanismus der Bildung des aktiven Zentrums in Proteinen und seine spezifische Wechselwirkung mit dem Liganden, der der Funktion von Proteinen zugrunde liegt.
4. Beispiele für den Einfluss exogener Liganden (Medikamente, Toxine, Gifte) auf die Konformation und funktionelle Aktivität von Proteinen.
5. Ursachen und Folgen der Proteindenaturierung, Faktoren, die die Denaturierung verursachen.
6. Beispiele für den Einsatz denaturierender Faktoren in der Medizin als Antiseptika und Mittel zur Sterilisation medizinischer Instrumente.
THEMA 1.1. STRUKTURELLE ORGANISATION VON PROTEINEN. STUFEN DER EINHEIMISCHEN BILDUNG
PROTEINKONFORMATIONEN
Proteine sind Polymermoleküle, deren Monomere nur aus 20 α-Aminosäuren bestehen. Der Satz und die Reihenfolge der Kombination von Aminosäuren in einem Protein wird durch die Struktur der Gene in der DNA von Individuen bestimmt. Jedes Protein erfüllt entsprechend seiner spezifischen Struktur seine charakteristische Funktion. Der Proteinsatz eines bestimmten Organismus bestimmt seine phänotypischen Eigenschaften sowie das Vorhandensein von Erbkrankheiten oder eine Veranlagung zu deren Entwicklung.
1. Aminosäuren, aus denen Proteine bestehen. Peptidbindung. Proteine sind Polymere, die aus Monomeren aufgebaut sind – 20 α-Aminosäuren, deren allgemeine Formel lautet
Aminosäuren unterscheiden sich in Struktur, Größe und physikalisch-chemischen Eigenschaften der an das α-Kohlenstoffatom gebundenen Radikale. Die funktionellen Gruppen der Aminosäuren bestimmen die Eigenschaften der verschiedenen α-Aminosäuren. Die in α-Aminosäuren vorkommenden Reste lassen sich in mehrere Gruppen einteilen:
Prolin, Im Gegensatz zu den anderen 19 Proteinmonomeren handelt es sich nicht um eine Aminosäure, sondern um eine Iminosäure; der Rest in Prolin ist sowohl mit dem α-Kohlenstoffatom als auch mit der Iminogruppe verbunden
 Aminosäuren unterscheiden sich in ihrer Löslichkeit in Wasser. Dies liegt an der Fähigkeit von Radikalen, mit Wasser (Hydrat) zu interagieren.
Aminosäuren unterscheiden sich in ihrer Löslichkeit in Wasser. Dies liegt an der Fähigkeit von Radikalen, mit Wasser (Hydrat) zu interagieren.
ZU hydrophil umfassen Radikale, die anionische, kationische und polare ungeladene funktionelle Gruppen enthalten.
ZU hydrophob umfassen Reste, die Methylgruppen, aliphatische Ketten oder Ringe enthalten.
2. Peptidbindungen verbinden Aminosäuren zu Peptiden. Während der Peptidsynthese interagiert die α-Carboxylgruppe einer Aminosäure mit der α-Aminogruppe einer anderen Aminosäure unter Bildung Peptidbindung:
Proteine sind Polypeptide, d.h. lineare Polymere aus α-Aminosäuren, die durch eine Peptidbindung verbunden sind (Abb. 1.1.)
 Reis. 1.1. Begriffe zur Beschreibung der Struktur von Peptiden
Reis. 1.1. Begriffe zur Beschreibung der Struktur von Peptiden
Die Monomere der Aminosäuren, aus denen Polypeptide bestehen, werden genannt Aminosäurereste. Eine Kette sich wiederholender Gruppen - NH-CH-CO- Formen Peptidrückgrat. Ein Aminosäurerest mit einer freien α-Aminogruppe wird als N-terminal bezeichnet, ein Aminosäurerest mit einer freien α-Carboxylgruppe wird als C-terminal bezeichnet. Peptide werden vom N-Terminus bis zum C-Terminus geschrieben und gelesen.
Die durch die Iminogruppe von Prolin gebildete Peptidbindung unterscheidet sich von anderen Peptidbindungen: Dem Stickstoffatom der Peptidgruppe fehlt ein Wasserstoffatom,
Stattdessen kommt es zu einer Bindung mit einem Rest, wodurch eine Seite des Rings in das Peptidrückgrat eingebunden wird:
 Peptide unterscheiden sich in der Aminosäurezusammensetzung, der Anzahl der Aminosäuren und der Reihenfolge der Aminosäureverbindungen. Beispielsweise sind Ser-Ala-Glu-Gis und His-Glu-Ala-Ser zwei verschiedene Peptide.
Peptide unterscheiden sich in der Aminosäurezusammensetzung, der Anzahl der Aminosäuren und der Reihenfolge der Aminosäureverbindungen. Beispielsweise sind Ser-Ala-Glu-Gis und His-Glu-Ala-Ser zwei verschiedene Peptide.
Peptidbindungen sind sehr stark und ihre chemische, nicht-enzymatische Hydrolyse erfordert raue Bedingungen: Das zu analysierende Protein wird 24 Stunden lang in konzentrierter Salzsäure bei einer Temperatur von etwa 110 °C hydrolysiert. In einer lebenden Zelle können Peptidbindungen aufgebrochen werden proteolytische Enzyme, angerufen Proteasen oder Peptidhydrolasen.
3. Primärstruktur von Proteinen. Aminosäurereste in den Peptidketten verschiedener Proteine wechseln sich nicht zufällig ab, sondern sind in einer bestimmten Reihenfolge angeordnet. Die lineare Abfolge oder Wechselreihenfolge von Aminosäureresten in einer Polypeptidkette wird als bezeichnet Primärstruktur des Proteins.
Die Primärstruktur jedes einzelnen Proteins ist im DNA-Molekül (in einer Region namens Gen) kodiert und wird während der Transkription (Kopieren von Informationen auf mRNA) und Translation (Synthese der Primärstruktur des Proteins) realisiert. Folglich handelt es sich bei der Primärstruktur der Proteine einer einzelnen Person um von den Eltern an die Kinder vererbte Informationen, die die Strukturmerkmale der Proteine eines bestimmten Organismus bestimmen, von denen die Funktion der vorhandenen Proteine abhängt (Abb. 1.2.).
 Reis. 1.2. Die Beziehung zwischen dem Genotyp und der Konformation der im Körper des Individuums synthetisierten Proteine
Reis. 1.2. Die Beziehung zwischen dem Genotyp und der Konformation der im Körper des Individuums synthetisierten Proteine
Jedes der rund 100.000 einzelnen Proteine im menschlichen Körper besitzt einzigartig Primärstruktur. Moleküle desselben Proteintyps (z. B. Albumin) weisen den gleichen Wechsel der Aminosäurereste auf, was Albumin von jedem anderen einzelnen Protein unterscheidet.
Die Reihenfolge der Aminosäurereste in einer Peptidkette kann als eine Form der Informationsaufzeichnung betrachtet werden. Diese Informationen bestimmen die räumliche Anordnung der linearen Peptidkette in eine kompaktere dreidimensionale Struktur namens Konformation Eichhörnchen. Der Prozess der Bildung einer funktionell aktiven Proteinkonformation wird aufgerufen falten
4. Proteinkonformation. Zwischen dem Stickstoffatom der Peptidgruppe und dem benachbarten α-Kohlenstoffatom sowie zwischen dem α-Kohlenstoffatom und dem Kohlenstoff der Carbonylgruppe ist eine freie Rotation im Peptidrückgrat möglich. Durch das Zusammenspiel funktioneller Gruppen von Aminosäureresten kann die Primärstruktur von Proteinen komplexere räumliche Strukturen annehmen. In globulären Proteinen gibt es zwei Hauptfaltungsebenen der Konformation von Peptidketten: sekundär Und Tertiärstruktur.
Sekundärstruktur von Proteinen ist eine räumliche Struktur, die durch die Bildung von Wasserstoffbrückenbindungen zwischen den funktionellen Gruppen -C=O und -NH- des Peptidrückgrats entsteht. In diesem Fall kann die Peptidkette zwei Arten regelmäßiger Strukturen annehmen: α-Helices Und β-Strukturen.
IN α-Helices Wasserstoffbrücken werden zwischen dem Sauerstoffatom der Carbonylgruppe und dem Wasserstoff des Amidstickstoffs der 4. Aminosäure gebildet; Seitenketten von Aminosäureresten
befinden sich entlang der Peripherie der Spirale, ohne an der Bildung der Sekundärstruktur beteiligt zu sein (Abb. 1.3.).
Massenradikale oder Radikale mit gleicher Ladung verhindern die Bildung einer α-Helix. Der Prolinrest, der eine Ringstruktur aufweist, unterbricht die α-Helix, da aufgrund des fehlenden Wasserstoffs am Stickstoffatom in der Peptidkette die Bildung einer Wasserstoffbrücke unmöglich ist. Die Bindung zwischen Stickstoff und dem α-Kohlenstoffatom ist Teil des Prolinrings, sodass das Peptidrückgrat an dieser Stelle gebogen wird.
β-Struktur wird zwischen den linearen Regionen des Peptidrückgrats einer Polypeptidkette gebildet und bildet dadurch gefaltete Strukturen. Es können sich Polypeptidketten oder Teile davon bilden parallel oder antiparallele β-Strukturen. Im ersten Fall fallen die N- und C-Termini der interagierenden Peptidketten zusammen, im zweiten Fall haben sie die entgegengesetzte Richtung (Abb. 1.4).
Reis. 1.3. Sekundärstruktur des Proteins - α-Helix
 Reis. 1.4. Parallele und antiparallele β-Faltblattstrukturen
Reis. 1.4. Parallele und antiparallele β-Faltblattstrukturen
β-Strukturen werden durch breite Pfeile angezeigt: A – Antiparallele β-Struktur. B – Parallele β-Faltblattstrukturen
In einigen Proteinen können β-Strukturen aufgrund der Bildung von Wasserstoffbrückenbindungen zwischen Atomen des Peptidrückgrats verschiedener Polypeptidketten gebildet werden.
Kommt auch in Proteinen vor Bereiche mit unregelmäßiger Sekundarstufe Struktur, die Biegungen, Schleifen und Windungen des Polypeptidrückgrats umfasst. Sie befinden sich häufig an Stellen, an denen sich die Richtung der Peptidkette ändert, beispielsweise wenn eine parallele β-Faltblattstruktur gebildet wird.
Basierend auf dem Vorhandensein von α-Helices und β-Strukturen können globuläre Proteine in vier Kategorien eingeteilt werden.
Reis. 1.5. Sekundärstruktur der Myoglobin- (A) und Hämoglobin-β-Kette (B), die acht α-Helices enthält

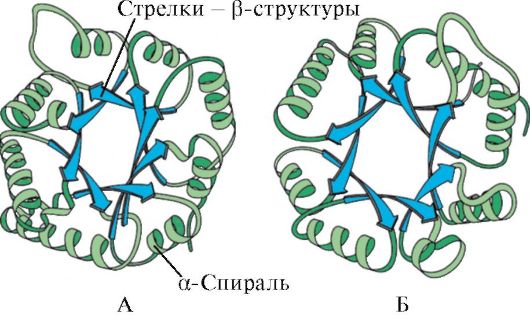 Reis. 1.6. Sekundärstruktur der Triosephosphat-Isomerase- und Pyruvatkinase-Domäne
Reis. 1.6. Sekundärstruktur der Triosephosphat-Isomerase- und Pyruvatkinase-Domäne
 Reis. 1.7. Sekundärstruktur der konstanten Domäne von Immunglobulin (A) und dem Enzym Superoxiddismutase (B)
Reis. 1.7. Sekundärstruktur der konstanten Domäne von Immunglobulin (A) und dem Enzym Superoxiddismutase (B)
IN vierte Kategorie Dazu gehörten Proteine, die eine kleine Menge regelmäßiger Sekundärstrukturen enthalten. Zu diesen Proteinen gehören kleine cysteinreiche Proteine oder Metalloproteine.
Protein-Tertiärstruktur- eine Art Konformation, die durch Wechselwirkungen zwischen Aminosäureresten entsteht, die in der Peptidkette weit voneinander entfernt sein können. Die meisten Proteine bilden eine räumliche Struktur, die einer Kugel ähnelt (globuläre Proteine).
Da hydrophobe Aminosäurereste dazu neigen, sich durch sogenannte zu verbinden Hydrophobe Wechselwirkungen und intermolekularen Van-der-Waals-Kräften bildet sich im Proteinkügelchen ein dichter hydrophober Kern. Hydrophile ionisierte und nichtionisierte Radikale befinden sich hauptsächlich auf der Oberfläche des Proteins und bestimmen dessen Löslichkeit in Wasser.
 Reis. 1.8. Arten von Bindungen, die zwischen Aminosäureresten während der Bildung der Tertiärstruktur eines Proteins entstehen
Reis. 1.8. Arten von Bindungen, die zwischen Aminosäureresten während der Bildung der Tertiärstruktur eines Proteins entstehen
1 - Ionenverbindung- kommt zwischen positiv und negativ geladenen funktionellen Gruppen vor;
2 - Wasserstoffverbindung– tritt zwischen einer hydrophilen ungeladenen Gruppe und jeder anderen hydrophilen Gruppe auf;
3 - Hydrophobe Wechselwirkungen- entstehen zwischen hydrophoben Radikalen;
4 - Disulfidbindung- entsteht durch Oxidation der SH-Gruppen von Cysteinresten und deren Wechselwirkung untereinander
Hydrophile Aminosäurereste, die sich im hydrophoben Kern befinden, können untereinander interagieren ionisch Und Wasserstoffbrücken(Abb. 1.8).
Ionen- und Wasserstoffbindungen sowie hydrophobe Wechselwirkungen sind schwach: Ihre Energie ist nicht viel höher als die Energie der thermischen Bewegung von Molekülen bei Raumtemperatur. Die Konformation des Proteins wird durch die Bildung vieler solcher schwacher Bindungen aufrechterhalten. Da die Atome, aus denen ein Protein besteht, ständig in Bewegung sind, ist es möglich, einige schwache Bindungen aufzubrechen und andere zu bilden, was zu leichten Bewegungen einzelner Abschnitte der Polypeptidkette führt. Diese Eigenschaft von Proteinen, ihre Konformation zu ändern, indem einige Bindungen aufgebrochen und andere schwache Bindungen gebildet werden, wird als schwache Bindungen bezeichnet Konformationslabilität.
Der menschliche Körper verfügt über unterstützende Systeme Homöostase- Konstanz der inneren Umgebung innerhalb bestimmter akzeptabler Grenzen für einen gesunden Körper. Unter Homöostasebedingungen stören kleine Konformationsänderungen die Gesamtstruktur und Funktion von Proteinen nicht. Die funktionell aktive Konformation eines Proteins nennt man native Konformation. Veränderungen in der inneren Umgebung (z. B. die Konzentration von Glukose, Ca-Ionen, Protonen usw.) führen zu Konformationsänderungen und Störungen der Proteinfunktionen.
Die Tertiärstruktur einiger Proteine wird stabilisiert Disulfidbindungen, entsteht durch die Wechselwirkung der -SH-Gruppen zweier Reste
 Reis. 1.9. Bildung einer Disulfidbindung in einem Proteinmolekül
Reis. 1.9. Bildung einer Disulfidbindung in einem Proteinmolekül
Cystein (Abb. 1.9). Die meisten intrazellulären Proteine weisen in ihrer Tertiärstruktur keine kovalenten Disulfidbindungen auf. Ihr Vorhandensein ist charakteristisch für die von der Zelle abgesonderten Proteine, was ihre größere Stabilität unter extrazellulären Bedingungen gewährleistet. Daher sind in den Molekülen von Insulin und Immunglobulinen Disulfidbindungen vorhanden.
Insulin- ein Proteinhormon, das in den β-Zellen der Bauchspeicheldrüse synthetisiert und als Reaktion auf einen Anstieg der Glukosekonzentration im Blut ins Blut ausgeschüttet wird.
 In der Struktur von Insulin gibt es zwei Disulfidbindungen, die die A- und B-Ketten des Polypeptids verbinden, und eine Disulfidbindung innerhalb der A-Kette (Abb. 1.10).
In der Struktur von Insulin gibt es zwei Disulfidbindungen, die die A- und B-Ketten des Polypeptids verbinden, und eine Disulfidbindung innerhalb der A-Kette (Abb. 1.10).
Reis. 1.10. Disulfidbindungen in der Struktur von Insulin 5. Supersekundäre Struktur von Proteinen. In Proteinen mit unterschiedlicher Primärstruktur und Funktion werden sie manchmal nachgewiesenähnliche Kombinationen und relative Positionen von Sekundärstrukturen,
die als Supersekundärstruktur bezeichnet werden. Es nimmt eine Zwischenstellung zwischen Sekundär- und Tertiärstruktur ein, da es sich um eine spezifische Kombination von Elementen der Sekundärstruktur bei der Bildung der Tertiärstruktur des Proteins handelt. Supersekundäre Strukturen haben spezifische Namen, wie „α-Helix-Turn-a-Helix“, „Leucin-Reißverschluss“, „Zinkfinger“ usw. Solche supersekundären Strukturen sind charakteristisch für DNA-bindende Proteine.„Leucin-Reißverschluss.“
 Diese Art von Supersekundärstruktur wird verwendet, um zwei Proteine miteinander zu verbinden. Auf der Oberfläche interagierender Proteine befinden sich α-helikale Regionen, die mindestens vier Leucinreste enthalten. Leucinreste in der α-Helix liegen sechs Aminosäuren voneinander entfernt. Da jede Windung der α-Helix 3,6 Aminosäurereste enthält, befinden sich Leucinradikale auf der Oberfläche jeder zweiten Windung. Leucinreste der α-Helix eines Proteins können mit Leucinresten eines anderen Proteins interagieren (hydrophobe Wechselwirkungen) und diese miteinander verbinden (Abb. 1.11.). Viele DNA-bindende Proteine funktionieren in oligomeren Komplexen, in denen die einzelnen Untereinheiten durch „Leucin-Reißverschlüsse“ miteinander verbunden sind.
Diese Art von Supersekundärstruktur wird verwendet, um zwei Proteine miteinander zu verbinden. Auf der Oberfläche interagierender Proteine befinden sich α-helikale Regionen, die mindestens vier Leucinreste enthalten. Leucinreste in der α-Helix liegen sechs Aminosäuren voneinander entfernt. Da jede Windung der α-Helix 3,6 Aminosäurereste enthält, befinden sich Leucinradikale auf der Oberfläche jeder zweiten Windung. Leucinreste der α-Helix eines Proteins können mit Leucinresten eines anderen Proteins interagieren (hydrophobe Wechselwirkungen) und diese miteinander verbinden (Abb. 1.11.). Viele DNA-bindende Proteine funktionieren in oligomeren Komplexen, in denen die einzelnen Untereinheiten durch „Leucin-Reißverschlüsse“ miteinander verbunden sind.
Reis. 1.11. „Leucin-Reißverschluss“ zwischen α-helikalen Regionen zweier Proteine Ein Beispiel für solche Proteine sind Histone. Histone
„Zinkfinger“- eine für DNA-bindende Proteine charakteristische Variante der Supersekundärstruktur, hat die Form eines länglichen Fragments auf der Oberfläche des Proteins und enthält etwa 20 Aminosäurereste (Abb. 1.12). Die Form des „verlängerten Fingers“ wird durch ein Zinkatom unterstützt, das an vier Aminosäurereste gebunden ist – zwei Cysteinreste und zwei Histidinreste. In einigen Fällen gibt es anstelle von Histidinresten Cysteinreste. Zwei eng beieinander liegende Cysteinreste sind von den beiden anderen Gisili-Resten durch eine Cys-Sequenz bestehend aus etwa 12 Aminosäureresten getrennt. Diese Region des Proteins bildet eine α-Helix, deren Reste spezifisch an die regulatorischen Regionen der großen DNA-Furche binden können. Individuelle Bindungsspezifität
 Reis. 1.12. Die Primärstruktur der Region der DNA-bindenden Proteine, die die „Zinkfinger“-Struktur bilden (die Buchstaben geben die Aminosäuren an, aus denen diese Struktur besteht)
Reis. 1.12. Die Primärstruktur der Region der DNA-bindenden Proteine, die die „Zinkfinger“-Struktur bilden (die Buchstaben geben die Aminosäuren an, aus denen diese Struktur besteht)
Das regulatorische DNA-Bindungsprotein hängt von der Sequenz der Aminosäurereste ab, die sich in der Zinkfingerregion befinden. Solche Strukturen enthalten insbesondere Rezeptoren für Steroidhormone, die an der Regulierung der Transkription (Lesen von Informationen von DNA zu RNA) beteiligt sind.
THEMA 1.2. GRUNDLAGEN DER PROTEINFUNKTION. ARZNEIMITTEL ALS LIGANDEN, DIE DIE PROTEINFUNKTION BEEINFLUSSEN
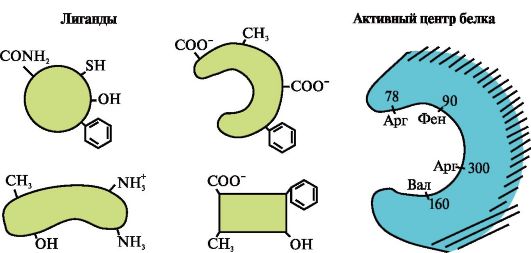
1. Das aktive Zentrum des Proteins und seine Wechselwirkung mit dem Liganden. Bei der Bildung der Tertiärstruktur entsteht auf der Oberfläche eines funktionell aktiven Proteins, meist in einer Vertiefung, ein Bereich, der durch in der Primärstruktur weit voneinander entfernte Aminosäurereste gebildet wird. Diese Region, die eine einzigartige Struktur für ein bestimmtes Protein aufweist und in der Lage ist, spezifisch mit einem bestimmten Molekül oder einer Gruppe ähnlicher Moleküle zu interagieren, wird als Protein-Ligand-Bindungsstelle oder aktive Stelle bezeichnet. Liganden sind Moleküle, die mit Proteinen interagieren.
Hohe Spezifität Die Wechselwirkung des Proteins mit dem Liganden wird durch die Komplementarität der Struktur des aktiven Zentrums zur Struktur des Liganden gewährleistet.
Komplementarität- das ist die räumliche und chemische Entsprechung interagierender Oberflächen. Das aktive Zentrum muss nicht nur räumlich dem darin enthaltenen Liganden entsprechen, sondern es müssen auch Bindungen (ionische, Wasserstoff- und hydrophobe Wechselwirkungen) zwischen den funktionellen Gruppen der im aktiven Zentrum enthaltenen Radikale und dem Liganden gebildet werden, die den Liganden halten im aktiven Zentrum (Abb. 1.13).
 Reis. 1.13. Komplementäre Wechselwirkung von Protein mit Ligand
Reis. 1.13. Komplementäre Wechselwirkung von Protein mit Ligand
Einige Liganden spielen, wenn sie an das aktive Zentrum eines Proteins gebunden sind, eine Hilfsrolle bei der Funktion von Proteinen. Solche Liganden werden als Cofaktoren bezeichnet, und Proteine, die einen Nicht-Protein-Anteil enthalten, werden als Cofaktoren bezeichnet komplexe Proteine(im Gegensatz zu einfachen Proteinen, die nur aus dem Proteinteil bestehen). Der Nicht-Protein-Anteil, der fest mit dem Protein verbunden ist, wird genannt Prothetische Gruppe. Beispielsweise enthalten Myoglobin, Hämoglobin und Cytochrome eine prosthetische Gruppe, Häm, die ein Eisenion enthält, das fest mit dem aktiven Zentrum verbunden ist. Komplexe Proteine, die Häm enthalten, werden Hämoproteine genannt.
Wenn bestimmte Liganden an Proteine gebunden werden, kommt die Funktion dieser Proteine zum Ausdruck. So erfüllt Albumin, das wichtigste Protein im Blutplasma, seine Transportfunktion, indem es hydrophobe Liganden wie Fettsäuren, Bilirubin, einige Medikamente usw. an das aktive Zentrum bindet (Abb. 1.14).
Liganden, die mit der dreidimensionalen Struktur der Peptidkette interagieren, können nicht nur niedermolekulare organische und anorganische Moleküle, sondern auch Makromoleküle sein:
DNA (Beispiele mit oben diskutierten DNA-bindenden Proteinen);
Polysaccharide;
 Reis. 1.14. Zusammenhang zwischen Genotyp und Phänotyp
Reis. 1.14. Zusammenhang zwischen Genotyp und Phänotyp
Die einzigartige Primärstruktur menschlicher Proteine, die im DNA-Molekül kodiert ist, wird in Zellen in Form einer einzigartigen Konformation, aktiven Zentrumsstruktur und Proteinfunktionen realisiert
In diesen Fällen erkennt das Protein eine spezifische Region des Liganden, die der Bindungsstelle entspricht und komplementär ist. So befinden sich auf der Oberfläche von Hepatozyten Rezeptorproteine für das Hormon Insulin, das ebenfalls eine Proteinstruktur aufweist. Die Wechselwirkung von Insulin mit dem Rezeptor führt zu einer Änderung seiner Konformation und Aktivierung von Signalsystemen, was zur Speicherung von Nährstoffen in Hepatozyten nach den Mahlzeiten führt.
Auf diese Weise, Die Funktion von Proteinen basiert auf der spezifischen Interaktion des aktiven Zentrums des Proteins mit dem Liganden.
2. Domänenstruktur und ihre Rolle bei der Funktion von Proteinen. Lange Polypeptidketten globulärer Proteine falten sich häufig in mehrere kompakte, relativ unabhängige Regionen. Sie haben eine eigenständige Tertiärstruktur, die an die globulärer Proteine erinnert, und werden „ Domänen. Aufgrund der Domänenstruktur von Proteinen lässt sich ihre Tertiärstruktur leichter ausbilden.
In Domänenproteinen befinden sich Ligandenbindungsstellen häufig zwischen Domänen. Somit ist Trypsin ein proteolytisches Enzym, das vom exokrinen Teil der Bauchspeicheldrüse produziert wird und für die Verdauung von Nahrungsproteinen notwendig ist. Es hat eine Zwei-Domänen-Struktur und das Bindungszentrum von Trypsin mit seinem Liganden – dem Nahrungsprotein – befindet sich in der Furche zwischen den beiden Domänen. Im aktiven Zentrum werden die Bedingungen geschaffen, die für eine wirksame Bindung einer bestimmten Stelle des Nahrungsproteins und die Hydrolyse seiner Peptidbindungen erforderlich sind.
Verschiedene Domänen in einem Protein können sich relativ zueinander bewegen, wenn das aktive Zentrum mit dem Liganden interagiert (Abb. 1.15).
Hexokinase– ein Enzym, das die Phosphorylierung von Glucose mithilfe von ATP katalysiert. Das aktive Zentrum des Enzyms befindet sich in der Spalte zwischen den beiden Domänen. Wenn Hexokinase an Glucose bindet, schließen sich die sie umgebenden Domänen und das Substrat wird eingefangen, wo die Phosphorylierung stattfindet (siehe Abb. 1.15).
 Reis. 1.15. Bindung von Hexokinase-Domänen an Glucose
Reis. 1.15. Bindung von Hexokinase-Domänen an Glucose
In einigen Proteinen erfüllen Domänen unabhängige Funktionen, indem sie an verschiedene Liganden binden. Solche Proteine werden als multifunktional bezeichnet.
3. Medikamente sind Liganden, die die Funktion von Proteinen beeinflussen. Die Wechselwirkung von Proteinen mit Liganden ist spezifisch. Aufgrund der Konformationslabilität des Proteins und seines aktiven Zentrums ist es jedoch möglich, eine andere Substanz auszuwählen, die ebenfalls mit dem Protein im aktiven Zentrum oder einem anderen Teil des Moleküls interagieren könnte.
Eine Substanz, die in ihrer Struktur einem natürlichen Liganden ähnelt, wird genannt Strukturanalogon des Liganden oder ein nichtnatürlicher Ligand. Es interagiert auch mit dem Protein im aktiven Zentrum. Ein strukturelles Analogon eines Liganden kann beide die Proteinfunktion verbessern (Agonist), und reduzieren Sie es (Gegner). Der Ligand und seine Strukturanaloga konkurrieren miteinander um die Bindung an das Protein an derselben Stelle. Solche Stoffe nennt man wettbewerbsfähige Modulatoren(Regulatoren) von Proteinfunktionen. Viele Medikamente wirken als Proteinhemmer. Einige davon werden durch chemische Modifikation natürlicher Liganden gewonnen. Inhibitoren von Proteinfunktionen können Medikamente und Gifte sein.
Atropin ist ein kompetitiver Inhibitor der M-cholinergen Rezeptoren. Acetylcholin ist ein Neurotransmitter zur Übertragung von Nervenimpulsen über cholinerge Synapsen. Um eine Erregung durchzuführen, muss das in den synaptischen Spalt freigesetzte Acetylcholin mit dem Rezeptorprotein der postsynaptischen Membran interagieren. Zwei Typen gefunden cholinerge Rezeptoren:
M-Rezeptor Zusätzlich zu Acetylcholin interagiert es selektiv mit Muscarin (Fliegenpilztoxin). M – cholinerge Rezeptoren sind auf der glatten Muskulatur vorhanden und bewirken bei Wechselwirkung mit Acetylcholin deren Kontraktion;
H-Rezeptor spezifisch an Nikotin binden. N-cholinerge Rezeptoren finden sich an den Synapsen der quergestreiften Skelettmuskulatur.
Spezifischer Inhibitor M-cholinerge Rezeptoren ist Atropin. Es kommt in Belladonna- und Bilsenkrautpflanzen vor.
 Atropin verfügt über funktionelle Gruppen, die in ihrer Struktur und räumlichen Anordnung Acetylcholin ähneln, und ist daher ein kompetitiver Inhibitor der M-cholinergen Rezeptoren. Da die Bindung von Acetylcholin an M-cholinerge Rezeptoren eine Kontraktion der glatten Muskulatur verursacht, wird Atropin als Arzneimittel zur Linderung ihrer Krämpfe eingesetzt (krampflösend). So ist es bekannt, Atropin zur Entspannung der Augenmuskulatur beim Betrachten des Augenhintergrundes sowie zur Linderung von Krämpfen bei Magen-Darm-Koliken einzusetzen. M-cholinerge Rezeptoren sind auch im Zentralnervensystem (ZNS) vorhanden, sodass große Atropindosen eine unerwünschte Reaktion des Zentralnervensystems hervorrufen können: motorische und geistige Unruhe, Halluzinationen, Krämpfe.
Atropin verfügt über funktionelle Gruppen, die in ihrer Struktur und räumlichen Anordnung Acetylcholin ähneln, und ist daher ein kompetitiver Inhibitor der M-cholinergen Rezeptoren. Da die Bindung von Acetylcholin an M-cholinerge Rezeptoren eine Kontraktion der glatten Muskulatur verursacht, wird Atropin als Arzneimittel zur Linderung ihrer Krämpfe eingesetzt (krampflösend). So ist es bekannt, Atropin zur Entspannung der Augenmuskulatur beim Betrachten des Augenhintergrundes sowie zur Linderung von Krämpfen bei Magen-Darm-Koliken einzusetzen. M-cholinerge Rezeptoren sind auch im Zentralnervensystem (ZNS) vorhanden, sodass große Atropindosen eine unerwünschte Reaktion des Zentralnervensystems hervorrufen können: motorische und geistige Unruhe, Halluzinationen, Krämpfe.
Ditilin ist ein kompetitiver Agonist von H-cholinergen Rezeptoren, der die Funktion neuromuskulärer Synapsen hemmt.
 Neuromuskuläre Synapsen der Skelettmuskulatur enthalten H-cholinerge Rezeptoren. Ihre Wechselwirkung mit Acetylcholin führt zu Muskelkontraktionen. Bei einigen chirurgischen Eingriffen sowie bei endoskopischen Untersuchungen werden Medikamente eingesetzt, die eine Entspannung der Skelettmuskulatur bewirken (Muskelrelaxantien). Dazu gehört Dithilin, ein Strukturanalogon von Acetylcholin. Es bindet an H-cholinerge Rezeptoren, wird aber im Gegensatz zu Acetylcholin sehr langsam durch das Enzym Acetylcholinesterase zerstört. Durch längeres Öffnen der Ionenkanäle und anhaltende Depolarisation der Membran wird die Weiterleitung von Nervenimpulsen gestört und es kommt zu einer Muskelentspannung. Ursprünglich wurden diese Eigenschaften im Curare-Gift entdeckt, weshalb solche Medikamente genannt werden Curare-artig.
Neuromuskuläre Synapsen der Skelettmuskulatur enthalten H-cholinerge Rezeptoren. Ihre Wechselwirkung mit Acetylcholin führt zu Muskelkontraktionen. Bei einigen chirurgischen Eingriffen sowie bei endoskopischen Untersuchungen werden Medikamente eingesetzt, die eine Entspannung der Skelettmuskulatur bewirken (Muskelrelaxantien). Dazu gehört Dithilin, ein Strukturanalogon von Acetylcholin. Es bindet an H-cholinerge Rezeptoren, wird aber im Gegensatz zu Acetylcholin sehr langsam durch das Enzym Acetylcholinesterase zerstört. Durch längeres Öffnen der Ionenkanäle und anhaltende Depolarisation der Membran wird die Weiterleitung von Nervenimpulsen gestört und es kommt zu einer Muskelentspannung. Ursprünglich wurden diese Eigenschaften im Curare-Gift entdeckt, weshalb solche Medikamente genannt werden Curare-artig.
THEMA 1.3. DENATURIERUNG VON PROTEINEN UND DIE MÖGLICHKEIT IHRER SPONTANEN RENATIVIERUNG
1. Da die native Konformation von Proteinen aufgrund schwacher Wechselwirkungen aufrechterhalten wird, führen Änderungen in der Zusammensetzung und den Eigenschaften der das Protein umgebenden Umgebung sowie die Einwirkung chemischer Reagenzien und physikalischer Faktoren zu einer Änderung ihrer Konformation (die Eigenschaft der Konformationslabilität). Das Aufbrechen einer großen Anzahl von Bindungen führt zur Zerstörung der nativen Konformation und zur Denaturierung von Proteinen.
Denaturierung von Proteinen- Dies ist die Zerstörung ihrer nativen Konformation unter dem Einfluss von Denaturierungsmitteln, die durch den Bruch schwacher Bindungen verursacht wird, die die räumliche Struktur des Proteins stabilisieren. Die Denaturierung geht mit der Zerstörung der einzigartigen dreidimensionalen Struktur und des aktiven Zentrums des Proteins sowie dem Verlust seiner biologischen Aktivität einher (Abb. 1.16).
Alle denaturierten Moleküle eines Proteins nehmen eine zufällige Konformation an, die sich von anderen Molekülen desselben Proteins unterscheidet. Es stellt sich heraus, dass die Aminosäurereste, die das aktive Zentrum bilden, räumlich voneinander entfernt sind, d. h. die spezifische Bindungsstelle des Proteins mit dem Liganden wird zerstört. Bei der Denaturierung bleibt die Primärstruktur der Proteine unverändert.
Anwendung von Denaturierungsmitteln in der biologischen Forschung und Medizin. Bei biochemischen Untersuchungen werden vor der Bestimmung niedermolekularer Verbindungen in biologischem Material in der Regel zunächst Proteine aus der Lösung entfernt. Zu diesem Zweck wird am häufigsten Trichloressigsäure (TCA) verwendet. Nach Zugabe von TCA zur Lösung fallen denaturierte Proteine aus und lassen sich leicht durch Filtration entfernen (Tabelle 1.1).
In der Medizin werden Denaturierungsmittel häufig zur Sterilisation medizinischer Instrumente und Materialien in Autoklaven (Denaturierungsmittel bei hoher Temperatur) und als Antiseptika (Alkohol, Phenol, Chloramin) zur Behandlung kontaminierter Oberflächen mit pathogener Mikroflora eingesetzt.
2. Spontane Reaktivierung von Proteinen- Nachweis des Determinismus der Primärstruktur, Konformation und Funktion von Proteinen. Einzelne Proteine sind Produkte eines Gens, die eine identische Aminosäuresequenz haben und in der Zelle die gleiche Konformation annehmen. Die grundlegende Schlussfolgerung, dass die Primärstruktur eines Proteins bereits Informationen über seine Konformation und Funktion enthält, wurde auf der Grundlage der Fähigkeit einiger Proteine (insbesondere Ribonuklease und Myoglobin) gezogen, sich spontan zu renativieren – also ihre native Konformation nach der Denaturierung wiederherzustellen.
Die Bildung räumlicher Proteinstrukturen erfolgt durch die Methode der Selbstorganisation – ein spontaner Prozess, bei dem eine Polypeptidkette, die eine einzigartige Primärstruktur aufweist, dazu neigt, in Lösung eine Konformation mit der niedrigsten freien Energie anzunehmen. Die Fähigkeit, Proteine, die nach der Denaturierung ihre Primärstruktur behalten, zu renativieren, wurde in einem Experiment mit dem Enzym Ribonuklease beschrieben.
Ribonuklease ist ein Enzym, das die Bindungen zwischen einzelnen Nukleotiden in einem RNA-Molekül auflöst. Dieses kugelförmige Protein verfügt über eine Polypeptidkette, deren Tertiärstruktur durch viele schwache und vier Disulfidbindungen stabilisiert wird.
Die Behandlung von Ribonuklease mit Harnstoff, der Wasserstoffbrückenbindungen im Molekül aufbricht, und einem Reduktionsmittel, das Disulfidbindungen aufbricht, führt zur Denaturierung des Enzyms und zum Verlust seiner Aktivität.
Die Entfernung von Denaturierungsmitteln durch Dialyse führt zur Wiederherstellung der Proteinkonformation und -funktion, d. h. zur Wiedergeburt. (Abb. 1.17).
 Reis. 1.17. Denaturierung und Renativierung der Ribonuklease
Reis. 1.17. Denaturierung und Renativierung der Ribonuklease
A – native Konformation der Ribonuklease, in deren Tertiärstruktur vier Disulfidbindungen vorhanden sind; B – denaturiertes Ribonukleasemolekül;
B – reaktiviertes Ribonuklease-Molekül mit wiederhergestellter Struktur und Funktion
1. Füllen Sie Tabelle 1.2 aus.
Tabelle 1.2. Klassifizierung von Aminosäuren nach der Polarität der Radikale
2. Schreiben Sie die Formel des Tetrapeptids:
Rapfen – Profi – Fen – Liz
a) Markieren Sie die sich wiederholenden Gruppen im Peptid, die das Peptidrückgrat bilden, und die variablen Gruppen, die durch Aminosäurereste dargestellt werden;
b) die N- und C-Termini beschriften;
c) Markieren Sie die Peptidbindungen;
d) ein weiteres Peptid schreiben, das aus denselben Aminosäuren besteht;
e) Zählen Sie die Anzahl möglicher Varianten eines Tetrapeptids mit ähnlicher Aminosäurezusammensetzung.
3. Erklären Sie die Rolle der Primärstruktur von Proteinen am Beispiel einer vergleichenden Analyse zweier strukturell ähnlicher und evolutionär ähnlicher Peptidhormone der Neurohypophyse von Säugetieren – Oxytocin und Vasopressin (Tabelle 1.3).
Tabelle 1.3. Struktur und Funktionen von Oxytocin und Vasopressin
 Dafür:
Dafür:
a) Vergleichen Sie die Zusammensetzung und Aminosäuresequenz zweier Peptide;
b) Finden Sie die Ähnlichkeit der Primärstruktur der beiden Peptide und die Ähnlichkeit ihrer biologischen Wirkung;
c) Unterschiede in der Struktur zweier Peptide und Unterschiede in ihren Funktionen finden;
d) eine Schlussfolgerung über den Einfluss der Primärstruktur von Peptiden auf ihre Funktionen ziehen.
4. Beschreiben Sie die Hauptstadien der Konformationsbildung globulärer Proteine (Sekundär-, Tertiärstrukturen, Konzept der Supersekundärstruktur). Geben Sie die Arten von Bindungen an, die an der Bildung von Proteinstrukturen beteiligt sind. Welche Aminosäurereste können an der Bildung hydrophober Wechselwirkungen, ionischer Wasserstoffbrückenbindungen, beteiligt sein?
Nenne Beispiele.
5. Definieren Sie das Konzept der „Konformationslabilität von Proteinen“ und geben Sie die Gründe für seine Existenz und Bedeutung an.
6. Erweitern Sie die Bedeutung des folgenden Satzes: „Die Funktion von Proteinen basiert auf ihrer spezifischen Interaktion mit dem Liganden“, indem Sie die Begriffe verwenden und ihre Bedeutung erklären: Proteinkonformation, aktives Zentrum, Ligand, Komplementarität, Proteinfunktion.
7. Erklären Sie anhand eines Beispiels, was Domänen sind und welche Rolle sie bei der Funktion von Proteinen spielen.
AUFGABEN ZUR SELBSTKONTROLLE
1. Übereinstimmen.
Funktionelle Gruppe im Aminosäurerest:
A. Carboxylgruppe B. Hydroxylgruppe C Guanidingruppe D. Thiolgruppe E. Aminogruppe
2. Wähle die richtigen Antworten.
Aminosäuren mit polaren ungeladenen Radikalen sind:
A. Cis B. Asn
B. Glu G. Drei
3. Wähle die richtigen Antworten.
Aminosäureradikale:
A. Spezifität der Primärstruktur bereitstellen B. An der Bildung der Tertiärstruktur beteiligt sein
B. Sie befinden sich auf der Oberfläche des Proteins und beeinflussen dessen Löslichkeit. D. Sie bilden das aktive Zentrum
D. Beteiligen Sie sich an der Bildung von Peptidbindungen
4. Wähle die richtigen Antworten.
Zwischen Aminosäureresten können sich hydrophobe Wechselwirkungen bilden:
A. Tre Lay B. Pro Three
B. Met Ile G. Tir Ala D. Val Fen
5. Wähle die richtigen Antworten.
Zwischen Aminosäureresten können sich ionische Bindungen bilden:
A. Gln Asp B. Apr Liz
B. Liz Glu G. Gis Asp D. Asn Apr
6. Wähle die richtigen Antworten.
Zwischen Aminosäureresten können sich Wasserstoffbrückenbindungen bilden:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Übereinstimmen.
Art der Bindung, die an der Bildung der Proteinstruktur beteiligt ist:
A. Primärstruktur B. Sekundärstruktur
B. Tertiärstruktur
D. Supersekundäre Struktur E. Konformation.
1. Wasserstoffbrückenbindungen zwischen Atomen des Peptidrückgrats
2. Schwache Bindungen zwischen funktionellen Gruppen von Aminosäureresten
3. Bindungen zwischen α-Amino- und α-Carboxylgruppen von Aminosäuren
8. Wähle die richtigen Antworten. Trypsin:
A. Proteolytisches Enzym B. Enthält zwei Domänen
B. Hydrolysiert Stärke
D. Die aktive Seite befindet sich zwischen den Domänen. D. Besteht aus zwei Polypeptidketten.
9. Wähle die richtigen Antworten. Atropin:
A. Neurotransmitter
B. Strukturanalogon von Acetylcholin
B. Interagiert mit H-cholinergen Rezeptoren
D. Stärkt die Weiterleitung von Nervenimpulsen durch cholinerge Synapsen
D. Kompetitiver Inhibitor von M-cholinergen Rezeptoren
10. Wählen Sie die richtigen Aussagen. In Proteinen:
A. Die Primärstruktur enthält Informationen über die Struktur ihres aktiven Zentrums
B. Das aktive Zentrum wird auf der Ebene der Primärstruktur gebildet
B. Die Konformation ist durch kovalente Bindungen starr fixiert
D. Das aktive Zentrum kann mit einer Gruppe ähnlicher Liganden interagieren
aufgrund der Konformationslabilität von Proteinen D. Veränderungen in der Umgebung können die Affinität des Wirkstoffs beeinflussen
Zentrum zum Liganden
1. 1-B, 2-G, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
GRUNDLEGENDE BEGRIFFE UND KONZEPTE
1. Protein, Polypeptid, Aminosäuren
2. Primäre, sekundäre, tertiäre Proteinstrukturen
3. Konformation, native Proteinkonformation
4. Kovalente und schwache Bindungen im Protein
5. Konformationslabilität
6. Aktives Zentrum des Proteins
7. Liganden
8. Proteinfaltung
9. Strukturanaloge von Liganden
10. Domänenproteine
11. Einfache und komplexe Proteine
12. Proteindenaturierung, Denaturierungsmittel
13. Reaktivierung von Proteinen
Probleme lösen
„Strukturelle Organisation von Proteinen und die Grundlagen ihrer Funktion“
1. Die Hauptfunktion des Proteins Hämoglobin A (HbA) ist der Transport von Sauerstoff zu Geweben. In der menschlichen Bevölkerung sind mehrere Formen dieses Proteins mit veränderten Eigenschaften und Funktionen bekannt – die sogenannten abnormalen Hämoglobine. Beispielsweise wurde festgestellt, dass Hämoglobin S, das in den roten Blutkörperchen von Patienten mit Sichelzellenanämie (HbS) vorkommt, unter Bedingungen eines niedrigen Sauerstoffpartialdrucks (wie es im venösen Blut der Fall ist) eine geringe Löslichkeit aufweist. Dies führt zur Bildung von Aggregaten dieses Proteins. Das Protein verliert seine Funktion, fällt aus und die roten Blutkörperchen nehmen eine unregelmäßige Form an (einige von ihnen bilden eine Sichelform) und werden in der Milz schneller als gewöhnlich zerstört. Infolgedessen entwickelt sich eine Sichelzellenanämie.
Der einzige Unterschied in der Primärstruktur von HbA wurde im N-terminalen Bereich der Hämoglobin-β-Kette gefunden. Vergleichen Sie die N-terminalen Regionen des β-Strangs und zeigen Sie, wie sich Änderungen in der Primärstruktur des Proteins auf seine Eigenschaften und Funktionen auswirken.
 Dafür:
Dafür:
a) Schreiben Sie die Formeln der Aminosäuren auf, durch die sich HbA unterscheidet, und vergleichen Sie die Eigenschaften dieser Aminosäuren (Polarität, Ladung).
b) eine Schlussfolgerung über den Grund für die Abnahme der Löslichkeit und die Störung des Sauerstofftransports im Gewebe ziehen.
2. Die Abbildung zeigt schematisch die Struktur eines Proteins, das über ein Bindungszentrum mit einem Liganden (aktives Zentrum) verfügt. Erklären Sie, warum das Protein bei der Wahl des Liganden selektiv ist. Dafür:
a) Merken Sie sich, was das aktive Zentrum eines Proteins ist, und betrachten Sie die in der Abbildung gezeigte Struktur des aktiven Zentrums des Proteins.
b) Schreiben Sie die Formeln der Aminosäurereste, aus denen das aktive Zentrum besteht;
c) Zeichnen Sie einen Liganden, der spezifisch mit dem aktiven Zentrum des Proteins interagieren könnte. Geben Sie darauf die funktionellen Gruppen an, die Bindungen mit den Aminosäureresten eingehen können, aus denen das aktive Zentrum besteht.
d) geben Sie die Arten der Bindungen an, die zwischen den Liganden- und Aminosäureresten des aktiven Zentrums entstehen;
e) Erklären Sie, worauf die Spezifität der Protein-Ligand-Wechselwirkung beruht.
 3.
Die Abbildung zeigt das aktive Zentrum des Proteins und mehrere Liganden.
3.
Die Abbildung zeigt das aktive Zentrum des Proteins und mehrere Liganden.
Bestimmen Sie, welcher Ligand am wahrscheinlichsten mit dem aktiven Zentrum des Proteins interagiert und warum.
 Welche Arten von Bindungen entstehen bei der Bildung eines Protein-Ligand-Komplexes?
Welche Arten von Bindungen entstehen bei der Bildung eines Protein-Ligand-Komplexes?
4. Strukturanaloge natürlicher Proteinliganden können als Arzneimittel zur Modifizierung der Aktivität von Proteinen eingesetzt werden.
Acetylcholin ist ein Mediator der Erregungsübertragung an neuromuskulären Synapsen. Wenn Acetylcholin mit Proteinen – Rezeptoren der postsynaptischen Membran der Skelettmuskulatur – interagiert, öffnen sich Ionenkanäle und es kommt zu einer Muskelkontraktion. Ditilin ist ein Arzneimittel, das bei einigen Operationen zur Muskelentspannung eingesetzt wird, da es die Übertragung von Nervenimpulsen durch neuromuskuläre Synapsen stört. Erklären Sie den Wirkungsmechanismus von Ditilin als Muskelrelaxans. Dafür:
a) Schreiben Sie die Formeln von Acetylcholin und Dithilin und vergleichen Sie ihre Strukturen;
b) Beschreiben Sie den Mechanismus der entspannenden Wirkung von Ditilin.
5. Bei manchen Erkrankungen steigt die Körpertemperatur des Patienten an, was als Schutzreaktion des Körpers gewertet wird. Hohe Temperaturen wirken sich jedoch schädlich auf die Körperproteine aus. Erklären Sie, warum bei Temperaturen über 40 °C die Funktion von Proteinen gestört wird und eine Gefahr für das menschliche Leben entsteht. Denken Sie dazu daran:
1) Die Struktur von Proteinen und die Bindungen, die ihre Struktur in der nativen Konformation halten;
2) Wie verändern sich Struktur und Funktion von Proteinen mit steigender Temperatur?;
3) Was ist Homöostase und warum ist sie für die Erhaltung der menschlichen Gesundheit wichtig?
Moduleinheit 2 OLIGOMERICKE PROTEINE ALS ZIELE REGULATORISCHER EINFLÜSSE. STRUKTURELLE UND FUNKTIONELLE VIELFALT VON PROTEINEN. VERFAHREN ZUR TRENNUNG UND REINIGUNG VON PROTEINEN
Lernziele: In der Lage sein:
1. Nutzen Sie Kenntnisse über die Merkmale der Struktur und Funktionen oligomerer Proteine, um die adaptiven Mechanismen der Regulierung ihrer Funktionen zu verstehen.
2. Erklären Sie die Rolle von Chaperonen bei der Synthese und Aufrechterhaltung der Proteinkonformation unter zellulären Bedingungen.
3. Erklären Sie die Vielfalt der Erscheinungsformen des Lebens anhand der Vielfalt der Strukturen und Funktionen der im Körper synthetisierten Proteine.
4. Analysieren Sie den Zusammenhang zwischen der Struktur von Proteinen und ihrer Funktion anhand von Vergleichsbeispielen verwandter Hämoproteine – Myoglobin und Hämoglobin – sowie Vertretern der fünf Proteinklassen der Immunglobulinfamilie.
5. Wenden Sie Kenntnisse über die Besonderheiten der physikalischen und chemischen Eigenschaften von Proteinen an, um Methoden zu ihrer Reinigung von anderen Proteinen und Verunreinigungen auszuwählen.
6. Interpretieren Sie die Ergebnisse der quantitativen und qualitativen Zusammensetzung von Blutplasmaproteinen, um die klinische Diagnose zu bestätigen oder zu klären.
Wissen:
1. Merkmale der Struktur oligomerer Proteine und adaptive Mechanismen zur Regulierung ihrer Funktionen am Beispiel von Hämoglobin.
2. Die Struktur und Funktionen von Chaperonen und ihre Bedeutung für die Aufrechterhaltung der nativen Konformation von Proteinen unter zellulären Bedingungen.
3. Prinzipien der Zusammenfassung von Proteinen zu Familien aufgrund der Ähnlichkeit ihrer Konformation und Funktion am Beispiel von Immunglobulinen.
4. Methoden zur Trennung von Proteinen anhand der Merkmale ihrer physikalisch-chemischen Eigenschaften.
5. Elektrophorese von Blutplasma als Methode zur Beurteilung der qualitativen und quantitativen Zusammensetzung von Proteinen.
THEMA 1.4. Merkmale der Struktur und Funktion oligomerer Proteine am Beispiel von Hämoglobin
1. Viele Proteine enthalten mehrere Polypeptidketten. Solche Proteine heißen oligomer, und einzelne Ketten - Protomer. Protomere in oligomeren Proteinen sind durch viele schwache nichtkovalente Bindungen (hydrophob, ionisch, Wasserstoff) verbunden. Interaktion
Protomer werden dank durchgeführt Komplementarität deren Kontaktflächen.
Die Anzahl der Protomeren in oligomeren Proteinen kann stark variieren: Hämoglobin enthält 4 Protomere, das Enzym Aspartataminotransferase hat 12 Protomere und das Protein des Tabakmosaikvirus enthält 2120 Protomere, die durch nichtkovalente Bindungen verbunden sind. Folglich können oligomere Proteine sehr hohe Molekulargewichte aufweisen.
Die Wechselwirkung eines Protomers mit anderen kann als Sonderfall der Protein-Ligand-Wechselwirkung angesehen werden, da jedes Protomer als Ligand für andere Protomere dient. Die Anzahl und Art der Verbindung von Protomeren in einem Protein wird genannt Quartäre Proteinstruktur.
Proteine können Protomeren gleicher oder unterschiedlicher Struktur enthalten. Homodimere sind beispielsweise Proteine, die zwei identische Protomere enthalten, und Heterodimere sind Proteine, die zwei unterschiedliche Protomere enthalten.
Enthalten Proteine unterschiedliche Protomere, so können sich auf ihnen Bindungszentren mit unterschiedlichen Liganden ausbilden, die sich in ihrer Struktur unterscheiden. Wenn ein Ligand an das aktive Zentrum bindet, wird die Funktion dieses Proteins manifestiert. Ein Zentrum, das sich auf einem anderen Protomer befindet, wird als allosterisch (anders als das aktive) bezeichnet. Kontaktaufnahme es übt eine regulatorische Funktion aus (Abb. 1.18). Die Wechselwirkung des allosterischen Zentrums mit dem Effektor führt aufgrund seiner Konformationslabilität zu Konformationsänderungen in der Struktur des gesamten oligomeren Proteins. Dies beeinflusst die Affinität des aktiven Zentrums für einen bestimmten Liganden und reguliert die Funktion dieses Proteins. Eine Änderung der Konformation und Funktion aller Protomere während der Wechselwirkung eines oligomeren Proteins mit mindestens einem Liganden wird als kooperative Konformationsänderung bezeichnet. Es werden Effektoren genannt, die die Funktion von Proteinen verbessern Aktivatoren, und Effektoren, die seine Funktion hemmen - Inhibitoren.
Somit haben oligomere Proteine, aber auch Proteine mit Domänenstruktur, im Vergleich zu monomeren Proteinen eine neue Eigenschaft – die Fähigkeit, Funktionen allosterisch zu regulieren (Regulierung durch Anheften verschiedener Liganden an das Protein). Dies lässt sich anhand eines Vergleichs der Strukturen und Funktionen zweier eng verwandter komplexer Proteine, Myoglobin und Hämoglobin, erkennen.
 Reis. 1.18. Schema der Struktur eines dimeren Proteins
Reis. 1.18. Schema der Struktur eines dimeren Proteins
2. Bildung räumlicher Strukturen und Funktion von Myoglobin.
Myoglobin (Mb) ist ein Protein, das in roten Muskeln vorkommt und dessen Hauptfunktion darin besteht, O2-Reserven zu schaffen, die für intensive Muskelarbeit notwendig sind. Mb ist ein komplexes Protein, das einen Proteinteil – ApoMb – und einen Nicht-Proteinteil – Häm – enthält. Die Primärstruktur von apoMB bestimmt seine kompakte kugelförmige Konformation und die Struktur des aktiven Zentrums, an das der Nicht-Protein-Teil von Myoglobin, Häm, gebunden ist. Sauerstoff, der aus dem Blut zu den Muskeln gelangt, bindet an Fe+2-Häme im Myoglobin. Mb ist ein monomeres Protein mit einer sehr hohen Affinität zu O 2, daher erfolgt die Freisetzung von Sauerstoff durch Myoglobin nur bei intensiver Muskelarbeit, wenn der Partialdruck von O 2 stark abnimmt.
Bildung der Mv-Konformation. In roten Muskeln wird auf Ribosomen während der Translation die Primärstruktur von MB synthetisiert, dargestellt durch eine spezifische Sequenz von 153 Aminosäureresten. Die Sekundärstruktur von Mb enthält acht α-Helices, die in lateinischen Buchstaben von A bis H bezeichnet werden und zwischen denen sich nicht-helikale Bereiche befinden. Die Tertiärstruktur von Mb hat die Form einer kompakten Kugel, in deren Vertiefung sich das aktive Zentrum zwischen den F- und E-α-Helices befindet (Abb. 1.19).
 Reis. 1.19. Myoglobinstruktur
Reis. 1.19. Myoglobinstruktur
3. Merkmale der Struktur und Funktionsweise des aktiven Zentrums von MV. Das aktive Zentrum von Mb wird überwiegend durch hydrophobe Aminosäurereste gebildet, die in der Primärstruktur weit voneinander entfernt sind (z. B. Tri). 3 9 und Fen 138) In Wasser schlecht lösliche Liganden – Häm und O 2 – binden an das aktive Zentrum. Häm ist ein spezifischer Ligand von apoMB (Abb. 1.20), dessen Basis aus vier durch Methenylbrücken verbundenen Pyrrolringen besteht; Im Zentrum befindet sich ein Fe+2-Atom, das über vier Koordinationsbindungen mit den Stickstoffatomen der Pyrrolringe verbunden ist. Im aktiven Zentrum von Mb befinden sich neben hydrophoben Aminosäureresten auch Reste von zwei Aminosäuren mit hydrophilen Resten – Gis E 7(Gis 64) und
 GIS F 8
GIS F 8
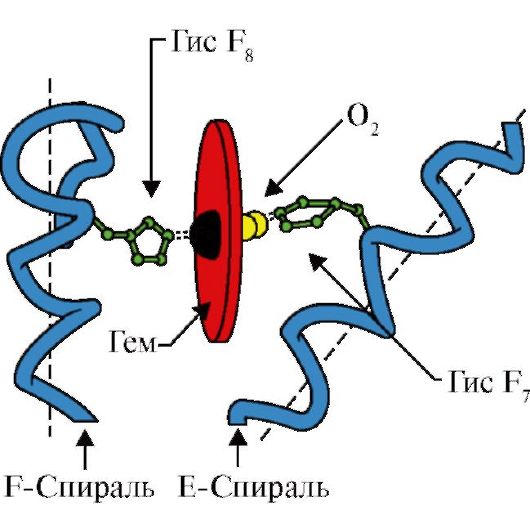 (His 93) (Abb. 1.21).
(His 93) (Abb. 1.21).
Reis. 1,20. Die Struktur von Häm – dem Nicht-Protein-Teil von Myoglobin und Hämoglobin
Gis E 7 Reis. 1.21. Lage von Häm und O2 im aktiven Zentrum von Apomyoglobin und Hämoglobin-Protomeren Häm ist ein spezifischer Ligand von apoMB (Abb. 1.20), dessen Basis aus vier durch Methenylbrücken verbundenen Pyrrolringen besteht; Häm ist über ein Eisenatom kovalent an His F8 gebunden. O 2 bindet sich an das Eisen auf der anderen Seite der Hämebene. Sein E 7 ist für die korrekte Ausrichtung von O 2 notwendig und erleichtert die Anlagerung von Sauerstoff an Fe + 2-Häm
bildet eine Koordinationsbindung mit Fe+ 2 und fixiert Häm fest im aktiven Zentrum.
notwendig für die korrekte Orientierung im aktiven Zentrum eines anderen Liganden – O 2 während seiner Wechselwirkung mit Fe + 2 Häm. Die Mikroumgebung von Häm schafft Bedingungen für eine starke, aber reversible Bindung von O 2 an Fe + 2 und verhindert, dass Wasser in das hydrophobe aktive Zentrum eindringt, was zu seiner Oxidation zu Fe + 3 führen kann. Die Monomerstruktur von Mb und seinem aktiven Zentrum bestimmt die hohe Affinität des Proteins für O 2. 4. Oligomerstruktur von Hb und Regulierung der Affinität von Hb zu O 2 -Liganden. Menschliche Hämoglobine- eine Familie von Proteinen wie Myoglobin, die mit komplexen Proteinen (Hämoproteinen) verwandt sind. Sie haben eine tetramere Struktur und enthalten zwei α-Ketten, unterscheiden sich jedoch in der Struktur der beiden anderen Polypeptidketten (2α-, 2x-Ketten). Die Struktur der zweiten Polypeptidkette bestimmt die Funktionsmerkmale dieser Hb-Formen. Etwa 98 % des Hämoglobins in den roten Blutkörperchen eines Erwachsenen besteht aus
Hämoglobin A (2α-, 2p-Ketten). Während der fetalen Entwicklung funktionieren zwei Haupttypen von Hämoglobinen: embryonales Hb(2α, 2ε), das in den frühen Stadien der fetalen Entwicklung vorkommt, und
HB A ist ein mit Myoglobin (MB) verwandtes Protein, das in erwachsenen menschlichen roten Blutkörperchen vorkommt. Die Struktur seiner einzelnen Protomere ähnelt der von Myoglobin. Die Sekundär- und Tertiärstrukturen von Myoglobin und Hämoglobin-Protomeren sind sehr ähnlich, trotz der Tatsache, dass in der Primärstruktur ihrer Polypeptidketten nur 24 Aminosäurereste identisch sind (die Sekundärstruktur von Hämoglobin-Protomeren enthält wie Myoglobin acht α-Helices, bezeichnet mit lateinischen Buchstaben von A bis H, und die Tertiärstruktur hat die Form einer kompakten Kugel). Aber im Gegensatz zu Myoglobin hat Hämoglobin eine oligomere Struktur, die aus vier Polypeptidketten besteht, die durch nichtkovalente Bindungen verbunden sind (Abbildung 1.22).
Jedes Hb-Protomer ist mit einem Nicht-Protein-Teil verbunden – Häm und benachbarten Protomeren. Die Verbindung des Proteinteils von Hb mit Häm ähnelt der von Myoglobin: Im aktiven Zentrum des Proteins sind die hydrophoben Teile des Häms mit Ausnahme von His F 8 und His E 7 von hydrophoben Aminosäureresten umgeben. die sich auf beiden Seiten der Hämebene befinden und eine ähnliche Rolle bei der Funktion des Proteins und seiner Bindung an Sauerstoff spielen (siehe Struktur von Myoglobin).
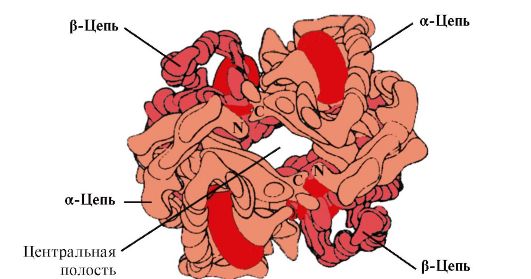 Reis. 1.22. Oligomerstruktur von Hämoglobin
Reis. 1.22. Oligomerstruktur von Hämoglobin
Außerdem, Häm ist ein spezifischer Ligand von apoMB (Abb. 1.20), dessen Basis aus vier durch Methenylbrücken verbundenen Pyrrolringen besteht; erfüllt eine wichtige Aufgabe zusätzliche Rolle in der Funktionsweise von Nv. Freies Häm hat eine 25.000-mal höhere Affinität zu CO als zu O2.
5. CO wird im Körper in geringen Mengen gebildet und könnte aufgrund seiner hohen Affinität zu Häm den für das Zellleben notwendigen O 2 -Transport stören. Allerdings übersteigt in der Zusammensetzung des Hämoglobins die Affinität von Häm zu Kohlenmonoxid die Affinität zu O 2 aufgrund der Anwesenheit von His E 7 im aktiven Zentrum nur um das 200-fache. Der Rest dieser Aminosäure schafft optimale Bedingungen für die Bindung von Häm an O 2 und schwächt die Wechselwirkung von Häm mit CO. Die Hauptfunktion von HB ist der Transport von O2 von der Lunge zum Gewebe.
Im Gegensatz zu monomerem Myoglobin, das eine sehr hohe Affinität zu O2 hat und die Funktion der Sauerstoffspeicherung in roten Muskeln übernimmt, sorgt die oligomere Struktur von Hämoglobin für Folgendes:
1) schnelle Sättigung von HB mit Sauerstoff in der Lunge;
2) die Fähigkeit von HB, Sauerstoff in Geweben bei einem relativ hohen O 2 -Partialdruck (20–40 mm Hg) freizusetzen;
6. Kooperative Veränderungen in der Konformation von Hämoglobin-Protomeren beschleunigen die Bindung von O 2 in der Lunge und seine Freisetzung in Gewebe. In der Lunge fördert der hohe Partialdruck von O 2 dessen Bindung an Hb im aktiven Zentrum von vier Protomeren (2α und 2β). Das aktive Zentrum jedes Protomers befindet sich wie bei Myoglobin zwischen zwei α-Helices (F und E) in einer hydrophoben Tasche. Es enthält einen Nicht-Protein-Teil – Häm, der durch viele schwache hydrophobe Wechselwirkungen und eine starke Bindung zwischen Fe 2 + Häm und His F 8 an den Proteinteil gebunden ist (siehe Abb. 1.21).
Im Desoxyhämoglobin ragt aufgrund dieser Bindung mit His F 8 das Fe 2 + -Atom aus der Hämebene in Richtung Histidin hervor. Die Bindung von O 2 an Fe 2 + erfolgt auf der anderen Seite des Häms in der His-E 7-Region über eine einzelne freie Koordinationsbindung. Sein E 7 bietet optimale Bedingungen für die Bindung von O 2 an Hämeisen.
Die Addition von O 2 an das Fe + 2-Atom eines Protomers bewirkt dessen Bewegung in die Hämebene, gefolgt von dem damit verbundenen Histidinrest
 Reis. 1.23. Änderung der Konformation des Hämoglobin-Protomers in Kombination mit O 2
Reis. 1.23. Änderung der Konformation des Hämoglobin-Protomers in Kombination mit O 2
Dies führt aufgrund ihrer Konformationslabilität zu einer Konformationsänderung aller Polypeptidketten. Die Änderung der Konformation anderer Ketten erleichtert deren Wechselwirkung mit nachfolgenden O 2 -Molekülen.
Das vierte O 2 -Molekül bindet 300-mal leichter an Hämoglobin als das erste (Abb. 1.24).
 Reis. 1.24. Kooperative Änderungen in der Konformation von Hämoglobin-Protomeren während ihrer Wechselwirkung mit O2
Reis. 1.24. Kooperative Änderungen in der Konformation von Hämoglobin-Protomeren während ihrer Wechselwirkung mit O2
In Geweben wird jedes nachfolgende O 2 -Molekül leichter abgespalten als das vorherige, auch aufgrund kooperativer Änderungen in der Konformation von Protomeren.
7. CO 2 und H+, die beim Abbau organischer Substanzen entstehen, verringern proportional zu ihrer Konzentration die Affinität von Hämoglobin zu O 2. Die für die Zellfunktion erforderliche Energie wird hauptsächlich in Mitochondrien bei der Oxidation organischer Substanzen mithilfe von O 2 erzeugt, das von Hämoglobin aus der Lunge geliefert wird. Durch die Oxidation organischer Stoffe entstehen die Endprodukte ihrer Zersetzung: CO 2 und K 2 O, deren Menge proportional zur Intensität der laufenden Oxidationsprozesse ist.
CO 2 diffundiert aus den Zellen ins Blut und dringt in die roten Blutkörperchen ein, wo es unter der Wirkung des Enzyms Carbanhydrase in Kohlensäure umgewandelt wird. Diese schwache Säure zerfällt in ein Proton und ein Bicarbonation.
H+ ist in der Lage, sich seinen Radikalen anzuschließen 14 6 in den α- und β-Ketten des Hämoglobins, d.h. in vom Häm entfernten Gebieten. Die Protonierung von Hämoglobin verringert seine Affinität zu O 2, fördert die Entfernung von O 2 aus OxyHb, die Bildung von DesoxyHb und erhöht die Sauerstoffversorgung des Gewebes proportional zur Anzahl der gebildeten Protonen (Abb. 1.25).
Eine Erhöhung der Menge an freigesetztem Sauerstoff in Abhängigkeit von der Erhöhung der H+-Konzentration in den roten Blutkörperchen wird als Bohr-Effekt bezeichnet (benannt nach dem dänischen Physiologen Christian Bohr, der diesen Effekt als Erster entdeckte).
In der Lunge fördert ein hoher Sauerstoffpartialdruck dessen Bindung an DesoxyHb, wodurch die Affinität des Proteins zu H+ verringert wird. Die unter Einwirkung von Kohlensäure freigesetzten Protonen reagieren mit Bicarbonaten zu CO 2 und H 2 O
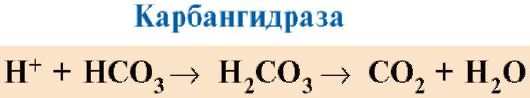
 Reis. 1,25. Abhängigkeit der Affinität von Hb zu O 2 von der Konzentration von CO 2 und Protonen (Bohr-Effekt):
Reis. 1,25. Abhängigkeit der Affinität von Hb zu O 2 von der Konzentration von CO 2 und Protonen (Bohr-Effekt):
A- Einfluss der CO 2- und H+-Konzentration auf die Freisetzung von O 2 aus dem Komplex mit HB (Bohr-Effekt); B- Sauerstoffanreicherung von Desoxyhämoglobin in der Lunge, Bildung und Freisetzung von CO 2.
Das entstehende CO 2 gelangt in den Alveolarraum und wird mit der Ausatemluft entfernt. So wird die vom Hämoglobin im Gewebe freigesetzte Sauerstoffmenge durch die Abbauprodukte organischer Stoffe reguliert: Je intensiver der Stoffabbau, beispielsweise bei körperlicher Anstrengung, desto höher die Konzentration von CO 2 und H+ und desto mehr Sauerstoff die Gewebe erhalten als Folge einer Abnahme der Affinität von Hb zu O 2.
8. Allosterische Regulierung der Affinität von Hb zu O2 durch den Liganden – 2,3-Bisphosphoglycerat. In Erythrozyten wird der allosterische Ligand des Hämoglobins, 2,3-Bisphosphoglycerat (2,3-BPG), aus dem Produkt der Glucoseoxidation – 1,3-Bisphosphoglycerat – synthetisiert. Unter normalen Bedingungen ist die Konzentration von 2,3-BPG hoch und vergleichbar mit der Konzentration von Hb. 2,3-BPG hat eine starke negative Ladung von -5.
 Bisphosphoglycerat in Gewebekapillaren erhöht durch die Bindung an Desoxyhämoglobin die Freisetzung von Sauerstoff in das Gewebe und verringert die Affinität von Hb zu O 2.
Bisphosphoglycerat in Gewebekapillaren erhöht durch die Bindung an Desoxyhämoglobin die Freisetzung von Sauerstoff in das Gewebe und verringert die Affinität von Hb zu O 2.
Im Zentrum des tetrameren Hämoglobinmoleküls befindet sich ein Hohlraum. Es wird aus Aminosäureresten aller vier Protomeren gebildet (siehe Abb. 1.22). In Gewebekapillaren führt die Protonierung von Hb (Bohr-Effekt) zum Aufbrechen der Bindung zwischen Häm-Eisen und O2. In einem Molekül
Desoxyhämoglobins treten im Vergleich zu Oxyhämoglobin zusätzliche ionische Bindungen auf, die die Protomere verbinden, wodurch die Abmessungen des zentralen Hohlraums im Vergleich zu Oxyhämoglobin zunehmen. Der zentrale Hohlraum ist der Ort der Bindung von 2,3-BPG an Hämoglobin. Aufgrund der unterschiedlichen Größe des zentralen Hohlraums kann sich 2,3-BPG nur an Desoxyhämoglobin binden.
2,3-BPG interagiert mit Hämoglobin an einer Stelle, die von den aktiven Zentren des Proteins entfernt liegt und dazu gehört allosterisch(regulatorische) Liganden und der zentrale Hohlraum von Hb ist allosterisches Zentrum. 2,3-BPG hat eine starke negative Ladung und interagiert mit fünf positiv geladenen Gruppen der beiden β-Ketten von Hb: der N-terminalen α-Aminogruppe von Val und den Lys 82 His 143-Radikalen (Abb. 1.26).
 Reis. 1.26. BPG in der zentralen Höhle des Desoxyhämoglobins
Reis. 1.26. BPG in der zentralen Höhle des Desoxyhämoglobins
BPG bindet an drei positiv geladene Gruppen an jedem β-Strang.
In Gewebekapillaren interagiert das entstehende Desoxyhämoglobin mit 2,3-BPG und es bilden sich ionische Bindungen zwischen den positiv geladenen Radikalen der β-Ketten und dem negativ geladenen Liganden, die die Konformation des Proteins verändern und die Affinität von Hb zu O2 verringern . Eine Verringerung der Affinität von Hb zu O 2 trägt zu einer effizienteren Freisetzung von O 2 in das Gewebe bei.
In der Lunge interagiert Sauerstoff bei hohem Partialdruck mit Hb und verbindet sich mit Häm-Eisen; in diesem Fall ändert sich die Konformation des Proteins, der zentrale Hohlraum verkleinert sich und 2,3-BPG wird aus dem allosterischen Zentrum verdrängt
Somit weisen oligomere Proteine im Vergleich zu monomeren Proteinen neue Eigenschaften auf. Bindung von Liganden an Stellen
räumlich voneinander entfernt (allosterisch) können Konformationsänderungen im gesamten Proteinmolekül verursachen. Durch die Wechselwirkung mit regulatorischen Liganden kommt es zu einer Konformationsänderung und Anpassung der Funktion des Proteinmoleküls an Umweltveränderungen.
THEMA 1.5. Aufrechterhaltung der nativen Konformation von Proteinen unter Zellbedingungen
In Zellen während der Synthese von Polypeptidketten, ihrem Transport durch Membranen zu den entsprechenden Teilen der Zelle, während des Faltungsprozesses (Bildung der nativen Konformation) und während des Zusammenbaus oligomerer Proteine sowie während ihrer Funktion als Zwischenprodukt Aggregationsanfällige, instabile Konformationen in der Proteinstruktur entstehen. Hydrophobe Radikale, die normalerweise in der nativen Konformation im Proteinmolekül verborgen sind, erscheinen auf der Oberfläche in einer instabilen Konformation und neigen dazu, sich mit Gruppen anderer Proteine zu verbinden, die in Wasser schlecht löslich sind. In den Zellen aller bekannten Organismen wurden spezielle Proteine gefunden, die für eine optimale Faltung der Zellproteine sorgen, ihre native Konformation während der Funktion stabilisieren und vor allem die Struktur und Funktionen intrazellulärer Proteine aufrechterhalten, wenn die Homöostase gestört ist. Diese Proteine heißen „Begleiter“ was auf Französisch „Kindermädchen“ bedeutet.
1. Molekulare Chaperone und ihre Rolle bei der Verhinderung der Denaturierung von Proteinen.
Chaperone (CH) werden nach der Masse ihrer Untereinheiten klassifiziert. Chaperone mit hohem Molekulargewicht haben eine Masse von 60 bis 110 kDa. Unter ihnen wurden drei Klassen am meisten untersucht: Sh-60, Sh-70 und Sh-90. Jede Klasse umfasst eine Familie verwandter Proteine. Somit enthält Sh-70 Proteine mit einem Molekulargewicht von 66 bis 78 kDa. Chaperone mit niedrigem Molekulargewicht haben ein Molekulargewicht von 40 bis 15 kDa.
Unter den Begleitpersonen gibt es konstitutiv Proteine, deren hohe Grundsynthese nicht von Stresseinwirkungen auf die Körperzellen abhängt, und induzierbar, dessen Synthese unter normalen Bedingungen schwach ist, unter Stress jedoch stark ansteigt. Induzierbare Chaperone werden auch „Hitzeschockproteine“ genannt, weil sie erstmals in Zellen entdeckt wurden, die hohen Temperaturen ausgesetzt waren. In Zellen ist aufgrund der hohen Proteinkonzentration eine spontane Reaktivierung teilweise denaturierter Proteine schwierig. Sh-70 kann den Beginn der Denaturierung verhindern und dabei helfen, die native Konformation von Proteinen wiederherzustellen. Molekulare Chaperone-70- eine hochkonservierte Klasse von Proteinen, die in allen Teilen der Zelle vorkommt: Zytoplasma, Zellkern, endoplasmatisches Retikulum, Mitochondrien. Am Carboxylende der einzelnen Polypeptidkette Sh-70 befindet sich eine Region, die eine Furche darstellt, die mit Längenpeptiden interagieren kann
7 bis 9 Aminosäurereste, angereichert mit hydrophoben Resten. Solche Regionen kommen in globulären Proteinen etwa alle 16 Aminosäuren vor. Sh-70 ist in der Lage, Proteine vor der Inaktivierung durch Temperatur zu schützen und die Konformation und Aktivität teilweise denaturierter Proteine wiederherzustellen.
2. Die Rolle von Chaperonen bei der Proteinfaltung. Bei der Proteinsynthese am Ribosom wird die N-terminale Region des Polypeptids vor der C-terminalen synthetisiert. Zur Bildung der nativen Konformation ist die vollständige Aminosäuresequenz des Proteins erforderlich. Im Prozess der Proteinsynthese sind Chaperone-70 aufgrund der Struktur ihres aktiven Zentrums in der Lage, zur Aggregation neigende Bereiche des Polypeptids zu schließen, angereichert mit hydrophoben Aminosäureresten, bis die Synthese abgeschlossen ist (Abbildung 1.27, A ).
 Reis. 1.27. Beteiligung von Chaperonen an der Proteinfaltung
Reis. 1.27. Beteiligung von Chaperonen an der Proteinfaltung
A – Beteiligung von Chaperonen-70 an der Verhinderung hydrophober Wechselwirkungen zwischen Abschnitten des synthetisierten Polypeptids; B – Bildung der nativen Konformation des Proteins im Chaperonkomplex
Viele hochmolekulare Proteine, die eine komplexe Konformation, beispielsweise eine Domänenstruktur, aufweisen, falten sich in einem speziellen, durch Sh-60 gebildeten Raum. Ø-60 fungieren als oligomerer Komplex bestehend aus 14 Untereinheiten. Sie bilden zwei Hohlringe, die jeweils aus sieben Untereinheiten bestehen, diese Ringe sind miteinander verbunden. Jede Sh-60-Untereinheit besteht aus drei Domänen: apikal (apikal), angereichert mit hydrophoben Radikalen, die dem Hohlraum des Rings zugewandt sind, intermediär und äquatorial (Abb. 1.28).
 Reis. 1.28. Struktur des Chaperonin-Komplexes bestehend aus 14 Ш-60
Reis. 1.28. Struktur des Chaperonin-Komplexes bestehend aus 14 Ш-60
A – Seitenansicht; B – Draufsicht
In den Hohlraum der Chaperonringe gelangen synthetisierte Proteine, die auf der Oberfläche für ungefaltete Moleküle charakteristische Elemente, insbesondere hydrophobe Radikale, aufweisen. In der spezifischen Umgebung dieser Hohlräume werden mögliche Konformationen aufgezählt, bis die einzige gefunden wird, die energetisch am günstigsten ist (Abb. 1.27, B). Die Bildung von Konformationen und die Proteinfreisetzung gehen mit einer ATP-Hydrolyse in der Äquatorregion einher. Typischerweise erfordert eine solche Chaperon-abhängige Faltung einen erheblichen Energieaufwand.
Chaperone sind nicht nur an der Bildung der dreidimensionalen Struktur von Proteinen und der Renativierung teilweise denaturierter Proteine beteiligt, sondern auch für den Ablauf grundlegender Prozesse wie dem Zusammenbau oligomerer Proteine, der Erkennung und dem Transport denaturierter Proteine in Lysosomen, Transport von Proteinen durch Membranen und Beteiligung an der Regulierung der Aktivität von Proteinkomplexen.
THEMA 1.6. VIELFALT AN PROTEINEN. PROTEINFAMILIEN: BEISPIEL FÜR IMMUNGLOBULINE
1. Proteine spielen eine entscheidende Rolle im Leben einzelner Zellen und des gesamten vielzelligen Organismus und ihre Funktionen sind überraschend vielfältig. Dies wird durch die Eigenschaften der Primärstruktur und Konformation von Proteinen, die einzigartige Struktur des aktiven Zentrums und die Fähigkeit zur Bindung spezifischer Liganden bestimmt.
Nur ein sehr kleiner Teil aller möglichen Varianten von Peptidketten kann eine stabile räumliche Struktur annehmen; mehrheitlich
von ihnen können viele Konformationen mit ungefähr derselben Gibbs-Energie, aber unterschiedlichen Eigenschaften annehmen. Die durch die biologische Evolution ausgewählte Primärstruktur der meisten bekannten Proteine gewährleistet eine außergewöhnliche Stabilität einer der Konformationen, die die Funktionsmerkmale dieses Proteins bestimmt.
2. Proteinfamilien. Innerhalb derselben biologischen Spezies können Substitutionen von Aminosäureresten zur Entstehung verschiedener Proteine führen, die verwandte Funktionen erfüllen und homologe Aminosäuresequenzen aufweisen. Solche verwandten Proteine weisen auffallend ähnliche Konformationen auf: Die Anzahl und relative Position der α-Helices und/oder β-Strukturen, die meisten Windungen und Biegungen der Polypeptidketten sind ähnlich oder identisch. Proteine mit homologen Regionen der Polypeptidkette, ähnlicher Konformation und verwandten Funktionen werden in Proteinfamilien eingeteilt. Beispiele für Proteinfamilien: Serinproteinasen, Immunglobulinfamilie, Myoglobinfamilie.
Serinproteinasen- eine Familie von Proteinen, die die Funktion proteolytischer Enzyme erfüllen. Dazu gehören Verdauungsenzyme – Chymotrypsin, Trypsin, Elastase und viele Blutgerinnungsfaktoren. Diese Proteine haben in 40 % ihrer Positionen identische Aminosäuren und eine sehr ähnliche Konformation (Abb. 1.29).

Reis. 1.29. Räumliche Strukturen von Elastase (A) und Chymotrypsin (B)
Einige Aminosäuresubstitutionen haben zu Veränderungen in der Substratspezifität dieser Proteine und zur Entstehung einer funktionellen Diversität innerhalb der Familie geführt.
3. Immunglobulin-Familie. Proteine der Immunglobulin-Superfamilie, zu der drei Proteinfamilien gehören, spielen eine große Rolle für die Funktion des Immunsystems:
Antikörper (Immunglobuline);
T-Lymphozyten-Rezeptoren;
Proteine des Haupthistokompatibilitätskomplexes – MHC-Klassen 1 und 2 (Major Histocompatibility Complex).
Alle diese Proteine haben eine Domänenstruktur, bestehen aus homologen immunähnlichen Domänen und erfüllen ähnliche Funktionen: Sie interagieren mit fremden Strukturen, die entweder im Blut, in der Lymphe oder in der Interzellularflüssigkeit gelöst sind (Antikörper) oder sich auf der Oberfläche von Zellen befinden (eigene bzw ausländisch).
4. Antikörper- spezifische Proteine, die von B-Lymphozyten als Reaktion auf das Eindringen einer Fremdstruktur in den Körper produziert werden, genannt Antigen.
Merkmale der Struktur von Antikörpern
Die einfachsten Antikörpermoleküle bestehen aus vier Polypeptidketten: zwei identischen leichten – L, die etwa 220 Aminosäuren enthalten, und zwei identischen schweren – H, bestehend aus 440–700 Aminosäuren. Alle vier Ketten im Antikörpermolekül sind durch viele nichtkovalente Bindungen und vier Disulfidbindungen verbunden (Abb. 1.30).
Leichte Antikörperketten bestehen aus zwei Domänen: einer variablen Domäne (VL), die sich im N-terminalen Bereich der Polypeptidkette befindet, und einer konstanten Domäne (CL), die sich am C-Terminus befindet. Schwere Ketten haben normalerweise vier Domänen: eine variable (VH), die sich am N-Terminus befindet, und drei konstante Domänen (CH1, CH2, CH3) (siehe Abb. 1.30). Jede Immunglobulindomäne verfügt über eine β-Faltblatt-Überstruktur, in der zwei Cysteinreste durch eine Disulfidbindung verbunden sind.
Zwischen den beiden konstanten Domänen CH1 und CH2 befindet sich eine Region mit einer Vielzahl von Prolinresten, die die Bildung einer Sekundärstruktur und die Interaktion benachbarter H-Ketten in diesem Segment verhindern. Diese Gelenkregion verleiht dem Antikörpermolekül Flexibilität. Zwischen den variablen Domänen der schweren und leichten Ketten gibt es zwei identische Antigen-Bindungsstellen (aktive Stellen zur Bindung von Antigenen), daher werden solche Antikörper oft als Antikörper bezeichnet Bivalente. An der Bindung des Antigens an den Antikörper ist nicht die gesamte Aminosäuresequenz der variablen Regionen beider Ketten beteiligt, sondern nur 20–30 Aminosäuren, die sich in den hypervariablen Regionen jeder Kette befinden. Es sind diese Regionen, die die einzigartige Fähigkeit jedes Antikörpertyps bestimmen, mit dem entsprechenden komplementären Antigen zu interagieren.
Antikörper sind eine der Verteidigungslinien des Körpers gegen eindringende Fremdorganismen. Ihre Funktionsweise kann in zwei Phasen unterteilt werden: Die erste Phase ist die Erkennung und Bindung von Antigenen auf der Oberfläche fremder Organismen, was aufgrund des Vorhandenseins von Antigen-Bindungsstellen in der Antikörperstruktur möglich ist; Die zweite Stufe ist die Einleitung des Prozesses der Inaktivierung und Zerstörung des Antigens. Die Spezifität der zweiten Stufe hängt von der Klasse der Antikörper ab. Es gibt fünf Klassen schwerer Ketten, die sich in der Struktur konstanter Domänen voneinander unterscheiden: α, δ, ε, γ und μ, nach denen fünf Klassen von Immunglobulinen unterschieden werden: A, D, E, G und M.
Die Strukturmerkmale der schweren Ketten verleihen den Gelenkregionen und C-terminalen Regionen der schweren Ketten eine für jede Klasse charakteristische Konformation. Nachdem das Antigen an einen Antikörper gebunden ist, bestimmen Konformationsänderungen in den konstanten Domänen den Weg zur Antigenentfernung.
 Reis. 1. 30. Domänenstruktur von IgG
Reis. 1. 30. Domänenstruktur von IgG
Immunglobuline M
Immunglobuline M gibt es in zwei Formen.
Monomere Form- 1. Klasse von Antikörpern, die von sich entwickelnden B-Lymphozyten produziert werden. Anschließend stellen viele B-Zellen auf die Produktion anderer Antikörperklassen um, jedoch mit derselben Antigenbindungsstelle. IgM ist in die Membran eingebettet und fungiert als Antigenerkennungsrezeptor. Die Integration von IgM in die Zellmembran ist aufgrund des Vorhandenseins von 25 hydrophoben Aminosäureresten im Schwanzteil der Region möglich.
Sekretorische Form von IgM enthält fünf durch Disulfidbrücken miteinander verbundene Monomeruntereinheiten und eine zusätzliche Polypeptid-J-Kette (Abb. 1.31). Die schweren Ketten von Monomeren dieser Form enthalten keinen hydrophoben Schwanz. Das Pentamer verfügt über 10 Antigenbindungsstellen und ist daher wirksam bei der Erkennung und Entfernung des Antigens, das zuerst in den Körper gelangt. Die sekretorische Form von IgM ist die Hauptklasse von Antikörpern, die während der primären Immunantwort ins Blut sezerniert werden. Die Bindung von IgM an ein Antigen verändert die Konformation von IgM und induziert seine Bindung an die erste Proteinkomponente des Komplementsystems (das Komplementsystem ist eine Reihe von Proteinen, die an der Zerstörung des Antigens beteiligt sind) und die Aktivierung dieses Systems. Befindet sich das Antigen auf der Oberfläche eines Mikroorganismus, führt das Komplementsystem zu einer Störung der Integrität der Zellmembran und zum Tod der Bakterienzelle.
Immunglobuline G
Quantitativ überwiegt diese Klasse von Immunglobulinen im Blut (75 % aller Igs). IgG – Monomere, die Hauptklasse von Antikörpern, die während einer sekundären Immunantwort ins Blut ausgeschieden werden. Nach der Interaktion von IgG mit den Oberflächenantigenen von Mikroorganismen ist der Antigen-Antikörper-Komplex in der Lage, Proteine des Komplementsystems zu binden und zu aktivieren oder mit spezifischen Rezeptoren von Makrophagen und Neutrophilen zu interagieren. Interaktion mit Phagozyten führt
 Reis. 1.31. Struktur der sekretorischen Form von IgM
Reis. 1.31. Struktur der sekretorischen Form von IgM
zur Aufnahme von Antigen-Antikörper-Komplexen und deren Zerstörung in Zellphagosomen. IgG ist die einzige Klasse von Antikörpern, die die Plazentaschranke durchdringen und den Fötus intrauterin vor Infektionen schützen können.
Immunglobuline A
Die Hauptklasse der Antikörper, die in Sekreten (Milch, Speichel, Sekrete der Atemwege und des Darmtrakts) vorkommen. IgA wird hauptsächlich in dimerer Form sezerniert, wobei die Monomere über eine zusätzliche J-Kette miteinander verbunden sind (Abb. 1.32).
IgA interagiert nicht mit dem Komplementsystem und phagozytischen Zellen, aber durch die Bindung an Mikroorganismen verhindern Antikörper deren Anlagerung an Epithelzellen und das Eindringen in den Körper.
Immunglobuline E
Immunglobuline E werden durch Monomere dargestellt, in denen die schweren ε-Ketten, wie die μ-Ketten der Immunglobuline M, eine variable und vier konstante Domänen enthalten. Nach der Sekretion bindet IgE daran
 Reis. 1.32. Struktur von IgA
Reis. 1.32. Struktur von IgA
C-terminale Regionen mit entsprechenden Rezeptoren auf der Oberfläche von Mastzellen und Basophilen. Dadurch werden sie zu Rezeptoren für Antigene auf der Oberfläche dieser Zellen (Abb. 1.33).
 Reis. 1.33. Wechselwirkung von IgE mit Antigen auf der Oberfläche einer Mastzelle
Reis. 1.33. Wechselwirkung von IgE mit Antigen auf der Oberfläche einer Mastzelle
Nach der Bindung des Antigens an die entsprechenden Antigenbindungsstellen des IgE erhalten die Zellen das Signal, biologisch aktive Substanzen (Histamin, Serotonin) abzusondern, die maßgeblich für die Entstehung der Entzündungsreaktion und für die Manifestation allergischer Reaktionen wie z Asthma, Urtikaria, Heuschnupfen.
Immunglobuline D
Immunglobuline D kommen in sehr geringen Mengen im Serum vor; sie sind Monomere. Schwere δ-Ketten haben eine variable und drei konstante Domänen. IgDs fungieren als Rezeptoren für B-Lymphozyten; andere Funktionen sind noch unbekannt. Die Interaktion spezifischer Antigene mit Rezeptoren auf der Oberfläche von B-Lymphozyten (IgD) führt zur Übertragung dieser Signale in die Zelle und zur Aktivierung von Mechanismen, die die Proliferation eines bestimmten Lymphozytenklons sicherstellen.
THEMA 1.7. PHYSIKALISCHE UND CHEMISCHE EIGENSCHAFTEN VON PROTEINEN UND METHODEN ZU IHRER TRENNUNG
1. Einzelne Proteine unterscheiden sich in ihren physikalischen und chemischen Eigenschaften:
Form von Molekülen;
Molekulargewicht;
Die Gesamtladung, deren Größe vom Verhältnis der anionischen und kationischen Gruppen der Aminosäuren abhängt;
Das Verhältnis polarer und unpolarer Aminosäurereste auf der Oberfläche von Molekülen;
Widerstandsgrade gegenüber verschiedenen Denaturierungsmitteln.
2. Die Proteinlöslichkeit hängt davon ab von den Eigenschaften der oben aufgeführten Proteine sowie von der Zusammensetzung des Mediums, in dem das Protein gelöst ist (pH-Werte, Salzzusammensetzung, Temperatur, Vorhandensein anderer organischer Substanzen, die mit dem Protein interagieren können). Die Ladungsmenge von Proteinmolekülen ist einer der Faktoren, die ihre Löslichkeit beeinflussen. Wenn die Ladung am isoelektrischen Punkt verloren geht, können Proteine leichter aggregieren und ausfallen. Dies ist besonders typisch für denaturierte Proteine, bei denen hydrophobe Aminosäurereste auf der Oberfläche auftreten.
Auf der Oberfläche eines Proteinmoleküls befinden sich sowohl positiv als auch negativ geladene Aminosäurereste. Die Anzahl dieser Gruppen und damit die Gesamtladung der Proteine hängt vom pH-Wert des Mediums ab, d. h. Verhältnis der Konzentrationen von H+- und OH-Gruppen. In einer sauren Umgebung Eine Erhöhung der H+-Konzentration führt zu einer Unterdrückung der Dissoziation der Carboxylgruppen -COO - + H+ > - COOH und einer Verringerung der negativen Ladung von Proteinen. In einer alkalischen Umgebung führt die Bindung von überschüssigem OH - durch Protonen, die bei der Dissoziation der Aminogruppen -NH 3 + + OH - - NH 2 + H 2 O unter Bildung von Wasser entstehen, zu einer Abnahme der positiven Ladung von Proteinen . Als pH-Wert bezeichnet man den pH-Wert, bei dem ein Protein eine Netto-Nullladung aufweist isoelektrischer Punkt (IEP). Bei IET ist die Anzahl der positiv und negativ geladenen Gruppen gleich, d. h. Das Protein befindet sich in einem isoelektrischen Zustand.
3. Trennung einzelner Proteine. Merkmale der Struktur und Funktion des Körpers hängen von der Menge der darin synthetisierten Proteine ab. Die Untersuchung der Struktur und Eigenschaften von Proteinen ist unmöglich, ohne sie aus der Zelle zu isolieren und von anderen Proteinen und organischen Molekülen zu reinigen. Stufen der Isolierung und Reinigung einzelner Proteine:
Zellzerstörung das zu untersuchende Gewebe und die Gewinnung eines Homogenats.
Trennung des Homogenats in Fraktionen Durch Zentrifugieren erhält man eine nukleare, mitochondriale, zytosolische oder andere Fraktion, die das gewünschte Protein enthält.
Selektive thermische Denaturierung- kurzzeitiges Erhitzen einer Proteinlösung, bei dem ein Teil der denaturierten Proteinverunreinigungen entfernt werden kann (sofern das Protein relativ hitzestabil ist).
Aussalzen. Verschiedene Proteine fallen bei unterschiedlichen Salzkonzentrationen in Lösung aus. Durch schrittweise Erhöhung der Salzkonzentration ist es möglich, mehrere separate Fraktionen zu erhalten, wobei eine davon den überwiegenden Anteil an isoliertem Protein aufweist.
Ammoniumsulfat wird am häufigsten zur Proteinfraktionierung verwendet. Proteine mit der geringsten Löslichkeit fallen bei niedrigen Salzkonzentrationen aus. Gelfiltration
- eine Methode zum Sieben von Molekülen durch gequollene Sephadex-Körner (dreidimensionale Polysaccharidketten von Dextran mit Poren). Die Geschwindigkeit, mit der Proteine eine mit Sephadex gefüllte Säule passieren, hängt von ihrem Molekulargewicht ab: Je kleiner die Masse der Proteinmoleküle, desto leichter dringen sie in das Granulat ein und bleiben dort länger. Je größer die Masse, desto schneller eluieren sie aus dem Granulat Spalte. Ultrazentrifugation
- eine Methode, bei der Proteine in einem Zentrifugenröhrchen in den Rotor einer Ultrazentrifuge gegeben werden. Wenn sich der Rotor dreht, ist die Sedimentationsrate der Proteine proportional zu ihrem Molekulargewicht: Fraktionen schwererer Proteine befinden sich näher am Boden des Reagenzglases, leichtere – näher an der Oberfläche. Elektrophorese
- eine Methode, die auf Unterschieden in der Bewegungsgeschwindigkeit von Proteinen in einem elektrischen Feld basiert. Dieser Wert ist proportional zur Ladung der Proteine. Die Elektrophorese von Proteinen wird auf Papier (in diesem Fall ist die Geschwindigkeit der Proteinbewegung nur proportional zu ihrer Ladung) oder in einem Polyacrylamidgel mit einer bestimmten Porengröße (die Geschwindigkeit der Proteinbewegung ist proportional zu ihrer Ladung und ihrem Molekulargewicht) durchgeführt. .- eine Fraktionierungsmethode, die auf der Bindung ionisierter Proteingruppen mit entgegengesetzt geladenen Gruppen von Ionenaustauscherharzen (unlöslichen Polymermaterialien) basiert. Die Stärke der Bindung des Proteins an das Harz ist proportional zur Ladung des Proteins. An das Ionenaustauscherpolymer adsorbierte Proteine können mit steigenden Konzentrationen von NaCl-Lösungen weggewaschen werden; Je niedriger die Proteinladung, desto geringer ist die NaCl-Konzentration, die erforderlich ist, um das an die ionischen Gruppen des Harzes gebundene Protein abzuwaschen.
Affinitätschromatographie- die spezifischste Methode zur Isolierung einzelner Proteine. Ein Ligand eines Proteins wird kovalent an ein inertes Polymer gebunden. Wenn eine Proteinlösung durch eine Säule mit einem Polymer geleitet wird, wird aufgrund der komplementären Bindung des Proteins an den Liganden nur das für einen bestimmten Liganden spezifische Protein an der Säule adsorbiert.
Dialyse- eine Methode zur Entfernung niedermolekularer Verbindungen aus einer Lösung isolierten Proteins. Die Methode basiert auf der Unfähigkeit von Proteinen, im Gegensatz zu Substanzen mit niedrigem Molekulargewicht, eine semipermeable Membran zu passieren. Es wird verwendet, um Proteine nach dem Aussalzen von niedermolekularen Verunreinigungen, beispielsweise Salzen, zu reinigen.
AUFGABEN FÜR AUSSERGEWÖHNLICHE ARBEITEN
1. Füllen Sie die Tabelle aus. 1.4.
Tabelle 1.4. Vergleichende Analyse der Struktur und Funktionen verwandter Proteine – Myoglobin und Hämoglobin
a) Erinnern Sie sich an die Struktur des aktiven Zentrums von Mb und Hb. Welche Rolle spielen hydrophobe Aminosäurereste bei der Bildung der aktiven Zentren dieser Proteine? Beschreiben Sie die Struktur des aktiven Zentrums von Mb und Hb und die Mechanismen der Bindung von Liganden daran. Welche Rolle spielen die Reste His F 8 und His E 7 bei der Funktion des aktiven Zentrums von Mb und Hb?
b) Welche neuen Eigenschaften hat das eng verwandte oligomere Protein Hämoglobin im Vergleich zum monomeren Myoglobin? Erklären Sie die Rolle kooperativer Änderungen in der Konformation von Protomeren im Hämoglobinmolekül, die Wirkung von CO 2 und Protonenkonzentrationen auf die Affinität von Hämoglobin zu Sauerstoff sowie die Rolle von 2,3-BPG bei der allosterischen Regulierung der Hb-Funktion .
2. Charakterisieren Sie molekulare Chaperone und achten Sie dabei auf die Beziehung zwischen ihrer Struktur und Funktion.
3. Welche Proteine werden in Familien eingeteilt? Identifizieren Sie am Beispiel der Immunglobulinfamilie ähnliche Strukturmerkmale und verwandte Funktionen der Proteine dieser Familie.
4. Für biochemische und medizinische Zwecke werden häufig gereinigte Einzelproteine benötigt. Erklären Sie, auf welchen physikalisch-chemischen Eigenschaften von Proteinen die Methoden zu ihrer Trennung und Reinigung basieren.
AUFGABEN ZUR SELBSTKONTROLLE
1. Wähle die richtigen Antworten.
Funktionen von Hämoglobin:
A. Transport von O 2 von der Lunge zum Gewebe B. Transport von H + vom Gewebe zur Lunge
B. Aufrechterhaltung eines konstanten Blut-pH-Werts. D. Transport von CO 2 von der Lunge zum Gewebe
D. Transport von CO 2 vom Gewebe zur Lunge
2. Wähle die richtigen Antworten. Ligandα -protomer Hb ist: A. Häm
B. Sauerstoff
B. CO G. 2,3-BPG
D. β-Protomer
3. Wähle die richtigen Antworten.
Hämoglobin im Gegensatz zu Myoglobin:
A. Hat eine Quartärstruktur
B. Die Sekundärstruktur wird nur durch α-Helices dargestellt
B. Gehört zu komplexen Proteinen
D. Interagiert mit einem allosterischen Liganden. D. Kovalent an Häm gebunden
4. Wähle die richtigen Antworten.
Die Affinität von Hb zu O2 nimmt ab:
A. Wenn ein O2-Molekül hinzugefügt wird. B. Wenn ein O2-Molekül entfernt wird
B. Bei der Interaktion mit 2,3-BPG
D. Bei Bindung an die Protomer H + D. Wenn die Konzentration von 2,3-BPG abnimmt
5. Übereinstimmen.
Typ HB zeichnet sich aus durch:
A. In Desoxyform bildet es fibrilläre Aggregate. B. Enthält zwei α- und zwei δ-Ketten
B. Die vorherrschende Form von Hb in erwachsenen Erythrozyten. D. Enthält Häm mit Fe+ 3 im aktiven Zentrum
D. Enthält zwei α- und zwei γ-Ketten 1. HbA 2.
6. Übereinstimmen.
Hb-Liganden:
A. Bindet im allosterischen Zentrum an Hb
B. Hat eine sehr hohe Affinität zum aktiven Zentrum von Hb
B. Durch die Bindung wird die Affinität von Hb zu O 2 G erhöht. Fe+ 2 wird zu Fe+ 3 oxidiert
D. Bildet eine kovalente Bindung mit hisF8
7. Wähle die richtigen Antworten.
Begleitpersonen:
A. Proteine, die in allen Teilen der Zelle vorhanden sind
B. Die Synthese nimmt unter Stress zu
B. An der Hydrolyse denaturierter Proteine beteiligt sein
D. Beteiligen Sie sich an der Aufrechterhaltung der nativen Konformation von Proteinen
D. Erstellen Sie Organellen, in denen die Konformation von Proteinen gebildet wird
8. Spiel. Immunglobuline:
A. Die sekretorische Form ist pentamer.
B. Klasse Ig, die die Plazentaschranke durchdringt
B. Ig – Mastzellrezeptor
D. Die Hauptklasse von Ig, die in den Sekreten von Epithelzellen vorkommt. D. B-Lymphozyten-Rezeptor, dessen Aktivierung die Zellproliferation gewährleistet
9. Wähle die richtigen Antworten.
Immunglobuline E:
A. Wird von Makrophagen produziert. B. Sie haben schwere ε-Ketten.
B. Eingebettet in die Membran von T-Lymphozyten
D. Wirken als Membranantigenrezeptoren auf Mastzellen und Basophilen
D. Verantwortlich für allergische Reaktionen
10. Wähle die richtigen Antworten.
Die Methode zur Trennung von Proteinen basiert auf Unterschieden in ihrem Molekulargewicht:
A. Gelfiltration
B. Ultrazentrifugation
B. Polyacrylamid-Gelelektrophorese D. Ionenaustauschchromatographie
D. Affinitätschromatographie
11. Wähle die richtige Antwort.
Die Methode zur Trennung von Proteinen basiert auf Unterschieden in ihrer Löslichkeit in Wasser:
A. Gelfiltration B. Aussalzen
B. Ionenaustauschchromatographie D. Affinitätschromatographie
D. Elektrophorese im Polyacrylamidgel
STANDARDS FÜR ANTWORTEN AUF „SELBSTKONTROLLAUFGABEN“
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
GRUNDLEGENDE BEGRIFFE UND KONZEPTE
1. Oligomere Proteine, Protomer, Quartärstruktur von Proteinen
2. Kooperative Änderungen in der Protomerkonformation
3. Bohr-Effekt
4. Allosterische Regulierung von Proteinfunktionen, allosterischem Zentrum und allosterischem Effektor
5. Molekulare Chaperone, Hitzeschockproteine
6. Proteinfamilien (Serinproteasen, Immunglobuline)
7. IgM-, G-, E-, A-Struktur-Funktions-Beziehung
8. Gesamtladung von Proteinen, isoelektrischer Punkt von Proteinen
9. Elektrophorese
10. Aussalzen
11. Gelfiltration
12. Ionenaustauschchromatographie
13. Ultrazentrifugation
14. Affinitätschromatographie
15. Elektrophorese von Blutplasmaproteinen
AUFGABEN FÜR DIE ARBEIT IM KLASSENZIMMER
1. Vergleichen Sie die Abhängigkeit des Sättigungsgrads von Hämoglobin (Hb) und Myoglobin (Mb) mit Sauerstoff von seinem Partialdruck im Gewebe
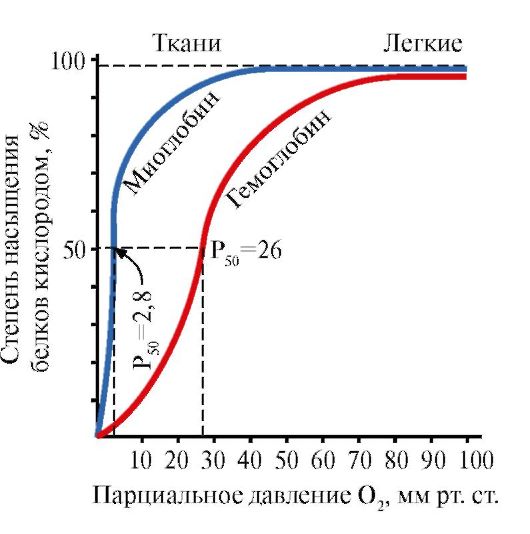 Reis. 1,34. Abhängigkeit der Sättigung Mv undNHSauerstoff aus seinem Partialdruck
Reis. 1,34. Abhängigkeit der Sättigung Mv undNHSauerstoff aus seinem Partialdruck
Bitte beachten Sie, dass die Form der Proteinsauerstoffsättigungskurven unterschiedlich ist: für Myoglobin – eine Hyperbel, für Hämoglobin – eine Sigmoidform.
1. Vergleichen Sie die Werte des Sauerstoffpartialdrucks, bei dem Mb und Hb zu 50 % mit O 2 gesättigt sind. Welches dieser Proteine hat eine höhere Affinität zu O 2?
2. Welche Strukturmerkmale von Mb bestimmen seine hohe Affinität zu O 2?
3. Welche Strukturmerkmale von HB ermöglichen es ihm, O2 in den Kapillaren ruhender Gewebe freizusetzen (bei einem relativ hohen O2-Partialdruck) und diese Freisetzung in arbeitenden Muskeln stark zu steigern? Welche Eigenschaft oligomerer Proteine sorgt für diesen Effekt?
4. Berechnen Sie, wie viel O 2 (in %) sauerstoffhaltiges Hämoglobin ruhenden und arbeitenden Muskeln gibt?
5. Rückschlüsse auf den Zusammenhang zwischen der Struktur eines Proteins und seiner Funktion ziehen.
2. Die vom Hämoglobin in den Kapillaren freigesetzte Sauerstoffmenge hängt von der Intensität der katabolen Prozesse im Gewebe ab (Bohr-Effekt). Wie regulieren Veränderungen im Gewebestoffwechsel die Affinität von Hb zu O2? Einfluss von CO 2 und H+ auf die Affinität von Hb zu O 2
 1. Beschreiben Sie den Bohr-Effekt.
1. Beschreiben Sie den Bohr-Effekt.
2. In welche Richtung verläuft der im Diagramm dargestellte Prozess:
a) in den Kapillaren der Lunge;
b) in Gewebekapillaren?
3. Welche physiologische Bedeutung hat der Bohr-Effekt?
4. Warum verändert die Wechselwirkung von Hb mit H+ an vom Häm entfernten Stellen die Affinität des Proteins für O 2?
3. Die Affinität von Hb zu O2 hängt von der Konzentration seines Liganden – 2,3-Bisphosphoglycerat – ab, der ein allosterischer Regulator der Affinität von Hb zu O2 ist.
Warum beeinflusst die Ligandeninteraktion an einer vom aktiven Zentrum entfernten Stelle die Proteinfunktion? Wie reguliert 2,3-BPG die Affinität von Hb zu O2? Um das Problem zu lösen, beantworten Sie die folgenden Fragen:
1. Wo und woraus wird 2,3-Bisphosphoglycerat (2,3-BPG) synthetisiert? Schreiben Sie die Formel auf und geben Sie die Ladung dieses Moleküls an.
2. Mit welcher Form von Hämoglobin (Oxy oder Desoxy) interagiert BPG und warum? In welchem Teil des Hb-Moleküls findet die Wechselwirkung statt?
3. In welche Richtung verläuft der im Diagramm dargestellte Prozess?
a) in Gewebekapillaren;
b) in den Kapillaren der Lunge?
4. wo die Konzentration des Komplexes höher sein sollte
Nv-2,3-BFG:
a) in den Kapillaren ruhender Muskeln,
b) in den Kapillaren arbeitender Muskeln (bei gleicher BPG-Konzentration in Erythrozyten)?
4. 5. Wie ändert sich die Affinität von HB zu Sauerstoff, wenn sich eine Person an die Bedingungen in großer Höhe anpasst, wenn die Konzentration von BPG in Erythrozyten zunimmt? Welche physiologische Bedeutung hat dieses Phänomen?
5. Die Zerstörung von 2,3-BPG während der Lagerung von Blutkonserven beeinträchtigt die Funktionen von HB. Wie verändert sich die Affinität von HB zu O 2 in konserviertem Blut, wenn die Konzentration von 2,3-BPG in Erythrozyten von 8 auf 0,5 mmol/l sinken kann? Ist eine Transfusion dieses Blutes an schwerkranke Patienten möglich, wenn die Konzentration von 2,3-BPG frühestens nach drei Tagen wiederhergestellt ist? Ist es möglich, die Funktionen der roten Blutkörperchen durch Zugabe von 2,3-BPG zum Blut wiederherzustellen?
Erinnern Sie sich an die Struktur der einfachsten Immunglobulinmoleküle. Welche Rolle spielen Immunglobuline im Immunsystem? Warum werden Igs oft als bivalente Substanzen bezeichnet? Wie hängt die Struktur von Igs mit ihrer Funktion zusammen? (Beschreiben Sie anhand eines Beispiels eine Klasse von Immunglobulinen.)
6. Physikochemische Eigenschaften von Proteinen und Methoden zu ihrer Trennung.
Wie beeinflusst die Nettoladung eines Proteins seine Löslichkeit?
a) Bestimmen Sie die Gesamtladung des Peptids bei pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis<7, рН <<7?
b) Wie ändert sich die Ladung dieses Peptids bei pH >7, pH
c) Was ist der isoelektrische Punkt eines Proteins (IEP) und in welcher Umgebung liegt es?
IET dieses Peptids?
7. d) bei welchem pH-Wert wird die geringste Löslichkeit dieses Peptids beobachtet?
8. Die Gelfiltration dient der Trennung einzelner Proteine. Eine Mischung, die die Proteine A, B, C mit Molekulargewichten von 160.000, 80.000 bzw. 60.000 enthielt, wurde durch Gelfiltration analysiert (Abb. 1.35). Das gequollene Gelgranulat ist für Proteine mit einem Molekulargewicht von weniger als 70.000 durchlässig. Welches Prinzip liegt dieser Trennmethode zugrunde? Welcher Graph gibt die Ergebnisse der Fraktionierung korrekt wieder? Geben Sie die Reihenfolge an, in der die Proteine A, B und C aus der Säule freigesetzt werden.
 Reis. 1,35. Verwendung von Gelfiltration zur Trennung von Proteinen
Reis. 1,35. Verwendung von Gelfiltration zur Trennung von Proteinen
9. In Abb. 1.36, A zeigt ein Diagramm der Elektrophorese auf Papier von Blutserumproteinen einer gesunden Person. Die relativen Mengen der mit dieser Methode erhaltenen Proteinfraktionen sind: Albumine 54–58 %, α 1 -Globuline 6–7 %, α 2 -Globuline 8–9 %, β-Globuline 13 %, γ-Globuline 11–12 %.
 Reis. 1.36 Elektrophorese auf Papier von Blutplasmaproteinen einer gesunden Person (A) und eines Patienten (B)
Reis. 1.36 Elektrophorese auf Papier von Blutplasmaproteinen einer gesunden Person (A) und eines Patienten (B)
I – γ-Globuline; II – β-Globuline; III -α 2-Globulin; IV -α 2-Globulin; V – Albumine
Viele Erkrankungen gehen mit quantitativen Veränderungen der Zusammensetzung der Serumproteine einher (Dysproteinämie). Die Art dieser Veränderungen wird bei der Diagnosestellung und der Beurteilung der Schwere und des Stadiums der Erkrankung berücksichtigt.
Verwendung der in der Tabelle angegebenen Daten. 1.5, machen Sie eine Vermutung über die Krankheit, die durch das in Abb. dargestellte elektrophoretische Profil gekennzeichnet ist. 1,36.
Tabelle 1.5. Veränderungen der Konzentration von Serumproteinen in der Pathologie
Eichhörnchen- organische Verbindungen mit hohem Molekulargewicht, bestehend aus α-Aminosäureresten.
IN Proteinzusammensetzung umfasst Kohlenstoff, Wasserstoff, Stickstoff, Sauerstoff, Schwefel. Einige Proteine bilden Komplexe mit anderen Molekülen, die Phosphor, Eisen, Zink und Kupfer enthalten.
Proteine haben ein großes Molekulargewicht: Eialbumin – 36.000, Hämoglobin – 152.000, Myosin – 500.000. Zum Vergleich: Das Molekulargewicht von Alkohol beträgt 46, Essigsäure – 60, Benzol – 78.
Aminosäurezusammensetzung von Proteinen
Eichhörnchen- nichtperiodische Polymere, deren Monomere sind α-Aminosäuren. Typischerweise werden 20 Arten von α-Aminosäuren als Proteinmonomere bezeichnet, obwohl über 170 davon in Zellen und Geweben vorkommen.
Je nachdem, ob Aminosäuren im Körper von Menschen und anderen Tieren synthetisiert werden können, werden sie unterschieden: nichtessentielle Aminosäuren- kann synthetisiert werden; essentielle Aminosäuren- kann nicht synthetisiert werden. Essentielle Aminosäuren müssen dem Körper über die Nahrung zugeführt werden. Pflanzen synthetisieren alle Arten von Aminosäuren.
Abhängig von der Aminosäurezusammensetzung Proteine sind: vollständig- den gesamten Satz an Aminosäuren enthalten; defekt- Einige Aminosäuren fehlen in ihrer Zusammensetzung. Bestehen Proteine nur aus Aminosäuren, nennt man sie einfach. Wenn Proteine neben Aminosäuren auch einen Nicht-Aminosäure-Anteil (prosthetische Gruppe) enthalten, werden sie aufgerufen Komplex. Die prothetische Gruppe kann durch Metalle (Metalloproteine), Kohlenhydrate (Glykoproteine), Lipide (Lipoproteine) und Nukleinsäuren (Nukleoproteine) repräsentiert werden.
Alle Aminosäuren enthalten: 1) Carboxylgruppe (-COOH), 2) Aminogruppe (-NH 2), 3) Radikal oder R-Gruppe (der Rest des Moleküls). Die Struktur des Restes ist für verschiedene Arten von Aminosäuren unterschiedlich. Abhängig von der Anzahl der Aminogruppen und Carboxylgruppen, aus denen die Aminosäuren bestehen, werden sie unterschieden: neutrale Aminosäuren mit einer Carboxylgruppe und einer Aminogruppe; basische Aminosäuren mit mehr als einer Aminogruppe; saure Aminosäuren mit mehr als einer Carboxylgruppe.
Aminosäuren sind amphotere Verbindungen, da sie in Lösung sowohl als Säuren als auch als Basen wirken können. In wässrigen Lösungen liegen Aminosäuren in verschiedenen ionischen Formen vor.
Peptidbindung
Peptide- organische Substanzen, die aus Aminosäureresten bestehen, die durch Peptidbindungen verbunden sind.
Die Bildung von Peptiden erfolgt durch die Kondensationsreaktion von Aminosäuren. Wenn die Aminogruppe einer Aminosäure mit der Carboxylgruppe einer anderen interagiert, entsteht zwischen ihnen eine kovalente Stickstoff-Kohlenstoff-Bindung, die sogenannte Peptid. Abhängig von der Anzahl der im Peptid enthaltenen Aminosäurereste gibt es Dipeptide, Tripeptide, Tetrapeptide usw. Die Bildung einer Peptidbindung kann viele Male wiederholt werden. Dies führt zur Bildung Polypeptide. An einem Ende des Peptids befindet sich eine freie Aminogruppe (N-Terminus genannt) und am anderen Ende eine freie Carboxylgruppe (C-Terminus genannt).

Räumliche Organisation von Proteinmolekülen
Die Erfüllung bestimmter spezifischer Funktionen durch Proteine hängt von der räumlichen Konfiguration ihrer Moleküle ab. Darüber hinaus ist es für die Zelle energetisch ungünstig, Proteine in einer ungefalteten Form, in Form einer Kette, zu halten. Daher unterliegen Polypeptidketten einer Faltung und erhalten eine bestimmte dreidimensionale Struktur oder Konformation. Es gibt 4 Ebenen räumliche Organisation von Proteinen.
Primäre Proteinstruktur- die Reihenfolge der Anordnung der Aminosäurereste in der Polypeptidkette, aus der das Proteinmolekül besteht. Die Bindung zwischen Aminosäuren ist eine Peptidbindung.

Wenn ein Proteinmolekül nur aus 10 Aminosäureresten besteht, beträgt die Anzahl der theoretisch möglichen Varianten von Proteinmolekülen, die sich in der Reihenfolge des Aminosäurewechsels unterscheiden, 10 20. Mit 20 Aminosäuren können Sie noch mehr unterschiedliche Kombinationen daraus erstellen. Im menschlichen Körper wurden etwa zehntausend verschiedene Proteine gefunden, die sich sowohl untereinander als auch von den Proteinen anderer Organismen unterscheiden.
Es ist die Primärstruktur des Proteinmoleküls, die die Eigenschaften der Proteinmoleküle und ihre räumliche Konfiguration bestimmt. Der Austausch lediglich einer Aminosäure durch eine andere in einer Polypeptidkette führt zu einer Veränderung der Eigenschaften und Funktionen des Proteins. Beispielsweise führt der Ersatz der sechsten Glutaminaminosäure in der β-Untereinheit des Hämoglobins durch Valin dazu, dass das Hämoglobinmolekül als Ganzes seine Hauptfunktion – den Sauerstofftransport – nicht erfüllen kann; In solchen Fällen entwickelt die Person eine Krankheit, die Sichelzellenanämie genannt wird.
Sekundärstruktur- geordnete Faltung der Polypeptidkette zu einer Spirale (sieht aus wie eine verlängerte Feder). Die Windungen der Helix werden durch Wasserstoffbrückenbindungen verstärkt, die zwischen Carboxylgruppen und Aminogruppen entstehen. Fast alle CO- und NH-Gruppen sind an der Bildung von Wasserstoffbrückenbindungen beteiligt. Sie sind schwächer als Peptide, verleihen dieser Konfiguration jedoch bei mehrfacher Wiederholung Stabilität und Steifigkeit. Auf der Ebene der Sekundärstruktur befinden sich Proteine: Fibroin (Seide, Spinnennetz), Keratin (Haare, Nägel), Kollagen (Sehnen).



Tertiärstruktur- Packung von Polypeptidketten in Kügelchen, die aus der Bildung chemischer Bindungen (Wasserstoff, ionisch, Disulfid) und der Etablierung hydrophober Wechselwirkungen zwischen den Resten von Aminosäureresten resultiert. Die Hauptrolle bei der Bildung der Tertiärstruktur spielen hydrophil-hydrophobe Wechselwirkungen. In wässrigen Lösungen neigen hydrophobe Radikale dazu, sich vor dem Wasser zu verstecken und sich innerhalb der Kügelchen zu gruppieren, während hydrophile Radikale aufgrund der Hydratation (Wechselwirkung mit Wasserdipolen) dazu neigen, auf der Oberfläche des Moleküls zu erscheinen. In einigen Proteinen wird die Tertiärstruktur durch kovalente Disulfidbindungen stabilisiert, die zwischen den Schwefelatomen zweier Cysteinreste gebildet werden. Auf der Tertiärstrukturebene befinden sich Enzyme, Antikörper und einige Hormone.

Quartärstruktur charakteristisch für komplexe Proteine, deren Moleküle aus zwei oder mehr Kügelchen bestehen. Die Untereinheiten werden durch ionische, hydrophobe und elektrostatische Wechselwirkungen im Molekül gehalten. Manchmal treten während der Bildung einer Quartärstruktur Disulfidbindungen zwischen Untereinheiten auf. Das am besten untersuchte Protein mit Quartärstruktur ist Hämoglobin. Es besteht aus zwei α-Untereinheiten (141 Aminosäurereste) und zwei β-Untereinheiten (146 Aminosäurereste). Mit jeder Untereinheit ist ein Hämmolekül verbunden, das Eisen enthält.
Wenn die räumliche Konformation von Proteinen aus irgendeinem Grund vom Normalzustand abweicht, kann das Protein seine Funktionen nicht erfüllen. Die Ursache des „Rinderwahnsinns“ (spongiforme Enzephalopathie) ist beispielsweise die abnormale Konformation von Prionen, den Oberflächenproteinen von Nervenzellen.
Eigenschaften von Proteinen
Die Aminosäurezusammensetzung und Struktur des Proteinmoleküls bestimmen es Eigenschaften. Proteine vereinen basische und saure Eigenschaften, die durch Aminosäurereste bestimmt werden: Je mehr saure Aminosäuren in einem Protein enthalten sind, desto ausgeprägter sind seine sauren Eigenschaften. Die Fähigkeit zur Abgabe und Zugabe von H+ wird bestimmt Puffereigenschaften von Proteinen; Einer der stärksten Puffer ist Hämoglobin in den roten Blutkörperchen, das den pH-Wert des Blutes auf einem konstanten Niveau hält. Es gibt lösliche Proteine (Fibrinogen) und es gibt unlösliche Proteine, die mechanische Funktionen erfüllen (Fibroin, Keratin, Kollagen). Es gibt chemisch aktive Proteine (Enzyme), es gibt chemisch inaktive Proteine, die gegenüber verschiedenen Umwelteinflüssen resistent sind, und solche, die äußerst instabil sind.
Äußere Faktoren (Hitze, ultraviolette Strahlung, Schwermetalle und deren Salze, pH-Änderungen, Strahlung, Dehydrierung)
kann zu Störungen der strukturellen Organisation des Proteinmoleküls führen. Der Prozess des Verlusts der dreidimensionalen Konformation, die einem bestimmten Proteinmolekül innewohnt, wird als bezeichnet Denaturierung. Die Ursache der Denaturierung ist das Aufbrechen von Bindungen, die eine bestimmte Proteinstruktur stabilisieren. Zunächst werden die schwächsten Bindungen zerbrochen, und je strenger die Bedingungen werden, desto stärkere werden zerbrochen. Daher gehen zunächst die Quartär-, dann die Tertiär- und Sekundärstrukturen verloren. Eine Änderung der räumlichen Konfiguration führt zu einer Änderung der Eigenschaften des Proteins und macht es dadurch für das Protein unmöglich, seine inhärenten biologischen Funktionen zu erfüllen. Wenn die Denaturierung nicht mit einer Zerstörung der Primärstruktur einhergeht, kann dies der Fall sein reversibel In diesem Fall kommt es zu einer Selbstwiederherstellung der für das Protein charakteristischen Konformation. Beispielsweise unterliegen Membranrezeptorproteine einer solchen Denaturierung. Der Prozess der Wiederherstellung der Proteinstruktur nach der Denaturierung wird als bezeichnet Renaturierung. Wenn eine Wiederherstellung der räumlichen Konfiguration des Proteins nicht möglich ist, spricht man von Denaturierung irreversibel.
Funktionen von Proteinen
| Funktion | Beispiele und Erklärungen |
|---|---|
| Konstruktion | Proteine sind an der Bildung zellulärer und extrazellulärer Strukturen beteiligt: Sie sind Teil von Zellmembranen (Lipoproteine, Glykoproteine), Haaren (Keratin), Sehnen (Kollagen) usw. |
| Transport | Das Blutprotein Hämoglobin bindet Sauerstoff und transportiert ihn von der Lunge zu allen Geweben und Organen und von dort aus Kohlendioxid in die Lunge; Zur Zusammensetzung der Zellmembranen gehören spezielle Proteine, die für den aktiven und streng selektiven Transfer bestimmter Stoffe und Ionen von der Zelle in die äußere Umgebung und zurück sorgen. |
| Regulatorisch | Proteinhormone sind an der Regulierung von Stoffwechselprozessen beteiligt. Beispielsweise reguliert das Hormon Insulin den Blutzuckerspiegel, fördert die Glykogensynthese und steigert die Bildung von Fetten aus Kohlenhydraten. |
| Schützend | Als Reaktion auf das Eindringen fremder Proteine oder Mikroorganismen (Antigene) in den Körper werden spezielle Proteine gebildet – Antikörper, die diese binden und neutralisieren können. Fibrin, das aus Fibrinogen gebildet wird, hilft, Blutungen zu stoppen. |
| Motor | Die kontraktilen Proteine Aktin und Myosin sorgen bei mehrzelligen Tieren für die Muskelkontraktion. |
| Signal | In die Oberflächenmembran der Zelle sind Proteinmoleküle eingebaut, die in der Lage sind, ihre Tertiärstruktur als Reaktion auf Umweltfaktoren zu ändern und so Signale von der äußeren Umgebung zu empfangen und Befehle an die Zelle zu übermitteln. |
| Lagerung | Im Körper von Tieren werden Proteine mit Ausnahme von Eialbumin und Milchkasein in der Regel nicht gespeichert. Dank Proteinen können einige Substanzen jedoch im Körper gespeichert werden. Beim Abbau von Hämoglobin wird beispielsweise Eisen nicht aus dem Körper entfernt, sondern gespeichert und bildet einen Komplex mit dem Protein Ferritin. |
| Energie | Wenn 1 g Protein in Endprodukte zerfällt, werden 17,6 kJ freigesetzt. Zuerst zerfallen Proteine in Aminosäuren und dann in die Endprodukte – Wasser, Kohlendioxid und Ammoniak. Allerdings werden Proteine erst dann als Energiequelle genutzt, wenn andere Quellen (Kohlenhydrate und Fette) aufgebraucht sind. |
| Katalytisch | Eine der wichtigsten Funktionen von Proteinen. Bereitgestellt durch Proteine – Enzyme, die biochemische Reaktionen in Zellen beschleunigen. Beispielsweise katalysiert Ribulosebiphosphat-Carboxylase die Fixierung von CO 2 während der Photosynthese. |
Enzyme
Enzyme, oder Enzyme sind eine spezielle Klasse von Proteinen, die biologische Katalysatoren sind. Dank Enzymen laufen biochemische Reaktionen mit enormer Geschwindigkeit ab. Die Geschwindigkeit enzymatischer Reaktionen ist Zehntausende Male (und manchmal Millionen) höher als die Geschwindigkeit von Reaktionen, die unter Beteiligung anorganischer Katalysatoren ablaufen. Der Stoff, auf den das Enzym einwirkt, wird genannt Substrat.
Enzyme sind kugelförmige Proteine, Strukturmerkmale Enzyme können in zwei Gruppen eingeteilt werden: einfache und komplexe. Einfache Enzyme sind einfache Proteine, d.h. bestehen nur aus Aminosäuren. Komplexe Enzyme sind komplexe Proteine, d.h. Zusätzlich zum Proteinanteil enthalten sie eine Gruppe nicht proteinischer Natur - Cofaktor. Einige Enzyme nutzen Vitamine als Cofaktoren. Das Enzymmolekül enthält einen speziellen Teil, der als aktives Zentrum bezeichnet wird. Aktives Zentrum- ein kleiner Abschnitt des Enzyms (drei bis zwölf Aminosäurereste), in dem die Bindung des Substrats oder der Substrate unter Bildung eines Enzym-Substrat-Komplexes erfolgt. Nach Abschluss der Reaktion zerfällt der Enzym-Substrat-Komplex in das Enzym und das/die Reaktionsprodukt(e). Einige Enzyme haben (außer aktive) allosterische Zentren- Bereiche, an denen Enzymgeschwindigkeitsregulatoren angebracht sind ( allosterische Enzyme).

Reaktionen der enzymatischen Katalyse zeichnen sich aus durch: 1) hohe Effizienz, 2) strenge Selektivität und Wirkungsrichtung, 3) Substratspezifität, 4) feine und präzise Regulierung. Die Substrat- und Reaktionsspezifität enzymatischer Katalysereaktionen wird durch die Hypothesen von E. Fischer (1890) und D. Koshland (1959) erklärt.
E. Fisher (Schlüssel-Schloss-Hypothese) schlugen vor, dass die räumlichen Konfigurationen des aktiven Zentrums des Enzyms und des Substrats genau einander entsprechen müssen. Das Substrat wird mit dem „Schlüssel“ verglichen, das Enzym mit dem „Schloss“.
D. Koshland (Handschuh-Hypothese) schlugen vor, dass die räumliche Korrespondenz zwischen der Struktur des Substrats und dem aktiven Zentrum des Enzyms erst im Moment ihrer Wechselwirkung miteinander entsteht. Diese Hypothese wird auch genannt induzierte Korrespondenzhypothese.
Die Geschwindigkeit enzymatischer Reaktionen hängt ab von: 1) der Temperatur, 2) der Enzymkonzentration, 3) der Substratkonzentration, 4) dem pH-Wert. Da es sich bei Enzymen um Proteine handelt, ist zu betonen, dass ihre Aktivität unter physiologisch normalen Bedingungen am höchsten ist.
Die meisten Enzyme können nur bei Temperaturen zwischen 0 und 40 °C wirken. Innerhalb dieser Grenzen erhöht sich die Reaktionsgeschwindigkeit mit jedem Temperaturanstieg um 10 °C etwa um das Zweifache. Bei Temperaturen über 40 °C kommt es zur Denaturierung des Proteins und die Enzymaktivität nimmt ab. Bei Temperaturen nahe dem Gefrierpunkt werden Enzyme inaktiviert.
Mit zunehmender Substratmenge nimmt die Geschwindigkeit der enzymatischen Reaktion zu, bis die Anzahl der Substratmoleküle der Anzahl der Enzymmoleküle entspricht. Bei einer weiteren Erhöhung der Substratmenge erhöht sich die Geschwindigkeit nicht, da die aktiven Zentren des Enzyms gesättigt sind. Eine Erhöhung der Enzymkonzentration führt zu einer erhöhten katalytischen Aktivität, da eine größere Anzahl von Substratmolekülen pro Zeiteinheit Umwandlungen durchläuft.

Für jedes Enzym gibt es einen optimalen pH-Wert, bei dem es seine maximale Aktivität zeigt (Pepsin – 2,0, Speichelamylase – 6,8, Pankreaslipase – 9,0). Bei höheren oder niedrigeren pH-Werten nimmt die Enzymaktivität ab. Bei plötzlichen pH-Änderungen denaturiert das Enzym.
Die Geschwindigkeit allosterischer Enzyme wird durch Substanzen reguliert, die an allosterische Zentren binden. Wenn diese Stoffe eine Reaktion beschleunigen, werden sie aufgerufen Aktivatoren, wenn sie langsamer werden - Inhibitoren.
Klassifizierung von Enzymen
Je nach Art der von ihnen katalysierten chemischen Umwandlungen werden Enzyme in 6 Klassen eingeteilt:
- Oxireduktasen(Übertragung von Wasserstoff-, Sauerstoff- oder Elektronenatomen von einem Stoff auf einen anderen – Dehydrogenase),
- Transferasen(Übertragung einer Methyl-, Acyl-, Phosphat- oder Aminogruppe von einer Substanz auf eine andere – Transaminase),
- Hydrolasen(Hydrolysereaktionen, bei denen aus dem Substrat zwei Produkte gebildet werden – Amylase, Lipase),
- Lyasen(nichthydrolytische Addition an das Substrat oder Ablösung einer Atomgruppe davon, wobei C-C-, C-N-, C-O-, C-S-Bindungen aufgebrochen werden können – Decarboxylase),
- Isomerasen(intramolekulare Umlagerung - Isomerase),
- Ligasen(die Verbindung zweier Moleküle durch Bildung von C-C-, C-N-, C-O-, C-S-Bindungen – Synthetase).
Klassen sind wiederum in Unterklassen und Unterunterklassen unterteilt. In der aktuellen internationalen Klassifikation hat jedes Enzym einen spezifischen Code, der aus vier durch Punkte getrennten Zahlen besteht. Die erste Zahl ist die Klasse, die zweite ist die Unterklasse, die dritte ist die Unterunterklasse, die vierte ist die Seriennummer des Enzyms in dieser Unterklasse, zum Beispiel lautet der Arginase-Code 3.5.3.1.
Gehe zu Vorträge Nr. 2„Struktur und Funktionen von Kohlenhydraten und Lipiden“
Gehe zu Vorträge Nr. 4„Struktur und Funktionen von ATP-Nukleinsäuren“
Proteine (Proteine) sind hochmolekulare Polymerverbindungen peptidischer Natur (Polyheteroaminosäuren).
Die Primärstruktur von Proteinen ist die Abfolge alternierender Aminosäurereste in der Polypeptidkette (PPC).
Die Primärstruktur von Proteinen ist kovalent Struktur, da es darauf basiert Peptid Verbindung zwischen a-Amino- und a-Carboxylgruppen von Aminosäuren. Dadurch sind die Polypeptidketten unverzweigt.
Das Grundgerüst (Rückgrat, Grundgerüst) einer Polypeptidkette besteht aus sich regelmäßig wiederholenden Strukturelementen
Die Polypeptidkette weist Vektorialität auf, die Richtung der Kette verläuft vom N-Terminus (Anfang der Kette) zum C-Terminus (Ende der Kette), der N-Terminus ist das Ende, an dem sich die freie a-Aminogruppe befindet befindet sich. Der C-Terminus ist das Ende, das die freie a-Carboxylgruppe enthält. Die Aminosäuresequenz von Proteinen wird ausgehend vom N-Terminus mit aus drei Buchstaben abgekürzten Aminosäurenamen bezeichnet, zum Beispiel: gly-ala-cis-pro. Es kann auch eine aus einem Buchstaben bestehende Bezeichnung für Aminosäurereste in einem Protein verwendet werden.
Die N- und C-Termini von Proteinen können verändert werden. Die Aminogruppe am N-Terminus kann acetyliert, formyliert oder methyliert sein. In einer Reihe von Proteinen ist der N-terminale Rest ein Pyrrolidoncarbonat (Pyroglutamat), das keine freie Aminogruppe enthält. Der C-Terminus kann amidiert sein. C-terminale Modifikationen sind im Vergleich zu N-terminalen Modifikationen seltener.
Der Polykondensationskoeffizient von Proteinen variiert im Bereich von 50 bis 2500. Typischerweise enthält ein Protein 100–300 Aminosäurereste. Da das durchschnittliche Molekulargewicht eines einzelnen Aminosäurerests etwa 110 Da beträgt, variiert das Molekulargewicht von Proteinen zwischen 6000 und Millionen Da.
Jedes einzelne Protein hat eine einzigartige Primärstruktur. Das erste Protein, dessen Primärstruktur etabliert wurde, war Insulin. Sanger hat dies geschafft. Seine Strategie war wie folgt. Er trennte zunächst die beiden Polypeptidketten und führte dann deren spezifische enzymatische Spaltung in kleine Peptide mit überlappenden Sequenzen durch. Die N-terminalen Reste wurden dann mit 1-Fluor-2,4-dinitrobenzol identifiziert. Darüber hinaus bestimmte er die Aminosäurezusammensetzung der Peptide und konnte schließlich deren Struktur durch Vergleich der Sequenzen überlappender Peptide bestimmen. Generell hat Sangers Strategie bis heute ihre Bedeutung behalten. Es wurden jedoch auch andere Ansätze vorgeschlagen. Edman entwickelte eine Methode für ein automatisches Verfahren zur sequentiellen Spaltung und Identifizierung N-terminaler Aminosäurereste. Mithilfe der Röntgenbeugungsanalyse kann die Primärstruktur entschlüsselt werden. Die Reihenfolge der Aminosäurereste kann aus der Nukleotidsequenz der Messenger-RNA bestimmt werden.
Derzeit ist die Primärstruktur von mehr als 2000 Proteinen bekannt. Theoretisch ist die Zahl der unterschiedlichen Varianten der Primärstruktur von Proteinen unbegrenzt. Selbst für ein Polypeptid aus 20 verschiedenen Aminosäuren beträgt die Anzahl möglicher Sequenzen 20´10 18 . In der belebten Natur wird nur ein kleiner Bruchteil der möglichen Sequenzen realisiert, deren Gesamtzahl bei allen Arten lebender Organismen auf 10 10 -10 12 geschätzt wird.
Die Primärstruktur von Proteinen ist genetisch bedingt, d.h. Die Reihenfolge der Aminosäuren in einem Protein wird durch die Reihenfolge der Nukleotide bestimmt DNA. Verzerrungen der DNA-Nukleotidsequenz führen zur Entstehung abnormaler Proteine mit veränderten biologischen Eigenschaften, was die Ursache für molekulare Pathologien ist. Insbesondere wird die Sichelzellenanämie durch eine Punktmutation im Gen verursacht, das die Hämoglobin-b-Kette steuert. Die Folge davon ist der Ersatz des Glutamatrestes an der 6. Position der b-Kette durch Valin. Diese Substitution führt zum Verlust einer negativen Ladung in jeder der beiden b-Ketten, was zu einer Änderung der Konformation des Hämoglobins und zum Verlust seiner biologischen Funktion führt.
Homologe Proteine sind Proteine, die in verschiedenen Spezies die gleichen Funktionen erfüllen. Ein Beispiel ist Hämoglobin: Bei allen Wirbeltieren erfüllt es die gleiche Funktion im Zusammenhang mit dem Sauerstofftransport. Homologe Proteine zeichnen sich durch das Vorhandensein der gleichen Aminosäuren an vielen Positionen aus. Wie sich herausstellte, ist die Anzahl der Aminosäurereste, an denen sich homologe Proteine unterscheiden, proportional zum phylogenetischen Unterschied zwischen diesen Arten. Beispielsweise unterscheiden sich die Cytochrom-C-Moleküle von Pferd und Hefe in 48 Aminosäureresten, während sich die gleichen Moleküle von Huhn und Ente nur in zwei Resten unterscheiden. Cytochrome C von Huhn und Truthahn weisen identische Aminosäuresequenzen auf. Informationen über die Anzahl der Unterschiede in den Aminosäuresequenzen homologer Proteine verschiedener Arten werden zur Erstellung von Evolutionskarten verwendet, die die aufeinanderfolgenden Stadien der Entstehung und Entwicklung verschiedener Tier- und Pflanzenarten im Evolutionsprozess widerspiegeln.
Vorlesung 3. Proteinstruktur
Definition:
Proteine sind unregelmäßige Polymere, deren Monomere bestehen L- Aminosäuren.
Aminosäuren
In der Natur gibt es zwei Formen von Stereoisomeren: L (Linkshänder) und D (rechtsdrehend). Außerdem L -Aminosäuren, aus denen Proteine bestehen, sind im Körper vorhanden D -Aminosäuren, die nicht in Proteinen enthalten sind.
Die allgemeine Formel einer Aminosäure ist in der Abbildung dargestellt.
Dies gilt für 19 der 20 Aminosäuren, die in Proteinen vorkommen. Zusätzlich zu diesen 19 Aminosäuren enthalten Proteine eine Iminosäure - Prolin
Alle Aminosäuren enthalten α -Aminogruppe. Daher der Name – „α-Aminosäuren“. In Prolin – α- imino Gruppe.
Klassifizierung der Aminosäuren, aus denen Proteine bestehen, nach dem Prinzip der Polarität (Unpolarität) des Rests.
1. Unpolare oder hydrophobe Radikale.
Aliphatisch - Alanin, Valin, Leucin, Isoleucin. Schwefelhaltig Methionin Aromatisch - Phenylalanin, Tryptophan. Iminosäure Prolin
2. Polare, aber ungeladene Radikale. Glycin.
Hydroxyaminosäuren - Serin, Threonin, Tyrosin. Enthält eine Sulfhydrylgruppe Cystein. Enthält eine Amidgruppe: Asparagin, Glutamin.
3. Negativ geladene Radikale. Asparaginsäure, Glutaminsäure.
4. Positiv geladene Radikale. Lysin, Arginin, Histidin.
Primäre Proteinstruktur
Definition:
Die Primärstruktur eines Proteins ist die Abfolge von Aminosäureresten in der Polypeptidkette.
Aminosäuren werden über kovalente (Amid-)Bindungen zu einem Polypeptid verbunden.
Ein Tripeptid, das aus drei verschiedenen Aminosäuren besteht, hat möglicherweise 3! = 6 verschiedene Primärstrukturen.
Ein aus zwanzig verschiedenen Aminosäuren bestehendes Oligopeptid hat eine Primärstrukturvielfalt von 20!, also 2x10 18.
Die Vielfalt der Primärstrukturen eines durchschnittlich großen Proteins (ca. 500 Aminosäuren) beträgt bereits etwa 20.500 Varianten (wenn alle Aminosäuren in äquimolaren Verhältnissen vorliegen).
AnEs gab, gibt und wird nicht zwei Menschen auf der Erde geben, die genau den gleichen Satz an Proteinen haben.
Sekundärstruktur von Proteinen
Definition:
Die Sekundärstruktur eines Proteins ist die geordnete Struktur von Polypeptidketten, die durch Wasserstoffbrückenbindungen zwischen C=O und entsteht N- Hverschiedene Aminosäuren.
Die Sekundärstruktur kann regelmäßig (α-Helix) oder unregelmäßig (β-Faltblattstruktur) sein. In α-Helix NH-Gruppe n Der Aminosäurerest interagiert mit der C=O-Gruppe des (n-4)-ten Aminosäurerests. Es gibt 3,6 Aminosäurereste pro Windung einer β-Helix mit einem Durchmesser von 10,1 Å. Die Identitätsperiode einer regulären α-Helix beträgt 18 Aminosäuren (5 Windungen). Der Disruptor einer regulären α-Helix ist in erster Linie Prolin. Den zweitwichtigsten Effekt haben gleichgeladene Radikale in der Nähe.
β-Falten können nicht nur von einzelnen, sondern auch von benachbarten Polypeptiden gebildet werden, die Teil eines Proteins sind.
Rein natürliches Alpha oder Beta - Proteine existieren nicht.
Protein-Tertiärstruktur
Definition
Die Tertiärstruktur eines Proteins ist die räumliche Konformation eines Polypeptids, das eine Sekundärstruktur aufweist und durch Wechselwirkungen zwischen Radikalen bestimmt wird.
Es gibt vier Arten von Wechselwirkungen zwischen Radikalen.
Arten von Wechselwirkungen zwischen Radikalen
1 . Kovalent Kommunikation zwischen Reste zwei Cysteine (Disulfid Brücken).
2. Ionische (elektrostatische) Wechselwirkungen zwischen entgegengesetzt geladenen Aminosäureresten (drei Radikale mit einem „+“-Zeichen und zwei mit einem „-“-Zeichen).
Beispielsweise ist die positiv geladene ε-Aminogruppe von Lysin (- NH 3 +) wird von der negativ geladenen Carboxylgruppe – (COO-) Glutamin- oder Asparaginsäure – angezogen.
3. Wasserstoffbrücken.
Beteiligt sind alle Aminosäuren mit Hydroxyl-, Amid- oder Carboxylgruppen.
4. Hydrophobe Wechselwirkungen . Wird zwischen unpolaren Radikalen in einer wässrigen Umgebung gebildet. Es sind 8 Aminosäuren beteiligt (erste Klasse).
Die Tertiärstruktur eines Proteins wird vollständig durch seine Primärstruktur bestimmt, d. h. Abfolge von Aminosäuren, die wiederum durch den genetischen Code bestimmt wird.
Hydrophobe Wechselwirkungen sind aufgrund ihrer Nichtselektivität (Unspezifität) und Vielfältigkeit entscheidend.
Die meisten Proteine haben einen hydrophoben Kern.
Quartäre Proteinstruktur
Definition: Die Quartärstruktur eines Proteins ist die Aggregation von zwei oder mehr Polypeptidketten mit Tertiärstruktur zu einer oligomeren, funktionell bedeutsamen Zusammensetzung.
Die Bindungen, die die Quartärstruktur bilden und aufrechterhalten, sind mit Ausnahme der hydrophoben die gleichen wie diejenigen, die die Tertiärstruktur bilden.
N Das -Ende der Betakette enthält polare Glutaminsäure („-“ geladen). Bei Patienten mit Sichelzellenanämie wird stattdessen unpolares Valin verwendet.Von den 574 Aminosäuren wurden 2 ersetzt.
Dieses Hämoglobin verliert seine Löslichkeit und es bildet sich ein faseriger Niederschlag, der die roten Blutkörperchen deformiert.
Sichelzellenanämie ist eine genetische Erkrankung. Der Grund ist der Ersatz nur eines Nukleotids im Gen, das für die Hämoglobin-B-Kette kodiert. Kinder, die für dieses Allel rezessiv homozygot sind, werden nicht älter als zwei Jahre. Heterozygoten haben 85 % normale und 15 % defekte rote Blutkörperchen. Dominante Homozygote erkranken an Malaria, Heterozygote erkranken nicht an Malaria.
Kugelförmige und fibrilläre Proteine
95 % der Proteine haben einen hydrophoben Kern. 5 % fibrilläre Proteine.
Die überwiegende Mehrheit der globulären Proteine ist löslich. Die meisten Fibrillen sind unlöslich (α-Keratine – sie machen fast das gesamte Trockengewicht von Haaren, Wolle, Hörnern, Hufen, Nägeln, Schuppen, Federn aus; Kollagen – das Protein von Sehnen, Knorpel; Fibroin – Seidenprotein).
Fibrilläre Proteine enthalten einen größeren Anteil geladener Aminosäuren als globuläre – einzelne Ketten sind löslich und ihre Komplexe sind unpolar und unlöslich.
