Ensisijainen rakenne - aminohapposekvenssi polypeptidiketjussa. Proteiinimolekyylissä, kun jäykkiä (peptidisidos) ja taipuisia (a-hiiliatomi) osia vaihdetaan, muodostuu ketjun kompakti järjestely avaruudessa.
Akoborin menetelmä on käyttää fenyylihydratsiini. Fenyylihydratsiini katkaisee peptidisidokset proteiinissa ja kiinnittyy kaikkiin aminohappoihin paitsi C-terminaaliseen. Myöhempi kromatografinen analyysi mahdollistaa proteiinin C-terminaalisen aminohapon tunnistamisen.
Primäärirakenteen tutkiminen on tärkeää yleinen biologinen ja lääketieteellinen merkitys:
- primäärirakenne määrittää myöhemmät proteiinirakenteet.
- proteiinin primaarisen rakenteen tuntemus on välttämätöntä proteiinien keinotekoiselle synteesille.
- primäärirakenne määrää lajispesifisyyden, esimerkiksi insuliiniproteiinissa, yleensä eri eläinlajeissa ja ihmisissä molekyylin keskellä, korvautuu pääsääntöisesti 3 ekvivalenttia aminohappoa.
- primaarirakenteen muutokset voivat johtaa moniin sairauksiin, esimerkiksi sirppisoluanemiaan, jossa β-ketjun hemoglobiinissa asemassa 6 glutamiinihappo korvataan valiinilla. Tämä korvaaminen epätasaisella aminohapolla johtaa hemoglobiinin toiminnan häiriintymiseen ja sirpin muotoisten punasolujen ilmaantumista.
Toissijainen rakenne - polypeptidiketjun säännöllisesti toistuva järjestysmalli avaruudessa. Useimmiten proteiineista löytyy 2 tyyppistä sekundaarirakennetta: α - heliksi ja β - rakenne.
α - heliksi vuonna 1951 sitä tutki L. Pauling röntgendiffraktiomenetelmällä. Se on oikeakätinen kierukkarakenne, jonka yhteen kierrokseen mahtuu 3,6 aminohappoa. Kierteen nousu (viereisten kierrosten välinen etäisyys) on 0,54 n.m. α-heliksi on kiinnitetty vetysidoksilla, jotka sulkeutuvat jokaisen neljännen aminohapon muodostamien peptidisidosten välissä. Toissijainen a-rakenne taittuu spontaanisti ja sen määrää proteiinin primäärirakenne. Kierteiseen rakenteeseen järjestettyjen alueiden osuus vaihtelee eri proteiineissa. Esimerkiksi hemoglobiinissa ja myoglobiinissa α - rakenteellinen laskos on hallitseva, mikä pienentää proteiinimolekyylin kokoa 4 kertaa.
β-rakenne sillä on "haitarin" muoto ja se on stabiloitunut vetysidoksilla yhden polypeptidiketjun etäisten osien tai useiden proteiinimolekyylien välillä. On olemassa rinnakkaisia β-rakenteita, joissa N- ja C-päät vastaavat toisiaan, ja antirinnakkaisrakenteita. Esimerkki proteiineista, jotka sisältävät pääasiassa β-rakenteita, ovat immunoglobuliinit.
Toissijaista rakennetta tutkitaan röntgendiffraktioanalyysillä ja ultraviolettisäteiden absorptiolla proteiiniin (mitä suurempi α-rakenteiden osuus, sitä suurempi absorptio).
Toissijainen rakenne tuhoutuu denaturoinnin aikana.
Tertiäärinen rakenne - kanssa polypeptidiketjun tilajärjestelyn muoto, joka on spesifinen kullekin proteiinille. Tämä rakenne muodostuu spontaanisti ja sen määrää primäärirakenne. Tertiäärinen rakenne lisää merkittävästi proteiinin tiiviyttä kymmenillä. Ei-kovalenttiset sidokset (hydrofobiset, ioniset) ja kovalenttiset (disulfidi) sidokset ovat mukana tertiaarisen rakenteen muodostumisessa.
Tertiäärinen rakenne määrää proteiinien biologisen aktiivisuuden ja fysikaalis-kemialliset ominaisuudet. Jos tertiäärinen rakenne häiriintyy, proteiini menettää biologisen aktiivisuutensa.
Tertiaarisen rakenteen tutkimisen menetelmiä ovat röntgendiffraktioanalyysi ja proteiinin yksittäisten aminohapporadikaalien kemiallisen aktiivisuuden määrittäminen. Myoglobiiniproteiinin tertiääristä rakennetta tutki ensimmäisenä J. Kendrew (1957). M. Perutz (1959) tutki hemoglobiinin rakennetta.
Proteiinien tertiääriseen rakenteeseen kuuluvat α-kierteiset, β-laskostuneet rakenteet, β-silmukat (joissa polypeptidiketju on taipunut 180 0) ja ns. epäjärjestynyt kela. Esimerkiksi insuliiniproteiini sisältää 57 % a-kierteisiä alueita, 6 % p-poimutettuja rakenteita, 10 % molekyylistä on järjestetty p-silmukoiksi ja 27 % molekyylistä on epäjärjestynyt kierukka.
Ensisijaisen, toissijaisen ja tertiaarisen kokonaisuus on konformaatio proteiinimolekyyli. Elinikäinen (natiivi) konformaatio muodostuu spontaanisti ja sen muodostumista kutsutaan taittumaksi. Proteiinien konformaatio on erittäin epävakaa ja muodostuu erityisten proteiinien osallistuessa - saattajia(kumppanit). Chaperonit pystyvät sitoutumaan osittain denaturoituihin proteiineihin, jotka ovat epävakaassa tilassa ja palauttavat alkuperäisen konformaationsa. Chaperonit luokitellaan molekyylipainon (60 - 100 cd.) mukaan. Tutkituimmat ovat Sh-60, Sh-70 ja Sh-90. Esimerkiksi Sh-70 on vuorovaikutuksessa proteiinien kanssa, joissa on runsaasti hydrofobisia radikaaleja ja suojaa niitä korkean lämpötilan denaturaatiolta. Yleensä kaperonit suojaavat kehon pääproteiineja, estävät denaturoitumista ja edistävät konformaation muodostumista, helpottavat denaturoituneiden proteiinien kuljetusta lysosomeihin ja osallistuvat proteiinisynteesiin.
Konformaation mukaan kaikki proteiinit jaetaan kolmeen ryhmään:
- fibrillaariset proteiinit: kollageeni, elastiini, fibroiini.
- Globulaariset proteiinit: hemoglobiini, albumiini, globuliini.
- Sekaproteiinit: myosiini.
Tertiäärinen rakenne on luontainen kaikille proteiineille. Vain oligomeerisillä proteiineilla, jotka sisältävät useita alayksiköitä, protomeerejä, on kvaternäärinen rakenne. Protomeeriä pidetään erillisenä polypeptidiketjuna, alayksikkö on oligomeerisen proteiinin toiminnallisesti aktiivinen osa. Alayksikkö voi sisältää joko yhden protomeerin tai useita.
Kvaternaarirakenne - oligomeeristen proteiinien alayksiköiden lukumäärä ja suhteellinen järjestys. Vain oligomeerisillä proteiineilla, jotka sisältävät useita alayksiköitä, protomeerejä, on kvaternäärinen rakenne. Protomeeriä pidetään erillisenä polypeptidiketjuna, alayksikkö on oligomeerisen proteiinin toiminnallisesti aktiivinen osa. Alayksikkö voi sisältää yhden protomeerin tai useita protomeerejä.
Kvaternaarisen rakenteen muodostumiseen liittyy heikkoja ei-kovalenttisia sidoksia (hydrofobinen, ioni, vety). Proteiinien kvaternäärinen rakenne muodostuu spontaanisti ja hajoaa helposti denaturoituessaan. Oligomeerisen proteiinin yksittäiset alayksiköt ovat vuorovaikutuksessa toistensa kanssa, mikä johtaa muutoksiin yksittäisten protomeerien tertiaarisessa rakenteessa. Tätä ilmiötä kutsutaan yhteistoiminnallisiksi muutoksiksi protomeerin konformaatiossa, ja siihen liittyy yleensä proteiiniaktiivisuuden lisääntyminen.
Oligomeerisillä proteiineilla on monia ominaisuuksia verrattuna monomeerisiin proteiineihin.
- Niillä on erittäin kompakti pakkaus ja suhteellisen pieni rajapinta, joten solunsisäisesti sijaitsevien ne sitovat vähemmän vettä
- Niiden toimintaa säädellään kehossa. Protomeerit ovat yleensä inaktiivisia, mutta oligomeeriset proteiinit ovat paljon aktiivisempia.
- Jos samantyyppiset protomeerit ovat mukana oligomeerisen proteiinin synteesissä, tämä säästää geneettistä materiaalia (useita identtisiä protomeerejä "leimataan" lyhyeen DNA-osaan)
- Ne ovat toiminnallisesti paremmin mukautettuja kehon olosuhteisiin.
Oligomeeristen proteiinien toiminnallisuutta havainnollistetaan vertaamalla proteiineja hemoglobiinia ja myoglobiinia, jotka osallistuvat hapen kuljettamiseen kudoksiin. Punasolujen hemoglobiini on oligomeerinen proteiini, joka sisältää 4 polypeptidiketjua. Lihasmyoglobiini on monomeerinen proteiini, joka sisältää yhden polypeptidiketjun. Myoglobiinin happisaturaatiokäyrä osoittaa sen suoran riippuvuuden happipitoisuudesta. Hemoglobiinin happisaturaatiokäyrä on S-muotoinen. Tämä johtuu asteittaisesta peräkkäisestä muutoksesta hemoglobiinin koostumuksen kunkin neljän protomeerin rakenteessa (konformaatiossa), minkä seurauksena hemoglobiinin affiniteetti happea kohtaan kasvaa jyrkästi. Tämä hemoglobiinin kyllästymisen luonne hapella lisää jyrkästi sen happikapasiteettia myoglobiiniin verrattuna.
Erityinen asema proteiinien joukossa on domeenin proteiinit .
Domeenit ovat rakenteellisesti ja toiminnallisesti erillisiä osia yhdestä polypeptidiketjusta. Domeenit voivat olla vastuussa proteiinin vuorovaikutuksesta erilaisten aineiden - ligandien (pienmolekyyliset aineet, DNA, RNA, polysakkaridit jne.) kanssa. Esimerkkejä domeeniproteiineista ovat seerumin albumiini, immunoglobuliinit ja jotkut entsyymit (haiman trypsiini).
Proteiinien korkean selektiivisyyden vuoksi ne voidaan yhdistää komplekseiksi, joita kutsutaan useimmiten monientsyymikomplekseiksi - nämä ovat useiden entsyymien rakenteellisia assosiaatioita, jotka katalysoivat monimutkaisen kemiallisen prosessin yksittäisiä vaiheita. Esimerkki: p(PDC), kolmen tyyppisen entsyymin kompleksi, joka katalysoi pyruvaattihapon (PVA) hapettumista.
On mahdollista spesifisesti yhdistää yksittäisten proteiinien lisäksi myös proteiineja lipidien (rasvojen) kanssa solukalvojen muodostumisen aikana ja proteiineja nukleiinihappojen kanssa kromatiinin muodostumisen aikana.
Proteiinien fysikaalis-kemialliset ominaisuudet.
Ne määräytyvät suurelta osin proteiinimolekyylin konformaatiosta (proteiinin primääri-tertiaarinen rakenne). Proteiinien fysikaalis-kemialliset ominaisuudet näkyvät liuoksissa.
Liukoisuus proteiinit vaihtelevat proteiinista toiseen.
Yleensä proteiinien liukoisuus on korkea, mutta vaihtelee eri tyyppisten proteiinien välillä. Siihen vaikuttavat seuraavat tekijät:
- proteiinimolekyylin muoto (pallomaiset proteiinit ovat liukoisempia kuin säikeiset proteiinit)
- proteiinin aminohapporadikaalin luonne, polaaristen ei-polaaristen radikaalien suhde (mitä enemmän polaarisia hydrofiilisiä radikaaleja proteiinissa on, sitä parempi sen liukoisuus)
- liuotinominaisuudet, suolojen läsnäolo. Pieni suolapitoisuus (KCL, NaCl) lisää joskus proteiinien liukoisuutta. Esimerkiksi albumiinit liukenevat paremmin puhtaaseen tislattuun veteen, globuliinit liukenevat vain 10 % suolojen (KCL, NaCl) läsnä ollessa. Sidekudoksen proteiinit kollageeni ja elastiini ovat liukenemattomia veteen tai suolaliuokseen.
Molekyylimassa proteiinit ovat melko suuria, vaihtelevat välillä 6 000 - 1 000 000. Esimerkiksi hemoglobiinin molekyylipaino on 68 000, albumiinin 100 000, ribonukleaasin noin 14 000 ja myosiinin 500 000.
Proteiinien moolimassan määritysmenetelmät on oltava lempeä, eikä se saa tuhota proteiinimolekyylejä. Esimerkiksi ebullioskooppinen menetelmä, joka perustuu liuosten kiehumispisteen mittaamiseen, ei sovellu proteiineihin. Tarkimmat menetelmät proteiinien molekyylipainon määrittämiseen ovat ultrasentrifugointimenetelmä ja röntgendiffraktiomenetelmä.
Ultrasentrifugointimenetelmä(sedimentaatio) perustuu muutokseen eri molekyylipainoisten proteiinien sedimentaationopeudessa pyöritettäessä proteiiniliuoksia suurella nopeudella. Tällä menetelmällä löydettyjen proteiinien molekyylipaino on merkitty Svedbergin yksiköllä (S = 10 -13 c.)
Röntgendiffraktiomenetelmä voit laskea molekyylipainon analysoimalla useita röntgenkuvia proteiinimolekyylistä.
Elektroforeettinen menetelmä perustuu proteiinin liikkumisnopeuden riippuvuuteen jatkuvassa sähkökentässä proteiinin molekyylipainosta (elektroforeettinen liikkuvuus on suurempi proteiineille, joilla on pienempi molekyylipaino)
Kromatografinen menetelmä perustuu eri proteiinien eri nopeuksiin molekyyligeelin "seulojen" läpi.
Suuret molekyylit, jotka ovat suuremmat kuin geelin huokoskoko, kulkevat geelin läpi nopeammin kuin pienemmät proteiinimolekyylit, jotka jäävät geelirakeiden sisään.
Elektronimikroskooppinen menetelmä suoritetaan vertaamalla proteiinimolekyylin kokoa tunnetun massan omaaviin vertailunäytteisiin.
Kemialliset menetelmät liittyvät proteiinien kemiallisen koostumuksen ominaisuuksiin
Proteiinimolekyylien muoto eri. Proteiinimolekyylit voivat olla muodoltaan säikeisiä tai pallomaisia. Fibrillaarisilla proteiineilla on lankamainen molekyylimuoto. Ne ovat yleensä liukenemattomia veteen ja laimeaan suolaliuokseen. Fibrillaarisia proteiineja ovat sidekudoksen tärkeimmät rakenneproteiinit: kollageeni, keratiini, elastiini. Globulaarisissa proteiineissa polypeptidiketjut on kietoutunut tiiviiksi tiiviiksi pallomaiseksi rakenteeksi. Useimmat pallomaiset proteiinit liukenevat hyvin veteen ja heikkoihin suolaliuoksiin. Globulaarisia proteiineja ovat entsyymit, vasta-aineet, albumiinit ja hemoglobiini. Joillakin proteiineilla on välityyppinen molekyyli, joka sisältää sekä lankamaisia että pallomaisia alueita. Esimerkki tällaisista proteiineista on lihasproteiini myosiini, joka liukenee suolaliuoksiin.
Proteiinimolekyylien koot ovat välillä 1-100 nm, lähellä kolloidisten hiukkasten kokoa. Tämän vuoksi proteiiniliuoksilla on sekä todellisten liuosten että kolloidisten liuosten ominaisuuksia.
Monet proteiiniliuosten molekyylikineettiset ominaisuudet ovat samanlaisia kolloidisten liuosten ominaisuudet .
- Niiden vaihtoon tarvittavien proteiinien hidas diffuusionopeus.
- Proteiinien kyvyttömyys kulkea puoliläpäisevien kalvojen läpi. Osastoissa, joissa on korkea proteiinipitoisuus, syntyy ylimääräistä hydrostaattista painetta, koska vesimolekyylit liikkuvat yksisuuntaisesti puoliläpäisevän kalvon läpi kohti korkeaa proteiinipitoisuutta. Proteiinien aiheuttamaa ylipainetta kutsutaan onkoottiseksi paineeksi. Se on tärkeä tekijä, joka määrää veden liikkeen kudosten, veren ja suoliston välillä.
- Proteiinien korkea viskositeetti johtuu suurten proteiinimolekyylien erilaisista molekyylien välisistä vuorovaikutuksista. Erityisesti lisääntynyt veren viskositeetti lisää sydänlihaksen kuormitusta.
- Jotkut proteiinit pystyvät muodostamaan geelejä, mikä lisää proteiinien vahvuutta (esim. kollageeni).
Proteiinien optiset ominaisuudet määräytyy proteiinimolekyylien koon, proteiinien aminohapporadikaalien rakenteen, peptidisidosten ja proteiinien alfakierteisten alueiden läsnäolon perusteella.
- Proteiiniliuokset ovat valon taittumisen (taittumisen) ja valonsirontavaikutus. Nämä ominaisuudet johtuvat proteiinimolekyylien suuresta koosta, joka on oikeassa suhteessa spektrin näkyvän osan aallonpituuteen. Tässä tapauksessa lyhyet siniset säteet ovat sironneet enemmän kuin pitkän aallonpituuden punaiset säteet. Taittumisaste on verrannollinen proteiiniliuoksen pitoisuuteen.
- Proteiiniliuokset absorboivat ultraviolettisäteilyä alueella 190-230 nm johtuen peptidisidosten läsnäolosta ja alueella 260-280 nm johtuen syklisten aminohappojen läsnäolosta proteiineissa. UV-absorptioaste on verrannollinen liuoksen proteiinipitoisuuteen.
- Proteiiniliuokset voivat kiertää polarisoidun valon tasoa, mikä johtuu proteiinin sisältämien aminohappojen optisesta aktiivisuudesta ja alfakierteisten osien läsnäolosta siinä. Valon polarisaation ja liuoksen proteiinipitoisuuden välillä on suora yhteys.
Proteiineilla, jotka ovat molekyyliliuoksia, on todellisten ratkaisujen ominaisuuksia . Todellisina ratkaisuina proteiiniliuokset ovat erittäin stabiileja.
MODUULI 1 PROTEIINIEN RAKENNE, OMINAISUUDET JA TOIMINNOT
MODUULI 1 PROTEIINIEN RAKENNE, OMINAISUUDET JA TOIMINNOT
Moduulin rakenne | Teemat |
Modulaarinen yksikkö 1 | 1.1. Proteiinien rakenneorganisaatio. Natiiviproteiinin konformaation muodostumisvaiheet 1.2. Proteiinin toiminnan perusteet. Lääkkeet ligandeina, jotka vaikuttavat proteiinien toimintaan 1.3. Proteiinien denaturoituminen ja niiden spontaanin uudelleensyntymisen mahdollisuus |
Moduuliyksikkö 2 | 1.4. Oligomeeristen proteiinien rakenteen ja toiminnan ominaisuudet hemoglobiinin esimerkin avulla 1.5. Natiiviproteiinin konformaation ylläpito soluolosuhteissa 1.6. Erilaisia proteiineja. Proteiiniperheet käyttämällä esimerkkiä immunoglobuliinista 1.7. Proteiinien fysikaalis-kemialliset ominaisuudet ja menetelmät niiden erottamiseen |
Moduuliyksikkö 1 MONOMEERISTEN PROTEIINIEN RAKENNEJÄRJESTELY JA NIIDEN TOIMINNAN PERUSTEET
Oppimistavoitteet Pystyy:
1. Käytä tietoa proteiinien rakenteellisista ominaisuuksista ja proteiinien toimintojen riippuvuudesta niiden rakenteesta ymmärtääksesi perinnöllisten ja hankittujen proteinopatioiden kehittymismekanismeja.
2. Selitä joidenkin lääkkeiden terapeuttisen vaikutuksen mekanismeja ligandeina, jotka ovat vuorovaikutuksessa proteiinien kanssa ja muuttavat niiden aktiivisuutta.
3. Käytä tietoa proteiinien rakenteesta ja konformationaalisesta labilisuudesta ymmärtääksesi niiden rakenteellista ja toiminnallista epästabiilisuutta ja taipumusta denaturoitua muuttuvissa olosuhteissa.
4. Selitä denaturointiaineiden käyttö lääketieteellisten materiaalien ja instrumenttien steriloinnissa sekä antiseptisinä aineina.
Tietää:
1. Proteiinien rakenteellisen organisoinnin tasot.
2. Proteiinien primäärirakenteen merkitys, joka määrää niiden rakenteellisen ja toiminnallisen monimuotoisuuden.
3. Proteiinien aktiivisen keskuksen muodostumismekanismi ja sen spesifinen vuorovaikutus ligandin kanssa, joka on proteiinien toiminnan taustalla.
4. Esimerkkejä eksogeenisten ligandien (lääkkeet, toksiinit, myrkyt) vaikutuksesta proteiinien konformaatioon ja toiminnalliseen aktiivisuuteen.
5. Proteiinien denaturoitumisen syyt ja seuraukset, denaturaatiota aiheuttavat tekijät.
6. Esimerkkejä denaturoivien tekijöiden käytöstä lääketieteessä antiseptisinä aineina ja keinoina lääketieteellisten instrumenttien sterilointiin.
AIHE 1.1. PROTEIINIEN RAKENNEJÄRJESTELY. ALKUPERÄISEN MUODOSTUSVAIHEET
PROTEIINIKONFORMAATIOT
Proteiinit ovat polymeerimolekyylejä, joiden monomeerit ovat vain 20 α-aminohappoa. Aminohappojen yhdistelmä ja järjestys proteiinissa määräytyy yksilöiden DNA:ssa olevien geenien rakenteen perusteella. Jokainen proteiini suorittaa oman tehtävänsä erityisrakenteensa mukaisesti. Tietyn organismin proteiinien joukko määrittää sen fenotyyppiset ominaisuudet, samoin kuin perinnöllisten sairauksien esiintymisen tai taipumuksen niiden kehittymiseen.
1. Aminohapot, jotka muodostavat proteiineja. Peptidisidos. Proteiinit ovat polymeerejä, jotka on rakennettu monomeereistä - 20 α-aminohaposta, joiden yleinen kaava on
Aminohapot eroavat α-hiiliatomiin kiinnittyneiden radikaalien rakenteesta, koosta ja fysikaalis-kemiallisista ominaisuuksista. Aminohappojen funktionaaliset ryhmät määräävät eri α-aminohappojen ominaisuuksien ominaisuudet. α-aminohapoissa esiintyvät radikaalit voidaan jakaa useisiin ryhmiin:
proliini, Toisin kuin muut 19 proteiinimonomeeriä, se ei ole aminohappo, vaan iminohappo, proliinissa oleva radikaali liittyy sekä a-hiiliatomiin että iminoryhmään
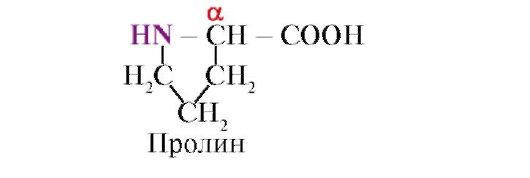 Aminohappojen liukoisuus veteen vaihtelee. Tämä johtuu radikaalien kyvystä olla vuorovaikutuksessa veden (hydraatin) kanssa.
Aminohappojen liukoisuus veteen vaihtelee. Tämä johtuu radikaalien kyvystä olla vuorovaikutuksessa veden (hydraatin) kanssa.
TO hydrofiilinen Käsitteet sisältävät anionisia, kationisia ja polaarisia varautumattomia funktionaalisia ryhmiä sisältävät radikaalit.
TO hydrofobinen Käsitteet sisältävät radikaaleja, jotka sisältävät metyyliryhmiä, alifaattisia ketjuja tai renkaita.
2. Peptidisidokset yhdistävät aminohappoja muodostaen peptidejä. Peptidisynteesin aikana yhden aminohapon α-karboksyyliryhmä on vuorovaikutuksessa toisen aminohapon α-aminoryhmän kanssa muodostaen peptidisidos:
Proteiinit ovat polypeptidejä, ts. α-aminohappojen lineaariset polymeerit, jotka on yhdistetty peptidisidoksella (kuva 1.1.)
 Riisi. 1.1. Termit, joita käytetään kuvaamaan peptidien rakennetta
Riisi. 1.1. Termit, joita käytetään kuvaamaan peptidien rakennetta
Aminohappojen monomeerejä, jotka muodostavat polypeptidejä, kutsutaan aminohappotähteet. Toistuvien ryhmien ketju - NH-CH-CO- lomakkeet peptidirunko. Aminohappotähdettä, jossa on vapaa a-aminoryhmä, kutsutaan N-päätteeksi, ja aminohappotähdettä, jossa on vapaa a-karboksyyliryhmä, kutsutaan C-päätteeksi. Peptidit kirjoitetaan ja luetaan N-päästä C-päähän.
Proliinin iminoryhmän muodostama peptidisidos eroaa muista peptidisidoksista: peptidiryhmän typpiatomista puuttuu vety,
sen sijaan on olemassa sidos radikaalin kanssa, jonka seurauksena renkaan toinen puoli sisältyy peptidirunkoon:
 Peptidit eroavat aminohappokoostumuksesta, aminohappojen lukumäärästä ja aminohappoliitoksen järjestyksestä, esimerkiksi Ser-Ala-Glu-Gis ja His-Glu-Ala-Ser ovat kaksi eri peptidiä.
Peptidit eroavat aminohappokoostumuksesta, aminohappojen lukumäärästä ja aminohappoliitoksen järjestyksestä, esimerkiksi Ser-Ala-Glu-Gis ja His-Glu-Ala-Ser ovat kaksi eri peptidiä.
Peptidisidokset ovat erittäin vahvoja ja niiden kemiallinen ei-entsymaattinen hydrolyysi vaatii ankaria olosuhteita: analysoitava proteiini hydrolysoituu väkevässä suolahapossa noin 110°:n lämpötilassa 24 tunnin ajan. Elävässä solussa peptidisidokset voivat katketa proteolyyttiset entsyymit, nimeltään proteaasit tai peptidihydrolaasit.
3. Proteiinien perusrakenne. Aminohappotähteet eri proteiinien peptidiketjuissa eivät vuorottele satunnaisesti, vaan ne on järjestetty tiettyyn järjestykseen. Polypeptidiketjun aminohappotähteiden lineaarista sekvenssiä tai vuorottelujärjestystä kutsutaan proteiinin primaarirakenne.
Kunkin yksittäisen proteiinin primäärirakenne on koodattu DNA-molekyylissä (alueella, jota kutsutaan geeniksi) ja se toteutuu transkription (informaation kopioiminen mRNA:lle) ja translaation (proteiinin primäärirakenteen synteesi) aikana. Näin ollen yksittäisen ihmisen proteiinien primäärirakenne on vanhemmilta lapsille perinnöllisesti välittyvä tieto, joka määrää tietyn organismin proteiinien rakenteelliset ominaisuudet, joista olemassa olevien proteiinien toiminta riippuu (kuva 1.2.).
 Riisi. 1.2. Genotyypin ja yksilön kehossa syntetisoitujen proteiinien konformaation välinen suhde
Riisi. 1.2. Genotyypin ja yksilön kehossa syntetisoitujen proteiinien konformaation välinen suhde
Jokaisella ihmiskehon noin 100 000 yksittäisestä proteiinista on ainutlaatuinen ensisijainen rakenne. Samantyyppisten proteiinien (esimerkiksi albumiinin) molekyyleillä on sama aminohappotähteiden vuorottelu, mikä erottaa albumiinin kaikista muista yksittäisistä proteiineista.
Peptidiketjun aminohappotähteiden sekvenssiä voidaan pitää tiedon tallennusmuotona. Tämä tieto määrittää lineaarisen peptidiketjun tilajärjestelyn kompaktimmaksi kolmiulotteiseksi rakenteeksi ns. konformaatio orava. Funktionaalisesti aktiivisen proteiinikonformaation muodostumisprosessia kutsutaan taitettava
4. Proteiinin rakenne. Vapaa pyöriminen peptidirungossa on mahdollista peptidiryhmän typpiatomin ja viereisen a-hiiliatomin välillä sekä a-hiiliatomin ja karbonyyliryhmän hiilen välillä. Aminohappotähteiden funktionaalisten ryhmien vuorovaikutuksen ansiosta proteiinien primäärirakenne voi saada monimutkaisempia tilarakenteita. Globulaarisissa proteiineissa on kaksi pääasiallista peptidiketjujen konformaation laskostumistasoa: toissijainen Ja tertiäärinen rakenne.
Proteiinien toissijainen rakenne on spatiaalinen rakenne, joka muodostuu vetysidosten muodostumisen seurauksena peptidirungon funktionaalisten ryhmien -C=O ja -NH- välille. Tässä tapauksessa peptidiketju voi hankkia kahden tyyppisiä säännöllisiä rakenteita: α-heliksit Ja β-rakenteet.
SISÄÄN α-heliksit vetysidoksia muodostuu karbonyyliryhmän happiatomin ja 4. aminohapon amiditypen vedyn välille; aminohappotähteiden sivuketjut
sijaitsevat spiraalin kehällä osallistumatta toisiorakenteen muodostukseen (kuva 1.3.).
Bulkkiradikaalit tai radikaalit, joissa on samat varaukset, estävät α-kierteen muodostumisen. Proliinitähde, jolla on rengasrakenne, katkaisee a-heliksin, koska vedyn puuttumisen vuoksi peptidiketjun typpiatomissa on mahdotonta muodostaa vetysidosta. Typen ja a-hiiliatomin välinen sidos on osa proliinirengasta, joten peptidirunko taipuu tässä vaiheessa.
β-rakenne muodostuu yhden polypeptidiketjun peptidirungon lineaaristen alueiden väliin, jolloin muodostuu laskostettuja rakenteita. Polypeptidiketjuja tai niiden osia voi muodostua rinnakkain tai antirinnakkaiset β-rakenteet. Ensimmäisessä tapauksessa vuorovaikutuksessa olevien peptidiketjujen N- ja C-päät ovat yhtenevät, ja toisessa niillä on vastakkainen suunta (kuva 1.4).
Riisi. 1.3. Proteiinin sekundaarirakenne - α-heliksi
 Riisi. 1.4. Rinnakkaiset ja vastasuuntaiset β-levyrakenteet
Riisi. 1.4. Rinnakkaiset ja vastasuuntaiset β-levyrakenteet
β-rakenteet on merkitty leveillä nuolilla: A - Vastasuuntainen β-rakenne. B - Rinnakkaiset β-levyrakenteet
Joissakin proteiineissa β-rakenteita voi muodostua johtuen vetysidosten muodostumisesta eri polypeptidiketjujen peptidirungon atomien välille.
Löytyy myös proteiineista alueet, joilla on epäsäännöllinen toissijainen rakenne, joka sisältää polypeptidirungon mutkia, silmukoita ja käänteitä. Ne sijaitsevat usein paikoissa, joissa peptidiketjun suunta muuttuu, esimerkiksi kun muodostuu yhdensuuntainen β-levyrakenne.
α-heliksien ja β-rakenteiden läsnäolon perusteella pallomaiset proteiinit voidaan jakaa neljään luokkaan.
Riisi. 1.5. Myoglobiinin (A) ja hemoglobiinin β-ketjun (B) toissijainen rakenne, joka sisältää kahdeksan α-heliksiä

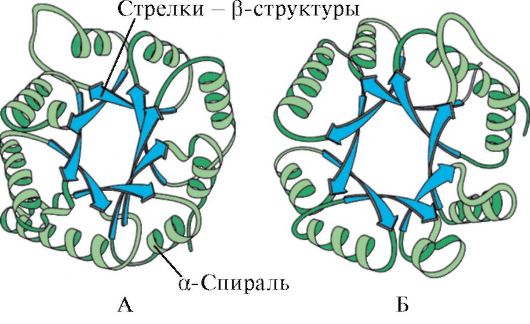 Riisi. 1.6. Triosefosfaatti-isomeraasin ja pyruvaattikinaasidomeenin toissijainen rakenne
Riisi. 1.6. Triosefosfaatti-isomeraasin ja pyruvaattikinaasidomeenin toissijainen rakenne
 Riisi. 1.7. Immunoglobuliinin (A) vakiodomeenin ja superoksididismutaasientsyymin (B) toissijainen rakenne
Riisi. 1.7. Immunoglobuliinin (A) vakiodomeenin ja superoksididismutaasientsyymin (B) toissijainen rakenne
SISÄÄN neljäs luokka sisälsi proteiineja, jotka sisältävät pienen määrän säännöllisiä sekundaarisia rakenteita. Nämä proteiinit sisältävät pieniä kysteiinipitoisia proteiineja tai metalloproteiineja.
Proteiinin tertiäärinen rakenne- aminohapporadikaalien välisistä vuorovaikutuksista muodostuva konformaatiotyyppi, joka voi sijaita huomattavan etäisyyden päässä toisistaan peptidiketjussa. Useimmat proteiinit muodostavat spatiaalisen rakenteen, joka muistuttaa palloa (globulaariset proteiinit).
Koska hydrofobiset aminohapporadikaalit pyrkivät yhdistymään ns hydrofobisia vuorovaikutuksia ja molekyylien välisten van der Waalsin voimien vuoksi proteiinipallon sisään muodostuu tiheä hydrofobinen ydin. Hydrofiiliset ionisoidut ja ionisoimattomat radikaalit sijaitsevat pääasiassa proteiinin pinnalla ja määrittävät sen vesiliukoisuuden.
 Riisi. 1.8 Sidostyypit, jotka syntyvät aminohapporadikaalien välille proteiinin tertiaarisen rakenteen muodostumisen aikana
Riisi. 1.8 Sidostyypit, jotka syntyvät aminohapporadikaalien välille proteiinin tertiaarisen rakenteen muodostumisen aikana
1 - ionisidos- esiintyy positiivisesti ja negatiivisesti varautuneiden funktionaalisten ryhmien välillä;
2 - vetysidos- esiintyy hydrofiilisen varaamattoman ryhmän ja minkä tahansa muun hydrofiilisen ryhmän välillä;
3 - hydrofobisia vuorovaikutuksia- syntyvät hydrofobisten radikaalien välillä;
4 - disulfidisidos- muodostuu kysteiinitähteiden SH-ryhmien hapettumisesta ja niiden vuorovaikutuksesta keskenään
Hydrofiiliset aminohappotähteet, jotka sijaitsevat hydrofobisen ytimen sisällä, voivat olla vuorovaikutuksessa toistensa kanssa käyttämällä ioninen Ja vetysidoksia(Kuva 1.8).
Ioni- ja vetysidokset sekä hydrofobiset vuorovaikutukset ovat heikkoja: niiden energia ei ole paljon suurempi kuin molekyylien lämpöliikkeen energia huoneenlämpötilassa. Proteiinin konformaatiota ylläpidetään muodostamalla monia tällaisia heikkoja sidoksia. Koska proteiinin muodostavat atomit ovat jatkuvassa liikkeessä, on mahdollista rikkoa joitakin heikkoja sidoksia ja muodostaa muita, mikä johtaa polypeptidiketjun yksittäisten osien vähäisiin liikkeisiin. Tätä proteiinien ominaisuutta muuttaa konformaatiota joidenkin hajoamisen ja muiden heikkojen sidosten muodostumisen seurauksena kutsutaan ns. konformationaalinen labilisuus.
Ihmiskehossa on järjestelmät, jotka tukevat homeostaasi- sisäisen ympäristön pysyvyys tietyissä terveen kehon hyväksyttävissä rajoissa. Homeostaasin olosuhteissa pienet muutokset konformaatiossa eivät häiritse proteiinien yleistä rakennetta ja toimintaa. Proteiinin toiminnallisesti aktiivista konformaatiota kutsutaan syntyperäinen rakenne. Muutokset sisäisessä ympäristössä (esim. glukoosin, Ca-ionien, protonien jne. pitoisuus) johtavat konformaatiomuutoksiin ja proteiinien toimintojen häiriintymiseen.
Joidenkin proteiinien tertiäärinen rakenne on stabiloitunut disulfidisidokset, muodostuu kahden tähteen -SH-ryhmien vuorovaikutuksesta
 Riisi. 1.9. Disulfidisidoksen muodostuminen proteiinimolekyylissä
Riisi. 1.9. Disulfidisidoksen muodostuminen proteiinimolekyylissä
kysteiini (kuva 1.9). Useimpien solunsisäisten proteiinien tertiäärisessä rakenteessa ei ole kovalenttisia disulfidisidoksia. Niiden läsnäolo on ominaista solun erittämille proteiineille, mikä varmistaa niiden paremman stabiilisuuden solunulkoisissa olosuhteissa. Siten disulfidisidoksia on läsnä insuliinin ja immunoglobuliinien molekyyleissä.
Insuliini- proteiinihormoni, joka syntetisoituu haiman β-soluissa ja erittyy vereen vasteena veren glukoosipitoisuuden nousulle. Insuliinin rakenteessa on kaksi disulfidisidosta, jotka yhdistävät polypeptidin A- ja B-ketjut, ja yksi disulfidisidos A-ketjussa (kuva 1.10).
 Riisi. 1.10. Disulfidisidokset insuliinin rakenteessa
Riisi. 1.10. Disulfidisidokset insuliinin rakenteessa
5. Proteiinien ylisekundaarinen rakenne. Proteiineissa, joilla on erilainen primäärirakenne ja -toiminnot, niitä joskus havaitaan samanlaiset yhdistelmät ja toissijaisten rakenteiden suhteelliset sijainnit, joita kutsutaan ylisekundaariseksi rakenteeksi. Se on väliasemassa sekundaari- ja tertiaarirakenteiden välillä, koska se on sekundaarirakenteen elementtien spesifinen yhdistelmä proteiinin tertiaarisen rakenteen muodostuksessa. Supersekundaarisilla rakenteilla on erityiset nimet, kuten "α-helix-turn-a-helix", "leusiinivetoketju", "sinkkisormet" jne. Tällaiset supersekundaariset rakenteet ovat ominaisia DNA:ta sitoville proteiineille.
"Leusiinivetoketju." Tämän tyyppistä ylisekundaarista rakennetta käytetään yhdistämään kaksi proteiinia yhteen. Vuorovaikutteisten proteiinien pinnalla on α-kierteisiä alueita, jotka sisältävät vähintään neljä leusiinitähdettä. Leusiinitähteet α-heliksissä sijaitsevat kuuden aminohapon päässä toisistaan. Koska jokainen α-heliksin kierros sisältää 3,6 aminohappotähdettä, leusiiniradikaalit sijaitsevat jokaisen toisen kierroksen pinnalla. Yhden proteiinin α-heliksin leusiinitähteet voivat olla vuorovaikutuksessa toisen proteiinin leusiinitähteiden kanssa (hydrofobiset vuorovaikutukset), jotka yhdistävät ne toisiinsa (kuva 1.11.). Monet DNA:ta sitovat proteiinit toimivat oligomeerisissa komplekseissa, joissa yksittäiset alayksiköt on liitetty toisiinsa "leusiinivetoketjuilla".
 Riisi. 1.11. "Leusiinivetoketju" kahden proteiinin α-kierteisten alueiden välissä
Riisi. 1.11. "Leusiinivetoketju" kahden proteiinin α-kierteisten alueiden välissä
Esimerkki sellaisista proteiineista ovat histonit. Histonit- ydinproteiinit, jotka sisältävät suuren määrän positiivisesti varautuneita aminohappoja - arginiinia ja lysiiniä (jopa 80%). Histonimolekyylit yhdistetään oligomeerisiksi komplekseiksi, jotka sisältävät kahdeksan monomeeriä käyttämällä "leusiinivetoketjuja", huolimatta näiden molekyylien merkittävästä homonyymistä varauksesta.
"sinkkisormi"- DNA:ta sitoville proteiineille tyypillinen supersekundäärisen rakenteen variantti on proteiinin pinnalla pitkänomaisen fragmentin muotoinen ja sisältää noin 20 aminohappotähdettä (kuva 1.12). "Jatketun sormen" muotoa tukee sinkkiatomi, joka on sitoutunut neljään aminohapporadikaaliin - kahteen kysteiinitähteeseen ja kahteen histidiinitähteeseen. Joissakin tapauksissa histidiinitähteiden sijasta on kysteiinijäämiä. Kaksi lähellä olevaa kysteiinitähdettä on erotettu kahdesta muusta Gisili-tähteestä Cys-sekvenssillä, joka koostuu noin 12 aminohappotähteestä. Tämä proteiinin alue muodostaa a-heliksin, jonka radikaalit voivat sitoutua spesifisesti DNA:n suuren uran säätelyalueisiin. Yksilöllinen sitoutumisspesifisyys
 Riisi. 1.12. DNA:ta sitovien proteiinien alueen ensisijainen rakenne, joka muodostaa "sinkkisormen" rakenteen (kirjaimet osoittavat aminohappoja, jotka muodostavat tämän rakenteen)
Riisi. 1.12. DNA:ta sitovien proteiinien alueen ensisijainen rakenne, joka muodostaa "sinkkisormen" rakenteen (kirjaimet osoittavat aminohappoja, jotka muodostavat tämän rakenteen)
Säätelevä DNA:ta sitova proteiini riippuu sinkkisormen alueella sijaitsevien aminohappotähteiden sekvenssistä. Tällaiset rakenteet sisältävät erityisesti steroidihormonireseptoreita, jotka osallistuvat transkription säätelyyn (informaation lukeminen DNA:sta RNA:ksi).
AIHE 1.2. PROTEIININ TOIMINNAN PERUSTEET. LAIMET LIGANDEINA, JOTKA VAIKUTTAVAT PROTEIIINITOIMINTOA
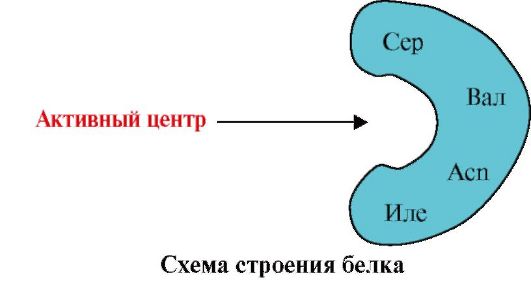
1. Proteiinin aktiivinen keskus ja sen vuorovaikutus ligandin kanssa. Tertiäärisen rakenteen muodostumisen aikana toiminnallisesti aktiivisen proteiinin pinnalle muodostuu yleensä syvennykseen alue, jonka muodostavat primäärirakenteessa kaukana toisistaan olevat aminohapporadikaalit. Tätä aluetta, jolla on ainutlaatuinen rakenne tietylle proteiinille ja joka pystyy spesifisesti vuorovaikuttamaan tietyn molekyylin tai samankaltaisten molekyylien ryhmän kanssa, kutsutaan proteiini-ligandin sitomispaikaksi tai aktiiviseksi paikaksi. Ligandit ovat molekyylejä, jotka ovat vuorovaikutuksessa proteiinien kanssa.
Korkea spesifisyys Proteiinin vuorovaikutus ligandin kanssa varmistetaan aktiivisen keskuksen rakenteen komplementaarisella ligandin rakenteen kanssa.
Täydentävyys- tämä on vuorovaikutuksessa olevien pintojen avaruudellinen ja kemiallinen vastaavuus. Aktiivisen keskuksen tulee paitsi avaruudellisesti vastata siihen kuuluvaa ligandia, myös sidoksia (ioni-, vety- ja hydrofobisia vuorovaikutuksia) tulee muodostua aktiiviseen keskukseen sisältyvien radikaalien funktionaalisten ryhmien ja ligandin pitävän ligandin välille. aktiivisessa keskustassa (kuva 1.13 ).
 Riisi. 1.13. Proteiinin täydentävä vuorovaikutus ligandin kanssa
Riisi. 1.13. Proteiinin täydentävä vuorovaikutus ligandin kanssa
Joillakin ligandeilla, kun ne ovat kiinnittyneet proteiinin aktiiviseen keskukseen, on apurooli proteiinien toiminnassa. Tällaisia ligandeja kutsutaan kofaktoreiksi, ja proteiineja, jotka sisältävät ei-proteiiniosan, kutsutaan monimutkaiset proteiinit(toisin kuin yksinkertaiset proteiinit, jotka koostuvat vain proteiiniosasta). Ei-proteiiniosaa, joka on kiinteästi liitetty proteiiniin, kutsutaan prosteettinen ryhmä. Esimerkiksi myoglobiini, hemoglobiini ja sytokromit sisältävät proteettisen ryhmän, hemin, joka sisältää rautaionin, joka on kiinteästi kiinnittynyt aktiiviseen keskustaan. Hemeä sisältäviä monimutkaisia proteiineja kutsutaan hemoproteiineiksi.
Kun spesifisiä ligandeja kiinnittyy proteiineihin, näiden proteiinien toiminta ilmenee. Siten albumiini, veriplasman tärkein proteiini, osoittaa kuljetustoimintonsa kiinnittämällä aktiiviseen keskukseen hydrofobisia ligandeja, kuten rasvahappoja, bilirubiinia, joitain lääkkeitä jne. (Kuva 1.14)
Peptidiketjun kolmiulotteisen rakenteen kanssa vuorovaikutuksessa olevat ligandit voivat olla paitsi pienimolekyylisiä orgaanisia ja epäorgaanisia molekyylejä, myös makromolekyylejä:
DNA (esimerkkejä edellä käsitellyistä DNA:ta sitovista proteiineista);
polysakkaridit;
 Riisi. 1.14. Genotyypin ja fenotyypin välinen suhde
Riisi. 1.14. Genotyypin ja fenotyypin välinen suhde
DNA-molekyyliin koodattu ihmisen proteiinien ainutlaatuinen primäärirakenne toteutuu soluissa ainutlaatuisen konformaation, aktiivisen keskusrakenteen ja proteiinitoimintojen muodossa.
Näissä tapauksissa proteiini tunnistaa ligandin spesifisen alueen, joka on suhteessa sitoutumiskohtaan ja komplementaarinen. Siten maksasolujen pinnalla on insuliinihormonin reseptoriproteiineja, jolla on myös proteiinirakenne. Insuliinin vuorovaikutus reseptorin kanssa aiheuttaa muutoksen sen konformaatiossa ja signaalijärjestelmien aktivoitumisen, mikä johtaa ravinteiden varastoimiseen maksasoluihin aterioiden jälkeen.
Täten, Proteiinien toiminta perustuu proteiinin aktiivisen keskuksen spesifiseen vuorovaikutukseen ligandin kanssa.
2. Domeenirakenne ja sen rooli proteiinien toiminnassa. Globulaaristen proteiinien pitkät polypeptidiketjut laskostuvat usein useiksi kompakteiksi, suhteellisen itsenäisiksi alueiksi. Niillä on itsenäinen tertiäärinen rakenne, joka muistuttaa globulaaristen proteiinien rakennetta, ja niitä kutsutaan verkkotunnuksia. Proteiinien domeenirakenteen ansiosta niiden tertiäärinen rakenne on helpompi muodostaa.
Domeeniproteiineissa ligandia sitovat kohdat sijaitsevat usein domeenien välissä. Siten trypsiini on proteolyyttinen entsyymi, jota haiman eksokriininen osa tuottaa ja joka on välttämätön ruokaproteiinien sulamiselle. Sillä on kaksidomeenirakenne, ja trypsiinin ja sen ligandin - ruokaproteiinin - sitoutumiskeskus sijaitsee näiden kahden domeenin välisessä urassa. Aktiivisessa keskustassa luodaan olosuhteet, jotka ovat välttämättömiä ruokaproteiinin tietyn kohdan tehokkaalle sitoutumiselle ja sen peptidisidosten hydrolyysille.
Proteiinin eri domeenit voivat liikkua suhteessa toisiinsa, kun aktiivinen keskus on vuorovaikutuksessa ligandin kanssa (kuva 1.15).
Heksokinaasi- entsyymi, joka katalysoi glukoosin fosforylaatiota ATP:n avulla. Entsyymin aktiivinen kohta sijaitsee kahden domeenin välisessä rakossa. Kun heksokinaasi sitoutuu glukoosiin, sitä ympäröivät domeenit sulkeutuvat ja substraatti jää loukkuun, jossa tapahtuu fosforylaatiota (katso kuva 1.15).
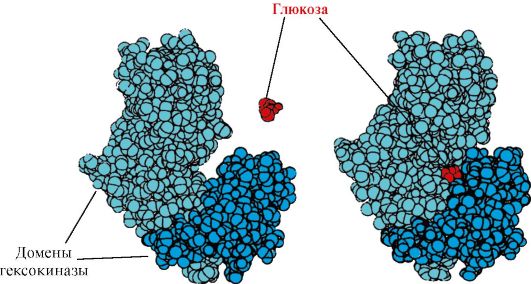 Riisi. 1.15. Heksokinaasidomeenien sitoutuminen glukoosiin
Riisi. 1.15. Heksokinaasidomeenien sitoutuminen glukoosiin
Joissakin proteiineissa domeenit suorittavat itsenäisiä toimintoja sitoutumalla erilaisiin ligandeihin. Tällaisia proteiineja kutsutaan monitoimisiksi.
3. Lääkkeet ovat ligandeja, jotka vaikuttavat proteiinien toimintaan. Proteiinien vuorovaikutus ligandien kanssa on spesifistä. Proteiinin ja sen aktiivisen keskuksen konformaatiolabiteetin vuoksi on kuitenkin mahdollista valita toinen aine, joka voisi myös olla vuorovaikutuksessa proteiinin kanssa aktiivisessa keskustassa tai muussa molekyylin osassa.
Ainetta, joka on rakenteeltaan samanlainen kuin luonnollinen ligandi, kutsutaan ligandin rakenteellinen analogi tai ei-luonnollinen ligandi. Se on myös vuorovaikutuksessa proteiinin kanssa aktiivisessa kohdassa. Ligandin rakenteellinen analogi voi molemmat parantaa proteiinin toimintaa (agonisti), ja vähentää sitä (antagonisti). Ligandi ja sen rakenteelliset analogit kilpailevat toistensa kanssa sitoutumisesta proteiiniin samassa kohdassa. Tällaisia aineita kutsutaan kilpailukykyiset modulaattorit proteiinitoimintojen säätelijät. Monet lääkkeet toimivat proteiinin estäjinä. Jotkut niistä saadaan luonnollisten ligandien kemiallisella modifioinnilla. Proteiinitoimintojen estäjiä voivat olla lääkkeet ja myrkyt.
Atropiini on M-kolinergisten reseptorien kilpaileva estäjä. Asetyylikoliini on välittäjäaine hermoimpulssien välittämiseen kolinergisten synapsien kautta. Herätyksen suorittamiseksi synaptiseen rakoon vapautuvan asetyylikoliinin on oltava vuorovaikutuksessa postsynaptisen kalvon reseptoriproteiinin kanssa. Kaksi tyyppiä löytyi kolinergiset reseptorit:
M-reseptori asetyylikoliinin lisäksi se vuorovaikuttaa selektiivisesti muskariinin (kärpäshelttatoksiinin) kanssa. M - kolinergiset reseptorit ovat läsnä sileissä lihaksissa ja vuorovaikutuksessa asetyylikoliinin kanssa aiheuttavat niiden supistumisen;
H-reseptori sitoutuvat erityisesti nikotiiniin. N-kolinergisiä reseptoreita löytyy poikkijuovaisten luustolihasten synapseista.
Spesifinen estäjä M-kolinergiset reseptorit on atropiini. Sitä löytyy belladonna- ja henbane-kasveista.
 Atropiinissa on rakenteeltaan samanlaisia funktionaalisia ryhmiä kuin asetyylikoliinilla ja niiden tilajärjestelyllä, joten se on M-kolinergisten reseptorien kilpaileva estäjä. Koska asetyylikoliinin sitoutuminen M-kolinergisiin reseptoreihin aiheuttaa sileiden lihasten supistumista, atropiinia käytetään lääkkeenä, joka lievittää niiden kouristuksia. (spasmodinen). Siten on tunnettua käyttää atropiinia silmälihasten rentouttamiseen silmänpohjaa tarkasteltaessa sekä kouristuksen lievittämiseen maha-suolikanavan koliikkien aikana. M-kolinergisiä reseptoreita on myös keskushermostossa (CNS), joten suuret annokset atropiinia voivat aiheuttaa keskushermoston ei-toivotun reaktion: motorista ja henkistä kiihtyneisyyttä, hallusinaatioita, kouristuksia.
Atropiinissa on rakenteeltaan samanlaisia funktionaalisia ryhmiä kuin asetyylikoliinilla ja niiden tilajärjestelyllä, joten se on M-kolinergisten reseptorien kilpaileva estäjä. Koska asetyylikoliinin sitoutuminen M-kolinergisiin reseptoreihin aiheuttaa sileiden lihasten supistumista, atropiinia käytetään lääkkeenä, joka lievittää niiden kouristuksia. (spasmodinen). Siten on tunnettua käyttää atropiinia silmälihasten rentouttamiseen silmänpohjaa tarkasteltaessa sekä kouristuksen lievittämiseen maha-suolikanavan koliikkien aikana. M-kolinergisiä reseptoreita on myös keskushermostossa (CNS), joten suuret annokset atropiinia voivat aiheuttaa keskushermoston ei-toivotun reaktion: motorista ja henkistä kiihtyneisyyttä, hallusinaatioita, kouristuksia.
Ditiliini on H-kolinergisten reseptorien kilpaileva agonisti, joka estää neuromuskulaaristen synapsien toimintaa.
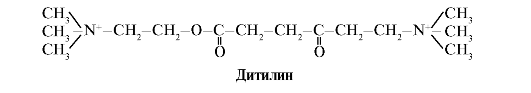 Luustolihasten neuromuskulaariset synapsit sisältävät H-kolinergisiä reseptoreita. Niiden vuorovaikutus asetyylikoliinin kanssa johtaa lihasten supistuksiin. Joissakin kirurgisissa leikkauksissa sekä endoskooppisissa tutkimuksissa käytetään lääkkeitä, jotka aiheuttavat luurankolihasten rentoutumista (lihasrelaksantit). Näitä ovat ditiliini, joka on asetyylikoliinin rakenteellinen analogi. Se kiinnittyy H-kolinergisiin reseptoreihin, mutta toisin kuin asetyylikoliini, tuhoaa sen hyvin hitaasti. Ionikanavien pitkittyneen avautumisen ja kalvon jatkuvan depolarisaation seurauksena hermoimpulssien johtuminen häiriintyy ja lihasten rentoutuminen tapahtuu. Aluksi nämä ominaisuudet löydettiin curare-myrkystä, minkä vuoksi tällaisia lääkkeitä kutsutaan curare kaltainen.
Luustolihasten neuromuskulaariset synapsit sisältävät H-kolinergisiä reseptoreita. Niiden vuorovaikutus asetyylikoliinin kanssa johtaa lihasten supistuksiin. Joissakin kirurgisissa leikkauksissa sekä endoskooppisissa tutkimuksissa käytetään lääkkeitä, jotka aiheuttavat luurankolihasten rentoutumista (lihasrelaksantit). Näitä ovat ditiliini, joka on asetyylikoliinin rakenteellinen analogi. Se kiinnittyy H-kolinergisiin reseptoreihin, mutta toisin kuin asetyylikoliini, tuhoaa sen hyvin hitaasti. Ionikanavien pitkittyneen avautumisen ja kalvon jatkuvan depolarisaation seurauksena hermoimpulssien johtuminen häiriintyy ja lihasten rentoutuminen tapahtuu. Aluksi nämä ominaisuudet löydettiin curare-myrkystä, minkä vuoksi tällaisia lääkkeitä kutsutaan curare kaltainen.
AIHE 1.3. PROTEIINIEN DENATUROINTI JA NIIDEN SPONTAANI UUDISTUMISEN MAHDOLLISUUS
1. Koska proteiinien natiivi konformaatio säilyy heikkojen vuorovaikutusten vuoksi, proteiinia ympäröivän ympäristön koostumuksen ja ominaisuuksien muutokset, altistuminen kemiallisille reagensseille ja fysikaalisille tekijöille aiheuttavat muutoksen niiden konformaatiossa (konformationaalisuuden ominaisuus). Suuren määrän sidosten katkeaminen johtaa proteiinien luonnollisen konformaation tuhoutumiseen ja denaturoitumiseen.
Proteiinien denaturaatio- tämä on niiden luonnollisen konformaation tuhoaminen denaturoivien aineiden vaikutuksesta, mikä johtuu heikkojen sidosten katkeamisesta, jotka stabiloivat proteiinin avaruudellista rakennetta. Denaturoitumiseen liittyy proteiinin ainutlaatuisen kolmiulotteisen rakenteen ja aktiivisen keskuksen tuhoutuminen ja sen biologisen aktiivisuuden menetys (kuva 1.16).
Kaikki yhden proteiinin denaturoidut molekyylit saavat satunnaisen konformaation, joka eroaa saman proteiinin muista molekyyleistä. Aktiivisen keskuksen muodostavat aminohapporadikaalit osoittautuvat avaruudellisesti etäämmiksi toisistaan, ts. proteiinin spesifinen sitoutumiskohta ligandin kanssa tuhoutuu. Denaturaation aikana proteiinien primäärirakenne pysyy muuttumattomana.
Denaturointiaineiden käyttö biologisessa tutkimuksessa ja lääketieteessä. Biokemiallisissa tutkimuksissa proteiinit poistetaan yleensä ensin liuoksesta ennen alhaisen molekyylipainon yhdisteiden määrittämistä biologisesta materiaalista. Trikloorietikkahappoa (TCA) käytetään useimmiten tähän tarkoitukseen. Kun TCA on lisätty liuokseen, denaturoidut proteiinit saostuvat ja ne poistetaan helposti suodattamalla (taulukko 1.1.)
Lääketieteessä denaturoivia aineita käytetään usein lääketieteellisten instrumenttien ja materiaalien sterilointiin autoklaaveissa (denaturoiva aine on korkea lämpötila) ja antiseptisinä aineina (alkoholi, fenoli, kloramiini) patogeenista mikroflooraa sisältävien saastuneiden pintojen käsittelyyn.
2. Spontaani proteiinin uudelleenaktivoituminen- todiste proteiinien primäärirakenteen, konformaation ja toiminnan determinismistä. Yksittäiset proteiinit ovat yhden geenin tuotteita, joilla on identtinen aminohapposekvenssi ja jotka saavat saman konformaation solussa. Peruspäätelmä, että proteiinin primaarirakenne sisältää jo tietoa sen konformaatiosta ja toiminnasta, tehtiin joidenkin proteiinien (erityisesti ribonukleaasin ja myoglobiinin) kyvyn perusteella spontaani renativoitua - palauttaa natiivi konformaatio denaturoinnin jälkeen.
Avaruusproteiinirakenteiden muodostuminen tapahtuu itsekokoamismenetelmällä - spontaanilla prosessilla, jossa polypeptidiketjulla, jolla on ainutlaatuinen primäärirakenne, on taipumus omaksua konformaatio, jossa on alhaisin vapaaenergia liuoksessa. Kyky renativoida proteiineja, jotka säilyttävät primäärirakenteensa denaturoinnin jälkeen, kuvattiin kokeessa ribonukleaasientsyymillä.
Ribonukleaasi on entsyymi, joka hajottaa yksittäisten nukleotidien välisiä sidoksia RNA-molekyylissä. Tällä pallomaisella proteiinilla on yksi polypeptidiketju, jonka tertiääristä rakennetta stabiloivat monet heikot ja neljä disulfidisidosta.
Ribonukleaasin käsittely urealla, joka katkaisee vetysidoksia molekyylissä, ja pelkistimellä, joka katkaisee disulfidisidoksia, johtaa entsyymin denaturoitumiseen ja sen aktiivisuuden menettämiseen.
Denaturoivien aineiden poistaminen dialyysillä johtaa proteiinin konformaation ja toiminnan palautumiseen, ts. uudestisyntymiseen. (Kuva 1.17).
 Riisi. 1.17. Ribonukleaasin denaturaatio ja renaturaatio
Riisi. 1.17. Ribonukleaasin denaturaatio ja renaturaatio
A - ribonukleaasin natiivi konformaatio, jonka tertiäärisessä rakenteessa on neljä disulfidisidosta; B - denaturoitu ribonukleaasimolekyyli;
B - uudelleenaktivoitu ribonukleaasimolekyyli, jolla on palautettu rakenne ja toiminta
1. Täytä taulukko 1.2.
Taulukko 1.2. Aminohappojen luokitus radikaalien polariteetin mukaan
2. Kirjoita tetrapeptidin kaava:
Asp - Pro - Fen - Liz
a) korostaa peptidin toistuvia ryhmiä, jotka muodostavat peptidirungon, ja aminohapporadikaalien edustamat muuttuvat ryhmät;
b) leimaa N- ja C-päät;
c) korostaa peptidisidoksia;
d) kirjoittaa toinen peptidi, joka koostuu samoista aminohapoista;
e) laskea tetrapeptidin mahdollisten varianttien lukumäärä, joilla on samanlainen aminohappokoostumus.
3. Selitä proteiinien primäärirakenteen rooli nisäkkään neurohypofyysin kahden rakenteellisesti samanlaisen ja evoluutionaalisesti läheisen peptidihormonin - oksitosiinin ja vasopressiinin - vertailevan analyysin esimerkillä (taulukko 1.3).
Taulukko 1.3. Oksitosiinin ja vasopressiinin rakenne ja toiminnot
 Tätä varten:
Tätä varten:
a) vertaa kahden peptidin koostumusta ja aminohapposekvenssiä;
b) löytää näiden kahden peptidin primäärirakenteen samankaltaisuus ja niiden biologisen vaikutuksen samankaltaisuus;
c) löytää eroja kahden peptidin rakenteessa ja eroja niiden toiminnassa;
d) tehdä johtopäätös peptidien primäärirakenteen vaikutuksesta niiden toimintoihin.
4. Kuvaa globulaaristen proteiinien konformaation muodostumisen päävaiheet (sekundaariset, tertiaariset rakenteet, supersekundaarisen rakenteen käsite). Ilmoita proteiinirakenteiden muodostumiseen osallistuvien sidostyypit. Mitkä aminohapporadikaalit voivat osallistua hydrofobisten vuorovaikutusten, ionisten, vetysidosten muodostumiseen.
Antaa esimerkkejä.
5. Määrittele käsite "proteiinien konformationaalinen labilisuus", osoita sen olemassaolon syyt ja merkitys.
6. Laajenna seuraavan lauseen merkitystä: "Proteiinien toiminta perustuu niiden spesifiseen vuorovaikutukseen ligandin kanssa" käyttämällä termejä ja selittämällä niiden merkitys: proteiinin konformaatio, aktiivinen keskus, ligandi, komplementaarisuus, proteiinin toiminta.
7. Selitä yhden esimerkin avulla, mitä domeenit ovat ja mikä niiden rooli on proteiinien toiminnassa.
ITSEVALTOTEHTÄVÄT
1. Ottelu.
Funktionaalinen ryhmä aminohapporadikaalissa:
A. Karboksyyliryhmä B. Hydroksyyliryhmä C Guanidiiniryhmä D. Tioliryhmä E. Aminoryhmä
2. Valitse oikeat vastaukset.
Aminohapot, joissa on polaarisia varautumattomia radikaaleja, ovat:
A. Cis B. Asn
B. Glu G. Kolme
3. Valitse oikeat vastaukset.
Aminohapporadikaalit:
A. Tarjoa primaarirakenteen spesifisyys B. Osallistu tertiaarisen rakenteen muodostukseen
B. Proteiinin pinnalla sijaitsevat ne vaikuttavat sen liukoisuuteen D. Muodostavat aktiivisen keskuksen
D. Osallistu peptidisidosten muodostukseen
4. Valitse oikeat vastaukset.
Hydrofobisia vuorovaikutuksia voi muodostua aminohapporadikaalien välillä:
A. Tre Lay B. Pro Three
B. Met Ile G. Tir Ala D. Val Fen
5. Valitse oikeat vastaukset.
Ionisidoksia voi muodostua aminohapporadikaalien välille:
A. Gln Asp B. Apr Liz
B. Liz Glu G. Gis Asp D. Asn Apr
6. Valitse oikeat vastaukset.
Aminohapporadikaalien välille voi muodostua vetysidoksia:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Ottelu.
Proteiinirakenteen muodostumiseen osallistuva sidostyyppi:
A. Ensisijainen rakenne B. Toissijainen rakenne
B. Tertiäärinen rakenne
D. Ylisekundaarinen rakenne E. Konformaatio.
1. Peptidirungon atomien väliset vetysidokset
2. Heikot sidokset aminohapporadikaalien funktionaalisten ryhmien välillä
3. Sidot aminohappojen α-amino- ja α-karboksyyliryhmien välillä
8. Valitse oikeat vastaukset. Trypsiini:
A. Proteolyyttinen entsyymi B. Sisältää kaksi domeenia
B. Hydrolysoi tärkkelystä
D. Aktiivinen paikka sijaitsee verkkotunnusten välissä. D. Koostuu kahdesta polypeptidiketjusta.
9. Valitse oikeat vastaukset. Atropiini:
A. Neurotransmitteri
B. Asetyylikoliinin rakenteellinen analogi
B. Vuorovaikuttaa H-kolinergisten reseptorien kanssa
D. Vahvistaa hermoimpulssien johtumista kolinergisten synapsien kautta
D. M-kolinergisten reseptorien kilpaileva estäjä
10. Valitse oikeat väitteet. Proteiinissa:
V. Ensisijainen rakenne sisältää tietoa aktiivisen sivustonsa rakenteesta
B. Aktiivinen keskus muodostuu primäärirakenteen tasolle
B. Konformaatio on jäykästi kiinnitetty kovalenttisilla sidoksilla
D. Aktiivinen kohta voi olla vuorovaikutuksessa samanlaisten ligandien ryhmän kanssa
johtuu proteiinien D konformationaalisesta labiiteetista. Muutokset ympäristössä voivat vaikuttaa aktiivisen proteiinin affiniteettiin
keskustasta ligandiin
1. 1-B, 2-G, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
PERUSKÄSITTEET JA KÄSITTEET
1. Proteiini, polypeptidi, aminohapot
2. Primaariset, sekundaariset, tertiääriset proteiinirakenteet
3. Konformaatio, natiivi proteiinin konformaatio
4. Proteiinin kovalenttiset ja heikot sidokset
5. Konformationaalinen labilisuus
6. Proteiinin aktiivinen kohta
7. Ligandit
8. Proteiinin taittaminen
9. Ligandien rakenteelliset analogit
10. Domain-proteiinit
11. Yksinkertaiset ja monimutkaiset proteiinit
12. Proteiinien denaturointi, denaturointiaineet
13. Proteiinien uudelleenaktivointi
Ratkaista ongelmia
"Proteiinien rakenneorganisaatio ja niiden toiminnan perusta"
1. Proteiinin - hemoglobiini A (HbA) - päätehtävä on hapen kuljettaminen kudoksiin. Ihmispopulaatiossa tunnetaan tämän proteiinin useita muotoja, joiden ominaisuudet ja toiminta ovat muuttuneet – niin sanotut epänormaalit hemoglobiinit. Esimerkiksi hemoglobiini S:llä, jota löytyy sirppisolusairautta (HbS) sairastavien potilaiden punasoluista, on havaittu olevan alhainen liukoisuus olosuhteissa, joissa hapen osapaine on alhainen (kuten laskimoveren tapauksessa). Tämä johtaa tämän proteiinin aggregaattien muodostumiseen. Proteiini menettää toimintansa, saostuu ja punasolut muuttuvat epäsäännöllisiksi (jotkut niistä muodostavat sirpin muodon) ja tuhoutuvat tavallista nopeammin pernassa. Seurauksena kehittyy sirppisoluanemia.
Ainoa ero HbA:n primäärirakenteessa havaittiin hemoglobiinin p-ketjun N-pään alueella. Vertaa β-juosteen N-terminaalisia alueita ja näytä kuinka muutokset proteiinin primäärirakenteessa vaikuttavat sen ominaisuuksiin ja toimintoihin.
 Tätä varten:
Tätä varten:
a) Kirjoita aminohappojen kaavat, joilla HbA eroaa, ja vertaa näiden aminohappojen ominaisuuksia (polariteetti, varaus).
b) tehdä johtopäätös siitä, miksi liukoisuus heikkenee ja hapen kuljetus kudoksiin häiriintyy.
2. Kuvassa on kaavio proteiinin rakenteesta, jossa on sitoutumiskeskus ligandin kanssa (aktiivinen keskus). Selitä, miksi proteiini on valikoiva ligandin valinnassa. Tätä varten:
a) muista mikä on proteiinin aktiivinen keskus ja tarkastele kuvassa näkyvän proteiinin aktiivisen keskuksen rakennetta;
b) kirjoittaa aktiivisen keskuksen muodostavien aminohapporadikaalien kaavat;
c) piirtää ligandi, joka voisi spesifisesti olla vuorovaikutuksessa proteiinin aktiivisen kohdan kanssa. Merkitse siihen funktionaaliset ryhmät, jotka voivat muodostaa sidoksia aminohapporadikaalien kanssa, jotka muodostavat aktiivisen keskuksen;
d) osoittavat sidostyypit, joita syntyy aktiivisen keskuksen ligandin ja aminohapporadikaalien välillä;
e) selittää, mihin proteiini-ligandi-vuorovaikutuksen spesifisyys perustuu.
 3.
Kuvassa näkyy proteiinin aktiivinen kohta ja useita ligandeja.
3.
Kuvassa näkyy proteiinin aktiivinen kohta ja useita ligandeja.
Määritä, mikä ligandi on todennäköisimmin vuorovaikutuksessa proteiinin aktiivisen kohdan kanssa ja miksi.
 Millaisia sidoksia syntyy proteiini-ligandikompleksin muodostumisen aikana?
Millaisia sidoksia syntyy proteiini-ligandikompleksin muodostumisen aikana?
4. Luonnollisten proteiiniligandien rakenteellisia analogeja voidaan käyttää lääkkeinä proteiinien aktiivisuuden modifioimiseksi.
Asetyylikoliini on hermo-muskulaaristen synapsien viritysvälityksen välittäjä. Kun asetyylikoliini on vuorovaikutuksessa proteiinien kanssa - luurankolihasten postsynaptisen kalvon reseptorit, ionikanavat avautuvat ja lihasten supistuminen tapahtuu. Ditiliin on lääke, jota käytetään joissakin leikkauksissa lihasten rentouttamiseen, koska se häiritsee hermoimpulssien siirtymistä hermo-lihassynapsien kautta. Selitä ditiilin vaikutusmekanismi lihasrelaksanttina. Tätä varten:
a) kirjoittaa asetyylikoliinin ja ditiliinin kaavat ja vertailla niiden rakenteita;
b) kuvaile ditiilin rentouttavan vaikutuksen mekanismia.
5. Joissakin sairauksissa potilaan ruumiinlämpö nousee, mitä pidetään kehon suojaavana reaktiona. Korkeat lämpötilat ovat kuitenkin haitallisia kehon proteiineille. Selitä, miksi yli 40 °C:n lämpötiloissa proteiinien toiminta häiriintyy ja ihmishenkiä uhkaa. Muista tehdäksesi tämän:
1) Proteiinien rakenne ja sidokset, jotka pitävät sen rakenteen alkuperäisessä konformaatiossa;
2) Miten proteiinien rakenne ja toiminta muuttuvat lämpötilan noustessa?;
3) Mikä on homeostaasi ja miksi se on tärkeää ihmisten terveyden ylläpitämiselle.
Moduuliyksikkö 2 OLIGOMERICK PROTEINIT SÄÄNTELYVAIKUTTEIDEN KOHTEENA. PROTEIINIEN RAKENNE JA TOIMINNALLINEN MONIMUOTOisuus. PROTEIINIEN EROTUS- JA PUHDISTUSMENETELMÄT
Oppimistavoitteet Pystyy:
1. Käytä tietoa oligomeeristen proteiinien rakenteen ja toiminnan ominaisuuksista ymmärtääksesi niiden toimintojen säätelyn adaptiivisia mekanismeja.
2. Selitä kaperonien rooli proteiinien konformaation synteesissä ja ylläpitämisessä soluolosuhteissa.
3. Selitä elämän ilmenemismuotojen monimuotoisuus kehossa syntetisoitujen proteiinien rakenteiden ja toimintojen moninaisuuden avulla.
4. Analysoi proteiinien rakenteen ja niiden toiminnan välistä suhdetta käyttämällä esimerkkejä vastaavien hemoproteiinien - myoglobiinin ja hemoglobiinin - vertailusta sekä immunoglobuliiniperheen viiden proteiiniluokan edustajista.
5. Soveltaa tietoa proteiinien fysikaalis-kemiallisten ominaisuuksien erityispiirteistä valittaessa menetelmiä niiden puhdistamiseksi muista proteiineista ja epäpuhtauksista.
6. Tulkitse veriplasman proteiinien kvantitatiivisen ja laadullisen koostumuksen tuloksia kliinisen diagnoosin vahvistamiseksi tai selkeyttämiseksi.
Tietää:
1. Oligomeeristen proteiinien rakenteen ominaisuudet ja adaptiiviset mekanismit niiden toimintojen säätelyyn hemoglobiinin esimerkin avulla.
2. Kaperonien rakenne ja toiminnot sekä niiden merkitys proteiinien natiivin konformaation ylläpitämisessä soluolosuhteissa.
3. Periaatteet proteiinien yhdistämisestä perheiksi niiden konformaation ja toimintojen samankaltaisuuden perusteella käyttäen esimerkkiä immunoglobuliinista.
4. Menetelmät proteiinien erottamiseksi niiden fysikaalis-kemiallisten ominaisuuksien ominaisuuksien perusteella.
5. Veriplasman elektroforeesi menetelmänä proteiinien laadullisen ja kvantitatiivisen koostumuksen arvioimiseksi.
AIHE 1.4. OLIGOMEERISTEN PROTEIINIEN RAKENTEEN JA TOIMINNAN OMINAISUUDET KÄYTTÄEN HEMOGLOBIIIN ESIMERKKIÄ
1. Monet proteiinit sisältävät useita polypeptidiketjuja. Tällaisia proteiineja kutsutaan oligomeerinen, ja yksittäiset ketjut - protomeerit. Oligomeeristen proteiinien protomeerit on yhdistetty monilla heikoilla ei-kovalenttisilla sidoksilla (hydrofobinen, ioninen, vety). Vuorovaikutus
protomeerit suoritetaan ansiosta täydentävyyttä niiden koskettavat pinnat.
Oligomeeristen proteiinien protomeerien määrä voi vaihdella suuresti: hemoglobiini sisältää 4 protomeeriä, aspa12 protomeeriä ja tupakan mosaiikkiviruksen proteiini sisältää 2120 ei-kovalenttisilla sidoksilla yhdistettyä protomeeriä. Tästä johtuen oligomeeristen proteiinien molekyylipainot voivat olla erittäin korkeat.
Yhden protomeerin vuorovaikutusta muiden kanssa voidaan pitää proteiini-ligandivuorovaikutuksen erikoistapauksena, koska jokainen protomeeri toimii ligandina muille protomeereille. Proteiinissa olevien protomeerien lukumäärää ja yhdistämismenetelmää kutsutaan kvaternäärinen proteiinirakenne.
Proteiinit voivat sisältää protomeerejä, joilla on sama tai erilainen rakenne, esimerkiksi homodimeerit ovat proteiineja, jotka sisältävät kaksi identtistä protomeeriä, ja heterodimeerit ovat proteiineja, jotka sisältävät kaksi eri protomeeriä.
Jos proteiinit sisältävät erilaisia protomeerejä, niihin voi muodostua sitoutumiskeskuksia erilaisilla ligandeilla, jotka ovat rakenteeltaan erilaisia. Kun ligandi sitoutuu aktiiviseen kohtaan, tämän proteiinin toiminta ilmenee. Eri protomeerissä sijaitsevaa keskustaa kutsutaan allosteeriseksi (eri kuin aktiivinen). Yhteydenotto allosteerinen ligandi tai efektori, se suorittaa säätelytoiminnon (kuva 1.18). Allosteerisen keskuksen vuorovaikutus efektorin kanssa aiheuttaa konformaatiomuutoksia koko oligomeerisen proteiinin rakenteessa sen konformationaalisuuden vuoksi. Tämä vaikuttaa aktiivisen kohdan affiniteettiin spesifiseen ligandiin ja säätelee kyseisen proteiinin toimintaa. Muutosta kaikkien protomeerien konformaatiossa ja toiminnassa oligomeerisen proteiinin vuorovaikutuksessa vähintään yhden ligandin kanssa kutsutaan yhteistoiminnallisiksi konformaatiomuutoksiksi. Proteiinin toimintaa tehostavia efektejä kutsutaan aktivaattorit, ja sen toimintaa estävät efektorit ovat estäjät.
Siten oligomeerisillä proteiineilla, samoin kuin proteiineilla, joilla on domeenirakenne, on uusi ominaisuus verrattuna monomeerisiin proteiineihin - kyky allosteerisesti säädellä toimintoja (säätely kiinnittämällä proteiiniin erilaisia ligandeja). Tämä voidaan nähdä vertaamalla kahden läheisesti toisiinsa liittyvän monimutkaisen proteiinin, myoglobiinin ja hemoglobiinin, rakenteita ja toimintoja.
 Riisi. 1.18. Kaavio dimeerisen proteiinin rakenteesta
Riisi. 1.18. Kaavio dimeerisen proteiinin rakenteesta
2. Avaruusrakenteiden muodostuminen ja myoglobiinin toiminta.
Myoglobiini (Mb) on punaisissa lihaksissa esiintyvä proteiini, jonka päätehtävänä on luoda intensiiviseen lihastyöhön tarvittavia O 2 -varastoja. Mb on monimutkainen proteiini, joka sisältää proteiiniosan - apoMb:n ja ei-proteiiniosan - hemin. ApoMB:n primäärirakenne määrää sen kompaktin pallomaisen konformaation ja aktiivisen keskuksen rakenteen, johon myoglobiinin ei-proteiiniosa, hemi, on kiinnittynyt. Verestä lihaksiin tuleva happi sitoutuu myoglobiinin Fe+ 2 -hemeihin. Mb on monomeerinen proteiini, jolla on erittäin korkea affiniteetti O 2:een, joten myoglobiinin hapen vapautuminen tapahtuu vain intensiivisen lihastyön aikana, kun O 2:n osapaine laskee jyrkästi.
Mv-konformaation muodostuminen. Punaisissa lihaksissa, ribosomeissa translaation aikana, syntetisoidaan MB:n primäärirakenne, jota edustaa spesifinen 153 aminohappotähteen sekvenssi. Mb:n toissijainen rakenne sisältää kahdeksan α-heliksiä, joita kutsutaan latinalaisilla kirjaimilla A:sta H:hen, joiden välissä on ei-kierteisiä alueita. Mb:n tertiäärinen rakenne on muodoltaan kompakti palloma, jonka syvennyksessä aktiivinen keskus sijaitsee F- ja E α-heliksien välissä (kuva 1.19).
 Riisi. 1.19. Myoglobiinin rakenne
Riisi. 1.19. Myoglobiinin rakenne
3. MV:n aktiivisen keskuksen rakenteen ja toiminnan piirteet. Mb:n aktiivinen keskus muodostuu pääasiassa hydrofobisista aminohapporadikaaleista, jotka ovat kaukana toisistaan primäärirakenteessa (esim. 3 9 ja Fen 138) Heikosti veteen liukeneva ligandit - hemi ja O 2 - kiinnittyvät aktiiviseen keskustaan. Hemi on apoMB:n spesifinen ligandi (kuva 1.20), jonka perusta koostuu neljästä pyrrolirenkaasta, jotka on yhdistetty metenyylisilloilla; keskellä on Fe+ 2 -atomi, joka on yhdistetty pyrrolirenkaiden typpiatomeihin neljällä koordinaatiosidoksella. Mb:n aktiivisessa keskustassa on hydrofobisten aminohapporadikaalien lisäksi myös kahden aminohapon tähteitä, joissa on hydrofiilisiä radikaaleja - Gis E 7(Gis 64) ja GIS F 8(Hänen 93) (Kuva 1.21).
 Riisi. 1.20. Hemin rakenne - myoglobiinin ja hemoglobiinin ei-proteiiniosa
Riisi. 1.20. Hemin rakenne - myoglobiinin ja hemoglobiinin ei-proteiiniosa
 Riisi. 1.21. Hemin ja O2:n sijainti apomyoglobiinin ja hemoglobiinin protomeerien aktiivisessa kohdassa
Riisi. 1.21. Hemin ja O2:n sijainti apomyoglobiinin ja hemoglobiinin protomeerien aktiivisessa kohdassa
Hemi on kovalenttisesti sidottu Hänen F8:aan rautaatomin kautta. O 2 kiinnittyy rautaan hemitason toisella puolella. Hänen E 7 on välttämätön O 2:n oikealle suuntaukselle ja helpottaa hapen lisäämistä Fe + 2 -heemiin
GIS F 8 muodostaa koordinaatiosidoksen Fe+ 2:n kanssa ja kiinnittää hemin tiukasti aktiiviseen keskustaan. Gis E 7 tarvitaan oikean suuntautumiseen toisen ligandin - O 2:n aktiivisessa keskustassa sen vuorovaikutuksessa Fe + 2 -hemin kanssa. Hemin mikroympäristö luo olosuhteet O 2:n vahvalle mutta palautuvalle sitoutumiselle Fe + 2:een ja estää vettä pääsemästä hydrofobiseen aktiiviseen kohtaan, mikä voi johtaa sen hapettumiseen Fe + 3:ksi.
Mb:n ja sen aktiivisen keskuksen monomeerinen rakenne määrää proteiinin korkean affiniteetin 02:lle.
4. Hb:n oligomeerinen rakenne ja Hb:n affiniteetin säätely 02-ligandeihin. Ihmisen hemoglobiinit- proteiiniperhe, kuten myoglobiini, joka liittyy monimutkaisiin proteiineihin (hemoproteiineihin). Niillä on tetrameerinen rakenne ja ne sisältävät kaksi a-ketjua, mutta eroavat kahden muun polypeptidiketjun (2a-, 2x-ketjut) rakenteesta. Toisen polypeptidiketjun rakenne määrittää näiden Hb-muotojen toiminnan piirteet. Noin 98 % aikuisen punasolujen hemoglobiinista on hemoglobiini A(2a-, 2p-ketjut).
Sikiön kehityksen aikana hemoglobiinien kaksi päätyyppiä toimivat: alkion Hb(2α, 2ε), joka löytyy sikiön kehityksen alkuvaiheista, ja hemoglobiini F (sikiö)- (2α, 2γ), joka korvaa varhaisen sikiön hemoglobiinin kohdunsisäisen kehityksen kuudentena kuukautena ja vasta syntymän jälkeen korvataan Hb A:lla.
HB A on myoglobiiniin (MB) liittyvä proteiini, jota löytyy aikuisen ihmisen punasoluista. Sen yksittäisten protomeerien rakenne on samanlainen kuin myoglobiinin. Myoglobiinin ja hemoglobiinin protomeerien sekundääriset ja tertiaariset rakenteet ovat hyvin samankaltaisia huolimatta siitä, että niiden polypeptidiketjujen primäärirakenteessa vain 24 aminohappotähdettä on identtisiä (hemoglobiinin protomeerien sekundaarirakenne, kuten myoglobiini, sisältää kahdeksan α-heliksiä, merkitty latinalaisilla kirjaimilla A:sta H:hen, ja tertiäärinen rakenne on tiiviin globulin muotoinen). Mutta toisin kuin myoglobiinilla, hemoglobiinilla on oligomeerinen rakenne, joka koostuu neljästä polypeptidiketjusta, jotka on yhdistetty ei-kovalenttisilla sidoksilla (kuva 1.22).
Jokainen Hb-protomeeri liittyy ei-proteiiniosaan - hemiin ja viereisiin protomeereihin. Hb:n proteiiniosan yhteys hemiin on samanlainen kuin myoglobiinin: proteiinin aktiivisessa keskustassa hemin hydrofobisia osia ympäröivät hydrofobiset aminohapporadikaalit paitsi His F 8 ja His E 7, jotka sijaitsevat molemmilla puolilla hemitasoa ja niillä on samanlainen rooli proteiinin toiminnassa ja sen sitoutumisessa happeen (katso myoglobiinin rakenne).
 Riisi. 1.22. Hemoglobiinin oligomeerinen rakenne
Riisi. 1.22. Hemoglobiinin oligomeerinen rakenne
Sitä paitsi, Gis E 7 suorittaa tärkeän lisärooli Nv:n toiminnassa. Vapaalla hemellä on 25 000 kertaa suurempi affiniteetti CO:lle kuin O2:lle. CO:ta muodostuu elimistössä pieniä määriä, ja sen korkean affiniteetin hemiin vuoksi se voi häiritä solujen elämälle välttämättömän O 2:n kuljetusta. Hemoglobiinin koostumuksessa hemin affiniteetti hiilimonoksidiin ylittää kuitenkin affiniteetin O 2:lle vain 200 kertaa, koska aktiivisessa keskustassa on His E 7. Loppuosa tästä aminohaposta luo optimaaliset olosuhteet hemin sitoutumiselle O 2:een ja heikentää hemin vuorovaikutusta CO:n kanssa.
5. HB:n päätehtävä on O2:n kuljettaminen keuhkoista kudoksiin. Toisin kuin monomeerinen myoglobiini, jolla on erittäin korkea affiniteetti O2:een ja joka varastoi happea punaisiin lihaksiin, hemoglobiinin oligomeerinen rakenne tarjoaa:
1) HB:n nopea kyllästyminen hapella keuhkoissa;
2) HB:n kyky vapauttaa happea kudoksissa suhteellisen korkeassa 02:n osapaineessa (20-40 mm Hg);
3) mahdollisuus säätää Hb:n affiniteettia O 2:een.
6. Yhteistoiminnalliset muutokset hemoglobiinin protomeerien konformaatiossa nopeuttavat O 2:n sitoutumista keuhkoissa ja sen vapautumista kudoksiin. Keuhkoissa 02:n korkea osapaine edistää sen sitoutumista Hb:hen neljän protomeerin (2α ja 2β) aktiivisessa kohdassa. Kunkin protomeerin aktiivinen keskus, kuten myoglobiinissa, sijaitsee kahden α-kierteen (F ja E) välissä hydrofobisessa taskussa. Se sisältää ei-proteiiniosan - hemin, joka on kiinnittynyt proteiiniosaan monilla heikkoilla hydrofobisilla vuorovaikutuksilla ja yhden vahvan sidoksen Fe 2 + -hemin ja His F 8:n välillä (katso kuva 1.21).
Deoksihemoglobiinissa, johtuen tästä sidoksesta His F 8:n kanssa, Fe 2+ -atomi työntyy esiin hemitasosta kohti histidiiniä. O2:n sitoutuminen Fe2+:aan tapahtuu hemin toisella puolella His E7 -alueella käyttämällä yhtä vapaata koordinaatiosidosta. His E 7 tarjoaa optimaaliset olosuhteet O 2:n sitoutumiselle hemiraudaan.
O 2:n lisääminen yhden protomeerin Fe + 2 -atomiin aiheuttaa sen siirtymisen hemitasoon, jota seuraa siihen liittyvä histidiinijäännös
 Riisi. 1.23. Muutos hemoglobiinin protomeerin konformaatiossa yhdistettynä O 2:een
Riisi. 1.23. Muutos hemoglobiinin protomeerin konformaatiossa yhdistettynä O 2:een
Tämä johtaa muutokseen kaikkien polypeptidiketjujen konformaatiossa niiden konformationaalisuuden vuoksi. Muiden ketjujen konformaation muuttaminen helpottaa niiden vuorovaikutusta myöhempien O2-molekyylien kanssa.
Neljäs O 2 -molekyyli kiinnittyy hemoglobiiniin 300 kertaa helpommin kuin ensimmäinen (kuva 1.24).
 Riisi. 1.24. Yhteistoiminnalliset muutokset hemoglobiinin protomeerien konformaatiossa sen vuorovaikutuksessa O2:n kanssa
Riisi. 1.24. Yhteistoiminnalliset muutokset hemoglobiinin protomeerien konformaatiossa sen vuorovaikutuksessa O2:n kanssa
Kudoksissa jokainen seuraava O 2 -molekyyli lohkeaa helpommin kuin edellinen, myös protomeerien konformaation yhteistoiminnallisten muutosten vuoksi.
7. Orgaanisten aineiden hajoamisen aikana muodostuneet CO 2 ja H+ vähentävät hemoglobiinin affiniteettia O 2:lle suhteessa pitoisuuteensa. Solujen toimintaan tarvittava energia tuotetaan pääasiassa mitokondrioissa orgaanisten aineiden hapettumisen aikana hemoglobiinin keuhkoista kuljettaman O 2:n avulla. Orgaanisten aineiden hapettumisen seurauksena muodostuu niiden hajoamisen lopputuotteita: CO 2 ja K 2 O, joiden määrä on verrannollinen käynnissä olevien hapetusprosessien intensiteettiin.
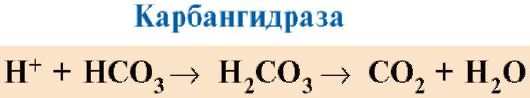
CO 2 diffundoituu soluista vereen ja tunkeutuu punasoluihin, joissa se muuttuu karbanhydraasientsyymin vaikutuksesta hiilihapoksi. Tämä heikko happo hajoaa protoniksi ja bikarbonaatti-ioniksi.
H+ pystyvät liittymään Hänen radikaaleihinsa 14 6 hemoglobiinin a- ja p-ketjuissa, ts. alueilla, jotka ovat kaukana hemistä. Hemoglobiinin protonoituminen vähentää sen affiniteettia O 2:een, edistää O 2:n poistumista oksiHb:stä, deoksiHb:n muodostumista ja lisää kudosten hapen saantia suhteessa muodostuneiden protonien määrään (kuva 1.25).
Vapautuneen hapen määrän lisääntymistä punasolujen H+-pitoisuuden kasvusta riippuen kutsutaan Bohrin efektiksi (nimetty tanskalaisen fysiologin Christian Bohrin mukaan, joka löysi tämän vaikutuksen ensimmäisenä).
Keuhkoissa korkea hapen osapaine edistää sen sitoutumista deoksiHb:hen, mikä vähentää proteiinin affiniteettia H+:aan. Hiilihapon vaikutuksesta vapautuneet protonit reagoivat bikarbonaattien kanssa muodostaen CO 2:ta ja H 2 O:ta
 Riisi. 1.25. Hb:n O 2 -affiniteetin riippuvuus CO 2:n ja protonien pitoisuudesta (Bohr-ilmiö):
Riisi. 1.25. Hb:n O 2 -affiniteetin riippuvuus CO 2:n ja protonien pitoisuudesta (Bohr-ilmiö):
A- CO 2- ja H+-konsentraation vaikutus 02:n vapautumiseen HB-kompleksista (Bohr-ilmiö); B- deoksihemoglobiinin hapetus keuhkoissa, hiilidioksidin muodostuminen ja vapautuminen.
Tuloksena oleva CO 2 tulee keuhkorakkuloihin ja poistuu uloshengitetyn ilman mukana. Siten hemoglobiinin vapauttaman hapen määrää kudoksissa säätelevät orgaanisten aineiden hajoamistuotteet: mitä voimakkaammin aineet hajoavat esimerkiksi fyysisen harjoituksen aikana, sitä korkeampi on CO 2 - ja H + -pitoisuus ja sitä enemmän happea. kudokset saavat Hb:n affiniteetin heikkenemisen seurauksena O 2:ta kohtaan.
8. Hb:n affiniteetin allosteerinen säätely 02:lle ligandilla - 2,3-bisfosfoglyseraatilla. Punasoluissa hemoglobiinin allosteerinen ligandi, 2,3-bisfosfoglyseraatti (2,3-BPG), syntetisoidaan glukoosin hapettumistuotteesta - 1,3-bisfosfoglyseraatista. Normaaleissa olosuhteissa 2,3-BPG:n pitoisuus on korkea ja verrattavissa Hb-pitoisuuteen. 2,3-BPG:llä on vahva negatiivinen varaus -5.
 Bisfosfoglyseraatti kudoskapillaareissa sitoutuen deoksihemoglobiiniin lisää hapen vapautumista kudoksiin, mikä vähentää Hb:n affiniteettia O 2:een.
Bisfosfoglyseraatti kudoskapillaareissa sitoutuen deoksihemoglobiiniin lisää hapen vapautumista kudoksiin, mikä vähentää Hb:n affiniteettia O 2:een.
Tetrameerisen hemoglobiinimolekyylin keskellä on ontelo. Se muodostuu kaikkien neljän protomeerin aminohappotähteistä (katso kuva 1.22). Kudoskapillaareissa Hb:n protonoituminen (Bohr-ilmiö) johtaa hemiraudan ja O2:n välisen sidoksen katkeamiseen. Molekyylissä
deoksihemoglobiini, verrattuna oksihemoglobiiniin, ilmaantuu ylimääräisiä ionisidoksia, jotka yhdistävät protomeerit, minkä seurauksena keskusontelon mitat kasvavat oksihemoglobiiniin verrattuna. Keskusontelo on paikka, jossa 2,3-BPG kiinnittyy hemoglobiiniin. Keskusontelon koon eroista johtuen 2,3-BPG voi kiinnittyä vain deoksihemoglobiiniin.
2,3-BPG on vuorovaikutuksessa hemoglobiinin kanssa paikassa, joka on kaukana proteiinin aktiivisista keskuksista ja kuuluu allosteerinen(säätely) ligandeja, ja Hb:n keskusontelo on allosteerinen keskus. 2,3-BPG:llä on voimakas negatiivinen varaus ja se on vuorovaikutuksessa Hb:n kahden β-ketjun viiden positiivisesti varautuneen ryhmän kanssa: Val:n N-terminaalisen α-aminoryhmän ja Lys 82 His 143 -radikaalin (kuva 1.26).
 Riisi. 1.26. BPG deoksihemoglobiinin keskusontelossa
Riisi. 1.26. BPG deoksihemoglobiinin keskusontelossa
BPG sitoutuu kolmeen positiivisesti varautuneeseen ryhmään kussakin β-juosteessa.
Kudoskapillaareissa muodostuva deoksihemoglobiini on vuorovaikutuksessa 2,3-BPG:n kanssa ja β-ketjujen positiivisesti varautuneiden radikaalien ja negatiivisesti varautuneen ligandin välille muodostuu ionisidoksia, jotka muuttavat proteiinin konformaatiota ja vähentävät Hb:n affiniteettia O2:een. . Hb:n affiniteetin lasku 02:lle edistää O 2:n tehokkaampaa vapautumista kudokseen.
Keuhkoissa korkeassa osapaineessa happi on vuorovaikutuksessa Hb:n kanssa liittyen hemiraudaan; tässä tapauksessa proteiinin konformaatio muuttuu, keskusontelo pienenee ja 2,3-BPG siirtyy allosteerisesta keskustasta
Siten oligomeerisillä proteiineilla on uusia ominaisuuksia verrattuna monomeerisiin proteiineihin. Ligandien kiinnittyminen paikkoihin
avaruudellisesti kaukana toisistaan (allosteerinen), voivat aiheuttaa konformaatiomuutoksia koko proteiinimolekyylissä. Vuorovaikutuksesta säätelyligandien kanssa tapahtuu muutos konformaatiossa ja proteiinimolekyylin toiminnan mukautuminen ympäristön muutoksiin.
AIHE 1.5. PROTEIINIEN NATIIVIN KONFORMAATION YLLÄPITO SOLU-OLLOISSA
Soluissa polypeptidiketjujen synteesin aikana niiden kulkeutuminen kalvojen läpi solun vastaaviin osiin, laskostumisprosessin aikana (natiivikonformaation muodostuminen) ja oligomeeristen proteiinien kokoamisen aikana sekä niiden toiminnan aikana, välituote , aggregaatiolle alttiita, epästabiileja konformaatioita syntyy proteiinirakenteessa. Hydrofobiset radikaalit, jotka ovat yleensä piilossa proteiinimolekyylin sisällä natiivissa konformaatiossa, ilmestyvät pinnalle epävakaassa konformaatiossa ja pyrkivät yhdistymään muiden proteiinien ryhmiin, jotka ovat huonosti veteen liukenevia. Kaikkien tunnettujen organismien soluista on löydetty erityisiä proteiineja, jotka varmistavat soluproteiinien optimaalisen laskostumisen, stabiloivat niiden natiivia konformaatiota toiminnan aikana ja mikä tärkeintä, ylläpitävät solunsisäisten proteiinien rakennetta ja toimintoja homeostaasin häiriintyessä. Näitä proteiineja kutsutaan "seuraajat" joka tarkoittaa ranskaksi lastenhoitajaa.
1. Molekyylikaperonit ja niiden rooli proteiinien denaturoitumisen estämisessä.
Chaperonit (CH) luokitellaan niiden alayksiköiden massan mukaan. Suuren molekyylipainon omaavien chaperonien massa on 60 - 110 kDa. Niistä kolmea luokkaa on eniten tutkittu: Sh-60, Sh-70 ja Sh-90. Jokainen luokka sisältää sukulaisia proteiineja. Siten Sh-70 sisältää proteiineja, joiden molekyylipaino on 66 - 78 kDa. Pienen molekyylipainon omaavien chaperonien molekyylipaino on 40 - 15 kDa.
Ohjaajien joukossa on konstitutiivinen proteiineja, joiden korkea perussynteesi ei riipu stressin vaikutuksista kehon soluihin, ja indusoituva, jonka synteesi normaaleissa olosuhteissa on heikkoa, mutta lisääntyy voimakkaasti rasituksessa. Indusoituvia chaperoneja kutsutaan myös "lämpösokkiproteiineiksi", koska ne löydettiin ensimmäisen kerran korkeille lämpötiloille altistetuista soluista. Soluissa proteiinien suuren pitoisuuden vuoksi osittain denaturoituneiden proteiinien spontaani uudelleenaktivointi on vaikeaa. Sh-70 voi estää denaturaation alkamista ja auttaa palauttamaan proteiinien alkuperäisen konformaation. Molekyyliset chaperonit-70- erittäin konservoitunut proteiiniluokka, jota löytyy solun kaikista osista: sytoplasmasta, tumasta, endoplasmisesta retikulumista, mitokondrioista. Yksittäisen polypeptidiketjun Ш-70 karboksyylipäässä on alue, joka on ura, joka pystyy olemaan vuorovaikutuksessa pitkien peptidien kanssa
7 - 9 aminohappotähdettä, jotka on rikastettu hydrofobisilla radikaaleilla. Tällaisia alueita pallomaisissa proteiineissa esiintyy suunnilleen joka 16. aminohappo. Sh-70 pystyy suojaamaan proteiineja lämpötilan inaktivoitumiselta ja palauttamaan osittain denaturoituneiden proteiinien konformaatiota ja aktiivisuutta.
2. Chaperonien rooli proteiinin laskostumisessa. Proteiinisynteesin aikana ribosomissa polypeptidin N-terminaalinen alue syntetisoituu ennen C-terminaalista aluetta. Luonnollisen konformaation muodostamiseksi tarvitaan proteiinin täydellinen aminohapposekvenssi. Proteiinisynteesiprosessissa kaperonit-70 pystyvät aktiivisen keskuksensa rakenteesta johtuen sulkemaan polypeptidin alueita, jotka ovat alttiita aggregaatiolle ja jotka ovat rikastuneet hydrofobisilla aminohapporadikaaleilla, kunnes synteesi on valmis (Kuva 1.27, A ).
 Riisi. 1.27. Chaperonien osallistuminen proteiinien laskostukseen
Riisi. 1.27. Chaperonien osallistuminen proteiinien laskostukseen
A - chaperons-70:n osallistuminen hydrofobisten vuorovaikutusten estämiseen syntetisoidun polypeptidin osien välillä; B - proteiinin luonnollisen konformaation muodostuminen chaperonikompleksissa
Monet korkeamolekyyliset proteiinit, joilla on monimutkainen konformaatio, kuten domeenirakenne, laskostuvat Sh-60:n muodostamaan erityiseen tilaan. Ш-60 toimivat oligomeerisena kompleksina, joka koostuu 14 alayksiköstä. Ne muodostavat kaksi onttoa rengasta, joista jokainen koostuu seitsemästä alayksiköstä, nämä renkaat ovat yhteydessä toisiinsa. Jokainen Sh-60-alayksikkö koostuu kolmesta domeenista: apikaalinen (apikaalinen), rikastettu hydrofobisilla radikaaleilla renkaan onteloa kohti, väli- ja ekvatoriaalinen (kuva 1.28).
 Riisi. 1.28. 14 Ш-60 koostuvan chaperoniinikompleksin rakenne
Riisi. 1.28. 14 Ш-60 koostuvan chaperoniinikompleksin rakenne
A - sivukuva; B - ylhäältä katsottuna
Syntetisoidut proteiinit, joiden pinnalla on laskostumattomille molekyyleille ominaisia elementtejä, erityisesti hydrofobisia radikaaleja, tulevat chaperonirenkaiden onteloon. Näiden onteloiden tietyssä ympäristössä mahdollisia konformaatioita etsitään, kunnes löydetään ainoa, joka on energeettisesti edullisin (kuva 1.27, B). Konformaatioiden muodostumiseen ja proteiinin vapautumiseen liittyy ATP-hydrolyysi ekvatoriaalisella alueella. Tyypillisesti tällainen chaperone-riippuvainen taittaminen vaatii huomattavan määrän energiaa.
Sen lisäksi, että chaperonit osallistuvat proteiinien kolmiulotteisen rakenteen muodostukseen ja osittain denaturoituneiden proteiinien renativoimiseen, ne ovat välttämättömiä myös perusprosessien, kuten oligomeeristen proteiinien kokoamisen, denaturoituneiden proteiinien tunnistamisen ja kuljetuksen lysosomeihin, esiintymiseen, proteiinien kuljettaminen kalvojen läpi ja osallistuminen proteiinikompleksien toiminnan säätelyyn.
AIHE 1.6. ERILAISIA PROTEIINEJA. PROTEIINIPERHEET: ESIMERKKI IMMUNOGLOBULIINEISTA
1. Proteiineilla on ratkaiseva rooli yksittäisten solujen ja koko monisoluisen organismin elämässä, ja niiden toiminnot ovat yllättävän monipuoliset. Tämän määräävät proteiinien primäärirakenteen ja konformaatioiden ominaisuudet, aktiivisen keskuksen ainutlaatuinen rakenne ja kyky sitoa spesifisiä ligandeja.
Vain hyvin pieni osa kaikista mahdollisista peptidiketjujen varianteista voi omaksua vakaan avaruudellisen rakenteen; suurin osa
niistä voi olla monia konformaatioita suunnilleen samalla Gibbsin energialla, mutta eri ominaisuuksilla. Useimpien biologisen evoluution perusteella valittujen proteiinien primäärirakenne varmistaa yhden konformaation poikkeuksellisen stabiilisuuden, mikä määrää tämän proteiinin toiminnan ominaisuudet.
2. Proteiiniperheet. Saman biologisen lajin sisällä aminohappotähteiden substituutiot voivat johtaa erilaisten proteiinien syntymiseen, jotka suorittavat toisiinsa liittyviä tehtäviä ja joilla on homologiset aminohapposekvenssit. Tällaisilla sukulaisproteiineilla on hämmästyttävän samankaltaiset konformaatiot: a-heliksien ja/tai p-rakenteiden lukumäärä ja suhteellinen sijainti, useimmat polypeptidiketjujen käännökset ja taivutukset ovat samanlaisia tai identtisiä. Proteiinit, joilla on homologisia polypeptidiketjun alueita, samanlainen konformaatio ja vastaavat toiminnot, luokitellaan proteiiniperheisiin. Esimerkkejä proteiiniperheistä: seriiniproteinaasit, immunoglobuliiniperhe, myoglobiiniperhe.
Seriiniproteinaasit- proteiiniperhe, joka suorittaa proteolyyttisten entsyymien tehtävää. Näitä ovat ruoansulatusentsyymit - kymotrypsiini, trypsiini, elastaasi ja monet veren hyytymistekijät. Näillä proteiineilla on identtiset aminohapot 40 %:ssa asemistaan ja hyvin samanlainen konformaatio (kuva 1.29).

Riisi. 1.29. Elastaasin (A) ja kymotrypsiinin (B) spatiaaliset rakenteet
Jotkut aminohapposubstituutiot ovat johtaneet muutoksiin näiden proteiinien substraattispesifisyydessä ja toiminnallisen monimuotoisuuden syntymiseen perheen sisällä.
3. Immunoglobuliiniperhe. Immuunijärjestelmän toiminnassa immunoglobuliinien superperheen proteiineilla on valtava rooli, joka sisältää kolme proteiiniperhettä:
Vasta-aineet (immunoglobuliinit);
T-lymfosyyttireseptorit;
Tärkeimmän proteiinit - MHC-luokat 1 ja 2 (Major Histocompatibility Complex).
Kaikilla näillä proteiineilla on domeenirakenne, ne koostuvat homologisista immuunityyppisistä domeeneista ja suorittavat samanlaisia toimintoja: ne ovat vuorovaikutuksessa vieraiden rakenteiden kanssa, joko liuenneena vereen, imusolmukkeisiin tai solujen väliseen nesteeseen (vasta-aineet) tai solujen pinnalla (omissa tai ulkomaalainen).
4. Vasta-aineet- B-lymfosyyttien tuottamat spesifiset proteiinit vasteena vieraan rakenteen pääsylle kehoon, ns antigeeni.
Vasta-aineiden rakenteen ominaisuudet
Yksinkertaisimmat vasta-ainemolekyylit koostuvat neljästä polypeptidiketjusta: kahdesta identtisestä kevyestä - L, joka sisältää noin 220 aminohappoa, ja kahdesta identtisestä raskaasta - H, joka koostuu 440-700 aminohaposta. Kaikki neljä vasta-ainemolekyylin ketjua on yhdistetty useilla ei-kovalenttisilla sidoksilla ja neljällä disulfidisidoksella (kuva 1.30).
Vasta-aineen kevyet ketjut koostuvat kahdesta domeenista: variaabelista domeenista (VL), joka sijaitsee polypeptidiketjun N-terminaalisella alueella, ja vakiodomeenista (CL), joka sijaitsee C-päässä. Raskaissa ketjuissa on yleensä neljä domeenia: yksi muuttuja (VH), joka sijaitsee N-päässä, ja kolme vakiodomeenia (CH1, CH2, CH3) (katso kuva 1.30). Jokaisella immunoglobuliinidomeenilla on β-levysuperrakenne, jossa kaksi kysteiinitähdettä on liitetty toisiinsa disulfidisidoksella.
Kahden vakiodomeenin CH1 ja CH2 välissä on alue, joka sisältää suuren määrän proliinitähteitä, jotka estävät sekundaarirakenteen muodostumisen ja viereisten H-ketjujen vuorovaikutuksen tässä segmentissä. Tämä sarana-alue antaa vasta-ainemolekyylille joustavuuden. Raskaan ja kevyen ketjun vaihtelevien domeenien välissä on kaksi identtistä antigeeniä sitovaa kohtaa (aktiivisia kohtia antigeenien sitomiselle), joten tällaisia vasta-aineita kutsutaan usein ns. bivalentteja. Molempien ketjujen vaihtelevien alueiden koko aminohapposekvenssi ei ole osallisena antigeenin sitoutumisessa vasta-aineeseen, vaan vain 20-30 aminohappoa, jotka sijaitsevat kunkin ketjun hypervariaabelialueilla. Juuri nämä alueet määrittävät kunkin vasta-ainetyypin ainutlaatuisen kyvyn olla vuorovaikutuksessa vastaavan komplementaarisen antigeenin kanssa.
Vasta-aineet ovat yksi kehon puolustuslinjoista tunkeutuvia vieraita organismeja vastaan. Niiden toiminta voidaan jakaa kahteen vaiheeseen: ensimmäinen vaihe on antigeenin tunnistaminen ja sitoutuminen vieraiden organismien pinnalle, mikä on mahdollista, koska vasta-ainerakenteessa on antigeeniä sitovia kohtia; toinen vaihe on antigeenin inaktivaatio- ja tuhoutumisprosessin käynnistäminen. Toisen vaiheen spesifisyys riippuu vasta-aineluokasta. Raskasketjuja on viisi luokkaa, jotka eroavat toisistaan vakiodomeenien rakenteessa: α, δ, ε, γ ja μ, joiden mukaan erotetaan viisi immunoglobuliiniluokkaa: A, D, E, G ja M.
Raskaiden ketjujen rakenteelliset ominaisuudet antavat raskaiden ketjujen sarana-alueille ja C-terminaalisille alueille kullekin luokalle ominaisen konformaation. Kun antigeeni on sitoutunut vasta-aineeseen, konformaatiomuutokset vakiodomeeneissa määrittävät antigeenin poistumisreitin.
 Riisi. 1. 30. IgG:n domeenirakenne
Riisi. 1. 30. IgG:n domeenirakenne
Immunoglobuliinit M
Immunoglobuliineilla M on kaksi muotoa.
Monomeerinen muoto- 1. luokka B-lymfosyyttien tuottamia vasta-aineita. Myöhemmin monet B-solut siirtyvät tuottamaan muita vasta-aineluokkia, mutta joilla on sama antigeenia sitova kohta. IgM on upotettu kalvoon ja toimii antigeenintunnistusreseptorina. IgM:n integroituminen solukalvoon on mahdollista, koska alueen häntäosassa on 25 hydrofobista aminohappotähdettä.
IgM:n eritysmuoto sisältää viisi monomeerista alayksikköä, jotka on liitetty toisiinsa disulfidisidoksilla, ja ylimääräisen polypeptidi-J-ketjun (kuva 1.31). Tämän muodon monomeerien raskaat ketjut eivät sisällä hydrofobista häntää. Pentameerissä on 10 antigeenin sitoutumiskohtaa, ja siksi se tunnistaa ja poistaa tehokkaasti antigeenin, joka tulee ensimmäisenä kehoon. IgM:n eritysmuoto on pääasiallinen vasta-aineiden luokka, joka erittyy vereen primaarisen immuunivasteen aikana. IgM:n sitoutuminen antigeeniin muuttaa IgM:n konformaatiota ja indusoi sen sitoutumisen komplementtijärjestelmän ensimmäiseen proteiinikomponenttiin (komplementtijärjestelmä on joukko proteiineja, jotka osallistuvat antigeenin tuhoamiseen) ja tämän järjestelmän aktivoitumisen. Jos antigeeni sijaitsee mikro-organismin pinnalla, komplementtijärjestelmä aiheuttaa solukalvon eheyden häiriöitä ja bakteerisolun kuoleman.
Immunoglobuliinit G
Määrällisesti tämä immunoglobuliiniluokka on vallitseva veressä (75 % kaikista Ig:istä). IgG - monomeerit, pääasiallinen vasta-aineiden luokka, jotka erittyvät vereen sekundaarisen immuunivasteen aikana. IgG:n vuorovaikutuksen jälkeen mikro-organismien pinta-antigeenien kanssa antigeeni-vasta-ainekompleksi pystyy sitomaan ja aktivoimaan komplementtijärjestelmän proteiineja tai voi olla vuorovaikutuksessa makrofagien ja neutrofiilien spesifisten reseptorien kanssa. Vuorovaikutus fagosyyttien kanssa johtaa
 Riisi. 1.31. IgM:n erittävän muodon rakenne
Riisi. 1.31. IgM:n erittävän muodon rakenne
antigeeni-vasta-ainekompleksien imeytymiseen ja niiden tuhoutumiseen solun fagosomeissa. IgG on ainoa vasta-aineluokka, joka pystyy tunkeutumaan istukan esteen läpi ja suojaamaan sikiötä kohdunsisäisesti infektioilta.
Immunoglobuliinit A
Eritteissä (maito, sylki, hengitysteiden ja suoliston eritteet) olevien vasta-aineiden pääluokka. IgA erittyy ensisijaisesti dimeerisessä muodossa, jossa monomeerit ovat liittyneet toisiinsa ylimääräisen J-ketjun kautta (kuva 1.32).
IgA ei ole vuorovaikutuksessa komplementtijärjestelmän ja fagosyyttisolujen kanssa, mutta sitoutumalla mikro-organismeihin vasta-aineet estävät niiden kiinnittymisen epiteelisoluihin ja tunkeutumisen kehoon.
Immunoglobuliinit E
Immunoglobuliineja E edustavat monomeerit, joissa raskaat e-ketjut sisältävät, kuten immunoglobuliinien M μ-ketjut, yhden vaihtelevan ja neljä vakiodomeenia. Erittymisen jälkeen IgE sitoutuu sen kanssa
 Riisi. 1.32. IgA:n rakenne
Riisi. 1.32. IgA:n rakenne
C-terminaaliset alueet, joissa on vastaavat reseptorit syöttösolujen ja basofiilien pinnalla. Tämän seurauksena niistä tulee näiden solujen pinnalla olevien antigeenien reseptoreita (kuva 1.33).
 Riisi. 1.33. IgE:n vuorovaikutus antigeenin kanssa syöttösolun pinnalla
Riisi. 1.33. IgE:n vuorovaikutus antigeenin kanssa syöttösolun pinnalla
Kun antigeeni on kiinnittynyt vastaaviin IgE:n antigeeniä sitoviin kohtiin, solut saavat signaalin erittää biologisesti aktiivisia aineita (histamiinia, serotoniinia), jotka ovat suurelta osin vastuussa tulehdusreaktion kehittymisestä ja allergisten reaktioiden, kuten esim. astma, urtikaria, heinänuha.
Immunoglobuliinit D
Immunoglobuliineja D löytyy hyvin pieninä määrinä seerumista, ne ovat monomeerejä. Raskaissa δ-ketjuissa on yksi muuttuva ja kolme vakiodomeenia. IgD:t toimivat B-lymfosyyttien reseptoreina. Muita toimintoja ei vielä tunneta. Spesifisten antigeenien vuorovaikutus B-lymfosyyttien (IgD) pinnalla olevien reseptoreiden kanssa johtaa näiden signaalien siirtymiseen soluun ja sellaisten mekanismien aktivoitumiseen, jotka varmistavat tietyn lymfosyyttikloonin lisääntymisen.
AIHE 1.7. PROTEIINIEN FYSIKAALISET JA KEMIALLISET OMINAISUUDET JA NIIDEN EROTUSMENETELMÄT
1. Yksittäiset proteiinit eroavat fysikaalisista ja kemiallisista ominaisuuksista:
Molekyylien muoto;
Molekyylipaino;
Kokonaisvaraus, jonka suuruus riippuu aminohappojen anionisten ja kationisten ryhmien suhteesta;
Polaaristen ja ei-polaaristen aminohapporadikaalien suhde molekyylien pinnalla;
Erilaisten denaturointiaineiden kestävyysasteet.
2. Proteiinien liukoisuus riippuu edellä lueteltujen proteiinien ominaisuuksista sekä alustan koostumuksesta, johon proteiini on liuennut (pH-arvot, suolakoostumus, lämpötila, muiden orgaanisten aineiden läsnäolo, jotka voivat olla vuorovaikutuksessa proteiinin kanssa). Proteiinimolekyylien varausmäärä on yksi niiden liukoisuuteen vaikuttavista tekijöistä. Kun isoelektrisen pisteen varaus katoaa, proteiinit aggregoituvat ja saostuvat helpommin. Tämä on erityisen tyypillistä denaturoiduille proteiineille, joiden pinnalle ilmestyy hydrofobisia aminohapporadikaaleja.
Proteiinimolekyylin pinnalla on sekä positiivisesti että negatiivisesti varautuneita aminohapporadikaaleja. Näiden ryhmien lukumäärä ja siten proteiinien kokonaisvaraus riippuu väliaineen pH:sta, ts. H+ - ja OH - ryhmien pitoisuuksien suhde. Happamassa ympäristössä H+-konsentraation kasvu johtaa karboksyyliryhmien -COO - + H+ > -COOH dissosiaation suppressioon ja proteiinien negatiivisen varauksen vähenemiseen. Alkalisessa ympäristössä ylimääräisen OH - sitoutuminen protoneilla, jotka muodostuvat aminoryhmien -NH 3 + + OH - - NH 2 + H 2 O hajoamisen aikana veden muodostumisen kanssa, johtaa proteiinien positiivisen varauksen vähenemiseen . pH-arvoa, jossa proteiinin nettovaraus on nolla, kutsutaan isoelektrinen piste (IEP). IET:ssä positiivisesti ja negatiivisesti varautuneiden ryhmien määrä on sama, ts. proteiini on isoelektrisessä tilassa.
3. Yksittäisten proteiinien erottaminen. Kehon rakenteen ja toiminnan ominaisuudet riippuvat siinä syntetisoitujen proteiinien joukosta. Proteiinien rakenteen ja ominaisuuksien tutkiminen on mahdotonta ilman niiden eristämistä solusta ja puhdistamista muista proteiineista ja orgaanisista molekyyleistä. Yksittäisten proteiinien eristyksen ja puhdistamisen vaiheet:
Solujen tuhoutuminen tutkittava kudos ja homogenaatin saaminen.
Homogenaatin erottaminen fraktioiksi sentrifugoimalla saadaan tuma-, mitokondrio-, sytosolinen tai muu fraktio, joka sisältää halutun proteiinin.
Valikoiva lämpödenaturointi- proteiiniliuoksen lyhytaikainen kuumennus, jonka aikana osa denaturoituneista proteiiniepäpuhtauksista voidaan poistaa (jos proteiini on suhteellisen lämmönkestävää).
Suolaus pois. Eri proteiinit saostuvat eri suolapitoisuuksilla liuoksessa. Vähitellen suolapitoisuutta lisäämällä on mahdollista saada useita erillisiä fraktioita, joissa yhdessä niistä on vallitseva pitoisuus eristettyä proteiinia. Ammoniumsulfaattia käytetään useimmiten proteiinien fraktiointiin. Vähiten liukenevat proteiinit saostuvat alhaisilla suolapitoisuuksilla.
Geelisuodatus- menetelmä molekyylien seulomiseksi turvonneiden Sephadex-rakeiden läpi (kolmiulotteiset dekstraanin polysakkaridiketjut, joissa on huokoset). Nopeus, jolla proteiinit kulkevat Sephadexillä täytetyn kolonnin läpi, riippuu niiden molekyylipainosta: mitä pienempi proteiinimolekyylien massa on, sitä helpommin ne tunkeutuvat rakeisiin ja pysyvät siellä pidempään, sitä nopeammin ne eluoituvat sarakkeessa.
Ultrasentrifugointi- menetelmä, jossa proteiinit asetetaan sentrifugiputkessa ultrasentrifugin roottoriin. Kun roottori pyörii, proteiinien sedimentaationopeus on verrannollinen niiden molekyylipainoon: raskaampien proteiinien fraktiot sijaitsevat lähempänä koeputken pohjaa, kevyemmät - lähempänä pintaa.
Elektroforeesi- menetelmä, joka perustuu eroihin proteiinien liikkumisnopeudessa sähkökentässä. Tämä arvo on verrannollinen proteiinien varaukseen. Proteiinien elektroforeesi suoritetaan paperilla (tässä tapauksessa proteiinin liikkeen nopeus on verrannollinen vain niiden varaukseen) tai polyakryyliamidigeelissä, jolla on tietty huokoskoko (proteiinin liikkeen nopeus on verrannollinen niiden varaukseen ja molekyylipainoon) .
Ioninvaihtokromatografia- fraktiointimenetelmä, joka perustuu ionisoituneiden proteiiniryhmien sitomiseen vastakkaisesti varautuneisiin ioninvaihtohartsiryhmiin (liukenemattomat polymeerimateriaalit). Proteiinin hartsiin sitoutumisen voimakkuus on verrannollinen proteiinin varaukseen. Ioninvaihtopolymeeriin adsorboituneet proteiinit voidaan pestä pois NaCl-liuosten kasvavilla pitoisuuksilla; mitä pienempi proteiinivaraus, sitä pienempi NaCl-pitoisuus vaaditaan hartsin ioniryhmiin sitoutuneen proteiinin pesemiseksi pois.
Affiniteettikromatografia- spesifisin menetelmä yksittäisten proteiinien eristämiseksi. Proteiinin ligandi kiinnitetään kovalenttisesti inerttiin polymeeriin. Kun proteiiniliuos johdetaan kolonnin läpi polymeerin kanssa, vain tietylle ligandille spesifinen proteiini adsorboituu pylvääseen johtuen proteiinin komplementaarisesta sitoutumisesta ligandiin.
Dialyysi- menetelmä, jota käytetään pienen molekyylipainon yhdisteiden poistamiseen eristetyn proteiinin liuoksesta. Menetelmä perustuu proteiinien kyvyttömyyteen kulkea puoliläpäisevän kalvon läpi, toisin kuin pienimolekyylipainoiset aineet. Sitä käytetään proteiinien puhdistamiseen pienimolekyylisistä epäpuhtauksista, esimerkiksi suoloista suolauksen jälkeen.
TEHTÄVÄT OPIN ULKOPUOLISESTI
1. Täytä taulukko. 1.4.
Taulukko 1.4. Sukulaisten proteiinien - myoglobiinin ja hemoglobiinin - rakenteen ja toimintojen vertaileva analyysi
a) muista Mb:n ja Hb:n aktiivisen keskuksen rakenne. Mikä rooli hydrofobisilla aminohapporadikaaleilla on näiden proteiinien aktiivisten keskusten muodostumisessa? Kuvaile Mb:n ja Hb:n aktiivisen keskuksen rakennetta ja ligandien kiinnittymismekanismeja. Mikä rooli His F 8- ja His E 7 -tähteillä on Mv iHv:n aktiivisen keskuksen toiminnassa?
b) mitä uusia ominaisuuksia monomeeriseen myoglobiiniin verrattuna on läheistä sukua olevalla oligomeeriproteiinilla, hemoglobiinilla? Selitä kooperatiivisten muutosten rooli hemoglobiinimolekyylin protomeerien konformaatiossa, CO 2 - ja protonipitoisuuksien vaikutus hemoglobiinin happiaffiniteettiin sekä 2,3-BPG:n rooli Hb-toiminnan allosteerisessa säätelyssä. .
2. Kuvaile molekyylikaperoneja kiinnittäen huomiota niiden rakenteen ja toiminnan väliseen suhteeseen.
3. Mitkä proteiinit on ryhmitelty perheisiin? Tunnista immunoglobuliiniperheen esimerkkiä käyttäen tämän perheen proteiinien samanlaiset rakenteelliset piirteet ja niihin liittyvät toiminnot.
4. Puhdistettuja yksittäisiä proteiineja tarvitaan usein biokemiallisiin ja lääketieteellisiin tarkoituksiin. Selitä, mihin proteiinien fysikaalis-kemiallisiin ominaisuuksiin niiden erottamiseen ja puhdistamiseen käytetyt menetelmät perustuvat.
ITSEVALTOTEHTÄVÄT
1. Valitse oikeat vastaukset.
Hemoglobiinin tehtävät:
A. O 2:n kuljetus keuhkoista kudokseen B. H +:n kuljetus kudoksista keuhkoihin
B. Veren pH-arvon ylläpitäminen D. CO 2:n kuljetus keuhkoista kudoksiin
D. CO 2:n kuljetus kudoksista keuhkoihin
2. Valitse oikeat vastaukset. Ligandiα -protomeeri Hb on: A. Heme
B. Happi
B. CO G. 2,3-BPG
D. P-protomeeri
3. Valitse oikeat vastaukset.
Hemoglobiini toisin kuin myoglobiini:
A. Sillä on kvaternäärinen rakenne
B. Toissijaista rakennetta edustavat vain α-heliksit
B. Kuuluu monimutkaisiin proteiineihin
D. Vuorovaikuttaa allosteerisen ligandin kanssa D. Kovalenttisesti sitoutunut hemiin
4. Valitse oikeat vastaukset.
Hb:n affiniteetti O2:lle laskee:
A. Kun yksi O 2 -molekyyli lisätään B. Kun yksi O 2 -molekyyli poistetaan
B. Kun olet vuorovaikutuksessa 2,3-BPG:n kanssa
D. Kiinnitettynä protomeereihin H + D. Kun 2,3-BPG:n pitoisuus pienenee
5. Ottelu.
HB-tyypeille on ominaista:
A. Deoksimuodossa se muodostaa fibrillaarisia aggregaatteja B. Sisältää kaksi α- ja kaksi δ-ketjua
B. Hb:n vallitseva muoto aikuisten punasoluissa D. Sisältää hemiä, jonka aktiivisessa keskustassa on Fe+ 3
D. Sisältää kaksi α- ja kaksi γ-ketjua 1. HbA 2.
6. Ottelu.
Hb-ligandit:
A. Sitoutuu Hb:hen allosteerisessa keskustassa
B. Sillä on erittäin korkea affiniteetti Hb:n aktiiviseen kohtaan
B. Liittymällä se lisää Hb:n affiniteettia O 2 G:n suhteen. Hapettaa Fe+ 2:n Fe+ 3:ksi
D. Muodostaa kovalenttisen sidoksen hisF8:n kanssa
7. Valitse oikeat vastaukset.
Saattajat:
A. Proteiinit, joita on solun kaikissa osissa
B. Synteesi lisääntyy stressissä
B. Osallistu denaturoitujen proteiinien hydrolyysiin
D. Osallistu proteiinien luonnollisen konformaation ylläpitämiseen
D. Ne luovat organelleja, joissa muodostuu proteiinien konformaatio.
8. Ottelu. Immunoglobuliinit:
A. Eritysmuoto on pentameerinen.
B. Ig-luokka, joka läpäisee istukan
B. Ig - syöttösolureseptori
D. Ig:n pääluokka, joka on läsnä epiteelisolujen eritteissä. D. B-lymfosyyttireseptori, jonka aktivointi varmistaa solujen lisääntymisen
9. Valitse oikeat vastaukset.
Immunoglobuliinit E:
A. Makrofagien tuottamat B. Niissä on raskaita e-ketjuja.
B. Upotettu T-lymfosyyttien kalvoon
D. Toimivat kalvoantigeenireseptoreina syöttösoluissa ja basofiileissä
D. Vastuussa allergisista reaktioista
10. Valitse oikeat vastaukset.
Proteiinien erottelumenetelmä perustuu niiden molekyylipainon eroihin:
A. Geelisuodatus
B. Ultrasentrifugointi
B. PolD. Ioninvaihtokromatografia
D. Affiniteettikromatografia
11. Valitse oikea vastaus.
Proteiinien erottelumenetelmä perustuu eroihin niiden vesiliukoisuudessa:
A. Geelisuodatus B. Suolaus
B. Ioninvaihtokromatografia D. Affiniteettikromatografia
D. Elektroforeesi polyakryyliamidigeelissä
VASTAUKSIEN STANDARDIT ”ITSEVALTOTEHTÄVÄIN”
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
PERUSKÄSITTEET JA KÄSITTEET
1. Oligomeeriset proteiinit, protomeeri, proteiinien kvaternäärinen rakenne
2. Yhteistoiminnalliset muutokset protomeerin konformaatiossa
3. Bohr-ilmiö
4. Proteiinitoimintojen allosteerinen säätely, allosteerinen keskus ja allosteerinen efektori
5. Molekyylikaperonit, lämpöshokkiproteiinit
6. Proteiiniperheet (seriiniproteaasit, immunoglobuliinit)
7. IgM-, G-, E-, A-rakenne-funktiosuhde
8. Proteiinien kokonaisvaraus, proteiinien isoelektrinen piste
9. Elektroforeesi
10. Suolaus
11. Geelisuodatus
12. Ioninvaihtokromatografia
13. Ultrasentrifugointi
14. Affiniteettikromatografia
15. Veriplasman proteiinien elektroforeesi
TEHTÄVÄT LUOKKATYÖHÄN
1. Vertaa hemoglobiinin (Hb) ja myoglobiinin (Mb) saturaatioasteiden riippuvuutta hapella sen osapaineesta kudoksissa
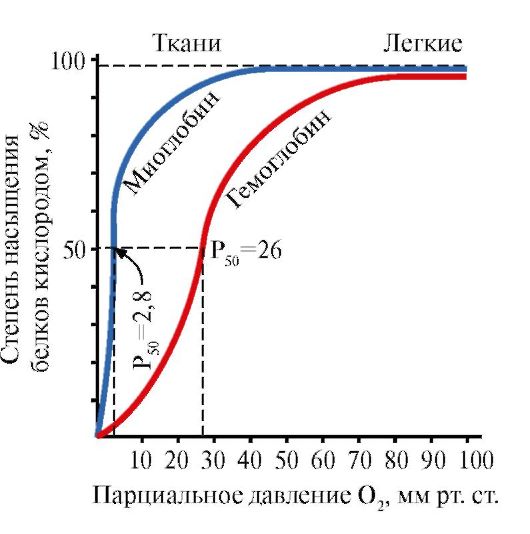 Riisi. 1.34. Kyllästymisen riippuvuus Mv jaNHhappea sen osapaineesta
Riisi. 1.34. Kyllästymisen riippuvuus Mv jaNHhappea sen osapaineesta
Huomaa, että proteiinin happisaturaatiokäyrien muoto on erilainen: myoglobiinilla - hyperbola, hemoglobiinilla - sigmoidimuoto.
1. Vertaa hapen osapaineen arvoja, joissa Mb ja Hb ovat kyllästyneet O 2:lla 50 %. Millä näistä proteiineista on suurempi affiniteetti O 2:een?
2. Mitkä Mb:n rakenteelliset ominaisuudet määräävät sen suuren affiniteetin O 2:lle?
3. Mitkä HB:n rakenteelliset ominaisuudet sallivat sen vapauttaa O2:ta lepäävien kudosten kapillaareista (suhteellisen korkealla O2:n osapaineella) ja lisätä tätä vapautumista jyrkästi toimivissa lihaksissa? Mikä oligomeeristen proteiinien ominaisuus tarjoaa tämän vaikutuksen?
4. Laske kuinka paljon O 2 (%) hapetettua hemoglobiinia antaa lepääville ja työskenteleville lihaksille?
5. tehdä johtopäätöksiä proteiinin rakenteen ja toiminnan välisestä suhteesta.
2. Hemoglobiinin kapillaareissa vapauttaman hapen määrä riippuu kudosten katabolisten prosessien voimakkuudesta (Bohr-ilmiö). Kuinka muutokset kudosten aineenvaihdunnassa säätelevät Hb:n affiniteettia O2:een? CO 2:n ja H+:n vaikutus Hb:n affiniteettiin O 2:ta kohtaan
 1. kuvaa Bohrin vaikutusta.
1. kuvaa Bohrin vaikutusta.
2. mihin suuntaan kaaviossa näkyvä prosessi etenee:
a) keuhkojen kapillaareissa;
b) kudoskapillaareissa?
3. Mikä on Bohr-ilmiön fysiologinen merkitys?
4. Miksi Hb:n ja H+:n vuorovaikutus hemistä kaukana olevissa kohdissa muuttaa proteiinin affiniteettia O 2:lle?
3. Hb:n affiniteetti 02:lle riippuu sen ligandin pitoisuudesta - 2,3-bisfosfoglyseraatti, joka on allosteerinen säätelijä Hb:n affiniteetille O2:lle. Miksi ligandin vuorovaikutus aktiivisesta kohdasta kaukana olevassa kohdassa vaikuttaa proteiinin toimintaan? Kuinka 2,3-BPG säätelee Hb:n affiniteettia O2:een? Voit ratkaista ongelman vastaamalla seuraaviin kysymyksiin:
1. mistä ja mistä 2,3-bisfosfoglyseraattia (2,3-BPG) syntetisoidaan? Kirjoita sen kaava, osoita tämän molekyylin varaus.
2. Minkä hemoglobiinimuodon (oksi tai deoksi) kanssa BPG on vuorovaikutuksessa ja miksi? Missä Hb-molekyylin osassa vuorovaikutus tapahtuu?
3. mihin suuntaan kaaviossa esitetty prosessi tapahtuu?
a) kudosten kapillaareissa;
b) keuhkojen kapillaareissa?
4. jossa kompleksin pitoisuuden tulisi olla suurempi
Nv-2,3-BFG:
a) levossa olevien lihasten kapillaareissa,
b) työskentelevien lihasten kapillaareissa (edellyttäen, että punasoluissa on sama BPG-pitoisuus)?
5. Miten HB:n affiniteetti hapen suhteen muuttuu, kun ihminen sopeutuu korkeisiin olosuhteisiin, jos punasolujen BPG-pitoisuus kasvaa? Mikä on tämän ilmiön fysiologinen merkitys?
4. 2,3-BPG:n tuhoutuminen säilötyn veren varastoinnin aikana heikentää HB:n toimintaa. Miten HB:n affiniteetti O 2:lle muuttuu säilyneessä veressä, jos 2,3-BPG:n pitoisuus punasoluissa voi laskea 8:sta 0,5 mmol/l:aan. Onko mahdollista siirtää tällaista verta vakavasti sairaille potilaille, jos 2,3-BPG-pitoisuus palautetaan aikaisintaan kolmen päivän kuluttua? Onko mahdollista palauttaa punasolujen toiminta lisäämällä vereen 2,3-BPG:tä?
5. Muista yksinkertaisimpien immunoglobuliinimolekyylien rakenne. Mikä rooli immunoglobuliineilla on immuunijärjestelmässä? Miksi iggejä kutsutaan usein bivalentteiksi? Miten Ig:n rakenne liittyy niiden toimintaan? (Kuvaile käyttämällä esimerkkiä immunoglobuliiniluokista.)
Proteiinien fysikaalis-kemialliset ominaisuudet ja menetelmät niiden erottamiseen.
6. Miten proteiinin nettovaraus vaikuttaa sen liukoisuuteen?
a) määritä peptidin kokonaisvaraus pH:ssa 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) kuinka tämän peptidin varaus muuttuu pH:ssa >7, pH<7, рН <<7?
c) mikä on proteiinin isoelektrinen piste (IEP) ja missä ympäristössä se sijaitsee?
Tämän peptidin IET?
d) missä pH-arvossa tämän peptidin pienin liukoisuus havaitaan.
7. Miksi piimä, toisin kuin tuore maito, "juostuu" keitettäessä (eli maitoproteiini kaseiini saostuu)? Tuoreessa maidossa kaseiinimolekyyleillä on negatiivinen varaus.
8. Geelisuodatusta käytetään yksittäisten proteiinien erottamiseen. Seos, joka sisälsi proteiineja A, B ja C, joiden molekyylipainot ovat 160 000, 80 000 ja 60 000, analysoitiin geelisuodatuksella (kuva 1.35). Turvonneet geelirakeet läpäisevät proteiineja, joiden molekyylipaino on alle 70 000. Mikä periaate on tämän erotusmenetelmän taustalla? Mikä kuvaaja heijastaa oikein fraktioinnin tuloksia? Ilmoita järjestys, jossa proteiinit A, B ja C vapautuvat kolonnista.
 Riisi. 1.35. Geelisuodatuksen käyttö proteiinien erottamiseen
Riisi. 1.35. Geelisuodatuksen käyttö proteiinien erottamiseen
9. Kuvassa 1.36, A esittää kaavion elektroforeesista paperille terveen ihmisen veren seerumin proteiineista. Tällä menetelmällä saatujen proteiinifraktioiden suhteelliset määrät ovat: albumiinit 54-58 %, a1-globuliinit 6-7 %, a2-globuliinit 8-9 %, p-globuliinit 13 %, y-globuliinit 11-12 %.
 Riisi. 1.36 Terveen ihmisen (A) ja potilaan (B) veriplasmaproteiinien elektroforeesi paperille
Riisi. 1.36 Terveen ihmisen (A) ja potilaan (B) veriplasmaproteiinien elektroforeesi paperille
I - y-globuliinit; II - p-globuliinit; III -α 2-globuliini; IV -α 2-globuliini; V - albumiinit
Moniin sairauksiin liittyy kvantitatiivisia muutoksia seerumin proteiinien koostumuksessa (dysproteinemia). Näiden muutosten luonne otetaan huomioon diagnoosia tehtäessä sekä taudin vaikeusastetta ja vaihetta arvioitaessa.
Käyttämällä taulukossa annettuja tietoja. 1.5, tee arvaus sairaudesta, jolle on tunnusomaista kuvassa 1 esitetty elektroforeettinen profiili. 1.36.
Taulukko 1.5. Muutokset seerumin proteiinien pitoisuudessa patologiassa
Proteiinit ovat orgaanisia aineita. Näille suurimolekyylisille yhdisteille on ominaista tietty koostumus, ja ne hajoavat hydrolysoituessaan aminohapoiksi. Proteiinimolekyylit voivat olla monissa eri muodoissa, joista monet koostuvat useista polypeptidiketjuista. Tieto proteiinin rakenteesta on koodattu DNA:han, ja proteiinimolekyylien synteesiprosessia kutsutaan translaatioksi.
Proteiinien kemiallinen koostumus
Keskimääräinen proteiini sisältää:
- 52 % hiiltä;
- 7 % vetyä;
- 12 % typpeä;
- 21 % happea;
- 3% rikkiä.
Proteiinimolekyylit ovat polymeerejä. Niiden rakenteen ymmärtämiseksi on tarpeen tietää, mitä niiden monomeerit - aminohapot - ovat.
Aminohappoja
Ne jaetaan yleensä kahteen luokkaan: jatkuvasti esiintyviin ja satunnaisiin. Ensimmäiset sisältävät 18 ja 2 muuta amidia: asparagiinihappo ja glutamiinihappo. Joskus löytyy vain kolme happoa.
Nämä hapot voidaan luokitella eri tavoin: sivuketjujen luonteen tai niiden radikaalien varauksen perusteella ne voidaan jakaa myös CN- ja COOH-ryhmien lukumäärällä.
Ensisijainen proteiinirakenne
Aminohappojen vuorottelujärjestys proteiiniketjussa määrittää sen myöhemmät organisaatiotasot, ominaisuudet ja toiminnot. Pääasiallinen monomeerien välillä on peptidi. Se muodostuu ottamalla vetyä yhdestä aminohaposta ja OH-ryhmää toisesta.
Proteiinimolekyylin ensimmäinen organisoitumistaso on siinä olevien aminohappojen sekvenssi, yksinkertaisesti ketju, joka määrittää proteiinimolekyylien rakenteen. Se koostuu "luurangosta", jolla on säännöllinen rakenne. Tämä on toistuva sekvenssi -NH-CH-CO-. Yksittäisiä sivuketjuja edustavat aminohapporadikaalit (R), niiden ominaisuudet määräävät proteiinirakenteen koostumuksen.
Vaikka proteiinimolekyylien rakenne on sama, ne voivat erota ominaisuuksiltaan vain siksi, että niiden monomeerien ketjussa on erilainen sekvenssi. Aminohappojen järjestys proteiinissa määräytyy geenien avulla, ja se sanelee proteiinille tietyt biologiset toiminnot. Monomeerien sekvenssi samasta tehtävästä vastaavissa molekyyleissä on usein samanlainen eri lajeissa. Tällaiset molekyylit ovat organisaatioltaan identtisiä tai samankaltaisia ja suorittavat samoja tehtäviä eri tyyppisissä organismeissa - homologisissa proteiineissa. Tulevien molekyylien rakenne, ominaisuudet ja toiminnot selviävät jo aminohappoketjun synteesivaiheessa.
Jotkut yhteiset ominaisuudet
Proteiinien rakennetta on tutkittu pitkään ja niiden primäärirakenteen analyysi on mahdollistanut yleistysten tekemisen. Suuremmalle määrälle proteiineja on ominaista kaikkien kahdenkymmenen aminohapon läsnäolo, joista erityisen paljon on glysiiniä, alaniinia, glutamiinia ja vähän tryptofaania, arginiinia, metioniinia ja histidiiniä. Ainoat poikkeukset ovat jotkin proteiiniryhmät, esimerkiksi histonit. Niitä tarvitaan DNA:n pakkaamiseen ja ne sisältävät paljon histidiiniä.
Kaikenlaiset organismien liikkeet (lihastyö, protoplasman liike solussa, värekkojen välkkyminen alkueläimissä jne.) suoritetaan proteiinien avulla. Proteiinien rakenteen ansiosta ne voivat liikkua, muodostaa kuituja ja renkaita.

Kuljetustoiminto on, että monet aineet kuljetetaan solukalvon läpi erityisten kantajaproteiinien avulla.
Näiden polymeerien hormonaalinen rooli on heti selvä: monet hormonit ovat rakenteeltaan proteiineja, esimerkiksi insuliini, oksitosiini.
Varastofunktio määräytyy sen perusteella, että proteiinit pystyvät muodostamaan kerrostumia. Esimerkiksi munavalgumiini, maitokeiini, kasvien siemenproteiinit - ne varastoivat suuren määrän ravintoaineita.
Kaikki jänteet, nivelnivelet, luuston luut ja kaviot muodostuvat proteiineista, mikä vie meidät niiden seuraavaan tehtävään - tukeen.
Proteiinimolekyylit ovat reseptoreita, jotka tunnistavat selektiivisesti tiettyjä aineita. Glykoproteiinit ja lektiinit ovat erityisen tunnettuja tästä roolista.
Tärkeimmät immuniteetin tekijät ovat vasta-aineet ja ne ovat peräisin proteiineista. Esimerkiksi veren hyytymisprosessi perustuu fibrinogeeniproteiinin muutoksiin. Ruokatorven ja mahan sisäseinämät on vuorattu suojaavalla kerroksella limakalvoproteiineja - lysiinejä. Toksiinit ovat myös alkuperältään proteiineja. Eläinten kehoa suojaavan ihon perusta on kollageeni. Kaikki nämä proteiinitoiminnot ovat suojaavia.

No, viimeinen toiminto on säätely. On proteiineja, jotka säätelevät genomin toimintaa. Eli ne säätelevät transkriptiota ja kääntämistä.
Huolimatta siitä, kuinka tärkeä rooli proteiinilla on, tiedemiehet ovat selvittäneet proteiinien rakenteen jo kauan sitten. Ja nyt he löytävät uusia tapoja käyttää tätä tietoa.
Lippu 2. 1. Lipidiluonnon olennaiset ravitsemukselliset tekijät. Joitakin lipidejä ei syntetisoidu ihmiskehossa ja ne ovat siksi tärkeitä ravitsemuksellisia tekijöitä. Näitä ovat rasvahapot, joissa on kaksi tai useampi kaksoissidos (polyeeni) - välttämättömiä rasvahappoja. Jotkut näistä hapoista ovat substraatteja paikallisten hormonien - eikosanoidien - synteesiin (aihe 8.10).
Rasvaliukoiset vitamiinit suorittaa erilaisia toimintoja: A-vitamiini osallistuu näköprosessiin sekä solujen kasvuun ja erilaistumiseen; sen kyky estää tietyntyyppisten kasvainten kasvua on todistettu; K-vitamiinia osallistuu veren hyytymiseen; D-vitamiini osallistuu kalsiumin aineenvaihdunnan säätelyyn; E-vitamiinia- antioksidantti, estää vapaiden radikaalien muodostumista ja siten ehkäisee lipidiperoksidaatiosta johtuvia soluvaurioita.
Asiakirja
2. Proteiinien rakenne ja rakenteellisen organisoinnin tasot
Proteiinien rakenteellisessa organisaatiossa on neljä tasoa: primaarinen, sekundaarinen, tertiäärinen ja kvaternäärinen. Jokaisella tasolla on omat ominaisuutensa.
Ensisijainen proteiinirakenne
Proteiinien päärakenne on lineaarinen polypeptidiketju aminohapoista, jotka on yhdistetty peptidisidoksilla. Primäärirakenne on proteiinimolekyylin rakenteellisen organisoinnin yksinkertaisin taso. Korkean stabiilisuuden antavat sille kovalenttiset peptidisidokset yhden aminohapon a-aminoryhmän ja toisen aminohapon a-karboksyyliryhmän välillä. [näytä].
Jos proliinin tai hydroksiproliinin iminoryhmä on osallisena peptidisidoksen muodostumisessa, sillä on eri muoto [näytä].
Kun soluihin muodostuu peptidisidoksia, yhden aminohapon karboksyyliryhmä aktivoituu ensin, ja sitten se yhdistyy toisen aminoryhmän kanssa. Polypeptidien laboratorio synteesi suoritetaan suunnilleen samalla tavalla.
Peptidisidos on polypeptidiketjun toistuva fragmentti. Sillä on useita ominaisuuksia, jotka eivät vaikuta vain primaarirakenteen muotoon, vaan myös polypeptidiketjun korkeampiin organisoitumistasoihin:

samantasoisuus - kaikki peptidiryhmään sisältyvät atomit ovat samassa tasossa;
kyky esiintyä kahdessa resonanssimuodossa (keto- tai enolimuoto);
substituenttien trans-asema suhteessa C-N-sidokseen;
kyky muodostaa vetysidoksia, ja jokainen peptidiryhmä voi muodostaa kaksi vetysidosta muiden ryhmien kanssa, mukaan lukien peptidiryhmät.
Poikkeuksen muodostavat peptidiryhmät, joissa on proliinin tai hydroksiproliinin aminoryhmä. Ne pystyvät muodostamaan vain yhden vetysidoksen (katso edellä). Tämä vaikuttaa proteiinin sekundaarirakenteen muodostumiseen. Polypeptidiketju alueella, jossa proliini tai hydroksiproliini sijaitsee, taipuu helposti, koska sitä ei pidä, kuten tavallista, toinen vetysidos.
Peptidien ja polypeptidien nimikkeistö. Peptidien nimi koostuu niiden aminohappojen nimistä. Kaksi aminohappoa muodostaa dipeptidin, kolme muodostaa tripeptidin, neljä muodostavat tetrapeptidin jne. Jokaisessa minkä tahansa pituisessa peptidissä tai polypeptidiketjussa on N-terminaalinen aminohappo, joka sisältää vapaan aminoryhmän, ja C-terminaalinen aminohappo, joka sisältää vapaan karboksyylin ryhmä. Polypeptidien nimeämisessä kaikki aminohapot luetellaan peräkkäin alkaen N-päästä ja korvataan niiden nimissä C-terminaalista lukuun ottamatta -in -liite -yylillä (koska peptidien aminohapoilla ei enää ole karboksyyliryhmä, mutta karbonyyliryhmä). Esimerkiksi kuvassa näkyvä nimi. 1 tripeptidi - leuc lietettä fenyylialaani lietettä treoni sisään.
Proteiinin perusrakenteen ominaisuudet. Polypeptidiketjun rungossa jäykät rakenteet (litteät peptidiryhmät) vuorottelevat suhteellisen liikkuvien alueiden (-CHR) kanssa, jotka pystyvät pyörimään sidosten ympärillä. Sellaiset polypeptidiketjun rakenteelliset piirteet vaikuttavat sen tilajärjestelyyn.
Proteiinin sekundaarinen rakenne
Toissijainen rakenne on tapa laskostaa polypeptidiketju järjestetyksi rakenteeksi johtuen vetysidosten muodostumisesta saman ketjun peptidiryhmien tai vierekkäisten polypeptidiketjujen välille. Toissijaiset rakenteet jaetaan konfiguraationsa mukaan kierteisiin (α-heliksi) ja kerroslaskostettuihin (β-rakenne ja poikki-β-muoto).


α-helix. Tämä on eräänlainen sekundäärinen proteiinirakenne, joka näyttää tavalliselta kierteeltä, joka muodostuu yhden polypeptidiketjun sisällä olevien peptidien välisten vetysidosten vuoksi. Pauling ja Corey ehdottivat mallia a-heliksin rakenteesta (kuvio 2), joka ottaa huomioon kaikki peptidisidoksen ominaisuudet. α-heliksin tärkeimmät ominaisuudet:
polypeptidiketjun kierteinen konfiguraatio, jolla on helikaalinen symmetria;
vetysidosten muodostuminen kunkin ensimmäisen ja neljännen aminohappotähteen peptidiryhmien välille;
spiraalikierrosten säännöllisyys;
kaikkien α-kierteen aminohappotähteiden vastaavuus niiden sivuradikaalien rakenteesta riippumatta;
aminohappojen sivuradikaalit eivät osallistu α-heliksin muodostumiseen.
Ulkoisesti α-helix näyttää sähköliesi hieman venyneeltä spiraalilta. Vetysidosten säännöllisyys ensimmäisen ja neljännen peptidiryhmän välillä määrää polypeptidiketjun käänteiden säännöllisyyden. Yhden kierroksen korkeus tai a-heliksin nousu on 0,54 nm; se sisältää 3,6 aminohappotähdettä, eli jokainen aminohappotähde liikkuu akselia pitkin (yhden aminohappotähteen korkeus) 0,15 nm (0,54:3,6 = 0,15 nm), mikä antaa mahdollisuuden puhua kaikkien aminohappotähteiden vastaavuudesta α-kierteessä. α-heliksin säännöllisyysjakso on 5 kierrosta tai 18 aminohappotähdettä; yhden jakson pituus on 2,7 nm. Riisi. 3. Pauling-Corey a-helix malli
β-rakenne. Tämä on eräänlainen sekundaarirakenne, jolla on hieman kaareva polypeptidiketjun konfiguraatio ja jonka muodostavat peptidien väliset vetysidokset yhden polypeptidiketjun yksittäisissä osissa tai vierekkäisissä polypeptidiketjuissa. Sitä kutsutaan myös kerrokselliseksi taittorakenteeksi. On olemassa erilaisia β-rakenteita. Proteiinin yhden polypeptidiketjun muodostamia rajoitettuja kerrosalueita kutsutaan cross-β-muodoksi (lyhyt p-rakenne). Polypeptidiketjun silmukoiden peptidiryhmien väliin muodostuu vetysidoksia ristiin-p-muodossa. Toinen tyyppi - täydellinen p-rakenne - on ominaista koko polypeptidiketjulle, jolla on pitkänomainen muoto ja jota pitävät peptidien väliset vetysidokset vierekkäisten rinnakkaisten polypeptidiketjujen välillä (kuvio 3). Tämä rakenne muistuttaa harmonikan palkea. Lisäksi β-rakenteiden variantit ovat mahdollisia: ne voivat muodostua rinnakkaisista ketjuista (polypeptidiketjujen N-terminaaliset päät on suunnattu samaan suuntaan) ja antirinnakkaisina (N-terminaaliset päät on suunnattu eri suuntiin). Yhden kerroksen sivuradikaalit sijoitetaan toisen kerroksen sivuradikaalien väliin.
Proteiineissa siirtymät α-rakenteista β-rakenteisiin ja takaisin ovat mahdollisia vetysidosten uudelleenjärjestelyn vuoksi. Sen sijaan, että ketjussa olisi säännöllisiä peptidien välisiä vetysidoksia (jonka ansiosta polypeptidiketju on kierretty spiraaliksi), kierteiset osiot purkautuvat ja vetysidokset sulkeutuvat polypeptidiketjujen pitkänomaisten fragmenttien väliin. Tämä siirtymä löytyy keratiinista, hiusten proteiinista. Kun hiuksia pestään emäksisellä pesuaineella, β-keratiinin kierteinen rakenne tuhoutuu helposti ja muuttuu α-keratiiniksi (kiharat hiukset suoristuvat).
Proteiinien säännöllisten toissijaisten rakenteiden (α-heliksit ja β-rakenteet) tuhoutumista, analogisesti kiteen sulamisen kanssa, kutsutaan polypeptidien "sulatukseksi". Tässä tapauksessa vetysidokset katkeavat ja polypeptidiketjut muodostuvat satunnaisen vyyhdin muodossa. Siten sekundäärirakenteiden stabiilisuus määräytyy peptidien välisten vetysidosten avulla. Muuntyyppiset sidokset eivät osallistu tähän juuri lainkaan, lukuun ottamatta disulfidisidoksia polypeptidiketjussa kysteiinitähteiden kohdissa. Lyhyet peptidit sulkeutuvat sykleihin disulfidisidosten vuoksi. Monet proteiinit sisältävät sekä α-kierteisiä alueita että β-rakenteita. 100-prosenttisesti α-kierteestä koostuvia luonnollisia proteiineja ei juurikaan ole (poikkeus on paramyosiini, lihasproteiini, joka on 96-100-prosenttisesti α-kierre), kun taas synteettisissä polypeptideissä on 100-prosenttinen heliksi.
Muilla proteiineilla on vaihteleva kiertymisaste. Paramyosiinissa, myoglobiinissa ja hemoglobiinissa havaitaan suuri taajuus a-kierteisiä rakenteita. Sitä vastoin trypsiinissä, ribonukleaasissa, merkittävä osa polypeptidiketjusta on laskostunut kerrosrakenteisiksi p-rakenteiksi. Tukikudosten proteiinit: keratiinilla (hiusten, villan proteiinilla), kollageenilla (jänteiden, ihon proteiinilla), fibroiinilla (luonnonsilkin proteiinilla) on polypeptidiketjujen β-konfiguraatio. Proteiinien polypeptidiketjujen erilaiset helisiteetin asteet osoittavat, että ilmeisesti on olemassa voimia, jotka osittain häiritsevät helisitettä tai "katkaisevat" polypeptidiketjun säännöllisen laskostumisen. Syynä tähän on proteiinipolypeptidiketjun tiiviimpi laskostuminen tietyssä tilavuudessa, eli tertiääriseen rakenteeseen.
Proteiinin tertiäärinen rakenne
Proteiinin tertiäärinen rakenne on tapa, jolla polypeptidiketju järjestetään avaruudessa. Tertiaarisen rakenteensa muodon perusteella proteiinit jaetaan pääasiassa pallomaisiin ja säikeisiin. Globulaarisilla proteiineilla on useimmiten ellipsoidimuoto, ja säikeisillä (langan kaltaisilla) proteiineilla on pitkänomainen muoto (sauvan tai karan muoto).
Proteiinien tertiaarisen rakenteen konfiguraatio ei kuitenkaan vielä anna aihetta olettaa, että fibrillaarisilla proteiineilla on vain β-rakenne ja globulaarisilla proteiineilla α-kierteinen rakenne. On fibrillaarisia proteiineja, joilla on kierteinen, ei kerroksellinen, taitettu sekundaarinen rakenne. Esimerkiksi α-keratiini ja paramyosiini (nilviäisten sulkijalihaksen proteiini), tropomyosiinit (luurankolihasten proteiinit) kuuluvat fibrillaarisiin proteiineihin (on sauvan muotoinen), ja niiden toissijainen rakenne on α-heliksi; sitä vastoin pallomaiset proteiinit voivat sisältää suuren määrän p-rakenteita.
Lineaarisen polypeptidiketjun spiralisoituminen pienentää sen kokoa noin 4 kertaa; ja pakkaaminen tertiääriseen rakenteeseen tekee siitä kymmeniä kertoja kompaktimman kuin alkuperäinen ketju.
Sidokset, jotka stabiloivat proteiinin tertiaarista rakennetta. Aminohappojen sivuradikaalien välisillä sidoksilla on rooli tertiaarisen rakenteen stabiloinnissa. Nämä liitännät voidaan jakaa:
vahva (kovalenttinen) [näytä].
heikko (polaarinen ja van der Waals) [näytä].
Lukuisat sidokset aminohappojen sivuradikaalien välillä määräävät proteiinimolekyylin avaruudellisen konfiguraation.
Proteiinin tertiaarisen rakenteen organisoinnin piirteet. Polypeptidiketjun tertiaarisen rakenteen konformaation määräävät siihen sisältyvien aminohappojen sivuradikaalien ominaisuudet (joilla ei ole havaittavissa olevaa vaikutusta primääri- ja sekundaaristen rakenteiden muodostumiseen) ja mikroympäristöstä, ts. ympäristöön. Poimutettuna proteiinin polypeptidiketjulla on taipumus saada energeettisesti suotuisa muoto, jolle on ominaista vähimmäismäärä vapaata energiaa. Siksi ei-polaariset R-ryhmät, "välten" vettä, muodostavat ikään kuin proteiinin tertiaarisen rakenteen sisäisen osan, jossa suurin osa polypeptidiketjun hydrofobisista tähteistä sijaitsee. Proteiinipallon keskellä ei ole juuri lainkaan vesimolekyylejä. Aminohapon polaariset (hydrofiiliset) R-ryhmät sijaitsevat tämän hydrofobisen ytimen ulkopuolella ja niitä ympäröivät vesimolekyylit. Polypeptidiketju on monimutkaisesti taivutettu kolmiulotteisessa tilassa. Kun se taipuu, sekundaarinen kierteinen konformaatio häiriintyy. Ketju "katkaisee" heikoista kohdista, joissa proliini tai hydroksiproliini sijaitsevat, koska nämä aminohapot ovat liikkuvampia ketjussa muodostaen vain yhden vetysidoksen muiden peptidiryhmien kanssa. Toinen taivutuskohta on glysiini, jossa on pieni R-ryhmä (vety). Siksi muiden aminohappojen R-ryhmät pinottuna pyrkivät miehittämään vapaan tilan glysiinin sijainnissa. Useat aminohapot - alaniini, leusiini, glutamaatti, histidiini - edistävät stabiilien kierukkarakenteiden säilymistä proteiinissa, ja kuten metioniini, valiini, isoleusiini ja asparagiinihappo edistävät β-rakenteiden muodostumista. Tertiäärisen konfiguraation omaavassa proteiinimolekyylissä on alueita α-heliksien (kierteisen), β-rakenteiden (kerroksisen) ja satunnaisen kierteen muodossa. Vain proteiinin oikea avaruudellinen laskostuminen tekee siitä aktiivisen; sen rikkominen johtaa muutoksiin proteiinin ominaisuuksissa ja biologisen aktiivisuuden menettämiseen.
Kvaternäärinen proteiinirakenne
Yhdestä polypeptidiketjusta koostuvilla proteiineilla on vain tertiäärinen rakenne. Näitä ovat myoglobiini - lihaskudosproteiini, joka osallistuu hapen sitomiseen, useita entsyymejä (lysotsyymi, pepsiini, trypsiini jne.). Jotkut proteiinit on kuitenkin rakennettu useista polypeptidiketjuista, joista jokaisella on tertiäärinen rakenne. Tällaisille proteiineille on otettu käyttöön kvaternaarisen rakenteen käsite, joka tarkoittaa useiden tertiäärisen rakenteen omaavien polypeptidiketjujen järjestämistä yhdeksi toiminnalliseksi proteiinimolekyyliksi. Tällaista kvaternaarisen rakenteen omaavaa proteiinia kutsutaan oligomeeriksi ja sen tertiäärisen rakenteen omaavia polypeptidiketjuja kutsutaan protomeereiksi tai alayksiköiksi (kuvio 4).

Organisaation kvaternäärisellä tasolla proteiinit säilyttävät tertiäärisen rakenteen peruskonfiguraation (pallomainen tai fibrillaarinen). Esimerkiksi hemoglobiini on proteiini, jolla on kvaternäärinen rakenne ja joka koostuu neljästä alayksiköstä. Jokainen alayksikkö on pallomainen proteiini, ja yleensä hemoglobiinilla on myös pallomainen konfiguraatio. Hius- ja villaproteiinit - keratiinit, jotka liittyvät tertiäärisessä rakenteessa säikeisiin proteiineihin, ovat fibrillaarista konformaatiota ja kvaternääristä rakennetta.
Proteiinin kvaternaarisen rakenteen stabilointi. Kaikki proteiinit, joilla on kvaternäärinen rakenne, eristetään yksittäisten makromolekyylien muodossa, jotka eivät hajoa alayksiköiksi. Kosketukset alayksiköiden pintojen välillä ovat mahdollisia vain aminohappotähteiden polaaristen ryhmien ansiosta, koska kunkin polypeptidiketjun tertiaarisen rakenteen muodostumisen aikana ei-polaaristen aminohappojen sivuradikaalit (jotka muodostavat suurimman osan kaikki proteinogeeniset aminohapot) ovat piilossa alayksikön sisällä. Polaaristen ryhmiensä välille muodostuu lukuisia ionisia (suola-), vety- ja joissakin tapauksissa disulfidisidoksia, jotka pitävät alayksiköt lujasti organisoidun kompleksin muodossa. Vetysidoksia rikkovien tai disulfidisiltoja vähentävien aineiden käyttö aiheuttaa protomeerien hajoamista ja proteiinin kvaternaarisen rakenteen tuhoutumista. Taulukossa Kuvassa 1 on yhteenveto tiedoista sidoksista, jotka stabiloivat proteiinimolekyylin eri organisoitumistasoja [näytä].
Joidenkin fibrillaaristen proteiinien rakenteellisen organisaation piirteet
Fibrillaaristen proteiinien rakenteellisella organisaatiolla on useita piirteitä verrattuna pallomaisiin proteiineihin. Nämä ominaisuudet voidaan nähdä esimerkiksi keratiinista, fibroiinista ja kollageenista. Keratiinit esiintyvät α- ja β-konformaatioina. α-Keratiineilla ja fibroiinilla on kerros-laskostettu sekundaarinen rakenne, mutta keratiinissa ketjut ovat yhdensuuntaisia ja fibroiinissa ne ovat vastasuuntaisia (katso kuva 3); Lisäksi keratiini sisältää ketjujen välisiä disulfidisidoksia, kun taas fibroiinissa niitä ei ole. Disulfidisidosten katkeaminen johtaa keratiinien polypeptidiketjujen erottumiseen. Päinvastoin, maksimimäärän disulfidisidosten muodostuminen keratiineihin altistuessaan hapettimille luo vahvan tilarakenteen. Yleensä fibrillaarisissa proteiineissa, toisin kuin pallomaisissa proteiineissa, on joskus vaikea tehdä tiukkaa eroa organisaation eri tasojen välillä. Jos hyväksymme (kuten globulaarisen proteiinin kohdalla), että tertiäärinen rakenne muodostuu asettamalla yksi polypeptidiketju avaruuteen ja kvaternäärinen rakenne useilla ketjuilla, niin fibrillaarisissa proteiineissa on mukana useita polypeptidiketjuja jo sekundaarirakenteen muodostumisen aikana. . Tyypillinen esimerkki fibrillaarisesta proteiinista on kollageeni, joka on yksi ihmiskehon runsaimmista proteiineista (noin 1/3 kaikkien proteiinien massasta). Sitä löytyy kudoksista, joilla on suuri lujuus ja alhainen venyvyys (luut, jänteet, iho, hampaat jne.). Kollageenissa kolmasosa aminohappotähteistä on glysiiniä ja noin neljäsosa tai hieman enemmän proliinia tai hydroksiproliinia.

Kollageenin eristetty polypeptidiketju (primäärirakenne) näyttää katkoviivalta. Se sisältää noin 1000 aminohappoa ja sen molekyylipaino on noin 105 (kuvio 5, a, b). Polypeptidiketju rakennetaan toistuvasta aminohappokolmiosta (tripletti), jonka koostumus on seuraava: gly-A-B, jossa A ja B ovat mitä tahansa muita aminohappoja kuin glysiiniä (useimmiten proliinia ja hydroksiproliinia). Kollageenipolypeptidiketjut (tai a-ketjut) sekundaaristen ja tertiääristen rakenteiden muodostumisen aikana (kuvio 5, c ja d) eivät voi tuottaa tyypillisiä a-heliksejä, joilla on kierteinen symmetria. Proliini, hydroksiproliini ja glysiini (antihelikaaliset aminohapot) häiritsevät tätä. Siksi kolme α-ketjua muodostavat ikään kuin kierrettyjä spiraaleja, kuten kolme lankaa, jotka kiertyvät sylinterin ympärille. Kolme kierteistä a-ketjua muodostavat toistuvan kollageenirakenteen, jota kutsutaan tropokollageeniksi (kuva 5d). Tropokollageeni on organisaatiossaan kollageenin tertiäärinen rakenne. Proliinin ja hydroksiproliinin litteät renkaat, jotka vuorottelevat säännöllisesti pitkin ketjua, antavat sille jäykkyyttä, samoin kuin ketjujen väliset sidokset tropokollageenin α-ketjujen välillä (siksi kollageeni kestää venymistä). Tropokollageeni on pohjimmiltaan kollageenifibrillien alayksikkö. Tropokollageenialayksiköiden asettuminen kollageenin kvaternaariseen rakenteeseen tapahtuu vaiheittain (kuva 5e).
Kollageenirakenteiden stabiloituminen tapahtuu ketjujen välisten vety-, ioni- ja van der Waalsin sidosten sekä pienen määrän kovalenttisten sidosten ansiosta.
Kollageenin α-ketjuilla on erilaiset kemialliset rakenteet. On olemassa erilaisia α1-ketjuja (I, II, III, IV) ja α2-ketjuja. Riippuen siitä, mitkä α 1 - ja α 2 -ketjut osallistuvat tropokollageenin kolmijuosteisen heliksin muodostumiseen, kollageenia erotetaan neljää tyyppiä:
ensimmäinen tyyppi - kaksi a1 (I) ja yksi a2-ketju;
toinen tyyppi - kolme a1(II)-ketjua;
kolmas tyyppi - kolme a1(III)-ketjua;
neljäs tyyppi - kolme α1 (IV) -ketjua.
Yleisin kollageeni on ensimmäinen tyyppi: sitä löytyy luukudoksesta, ihosta, jänteistä; Toisen tyypin kollageenia löytyy rustokudoksesta jne. Yhdessä kudostyypissä voi olla erityyppistä kollageenia.
Kollageenirakenteiden järjestetty aggregaatio, niiden jäykkyys ja inertisyys takaavat kollageenikuitujen korkean lujuuden. Kollageeniproteiinit sisältävät myös hiilihydraattikomponentteja, eli ne ovat proteiini-hiilihydraattikomplekseja.
Kollageeni on solunulkoinen proteiini, jota muodostavat kaikissa elimissä esiintyvät sidekudossolut. Siksi kollageenin vaurioituessa (tai sen muodostumisen häiriintymisessä) esiintyy useita elinten sidekudoksen tukitoimintojen rikkomuksia.
Alfa-polypeptidiketju päättyy aminohappoyhdistelmään valiini-leusiini ja beeta-polypeptidiketju päättyy valiini-histidiini-leusiini-yhdistelmään. Alfa- ja beeta-polypeptidiketjut hemoglobiinimolekyylissä eivät ole järjestetty lineaarisesti, tämä on ensisijainen rakenne. Molekyylisisäisten voimien olemassaolon vuoksi polypeptidiketjut kiertyvät proteiineille tyypillisen alfa-heliksikierteen (sekundaarirakenteen) muodossa. Alfa-heliksikierre itse taipuu avaruudellisesti kunkin alfa- ja beeta-polypeptidiketjun kohdalla muodostaen munanmuotoisia plexuksia (tertiäärinen rakenne). Polypeptidiketjujen alfa-heliksikierteiden yksittäiset osat on merkitty latinalaisilla kirjaimilla A:sta H:hen. Kaikki neljä tertiääristä kaarevaa alfa- ja beeta-polypeptidiketjua sijaitsevat avaruudellisesti tietyssä suhteessa - kvaternaarisessa rakenteessa. Niitä eivät yhdistä todelliset kemialliset sidokset, vaan molekyylien väliset voimat.
Kävi ilmi, että ihmisillä on kolme päätyyppiä normaalia hemoglobiinia: alkion - U, sikiön - F ja aikuisen hemoglobiini - A. HbU (nimetty sanan kohtu alkukirjaimen mukaan) esiintyy alkiossa 7-12 elinviikon välillä. sitten se katoaa ja ilmestyy sikiön hemoglobiini, joka kolmannen kuukauden jälkeen on sikiön tärkein hemoglobiini. Tämän jälkeen tavallinen aikuisen hemoglobiini ilmestyy vähitellen, nimeltään HbA, englannin sanan "adult" alkukirjaimen jälkeen. Sikiön hemoglobiinin määrä vähenee vähitellen niin, että syntymähetkellä hemoglobiinista 80 % on HbA:ta ja vain 20 % HbF:ää. Synnytyksen jälkeen sikiön hemoglobiini jatkaa laskuaan ja 2–3 elinvuoden jälkeen se on enää 1–2 %. Sama määrä sikiön hemoglobiinia on aikuisella. Yli 2 % HbF:n määrää pidetään patologisena aikuisella ja yli 3-vuotiailla lapsilla.
Normaalien hemoglobiinityyppien lisäksi tunnetaan tällä hetkellä yli 50 patologista muunnelmaa. Ne nimettiin ensin latinalaisilla kirjaimilla. B-kirjain puuttuu hemoglobiinityyppien nimeämisestä, koska se merkitsi alun perin HbS:ää.
Hemoglobiini (Hb)- punasoluissa oleva kromoproteiini, joka osallistuu hapen kuljettamiseen kudoksiin. Aikuisten hemoglobiinia kutsutaan hemoglobiini A:ksi (Hb A). Sen molekyylipaino on noin 65 000 Da. Hb A -molekyylillä on kvaternäärinen rakenne ja se sisältää neljä alayksikköä - polypeptidiketjut (nimetty α1, α2, β1 ja β2, joista jokainen liittyy hemiin.
Muista, että hemoglobiini on allosteerinen proteiini, jonka molekyylit voivat muuttua palautuvasti yhdestä konformaatiosta toiseen. Tämä muuttaa proteiinin affiniteettia ligandeihin. Konformaatiota, jolla on vähiten affiniteetti ligandiin, kutsutaan jännittyneeksi tai T-konformaatioksi. Konformaatiota, jolla on suurin affiniteetti ligandiin, kutsutaan relaksoiduksi tai R-konformaatioksi.
Hemoglobiinimolekyylin R- ja T-konformaatiot ovat dynaamisen tasapainon tilassa:
Erilaiset ympäristötekijät voivat muuttaa tätä tasapainoa suuntaan tai toiseen. Allosteeriset säätelijät, jotka vaikuttavat Hb:n affiniteettiin O2:n suhteen, ovat: 1) happi; 2) H+-pitoisuus (keskimääräinen pH); 3) hiilidioksidi (CO2); 4) 2,3-difosfoglyseraatti (DPG). Happimolekyylin kiinnittyminen johonkin hemoglobiinialayksiköstä edistää jännittyneen konformaation siirtymistä relaksaatioon ja lisää saman hemoglobiinimolekyylin muiden alayksiköiden affiniteettia happea kohtaan. Tätä ilmiötä kutsutaan yhteistyövaikutukseksi. Hemoglobiinin happeen sitoutumisen monimutkainen luonne heijastuu hemoglobiini O2:n saturaatiokäyrästä, jolla on S-muotoinen (kuva 3.1).

Kuva 3.1. Myoglobiinin (1) ja hemoglobiinin (2) happisaturaatiokäyrät.
Molekyylimuodot hemoglobiini eroaa toisistaan polypeptidiketjujen rakenteessa. Esimerkki tällaisesta fysiologisissa olosuhteissa esiintyvästä hemoglobiinin vaihtelusta on sikiön hemoglobiini (HbF), joita esiintyy veressä ihmisen alkiovaiheessa. Toisin kuin HbA, sen molekyyli sisältää 2 α- ja 2 y-ketjua (eli β-ketjut on korvattu y-ketjuilla). Tällaisella hemoglobiinilla on suurempi affiniteetti happea kohtaan. Tämän ansiosta alkio voi vastaanottaa happea äidin verestä istukan kautta. Pian syntymän jälkeen vauvan veressä oleva HbF korvataan HbA:lla.
Esimerkki epänormaalista tai patologisesta hemoglobiinista on jo mainittu (katso 2.4.) hemoglobiini S, joka löytyy potilailta, joilla on sirppisoluanemia. Kuten jo tiedät, se eroaa hemoglobiini A:sta korvaamalla glutamaatin valiinilla β-ketjuissa. Tämä aminohapposubstituutio vähentää HbS:n liukoisuutta veteen ja vähentää sen affiniteettia 02:een.
Proteiinien kemiallista rakennetta edustavat alfa-aminohapot, jotka on liitetty ketjuun peptidisidoksella. Elävissä organismeissa koostumus määräytyy geneettisen koodin mukaan. Synteesiprosessissa käytetään useimmissa tapauksissa 20 standardityypin aminohappoa. Niiden monet yhdistelmät muodostavat proteiinimolekyylejä, joilla on laaja valikoima ominaisuuksia. Aminohappotähteet ovat usein alttiina translaation jälkeisille modifikaatioille. Ne voivat syntyä ennen kuin proteiini alkaa suorittaa tehtävänsä ja sen toiminnan aikana solussa. Elävissä organismeissa useat molekyylit muodostavat usein monimutkaisia komplekseja. Esimerkki on fotosynteettinen assosiaatio.
Yhteyksien tarkoitus
Proteiineja pidetään tärkeänä osana ihmisten ja eläinten ravintoa, koska niiden elimistö ei pysty syntetisoimaan kaikkia tarvittavia aminohappoja. Joidenkin niistä pitäisi tulla proteiiniruokaa. Tärkeimmät yhdisteiden lähteet ovat liha, pähkinät, maito, kala ja viljat. Vähäisemmässä määrin proteiineja on vihanneksissa, sienissä ja marjoissa. Entsyymien kautta tapahtuvan ruoansulatuksen aikana kulutetut proteiinit hajoavat aminohapoiksi. Niitä käytetään jo omien proteiiniensa biosynteesissä kehossa tai ne hajoavat edelleen energian saamiseksi.

Historiallinen viittaus
Frederij Senger määritti ensin insuliiniproteiinirakenteen sekvenssin. Työstään hän sai Nobel-palkinnon vuonna 1958. Sanger käytti sekvensointimenetelmää. Röntgendiffraktiota käyttämällä saatiin myöhemmin (1950-luvun lopulla) myoglobiinin ja hemoglobiinin kolmiulotteiset rakenteet. Työn suorittivat John Kendrew ja Max Perutz.
Proteiinimolekyylin rakenne
Se sisältää lineaarisia polymeerejä. Ne puolestaan koostuvat alfa-aminohappotähteistä, jotka ovat monomeerejä. Lisäksi proteiinirakenne voi sisältää ei-aminohappoluonteisia komponentteja ja modifioituja aminohappotähteitä. Komponentteja määritettäessä käytetään 1- tai 3-kirjaimia lyhenteitä. Yhdistettä, joka sisältää kahdesta useaan tusinaan jäännöksiä, kutsutaan usein "polypeptidiksi". Yhden aminohapon alfa-karboksyyliryhmän vuorovaikutuksen seurauksena toisen alfa-aminoryhmän kanssa syntyy sidoksia (proteiinirakenteen muodostumisen aikana). Yhdisteen C- ja N-terminaaliset päät erotetaan sen mukaan, mikä aminohappotähteen ryhmä on vapaa: -COOH tai -NH2. Proteiinisynteesin prosessissa ribosomissa ensimmäinen terminaalinen tähde on yleensä metioniinitähde; seuraavat on kiinnitetty edellisten C-päähän.

Organisaation tasot
Niitä ehdotti Lindrem-Lang. Huolimatta siitä, että tätä jakoa pidetään jonkin verran vanhentuneena, sitä käytetään edelleen. Yhteysorganisaatiossa ehdotettiin erottavan neljä tasoa. Proteiinimolekyylin primäärirakenne määräytyy geenin geneettisen koodin ja ominaisuuksien mukaan. Korkeammille tasoille on ominaista muodostuminen proteiinin laskostumisen aikana. Proteiinin avaruudellinen rakenne määräytyy kokonaisuutena aminohappoketjun mukaan. Siitä huolimatta se on melko labiili. Siihen voivat vaikuttaa ulkoiset tekijät. Tässä suhteessa on oikein puhua yhdisteen konformaatiosta, joka on edullisin ja energeettisesti parempi.
Taso 1
Sitä edustaa polypeptidiketjun aminohappotähteiden sekvenssi. Pääsääntöisesti se kuvataan yksi- tai kolmikirjaimilla merkinnöillä. Proteiinien primaariselle rakenteelle on tunnusomaista stabiilit aminohappotähteiden yhdistelmät. He suorittavat tiettyjä tehtäviä. Tällaiset "konservatiiviset motiivit" säilyvät lajin evoluution aikana. Niitä voidaan usein käyttää ennustamaan tuntemattoman proteiinin ongelma. Arvioimalla eri organismien aminohappoketjujen samankaltaisuuden astetta (homologiaa) on mahdollista määrittää näiden organismien muodostavien taksonien välinen evoluutionaalinen etäisyys. Proteiinien primäärirakenne määritetään sekvensoimalla tai sen mRNA:n alkuperäisellä kompleksilla käyttämällä geneettistä kooditaulukkoa.

Paikallinen ketjuosan tilaus
Tämä on seuraava organisaatiotaso - proteiinien toissijainen rakenne. Sitä on useita tyyppejä. Polypeptidiketjun osan paikallinen järjestys stabiloidaan vetysidoksilla. Suosituimmat tyypit ovat:

Tilarakenne
Proteiinien tertiäärinen rakenne sisältää edellisen tason elementtejä. Ne vakautuvat erilaisilla vuorovaikutuksilla. Hydrofobiset sidokset ovat äärimmäisen tärkeitä. Stabilointi sisältää:
- Kovalenttiset vuorovaikutukset.
- Ionisidokset muodostuvat aminohapposivuryhmien välille, joilla on vastakkaiset varaukset.
- Vedyn vuorovaikutukset.
- Hydrofobiset sidokset. Vuorovaikutuksessa ympäröivien elementtien H20 kanssa proteiini laskostuu siten, että sivupuoliset ei-polaariset aminohapporyhmät eristetään vesiliuoksesta. Hydrofiilisiä ryhmiä (polaarisia) ilmestyy molekyylin pinnalle.
Proteiinien tertiäärinen rakenne määritetään magneetti- (ydin)resonanssimenetelmillä, tietyntyyppisillä mikroskopialla ja muilla menetelmillä.
Asennusperiaate
Tutkimus on osoittanut, että on kätevää tunnistaa vielä yksi taso tasojen 2 ja 3 väliltä. Sitä kutsutaan "arkkitehtuuriksi", "asennusaiheeksi". Se määräytyy sekundaarirakenteen komponenttien (beeta-säikeiden ja alfaheliksien) suhteellisesta sijainnista kompaktin globulin - proteiinidomeenin - rajoissa. Se voi esiintyä itsenäisesti tai sisällyttää suurempaan proteiiniin yhdessä muiden samankaltaisten proteiinien kanssa. On todettu, että tyylin motiivit ovat varsin konservatiivisia. Niitä löytyy proteiineista, joilla ei ole evoluutio- tai toiminnallisia suhteita. Arkkitehtuurin määritelmä on rationaalisen (fyysisen) luokittelun perusta.

Verkkotunnuksen organisaatio
Useiden polypeptidiketjujen keskinäisen järjestelyn avulla yhdessä proteiinikompleksissa muodostuu proteiinien kvaternäärinen rakenne. Sen muodostavat alkuaineet muodostuvat erikseen ribosomeihin. Tämä proteiinirakenne alkaa muodostua vasta synteesin päätyttyä. Se voi sisältää sekä erilaisia että identtisiä polypeptidiketjuja. Proteiinien kvaternäärinen rakenne on stabiloitunut samojen vuorovaikutusten ansiosta kuin edellisellä tasolla. Jotkut kompleksit voivat sisältää useita kymmeniä proteiineja.
Proteiinin rakenne: suojatehtävät
Sytoskeleton polypeptidit, jotka toimivat jollain tavalla vahvistuksena, antavat monille organelleille muodon ja osallistuvat sen muutokseen. Rakenteelliset proteiinit suojaavat kehoa. Esimerkiksi kollageeni on tällainen proteiini. Se muodostaa perustan sidekudosten solujen väliselle aineelle. Keratiinilla on myös suojaava tehtävä. Se muodostaa sarvien, höyhenten, hiusten ja muiden orvaskeden johdannaisten perustan. Kun proteiinit sitovat myrkkyjä, tapahtuu monissa tapauksissa vieroitus. Näin kehon kemiallisen suojan tehtävä suoritetaan. Maksaentsyymeillä on erityisen tärkeä rooli myrkkyjen neutralointiprosessissa ihmiskehossa. Ne pystyvät hajottamaan myrkkyjä tai muuttamaan ne liukoiseen muotoon. Tämä helpottaa nopeampaa kuljetusta kehosta. Veressä ja muissa kehon nesteissä olevat proteiinit tarjoavat immuunipuolustusta laukaisemalla vasteen sekä taudinaiheuttajahyökkäykselle että vauriolle. Immunoglobuliinit (vasta-aineet ja komplementtijärjestelmän komponentit) pystyvät neutraloimaan bakteereja, vieraita proteiineja ja viruksia.
Sääntelymekanismi
Proteiinimolekyylit, jotka eivät toimi energialähteenä eivätkä rakennusmateriaalina, ohjaavat monia solunsisäisiä prosesseja. Siten niiden ansiosta translaatiota, transkriptiota, viipalointia ja muiden polypeptidien aktiivisuutta säädellään. Säätelymekanismi perustuu entsymaattiseen aktiivisuuteen tai ilmenee spesifisen sitoutumisen seurauksena muihin molekyyleihin. Esimerkiksi transkriptiotekijät, aktivaattoripolypeptidit ja repressoriproteiinit pystyvät säätelemään geenin transkription intensiteettiä. Näin tehdessään ne ovat vuorovaikutuksessa geenisäätelysekvenssien kanssa. Tärkein rooli solunsisäisten prosessien kulun säätelyssä on proteiinifosfataaseille ja proteiinikinaaseille. Nämä entsyymit laukaisevat tai estävät muiden proteiinien toimintaa lisäämällä tai poistamalla niistä fosfaattiryhmiä.

Signaalitehtävä
Se yhdistetään usein säätelytoimintoon. Tämä johtuu siitä tosiasiasta, että monet solunsisäiset ja solunulkoiset polypeptidit voivat välittää signaaleja. Kasvutekijöillä, sytokiineilla, hormoneilla ja muilla yhdisteillä on tämä kyky. Steroidit kulkeutuvat veren kautta. Hormonin vuorovaikutus reseptorin kanssa toimii signaalina, joka laukaisee soluvasteen. Steroidit säätelevät yhdisteiden pitoisuutta veressä ja soluissa, lisääntymistä, kasvua ja muita prosesseja. Esimerkki on insuliini. Se säätelee glukoositasoja. Solujen vuorovaikutus tapahtuu signaaliproteiiniyhdisteiden kautta, jotka välittyvät solujen välisen aineen kautta.
Elementtien kuljetus
Pienten molekyylien liikkumiseen osallistuvilla liukoisilla proteiineilla on korkea affiniteetti substraattiin, jota on läsnä lisääntyneenä pitoisuutena. Heillä on myös kyky vapauttaa se helposti alueilla, joilla sen pitoisuus on alhainen. Esimerkki on kuljetusproteiini hemoglobiini. Se siirtää happea keuhkoista muihin kudoksiin ja siirtää niistä hiilidioksidia. Jotkut kalvoproteiinit osallistuvat myös pienten molekyylien kuljettamiseen soluseinien läpi ja muuttavat niitä. Sytoplasman lipidikerros on vedenpitävä. Tämä estää varautuneiden tai polaaristen molekyylien diffuusion. Kalvokuljetusyhteydet jaetaan yleensä kantajien ja kanavien välillä.
Varaliitännät
Nämä proteiinit muodostavat niin sanottuja varantoja. Niitä kertyy esimerkiksi kasvien siemeniin ja eläinten muniin. Tällaiset proteiinit toimivat aineen ja energian varalähteenä. Joitakin yhdisteitä elimistö käyttää aminohappovarastona. Ne puolestaan ovat aineenvaihdunnan säätelyyn osallistuvien vaikuttavien aineiden esiasteita.
Solujen reseptorit
Tällaiset proteiinit voivat sijaita joko suoraan sytoplasmassa tai upotettuna seinämään. Yksi yhteyden osa vastaanottaa signaalin. Yleensä se on kemiallinen aine, ja joissakin tapauksissa mekaaninen vaikutus (esimerkiksi venyttely), valo ja muut ärsykkeet. Signaalin altistumisprosessissa tietylle molekyylin fragmentille - polypeptidireseptorille - sen konformaatiomuutokset alkavat. Ne aiheuttavat muutoksen muun osan konformaatiossa, joka välittää ärsykkeen solun muihin osiin. Signaalin lähettäminen voidaan tehdä eri tavoin. Jotkut reseptorit pystyvät katalysoimaan kemiallista reaktiota, kun taas toiset toimivat ionikanavina, jotka sulkeutuvat tai avautuvat ärsykkeen vaikutuksesta. Jotkut yhdisteet sitovat spesifisesti lähettimolekyylejä solussa.
Motoriset polypeptidit
On olemassa kokonainen luokka proteiineja, jotka tarjoavat keholle liikettä. Motoriset proteiinit osallistuvat lihasten supistumiseen, solujen liikkeisiin sekä siimojen ja värekärpien toimintaan. Ne tarjoavat myös suunnatun ja aktiivisen kuljetuksen. Kinesiinit ja dyneiinit kuljettavat molekyylejä mikrotubuluksia pitkin käyttämällä ATP-hydrolyysiä energialähteenä. Jälkimmäiset siirtävät organelleja ja muita elementtejä kohti senrosomia perifeerisiltä solualueilta. Kinesiinit liikkuvat vastakkaiseen suuntaan. Dyneiinit ovat vastuussa myös siima- ja värekärkeiden toiminnasta.
