კომპლექსური მარტივი ენის შესახებ.
ეს თემა რთული და კომპლექსურია, რომელიც დაუყოვნებლივ მოქმედებს ჩვენს ორგანიზმში მიმდინარე ბიოქიმიურ პროცესებზე. მაგრამ მაინც შევეცადოთ გაერკვნენ, რა არის მიტოქონდრია და როგორ მუშაობს ისინი.
ასე რომ, მიტოქონდრია ცოცხალი უჯრედის ერთ-ერთი ყველაზე მნიშვნელოვანი კომპონენტია. მარტივი სიტყვებით, შეგვიძლია ვთქვათ, რომ ეს უჯრედის ელექტროსადგური. მათი აქტივობა ეფუძნება ორგანული ნაერთების დაჟანგვას და ელექტრული პოტენციალის წარმოქმნას (ატფ-ის მოლეკულის დაშლის დროს გამოთავისუფლებული ენერგია) კუნთების შეკუმშვის განსახორციელებლად.
ჩვენ ყველამ ვიცით, რომ ჩვენი სხეული მუშაობს თერმოდინამიკის პირველი კანონის მკაცრად დაცვით. ენერგია ჩვენს სხეულში არ იქმნება, არამედ მხოლოდ გარდაიქმნება. ორგანიზმი მხოლოდ ენერგიის გარდაქმნის ფორმას ირჩევს, მისი წარმოქმნის გარეშე, ქიმიურიდან მექანიკურ და თერმულში. დედამიწის პლანეტაზე მთელი ენერგიის მთავარი წყარო მზეა. სინათლის სახით მოდის ჩვენთან, ენერგია შეიწოვება მცენარეების ქლოროფილის მიერ, სადაც ის აღაგზნებს წყალბადის ატომის ელექტრონს და ამით აძლევს ენერგიას ცოცხალ მატერიას.
ჩვენი სიცოცხლე მცირე ელექტრონის ენერგიას გვმართებს.
მიტოქონდრიის მუშაობა მოიცავს წყალბადის ელექტრონის ენერგიის ეტაპობრივ გადაცემას ლითონის ატომებს შორის, რომლებიც გვხვდება რესპირატორული ჯაჭვის ცილოვანი კომპლექსების ჯგუფებში (ცილების ელექტრონის სატრანსპორტო ჯაჭვი), სადაც ყოველ მომდევნო კომპლექსს აქვს უფრო მაღალი მიდრეკილება ელექტრონის მიმართ, იზიდავს მას, ვიდრე წინა, სანამ ელექტრონი არ გაერთიანდება მოლეკულურ ჟანგბადთან, რომელსაც აქვს ელექტრონების ყველაზე მაღალი აფინურობა.
ყოველ ჯერზე, როდესაც ელექტრონი გადის წრედში, გამოიყოფა ენერგია, რომელიც გროვდება ელექტროქიმიური გრადიენტის სახით და შემდეგ რეალიზდება კუნთების შეკუმშვისა და სითბოს გათავისუფლების სახით.
მიტოქონდრიებში ჟანგვითი პროცესების სერია, რომელიც საშუალებას გაძლევთ გადაიტანოთ ელექტრონის ენერგეტიკული პოტენციალი, ე.წ. "უჯრედშიდა სუნთქვა"ან ხშირად "სასუნთქი ჯაჭვი", ვინაიდან ელექტრონი გადადის ჯაჭვის გასწვრივ ატომიდან ატომამდე, სანამ არ მიაღწევს საბოლოო მიზანს ჟანგბადის ატომს.
მიტოქონდრიებს ჟანგბადი სჭირდებათ ენერგიის გადასატანად დაჟანგვის პროცესში.
მიტოქონდრია მოიხმარს ჟანგბადის 80%-მდე, რომელსაც ჩვენ ვსუნთქავთ.
მიტოქონდრია არის მუდმივი უჯრედის სტრუქტურა, რომელიც მდებარეობს მის ციტოპლაზმაში. მიტოქონდრიები, როგორც წესი, 0,5-დან 1 მკმ დიამეტრშია. ფორმის მიხედვით, მას აქვს მარცვლოვანი სტრუქტურა და შეუძლია დაიკავოს უჯრედის მოცულობის 20%. უჯრედის ამ მუდმივ ორგანულ სტრუქტურას ორგანელა ეწოდება. ორგანელებში ასევე შედის მიოფიბრილები - კუნთოვანი უჯრედის კონტრაქტული ერთეულები; და უჯრედის ბირთვი ასევე ორგანელაა. ზოგადად, ნებისმიერი მუდმივი უჯრედის სტრუქტურა არის ორგანელა-ორგანელა.
მიტოქონდრია აღმოაჩინა და პირველად აღწერა გერმანელმა ანატომისტმა და ჰისტოლოგმა რიჩარდ ალტმანმა 1894 წელს, ხოლო ამ ორგანელის სახელი დაარქვა სხვა გერმანელმა ჰისტოლოგმა კ.ბენდმა 1897 წელს. მაგრამ მხოლოდ 1920 წელს, ისევ გერმანელმა ბიოქიმიკოსმა ოტო ვაგბურგმა დაამტკიცა, რომ უჯრედული სუნთქვის პროცესები დაკავშირებულია მიტოქონდრიასთან.
არსებობს თეორია, რომ მიტოქონდრია გაჩნდა პრიმიტიული უჯრედების მიერ დაჭერის შედეგად, უჯრედები, რომლებსაც თავად არ შეეძლოთ ჟანგბადის გამოყენება ენერგიის გამომუშავებისთვის, პროტოგენოტური ბაქტერიები, რომლებსაც ამის გაკეთება შეეძლოთ. ზუსტად იმიტომ, რომ მიტოქონდრიონი ადრე ცალკე ცოცხალი ორგანიზმი იყო, მას დღემდე აქვს საკუთარი დნმ.
ადრე მიტოქონდრია დამოუკიდებელი ცოცხალი ორგანიზმი იყო.
ევოლუციის მსვლელობისას, პროგენოტებმა თავიანთი მრავალი გენი უღალატა ბირთვს, რომელიც წარმოიქმნა გაზრდილი ენერგოეფექტურობის გამო და შეწყვიტა დამოუკიდებელი ორგანიზმები. მიტოქონდრია ყველა უჯრედშია. სპერმაც კი შეიცავს მიტოქონდრიებს. სწორედ მათი წყალობით მოძრაობს სპერმატოზოვას კუდი, რომელიც ახორციელებს მის მოძრაობას. მაგრამ განსაკუთრებით ბევრია მიტაქონდრია იმ ადგილებში, სადაც ენერგია საჭიროა ნებისმიერი ცხოვრების პროცესისთვის. და რა თქმა უნდა, ეს, პირველ რიგში, კუნთების უჯრედებია.
კუნთოვან უჯრედებში მიტოქონდრიები შეიძლება გაერთიანდეს გიგანტური განშტოებული მიტოქონდრიების ჯგუფებად, რომლებიც დაკავშირებულია ერთმანეთთან ინტერმიტოქონდრიული კონტაქტების გამოყენებით, რომელშიც ისინი კოორდინირებული სამუშაო კოოპერატიული სისტემის შექმნა. ასეთ ზონაში სივრცეს აქვს გაზრდილი ელექტრონის სიმკვრივე. ახალი მიტოქონდრია წარმოიქმნება წინა ორგანელების მარტივი გაყოფით. ენერგიის მიწოდების ყველაზე "მარტივ" და ყველა უჯრედისთვის მისაწვდომ მექანიზმს ყველაზე ხშირად გლიკოლიზის ზოგად კონცეფციას უწოდებენ.
ეს არის გლუკოზის თანმიმდევრული დაშლის პროცესი პირუვინის მჟავამდე. თუ ეს პროცესი ხდება მოლეკულური ჟანგბადის მონაწილეობის გარეშეან არასაკმარისი ყოფნით, მაშინ ე.წ ანაერობული გლიკოლიზი. ამ შემთხვევაში, გლუკოზა იშლება არა საბოლოო პროდუქტებად, არამედ ლაქტურ და პირუვინ მჟავად, რომელიც შემდგომ ტრანსფორმაციას განიცდის დუღილის დროს. აქედან გამომდინარე, გამოთავისუფლებული ენერგია ნაკლებია, მაგრამ ენერგიის წარმოების სიჩქარე უფრო სწრაფია. ანაერობული გლიკოლიზის შედეგად უჯრედი ერთი გლუკოზის მოლეკულისგან იღებს 2 ATP მოლეკულას და 2 რძემჟავას მოლეკულას. ასეთი „ძირითადი“ ენერგეტიკული პროცესი შეიძლება მოხდეს ნებისმიერი უჯრედის შიგნით. მიტოქონდრიის გარეშე.
AT მოლეკულური ჟანგბადის არსებობახორციელდება მიტოქონდრიის შიგნით აერობული გლიკოლიზირესპირატორული ჯაჭვის შიგნით. პირუვიკის მჟავა აერობულ პირობებში მონაწილეობს ტრიკარბოქსილის მჟავას ციკლში ან კრებსის ციკლში. ამ მრავალსაფეხურიანი პროცესის შედეგად ერთი გლუკოზის მოლეკულისგან წარმოიქმნება 36 ATP მოლეკულა. უჯრედის ენერგეტიკული ბალანსის შედარება განვითარებული მიტოქონდრიებით და უჯრედებით, სადაც ისინი არ არის განვითარებული, აჩვენებს(საკმარისი ჟანგბადით) უჯრედის შიგნით გლუკოზის ენერგიის გამოყენების სისრულეში განსხვავება თითქმის 20-ჯერ არის!
ადამიანებში ჩონჩხის კუნთების ბოჭკოებს შეუძლიათ პირობითადმექანიკური და მეტაბოლური თვისებების მიხედვით იყოფა სამ ტიპად: - ნელი დაჟანგვა; - სწრაფი გლიკოლიზური; - სწრაფი ჟანგვის-გლიკოლიზური.

სწრაფი კუნთების ბოჭკოებიშექმნილია სწრაფი და მძიმე სამუშაოსთვის. მათი შეკუმშვისთვის ისინი ძირითადად იყენებენ ენერგიის სწრაფ წყაროებს, კერძოდ, კრიატინის ფოსფატს და ანაერობულ გლიკოლიზს. ამ ტიპის ბოჭკოებში მიტოქონდრიის შემცველობა გაცილებით ნაკლებია, ვიდრე ნელი კუნთების ბოჭკოებში.
ნელი კუნთების ბოჭკოებიშეასრულეთ ნელი შეკუმშვა, მაგრამ შეუძლიათ იმუშაონ დიდი ხნის განმავლობაში. ისინი იყენებენ აერობულ გლიკოლიზს და ენერგიის სინთეზს ცხიმებიდან ენერგიად. ეს უზრუნველყოფს ბევრად მეტ ენერგიას, ვიდრე ანაერობული გლიკოლიზი, მაგრამ სანაცვლოდ მეტ დროს მოითხოვს, რადგან გლუკოზის დეგრადაციის ჯაჭვი უფრო რთულია და მოითხოვს ჟანგბადის არსებობას, რომელსაც ასევე დრო სჭირდება ენერგიის გარდაქმნის ადგილზე ტრანსპორტირებას. ნელი კუნთების ბოჭკოებს წითელს უწოდებენ მიოგლობინის გამო, ცილა, რომელიც პასუხისმგებელია ბოჭკოში ჟანგბადის შეტანაზე. ნელი კუნთების ბოჭკოები შეიცავს მიტოქონდრიის მნიშვნელოვან რაოდენობას.
ჩნდება კითხვა, როგორ და რა ვარჯიშების დახმარებით შეიძლება განვითარდეს მიტოქონდრიების ფართო ქსელი კუნთოვან უჯრედებში? არსებობს ტრენინგის სხვადასხვა თეორიები და მეთოდები და მათ შესახებ მასალაში.
დამახასიათებელია უჯრედების დიდი უმრავლესობისთვის. მთავარი ფუნქციაა ორგანული ნაერთების დაჟანგვა და გამოთავისუფლებული ენერგიისგან ATP მოლეკულების გამომუშავება. პატარა მიტოქონდრიონი არის მთელი ორგანიზმის მთავარი ენერგეტიკული სადგური.
მიტოქონდრიის წარმოშობა
დღეს მეცნიერებს შორის ძალიან პოპულარულია მოსაზრება, რომ ევოლუციის პროცესში მიტოქონდრია უჯრედში დამოუკიდებლად არ გამოჩენილა. სავარაუდოდ, ეს მოხდა პრიმიტიული უჯრედის მიერ დაჭერის გამო, რომელსაც იმ დროს არ შეეძლო ჟანგბადის დამოუკიდებლად გამოყენება, ბაქტერია, რომელმაც იცოდა ამის გაკეთება და, შესაბამისად, ენერგიის შესანიშნავი წყარო იყო. ასეთი სიმბიოზი წარმატებული იყო და დაიმკვიდრა შემდგომ თაობებში. ამ თეორიას ადასტურებს მიტოქონდრიებში საკუთარი დნმ-ის არსებობა.
როგორ არის მოწყობილი მიტოქონდრია?
მიტოქონდრიას აქვს ორი გარსი: გარე და შიდა. გარე მემბრანის მთავარი ფუნქციაა ორგანოიდის გამოყოფა უჯრედის ციტოპლაზმისგან. იგი შედგება ბილიპიდური შრისა და მასში შემავალი ცილებისგან, რომლის მეშვეობითაც ხორციელდება სამუშაოსთვის აუცილებელი მოლეკულებისა და იონების ტრანსპორტირება. მიუხედავად იმისა, რომ გლუვი, შიდა ერთი ქმნის უამრავ ნაკეცს - cristae, რაც მნიშვნელოვნად ზრდის მის ფართობს. შიდა მემბრანა ძირითადად შედგება ცილებისგან, რომელთა შორისაა რესპირატორული ჯაჭვის ფერმენტები, სატრანსპორტო ცილები და დიდი ATP სინთეზის კომპლექსები. სწორედ აქ ხდება ATP სინთეზი. გარე და შიდა გარსებს შორის არის მემბრანთაშორისი სივრცე თავისი თანდაყოლილი ფერმენტებით.
მიტოქონდრიის შიდა ნაწილს მატრიცა ეწოდება. აქ არის ცხიმოვანი მჟავების და პირუვატის დაჟანგვის ფერმენტული სისტემები, კრებსის ციკლის ფერმენტები, ასევე მიტოქონდრიის მემკვიდრეობითი მასალა - დნმ, რნმ და ცილის სინთეზის აპარატი.
რისთვის არის მიტოქონდრია?
მიტოქონდრიის მთავარი ფუნქციაა ქიმიური ენერგიის უნივერსალური ფორმის - ატფ-ის სინთეზი. ისინი ასევე მონაწილეობენ ტრიკარბოქსილის მჟავას ციკლში, პირუვატსა და ცხიმოვან მჟავებს გარდაქმნიან აცეტილ-CoA-ად და შემდეგ აჟანგდებიან. ამ ორგანოოიდში მიტოქონდრიული დნმ ინახება და მემკვიდრეობით მიიღება, რომელიც კოდირებს tRNA, rRNA და ზოგიერთი ცილის რეპროდუქციას, რომელიც აუცილებელია მიტოქონდრიის ნორმალური ფუნქციონირებისთვის.
მიტოქონდრიაარის სპირალური, მომრგვალებული, წაგრძელებული ან განშტოებული ორგანელა.
მიტოქონდრიის კონცეფცია პირველად შემოგვთავაზა ბენდამ 1897 წელს. მიტოქონდრიების აღმოჩენა შესაძლებელია ცოცხალ უჯრედებში ფაზური კონტრასტული და ინტერფერენციული მიკროსკოპის გამოყენებით მარცვლების, გრანულების ან ძაფების სახით. ეს არის საკმაოდ მობილური სტრუქტურები, რომლებსაც შეუძლიათ გადაადგილება, ერთმანეთთან შერწყმა, გაზიარება. სინათლის მიკროსკოპით მკვდარ უჯრედებში სპეციალური მეთოდებით შეღებვისას, მიტოქონდრია ჰგავს ციტოპლაზმაში დიფუზურად განაწილებულ ან მის ზოგიერთ კონკრეტულ უბანში კონცენტრირებულ პატარა მარცვლებს (გრანულებს).
ჟანგბადის თანდასწრებით გლუკოზისა და ცხიმების განადგურების შედეგად მიტოქონდრიებში წარმოიქმნება ენერგია, ხოლო ორგანული ნივთიერებები გარდაიქმნება წყალში და ნახშირორჟანგად. სწორედ ამ გზით იღებენ ცხოველური ორგანიზმები სიცოცხლისთვის აუცილებელ ძირითად ენერგიას. ენერგია ინახება ადენოზინტრიფოსფატში (ATP), უფრო სწორად, მის მაკროერგიულ ობლიგაციებში. მიტოქონდრიების ფუნქცია მჭიდროდ არის დაკავშირებული ორგანული ნაერთების დაჟანგვასთან და მათი დაშლის დროს გამოთავისუფლებული ენერგიის გამოყენებასთან ATP მოლეკულების სინთეზისთვის. ამიტომ, მიტოქონდრიებს ხშირად უწოდებენ უჯრედის ენერგეტიკულ სადგურებს, ან უჯრედული სუნთქვის ორგანელებს. ATP მოქმედებს როგორც ენერგიის მიმწოდებელი თავისი ენერგიით მდიდარი ბოლო ფოსფატის ჯგუფის სხვა მოლეკულაზე გადაცემით და ამით გარდაიქმნება ADP-ად.
ვარაუდობენ, რომ ევოლუციაში მიტოქონდრია იყო პროკარიოტული მიკროორგანიზმები, რომლებიც გახდნენ სიმბიოტები უძველესი უჯრედის სხეულში. შემდგომში ისინი სასიცოცხლო მნიშვნელობის გახდა, რაც დაკავშირებული იყო დედამიწის ატმოსფეროში ჟანგბადის შემცველობის მატებასთან. ერთის მხრივ, მიტოქონდრიები აშორებდნენ უჯრედს ზედმეტ ტოქსიკურ ჟანგბადს, მეორე მხრივ კი ენერგიას აძლევდნენ.
მიტოქონდრიის გარეშე უჯრედს პრაქტიკულად არ შეუძლია გამოიყენოს ჟანგბადი, როგორც ნივთიერება, რომელიც უზრუნველყოფს ენერგომომარაგებას და შეუძლია მხოლოდ ანაერობული პროცესების მეშვეობით დააკმაყოფილოს მისი ენერგეტიკული საჭიროებები. ამრიგად, ჟანგბადი შხამია, მაგრამ უჯრედისთვის სასიცოცხლოდ აუცილებელი შხამი და ჟანგბადის სიჭარბე ისეთივე მავნეა, როგორც მისი დეფიციტი.
მიტოქონდრიებს შეუძლიათ შეცვალონ ფორმა და გადავიდნენ უჯრედის იმ ადგილებში, სადაც ისინი ყველაზე მეტად საჭიროა. ასე რომ, კარდიომიოციტებში მიტოქონდრია განლაგებულია მიოფიბრილების მახლობლად, თირკმლის მილაკების უჯრედებში, ბაზალური გამონაზარდების მახლობლად და ა.შ. უჯრედი შეიცავს ათასამდე მიტოქონდრიას და მათი რაოდენობა დამოკიდებულია უჯრედის აქტივობაზე.
მიტოქონდრიებს აქვთ საშუალო განივი ზომები 0,5…3 მკმ. ზომის მიხედვით იზოლირებულია მცირე, საშუალო, დიდი და გიგანტური მიტოქონდრია (ისინი ქმნიან ვრცელ ქსელს - მიტოქონდრიულ რეტიკულუმს). მიტოქონდრიების ზომა და რაოდენობა მჭიდრო კავშირშია უჯრედების აქტივობასთან და ენერგიის მოხმარებასთან. ისინი უკიდურესად ცვალებადია და, უჯრედის აქტივობის, ჟანგბადის შემცველობისა და ჰორმონალური ზემოქმედების მიხედვით, მათ შეუძლიათ შეშუპება, შეცვალონ კრისტალების რაოდენობა და სტრუქტურა, განსხვავდებოდეს რიცხვით, ფორმით და ზომით, ასევე ფერმენტული აქტივობით.
მიტოქონდრიების მოცულობის სიმკვრივე, მათი შიდა ზედაპირის განვითარების ხარისხი და სხვა მაჩვენებლები დამოკიდებულია უჯრედის ენერგეტიკულ საჭიროებებზე. ლიმფოციტებში მხოლოდ რამდენიმე მიტოქონდრიაა, ხოლო ღვიძლის უჯრედებში 2-3 ათასი მათგანია.
მიტოქონდრია შედგება მატრიქსის, შიდა მემბრანის, პერიმიტოქონდრიული სივრცისა და გარე გარსისგან. გარე მიტოქონდრიული მემბრანა გამოყოფს ორგანელას ჰიალოპლაზმისგან. ჩვეულებრივ მას აქვს თანაბარი კონტურები და დახურულია ისე, რომ ეს არის მემბრანული ჩანთა.
გარე მემბრანა გამოყოფილია შიდა მემბრანისგან 10-20 ნმ სიგანის პერიმიტოქონდრიული სივრცით. შიდა მიტოქონდრიული მემბრანა ზღუდავს მიტოქონდრიის - მატრიქსის რეალურ შინაგან შინაარსს. შიდა მემბრანა ქმნის უამრავ გამონაყარს მიტოქონდრიის შიგნით, რომლებიც ჰგავს ბრტყელ ქედებს ან კრისტალებს.
კრისტაების ფორმა შეიძლება იყოს ფირფიტების (ტრაბეკულარული) და მილაკების (მულტისიკულარული ჭრილზე) სახით და ისინი მიმართულია გრძივად ან განივი მიტოქონდრიის მიმართ.
თითოეული მიტოქონდრიონი ივსება მატრიცით, რომელიც ელექტრონულ მიკროგრაფებზე უფრო მკვრივი ჩანს, ვიდრე მიმდებარე ციტოპლაზმა. მიტოქონდრიული მატრიცა არის ერთგვაროვანი (ერთგვაროვანი), ზოგჯერ წვრილმარცვლოვანი, სხვადასხვა ელექტრონული სიმკვრივის. იგი ავლენს თხელ ძაფებს, რომელთა სისქეა დაახლოებით 2...3 ნმ და გრანულები ზომით დაახლოებით 15...20 ნმ. მატრიქსის ძაფები დნმ-ის მოლეკულებია, ხოლო მცირე გრანულები მიტოქონდრიული რიბოსომებია. მატრიცა შეიცავს ფერმენტებს, ერთჯაჭვიან, ციკლურ დნმ-ს, მიტოქონდრიულ რიბოზომებს, Ca 2+ იონს.
მიტოქონდრიის ცილის სინთეზის ავტონომიური სისტემა წარმოდგენილია ჰისტონებისგან თავისუფალი დნმ-ის მოლეკულებით. დნმ არის მოკლე, რგოლისებრი (ციკლური) და შეიცავს 37 გენს. ბირთვული დნმ-ისგან განსხვავებით, მასში პრაქტიკულად არ არსებობს არაკოდიციური ნუკლეოტიდური თანმიმდევრობები. სტრუქტურისა და ორგანიზაციის თავისებურებები აერთიანებს მიტოქონდრიის დნმ-ს ბაქტერიული უჯრედების დნმ-სთან. მიტოქონდრიის დნმ-ზე სინთეზირებულია სხვადასხვა ტიპის რნმ-ის მოლეკულები: ინფორმაციული, გადამტანი (სატრანსპორტო) და რიბოსომული. მიტოქონდრიული მესინჯერი რნმ არ განიცდის შერწყმას (იჭრება უბნები, რომლებიც არ ატარებენ ინფორმაციულ დატვირთვას). მიტოქონდრიული დნმ-ის მოლეკულების მცირე ზომა ვერ განსაზღვრავს ყველა მიტოქონდრიული ცილის სინთეზს. მიტოქონდრიული ცილების უმეტესობა უჯრედის ბირთვის გენეტიკური კონტროლის ქვეშ იმყოფება და სინთეზირდება ციტოპლაზმაში, რადგან მიტოქონდრიული დნმ სუსტად არის გამოხატული და შეუძლია უზრუნველყოს ჟანგვითი ფოსფორილირების ჯაჭვის ფერმენტების მხოლოდ ნაწილის ფორმირება. მიტოქონდრიული დნმ კოდირებს არაუმეტეს ათი ცილას, რომლებიც ლოკალიზებულია მემბრანებში და წარმოადგენს სტრუქტურულ ცილებს, რომლებიც პასუხისმგებელნი არიან მიტოქონდრიული მემბრანების ინდივიდუალური ფუნქციური ცილოვანი კომპლექსების სწორ ინტეგრაციაზე. ასევე სინთეზირებულია ცილები, რომლებიც ასრულებენ სატრანსპორტო ფუნქციებს. ცილის სინთეზის ასეთი სისტემა არ უზრუნველყოფს მიტოქონდრიის ყველა ფუნქციას, ამიტომ მიტოქონდრიის ავტონომია შეზღუდული და შედარებითია.
ძუძუმწოვრებში მიტოქონდრიები განაყოფიერებისას მხოლოდ კვერცხუჯრედის მეშვეობით გადადის, ხოლო სპერმატოზოიდები ახალ ორგანიზმში ბირთვული დნმ-ს მოაქვს.
მიტოქონდრიის მატრიცაში წარმოიქმნება რიბოსომები, რომლებიც განსხვავდება ციტოპლაზმის რიბოსომებისგან. ისინი მონაწილეობენ რიგი მიტოქონდრიული ცილების სინთეზში, რომლებიც არ არის კოდირებული ბირთვით. მიტოქონდრიულ რიბოსომებს აქვთ დანალექი რიცხვი 60 (განსხვავებით ციტოპლაზმური რიბოზომებისგან, რომელთა დანალექი რიცხვი 80-ია). დალექვის ნომერი არის დალექვის სიჩქარე ცენტრიფუგაციისა და ულტრაცენტრფუგაციის დროს. სტრუქტურის მიხედვით, მიტოქონდრიული რიბოსომები მსგავსია პროკარიოტული ორგანიზმების, მაგრამ უფრო მცირეა და განსხვავდება გარკვეული ანტიბიოტიკების მიმართ მგრძნობელობით (ლევომიცეტინი, ტეტრაციკლინი და ა.შ.).
შიდა მიტოქონდრიულ მემბრანას აქვს მაღალი სელექციურობის ხარისხი ნივთიერებების ტრანსპორტირებაში. მის შიდა ზედაპირზე მიმაგრებულია ოქსიდაციური ფოსფორილირების ჯაჭვის მჭიდროდ მიმდებარე ფერმენტები, ელექტრონის გადამზიდავი ცილები, სატრანსპორტო სისტემები ATP, ADP, პირუვატი და ა.შ.. კატალიზური პროცესების ეფექტურობა.
ელექტრონულმა მიკროსკოპმა გამოავლინა სოკოს ფორმის ნაწილაკები, რომლებიც ამოდიოდა მატრიცის სანათურში. მათ აქვთ ATP-სინთეზური (აყალიბებს ATP-ს ADP-დან) აქტივობა. ელექტრონების ტრანსპორტირება მიდის რესპირატორული ჯაჭვის გასწვრივ, ლოკალიზებულია შიდა მემბრანაში, რომელიც შეიცავს ოთხ დიდ ფერმენტულ კომპლექსს (ციტოქრომები). როდესაც ელექტრონები გადიან რესპირატორულ ჯაჭვში, წყალბადის იონები მატრიციდან ამოტუმბულია პერიმიტოქონდრიულ სივრცეში, რაც უზრუნველყოფს პროტონული გრადიენტის (ტუმბოს) წარმოქმნას. ამ გრადიენტის ენერგია (განსხვავებები ნივთიერებების კონცენტრაციაში და მემბრანული პოტენციალის ფორმირებაში) გამოიყენება ატფ-ის სინთეზისთვის და მეტაბოლიტებისა და არაორგანული იონების ტრანსპორტირებისთვის. გადამზიდავი ცილები, რომლებიც შეიცავს შიდა მემბრანას, გადააქვს ორგანულ ფოსფატებს, ATP, ADP, ამინომჟავებს, ცხიმოვან მჟავებს, ტრი- და დიკარბოქსილის მჟავებს.
მიტოქონდრიის გარე მემბრანა უფრო გამტარია დაბალმოლეკულური წონის ნივთიერებებისთვის, რადგან შეიცავს ბევრ ჰიდროფილურ ცილის არხს. სპეციფიკური რეცეპტორული კომპლექსები განლაგებულია გარე მემბრანაზე, რომლის მეშვეობითაც მატრიციდან ცილები ტრანსპორტირდება პერიმიტოქონდრიულ სივრცეში.
მისი ქიმიური შემადგენლობისა და თვისებების მიხედვით, გარე მემბრანა ახლოსაა სხვა უჯრედშიდა გარსებთან და პლაზმალემასთან. იგი შეიცავს ფერმენტებს, რომლებიც ახდენენ ცხიმების მეტაბოლიზებას, ააქტიურებენ (კატალიზებენ) ამინების, ამინოოქსიდაზას ტრანსფორმაციას. თუ გარე მემბრანის ფერმენტები აქტიური რჩება, მაშინ ეს არის მიტოქონდრიის ფუნქციური უსაფრთხოების მაჩვენებელი.
მიტოქონდრიას აქვს ორი ავტონომიური ქვეგანყოფილება. მაშინ, როცა პერიმიტოქონდრიული სივრცე, ან მიტოქონდრიის გარე კამერა (გარე ქვეგანყოფილება) იქმნება ჰიალოპლაზმური ცილის კომპლექსების შეღწევის გამო, შიდა ქვეგანყოფილება (მიტოქონდრიული მატრიცა) ნაწილობრივ წარმოიქმნება მიტოქონდრიული დნმ-ის სინთეზური აქტივობის გამო. შიდა ქვეგანყოფილება (მატრიცა) შეიცავს დნმ-ს, რნმ-ს და რიბოზომებს. გამოირჩევა Ca 2+ იონების მაღალი დონით ჰიალოპლაზმასთან შედარებით. წყალბადის იონები გროვდება გარე ქვეგანყოფილებაში. გარე და შიდა ქვეგანყოფილებების ფერმენტული აქტივობა, ცილების შემადგენლობა მნიშვნელოვნად განსხვავდება. შიდა ქვეგანყოფილებას აქვს უფრო მაღალი ელექტრონის სიმკვრივე, ვიდრე გარე.
მიტოქონდრიის სპეციფიკური მარკერებია ფერმენტები ციტოქრომ ოქსიდაზა და სუქცინატდეჰიდროგენაზა, რომელთა გამოვლენა შესაძლებელს ხდის მიტოქონდრიაში ენერგეტიკული პროცესების რაოდენობრივ განსაზღვრას.
მიტოქონდრიის ძირითადი ფუნქცია- ატფ სინთეზი. თავდაპირველად, შაქრები (გლუკოზა) ნადგურდება ჰიალოპლაზმაში რძის და პირუვიტის მჟავებამდე (პირუვატი) მცირე რაოდენობით ატფ-ის ერთდროული სინთეზით. ერთი გლუკოზის მოლეკულის გლიკოლიზის შედეგად გამოიყენება ორი ATP მოლეკულა და იქმნება ოთხი. ამრიგად, მხოლოდ ორი ATP მოლეკულა ქმნის დადებით ბალანსს. ეს პროცესები მიმდინარეობს ჟანგბადის გარეშე (ანაერობული გლიკოლიზი).
ენერგიის წარმოების ყველა შემდგომი ეტაპი ხდება აერობული დაჟანგვის პროცესში, რაც უზრუნველყოფს დიდი რაოდენობით ატფ-ის სინთეზს. ამ შემთხვევაში, ორგანული ნივთიერებები განადგურებულია CO 2-მდე და წყალში. დაჟანგვას თან ახლავს პროტონების გადაცემა მათ მიმღებებში. ეს რეაქციები ტარდება ტრიკარბოქსილის მჟავას ციკლის მთელი რიგი ფერმენტების დახმარებით, რომლებიც განლაგებულია მიტოქონდრიულ მატრიქსში.
ელექტრონების გადაცემის და მასთან დაკავშირებული ADP ფოსფორილირების სისტემები (ჟანგვითი ფოსფორილირება) ჩაშენებულია კრისტალების გარსებში. ამ შემთხვევაში, ელექტრონები გადადის ერთი ელექტრონის მიმღები ცილიდან მეორეში და ბოლოს, ისინი ჟანგბადს უკავშირდება, რის შედეგადაც წარმოიქმნება წყალი. ამავდროულად, ელექტრონის სატრანსპორტო ჯაჭვში ასეთი დაჟანგვის დროს გამოთავისუფლებული ენერგიის ნაწილი ინახება მაკროერგიული ბმის სახით ADP ფოსფორილირების დროს, რაც იწვევს დიდი რაოდენობით ATP მოლეკულების წარმოქმნას - მთავარი უჯრედშიდა ენერგიის ეკვივალენტს. მიტოქონდრიული კრისტალების მემბრანებზე აქ მდებარე ჟანგვის ჯაჭვის ცილების და ADP-ATP სინთეტაზას ფოსფორილირების ფერმენტის დახმარებით მიმდინარეობს ჟანგვითი ფოსფორილირების პროცესი. ჟანგვითი ფოსფორილირების შედეგად ერთი გლუკოზის მოლეკულისგან წარმოიქმნება ატფ-ის 36 მოლეკულა.
მიტოქონდრიებს აქვთ სპეციალიზებული (აფინირებული) რეცეპტორები გარკვეული ჰორმონებისა და ნივთიერებების მიმართ. ტრიიოდთირონინი ჩვეულებრივ აჩქარებს მიტოქონდრიის სინთეზურ აქტივობას. ინტერლეუკინ-1 და ტრიიოდოთირონინის მაღალი კონცენტრაცია წყვეტს ჟანგვითი ფოსფორილირების ჯაჭვებს, იწვევს მიტოქონდრიის შეშუპებას, რასაც თან ახლავს თერმული ენერგიის გამომუშავების ზრდა.
ახალი მიტოქონდრიები წარმოიქმნება გაყოფით, შეკუმშვით ან კვირტით. ამ უკანასკნელ შემთხვევაში წარმოიქმნება პროტომიტოქონდრიონი, რომელიც თანდათან იზრდება ზომაში.
პროტომიტოქონდრიონი არის პატარა ორგანელა გარე და შიდა გარსებით. შიდა გარსს არ აქვს ან შეიცავს განუვითარებელ კრისტას. ორგანელას ახასიათებს აერობული ფოსფორილირების დაბალი დონე. როდესაც შეკუმშვა იქმნება, მიტოქონდრიის შიგთავსი ნაწილდება ორ ახალ საკმაოდ დიდ ორგანელებს შორის. გამრავლების ნებისმიერი მეთოდით, თითოეულ ახლად წარმოქმნილ მიტოქონდრიას აქვს საკუთარი გენომი.
ძველი მიტოქონდრია ნადგურდება აუტოლიზით (უჯრედის მიერ თვითმონელება ლიზოსომების დახმარებით) აუტოლიზოსომების წარმოქმნით. ნარჩენი სხეული იქმნება აუტოლიზოსომისგან. სრული მონელებით, ნარჩენი სხეულის შიგთავსი, რომელიც შედგება დაბალი მოლეკულური წონის ორგანული ნივთიერებებისგან, გამოიყოფა ეგზოციტოზით. არასრული მონელების შემთხვევაში, მიტოქონდრიული ნარჩენები შეიძლება დაგროვდეს უჯრედში ფენოვანი სხეულების ან გრანულების სახით ნიპოფუსცინით. მიტოქონდრიის ნაწილში უხსნადი კალციუმის მარილები გროვდება კრისტალების - კალციფიკაციების წარმოქმნით. მიტოქონდრიული დეგენერაციის პროდუქტების დაგროვებამ შეიძლება გამოიწვიოს უჯრედების დისტროფია.
მიტოქონდრია (MT) ჩემთვის კვლევის ერთ-ერთი ყველაზე საინტერესო სფეროა. დაახლოებით 1,6 მილიარდი წლის წინ ენდოსიმბიოზის დროს მიტოქონდრიის სხვა უჯრედთან კავშირი გახდა რთული სტრუქტურის მქონე ყველა მრავალუჯრედიანი ევკარიოტის საფუძველი. სავარაუდოდ, მიტოქონდრია წარმოიშვა α-პროტეობაქტერიების მსგავსი უჯრედებიდან.
მიტოქონდრიის საუკეთესო ბოლო მიმოხილვა არის ვერნერ კულბრანდტის "მიტოქონდრიული მემბრანის ცილოვანი კომპლექსების სტრუქტურა და ფუნქცია". თუ იცით ინგლისური და გაინტერესებთ ამ ორგანელების აგებულება, გირჩევთ წაიკითხოთ. ეს სტატია იმდენად კარგია, რომ ადვილად შეიძლება იყოს კარგი მოლეკულური ბიოლოგიის სახელმძღვანელოს ხელმძღვანელი. თავიდან მთელი სტატიის თარგმნა მინდოდა, მაგრამ ამას უპატიებლად დიდი დრო დასჭირდებოდა და სხვა რამეს მაშორებდა. ამიტომ შემოვიფარგლები თეზისებითა და სურათებით. პერიოდულად გააზავეთ ყველაფერი თქვენი ფიქრებით.
თავად მიტოქონდრია აკოდირებს მხოლოდ 13 ცილას, მიუხედავად უჯრედისგან განცალკევებული დნმ-ის (mtDNA) არსებობისა და ცილის ტრანსკრიფციისთვის მთელი „წარმოების“ ციკლისთვის. იზოლირებულ მიტოქონდრის შეუძლია შეინარჩუნოს თავისი შემადგენლობა და ფუნქცია გარკვეული დროის განმავლობაში.
სურათი 1. მიტოქონდრიული მემბრანის კომპონენტები. გარე მემბრანა გამოყოფს მიტოქონდრიას ციტოპლაზმისგან. ის გარს აკრავს შიდა მემბრანას, რომელიც გამოყოფს მემბრანთაშორის სივრცეს ცილებით მდიდარი ცენტრალური მატრიცისგან. შიდა მემბრანა იყოფა შიდა შემზღუდველ გარსად და კრისტაებად. ეს ორი ნაწილი უწყვეტია კრისტას მიმაგრების წერტილებში (cristae შეერთება). კრისტაები მეტ-ნაკლებად ღრმად ვრცელდება მატრიქსში და წარმოადგენს მიტოქონდრიული ენერგიის გადაქცევის მთავარ ადგილს. მცირე პროტონული გრადიენტი მემბრანთაშორის სივრცეში (pH7.2-7.4) და მატრიცა (pH7.9-8.0) იწვევს ატფ-ს წარმოქმნას ატფ-სინთაზას მიერ კრისტალების მემბრანებში.
გარე მემბრანა ფოროვანია და ციტოპლაზმიდან ნივთიერებებს გავლის საშუალებას აძლევს. შიდა მემბრანა მკვრივია, მის გადაკვეთას სატრანსპორტო ცილებია საჭირო [გილბერტ ლინგი გონივრულად არ ეთანხმება], ბარიერის უწყვეტობა საშუალებას აძლევს შიდა მემბრანას ჰქონდეს ელექტროქიმიური პოტენციალი -180 მვ. მატრიცას აქვს საკმაოდ მაღალი pH (7.9-8). კიდევ ერთხელ ჩავუღრმავდები ლინგს. ტუტე (7-ზე მეტი) pH ხელს უწყობს უფრო გაშლილი ცილის კონფორმაციას. მაღალი pH არღვევს წყალბადისა და მარილის ობლიგაციებს, აქცევს პოლარიზებულ CO და NH-ს წყლის მოლეკულებისთვის ხელმისაწვდომს, რითაც იზრდება ყველა უჯრედშიდა წყლის დიპოლური მომენტი და აკავშირებს მას. ამ ვენაში მემბრანის არსებობა არ არის საჭირო უჯრედის შიგნით პროტოპლაზმის „შესანარჩუნებლად“ (ამას თავად ცილები აკეთებენ მაღალი pH-ზე), არამედ პოტენციალის ქონა.
mtDNA გვხვდება ნუკლეოტიდებში, რომლებიც შეადგენს დაახლოებით 1000 უჯრედს. მატრიქსის ცილის სიმკვრივე საკმაოდ მაღალია (500 მგ/მლ-მდე), რაც ახლოსაა კრისტალიზებულ ცილებთან.
შიდა მემბრანა აყალიბებს ინვაგინაციებს, სახელწოდებით cristae, რომლებიც ღრმად აღწევს მატრიქსში. cristae განსაზღვრავს მიტოქონდრიის მესამე "კუპეს" - cristae-ის სანათურს (cristae lumen). კრისტას მემბრანები შეიცავს ელექტრონის ტრანსპორტირების ჯაჭვისა და ატფ სინთაზას, თუ არა ყველა, სრულად „აწყობილ“ კომპლექსებს. კრისტას სანათური შეიცავს დიდი რიცხვიმცირე ხსნადი ცილის ელექტრონის მატარებელი (ციტოქრომი c). ამრიგად, მიტოქონდრიული კრისტაები არის ბიოლოგიური ენერგიის გარდაქმნის მთავარი ადგილი ყველა არაფოტოსინთეზურ ევკარიოტში.
ბევრი საინტერესო რამ არის კრისტალებთანაც. კრისტას ოპტიკური თვისებები გავლენას ახდენს ქსოვილებში სინათლის გავრცელებასა და წარმოქმნაზე. მე კი წავაწყდი იდეებს, რომ კრისტასების ზედაპირი მსგავსია (ვარაუდი) ტოპოლოგიური იზოლატორების ზედაპირების (ზეგამტარობა მუხტის გაფანტვის გარეშე იგულისხმებოდა).

სურათი 2. რესპირატორული ჯაჭვის მემბრანული ცილოვანი კომპლექსები. კომპლექსიმე (NADH/უბიქინონ ოქსიდორედუქტაზა, ლურჯი), კომპლექსიII(სუქცინატდეჰიდროგენაზა, ვარდისფერი), კომპლექსიIII(ციტოქრომ C რედუქტაზა, ფორთოხალი), კომპლექსიIV(ციტოქრომ C ოქსიდაზა, მწვანე) და მიტოქონდრიული ატფ სინთაზა (ცნობილია როგორც კომპლექსივ, კრემისფერი) ერთად მუშაობენ ჟანგვითი ფოსფორილირების დროს, რათა უჯრედებმა ენერგია გამოიყენონ. კომპლექსებიმე, III, IVამოტუმბავს პროტონებს კრისტა მემბრანის გასწვრივ, რაც ქმნის პროტონულ გრადიენტს, რომელიც ასტიმულირებს ATP სინთეზს.
ახლა ცოტა ყურადღება II კომპლექსს. გახსოვთ, რომ ცხიმი (კეტო) ხაზს უსვამს FADH2 და II კომპლექსს მეტაბოლიზმში. ისინი აღადგენენ CoQ წყვილს, რაღაც მომენტში დაჟანგული CoQ არ არის საკმარისი ელექტრონების III კომპლექსში გადასატანად და აყალიბებს ელექტრონების საპირისპირო ნაკადს I კომპლექსში სუპეროქსიდის წარმოქმნით. ხანგრძლივი HFLC კვებით, I კომპლექსი შექცევადად განადგურდება, მაშინ როცა ეს ნორმალური ფიზიოლოგიური ოპტიმიზაციაა.
ასევე გთხოვთ, გაითვალისწინოთ, რომ II კომპლექსი არ ამოტუმბავს პროტონებს. რა ანაწილებს ჩვენში პროტონულ გრადიენტს, არღვევს ფოსფორილირებას და ასტიმულირებს ცხიმების დაწვას სითბოსთვის? მართალია, ცივი სტრესი. თერმოგენეზი დაკავშირებულია მეტაბოლიზმთან კომპლექსის საშუალებით, რომელიც არ შლის პროტონებს, რითაც არ იძლევა დამატებით პროტონებს ATP სინთეზისთვის. შეიძლება მხოლოდ გაინტერესებდეს, რამდენად საოცრად არის გააზრებული ჩვენი ორგანიზმი.
კრისტალის სამაგრები დაMICOS
Cristae-ის შეერთებები არის პატარა მრგვალი ხვრელები, რომელთა დიამეტრი დაახლოებით 25 ნმ. ყველა ორგანიზმის მიტოქონდრიას აქვს MICOS (მიტოქონდრიის კონტაქტის ადგილი და კრისტაები გარე მემბრანასთან) სისტემა, ხუთი მემბრანის და ერთი ხსნადი ცილის ერთობლიობა, რომლებიც ამაგრებენ კრისტას გარე მემბრანაზე.
გაზრდილი ენერგიის მოთხოვნილების მქონე უჯრედებში, როგორიცაა ჩონჩხი და გულის კუნთები, კრისტაები მჭიდროდ ავსებენ მიტოქონდრიული მოცულობის უმეტეს ნაწილს. ქვედა ენერგეტიკული მოთხოვნილების მქონე ქსოვილებში, როგორიცაა ღვიძლი და თირკმელები, კრისტაები არ არის ერთმანეთთან მჭიდროდ შეფუთული. მატრიცაში მეტი ადგილია ბიოსინთეზური ფერმენტებისთვის.
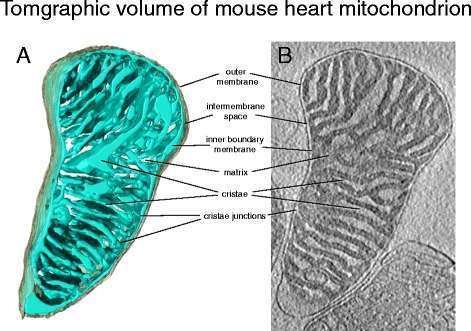
სურათი 3. თაგვის გულის მიტოქონდრიის ტომოგრაფიული მოცულობა. ა) თაგვის გულის მიტოქონდრიის სამგანზომილებიანი მოცულობა, დაფიქსირებული კრიო-ET-ით. გარე მემბრანა (ნაცრისფერი) ფარავს შიდა გარსს (ღია ლურჯი). შიდა გარსი მჭიდროდ ივსება კრისტალებით ბ) მოცულობის ტომოგრაფიული მონაკვეთი. მჭიდროდ შევსებული მატრიცა, რომელიც შეიცავს მიტოქონდრიული ცილების უმეტესობას, ბნელი ჩანს ელექტრონულ მიკროსკოპზე. მიუხედავად იმისა, რომ მემბრანთაშორისი სივრცე და კრისტას სანათურები მსუბუქად გამოიყურება ცილების დაბალი კონცენტრაციის გამო.
ატფ სინთაზას დიმერები
მიტოქონდრიული F1-F0 ატფ სინთაზა არის კრისტას ყველაზე გამორჩეული ცილოვანი კომპლექსი. ATP synthase არის უძველესი ნანომანქანა, რომელიც იყენებს ელექტროქიმიურ პროტონულ გრადიენტს შიდა მემბრანის გარშემო, რათა შექმნას AFT ბრუნვითი კატალიზის გზით. მემბრანის F0 კომპლექსში მოძრავი პროტონები ატრიალებენ 8 (ძუძუმწოვრებში) ან 10 (საფუარში) c-კვანძის როტორს. ცენტრალური ღერო გადასცემს c-rotor ბრუნვას F1 კატალიზურ თავში, სადაც ATP წარმოიქმნება ADP-დან და ფოსფატიდან კონფორმაციული ცვლილებების თანმიმდევრობით. პერიფერიული ღერო ხელს უშლის F1 Head-ის არაპროდუქტიულ ბრუნვას F0 კომპლექსის წინააღმდეგ.
მრავალი წლის განმავლობაში ითვლებოდა, რომ ატფ სინთაზა შემთხვევით მდებარეობდა შიდა მემბრანაზე. მაგრამ აღმოჩნდა რომ ატფ სინთაზა განლაგებულია ორმაგი რიგებად. უფრო მეტიც, ATP სინთაზას ხაზოვანი რიგები ყველა ცოცხალი მიტოქონდრიის ფუნდამენტური ატრიბუტია.

სურათი 4. ATP სინთაზას ორმაგი რიგები შვიდ სხვადასხვა სახეობაში.
ატფ სინთაზას რიგები ძირითადად განლაგებულია კრისტალების ქედების გასწვრივ. დიმერები ახვევენ ლიპიდურ ორშრს და შედეგად თვითორგანიზება ხდება რიგებად. როდესაც APT სინთაზას e და g კვანძები ამოვარდნენ საფუარის მიტოქონდრიებში, შტამი იზრდებოდა 60%-ით ნელა, ვიდრე მისი ველური კოლეგები და მათი მიტოქონდრიების მემბრანული პოტენციალი განახევრდა. პროკარიოტულ APT სინთაზას აკლია რამდენიმე დიმერთან ასოცირებული კვანძი და დიმერის რიგები არ არის ნაპოვნი ბაქტერიებსა და არქეებში. ამგვარად, AFT სინთაზას დიმერების კრისტაები და რიგები ადაპტაციას წარმოადგენს სხეულის მაღალი ენერგიის მოთხოვნილებებთან.

სურათი 5. პოლიმელა სპ მიტოქონდრიის ATP სინთაზას დიმერის სტრუქტურა. ATP სინთაზას V- ფორმის დიმერის გვერდითი ხედი.
რესპირატორული ჯაჭვის კომპლექსები და სუპერკომპლექსები
პროტონის გრადიენტი შიდა მემბრანის ირგვლივ იქმნება სამი დიდი მემბრანის კომპლექსით, რომლებიც ცნობილია როგორც I, კომპლექსი III და IV კომპლექსი (იხ. სურათი 2). I კომპლექსი იკვებება NADH-ის ელექტრონებით, ენერგია, რომელიც გამოიყოფა ელექტრონის შემოწირულობისას, გამოყოფს ოთხ პროტონს. III კომპლექსი იღებს ელექტრონს შემცირებული ქინოლიდან და გადასცემს მას ელექტრონის მატარებელში (ციტოქრომი c), რის შედეგადაც ამოტუმბავს ერთ პროტონს. IV კომპლექსი იღებს ელექტრონს ციტოქრომ c-დან და ჩუქნის მას მოლეკულურ ჟანგბადს, ამოტუმბავს 4 პროტონს წყალში გარდაქმნილი ჟანგბადის თითოეული მოლეკულისთვის. კომპლექსი II არ ამოტუმბავს პროტონებს, პირდაპირ აძლევს ელექტრონებს ქინოლს. როგორ უკავშირდება ელექტრონის გადატანა NADH-დან ქინოლში პროტონის ტრანსლოკაციას, ჯერჯერობით უცნობია. I კომპლექსი უფრო დიდია ვიდრე III და IV ერთად.

სურათი 6. მსხვილფეხა რქოსანი გულის მიტოქონდრიების I კომპლექსი. მატრიცის ნაწილი შეიცავს რვა რკინა-გოგირდის (Fe-S) ჯგუფს, რომლებიც მიმართავენ ელექტრონებს NADH-დან ქინოლში მატრიცა-მემბრანის შეერთებაზე. მემბრანის ნაწილი შედგება 78 პირისგან, მოლეკულების ჩათვლით, რომლებიც ამოტუმბავს პროტონებს.
კომპლექსებიმეIII დაIV გაერთიანდება სუპერკომპლექსებადან რესპირაზომი. პურის საფუარში ( საქარომიცებიcerevisiae) არ არსებობს I კომპლექსი, მათი სუპერკომპლექსები შედგება III და IV-სგან. სუპერკომპლექსების როლი ჯერ არ არის ნათელი. ითვლება, რომ ეს ელექტრონების ტრანსპორტირებას უფრო ეფექტურს ხდის, მაგრამ ამის პირდაპირი მტკიცებულება ჯერ არ არსებობს.

სურათი 7. მსხვილფეხა რქოსანი გულის მიტოქონდრიული სუპერკომპლექსი. ყურადღება მიაქციეთ მანძილს I და III კომპლექსებს შორის, რომელიც აუცილებლად უნდა გაკეთდეს ქინოლით. ისრები აჩვენებს ელექტრონის მოძრაობას სუპერკომპლექსში.
კრისტას სანათურში მთავარი ცილაა ციტოქრომი c, რომელიც ელექტრონს III კომპლექსიდან IV კომპლექსში გადააქვს. თუ ციტოქრომი c გამოიყოფა უჯრედის ციტოპლაზმაში, ეს იწვევს აპოპტოზს.

სურათი 8. ატფ სინთაზას დიმერების რიგები განსაზღვრავს კრისტას ფორმას. კრისტას ქედზე AFT სინთაზა (ყვითელი) ქმნის პროტონების შერწყმას (წითელი), ელექტრონული წრედის პროტონული ტუმბოები (მწვანე) არის დიმერების რიგების ორივე მხარეს. პროტონების წყაროდან ატფ-სინთაზასკენ მიმართვით, კრისტაები მოქმედებენ როგორც პროტონების სახელმძღვანელო, რათა უზრუნველყონ ATP ეფექტური წარმოქმნა. წითელი ისრები აჩვენებს პროტონული ნაკადის მიმართულებას.
მემბრანის რეორგანიზაცია დაბერების დროს
დაბერება ფუნდამენტური და ცუდად გაგებული პროცესია ყველა ევკარიოტში. სოკოებზე შეისწავლეს მიტოქონდრიული დაბერება პოდოსპორა ანსერინარომლებიც ცხოვრობენ მხოლოდ 18 დღე. ნორმალურ მიტოქონდრიებში კრისტაები ღრმად ვრცელდება მატრიქსში. ამისათვის საჭიროა ATP სინთაზას დიმერების რიგები და MICOS კომპლექსი კრისტალების მიმაგრების ადგილებში. ასაკთან ერთად, კრისტაები იწყებენ მემბრანის ზედაპირთან მიახლოებას, APT სინთაზას დიმერები გადაიქცევა მონომერებად და ყველაფერი მთავრდება ციტოქრომის c გამოთავისუფლებით და უჯრედის სიკვდილით.
ელექტრონის ტრანსპორტი ქმნის სუპეროქსიდს I და III კომპლექსებში. ეს არის ნივთიერებათა ცვლის გვერდითი პროდუქტი. აუცილებელი და ამავე დროს სასიკვდილო. დაბერების დროს, დაშლა იწყებს უპირატესობას შერწყმაზე. ეს ხელს უშლის დაზიანებული მიტოქონდრიების "გადარჩენას" შერწყმის შედეგად და აჩქარებს გარდაუვალს.

სურათი 9. ცვლილებები შიდა მემბრანის მორფოლოგიაში და ატფ-სინთაზას დიმერების მიტოქონდრიული დაბერების დროს.
როგორც ხედავთ, მომავალი კვლევისთვის ბევრი სფეროა. მე ვვარაუდობ ფიზიკისა და ბიოლოგიის კვეთაზე, სადაც ფიზიკოსები შეეცდებიან ახსნან, რატომ არის ასეთი სტრუქტურა უფრო ენერგოეფექტური. განსაკუთრებით პუბლიკაციები კვანტურ ბიოლოგიაზე
მიტოქონდრია
მიტოქონდრია არის ღეროს ფორმის ან ოვალური ფორმის სტრუქტურები (გრ. მიტოები- ძაფი, ქონდროსი- გრანულები). ისინი გვხვდება ყველა ცხოველურ უჯრედში (მომწიფებული ერითროციტების გამოკლებით): მაღალ მცენარეებში, წყალმცენარეებში და პროტოზოებში. ისინი არ არის მხოლოდ ბაქტერიების პროკარიოტებში.
ეს ორგანელები პირველად აღმოაჩინა და აღწერა გასული საუკუნის ბოლოს ალტმანმა. ცოტა მოგვიანებით, ამ სტრუქტურებს მიტოქონდრია უწოდეს. 1948 წელს ჰოგებუმმა მიუთითა მიტოქონდრიის, როგორც უჯრედული სუნთქვის ცენტრის მნიშვნელობაზე, ხოლო 1949 წელს კენედიმ და ლენინგერმა დაადგინეს, რომ მიტოქონდრიებში ხდება ოქსიდაციური ფოსფორილირების ციკლი. ასე რომ, დადასტურდა, რომ მიტოქონდრია ემსახურება ენერგიის წარმოქმნის ადგილს.
მიტოქონდრია ხილულია ჩვეულებრივი სინათლის მიკროსკოპით სპეციალური შეღებვის მეთოდებით. ფაზაში - კონტრასტული მიკროსკოპით და "ბნელ ველში" მათი დაკვირვება შესაძლებელია ცოცხალ უჯრედებში.
სტრუქტურა, ზომები, ფორმა მიტოქონდრია ძალიან ცვალებადია. ეს დამოკიდებულია პირველ რიგში უჯრედების ფუნქციურ მდგომარეობაზე. მაგალითად, დადგინდა, რომ ბუზების მოძრაობის ნეირონებში, რომლებიც 2 საათის განმავლობაში უწყვეტად დაფრინავენ, ჩნდება სფერული მიტოქონდრიების უზარმაზარი რაოდენობა, ხოლო წებოვანი ფრთებით ბუზებში, მიტოქონდრიების რაოდენობა გაცილებით ნაკლებია და მათ აქვთ ღეროს ფორმის ( ლ.ბ.ლევინსონი). ფორმის მიხედვით, ისინი შეიძლება იყოს ძაფისებრი, ღეროს ფორმის, მრგვალი და ჰანტელის ფორმის, თუნდაც იმავე უჯრედში.
მიტოქონდრია ლოკალიზებულია უჯრედში, როგორც წესი, ან იმ ადგილებში, სადაც ენერგია მოიხმარება, ან სუბსტრატის დაგროვების მახლობლად (მაგალითად, ლიპიდური წვეთები), ასეთის არსებობის შემთხვევაში.
მიტოქონდრიების მკაცრი ორიენტაცია გვხვდება სპერმატოზოიდების დროშების გასწვრივ, განივზოლიან კუნთოვან ქსოვილში, სადაც ისინი განლაგებულია მიოფიბრილების გასწვრივ, თირკმლის მილაკების ეპითელიუმში ისინი ლოკალიზებულია სარდაფის მემბრანის ინვაგინაციებში და ა.
უჯრედებში მიტოქონდრიების რაოდენობას აქვს ორგანოს მახასიათებლები, მაგალითად, ვირთხის ღვიძლის უჯრედები შეიცავს 100-დან 2500-მდე მიტოქონდრიას, ხოლო თირკმლის შემგროვებელი სადინარების უჯრედებში - 300, სპერმატოზოიდების სხვადასხვა სახეობის ცხოველებში 20-დან 72-მდე. გიგანტური ამება ქაოსი ქაოსიმათი რიცხვი 500 000-ს აღწევს.მიტოქონდრიების ზომები მერყეობს 1-დან 10 მიკრონიმდე.
მიტოქონდრიების ულტრამიკროსკოპული სტრუქტურა იგივეა, მიუხედავად მათი ფორმისა და ზომისა. ისინი დაფარულია ორი ლიპოპროტეინის გარსით: გარე და შიდა. მათ შორის არის მემბრანთაშორისი სივრცე.
მიტოქონდრიულ სხეულში ამოვარდნილი შიდა მემბრანის ინვაგინაციები ეწოდება cristae. კრისტალების განლაგება მიტოქონდრიაში შეიძლება იყოს განივი ან გრძივი. კრისტას ფორმა შეიძლება იყოს მარტივი ან განშტოებული. ზოგჯერ ისინი ქმნიან რთულ ქსელს. ზოგიერთ უჯრედში, მაგალითად, თირკმელზედა ჯირკვლის გლომერულური ზონის უჯრედებში, კრისტაები ტუბულებს ჰგავს. კრისტატების რაოდენობა პირდაპირპროპორციულია მიტოქონდრიებში მიმდინარე ჟანგვითი პროცესების ინტენსივობისა. მაგალითად, კარდიომიოციტების მიტოქონდრიებში მათი რაოდენობა რამდენჯერმე მეტია, ვიდრე ჰეპატოციტების მიტოქონდრიაში. შიდა მემბრანით შემოსაზღვრული სივრცე ქმნის მიტოქონდრიის შიდა კამერას. მასში, კრისტებს შორის, არის მიტოქონდრიული მატრიცა - შედარებით ელექტრონულად მკვრივი ნივთიერება.
შიდა მემბრანის ცილები სინთეზირდება მიტორიბოსომებით, ხოლო გარე მემბრანის ცილები სინთეზირებულია ციტორიბოსომებით.
"მიტოქონდრიის გარე მემბრანა მრავალი თვალსაზრისით მსგავსია EPS მემბრანების. ის ღარიბია ჟანგვითი ფერმენტებით. მემბრანულ სივრცეში ისინი ცოტაა. მაგრამ შიდა მემბრანა და მიტოქონდრიული მატრიცა ფაქტიურად გაჯერებულია მათით. ამრიგად, კრებსის ციკლის ფერმენტები და ცხიმოვანი მჟავების დაჟანგვა კონცენტრირებულია მიტოქონდრიულ მატრიქსში, შიგნიდან მემბრანა ლოკალიზებულია ელექტრონული სატრანსპორტო ჯაჭვით, ფოსფორილირების ფერმენტებით (ატფ-ის ფორმირება ADP-დან), მრავალი სატრანსპორტო სისტემა.
ცილისა და ლიპიდების გარდა, მიტოქონდრიული მემბრანების შემადგენლობაში შედის რნმ, დნმ, ამ უკანასკნელს აქვს გენეტიკური სპეციფიკა და განსხვავდება ბირთვული დნმ-ისგან თავისი ფიზიკურ-ქიმიური თვისებებით.
ელექტრონულ მიკროსკოპულმა კვლევებმა აჩვენა, რომ გარე მემბრანის ზედაპირი დაფარულია პატარა სფერული ელემენტარული ნაწილაკებით. შიდა გარსი და კრისტაები შეიცავს მსგავს ელემენტარულ ნაწილაკებს „ფეხებზე“, ეგრეთ წოდებულ სოკოს სხეულებზე. ისინი შედგება სამი ნაწილისგან: სფერული თავი (დიამეტრი 90-100 A°), ცილინდრული ფეხები, 5 ნმ სიგრძისა და 3-4 ნმ სიგანისა და ფუძის ზომები 4 11 ნმ. სოკოს სხეულის თავები დაკავშირებულია ფოსფორილირებასთან, შემდეგ აღმოჩენილია, რომ თავები შეიცავს ATP-id აქტივობის მქონე ფერმენტს.
მემბრანთაშორის სივრცეში არის ნივთიერება, რომლის ელექტრონის სიმკვრივე უფრო დაბალია, ვიდრე მატრიცა. იგი უზრუნველყოფს კომუნიკაციას მემბრანებს შორის და ამარაგებს ორივე მემბრანაში მდებარე ფერმენტებს დამხმარე კატალიზატორები-კოფერმენტებით.
ახლა ცნობილია, რომ მიტოქონდრიის გარე მემბრანა ძალზე გამტარია დაბალი მოლეკულური წონის მქონე ნივთიერებებისთვის, კერძოდ, ცილოვანი ნაერთებისთვის. მიტოქონდრიის შიდა მემბრანა შერჩევითად გამტარია. იგი პრაქტიკულად შეუვალია ანიონებისთვის (Cl-1, Br-1, SO 4-2, HCO 3-1, Sn +2, Mg +2 კატიონები, მრავალი შაქარი და ამინომჟავების უმეტესობა, ხოლო Ca 2+, Mn2 + , ფოსფატი , პოლიკარბოქსილის მჟავები ადვილად შეაღწევს მასში. არსებობს მტკიცებულება, რომ შიდა მემბრანაში არსებობს შეღწევადი ანიონებისა და კათიონების ცალკეული ჯგუფებისთვის დამახასიათებელი რამდენიმე მატარებელი. მემბრანებში ნივთიერებების აქტიური ტრანსპორტირება ხდება ენერგიის გამოყენების გამო. ატფ-აზა სისტემის ან მემბრანაზე წარმოქმნილი ელექტრული პოტენციალის მუშაობის რესპირატორული ჯაჭვის შედეგად.მიტოქონდრიებში სინთეზირებულ ატფ-საც კი შეუძლია გამოვიდეს გადამზიდველის (დაწყვილებული ტრანსპორტი) დახმარებით.
მიტოქონდრიული მატრიცა წარმოდგენილია წვრილმარცვლოვანი ელექტრონულად მკვრივი ნივთიერებით. იგი შეიცავს მიტორიბოზომებს, ფიბრილარულ სტრუქტურებს, რომლებიც შედგება დნმ-ის მოლეკულებისგან და 200A-ზე მეტი დიამეტრის მქონე გრანულებისაგან ◦ წარმოქმნილი მარილებით: Ca 3 (PO 4), Ba 3 (PO 4) 2, Mg 3 (PO 4) . ითვლება, რომ გრანულები ემსახურება Ca +2 და Mg +2 იონების რეზერვუარს. მათი რიცხვი იზრდება მიტოქონდრიული მემბრანების გამტარიანობის ცვლილებებით.
მიტოქონდრიებში დნმ-ის არსებობა უზრუნველყოფს მიტოქონდრიის მონაწილეობას რნმ-ის და სპეციფიკური ცილების სინთეზში და ასევე მიუთითებს ციტოპლაზმური მემკვიდრეობის არსებობაზე. თითოეული მიტოქონდრიონი შეიცავს, მისი ზომის მიხედვით, დნმ-ის ერთ ან მეტ მოლეკულას (2-დან 10-მდე). მიტოქონდრიული დნმ-ის მოლეკულური წონა არის დაახლოებით (30-40) * 10 6 პროტოზოებში, საფუარებში, სოკოებში. მაღალ ცხოველებში, დაახლოებით (9–10) * 10 6.
მისი სიგრძე საფუარში დაახლოებით 5 მიკრონი უდრის, მცენარეებში - 30 მიკრონი. მიტოქონდრიულ დნმ-ში შემავალი გენეტიკური ინფორმაციის რაოდენობა მცირეა: იგი შედგება 15-75 ათასი ბაზის წყვილისგან, რომლებსაც შეუძლიათ დაშიფვრონ საშუალოდ 25-125 ცილოვანი ჯაჭვი, რომელთა მოლეკულური წონა დაახლოებით 40000-ს შეადგენს.
მიტოქონდრიული დნმ განსხვავდება ბირთვული დნმ-ისგან რამდენიმე მხრივ: მას აქვს სინთეზის უფრო მაღალი მაჩვენებელი (5-7-ჯერ), უფრო მდგრადია DNase-ს მოქმედების მიმართ, არის ორრგოლიანი მოლეკულა, შეიცავს მეტ გუანინს და ციტოზინი, ის დენატურდება მაღალ ტემპერატურაზე და უფრო ადვილად აღდგება. თუმცა, ყველა მიტოქონდრიული ცილა არ არის სინთეზირებული მიტოქონდრიული სისტემის მიერ. ამრიგად, ციტოქრომ C-ისა და სხვა ფერმენტების სინთეზს უზრუნველყოფს ბირთვში არსებული ინფორმაცია. მიტოქონდრიის მატრიცაში ლოკალიზებულია ვიტამინები A, B 2, B 12, K, E, ასევე გლიკოგენი.
მიტოქონდრიული ფუნქცია იგი შედგება უჯრედების სასიცოცხლო აქტივობისთვის აუცილებელი ენერგიის ფორმირებაში. უჯრედში ენერგიის წყაროდ შეიძლება გახდეს სხვადასხვა ნაერთები: ცილები, ცხიმები, ნახშირწყლები. თუმცა, ერთადერთი სუბსტრატი, რომელიც დაუყოვნებლივ მონაწილეობს ენერგეტიკულ პროცესებში, არის გლუკოზა.
ბიოლოგიური პროცესები, რის შედეგადაც მიტოქონდრიებში წარმოიქმნება ენერგია, შეიძლება დაიყოს 3 ჯგუფად: I ჯგუფი - ჟანგვითი რეაქციები, მათ შორის ორი ფაზა: ანაერობული (გლიკოლიზი) და აერობული. II ჯგუფი - დეფოსფორილირება, ატფ-ის გაყოფა და ენერგიის გამოყოფა. III ჯგუფი - ჟანგვის პროცესთან დაკავშირებული ფოსფორილირება.
გლუკოზის დაჟანგვის პროცესი პირველ რიგში ხდება ჟანგბადის (ანაერობული ან გლიკოლიზური გზით) პირუვინის ან რძემჟავას მონაწილეობის გარეშე.
თუმცა, ენერგიის მხოლოდ მცირე რაოდენობა გამოიყოფა. მომავალში, ეს მჟავები მონაწილეობენ დაჟანგვის პროცესებში, რომლებიც ხდება ჟანგბადის მონაწილეობით, ანუ ისინი აერობული არიან. პირუვიკის და რძემჟავას დაჟანგვის პროცესის შედეგად, რომელსაც კრებსის ციკლს უწოდებენ, წარმოიქმნება ნახშირორჟანგი, წყალი და დიდი რაოდენობით ენერგია.
მიღებული ენერგია არ გამოიყოფა სითბოს სახით, რაც გამოიწვევს უჯრედების გადახურებას და მთელი ორგანიზმის სიკვდილს, მაგრამ გროვდება შესანახად და ტრანსპორტირებისთვის ხელსაყრელ ფორმაში ადენოზინტრიფოსფორის მჟავას (ATP) სახით. ATP-ის სინთეზი მოდის ADP-დან და ფოსფორის მჟავიდან და ამიტომ ე.წ ფოსფორილირება.
ჯანსაღ უჯრედებში ფოსფორილირება დაკავშირებულია დაჟანგვასთან. დაავადებებში, კონიუგაცია შეიძლება დაწყვილდეს, ამიტომ სუბსტრატი იჟანგება და ფოსფორილირება არ ხდება და დაჟანგვა გადაიქცევა სითბოდ, ხოლო ატფ-ის შემცველობა უჯრედებში მცირდება. შედეგად, ტემპერატურა იზრდება და უჯრედების ფუნქციური აქტივობა მცირდება.
ამრიგად, მიტოქონდრიის მთავარი ფუნქციაა უჯრედის თითქმის მთელი ენერგიის გამომუშავება და ხდება თავად ორგანოიდის აქტივობისთვის აუცილებელი კომპონენტების, "რესპირატორული ანსამბლის" ფერმენტების, ფოსფოლიპიდების და ცილების სინთეზი.
მიტოქონდრიების აქტივობის კიდევ ერთი მხარეა მათი მონაწილეობა სპეციფიკურ სინთეზებში, მაგალითად, სტეროიდული ჰორმონების და ცალკეული ლიპიდების სინთეზში. სხვადასხვა ცხოველის კვერცხუჯრედებში იღლიის დაგროვება იქმნება მიტოქონდრიებში, ხოლო ისინი კარგავენ ძირითად სისტემას. ამოწურულ მიტოქონდრიებს ასევე შეუძლიათ გამოყოფის პროდუქტების დაგროვება.
ზოგიერთ შემთხვევაში (ღვიძლი, თირკმელები) მიტოქონდრიებს შეუძლიათ უჯრედში შემავალი მავნე ნივთიერებების და შხამების დაგროვება, მათი იზოლირება ძირითადი ციტოპლაზმიდან და ნაწილობრივ ბლოკავს ამ ნივთიერებების მავნე ზემოქმედებას. ამრიგად, მიტოქონდრიას შეუძლია აითვისოს სხვა უჯრედის ორგანელების ფუნქციები, როდესაც ეს საჭიროა კონკრეტული პროცესის სრულად მხარდასაჭერად ნორმალურ ან ექსტრემალურ პირობებში.
მიტოქონდრიების ბიოგენეზი. მიტოქონდრიები განახლებული სტრუქტურებია საკმაოდ მოკლე სასიცოცხლო ციკლით (მაგალითად, ვირთხების ღვიძლის უჯრედებში, მიტოქონდრიის ნახევარგამოყოფის პერიოდი დაახლოებით 10 დღეა). მიტოქონდრია წარმოიქმნება წინა მიტოქონდრიების ზრდისა და გაყოფის შედეგად. მათი დაყოფა შეიძლება მოხდეს სამი გზით: შეკუმშვა, მცირე უბნების გაჩენა და ქალიშვილის მიტოქონდრიების გაჩენა დედის შიგნით. მიტოქონდრიების დაყოფას (გამრავლებას) წინ უძღვის საკუთარი გენეტიკური სისტემის - მიტოქონდრიული დნმ-ის რეპროდუქცია.
ასე რომ, მკვლევართა უმრავლესობის შეხედულებისამებრ, მიტოქონდრიების ფორმირება ძირითადად ხდება მათი თვითრეპროდუქციის de novo გზით.
