Esej
na tému: "Rozmnožovanie"
Úvod 3
1. Typy reprodukcie 4
1.1 Nepohlavné rozmnožovanie 4
1.2 Pohlavné rozmnožovanie 6
2. Individuálny vývoj organizmov 10
2.1 Embryonálne obdobie vývoja 10
2.2 Postembryonálne obdobie vývoja 13
2.3 Všeobecné vzorce vývoja. Biogenetický zákon 15
Záver 18
Referencie 18
Úvod
Schopnosť rozmnožovania, t.j. produkcia novej generácie jedincov toho istého druhu je jednou z hlavných charakteristík živých organizmov. V procese rozmnožovania dochádza k prenosu genetického materiálu z rodičovskej generácie na ďalšiu generáciu, čo zabezpečuje reprodukciu vlastností nielen daného druhu, ale konkrétnych rodičovských jedincov. Pre druh je zmyslom rozmnožovania nahradiť tých jeho zástupcov, ktorí uhynú, čo zabezpečuje kontinuitu existencie druhu; rozmnožovanie navyše za vhodných podmienok umožňuje zvýšiť celkový počet druhov.
Každý nový jedinec, kým dosiahne štádium, v ktorom je schopný reprodukcie, musí prejsť niekoľkými štádiami rastu a vývoja. Niektorí jedinci umierajú pred dosiahnutím reprodukčného štádia (alebo sexuálnej zrelosti) v dôsledku ničenia predátormi, chorobami a rôznymi náhodnými udalosťami; preto tento druh môže prežiť len pod podmienkou, že každá generácia vyprodukuje viac potomkov, ako bolo rodičov, ktorí sa podieľali na reprodukcii. Veľkosť populácie kolíše v závislosti od rovnováhy medzi rozmnožovaním a vymieraním jedincov. Existuje množstvo rôznych stratégií šírenia, z ktorých každá má odlišné výhody a nevýhody; všetky budú opísané v tomto abstrakte.
1. Typy reprodukcie
Sú známe rôzne formy rozmnožovania, ale všetky sa dajú kombinovať do dvoch typov: sexuálne a asexuálne.
Sexuálna reprodukcia znamená zmenu generácií a vývoj organizmov zo špecializovaných pohlavných buniek vytvorených v gonádach. V tomto prípade sa nový organizmus vyvinie ako výsledok fúzie dvoch zárodočných buniek tvorených rôznymi rodičmi. U bezstavovcov sa však spermie a vajíčka často tvoria v tele jedného organizmu. Tento jav, bisexualita, sa nazýva hermafroditizmus. Kvitnúce rastliny sú tiež obojpohlavné. Vo väčšine druhov krytosemenných (kvitnúcich) rastlín obsahuje obojpohlavný kvet tak tyčinky, ktoré tvoria samčie pohlavné bunky (spermie), ako aj piestiky, ktoré obsahujú vajíčka. Približne u štvrtiny druhov sa samčie (tyčinkové) a samičie (peštikové) kvety vyvíjajú nezávisle, t.j. ich kvety sú jednopohlavné. Príkladom je konope. U niektorých rastlín (kukurica, breza) sa na tom istom jedincovi objavujú samčie aj samičie kvety.
Vyvíjajú sa niektoré druhy zvierat a rastlín
neoplodnené vajíčko. Tento typ reprodukcie sa nazýva panenský alebo partenogenetický.
Nepohlavné rozmnožovanie je charakteristické tým, že sa z nepohlavných, somatických (telesných) buniek vyvinie nový jedinec.
1.1 Nepohlavné rozmnožovanie
Pri nepohlavnom rozmnožovaní môže vzniknúť nový organizmus z jednej bunky alebo z niekoľkých nepohlavných (somatických) buniek matky. Asexuálna reprodukcia zahŕňa iba jedného rodiča. Keďže bunky, z ktorých vznikajú dcérske organizmy, vznikajú v dôsledku mitózy, všetci potomkovia budú mať podobné dedičné vlastnosti ako materská osoba.
Ryža. 1. Reprodukcia zeleného euglena
Mitotickým delením buniek sa rozmnožujú mnohé prvoky (améba, zelená euglena atď.), jednobunkové riasy (Chlamydomonas) (obr. 1). Iné jednobunkové niektoré nižšie huby, riasy (chlorella), živočíchy, napríklad pôvodca malárie, malarické plazmodium, sa vyznačujú sporuláciou. V tomto prípade sa bunka rozpadne na veľký počet jedincov, ktorý sa rovná počtu jadier, ktoré sa predtým vytvorili v rodičovskej bunke v dôsledku opakovaného delenia jej jadra. Mnohobunkové organizmy sú tiež schopné sporulácie: sú to machy, vyššie huby, mnohobunkové riasy, pteridofyty a niektoré ďalšie.
U jednobunkových aj mnohobunkových organizmov je pučenie tiež metódou nepohlavného rozmnožovania. Napríklad u kvasinkových húb a niektorých nálevníkov (ciciakov) sa pri pučaní na materskej bunke spočiatku vytvorí malý hrbolček obsahujúci jadro, púčik. Rastie, dosahuje veľkosť blízku veľkosti tela matky a potom sa oddeľuje a prechádza do nezávislej existencie. U mnohobunkových organizmov (sladkovodná hydra) oblička pozostáva zo skupiny buniek z oboch vrstiev steny tela. Púčik rastie, predlžuje sa a na jeho prednom konci sa objaví ústny otvor obklopený chápadlami. Pučanie končí vytvorením malej hydry, ktorá sa potom oddelí od materského organizmu.
U mnohobunkových živočíchov dochádza k nepohlavnému rozmnožovaniu rovnakým spôsobom (medúzy, annelids, ploskavce, ostnatokožce). Z každej takejto časti sa vyvinie plnohodnotný jedinec.
V rastlinách je rozšírené vegetatívne rozmnožovanie, t.j. časti tela odrezky, úponky, hľuzy. Zemiaky sa teda rozmnožujú upravenými podzemnými časťami stonky – hľuzami. Výhonky a odrezky jazmínu a vŕby sa ľahko zakoreňujú. Hrozno, ríbezle a egreše sa rozmnožujú pomocou odrezkov.
Dlhé plazivé stonky úponkov jahôd tvoria púčiky, ktoré po zakorenení dávajú vznik novej rastline. Len málo rastlín, ako napríklad begónia, sa dá rozmnožovať odrezkami listov (listová čepeľ a stopka). Na spodnej strane listu, v miestach, kde sa rozvetvujú veľké žily, sa objavujú korene, na hornej strane sú puky a potom výhonky.
Koreň sa používa aj na vegetatívne rozmnožovanie. V záhradníctve sa maliny, čerešne, slivky a ruže rozmnožujú pomocou odrezkov z bočných koreňov. Dahlia sa rozmnožujú pomocou koreňových hľúz. Úpravou podzemnej časti oddenky stonky vznikajú aj nové rastliny. Napríklad bodliak siaty s pomocou podzemkov dokáže vyprodukovať viac ako tisíc nových jedincov na 1 m2 pôdy.
1.2 Pohlavné rozmnožovanie
Sexuálne rozmnožovanie má v porovnaní s nepohlavným rozmnožovaním veľmi veľké evolučné výhody. Je to spôsobené tým, že genotyp potomstva vzniká spojením génov patriacich obom rodičom. V dôsledku toho sa zvyšuje schopnosť organizmov prispôsobiť sa podmienkam prostredia. Keďže v každej generácii sa uskutočňujú nové kombinácie, oveľa väčší počet jedincov sa môže prispôsobiť novým podmienkam existencie ako počas nepohlavného rozmnožovania. Vznik nových génových kombinácií zabezpečuje úspešnejšiu a rýchlejšiu adaptáciu druhu na meniace sa podmienky prostredia.
Podstata pohlavného rozmnožovania teda spočíva v spojení v dedičnom materiáli potomka genetickej informácie z dvoch rôznych zdrojov – rodičov.
V pohlavných žľazách sa vyvíjajú pohlavné bunky: mužské spermie, ženské vajíčka (alebo vajíčka). V prvom prípade sa ich vývoj nazýva spermatogenéza, v druhom - oogenéza (z latinského ovo - vajíčko).
V procese tvorby zárodočných buniek sa rozlišuje niekoľko štádií. Prvým štádiom je obdobie rozmnožovania, v ktorom sa prvotné zárodočné bunky delia mitózou, čo vedie k zvýšeniu ich počtu.
Druhou fázou je obdobie rastu. U nezrelých mužských gamét nie je výrazný. Ich veľkosť sa mierne zväčšuje. Naopak, oocyty budúcich vajíčok sa zväčšujú, niekedy stovky a častejšie tisíckrát alebo dokonca miliónkrát. Rast oocytov sa uskutočňuje v dôsledku látok tvorených inými bunkami tela. U rýb, obojživelníkov a vo väčšej miere aj u plazov a vtákov je teda hlavnou časťou vajíčka žĺtok. Syntetizuje sa v pečeni, v špeciálnej rozpustnej forme je transportovaný krvou do vaječníkov, preniká do rastúcich oocytov a tam sa ukladá vo forme žĺtkových doštičiek. Okrem toho sa v samotnej budúcej reprodukčnej bunke syntetizuje množstvo proteínov a veľké množstvo rôznych RNA: transportné, ribozomálne a informačné. Žĺtok je súbor živín (tuky, bielkoviny, uhľohydráty, vitamíny atď.) nevyhnutných na výživu vyvíjajúceho sa embrya a RNA zabezpečuje syntézu bielkovín v ranom štádiu vývoja, keď ešte nie sú využité jej vlastné katastrofálne informácie.
Ďalšia fáza, obdobie dozrievania alebo meióza, je znázornené na obrázku 2. Bunky vstupujúce do obdobia dozrievania obsahujú diploidnú sadu chromozómov a už zdvojnásobené množstvo DNA.

Ryža. 2. Dozrievanie zárodočných buniek (meióza)
Podstatou meiózy je, že každá pohlavná bunka dostane jeden haploidný súbor chromozómov. Meióza je však zároveň štádiom, počas ktorého sa spojením rôznych materských a otcovských chromozómov vytvárajú nové kombinácie génov, dochádza aj k rekombinácii dedičných sklonov v dôsledku kríženia - výmeny úsekov medzi homológnymi chromozómami počas procesu. meiózy.
Meióza zahŕňa dve po sebe nasledujúce divízie. Rovnako ako v mitóze má každé meiotické delenie štyri štádiá: profázu, metafázu, anafázu a telofázu.
Prvé (I) meiotické delenie. Profáza I začína špirálovitosťou chromozómov. Ako si pamätáte, každý chromozóm pozostáva z dvoch chromatidov spojených centromérou. Potom sa homológne chromozómy priblížia k sebe, každý bod každej chromatidy jedného chromozómu sa spojí so zodpovedajúcim bodom chromatidy iného, homológneho chromozómu. Tento proces presného a úzkeho spojenia homológnych chromozómov v meióze sa nazýva konjugácia. V budúcnosti môže medzi týmito chromozómami dôjsť k prekríženiu - výmene identických alebo homológnych, t.j. obsahujúcich rovnaké gény, oblasti. Ku koncu profázy vznikajú medzi homológnymi chromozómami odpudivé sily. Najprv sa objavia v oblasti centroméry a potom v iných oblastiach.
V metafáze I je chromozómová špirála maximálna. Konjugované chromozómy sú umiestnené pozdĺž rovníka, pričom centroméry homológnych chromozómov smerujú k rôznym pólom bunky. K nim sú pripevnené závity vretena.
V anafáze I sa ramená homológnych chromozómov nakoniec oddelia a chromozómy sa presunú na rôzne póly. Z každého páru homológnych chromozómov sa teda do dcérskej bunky dostane iba jeden. Počet chromozómov sa zníži na polovicu, chromozómová sada sa stáva haploidnou. Každý chromozóm sa však skladá z dvoch chromatidov, t.j. stále obsahuje dvojnásobné množstvo DNA.
V telofáze I sa na krátky čas vytvorí jadrový obal. Počas medzifázy medzi prvým a druhým delením meiózy nedochádza k reduplikácii DNA. Bunky vytvorené v dôsledku prvého delenia dozrievania sa líšia v zložení otcovských a materských chromozómov a následne v súbore génov.
Napríklad všetky ľudské bunky, vrátane primordiálnych zárodočných buniek, obsahujú 46 chromozómov. Z toho 23 od otca a 23 od matky. Keď sa zárodočné bunky vytvoria po prvom meiotickom delení, spermatocyty a oocyty tiež dostanú 23 chromozómov. Avšak v dôsledku náhodnosti divergencie otcovských a materských chromozómov v anafáze I dostávajú výsledné bunky širokú škálu kombinácií rodičovských chromozómov. Napríklad jeden z nich môže mať 3 otcovské a 20 materských chromozómov, ďalší 10 otcovských a 13 materských chromozómov, tretí 20 otcovských a 3 materské atď. Počet možných kombinácií je veľmi veľký. Ak zoberieme do úvahy aj výmenu homológnych úsekov chromozómov v profáze prvého delenia meiózy, tak je celkom zrejmé, že každá výsledná zárodočná bunka je geneticky jedinečná, keďže nesie svoj vlastný unikátny súbor génov.
V dôsledku toho je meióza základom kombinovanej genotypovej variability.
Druhé (II.) meiotické delenie. Druhé delenie meiózy vo všeobecnosti prebieha rovnakým spôsobom ako bežné mitotické delenie, len s tým rozdielom, že deliaca sa bunka je haploidná. V anafáze II sa centroméry spájajúce sesterské chromatidy v každom chromozóme delia a chromatidy, ako pri mitóze, sa od tohto momentu stávajú nezávislými chromozómami. S dokončením telofázy II sa celý proces meiózy končí: z pôvodnej primárnej zárodočnej bunky sa vytvoria štyri haploidné bunky.
U mužov sa všetky premieňajú na gaméty - spermie. U žien v dôsledku nerovnomernej meiózy iba jedna bunka produkuje životaschopné vajíčko. Ďalšie tri dcérske bunky sú oveľa menšie, menia sa na takzvané vodiace alebo redukčné telá, ktoré čoskoro odumierajú. Z biologického hľadiska je vznik iba jedného vajíčka a smrť troch geneticky kompletných vodiacich teliesok spôsobená potrebou zachovať v jednej bunke všetky rezervné živiny, ktoré budú potrebné pre vývoj budúceho embrya.
Obdobie formovania pozostáva z buniek, ktoré nadobúdajú určitý tvar a veľkosť zodpovedajúcu ich funkcii.
Počas procesu dozrievania sa ženské zárodočné bunky pokrývajú membránami a sú pripravené na oplodnenie ihneď po ukončení meiózy. V mnohých prípadoch, napríklad u plazov, vtákov a cicavcov, sa v dôsledku aktivity buniek obklopujúcich vajíčko okolo vajíčka objaví množstvo ďalších membrán. Ich funkciou je chrániť vajíčko a vyvíjajúce sa embryo pred vonkajšími nepriaznivými vplyvmi. Spermie môžu mať rôzne veľkosti a tvary.
Funkciou spermií je dodávať do vajíčka genetickú informáciu a stimulovať jeho vývoj. Vytvorená spermia obsahuje mitochondrie, Golgiho aparát, ktorý vylučuje enzýmy, ktoré rozpúšťajú membránu vajíčka počas oplodnenia, t.j. počas splynutia spermie a vajíčka. Výsledná diploidná bunka sa nazýva zygota.
2. Individuálny vývoj organizmov
Individuálny vývoj alebo ontogenéza sa vzťahuje na celé obdobie života jedinca, od okamihu splynutia spermie s vajíčkom a vytvorením zygoty až po smrť organizmu. Ontogenéza je rozdelená do dvoch období: 1) embryonálna od vytvorenia zygoty po pôrod alebo výstup z vaječných membrán; 2) postembryonálne od výstupu z vaječných membrán alebo narodenia až po smrť organizmu.
Veda, ktorá študuje zákonitosti individuálneho vývoja organizmov v embryonálnom štádiu, sa nazýva embryológia (z gréckeho embryo embryo).
2.1 Embryonálne obdobie vývoja
U väčšiny mnohobunkových živočíchov, bez ohľadu na zložitosť ich organizácie, sú štádiá embryonálneho vývoja, ktorými embryo prechádza, rovnaké. V embryonálnom období existujú tri hlavné štádiá: štiepenie, gastrulácia a primárna organogenéza.
Rozdelenie. Vývoj organizmu začína v štádiu jednej bunky. Oplodnené vajíčko je bunka a zároveň organizmus v najskoršom štádiu svojho vývoja. V dôsledku opakovaného delenia sa jednobunkový organizmus mení na mnohobunkový. Diploidné jadro, ktoré vzniká pri oplodnení splynutím spermie a vajíčka, sa v priebehu niekoľkých minút začne deliť a s ním sa delí aj cytoplazma. Výsledné bunky sa každým delením zmenšujú, preto sa proces delenia nazýva štiepenie. Počas obdobia fragmentácie sa bunkový materiál hromadí pre ďalší vývoj. Fragmentácia končí vytvorením mnohobunkového embrya, blastuly. Blatula má dutinu vyplnenú tekutinou, takzvanú primárnu telovú dutinu.
V prípadoch, keď je v cytoplazme vajíčka málo žĺtka (ako v lancelete) alebo relatívne málo (ako u žaby), je fragmentácia úplná, t. j. vajce sa úplne rozdelí.
V opačnom prípade nastáva obdobie fragmentácie u vtákov. Cytoplazma bez žĺtka tvorí iba 1% z celkového objemu kuracieho vajca; celá zostávajúca cytoplazma vajíčka, a teda aj zygota, je naplnená hmotou žĺtka. Ak sa pozriete pozorne na kuracie vajce, na jednom z jeho pólov priamo na žĺtku môžete vidieť malú škvrnu - blastulu alebo zárodočný disk, ktorý vznikol v dôsledku rozdrvenia časti cytoplazmy bez žĺtka obsahujúcej jadro. V takýchto prípadoch sa drvenie nazýva neúplné. Neúplná fragmentácia je charakteristická aj pre niektoré ryby a plazy.
Vo všetkých prípadoch, tak v lancelete, ako aj u obojživelníkov a vtákov, ako aj u iných zvierat, celkový objem buniek v štádiu blastuly nepresahuje objem zygoty. Inými slovami, mitotické delenie zygoty nie je sprevádzané rastom výsledných dcérskych buniek na objem matky a ich veľkosť v dôsledku série postupných delení sa postupne zmenšuje. Táto vlastnosť delenia mitotických buniek počas štiepenia sa pozoruje počas vývoja oplodnených vajíčok u všetkých zvierat.
Pre rôzne druhy zvierat sú charakteristické aj niektoré ďalšie znaky drvenia. Napríklad všetky bunky v blastule majú diploidnú sadu chromozómov, majú identickú štruktúru a líšia sa od seba najmä množstvom žĺtka, ktoré obsahujú. Takéto bunky, ktorým chýbajú znaky špecializácie na vykonávanie určitých funkcií, sa nazývajú nešpecializované (alebo nediferencované) bunky. Ďalším znakom štiepenia je extrémne krátky mitotický cyklus blastomérov v porovnaní s bunkami dospelého organizmu. Počas veľmi krátkej interfázy dochádza len k duplikácii DNA.
Gastrulácia. Blastula, spravidla pozostávajúca z veľkého počtu blastomérov (napríklad v lancelete z 3000 buniek), počas vývojového procesu prechádza do nového štádia, ktoré sa nazýva gastrula (z gréckeho gaster žalúdok). Embryo v tomto štádiu pozostáva z jasne rozlíšiteľných vrstiev buniek - takzvaných zárodočných vrstiev: vonkajšej alebo ektodermy (z gréckeho ectos - nachádza sa vonku) a vnútornej alebo endodermy (z gréckeho entos - nachádza sa vo vnútri). . Súbor procesov vedúcich k vzniku gastruly sa nazýva gastrulácia.
V lancelete sa gastrulácia uskutočňuje invagináciou jedného z pólov blastuly dovnútra, smerom k druhému, u iných zvierat buď delamináciou steny blastuly, alebo prerastaním masívneho vegetatívneho pólu malými bunkami blastuly. zvieracia tyč.
U mnohobunkových živočíchov, s výnimkou coelenterátov, sa paralelne s gastruláciou alebo, ako v lancelete, po nej objavuje tretia zárodočná vrstva mezoderm (z gréckeho mesos umiestnená v strede), čo je súbor bunkových prvkov umiestnených medzi ekto- a endoderm v blastocéle primárnej telesnej dutiny. S výskytom mezodermu sa embryo stáva trojvrstvovým.
Podstatou gastrulačného procesu je teda pohyb bunkových hmôt. Bunky embrya sa prakticky delia a nerastú. V tomto štádiu však začína využívanie genetickej informácie embryonálnych buniek a objavujú sa prvé známky diferenciácie.
Diferenciácia, alebo diferenciácia, je proces jej vzniku a zväčšovania štrukturálnych a funkčných rozdielov medzi jednotlivými bunkami a časťami embrya. Z morfologického hľadiska je diferenciácia vyjadrená tvorbou niekoľkých stoviek typov buniek špecifickej štruktúry, ktoré sa navzájom líšia. Z nešpecializovaných buniek blastuly postupne vznikajú epitelové bunky kože, črevného epitelu, pľúc, vznikajú nervové a svalové bunky atď. Z biochemického hľadiska bunková špecializácia spočíva v schopnosti syntetizovať určité proteíny, ktoré sú charakteristické len pre daný typ bunky. Lymfocyty syntetizujú ochranné proteíny protilátky, svalové bunky kontraktilný proteín myozín. Každý typ bunky produkuje svoje vlastné proteíny, ktoré sú pre ňu jedinečné. Biochemická špecializácia buniek je zabezpečená selektívnou, rozdielnou aktivitou génov, t.j. v bunkách rôznych zárodočných vrstiev, základoch určitých orgánov a systémov, začínajú fungovať rôzne skupiny génov.
U rôznych živočíšnych druhov z rovnakých zárodočných vrstiev vznikajú rovnaké orgány a tkanivá. To znamená, že sú homológne. Z buniek vonkajšej zárodočnej vrstvy – ektodermy – sa teda u článkonožcov, strunatcov vrátane rýb, obojživelníkov, plazov, vtákov a cicavcov tvorí koža a ich deriváty, ale aj nervový systém a zmyslové orgány. Homológia zárodočných vrstiev veľkej väčšiny živočíchov je jedným z dôkazov jednoty živočíšneho sveta.
Organogenéza. Po dokončení gastrulácie tvorí embryo komplex axiálnych orgánov: nervová trubica, notochorda a črevná trubica. V lancelete sú axiálne orgány tvorené nasledovne: ektoderm na dorzálnej strane embrya sa ohýba pozdĺž stredovej čiary a mení sa na drážku a na jej okrajoch začína rásť ektoderm, ktorý sa nachádza vpravo a vľavo od nej. Drážka, základ nervového systému, klesá pod ektoderm a jej okraje sa uzatvárajú. Vytvára sa nervová trubica. Zvyšok ektodermu je základom kožného epitelu.
Dorzálna časť endodermu, ktorá sa nachádza priamo pod nervovým rudimentom, je oddelená od zvyšku endodermy a skladá sa do hustej šnúry nazývanej notochord. Zo zvyšnej časti endodermu sa vyvíja mezoderm a črevný epitel. Ďalšia diferenciácia embryonálnych buniek vedie k vzniku mnohých derivátov zárodočných vrstiev - orgánov a tkanív. V procese špecializácie buniek, ktoré tvoria zárodočné vrstvy, sa z ektodermy vytvára nervový systém, zmyslové orgány, kožný epitel a zubná sklovina; z endodermu črevný epitel, tráviace žľazy pečeň a pankreas, epitel žiabier a pľúc; z mezodermového svalového tkaniva, spojivového tkaniva vrátane voľného spojivového tkaniva, chrupavkového a kostného tkaniva, krvi a lymfy, ako aj obehového systému, obličiek, pohlavných žliaz.
2.2 Postembryonálne obdobie vývoja
V momente pôrodu alebo uvoľnenia organizmu z vaječných škrupín končí embryonálne obdobie a začína postembryonálne obdobie vývoja. Postembryonálny vývoj môže byť priamy alebo sprevádzaný transformáciou (metamorfózou).
Počas priameho vývoja (u plazov, vtákov, cicavcov) sa z vaječných škrupín alebo z tela matky vynorí malý organizmus, ale už sú vytvorené všetky hlavné orgány charakteristické pre dospelého zvieraťa. Postembryonálny vývoj je v tomto prípade redukovaný hlavne na rast a pubertu.
Počas vývoja s metamorfózou sa z vajíčka vynorí larva, ktorá má zvyčajne jednoduchšiu štruktúru ako dospelé zviera, so špeciálnymi larválnymi orgánmi, ktoré v dospelom stave chýbajú. Larva sa živí, rastie a po čase sú orgány lariev nahradené orgánmi charakteristickými pre dospelých jedincov. V dôsledku toho sa počas metamorfózy zničia orgány lariev a objavia sa orgány charakteristické pre dospelé zvieratá.
Pozrime sa na niekoľko príkladov nepriameho postembryonálneho vývoja. Ascidiánska larva (kmeň Chordata, podkmeň Larval-Chordata) má všetky hlavné charakteristiky strunatcov: notochord, nervovú trubicu a žiabrové štrbiny v hltane. Voľne pláva, potom sa prichytí k nejakému pevnému povrchu na dne mora, kde dôjde k metamorfóze: jeho chvost, notochorda a svaly zmiznú a nervová trubica sa rozpadne na jednotlivé bunky, z ktorých väčšina je fagocytovaná. Z nervového systému lariev zostáva len skupina buniek, z ktorých vzniká nervový ganglion. Štruktúra tela dospelého ascidiána, ktorý vedie pripútaný životný štýl, sa vôbec nepodobá obvyklým znakom organizácie strunatcov. Iba znalosť znakov ontogenézy umožňuje určiť systematické postavenie ascidiánov: štruktúra lariev naznačuje ich pôvod z strunatcov, ktorí viedli voľný životný štýl. V procese metamorfózy prechádzajú ascidíny na sedavý spôsob života, a preto je ich organizácia zjednodušená.
Larválna forma obojživelníkov je pulec, ktorý sa vyznačuje žiabrovými štrbinami, bočnou líniou, dvojkomorovým srdcom a jedným kruhom krvného obehu. Počas procesu metamorfózy, ku ktorému dochádza pod vplyvom hormónu štítnej žľazy, sa chvost vyrieši, objavia sa končatiny, zmizne bočná línia, rozvinú sa pľúca a druhý kruh krvného obehu. Pozoruhodná je podobnosť mnohých štrukturálnych znakov pulcov a rýb (laterálna línia, štruktúra srdca a obehového systému, žiabrové štrbiny).
Ako príklad metamorfózy môže slúžiť aj vývoj hmyzu. Húsenice motýľov alebo larvy vážok sa výrazne líšia v štruktúre, životnom štýle a biotope od dospelých zvierat a pripomínajú ich predkov, annelids.
Postembryonálne obdobie vývoja má rôzne trvanie. Napríklad májky žijú 2-3 roky v larválnom stave a od 2-3 hodín do 2-3 dní v zrelom stave, v závislosti od druhu. Vo väčšine prípadov je postembryonálne obdobie dlhšie. U ľudí zahŕňa štádium puberty, štádium zrelosti a štádium staroby.
U cicavcov a ľudí je známa závislosť dĺžky života od trvania puberty a tehotenstva. Priemerná dĺžka života zvyčajne presahuje
predreprodukčné obdobie ontogenézy o 5-8 krát.
Postembryonálny vývoj je sprevádzaný rastom. Rozlišuje sa neurčitý rast, ktorý pokračuje počas celého života, a rast určitý, obmedzený na určité obdobie. Neobmedzený rast sa pozoruje u drevnatých foriem rastlín, niektorých mäkkýšov, stavovcov, rýb a potkanov.
U mnohých zvierat sa rast zastaví krátko po dosiahnutí pohlavnej dospelosti. U ľudí sa rast končí vo veku 20-25 rokov.
2.3 Všeobecné vzorce vývoja. Biogenetický zákon
Všetky mnohobunkové organizmy sa vyvíjajú z oplodneného vajíčka. Vývoj embryí u zvierat patriacich do rovnakého typu je do značnej miery podobný. U všetkých strunatcov sa v embryonálnom období vytvorí notochord axiálneho skeletu, objaví sa nervová trubica a v prednej časti hltana sa vytvoria žiabrové štrbiny. Štrukturálny plán strunatcov je tiež rovnaký. V počiatočných štádiách vývoja sú embryá stavovcov veľmi podobné (obr. 3). Tieto fakty potvrdzujú platnosť zákona embryonálnej podobnosti formulovaného K. Baerom: „Embryá vykazujú už od najranejších štádií určitú všeobecnú podobnosť v rámci typu.“ Podobnosť embryí rôznych systematických skupín naznačuje ich spoločný pôvod. Následne štruktúra embryí odhaľuje charakteristiky triedy, rodu, druhu a nakoniec vlastnosti charakteristické pre daného jedinca. Rozdielnosť charakteristík embryí počas vývoja sa nazýva embryonálna divergencia a odráža vývoj konkrétnej systematickej skupiny zvierat, históriu vývoja daného druhu.

Ryža. 3. Zárodočná podobnosť u stavovcov: 1 monotremes (echidna), 2 vačkovce (klokany), 3 artiodaktyly (jeleň), 4 mäsožravce (mačka), 5 primátov (opice), 6 - ľudia
Veľká podobnosť medzi embryami v počiatočných štádiách vývoja a
fenomén rozdielov v neskorších štádiách má svoje vysvetlenie.
Telo podlieha variabilite počas celého vývoja.
Mutačný proces ovplyvňuje gény, ktoré určujú štrukturálne a metabolické vlastnosti najmladších embryí. Ale štruktúry, ktoré v nich vznikajú (starodávne charakteristiky charakteristické pre vzdialených predkov), zohrávajú veľmi dôležitú úlohu v procesoch ďalšieho vývoja. Ako je uvedené, notochord primordium indukuje tvorbu nervovej trubice a jej strata vedie k zastaveniu vývoja. Preto zmeny v počiatočných štádiách zvyčajne vedú k nedostatočnému rozvoju a smrti jedinca. Naopak, zmeny v neskorších štádiách, ovplyvňujúce menej významné vlastnosti, môžu byť pre organizmus prospešné a v takýchto prípadoch sú zachytené prirodzeným výberom.
Vzhľad v embryonálnom období vývoja moderných zvierat s vlastnosťami charakteristickými pre ich vzdialených predkov odráža evolučné transformácie v štruktúre orgánov.
Organizmus vo svojom vývoji prechádza jednobunkovým štádiom (štádium zygoty), ktoré možno považovať za opakovanie fylogenetického štádia primitívnej améby. U všetkých stavovcov, vrátane ich najvyšších predstaviteľov, vzniká notochorda, ktorá je následne nahradená chrbticou a u ich predkov, súdiac podľa lanceletu, notochorda zostala na celý život. Počas embryonálneho vývoja vtákov a cicavcov, vrátane ľudí, sa v hltane objavujú žiabrové štrbiny a zodpovedajúce septa. Skutočnosť vzniku častí žiabrového aparátu v embryách suchozemských stavovcov sa vysvetľuje ich pôvodom z rybích predkov, ktorí dýchali žiabrami. Štruktúra srdca ľudského embrya v ranom období formovania sa podobá štruktúre tohto orgánu u rýb: má jednu predsieň a jednu komoru. Bezzubé veľryby vyvíjajú zuby počas embryonálneho obdobia. Tieto zuby nevyrážajú, sú zničené a rozpustené.
Tu uvedené a mnohé ďalšie príklady poukazujú na hlbokú súvislosť medzi individuálnym vývojom organizmov a ich historickým vývojom. Táto súvislosť je vyjadrená v biogenetickom zákone, ktorý sformulovali F. Müller a E. Haeckel v 19. storočí: ontogenéza (individuálny vývoj) každého jedinca je krátkym a rýchlym opakovaním fylogenézy (historického vývoja) druhu, ku ktorému tento jedinec patrí. .
Záver
Dokončením práce na abstrakte môžeme dospieť k záveru, že schopnosť rozmnožovania, čiže sebarozmnožovania, je jednou z najdôležitejších charakteristík organickej prírody. Rozmnožovanie je vlastnosť vlastná všetkým živým organizmom bez výnimky, od baktérií po cicavce.
Existencia akýchkoľvek druhov živočíchov a rastlín, baktérií a húb, kontinuita medzi rodičovskými jedincami a ich potomkami je zachovaná len prostredníctvom rozmnožovania. S vlastnou reprodukciou úzko súvisí ďalšia vlastnosť živých vecí.vývoj organizmov. toJe tiež súčasťou všetkého života na Zemi: najmenších jednobunkových organizmov a mnohobunkových rastlín a živočíchov.
Bibliografia
Bogen G. Moderná biológia. - M.: Mir, 1970.
Green N., Stout W., Taylor D. Biology: v 3 zväzkoch T. 3: trans. z angličtiny/ed. R. Soper. - M.: Mir, 1990.
Mamontov S.G. Biológia. Všeobecné vzory. M.: Drop, 2002.
Od zvierat k ľuďom. M.: Nauka, 1971.
Slyusarev A.A. Biológia so všeobecnou genetikou. - M.: Medicína, 1978.
Prednáška č. 3 Ontogenéza
1. Gametogenéza
2. Embryonálne obdobie
3. Postembryonálne obdobie
Ontogenéza– individuálny vývoj organizmu od zygoty po biologickú smrť. Progenéza– obdobie pred ontogenézou, zahŕňa gametogenézu a oplodnenie. Fylogenéza– evolučný vývoj druhu.
Pre človeka je charakteristické pohlavné rozmnožovanie, ktoré je charakterizované: prítomnosťou pohlavných žliaz, gamét, procesom oplodnenia a sexuálnym dimorfizmom.
Gametogenéza - proces tvorby zárodočných buniek ovogenéza - dozrievanie oocytov, spermatogenéza- spermie. Gametogenéza sa vyskytuje v pohlavných žľazách tela. Gaméty sa tvoria z embryonálnych epitelových buniek, ktoré vznikajú počas embryonálneho obdobia vývoja organizmu.
V procese formovania prechádzajú zárodočné bunky tromi štádiami:
1. Reprodukčné obdobie (bunky embryonálneho epitelu sa delia mitózou);
2. Obdobie rastu;
3. Obdobie dozrievania, bunky sa delia meiózou, výsledkom čoho je tvorba gamét (obr. 5).
Ryža. 5. Delenie zárodočných buniek
Spermie dozrievajú v semenných tubuloch semenníkov. Nachádza sa medzi semennými tubulmi intersticiálneho tkaniva produkujúci mužský pohlavný hormón - testosterónu. Mužské pohlavné hormóny - androgény regulujú reprodukčnú funkciu, gametogenézu a tvorbu sekundárnych sexuálnych charakteristík. Spermie dozrievajú za 70 dní. Semenné tubuly obsahujú gaméty v rôznych štádiách dozrievania. V 5 ml. Ľudská semenná tekutina obsahuje 12 miliónov spermií. Sú negatívne nabité, náboj bráni ich zlepeniu. Veľký počet samčích gamét je biologicky účelný, zníženie ich počtu o 40% znemožňuje proces oplodnenia. Spermie sú malé pohyblivé bunky pozostávajúce z hlavy, krku a chvosta. Na hlave je akrozóm, modifikovaný lyzozóm, obsahuje enzýmy, ktoré rozpúšťajú membránu vajíčka počas oplodnenia. Predpokladaná dĺžka života je od 6 do 72 hodín.
Oogenéza začína v embryonálnom období vývoja ženského tela. V profáznom štádiu prvého delenia meiózy, keď dôjde ku konjugácii a prekríženiu, nastáva dočasná pauza. Ďalší vývoj vajíčka sa obnoví počas puberty dievčaťa. Vplyvom ženských pohlavných hormónov – estrogénov sa okolo oocytu prvého rádu vytvorí folikul, bunkový mechúrik, ktorý bunku chráni a vyživuje.
Ako folikul dozrieva, presúva sa na okraj vaječníka a potom praskne; toto štádium vývoja vajíčka sa nazýva vaječník alebo folikul, trvá 12 dní. Prasknutie folikulu a uvoľnenie vajíčka z vaječníka do brušnej dutiny sa nazýva ovulácia.
Po ovulácii je možné oplodnenie. Ženská gaméta je zachytená fimbriami vajcovodu. Pomocou ciliárneho epitelu sa presúva do maternice, štádium maternice trvá 12-14 dní. Do tejto doby sa pod vplyvom ženských pohlavných hormónov uvoľní maternicový epitel. Ak nedôjde k oplodneniu, epitel a vajíčka sa vylučujú z tela v dôsledku kontrakcie hladkých svalov stien maternice. Tento proces sa nazýva mensis a trvá 3-4 dni. Čas od začiatku tvorby folikulov po nástup menštruácie je teda 20-30 dní a nazýva sa ovariálny-menštruačný cyklus (obr. 6).

Ryža. 6. Ovulácia a implantácia vajíčka do maternice
Obsahuje ovariálne štádium, maternica a menzes. Počas ovariálno-menštruačného cyklu sa rekonštruuje celý reprodukčný systém ženy, mení sa kvalitatívne a kvantitatívne zloženie hormónov, mení sa výkonnosť, sila, flexibilita, vytrvalosť. Pri organizovaní tréningového procesu sa berú do úvahy fyziologické zmeny v ženskom tele.
Hnojenie- Toto je splynutie vajíčka a spermie. Existujú tri fázy oplodnenia:
1) akrozomálna reakcia (20 sekúnd) – rozpustenie škrupiny vajíčka akrozómovými enzýmami a prienik spermie do vajíčka. Predpokladá sa, že spermie, ktoré vytvárajú najväčší penetračný potenciál (vzrušenie) na membráne vajíčka, prenikajú do vajíčka;
2) dočasná pauza – vo vajíčku nedochádza k viditeľným zmenám, metabolizmus sa zvyšuje;
3) fúzia jadier vajíčka a spermie, obnovenie diploidnej sady chromozómov.
K oplodneniu dochádza v hornej tretine vajíčkovodu. Oplodnené vajíčko je tzv zygota. Vytvorením zygoty sa začína ontogenéza človeka.
2. Ontogenézačlovek má dve fázy: embryonálne a postembryonálne.
Embryonálne štádium vývoja pokračuje od zygoty po pôrod. Zahŕňa štádiá: zygota, blastula, gastrula, histogenéza, organogenéza.
Zygota– opakovane sa delí mitózou, čo vedie k vytvoreniu mnohobunkového jednovrstvového embrya – blastula. Blastula sa nezväčšuje, pretože v tomto čase je vo vajíčkovode a mala by sa voľne pohybovať do maternice. Šiesty deň vstúpi do maternice a prichytí sa k jej stene, pričom sa ponorí do uvoľneného epitelu. Tento proces sa nazýva implantácia. Embryo pokračuje vo vývoji a stáva sa dvojvrstvovým, potom trojvrstvovým. V tomto štádiu vývoja je tzv gastrula. V dôsledku toho sa vytvárajú tri zárodočné vrstvy: ektoderm, endoderm, mezoderm. Tkanivá a orgány sa tvoria z embryonálnych systémov.
Takto: blastula– jednovrstvové embryo, gastrula– dvoj a trojvrstvové embryo. Histogenéza– kladenie tkanív zo zárodočných vrstiev. Organogenéza- kladenie orgánov. Vo veku 8 týždňov je hmotnosť ľudského embrya 4 g, veľkosť 5 mm. Do tejto doby prešiel formačnými procesmi a získal obrysy ľudského tela. Po ôsmich týždňoch tehotenstva sa ľudské embryo nazýva plod.
Počas vývoja ľudského embrya dočasné orgány, ktoré po narodení strácajú svoje funkcie. Medzi dočasné orgány patria: zárodočný mušle, chorion, amnion a placenta, pupocna snura. Placenta zabezpečuje úzky kontakt medzi kapilárami dieťaťa a matky. Vyvíjajúci sa organizmus dostáva cez placentu živiny, kyslík, protilátky, odvádza produkty látkovej výmeny. Placenta plní bariérovú funkciu, chráni embryo pred mikroorganizmami, niektorými liekmi a toxínmi (obr. 7).

Ryža. 7. Placenta
1 – maternica; 2 – amnion; 3 – embryo; 4 – plodová voda; 5 – placenta; 6 – kapiláry placenty; 7 – žily; 8 – tepny
Jemné mechanizmy tvorby orgánov a tkanív dieťaťa môžu byť narušené pri vystavení toxickým, omamným látkam, alkoholu, nikotínu obsiahnutým v krvi matky. Poruchy ľudského embryonálneho vývoja sa nazývajú vrodené deformácie a malformácie. Patria sem: „rázštep podnebia“ (neprítomnosť horného podnebia), „rázštep pery“ (nezrastenie maxilárnych kostí), polydaktýlia (prsty navyše), syndaktýlia (zrastené prsty). Nepriaznivé faktory prostredia, ktoré spôsobujú vrodené vývojové chyby, sa nazývajú teratogénny(spôsobujúce deformácie) (obr. 8).

Ryža. 8. Faktory prostredia ovplyvňujúce vývoj ľudského tela
Embryonálne obdobie ľudského vývoja trvá 36 týždňov.
3. Obdobie po narodení a pred biologickou smrťou sa nazýva postembryonálne. Obsahuje mladistvý, puberta obdobia vývoja a starnutia.
Mládež – bodka pred pubertou. Puberta– obdobie puberty.
V športovej praxi, fyziológii, pedagogike, psychológii sa používa nasledovná veková periodizácia postembryonálna ontogenéza človeka.
V období od 1 do 10 dní je dieťa tzv novorodenca;
Od 10 dní do 1 roka - hrudník;
od 1 roka do troch – obdobie rané detstvo;
od 4 do 7 rokov - prvé detstvo;
od 8 rokov do 12 rokov pre dievčatá a do 13 rokov pre chlapcov druhé detstvo;
od 12 do 15 rokov pre dievčatá a od 13 do 16 pre chlapcov - tínedžerov;
17-21 (chlapci), 16-20 rokov (dievčatá) – dospievania;
20-55 pre ženy a 21-60 pre mužov obdobie puberta(puberta);
od 55 a 60 do 70 - starí ľudia;
od 70 do 90 starci;
viac ako 90 storočných.
V ľudskej ontogenéze existujú citlivý a kritický obdobia.
Kritické obdobia sa vyznačujú zvýšenou aktivitou jednotlivých génov a ich komplexov, ktoré riadia vývoj akýchkoľvek znakov organizmu. V týchto obdobiach dochádza k výraznej reštrukturalizácii regulačných procesov, ku kvalitatívnemu a kvantitatívnemu skoku vo vývoji jednotlivých orgánov a funkčných systémov. V týchto obdobiach je telo najcitlivejšie na účinky environmentálnych faktorov. Za kritické obdobia ontogenézy sa považuje implantácia, placentácia, tvorba osových orgánov, nervových a črevných trubíc, notochordov, tvorba srdca a iné.
Ak sa žena nakazí rubeolou medzi 3. a 9. týždňom tehotenstva, existuje riziko, že sa u plodu vyvinú chyby, ako sú srdcové choroby, šedý zákal a hluchota. Inokedy rubeola nespôsobuje malformácie plodu.
Citlivé obdobia– ide o obdobia zníženej genetickej kontroly a zvýšenej citlivosti individuálnych charakteristík tela na vplyvy prostredia, vrátane pedagogických a trénerských. Citlivé obdobie na prejavenie rôznych ukazovateľov kvality rýchlosti teda nastáva vo veku 11-14 rokov a do 15. roku života je dosiahnutá jeho maximálna úroveň, kedy sú možné vysoké športové výkony. Podobný obraz sa pozoruje v ontogenéze pre prejav kvalít obratnosti a flexibility.
Pre trénerov a učiteľov pôsobiacich v oblasti telesnej a športovej výchovy má znalosť citlivých období veľký praktický význam, keďže v citlivých obdobiach má najväčší význam tréningový efekt.
Možno ich rozdeliť podľa toho, koľko buniek sa podieľa na procese nepohlavného rozmnožovania: nepohlavné rozmnožovanie, pri ktorom z jednej bunky vzniká dcérska generácia: bunkové delenie viacnásobné delenie buniek schizogónia sporulácia sporulácia pučanie v jednobunkových kvasinkách...
Zdieľajte svoju prácu na sociálnych sieťach
Ak vám táto práca nevyhovuje, v spodnej časti stránky je zoznam podobných prác. Môžete tiež použiť tlačidlo vyhľadávania
Rozmnožovanie a vývoj organizmov
Plán
- Pojem a význam reprodukcie.
- Formy a typy reprodukcie.
- Bunkový cyklus. Mitóza. meióza.
- Štruktúra gamét. Gametogenéza.
1. Pojem a význam reprodukcie
Jednou z vlastností živých vecí je diskrétnosť, tie. na akejkoľvek úrovni organizácie je živá hmota reprezentovaná elementárnymi štruktúrnymi jednotkami. Každý jedinec daného druhu je smrteľný a existencia druhu je zachovaná reprodukcie organizmov. Diskrétnosť života teda predpokladá jeho reprodukciu, t.j. reprodukčný proces.
Reprodukcia toto je schopnosť živých bytostí reprodukovať svoj vlastný druh. Tým je zabezpečená kontinuita a kontinuita života. Kontinuita je určená skutočnosťou, že počas procesu reprodukcie sa genetický materiál prenáša z rodičov na potomkov, v dôsledku čoho sa v dcérskych organizmoch prejavujú rodičovské vlastnosti v tej či onej miere.
Reprodukčná hodnota:
- Zvyšovanie alebo udržiavanie počtu druhov. V dôsledku reprodukcie dochádza nielen k reprodukcii (t. j. reprodukcii vlastného druhu), ale aj k zvýšeniu počtu živých organizmov.
- Kontinuita života. Vďaka rozmnožovaniu môžu viac či menej veľké skupiny jedincov (napríklad populácie a druhy) existovať pomerne dlho, pretože pokles ich počtu v dôsledku prirodzeného úhynu jedincov je kompenzovaný neustálym rozmnožovaním organizmov a nahradenie mŕtvych novorodencami.
- Kontinuita medzi generáciami. V procese rozmnožovania sa od jedincov rodičovskej generácie prenáša genetická informácia, ktorá zabezpečuje reprodukciu vlastností konkrétnych rodičov a celého druhu, ku ktorému organizmus patrí.
- Realizujú sa také vlastnosti živých vecí, ako je dedičnosť a variabilita.
2. Formy a druhy reprodukcie
Existujú dve hlavné formy reprodukcie: sexuálna a asexuálna.
Asexuálna reprodukcia
Pri nepohlavnom rozmnožovaní vzniká nový jedinec z nešpecializovaných nereprodukčných somatických buniek tela. Preto asexuálna reprodukcia zahŕňa jedného jedinca. Nepohlavné rozmnožovanie zabezpečuje rozmnožovanie veľkého počtu identických jedincov, čo je prospešné pre druhy žijúce v stálych podmienkach. Rýchle a početné rozmnožovanie potomstva identického s materským je biologickým významom nepohlavného rozmnožovania.
V prírode existujú rôzne možnosti asexuálnej reprodukcie:delenie, sporulácia, fragmentácia, pučanie, vegetatívne rozmnožovanie, klonovanie.
Môžu byť rozdelené podľa toho, koľko buniek sa podieľa na procese asexuálnej reprodukcie:
- asexuálna reprodukcia, pri ktorej dcérska generácia vzniká z jednej bunky:
- bunkové delenie
- viacnásobné bunkové delenie (schizogónia)
- sporulácia (sporulácia)
- pučanie v jednobunkových organizmoch (kvasinky)
2) asexuálna reprodukcia, ktorá je založená na rozdelení skupiny buniek:
- vegetatívny
- fragmentácia
- pučanie v mnohobunkových organizmoch (hydra)
Existuje ďalšia klasifikácia, podľa ktorej sú možnosti asexuálnej reprodukcie:
- vegetatívne rozmnožovanie (t. j. delenie podľa častí tela matky):
divízie
Viacnásobné štiepenie (schizogónia)
Pučanie
Fragmentácia
Rozmnožovanie rastlín odrezkami, cibuľkami, listami, podzemkami
- rozmnožovanie spórami, t.j. sporulácia
Pozrime sa na typy nepohlavného rozmnožovania podrobnejšie.
divízie. Najjednoduchšia forma nepohlavného rozmnožovania. Existujú rôzne možnosti rozdelenia:
- Delenie jednoduchým zovretím s vytvorením dvoch dcérskych organizmov z jedného rodičovského organizmu. Charakteristické pre baktérie a cyanobaktérie.
- Delenie mitotickým delením jadra, po ktorom nasleduje separácia cytoplazmy. Charakteristické pre jednobunkové organizmy (veľa prvokov - améby, zelené euglena atď.; jednobunkové riasy - chlamydomonas atď.).
V oboch prípadoch sa to stane binárne rozdelenie, t.j. do dvoch buniek. Je však možná aj iná možnosť:
- Viacnásobné štiepenie (schizogónia). Najprv sa opakovane rozdelí jadro a potom sa cytoplazma rozdelí na časti. Napríklad rozmnožovanie Plasmodium falciparum (pôvodca malárie) v ľudských erytrocytoch. V tomto prípade plazmódia mnohokrát podliehajú opakovanému deleniu jadra, po ktorom sa cytoplazma delí. Výsledkom je, že z 1 plazmódia vznikne 12-24 dcérskych organizmov.
Tvorba spór (sporulácia). Veľmi rozšírený spôsob nepohlavného rozmnožovania medzi živými organizmami a nachádza sa takmer vo všetkých rastlinách, hubách a niektorých prvokoch (napríklad typ sporozoan), ako aj v prokaryotických organizmoch (veľa baktérií, modrozelených rias).
Spore Ide o bunku pokrytú vyvinutým ochranným obalom - spórovou membránou, ktorá jej umožňuje odolávať účinkom rôznych nepriaznivých faktorov prostredia.Pre svoju malú veľkosť obsahuje výtrus zvyčajne len minimálne zásoby živín. V mnohých organizmoch je hlavnou jednotkou šírenia, pretože veľké množstvo vytvorených svetelných spór sa voľne prenáša na veľké vzdialenosti pohybom vzdušných hmôt a vodných tokov.Spóry sa často tvoria vo veľkých množstvách a majú zanedbateľnú hmotnosť, čo uľahčuje ich šírenie vetrom, ako aj zvieratami, najmä hmyzom.
Okrem toho v niektorých formách (protozoá, huby) môžu spóry pôsobiť ako špeciálne štádium životného cyklu, čo im umožňuje „prežiť“ účinky nepriaznivých faktorov prostredia.Bakteriálne spóry, prísne vzaté, neslúžia na rozmnožovanie, ale na prežitie v nepriaznivých podmienkach, keďže každá baktéria produkuje len jednu spóru. Bakteriálne spóry patria medzi najodolnejšie: často znesú napríklad ošetrenie silnými dezinfekčnými prostriedkami a vyváranie vo vode.
V mnohých rastlinách proces tvorby spór(sporogenéza) prebieha v špeciálnych vakovitých štruktúrach nazývaných sporangia. Spóry môžu byť pohyblivé v dôsledku prítomnosti bičíkového aparátu (v tomto prípade sa nazývajú zoospóry), a nehybné, zbavené schopnosti aktívneho pohybu.
Zoospóry sú pozorované v niektorých zelených riasach. Medzi zvieratami sa sporulácia pozoruje v malarickom plazmódiu a celej skupine sporozoanov (jednobunkových organizmov).
Pučanie. Nachádza sa v jednobunkových organizmoch, napríklad v kvasinkách a niektorých typoch nálevníkov, ako aj v mnohobunkových organizmoch, napríklad v zástupcoch coelenterátneho typu (hydra), ako aj v plášťovcoch (trieda ascidiánov).
U jednobunkových organizmov táto metóda spočíva vo vytvorení tuberkulózy (výrastku) s jadrom na materskej bunke, ktorá sa následne oddelí a stane sa samostatným organizmom.
Pre mnohobunkové organizmy táto metóda spočíva v tom, že najprv sa na tele materského jedinca objaví malý tuberkul, ktorý sa zväčšuje, potom sa objavia základy všetkých štruktúr a orgánov charakteristických pre materský organizmus. Potom nastáva oddelenie (pučanie) dcérskeho jedinca. Potom mladý, novo oddelený organizmus rastie a dosahuje veľkosť pôvodného exempláru.
Ryža. Pučanie v koelenterátoch (hydra), 1 dospelý organizmus, 2 dcérske pučiace organizmy.
Neobvyklá forma pučania je opísaná u sukulentnej rastliny Bryophyllum - xerofyt, často pestovaný ako okrasná izbová rastlina: pozdĺž okrajov jeho listov sa vyvíjajú miniatúrne rastliny vybavené malými koreňmi; tieto „púčiky“ nakoniec odpadnú a začnú existovať ako samostatné rastliny.
Fragmentácia rozdelenie jedinca na dve alebo viac častí, z ktorých každá sa vyvinie v nový organizmus. Fragmentácia sa vyskytuje napríklad u vláknitých rias, ako je Spirogyra. Vlákno Spirogyra sa môže na akomkoľvek mieste rozpadnúť na dve časti, z ktorých sa následne vytvoria dva organizmy.
Fragmentáciu pozorujeme aj u niektorých nižších živočíchov, ktoré si na rozdiel od viac organizovaných foriem zachovávajú významnú schopnosť regenerácie z relatívne slabo diferencovaných buniek. Napríklad telo nemerteanov (skupina primitívnych červov, najmä morských) sa obzvlášť ľahko roztrhne na mnoho častí, z ktorých každá môže v dôsledku regenerácie dať vzniknúť novému jedincovi. V tomto prípade je regenerácia normálnym a regulovaným procesom; u niektorých živočíchov (napríklad hviezdice) však k obnove z jednotlivých častí dochádza až po náhodnej fragmentácii. Fragmentácia sa pozoruje u špongií, coelenterátov (hydras), medúz, ostnokožcov, annelidov a plochých červov. Niekedy je táto schopnosť tak dobre vyvinutá, že sa zo samostatného fragmentu obnoví celý jedinec.
Ryža. . Regenerácia hviezdice z jedného lúča. A, B, C postupné fázy regenerácie
Vegetatívne rozmnožovanieide o rozmnožovanie, pri ktorom sa z časti oddelenej od materského organizmu vyvinie nový dcérsky organizmus. V tomto prípade sa od materského exempláru oddelí pomerne dobre diferencovaná časť, ktorá sa ďalej vyvíja v samostatnú rastlinu. Alebo rastlina vytvára špeciálne štruktúry špeciálne určené na vegetatívne rozmnožovanie. Napríklad cibuľky, hľuzy, hľuzy, pakorene, úponky, púčiky. Niektoré z týchto štruktúr slúžia aj na ukladanie živín, čo umožňuje rastline prežiť obdobia nepriaznivých podmienok, ako je chlad alebo sucho. Vegetatívne rozmnožovanie je charakteristické pre mnohé skupiny rastlín, od rias až po kvitnúce rastliny.
V zásade sa vegetatívne rozmnožovanie prakticky nelíši od fragmentácie alebo pučania, ale tradične sa tento termín používa na rastlinné organizmy a len niekedy na zvieratá, na rozdiel od fragmentácie a pučania.
Klonovanie. Ide o umelú metódu rozmnožovania, ktorá sa prirodzene nevyskytuje. Rozšíril sa až v posledných 30-40 rokoch a čoraz viac sa využíva na hospodárske účely. Existuje množstvo špeciálnych techník, ktoré umožňujú klonovať niektoré rastliny a živočíchy. Klonovať Výrazom "nepohlavné rozmnožovanie" sa rozumie geneticky identické potomstvo získané od jedného jedinca v dôsledku jedného alebo druhého spôsobu nepohlavného rozmnožovania. Odtiaľ pochádza aj názov tejto metódy.
Klonovacie experimenty sa obzvlášť často vykonávajú na rastlinách, čo je spôsobené ich vysokou schopnosťou regenerácie. Jednotlivé bunky sa umiestnia na živné pôdy, kde sa delia a pomocou špeciálnych techník sa získa neorganizovaná masa buniek, príp. kalus. Potom spôsobujú diferenciáciu primárnych homogénnych kalusov a tvorbu rôznych tkanív a orgánov a v konečnom dôsledku celého rastlinného organizmu, ktorý má všetky rovnaké vlastnosti ako ten pôvodný, z ktorého boli bunky odobraté.
Pomocou metódy klonovania je možné získať rôzne hybridné formy. Pomocou enzýmov alebo ultrazvuku sa teda odstraňujú bunkové steny rastlinných buniek, po čom môžu výsledné „nahé“ protoplasty splývať, čím vznikajú hybridné bunky (napríklad hybridy paradajka-zemiak alebo tabak-petúnia). Potom sa obnovia bunkové steny, vytvorí sa kalus a potom celá hybridná rastlina.
V prípade zvierat sa používa nasledujúci spôsob: jadro vajíčka sa odstráni alebo zničí a na jeho miesto sa umiestni jadro somatickej bunky (napríklad epiteliálna bunka). V budúcnosti sa z takéhoto vajíčka dá získať organizmus, ktorý má identické vlastnosti s darcom zvieracieho jadra. Týmto spôsobom môžete získať klony niektorých zvierat pazúrových žiab(Xenopus), mloky (Triturus). V súčasnosti sa podarilo získať dokonca klony cicavcov, ako je napríklad známa ovca Dolly. Vývoj klonovacích techník vykonáva špeciálny odbor biológie - biotechnológia, ktorého úlohy nie sú obmedzené na toto.
Sexuálna reprodukcia
Sexuálna reprodukcianazývaná výmena generácií a vývoj organizmov založený na fúzii špecializovaných zárodočných buniek ( gaméty ) a vznik zygoty. V pohlavných žľazách dochádza k tvorbe gamét (ženské a mužské, vajíčka a spermie).
Pohlavné rozmnožovanie je typické pre veľkú väčšinu živých bytostí, je progresívnejšie v porovnaní s nepohlavným rozmnožovaním a má obrovské genetické výhody. Pohlavné rozmnožovanie najlepšie zabezpečuje genetickú diverzitu potomstva, pretože existuje kombinácia génov, ktoré predtým patrili obom rodičom. Rôznorodosť genotypov jedincov, ktorí tvoria druh, poskytuje možnosť úspešnejšej a rýchlejšej adaptácie druhu na meniace sa podmienky prostredia.
Počas oplodnenia sa gaméty spájajú a vytvárajú diploid zygota z ktorých sa v procese vývoja získava zrelý organizmus. Gaméty sú haploidné – obsahujú jednu sadu chromozómov, zygota je diploidná, obsahuje dvojitú sadu chromozómov. Toto je prvá bunka budúceho organizmu.
Gamety sú zvyčajne dvoch typov - mužské a ženské. Sú produkované mužskými a ženskými rodičmi, ak ide o druh dvojdomý; ( také sú niektoré kvitnúce rastliny, väčšina zvierat a ľudia) alebo tým istým jedincom (harmafroditizmus).
Charakteristické pre samce a samice zvieratpohlavný dimorfizmus- t.j. sexuálne rozdiely v štruktúre, vzhľade (veľkosť, farba a iné vlastnosti), ako aj v správaní. U živočíchov sa vyskytuje už v nižších štádiách evolučného vývoja, napríklad u okrúhlych helmintov a článkonožcov, a najväčší prejav dosahuje u stavovcov, u ktorých sú vonkajšie rozdiely medzi samcami a samicami veľmi výrazné. V rastlinách tých druhov, ktoré sa vyznačujú prítomnosťou samčích a samičích jedincov, sa vyskytuje aj sexuálny dimorfizmus, ktorý je však veľmi mierne vyjadrený.
Porovnanie pohlavného a nepohlavného rozmnožovania je uvedené v tabuľke. 5
Tabuľka 5
Porovnanie nepohlavného a sexuálneho rozmnožovania
|
Asexuálna reprodukcia |
Sexuálna reprodukcia (okrem baktérií) |
|
|
Jeden rodič |
Zvyčajne dvaja rodičia |
|
|
Netvoria sa gaméty |
Vznikajú haploidné gaméty, ktorých jadrá sa spájajú (oplodnenie) a vytvárajú diploidnú zygotu |
|
|
Žiadna meióza |
V určitom štádiu životného cyklu nastáva meióza, ktorá zabraňuje zdvojeniu chromozómov v každej generácii. |
|
|
Potomkovia sú identickí so svojimi rodičmi. Jediným zdrojom genetických variácií sú náhodné mutácie |
Potomkovia nie sú identickí rodičovských jedincov. Vykazujú genetické variácie vyplývajúce z genetickej rekombinácie. To podporuje prirodzený výber najsilnejších a najschopnejších jedincov a následne evolúciu. |
|
|
Charakteristické pre rastliny, niektoré nižšie živočíchy a mikroorganizmy. Nenachádza sa u vyšších zvierat |
Charakteristické pre väčšinu rastlín a živočíchov |
|
|
Často vedie k rýchlemu vytvoreniu veľkého počtu potomkov |
Menej rýchly nárast počtu |
Sexuálna reprodukcia, v závislosti od pomeru veľkosti a funkcie zárodočných buniek, môže byť v troch možnostiach:
Izogamia (riasy, prvoky). Mužské a ženské reprodukčné bunky majú rovnakú veľkosť, štruktúru a pohyblivosť.
Heterogamia. Bunky sa líšia veľkosťou a štruktúrou.
Oogamia. Toto je jeden z variantov heterogamie, keď je vajíčko veľké a nehybné a spermie je menšia, má orgán pohybu a má pohyblivosť.
Existujú špeciálne formy sexuálneho rozmnožovania, ako je hermafroditizmus a partenogenéza.
Hermafroditizmus. Pojem „hermafroditizmus“ je spojením gréckych mien Hermes (boh mužskej krásy) a Afrodita (bohyňa ženskej krásy).
Vo väčšine hermafroditných druhov však oplodnenie zahŕňa gaméty pochádzajúce z rôznych jedincov a majú početné genetické, morfologické a fyziologické úpravy, ktoré bránia samooplodneniu a podporujú krížové oplodnenie. Napríklad u mnohých prvokov bráni samooplodneniu genetická inkompatibilita, u mnohých kvitnúcich rastlín štruktúra androecium a gynoecium a u mnohých zvierat skutočnosť, že vajíčka a spermie sa tvoria u toho istého jedinca v rôznych časoch.
Fenomén skutočného hermafroditizmu sa vyskytuje aj u organizovanejších tvorov. Vyskytuje sa najmä u cicavcov. Napríklad u ošípaných sa niekedy na jednej strane tela pozoruje vývoj vaječníkov a na druhej vývoj semenníkov (semenníkov) alebo vývoj kombinovaných štruktúr (ovotestis), v oboch prípadoch syntéza funkčne prebiehajú aktívne vajíčka a spermie. Takéto zvieratá sú klasifikované ako „stredné“ pohlavie, pričom väčšina jedincov stredného sexuálneho typu sú samice s dvoma chromozómami XX. Podobný jav bol zaznamenaný u kôz.
Skutočný hermafroditizmus sa vyskytuje aj u ľudí, ktorý je výsledkom vývojových porúch. Genotypy hermafroditov sú 46XX alebo 46 XY , pričom väčšina prípadov odkazuje na XX (asi 60 %). Známy je aj falošný hermafroditizmus, kedy jedinci majú vonkajšie pohlavné orgány a sekundárne pohlavné znaky charakteristické pre obe pohlavia, no produkujú zárodočné bunky len jedného typu – mužského alebo ženského.
Väčšina kvitnúcich rastlín má hermafroditné kvety, ktoré sa zvyčajne nazývajú obojpohlavné, pretože každý kvet obsahuje piestik a tyčinky. Z tohto dôvodu sa plody vyvíjajú zo všetkých kvetov. Pšenica, čerešňa, jablko a mnoho ďalších druhov rastlín sú obojpohlavné. Okrem bisexuálov sa počas evolúcie vyvinuli rastliny s oddelením pohlaví v rámci toho istého druhu, teda vznikli jednodomé a dvojdomé rastliny. Rastliny obsahujúce piestikové (samičie) aj stonkové (samčie) kvety sa nazývajú jednodomé. V jednodomých rastlinách sa plody vyvíjajú iba z piestikových kvetov. Kukurica je jednodomáuhorka, tekvica a iné. Na rozdiel od toho, dvojdomé rastliny sú rastliny, ktoré obsahujú buď piestikové alebo staminátové kvety (v rámci toho istého druhu). V dvojdomých rastlinách plodia len tie, ktoré majú piestikové kvety (samičie). Topoľ, jahoda a iné druhy drevín a bylín sú dvojdomé.
Partenogenéza (z gréčtiny. parthenos panna a genéza pôrod) je jednou z modifikácií pohlavného rozmnožovania, pri ktorej sa zo samičej gaméty vyvinie nový jedinec bez oplodnenia samčou gamétou. Výhodou partenogenézy je, že v niektorých prípadoch zvyšuje rýchlosť rozmnožovania a u sociálneho hmyzu umožňuje regulovať počet potomkov každého typu. Partenogenéza môže byť obligátna (povinná) a fakultatívna (voliteľná).
Napríklad vo včele medonosnej(Apis mellifera) kráľovná kladie oplodnené vajíčka(2п = 32), z ktorých vo vývoji vznikajú samice (kráľovné alebo robotnice) a neoplodnené vajíčka(P = 16), ktoré produkujú samce (drony), ktoré produkujú spermín skôr mitózou ako meiózou.
Ryža. Schéma typov partenogenézy.
- Bunkový cyklus. Mitóza. meióza
Bunkové delenie je základom rozmnožovania a individuálneho vývoja organizmov.
Všetky živé organizmy sa skladajú z buniek. Vývoj, rast a formovanie typickej stavby tela sa uskutočňuje rozmnožovaním jednej alebo skupiny pôvodných buniek. V procese života sa niektoré bunky tela opotrebúvajú, starnú a odumierajú. Na udržanie štruktúry a normálneho fungovania musí telo produkovať nové bunky, ktoré nahradia staré. Jediný spôsob, ako vytvoriť bunky, je rozdeliť predchádzajúce bunky.
Bunkové delenie životne dôležitý proces pre všetky organizmy. V ľudskom tele sa skladá z približne 10 13 bunky, niekoľko miliónov z nich sa musí rozdeliť každú sekundu.
Boli opísané tri spôsoby delenia eukaryotických buniek: amitóza (priame rozdelenie), mitóza (nepriame delenie) a meióza (redukčné delenie).
Amitóza pomerne zriedkavý a málo prebádaný spôsob delenia buniek. Opisuje sa pri starnúcich a patologicky zmenených bunkách. Pri amitóze je medzifázové jadro rozdelené zúžením a nie je zabezpečená rovnomerná distribúcia dedičného materiálu. Často sa jadro delí bez následnej separácie cytoplazmy a vznikajú dvojjadrové bunky. Bunka, ktorá prešla amitózou, nie je následne schopná vstúpiť do normálneho mitotického cyklu. Preto sa amitóza spravidla vyskytuje v bunkách a tkanivách odsúdených na smrť, napríklad v bunkách embryonálnych membrán cicavcov a v nádorových bunkách.
Mitóza univerzálna metóda delenia eukaryotických buniek. Jej trvanie v živočíšnych bunkách je asi 1 hod. Mitóza je kontinuálny proces, ktorý sa bežne delí na štyri fázy: profázu, metafázu, anafázu a telofázu.
Postupnosť dejov, ktorá nastáva medzi vznikom danej bunky a jej rozdelením na dcérske bunky, sa nazývabunkový cyklus. Tento cyklus pozostáva z troch hlavných etáp:
1. Medzifáza. Obdobie intenzívnej syntézy arast. Bunka syntetizuje mnoho látok potrebných pre jej rast a realizáciu všetkých jej vlastných funkcií. Počas interfázy dochádza k replikácii DNA.
2. Mitóza. Toto je proces jadrového delenia (karyokinéza), pri ktorom sú chromatidy od seba oddelené a redistribuované ako chromozómy medzi dcérske bunky.
3. Cytokinéza je proces delenia cytoplazmy (cytokinéza) medzi dve dcérske bunky.
Dĺžka bunkového cyklu závisí od typu bunky a od vonkajších faktorov, akými sú teplota, živiny a kyslík. Bakteriálne bunky sa môžu deliť každých 20 minút, bunky črevného epitelu - každých 8-10, bunky v špičke koreňa cibule - každých 20 hodín a mnohé bunky nervového systému sa nikdy nedelia.
Vzťah medzi procesmi v čase je znázornený v ryža.

Ryža . Fázy bunkového cyklu
Medzifáza pozostáva z niekoľkých období: G1, S, G2.
Obdobie G 1 nazývané presyntetické. Najviac variabilné v trvaní. V tomto čase sa v bunke aktivujú procesy biologickej syntézy, predovšetkým štruktúrne a funkčné proteíny. Bunka rastie a pripravuje sa na ďalšie obdobie. Počas tohto obdobia prebiehajú intenzívne procesy biosyntézy. Tvorba mitochondrií, chloroplastov (v rastlinách), endoplazmatického retikula, lyzozómov, Golgiho aparátu, vakuol a vezikúl. Jadierko produkuje rRNA, mRNA a tRNA; tvoria sa ribozómy; bunka syntetizuje štrukturálne a funkčné proteíny. Intenzívny bunkový metabolizmus riadený enzýmami. Bunkový rast. Vznik látok, ktoré potláčajú alebo stimulujú nástup ďalšej fázy.
Obdobie S nazývané syntetické. Toto je hlavné obdobie mitotického cyklu. V deliacich sa cicavčích bunkách trvá asi 6 x 10 hod. Tu dochádza k replikácii DNA. Syntéza proteínových molekúl nazývaných históny, ktoré sa viažu na každé vlákno DNA. Každý chromozóm sa zmení na dve chromatidy.
Obdobie G2 sa nazýva postsyntetické.Je pomerne krátka, v bunkách cicavcov je to asi 2 x 5 hodín.V tomto čase sa zdvojnásobuje počet centriol, mitochondrií a plastidov, prebiehajú aktívne metabolické procesy, akumulujú sa bielkoviny a energia pre nastávajúce delenie. Bunka sa začína deliť. Vyskytujú sa intenzívne procesy biosyntézy. Rozdelenie mitochondrií a chloroplastov. Zvýšené zásoby energie. Replikácia centriolov (v tých bunkách, kde sú prítomné) a začiatok tvorby vretienka
Mitóza podmienene rozdelené do štyroch fáz: profáza, metafáza, anafáza a telofáza
Profáza . Špiralizácia DNA začína a postupne sa zvyšuje v jadre. Chromozómy sa skracujú, zhrubnú, zviditeľnia sa a získajú typickú bichromatidovú štruktúru. Jadierko postupne mizne. V cytoplazme sú mikrotubuly orientované okolo každého páru centriolov a tvoria vretienkové centrá. Centrioly sa pohybujú k rôznym pólom, mikrotubuly sa rozprestierajú pozdĺž bunkovej osi a začína sa tvorba achromatínového vretienka. Jadrový obal sa rozpadá na samostatné malé fragmenty. Chromozómy sa pohybujú smerom k stredu bunky
Metafáza . Chromozómy sú maximálne špirálovité a usporiadané tak, že ich centroméry ležia v rovnakej rovine – rovine bunkového rovníka. Vzniká metafázová platnička Dokončuje sa tvorba mitotického vretienka. Centrioly sú umiestnené v pároch na opačných póloch a vlákna vretienka z rôznych pólov sú pripojené k centromére každého chromozómu.
Anaphase . Toto je najkratšia fáza mitózy. Tu dochádza k pozdĺžnemu štiepeniu každého chromozómu, redukcii vlákienvretienka a divergencia chromatíd (dcérskych chromozómov) smerom k pólom bunky.
Telofáza . Dcérske chromozómy, pozostávajúce z jednej chromatidy, dosahujú póly bunky. DNA, ktorá ich tvorí, sa začne despirovať, objaví sa jadierko, okolo každej skupiny dcérskych chromozómov sa vytvorí jadrová membrána a postupne sa rozpadnú vlákna achromatínového vretienka. Jadrové štiepenie je dokončené.
Začína sa cytoplazmatické delenie(cytotómia) a vytvorenie priehradky medzi dcérskymi bunkami. Živočíšne bunky vykonávajú cytotómiu stiahnutím cytoplazmatickej membrány. U rastlín sa v rovine bunkového rovníka vytvára membránová priehradka, ktorá rastie laterálne a dosahuje bunkovú stenu. V dôsledku toho sa vytvoria dve úplne oddelené dcérske bunky.
Poďme na to zmena dedičného materiálupočas mitotického cyklu. Hlavnými udalosťami mitotického cyklu sú replikácia DNA vyskytujúce sa v interfáze a vedúce k zdvojnásobeniu množstva dedičnej informácie, asegregácia chromatíd,vyskytujúce sa v anafáze mitózy a zabezpečujúce rovnomernú distribúciu dedičnej informácie medzi dcérskymi bunkami. Dedičný materiál vykonáva tieto procesy, pričom má rôzne štrukturálne formy. Prebieha replikatívna syntézamedzifázový chromatín, inv ktorom je molekula DNA v relatívne despiralizovanom stave. Distribúcia genetickej informácie sa uskutočňujemitotické chromozómy,v ktorom je DNA maximálne špirálovitá.
V mitotickom cykle sa mení aj množstvo dedičného materiálu. Ak je počet chromozómov v haploidnej sade označený písmenom p (v diploidnom súbore 2n) a počet molekúl DNA je označený písmenom s, potom je možné sledovať zmenu vzorca jadra somatickej bunky v rôznych štádiách mitotického cyklu. Predtým S -obdobie, keď každý chromozóm pozostáva z jednej molekuly DNA, celkové množstvo DNA v jadre zodpovedá počtu chromozómov v ňom a vzorec diploidnej bunky má tvar 2p2s. Po replikácii, keď sa DNA každého chromozómu zdvojnásobí, celkové množstvo DNA v jadre sa zdvojnásobí a vzorec bunky nadobudne formu 2n4c. V dôsledku separácie chromatidov v anafáze mitózy dcérske jadrá dostávajú diploidnú sadu jednochromatidových chromozómov. Vzorec dcérskych buniek sa opäť stáva 2p2s.
Biologický význam mitózyje, že v dôsledku tohto spôsobu delenia vznikajú bunky s dedičnou informáciou, ktorá je kvalitatívne a kvantitatívne identická s informáciou materskej bunky. Rovnomerná distribúcia dedičného materiálu je zabezpečená procesmi replikácie DNA a zdvojenia chromozómov v interfáze mitotického cyklu, ako aj spiralizáciou a rovnomernou distribúciou chromatíd medzi dcérskymi bunkami počas mitózy. Mitóza zabezpečuje udržanie stálosti karyotypu počas niekoľkých bunkových generácií a slúži ako bunkový mechanizmus pre procesy rastu a vývoja tela, ako aj regeneráciu a asexuálnu reprodukciu.
Pôsobenie množstva faktorov prostredia môže narušiť normálny priebeh mitózy a viesť k poškodeniu chromozómov, ako aj k zmenám v počte jednotlivých chromozómov alebo celých chromozómových sád v somatických bunkách tela. Patologické mitózy môžu spôsobiť množstvo chromozomálnych ochorení. Patologické mitózy sa pozorujú obzvlášť často v nádorových bunkách.
Schéma mitózy:
MITÓZA
|
1. Medzifáza Často sa nesprávne nazýva pokojová fáza. Trvanie interfázy sa mení a závisí od funkcie danej bunky. Toto je obdobie, počas ktorého bunka normálne syntetizuje organely a zväčšuje svoju veľkosť. Jadierka sú jasne viditeľné a aktívne syntetizujú ribozomálny materiál. Tesne pred delením buniek sa replikuje DNA a históny každého chromozómu. Každý chromozóm je teraz reprezentovaný párom chromatidov, ktoré sú navzájom spojené centromérou. Látka chromozómov je zafarbená a nazýva sa chromatín, ale tieto štruktúry samotné sú ťažko viditeľné. |
|
|
2. Profázujte Zvyčajne ide o najdlhšiu fázu bunkového delenia. Chromatidy sa skracujú (až o 4 % svojej pôvodnej dĺžky) a hrubnú v dôsledku ich špirálovitosti a kondenzácie. Pri farbení sú chromatidy jasne viditeľné, ale centroméry nie sú viditeľné. V rôznych pároch chromatidov je centroméra umiestnená inak. V živočíšnych bunkách a v nižších rastlinách sa centrioly rozchádzajú k opačným pólom bunky. Z každého centriolu vychádzajú krátke mikrotubuly vo forme lúčov, ktoré spoločne vytvárajú hviezdu. Jadierka sa zmenšujú, pretože ich nukleová kyselina je čiastočne prenesená do určitých párov chromatidov. Ku koncu profázy sa jadrová membrána rozpadne a vytvorí sa štiepne vreteno. |
|
|
3. Metafáza Páry chromatidov sú svojimi centromérmi pripevnené k vláknam vretena (mikrotubuly) a pohybujú sa hore a dole po vretene, kým sa ich centroméry nezarovnajú pozdĺž rovníka vretena kolmo na jeho os. |
|
|
4. Anafáza Toto je veľmi krátka etapa. Každá centroméra sa rozdelí na dve a vretenové vlákna ťahajú dcérske centroméry k opačným pólom. Centroméry ťahajú za sebou chromatidy, ktoré sa od seba oddelili a ktoré sa teraz nazývajú chromozómy. |
|
|
5. Telofáza Chromozómy dosahujú bunkové póly, despirujú, predlžujú sa a už sa nedajú jasne rozlíšiť. Vretenové vlákna sú zničené a centrioly sú replikované. Okolo chromozómov na každom póle sa vytvorí jadrová membrána. Znovu sa objaví jadierko. Po telofáze môže bezprostredne nasledovať cytokinéza (rozdelenie celej bunky na dve). |
|





Meiosis (z gréckeho meiosis redukcia) zvláštny spôsob delenia buniek, čo vedie k zníženiu počtu chromozómov v nich na polovicu. Meióza je ústredným článkom gametogenéza u zvierat a sporogenéza v rastlinách. Meióza pozostáva z dvoch po sebe nasledujúcich delení, ktorým predchádza jediná replikácia DNA. Všetky látky a energia potrebné pre obe delenia sa ukladajú počas medzifázy predchádzajúcej meióze ja Medzifáza II prakticky chýbajú a divízie rýchlo nasledujú jedno za druhým. V každom z meiotických delení sa rozlišujú rovnaké štyri štádiá: profáza, metafáza, anafáza a telofáza, ktoré sú charakteristické pre mitózu, ale líšia sa mnohými znakmi.
Prvé meiotické delenie (meióza ja ) vedie k zníženiu počtu chromozómov na polovicu a nazýva sa redukcia. V dôsledku toho z jednej diploidnej bunky(2p 4c) vznikajú dve haploidné bunky(p 2c) bunky.
Profáza I meióza je najdlhšia a najkomplexnejšia. Okrem procesov helixácie DNA a tvorby vretienka typických pre profázu mitózy, ja Vyskytujú sa dve mimoriadne dôležité biologické udalosti: konjugácia alebo synapsia homológne chromozómy a prejsť.
Konjugácia je proces tesnej blízkosti homológnych chromozómov. Vznikajú tieto párové chromozómy bivalentný a sú zachované vo svojom zložení pomocou špeciálnych bielkovín. Keďže každý chromozóm pozostáva z dvoch chromatidov, bivalentný zahŕňa štyri chromatidy a nazýva sa tiež notebook. Diploidná bunka produkuje P bivalenty. Po konjugácii získa vzorec bunky formu prejsť.
Na niektorých miestach bivalentu sa chromatidy konjugovaných chromozómov pretínajú, lámu a vymieňajú si zodpovedajúce úseky. Tento proces výmeny fragmentov homológnych chromozómov sa nazýva crossover. Zabezpečuje tvorbu nových kombinácií otcovských a materských génov v chromozómoch budúcich gamét. Kríženie môže nastať v niekoľkých oblastiach (viacnásobné prekríženie), čo poskytuje vyšší stupeň rekombinácie dedičnej informácie v gamétach. Do konca profázy ja Stupeň spiralizácie chromozómov sa zvyšuje, chromatidy sa stávajú jasne rozlíšiteľnými, závity vretienka z každého pólu sú pripojené k centromére jedného z bivalentných chromozómov. Jadrový obal je zničený a bivalenty sú nasmerované k rovníkovej rovine bunky.
V metafáze I Meióza dokončuje tvorbu vretena, bivalenty sú inštalované v rovníkovej rovine bunky. Vretienkové vlákna z jedného pólu sú pripojené k centromére každého chromozómu.
V anafáze I Pri meióze sa homológne chromozómy pod pôsobením vretenových nití od seba vzďaľujú a smerujú k opačným pólom bunky. Výsledkom je, že na každom póle bunky ahaploidný súborchromozómy, obsahujúce jeden bichromatidový chromozóm z každého páru homológnych chromozómov. V anafáze ja chromozómy rôznych párov, t.j. nehomologické chromozómy sa správajú úplne nezávisle od seba, čím zabezpečujú tvorbu širokej škály kombinácie otcovské a materské chromozómy v haploidnom súbore budúcich gamét. Počet takýchto kombinácií zodpovedá vzorcu 2 P, kde p počet párov homológnych chromozómov. U ľudí sa táto hodnota rovná 2, t.j. 8.4 V ľudských gamétach je možných 10 variantov kombinácií otcovských a materských chromozómov.
Takže divergencia homológnych chromozómov v anafáze ja meióza zaisťuje nielen zníženie počtu chromozómov v budúcich zárodočných bunkách, ale aj obrovskú rozmanitosť zárodočných buniek v dôsledku náhodnej kombinácie otcovských a materských chromozómov rôznych párov.
V telofáze I K meióze dochádza pri tvorbe buniek, ktorých jadrá majú haploidnú sadu chromozómov a dvojnásobné množstvo DNA, pretože každý chromozóm pozostáva z dvoch chromatidov. Bunky, ktoré sú výsledkom prvého meiotického delenia, majú vzorec p2s a po krátkej medzifáze začnú ďalšie delenie.
Druhé meiotické delenie (meióza II ) prebieha ako typická mitóza (obr. 5.3), ale líši sa tým, že bunky, ktoré do nej vstupujú, obsahujú haploidnú sadu chromozómov. V dôsledku tohto rozdelenia P bichromatidové chromozómy (r2c), štiepenie, form P jednochromatidové chromozómy(ps). Toto rozdelenie sa nazýva rovnicové (alebo vyrovnávanie).
Po dvoch po sebe nasledujúcich meiotických deleniach tak z jednej bunky s diploidnou sadou dvojchromatidových chromozómov (2x4c) vzniknú štyri bunky s haploidnou sadou jednochromatidových chromozómov.(ps).
Biologický význam meiózyspočíva vo vytváraní buniek s redukovaným súborom chromozómov a udržiavaní stálosti karyotypu v množstve generácií organizmov, ktoré sa rozmnožujú. sexuálne prenosné. Meióza slúži ako základ pre kombinačnú variabilitu, ktorá poskytuje genetickú diverzitu gamét prostredníctvom procesov kríženia, divergencie a kombinatoriky otcovských a materských chromozómov. Zmeny v štruktúre chromozómov v dôsledku nerovnomerného kríženia, narušenie divergencie všetkých alebo jednotlivých chromozómov v anafáze I a II meiotické delenia vedú k tvorbe abnormálnych gamét a môžu slúžiť ako základ pre smrť organizmu alebo rozvoj množstva chromozomálnych syndrómov u potomkov.
Schéma a stručné opisy po sebe nasledujúcich štádií meiózy v živočíšnej bunke.
|
1. Medzifáza Trvanie sa líši medzi rôznymi druhmi. Dochádza k replikácii organel a bunka sa zväčšuje. Replikácia DNA a histónov končí hlavne v premeiotickej interfáze, ale čiastočne zahŕňa aj začiatok profázy. Každý chromozóm je teraz reprezentovaný párom chromatidov spojených centromérou. Chromozomálny materiál je zafarbený, ale zo všetkých štruktúr sú jasne viditeľné iba jadierka. |
A. Skorá profáza ja |
|
2. Profázujte Najdlhšia fáza. Často sa delí na päť štádií (leptotéma, zygonéma, pachynéma, diplonéma a diakinéza), ale tu sa bude považovať za súvislú sekvenciu chromozómových zmien. A . Chromozómy sa skracujú a stávajú sa viditeľnými ako samostatné štruktúry. V niektorých organizmoch vyzerajú ako šnúry guľôčok: oblasti intenzívne zafarbeného materiálu - chromoméry - sa striedajú s oblasťami bez zafarbenia. Chromoméry sú miesta, kde je chromozomálny materiál vysoko stočený. |
|
|
B. Homologické chromozómy pochádzajúce z jadier materských a otcovských gamét sa k sebe približujú a konjugujú. Tieto chromozómy sú rovnako dlhé, ich centroméry sú na rovnakej pozícii a zvyčajne obsahujú rovnaký počet génov usporiadaných v rovnakej lineárnej sekvencii. Chromoméry homológnych chromozómov ležia vedľa seba. Proces konjugácie môže začať v niekoľkých bodoch na chromozómoch, ktoré sú potom spojené po celej dĺžke (akoby zipsom). Páry konjugovaných homológnych chromozómov sa často nazývajú bivalenty. Bivalenty sa skracujú a zahusťujú. V tomto prípade nastáva hustejšie balenie na molekulárnej úrovni a navonok viditeľné skrútenie (špiralizácia). Teraz je jasne viditeľný každý chromozóm so svojou centromérou. |
B. Profáza I |
|
IN. Homologické chromozómy, ktoré tvoria bivalent, sú čiastočne oddelené, akoby sa od seba odtláčali. Teraz môžete vidieť, že každý chromozóm pozostáva z dvoch chromatidov. Chromozómy sú stále navzájom spojené v niekoľkých bodoch. Tieto body sa nazývajú chiasmata (z gréčtiny. chiasma - kríž). V každej chiazme dochádza k výmene úsekov chromatíd v dôsledku zlomov a opätovných spojení, do ktorých sú zapojené dve zo štyroch vlákien prítomných v každej chiazme. V dôsledku toho sú gény z jedného chromozómu (napríklad otcovské - A, B, C) spojené s génmi z iného chromozómu (materského a, b , c), čo vedie k novým génovým kombináciám vo výsledných chromatidoch. Tento proces sa nazýva prekríženie. Homologické chromozómy sa po prekrížení neoddelia, pretože sesterské chromatidy (oboch chromozómov) zostávajú pevne spojené až do anafázy. |
B. Prekročenie počas profázy ja |
|
G . Chromatidy homológnych chromozómov sa naďalej navzájom odpudzujú a bivalenty nadobúdajú určitú konfiguráciu v závislosti od počtu chiazmat. Bivalenty s jednou chiazmou majú krížový tvar, s dvoma chiazmatami sú prstencovité a s tromi a viacerými tvoria na seba kolmo ležiace slučky. Na konci profázy sú všetky chromozómy úplne kondenzované a intenzívne zafarbené. V bunke nastávajú ďalšie zmeny: migrácia centriol (ak existujú) k pólom, deštrukcia jadierok a jadrovej membrány a potom tvorba vretenových filamentov. |
|
|
2. Metafáza Bivalenty sa zoraďujú v rovníkovej rovine a vytvárajú metafázovú platňu. Ich centroméry sa správajú ako jednotlivé štruktúry (hoci sa často javia ako dvojité) a organizujú k nim pripojené vretenové vlákna, z ktorých každé smeruje iba k jednému z pólov. V dôsledku slabej ťažnej sily týchto nití sa každá bivalentná nachádza v oblasti rovníka a obe jej centroméry sú od nej v rovnakej vzdialenosti, jedna pod ňou a druhá nad ňou. |
D. Neskorá metafáza ja |
|
3. Anafáza Dve centroméry prítomné v každej bivalentnej sa ešte nerozdelili, ale sesterské chromatidy už navzájom nesusedia. Vlákna vretena ťahajú centroméry, z ktorých každá je spojená s dvoma chromatidami, smerom k opačným pólom vretena. Výsledkom je, že chromozómy sú rozdelené do dvoch haploidných súborov, ktoré končia v dcérskych bunkách. |
E. Anafáza I |
|
4. Telofáza Divergencia homológnych centromér a pridružených chromatíd k opačným pólom znamená dokončenie prvého meiotického delenia. Počet chromozómov v jednej sade sa zväčšil o polovicu, ale chromozómy na každom póle pozostávajú z dvoch chromatidov. V dôsledku prekríženia pri tvorbe chiazmat sú tieto chromatidy geneticky neidentické a pri druhom meiotickom delení sa oddelia. Vretená a ich závity zvyčajne zmiznú. U živočíchov a niektorých rastlín chromatidy despirujú, na každom póle sa okolo nich vytvorí jadrová membrána a vzniknuté jadro sa dostane do interfázy. Potom začína delenie cytoplazmy (u zvierat) alebo tvorba deliacej sa bunkovej steny (u rastlín), ako pri mitóze. V mnohých rastlinách nie je pozorovaná ani telofáza, ani tvorba bunkovej steny, ani interfáza a bunka priamo prechádza z anafázy I k profáze II. |
G. Telofáza I v živočíšnej bunke |







|
Medzifáza II Toto štádium sa zvyčajne pozoruje iba v živočíšnych bunkách: jeho trvanie sa mení. Fáza S chýba a nenastáva žiadna ďalšia replikácia DNA. Procesy zapojené do druhého delenia meiózy sú svojím mechanizmom podobné tým, ktoré sa vyskytujú pri mitóze. Zahŕňajú separáciu chromatidov v oboch dcérskych bunkách, ktoré sú výsledkom prvého meiotického delenia. Druhé delenie meiózy sa od mitózy líši najmä dvoma spôsobmi: 1) v metafáze II meióza, sesterské chromatidy sú často navzájom silne oddelené; 2) počet chromozómov je haploidný. |
|
|
Profáza II V bunkách, ktoré strácajú medzifázu II , táto etapa tiež chýba. Trvanie profázy II nepriamo úmerné trvaniu telofázy ja . Jadierka a jadrové membrány sú zničené a chromatidy sú skrátené a zhrubnuté. Centrioly, ak sú prítomné, sa pohybujú k opačným pólom buniek; objavujú sa vretenové vlákna. Chromatidy sú usporiadané tak, že ich dlhé osi sú kolmé na os vretienka prvého meiotického delenia. |
Z. Profáza P |
|
Metafáza II Pri druhom delení sa centroméry správajú ako dvojité štruktúry. Usporadúvajú závity vretena smerujúce k obom pólom, a tak vyrovnávajú vreteno na rovníku. |
Metafáza II |
|
Anafáza II centrum Omer sa delí a závity vretena ich ťahajú k opačným pólom. Centroméry ťahajú pozdĺž oddelených chromatidov, ktoré sa teraz nazývajú chromozómy. Telofáza II Toto štádium je veľmi podobné telofáze mitózy. Chromozómy sa despirujú, naťahujú a potom je ťažké ich rozlíšiť. Vretenové vlákna zmiznú a centrioly sa replikujú. Okolo každého jadra, ktoré teraz obsahuje polovicu (haploidného) počtu chromozómov pôvodnej rodičovskej bunky, sa opäť vytvorí jadrová membrána. Následným delením cytoplazmy (u zvierat) alebo vytvorením bunkovej steny (u rastlín) sa z jednej pôvodnej rodičovskej bunky získajú štyri dcérske bunky. |
|


Rozdiely medzi štádiami mitózy a meiózy
|
Etapa |
Mitóza |
meióza |
|
Profáza |
Chromoméry nie sú viditeľné Homologické chromozómy sú oddelené Chiasmata sa netvoria Priechody sa nevyskytujú |
Viditeľné chromoméry Homológne chromozómy sú konjugované. Vznikajú chiasmata. Môže dôjsť k prekročeniu |
|
Metafáza |
Páry chromatíd sa nachádzajú na vretenovom rovníku Centroméry sú usporiadané v rovnakej rovine na vretenovom rovníku |
Páry chromatíd sa nachádzajú na rovníku vretena iba v druhej divízii meiózy Centroméry v prvej divízii meiózy sa nachádzajú nad a pod rovníkom v rovnakých vzdialenostiach od neho |
|
Anaphase |
Centroméry sa delia. Chromatidy sa oddelia. Divergentné chromatidy sú identické |
Centroméry sa delia až v druhom štádiu meiózy. Počas druhého delenia meiózy sa chromatidy oddelia. Pri prvom delení sa oddelia celé chromozómy. Divergujúce chromozómy nemusia byť identické v dôsledku kríženia |
|
Telofáza |
Počet chromozómov v dcérskych bunkách je rovnaký ako v rodičovských bunkách Dcérske bunky obsahujú oba homológne chromozómy (v diploidoch) |
Počet chromozómov v dcérskych bunkách je polovičný ako v rodičovských bunkách Dcérske bunky obsahujú iba jeden z každého páru homológnych chromozómov |
|
Kde sa vyskytuje tento typ delenia? |
Možné v haploidných, diploidných a polyploidných bunkách Vyskytuje sa pri tvorbe somatických buniek a niektorých spór, ako aj pri tvorbe gamét v rastlinách, v ktorých dochádza k striedaniu generácií |
Len v diploidných a polyploidných bunkách Počas gameto- alebo sporogenézy |
- Štruktúra gamét. Gametogenéza
Sexuálna reprodukcia sa uskutočňuje pomocou špecializovaných zárodočných buniek nazývaných gaméty. Ženské pohlavné bunky sú tzv vajcia, samec spermie.Gamety sa od somatických buniek líšia predovšetkým tým, že majú polovičný počet chromozómov, ako aj nízku úroveň metabolických procesov.
Vajíčka relatívne veľké nepohyblivé bunky, zvyčajne okrúhleho tvaru; Okrem typických organel obsahuje cytoplazma inklúzie rezervných živín vo forme žĺtka ( ryža .). V jadrách vajíčok sa tvorí veľa kópií ribozomálnych génov a mRNA, ktoré zabezpečujú syntézu životne dôležitých bielkovín budúceho embrya. Vajcia rôznych organizmov sa líšia množstvom a povahou distribúcie žĺtka v nich. Existuje niekoľko druhov vajec.

Isolecithalsa nazývajú relatívne malé vajcia s malým množstvom rovnomerne rozloženého žĺtka. Jadro v nich je umiestnené bližšie k stredu. Takéto vajíčka sa nachádzajú u červov, lastúrnikov a ulitníkov, ostnokožcov a lancetov.Stredne telolecitálnyVajíčka jesetera a obojživelníkov majú priemer asi 1,5 × 2 mm a obsahujú priemerné množstvo žĺtka, ktorého väčšina je sústredená na jednom z pólov (vegetatívna). Na opačnom póle (zviera), kde je málo žĺtka, je jadro vajíčka.
Silne telolecitálneVajíčka niektorých rýb, plazov, vtákov a vajcorodých cicavcov obsahujú veľa žĺtka, ktorý zaberá takmer celý objem cytoplazmy vajíčka. Na zvieracom póle je zárodočný disk s aktívnou cytoplazmou bez žĺtka. Veľkosti týchto vajec sú veľké, 10 15 mm alebo viac.
Alecital vajíčka prakticky neobsahujú žĺtok, majú mikroskopicky malé rozmery (0,1 × 0,3 mm) a sú charakteristické pre placentárne cicavce vrátane ľudí.
Spermie alebo spermie sú veľmi malé pohyblivé samčie gaméty (napríklad ľudské spermie sú 50 x 70 µm dlhé a krokodílie 20 µm) tvorené mužskými gonádami – semenníkmi; ich počet sa pohybuje v miliónoch. Tvar spermií sa u rôznych zvierat líši, ale ich štruktúra je rovnaká, väčšina z nich má hlavu a krk. V hlave spermie je jadro obsahujúce haploidný počet chromozómov a pokryté akrozóm. Akrozóm je špeciálna štruktúra, modifikovaný Golgiho komplex, ktorý obsahuje enzýmy na rozpustenie membrány vajíčka pri oplodnení, ohraničené membránou. Krk obsahuje početné mitochondrie a dva centrioly. Chvost tvorený mikrotubulami vyrastá z krku a zabezpečuje pohyblivosť spermií. Stredná časť je rozšírená vďaka početným mitochondriám, ktoré obsahuje, zostavených do špirály okolo bičíka. Tieto mitochondrie poskytujú energiu pre kontraktilné mechanizmy, ktoré zabezpečujú pohyb bičíka. Hlavné a chvostové časti spermií majú štruktúru charakteristickú pre bičíky.

Ak sa pozriete na hlavu ľudskej spermie zhora, zdá sa, že je okrúhla a pri pohľade zboku sa zdá byť sploštená. Samotný bičíkový pohyb nestačí na to, aby spermie prekonali vzdialenosť od vagíny k miestu, kde dochádza k oplodneniu. Hlavnou lokomočnou úlohou spermií je zhrnúť sa okolo oocytu a orientovať sa špecifickým spôsobom predtým, ako preniknú cez membrány oocytu.
Tvorba gamét (gametogenéza).Bunky germinálneho epitelu u mužov A Ženské pohlavné žľazy prechádzajú radom postupných mitotických a meiotických delení, ktoré sa súhrnne nazývajú gametogenéza, čo vedie k tvorbe zrelých mužských gamét (spermatogenéza) a ženských gamét (oogenéza). V oboch prípadoch je proces rozdelený do troch fáz – reprodukčná fáza, rastová fáza a fáza dozrievania. Reprodukčná fáza zahŕňa opakované mitotické delenia vedúce k tvorbe mnohých spermatogónií alebo oogónií. Každý z nich prechádza obdobím rastu v rámci prípravy na prvé meiotické delenie a následnú cytokinézu. Potom začína fáza dozrievania, počas ktorej dochádza k prvému a druhému meiotickému deleniu s následná diferenciácia haploidných buniek A tvorba zrelých gamét.
Vývoj spermií (spermatogenéza).Spermie (spermie) sa tvoria ako výsledok série postupných delení buniek, súhrnne tzv.spermatogenéza,nasleduje zložitý diferenciačný proces tzv spermiogenéza (obr. 20.31 ). Proces tvorby spermií trvá približne 70 dní; na 1 g hmotnosti semenníka sa tvorí 10 7 spermií za deň. Epitel semenného tubulu pozostáva z vonkajšej vrstvy buniekzárodočný epitela asi šesť vrstiev buniek vytvorených v dôsledku opakovaného delenia buniek tejto vrstvy (obr. 20.32 a 20.33); tieto vrstvy zodpovedajú po sebe nasledujúcim štádiám vývoja spermií. Po prvé, delenie zárodočných epiteliálnych buniek vedie k početným spermatogónie ktoré sa zväčšujú a stávajú saspermatocyty prvého rádu.V dôsledku prvého meiotického delenia tieto spermatocyty tvoria haploidspermatocyty druhého rádu,po ktorom prechádzajú druhým meiotickým delením a menia sa na spermatidy. Medzi „vláknami“ vyvíjajúcich sa buniek sú veľké Sertoliho bunky, príp trofické bunky,umiestnené v celom priestore od vonkajšej vrstvy tubulu po jeho lúmen.

Spermatocyty sa nachádzajú v početných invagináciách na bočných povrchoch Sertoliho buniek; tu sa zmenia na spermatidy a potom sa presunú k okraju Sertoliho bunky, ktorý je obrátený k lúmenu semenného tubulu, kde dozrievajú a tvoria spermín (obr. 20.33 ). Zdá sa, že Sertoliho bunky poskytujú mechanickú podporu, ochranu a výživu dozrievajúcim spermiám. Všetky živiny a kyslík dodávané do vyvíjajúcich sa gamét cez krvné cievy obklopujúce semenné tubuly a metabolický odpad uvoľnený do krvi prechádzajú cez Sertoliho bunky. Tieto bunky tiež vylučujú tekutinu, s ktorou spermie prechádzajú cez tubuly.
Vývoj vajíčok u ľudí (oogenéza).Na rozdiel od tvorby spermií, ktorá začína u mužov až v puberte, tvorba vajíčok u žien začína ešte pred ich narodením a je ukončená pre každé dané vajíčko až po jeho oplodnení. Štádiá oogenézy sú znázornené na obr. 20.36. Počas vývoja plodu sa primordiálne zárodočné bunky opakovane delia mitózou, pričom vzniká mnoho veľkých buniek tzv oogónia. Oogonia opäť prechádza mitózou a formuje saoocygy prvého rádu,ktoré zostávajú v profáznom štádiu takmer až do ovulácie. Oocyty prvého rádu sú obklopené jednou vrstvou buniek- granulózová membrána-a tvoria tzvprimordiálne folikuly.Ženský plod bezprostredne pred pôrodom obsahuje asi 2 10 6 tieto folikuly, ale len asi 450 z nich dosiahne štádium oocytov druhého rádu a opustí vaječník (ovuláciu). Pred ovuláciou sa oocyt prvého rádu podrobuje prvému meiotickému deleniu a vytvára haploidoocyt druhého rádu A prvé polárne teleso.Druhé meiotické delenie dosiahne štádium metafázy, ale nepokračuje, kým sa oocyt nespojí so spermiou. Počas oplodnenia sa oocyt druhého rádu podrobí druhému meiotickému deleniu, čím sa vytvorí veľká bunka - vajíčko a tiež druhé polárne teleso.Všetky polárne telá sú malé bunky; nehrajú žiadnu úlohu v oogenéze a nakoniec sú zničené.

partenogenéza
povinný
voliteľné
Vývoj organizmov len z neoplodneného vajíčka.
Tvoria sa len samice.
Napríklad skalník kaukazský
letlety sa môžu vyvíjať bez oplodnenia aj s oplodnením
Ženský typ
Samce sa vyvíjajú z neoplodnených vajíčok
Mužský typ
V niektorých izogamných riasach
Ďalšie podobné diela, ktoré by vás mohli zaujímať.vshm> |
|||
| 6644. | Individuálny vývoj organizmov (ontogenéza) | 78,69 kB | |
| Predpokladá sa, že jadrový obsah iba jednej spermie preniká do vajíčka zvierat. V prípade ľudí a niekedy vyšších zvierat sa obdobie vývoja pred narodením často nazýva prenatálne po narodení a postnatálne. U väčšiny mnohobunkových živočíchov, bez ohľadu na zložitosť ich organizácie, sú štádiá embryonálneho vývoja, ktorými embryo prechádza, rovnaké. Povaha drvenia a typy blastúl u rôznych stavovcov... | |||
| 13714. | Individuálny vývoj organizmov a ich správanie. Ontogenéza. Životný cyklus rastlín a živočíchov | 9,96 kB | |
| Ontogenéza vznik osobnosti je individuálny vývoj organizmu od okamihu vytvorenia zygoty až po jej smrť. Gamety sú pohlavné bunky, ktoré nesú dedičnú informáciu a majú haploidnú sadu. Hnojenie je proces splynutia samčích a samičích reprodukčných buniek rastlín alebo zvierat a je základom pohlavného procesu. Embryonálne zárodočné obdobie je obdobím ontogenézy od vytvorenia zygoty po pôrod alebo vzídenie z vaječných membrán alebo klíčenie. | |||
| 10427. | Reprodukcia | 6,75 kB | |
| S obrovskou rozmanitosťou foriem rozmnožovania organizmov sa dajú všetky zredukovať na dva hlavné typy: asexuálne a sexuálne. Pri nepohlavnom rozmnožovaní dochádza k rozmnožovaniu potomstva od jedného rodiča tvorbou spór alebo vegetatívne. Počas vegetatívneho rozmnožovania potomstvo vzniká z častí tela oddelených od matky. Počas vegetatívneho rozmnožovania si rastliny zachovávajú heterozygotnosť po mnoho generácií. | |||
| 21332. | Obmedzujúce faktory. Adaptácia organizmov na faktory | 303,8 kB | |
| Niektoré zvieratá milujú intenzívne teplo, iné lepšie znášajú mierne teploty prostredia atď. Okrem toho sa živé organizmy delia na tie, ktoré sú schopné existovať v širokom alebo úzkom rozsahu zmien akéhokoľvek faktora prostredia. Ak vplyv podmienok prostredia nedosahuje extrémne hodnoty, živé organizmy naň reagujú určitým konaním alebo zmenami svojho stavu, čo v konečnom dôsledku vedie k prežitiu druhu. Predmetom štúdia v tejto práci sú faktory prostredia, predmetom sú limitujúce faktory a adaptácia organizmov na... | |||
| 8875. | Všeobecné vzorce interakcie medzi organizmami a environmentálnymi faktormi | 193,58 kB | |
| Biotické faktory suchozemského a vodného prostredia pôd Biologicky aktívne látky živých organizmov Antropogénne faktory Všeobecné vzorce interakcie medzi organizmami a faktormi prostredia Pojem limitujúci faktor. Liebigov minimálny zákon Shelfordov zákon Špecifiká vplyvu antropogénnych faktorov na organizmus Klasifikácia organizmov vo vzťahu k faktorom prostredia 1. Podmienky perinových stepí predstavujú úplne odlišné režimy abiotických faktorov. | |||
| 12700. | Biologické vlastnosti škodcov a opatrenia na boj proti nim | 62,79 kB | |
| K výrazným stratám na úrode dochádza najmä v dôsledku prítomnosti burín, ktoré odstraňujú z pôdy živiny a vlahu, tienia kultúrne rastliny a v mnohých prípadoch kontaminujú produkty toxickými látkami a semenami, ktoré spôsobujú otravy ľudí a zvierat. Hlavné smery chemizácie poľnohospodárstva: používanie hnojív, chemikálií, ochrana rastlín pred škodcami, chorobami a burinou, používanie chemických prípravkov v chove hospodárskych zvierat, konzervovanie poľnohospodárskych produktov a... | |||
| 13403. | Vírusy ako nebunkové formy života. Štruktúra, klasifikácia, interakcia vírusu s bunkami rôznych organizmov | 12,75 kB | |
| Sú to biologické objekty, genómy zložené z nukleových kyselín DNA alebo RNA, produkované v živých bunkách pomocou ich biosyntetického aparátu. Rozdiely medzi vírusmi a inými formami života: nemajú bunkovú štruktúru; 1 typ nukleových kyselín; iba DNA alebo RNA; nemajú vlastný metabolizmus. Hypotéza pôvodu vírusov: vírusy vznikli z komponentov normálnej bunky, ktoré sa vymkli spod kontroly regulačných mechanizmov a zmenili sa na samostatnú jednotku, pravdepodobne došlo k sérii genetických zmien na úseku DNA, ktorý... | |||
| 18798. | Štrukturálne a funkčné ukazovatele organizmov pôdnej mezofauny v lesných a otvorených biotopoch údolia rieky Pozim | 61,54 kB | |
| Rozvoj tohto územia je však dnes dôsledkom viacerých environmentálnych problémov. Najmä v doline sa intenzívne pracuje na odvodnení lužných lúk, čo priamo ovplyvňuje prírodné biotopy. | |||
| 19386. | Vývoj liberalizmu v SNŠ | 35,55 kB | |
| Komplexný systémový charakter uvažovaných zmien v procesoch evolúcie zahraničnoekonomických vzťahov determinuje revíziu a zmenu celej axiomatiky tradičných predstáv o identifikácii, hraniciach a suverenite národného hospodárstva... | |||
| 8867. | VZDELÁVANIE A ROZVOJ | 162,32 kB | |
| Podstata problému vzťahu tréningu a rozvoja. Základné prístupy k riešeniu problému vzťahu tréningu a rozvoja. Koncept zóny proximálneho rozvoja L. Podstata problému vzťahu tréningu a rozvoja. | |||
Reprodukcia je vlastnosť organizmov produkovať potomstvo alebo schopnosť organizmov reprodukovať sa. Reprodukcia, ktorá je najdôležitejšou vlastnosťou živých vecí, zabezpečuje kontinuitu života a pokračovanie druhov.
Proces rozmnožovania je mimoriadne zložitý a súvisí nielen s prenosom genetickej informácie z rodičov na potomstvo, ale aj s anatomickými a fyziologickými vlastnosťami organizmov, s ich správaním a hormonálnym riadením. Reprodukcia organizmov je sprevádzaná procesmi ich rastu a vývoja.
Živé tvory sa vyznačujú extrémnou rozmanitosťou v spôsoboch rozmnožovania. Napriek tomu existujú dva hlavné spôsoby rozmnožovania – asexuálne a sexuálne (obr. 16). Nepohlavné rozmnožovanie alebo apomixis (z gréčtiny. aro - bez, mixis - miešanie) je proces, na ktorom sa zúčastňuje iba jeden rodič (bunka alebo mnohobunkový organizmus). Naproti tomu sexuálna reprodukcia zahŕňa dvoch rodičov, z ktorých každý má svoj vlastný reprodukčný systém a produkuje pohlavné bunky (gaméty), ktoré po splynutí vytvoria zygotu (oplodnené vajíčko), ktoré sa potom diferencuje na embryo. V dôsledku toho dochádza počas sexuálneho rozmnožovania k zmesi dedičných faktorov, t. j. procesu nazývanému amphimixis (z gréčtiny. amfi - na oboch stranách, mixis - miešanie).
Asexuálna reprodukcia
Nepohlavné rozmnožovanie je charakteristické pre organizmy mnohých druhov, rastlín aj živočíchov. Nachádza sa vo vírusoch, baktériách, riasach, hubách, cievnatých rastlinách, prvokoch, hubách, coelenterátoch, machorastoch a plášťovcoch.
Najjednoduchšia forma nepohlavného rozmnožovania je charakteristická pre vírusy. Ich reprodukčný proces je spojený s molekulami nukleových kyselín, so schopnosťou týchto molekúl sa samoduplikovať
a je založený na špecifickosti relatívne slabých vodíkových väzieb medzi nukleotidmi.
Ryža. 16.Metódy rozmnožovania organizmov
Vo vzťahu k iným organizmom, ktoré sa rozmnožujú nepohlavne, sa rozlišuje vegetatívne rozmnožovanie a rozmnožovanie sporuláciou.
Vegetatívne rozmnožovanie je rozmnožovanie, pri ktorom sa z časti oddelenej od materského organizmu vyvíja nový organizmus. Tento typ reprodukcie je charakteristický pre jednobunkové aj mnohobunkové organizmy, ale má v nich rôzne prejavy.
V jednobunkových organizmoch je vegetatívna reprodukcia reprezentovaná takými formami, ako je delenie, viacnásobné štiepenie a pučanie. Rozdelenie jednoduchým zovretím s vytvorením dvoch dcérskych organizmov z jedného rodičovského organizmu je charakteristické pre baktérie a modrozelené riasy (sinice). Naopak, rozmnožovanie delením hnedých a zelených rias, ako aj jednobunkových živočíchov (sarkódy, bičíkovce a nálevníky) prebieha mitotickým delením jadra s následným zovretím cytoplazmy.
Reprodukcia viacnásobným štiepením (schizogónia) zahŕňa rozdelenie jadra, po ktorom nasleduje rozdelenie cytoplazmy na časti. V dôsledku tohto delenia vzniká z jednej bunky niekoľko dcérskych organizmov. Príkladom viacnásobného delenia je rozmnožovanie falciparum plasmodium (P. vivax) v ľudských erytrocytoch. V tomto prípade v plazmódiách dochádza mnohokrát k opakovanému deleniu jadra bez cytokinézy, po ktorej nasleduje cytokinéza. Výsledkom je, že z jedného plazmódia vznikne 12-24 dcérskych organizmov.
V mnohobunkových rastlinných organizmoch sa vegetatívne rozmnožovanie delením uskutočňuje odrezkami, cibuľkami, listami a podzemkami. Ale to je v podstate umelé rozmnožovanie používané v poľnohospodárskej praxi. Rozmnožovanie vyšších rastlín v umelých podmienkach je možné aj z jednej bunky. Organizmy, ktoré sa vyvinú z jednej bunky (klonu), majú všetky vlastnosti pôvodného mnohobunkového organizmu. Toto rozmnožovanie sa nazýva klonálna mikropropagácia. Jednou z foriem vegetatívneho rozmnožovania môže byť štepenie alebo presádzanie mnohých kultúrnych rastlín, ktoré spočíva v presadení púčika alebo časti výhonku z jednej rastliny na druhú. Samozrejme, ide aj o spôsob rozmnožovania, ktorý sa síce v prírode nevyskytuje, ale v poľnohospodárstve sa využíva veľmi široko.
U mnohobunkových zvierat dochádza k vegetatívnej reprodukcii fragmentáciou ich tela na časti, po ktorých každá
časť sa vyvinie v nové zviera. Takáto reprodukcia je typická pre huby, coelenteráty (hydry), nemerteany, ploché červy, ostnatokožce (hviezdice) a niektoré ďalšie organizmy. Blízkou formou fragmentácie k vegetatívnej reprodukcii zvierat je zvieracia polyembryónia, ktorá spočíva v tom, že embryo je v určitom štádiu vývoja rozdelené na niekoľko častí, z ktorých každá sa vyvinie do samostatného organizmu. Polyembryónia sa vyskytuje u pásavcov. Tí druhí sa však rozmnožujú sexuálne. Preto je polyembryónia skôr jedinečným štádiom sexuálneho rozmnožovania a potomstvo, ktoré je výsledkom polyembryónie, predstavujú jednovaječné dvojčatá.
Pučanie spočíva vo vytvorení tuberkulózy (výrastku) s jadrom na materskej bunke, ktorá sa potom oddelí a stane sa samostatným organizmom. Pučanie sa vyskytuje tak u jednobunkových rastlín, napríklad u kvasiniek, ako aj u jednobunkových živočíchov, napríklad u niektorých druhov nálevníkov.
Rozmnožovanie sporuláciou je spojené s tvorbou špecializovaných buniek - spór, ktoré obsahujú jadro, cytoplazmu, sú pokryté hustou membránou a sú schopné dlhodobej existencie v nepriaznivých podmienkach, čo navyše prispieva k ich šíreniu. Najčastejšie sa takáto reprodukcia vyskytuje u baktérií, rias, húb, machov a papradí. V niektorých zelených riasach sa z jednotlivých buniek môžu vytvárať takzvané zoospóry.
Medzi živočíchmi sa rozmnožovanie sporuláciou pozoruje u sporozoánov, najmä u falciparum plasmodium.
V organizmoch mnohých druhov sa nepohlavné rozmnožovanie môže striedať so pohlavným rozmnožovaním.
SEXUÁLNA REPRODUKCIA
K pohlavnému rozmnožovaniu dochádza u jednobunkových aj mnohobunkových rastlín a živočíchov.
Ako je uvedené v kapitolách V a XIII, sexuálna reprodukcia v baktériách sa uskutočňuje konjugáciou, ktorá slúži ako analóg pohlavného procesu a je systémom rekombinácie týchto organizmov, zatiaľ čo u prvokov dochádza k sexuálnej reprodukcii aj konjugáciou alebo syngamiou. a autogamie.
V mnohobunkových organizmoch (rastliny a zvieratá) je pohlavné rozmnožovanie spojené s tvorbou zárodočných alebo pohlavných buniek (gamét), oplodnením a tvorbou zygot.
Pohlavné rozmnožovanie je významnou evolučnou akvizíciou organizmov. Na druhej strane prispieva k preskupovaniu génov, vzniku diverzity organizmov a zvyšovaniu ich konkurencieschopnosti v neustále sa meniacich podmienkach prostredia.
V jednobunkových organizmoch sa pohlavné rozmnožovanie vyskytuje v niekoľkých formách. V baktériách môže byť sexuálna reprodukcia analogická s konjugáciou, ktorá sa v nich vyskytuje, ktorá spočíva v prenose plazmidu alebo chromozomálnej DNA z donorových buniek (obsahujúcich plazmidy) do buniek príjemcu (neobsahujúcich plazmidy), ako aj s transdukciou baktérií. , ktorá spočíva v prenose genetického materiálu z niektorých bakteriálnych buniek do iných fágov. Konjugácia sa nachádza aj u nálevníkov, v ktorých počas tohto procesu dochádza k prenosu jadier z jedného jedinca na druhého, po ktorom nasleduje rozdelenie druhého.
V mnohobunkových rastlinách a živočíchoch dochádza k sexuálnemu rozmnožovaniu buniek prostredníctvom tvorby samičích a samčích zárodočných buniek (vajíčok a spermií), následného oplodnenia vajíčka spermiou a tvorby zygoty. V rastlinách sa pohlavné bunky produkujú v špecializovaných reprodukčných štruktúrach; u zvierat sú produkované v pohlavných žľazách nazývaných gonády (z gréčtiny. preč - semeno).
Medzi somatickými a reprodukčnými bunkami zvierat je dôležitý rozdiel. Spočíva v tom, že somatické bunky sú schopné delenia, teda samy sa rozmnožujú a navyše z nich vznikajú zárodočné bunky. Naopak, pohlavné bunky sa nedelia, ale „začínajú“ reprodukciu celého organizmu.
Diploidné somatické bunky, v ktorých sa tvoria mužské zárodočné bunky, sa nazývajú spermatogónie a v ktorých sa tvoria ženské zárodočné bunky - oogónia. Proces tvorby (rastu a diferenciácie) mužských a ženských zárodočných buniek sa nazýva gametogenéza.
Gametogenéza je založená na meióze (z gréčtiny. meióza - redukovať), čo je redukčné delenie bunkových jadier sprevádzané poklesom počtu chromozómov na jadro. Meióza sa vyskytuje v špecializovaných bunkách reprodukčných orgánov živých tvorov, ktoré sa rozmnožujú sexuálne (obr. 17). Napríklad v pteridofytoch sa meióza vyskytuje v špecializovaných sporangiálnych bunkách umiestnených na spodnom povrchu listov týchto rastlín a vyvíja sa do spór a potom do gametofytov. Posledne menované existujú oddelene a v konečnom dôsledku produkujú samčie a samičie gaméty. V kvitnúcich rastlinách sa meióza vyskytuje v špecializovaných bunkách vajíčok, z ktorých sa vyvinú spóry. Posledne menované produkujú gametofyt s jedným vajcom. Okrem toho sa v týchto rastlinách meióza vyskytuje aj v špecializovaných bunkách prašníkov, z ktorých sa tiež vyvinú spóry, ktoré nakoniec produkujú peľ s dvoma samčími gamétami. U dážďoviek, ktoré sú hermafroditmi a obsahujú samčie reprodukčné orgány v jednom segmente tela a samičie orgány v druhom a ktoré sa vyznačujú
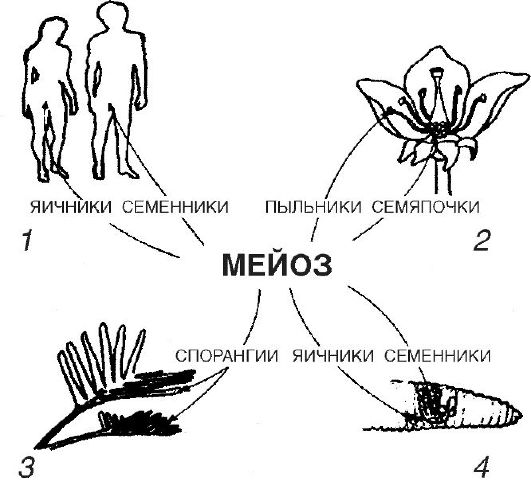
Ryža. 17. Meióza v rôznych organizmoch: 1 - človek; 2 - kvitnúce rastliny; 3 - papraď; 4 - dážďovka
Majú schopnosť krížového oplodnenia medzi rôznymi jedincami, majú schopnosť súčasne vykonávať spermatogenézu a oogenézu.
Meióza sa vyskytuje v špecializovaných bunkách semenníkov a vaječníkov, ktoré produkujú mužské a ženské gaméty. Boli identifikované proteíny, ktoré sú induktormi meiózy.
Počas procesu meiózy sa diploidný počet chromozómov (2n), ktorý je charakteristický pre somatické bunky (bunkové jadrá) a nezrelé bunkové zárodky, mení na haploidný počet (n), charakteristický pre zrelé zárodočné bunky. Zárodočné bunky teda v dôsledku gametogenézy dostávajú len polovicu chromozómov somatických buniek (obr. 18).

Ryža. 18.
Správanie chromozómov počas gametogenézy u zvierat je rovnaké u samcov aj samíc. Pohlavia sa však líšia v čase vzniku jednotlivých štádií meiózy, čo je najmä
obzvlášť viditeľné u ľudí. U postpubertálnych mužov je úplný proces meiózy ukončený približne za dva mesiace, zatiaľ čo u žien sa prvé meiotické delenie začína vo fetálnom vaječníku a je ukončené až po začatí ovulácie, ktorá nastáva približne vo veku 15 rokov.
U vyšších živočíchov je v prípade samcov meióza sprevádzaná tvorbou štyroch funkčne aktívnych gamét.
Naopak, u žien produkuje každý oocyt druhého rádu iba jedno vajíčko. Ďalšími jadrovými produktmi ženskej meiózy sú tri redukčné telieska, ktoré sa nezúčastňujú reprodukcie a degenerujú.
Meióza pozostáva z dvoch jadrových divízií. Prvé meiotické jadrové delenie oddeľuje členov každého páru homológnych chromozómov potom, čo sa navzájom spárovali (synapsia) a vymenili si genetický materiál (crossing). V dôsledku tohto delenia vznikajú dve haploidné jadrá. Druhé meiotické delenie oddeľuje dve pozdĺžne polovice chromozómov (chromatidy) v každom z týchto jadier, čím vznikajú štyri haploidné jadrá.
V procese gametogenézy dochádza aj k diferenciácii vajíčok (ovogenéza) a spermií (spermatogenéza), čo je predpokladom ich funkcií. Živočíšne vajíčka sú oveľa väčšie ako spermie, sú zvyčajne nepohyblivé a obsahujú nutričný materiál, ktorý zabezpečuje vývoj embrya v počiatočnom období po oplodnení. Spermie väčšiny zvierat majú bičík, ktorý zabezpečuje ich nezávislosť v pohybe.
Meióza má vynikajúci biologický význam. Vďaka meióze sa v bunkách organizmov udržiava konštantný počet chromozómov bez ohľadu na počet generácií. Meióza preto zachováva stálosť druhov. Nakoniec v meióze v dôsledku kríženia dochádza k rekombinácii génov, čo je jeden z faktorov evolúcie.
SPERMATOGENÉZA A OVOGENÉZA
Spermatogenéza je proces tvorby zrelých mužských zárodočných buniek. Spermie sa vyvíjajú v mužských pohlavných žľazách (semenníky alebo semenníky) zo špecializovaných somatických buniek (obr. 19). Takéto špecializované bunky slúžia takto:
nazývané primordiálne zárodočné bunky, ktoré migrujú do semenníkov v ranom období embryogenézy mužského jedinca. V dôsledku toho sú primordiálne bunky progenitormi (prekurzormi) zrelých zárodočných buniek.
Ľudské semenníky pozostávajú z mnohých tubulov, ktorých steny sú tvorené vrstvami buniek, ktoré sú v rôznych štádiách vývoja spermií. Vonkajšia vrstva tubulov je tvorená veľkými bunkami nazývanými spermatogónia. Tieto bunky obsahujú diploidnú sadu chromozómov a sú potomkami primordiálnych zárodočných buniek v semenníkoch. V období puberty jedinca sa časť spermatogónie presúva do vnútornej vrstvy tubulov, kde sa v dôsledku meiózy vyvinú na bunky nazývané spermatocyty prvého rádu (spermatocyty I), potom na spermatocyty druhého rádu. (spermatocyty II) a nakoniec

Ryža. 19.Spermatogenéza a oogenéza
na spermatidy, čo sú haploidné zárodočné bunky, ktoré sa nakoniec diferencujú na zrelé spermie. Vo všeobecnosti teda môžeme povedať, že spermatogenéza je iniciovaná v diploidných somatických bunkách (spermatogónia), po ktorej nasleduje obdobie dozrievania zárodočných buniek, v ktorom dochádza k dvom deleniu jadra prostredníctvom meiózy, čo vedie k tvorbe spermatíd. Toto je obraz osoby.
Meióza (obr. 20) v spermatogenéze prebieha v niekoľkých štádiách (fázach). Medzi deleniami sú dve medzifázy. Meiotické delenie teda možno znázorniť ako sériu dejov nasledujúcich za sebou, a to: interfáza I - prvé meiotické delenie (skorá profáza I, neskorá profáza I, metafáza I, anafáza I, telofáza I) - interfáza II (interokinéza) - druhé meiotické delenie (profáza I, metafáza II, anafáza II, telofáza II). Proces meiózy je veľmi dynamický, takže mikroskopické rozdiely medzi rôznymi štádiami neodrážajú povahu samotných štádií, ale skôr vlastnosti chromozómov v rôznych štádiách. Medzifáza I

Ryža. 20.Fázy meiózy: 1 - prvé meiotické delenie; 2 - druhé meiotické delenie
charakterizovaný tým, že v ňom prebieha replikácia chromozómov (zdvojenie DNA), ktorá je takmer úplne ukončená začiatkom ranej profázy I.
Prvé meiotické delenie začína v primárnom spermatocyte a je charakterizované dlhou profázou, ktorá pozostáva z prechodu profázy I a profázy II do seba. V profáze I je päť hlavných štádií – leptonéma, zygonéma, pachynéma, diplonéma a diakinéza.
V štádiu leptonómu sú chromozómy v jadre prezentované vo forme tenkých špirálových vlákien obsahujúcich početné tmavo sfarbené granuly (chromoméry). Rozdelenie chromomérov a vlákien nie je zaznamenané, ale predpokladá sa, že chromozómy v tomto štádiu sú dvojité, t.j. diploidné. Homológy každého chromozómového páru sú spojené chromomérmi pozdĺž ich dĺžky podľa princípu spony.
Štádium zygonému je charakterizované vytvorením synapsií medzi homológnymi chromozómami, čo vedie k tvorbe párových chromozómov (bivalentov). Chromozómy X a Y sa v porovnaní s autozómami správajú trochu inak. Kondenzujú do tmavo sfarbených heterochromatických teliesok, ktoré sú spárované v dôsledku homológnych oblastí na ich koncoch.
V štádiu pachynémy, čo je najdlhšie štádium meiotickej profázy, dochádza ku kondenzácii bivalentov a k rozdeleniu každej chromatídy na dve, v dôsledku čoho je každá bivalentná zložitá špirálovitá štruktúra pozostávajúca zo štyroch sesterských chromatíd (tetrád). Na konci tohto štádia začína separácia párových bivalentných chromozómov. Teraz je možné pozorovať homológne chromozómy vedľa seba. Preto v niektorých prípravkoch možno vidieť štyri chromozómy, ktoré sa tvoria ako výsledok duplikácie každého homológu, čím sa vytvárajú sesterské chromatidy. V tomto štádiu dochádza k výmenám medzi homológmi a vytváraniu chiazmat.
V štádiu diplonémy dochádza ku skracovaniu, zhrubnutiu a vzájomnému odpudzovaniu sesterských chromatidov, v dôsledku čoho sú chromatidy v bivalente takmer oddelené. Separácia sa považuje za neúplnú, pretože centroméra ešte nebola rozdelená v každom páre chromozómov. Pokiaľ ide o bivalenty, na rôznych miestach pozdĺž ich dĺžky sú držané chiazmatami, čo sú štruktúry vytvorené medzi homológnymi chromatidami.
predchádzajúci prechod medzi synapticky spojenými homológmi. V dobrých prípravkoch možno pozorovať jednu až niekoľko chiazmat v závislosti od dĺžky bivalentu. Každá chiazma pozorovaná v tomto štádiu predstavuje výsledok výmeny, ku ktorej došlo medzi nesesterskými chromatidami počas štádia pachynémy. Keď sa stláčanie a odpudzovanie bivalentov zintenzívňuje, chiazmata sa posúvajú ku koncom chromozómov, t.j. dochádza k chromozómovej terminácii. Na konci diplonému dochádza k despiralizácii chromozómov; homológy sa naďalej navzájom odpudzujú.
V štádiu diakinézy, ktorá je podobná diplonéme, pokračuje skracovanie bivalentov a slabnú (ubúdajú) chiazmata, v dôsledku čoho vznikajú diskrétne jednotky vo forme chromatíd (štyri). Bezprostredne po dokončení tejto fázy nastáva rozpustenie jadrovej membrány.
V metafáze I dosahujú bivalenty svoju najvyššiu koncentráciu. Stávajú sa oválne a nachádzajú sa v rovníkovej časti jadra, kde tvoria rovníkové platne meiotickej metafázy I. Tvar každého bivalentu je určený počtom a umiestnením chiazmat. U mužov je počet chiazmat na bivalent v metafáze I zvyčajne 1-5. Bivalent XY sa stáva tyčinkovitým v dôsledku jedinej koncovej chiazmy.
V anafáze I začína pohyb opačných centromér k opačným pólom bunky. Výsledkom je oddelenie homológnych chromozómov. Každý chromozóm teraz pozostáva z dvoch chromatidov držaných pohromade centromérou, ktorá sa nedelí a zostáva nedotknutá. To odlišuje anafázu I meiózy od anafázy mitózy, v ktorej sa centroméra delí. Je dôležité poznamenať, že crossover robí každú chromatídu geneticky odlišnou.
V štádiu telofázy I sa chromozómy dostanú k pólom, čím sa ukončí prvé meiotické delenie. Po telofáze I nastáva krátka interfáza (interkinéza), v ktorej chromozómy despirujú a stávajú sa difúznymi, alebo telofáza I prechádza priamo do profázy II druhého meiotického delenia. Ani v jednom prípade nie je pozorovaná replikácia DNA. Po prvom meiotickom delení sa bunky nazývajú spermatocyty druhého rádu. Počet chromozómov v každej takejto bunke klesá z 2n na 1n, ale obsah DNA sa ešte nemení.
Druhé meiotické delenie prebieha v niekoľkých fázach (profáza II, metafáza II, anafáza II, telofáza II) a je podobné mitotickému deleniu. V profáze II zostávajú chromozómy sekundárnych spermatocytov na póloch. V metafáze II sa centroméra každého z dvojitých chromozómov delí a poskytuje každému novému chromozómu vlastnú centroméru. Začína sa tvorba vretienka, k pólu ktorého sa pohybujú nové chromozómy. V telofáze II končí druhé meiotické delenie, v dôsledku čoho každý spermatocyt druhého rádu produkuje dve spermatídy, z ktorých sa potom diferencujú spermie. Rovnako ako v sekundárnom spermatocyte je počet chromozómov v spermatíde haploidný (1n). Chromozómy spermatíd sú však jednoduché, zatiaľ čo chromozómy sekundárnych spermatocytov II sú dvojité a sú postavené z dvoch chromatidov. V dôsledku toho má jadro každej spermatídy jednu sadu nehomologických chromozómov. Sekundárne meiotické delenie je delenie mitotického typu (ekvatoriálne delenie). Oddeľuje dvojité sesterské chromatidy a líši sa od redukčného delenia, pri ktorom sa oddeľujú homológne chromozómy. Jediný významný rozdiel od klasickej mitózy je v tom, že existuje haploidná sada chromozómov.
Takže prvé meiotické delenie spermatocytov prvého rádu vedie k vytvoreniu dvoch sekundárnych spermatocytov (druhého rádu). Obidve chromatidy štruktúr vytvorených v dôsledku redukčného delenia sú sesterské chromatidy. Posledne menované vznikajú ako výsledok replikácie predchádzajúcej prvému meiotickému deleniu. Druhé meiotické delenie každého sekundárneho spermatocytu vedie k vytvoreniu štyroch spermatíd. Pri typickej meióze sa teda bunky delia dvakrát, zatiaľ čo chromozómy sa delia iba raz (obr. 21).
Konečné štádium spermatogenézy je spojené s diferenciáciou, ktorá končí tým, že sa každá z relatívne veľkých, sféricky nepohyblivých spermatíd zmení na malú predĺženú pohyblivú spermiu.
U väčšiny dospelých (pohlavne zrelých) samcov sa spermatogenéza vyskytuje v semenníkoch neustále alebo periodicky (sezónne). Napríklad u hmyzu trvá dokončenie cyklu spermatogenézy len niekoľko dní, zatiaľ čo u cicavcov tento cyklus trvá týždne alebo dokonca

Ryža. 21.Distribúcia chromozómov počas gametogenézy
mesiacov. U dospelých sa spermatogenéza vyskytuje počas celého roka. Čas vývoja primitívnej spermatogónie na zrelé spermie je asi 74 dní.
Samčie reprodukčné bunky, produkované organizmami rôznych druhov, sa vyznačujú pohyblivosťou a extrémnou diverzitou vo veľkosti a štruktúre (obr. 22). Každá ľudská spermia pozostáva z troch častí – hlavy, strednej časti a chvosta (obr. 23). Hlava spermie obsahuje jadro, ktoré obsahuje haploidnú sadu chromozómov.
Hlavička je vybavená akrozómom, ktorý obsahuje lytické enzýmy potrebné na to, aby sa obsah spermií dostal do vajíčka. V hlave sú lokalizované aj dve centrioly - proc-

Ryža. 22. Tvary spermií

Ryža. 23. Štruktúra spermií: A - svetelný mikroskopický obraz v rôznych rovinách: 1 - hlava, 2 - stredná časť, 3 - chvost; B - schematická rekonštrukcia obrazu z elektrónového mikroskopu: 1 - jadro, 2 - akrozóm, 3 - centrozóm (proximálny centriol), 4 - centrálny kruh, 5 - mitochondriálna špirála,
6 - axiálny závit
Simal, ktorý stimuluje delenie vajíčka oplodneného spermiou, a distálny, ktorý dáva vznik axiálnej drieku chvosta. Stredná časť spermií obsahuje bazálne telo chvosta a mitochondrie. Chvost (proces) spermie je tvorený vnútornou aximálnou tyčinkou a vonkajším plášťom, ktorý je cytoplazmatického pôvodu. Ľudské spermie sa vyznačujú výraznou pohyblivosťou.
OVOGENÉZA A OGGLE BUNKY
Proces tvorby vajíčok sa nazýva oogenéza. Jeho funkciami je zabezpečiť haploidnú sadu chromozómov v jadre vajíčka a nutričné potreby zygoty. Oogenéza je vo svojom prejave v podstate porovnateľná so spermatogenézou.
U cicavcov a ľudí sa oogenéza začína v prenatálnom období (pred narodením). Oogónia, čo sú malé bunky s dosť veľkým jadrom a lokalizované vo ovariálnych folikuloch, sa začínajú diferencovať na primárne oocyty. Tie sa tvoria už v treťom mesiaci vnútromaternicového vývoja, po ktorom vstupujú do profázy prvého meiotického delenia. Keď sa dievča narodí, všetky primárne oocyty sú už v profáze prvého meiotického delenia. Primárne oocyty zostávajú v profáze až do nástupu puberty u ženského jedinca. Jeden oocyt obsahuje až 100 000 mitochondrií. Keď ovariálne folikuly dozrievajú na začiatku puberty, obnoví sa meiotická profáza v primárnych oocytoch. Prvé meiotické delenie pre každé vyvíjajúce sa vajíčko sa dokončí krátko pred časom ovulácie daného vajíčka. V dôsledku prvého meiotického delenia a nerovnomerného rozdelenia cytoplazmy sa jedna výsledná bunka stáva sekundárnym oocytom, druhá - polárnym (redukčným) telom.
Sekundárne meiotické delenie nastáva, keď sekundárny oocyt (vyvíjajúce sa vajíčko) prechádza z vaječníka do vajíčkovodu. Toto delenie sa však nedokončí, kým spermie neprenikne do sekundárneho oocytu, ktorý sa zvyčajne vyskytuje vo vajíčkovode. Keď spermie prenikne do sekundárneho oocytu, tento sa rozdelí, čo vedie k vytvoreniu ovote (zrelé vajíčko) s pronukleom obsahujúcim jednu sadu
z 23 materských chromozómov. Ďalšia bunka, ktorá je výsledkom tohto delenia, je druhé polárne teleso, ktoré nie je schopné ďalšieho vývoja. V tomto čase dochádza aj k deleniu polárneho (redukčného) telesa na dve časti. Vývoj jedného oocytu prvého rádu je teda sprevádzaný tvorbou jedného ovotidu a troch redukčných teliesok. Vo vaječníkoch takto dozrieva zvyčajne 300 – 400 oocytov počas celého života, no za mesiac dozrieva len jeden oocyt. Pri diferenciácii vajíčok sa vytvárajú blany a ich jadro sa zmenšuje.
U niektorých živočíšnych druhov prebieha oogenéza rýchlo a nepretržite a vedie k produkcii veľkého počtu vajíčok.
Napriek podobnosti so spermatogenézou sa oogenéza vyznačuje niektorými špecifickými znakmi. Výživový materiál (žĺtok) primárneho oocytu nie je rovnomerne rozdelený medzi štyri bunky, ktoré sa tvoria ako výsledok meiotického delenia. Hlavné množstvo žĺtka je uložené v jednej veľkej bunke, zatiaľ čo polárne telieska obsahujú veľmi málo tejto látky. V dôsledku delenia prvé a druhé polárne telieska dostávajú rovnaké sady chromozómov ako sekundárne oocyty, ale nestávajú sa zárodočnými bunkami. Preto sú vajíčka oveľa bohatšie na nutričný materiál v porovnaní so spermiami. Tento rozdiel je obzvlášť výrazný v prípade zvierat, ktoré znášajú vajcia.
Vajíčka cicavcov majú oválny alebo trochu pretiahnutý tvar (obr. 24) a vyznačujú sa typickými znakmi bunkovej stavby. Obsahujú všetky štruktúry charakteristické pre somatické bunky, avšak vnútrobunková organizácia vajíčka je veľmi špecifická a je daná tým, že vajíčko je zároveň prostredím, ktoré zabezpečuje vývoj zygoty. Jednou z charakteristických vlastností vajec je zložitosť štruktúry ich membrán. U mnohých zvierat sa rozlišujú primárne, sekundárne a terciárne membrány vajíčok. Primárna škrupina (vnútorná) sa tvorí v štádiu oocytov. Predstavuje povrchovú vrstvu oocytu a má zložitú štruktúru, pretože je preniknutá výrastkami susediacich folikulárnych buniek. Sekundárna (stredná) škrupina je úplne tvorená folikulárnymi bunkami a terciárna (vonkajšia) škrupina je tvorená látkami, ktoré sú sekrečnými produktmi žliaz vajcovodov, ktorými prechádzajú vajíčka. U vtákov napr.
Terciárne membrány vajec sú bielka, podškrupina a škrupina. Vajcia cicavcov sa vyznačujú prítomnosťou dvoch membrán. Štruktúra vnútrobunkových zložiek vajíčok je druhovo špecifická a niekedy má dokonca individuálne vlastnosti.

Ryža. 24. Štruktúra vajíčka: 1 - folikulárne bunky; 2 - škrupina; 3 - cytoplazma; 4- jadro
HNOJENIE
Hnojenie je proces spájania mužských a ženských gamét, ktorý vedie k vytvoreniu zygoty a následnému vývoju nového organizmu. Počas procesu oplodnenia sa v zygote vytvorí diploidný súbor chromozómov, ktorý určuje výnimočný biologický význam tohto procesu.
V závislosti od druhu organizmov, ktoré sa rozmnožujú sexuálne, sa rozlišuje vonkajšie a vnútorné oplodnenie. K vonkajšiemu oplodneniu dochádza v prostredí, do ktorého vstupujú samčie a samičie reprodukčné bunky. Napríklad oplodnenie u rýb je vonkajšie. Nimi uvoľnené samčie (mlieko) a samičie (kaviár) reprodukčné bunky vstupujú do vody, kde sa stretávajú a spájajú.
Vnútorné oplodnenie je zabezpečené prenosom spermií z mužského tela do ženského tela. K takémuto oplodneniu dochádza u cicavcov, ktorých ústredným bodom je splynutie jadier zárodočných buniek. Predpokladá sa, že obsah jednej spermie preniká do vajíčka. V mechanizmoch oplodnenia
dokončenia, stále zostáva veľa nejasností. Údaje o oplodnení morských ježkov naznačujú, že už 2 sekundy po kontakte spermie a vajíčka nastanú zmeny v elektrických vlastnostiach plazmatickej membrány týchto vajíčok. K fúzii gamét dochádza do 7 sekúnd. Predpokladá sa, že prienik obsahu len jednej z mnohých spermií do vajíčka sa vysvetľuje zmenami elektrických vlastností jej plazmatickej membrány. Z oplodneného vajíčka vzniká zygota. Existujú dva názory na dôvody aktivácie metabolizmu vajíčok spermiami. Niektorí veria, že väzba spermií na vonkajšie receptory na povrchu buniek je signálom, ktorý cez membránu vstupuje do vajíčka a aktivuje tam inozitoltrifosfát a ióny vápnika. Iní veria, že spermie obsahujú špeciálny iniciačný faktor.
Experimentálny vývoj uskutočnený v posledných rokoch ukázal, že oplodnenie vajíčok cicavcov, vrátane ľudí, je možné in vitro. Okrem toho embryá vyvinuté in vitro môžu byť implantované do maternice ženy, kde prechádzajú ďalším normálnym vývojom. Existuje mnoho prípadov narodenia detí zo „skúmavky“.
Na rozdiel od zvierat, ktoré sa rozmnožujú zygogenézou, mnohé organizmy sú schopné rozmnožovania partenogenézou (z gr. parthenos - panna a genos - pôrod), ktorý označuje rozmnožovanie organizmov z neoplodneného vajíčka. Existuje obligátna a fakultatívna partenogenéza. Obligátna partenogenéza sa stala hlavnou metódou reprodukcie organizmov určitých druhov, napríklad jašterice kaukazskej. Zvieratá tohto druhu sú len samice. Naopak, fakultatívna partenogenéza znamená, že vajíčka sú schopné vyvíjať sa bez oplodnenia aj po oplodnení. Fakultatívna partenogenéza je zasa ženská a mužská. Samičia partenogenéza sa často pozoruje u včiel, mravcov a vírnikov, pri ktorých sa samce vyvíjajú z neoplodnených vajíčok. Samčia partenogenéza sa vyskytuje u niektorých izogamných rias.
Partenogenéza môže byť prirodzená aj umelá (indukovaná). Mechanizmus umelej partenogenézy je dráždenie vajíčok pomocou fyzikálnych alebo chemických
faktorov, čo vedie k aktivácii vajíčok a v dôsledku toho k vývoju neoplodnených vajíčok. Umelá partenogenéza bola pozorovaná u zvierat mnohých systematických skupín - ostnatokožcov, červov, mäkkýšov a dokonca aj cicavcov.
Známa forma partenogenézy sa nazýva androgenéza (z gréčtiny. andros - muž, genéza - pôvod). Ak je jadro vo vajíčku inaktivované a následne doň prenikne niekoľko spermií, potom sa z takého vajíčka v dôsledku splynutia jadier samcov (spermií) vyvinie mužský organizmus.
Existujú prípady, kedy partenogenéza prebieha cyklicky, v závislosti od ročného obdobia. Napríklad vírniky, dafnie a vošky sa rozmnožujú v lete partenogenézou a na jeseň oplodnením vajíčok a tvorbou zygot, t.j. zygogenézou.
Úloha partenogenézy a jej foriem v prírode je malá, pretože neposkytuje široké adaptačné schopnosti organizmov.
Na rozdiel od zygogenézy a partenogenézy existuje gynogenéza, čo je pseudogamia, keď spermia vstúpi do vajíčka a aktivuje ho, ale jadro spermie sa nespojí s jadrom vajíčka. V tomto prípade výsledné potomstvo pozostáva iba zo samíc. Gynogenéza sa prirodzene vyskytuje u háďatiek a rýb, ale môže byť vyvolaná aj umelo. Boli opísané najmä prípady umelej gynogenézy u priadky morušovej, rýb a obojživelníkov.
STRIEDAJÚCI SA HAPLOIDNÝ A DIPLOIDNÝ.
STRIEDAJÚCE SA GENERÁCIE
Pre organizmy, ktoré sa rozmnožujú sexuálne, je charakteristické striedanie haploidných a diploidných fáz vo svojom vývoji. U mnohých organizmov, vrátane cicavcov, je toto striedanie pravidelné a je na ňom založené zachovanie druhovej charakteristiky organizmov.
Pre mnohé organizmy je charakteristická aj výmena generácií, kedy generácie jedincov rozmnožujúcich sa nepohlavne nahrádzajú generácie jedincov rozmnožujúcich sa pohlavne s tvorbou gamét. Tomu sa hovorí primárna výmena generácií. Nachádza sa v sporozoánoch, bičíkovcoch a mnohých rastlinách. Primárna výmena generácií je pravidelná a je prítomná
To, čo sa nachádza v prírode, naznačuje zachovanie vo fylogenéze mnohých organizmov nepohlavného aj sexuálneho rozmnožovania. U iných organizmov dochádza k striedaniu pohlavného rozmnožovania s partenogenézou. Hovorí sa tomu sekundárna generačná výmena. Napríklad u trematód je sexuálna reprodukcia pravidelne nahradená partenogenézou. Toto sekundárne striedanie generácií sa nazýva heterogónia. U koelenterátov v niektorých štádiách vývoja dochádza k prechodu zo sexuálneho rozmnožovania na nepohlavné (vegetatívne). Táto forma sekundárneho striedania generácií sa nazýva metagenéza.
SEXUÁLNY DIMORFIZMUS. HERMAFRODITIZMUS
Muži a ženy sa vyznačujú špecifickými fenotypovými znakmi. Rozdiely medzi ženami a mužmi v ich vlastnostiach sa nazývajú sexuálny dimorfizmus. U živočíchov sa vyskytuje už v nižších štádiách evolučného vývoja, napríklad u okrúhlych helmintov a článkonožcov, a najväčší prejav dosahuje u stavovcov.
Ak mužské a ženské reprodukčné bunky produkuje ten istý jedinec, ktorý má mužské aj ženské pohlavné žľazy, potom sa tento jav nazýva skutočný hermafroditizmus. Nachádza sa v plochých červoch, annelidoch a mäkkýšoch. U plochých červov fungujú samčie a samičie pohlavné žľazy počas celého života jedinca. Naproti tomu u mäkkýšov gonády produkujú vajíčka a spermie striedavo.
Skutočný hermafroditizmus sa vyskytuje aj u ľudí, ktorý je výsledkom vývojových porúch. Genotypy hermafroditov sú 46 XX alebo 46 XY, pričom väčšina prípadov je XX (asi 60 %). Genotyp XX sa najčastejšie vyskytuje u hermafroditov černošských afrických populácií, zatiaľ čo XY sa najčastejšie vyskytuje u Japoncov. U hermafroditov oboch typov bola zaznamenaná tendencia k bilaterálnej gonadálnej asymetrii. Medzi skutočnými hermafroditmi existujú aj chromozomálne mozaiky, ktoré v niektorých somatických bunkách majú pár chromozómov XX, v iných - pár XY.
Známy je aj falošný hermafroditizmus, kedy jedinci majú vonkajšie pohlavné orgány a sekundárne pohlavné znaky charakteristické pre obe pohlavia, no produkujú len jeden typ zárodočných buniek – mužské alebo ženské.
PÔVOD REPRODUKČNÝCH METÓD
Predpokladá sa, že najstaršia je asexuálna reprodukcia, najmä vegetatívna reprodukcia. Z toho posledného sa vyvinulo rozmnožovanie tvorbou spór, ktorej nepochybnou výhodou je, že poskytuje lepšie možnosti na zachovanie druhov a najmä na ich šírenie.
Sexuálne rozmnožovanie je najefektívnejším spôsobom rozmnožovania organizmov. Predpokladá sa, že sa vyvinul z asexuálneho rodu, ktorý sa objavil asi pred 1 miliardou rokov, a prvé štádiá boli spojené s komplikáciou vo vývoji gamét. Primitívne gaméty sa vyznačovali nedostatočnou morfologickou diferenciáciou, v dôsledku ktorej vznikla izogamia (z gr. isos- rovný, gamos - manželstvo), keď pohlavné bunky boli pohyblivé izogaméty, nerozlíšené na mužské a ženské formy.
Následne sa vyvinula anizogamia (z gréčtiny. anisos- nerovný, gamos - manželstvo), charakterizované prítomnosťou diferencovaných gamét, ktoré sa líšia iba veľkosťou. V neskorších štádiách evolúcie vznikli ostré rozdiely v pohyblivosti, tvare a veľkosti gamét. V procese evolúcie stavovce vyvinuli aj množstvo prídavných zariadení, ktoré uľahčujú prenos mužských spermií do ženského reprodukčného traktu a vytvárajú podmienky pre vývoj oplodneného vajíčka. Počas evolúcie sa tieto zariadenia vyvinuli z vylučovacieho systému, čo viedlo k vytvoreniu urogenitálneho systému.
Diploidný stav poskytuje organizmom extrémne výhody, pretože v tomto stave dochádza k akumulácii rôznych alel. Sexuálne rozmnožovanie má preto aj tú výhodu, že poskytuje organizmomOväčšia možnosť variability v porovnaní s asexuálom, a to hrá v evolúcii zásadnú úlohu.
Partenogenéza má jasné reprodukčné výhody, pretože produkuje iba samičie potomstvo. Je to však zriedkavé. Na vysvetlenie nízkej frekvencie substitúcie pohlavného rozmnožovania partenogenézou v prirodzených populáciách organizmov sa používajú dve hypotézy. Podľa jednej z nich (mutačno-kumulatívna) je pohlavie adaptívnou adaptáciou, keďže „čistí“ genóm od mutácií, ktoré sa časom opakujú, zatiaľ čo podľa inej hypotézy (ekologickej) je pohlavie
Prvé myšlienky o raste a rozvoji sa datujú do starovekého sveta. Už Hippokrates (460-377 pred n. l.) predpokladal, že vajíčka už obsahujú plne sformovaný organizmus, ale vo veľmi redukovanej forme. Táto myšlienka sa potom rozvinula do doktríny preformationizmu (z lat. predformácia - preformácia), ktorý sa ukázal byť obzvlášť populárny v 17.-18. Podporovateľmi preformationizmu boli Harvey, Malpighi a mnohí ďalší významní biológovia a lekári tej doby. Pre preformationistov bola kontroverzná len otázka, v ktorých pohlavných bunkách bol organizmus preformovaný – ženské alebo mužské. Tí, ktorí uprednostňovali vajíčka, sa nazývali ovisti a tí, ktorí pripisovali väčšiu dôležitosť mužským reprodukčným bunkám, sa nazývali animalculists. Preformationizmus je metafyzická doktrína od začiatku do konca, pretože popieral vývoj. Rozhodujúci úder preformacionizmu zasadil C. Bonnet (1720-1793), ktorý partenogenézu objavil v roku 1745 na príklade vývoja vošiek z neoplodnených vajíčok. Potom sa preformacionizmus už nemohol zotaviť a začal strácať svoj význam.
V starovekom svete vznikla ďalšia doktrína, ktorá bola v protiklade k preformationizmu a neskôr dostala názov epigenéza (z gréčtiny. epi- po, genéza - rozvoj). Podobne ako preformacionizmus, aj epigenéza sa rozšírila v 17. – 18. storočí. Pri šírení epigenézy mali veľký význam názory K.F. Wolf (1733-1794), zhrnuté vo svojej knihe „Teória vývoja“ (1759). K.F. Wolf veril, že vajce neobsahuje ani predtvarovaný organizmus, ani jeho časti a že pozostáva z pôvodne homogénnej hmoty. Na rozdiel od preformistov sú názory K.F. Wolf a ďalší priaznivci epigenézy boli na svoju dobu pokrokoví, pretože obsahovali myšlienku rozvoja. Neskôr sa však objavili nové momenty. Najmä v roku 1828 K. Baer publikoval svoju prácu „História vývoja zvierat“, v ktorej ukázal, že obsah vajíčka je heterogénny, teda štruktúrovaný a stupeň štruktúry sa zvyšuje s vývojom embrya. K. Baer teda ukázal nekonzistentnosť preformationizmu aj epigenézy.
Rast a vývoj sú najdôležitejšie vlastnosti živých vecí. Rast je nárast hmotnosti celého organizmu v dôsledku zvýšenia počtu buniek, zatiaľ čo vývoj sú kvalitatívne zmeny v tele, ktoré sú determinované bunkovou diferenciáciou a morfogenézou, zabezpečujúce progresívne zmeny u jednotlivcov, počnúc vajíčkami. a končiac ich stavom dospelosti.
ontogenéza (z gréčtiny. na - stvorenie, genéza - vývoj) je história (cyklus) vývoja jednotlivca, počnúc vytvorením zárodočných buniek, z ktorých vznikol, a končiac jeho smrťou. Predstavy o ontogenéze sú založené na údajoch o raste, vývoji a diferenciácii. Štúdium základných princípov ontogenézy je dôležité pre pochopenie biológie človeka.
Moderné údaje o vývoji organizmov odmietajú preformacionizmus aj epigenézu. V rámci moderných koncepcií sa vývoj organizmu chápe ako proces, v ktorom skôr vytvorené štruktúry stimulujú vývoj štruktúr nasledujúcich. Proces vývoja je daný geneticky a úzko súvisí s prostredím. V dôsledku toho je vývoj určený jednotou vnútorných a vonkajších faktorov.
JEDNOTA RASTU A VÝVOJA
Rast organizmu je postupné zväčšovanie jeho hmoty a zmena tvaru v dôsledku zvyšovania počtu buniek a ich diferenciácie, tvorby tkanív a orgánov a biochemických zmien v bunkách a tkanivách. Rast je teda výsledkom kvantitatívnych zmien v podobe nárastu počtu buniek (telesnej hmotnosti) a kvalitatívnych zmien v podobe diferenciácie a morfogenézy buniek. Bunková diferenciácia je proces, pri ktorom sa niektoré bunky morfologicky, biochemicky a funkčne líšia od iných buniek. Reprodukcia a diferenciácia niektorých buniek je vždy koordinovaná s rastom a diferenciáciou iných. Oba tieto procesy prebiehajú počas celého životného cyklu organizmu. Keďže diferencované bunky menia svoj tvar a skupiny buniek sa podieľajú na zmenách tvaru, je to sprevádzané morfogenézou, čo je súbor procesov, ktoré určujú štruktúrnu organizáciu buniek a tkanív, ako aj všeobecnú morfológiu organizmov.
Rast možno merať zostrojením kriviek veľkosti tela, hmotnosti, sušiny, počtu buniek, obsahu dusíka a iných ukazovateľov na základe výsledkov merania.
ONTOGENÉZA A JEJ TYPY. PERIODIZÁCIA ONTOGENÉZY
Ontogenéza sa v závislosti od charakteru vývoja organizmov delí na priamy a nepriamy, a preto sa rozlišuje priamy a nepriamy vývoj. Priamy vývoj organizmov v prírode prebieha vo forme nelarválneho a vnútromaternicového vývoja, zatiaľ čo nepriamy vývoj sa pozoruje vo forme larválneho vývoja. Na rozdiel od ontogenézy je druhovou kategóriou fylogenéza.
Vývoj lariev. Tento vývoj sa chápe ako nepriamy vývoj, pretože organizmy majú vo svojom vývoji jedno alebo viac larválnych štádií. Vývoj lariev je typický pre hmyz, obojživelníky a ostnokožce. Larvy týchto zvierat vedú nezávislý životný štýl a potom prechádzajú transformáciou. Preto sa tento vývoj nazýva aj vývoj s metamorfózou (pozri nižšie).
Nelarválny vývoj. Táto forma vývoja je charakteristická pre organizmy, ktoré sa vyvíjajú priamo, napríklad ryby, plazy a vtáky, ktorých vajcia sú bohaté na žĺtok (živný materiál). Značná časť ontogenézy sa vďaka tomu odohráva vo vajíčkach znesených vo vonkajšom prostredí. Metabolizmus embryí je zabezpečený vyvíjajúcimi sa provizórnymi orgánmi, ktorými sú embryonálne membrány (žĺtkový vak, amnion, alantois).
Vnútromaternicový vývoj. Tento vývoj je charakteristický aj pre priamo sa vyvíjajúce organizmy, ako sú cicavce vrátane človeka. Keďže vajíčka týchto organizmov sú veľmi chudobné na živiny, všetky vitálne funkcie embryí zabezpečuje materské telo tvorbou provizórnych orgánov z tkanív matky a embrya, medzi ktorými je hlavná placenta. Evolučne je vnútromaternicový vývoj najnovšou formou, no pre embryá je najprospešnejší, pretože efektívne zabezpečuje ich prežitie.
Ontogenéza sa delí na proembryonálne, embryonálne a postembryonálne obdobia. V prípade ľudí sa obdobie vývoja pred narodením nazýva prenatálne alebo antenatálne, po narodení - postnatálne. Vyvíjajúce sa embryo pred vytvorením orgánových rudimentov sa nazýva embryo, po vytvorení orgánových rudimentov - plod.
PROEMBRYONÁLNY VÝVOJ
Toto obdobie v individuálnom vývoji organizmov je spojené s tvorbou gamét v procese gametogenézy. Mužské reprodukčné bunky nemajú významné rozdiely od iných buniek, zatiaľ čo vajcia sa líšia tým, že obsahujú veľa žĺtka. Vzhľadom na množstvo žĺtka a jeho distribúciu vo vajciach sú tieto rozdelené do troch typov:
1) izolecitálne vajcia, obsahujúci trochu žĺtka, ktorý je lokalizovaný rovnomerne v celej bunke. Tieto vajíčka produkujú ostnokožce (morské ježovky), nižšie strunatce (lancelety) a cicavce;
2) telolecitné vajíčka obsahujú veľké množstvo žĺtka, ktorý sa koncentruje na jednom z pólov – vegetatívnom. Takéto vajcia produkujú mäkkýše, obojživelníky, plazy a vtáky. Napríklad žabie vajcia pozostávajú z 50% žĺtka, kuracie vajcia (bežne kuracie vajcia) - 95%. Na druhom póle (živočích) telolecitálnych vajíčok sa koncentruje cytoplazma a jadro;
3) centrolecitálne oocyty, v ktorej je málo žĺtka a zaujíma centrálnu polohu. Cytoplazma sa nachádza na periférii takýchto vajíčok. Centrolecitálne vajíčka produkujú článkonožce.
Proembryonálne obdobie je charakteristické aj tým, že v tomto období prebiehajú v gamétach metabolické procesy spojené s akumuláciou molekúl DNA.
EMBRYONÁLNY VÝVOJ
Embryogenéza (z gréčtiny. etbryop - zárodok), alebo embryonálne obdobie, začína splynutím samčích a samičích zárodočných buniek, ktoré predstavujú proces oplodnenia vajíčok.
U organizmov charakterizovaných vnútromaternicovým vývojom sa embryonálne obdobie končí narodením a u organizmov charakterizovaných larválnym a nelarválnym typom vývoja sa embryonálne obdobie končí uvoľnením organizmu z vajíčka, resp. embryonálnych membrán. V rámci embryonálneho obdobia sa rozlišujú štádiá zygota, štiepenie, blastula, tvorba zárodočných vrstiev, histogenéza a organogenéza.
Zygota.Hnojenie zahŕňa sériu procesov, pri ktorých samčia reprodukčná bunka iniciuje vývoj vajíčka. Vo vajíčku aktivovanom mužskou gamétou prebieha množstvo fyzikálnych a chemických procesov, vrátane zvýšenej syntézy bielkovín. Pohyb protoplazmy vedie k vytvoreniu bilaterálnej symetrie vajíčka. Jadrá sa spoja a obnoví sa diploidná sada chromozómov. Vznikne tak jednobunkový organizmus.
Rozdelenie.Predstavuje počiatočné obdobie vývoja zygoty (oplodneného vajíčka), ktoré pozostáva z delenia zygoty mitózou. Rozdelenie začína objavením sa brázdy na povrchu vajíčka. Prvá brázda vedie k vytvoreniu dvoch buniek - dve blastoméry, druhá - štyri blastoméry, tretia - osem blastomérov (obr. 25). Skupina buniek vytvorená v dôsledku postupných fragmentácií sa nazýva morula (z lat. morum- moruše).
Biologický význam tohto štádia spočíva v tom, že z veľkej bunky, ktorou je vajíčko, vznikajú menšie bunky, v ktorých sa znižuje pomer cytoplazmy k jadru.
Fragmentácia zygoty končí vytvorením mnohobunkovej štruktúry nazývanej blastula (z gréčtiny. blastos - klíčiť). Táto štruktúra má tvar vezikuly nazývanej blastoderm, ktorá pozostáva z jednej vrstvy buniek. Teraz sa tieto bunky nazývajú embryonálne. Blatula má podobnú veľkosť ako vajíčko. Počas obdobia fragmentácie sa zvyšuje počet jadier a celkový počet DNA. Syntetizuje sa aj malé množstvo mRNA a tRNA, zatiaľ čo ribozomálna RNA ešte nie je detegovateľná.
Všetky zvieratá prechádzajú štádiom blastuly, ale každý prípad má svoje vlastné charakteristiky. U cicavcov je delenie nerovnomerné, takže morule pozostávajú z rôzneho počtu buniek. Okrem toho časť buniek tvorí štruktúru nazývanú trofoblast,

Larva Blastocoel Pulec
formulár
Ryža. 25.Fragmentácia zygotov a tvorba blastúl v rôznych organizmoch: 1 - pôvodné vajíčko; 2 - dve blastoméry; 3 - štyri blastoméry; 4 - osem blastomérov; 5 - blastula; 6 - forma pre dospelých
bunky, ktoré vyživujú embryo a vďaka enzýmom zabezpečujú jeho prienik do steny maternice. Neskôr sa bunky trofoblastu z embrya odlúpnu a vytvoria vezikula, ktorá je naplnená tekutinou z tkaniva maternice.
Biologický význam tohto štádia spočíva v tom, že z veľkej bunky, akou je vajíčko, vznikajú menšie bunky, v ktorých sa zníži pomer cytoplazmy k jadru a jadro má nové cytoplazmatické prostredie.
Gastrulácia(z gréčtiny gastre - dutina cievy). Ide o proces pohybu embryonálnych buniek po vytvorení blastuly, ktorý je sprevádzaný tvorbou dvoch alebo troch (v závislosti od druhu zvieraťa) vrstiev embrya, alebo tzv. zárodočných vrstiev (obr. 26). ).
Vývoj (gastrulácia) izolecitálnych vajíčok nastáva invagináciou (invagináciou) vegetatívneho pólu vo vnútri blastuly, v dôsledku čoho sa opačné póly takmer spájajú a blastocoel (dutina blastuly) takmer alebo úplne zmizne. Vonkajšie
Ryža. 26.Gastrulácia v rôznych organizmoch: 1 - blastocoel; 2 - primárny mezenchým; 3, 4, 5 - primárne črevo
Vrstva embryonálnych buniek sa nazýva ektoderm (z gréčtiny. ektos - vonku, derma - koža), alebo vonkajšia zárodočná vrstva, zatiaľ čo vnútorná je endoderm (z gréčtiny. entos - vnútri), alebo vnútorná zárodočná vrstva. V tomto prípade vytvorená dutina sa nazýva gastrocoel alebo primárne črevo, ktorého vstup sa nazýva blastopór (primárne ústa).
Pre huby a koelenteráty je charakteristický vývoj dvoch zárodočných vrstiev. Strunatce v období gastrulácie sa však vyznačujú vývojom tretej zárodočnej vrstvy - mezodermu (z gréčtiny. Mesos - stredná), vytvorená medzi ektodermou a endodermou.
Gastrulácia je nevyhnutným predpokladom pre ďalšie štádiá vývoja, pretože privádza bunky do polohy, ktorá umožňuje tvorbu orgánov. Embryonálny materiál diferencovaný do troch embryonálnych anlage dáva vznik všetkým tkanivám a orgánom vyvíjajúceho sa embrya.
Vývoj (diferenciácia) zárodočných vrstiev je sprevádzaná tvorbou rôznych tkanív a orgánov z nich. Najmä epidermis kože, nechtov a vlasov, mazové a potné žľazy, nervový systém (mozog, miecha, gangliá, nervy), receptorové bunky zmyslových orgánov, očná šošovka, epitel ústnej dutiny , nosová dutina a konečník, zub -
naya smalt. Z endodermu sa vyvíja epitel pažeráka, žalúdka, čriev, žlčníka, priedušnice, priedušiek, pľúc, močovej trubice, ako aj pečene, pankreasu, štítnej žľazy, prištítnych teliesok a týmusu. Z mezodermu sa vyvíja hladké svalstvo, kostrové a srdcové svalstvo, dermis, spojivové tkanivo, kosti a chrupavky, zubovina, krv a cievy, mezentéria, obličky, semenníky a vaječníky. U ľudí sa ako prvé oddelí mozog a miecha. Po 2 mesiacoch sa objavia takmer všetky štruktúry tela. Spojenie embrya s prostredím sa uskutočňuje prostredníctvom provizórnych orgánov. Organogenéza končí na konci embryonálneho obdobia. Ak sa definitívny ústny otvor vytvorí v mieste primárneho ústia (blastopór), potom sa tieto zvieratá nazývajú protostómy (červy, mäkkýše, článkonožce).
Ak sa definitívna tlama vytvorí na opačnom mieste, potom sa tieto živočíchy nazývajú deuterostómy (ostnatokožce, strunatce).
Na zabezpečenie spojenia embrya s prostredím sa používajú takzvané provizórne orgány, ktoré existujú dočasne. V závislosti od typu vajíčok sú dočasné orgány rôzne štruktúry. U rýb, plazov a vtákov medzi dočasné orgány patrí žĺtkový vak. U cicavcov sa žĺtkový vak vytvára na začiatku embryogenézy, ale nevyvíja sa. Neskôr sa zníži. Vonkajšia membrána embrya sa nazýva chorion. Prerastá do maternice. Miesto najväčšieho vrastania do maternice sa nazýva placenta. Embryo je spojené s placentou cez pupočnú šnúru alebo pupočnú šnúru, ktorá obsahuje krvné cievy, ktoré zabezpečujú krvný obeh placenty. Metabolizmus plodu je zabezpečený cez placentu.
Formatívna interakcia častí embrya je založená na určitých koordinovaných metabolických procesoch. Vzorec vývoja je heterochrónia, ktorá sa chápe ako rôzna tvorba orgánových anlág v čase a rôzna intenzita ich vývoja. Tie orgány a systémy, ktoré by mali začať fungovať skôr, sa vyvíjajú rýchlejšie. Napríklad u ľudí sa rudimenty horných končatín vyvíjajú rýchlejšie ako dolné.
Embryá sú mimoriadne citlivé na rôzne vplyvy. Preto sa rozlišujú kritické obdobia, t. j. obdobia, kedy sú embryá najcitlivejšie na škodlivé faktory. V prípade ľudí kritické obdobia embryonálnej ontogenézy
sú prvé dni po oplodnení, čas tvorby placenty a pôrodu.
Jadrá somatických buniek sú schopné zabezpečiť normálny vývoj vajíčok, čo bolo objasnené v experimentoch na transplantácii jadier somatických buniek do vajíčok bez jadier.
Experimenty tiež ukázali, že transformácia jednej blastoméry z 8- a 16-bunkových embryí oviec jedného plemena na bezjadrovú polovicu vajíčka (po rozrezaní na polovicu) iného plemena bola sprevádzaná vývojom životaschopných embryí. a narodenie jahniat.
Od čias Hippokrata (5. storočie pred n. l.) sa diskutovalo o otázke dôvodov, ktoré iniciujú narodenie plodu. Najmä sám Hippokrates navrhol, že vývoj plodu iniciuje svoj vlastný pôrod. Najnovšia experimentálna práca anglických výskumníkov na ovciach ukázala, že u oviec je začatie jahňacieho mäsa riadené komplexom hypotalamus + hypofýza + nadobličky plodu. Poškodenie jadier hypotalamu, odstránenie predného laloka hypofýzy alebo nadobličiek predlžuje graviditu oviec. Naopak, podávanie adenokortikotropného hormónu (výlučok z predného laloku hypofýzy) alebo kortizolu (výlučok z nadobličiek) ovciam skracuje trvanie ich gravidity.
Takže v procese vývoja vyšších eukaryotov jediná oplodnená bunka zygoty počas ďalšieho vývoja v dôsledku mitózy dáva vznik bunkám rôznych typov - epitelové, nervové, kostné, krvinky a iné, ktoré sa vyznačujú rozmanitosť morfológie a makromolekulárneho zloženia. Pre bunky rôznych typov je však charakteristické aj to, že obsahujú rovnaké sady génov, ale sú vysoko špecializované a vykonávajú iba jednu alebo niekoľko špecifických funkcií, to znamená, že niektoré gény „fungujú“ v bunkách, zatiaľ čo iné sú neaktívne. Napríklad,

Ryža. 27. Vývoj jednovaječných dvojčiat
iba červené krvinky sú špecifické pri syntéze a skladovaní hemoglobínu. Rovnakým spôsobom iba epidermálne bunky syntetizujú keratín. Preto už dlho vznikajú otázky o genetickej identite jadier somatických buniek a o kontrolných mechanizmoch vývoja oplodnených vajíčok ako predpokladu pre pochopenie mechanizmov, ktoré sú základom diferenciácie buniek. Kmeňové bunky majú veľký význam pri diferenciácii buniek (pozri nižšie). Pokusy na myšiach ukázali, že bunky jedného typu sú schopné premeny na bunky iného typu. Najmä sa ukázalo, že diferencované alebo diferencované pečeňové bunky sa premieňajú na bunky pankreasu.
Od 50-tych rokov. XX storočia V mnohých laboratóriách sa robili pokusy s úspešnou transplantáciou jadier somatických buniek do vajíčok umelo zbavených vlastných jadier. Štúdia DNA z jadier rôznych diferencovaných buniek ukázala, že takmer vo všetkých prípadoch genómy obsahujú rovnaké sady sekvencií nukleotidových párov. Sú známe výnimky, kedy červené krvinky cicavcov strácajú svoje jadrá v poslednom štádiu diferenciácie. Ale v tomto čase bazény
Niektoré mRNA hemoglobínu už boli syntetizované, takže jadrá už nie sú potrebné pre červené krvinky. Ďalšie príklady zahŕňajú imunoglobulínové a T-bunkové gény, ktoré sú modifikované počas vývoja.
Jednou z hlavných etáp v smere pochopenia kontrolných mechanizmov embryonálnej ontogenézy boli výsledky experimentov uskutočnených v 60. – 70. rokoch 20. storočia. Anglický výskumník D. Gurdon s cieľom zistiť, či jadrá somatických buniek majú schopnosť zabezpečiť ďalší vývoj vajíčok, ak sa tieto jadrá zavedú do vajíčok, z ktorých boli predtým odstránené ich vlastné jadrá. Na obr. Obrázok 28 ukazuje diagram jedného z týchto experimentov, v ktorom boli jadrá somatických buniek pulcov transplantované do žabích vajíčok s predtým odstránenými jadrami. Tieto experimenty ukázali, že jadrá somatických buniek skutočne dokážu zabezpečiť ďalší vývoj vajíčok, keďže dokázali vajíčka oplodniť a „prinútiť“ ich k ďalšiemu vývoju. To dokázalo možnosť klonovania zvierat.

Ryža. 28.Schéma experimentu s transplantáciou jadier somatických buniek do bezjadrových vajíčok (D. Gurdon, 1968)
Neskôr ďalší výskumníci vykonali experimenty, v ktorých ukázali, že prenos jednotlivých blastomér z 8- a 16-dňových embryí oviec jedného plemena do bezjadrovej polovice vajíčka (po rozrezaní druhej na polovicu) iného plemena bol sprevádzaný tzv. tvorba životaschopných embryí s následným narodením jahniat.
Začiatkom roku 1997 anglickí autori zistili, že zavedenie jadier somatických buniek (buniek embryí, plodov alebo vemien dospelých oviec) do umelo zbavených ovčích vajec a následná implantácia takto oplodnených vajíčok do maternice oviec výskyt gravidity s následným narodením jahniat (obr. 29). Jedno z jahniat sa volalo Dolly. V roku 2003 Dolly zomrela. Počas tejto doby boli získané embryá myší, kráv, králikov, koní, potkanov a iných zvierat.

Zavedenie kultúry do tela matky
Ryža. 29.Transplantácia jadier somatických buniek do vajíčok bez jadier
Vyhodnotenie týchto výsledkov ukazuje, že cicavce sa môžu množiť nepohlavne, pričom produkujú potomstvo zvierat, ktorých bunky obsahujú jadrový materiál otcovského alebo materského pôvodu, v závislosti od pohlavia darcovskej ovce. V takýchto bunkách
len cytoplazma a mitochondrie sú materského pôvodu. Tento záver má mimoriadne dôležitý všeobecný biologický význam a rozširuje naše názory na reprodukčný potenciál zvierat. Dôležité je ale tiež dodať, že hovoríme o genetických manipuláciách, ktoré v prírode neexistujú. Na druhej strane z praktického hľadiska tieto genetické manipulácie predstavujú priamy spôsob klonovania vysoko organizovaných zvierat s požadovanými vlastnosťami, čo má veľký ekonomický význam. Z lekárskeho hľadiska môže byť táto cesta v budúcnosti použitá na prekonanie mužskej neplodnosti.
Takže genetická informácia potrebná pre normálny vývoj embrya sa nestráca počas bunkovej diferenciácie vďaka takzvaným kmeňovým bunkám, ktoré majú potenciál vyvinúť sa na rôzne typy telesných buniek. Keď sa kmeňové bunky delia, každá nová bunka má potenciál buď zostať kmeňovou bunkou, alebo sa stať bunkou so špecializovanejšou funkciou (svalové bunky, krvinky alebo mozgové bunky). Oplodnené vajíčko je totipotentné, pretože z neho v tele vznikajú rôzne typy buniek. Totipotentné kmeňové bunky môžu viesť k vzniku akéhokoľvek typu buniek okrem tých, ktoré sú potrebné pre vývoj plodu. Bežne sa nazývajú kmeňové bunky, ktoré môžu viesť k vzniku širokej škály typov buniek multitotipotentný bunky. Zistilo sa, že dospelé kmeňové bunky môžu produkovať diferencované bunky z embryonálnych nepríbuzných tkanív. Somatické bunky majú tiež vlastnosť nazývanú totipotencia, t.j. ich genóm obsahuje všetky informácie, ktoré dostali z oplodneného vajíčka, ktoré im umožnilo vznik v dôsledku diferenciácie. Prítomnosť týchto údajov nepochybne znamená, že diferenciácia buniek podlieha genetickej kontrole. Štúdium kmeňových buniek má dôsledky pre medicínu.
Zistilo sa, že intenzívna syntéza proteínov po oplodnení u väčšiny eukaryotov nie je sprevádzaná syntézou mRNA. Štúdium oogenézy u stavovcov, najmä u obojživelníkov, ukázalo, že intenzívna transkripcia prebieha aj počas profázy I (najmä diplonémy) meiózy. Preto sú génové transkripty vo forme molekúl mRNA alebo pro-mRNA uložené vo vajíčkach v kľudovom stave. Zistilo sa, že v embryonálnych bunkách existuje takzvaná asymetrickosť
ternárne delenie, ktoré spočíva v tom, že delením embryonálnej bunky vznikajú dve bunky, z ktorých len jedna dedí proteíny podieľajúce sa na transkripcii. Nerovnomerná distribúcia transkripčných faktorov medzi dcérskymi bunkami teda vedie k expresii rôznych súborov génov v nich po delení, teda k diferenciácii buniek.
U obojživelníkov a snáď väčšiny stavovcov sa genetické programy, ktoré riadia skorý vývoj (pred štádiom blastuly), vytvárajú počas oogenézy. Neskoršie štádiá vývoja, keď začína bunková diferenciácia (približne od štádia gastruly), vyžadujú nové programy na génovú expresiu. Bunková diferenciácia je teda spojená s preprogramovaním genetickej informácie jedným alebo druhým smerom.
Charakteristickým znakom diferenciácie buniek je, že nezvratne vedie k jednému alebo druhému typu bunky. Tento proces sa nazýva rozhodnosť a je tiež pod genetickou kontrolou, a ako sa teraz predpokladá, bunková diferenciácia a determinácia sú regulované interakciou buniek na základe signálov uskutočňovaných peptidovými rastovými faktormi prostredníctvom tyrozínkinázových receptorov. Pravdepodobne existuje veľa takýchto systémov. Jedným z nich je, že diferenciácia svalových a nervových buniek je regulovaná neuroregulínmi, čo sú membránové proteíny, ktoré pôsobia prostredníctvom jedného alebo viacerých tyrozínkinázových receptorov.
Genetická kontrola determinácie je demonštrovaná aj existenciou takzvaných homeiotropných alebo homeotických mutácií, ktoré u hmyzu spôsobujú zmeny v determinácii špecifických imaginárnych diskov. Výsledkom je, že niektoré časti tela sa vyvíjajú nesprávne. Napríklad u Drosophila mutácie transformujú určenie anténneho disku na disk, ktorý sa vyvinie do apendixu končatiny siahajúceho od hlavy. U hmyzu rodu Ophthalmoptera krídlové štruktúry sa môžu vyvinúť z očného disku. U myší bola preukázaná existencia génového zhluku (komplexu) Hox, ktorý pozostáva z 38 génov a riadi vývoj končatín.
Samostatný význam má otázka regulácie génovej aktivity počas embryonálneho vývoja. Predpokladá sa, že počas diferenciácie gény pôsobia v rôznych časoch, čo sa prejavuje
pri transkripcii v rôznych diferencovaných bunkách rôznych mRNA, t.j. dochádza k represii a derepresii génov. Napríklad počet génov prepísaných do RNA v blastocytoch morského ježka je 10 %, v pečeňových bunkách potkanov je to tiež 10 % a v bunkách týmusu hovädzieho dobytka je to 15 %. Predpokladá sa, že nehistónové proteíny sa podieľajú na kontrole transkripčného stavu génov. Nasledujúce údaje podporujú tento predpoklad. Keď je bunkový chromatín vo fáze S prepísané v systéme in vitro, potom sa syntetizuje len histónová mRNA, po ktorej nasledujú históny. Naopak, pri použití chromatínu buniek z β-fázy nedochádza k syntéze histónovej mRNA, keď sú nepiestónové proteíny z chromatínu β-fázy odstránené a sú nahradené nehistónovými chromozomálnymi proteínmi syntetizovanými v fáza S, potom po transkripcii takéhoto chromatínu in vitro syntetizuje sa histónová mRNA. Navyše, keď nehistónové proteíny pochádzajú z 1-fázových buniek a DNA a históny pochádzajú z buniek S-fázy, nesyntetizuje sa žiadna histónová mRNA. Tieto výsledky naznačujú, že nehistónové proteíny obsiahnuté v chromatíne určujú schopnosť prepisovať gény kódujúce históny. Preto sa domnievame, že nehistónové chromozomálne proteíny môžu hrať dôležitú úlohu pri regulácii a expresii génov v eukaryotoch.
Dostupné údaje naznačujú, že proteínové a steroidné hormóny sa podieľajú na regulácii transkripcie u zvierat. Proteínové (inzulín) a steroidné (estrogén a testosterón) hormóny sú dva signalizačné systémy používané v medzibunkovej komunikácii. U vyšších živočíchov sa hormóny syntetizujú v špecializovaných sekrečných bunkách. Uvoľnené do krvného obehu vstupujú do tkanív. Keďže molekuly proteínových hormónov sú relatívne veľké, neprenikajú do buniek, takže ich účinky zabezpečujú receptorové proteíny lokalizované v membránach cieľových buniek a intracelulárne hladiny cyklického AMP (cAMP). Naopak, steroidné hormóny sú malé molekuly, v dôsledku čoho ľahko prenikajú do buniek cez membrány. Keď sú vo vnútri buniek, viažu sa na špecifické receptorové proteíny, ktoré sa nachádzajú iba v cytoplazme cieľových buniek. Zvažujú sa komplexy hormón - proteínový receptor, koncentrujú sa v jadrách cieľových buniek a aktivujú transkripciu špecifických génov prostredníctvom interakcie s určitými
mi nehistónové proteíny, ktoré sa viažu na promótorové oblasti špecifických génov. V dôsledku toho väzba komplexu hormón + proteín (proteínový receptor) s nehistónovými proteínmi uvoľňuje promótorové oblasti pre pohyb RNA polymerázy. Zhrnutím údajov o genetickom riadení embryonálneho obdobia v ontogenéze organizmov môžeme konštatovať, že jeho priebeh je riadený rozdielnym zapínaním a vypínaním pôsobenia génov v rôznych bunkách (tkanive) prostredníctvom ich derepresie a represie.
POSTEMBRYONÁLNY VÝVOJ
Po narodení organizmu sa začína jeho postembryonálny vývoj (u človeka postnatálny), ktorý u rôznych organizmov trvá od niekoľkých dní až po stovky rokov v závislosti od ich druhu. V dôsledku toho je priemerná dĺžka života druh charakteristický pre organizmy, ktorý nezávisí od úrovne ich organizácie (pozri nižšie).
V postembryonálnej ontogenéze sa rozlišuje juvenilné a pubertálne obdobie, ako aj obdobie staroby, končiace smrťou.
Juvenilné obdobie. Toto obdobie (od lat. juvenilis- mláďa) je určené časom od narodenia organizmu do puberty. Vyskytuje sa rôznymi spôsobmi a závisí od typu ontogenézy organizmov. Toto obdobie je charakteristické buď priamym alebo nepriamym vývojom.
V prípade organizmov, ktoré sa vyznačujú priamym vývojom (veľa bezstavovcov, rýb, plazov, vtákov, cicavcov, ľudí), sú organizmy vyliahnuté z vaječných škrupín alebo novorodencov podobné dospelým formám, líšia sa od nich len menšími veľkosťami. ako nevyvinutosť jednotlivých orgánov a nedokonalé telesné proporcie (obr. 30).
Charakteristickým znakom rastu v juvenilnom období organizmov podliehajúcich priamemu vývoju je zvýšenie počtu a veľkosti buniek a zmena proporcií tela. Rast človeka počas rôznych období jeho ontogenézy je znázornený na obr. 31. Rast rôznych ľudských orgánov je nerovnomerný. Napríklad rast hlavy končí v detstve, nohy dosahujú proporcionálnu veľkosť asi o 10 rokov. Vonkajšie pohlavné orgány rastú veľmi rýchlo vo veku 12 až 14 rokov. Rozlišuje sa určitý a neurčitý rast. Určitý rast je charakteristický pre organizmy, ktoré v určitom veku prestanú rásť,
napríklad hmyz, cicavce, ľudia. Neobmedzený rast je charakteristický pre organizmy, ktoré rastú počas svojho života, napríklad mäkkýše, ryby, obojživelníky, plazy a mnohé druhy rastlín.

Ryža. tridsať.Priamy a nepriamy vývoj organizmov rôznych druhov
V prípade nepriameho vývoja organizmy prechádzajú premenami nazývanými metamorfózy (z lat. metamorfóza - transformácia).

Ryža. 31.Rast a vývoj v rôznych obdobiach ľudskej ontogenézy
Predstavujú modifikácie organizmov počas vývoja. Metamorfózy sa bežne vyskytujú v coelenterátoch (hydra, medúzy, koralové polypy), plochých červoch (fasciola), škrkavkách (škrkavkách), mäkkýšoch (ustrice, mušle, chobotnice), článkonožcoch (raky, riečne kraby, homáre, krevety, škorpióny, pavúky, roztoče , hmyz) a dokonca aj u niektorých strunatcov (plášteníkov a obojživelníkov). V tomto prípade sa rozlišujú úplné a neúplné metamorfózy. Najvýraznejšie formy metamorfózy sa pozorujú u hmyzu, ktorý prechádza neúplnou aj úplnou metamorfózou.
Neúplná premena je vývoj, pri ktorom z vaječných škrupín vzniká organizmus, ktorého štruktúra je podobná stavbe dospelého organizmu, ale jeho veľkosť je oveľa menšia. Takýto organizmus sa nazýva larva. Počas procesu rastu a vývoja sa veľkosť lariev zväčšuje, ale existujúci chitinizovaný obal bráni ďalšiemu zväčšovaniu veľkosti tela, čo vedie k línaniu, teda k odlupovaniu chitinizovaného obalu, pod ktorým je mäkká kutikula. Ten sa narovná a to je sprevádzané zväčšením veľkosti zvieraťa. Po niekoľkých moultoch zviera dosiahne zrelosť. Neúplná premena je typická napríklad v prípade vývoja ploštice domácej.
Úplná metamorfóza je vývoj, pri ktorom sa z vaječných škrupín uvoľní larva, výrazne odlišná
v štruktúre od dospelých jedincov. Napríklad u motýľov a mnohých druhov hmyzu sú larvy húsenice. Húsenice podliehajú línaniu a môžu sa niekoľkokrát pretaviť a potom sa premeniť na kukly. Z tých druhých sa vyvíjajú dospelé formy (imago), ktoré sa nelíšia od pôvodných.
U stavovcov sa metamorfóza vyskytuje medzi obojživelníkmi a kostnatými rybami. Larválne štádium je charakterizované prítomnosťou provizórnych orgánov, ktoré buď opakujú vlastnosti predkov, alebo majú jednoznačne adaptačný význam. Napríklad pulec, ktorý je larválnou formou žaby a opakuje vlastnosti pôvodnej formy, sa vyznačuje tvarom podobným rybe, prítomnosťou žiabrového dýchania a jedným kruhom krvného obehu. Adaptačnými znakmi pulcov sú ich prísavky a dlhé črevá. Pre larválne formy je charakteristické aj to, že v porovnaní s dospelými formami sú prispôsobené na život v úplne iných podmienkach, zaberajú inú ekologickú niku a iné miesto v potravinovom reťazci. Napríklad larvy žiab majú žiabrové dýchanie, zatiaľ čo dospelé formy majú pľúcne dýchanie. Na rozdiel od dospelých foriem, ktoré sú mäsožravce, sa larvy žiab živia rastlinnou potravou.
Postupnosť udalostí vo vývoji organizmov sa často nazýva životné cykly, ktoré môžu byť jednoduché alebo zložité. Najjednoduchšie vývojové cykly sú charakteristické napríklad pre cicavce, kedy sa z oplodneného vajíčka vyvinie organizmus, ktorý opäť produkuje vajíčka atď. Zložité biologické cykly sú cykly u živočíchov, ktoré sa vyznačujú vývojom s metamorfózou. Poznatky o biologických cykloch majú praktický význam najmä v prípadoch, keď sú organizmy patogénmi alebo prenášačmi patogénov u zvierat a rastlín.
Vývoj a diferenciácia spojená s metamorfózou sú výsledkom prirodzeného výberu, vďaka ktorému sú mnohé larválne formy, ako napríklad húsenice hmyzu a žaby, lepšie prispôsobené prostrediu ako dospelé sexuálne zrelé formy.
Puberta. Toto obdobie sa nazýva aj zrelé a súvisí s pohlavnou zrelosťou organizmov. Vývoj organizmov v tomto období dosahuje maximum.
Rast a vývoj v postembryonálnom období do značnej miery ovplyvňujú faktory prostredia. Pre rastliny sú rozhodujúce svetlo, vlhkosť, teplota, množstvo a kvalita živín v pôde. Pre zvieratá má prvoradý význam správne kŕmenie (prítomnosť bielkovín, sacharidov, lipidov, minerálnych solí, vitamínov, mikroelementov v krmive). Dôležitý je aj kyslík, teplota, svetlo (syntéza vitamínu D).
Rast a individuálny vývoj živočíšnych organizmov podlieha neurohumorálnej regulácii humorálnymi a nervovými regulačnými mechanizmami. V rastlinách boli objavené účinné látky podobné hormónom nazývané fytohormóny. Posledne menované ovplyvňujú životné funkcie rastlinných organizmov.
V živočíšnych bunkách sa počas ich životných procesov syntetizujú chemicky aktívne látky, ktoré ovplyvňujú životné procesy. Nervové bunky bezstavovcov a stavovcov produkujú látky nazývané neurosekréty. Endokrinné alebo vnútorné sekrečné žľazy tiež vylučujú látky nazývané hormóny. Endokrinné žľazy, najmä tie, ktoré súvisia s rastom a vývojom, sú regulované neurosekréciou. U článkonožcov je regulácia rastu a vývoja veľmi dobre znázornená vplyvom hormónov na prelínanie. Syntéza sekrétov lariev bunkami je regulovaná hormónmi, ktoré sa hromadia v mozgu. Špeciálna žľaza u kôrovcov produkuje hormón, ktorý inhibuje preliatie. Hladiny týchto hormónov určujú frekvenciu línania. U hmyzu bola stanovená hormonálna regulácia dozrievania vajíčok a diapauzy.
U stavovcov sú endokrinnými žľazami hypofýza, epifýza, štítna žľaza, prištítne telieska, pankreas, nadobličky a pohlavné žľazy, ktoré sú navzájom úzko spojené. Hypofýza u stavovcov produkuje gonadotropný hormón, ktorý stimuluje činnosť pohlavných žliaz. U ľudí hormón hypofýzy ovplyvňuje rast. Pri nedostatku vzniká nanizmus, pri nadbytku gigantizmus. Epifýza produkuje hormón, ktorý ovplyvňuje sezónne výkyvy sexuálnej aktivity zvierat. Hormón štítnej žľazy ovplyvňuje metamorfózu hmyzu a obojživelníkov. U cicavcov nedostatočný rozvoj štítnej žľazy vedie k spomaleniu rastu a nedostatočnému rozvoju pohlavných orgánov. U ľudí je v dôsledku defektu štítnej žľazy oneskorená osifikácia a rast.
(trpaslík), puberta nenastáva, duševný vývoj sa zastaví (kretenizmus). Nadobličky produkujú hormóny, ktoré ovplyvňujú metabolizmus, rast a diferenciáciu buniek. Gonády produkujú pohlavné hormóny, ktoré určujú sekundárne sexuálne charakteristiky. Odstránenie gonád vedie k nezvratným zmenám v rade charakteristík. Napríklad u kastrovaných kohútov sa zastaví rast hrebeňa a stratí sa sexuálny pud. Kastrovaný muž nadobudne vonkajšiu podobnosť so ženou (na koži nerastú fúzy a chlpy, na hrudi a panve sa ukladá tuk, farba hlasu je zachovaná atď.).
Rastlinné fytohormóny sú auxíny, cytokiníny a gibberelíny. Regulujú rast a delenie buniek, tvorbu nových koreňov, vývoj kvetov a ďalšie vlastnosti rastlín.
Vo všetkých obdobiach ontogenézy sú organizmy schopné obnoviť stratené alebo poškodené časti tela. Táto vlastnosť organizmov je tzv regenerácia, ktoré môžu byť fyziologické a reparatívne.
Fyziologická regenerácia - Ide o náhradu stratených častí tela počas životnosti tela. Regenerácie tohto typu sú vo svete zvierat veľmi bežné. Napríklad u článkonožcov je reprezentovaný línaním, ktoré je spojené s rastom. U plazov sa regenerácia prejavuje výmenou chvosta a šupín, u vtákov - perím, pazúrmi a ostrohami. U cicavcov je príkladom fyziologickej regenerácie každoročné zhadzovanie parožia jeleňovou zverou.
Reparatívna regenerácia - Ide o obnovu časti tela organizmu, ktorá bola násilne odtrhnutá. Regenerácia tohto typu je možná u mnohých zvierat, no jej prejavy sú rôzne. Napríklad je to bežné v hydrách a súvisí s ich reprodukciou, pretože celý organizmus sa regeneruje z časti. U iných organizmov sa regenerácia prejavuje v podobe schopnosti jednotlivých orgánov zotaviť sa po strate ktorejkoľvek časti. U ľudí majú epiteliálne, spojivové, svalové a kostné tkanivá dosť vysokú regeneračnú schopnosť.
Rastliny mnohých druhov sú tiež schopné regenerácie.
Údaje o regenerácii majú veľký význam nielen v biológii. Sú široko používané v poľnohospodárstve, medicíne, najmä v chirurgii.
Staroba ako štádium ontogenézy. Staroba je predposledným štádiom ontogenézy zvierat a určuje sa aj jej trvanie
celková dĺžka života, ktorá slúži ako druhová charakteristika a líši sa medzi rôznymi zvieratami. Staroba bola najpresnejšie skúmaná u ľudí.
Existujú rôzne definície ľudskej staroby. Jedna z najpopulárnejších definícií je, že staroba je hromadenie po sebe nasledujúcich zmien, ktoré sprevádzajú zvyšovanie veku organizmu a zvyšujú pravdepodobnosť jeho choroby alebo smrti. Veda o starnutí človeka sa nazýva gerontológia (z gréčtiny. Geron - starý muž, starý muž, logá - veda). Jeho úlohou je študovať vzorce vekového prechodu medzi zrelosťou a smrťou.
Vedecký výskum v gerontológii siaha do rôznych oblastí, od štúdií zmien aktivity bunkových enzýmov až po objasnenie vplyvu psychologických a sociologických zmierňovaní environmentálneho stresu na správanie starých ľudí.
V prípade ľudí sa rozlišuje fyziologická staroba; staroba spojená s kalendárnym vekom; a predčasné starnutie spôsobené sociálnymi faktormi a chorobami. V súlade s odporúčaniami WHO by sa staršia osoba mala považovať za osobu vo veku približne 60 - 75 rokov a za starú osobu vo veku 75 rokov alebo viac.
Ľudskú starobu charakterizuje množstvo vonkajších a vnútorných znakov.
Z vonkajších prejavov staroby sú najvýraznejšie zníženie plynulosti pohybov, zmeny držania tela, zníženie elasticity pokožky, telesnej hmotnosti, pevnosti a pružnosti svalov, vznik vrások na tvári a iných častiach tela. tela a vypadávanie zubov. Napríklad podľa zovšeobecnených údajov osoba vo veku 30 rokov stratí 2 zuby (v dôsledku straty), vo veku 40 rokov - 4 zuby, vo veku 50 rokov - 8 zubov a vo veku 60 rokov - už 11 zubov. Prvý signalizačný systém prechádza výraznými zmenami (ostrosť zmyslových orgánov je otupená). Napríklad maximálna vzdialenosť, na ktorú zdraví ľudia rozlíšia určité identické zvuky, je 12 m vo veku 20-30 rokov, 10 m vo veku 50 rokov, 7 m vo veku 60 rokov a iba 4 m vo veku 70 rokov. druhý signalizačný systém (intonácia reči sa mení, hlas otupí).
Z vnútorných znakov treba v prvom rade spomenúť znaky ako spätný vývoj (involúcia) orgánov. Dochádza k poklesu veľkosti pečene a obličiek, ako aj počtu
počet nefrónov v obličkách (do 80. roku života takmer polovica), čo znižuje funkčnosť obličiek a ovplyvňuje metabolizmus voda-elektrolyt. Znižuje sa elasticita ciev, znižuje sa prekrvenie tkanív a orgánov a zvyšuje sa periférna cievna rezistencia. V kostiach sa hromadia anorganické soli, chrupavka sa mení (vápenate), znižuje sa schopnosť regenerácie orgánov. V bunkách nastávajú výrazné zmeny, spomaľuje sa delenie a obnova ich funkčného tonusu, znižuje sa obsah vody, znižuje sa aktivita bunkových enzýmov, je narušená koordinácia medzi asimiláciou a disimiláciou. V mozgu je narušená syntéza bielkovín, čo vedie k tvorbe abnormálnych bielkovín. Zvyšuje sa viskozita bunkových membrán, je narušená syntéza a využitie pohlavných hormónov a dochádza k zmenám v štruktúre neurónov. V bielkovinách spojivového tkaniva a elasticite tohto tkaniva dochádza k štrukturálnym zmenám. Imunologické reakcie sú oslabené a zvyšuje sa možnosť autoimunitných reakcií. Funkcie endokrinných systémov, najmä pohlavných žliaz, sú znížené. Správanie ostatných znamení v starobe je znázornené na obr. 32.

Ryža. 32.Zmeny niektorých ľudských vlastností s vekom: 1 - rýchlosť nervových impulzov; 2 - úroveň bazálneho metabolizmu; 3 - srdcový index; 4 - úroveň renálnej filtrácie pre inzulín; 5 - dychové objemy pľúc; 6 - úroveň prietoku plazmy v obličkách
Túžba pochopiť podstatu starnutia tela existuje už dlho. V starovekom Grécku Hippokrates veril, že starnutie je spojené s nadbytkom jedla a nedostatočným pobytom na čerstvom vzduchu. Podľa Aristotela je starnutie spôsobené spotrebou tepelnej energie organizmom. Dôležitosť jedla ako faktora starnutia si všimol aj Galén. Dlho však nebolo dostatok vedeckých údajov na objektívne pochopenie tohto problému. Až v 19. storočí. V skúmaní starnutia sa dosiahol určitý pokrok a začali sa objavovať teórie starnutia.
Jednou z prvých najznámejších teórií starnutia ľudského tela je teória nemeckého lekára H. Hufelanda (1762-1836), ktorý pripisoval dôležitosť pracovnej činnosti v dlhovekosti. Počuli sme jeho výrok, že staroby sa nedožil ani jeden lenivec. Ešte slávnejšie endokrinná teória starnutia, ktorý pochádza z pokusov uskutočnených ešte v polovici predminulého storočia Bertholdom (1849), ktorý ukázal, že transplantácia semenníkov z jedného zvieraťa na druhé je sprevádzaná rozvojom sekundárnych pohlavných znakov. Neskôr francúzsky fyziológ C. Brown-Séquard (1818-1894) na základe výsledkov injekčného podávania extraktov zo semenníkov tvrdil, že tieto injekcie majú priaznivý a omladzujúci účinok. Na začiatku 20. stor. Existuje už názor, že nástup staroby je spojený so zánikom činnosti žliaz s vnútornou sekréciou, najmä pohlavných žliaz. V 20.-30. XX storočia Na základe tohto presvedčenia sa v rôznych krajinách vykonalo mnoho operácií na omladenie starších alebo starých ľudí. Napríklad G. Steinach v Rakúsku podviazal mužom semenné povrazce, čo viedlo k zastaveniu vonkajšej sekrécie pohlavných žliaz a údajne k istému omladeniu. S.A. Voronov vo Francúzsku transplantoval semenníky z mladých zvierat na staré a z opíc na mužov a Tushnov v ZSSR omladzoval kohútov injekciou histolyzátov gonád. Všetky tieto operácie viedli k určitým účinkom, ale len dočasným. Po týchto dopadoch proces starnutia pokračoval, a to ešte intenzívnejšie.
Na začiatku 20. stor. vznikol mikrobiologická teória starnutia, ktorého tvorcom bol I.I. Mečnikov, ktorý rozlišoval medzi fyziologickou a patologickou starobou. Veril, že ľudská staroba je patologická, t.j. predčasné. Základ myšlienok I.I Mečnikovova doktrína ortobiózy (ortos -
správne, bios -života), podľa ktorého hlavnou príčinou starnutia je poškodenie nervových buniek intoxikačnými produktmi vznikajúcimi v dôsledku hniloby v hrubom čreve. Rozvíjanie doktríny normálneho životného štýlu (dodržiavanie hygienických pravidiel, pravidelná práca, abstinencia od zlých návykov), I.I. Mečnikov tiež navrhol spôsob, ako potlačiť hnilobné črevné baktérie konzumáciou fermentovaných mliečnych výrobkov.
V 30-tych rokoch XX storočia sa rozšírilo teória o úlohe centrálneho nervového systému (CNS) pri starnutí. Tvorcom tejto teórie je I.P. Pavlov, ktorý stanovil integračnú úlohu centrálneho nervového systému v normálnom fungovaní organizmov. Stúpenci I.P. Pavlovove pokusy na zvieratách ukázali, že predčasné starnutie je spôsobené nervovými šokmi a dlhotrvajúcim nervovým prepätím.
Stojí za zmienku teória zmien v spojivovom tkanive súvisiacich s vekom, formulované v 30-tych rokoch. XX storočia A.A. Bogomolets (1881-1946). Veril, že fyziologickú činnosť organizmu zabezpečuje spojivové tkanivo (kostné tkanivo, chrupavka, šľachy, väzy a vláknité spojivové tkanivo) a že zmeny v koloidnom stave buniek, strata ich turgoru atď. určujú zmeny súvisiace s vekom. v organizmoch. Moderné údaje poukazujú na dôležitosť akumulácie vápnika v spojivových tkanivách, pretože prispieva k strate jeho elasticity, ako aj k kôrnateniu krvných ciev.
Moderné prístupy k pochopeniu podstaty a mechanizmov starnutia sa vyznačujú širokým využívaním údajov z fyzikálno-chemickej biológie a najmä úspechov molekulárnej genetiky. Najbežnejšie moderné predstavy o mechanizme starnutia sa scvrkávajú na skutočnosť, že počas života sa v bunkách tela hromadia somatické mutácie, v dôsledku ktorých dochádza k syntéze defektných proteínov alebo neopravených krížových väzieb DNA s proteínmi. Keďže defektné proteíny hrajú dezintegračnú úlohu v bunkovom metabolizme, vedie to k starnutiu. V prípade kultivovaných fibroblastov sa ukázalo, že proteíny a mRNA spojené so starými bunkami potláčajú syntézu DNA v mladých fibroblastoch.
Známa je aj hypotéza, podľa ktorej sa starnutie považuje za dôsledok zmien mitochondriálnych metabolitov s následnou dysfunkciou enzýmov.
U ľudí bola preukázaná existencia génov, ktoré určujú načasovanie vývoja dedičných degeneratívnych procesov spojených so starnutím. Viacerí vedci sa domnievajú, že príčinou starnutia sú zmeny v imunitnom obrannom systéme tela, najmä autoimunitné reakcie na telesné štruktúry, ktoré sú životne dôležité. Napokon, pri vysvetľovaní mechanizmov starnutia odborníci venujú veľkú pozornosť poškodeniu bielkovín spojenému s tvorbou voľných radikálov. Napokon, význam sa niekedy pripisuje hydrolázam uvoľneným po rozpade lyzozómov, ktoré ničia bunky.
Komplexná teória starnutia však ešte nebola vytvorená, keďže je zrejmé, že žiadna z týchto teórií nedokáže samostatne vysvetliť mechanizmy starnutia.
Smrť.Smrť je posledným štádiom ontogenézy. Otázka smrti v biológii zaujíma osobitné miesto, pretože pocit smrti „... je v ľudskej prirodzenosti úplne inštinktívny a vždy bol jednou z najväčších starostí človeka“ (I.I. Mechnikov, 1913). Navyše, otázka smrti bola a je v centre pozornosti všetkých filozofických a náboženských náuk, hoci filozofia smrti bola v rôznych historických dobách prezentovaná rôzne. V starovekom svete Sokrates a Platón argumentovali za nesmrteľnosť duše, zatiaľ čo Aristoteles popieral Platónovu myšlienku nesmrteľnosti duše a veril v nesmrteľnosť ľudského ducha, ktorý naďalej žije po smrti človeka. Cicero a Seneca tiež uznávali budúci život, ale Marcus Aurelius považoval smrť za prirodzený jav, ktorý by sa mal akceptovať bez sťažností. V 18. storočí I. Kant a I. Fichte (1762-1814) tiež verili v budúci život a G. Hegel sa držal presvedčenia, že dušu pohlcuje „absolútne bytie“, hoci povaha tohto „bytia“ nebola odhalená. .
V súlade so všetkými známymi náboženskými učeniami pozemský život človeka pokračuje aj po jeho smrti a človek sa musí neúnavne pripravovať na túto budúcu smrť. Prírodovedci a filozofi, ktorí neuznávajú nesmrteľnosť, však verili a stále veria, že smrť je, ako opakovane zdôraznil I.I. Mečnikov, prirodzený výsledok života organizmu. Viac obrazná definícia smrti je, že „...je jasným víťazstvom nezmyselnosti nad zmyslom, chaosu nad priestorom“ (V. Solovjov, 1894).
Vedecké dôkazy naznačujú, že v jednobunkových organizmoch (rastlinách a zvieratách) treba rozlišovať smrť od zastavenia.
vedomosti o ich existencii. Smrť je ich zničením, zatiaľ čo zánik existencie je spojený s ich rozdelením. V dôsledku toho je krehkosť jednobunkových organizmov kompenzovaná ich rozmnožovaním. U mnohobunkových rastlín a živočíchov je smrť v plnom zmysle slova ukončením života organizmu.
U ľudí sa pravdepodobnosť úmrtia zvyšuje počas puberty. Najmä vo vyspelých krajinách sa po 28. roku života zvyšuje pravdepodobnosť úmrtia takmer exponenciálne.
Existuje klinická a biologická smrť človeka. Klinická smrť sa prejavuje stratou vedomia, zastavením srdcového tepu a dýchania, ale väčšina buniek a orgánov zostáva nažive. Dochádza k samoobnoveniu buniek a pokračuje črevná peristaltika. Klinická smrť „nedosiahne“ biologickú smrť, pretože je reverzibilná, pretože v stave klinickej smrti sa človek môže „vrátiť“ k životu. Napríklad psi sa „vracajú“ k životu po 5-6 minútach, ľudia - po 6-7 minútach od začiatku klinickej smrti. Biologická smrť je charakteristická tým, že je nezvratná. Zastavenie tlkotu srdca a dýchania sprevádza zastavenie samoobnovovacích procesov, smrť a rozklad buniek. Bunková smrť však nezačína vo všetkých orgánoch súčasne. Najprv odumiera mozgová kôra, potom odumierajú epitelové bunky čriev, pľúc, pečene, svalových buniek a srdca.
Opatrenia na resuscitáciu (oživenie) organizmov sú založené na myšlienke klinickej smrti, ktorá má v modernej medicíne mimoriadny význam.
DĹŽKA ŽIVOTA
Porovnanie údajov o strednej dĺžke života predstaviteľov flóry a fauny ukazuje, že medzi rastlinami a zvieratami žijú rôzne organizmy rôzne časy. Napríklad bylinné rastliny (divoké a pestované) žijú jednu sezónu. Naopak, ich dreviny sa vyznačujú dlhšou životnosťou. Napríklad čerešňa žije 100 rokov, smrek - 1000 rokov, dub -
2000 rokov, borovica - až 2000-4000 rokov.
Ryby mnohých druhov žijú 55-80 rokov, žaby - 16 rokov, krokodíly - 50-60 rokov, vtáky niektorých druhov - až 100 rokov. Životnosť cicavcov je kratšia.
Napríklad malé hospodárske zvieratá žijú 20-25 rokov, dobytok - 30 rokov alebo viac, kone - do 30 rokov, psy - 20 rokov alebo viac, vlci - 15 rokov, medvede - 50 rokov, slony - 100 rokov. Medzi cicavcami sa človek dožíva najdlhšie. Veľa ľudí sa dožívalo 115-120 rokov aj viac.
V súlade s existujúcimi myšlienkami je priemerná dĺžka života druhovo špecifický kvantitatívny znak, ktorý podlieha kontrole genotypom. Predpokladá sa, že geneticky podmienená dĺžka života koreluje s obdobiami ontogenézy (tabuľka 5).
Tabuľka 5.Závislosť hladiny fibroblastov počas kultivácie od veku darcu
Predpokladá sa, že dĺžka života druhu je evolučnou akvizíciou druhu. Pokiaľ ide o dlhovekosť jedincov, možno ju vysvetliť predpokladom prítomnosti kombinácií určitých génov v ich genotypoch, prípadne malým počtom alebo úplnou absenciou mutácií v ich bunkách.
A.A. Bogomolets a I.I. Schmalhausen vypočítal, že prirodzená dĺžka života človeka by mala byť 120-150 rokov. Tohto veku sa však dožije len málokto. Preto sa skutočná dĺžka života nezhoduje s prirodzenou dĺžkou života.
Priemernú dĺžku života ovplyvňuje množstvo faktorov (pokles dojčenskej úmrtnosti, účinnosť kontroly infekcií, pokroky v chirurgii, zlepšenie výživy a všeobecných životných podmienok). Hlavnými príčinami poklesu strednej dĺžky života sú úmrtnosť na hlad, choroby a nedostatočná lekárska starostlivosť. V Rusku bola v posledných rokoch zaznamenaná negatívna populácia vo všetkých ruských mestách a vidieckych oblastiach v dôsledku poklesu pôrodnosti a vysokej úmrtnosti.
Z lekárskeho hľadiska je priemerná dĺžka života ukazovateľom zdravia národa. ZSSR sa umiestnil na prvom mieste na svete z hľadiska počtu starých ľudí. Napríklad na 1 milión obyvateľov pripadá 104 ľudí starších ako 90 rokov, kým v Anglicku - 6, vo Francúzsku - 7
a USA - 15 ľudí. V súčasnosti dochádza k zmene hraníc práceschopného obyvateľstva v porovnaní napríklad s 30-tymi rokmi. XX storočia Rozdiel medzi vekom odchodu do dôchodku a úrovňou aktivity ľudí sa tiež líši. V roku 1982 sa vo Viedni konalo Svetové zhromaždenie o problémoch starnutia svetovej populácie, na ktorom boli formulované prognózy k tejto problematike do roku 2025. Predpokladá sa, že počet ľudí vo veku 60 rokov a starších v porovnaní s rokom 1950 sa bude zvyšovať 5-krát, ľudia nad 80 rokov - 7-krát. Podľa tohto medzinárodného fóra existujú rozdiely v rýchlosti starnutia populácie medzi rôznymi národmi, krajinami a regiónmi.
Geriatria je veda, ktorej úlohou je vyvinúť spôsoby, ako normalizovať meniace sa funkcie starnúceho organizmu. Moderná medicína však zatiaľ nemá metódy a prostriedky na ovplyvnenie normálnych fyziologických procesov. Preto sa úloha geriatrie obmedzuje na liečbu ochorení, ktoré vznikajú v starobe a elimináciu (ak je to možné) rizikových faktorov, ktoré spôsobujú predčasné starnutie.
OTÁZKY NA DISKUSII
1. Čo rozumiete pod pohlavným rozmnožovaním organizmov a aká je jeho biologická úloha?
2. Opíšte nepohlavné rozmnožovanie a pomenujte jeho formy.
3. Opíšte znaky pohlavného procesu u jednobunkových a mnohobunkových organizmov.
4. Čo je gametogenéza?
5. Akú funkciu majú jednotlivé typy gamét?
6. Aké štádiá vývoja gamét poznáte?
7. Aké sú podobnosti a rozdiely medzi spermatogenézou a oogenézou?
8. Čo je meióza a aký je jej biologický význam?
9. Opíšte fázy meiózy.
10. Ovplyvňuje crossing over výsledky meiózy a ako?
11. Popíšte podstatu oplodnenia.
12. Aký je rozdiel medzi zygogenézou a partenogenézou?
13. Aká je biologická úloha striedania haploidie a diploidie?
14. Čo je sexuálny dimorfizmus? Čo rozumiete pod pojmom hermafroditizmus? Pozorujú sa prípady hermafroditizmu u ľudí a ako často?
15. Ako si predstavujete evolúciu metód rozmnožovania?
16. Čo rozumiete pod pojmom rast a vývoj organizmov? Aký je vzťah medzi bunkovým rastom a diferenciáciou?
17. Aké sú molekulárne základy diferenciácie buniek? Čo viete o kmeňových bunkách?
18. Formulujte pojem ontogenéza a pomenujte obdobia ontogenézy.
19. Aké sú rozdiely medzi priamym a nepriamym rozvojom?
20. Aký vplyv má oplodnenie na vajíčka?
21. V akom štádiu implementácie genetickej informácie sa uskutočňuje kontrola pôsobenia génov?
22. Ako sa z oplodneného vajíčka vyvinie mnohobunková štruktúra?
23. Ako sa vyvíjajúce sa bunky a tkanivá počas vývoja navzájom líšia?
24. Dajú sa stratené alebo poškodené časti tela obnoviť?
25. Aká je súvislosť v ontogenéze medzi starobou a strednou dĺžkou života?
26. Formulujte a určte rozdiel medzi prirodzenou (pravdepodobnou) a skutočnou dĺžkou života.
27. Aké teórie starnutia organizmov poznáte?
29. Aké faktory ovplyvňujú dĺžku života?
30. Na čom je založené klonovanie organizmov? Uveďte príklady klonovania organizmov.
31. Preniká spermia alebo jej obsah do bunky?
32. Čo naznačujú údaje o klonovaní?
33. Preformationizmus a epigenéza v dejinách biológie. Majú vedecký význam?
34. Aké sú rozdiely medzi klinickou a biologickou smrťou?
Reprodukcia je schopnosť organizmov vytvárať svoj vlastný druh. Reprodukcia je jednou z najdôležitejších vlastností života a je možná vďaka všeobecnej schopnosti organizmov produkovať potomstvo. Bezprostrední potomkovia však nie sú vždy podobní svojim rodičom. Napríklad z výtrusov papradí vyrastá početné potomstvo, reprezentované výhonkami, ktoré nie sú podobné materskej výtrusnej rastline. Na výrastku sa zase objaví rastlina na rozdiel od nej - sporofyt. Tento jav sa nazýva striedanie generácií.
Špeciálnou formou pohlavného rozmnožovania je partenogenéza alebo panenské rozmnožovanie, vývoj organizmu z neoplodneného vajíčka. Táto forma rozmnožovania je charakteristická najmä pre druhy s krátkym životným cyklom s výraznými sezónnymi zmenami. Partenogenéza môže byť haploidná alebo diploidná. Pri haploidnej (generatívnej) partenogenéze sa z haploidného vajíčka vyvinie nový organizmus. Výsledné jedince môžu byť iba muži, iba ženy alebo oboje. Závisí to od chromozomálneho určenia pohlavia.
Individuálny vývoj organizmu alebo ontogenéza je súbor postupných morfologických, fyziologických a biochemických premien, ktorými organizmus prechádza od okamihu svojho vzniku až po smrť. Počas ontogenézy dochádza k implementácii dedičných informácií, ktoré telo dostáva od svojich rodičov. V ontogenéze existujú dve hlavné obdobia – embryonálne a postembryonálne. V embryonálnom štádiu sa u zvierat vytvára embryo, v ktorom sa tvoria hlavné orgánové systémy. V postembryonálnom období sa dokončujú vývinové procesy, nastáva puberta, rozmnožovanie, starnutie a smrť.
