5. Sääntelytoiminto. Proteiinit suorittavat signalointiaineiden tehtäviä - jotkin hormonit, histohormonit ja välittäjäaineet ovat minkä tahansa rakenteen signaaliaineiden reseptoreita ja varmistavat signaalin edelleen siirtymisen solun biokemiallisissa signaaliketjuissa. Esimerkkejä ovat kasvuhormoni somatotropiini, hormoni insuliini, H- ja M-kolinergiset reseptorit.
6. Moottorin toiminta. Proteiinien avulla suoritetaan supistumisprosessit ja muut biologiset liikkeet. Esimerkkejä ovat tubuliini, aktiini ja myosiini.
7. Varatoiminto. Kasvit sisältävät varaproteiineja, jotka ovat arvokkaita ravintoaineita eläinkehoissa, lihasproteiinit toimivat vararavintoaineina, jotka mobilisoidaan, kun se on ehdottoman välttämätöntä.
Proteiineille on ominaista useiden rakenteellisten organisaatiotasojen läsnäolo.
Ensisijainen rakenne Proteiini on aminohappotähteiden sekvenssi polypeptidiketjussa. Peptidisidos on karboksamidisidos yhden aminohapon a-karboksyyliryhmän ja toisen aminohapon a-aminoryhmän välillä.
alanyylifenyylialanyylikysteyyliproliini
U p eptisidos ominaisuuksia on useita:
a) se on resonanssistabiloitu ja siksi sijaitsee käytännössä samassa tasossa - tasossa; pyöriminen C-N-sidoksen ympäri vaatii paljon energiaa ja on vaikeaa;
b) -CO-NH-sidoksella on erityinen luonne, se on pienempi kuin tavallinen, mutta suurempi kuin kaksinkertainen, eli siinä on keto-enolitautomerismi:

c) substituentit peptidisidoksen suhteen ovat sisällä transsi- asema;
d) peptidirunkoa ympäröivät eri luonteiset sivuketjut, jotka ovat vuorovaikutuksessa ympäröivien liuotinmolekyylien kanssa, vapaat karboksyyli- ja aminoryhmät ionisoituvat muodostaen proteiinimolekyylin kationisia ja anionisia keskuksia. Niiden suhteesta riippuen proteiinimolekyyli saa positiivisen tai negatiivisen kokonaisvarauksen, ja sille on tunnusomaista myös yksi tai toinen väliaineen pH-arvo saavuttaessaan proteiinin isoelektrisen pisteen. Radikaalit muodostavat suola-, eetteri- ja disulfidisiltoja proteiinimolekyylin sisällä ja määrittävät myös proteiineille ominaisen reaktioiden alueen.
Tällä hetkellä suostuivat pitämään proteiineina polymeerejä, jotka koostuvat 100 tai useammasta aminohappotähteestä, polypeptidejä - polymeerejä, jotka koostuvat 50-100 aminohappotähteestä, pienimolekyylipainoisia peptidejä - polymeerejä, jotka koostuvat alle 50 aminohappotähteestä.
Jonkin verran alhainen molekyylipaino peptideillä on itsenäinen biologinen rooli. Esimerkkejä joistakin näistä peptideistä:
Glutationi - γ-glu-cis-gly - yksi yksi yleisimmistä solunsisäisistä peptideistä, osallistuu solujen redox-prosesseihin ja aminohappojen siirtoon biologisten kalvojen läpi.
karnosiini - β-ala-his -peptidi, Eläinten lihaksissa oleva, eliminoi lipidiperoksidin hajoamistuotteet, nopeuttaa hiilihydraattien hajoamisprosessia lihaksissa ja osallistuu lihasten energia-aineenvaihduntaan fosfaatin muodossa.
Vasopressiini on aivolisäkkeen takalohkon hormoni, joka osallistuu veden aineenvaihdunnan säätelyyn kehossa:

Phalloidiini- myrkyllinen kärpäsherukkapolypeptidi, aiheuttaa vähäisinä pitoisuuksina kehon kuoleman entsyymien ja kalium-ionien vapautuessa soluista:

Gramisidiini - antibiootti, joka vaikuttaa moniin grampositiivisiin bakteereihin, muuttaa biologisten kalvojen läpäisevyyttä pienimolekyylisille yhdisteille ja aiheuttaa solukuoleman:

Meth-enkefaliini - tyr-gly-gly-phen-met - peptidi, joka syntetisoituu hermosoluissa ja vähentää kipua.
Proteiinin sekundaarinen rakenne on spatiaalinen rakenne, joka muodostuu peptidirungon funktionaalisten ryhmien välisten vuorovaikutusten tuloksena.
Peptidiketju sisältää monet peptidisidosten CO- ja NH-ryhmät, joista jokainen voi mahdollisesti osallistua vetysidosten muodostukseen. On olemassa kaksi päätyyppiä rakenteita, jotka mahdollistavat tämän tapahtumisen: α-kierre, jossa ketju on kierretty puhelinjohdon tavoin, ja taitettu β-rakenne, jossa yhden tai useamman ketjun pitkänomaiset osat asetetaan vierekkäin. puolella. Molemmat rakenteet ovat erittäin vakaita.
α-heliksille on ominaista kierretyn polypeptidiketjun erittäin tiheässä pakkauksessa oikeakätisen kierteen jokaista kierrosta kohden on 3,6 aminohappotähdettä, joiden radikaalit on aina suunnattu ulospäin ja hieman taaksepäin, eli polypeptidiketjun alkuun.
α-heliksin tärkeimmät ominaisuudet:
1) a-heliksi stabiloidaan vetysidoksilla peptidiryhmän typessä olevan vetyatomin ja tähteen karbonyylihapen välillä, jotka sijaitsevat neljässä asemassa ketjua pitkin;
2) kaikki peptidiryhmät osallistuvat vetysidoksen muodostumiseen, mikä varmistaa a-heliksin maksimaalisen stabiilisuuden;
3) kaikki peptidiryhmien typpi- ja happiatomit ovat mukana vetysidosten muodostumisessa, mikä vähentää merkittävästi a-kierteisten alueiden hydrofiilisyyttä ja lisää niiden hydrofobisuutta;
4) a-heliksi muodostuu spontaanisti ja on polypeptidiketjun stabiilin konformaatio, joka vastaa vähimmäisvapaata energiaa;
5) L-aminohappojen polypeptidiketjussa oikeakätinen heliksi, jota yleensä esiintyy proteiineissa, on paljon vakaampi kuin vasenkätinen.
α-heliksin muodostumisen mahdollisuus määräytyy proteiinin primäärirakenteen mukaan. Jotkut aminohapot estävät peptidirunkoa kiertymästä. Esimerkiksi glutamaatin ja aspartaatin vierekkäiset karboksyyliryhmät hylkivät toisiaan, mikä estää vetysidosten muodostumisen a-heliksiin. Samasta syystä ketjun helikalisaatio on vaikeaa paikoissa, joissa positiivisesti varautuneet lysiini- ja arginiinitähteet sijaitsevat lähellä toisiaan. Proliinilla on kuitenkin suurin rooli α-heliksin hajoamisessa. Ensinnäkin proliinissa typpiatomi on osa jäykkää rengasta, joka estää pyörimisen N-C-sidoksen ympäri, ja toiseksi proliini ei muodosta vetysidosta, koska typpiatomissa ei ole vetyä.
β-levy on kerrosrakenne muodostuu vetysidoksista lineaarisesti järjestettyjen peptidifragmenttien välillä. Molemmat ketjut voivat olla itsenäisiä tai kuulua samaan polypeptidimolekyyliin. Jos ketjut ovat samaan suuntaan, niin tällaista β-rakennetta kutsutaan rinnakkaiseksi. Vastakkaisten ketjusuuntien tapauksessa, eli kun yhden ketjun N-pää osuu yhteen toisen ketjun C-pään kanssa, β-rakennetta kutsutaan antirinnakkaiseksi. Antirinnakkaiset β-levyt, joissa on lähes lineaariset vetysillat, ovat energeettisesti edullisempia.


yhdensuuntainen β-levy antirinnakkais β-levy
Toisin kuin α-heliksi vetysidoksilla kyllästettynä jokainen β-levyketjun osa on avoin lisävetysidosten muodostumiselle. Aminohappojen sivuradikaalit on suunnattu lähes kohtisuoraan arkin tasoon nähden, vuorotellen ylös ja alas.
Niillä alueilla, joilla peptidiketju taipuu melko jyrkästi sisältäen usein β-silmukan. Tämä on lyhyt fragmentti, jossa 4 aminohappotähdettä on taivutettu 180° ja stabiloitu yhdellä vetysillalla ensimmäisen ja neljännen tähteen välillä. Suuret aminohapporadikaalit häiritsevät β-silmukan muodostumista, joten se sisältää useimmiten pienimmän aminohapon, glysiinin.
Proteiinin suprasekundaarinen rakenne- tämä on tietty sekundäärirakenteiden vuorottelujärjestys. Domeeni ymmärretään proteiinimolekyylin erilliseksi osaksi, jolla on tietty rakenteellinen ja toiminnallinen autonomia. Domaineja pidetään nykyään proteiinimolekyylien rakenteen peruselementteinä, ja α-heliksien ja β-levyjen järjestelyn suhde ja luonne mahdollistaa enemmän proteiinimolekyylien kehityksen ja fylogeneettisten suhteiden ymmärtämisen kuin primäärirakenteiden vertailun.
Evoluution päätehtävä on Suunnittelemalla yhä enemmän uusia proteiineja. On äärettömän pieni mahdollisuus syntetisoida vahingossa aminohapposekvenssi, joka täyttäisi pakkausehdot ja varmistaisi toiminnallisten tehtävien suorittamisen. Siksi on yleistä löytää proteiineja, joilla on erilaisia toimintoja, mutta rakenteeltaan niin samanlaisia, että niillä näyttää olleen yhteinen esi-isä tai ne ovat kehittyneet toisistaan. Näyttää siltä, että kun evoluutio kohtaa tarpeen ratkaista tietty ongelma, se ei mieluummin suunnittele proteiineja tähän tarkoitukseen alusta alkaen, vaan mukauttaa jo vakiintuneita rakenteita tähän tarkoitukseen ja mukauttaa niitä uusiin tarkoituksiin.
Esimerkkejä usein toistuvista suprasekundaarisista rakenteista:
1) αα’ - proteiinit, jotka sisältävät vain α-heliksiä (myoglobiini, hemoglobiini);
2) ββ’ - proteiinit, jotka sisältävät vain β-rakenteita (immunoglobuliinit, superoksididismutaasi);
3) βαβ’ - β-tynnyrirakenne, jokainen β-kerros sijaitsee tynnyrin sisällä ja on yhdistetty molekyylin pinnalla sijaitsevaan a-heliksiin (trioosifosfoisomeraasi, laktaattidehydrogenaasi);
4) "sinkkisormi" - proteiinifragmentti, joka koostuu 20 aminohappotähteestä, sinkkiatomi on kytketty kahteen kysteiinitähteeseen ja kahteen histidiinitähteeseen, jolloin muodostuu noin 12 aminohappotähteen "sormi", joka voi sitoutua DNA-molekyylin säätelyalueisiin;
5) "leusiinivetoketju" - vuorovaikutuksessa olevilla proteiineilla on α-kierteinen alue, joka sisältää vähintään 4 leusiinitähdettä, ne sijaitsevat 6 aminohapon päässä toisistaan, eli ne ovat joka toisen kierroksen pinnalla ja voivat muodostaa hydrofobisia sidoksia leusiinitähteiden kanssa toinen proteiini. Esimerkiksi leusiinivetoketjujen avulla voidaan kompleksoida vahvasti emäksisten histoniproteiinien molekyylejä positiivisen varauksen voittamiseksi.
Proteiinin tertiäärinen rakenne- tämä on proteiinimolekyylin avaruudellinen järjestely, jota stabiloivat aminohappojen sivuradikaalien väliset sidokset.
Sidostyypit, jotka stabiloivat proteiinin tertiaarista rakennetta:

sähköstaattinen vety hydrofobinen disulfidi vuorovaikutus sidokset vuorovaikutus sidokset
Taitosta riippuen Proteiinien tertiäärinen rakenne voidaan luokitella kahteen päätyyppiin - fibrillaariseen ja pallomaiseen.
Fibrillaariset proteiinit- pitkiä, lankamaisia, veteen liukenemattomia molekyylejä, joiden polypeptidiketjut ovat venyneet yhtä akselia pitkin. Nämä ovat pääasiassa rakenteellisia ja supistuvia proteiineja. Esimerkkejä yleisimmistä fibrillaarisista proteiineista:
1. α- Keratiinit. Epidermaalisolujen syntetisoima. Ne muodostavat lähes kaiken hiusten, turkisten, höyhenten, sarvien, kynsien, kynsien, kynsien, suomujen, kavioiden ja kilpikonnankuoren kuivapainon sekä merkittävän osan ihon ulkokerroksen painosta. Tämä on koko proteiiniperhe, ne ovat samankaltaisia aminohappokoostumuksessa, sisältävät monia kysteiinijäännöksiä ja niillä on sama polypeptidiketjujen avaruudellinen järjestely.
Hiussoluissa keratiinin polypeptidiketjut ensin organisoituu kuiduiksi, joista sitten muodostetaan rakenteita kuten köysi tai kierretty kaapeli, jotka lopulta täyttävät koko solun tilan. Karvasolut litistyvät ja lopulta kuolevat, ja soluseinämät muodostavat putkimaisen tupen, jota kutsutaan kynsinauhoksi jokaisen hiuksen ympärille. α-keratiinissa polypeptidiketjut ovat a-heliksin muotoisia, kierrettyinä toistensa ympärille kolmiytimiksi kaapeliksi muodostaen ristikkäisiä disulfidisidoksia.
N-pään tähteet sijaitsevat toisella puolella (rinnakkais). Keratiinit ovat veteen liukenemattomia, koska niiden koostumuksessa on vallitsevia aminohappoja ja ei-polaarisia sivuradikaaleja, jotka ovat vesifaasia päin. Permin aikana tapahtuu seuraavia prosesseja: ensin disulfidisillat tuhoutuvat pelkistämällä tioleilla, ja sitten, kun hiuksille on annettu tarvittava muoto, se kuivataan kuumentamalla, kun taas ilmakehän hapella tapahtuvan hapettumisen vuoksi muodostuu uusia disulfidisiltoja. , jotka säilyttävät kampauksen muodon.
2. β-keratiinit. Näitä ovat silkki- ja hämähäkinverkkofibroiini. Ne ovat antiparalleelisia β-laskostettuja kerroksia, joiden koostumuksessa on hallitseva osa glysiinistä, alaniinista ja seriinistä.
3. Kollageeni. Yleisin proteiini korkeammissa eläimissä ja sidekudosten tärkein fibrillaarinen proteiini. Kollageeni syntetisoituu fibroblasteissa ja kondrosyyteissä - erikoistuneissa sidekudossoluissa, joista se sitten poistuu. Kollageenikuituja löytyy ihosta, jänteistä, rustosta ja luista. Ne eivät veny, ovat vahvempia kuin teräslanka, ja kollageenisäikeille on ominaista poikittaisjuovaisuus.
Vedessä keitettynä kuitumainen, liukenematon ja sulamaton kollageeni muuttuu gelatiiniksi joidenkin kovalenttisten sidosten hydrolyysillä. Kollageeni sisältää 35 % glysiiniä, 11 % alaniinia, 21 % proliinia ja 4-hydroksiproliinia (kollageenille ja elastiinille ainutlaatuinen aminohappo). Tämä koostumus määrittää gelatiinin suhteellisen alhaisen ravintoarvon elintarvikeproteiinina. Kollageenifibrillit koostuvat toistuvista polypeptidialayksiköistä, joita kutsutaan tropokollageeniksi. Nämä alayksiköt on järjestetty fibrilliä pitkin yhdensuuntaisten nippujen muodossa päästä häntään -tavalla. Pään siirtyminen antaa tyypilliset poikittaisjuovat. Tämän rakenteen ontelot voivat tarvittaessa toimia paikkana hydroksiapatiitti Ca 5 (OH) (PO 4) 3 -kiteiden laskeutumiselle, jolla on tärkeä rooli luun mineralisaatiossa.


Tropokollageenin alayksiköt koostuvat kolmesta polypeptidiketjusta, jotka on kietoutunut tiukasti kolmijuosteiseksi köydeksi, erottuen a- ja β-keratiineista. Joissakin kollageeneissa kaikilla kolmella ketjulla on sama aminohapposekvenssi, kun taas toisissa vain kaksi ketjua ovat identtisiä ja kolmas on erilainen. Tropokollageenin polypeptidiketju muodostaa vasenkätisen kierteen, jossa on vain kolme aminohappotähdettä per kierros johtuen proliinin ja hydroksiproliinin aiheuttamista ketjun mutkista. Nämä kolme ketjua on liitetty toisiinsa vetysidosten lisäksi kovalenttisella sidoksella, joka muodostuu kahden vierekkäisissä ketjuissa sijaitsevan lysiinitähteen väliin:

Kun vanhenemme, tropokollageenialayksiköihin ja niiden väliin muodostuu yhä enemmän ristisidoksia, mikä tekee kollageenisäikeistä jäykempiä ja hauraampia, mikä muuttaa ruston ja jänteiden mekaanisia ominaisuuksia, tekee luista hauraampia ja vähentää sarveiskalvon läpinäkyvyyttä.
4. Elastiini. Sisältyy nivelsiteiden keltaiseen elastiseen kudokseen ja suurten valtimoiden seinämien elastiseen sidekudoskerrokseen. Elastiinifibrillien pääalayksikkö on tropoelastiini. Elastiini sisältää runsaasti glysiiniä ja alaniinia, sisältää paljon lysiiniä ja vähän proliinia. Elastiinin spiraaliosat venyvät jännityksen yhteydessä, mutta palautuvat alkuperäiseen pituuteensa, kun kuormitus poistetaan. Neljän eri ketjun lysiinitähteet muodostavat kovalenttisia sidoksia keskenään ja antavat elastiinin venyä palautuvasti kaikkiin suuntiin.
Globaalit proteiinit- proteiinit, joiden polypeptidiketju on laskostunut tiiviiksi palloksi, pystyvät suorittamaan monenlaisia toimintoja.
Globulaaristen proteiinien tertiäärinen rakenne On kätevintä käyttää esimerkkiä myoglobiinista. Myoglobiini on suhteellisen pieni happea sitova proteiini, jota löytyy lihassoluista. Se varastoi sitoutuneen hapen ja edistää sen siirtymistä mitokondrioihin. Myoglobiinimolekyyli sisältää yhden polypeptidiketjun ja yhden hemoryhmän (hemi) - protoporfyriinin ja raudan kompleksin.
Perusominaisuudet myoglobiini:
a) myoglobiinimolekyyli on niin kompakti, että sen sisään mahtuu vain 4 vesimolekyyliä;
b) kaikki polaariset aminohappotähteet kahta lukuun ottamatta sijaitsevat molekyylin ulkopinnalla, ja ne kaikki ovat hydratoituneessa tilassa;
c) suurin osa hydrofobisista aminohappotähteistä sijaitsee myoglobiinimolekyylin sisällä ja on siten suojattu kosketukselta veden kanssa;
d) jokainen myoglobiinimolekyylin neljästä proliinitähteestä sijaitsee polypeptidiketjun taivutuskohdassa, seriini-, treoniini- ja asparagiinitähteet sijaitsevat muissa taivutuskohdissa, koska tällaiset aminohapot estävät a-heliksin muodostumisen; sijaitsevat vierekkäin;
e) litteä hemiryhmä sijaitsee ontelossa (taskussa) lähellä molekyylin pintaa, rautaatomissa on kaksi hemitasoon nähden kohtisuoraan suunnattua koordinaatiosidosta, joista toinen on kytketty histidiinitähteeseen 93 ja toinen sitoutuu happimolekyyli.
Alkaen proteiinin tertiaarisesta rakenteesta kykenee suorittamaan luontaisia biologisia tehtäviä. Proteiinien toiminnan perusta on, että kun proteiinin pinnalle asetetaan tertiäärinen rakenne, muodostuu alueita, jotka voivat kiinnittää muita molekyylejä, joita kutsutaan ligandeiksi. Proteiinin ja ligandin vuorovaikutuksen korkea spesifisyys varmistaa aktiivisen keskuksen rakenteen komplementaarisuus ligandin rakenteen kanssa. Komplementaarisuus on vuorovaikutuksessa olevien pintojen avaruudellista ja kemiallista vastaavuutta. Useimmille proteiineille tertiäärinen rakenne on laskostumisen maksimitaso.
Kvaternäärinen proteiinirakenne- ominaisuus proteiineille, jotka koostuvat kahdesta tai useammasta polypeptidiketjusta, jotka on liitetty toisiinsa yksinomaan ei-kovalenttisilla, pääasiassa sähköstaattisilla ja vetysidoksilla. Useimmiten proteiinit sisältävät kaksi tai neljä alayksikköä, yleensä enemmän kuin neljä alayksikköä sisältävät säätelyproteiineja.
Kvaternäärisen rakenteen omaavat proteiinit, kutsutaan usein oligomeerisiksi. On homomeerisia ja heteromeerisiä proteiineja. Homomeerisiin proteiineihin kuuluvat proteiinit, joissa kaikilla alayksiköillä on sama rakenne, esimerkiksi katalaasientsyymi koostuu neljästä täysin identtisestä alayksiköstä. Heteromeerisissa proteiineissa on erilaisia alayksiköitä, esimerkiksi RNA-polymeraasientsyymi koostuu viidestä rakenteellisesti erilaisesta alayksiköstä, jotka suorittavat erilaisia tehtäviä.
Yhden alayksikön vuorovaikutus spesifinen ligandi aiheuttaa konformaatiomuutoksia koko oligomeerisessä proteiinissa ja muuttaa muiden alayksiköiden affiniteettia ligandeihin, tämä ominaisuus on oligomeeristen proteiinien allosteerisen säätelyn kyvyn taustalla.
Proteiinin kvaternaarista rakennetta voidaan tutkia hemoglobiinin esimerkkiä käyttäen. Sisältää neljä polypeptidiketjua ja neljä hemiproteesiryhmää, joissa rautaatomit ovat rautamuodossa Fe 2+. Molekyylin proteiiniosa - globiini - koostuu kahdesta α-ketjusta ja kahdesta β-ketjusta, jotka sisältävät jopa 70 % α-heliksiä. Jokaisella neljästä ketjusta on ominaista tertiäärinen rakenne, ja jokaiseen ketjuun liittyy yksi hemoryhmä. Eri ketjujen heemit sijaitsevat suhteellisen kaukana toisistaan ja niillä on erilaiset kaltevuuskulmat. Kahden a-ketjun ja kahden p-ketjun välille muodostuu vähän suoria kontakteja, kun taas a- ja p-ketjujen väliin muodostuu lukuisia hydrofobisten radikaalien muodostamia α1β1- ja α2β2-tyyppisiä kontakteja. α 1 β 1 ja α 2 β 2 väliin jää kanava.
Toisin kuin myoglobiini hemoglobiini luonnehdittu huomattavasti alhaisempi affiniteetti happea kohtaan, mikä mahdollistaa sen, kudoksissa olevilla alhaisilla hapen osapaineilla, antaa niille merkittävän osan sitoutuneesta hapesta. Hemoglobiinirauta sitoo happea helpommin korkeammilla pH-arvoilla ja alhaisilla CO 2 -pitoisuuksilla, jotka ovat tyypillisiä keuhkojen alveoleille; hapen vapautumista hemoglobiinista edistävät alhaisemmat pH-arvot ja kudoksille ominaiset korkeat CO 2 -pitoisuudet.
Hemoglobiini kuljettaa hapen lisäksi vetyioneja, jotka sitoutuvat ketjuissa oleviin histidiinitähteisiin. Hemoglobiini sisältää myös hiilidioksidia, joka kiinnittyy kunkin neljän polypeptidiketjun terminaaliseen aminoryhmään, mikä johtaa karbaminohemoglobiinin muodostumiseen:

SISÄÄN punasolut melko korkeissa pitoisuuksissa aine 2,3-difosfoglyseraatti (DPG) on läsnä, sen pitoisuus kasvaa noustessa korkeuksiin ja hypoksian aikana, mikä helpottaa hapen vapautumista hemoglobiinista kudoksissa. DPG sijaitsee kanavassa α1β1:n ja α2β2:n välillä ja on vuorovaikutuksessa positiivisesti kontaminoituneiden p-ketjuryhmien kanssa. Kun hemoglobiini sitoo happea, DPG pakotetaan ulos ontelosta. Joidenkin lintujen punasolut eivät sisällä DPG:tä, vaan inositoliheksafosfaattia, mikä edelleen vähentää hemoglobiinin happiaffiniteettia.
 2,3-difosfoglyseraatti (DPG)
2,3-difosfoglyseraatti (DPG)
HbA - normaali aikuisen hemoglobiini, HbF - sikiön hemoglobiini, sillä on suurempi affiniteetti O 2:een, HbS - hemoglobiiniin sirppisoluanemiassa. Sirppisoluanemia on vakava perinnöllinen sairaus, joka johtuu hemoglobiinin geneettisestä poikkeavuudesta. Sairaiden ihmisten veressä on epätavallisen paljon ohuita sirpin muotoisia punasoluja, jotka ensinnäkin repeytyvät helposti ja toiseksi tukkivat veren kapillaareja.
Molekyylitasolla hemoglobiini S on erilainen hemoglobiini A:sta on yksi aminohappotähde β-ketjun kohdassa 6, jossa glutamiinihappotähteen sijaan on valiini. Siten hemoglobiini S sisältää kaksi vähemmän negatiivista varausta, jolloin molekyylin pinnalle muodostuu "tahmea" hydrofobinen kontakti, minkä seurauksena deoksihemoglobiini S -molekyylit tarttuvat yhteen ja muodostavat liukenemattomia, epänormaalin pitkiä; lankamaisia aggregaatteja, jotka johtavat punasolujen muodonmuutokseen.
Ei ole mitään syytä ajatella, että proteiinin rakenteellisen organisoinnin tasojen muodostumiselle primaaritason yläpuolella on riippumaton geneettinen valvonta, koska primäärirakenne määrää sekundaarisen, tertiaarisen ja kvaternaarisen (jos sellainen on). Proteiinin natiivi konformaatio on termodynaamisesti stabiilin rakenne tietyissä olosuhteissa.
LUENTO 6
Proteiinilla on fysikaalisia, kemiallisia ja biologisia ominaisuuksia.
Proteiinien fysikaaliset ominaisuudet ovat molekyylipainon läsnäolo, kahtaistaitteisuus (muutos proteiiniliuoksen optisissa ominaisuuksissa liikkeessä verrattuna liuokseen levossa), proteiinien ei-pallomaisesta muodosta, liikkuvuus sähkökentässä, proteiinimolekyylien varauksesta johtuen . Lisäksi proteiineille on tunnusomaista optiset ominaisuudet, jotka koostuvat kyvystä kiertää valon polarisaatiotasoa, sirotella valonsäteitä proteiinihiukkasten suuren koon vuoksi ja absorboida ultraviolettisäteitä.
Yksi tyypillisistä fysikaalisista ominaisuuksista proteiinit ovat kyky adsorboitua pintaan ja joskus vangita molekyylejä, pienimolekyylisiä orgaanisia yhdisteitä ja ioneja sisällä.
Proteiinien kemialliset ominaisuudet vaihtelevat poikkeuksellinen monimuotoisuus, koska proteiineille ovat ominaisia kaikki aminohapporadikaalien reaktiot ja niille on ominaista peptidisidosten hydrolyysireaktio.
Siinä on huomattava määrä happamia ja emäksisiä ryhmiä, proteiineilla on amfoteerisia ominaisuuksia. Toisin kuin vapaat aminohapot, proteiinien happo-emäsominaisuudet eivät määräydy peptidisidosten muodostukseen osallistuvien α-amino- ja α-karboksiryhmien, vaan aminohappotähteiden varautuneiden radikaalien perusteella. Proteiinien pääominaisuudet määrittävät arginiini-, lysiini- ja histidiinitähteet. Happamat ominaisuudet johtuvat asparagiini- ja glutamiinihappojäämistä.
Proteiinititrauskäyrät ovat riittävät vaikea tulkita, koska missä tahansa proteiinissa on liikaa titrattavia ryhmiä, proteiinin ionisoituneiden ryhmien välillä on sähköstaattisia vuorovaikutuksia ja kunkin titrattavan ryhmän pK:aan vaikuttavat lähellä olevat hydrofobiset tähteet ja vetysidokset. Suurin käytännön sovellus on proteiinin isoelektrinen piste - pH-arvo, jossa proteiinin kokonaisvaraus on nolla. Isoelektrisessä pisteessä proteiini on mahdollisimman inertti, ei liiku sähkökentässä ja sillä on ohuin hydraatiokuori.
Proteiineilla on puskuroivia ominaisuuksia, mutta niiden puskurikapasiteetti on merkityksetön. Poikkeuksen muodostavat proteiinit, jotka sisältävät suuren määrän histidiinitähteitä. Esimerkiksi punasoluissa olevalla hemoglobiinilla on histidiinijäämien erittäin korkean pitoisuuden vuoksi merkittävä puskurikapasiteetti pH:ssa noin 7, mikä on erittäin tärkeää punasolujen roolin kannalta hapen ja hiilidioksidin kuljettamisessa veri.
Proteiineille on ominaista liukoisuus veteen, ja fysikaalisesta näkökulmasta katsottuna ne muodostavat todellisia molekyyliliuoksia. Proteiiniliuoksille on kuitenkin ominaista joitain kolloidisia ominaisuuksia: Tendahl-ilmiö (valonsirontailmiö), kyvyttömyys läpäistä puoliläpäiseviä kalvoja, korkea viskositeetti ja geelien muodostuminen.
Proteiinien liukoisuus riippuu suuresti suolojen konsentraatiosta eli liuoksen ionivahuudesta. Tislattuun veteen proteiinit ovat useimmiten huonosti liukenevia, mutta niiden liukoisuus kasvaa ionivahvuuden kasvaessa. Samalla lisääntyvä määrä hydratoituneita epäorgaanisia ioneja sitoutuu proteiinin pintaan ja siten sen aggregaatioaste laskee. Suurella ionivahvuudella suola-ionit ottavat pois hydraatiokuoren proteiinimolekyyleistä, mikä johtaa proteiinien aggregoitumiseen ja saostumiseen (suolautumisilmiö). Liukoisuuseroja käyttämällä on mahdollista erottaa proteiiniseos tavallisilla suoloilla.
Proteiinien biologisten ominaisuuksien joukossa sisältävät ensisijaisesti niiden katalyyttisen aktiivisuuden. Toinen tärkeä proteiinien biologinen ominaisuus on niiden hormonaalinen aktiivisuus, eli kyky vaikuttaa kokonaisiin reaktioryhmiin kehossa. Joillakin proteiineilla on myrkyllisiä ominaisuuksia, patogeenistä aktiivisuutta, suojaavia ja reseptoritoimintoja, ja ne ovat vastuussa solun adheesioilmiöistä.
Toinen ainutlaatuinen proteiinien biologinen ominaisuus- denaturaatio. Proteiineja niiden luonnollisessa tilassa kutsutaan natiiviksi. Denaturaatio on proteiinien spatiaalisen rakenteen tuhoamista denaturoivien aineiden vaikutuksesta. Proteiinien primäärirakenne ei vaurioidu denaturaation aikana, mutta niiden biologinen aktiivisuus sekä liukoisuus, elektroforeettinen liikkuvuus ja eräät muut reaktiot menetetään. Denaturoituessa proteiinin aktiivisen keskuksen muodostavat aminohapporadikaalit ovat avaruudellisesti etäällä toisistaan, eli proteiinin spesifinen sitoutumiskeskus ligandin kanssa tuhoutuu. Hydrofobiset radikaalit, jotka yleensä sijaitsevat pallomaisten proteiinien hydrofobisessa ytimessä, päätyvät denaturoituessaan molekyylin pinnalle, mikä luo olosuhteet saostuvien proteiinien aggregoitumiselle.
Reagenssit ja olosuhteet, jotka aiheuttavat proteiinien denaturoitumista:
Lämpötila yli 60 o C - heikkojen sidosten tuhoutuminen proteiinissa,
Hapot ja emäkset - ionogeenisten ryhmien ionisaatiomuutos, ioni- ja vetysidosten katkeaminen,
Urea - molekyylin sisäisten vetysidosten tuhoutuminen vetysidosten muodostumisen seurauksena urean kanssa,
Alkoholi, fenoli, kloramiini - hydrofobisten ja vetysidosten tuhoaminen,
Raskasmetallien suolat - proteiinien liukenemattomien suolojen muodostuminen raskasmetalli-ionien kanssa.
Kun denaturoivat aineet poistetaan, renativaatio on mahdollista, koska peptidiketjulla on taipumus omaksua konformaatio, jolla on pienin vapaa energia liuoksessa.
Soluolosuhteissa proteiinit voivat spontaanisti denaturoituu, vaikkakin hitaammin kuin korkeassa lämpötilassa. Proteiinien spontaani renativaatio solussa on vaikeaa, koska suuresta pitoisuudesta johtuen on suuri todennäköisyys osittain denaturoituneiden molekyylien aggregoitumiselle.
Solut sisältävät proteiineja- molekyylikaperoneja, joilla on kyky sitoutua osittain denaturoituihin proteiineihin, jotka ovat epävakaassa tilassa ja jotka ovat alttiita aggregaatiolle, ja palauttavat alkuperäisen konformaationsa. Aluksi nämä proteiinit löydettiin lämpösokkiproteiineiksi, koska niiden synteesi lisääntyi, kun solu altistui stressille, esimerkiksi lämpötilan noustessa. Chaperonit luokitellaan alayksikköjensä massan mukaan: hsp-60, hsp-70 ja hsp-90. Jokainen luokka sisältää sukulaisia proteiineja.
Molekyyliset chaperonit ( hsp-70) erittäin konservoitunut proteiiniluokka, jota löytyy solun kaikista osista: sytoplasmasta, tumasta, endoplasmisesta retikulumista, mitokondrioista. Yksittäisen polypeptidiketjun C-päässä hsp-70:llä on alue, joka on uurre, joka pystyy olemaan vuorovaikutuksessa 7-9 aminohappotähteen pituisten peptidien kanssa, jotka ovat rikastuneet hydrofobisilla radikaaleilla. Tällaisia alueita pallomaisissa proteiineissa esiintyy suunnilleen joka 16. aminohappo. Hsp-70 pystyy suojaamaan proteiineja lämpötilan inaktivoitumiselta ja palauttamaan osittain denaturoituneiden proteiinien konformaation ja aktiivisuuden.
Chaperones-60 (hsp-60) osallistua proteiinien tertiäärisen rakenteen muodostukseen. Hsp-60 toimii oligomeerisinä proteiineina, jotka koostuvat 14 alayksiköstä. Hsp-60 muodostaa kaksi rengasta, kukin rengas koostuu 7 alayksiköstä, jotka on kytketty toisiinsa.
Jokainen alayksikkö koostuu kolmesta alueesta:
Apikaalisessa domeenissa on useita hydrofobisia aminohappotähteitä alayksiköiden muodostaman ontelon sisäpuolelle päin;
Ekvatoriaalisella domeenilla on ATPaasiaktiivisuutta ja se on välttämätön proteiinin vapauttamiseksi kaperoniinikompleksista;
Välidomeeni yhdistää apikaaliset ja ekvatoriaaliset alueet.
Proteiini, jonka pinnalla on fragmentteja, rikastettu hydrofobisilla aminohapoilla, tulee chaperoniinikompleksin onteloon. Tämän onkalon spesifisessä ympäristössä, solun sytosolin muista molekyyleistä eristäytymisolosuhteissa, mahdollisten proteiinikonformaatioiden valinta tapahtuu, kunnes löydetään energeettisesti edullisempi konformaatio. Natiivin konformaation chaperonista riippuvainen muodostuminen liittyy huomattavan energiamäärän kulutukseen, jonka lähde on ATP.
MODUULI 1 PROTEIINIEN RAKENNE, OMINAISUUDET JA TOIMINNOT
MODUULI 1 PROTEIINIEN RAKENNE, OMINAISUUDET JA TOIMINNOT
Moduulin rakenne | Teemat |
Modulaarinen yksikkö 1 | 1.1. Proteiinien rakenneorganisaatio. Natiiviproteiinin konformaation muodostumisvaiheet 1.2. Proteiinin toiminnan perusteet. Lääkkeet ligandeina, jotka vaikuttavat proteiinien toimintaan 1.3. Proteiinien denaturoituminen ja niiden spontaanin uudelleensyntymisen mahdollisuus |
Modulaarinen yksikkö 2 | 1.4. Oligomeeristen proteiinien rakenteen ja toiminnan ominaisuudet hemoglobiinin esimerkin avulla 1.5. Natiiviproteiinin konformaation ylläpito soluolosuhteissa 1.6. Erilaisia proteiineja. Proteiiniperheet käyttämällä esimerkkiä immunoglobuliinista 1.7. Proteiinien fysikaalis-kemialliset ominaisuudet ja menetelmät niiden erottamiseen |
Moduuliyksikkö 1 MONOMEERISTEN PROTEIINIEN RAKENNEJÄRJESTELY JA NIIDEN TOIMINNAN PERUSTEET
Oppimistavoitteet Pystyy:
1. Käytä tietoa proteiinien rakenteellisista ominaisuuksista ja proteiinien toimintojen riippuvuudesta niiden rakenteesta ymmärtääksesi perinnöllisten ja hankittujen proteinopatioiden kehittymismekanismeja.
2. Selitä joidenkin lääkkeiden terapeuttisen vaikutuksen mekanismeja ligandeina, jotka ovat vuorovaikutuksessa proteiinien kanssa ja muuttavat niiden aktiivisuutta.
3. Käytä tietoa proteiinien rakenteesta ja konformationaalisesta labilisuudesta ymmärtääksesi niiden rakenteellista ja toiminnallista epästabiilisuutta ja taipumusta denaturoitua muuttuvissa olosuhteissa.
4. Selitä denaturointiaineiden käyttö lääketieteellisten materiaalien ja instrumenttien steriloinnissa sekä antiseptisinä aineina.
Tietää:
1. Proteiinien rakenteellisen organisoinnin tasot.
2. Proteiinien primäärirakenteen merkitys, joka määrää niiden rakenteellisen ja toiminnallisen monimuotoisuuden.
3. Proteiinien aktiivisen keskuksen muodostumismekanismi ja sen spesifinen vuorovaikutus ligandin kanssa, joka on proteiinien toiminnan taustalla.
4. Esimerkkejä eksogeenisten ligandien (lääkkeet, toksiinit, myrkyt) vaikutuksesta proteiinien konformaatioon ja toiminnalliseen aktiivisuuteen.
5. Proteiinien denaturoitumisen syyt ja seuraukset, denaturaatiota aiheuttavat tekijät.
6. Esimerkkejä denaturoivien tekijöiden käytöstä lääketieteessä antiseptisinä aineina ja keinoina lääketieteellisten instrumenttien sterilointiin.
AIHE 1.1. PROTEIINIEN RAKENNEJÄRJESTELY. NATIIVIN MUODOSTAMINEN
PROTEIINIKONFORMAATIOT
Proteiinit ovat polymeerimolekyylejä, joiden monomeerit ovat vain 20 α-aminohappoa. Aminohappojen yhdistelmä ja järjestys proteiinissa määräytyy yksilöiden DNA:ssa olevien geenien rakenteen perusteella. Jokainen proteiini suorittaa oman tehtävänsä erityisrakenteensa mukaisesti. Tietyn organismin proteiinien joukko määrittää sen fenotyyppiset ominaisuudet, samoin kuin perinnöllisten sairauksien esiintymisen tai taipumuksen niiden kehittymiseen.
1. Aminohapot, jotka muodostavat proteiineja. Peptidisidos. Proteiinit ovat polymeerejä, jotka on rakennettu monomeereistä - 20 α-aminohaposta, joiden yleinen kaava on
Aminohapot eroavat α-hiiliatomiin kiinnittyneiden radikaalien rakenteesta, koosta ja fysikaalis-kemiallisista ominaisuuksista. Aminohappojen funktionaaliset ryhmät määräävät eri α-aminohappojen ominaisuuksien ominaisuudet. α-aminohapoissa esiintyvät radikaalit voidaan jakaa useisiin ryhmiin:
proliini, Toisin kuin muut 19 proteiinimonomeeriä, se ei ole aminohappo, vaan iminohappo, proliinissa oleva radikaali liittyy sekä a-hiiliatomiin että iminoryhmään
 Aminohappojen liukoisuus veteen vaihtelee. Tämä johtuu radikaalien kyvystä olla vuorovaikutuksessa veden (hydraatin) kanssa.
Aminohappojen liukoisuus veteen vaihtelee. Tämä johtuu radikaalien kyvystä olla vuorovaikutuksessa veden (hydraatin) kanssa.
TO hydrofiilinen Käsitteet sisältävät anionisia, kationisia ja polaarisia varautumattomia funktionaalisia ryhmiä sisältävät radikaalit.
TO hydrofobinen Käsitteet sisältävät radikaaleja, jotka sisältävät metyyliryhmiä, alifaattisia ketjuja tai renkaita.
2. Peptidisidokset yhdistävät aminohappoja muodostaen peptidejä. Peptidisynteesin aikana yhden aminohapon α-karboksyyliryhmä on vuorovaikutuksessa toisen aminohapon α-aminoryhmän kanssa muodostaen peptidisidos:
Proteiinit ovat polypeptidejä, ts. α-aminohappojen lineaariset polymeerit, jotka on yhdistetty peptidisidoksella (kuva 1.1.)
 Riisi. 1.1. Termit, joita käytetään kuvaamaan peptidien rakennetta
Riisi. 1.1. Termit, joita käytetään kuvaamaan peptidien rakennetta
Aminohappojen monomeerejä, jotka muodostavat polypeptidejä, kutsutaan aminohappotähteet. Toistuvien ryhmien ketju - NH-CH-CO- lomakkeet peptidirunko. Aminohappotähdettä, jossa on vapaa a-aminoryhmä, kutsutaan N-päätteeksi, ja aminohappotähdettä, jossa on vapaa a-karboksyyliryhmä, kutsutaan C-päätteeksi. Peptidit kirjoitetaan ja luetaan N-päästä C-päähän.
Proliinin iminoryhmän muodostama peptidisidos eroaa muista peptidisidoksista: peptidiryhmän typpiatomista puuttuu vety,
sen sijaan on olemassa sidos radikaalin kanssa, jonka seurauksena renkaan toinen puoli sisältyy peptidirunkoon:
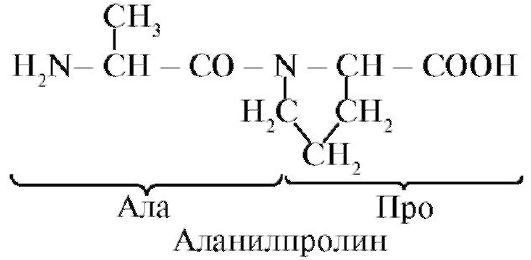 Peptidit eroavat aminohappokoostumuksesta, aminohappojen lukumäärästä ja aminohappoliitoksen järjestyksestä, esimerkiksi Ser-Ala-Glu-Gis ja His-Glu-Ala-Ser ovat kaksi eri peptidiä.
Peptidit eroavat aminohappokoostumuksesta, aminohappojen lukumäärästä ja aminohappoliitoksen järjestyksestä, esimerkiksi Ser-Ala-Glu-Gis ja His-Glu-Ala-Ser ovat kaksi eri peptidiä.
Peptidisidokset ovat erittäin vahvoja ja niiden kemiallinen ei-entsymaattinen hydrolyysi vaatii ankaria olosuhteita: analysoitava proteiini hydrolysoituu väkevässä suolahapossa noin 110°:n lämpötilassa 24 tunnin ajan. Elävässä solussa peptidisidokset voivat katketa proteolyyttiset entsyymit, nimeltään proteaasit tai peptidihydrolaasit.
3. Proteiinien perusrakenne. Aminohappotähteet eri proteiinien peptidiketjuissa eivät vuorottele satunnaisesti, vaan ne on järjestetty tiettyyn järjestykseen. Polypeptidiketjun aminohappotähteiden lineaarista sekvenssiä tai vuorottelujärjestystä kutsutaan proteiinin primaarirakenne.
Kunkin yksittäisen proteiinin primäärirakenne on koodattu DNA-molekyylissä (alueella, jota kutsutaan geeniksi) ja se toteutuu transkription (informaation kopioiminen mRNA:lle) ja translaation (proteiinin primäärirakenteen synteesi) aikana. Näin ollen yksittäisen ihmisen proteiinien primäärirakenne on vanhemmilta lapsille perinnöllisesti välittyvä tieto, joka määrää tietyn organismin proteiinien rakenteelliset ominaisuudet, joista olemassa olevien proteiinien toiminta riippuu (kuva 1.2.).
 Riisi. 1.2. Genotyypin ja yksilön kehossa syntetisoitujen proteiinien konformaation välinen suhde
Riisi. 1.2. Genotyypin ja yksilön kehossa syntetisoitujen proteiinien konformaation välinen suhde
Jokaisella ihmiskehon noin 100 000 yksittäisestä proteiinista on ainutlaatuinen ensisijainen rakenne. Samantyyppisten proteiinien (esimerkiksi albumiinin) molekyyleillä on sama aminohappotähteiden vuorottelu, mikä erottaa albumiinin kaikista muista yksittäisistä proteiineista.
Peptidiketjun aminohappotähteiden sekvenssiä voidaan pitää eräänä tiedon tallennusmuotona. Tämä tieto määrittää lineaarisen peptidiketjun tilajärjestelyn kompaktimmaksi kolmiulotteiseksi rakenteeksi ns. konformaatio orava. Funktionaalisesti aktiivisen proteiinikonformaation muodostumisprosessia kutsutaan taitettava
4. Proteiinin rakenne. Vapaa pyöriminen peptidirungossa on mahdollista peptidiryhmän typpiatomin ja viereisen a-hiiliatomin välillä sekä a-hiiliatomin ja karbonyyliryhmän hiilen välillä. Aminohappotähteiden funktionaalisten ryhmien vuorovaikutuksesta johtuen proteiinien primaarirakenne voi saada monimutkaisempia tilarakenteita. Globulaarisissa proteiineissa on kaksi pääasiallista peptidiketjujen konformaation laskostumistasoa: toissijainen Ja tertiäärinen rakenne.
Proteiinien toissijainen rakenne on spatiaalinen rakenne, joka muodostuu vetysidosten muodostumisen seurauksena peptidirungon funktionaalisten ryhmien -C=O ja -NH- välille. Tässä tapauksessa peptidiketju voi hankkia kahdenlaisia säännöllisiä rakenteita: α-heliksit Ja β-rakenteet.
SISÄÄN α-heliksit vetysidokset muodostuvat karbonyyliryhmän happiatomin ja siitä peräisin olevan 4. aminohapon amiditypen vedyn välille; aminohappotähteiden sivuketjut
sijaitsevat spiraalin kehällä osallistumatta toisiorakenteen muodostukseen (kuva 1.3.).
Bulkkiradikaalit tai radikaalit, joissa on samat varaukset, estävät α-kierteen muodostumisen. Proliinitähde, jolla on rengasrakenne, katkaisee a-heliksin, koska vedyn puuttumisen vuoksi peptidiketjun typpiatomissa on mahdotonta muodostaa vetysidosta. Typen ja a-hiiliatomin välinen sidos on osa proliinirengasta, joten peptidirunko taipuu tässä vaiheessa.
β-rakenne muodostuu yhden polypeptidiketjun peptidirungon lineaaristen alueiden väliin, jolloin muodostuu laskostettuja rakenteita. Polypeptidiketjuja tai niiden osia voi muodostua rinnakkain tai antirinnakkaiset β-rakenteet. Ensimmäisessä tapauksessa vuorovaikutuksessa olevien peptidiketjujen N- ja C-päät ovat yhtenevät, ja toisessa niillä on vastakkainen suunta (kuva 1.4).
Riisi. 1.3. Proteiinin sekundaarirakenne - α-heliksi
 Riisi. 1.4. Rinnakkaiset ja antirinnakkaiset β-levyrakenteet
Riisi. 1.4. Rinnakkaiset ja antirinnakkaiset β-levyrakenteet
β-rakenteet on merkitty leveillä nuolilla: A - Vastasuuntainen β-rakenne. B - Rinnakkaiset β-levyrakenteet
Joissakin proteiineissa β-rakenteita voi muodostua johtuen vetysidosten muodostumisesta eri polypeptidiketjujen peptidirungon atomien välille.
Löytyy myös proteiineista alueet, joilla on epäsäännöllinen toissijainen rakenne, joka sisältää polypeptidirungon mutkia, silmukoita ja käänteitä. Ne sijaitsevat usein paikoissa, joissa peptidiketjun suunta muuttuu, esimerkiksi kun muodostuu yhdensuuntainen β-levyrakenne.
α-heliksien ja β-rakenteiden läsnäolon perusteella pallomaiset proteiinit voidaan jakaa neljään luokkaan.
Riisi. 1.5. Myoglobiinin (A) ja hemoglobiinin β-ketjun (B) toissijainen rakenne, joka sisältää kahdeksan α-heliksiä

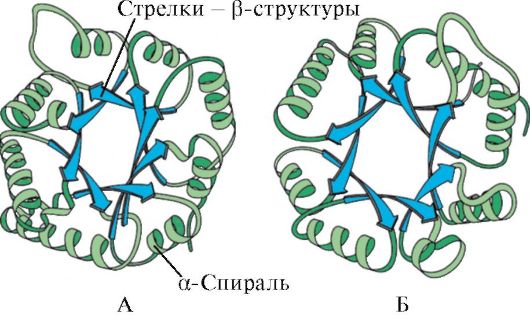 Riisi. 1.6. Triosefosfaatti-isomeraasin ja pyruvaattikinaasidomeenin toissijainen rakenne
Riisi. 1.6. Triosefosfaatti-isomeraasin ja pyruvaattikinaasidomeenin toissijainen rakenne
 Riisi. 1.7. Immunoglobuliinin (A) vakiodomeenin ja superoksididismutaasientsyymin (B) toissijainen rakenne
Riisi. 1.7. Immunoglobuliinin (A) vakiodomeenin ja superoksididismutaasientsyymin (B) toissijainen rakenne
SISÄÄN neljäs luokka sisälsi proteiineja, jotka sisältävät pienen määrän säännöllisiä sekundaarisia rakenteita. Nämä proteiinit sisältävät pieniä kysteiinipitoisia proteiineja tai metalloproteiineja.
Proteiinin tertiäärinen rakenne- aminohapporadikaalien välisistä vuorovaikutuksista muodostuva konformaatiotyyppi, joka voi sijaita huomattavan etäisyyden päässä toisistaan peptidiketjussa. Useimmat proteiinit muodostavat spatiaalisen rakenteen, joka muistuttaa palloa (globulaariset proteiinit).
Koska hydrofobiset aminohapporadikaalit pyrkivät yhdistymään ns hydrofobisia vuorovaikutuksia ja molekyylien välisten van der Waalsin voimien vuoksi proteiinipallon sisään muodostuu tiheä hydrofobinen ydin. Hydrofiiliset ionisoidut ja ionisoimattomat radikaalit sijaitsevat pääasiassa proteiinin pinnalla ja määrittävät sen vesiliukoisuuden.
 Riisi. 1.8. Sidostyypit, jotka syntyvät aminohapporadikaalien välille proteiinin tertiaarisen rakenteen muodostumisen aikana
Riisi. 1.8. Sidostyypit, jotka syntyvät aminohapporadikaalien välille proteiinin tertiaarisen rakenteen muodostumisen aikana
1 - ionisidos- esiintyy positiivisesti ja negatiivisesti varautuneiden funktionaalisten ryhmien välillä;
2 - vetysidos- esiintyy hydrofiilisen varaamattoman ryhmän ja minkä tahansa muun hydrofiilisen ryhmän välillä;
3 - hydrofobisia vuorovaikutuksia- syntyvät hydrofobisten radikaalien välillä;
4 - disulfidisidos- muodostuu kysteiinitähteiden SH-ryhmien hapettumisesta ja niiden vuorovaikutuksesta keskenään
Hydrofiiliset aminohappotähteet, jotka sijaitsevat hydrofobisen ytimen sisällä, voivat olla vuorovaikutuksessa toistensa kanssa käyttämällä ioninen Ja vetysidoksia(Kuva 1.8).
Ioni- ja vetysidokset sekä hydrofobiset vuorovaikutukset ovat heikkoja: niiden energia ei ole paljon suurempi kuin molekyylien lämpöliikkeen energia huoneenlämpötilassa. Proteiinin konformaatiota ylläpidetään muodostamalla monia tällaisia heikkoja sidoksia. Koska proteiinin muodostavat atomit ovat jatkuvassa liikkeessä, on mahdollista rikkoa joitakin heikkoja sidoksia ja muodostaa muita, mikä johtaa polypeptidiketjun yksittäisten osien vähäisiin liikkeisiin. Tätä proteiinien ominaisuutta muuttaa konformaatiota joidenkin hajoamisen ja muiden heikkojen sidosten muodostumisen seurauksena kutsutaan ns. konformationaalinen labilisuus.
Ihmiskehossa on järjestelmät, jotka tukevat homeostaasi- sisäisen ympäristön pysyvyys tietyissä terveen kehon hyväksyttävissä rajoissa. Homeostaasin olosuhteissa pienet muutokset konformaatiossa eivät häiritse proteiinien yleistä rakennetta ja toimintaa. Proteiinin toiminnallisesti aktiivista konformaatiota kutsutaan syntyperäinen rakenne. Muutokset sisäisessä ympäristössä (esim. glukoosin, Ca-ionien, protonien jne. pitoisuus) johtavat konformaatiomuutoksiin ja proteiinien toimintojen häiriintymiseen.
Joidenkin proteiinien tertiäärinen rakenne on stabiloitunut disulfidisidokset, muodostuu kahden tähteen -SH-ryhmien vuorovaikutuksesta
 Riisi. 1.9. Disulfidisidoksen muodostuminen proteiinimolekyylissä
Riisi. 1.9. Disulfidisidoksen muodostuminen proteiinimolekyylissä
kysteiini (kuva 1.9). Useimpien solunsisäisten proteiinien tertiäärisessä rakenteessa ei ole kovalenttisia disulfidisidoksia. Niiden läsnäolo on ominaista solun erittämille proteiineille, mikä varmistaa niiden paremman stabiilisuuden solunulkoisissa olosuhteissa. Siten disulfidisidoksia on läsnä insuliinin ja immunoglobuliinien molekyyleissä.
Insuliini- proteiinihormoni, joka syntetisoituu haiman β-soluissa ja erittyy vereen vasteena veren glukoosipitoisuuden nousulle. Insuliinin rakenteessa on kaksi disulfidisidosta, jotka yhdistävät polypeptidin A- ja B-ketjut, ja yksi disulfidisidos A-ketjussa (kuva 1.10).
 Riisi. 1.10. Disulfidisidokset insuliinin rakenteessa
Riisi. 1.10. Disulfidisidokset insuliinin rakenteessa
5. Proteiinien ylisekundaarinen rakenne. Proteiineissa, joilla on erilainen primäärirakenne ja toiminnot, niitä joskus havaitaan samanlaiset yhdistelmät ja toissijaisten rakenteiden suhteelliset sijainnit, joita kutsutaan ylisekundaariseksi rakenteeksi. Sillä on väliasema sekundaari- ja tertiaarirakenteiden välillä, koska se on sekundäärisen rakenteen elementtien spesifinen yhdistelmä proteiinin tertiaarisen rakenteen muodostuksessa. Supersekundaarisilla rakenteilla on erityiset nimet, kuten "α-helix-turn-a-helix", "leusiinivetoketju", "sinkkisormet" jne. Tällaiset supersekundaariset rakenteet ovat ominaisia DNA:ta sitoville proteiineille.
"Leusiinivetoketju." Tämän tyyppistä ylisekundaarista rakennetta käytetään yhdistämään kaksi proteiinia yhteen. Vuorovaikutteisten proteiinien pinnalla on α-kierteisiä alueita, jotka sisältävät vähintään neljä leusiinitähdettä. Leusiinitähteet α-heliksissä sijaitsevat kuuden aminohapon päässä toisistaan. Koska jokainen α-heliksin kierros sisältää 3,6 aminohappotähdettä, leusiiniradikaalit sijaitsevat jokaisen toisen kierroksen pinnalla. Yhden proteiinin α-heliksin leusiinitähteet voivat olla vuorovaikutuksessa toisen proteiinin leusiinitähteiden kanssa (hydrofobiset vuorovaikutukset), jotka yhdistävät ne toisiinsa (kuva 1.11.). Monet DNA:ta sitovat proteiinit toimivat oligomeerisissa komplekseissa, joissa yksittäiset alayksiköt on liitetty toisiinsa "leusiinivetoketjuilla".
 Riisi. 1.11. "Leusiinivetoketju" kahden proteiinin α-kierteisten alueiden välissä
Riisi. 1.11. "Leusiinivetoketju" kahden proteiinin α-kierteisten alueiden välissä
Esimerkki sellaisista proteiineista ovat histonit. Histonit- ydinproteiinit, jotka sisältävät suuren määrän positiivisesti varautuneita aminohappoja - arginiinia ja lysiiniä (jopa 80%). Histonimolekyylit yhdistetään oligomeerisiksi komplekseiksi, jotka sisältävät kahdeksan monomeeriä käyttämällä "leusiinivetoketjuja", huolimatta näiden molekyylien merkittävästä homonyymistä varauksesta.
"sinkkisormi"- DNA:ta sitoville proteiineille tyypillinen supersekundäärisen rakenteen variantti on proteiinin pinnalla pitkänomaisen fragmentin muotoinen ja sisältää noin 20 aminohappotähdettä (kuva 1.12). "Jatketun sormen" muotoa tukee sinkkiatomi, joka on sitoutunut neljään aminohapporadikaaliin - kahteen kysteiinitähteeseen ja kahteen histidiinitähteeseen. Joissakin tapauksissa histidiinitähteiden sijasta on kysteiinijäämiä. Kaksi lähellä olevaa kysteiinitähdettä on erotettu kahdesta muusta Gisili-tähteestä Cys-sekvenssillä, joka koostuu noin 12 aminohappotähteestä. Tämä proteiinin alue muodostaa a-heliksin, jonka radikaalit voivat sitoutua spesifisesti DNA:n suuren uran säätelyalueisiin. Yksilöllinen sitoutumisspesifisyys
 Riisi. 1.12. DNA:ta sitovien proteiinien alueen ensisijainen rakenne, joka muodostaa "sinkkisormen" rakenteen (kirjaimet osoittavat aminohappoja, jotka muodostavat tämän rakenteen)
Riisi. 1.12. DNA:ta sitovien proteiinien alueen ensisijainen rakenne, joka muodostaa "sinkkisormen" rakenteen (kirjaimet osoittavat aminohappoja, jotka muodostavat tämän rakenteen)
Säätelevä DNA:ta sitova proteiini riippuu sinkkisormen alueella sijaitsevien aminohappotähteiden sekvenssistä. Tällaiset rakenteet sisältävät erityisesti steroidihormonireseptoreita, jotka osallistuvat transkription säätelyyn (informaation lukeminen DNA:sta RNA:ksi).
AIHE 1.2. PROTEIININ TOIMINNAN PERUSTEET. LAIMET LIGANDEINA, JOTKA VAIKUTTAVAT PROTEIINITOIMINNASSA
1. Proteiinin aktiivinen keskus ja sen vuorovaikutus ligandin kanssa. Tertiäärisen rakenteen muodostumisen aikana toiminnallisesti aktiivisen proteiinin pinnalle muodostuu yleensä syvennykseen alue, jonka muodostavat primäärirakenteessa kaukana toisistaan olevat aminohapporadikaalit. Tätä aluetta, jolla on ainutlaatuinen rakenne tietylle proteiinille ja joka pystyy spesifisesti vuorovaikuttamaan tietyn molekyylin tai samankaltaisten molekyylien ryhmän kanssa, kutsutaan proteiini-ligandin sitomispaikaksi tai aktiiviseksi paikaksi. Ligandit ovat molekyylejä, jotka ovat vuorovaikutuksessa proteiinien kanssa.
Korkea spesifisyys Proteiinin vuorovaikutus ligandin kanssa varmistetaan aktiivisen keskuksen rakenteen komplementaarisella ligandin rakenteen kanssa.
Täydentävyys- tämä on vuorovaikutuksessa olevien pintojen avaruudellinen ja kemiallinen vastaavuus. Aktiivisen keskuksen tulee paitsi avaruudellisesti vastata siihen kuuluvaa ligandia, myös sidoksia (ioni-, vety- ja hydrofobisia vuorovaikutuksia) tulee muodostua aktiiviseen keskukseen sisältyvien radikaalien funktionaalisten ryhmien ja ligandin pitävän ligandin välille. aktiivisessa keskustassa (kuva 1.13 ).
 Riisi. 1.13. Proteiinin täydentävä vuorovaikutus ligandin kanssa
Riisi. 1.13. Proteiinin täydentävä vuorovaikutus ligandin kanssa
Joillakin ligandeilla, kun ne ovat kiinnittyneet proteiinin aktiiviseen keskukseen, on apurooli proteiinien toiminnassa. Tällaisia ligandeja kutsutaan kofaktoreiksi, ja proteiineja, jotka sisältävät ei-proteiiniosan, kutsutaan monimutkaiset proteiinit(toisin kuin yksinkertaiset proteiinit, jotka koostuvat vain proteiiniosasta). Ei-proteiiniosaa, joka on kiinteästi liitetty proteiiniin, kutsutaan prosteettinen ryhmä. Esimerkiksi myoglobiini, hemoglobiini ja sytokromit sisältävät proteettisen ryhmän, hemin, joka sisältää rautaionin, joka on kiinteästi kiinnittynyt aktiiviseen keskustaan. Hemeä sisältäviä monimutkaisia proteiineja kutsutaan hemoproteiineiksi.
Kun spesifisiä ligandeja kiinnittyy proteiineihin, näiden proteiinien toiminta ilmenee. Siten albumiini, veriplasman tärkein proteiini, osoittaa kuljetustoimintonsa kiinnittämällä aktiiviseen keskukseen hydrofobisia ligandeja, kuten rasvahappoja, bilirubiinia, joitain lääkkeitä jne. (Kuva 1.14)
Peptidiketjun kolmiulotteisen rakenteen kanssa vuorovaikutuksessa olevat ligandit voivat olla paitsi pienimolekyylisiä orgaanisia ja epäorgaanisia molekyylejä, myös makromolekyylejä:
DNA (esimerkkejä edellä käsitellyistä DNA:ta sitovista proteiineista);
polysakkaridit;
 Riisi. 1.14. Genotyypin ja fenotyypin välinen suhde
Riisi. 1.14. Genotyypin ja fenotyypin välinen suhde
DNA-molekyyliin koodattu ihmisen proteiinien ainutlaatuinen primäärirakenne toteutuu soluissa ainutlaatuisen konformaation, aktiivisen keskusrakenteen ja proteiinitoimintojen muodossa.
Näissä tapauksissa proteiini tunnistaa ligandin spesifisen alueen, joka on suhteessa sitoutumiskohtaan ja komplementaarinen. Siten maksasolujen pinnalla on insuliinihormonin reseptoriproteiineja, jolla on myös proteiinirakenne. Insuliinin vuorovaikutus reseptorin kanssa aiheuttaa muutoksen sen konformaatiossa ja signaalijärjestelmien aktivoitumisen, mikä johtaa ravinteiden varastoimiseen maksasoluihin aterioiden jälkeen.
Täten, Proteiinien toiminta perustuu proteiinin aktiivisen keskuksen spesifiseen vuorovaikutukseen ligandin kanssa.
2. Domeenirakenne ja sen rooli proteiinien toiminnassa. Globulaaristen proteiinien pitkät polypeptidiketjut laskostuvat usein useiksi kompakteiksi, suhteellisen itsenäisiksi alueiksi. Niillä on itsenäinen tertiäärinen rakenne, joka muistuttaa globulaaristen proteiinien rakennetta, ja niitä kutsutaan verkkotunnuksia. Proteiinien domeenirakenteen ansiosta niiden tertiäärinen rakenne on helpompi muodostaa.
Domeeniproteiineissa ligandia sitovat kohdat sijaitsevat usein domeenien välissä. Siten trypsiini on proteolyyttinen entsyymi, jota haiman eksokriininen osa tuottaa ja joka on välttämätön ruokaproteiinien sulamiselle. Sillä on kaksidomeenirakenne, ja trypsiinin ja sen ligandin - ruokaproteiinin - sitoutumiskeskus sijaitsee näiden kahden domeenin välisessä urassa. Aktiivisessa keskustassa luodaan olosuhteet, jotka ovat välttämättömiä ruokaproteiinin tietyn kohdan tehokkaalle sitoutumiselle ja sen peptidisidosten hydrolyysille.
Proteiinin eri domeenit voivat liikkua suhteessa toisiinsa, kun aktiivinen keskus on vuorovaikutuksessa ligandin kanssa (kuva 1.15).
Heksokinaasi- entsyymi, joka katalysoi glukoosin fosforylaatiota ATP:n avulla. Entsyymin aktiivinen kohta sijaitsee kahden domeenin välisessä rakossa. Kun heksokinaasi sitoutuu glukoosiin, sitä ympäröivät domeenit sulkeutuvat ja substraatti jää loukkuun, jossa tapahtuu fosforylaatiota (katso kuva 1.15).
 Riisi. 1.15. Heksokinaasidomeenien sitoutuminen glukoosiin
Riisi. 1.15. Heksokinaasidomeenien sitoutuminen glukoosiin
Joissakin proteiineissa domeenit suorittavat itsenäisiä toimintoja sitoutumalla erilaisiin ligandeihin. Tällaisia proteiineja kutsutaan monitoimisiksi.
3. Lääkkeet ovat ligandeja, jotka vaikuttavat proteiinien toimintaan. Proteiinien vuorovaikutus ligandien kanssa on spesifistä. Proteiinin ja sen aktiivisen keskuksen konformaatiolabiteetin vuoksi on kuitenkin mahdollista valita toinen aine, joka voisi myös olla vuorovaikutuksessa proteiinin kanssa aktiivisessa keskustassa tai muussa molekyylin osassa.
Ainetta, joka on rakenteeltaan samanlainen kuin luonnollinen ligandi, kutsutaan ligandin rakenteellinen analogi tai ei-luonnollinen ligandi. Se on myös vuorovaikutuksessa proteiinin kanssa aktiivisessa kohdassa. Ligandin rakenteellinen analogi voi molemmat parantaa proteiinin toimintaa (agonisti), ja vähentää sitä (antagonisti). Ligandi ja sen rakenteelliset analogit kilpailevat toistensa kanssa sitoutumisesta proteiiniin samassa kohdassa. Tällaisia aineita kutsutaan kilpailukykyiset modulaattorit proteiinitoimintojen säätelijät. Monet lääkkeet toimivat proteiinin estäjinä. Jotkut niistä saadaan luonnollisten ligandien kemiallisella modifioinnilla. Proteiinitoimintojen estäjiä voivat olla lääkkeet ja myrkyt.
Atropiini on M-kolinergisten reseptorien kilpaileva estäjä. Asetyylikoliini on välittäjäaine hermoimpulssien välittämiseen kolinergisten synapsien kautta. Herätyksen suorittamiseksi synaptiseen rakoon vapautuvan asetyylikoliinin on oltava vuorovaikutuksessa postsynaptisen kalvon reseptoriproteiinin kanssa. Kaksi tyyppiä löytyi kolinergiset reseptorit:
M-reseptori asetyylikoliinin lisäksi se vuorovaikuttaa selektiivisesti muskariinin (kärpäshelttatoksiinin) kanssa. M - kolinergiset reseptorit ovat läsnä sileissä lihaksissa ja vuorovaikutuksessa asetyylikoliinin kanssa aiheuttavat niiden supistumisen;
H-reseptori sitoutuvat erityisesti nikotiiniin. N-kolinergisiä reseptoreita löytyy poikkijuovaisten luustolihasten synapseista.
Spesifinen estäjä M-kolinergiset reseptorit on atropiini. Sitä löytyy belladonna- ja henbane-kasveista.
 Atropiinissa on rakenteeltaan samanlaisia funktionaalisia ryhmiä kuin asetyylikoliinilla ja niiden tilajärjestelyllä, joten se on M-kolinergisten reseptorien kilpaileva estäjä. Koska asetyylikoliinin sitoutuminen M-kolinergisiin reseptoreihin aiheuttaa sileiden lihasten supistumista, atropiinia käytetään lääkkeenä, joka lievittää niiden kouristuksia. (spasmodinen). Siten on tunnettua käyttää atropiinia silmälihasten rentouttamiseen silmänpohjaa tarkasteltaessa sekä kouristuksen lievittämiseen maha-suolikanavan koliikkien aikana. M-kolinergisiä reseptoreita on myös keskushermostossa (CNS), joten suuret annokset atropiinia voivat aiheuttaa keskushermoston ei-toivotun reaktion: motorista ja henkistä kiihtyneisyyttä, hallusinaatioita, kouristuksia.
Atropiinissa on rakenteeltaan samanlaisia funktionaalisia ryhmiä kuin asetyylikoliinilla ja niiden tilajärjestelyllä, joten se on M-kolinergisten reseptorien kilpaileva estäjä. Koska asetyylikoliinin sitoutuminen M-kolinergisiin reseptoreihin aiheuttaa sileiden lihasten supistumista, atropiinia käytetään lääkkeenä, joka lievittää niiden kouristuksia. (spasmodinen). Siten on tunnettua käyttää atropiinia silmälihasten rentouttamiseen silmänpohjaa tarkasteltaessa sekä kouristuksen lievittämiseen maha-suolikanavan koliikkien aikana. M-kolinergisiä reseptoreita on myös keskushermostossa (CNS), joten suuret annokset atropiinia voivat aiheuttaa keskushermoston ei-toivotun reaktion: motorista ja henkistä kiihtyneisyyttä, hallusinaatioita, kouristuksia.
Ditiliini on H-kolinergisten reseptorien kilpaileva agonisti, joka estää neuromuskulaaristen synapsien toimintaa.
 Luustolihasten neuromuskulaariset synapsit sisältävät H-kolinergisiä reseptoreita. Niiden vuorovaikutus asetyylikoliinin kanssa johtaa lihasten supistuksiin. Joissakin kirurgisissa leikkauksissa sekä endoskooppisissa tutkimuksissa käytetään lääkkeitä, jotka aiheuttavat luurankolihasten rentoutumista (lihasrelaksantit). Näitä ovat ditiliini, joka on asetyylikoliinin rakenteellinen analogi. Se kiinnittyy H-kolinergisiin reseptoreihin, mutta toisin kuin asetyylikoliini, tuhoaa sen hyvin hitaasti. Ionikanavien pitkittyneen avautumisen ja kalvon jatkuvan depolarisaation seurauksena hermoimpulssien johtuminen häiriintyy ja lihasten rentoutuminen tapahtuu. Aluksi nämä ominaisuudet löydettiin curare-myrkystä, minkä vuoksi tällaisia lääkkeitä kutsutaan curaremainen.
Luustolihasten neuromuskulaariset synapsit sisältävät H-kolinergisiä reseptoreita. Niiden vuorovaikutus asetyylikoliinin kanssa johtaa lihasten supistuksiin. Joissakin kirurgisissa leikkauksissa sekä endoskooppisissa tutkimuksissa käytetään lääkkeitä, jotka aiheuttavat luurankolihasten rentoutumista (lihasrelaksantit). Näitä ovat ditiliini, joka on asetyylikoliinin rakenteellinen analogi. Se kiinnittyy H-kolinergisiin reseptoreihin, mutta toisin kuin asetyylikoliini, tuhoaa sen hyvin hitaasti. Ionikanavien pitkittyneen avautumisen ja kalvon jatkuvan depolarisaation seurauksena hermoimpulssien johtuminen häiriintyy ja lihasten rentoutuminen tapahtuu. Aluksi nämä ominaisuudet löydettiin curare-myrkystä, minkä vuoksi tällaisia lääkkeitä kutsutaan curaremainen.
AIHE 1.3. Proteiinien DENATUROINTI JA NIIDEN SPONTAANI UUDISTUMISEN MAHDOLLISUUS
1. Koska proteiinien natiivi konformaatio säilyy heikkojen vuorovaikutusten vuoksi, proteiinia ympäröivän ympäristön koostumuksen ja ominaisuuksien muutokset, altistuminen kemiallisille reagensseille ja fysikaalisille tekijöille aiheuttavat muutoksen niiden konformaatiossa (konformationaalisuuden ominaisuus). Suuren määrän sidosten katkeaminen johtaa proteiinien luonnollisen konformaation tuhoutumiseen ja denaturoitumiseen.
Proteiinien denaturaatio- tämä on niiden luonnollisen konformaation tuhoaminen denaturoivien aineiden vaikutuksesta, mikä johtuu heikkojen sidosten katkeamisesta, jotka stabiloivat proteiinin avaruudellista rakennetta. Denaturoitumiseen liittyy proteiinin ainutlaatuisen kolmiulotteisen rakenteen ja aktiivisen keskuksen tuhoutuminen ja sen biologisen aktiivisuuden menetys (kuva 1.16).
Kaikki yhden proteiinin denaturoidut molekyylit saavat satunnaisen konformaation, joka eroaa saman proteiinin muista molekyyleistä. Aktiivisen keskuksen muodostavat aminohapporadikaalit osoittautuvat avaruudellisesti etäämmiksi toisistaan, ts. proteiinin spesifinen sitoutumiskohta ligandin kanssa tuhoutuu. Denaturaation aikana proteiinien primäärirakenne pysyy muuttumattomana.
Denaturointiaineiden käyttö biologisessa tutkimuksessa ja lääketieteessä. Biokemiallisissa tutkimuksissa proteiinit poistetaan yleensä ensin liuoksesta ennen alhaisen molekyylipainon yhdisteiden määrittämistä biologisesta materiaalista. Trikloorietikkahappoa (TCA) käytetään useimmiten tähän tarkoitukseen. Kun TCA on lisätty liuokseen, denaturoidut proteiinit saostuvat ja ne poistetaan helposti suodattamalla (taulukko 1.1.)
Lääketieteessä denaturoivia aineita käytetään usein lääketieteellisten instrumenttien ja materiaalien sterilointiin autoklaaveissa (denaturoiva aine on korkea lämpötila) ja antiseptisinä aineina (alkoholi, fenoli, kloramiini) patogeenista mikroflooraa sisältävien saastuneiden pintojen käsittelyyn.
2. Spontaani proteiinin uudelleenaktivoituminen- todiste proteiinien primäärirakenteen, konformaation ja toiminnan determinismistä. Yksittäiset proteiinit ovat yhden geenin tuotteita, joilla on identtinen aminohapposekvenssi ja jotka saavat saman konformaation solussa. Peruspäätelmä, että proteiinin primaarirakenne sisältää jo tietoa sen konformaatiosta ja toiminnasta, tehtiin joidenkin proteiinien (erityisesti ribonukleaasin ja myoglobiinin) kyvyn perusteella spontaani renativoitua - palauttaa natiivi konformaatio denaturoinnin jälkeen.
Avaruusproteiinirakenteiden muodostuminen tapahtuu itsekokoamismenetelmällä - spontaanilla prosessilla, jossa polypeptidiketjulla, jolla on ainutlaatuinen primäärirakenne, on taipumus omaksua konformaatio, jossa on alhaisin vapaaenergia liuoksessa. Kyky renativoida proteiineja, jotka säilyttävät primäärirakenteensa denaturoinnin jälkeen, kuvattiin kokeessa ribonukleaasientsyymillä.
Ribonukleaasi on entsyymi, joka hajottaa yksittäisten nukleotidien välisiä sidoksia RNA-molekyylissä. Tällä pallomaisella proteiinilla on yksi polypeptidiketju, jonka tertiääristä rakennetta stabiloivat monet heikot ja neljä disulfidisidosta.
Ribonukleaasin käsittely urealla, joka katkaisee vetysidoksia molekyylissä, ja pelkistimellä, joka katkaisee disulfidisidoksia, johtaa entsyymin denaturoitumiseen ja sen aktiivisuuden menettämiseen.
Denaturoivien aineiden poistaminen dialyysillä johtaa proteiinin konformaation ja toiminnan palautumiseen, ts. uudestisyntymiseen. (Kuva 1.17).
 Riisi. 1.17. Ribonukleaasin denaturaatio ja renaturaatio
Riisi. 1.17. Ribonukleaasin denaturaatio ja renaturaatio
A - ribonukleaasin natiivi konformaatio, jonka tertiäärisessä rakenteessa on neljä disulfidisidosta; B - denaturoitu ribonukleaasimolekyyli;
B - uudelleenaktivoitu ribonukleaasimolekyyli, jolla on palautettu rakenne ja toiminta
1. Täytä taulukko 1.2.
Taulukko 1.2. Aminohappojen luokittelu radikaalien polariteetin mukaan
2. Kirjoita tetrapeptidin kaava:
Asp - Pro - Fen - Liz
a) korostaa peptidin toistuvia ryhmiä, jotka muodostavat peptidirungon, ja aminohapporadikaalien edustamat muuttuvat ryhmät;
b) leimaa N- ja C-päät;
c) korostaa peptidisidoksia;
d) kirjoittaa toinen peptidi, joka koostuu samoista aminohapoista;
e) laskea tetrapeptidin mahdollisten varianttien lukumäärä, joilla on samanlainen aminohappokoostumus.
3. Selitä proteiinien primäärirakenteen rooli nisäkkään neurohypofyysin kahden rakenteellisesti samanlaisen ja evoluutionaalisesti läheisen peptidihormonin - oksitosiinin ja vasopressiinin - vertailevan analyysin esimerkillä (taulukko 1.3).
Taulukko 1.3. Oksitosiinin ja vasopressiinin rakenne ja toiminnot
 Tätä varten:
Tätä varten:
a) vertaa kahden peptidin koostumusta ja aminohapposekvenssiä;
b) löytää näiden kahden peptidin primäärirakenteen samankaltaisuus ja niiden biologisen vaikutuksen samankaltaisuus;
c) löytää eroja kahden peptidin rakenteessa ja eroja niiden toiminnassa;
d) tehdä johtopäätös peptidien primäärirakenteen vaikutuksesta niiden toimintoihin.
4. Kuvaa globulaaristen proteiinien konformaation muodostumisen päävaiheet (sekundaariset, tertiaariset rakenteet, supersekundaarisen rakenteen käsite). Ilmoita proteiinirakenteiden muodostumiseen osallistuvien sidostyypit. Mitkä aminohapporadikaalit voivat osallistua hydrofobisten vuorovaikutusten, ionisten, vetysidosten muodostumiseen.
Antaa esimerkkejä.
5. Määrittele käsite "proteiinien konformationaalinen labilisuus", osoita sen olemassaolon syyt ja merkitys.
6. Laajenna seuraavan lauseen merkitystä: "Proteiinien toiminta perustuu niiden spesifiseen vuorovaikutukseen ligandin kanssa" käyttämällä termejä ja selittämällä niiden merkitys: proteiinin konformaatio, aktiivinen keskus, ligandi, komplementaarisuus, proteiinin toiminta.
7. Selitä yhden esimerkin avulla, mitä domeenit ovat ja mikä niiden rooli on proteiinien toiminnassa.
ITSEVALTOTEHTÄVÄT
1. Ottelu.
Funktionaalinen ryhmä aminohapporadikaalissa:
A. Karboksyyliryhmä B. Hydroksyyliryhmä C Guanidiiniryhmä D. Tioliryhmä E. Aminoryhmä
2. Valitse oikeat vastaukset.
Aminohapot, joissa on polaarisia varautumattomia radikaaleja, ovat:
A. Cis B. Asn
B. Glu G. Kolme
3. Valitse oikeat vastaukset.
Aminohapporadikaalit:
A. Tarjoa primaarirakenteen spesifisyys B. Osallistu tertiaarisen rakenteen muodostukseen
B. Proteiinin pinnalla sijaitsevat ne vaikuttavat sen liukoisuuteen D. Muodostavat aktiivisen keskuksen
D. Osallistu peptidisidosten muodostukseen
4. Valitse oikeat vastaukset.
Hydrofobisia vuorovaikutuksia voi muodostua aminohapporadikaalien välillä:
A. Tre Lay B. Pro Three
B. Met Ile G. Tir Ala D. Val Fen
5. Valitse oikeat vastaukset.
Ionisidoksia voi muodostua aminohapporadikaalien välille:
A. Gln Asp B. Apr Liz
B. Liz Glu G. Gis Asp D. Asn Apr
6. Valitse oikeat vastaukset.
Aminohapporadikaalien välille voi muodostua vetysidoksia:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Ottelu.
Proteiinirakenteen muodostukseen osallistuva sidostyyppi:
A. Ensisijainen rakenne B. Toissijainen rakenne
B. Tertiäärinen rakenne
D. Ylisekundäärinen rakenne E. Konformaatio.
1. Peptidirungon atomien väliset vetysidokset
2. Heikot sidokset aminohapporadikaalien funktionaalisten ryhmien välillä
3. Sidot aminohappojen α-amino- ja α-karboksyyliryhmien välillä
8. Valitse oikeat vastaukset. Trypsiini:
A. Proteolyyttinen entsyymi B. Sisältää kaksi domeenia
B. Hydrolysoi tärkkelystä
D. Aktiivinen paikka sijaitsee verkkotunnusten välissä. D. Koostuu kahdesta polypeptidiketjusta.
9. Valitse oikeat vastaukset. Atropiini:
A. Neurotransmitteri
B. Asetyylikoliinin rakenteellinen analogi
B. Vuorovaikuttaa H-kolinergisten reseptorien kanssa
D. Vahvistaa hermoimpulssien johtumista kolinergisten synapsien kautta
D. M-kolinergisten reseptorien kilpaileva estäjä
10. Valitse oikeat väitteet. Proteiinissa:
V. Ensisijainen rakenne sisältää tietoa aktiivisen sivustonsa rakenteesta
B. Aktiivinen keskus muodostuu primäärirakenteen tasolle
B. Konformaatio on jäykästi kiinnitetty kovalenttisilla sidoksilla
D. Aktiivinen kohta voi olla vuorovaikutuksessa samanlaisten ligandien ryhmän kanssa
johtuu proteiinien D konformationaalisesta labiiteetista. Muutokset ympäristössä voivat vaikuttaa aktiivisen proteiinin affiniteettiin
keskustasta ligandiin
1. 1-B, 2-G, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
PERUSKÄSITTEET JA KÄSITTEET
1. Proteiini, polypeptidi, aminohapot
2. Primaariset, sekundaariset, tertiääriset proteiinirakenteet
3. Konformaatio, natiivi proteiinin konformaatio
4. Proteiinin kovalenttiset ja heikot sidokset
5. Konformaatiolabiteetti
6. Proteiinin aktiivinen kohta
7. Ligandit
8. Proteiinin taittaminen
9. Ligandien rakenteelliset analogit
10. Domain-proteiinit
11. Yksinkertaiset ja monimutkaiset proteiinit
12. Proteiinien denaturointi, denaturointiaineet
13. Proteiinien uudelleenaktivointi
Ratkaista ongelmia
"Proteiinien rakenneorganisaatio ja niiden toiminnan perusta"
1. Proteiinin - hemoglobiini A (HbA) - päätehtävä on hapen kuljettaminen kudoksiin. Ihmispopulaatiossa tunnetaan tämän proteiinin useita muotoja, joiden ominaisuudet ja toiminta ovat muuttuneet – niin sanotut epänormaalit hemoglobiinit. Esimerkiksi hemoglobiini S:llä, jota löytyy sirppisolusairautta (HbS) sairastavien potilaiden punasoluista, on havaittu olevan alhainen liukoisuus olosuhteissa, joissa hapen osapaine on alhainen (kuten laskimoveren tapauksessa). Tämä johtaa tämän proteiinin aggregaattien muodostumiseen. Proteiini menettää toimintansa, saostuu ja punasolut muuttuvat epäsäännöllisiksi (jotkut niistä muodostavat sirpin muodon) ja tuhoutuvat tavallista nopeammin pernassa. Seurauksena kehittyy sirppisoluanemia.
Ainoa ero HbA:n primäärirakenteessa havaittiin hemoglobiinin p-ketjun N-pään alueella. Vertaa β-juosteen N-terminaalisia alueita ja näytä kuinka muutokset proteiinin primäärirakenteessa vaikuttavat sen ominaisuuksiin ja toimintoihin.
 Tätä varten:
Tätä varten:
a) Kirjoita aminohappojen kaavat, joilla HbA eroaa, ja vertaa näiden aminohappojen ominaisuuksia (polariteetti, varaus).
b) tehdä johtopäätös siitä, miksi liukoisuus heikkenee ja hapen kuljetus kudoksiin häiriintyy.
2. Kuvassa on kaavio proteiinin rakenteesta, jossa on sitoutumiskeskus ligandin kanssa (aktiivinen keskus). Selitä, miksi proteiini on valikoiva ligandin valinnassa. Tätä varten:
a) muista mikä on proteiinin aktiivinen keskus ja tarkastele kuvassa näkyvän proteiinin aktiivisen keskuksen rakennetta;
b) kirjoittaa aktiivisen keskuksen muodostavien aminohapporadikaalien kaavat;
c) piirtää ligandi, joka voisi spesifisesti olla vuorovaikutuksessa proteiinin aktiivisen kohdan kanssa. Merkitse siihen funktionaaliset ryhmät, jotka voivat muodostaa sidoksia aminohapporadikaalien kanssa, jotka muodostavat aktiivisen keskuksen;
d) osoittavat sidostyypit, joita syntyy aktiivisen keskuksen ligandin ja aminohapporadikaalien välillä;
e) selittää, mihin proteiini-ligandi-vuorovaikutuksen spesifisyys perustuu.
 3.
Kuvassa näkyy proteiinin aktiivinen kohta ja useita ligandeja.
3.
Kuvassa näkyy proteiinin aktiivinen kohta ja useita ligandeja.
Määritä, mikä ligandi on todennäköisimmin vuorovaikutuksessa proteiinin aktiivisen kohdan kanssa ja miksi.
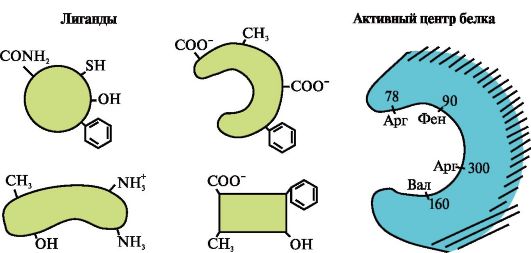 Millaisia sidoksia syntyy proteiini-ligandikompleksin muodostumisen aikana?
Millaisia sidoksia syntyy proteiini-ligandikompleksin muodostumisen aikana?
4. Luonnollisten proteiiniligandien rakenteellisia analogeja voidaan käyttää lääkkeinä proteiinien aktiivisuuden modifioimiseksi.
Asetyylikoliini on hermo-muskulaaristen synapsien viritysvälityksen välittäjä. Kun asetyylikoliini on vuorovaikutuksessa proteiinien kanssa - luurankolihasten postsynaptisen kalvon reseptorit, ionikanavat avautuvat ja lihasten supistuminen tapahtuu. Ditiliin on lääke, jota käytetään joissakin leikkauksissa lihasten rentouttamiseen, koska se häiritsee hermoimpulssien siirtymistä hermo-lihassynapsien kautta. Selitä ditiilin vaikutusmekanismi lihasrelaksanttina. Tätä varten:
a) kirjoittaa asetyylikoliinin ja ditiliinin kaavat ja vertailla niiden rakenteita;
b) kuvaile ditiilin rentouttavan vaikutuksen mekanismia.
5. Joissakin sairauksissa potilaan ruumiinlämpö nousee, mitä pidetään kehon suojaavana reaktiona. Korkeat lämpötilat ovat kuitenkin haitallisia kehon proteiineille. Selitä, miksi yli 40 °C:n lämpötiloissa proteiinien toiminta häiriintyy ja ihmishenkiä uhkaa. Muista tehdäksesi tämän:
1) Proteiinien rakenne ja sidokset, jotka pitävät sen rakenteen alkuperäisessä konformaatiossa;
2) Miten proteiinien rakenne ja toiminta muuttuvat lämpötilan noustessa?;
3) Mikä on homeostaasi ja miksi se on tärkeää ihmisten terveyden ylläpitämiselle.
Moduuliyksikkö 2 OLIGOMERICK PROTEINIT SÄÄNTELYN VAIKUTTAJIEN KOHTEINA. PROTEIINIEN RAKENNE JA TOIMINNALLINEN MONIMUOTOisuus. PROTEIINIEN EROTUS- JA PUHDISTUSMENETELMÄT
Oppimistavoitteet Pystyy:
1. Käytä tietoa oligomeeristen proteiinien rakenteen ja toiminnan ominaisuuksista ymmärtääksesi niiden toimintojen säätelyn adaptiivisia mekanismeja.
2. Selitä kaperonien rooli proteiinien konformaation synteesissä ja ylläpitämisessä soluolosuhteissa.
3. Selitä elämän ilmenemismuotojen monimuotoisuus kehossa syntetisoitujen proteiinien rakenteiden ja toimintojen moninaisuuden avulla.
4. Analysoi proteiinien rakenteen ja niiden toiminnan välistä suhdetta käyttämällä esimerkkejä vastaavien hemoproteiinien - myoglobiinin ja hemoglobiinin - vertailusta sekä immunoglobuliiniperheen viiden proteiiniluokan edustajista.
5. Soveltaa tietoa proteiinien fysikaalisten ja kemiallisten ominaisuuksien erityispiirteistä valitakseen menetelmiä niiden puhdistamiseksi muista proteiineista ja epäpuhtauksista.
6. Tulkitse veriplasman proteiinien kvantitatiivisen ja laadullisen koostumuksen tuloksia kliinisen diagnoosin vahvistamiseksi tai selkeyttämiseksi.
Tietää:
1. Oligomeeristen proteiinien rakenteen ominaisuudet ja adaptiiviset mekanismit niiden toimintojen säätelyyn hemoglobiinin esimerkin avulla.
2. Chaperonien rakenne ja toiminnot sekä niiden merkitys proteiinien natiivin konformaation ylläpitämisessä soluolosuhteissa.
3. Periaatteet proteiinien yhdistämisestä perheiksi niiden konformaation ja toimintojen samankaltaisuuden perusteella käyttäen esimerkkiä immunoglobuliinista.
4. Menetelmät proteiinien erottamiseksi niiden fysikaalis-kemiallisten ominaisuuksien ominaisuuksien perusteella.
5. Veriplasman elektroforeesi menetelmänä proteiinien laadullisen ja kvantitatiivisen koostumuksen arvioimiseksi.
AIHE 1.4. OLIGOMEERISTEN PROTEIINIEN RAKENTEEN JA TOIMINNAN OMINAISUUDET KÄYTTÄEN HEMOGLOBIIIN ESIMERKKIÄ
1. Monet proteiinit sisältävät useita polypeptidiketjuja. Tällaisia proteiineja kutsutaan oligomeerinen, ja yksittäiset ketjut - protomeerit. Oligomeeristen proteiinien protomeerit on yhdistetty monilla heikoilla ei-kovalenttisilla sidoksilla (hydrofobinen, ioninen, vety). Vuorovaikutus
protomeerit suoritetaan ansiosta täydentävyyttä niiden koskettavat pinnat.
Oligomeeristen proteiinien protomeerien määrä voi vaihdella suuresti: hemoglobiini sisältää 4 protomeeriä, aspa12 protomeeriä ja tupakan mosaiikkiviruksen proteiini sisältää 2120 ei-kovalenttisilla sidoksilla yhdistettyä protomeeriä. Tästä johtuen oligomeeristen proteiinien molekyylipainot voivat olla erittäin korkeat.
Yhden protomeerin vuorovaikutusta muiden kanssa voidaan pitää proteiini-ligandivuorovaikutuksen erikoistapauksena, koska jokainen protomeeri toimii ligandina muille protomeereille. Proteiinin protomeerien lukumäärää ja yhdistämismenetelmää kutsutaan kvaternäärinen proteiinirakenne.
Proteiinit voivat sisältää protomeerejä, joilla on sama tai erilainen rakenne, esimerkiksi homodimeerit ovat proteiineja, jotka sisältävät kaksi identtistä protomeeriä, ja heterodimeerit ovat proteiineja, jotka sisältävät kaksi eri protomeeriä.
Jos proteiinit sisältävät erilaisia protomeerejä, niihin voi muodostua sitoutumiskeskuksia erilaisilla ligandeilla, jotka ovat rakenteeltaan erilaisia. Kun ligandi sitoutuu aktiiviseen kohtaan, tämän proteiinin toiminta ilmenee. Eri protomeerissä sijaitsevaa keskustaa kutsutaan allosteeriseksi (eri kuin aktiivinen). Yhteydenotto allosteerinen ligandi tai efektori, se suorittaa säätelytoiminnon (kuva 1.18). Allosteerisen keskuksen vuorovaikutus efektorin kanssa aiheuttaa konformaatiomuutoksia koko oligomeerisen proteiinin rakenteessa sen konformationaalisuuden vuoksi. Tämä vaikuttaa aktiivisen kohdan affiniteettiin spesifiseen ligandiin ja säätelee kyseisen proteiinin toimintaa. Muutosta kaikkien protomeerien konformaatiossa ja toiminnassa oligomeerisen proteiinin vuorovaikutuksessa vähintään yhden ligandin kanssa kutsutaan yhteistoiminnallisiksi konformaatiomuutoksiksi. Proteiinin toimintaa tehostavia efektejä kutsutaan aktivaattorit, ja efektorit, jotka estävät sen toimintaa - estäjät.
Siten oligomeerisillä proteiineilla, samoin kuin proteiineilla, joilla on domeenirakenne, on uusi ominaisuus verrattuna monomeerisiin proteiineihin - kyky allosteerisesti säädellä toimintoja (säätely kiinnittämällä proteiiniin erilaisia ligandeja). Tämä voidaan nähdä vertaamalla kahden läheisesti toisiinsa liittyvän monimutkaisen proteiinin, myoglobiinin ja hemoglobiinin, rakenteita ja toimintoja.
 Riisi. 1.18. Kaavio dimeerisen proteiinin rakenteesta
Riisi. 1.18. Kaavio dimeerisen proteiinin rakenteesta
2. Avaruusrakenteiden muodostuminen ja myoglobiinin toiminta.
Myoglobiini (Mb) on punaisissa lihaksissa esiintyvä proteiini, jonka päätehtävänä on luoda intensiiviseen lihastyöhön tarvittavia O 2 -varastoja. Mb on monimutkainen proteiini, joka sisältää proteiiniosan - apoMb:n ja ei-proteiiniosan - hemin. ApoMB:n primäärirakenne määrää sen kompaktin pallomaisen konformaation ja aktiivisen keskuksen rakenteen, johon myoglobiinin ei-proteiiniosa, hemi, on kiinnittynyt. Verestä lihaksiin tuleva happi sitoutuu myoglobiinin Fe+ 2 -hemeihin. Mb on monomeerinen proteiini, jolla on erittäin korkea affiniteetti O 2:een, joten myoglobiinin hapen vapautuminen tapahtuu vain intensiivisen lihastyön aikana, kun O 2:n osapaine laskee jyrkästi.
Mv-konformaation muodostuminen. Punaisissa lihaksissa, ribosomeissa translaation aikana, syntetisoidaan MB:n primäärirakenne, jota edustaa spesifinen 153 aminohappotähteen sekvenssi. Mb:n toissijainen rakenne sisältää kahdeksan α-heliksiä, joita kutsutaan latinalaisilla kirjaimilla A:sta H:hen, joiden välissä on ei-kierteisiä alueita. Mb:n tertiäärinen rakenne on muodoltaan kompakti palloma, jonka syvennyksessä aktiivinen keskus sijaitsee F- ja E α-heliksien välissä (kuva 1.19).
 Riisi. 1.19. Myoglobiinin rakenne
Riisi. 1.19. Myoglobiinin rakenne
3. MV:n aktiivisen keskuksen rakenteen ja toiminnan piirteet. Mb:n aktiivinen keskus muodostuu pääasiassa hydrofobisista aminohapporadikaaleista, jotka ovat kaukana toisistaan primäärirakenteessa (esim. 3 9 ja Fen 138) Heikosti veteen liukeneva ligandit - hemi ja O 2 - kiinnittyvät aktiiviseen keskustaan. Hemi on apoMB:n spesifinen ligandi (kuva 1.20), jonka perusta koostuu neljästä pyrrolirenkaasta, jotka on yhdistetty metenyylisilloilla; keskellä on Fe+ 2 -atomi, joka on yhdistetty pyrrolirenkaiden typpiatomeihin neljällä koordinaatiosidoksella. Mb:n aktiivisessa keskustassa on hydrofobisten aminohapporadikaalien lisäksi myös kahden aminohapon tähteitä, joissa on hydrofiilisiä radikaaleja - Gis E 7(Gis 64) ja GIS F 8(Hänen 93) (Kuva 1.21).
 Riisi. 1.20. Hemin rakenne - myoglobiinin ja hemoglobiinin ei-proteiiniosa
Riisi. 1.20. Hemin rakenne - myoglobiinin ja hemoglobiinin ei-proteiiniosa
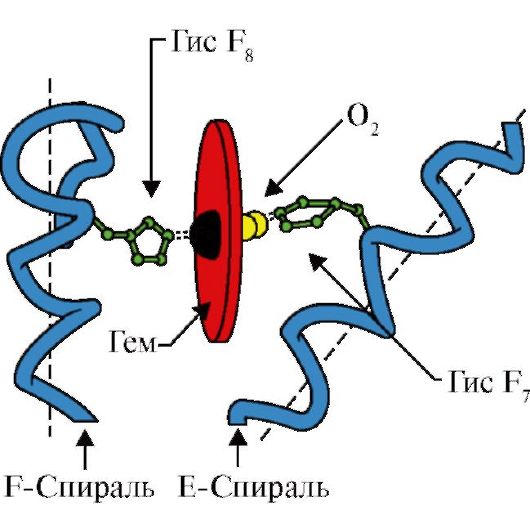 Riisi. 1.21. Hemin ja O2:n sijainti apomyoglobiinin ja hemoglobiinin protomeerien aktiivisessa kohdassa
Riisi. 1.21. Hemin ja O2:n sijainti apomyoglobiinin ja hemoglobiinin protomeerien aktiivisessa kohdassa
Hemi on kovalenttisesti sidottu Hänen F8:aan rautaatomin kautta. O 2 kiinnittyy rautaan hemitason toisella puolella. Hänen E 7 on välttämätön O 2:n oikealle suuntaukselle ja helpottaa hapen lisäämistä Fe + 2 -heemiin
GIS F 8 muodostaa koordinaatiosidoksen Fe+ 2:n kanssa ja kiinnittää hemin tiukasti aktiiviseen keskustaan. Gis E 7 tarvitaan oikean suuntautumiseen toisen ligandin - O 2:n aktiivisessa keskustassa sen vuorovaikutuksessa Fe + 2 -hemin kanssa. Hemin mikroympäristö luo olosuhteet O 2:n vahvalle mutta palautuvalle sitoutumiselle Fe + 2:een ja estää vettä pääsemästä hydrofobiseen aktiiviseen kohtaan, mikä voi johtaa sen hapettumiseen Fe + 3:ksi.
Mb:n ja sen aktiivisen keskuksen monomeerinen rakenne määrää proteiinin korkean affiniteetin 02:lle.
4. Hb:n oligomeerinen rakenne ja Hb:n affiniteetin säätely 02-ligandeihin. Ihmisen hemoglobiinit- proteiiniperhe, kuten myoglobiini, joka liittyy monimutkaisiin proteiineihin (hemoproteiineihin). Niillä on tetrameerinen rakenne ja ne sisältävät kaksi a-ketjua, mutta eroavat kahden muun polypeptidiketjun (2a-, 2x-ketjut) rakenteesta. Toisen polypeptidiketjun rakenne määrittää näiden Hb-muotojen toiminnan piirteet. Noin 98 % aikuisen punasolujen hemoglobiinista on hemoglobiini A(2a-, 2p-ketjut).
Sikiön kehityksen aikana kaksi hemoglobiinityyppiä toimii: alkion Hb(2α, 2ε), joka löytyy sikiön kehityksen alkuvaiheista, ja hemoglobiini F (sikiö)- (2α, 2γ), joka korvaa varhaisen sikiön hemoglobiinin kohdunsisäisen kehityksen kuudennessa kuukaudessa ja vasta syntymän jälkeen korvataan Hb A:lla.
HB A on myoglobiiniin (MB) liittyvä proteiini, jota löytyy aikuisen ihmisen punasoluista. Sen yksittäisten protomeerien rakenne on samanlainen kuin myoglobiinin. Myoglobiinin ja hemoglobiinin protomeerien sekundääriset ja tertiaariset rakenteet ovat hyvin samankaltaisia huolimatta siitä, että niiden polypeptidiketjujen primäärirakenteessa vain 24 aminohappotähdettä on identtisiä (hemoglobiinin protomeerien sekundaarirakenne, kuten myoglobiini, sisältää kahdeksan α-heliksiä, merkitty latinalaisilla kirjaimilla A:sta H:hen, ja tertiäärinen rakenne on muodoltaan tiivis pallo). Mutta toisin kuin myoglobiinilla, hemoglobiinilla on oligomeerinen rakenne, joka koostuu neljästä polypeptidiketjusta, jotka on yhdistetty ei-kovalenttisilla sidoksilla (kuva 1.22).
Jokainen Hb-protomeeri liittyy ei-proteiiniosaan - hemiin ja viereisiin protomeereihin. Hb:n proteiiniosan yhteys hemiin on samanlainen kuin myoglobiinin: proteiinin aktiivisessa keskustassa hemin hydrofobisia osia ympäröivät hydrofobiset aminohapporadikaalit paitsi His F 8 ja His E 7, jotka sijaitsevat molemmilla puolilla hemitasoa ja niillä on samanlainen rooli proteiinin toiminnassa ja sen sitoutumisessa happeen (katso myoglobiinin rakenne).
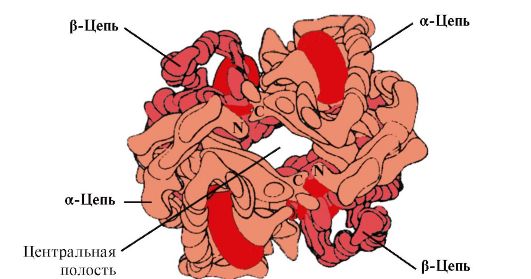 Riisi. 1.22. Hemoglobiinin oligomeerinen rakenne
Riisi. 1.22. Hemoglobiinin oligomeerinen rakenne
Sitä paitsi, Gis E 7 suorittaa tärkeän lisärooli Nv:n toiminnassa. Vapaalla hemellä on 25 000 kertaa suurempi affiniteetti CO:lle kuin O2:lle. CO:ta muodostuu elimistössä pieniä määriä, ja sen korkea affiniteetti heemille voi häiritä solun elämälle välttämättömän O 2:n kuljetusta. Hemoglobiinin koostumuksessa hemin affiniteetti hiilimonoksidiin ylittää kuitenkin affiniteetin O 2:lle vain 200 kertaa, koska aktiivisessa keskustassa on His E 7. Loppuosa tästä aminohaposta luo optimaaliset olosuhteet hemin sitoutumiselle O 2:een ja heikentää hemin vuorovaikutusta CO:n kanssa.
5. HB:n päätehtävä on O2:n kuljettaminen keuhkoista kudoksiin. Toisin kuin monomeerinen myoglobiini, jolla on erittäin korkea affiniteetti O2:een ja joka varastoi happea punaisiin lihaksiin, hemoglobiinin oligomeerinen rakenne tarjoaa:
1) HB:n nopea kyllästyminen hapella keuhkoissa;
2) HB:n kyky vapauttaa happea kudoksissa suhteellisen korkeassa 02:n osapaineessa (20-40 mm Hg);
3) mahdollisuus säädellä Hb:n affiniteettia O 2:een.
6. Yhteistoiminnalliset muutokset hemoglobiinin protomeerien konformaatiossa nopeuttavat O 2:n sitoutumista keuhkoissa ja sen vapautumista kudoksiin. Keuhkoissa 02:n korkea osapaine edistää sen sitoutumista Hb:hen neljän protomeerin (2α ja 2β) aktiivisessa kohdassa. Kunkin protomeerin aktiivinen keskus, kuten myoglobiinissa, sijaitsee kahden α-kierteen (F ja E) välissä hydrofobisessa taskussa. Se sisältää ei-proteiiniosan - hemin, joka on kiinnittynyt proteiiniosaan monilla heikkoilla hydrofobisilla vuorovaikutuksilla ja yhden vahvan sidoksen Fe 2 + -hemin ja His F 8:n välillä (katso kuva 1.21).
Deoksihemoglobiinissa, johtuen tästä sidoksesta His F 8:n kanssa, Fe 2+ -atomi työntyy esiin hemitasosta kohti histidiiniä. O2:n sitoutuminen Fe2+:aan tapahtuu hemin toisella puolella His E7 -alueella käyttämällä yhtä vapaata koordinaatiosidosta. His E 7 tarjoaa optimaaliset olosuhteet O 2:n sitoutumiselle hemiraudaan.
O 2:n lisääminen yhden protomeerin Fe + 2 -atomiin aiheuttaa sen siirtymisen hemitasoon, jota seuraa siihen liittyvä histidiinijäännös
 Riisi. 1.23. Muutos hemoglobiinin protomeerin konformaatiossa yhdistettynä O 2:een
Riisi. 1.23. Muutos hemoglobiinin protomeerin konformaatiossa yhdistettynä O 2:een
Tämä johtaa muutokseen kaikkien polypeptidiketjujen konformaatiossa niiden konformationaalisuuden vuoksi. Muiden ketjujen konformaation muuttaminen helpottaa niiden vuorovaikutusta myöhempien O 2 -molekyylien kanssa.
Neljäs O 2 -molekyyli kiinnittyy hemoglobiiniin 300 kertaa helpommin kuin ensimmäinen (kuva 1.24).
 Riisi. 1.24. Yhteistoiminnalliset muutokset hemoglobiinin protomeerien konformaatiossa sen vuorovaikutuksessa O2:n kanssa
Riisi. 1.24. Yhteistoiminnalliset muutokset hemoglobiinin protomeerien konformaatiossa sen vuorovaikutuksessa O2:n kanssa
Kudoksissa jokainen seuraava O 2 -molekyyli lohkeaa helpommin kuin edellinen, myös protomeerien konformaation yhteistoiminnallisten muutosten vuoksi.
7. Orgaanisten aineiden hajoamisen aikana muodostuneet CO 2 ja H+ vähentävät hemoglobiinin affiniteettia O 2:lle suhteessa pitoisuuteensa. Solujen toimintaan tarvittava energia tuotetaan pääasiassa mitokondrioissa orgaanisten aineiden hapettumisen aikana hemoglobiinin keuhkoista kuljettaman O 2:n avulla. Orgaanisten aineiden hapettumisen seurauksena muodostuu niiden hajoamisen lopputuotteita: CO 2 ja K 2 O, joiden määrä on verrannollinen käynnissä olevien hapetusprosessien intensiteettiin.
CO 2 diffundoituu soluista vereen ja tunkeutuu punasoluihin, joissa se muuttuu karbanhydraasientsyymin vaikutuksesta hiilihapoksi. Tämä heikko happo hajoaa protoniksi ja bikarbonaatti-ioniksi.
H+ pystyvät liittymään Hänen radikaaleihinsa 14 6 hemoglobiinin a- ja p-ketjuissa, ts. alueilla, jotka ovat kaukana hemistä. Hemoglobiinin protonoituminen vähentää sen affiniteettia O 2:een, edistää O 2:n poistumista oksiHb:stä, deoksiHb:n muodostumista ja lisää kudosten hapen saantia suhteessa muodostuneiden protonien määrään (kuva 1.25).
Vapautuneen hapen määrän lisääntymistä punasolujen H+-pitoisuuden kasvusta riippuen kutsutaan Bohrin efektiksi (nimetty tanskalaisen fysiologin Christian Bohrin mukaan, joka löysi tämän vaikutuksen ensimmäisenä).
Hapen korkea osapaine keuhkoissa edistää sen sitoutumista deoksiHb:hen, mikä vähentää proteiinin affiniteettia H+:aan. Hiilihapon vaikutuksesta vapautuneet protonit reagoivat bikarbonaattien kanssa muodostaen CO 2:ta ja H 2 O:ta
 Riisi. 1.25. Hb:n O 2 -affiniteetin riippuvuus CO 2:n ja protonien pitoisuudesta (Bohr-ilmiö):
Riisi. 1.25. Hb:n O 2 -affiniteetin riippuvuus CO 2:n ja protonien pitoisuudesta (Bohr-ilmiö):
A- CO 2- ja H+-konsentraation vaikutus 02:n vapautumiseen HB-kompleksista (Bohr-ilmiö); B- deoksihemoglobiinin hapetus keuhkoissa, hiilidioksidin muodostuminen ja vapautuminen.
Tuloksena oleva CO 2 tulee keuhkorakkuloihin ja poistuu uloshengitetyn ilman mukana. Siten hemoglobiinin vapauttaman hapen määrää kudoksissa säätelevät orgaanisten aineiden hajoamistuotteet: mitä voimakkaammin aineet hajoavat esimerkiksi fyysisen harjoituksen aikana, sitä korkeampi on CO 2 - ja H + -pitoisuus ja sitä enemmän happea. kudokset saavat Hb:n affiniteetin heikkenemisen seurauksena O 2:ta kohtaan.
8. Hb:n O2:n affiniteetin allosteerinen säätely ligandilla - 2,3-bisfosfoglyseraatilla. Punasoluissa hemoglobiinin allosteerinen ligandi, 2,3-bisfosfoglyseraatti (2,3-BPG), syntetisoidaan glukoosin hapettumistuotteesta - 1,3-bisfosfoglyseraatista. Normaaleissa olosuhteissa 2,3-BPG:n pitoisuus on korkea ja verrattavissa Hb-pitoisuuteen. 2,3-BPG:llä on vahva negatiivinen varaus -5.
 Kudoskapillaareissa oleva bisfosfoglyseraatti, joka sitoutuu deoksihemoglobiiniin, lisää hapen vapautumista kudoksiin, mikä vähentää Hb:n affiniteettia O 2:een.
Kudoskapillaareissa oleva bisfosfoglyseraatti, joka sitoutuu deoksihemoglobiiniin, lisää hapen vapautumista kudoksiin, mikä vähentää Hb:n affiniteettia O 2:een.
Tetrameerisen hemoglobiinimolekyylin keskellä on ontelo. Se muodostuu kaikkien neljän protomeerin aminohappotähteistä (katso kuva 1.22). Kudoskapillaareissa Hb:n protonoituminen (Bohr-ilmiö) johtaa hemiraudan ja O2:n välisen sidoksen katkeamiseen. Molekyylissä
deoksihemoglobiini, verrattuna oksihemoglobiiniin, ilmaantuu ylimääräisiä ionisidoksia, jotka yhdistävät protomeerit, minkä seurauksena keskusontelon mitat kasvavat oksihemoglobiiniin verrattuna. Keskusontelo on paikka, jossa 2,3-BPG kiinnittyy hemoglobiiniin. Keskusontelon koon eroista johtuen 2,3-BPG voi kiinnittyä vain deoksihemoglobiiniin.
2,3-BPG on vuorovaikutuksessa hemoglobiinin kanssa paikassa, joka on kaukana proteiinin aktiivisista keskuksista ja kuuluu allosteerinen(säätely) ligandit, ja Hb:n keskusontelo on allosteerinen keskus. 2,3-BPG:llä on voimakas negatiivinen varaus ja se on vuorovaikutuksessa Hb:n kahden β-ketjun viiden positiivisesti varautuneen ryhmän kanssa: Val:n N-terminaalisen α-aminoryhmän ja Lys 82 His 143 -radikaalin (kuva 1.26).
 Riisi. 1.26. BPG deoksihemoglobiinin keskusontelossa
Riisi. 1.26. BPG deoksihemoglobiinin keskusontelossa
BPG sitoutuu kolmeen positiivisesti varautuneeseen ryhmään kussakin β-juosteessa.
Kudoskapillaareissa muodostuva deoksihemoglobiini on vuorovaikutuksessa 2,3-BPG:n kanssa ja β-ketjujen positiivisesti varautuneiden radikaalien ja negatiivisesti varautuneen ligandin välille muodostuu ionisidoksia, jotka muuttavat proteiinin konformaatiota ja vähentävät Hb:n affiniteettia O2:een. . Hb:n affiniteetin heikkeneminen 02:lle edistää O 2:n tehokkaampaa vapautumista kudokseen.
Keuhkoissa korkeassa osapaineessa happi on vuorovaikutuksessa Hb:n kanssa liittyen hemiraudaan; tässä tapauksessa proteiinin konformaatio muuttuu, keskusontelo pienenee ja 2,3-BPG siirtyy allosteerisesta keskustasta
Siten oligomeerisillä proteiineilla on uusia ominaisuuksia verrattuna monomeerisiin proteiineihin. Ligandien kiinnittyminen paikkoihin
avaruudellisesti kaukana toisistaan (allosteerinen), voivat aiheuttaa konformaatiomuutoksia koko proteiinimolekyylissä. Vuorovaikutuksesta säätelyligandien kanssa tapahtuu muutos konformaatiossa ja proteiinimolekyylin toiminnan mukautuminen ympäristön muutoksiin.
AIHE 1.5. PROTEIINIEN NATIIVIN KONFORMAATION YLLÄPITO SOLU-OLLOISSA
Soluissa polypeptidiketjujen synteesin aikana niiden kulkeutuminen kalvojen läpi solun vastaaviin osiin, laskostumisprosessin aikana (natiivikonformaation muodostuminen) ja oligomeeristen proteiinien kokoamisen aikana sekä niiden toiminnan aikana, välituote , aggregaatiolle alttiita, epästabiileja konformaatioita syntyy proteiinirakenteessa. Hydrofobiset radikaalit, jotka ovat yleensä piilossa proteiinimolekyylin sisällä natiivissa konformaatiossa, ilmestyvät pinnalle epävakaassa konformaatiossa ja pyrkivät yhdistymään muiden proteiinien ryhmiin, jotka ovat huonosti veteen liukenevia. Kaikkien tunnettujen organismien soluista on löydetty erityisiä proteiineja, jotka varmistavat soluproteiinien optimaalisen laskostumisen, stabiloivat niiden natiivia konformaatiota toiminnan aikana ja mikä tärkeintä, ylläpitävät solunsisäisten proteiinien rakennetta ja toimintoja homeostaasin häiriintyessä. Näitä proteiineja kutsutaan "seuraajat" joka tarkoittaa ranskaksi lastenhoitajaa.
1. Molekyylikaperonit ja niiden rooli proteiinien denaturoitumisen estämisessä.
Chaperonit (CH) luokitellaan niiden alayksiköiden massan mukaan. Suuren molekyylipainon omaavien chaperonien massa on 60 - 110 kDa. Niistä kolmea luokkaa on eniten tutkittu: Sh-60, Sh-70 ja Sh-90. Jokainen luokka sisältää sukulaisia proteiineja. Siten Sh-70 sisältää proteiineja, joiden molekyylipaino on 66 - 78 kDa. Pienen molekyylipainon omaavien chaperonien molekyylipaino on 40 - 15 kDa.
Ohjaajien joukossa on konstitutiivinen proteiineja, joiden korkea perussynteesi ei riipu stressin vaikutuksista kehon soluihin, ja indusoituva, jonka synteesi normaaleissa olosuhteissa on heikkoa, mutta lisääntyy voimakkaasti rasituksessa. Indusoituvia chaperoneja kutsutaan myös "lämpösokkiproteiineiksi", koska ne löydettiin ensimmäisen kerran korkeille lämpötiloille altistetuista soluista. Soluissa proteiinien suuren pitoisuuden vuoksi osittain denaturoituneiden proteiinien spontaani renativaatio on vaikeaa. Sh-70 voi estää denaturaation alkamista ja auttaa palauttamaan proteiinien alkuperäisen konformaation. Molekyyliset chaperonit-70- erittäin konservoitunut proteiiniluokka, jota löytyy solun kaikista osista: sytoplasmasta, tumasta, endoplasmisesta retikulumista, mitokondrioista. Yhden polypeptidiketjun Sh-70 karboksyylipäässä on alue, joka on ura, joka pystyy olemaan vuorovaikutuksessa pitkien peptidien kanssa.
7 - 9 aminohappotähdettä, jotka on rikastettu hydrofobisilla radikaaleilla. Tällaisia alueita pallomaisissa proteiineissa esiintyy suunnilleen joka 16. aminohappo. Sh-70 pystyy suojaamaan proteiineja lämpötilan inaktivoitumiselta ja palauttamaan osittain denaturoituneiden proteiinien konformaatiota ja aktiivisuutta.
2. Chaperonien rooli proteiinien laskostumisessa. Proteiinisynteesin aikana ribosomissa polypeptidin N-terminaalinen alue syntetisoituu ennen C-terminaalista aluetta. Luonnollisen konformaation muodostamiseksi tarvitaan proteiinin täydellinen aminohapposekvenssi. Proteiinisynteesiprosessissa kaperonit-70 pystyvät aktiivisen keskuksensa rakenteesta johtuen sulkemaan polypeptidin alueita, jotka ovat alttiita aggregaatiolle ja jotka ovat rikastuneet hydrofobisilla aminohapporadikaaleilla, kunnes synteesi on valmis (Kuva 1.27, A ).
 Riisi. 1.27. Chaperonien osallistuminen proteiinien laskostukseen
Riisi. 1.27. Chaperonien osallistuminen proteiinien laskostukseen
A - chaperons-70:n osallistuminen hydrofobisten vuorovaikutusten estämiseen syntetisoidun polypeptidin osien välillä; B - proteiinin luonnollisen konformaation muodostuminen chaperonikompleksissa
Monet korkeamolekyyliset proteiinit, joilla on monimutkainen konformaatio, kuten domeenirakenne, laskostuvat Sh-60:n muodostamaan erityiseen tilaan. Ш-60 toimivat oligomeerisena kompleksina, joka koostuu 14 alayksiköstä. Ne muodostavat kaksi onttoa rengasta, joista jokainen koostuu seitsemästä alayksiköstä, nämä renkaat ovat yhteydessä toisiinsa. Jokainen Sh-60-alayksikkö koostuu kolmesta domeenista: apikaalinen (apikaalinen), rikastettu hydrofobisilla radikaaleilla, jotka ovat renkaan onteloa päin, väli- ja ekvatoriaalinen (kuva 1.28).
 Riisi. 1.28. 14 Ш-60 koostuvan chaperoniinikompleksin rakenne
Riisi. 1.28. 14 Ш-60 koostuvan chaperoniinikompleksin rakenne
A - sivukuva; B - ylhäältä katsottuna
Syntetisoidut proteiinit, joiden pinnalla on laskostumattomille molekyyleille ominaisia elementtejä, erityisesti hydrofobisia radikaaleja, tulevat chaperonirenkaiden onteloon. Näiden onteloiden tietyssä ympäristössä mahdollisia konformaatioita etsitään, kunnes löydetään ainoa, joka on energeettisesti edullisin (kuva 1.27, B). Konformaatioiden muodostumiseen ja proteiinin vapautumiseen liittyy ATP-hydrolyysi ekvatoriaalisella alueella. Tyypillisesti tällainen chaperone-riippuvainen taittaminen vaatii huomattavan määrän energiaa.
Sen lisäksi, että chaperonit osallistuvat proteiinien kolmiulotteisen rakenteen muodostukseen ja osittain denaturoituneiden proteiinien renativoimiseen, ne ovat välttämättömiä myös perusprosessien, kuten oligomeeristen proteiinien kokoamisen, denaturoituneiden proteiinien tunnistamisen ja kuljetuksen lysosomeihin, esiintymiseen, proteiinien kuljettaminen kalvojen läpi ja osallistuminen proteiinikompleksien toiminnan säätelyyn.
AIHE 1.6. ERILAISIA PROTEIINEJA. PROTEIINIPERHEET: ESIMERKKI IMMUNOGLOBULIINEISTA
1. Proteiineilla on ratkaiseva rooli yksittäisten solujen ja koko monisoluisen organismin elämässä, ja niiden toiminnot ovat yllättävän monipuoliset. Tämän määräävät proteiinien primäärirakenteen ja konformaatioiden ominaisuudet, aktiivisen keskuksen ainutlaatuinen rakenne ja kyky sitoa spesifisiä ligandeja.
Vain hyvin pieni osa kaikista mahdollisista peptidiketjujen varianteista voi omaksua vakaan avaruudellisen rakenteen; suurin osa
niistä voi olla useita konformaatioita suunnilleen samalla Gibbsin energialla, mutta eri ominaisuuksilla. Useimpien biologisen evoluution perusteella valittujen proteiinien primäärirakenne varmistaa yhden konformaation poikkeuksellisen stabiilisuuden, mikä määrää tämän proteiinin toiminnan ominaisuudet.
2. Proteiiniperheet. Saman biologisen lajin sisällä aminohappotähteiden substituutiot voivat johtaa erilaisten proteiinien syntymiseen, jotka suorittavat toisiinsa liittyviä tehtäviä ja joilla on homologiset aminohapposekvenssit. Tällaisilla sukulaisproteiineilla on hämmästyttävän samankaltaiset konformaatiot: a-heliksien ja/tai p-rakenteiden lukumäärä ja suhteellinen sijainti, useimmat polypeptidiketjujen käännökset ja taivutukset ovat samanlaisia tai identtisiä. Proteiinit, joilla on homologisia polypeptidiketjun alueita, samanlainen konformaatio ja vastaavat toiminnot, luokitellaan proteiiniperheisiin. Esimerkkejä proteiiniperheistä: seriiniproteinaasit, immunoglobuliiniperhe, myoglobiiniperhe.
Seriiniproteinaasit- proteiiniperhe, joka suorittaa proteolyyttisten entsyymien tehtävää. Näitä ovat ruoansulatusentsyymit - kymotrypsiini, trypsiini, elastaasi ja monet veren hyytymistekijät. Näillä proteiineilla on identtiset aminohapot 40 %:ssa asemistaan ja hyvin samanlainen konformaatio (kuva 1.29).

Riisi. 1.29. Elastaasin (A) ja kymotrypsiinin (B) spatiaaliset rakenteet
Jotkut aminohapposubstituutiot ovat johtaneet muutoksiin näiden proteiinien substraattispesifisyydessä ja toiminnallisen monimuotoisuuden syntymiseen perheen sisällä.
3. Immunoglobuliiniperhe. Immuunijärjestelmän toiminnassa immunoglobuliinien superperheen proteiineilla on valtava rooli, joka sisältää kolme proteiiniperhettä:
Vasta-aineet (immunoglobuliinit);
T-lymfosyyttireseptorit;
Tärkeimmän proteiinit - MHC-luokat 1 ja 2 (Major Histocompatibility Complex).
Kaikilla näillä proteiineilla on domeenirakenne, ne koostuvat homologisista immuunityyppisistä domeeneista ja suorittavat samanlaisia toimintoja: ne ovat vuorovaikutuksessa vieraiden rakenteiden kanssa, joko liuenneena vereen, imusolmukkeisiin tai solujen väliseen nesteeseen (vasta-aineet) tai solujen pinnalla (omissa tai ulkomaalainen).
4. Vasta-aineet- B-lymfosyyttien tuottamat spesifiset proteiinit vasteena vieraan rakenteen pääsylle kehoon, ns antigeeni.
Vasta-aineiden rakenteen ominaisuudet
Yksinkertaisimmat vasta-ainemolekyylit koostuvat neljästä polypeptidiketjusta: kahdesta identtisestä kevyestä - L, joka sisältää noin 220 aminohappoa, ja kahdesta identtisestä raskaasta - H, joka koostuu 440-700 aminohaposta. Kaikki neljä vasta-ainemolekyylin ketjua on yhdistetty useilla ei-kovalenttisilla sidoksilla ja neljällä disulfidisidoksella (kuva 1.30).
Vasta-aineen kevyet ketjut koostuvat kahdesta domeenista: variaabelista domeenista (VL), joka sijaitsee polypeptidiketjun N-terminaalisella alueella, ja vakiodomeenista (CL), joka sijaitsee C-päässä. Raskaissa ketjuissa on yleensä neljä domeenia: yksi muuttuja (VH), joka sijaitsee N-päässä, ja kolme vakiodomeenia (CH1, CH2, CH3) (katso kuva 1.30). Jokaisella immunoglobuliinidomeenilla on β-levyn ylärakenne, jossa kaksi kysteiinitähdettä on liitetty toisiinsa disulfidisidoksella.
Kahden vakiodomeenin CH1 ja CH2 välissä on alue, joka sisältää suuren määrän proliinitähteitä, jotka estävät sekundaarirakenteen muodostumisen ja viereisten H-ketjujen vuorovaikutuksen tässä segmentissä. Tämä sarana-alue antaa vasta-ainemolekyylille joustavuuden. Raskaan ja kevyen ketjun vaihtelevien domeenien välissä on kaksi identtistä antigeeniä sitovaa kohtaa (aktiivisia kohtia antigeenien sitomiselle), joten tällaisia vasta-aineita kutsutaan usein ns. bivalentteja. Molempien ketjujen vaihtelevien alueiden koko aminohapposekvenssi ei ole osallisena antigeenin sitoutumisessa vasta-aineeseen, vaan vain 20-30 aminohappoa, jotka sijaitsevat kunkin ketjun hypervariaabelialueilla. Juuri nämä alueet määrittävät kunkin vasta-ainetyypin ainutlaatuisen kyvyn olla vuorovaikutuksessa vastaavan komplementaarisen antigeenin kanssa.
Vasta-aineet ovat yksi kehon puolustuslinjoista tunkeutuvia vieraita organismeja vastaan. Niiden toiminta voidaan jakaa kahteen vaiheeseen: ensimmäinen vaihe on antigeenin tunnistaminen ja sitoutuminen vieraiden organismien pinnalle, mikä on mahdollista, koska vasta-ainerakenteessa on antigeeniä sitovia kohtia; toinen vaihe on antigeenin inaktivaatio- ja tuhoutumisprosessin käynnistäminen. Toisen vaiheen spesifisyys riippuu vasta-aineluokasta. Raskasketjuja on viisi luokkaa, jotka eroavat toisistaan vakiodomeenien rakenteessa: α, δ, ε, γ ja μ, joiden mukaan erotetaan viisi immunoglobuliiniluokkaa: A, D, E, G ja M.
Raskaiden ketjujen rakenteelliset ominaisuudet antavat raskaiden ketjujen sarana-alueille ja C-terminaalisille alueille kullekin luokalle ominaisen konformaation. Kun antigeeni on sitoutunut vasta-aineeseen, konformaatiomuutokset vakiodomeeneissa määrittävät antigeenin poistumisreitin.
 Riisi. 1. 30. IgG:n domeenirakenne
Riisi. 1. 30. IgG:n domeenirakenne
Immunoglobuliinit M
Immunoglobuliineilla M on kaksi muotoa.
Monomeerinen muoto- 1. luokka B-lymfosyyttien tuottamia vasta-aineita. Myöhemmin monet B-solut siirtyvät tuottamaan muita vasta-aineluokkia, mutta joilla on sama antigeenia sitova kohta. IgM on upotettu kalvoon ja toimii antigeenintunnistusreseptorina. IgM:n integroituminen solukalvoon on mahdollista, koska alueen häntäosassa on 25 hydrofobista aminohappotähdettä.
IgM:n eritysmuoto sisältää viisi monomeerista alayksikköä, jotka on liitetty toisiinsa disulfidisidoksilla, ja ylimääräisen polypeptidi-J-ketjun (kuva 1.31). Tämän muodon monomeerien raskaat ketjut eivät sisällä hydrofobista häntää. Pentameerissä on 10 antigeenin sitoutumiskohtaa, ja siksi se tunnistaa ja poistaa tehokkaasti antigeenin, joka tulee ensimmäisenä kehoon. IgM:n eritysmuoto on pääasiallinen vasta-aineiden luokka, joka erittyy vereen primaarisen immuunivasteen aikana. IgM:n sitoutuminen antigeeniin muuttaa IgM:n konformaatiota ja indusoi sen sitoutumisen komplementtijärjestelmän ensimmäiseen proteiinikomponenttiin (komplementtijärjestelmä on joukko proteiineja, jotka osallistuvat antigeenin tuhoamiseen) ja tämän järjestelmän aktivoitumisen. Jos antigeeni sijaitsee mikro-organismin pinnalla, komplementtijärjestelmä aiheuttaa solukalvon eheyden häiriöitä ja bakteerisolun kuoleman.
Immunoglobuliinit G
Määrällisesti tämä immunoglobuliiniluokka on vallitseva veressä (75 % kaikista Ig:istä). IgG - monomeerit, pääasiallinen vasta-aineiden luokka, jotka erittyvät vereen sekundaarisen immuunivasteen aikana. IgG:n vuorovaikutuksen jälkeen mikro-organismien pinta-antigeenien kanssa antigeeni-vasta-ainekompleksi pystyy sitomaan ja aktivoimaan komplementtijärjestelmän proteiineja tai voi olla vuorovaikutuksessa makrofagien ja neutrofiilien spesifisten reseptorien kanssa. Vuorovaikutus fagosyyttien kanssa johtaa
 Riisi. 1.31. IgM:n erittävän muodon rakenne
Riisi. 1.31. IgM:n erittävän muodon rakenne
antigeeni-vasta-ainekompleksien imeytymiseen ja niiden tuhoutumiseen solun fagosomeissa. IgG on ainoa vasta-aineluokka, joka pystyy tunkeutumaan istukan esteen läpi ja tarjoamaan sikiön kohdunsisäisen suojan infektioilta.
Immunoglobuliinit A
Eritteissä (maito, sylki, hengitysteiden ja suoliston eritteet) olevien vasta-aineiden pääluokka. IgA erittyy ensisijaisesti dimeerisessä muodossa, jossa monomeerit ovat liittyneet toisiinsa ylimääräisen J-ketjun kautta (kuva 1.32).
IgA ei ole vuorovaikutuksessa komplementtijärjestelmän ja fagosyyttisolujen kanssa, mutta sitoutumalla mikro-organismeihin vasta-aineet estävät niiden kiinnittymisen epiteelisoluihin ja tunkeutumisen kehoon.
Immunoglobuliinit E
Immunoglobuliineja E edustavat monomeerit, joissa raskaat e-ketjut sisältävät, kuten immunoglobuliinien M μ-ketjut, yhden vaihtelevan ja neljä vakiodomeenia. Erittymisen jälkeen IgE sitoutuu sen kanssa
 Riisi. 1.32. IgA:n rakenne
Riisi. 1.32. IgA:n rakenne
C-terminaaliset alueet, joissa on vastaavat reseptorit syöttösolujen ja basofiilien pinnalla. Tämän seurauksena niistä tulee näiden solujen pinnalla olevien antigeenien reseptoreita (kuva 1.33).
 Riisi. 1.33. IgE:n vuorovaikutus antigeenin kanssa syöttösolun pinnalla
Riisi. 1.33. IgE:n vuorovaikutus antigeenin kanssa syöttösolun pinnalla
Kun antigeeni on kiinnittynyt vastaaviin IgE:n antigeeniä sitoviin kohtiin, solut saavat signaalin erittää biologisesti aktiivisia aineita (histamiinia, serotoniinia), jotka ovat suurelta osin vastuussa tulehdusreaktion kehittymisestä ja allergisten reaktioiden, kuten esim. astma, urtikaria, heinänuha.
Immunoglobuliinit D
Immunoglobuliineja D löytyy hyvin pieninä määrinä seerumista, ne ovat monomeerejä. Raskaissa δ-ketjuissa on yksi muuttuva ja kolme vakiodomeenia. IgD:t toimivat B-lymfosyyttien reseptoreina. Muita toimintoja ei vielä tunneta. Spesifisten antigeenien vuorovaikutus B-lymfosyyttien (IgD) pinnalla olevien reseptoreiden kanssa johtaa näiden signaalien siirtymiseen soluun ja sellaisten mekanismien aktivoitumiseen, jotka varmistavat tietyn lymfosyyttikloonin lisääntymisen.
AIHE 1.7. PROTEIINIEN FYSIKAALISET JA KEMIALLISET OMINAISUUDET JA NIIDEN EROTUSMENETELMÄT
1. Yksittäiset proteiinit eroavat fysikaalis-kemiallisista ominaisuuksistaan:
Molekyylien muoto;
Molekyylipaino;
Kokonaisvaraus, jonka arvo riippuu aminohappojen anionisten ja kationisten ryhmien suhteesta;
Polaaristen ja ei-polaaristen aminohapporadikaalien suhde molekyylien pinnalla;
Erilaisten denaturointiaineiden kestävyysasteet.
2. Proteiinien liukoisuus riippuu edellä lueteltujen proteiinien ominaisuuksista sekä alustan koostumuksesta, johon proteiini on liuennut (pH-arvot, suolakoostumus, lämpötila, muiden orgaanisten aineiden läsnäolo, jotka voivat olla vuorovaikutuksessa proteiinin kanssa). Proteiinimolekyylien varausmäärä on yksi niiden liukoisuuteen vaikuttavista tekijöistä. Kun isoelektrisen pisteen varaus katoaa, proteiinit aggregoituvat ja saostuvat helpommin. Tämä on erityisen tyypillistä denaturoiduille proteiineille, joiden pinnalle ilmestyy hydrofobisia aminohapporadikaaleja.
Proteiinimolekyylin pinnalla on sekä positiivisesti että negatiivisesti varautuneita aminohapporadikaaleja. Näiden ryhmien lukumäärä ja siten proteiinien kokonaisvaraus riippuu väliaineen pH:sta, ts. H+ - ja OH - ryhmien pitoisuuksien suhde. Happamassa ympäristössä H+-konsentraation kasvu johtaa karboksyyliryhmien -COO - + H+ > -COOH dissosiaation suppressioon ja proteiinien negatiivisen varauksen vähenemiseen. Alkalisessa ympäristössä ylimääräisen OH - sitoutuminen protoneilla, jotka muodostuvat aminoryhmien -NH 3 + + OH - - NH 2 + H 2 O hajoamisen aikana veden muodostumisen kanssa, johtaa proteiinien positiivisen varauksen vähenemiseen . pH-arvoa, jossa proteiinin nettovaraus on nolla, kutsutaan isoelektrinen piste (IEP). IET:ssä positiivisesti ja negatiivisesti varautuneiden ryhmien määrä on sama, ts. proteiini on isoelektrisessä tilassa.
3. Yksittäisten proteiinien erottaminen. Kehon rakenteen ja toiminnan ominaisuudet riippuvat siinä syntetisoitujen proteiinien joukosta. Proteiinien rakenteen ja ominaisuuksien tutkiminen on mahdotonta ilman niiden eristämistä solusta ja puhdistamista muista proteiineista ja orgaanisista molekyyleistä. Yksittäisten proteiinien eristyksen ja puhdistamisen vaiheet:
Solujen tuhoutuminen tutkittava kudos ja homogenaatin saaminen.
Homogenaatin erottaminen fraktioiksi sentrifugoimalla saadaan tuma-, mitokondrio-, sytosolinen tai muu fraktio, joka sisältää halutun proteiinin.
Valikoiva lämpödenaturointi- proteiiniliuoksen lyhytaikainen kuumennus, jonka aikana osa denaturoituneista proteiiniepäpuhtauksista voidaan poistaa (jos proteiini on suhteellisen lämmönkestävää).
Suolaus pois. Eri proteiinit saostuvat eri suolapitoisuuksilla liuoksessa. Vähitellen suolapitoisuutta lisäämällä on mahdollista saada useita erillisiä fraktioita, joissa yhdessä niistä on hallitseva pitoisuus eristettyä proteiinia. Ammoniumsulfaattia käytetään useimmiten proteiinien fraktiointiin. Vähiten liukenevat proteiinit saostuvat alhaisilla suolapitoisuuksilla.
Geelisuodatus- menetelmä molekyylien seulomiseksi turvonneiden Sephadex-rakeiden läpi (kolmiulotteiset dekstraanin polysakkaridiketjut, joissa on huokoset). Nopeus, jolla proteiinit kulkevat Sephadexillä täytetyn kolonnin läpi, riippuu niiden molekyylipainosta: mitä pienempi proteiinimolekyylien massa on, sitä helpommin ne tunkeutuvat rakeisiin ja pysyvät siellä pidempään, sitä nopeammin ne eluoituvat sarakkeessa.
Ultrasentrifugointi- menetelmä, jossa proteiinit asetetaan sentrifugiputkessa ultrasentrifugin roottoriin. Kun roottori pyörii, proteiinien sedimentaationopeus on verrannollinen niiden molekyylipainoon: raskaampien proteiinien fraktiot sijaitsevat lähempänä koeputken pohjaa, kevyemmät - lähempänä pintaa.
Elektroforeesi- menetelmä, joka perustuu eroihin proteiinien liikkumisnopeudessa sähkökentässä. Tämä arvo on verrannollinen proteiinien varaukseen. Proteiinien elektroforeesi suoritetaan paperilla (tässä tapauksessa proteiinin liikkeen nopeus on verrannollinen vain niiden varaukseen) tai polyakryyliamidigeelissä, jolla on tietty huokoskoko (proteiinin liikkeen nopeus on verrannollinen niiden varaukseen ja molekyylipainoon) .
Ioninvaihtokromatografia- fraktiointimenetelmä, joka perustuu ionisoituneiden proteiiniryhmien sitomiseen vastakkaisesti varautuneisiin ioninvaihtohartsiryhmiin (liukenemattomat polymeerimateriaalit). Proteiinin hartsiin sitoutumisen voimakkuus on verrannollinen proteiinin varaukseen. Ioninvaihtopolymeeriin adsorboituneet proteiinit voidaan pestä pois NaCl-liuosten kasvavilla pitoisuuksilla; mitä pienempi proteiinivaraus, sitä pienempi NaCl-pitoisuus vaaditaan hartsin ioniryhmiin sitoutuneen proteiinin pesemiseksi pois.
Affiniteettikromatografia- spesifisin menetelmä yksittäisten proteiinien eristämiseksi. Proteiinin ligandi kiinnitetään kovalenttisesti inerttiin polymeeriin. Kun proteiiniliuos johdetaan kolonnin läpi polymeerin kanssa, vain tietylle ligandille spesifinen proteiini adsorboituu pylvääseen johtuen proteiinin komplementaarisesta sitoutumisesta ligandiin.
Dialyysi- menetelmä, jota käytetään pienen molekyylipainon yhdisteiden poistamiseen eristetyn proteiinin liuoksesta. Menetelmä perustuu proteiinien kyvyttömyyteen kulkea puoliläpäisevän kalvon läpi, toisin kuin pienimolekyylipainoiset aineet. Sitä käytetään proteiinien puhdistamiseen pienimolekyylisistä epäpuhtauksista, esimerkiksi suoloista suolauksen jälkeen.
TEHTÄVÄT OPIN ULKOPUOLISESTI
1. Täytä taulukko. 1.4.
Taulukko 1.4. Sukulaisten proteiinien - myoglobiinin ja hemoglobiinin - rakenteen ja toimintojen vertaileva analyysi
a) muista Mb:n ja Hb:n aktiivisen keskuksen rakenne. Mikä rooli hydrofobisilla aminohapporadikaaleilla on näiden proteiinien aktiivisten keskusten muodostumisessa? Kuvaile Mb:n ja Hb:n aktiivisen keskuksen rakennetta ja ligandien kiinnittymismekanismeja. Mikä rooli His F 8- ja His E 7 -tähteillä on Mv iHv:n aktiivisen keskuksen toiminnassa?
b) mitä uusia ominaisuuksia monomeeriseen myoglobiiniin verrattuna on läheistä sukua olevalla oligomeeriproteiinilla, hemoglobiinilla? Selitä kooperatiivisten muutosten rooli hemoglobiinimolekyylin protomeerien konformaatiossa, CO 2 - ja protonipitoisuuksien vaikutus hemoglobiinin happiaffiniteettiin sekä 2,3-BPG:n rooli Hb-toiminnan allosteerisessa säätelyssä. .
2. Kuvaile molekyylikaperoneja kiinnittäen huomiota niiden rakenteen ja toiminnan väliseen suhteeseen.
3. Mitkä proteiinit on ryhmitelty perheisiin? Tunnista immunoglobuliiniperheen esimerkkiä käyttäen tämän perheen proteiinien samanlaiset rakenteelliset piirteet ja niihin liittyvät toiminnot.
4. Puhdistettuja yksittäisiä proteiineja tarvitaan usein biokemiallisiin ja lääketieteellisiin tarkoituksiin. Selitä, mihin proteiinien fysikaalis-kemiallisiin ominaisuuksiin niiden erottamiseen ja puhdistamiseen käytetyt menetelmät perustuvat.
ITSEVALTOTEHTÄVÄT
1. Valitse oikeat vastaukset.
Hemoglobiinin tehtävät:
A. O 2:n kuljetus keuhkoista kudokseen B. H +:n kuljetus kudoksista keuhkoihin
B. Veren pH-arvon ylläpitäminen D. CO 2:n kuljetus keuhkoista kudoksiin
D. CO 2:n kuljetus kudoksista keuhkoihin
2. Valitse oikeat vastaukset. Ligandiα -protomeeri Hb on: A. Heme
B. Happi
B. CO G. 2,3-BPG
D. P-protomeeri
3. Valitse oikeat vastaukset.
Hemoglobiini toisin kuin myoglobiini:
A. Sillä on kvaternäärinen rakenne
B. Toissijaista rakennetta edustavat vain α-heliksit
B. Kuuluu monimutkaisiin proteiineihin
D. Vuorovaikuttaa allosteerisen ligandin kanssa D. Kovalenttisesti sitoutunut hemiin
4. Valitse oikeat vastaukset.
Hb:n affiniteetti O2:lle laskee:
A. Kun yksi O 2 -molekyyli lisätään B. Kun yksi O 2 -molekyyli poistetaan
B. Kun olet vuorovaikutuksessa 2,3-BPG:n kanssa
D. Kiinnitettynä protomeereihin H + D. Kun 2,3-BPG:n pitoisuus pienenee
5. Ottelu.
HB-tyypeille on ominaista:
A. Deoksimuodossa se muodostaa fibrillaarisia aggregaatteja B. Sisältää kaksi α- ja kaksi δ-ketjua
B. Hb:n vallitseva muoto aikuisten punasoluissa D. Sisältää hemiä, jonka aktiivisessa keskustassa on Fe+ 3
D. Sisältää kaksi α- ja kaksi γ-ketjua 1. HbA 2.
6. Ottelu.
Hb-ligandit:
A. Sitoutuu Hb:hen allosteerisessa keskustassa
B. Sillä on erittäin korkea affiniteetti Hb:n aktiiviseen kohtaan
B. Liittymällä se lisää Hb:n affiniteettia O 2 G:n suhteen. Hapettaa Fe+ 2:n Fe+ 3:ksi
D. Muodostaa kovalenttisen sidoksen hisF8:n kanssa
7. Valitse oikeat vastaukset.
Saattajat:
A. Proteiinit, joita on solun kaikissa osissa
B. Synteesi lisääntyy stressissä
B. Osallistu denaturoitujen proteiinien hydrolyysiin
D. Osallistu proteiinien luonnollisen konformaation ylläpitämiseen
D. Luo organelleja, joissa muodostuu proteiinien konformaatio
8. Ottelu. Immunoglobuliinit:
A. Eritysmuoto on pentameerinen.
B. Ig-luokka, joka läpäisee istukan
B. Ig - syöttösolureseptori
D. Ig:n pääluokka, joka on läsnä epiteelisolujen eritteissä. D. B-lymfosyyttireseptori, jonka aktivointi varmistaa solujen lisääntymisen
9. Valitse oikeat vastaukset.
Immunoglobuliinit E:
A. Makrofagien tuottamat B. Niissä on raskaita e-ketjuja.
B. Upotettu T-lymfosyyttien kalvoon
D. Toimivat kalvoantigeenireseptoreina syöttösoluissa ja basofiileissä
D. Vastuussa allergisista reaktioista
10. Valitse oikeat vastaukset.
Proteiinien erottelumenetelmä perustuu niiden molekyylipainon eroihin:
A. Geelisuodatus
B. Ultrasentrifugointi
B. PolD. Ioninvaihtokromatografia
D. Affiniteettikromatografia
11. Valitse oikea vastaus.
Proteiinien erottelumenetelmä perustuu eroihin niiden vesiliukoisuudessa:
A. Geelisuodatus B. Suolaus
B. Ioninvaihtokromatografia D. Affiniteettikromatografia
D. Elektroforeesi polyakryyliamidigeelissä
VASTAUKSIEN STANDARDIT ”ITSEVALTOTEHTÄVÄIN”
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
PERUSKÄSITTEET JA KÄSITTEET
1. Oligomeeriset proteiinit, protomeeri, proteiinien kvaternäärinen rakenne
2. Yhteistoiminnalliset muutokset protomeerin konformaatiossa
3. Bohr-ilmiö
4. Proteiinitoimintojen allosteerinen säätely, allosteerinen keskus ja allosteerinen efektori
5. Molekyylikaperonit, lämpöshokkiproteiinit
6. Proteiiniperheet (seriiniproteaasit, immunoglobuliinit)
7. IgM-, G-, E-, A-rakenne-funktiosuhde
8. Proteiinien kokonaisvaraus, proteiinien isoelektrinen piste
9. Elektroforeesi
10. Suolaus
11. Geelisuodatus
12. Ioninvaihtokromatografia
13. Ultrasentrifugointi
14. Affiniteettikromatografia
15. Veriplasman proteiinien elektroforeesi
TEHTÄVÄT LUOKKATYÖHÄN
1. Vertaa hemoglobiinin (Hb) ja myoglobiinin (Mb) saturaatioasteiden riippuvuutta hapella sen osapaineesta kudoksissa
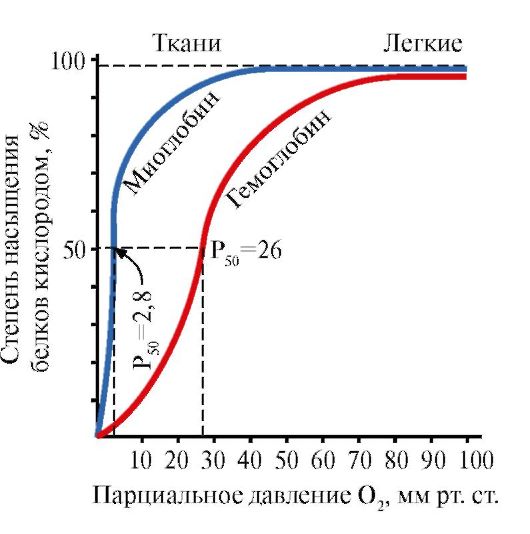 Riisi. 1.34. Kyllästymisen riippuvuus Mv jaNHhappea sen osapaineesta
Riisi. 1.34. Kyllästymisen riippuvuus Mv jaNHhappea sen osapaineesta
Huomaa, että proteiinin happisaturaatiokäyrien muoto on erilainen: myoglobiinilla - hyperbola, hemoglobiinilla - sigmoidimuoto.
1. Vertaa hapen osapaineen arvoja, joissa Mb ja Hb ovat kyllästyneet O 2:lla 50 %. Millä näistä proteiineista on suurempi affiniteetti O 2:een?
2. Mitkä Mb:n rakenteelliset ominaisuudet määräävät sen suuren affiniteetin O 2:lle?
3. Mitkä HB:n rakenteelliset ominaisuudet sallivat sen vapauttaa O2:ta lepäävien kudosten kapillaareista (suhteellisen korkealla O2:n osapaineella) ja lisätä tätä vapautumista jyrkästi toimivissa lihaksissa? Mikä oligomeeristen proteiinien ominaisuus tarjoaa tämän vaikutuksen?
4. Laske kuinka paljon O 2 (%) hapetettua hemoglobiinia antaa lepääville ja työskenteleville lihaksille?
5. tehdä johtopäätöksiä proteiinin rakenteen ja toiminnan välisestä suhteesta.
2. Hemoglobiinin kapillaareissa vapauttaman hapen määrä riippuu kudosten katabolisten prosessien voimakkuudesta (Bohr-ilmiö). Kuinka muutokset kudosten aineenvaihdunnassa säätelevät Hb:n affiniteettia O2:een? CO 2:n ja H+:n vaikutus Hb:n affiniteettiin O 2:ta kohtaan
 1. kuvaa Bohrin vaikutusta.
1. kuvaa Bohrin vaikutusta.
2. mihin suuntaan kaaviossa näkyvä prosessi etenee:
a) keuhkojen kapillaareissa;
b) kudoskapillaareissa?
3. Mikä on Bohr-ilmiön fysiologinen merkitys?
4. Miksi Hb:n ja H+:n vuorovaikutus hemistä kaukana olevissa kohdissa muuttaa proteiinin affiniteettia O 2:lle?
3. Hb:n affiniteetti 02:lle riippuu sen ligandin pitoisuudesta - 2,3-bisfosfoglyseraatti, joka on allosteerinen säätelijä Hb:n affiniteetille O2:lle. Miksi ligandin vuorovaikutus aktiivisesta kohdasta kaukana olevassa kohdassa vaikuttaa proteiinin toimintaan? Kuinka 2,3-BPG säätelee Hb:n affiniteettia O2:een? Voit ratkaista ongelman vastaamalla seuraaviin kysymyksiin:
1. mistä ja mistä 2,3-bisfosfoglyseraattia (2,3-BPG) syntetisoidaan? Kirjoita sen kaava, osoita tämän molekyylin varaus.
2. Minkä hemoglobiinimuodon (oksi tai deoksi) kanssa BPG on vuorovaikutuksessa ja miksi? Missä Hb-molekyylin osassa vuorovaikutus tapahtuu?
3. mihin suuntaan kaaviossa esitetty prosessi tapahtuu?
a) kudosten kapillaareissa;
b) keuhkojen kapillaareissa?
4. jossa kompleksin pitoisuuden tulisi olla suurempi
Nv-2,3-BFG:
a) levossa olevien lihasten kapillaareissa,
b) työskentelevien lihasten kapillaareissa (edellyttäen, että punasoluissa on sama BPG-pitoisuus)?
5. Miten HB:n affiniteetti hapen suhteen muuttuu, kun ihminen sopeutuu korkeisiin olosuhteisiin, jos BPG:n pitoisuus erytrosyyteissä kasvaa? Mikä on tämän ilmiön fysiologinen merkitys?
4. 2,3-BPG:n tuhoutuminen säilötyn veren varastoinnin aikana heikentää HB:n toimintaa. Miten HB:n affiniteetti O 2:lle muuttuu säilyneessä veressä, jos 2,3-BPG:n pitoisuus punasoluissa voi laskea 8:sta 0,5 mmol/l:aan. Onko mahdollista siirtää tällaista verta vakavasti sairaille potilaille, jos 2,3-BPG-pitoisuus palautuu aikaisintaan kolmen päivän kuluttua? Onko mahdollista palauttaa punasolujen toiminta lisäämällä vereen 2,3-BPG:tä?
5. Muista yksinkertaisimpien immunoglobuliinimolekyylien rakenne. Mikä rooli immunoglobuliineilla on immuunijärjestelmässä? Miksi iggejä kutsutaan usein bivalentteiksi? Miten Ig:n rakenne liittyy niiden toimintaan? (Kuvaile käyttämällä esimerkkiä immunoglobuliiniluokista.)
Proteiinien fysikaalis-kemialliset ominaisuudet ja menetelmät niiden erottamiseen.
6. Miten proteiinin nettovaraus vaikuttaa sen liukoisuuteen?
a) määritä peptidin kokonaisvaraus pH:ssa 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) kuinka tämän peptidin varaus muuttuu pH:ssa >7, pH<7, рН <<7?
c) mikä on proteiinin isoelektrinen piste (IEP) ja missä ympäristössä se sijaitsee?
Tämän peptidin IET?
d) missä pH-arvossa tämän peptidin pienin liukoisuus havaitaan.
7. Miksi piimä, toisin kuin tuore maito, "juostuu" keitettäessä (eli maitoproteiini kaseiini saostuu)? Tuoreessa maidossa kaseiinimolekyyleillä on negatiivinen varaus.
8. Geelisuodatusta käytetään yksittäisten proteiinien erottamiseen. Seos, joka sisälsi proteiineja A, B ja C, joiden molekyylipainot ovat 160 000, 80 000 ja 60 000, analysoitiin geelisuodatuksella (kuva 1.35). Turvonneet geelirakeet läpäisevät proteiineja, joiden molekyylipaino on alle 70 000. Mikä periaate on tämän erotusmenetelmän taustalla? Mikä kuvaaja heijastaa oikein fraktioinnin tuloksia? Ilmoita järjestys, jossa proteiinit A, B ja C vapautuvat kolonnista.
 Riisi. 1.35. Geelisuodatuksen käyttö proteiinien erottamiseen
Riisi. 1.35. Geelisuodatuksen käyttö proteiinien erottamiseen
9. Kuvassa 1.36, A esittää kaavion elektroforeesista paperille terveen ihmisen veren seerumin proteiineista. Tällä menetelmällä saatujen proteiinifraktioiden suhteelliset määrät ovat: albumiinit 54-58 %, a1-globuliinit 6-7 %, a2-globuliinit 8-9 %, p-globuliinit 13 %, y-globuliinit 11-12 %.
 Riisi. 1.36 Terveen ihmisen (A) ja potilaan (B) veriplasmaproteiinien elektroforeesi paperille
Riisi. 1.36 Terveen ihmisen (A) ja potilaan (B) veriplasmaproteiinien elektroforeesi paperille
I - y-globuliinit; II - p-globuliinit; III -α 2-globuliini; IV -α 2-globuliini; V - albumiinit
Moniin sairauksiin liittyy kvantitatiivisia muutoksia seerumin proteiinien koostumuksessa (dysproteinemia). Näiden muutosten luonne otetaan huomioon diagnoosia tehtäessä sekä taudin vaikeusastetta ja vaihetta arvioitaessa.
Käyttämällä taulukossa annettuja tietoja. 1.5, tee arvaus sairaudesta, jolle on tunnusomaista kuvassa 1 esitetty elektroforeettinen profiili. 1.36.
Taulukko 1.5. Muutokset seerumin proteiinien pitoisuudessa patologiassa
Oravat- suurimolekyylipainoiset orgaaniset yhdisteet, jotka koostuvat α-aminohappotähteistä.
SISÄÄN proteiinikoostumus sisältää hiilen, vedyn, typen, hapen, rikin. Jotkut proteiinit muodostavat komplekseja muiden fosforia, rautaa, sinkkiä ja kuparia sisältävien molekyylien kanssa.
Proteiineilla on suuri molekyylipaino: munaalbumiini - 36 000, hemoglobiini - 152 000, myosiini - 500 000 Vertailun vuoksi: alkoholin molekyylipaino on 46, etikkahappo - 60, bentseeni - 78.
Proteiinien aminohappokoostumus
Oravat- ei-jaksolliset polymeerit, joiden monomeerit ovat α-aminohapot. Tyypillisesti 20 tyyppistä α-aminohappoa kutsutaan proteiinimonomeereiksi, vaikka yli 170 niistä löytyy soluista ja kudoksista.
Sen mukaan, voidaanko aminohappoja syntetisoida ihmisten ja muiden eläinten kehossa, ne erotetaan: ei-välttämättömiä aminohappoja- voidaan syntetisoida; välttämättömiä aminohappoja- ei voida syntetisoida. Välttämättömät aminohapot on saatava elimistölle ruuan kautta. Kasvit syntetisoivat kaikenlaisia aminohappoja.
Riippuen aminohappokoostumuksesta, proteiinit ovat: täydellisiä- sisältää koko sarjan aminohappoja; viallinen- Jotkut aminohapot puuttuvat niiden koostumuksesta. Jos proteiinit koostuvat vain aminohapoista, niitä kutsutaan yksinkertainen. Jos proteiinit sisältävät aminohappojen lisäksi ei-aminohappokomponentin (proteesiryhmä), niitä kutsutaan ns. monimutkainen. Prosteettista ryhmää voivat edustaa metallit (metalliproteiinit), hiilihydraatit (glykoproteiinit), lipidit (lipoproteiinit), nukleiinihapot (nukleoproteiinit).
Kaikki aminohapot sisältävät 1) karboksyyliryhmä (-COOH), 2) aminoryhmä (-NH2), 3) radikaali tai R-ryhmä (muu molekyyli). Radikaalin rakenne on erilainen erityyppisille aminohappoille. Aminohappojen koostumukseen sisältyvien aminoryhmien ja karboksyyliryhmien lukumäärästä riippuen ne erotetaan: neutraaleja aminohappoja jossa on yksi karboksyyliryhmä ja yksi aminoryhmä; emäksiset aminohapot joissa on useampi kuin yksi aminoryhmä; happamat aminohapot joissa on useampi kuin yksi karboksyyliryhmä.
Aminohapot ovat amfoteeriset yhdisteet, koska liuoksessa ne voivat toimia sekä happoina että emäksinä. Vesiliuoksissa aminohapot ovat eri ionimuodoissa.
Peptidisidos
Peptidit- orgaaniset aineet, jotka koostuvat aminohappotähteistä, jotka on yhdistetty peptidisidoksilla.
Peptidien muodostuminen tapahtuu aminohappojen kondensaatioreaktion seurauksena. Kun yhden aminohapon aminoryhmä on vuorovaikutuksessa toisen aminohapon karboksyyliryhmän kanssa, niiden välille muodostuu kovalenttinen typpi-hiilisidos, joka on ns. peptidi. Peptidiin sisältyvien aminohappotähteiden lukumäärästä riippuen niitä on dipeptidit, tripeptidit, tetrapeptidit jne. Peptidisidoksen muodostuminen voidaan toistaa monta kertaa. Tämä johtaa muodostumiseen polypeptidit. Peptidin toisessa päässä on vapaa aminoryhmä (kutsutaan N-pääksi), ja toisessa on vapaa karboksyyliryhmä (kutsutaan C-pääksi).

Proteiinimolekyylien tilaorganisaatio
Proteiinien tiettyjen spesifisten toimintojen suorittaminen riippuu niiden molekyylien avaruudellisesta konfiguraatiosta. Lisäksi solun on energeettisesti epäedullista pitää proteiineja laskostumattomassa muodossa, ketjun muodossa, minkä vuoksi polypeptidiketjut laskostuvat, jolloin ne laskostuvat; tietty kolmiulotteinen rakenne tai konformaatio. Tasoja on 4 proteiinien tilajärjestely.
Ensisijainen proteiinirakenne- aminohappotähteiden järjestyksen sekvenssi polypeptidiketjussa, joka muodostaa proteiinimolekyylin. Aminohappojen välinen sidos on peptidisidos.

Jos proteiinimolekyyli koostuu vain 10 aminohappotähteestä, niin teoreettisesti mahdollisten proteiinimolekyylien varianttien lukumäärä, jotka eroavat aminohappojen vuorottelujärjestyksessä, on 10 20. Kun sinulla on 20 aminohappoa, voit tehdä niistä entistä monipuolisempia yhdistelmiä. Ihmiskehosta on löydetty noin kymmenentuhatta erilaista proteiinia, jotka eroavat sekä toisistaan että muiden organismien proteiineista.
Se on proteiinimolekyylin ensisijainen rakenne, joka määrittää proteiinimolekyylien ominaisuudet ja sen avaruudellisen konfiguraation. Vain yhden aminohapon korvaaminen toisella polypeptidiketjussa johtaa proteiinin ominaisuuksien ja toimintojen muutokseen. Esimerkiksi kuudennen glutamiiniaminohapon korvaaminen valiinilla hemoglobiinin β-alayksikössä johtaa siihen, että hemoglobiinimolekyyli kokonaisuutena ei voi suorittaa päätehtäväänsä - hapen kuljetusta; Tällaisissa tapauksissa henkilölle kehittyy sairaus, jota kutsutaan sirppisoluanemiaksi.
Toissijainen rakenne- määrätty polypeptidiketjun laskostuminen spiraaliksi (näyttää pidennetyltä jouselta). Kierteen käänteitä vahvistavat vetysidokset, jotka syntyvät karboksyyliryhmien ja aminoryhmien välillä. Lähes kaikki CO- ja NH-ryhmät osallistuvat vetysidosten muodostukseen. Ne ovat heikompia kuin peptidit, mutta useaan otteeseen toistettuna antavat tälle konfiguraatiolle stabiiliutta ja jäykkyyttä. Toissijaisen rakenteen tasolla on proteiineja: fibroiini (silkki, hämähäkinverkko), keratiini (hiukset, kynnet), kollageeni (jänteet).



Tertiäärinen rakenne- polypeptidiketjujen pakkaaminen palloiksi, jotka johtuvat kemiallisten sidosten (vety, ioni, disulfidi) muodostumisesta ja hydrofobisten vuorovaikutusten muodostumisesta aminohappotähteiden radikaalien välille. Päärooli tertiäärisen rakenteen muodostumisessa on hydrofiilis-hydrofobisilla vuorovaikutuksilla. Vesiliuoksissa hydrofobisilla radikaaleilla on taipumus piiloutua vedestä ryhmittymällä pallon sisään, kun taas hydrofiiliset radikaalit hydratoitumisen seurauksena (vuorovaikutus vesidipolien kanssa) pyrkivät ilmestymään molekyylin pinnalle. Joissakin proteiineissa tertiääristä rakennetta stabiloivat kovalenttiset disulfidisidokset, jotka muodostuvat kahden kysteiinitähteen rikkiatomien välille. Tertiäärisellä rakennetasolla on entsyymejä, vasta-aineita ja joitain hormoneja.

Kvaternaarirakenne ominaisuus monimutkaisille proteiineille, joiden molekyylit muodostuvat kahdesta tai useammasta pallosta. Alayksiköt pysyvät molekyylissä ionisten, hydrofobisten ja sähköstaattisten vuorovaikutusten avulla. Joskus kvaternaarisen rakenteen muodostumisen aikana alayksiköiden välillä tapahtuu disulfidisidoksia. Tutkituin kvaternäärisen rakenteen omaava proteiini on hemoglobiini. Se muodostuu kahdesta α-alayksiköstä (141 aminohappotähdettä) ja kahdesta β-alayksiköstä (146 aminohappotähdettä). Jokaiseen alayksikköön liittyy rautaa sisältävä hemimolekyyli.
Jos proteiinien avaruudellinen konformaatio jostain syystä poikkeaa normaalista, proteiini ei voi suorittaa tehtäviään. Esimerkiksi "hullun lehmän taudin" (spongiformisen enkefalopatian) syy on prionien, hermosolujen pintaproteiinien, epänormaali konformaatio.
Proteiinien ominaisuudet
Proteiinimolekyylin aminohappokoostumus ja rakenne määräävät sen ominaisuuksia. Proteiineissa yhdistyvät emäksiset ja happamat ominaisuudet, jotka määräytyvät aminohapporadikaalien avulla: mitä enemmän proteiinissa on happamia aminohappoja, sitä selvemmät sen happamat ominaisuudet ovat. Mahdollisuus lahjoittaa ja lisätä H + on määritetty proteiinien puskurointiominaisuudet; Yksi tehokkaimmista puskureista on punasoluissa oleva hemoglobiini, joka pitää veren pH:n vakiona. On liukoisia proteiineja (fibrinogeeni) ja on liukenemattomia proteiineja, jotka suorittavat mekaanisia toimintoja (fibroiini, keratiini, kollageeni). On proteiineja, jotka ovat kemiallisesti aktiivisia (entsyymejä), on kemiallisesti inaktiivisia proteiineja, jotka kestävät erilaisia ympäristöolosuhteita ja jotka ovat erittäin epästabiileja.
Ulkoiset tekijät (lämpö, ultraviolettisäteily, raskasmetallit ja niiden suolat, pH-muutokset, säteily, kuivuminen)
voi aiheuttaa häiriöitä proteiinimolekyylin rakenteellisessa organisaatiossa. Tietylle proteiinimolekyylille ominaisen kolmiulotteisen konformaation menetysprosessia kutsutaan denaturaatio. Denaturoitumisen syynä on tiettyä proteiinirakennetta stabiloivien sidosten katkeaminen. Aluksi heikoimmat siteet katkeavat, ja kun olosuhteet tiukentuvat, vielä vahvemmat katkeavat. Siksi ensin kvaternaariset, sitten tertiääri- ja sekundaarirakenteet menetetään. Muutos spatiaalisessa konfiguraatiossa johtaa muutokseen proteiinin ominaisuuksissa ja sen seurauksena tekee mahdottomaksi proteiinin suorittaa sen luontaisia biologisia tehtäviä. Jos denaturaatioon ei liity primaarirakenteen tuhoutumista, niin se voi olla käännettävä Tässä tapauksessa tapahtuu proteiinille ominaisen konformaation palautuminen itsestään. Esimerkiksi kalvoreseptoriproteiinit käyvät läpi tällaisen denaturoitumisen. Prosessia, jossa proteiinirakenne palautetaan denaturoinnin jälkeen, kutsutaan renaturaatio. Jos proteiinin spatiaalisen konfiguraation palauttaminen on mahdotonta, kutsutaan denaturaatiota peruuttamaton.
Proteiinien toiminnot
| Toiminto | Esimerkkejä ja selityksiä |
|---|---|
| Rakentaminen | Proteiinit osallistuvat solu- ja ekstrasellulaaristen rakenteiden muodostumiseen: ne ovat osa solukalvoja (lipoproteiinit, glykoproteiinit), hiuksia (keratiini), jänteitä (kollageeni) jne. |
| Kuljetus | Veren proteiini hemoglobiini kiinnittää happea ja kuljettaa sen keuhkoista kaikkiin kudoksiin ja elimiin ja siirtää niistä hiilidioksidia keuhkoihin; Solukalvojen koostumus sisältää erityisiä proteiineja, jotka varmistavat tiettyjen aineiden ja ionien aktiivisen ja tiukasti selektiivisen siirron solusta ulkoiseen ympäristöön ja takaisin. |
| Sääntely | Proteiinihormonit osallistuvat aineenvaihduntaprosessien säätelyyn. Esimerkiksi hormoni insuliini säätelee veren glukoositasoja, edistää glykogeenisynteesiä ja lisää rasvojen muodostumista hiilihydraateista. |
| Suojaava | Reaktiona vieraiden proteiinien tai mikro-organismien (antigeenien) tunkeutumiseen kehoon muodostuu erityisiä proteiineja - vasta-aineita, jotka voivat sitoa ja neutraloida ne. Fibrinogeenistä muodostuva fibriini auttaa pysäyttämään verenvuodon. |
| Moottori | Supistuvat proteiinit aktiini ja myosiini saavat aikaan lihasten supistumisen monisoluisissa eläimissä. |
| Signaali | Solun pintakalvoon on rakennettu proteiinimolekyylejä, jotka pystyvät muuttamaan tertiääristä rakennettaan vasteena ympäristötekijöille ja siten vastaanottamaan signaaleja ulkoisesta ympäristöstä ja lähettämään komentoja soluun. |
| Varastointi | Eläinten kehossa proteiineja ei yleensä varastoida, paitsi munaalbumiini ja maitokeiini. Mutta proteiinien ansiosta joitain aineita voidaan varastoida elimistöön, esimerkiksi hemoglobiinin hajoamisen aikana, rautaa ei poisteta kehosta, vaan se varastoituu muodostaen kompleksin ferritiiniproteiinin kanssa. |
| Energiaa | Kun 1 g proteiinia hajoaa lopputuotteiksi, vapautuu 17,6 kJ. Ensin proteiinit hajoavat aminohapoiksi ja sitten lopputuotteiksi - vedeksi, hiilidioksidiksi ja ammoniakiksi. Proteiineja käytetään kuitenkin energianlähteenä vasta, kun muut lähteet (hiilihydraatit ja rasvat) on käytetty loppuun. |
| Katalyyttinen | Yksi proteiinien tärkeimmistä toiminnoista. Tarjoaa proteiinit - entsyymit, jotka nopeuttavat soluissa tapahtuvia biokemiallisia reaktioita. Esimerkiksi rkatalysoi CO 2:n kiinnittymistä fotosynteesin aikana. |
Entsyymit
Entsyymit, tai entsyymejä, ovat erityinen proteiiniluokka, jotka ovat biologisia katalyyttejä. Entsyymien ansiosta biokemialliset reaktiot tapahtuvat valtavalla nopeudella. Entsymaattisten reaktioiden nopeus on kymmeniä tuhansia kertoja (ja joskus miljoonia) suurempi kuin epäorgaanisten katalyyttien kanssa tapahtuvien reaktioiden nopeus. Ainetta, johon entsyymi vaikuttaa, kutsutaan substraatti.
Entsyymit ovat pallomaisia proteiineja, rakenteellisia ominaisuuksia Entsyymit voidaan jakaa kahteen ryhmään: yksinkertaisiin ja monimutkaisiin. Yksinkertaiset entsyymit ovat yksinkertaisia proteiineja, ts. koostuvat vain aminohapoista. Monimutkaiset entsyymit ovat monimutkaisia proteiineja, ts. Proteiiniosan lisäksi ne sisältävät ryhmän ei-proteiinia - kofaktori. Jotkut entsyymit käyttävät vitamiineja kofaktoreina. Entsyymimolekyyli sisältää erityisen osan, jota kutsutaan aktiiviseksi keskukseksi. Aktiivinen keskus- pieni osa entsyymistä (kolmesta kahteentoista aminohappotähdettä), jossa substraatin tai substraattien sitoutuminen tapahtuu entsyymi-substraattikompleksin muodostamiseksi. Reaktion päätyttyä entsyymi-substraattikompleksi hajoaa entsyymiksi ja reaktiotuotteeksi (-tuotteiksi). Joillakin entsyymeillä (paitsi aktiivisella) allosteeriset keskukset- alueet, joihin on kiinnitetty entsyyminopeudensäätimet ( allosteeriset entsyymit).

Entsymaattisen katalyysin reaktiot ovat tunnusomaisia: 1) korkea hyötysuhde, 2) tiukka selektiivisyys ja toiminnan suunta, 3) substraattispesifisyys, 4) hieno ja tarkka säätely. Entsymaattisten katalyysireaktioiden substraatti- ja reaktiospesifisyys selittyvät E. Fischerin (1890) ja D. Koshlandin (1959) hypoteesilla.
E. Fisher (näppäinlukon hypoteesi) ehdotti, että entsyymin aktiivisen keskuksen ja substraatin avaruudellisten konfiguraatioiden on vastattava tarkasti toisiaan. Substraattia verrataan "avaimeen", entsyymiä "lukkoon".
D. Koshland (käsi-hansikas hypoteesi) ehdotti, että substraatin rakenteen ja entsyymin aktiivisen keskuksen välinen spatiaalinen vastaavuus syntyy vain niiden vuorovaikutuksen hetkellä. Tätä hypoteesia kutsutaan myös indusoitu vastaavuushypoteesi.
Entsymaattisten reaktioiden nopeus riippuu: 1) lämpötilasta, 2) entsyymipitoisuudesta, 3) substraattipitoisuudesta, 4) pH:sta. On syytä korostaa, että koska entsyymit ovat proteiineja, niiden aktiivisuus on suurin fysiologisesti normaaleissa olosuhteissa.
Useimmat entsyymit voivat toimia vain 0-40 °C:n lämpötiloissa. Näissä rajoissa reaktionopeus kasvaa noin 2 kertaa jokaisella 10 °C:n lämpötilan nousulla. Yli 40 °C:n lämpötiloissa proteiini denaturoituu ja entsyymiaktiivisuus laskee. Lähellä jäätymistä entsyymit inaktivoituvat.
Substraatin määrän kasvaessa entsymaattisen reaktion nopeus kasvaa, kunnes substraattimolekyylien lukumäärä on yhtä suuri kuin entsyymimolekyylien lukumäärä. Kun substraatin määrää kasvaa edelleen, nopeus ei kasva, koska entsyymin aktiiviset keskukset ovat kyllästyneet. Entsyymipitoisuuden kasvu johtaa lisääntyneeseen katalyyttiseen aktiivisuuteen, koska suurempi määrä substraattimolekyylejä käy läpi transformaatioita aikayksikköä kohti.

Jokaiselle entsyymille on optimaalinen pH-arvo, jossa se osoittaa maksimaalista aktiivisuutta (pepsiini - 2,0, syljen amylaasi - 6,8, haiman lipaasi - 9,0). Korkeammilla tai alhaisemmilla pH-arvoilla entsyymiaktiivisuus laskee. Äkillisten pH-muutosten myötä entsyymi denaturoituu.
Allosteeristen entsyymien nopeutta säätelevät aineet, jotka kiinnittyvät allosteerisiin keskuksiin. Jos nämä aineet nopeuttavat reaktiota, niitä kutsutaan aktivaattorit, jos ne hidastavat - estäjät.
Entsyymien luokittelu
Katalysoimiensa kemiallisten muutosten tyypin mukaan entsyymit jaetaan kuuteen luokkaan:
- oksireduktaasit(vety-, happi- tai elektroniatomien siirto aineesta toiseen - dehydrogenaasi),
- transferaasit(metyyli-, asyyli-, fosfaatti- tai aminoryhmän siirto aineesta toiseen - transaminaasi),
- hydrolaasit(hydrolyysireaktiot, joissa substraatista muodostuu kaksi tuotetta - amylaasi, lipaasi),
- lyaasit(ei-hydrolyyttinen lisäys substraattiin tai atomiryhmän irrottaminen siitä, jolloin C-C-, C-N-, C-O-, C-S-sidokset voivat katketa - dekarboksylaasi),
- isomeraaseja(molekyylinsisäinen uudelleenjärjestely - isomeraasi),
- ligaasit(kahden molekyylin yhdistäminen C-C-, C-N-, C-O-, C-S-sidosten muodostumisen seurauksena - syntetaasi).
Luokat puolestaan on jaettu alaluokkiin ja alaluokkiin. Nykyisessä kansainvälisessä luokituksessa jokaisella entsyymillä on oma koodi, joka koostuu neljästä pisteillä erotetusta numerosta. Ensimmäinen numero on luokka, toinen on alaluokka, kolmas on alaluokka, neljäs on tämän alaluokan entsyymin sarjanumero, esimerkiksi arginaasikoodi on 3.5.3.1.
Mene luennot nro 2"Hiilihydraattien ja lipidien rakenne ja toiminnot"
Mene luennot nro 4"ATP-nukleiinihappojen rakenne ja toiminnot"
Proteiinit (proteiinit) ovat peptidiluonteisia suurimolekyylisiä polymeeriyhdisteitä (polyheteroaminohappoja).
Proteiinien päärakenne on polypeptidiketjun (PPC) vuorottelevien aminohappotähteiden sekvenssi.
Proteiinien ensisijainen rakenne on kovalenttinen rakenne, koska se perustuu peptidi aminohappojen a-amino- ja a-karboksyyliryhmien välinen yhteys. Tämän seurauksena polypeptidiketjut ovat haaroittumattomia.
Polypeptidiketjun luuranko (runko, luuranko) koostuu säännöllisesti toistuvista rakenneelementeistä
Polypeptidiketjulla on vektoriteettia, ketjun suunta on N-päästä (ketjun alku) C-päähän (ketjun päähän), N-pää on se pää, jossa vapaa a-aminoryhmä sijaitsee. C-pää on pää, joka sisältää vapaan a-karboksyyliryhmän. Proteiinien aminohapposekvenssi merkitään N-päästä alkaen käyttämällä kolmikirjaimia lyhennettyjä aminohapponimiä, esimerkiksi: gly-ala-cis-pro. Proteiinin aminohappotähteiden yksikirjaimista nimitystä voidaan myös käyttää.
Proteiinien N- ja C-päät voidaan modifioida. N-päässä oleva aminoryhmä voi olla asetyloitu, formyloitu tai metyloitu. Monissa proteiineissa N-terminaalinen tähde on pyrrolidonikarbonaatti (pyroglutamaatti), joka ei sisällä vapaata aminoryhmää. C-pää voi olla amidoitu. C-pään modifikaatiot ovat harvinaisempia kuin N-terminaaliset modifikaatiot.
Proteiinien polykondensaatiokerroin vaihtelee välillä 50-2500. Tyypillisesti proteiini sisältää 100-300 aminohappotähdettä. Koska yhden aminohappotähteen keskimääräinen molekyylipaino on noin 110 Da, proteiinien molekyylipaino vaihtelee 6000:sta miljooniin Da.
Jokaisella yksittäisellä proteiinilla on ainutlaatuinen primäärirakenne. Ensimmäinen proteiini, jonka primäärirakenne vakiintui, oli insuliini. Sanger onnistui tekemään tämän. Hänen strategiansa oli seuraava. Hän erotti ensin kaksi polypeptidiketjua ja suoritti sitten niiden spesifisen entsymaattisen pilkkomisen pieniksi peptideiksi, jotka sisälsivät päällekkäisiä sekvenssejä. N-terminaaliset tähteet tunnistettiin sitten käyttämällä 1-fluori-2,4-dinitrobentseeniä. Lisäksi hän määritti peptidien aminohappokoostumuksen ja pystyi lopulta määrittämään niiden rakenteen vertaamalla päällekkäisten peptidien sekvenssejä. Yleisesti ottaen Sangerin strategia on säilyttänyt merkityksensä tähän päivään asti. Muitakin lähestymistapoja on kuitenkin ehdotettu. Edman kehitti menetelmän automaattiseen menettelyyn N-terminaalisten aminohappotähteiden peräkkäiseen pilkkomiseen ja tunnistamiseen. Röntgendiffraktioanalyysiä voidaan käyttää primäärirakenteen tulkitsemiseen. Aminohappotähteiden sekvenssi voidaan määrittää lähetti-RNA:n nukleotidisekvenssistä.
Tällä hetkellä yli 2000 proteiinin primäärirakenne on selvitetty. Teoreettisesti proteiinien primäärirakenteen eri varianttien määrä on rajoittamaton. Jopa 20 eri aminohapon polypeptidille mahdollisten sekvenssien lukumäärä on 20'1018. Elävässä luonnossa toteutuu vain pieni osa mahdollisista sekvensseistä, joiden kokonaismääräksi kaikentyyppisissä elävissä organismeissa on arviolta 10 10 -10 12.
Proteiinien primäärirakenne on geneettisesti määrätty, ts. proteiinin aminohapposekvenssi määräytyy nukleotidien sekvenssin mukaan DNA. DNA-nukleotidisekvenssin vääristymät johtavat epänormaalien proteiinien syntymiseen, joilla on muuttuneet biologiset ominaisuudet, mikä on molekyylipatologian syy. Erityisesti sirppisoluanemia johtuu hemoglobiinin b-ketjua säätelevän geenin pistemutaatiosta. Tämän seurauksena b-ketjun 6. asemassa oleva glutamaattijäännös korvataan valiinilla. Tämä substituutio johtaa yhden negatiivisen varauksen menettämiseen kummassakin kahdessa b-ketjussa, mikä johtaa muutokseen hemoglobiinin konformaatiossa ja sen biologisen toiminnan menettämiseen.
Homologiset proteiinit ovat proteiineja, jotka suorittavat samoja tehtäviä eri lajeissa. Esimerkki on hemoglobiini: kaikissa selkärankaisissa se suorittaa saman hapen kuljetukseen liittyvän toiminnon. Homologisille proteiineille on tunnusomaista samojen aminohappojen läsnäolo monissa asemissa. Kuten kävi ilmi, niiden aminohappotähteiden lukumäärä, joissa homologiset proteiinit eroavat toisistaan, on verrannollinen näiden lajien väliseen fylogeneettiseen eroon. Esimerkiksi hevosen ja hiivan sytokromi C -molekyylit eroavat 48 aminohappotähteen osalta, kun taas kanan ja ankan samat molekyylit eroavat vain kahdessa tähteessä. Mitä tulee kanan ja kalkkunan sytokromeihin C, niillä on identtiset aminohapposekvenssit. Tietoa eri lajien homologisten proteiinien aminohapposekvenssien erojen määrästä käytetään evoluutiokarttojen rakentamiseen, jotka kuvastavat eri eläin- ja kasvilajien syntymisen ja kehityksen peräkkäisiä vaiheita evoluutioprosessissa.
Luento 3. Proteiinin rakenne
Määritelmä:
Proteiinit ovat epäsäännöllisiä polymeerejä, joiden monomeerit ovat L- aminohappoja.
Aminohappoja
Luonnossa on kaksi stereoisomeerien muotoa: L (vasenkätinen) ja D (vapaapäin kiertävä). sitä paitsi L - aminohappoja, joista muodostuu proteiineja, on kehossa D -aminohapot, jotka eivät sisälly proteiineihin.
Aminohapon yleinen kaava on esitetty kuvassa.
Se koskee 19:ää proteiineissa olevasta 20 aminohaposta. Näiden 19 aminohapon lisäksi proteiinit sisältävät yhden iminohappo - proliini
Kaikki aminohapot sisältävät α -aminoryhmä. Tästä johtuu nimi - "α-aminohapot". proliinissa - α- imino ryhmä.
Proteiineja muodostavien aminohappojen luokitus radikaalin polaarisuuden (ei-polaarisuuden) periaatteen mukaan.
1. Ei-polaariset tai hydrofobiset radikaalit.
Alifaattinen - alaniini, valiini, leusiini, isoleusiini. Rikkiä sisältävä metioniini Aromaattinen - fenyylialaniini, tryptofaani. Iminohappo proliini
2. Polaariset, mutta varautumattomat radikaalit. Glysiini.
Hydroksiaminohapot - seriini, treoniini, tyrosiini. Sisältää sulfhydryyliryhmän kysteiini. Sisältää amidiryhmän: asparagiini, glutamiini.
3. Negatiivisesti varautuneet radikaalit. Asparagiinihappo, glutamiinihappo.
4. Positiivisesti varautuneita radikaaleja. Lysiini, arginiini, histidiini.
Ensisijainen proteiinirakenne
Määritelmä:
Proteiinin ensisijainen rakenne on aminohappotähteiden sekvenssi polypeptidiketjussa.
Aminohapot yhdistetään muodostamaan polypeptidi käyttämällä kovalenttisia (amidi)sidoksia.
Tripeptidillä, joka koostuu kolmesta eri aminohaposta, on mahdollista 3! = 6 erilaista primäärirakennetta.
Kahdestakymmenestä erilaisesta aminohaposta koostuvan oligopeptidin primäärirakenteiden monimuotoisuus on 20!, mikä tarkoittaa 2x10 18.
Keskikokoisen proteiinin primäärirakenteiden monimuotoisuus (noin 500 aminohappoa) on jo noin 20 500 muunnelmaa (jos kaikki aminohapot esitetään ekvimolaarisissa suhteissa).
PäälläMaapallolla ei ole ollut, ei ole eikä tule olemaan kahta ihmistä, joilla on täsmälleen sama proteiinisarja.
Proteiinin sekundaarinen rakenne
Määritelmä:
Proteiinin sekundäärinen rakenne on polypeptidiketjujen järjestynyt rakenne, jonka aiheuttavat vetysidokset C=O:n ja N- Herilaisia aminohappoja.
Toissijainen rakenne voi olla säännöllinen (α-heliksi) tai epäsäännöllinen (β-levyrakenne). α-heliksissä NH-ryhmä n Kolmas aminohappotähde on vuorovaikutuksessa (n-4) aminohappotähteen C=O-ryhmän kanssa. Halkaisijaltaan 10,1 Å:n β-heliksin kierrosta kohti on 3,6 aminohappotähdettä. Tavallisen α-heliksin identiteettijakso on 18 aminohappoa (5 kierrosta). Normaalin α-heliksin häiritsijä on ensisijaisesti proliini. Toiseksi tärkein vaikutus on lähellä sijaitsevat yhtä varautuneet radikaalit.
p-laskokset voivat muodostua ei vain yksittäisistä, vaan myös vierekkäisistä polypeptideistä, jotka ovat osa yhtä proteiinia.
Puhdas luonnollinen alfa tai beta - proteiineja ei ole olemassa.
Proteiinin tertiäärinen rakenne
Määritelmä
Proteiinin tertiäärinen rakenne on polypeptidin spatiaalinen konformaatio, jolla on sekundäärinen rakenne ja jonka määräävät radikaalien väliset vuorovaikutukset.
Radikaalien välillä on neljä vuorovaikutusta.
Radikaalien välisten vuorovaikutusten tyypit
1 . Kovalenttinen Viestintä välillä jäämiä kaksi kysteiinit (disulfidi sillat).
2. Ioniset (sähköstaattiset) vuorovaikutukset vastakkaisesti varautuneiden aminohappotähteiden välillä (kolme radikaalia "+"-merkillä ja kaksi "-"-merkillä).
Esimerkiksi lysiinin positiivisesti varautunut e-aminoryhmä (- NH3 +) vetää puoleensa negatiivisesti varautunut karboksyyliryhmä – (COO-) glutamiini- tai asparagiinihappo.
3. Vetysidokset.
Kaikki aminohapot, joissa on hydroksyyli-, amidi- tai karboksyyliryhmiä, ovat mukana.
4. Hydrofobiset vuorovaikutukset . Muodostuu ei-polaaristen radikaalien väliin vesipitoisessa ympäristössä. Mukana on 8 aminohappoa (ensimmäinen luokka).
Proteiinin tertiaarisen rakenteen määrää täysin sen primäärirakenne, ts. aminohapposekvenssi, jonka puolestaan määrää geneettinen koodi.
Hydrofobiset vuorovaikutukset ovat ratkaisevia niiden epäselektiivisyyden (epäspesifisyyden) ja moninkertaisuuden vuoksi.
Useimmissa proteiineissa on hydrofobinen ydin.
Kvaternäärinen proteiinirakenne
Määritelmä: Proteiinin kvaternäärinen rakenne on kahden tai useamman tertiäärisen rakenteen omaavan polypeptidiketjun aggregaatio oligomeeriseksi toiminnallisesti merkittäväksi koostumukseksi.
Kvaternaarisen rakenteen muodostavat ja ylläpitävät sidokset ovat samat kuin ne, jotka muodostavat tertiaarisen rakenteen, lukuun ottamatta hydrofobisia.
N Beetaketjun -pää sisältää polaarista glutamiinihappoa ("-" varautunut). Sirppisoluanemiaa sairastavilla potilailla käytetään sen sijaan ei-polaarista valiinia.574 aminohaposta 2 korvattiin.
Tällainen hemoglobiini menettää liukoisuuden ja muodostuu kuitumainen sakka, joka muuttaa punasoluja.
Sirppisoluanemia on geneettinen sairaus. Syynä on vain yhden nukleotidin korvaaminen hemoglobiini B -ketjua koodaavassa geenissä. Lapset, jotka ovat resessiivisiä homotsygootteja tälle alleelille, eivät elä kaksivuotiaiksi. Heterotsygooteilla on 85 % normaaleja ja 15 % viallisia punasoluja. Hallitsevat homotsygootit kärsivät malariasta, heterotsygootit eivät kärsi malariasta.
Globulaariset ja fibrillaariset proteiinit
95 prosentilla proteiineista on hydrofobinen ydin. 5% fibrillaarisia proteiineja.
Suurin osa pallomaisista proteiineista on liukoisia. Useimmat fibrillaarit ovat liukenemattomia (α-keratiinit - ne muodostavat melkein koko hiusten kuivapainon, villan, sarvet, sorkat, kynnet, suomukset, höyhenet; kollageeni - jänteiden, ruston proteiini; fibroiini - silkkiproteiini).
Fibrillaariset proteiinit sisältävät suuremman osan varautuneita aminohappoja kuin globulaariset - yksittäiset ketjut ovat liukoisia ja niiden kompleksit ovat polaarittomia ja liukenemattomia.
