მემბრანები ასრულებენ სხვადასხვა ფუნქციების დიდ რაოდენობას:
გარსები განსაზღვრავს ორგანელის ან უჯრედის ფორმას;
ბარიერი: აკონტროლებს ხსნადი ნივთიერებების (მაგალითად, იონების Na +, K +, Cl-) გაცვლას შიდა და გარე განყოფილებას შორის;
ენერგია: ატფ-ის სინთეზი მიტოქონდრიების შიდა მემბრანებზე და ფოტოსინთეზი ქლოროპლასტის მემბრანებში; ქიმიური რეაქციების წარმოქმნის ზედაპირს (ფოსფორილირება მიტოქონდრიულ გარსებზე);
არის სტრუქტურა, რომელიც უზრუნველყოფს ქიმიური სიგნალების ამოცნობას (მემბრანაზე განლაგებულია ჰორმონის და ნეიროტრანსმიტერული რეცეპტორები);
თამაშობენ როლს უჯრედშორის ურთიერთქმედებაში და ხელს უწყობენ უჯრედების მოძრაობას.
ტრანსპორტირება მემბრანის გასწვრივ. მემბრანას აქვს შერჩევითი გამტარიანობა ხსნადი ნივთიერებებისთვის, რაც აუცილებელია:
უჯრედის გამოყოფა უჯრედგარე გარემოდან;
უჯრედში შეღწევისა და მასში აუცილებელი მოლეკულების (როგორიცაა ლიპიდები, გლუკოზა და ამინომჟავები) შეკავების უზრუნველყოფა, აგრეთვე მეტაბოლური პროდუქტების (მათ შორის არასაჭირო) უჯრედიდან მოცილება;
ტრანსმემბრანული იონის გრადიენტის შენარჩუნება.
უჯრედშიდა ორგანელებს ასევე შეიძლება ჰქონდეთ შერჩევით გამტარი მემბრანა. მაგალითად, ლიზოსომებში მემბრანა ინარჩუნებს წყალბადის იონების კონცენტრაციას (H +) 1000-10000-ჯერ მეტი ვიდრე ციტოზოლში.
მემბრანის გასწვრივ ტრანსპორტი შეიძლება იყოს პასიური, მსუბუქიან აქტიური.
პასიური ტრანსპორტიარის მოლეკულების ან იონების მოძრაობა კონცენტრაციის ან ელექტროქიმიური გრადიენტის გასწვრივ. ეს შეიძლება იყოს მარტივი დიფუზია, როგორც გაზების (მაგალითად, O 2 და CO 2) ან მარტივი მოლეკულების (ეთანოლი) შეღწევის პლაზმურ მემბრანაში. მარტივი დიფუზიის დროს უჯრედგარე სითხეში გახსნილი მცირე მოლეკულები თანმიმდევრულად იხსნება მემბრანაში და შემდეგ უჯრედშიდა სითხეში. ეს პროცესი არასპეციფიკურია, ხოლო მემბრანის მეშვეობით შეღწევის სიჩქარე განისაზღვრება მოლეკულის ჰიდროფობიურობის ხარისხით, ანუ მისი ცხიმის ხსნადობით. დიფუზიის სიჩქარე ლიპიდურ ორ შრეში პირდაპირპროპორციულია ჰიდროფობიურობის, ასევე ტრანსმემბრანული კონცენტრაციის გრადიენტის ან ელექტროქიმიური გრადიენტის მიმართ.
გაადვილებული დიფუზია არის მოლეკულების სწრაფი მოძრაობა მემბრანაზე მემბრანის სპეციფიკური ცილების მიერ, რომელსაც ეწოდება პერმეაზები. ეს პროცესი სპეციფიკურია, ის უფრო სწრაფად მიმდინარეობს, ვიდრე მარტივი დიფუზია, მაგრამ აქვს ტრანსპორტირების სიჩქარის ლიმიტი.
გაადვილებული დიფუზია, როგორც წესი, დამახასიათებელია წყალში ხსნადი ნივთიერებებისთვის. მემბრანული გადამტანების უმეტესობა (თუ არა ყველა) არის ცილა. გაადვილებული დიფუზიის დროს მატარებლების ფუნქციონირების სპეციფიკური მექანიზმი საკმარისად არ არის შესწავლილი. მათ შეუძლიათ, მაგალითად, უზრუნველყონ გადატანა მემბრანაში ბრუნვითი მოძრაობით. ახლახან გამოჩნდა ინფორმაცია, რომ გადამზიდავი ცილები, ტრანსპორტირებულ ნივთიერებასთან შეხებისას, ცვლის მათ კონფორმაციას, რის შედეგადაც მემბრანაში იხსნება თავისებური „კარიბჭეები“ ან არხები. ეს ცვლილებები ხდება ენერგიის გამო გამოთავისუფლებული, როდესაც ტრანსპორტირებადი ნივთიერება აკავშირებს ცილას. შესაძლებელია რელეს ტიპის გადაცემაც. ამ შემთხვევაში, თავად გადამზიდავი რჩება უმოძრაო და იონები მის გასწვრივ მიგრირებენ ერთი ჰიდროფილური ბმადან მეორეში.
ანტიბიოტიკი გრამიციდინი შეიძლება იყოს მოდელი ამ ტიპის მატარებლისთვის. მემბრანის ლიპიდურ შრეში მისი გრძელი ხაზოვანი მოლეკულა იღებს სპირალის ფორმას და ქმნის ჰიდროფილურ არხს, რომლის მეშვეობითაც K იონს შეუძლია გრადიენტის გასწვრივ მიგრაცია.
მოპოვებულია ექსპერიმენტული მტკიცებულება ბიოლოგიურ მემბრანებში ბუნებრივი არხების არსებობის შესახებ. სატრანსპორტო პროტეინებს ახასიათებთ მაღალი სპეციფიკა მემბრანის მეშვეობით ტრანსპორტირებულ ნივთიერებასთან მიმართებაში, რომელიც ჰგავს ფერმენტებს მრავალი თვისებით. ისინი ძალიან მგრძნობიარენი არიან pH-ის მიმართ, კონკურენტულად ინჰიბირებულია გადატანილი ნივთიერების სტრუქტურით მსგავსი ნაერთებით და არაკონკურენტულად - აგენტებით, რომლებიც ცვლიან ცილების სპეციფიკურ ფუნქციურ ჯგუფებს.
გაადვილებული დიფუზია განსხვავდება ჩვეულებრივისგან არა მხოლოდ სიჩქარით, არამედ გაჯერების უნარით. ნივთიერებების გადაცემის სიჩქარის ზრდა ხდება კონცენტრაციის გრადიენტის ზრდის პროპორციულად მხოლოდ გარკვეულ ზღვრამდე. ამ უკანასკნელს გადამზიდველის „ძალაუფლება“ განსაზღვრავს.
აქტიური ტრანსპორტი არის იონების ან მოლეკულების მოძრაობა მემბრანის გასწვრივ კონცენტრაციის გრადიენტის წინააღმდეგ ATP ჰიდროლიზის ენერგიის გამო. აქტიური იონის ტრანსპორტირების სამი ძირითადი ტიპი არსებობს:
ნატრიუმ-კალიუმის ტუმბო - Na + /K + -ადენოზინტრიფოსფატაზა (ATPase), რომელიც ატარებს Na + გარეთ და K + შიგნით;
კალციუმის (Ca 2+) ტუმბო - Ca 2+ -ATPase, რომელიც გადააქვს Ca 2+ უჯრედიდან ან ციტოზოლიდან სარკოპლაზმურ რეტიკულუმში;
პროტონული ტუმბო - H + -ATPase. აქტიური ტრანსპორტით შექმნილი იონური გრადიენტები შეიძლება გამოყენებულ იქნას სხვა მოლეკულების აქტიური ტრანსპორტირებისთვის, როგორიცაა გარკვეული ამინომჟავები და შაქარი (მეორადი აქტიური ტრანსპორტი).
კოტრანსპორტი- ეს არის იონის ან მოლეკულის ტრანსპორტი, სხვა იონის გადაცემასთან ერთად. სიმპორტი- ორივე მოლეკულის ერთდროული გადატანა ერთი მიმართულებით; ანტიპორტი- ორივე მოლეკულის ერთდროული გადატანა საპირისპირო მიმართულებით. თუ ტრანსპორტი არ არის დაკავშირებული სხვა იონის გადაცემასთან, ამ პროცესს ე.წ უნიპორტი. კოტრანსპორტი შესაძლებელია როგორც გაადვილებული დიფუზიით, ასევე აქტიური ტრანსპორტის პროცესში.
გლუკოზის ტრანსპორტირება შესაძლებელია გაადვილებული დიფუზიით სიმპორტის გზით. იონები Cl - და HCO 3 - ტრანსპორტირდება ერითროციტების მემბრანის მეშვეობით, გაადვილებული დიფუზიით გადამზიდველის მიერ, რომელსაც ეწოდება ზოლი 3, ანტიპორტის ტიპის მიხედვით. ამ შემთხვევაში, Cl - და HCO 3 - გადადის საპირისპირო მიმართულებით, ხოლო გადაცემის მიმართულება განისაზღვრება გაბატონებული კონცენტრაციის გრადიენტით.
იონების აქტიური ტრანსპორტირება კონცენტრაციის გრადიენტთან მიმართებაში მოითხოვს ატფ-ის ADP-მდე ჰიდროლიზის დროს გამოთავისუფლებულ ენერგიას: ATP ADP + F (არაორგანული ფოსფატი). აქტიური ტრანსპორტი, ისევე როგორც გაადვილებული დიფუზია, ხასიათდება: სპეციფიკურობით, მაქსიმალური სიჩქარის შეზღუდვით (ანუ კინეტიკური მრუდი აღწევს პლატოზე) და ინჰიბიტორების არსებობით. მაგალითად არის პირველადი აქტიური ტრანსპორტი, რომელსაც ახორციელებს Na + /K + - ATPase. ამ ანტიპორტის ფრაგმენტული სისტემის ფუნქციონირებისთვის საჭიროა Na +, K + და მაგნიუმის იონების არსებობა. ის გვხვდება თითქმის ყველა ცხოველურ უჯრედში და მისი კონცენტრაცია განსაკუთრებით მაღალია აგზნებად ქსოვილებში (მაგალითად, ნერვებში და კუნთებში) და უჯრედებში, რომლებიც აქტიურად მონაწილეობენ ნატრიუმის მოძრაობაში პლაზმური მემბრანის მეშვეობით (მაგალითად, თირკმელების კორტიკალური შრე და სანერწყვე ჯირკვლები).
ATPase ფერმენტი თავისთავად არის ოლიგომერი, რომელიც შედგება 2 ქვედანაყოფი 110 kDa და 2 გლიკოპროტეინის ქვედანაყოფი 55 kDa თითოეული. ფოსფორილირებას ესაჭიროება Na + და Mg 2+, მაგრამ არა K +, ხოლო დეფოსფორილირებისთვის საჭიროა K +, მაგრამ არა Na + ან Mg 2+. აღწერილია ცილოვანი კომპლექსის ორი კონფორმაციული მდგომარეობა სხვადასხვა ენერგეტიკული დონით, რომლებიც ჩვეულებრივ აღინიშნება როგორც E 1 და E 2, ამიტომ ATPase ასევე ე.წ. ტიპის E გადამზიდავი 1 - ე 2 . მაგალითად, გულის გლიკოზიდები დიგოქსინიდა უაბაინიაინჰიბირებს ატფ-აზას აქტივობას.წყალში კარგი ხსნადობის გამო, ouabain ფართოდ გამოიყენება ექსპერიმენტულ კვლევებში ნატრიუმის ტუმბოს შესასწავლად.
ზოგადად მიღებული იდეა Na + /K + - ATPase-ს მუშაობის შესახებ ასეთია. Na და ATP იონები მიმაგრებულია ATPase მოლეკულაზე Mg 2+ თანდასწრებით. Na იონების შეერთება იწვევს ATP ჰიდროლიზის რეაქციას, რაც იწვევს ADP-ს და ფერმენტის ფოსფორილირებულ ფორმას. ფოსფორილირება იწვევს ფერმენტული ცილის გადასვლას ახალ კონფორმაციულ მდგომარეობაში და Na-ის შემცველი ადგილი ან ადგილები აღმოჩნდება გარე გარემოსკენ. აქ Na + იცვლება K+-ზე, ვინაიდან ფერმენტის ფოსფორილირებულ ფორმას ახასიათებს მაღალი აფინურობა K იონების მიმართ. ფერმენტის საპირისპირო კონფორმაციაზე გადასვლა იწყება ფოსფორილის ჯგუფის ჰიდროლიზური დაშლით. არაორგანული ფოსფატი და თან ახლავს K+-ის გამოყოფა უჯრედის შიგნით. ფერმენტის დეფოსფორილირებულ აქტიურ ადგილს შეუძლია ახალი ATP მოლეკულის მიმაგრება და ციკლი მეორდება.
ტუმბოს მუშაობის შედეგად უჯრედში შემავალი K და Na იონების რაოდენობა ერთმანეთის ტოლი არ არის. სამი გამოყოფილი Na იონისთვის არის ორი შეყვანილი K იონი ერთი ATP მოლეკულის ერთდროული ჰიდროლიზით. მემბრანის მოპირდაპირე მხარეს არხის გახსნა და დახურვა და Na-სა და K-ის შეკავშირების ეფექტურობის მონაცვლეობითი ცვლილება უზრუნველყოფილია ATP ჰიდროლიზის ენერგიით. ტრანსპორტირებული იონები - Na და K - ამ ფერმენტული რეაქციის კოფაქტორები. თეორიულად, შესაძლებელია წარმოვიდგინოთ ტუმბოების მრავალფეროვნება, რომლებიც მუშაობენ ამ პრინციპით, თუმცა ამჟამად მხოლოდ რამდენიმე მათგანია ცნობილი.
გლუკოზის ტრანსპორტირება.გლუკოზის ტრანსპორტირება შეიძლება მოხდეს როგორც გაადვილებული დიფუზიის, ასევე აქტიური ტრანსპორტის სახით, პირველ შემთხვევაში ის მიმდინარეობს როგორც უნიპორტი, მეორეში - როგორც სიმპორტი. გლუკოზის ტრანსპორტირება შესაძლებელია ერითროციტებში გაადვილებული დიფუზიით. ერითროციტებში გლუკოზის ტრანსპორტირებისთვის მიქაელის მუდმივა (კმ) არის დაახლოებით 1,5 მმოლ/ლ (ანუ, გლუკოზის ამ კონცენტრაციით, ხელმისაწვდომი პერმეაზას მოლეკულების დაახლოებით 50% უკავშირდება გლუკოზის მოლეკულებს). ვინაიდან ადამიანის სისხლში გლუკოზის კონცენტრაცია არის 4-6 მმოლ/ლ, მისი შეწოვა ერითროციტების მიერ თითქმის მაქსიმალური სიჩქარით ხდება. პერმეაზას სპეციფიკა უკვე გამოიხატება იმაში, რომ L-იზომერი თითქმის არ ტრანსპორტირდება ერითროციტებში, განსხვავებით D-გალაქტოზასა და D-მანოზასგან, მაგრამ მათი უფრო მაღალი კონცენტრაციაა საჭირო სატრანსპორტო სისტემის ნახევრად გაჯერების მისაღწევად. უჯრედში შესვლის შემდეგ გლუკოზა განიცდის ფოსფორილირებას და ვეღარ ტოვებს უჯრედს. გლუკოზის პერმეაზას ასევე უწოდებენ D-ჰექსოზას. ეს არის მემბრანული ცილა, რომლის მოლეკულური წონაა 45 კდ.
გლუკოზის ტრანსპორტირება ასევე შესაძლებელია Na +-დამოკიდებული სიმპორტის სისტემით, რომელიც გვხვდება მრავალი ქსოვილის პლაზმურ მემბრანებში, მათ შორის თირკმელების მილაკებში და ნაწლავის ეპითელიუმში. ამ შემთხვევაში, ერთი გლუკოზის მოლეკულა ტრანსპორტირდება გაადვილებული დიფუზიით კონცენტრაციის გრადიენტის წინააღმდეგ, ხოლო ერთი Na-იონი ტრანსპორტირდება კონცენტრაციის გრადიენტის გასწვრივ. მთლიანი სისტემა საბოლოოდ ფუნქციონირებს Na + /K + - ATPase-ს სატუმბი ფუნქციით. ამრიგად, სიმპორტი არის მეორადი აქტიური სატრანსპორტო სისტემა. ამინომჟავების ტრანსპორტირება ხდება მსგავსი გზით.
Ca 2+ -ტუმბოარის E 1 - E 2 ტიპის აქტიური სატრანსპორტო სისტემა, რომელიც შედგება ინტეგრალური მემბრანული ცილისგან, რომელიც Ca 2+ გადაცემის პროცესში ფოსფორილირდება ასპარტატის ნარჩენზე. ATP-ის თითოეული მოლეკულის ჰიდროლიზის დროს გადადის ორი Ca 2+ იონი. ევკარიოტულ უჯრედებში Ca 2+-ს შეუძლია დაუკავშირდეს კალციუმის შემაკავშირებელ ცილას ე.წ კალმოდულინიდა მთელი კომპლექსი უკავშირდება Ca 2+ ტუმბოს. Ca 2+ შემაკავშირებელ ცილებს ასევე შეიცავს ტროპონინს C და პარვალბუმინს.
Ca იონები, ისევე როგორც Na იონები, აქტიურად გამოიყოფა უჯრედებიდან Ca 2+ -ATPase. ენდოპლაზმური ბადის გარსები შეიცავს განსაკუთრებით დიდი რაოდენობით კალციუმის ტუმბოს პროტეინს. ქიმიური რეაქციების ჯაჭვი, რომელიც იწვევს ATP ჰიდროლიზს და Ca 2+ გადაცემას, შეიძლება დაიწეროს შემდეგი განტოლებით:
2Ca n + ATP + E 1 Ca 2 - E - P + ADP
Ca 2 - E - P 2Ca ext + PO 4 3- + E 2
სად არის San - Ca2 +, მდებარეობს გარეთ;
Ca ext - Ca 2+ მდებარეობს შიგნით;
E 1 და E 2 - გადამზიდავი ფერმენტის სხვადასხვა კონფორმაციები, რომელთა გადასვლა ერთიდან მეორეზე ასოცირდება ATP ენერგიის გამოყენებასთან.
ციტოპლაზმიდან H+-ის აქტიური მოცილების სისტემას მხარს უჭერს ორი ტიპის რეაქცია: ელექტრონის სატრანსპორტო ჯაჭვის აქტივობა (რედოქსის ჯაჭვი) და ATP ჰიდროლიზი. როგორც რედოქსი, ასევე ჰიდროლიზური H + ტუმბოები განლაგებულია მემბრანებში, რომლებსაც შეუძლიათ სინათლის ან ქიმიური ენერგიის H + ენერგიად გარდაქმნა (ანუ პროკარიოტების პლაზმური მემბრანები, ქლოროპლასტების და მიტოქონდრიების კონიუგირებული მემბრანები). H + ATPase და / ან რედოქსის ჯაჭვის მუშაობის შედეგად, პროტონები გადაადგილდებიან და მემბრანაზე ჩნდება პროტონ-მოძრავი ძალა (H +). წყალბადის იონების ელექტროქიმიური გრადიენტი, როგორც კვლევები აჩვენებს, შეიძლება გამოყენებულ იქნას დიდი რაოდენობით მეტაბოლიტების - ანიონების, ამინომჟავების, შაქრების და ა.შ. კონიუგირებული ტრანსპორტირებისთვის (მეორადი აქტიური ტრანსპორტისთვის).
პლაზმური მემბრანის აქტივობა დაკავშირებულია უჯრედის მიერ დიდი მოლეკულური წონის მქონე მყარი და თხევადი ნივთიერებების შეწოვასთან, - ფაგოციტოზიდა პინოციტოზი(გერჩ. ფაგოსები- იქ არის , პინოსი-სასმელი, ციტოსი- უჯრედი). უჯრედის მემბრანა აყალიბებს ჯიბეებს, ანუ ინვაგინაციებს, რომლებიც იზიდავს ნივთიერებებს გარედან. შემდეგ ასეთი ინვაგინაციები იჭრება და გარშემორტყმულია გარე გარემოს წვეთით (პინოციტოზი) ან მყარი ნაწილაკებით (ფაგოციტოზი). პინოციტოზი შეინიშნება უჯრედების მრავალფეროვნებაში, განსაკუთრებით იმ ორგანოებში, სადაც ხდება შთანთქმის პროცესები.
უჯრედის მემბრანა- ეს არის უჯრედის მემბრანა, რომელიც ასრულებს შემდეგ ფუნქციებს: უჯრედის შიგთავსის გამოყოფა და გარე გარემო, ნივთიერებების შერჩევითი ტრანსპორტირება (უჯრედისთვის გარე გარემოსთან გაცვლა), ზოგიერთი ბიოქიმიური რეაქციის ადგილი, უჯრედების ინტეგრაცია. ქსოვილებში და მიღებაში.
უჯრედის მემბრანები იყოფა პლაზმურ (უჯრედშიდა) და გარედ. ნებისმიერი მემბრანის მთავარი თვისებაა ნახევრად გამტარიანობა, ანუ მხოლოდ გარკვეული ნივთიერებების გავლის უნარი. ეს საშუალებას აძლევს შერჩევით გაცვლას უჯრედსა და გარე გარემოს შორის, ან გაცვლას უჯრედის განყოფილებებს შორის.
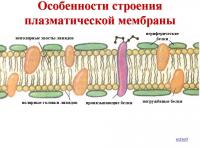
პლაზმური მემბრანები ლიპოპროტეინების სტრუქტურებია. ლიპიდები სპონტანურად ქმნიან ორ ფენას (ორმაგი შრე) და მასში მემბრანის ცილები „ცურავს“. მემბრანებში რამდენიმე ათასი სხვადასხვა ცილაა: სტრუქტურული, მატარებლები, ფერმენტები და ა.შ. ცილის მოლეკულებს შორის არის ფორები, რომლებშიც გადის ჰიდროფილური ნივთიერებები (ლიპიდური ორშრე ხელს უშლის მათ პირდაპირ შეღწევას უჯრედში). გლიკოზილის ჯგუფები (მონოსაქარიდები და პოლისაქარიდები) მიმაგრებულია მემბრანის ზედაპირზე არსებულ ზოგიერთ მოლეკულაზე, რომლებიც მონაწილეობენ ქსოვილის ფორმირებისას უჯრედების ამოცნობის პროცესში.
მემბრანები განსხვავდება მათი სისქეში, ჩვეულებრივ 5-დან 10 ნმ-მდე. სისქე განისაზღვრება ამფიფილური ლიპიდური მოლეკულის ზომით და არის 5,3 ნმ. მემბრანის სისქის შემდგომი ზრდა განპირობებულია მემბრანის ცილის კომპლექსების ზომით. გარე პირობებიდან გამომდინარე (ქოლესტერინი არის მარეგულირებელი), ორფენის სტრუქტურა შეიძლება შეიცვალოს ისე, რომ ის უფრო მკვრივი ან თხევადი გახდეს - მემბრანის გასწვრივ ნივთიერებების გადაადგილების სიჩქარე ამაზეა დამოკიდებული.
უჯრედის მემბრანებს მიეკუთვნება: პლაზმალემა, კარიოლემა, ენდოპლაზმური ბადის მემბრანები, გოლჯის აპარატი, ლიზოსომები, პეროქსიზომები, მიტოქონდრია, ინკლუზიები და ა.შ.
ლიპიდები წყალში უხსნადია (ჰიდროფობია), მაგრამ ადვილად ხსნადი ორგანულ გამხსნელებში და ცხიმებში (ლიპოფილურობა). ლიპიდების შემადგენლობა სხვადასხვა მემბრანაში არ არის იგივე. მაგალითად, პლაზმური მემბრანა შეიცავს უამრავ ქოლესტერინს. მემბრანის ლიპიდებიდან ყველაზე გავრცელებულია ფოსფოლიპიდები (გლიცეროფოსფატიდები), სფინგომიელინები (სფინგოლიპიდები), გლიკოლიპიდები და ქოლესტერინი.
ფოსფოლიპიდები, სფინგომიელინები, გლიკოლიპიდები შედგება ორი ფუნქციურად განსხვავებული ნაწილისგან: ჰიდროფობიური არაპოლარული, რომელიც არ ატარებს მუხტს - ცხიმოვანი მჟავებისგან შემდგარი "კუდები" და ჰიდროფილური, რომელიც შეიცავს დამუხტულ პოლარულ "თავებს" - ალკოჰოლის ჯგუფებს (მაგალითად, გლიცეროლს).
მოლეკულის ჰიდროფობიური ნაწილი ჩვეულებრივ შედგება ორი ცხიმოვანი მჟავისგან. ერთი მჟავა შემზღუდველია, მეორე კი უჯერი. ეს განსაზღვრავს ლიპიდების უნარს სპონტანურად შექმნან ორფენიანი (ბილიპიდური) მემბრანული სტრუქტურები. მემბრანული ლიპიდები ასრულებენ შემდეგ ფუნქციებს: ბარიერი, ტრანსპორტი, ცილების მიკროგარემო, მემბრანის ელექტრული წინააღმდეგობა.
მემბრანები ერთმანეთისგან განსხვავდება ცილის მოლეკულების ნაკრებით. ბევრი მემბრანის ცილა შედგება პოლარული (მუხტის მატარებელი) ამინომჟავებით მდიდარი რეგიონებისგან და არაპოლარული ამინომჟავებით (გლიცინი, ალანინი, ვალინი, ლეიცინი). მემბრანების ლიპიდურ ფენებში ასეთი ცილები განლაგებულია ისე, რომ მათი არაპოლარული უბნები, თითქოსდა, ჩაეფლო მემბრანის "ცხიმიან" ნაწილში, სადაც მდებარეობს ლიპიდების ჰიდროფობიური უბნები. ამ ცილების პოლარული (ჰიდროფილური) ნაწილი ურთიერთქმედებს ლიპიდურ თავებთან და მიმართულია წყლის ფაზისკენ.
ბიოლოგიურ მემბრანებს აქვთ საერთო თვისებები:
მემბრანები არის დახურული სისტემები, რომლებიც არ აძლევენ უჯრედის და მისი ნაწილების შერევის საშუალებას. მემბრანის მთლიანობის დარღვევამ შეიძლება გამოიწვიოს უჯრედის სიკვდილი;
ზედაპირული (გეგმური, გვერდითი) მობილურობა. მემბრანებში ხდება ნივთიერებების უწყვეტი მოძრაობა ზედაპირზე;
მემბრანის ასიმეტრია. გარე და ზედაპირული ფენების სტრუქტურა ქიმიურად, სტრუქტურულად და ფუნქციურად არაერთგვაროვანია.
Მოკლე აღწერა:
საზონოვი ვ.ფ. 1_1 უჯრედის მემბრანის სტრუქტურა [ელექტრონული რესურსი] // კინეზიოლოგი, 2009-2018: [ვებგვერდი]. განახლების თარიღი: 06.02.2018..__.201_). _აღწერილია უჯრედის მემბრანის აგებულება და ფუნქციონირება (სინონიმები: პლაზმალემა, პლაზმოლემა, ბიომემბრანა, უჯრედის მემბრანა, გარეთა უჯრედის მემბრანა, უჯრედის მემბრანა, ციტოპლაზმური მემბრანა). ეს საწყისი ინფორმაცია აუცილებელია როგორც ციტოლოგიისთვის, ასევე ნერვული აქტივობის პროცესების გასაგებად: ნერვული აგზნება, ინჰიბირება, სინაფსების და სენსორული რეცეპტორების მუშაობა.
უჯრედის მემბრანა (პლაზმა ალემა ან პლაზმა შესახებლემა)
კონცეფციის განმარტება
უჯრედის მემბრანა (სინონიმები: პლაზმალემა, პლაზმოლემა, ციტოპლაზმური მემბრანა, ბიომემბრანა) არის სამმაგი ლიპოპროტეინის (ანუ „ცხიმ-პროტეინის“) მემბრანა, რომელიც გამოყოფს უჯრედს გარემოსგან და ახორციელებს კონტროლირებად გაცვლასა და კომუნიკაციას უჯრედსა და მის გარემოს შორის.
ამ განმარტებაში მთავარი ის კი არ არის, რომ მემბრანა უჯრედს გარემოსგან ჰყოფს, არამედ მხოლოდ ის აკავშირებს უჯრედი გარემოსთან. მემბრანა არის აქტიური უჯრედის სტრუქტურა, ის მუდმივად მუშაობს.
ბიოლოგიური მემბრანა არის ფოსფოლიპიდების ულტრათხელი ბიმოლეკულური ფილმი, რომელიც დაფარულია ცილებით და პოლისაქარიდებით. ეს უჯრედული სტრუქტურა ეფუძნება ცოცხალი ორგანიზმის ბარიერულ, მექანიკურ და მატრიცულ თვისებებს (Antonov VF, 1996).
მემბრანის ფიგურული გამოსახულება
უჯრედის მემბრანა მეჩვენება როგორც გისოსიანი ღობე, რომელშიც ბევრი კარია, რომელიც აკრავს გარკვეულ ტერიტორიას. ნებისმიერ პატარა ცოცხალ არსებას თავისუფლად შეუძლია ამ ღობეზე წინ და უკან გადაადგილება. მაგრამ უფრო დიდ ვიზიტორებს მხოლოდ კარებიდან შეუძლიათ შესვლა და მაშინაც კი არა ყველას. სხვადასხვა ვიზიტორს აქვს მხოლოდ საკუთარი კარის გასაღები და მათ არ შეუძლიათ სხვისი კარების გავლა. ასე რომ, ამ ღობის გავლით მუდმივად ხდება ვიზიტორების ნაკადები წინ და უკან, რადგან მემბრანა-ღობის ძირითადი ფუნქცია ორმხრივია: ტერიტორიის მიმდებარე სივრცისგან გამოყოფა და ამავე დროს მიმდებარე სივრცესთან დაკავშირება. ამისათვის ღობეზე ბევრი ხვრელი და კარია - !
მემბრანის თვისებები
1. გამტარიანობა.
2. ნახევრად გამტარიანობა (ნაწილობრივი გამტარიანობა).
3. შერჩევითი (სინონიმი: შერჩევითი) გამტარიანობა.
4. აქტიური გამტარიანობა (სინონიმი: აქტიური ტრანსპორტი).
5. კონტროლირებადი გამტარიანობა.
როგორც ხედავთ, მემბრანის მთავარი თვისებაა მისი გამტარიანობა სხვადასხვა ნივთიერებების მიმართ.
6. ფაგოციტოზი და პინოციტოზი.
7. ეგზოციტოზი.
8. ელექტრული და ქიმიური პოტენციალების არსებობა, უფრო სწორედ, პოტენციური სხვაობა მემბრანის შიდა და გარე მხარეებს შორის. ფიგურალურად შეიძლება ითქვას "მემბრანა აქცევს უჯრედს "ელექტრო ბატარეად" იონური ნაკადების კონტროლით". დეტალები: .
9. ელექტრული და ქიმიური პოტენციალის ცვლილებები.
10. გაღიზიანებადობა. მემბრანაზე განლაგებულ სპეციალურ მოლეკულურ რეცეპტორებს შეუძლიათ დაუკავშირდნენ სასიგნალო (საკონტროლო) ნივთიერებებს, რის შედეგადაც შეიძლება შეიცვალოს მემბრანის მდგომარეობა და მთელი უჯრედი. მოლეკულური რეცეპტორები იწვევს ბიოქიმიურ რეაქციებს მათთან ლიგანდების (საკონტროლო ნივთიერებების) გაერთიანების საპასუხოდ. მნიშვნელოვანია აღინიშნოს, რომ სასიგნალო ნივთიერება რეცეპტორზე მოქმედებს გარედან, ხოლო ცვლილებები უჯრედის შიგნით გრძელდება. ირკვევა, რომ მემბრანა გადასცემდა ინფორმაციას გარემოდან უჯრედის შიდა გარემოში.
11. კატალიზური ფერმენტული აქტივობა. ფერმენტები შეიძლება იყოს ჩასმული მემბრანაში ან ასოცირებული იყოს მის ზედაპირთან (როგორც უჯრედის შიგნით, ისე მის გარეთ) და იქ ისინი ახორციელებენ ფერმენტულ აქტივობას.
12. ზედაპირის ფორმისა და მისი ფართობის შეცვლა. ეს საშუალებას აძლევს მემბრანას წარმოქმნას გამონაზარდები გარედან ან, პირიქით, უჯრედში შეღწევა.
13. სხვა უჯრედულ მემბრანებთან კონტაქტის დამყარების უნარი.
14. ადჰეზია - მყარ ზედაპირებზე შეწებების უნარი.
მემბრანის თვისებების მოკლე ჩამონათვალი
- გამტარიანობა.
- ენდოციტოზი, ეგზოციტოზი, ტრანსციტოზი.
- პოტენციალი.
- გაღიზიანებადობა.
- ფერმენტული აქტივობა.
- კონტაქტები.
- ადჰეზია.
მემბრანის ფუნქციები
1. შინაგანი შინაარსის არასრული იზოლაცია გარე გარემოდან.
2. უჯრედის მემბრანის მუშაობაში მთავარია გაცვლა სხვადასხვა ნივთიერებები უჯრედსა და უჯრედგარე გარემოს შორის. ეს გამოწვეულია მემბრანის ისეთი თვისებით, როგორიცაა გამტარიანობა. გარდა ამისა, მემბრანა არეგულირებს ამ გაცვლას მისი გამტარიანობის რეგულირებით.
3. მემბრანის კიდევ ერთი მნიშვნელოვანი ფუნქციაა ქმნის განსხვავებას ქიმიურ და ელექტრულ პოტენციალებში მის შიდა და გარე მხარეებს შორის. ამის გამო, უჯრედის შიგნით არის უარყოფითი ელექტრული პოტენციალი -.
4. მემბრანის მეშვეობით ასევე ხორციელდება ინფორმაციის გაცვლა უჯრედსა და მის გარემოს შორის. მემბრანაზე განლაგებულ სპეციალურ მოლეკულურ რეცეპტორებს შეუძლიათ დაუკავშირონ საკონტროლო ნივთიერებებს (ჰორმონები, შუამავლები, მოდულატორები) და გამოიწვიოს ბიოქიმიური რეაქციები უჯრედში, რაც იწვევს უჯრედში ან მის სტრუქტურებში სხვადასხვა ცვლილებებს.
ვიდეო:უჯრედის მემბრანის სტრუქტურა
ვიდეო ლექცია:დეტალები მემბრანის სტრუქტურისა და ტრანსპორტის შესახებ
მემბრანის სტრუქტურა
უჯრედის მემბრანას აქვს უნივერსალური სამ ფენიანი სტრუქტურა. მისი საშუალო ცხიმის ფენა უწყვეტია, ხოლო ზედა და ქვედა ცილის ფენები მას ფარავს ცალკეული ცილის უბნების მოზაიკის სახით. ცხიმოვანი ფენა არის საფუძველი, რომელიც უზრუნველყოფს უჯრედის გარემოსგან იზოლაციას, გარემოსგან იზოლირებას. თავისთავად, ის ძალიან ცუდად გადის წყალში ხსნად ნივთიერებებს, მაგრამ ადვილად გადის ცხიმში ხსნადს. ამიტომ, მემბრანის გამტარიანობა წყალში ხსნადი ნივთიერებებისთვის (მაგალითად, იონები) უნდა იყოს უზრუნველყოფილი სპეციალური ცილოვანი სტრუქტურებით - და.
ქვემოთ მოცემულია კონტაქტური უჯრედების რეალური უჯრედის მემბრანების მიკროფოტოები, რომლებიც მიღებულია ელექტრონული მიკროსკოპის გამოყენებით, ასევე სქემატური ნახატი, რომელიც გვიჩვენებს სამშრიანი მემბრანას და მისი ცილოვანი ფენების მოზაიკურ ხასიათს. სურათის გასადიდებლად დააწკაპუნეთ მასზე.

უჯრედის მემბრანის შიდა ლიპიდური (ცხიმოვანი) ფენის ცალკე გამოსახულება, გაჟღენთილი ინტეგრალური ჩაშენებული ცილებით. ზედა და ქვედა ცილის შრეები ამოღებულია ისე, რომ ხელი არ შეუშალოს ლიპიდური ორშრის განხილვას.

სურათი ზემოთ: უჯრედის მემბრანის (უჯრედის კედლის) არასრული სქემატური წარმოდგენა ვიკიპედიიდან.
გაითვალისწინეთ, რომ აქ გარსიდან ამოღებულია ცილის გარე და შიდა შრეები, რათა უკეთ დავინახოთ ცენტრალური ცხიმოვანი ორმაგი ლიპიდური შრე. რეალურ უჯრედულ მემბრანაში დიდი ცილის „კუნძულები“ ცურავს ცხიმოვანი ფილმის გასწვრივ ზემოთ და ქვემოთ (სურათზე პატარა ბურთები) და მემბრანა აღმოჩნდება უფრო სქელი, სამშრიანი: ცილა-ცხიმ-ცილა . ასე რომ, ეს რეალურად ჰგავს სენდვიჩს ორი ცილისგან შემდგარი „პურის ნაჭერით“, შუაში „კარაქის“ სქელი ფენით, ე.ი. აქვს სამ ფენიანი სტრუქტურა და არა ორფენიანი.
ამ ფიგურაში პატარა ლურჯი და თეთრი ბურთულები შეესაბამება ლიპიდების ჰიდროფილურ (დასველებად) „თავებს“, მათზე დამაგრებული „სტრიქონები“ ჰიდროფობიურ (არასველებად) „კუდებს“. ცილებიდან ნაჩვენებია მხოლოდ მემბრანის ინტეგრალური ცილები (წითელი გლობულები და ყვითელი ხვეულები). მემბრანის შიგნით ყვითელი ოვალური წერტილები არის ქოლესტერინის მოლეკულები გარსის გარეთა მძივების ყვითელ-მწვანე ჯაჭვები არის ოლიგოსაქარიდული ჯაჭვები, რომლებიც ქმნიან გლიკოკალიქსს. გლიკოკალიქსი ჰგავს ნახშირწყლების („შაქარის“) „ფუფუნებას“ მემბრანაზე, რომელიც წარმოიქმნება მისგან გამოსული გრძელი ნახშირწყალ-ცილოვანი მოლეკულებით.
ლაივინგი არის პატარა „ცილოვან-ცხიმიანი ტომარა“, რომელიც სავსეა ნახევრად თხევადი ჟელეს მსგავსი შიგთავსით, რომელიც შეაღწევს ფილმებს და მილებს.
ამ ტომრის კედლები წარმოიქმნება ორმაგი ცხიმოვანი (ლიპიდური) ფილმით, რომელიც დაფარულია შიგნიდან და გარედან ცილებით - უჯრედის მემბრანა. ამიტომ, მემბრანას ამბობენ, რომ აქვს სამ ფენიანი სტრუქტურა : ცილები-ცხიმები-პროტეინები. უჯრედის შიგნით არის ასევე მრავალი მსგავსი ცხიმოვანი მემბრანა, რომელიც მის შიდა სივრცეს ნაწილებად ყოფს. უჯრედულ ორგანელებს აკრავს ერთი და იგივე გარსები: ბირთვი, მიტოქონდრია, ქლოროპლასტები. ასე რომ, მემბრანა არის უნივერსალური მოლეკულური სტრუქტურა, რომელიც თან ახლავს ყველა უჯრედს და ყველა ცოცხალ ორგანიზმს.

მარცხნივ - აღარ არის ბიოლოგიური მემბრანის ნაწილის რეალური, არამედ ხელოვნური მოდელი: ეს არის ცხიმოვანი ფოსფოლიპიდური ორფენის (ანუ ორმაგი შრის) მყისიერი სურათი მისი მოლეკულური დინამიკის მოდელირების პროცესში. ნაჩვენებია მოდელის საანგარიშო უჯრედი - 96 PQ მოლეკულა ( ვოსფატიდილი Xოლინი) და 2304 წყლის მოლეკულა, სულ 20544 ატომი.
მარჯვნივ არის იგივე ლიპიდის ერთი მოლეკულის ვიზუალური მოდელი, საიდანაც აწყობილია მემბრანის ლიპიდური ორშრე. ზემოდან აქვს ჰიდროფილური (წყლის მოყვარული) თავი, ქვემოდან კი ორი ჰიდროფობიური (წყლის მოშიში) კუდი. ამ ლიპიდს აქვს მარტივი სახელი: 1-სტეროილ-2-დოკოზაჰექსაენოილ-Sn-გლიცერო-3-ფოსფატიდილქოლინი (18:0/22:6(n-3)cis PC), მაგრამ თქვენ არ გჭირდებათ მისი დამახსოვრება, თუ დაგეგმეთ თქვენი მასწავლებლის გაძარცვა თქვენი ცოდნის სიღრმით.
თქვენ შეგიძლიათ მიუთითოთ უჯრედის უფრო ზუსტი მეცნიერული განმარტება:
არის ბიოპოლიმერების მოწესრიგებული, სტრუქტურირებული ჰეტეროგენული სისტემა, რომელიც შემოიფარგლება აქტიური მემბრანით, რომელიც მონაწილეობს მეტაბოლური, ენერგეტიკული და საინფორმაციო პროცესების ერთ ნაკრებში და ასევე ინარჩუნებს და ამრავლებს მთელ სისტემას, როგორც მთლიანს.
უჯრედის შიგნით ასევე შეაღწევს მემბრანები, ხოლო გარსებს შორის არის არა წყალი, არამედ ცვლადი სიმკვრივის ბლანტი გელი/სოლი. ამრიგად, უჯრედში ურთიერთმოქმედი მოლეკულები თავისუფლად არ ცურავს, როგორც სინჯარაში წყალხსნარში, არამედ ძირითადად სხედან (იმობილიზებულნი) ციტოჩონჩხის ან უჯრედშიდა გარსების პოლიმერულ სტრუქტურებზე. და ამიტომ, ქიმიური რეაქციები ხდება უჯრედის შიგნით თითქმის ისე, როგორც მყარ სხეულში, და არა სითხეში. გარე მემბრანა, რომელიც გარს აკრავს უჯრედს, ასევე დაფარულია ფერმენტებითა და მოლეკულური რეცეპტორებით, რაც მას უჯრედის ძალიან აქტიურ ნაწილად აქცევს.
უჯრედის მემბრანა (პლაზმალემა, პლაზმოლემა) არის აქტიური გარსი, რომელიც გამოყოფს უჯრედს გარემოსგან და აკავშირებს მას გარემოსთან. © Sazonov V.F., 2016 წ.
მემბრანის ამ განმარტებიდან გამომდინარეობს, რომ ის უბრალოდ არ ზღუდავს უჯრედს, არამედ აქტიურად მუშაობსაკავშირებს მას მის გარემოსთან.
ცხიმი, რომელიც ქმნის გარსებს, განსაკუთრებულია, ამიტომ მის მოლეკულებს ჩვეულებრივ უწოდებენ არა მხოლოდ ცხიმს, არამედ ლიპიდები, ფოსფოლიპიდები, სფინგოლიპიდები. მემბრანის ფილმი ორმაგია, ანუ შედგება ორი ფირისგან, რომლებიც ერთმანეთთან არის დამაგრებული. ამიტომ, სახელმძღვანელოები წერენ, რომ უჯრედის მემბრანის საფუძველი შედგება ორი ლიპიდური ფენისგან (ან " ორფენიანი", ანუ ორმაგი ფენა). თითოეული ცალკეული ლიპიდური ფენისთვის, ერთი მხარე შეიძლება დატენიანდეს წყლით, მეორე კი არა. ასე რომ, ეს ფენები ერთმანეთს ეწებება ზუსტად მათი არასველი მხარეებით.
ბაქტერიების მემბრანა
გრამუარყოფითი ბაქტერიების პროკარიოტული უჯრედის გარსი შედგება რამდენიმე ფენისგან, რომლებიც ნაჩვენებია ქვემოთ მოცემულ ფიგურაში.
გრამუარყოფითი ბაქტერიების გარსის ფენები:
1. შიდა სამშრიანი ციტოპლაზმური მემბრანა, რომელიც კონტაქტშია ციტოპლაზმასთან.
2. უჯრედის კედელი, რომელიც შედგება მირეინისგან.
3. გარე სამშრიანი ციტოპლაზმური მემბრანა, რომელსაც აქვს იგივე ლიპიდების სისტემა ცილოვანი კომპლექსებით, როგორც შიდა მემბრანა.
გრამუარყოფითი ბაქტერიების უჯრედების კომუნიკაცია გარე სამყაროსთან ასეთი რთული სამსაფეხურიანი სტრუქტურის მეშვეობით არ აძლევს მათ უპირატესობას მძიმე პირობებში გადარჩენაში გრამდადებით ბაქტერიებთან შედარებით, რომლებსაც აქვთ ნაკლებად ძლიერი გარსი. ისინი ისევე ცუდად იტანენ მაღალ ტემპერატურას, მაღალ მჟავიანობას და წნევის ვარდნას.




ვიდეო ლექცია:პლაზმური მემბრანა. ე.ვ. შევალი, ფ.
ვიდეო ლექცია:მემბრანა, როგორც უჯრედის საზღვარი. ა.ილიასკინი
მემბრანული იონური არხების მნიშვნელობა
ადვილი გასაგებია, რომ მხოლოდ ცხიმში ხსნადი ნივთიერებები შეიძლება შევიდნენ უჯრედში მემბრანის ცხიმოვანი ფირის მეშვეობით. ეს არის ცხიმები, ალკოჰოლები, გაზები.მაგალითად, ერითროციტებში ჟანგბადი და ნახშირორჟანგი ადვილად შედიან და გამოდიან პირდაპირ მემბრანაში. მაგრამ წყალი და წყალში ხსნადი ნივთიერებები (მაგალითად, იონები) უბრალოდ ვერ გადადიან მემბრანის გავლით რომელიმე უჯრედში. ეს ნიშნავს, რომ მათ სპეციალური ხვრელები სჭირდებათ. მაგრამ თუ თქვენ უბრალოდ გააკეთებთ ხვრელს ცხიმოვან ფილმში, მაშინ ის დაუყოვნებლივ გამკაცრდება უკან. Რა უნდა ვქნა? ბუნებაში იპოვეს გამოსავალი: აუცილებელია სპეციალური ცილის სატრანსპორტო სტრუქტურების დამზადება და მემბრანის გავლით გაჭიმვა. ასე მიიღება ცხიმში უხსნადი ნივთიერებების გავლის არხები – უჯრედის მემბრანის იონური არხები.
ასე რომ, იმისათვის, რომ მის მემბრანას პოლარული მოლეკულებისთვის (იონები და წყალი) გამტარიანობის დამატებითი თვისებები მისცეს, უჯრედი სინთეზირებს ციტოპლაზმაში სპეციალურ ცილებს, რომლებიც შემდეგ მემბრანაში ინტეგრირდება. ისინი ორი ტიპისაა: გადამტანი ცილები (მაგალითად, სატრანსპორტო ATPases) და არხის წარმომქმნელი ცილები (არხის შემქმნელები). ეს ცილები ჩაშენებულია მემბრანის ორმაგ ცხიმოვან შრეში და ქმნიან სატრანსპორტო სტრუქტურებს გადამტანების სახით ან იონური არხების სახით. ამ სატრანსპორტო სტრუქტურებში შეიძლება გაიაროს წყალში ხსნადი სხვადასხვა ნივთიერებები, რომლებიც სხვაგვარად ვერ გაივლიან ცხიმოვანი მემბრანის ფილას.
ზოგადად მემბრანაში ჩადგმულ ცილებსაც ეძახიან განუყოფელი, ზუსტად იმიტომ, რომ ისინი, როგორც იქნა, შედიან მემბრანის შემადგენლობაში და შეაღწევენ მასში და მეშვეობით. სხვა ცილები, არა განუყოფელი, ქმნიან კუნძულებს, რომლებიც "ცურავს" მემბრანის ზედაპირზე: ან მისი გარე ზედაპირის გასწვრივ, ან მისი შიდა ზედაპირის გასწვრივ. ყოველივე ამის შემდეგ, ყველამ იცის, რომ ცხიმი კარგი ლუბრიკანტია და მასზე სრიალება ადვილია!
დასკვნები
1. ზოგადად მემბრანა სამფენიანია:
1) ცილის გარე ფენა "კუნძულები",
2) ცხიმოვანი ორფენიანი „ზღვა“ (ლიპიდური ორშრე), ე.ი. ორმაგი ლიპიდური ფილმი
3) ცილოვანი „კუნძულების“ შიდა ფენა.
მაგრამ ასევე არსებობს ფხვიერი გარე შრე - გლიკოკალიქსი, რომელიც წარმოიქმნება გარსიდან გამოსული გლიკოპროტეინებით. ისინი მოლეკულური რეცეპტორებია, რომლებზეც სიგნალის კონტროლის ბლოკებია დაკავშირებული.
2. მემბრანაში ჩაშენებულია სპეციალური ცილოვანი სტრუქტურები, რაც უზრუნველყოფს მის გამტარიანობას იონების ან სხვა ნივთიერებების მიმართ. არ უნდა დაგვავიწყდეს, რომ ზოგან ცხიმის ზღვა გაჟღენთილია ინტეგრალური ცილებით. და ეს არის განუყოფელი ცილები, რომლებიც ქმნიან განსაკუთრებულს სატრანსპორტო სტრუქტურები უჯრედის მემბრანა (იხ. განყოფილება 1_2 მემბრანის ტრანსპორტირების მექანიზმები). მათი მეშვეობით ნივთიერებები უჯრედში შედიან და ასევე უჯრედიდან გარედან გამოიყოფა.
3. ფერმენტული ცილები შეიძლება განთავსდეს მემბრანის ნებისმიერ მხარეს (გარე და შიდა), ასევე მემბრანის შიგნით, რაც გავლენას ახდენს როგორც თავად მემბრანის მდგომარეობაზე, ასევე მთელი უჯრედის სიცოცხლეზე.
ასე რომ, უჯრედის მემბრანა არის აქტიური ცვლადი სტრუქტურა, რომელიც აქტიურად მუშაობს მთელი უჯრედის ინტერესებიდან გამომდინარე და აკავშირებს მას გარე სამყაროსთან და არ არის მხოლოდ „დამცავი გარსი“. ეს არის ყველაზე მნიშვნელოვანი რამ, რაც უნდა იცოდეთ უჯრედის მემბრანის შესახებ.
მედიცინაში მემბრანის ცილებს ხშირად იყენებენ წამლების "სამიზნეებად". ასეთ სამიზნეებად მოქმედებენ რეცეპტორები, იონური არხები, ფერმენტები, სატრანსპორტო სისტემები. ბოლო დროს მემბრანის გარდა, ნარკოტიკების სამიზნე უჯრედის ბირთვში დამალული გენებიც გახდა.
ვიდეო:უჯრედის მემბრანის ბიოფიზიკის შესავალი: მემბრანის სტრუქტურა 1 (ვლადიმიროვი იუ.ა.)
ვიდეო:უჯრედის მემბრანის ისტორია, სტრუქტურა და ფუნქციები: მემბრანების სტრუქტურა 2 (ვლადიმიროვი იუ.ა.)
© 2010-2018 Sazonov V.F., © 2010-2016 kineziolog.bodhy.
მემბრანა არის ჰიპერწვრილი სტრუქტურა, რომელიც ქმნის ორგანელების ზედაპირს და მთლიანად უჯრედს. ყველა მემბრანას აქვს მსგავსი სტრუქტურა და დაკავშირებულია ერთ სისტემაში.
Ქიმიური შემადგენლობა
უჯრედის მემბრანები ქიმიურად ერთგვაროვანია და შედგება სხვადასხვა ჯგუფის ცილებისა და ლიპიდებისგან:
- ფოსფოლიპიდები;
- გალაქტოლიპიდები;
- სულფოლიპიდები.
ისინი ასევე შეიცავს ნუკლეინის მჟავებს, პოლისაქარიდებს და სხვა ნივთიერებებს.
ფიზიკური თვისებები
ნორმალურ ტემპერატურაზე მემბრანები თხევადკრისტალურ მდგომარეობაშია და მუდმივად იცვლება. მათი სიბლანტე ახლოს არის მცენარეული ზეთის სიბლანტესთან.
მემბრანა არის აღდგენითი, ძლიერი, ელასტიური და აქვს ფორები. მემბრანების სისქე 7 - 14 ნმ.
TOP 4 სტატიავინც ამას კითხულობს
დიდი მოლეკულებისთვის მემბრანა გაუვალია. მცირე მოლეკულებს და იონებს შეუძლიათ გაიარონ ფორებში და თავად მემბრანაში მემბრანის სხვადასხვა მხარეს კონცენტრაციის სხვაობის გავლენის ქვეშ, აგრეთვე სატრანსპორტო ცილების დახმარებით.
მოდელი
მემბრანების სტრუქტურა ჩვეულებრივ აღწერილია თხევადი მოზაიკის მოდელის გამოყენებით. მემბრანას აქვს ჩარჩო - ლიპიდური მოლეკულების ორი რიგი, მჭიდროდ, აგურის მსგავსად, ერთმანეთის მიმდებარედ.

ბრინჯი. 1. სენდვიჩის ტიპის ბიოლოგიური მემბრანა.
ორივე მხრიდან ლიპიდების ზედაპირი დაფარულია ცილებით. მოზაიკის ნიმუში იქმნება ცილის მოლეკულებით, რომლებიც არათანაბრად ნაწილდება მემბრანის ზედაპირზე.
ბილიპიდური შრეში ჩაძირვის ხარისხის მიხედვით ცილის მოლეკულები იყოფა სამი ჯგუფი:
- ტრანსმემბრანული;
- ჩაძირული;
- ზედაპირული.
ცილები უზრუნველყოფენ მემბრანის ძირითად თვისებას - მის შერჩევით გამტარიანობას სხვადასხვა ნივთიერებისთვის.
მემბრანის ტიპები
ყველა უჯრედის მემბრანა ლოკალიზაციის მიხედვით შეიძლება დაიყოს შემდეგი ტიპები:
- გარე;
- ბირთვული;
- ორგანული მემბრანები.
გარე ციტოპლაზმური მემბრანა, ანუ პლაზმოლემა, არის უჯრედის საზღვარი. ციტოჩონჩხის ელემენტებთან შეერთებით, ის ინარჩუნებს ფორმასა და ზომას.

ბრინჯი. 2. ციტოჩონჩხი.
ბირთვული მემბრანა, ანუ კარიოლემა, არის ბირთვული შინაარსის საზღვარი. იგი აგებულია ორი მემბრანისგან, რომელიც ძალიან ჰგავს გარე მემბრანას. ბირთვის გარე მემბრანა დაკავშირებულია ენდოპლაზმური ბადის მემბრანებთან (ER) და ფორების მეშვეობით შიდა მემბრანასთან.
EPS მემბრანები შეაღწევს მთელ ციტოპლაზმას, ქმნიან ზედაპირებს, რომლებზეც სინთეზირდება სხვადასხვა ნივთიერებები, მათ შორის მემბრანის ცილები.
ორგანული მემბრანები
ორგანელების უმეტესობას აქვს მემბრანული სტრუქტურა.
კედლები აგებულია ერთი გარსისგან:
- გოლგის კომპლექსი;
- ვაკუოლები;
- ლიზოსომები.
პლასტიდები და მიტოქონდრია აგებულია მემბრანის ორი ფენისგან. მათი გარე გარსი გლუვია, შიდა კი მრავალ ნაკეცს ქმნის.
ქლოროპლასტების ფოტოსინთეზური მემბრანების თავისებურებები არის ქლოროფილის მოლეკულები.
გარე გარსის ზედაპირზე ცხოველურ უჯრედებს აქვთ ნახშირწყლების ფენა, რომელსაც გლიკოკალიქსი ეწოდება.

ბრინჯი. 3. გლიკოკალიქსი.
გლიკოკალიქსი ყველაზე მეტად განვითარებულია ნაწლავის ეპითელიუმის უჯრედებში, სადაც ქმნის მონელების პირობებს და იცავს პლაზმოლემას.
ცხრილი "უჯრედის მემბრანის სტრუქტურა"
რა ვისწავლეთ?
ჩვენ შევისწავლეთ უჯრედის მემბრანის სტრუქტურა და ფუნქციები. მემბრანა არის უჯრედის, ბირთვისა და ორგანელების შერჩევითი (შერჩევითი) ბარიერი. უჯრედის მემბრანის სტრუქტურა აღწერილია სითხე-მოზაიკის მოდელით. ამ მოდელის მიხედვით, ცილის მოლეკულები ჩასმულია ბლანტი ლიპიდების ორმაგი ფენაში.
თემის ვიქტორინა
ანგარიშის შეფასება
Საშუალო რეიტინგი: 4.5. სულ მიღებული შეფასებები: 270.
ორგანიზმების, ისევე როგორც მცენარეების, ცხოველების და ადამიანების სტრუქტურის შესწავლა ბიოლოგიის დარგია, რომელსაც ციტოლოგია ეწოდება. მეცნიერებმა დაადგინეს, რომ უჯრედის შიგთავსი, რომელიც მასშია, საკმაოდ რთულია. მას აკრავს ეგრეთ წოდებული ზედაპირული აპარატი, რომელიც მოიცავს გარე უჯრედის მემბრანას, ზემემბრანულ სტრუქტურებს: გლიკოკალიქსს და მიკროფილამენტებს, პელიკულას და მიკროტუბულებს, რომლებიც ქმნიან მის ქვემემბრანულ კომპლექსს.
ამ სტატიაში ჩვენ შევისწავლით გარე უჯრედის მემბრანის სტრუქტურასა და ფუნქციებს, რომელიც წარმოადგენს სხვადასხვა ტიპის უჯრედების ზედაპირული აპარატის ნაწილს.
რა ფუნქციები აქვს გარე უჯრედის მემბრანას?
როგორც უკვე აღვნიშნეთ, გარე მემბრანა არის თითოეული უჯრედის ზედაპირული აპარატის ნაწილი, რომელიც წარმატებით გამოყოფს მის შიდა შიგთავსს და იცავს უჯრედის ორგანელებს არასასურველი გარემო პირობებისგან. კიდევ ერთი ფუნქციაა უჯრედის შიგთავსსა და ქსოვილის სითხეს შორის ნივთიერებების გაცვლის უზრუნველყოფა, შესაბამისად, უჯრედის გარე მემბრანა გადააქვს მოლეკულებს და იონებს, რომლებიც შედიან ციტოპლაზმაში, ასევე ხელს უწყობს უჯრედიდან ტოქსინებისა და ჭარბი ტოქსიკური ნივთიერებების ამოღებას.

უჯრედის მემბრანის სტრუქტურა
სხვადასხვა ტიპის უჯრედების მემბრანები, ანუ პლაზმალემები ძალიან განსხვავდება ერთმანეთისგან. ძირითადად, ქიმიური სტრუქტურა, აგრეთვე მათში ლიპიდების, გლიკოპროტეინების, ცილების ფარდობითი შემცველობა და, შესაბამისად, მათში არსებული რეცეპტორების ბუნება. გარეგანი, რომელიც ძირითადად განისაზღვრება გლიკოპროტეინების ინდივიდუალური შემადგენლობით, მონაწილეობს გარემო სტიმულის ამოცნობაში და თავად უჯრედის რეაქციებში მათ მოქმედებებზე. ზოგიერთი ტიპის ვირუსს შეუძლია ურთიერთქმედება უჯრედის მემბრანების ცილებთან და გლიკოლიპიდებთან, რის შედეგადაც ისინი შეაღწევენ უჯრედში. ჰერპესის და გრიპის ვირუსების გამოყენება შესაძლებელია მათი დამცავი გარსის შესაქმნელად.

ხოლო ვირუსები და ბაქტერიები, ეგრეთ წოდებული ბაქტერიოფაგები, მიმაგრებულია უჯრედის მემბრანაზე და სპეციალური ფერმენტის დახმარებით ხსნიან მას შეხების ადგილას. შემდეგ ვირუსული დნმ-ის მოლეკულა გადადის წარმოქმნილ ხვრელში.
ევკარიოტების პლაზმური მემბრანის სტრუქტურის თავისებურებები
შეგახსენებთ, რომ გარე უჯრედის მემბრანა ასრულებს ტრანსპორტირების ფუნქციას, ანუ ნივთიერებების გადატანას და მის გარეთ გარე გარემოში. ასეთი პროცესის განსახორციელებლად საჭიროა სპეციალური სტრუქტურა. მართლაც, პლაზმალემა არის ზედაპირის აპარატის მუდმივი, უნივერსალური სისტემა ყველასთვის. ეს არის თხელი (2-10 ნმ), მაგრამ საკმაოდ მკვრივი მრავალშრიანი ფილმი, რომელიც ფარავს მთელ უჯრედს. მისი სტრუქტურა 1972 წელს შეისწავლეს ისეთმა მეცნიერებმა, როგორებიც არიან დ.სინგერი და გ.ნიკოლსონი, მათ ასევე შექმნეს უჯრედის მემბრანის სითხე-მოზაიკური მოდელი.
ძირითადი ქიმიური ნაერთები, რომლებიც მას ქმნიან, არის ცილების და გარკვეული ფოსფოლიპიდების მოწესრიგებული მოლეკულები, რომლებიც ირევა თხევად ლიპიდურ გარემოში და წააგავს მოზაიკას. ამრიგად, უჯრედის მემბრანა შედგება ლიპიდების ორი შრისგან, რომელთა არაპოლარული ჰიდროფობიური „კუდები“ მემბრანის შიგნითაა განლაგებული, ხოლო პოლარული ჰიდროფილური თავები უჯრედის ციტოპლაზმისა და უჯრედშორისი სითხისკენაა მიმართული.
ლიპიდურ ფენაში შედიან ცილის დიდი მოლეკულები, რომლებიც ქმნიან ჰიდროფილურ ფორებს. სწორედ მათი მეშვეობით ხდება გლუკოზისა და მინერალური მარილების წყალხსნარების ტრანსპორტირება. ცილის ზოგიერთი მოლეკულა განლაგებულია როგორც პლაზმურის გარე, ისე შიდა ზედაპირებზე. ამრიგად, ბირთვების მქონე ყველა ორგანიზმის უჯრედებში გარე უჯრედის მემბრანაზე არის ნახშირწყლების მოლეკულები, რომლებიც დაკავშირებულია კოვალენტური ბმებით გლიკოლიპიდებთან და გლიკოპროტეინებთან. უჯრედის მემბრანებში ნახშირწყლების შემცველობა 2-დან 10%-მდე მერყეობს.

პროკარიოტული ორგანიზმების პლაზმალემის სტრუქტურა
პროკარიოტებში გარე უჯრედის მემბრანა ასრულებს ბირთვული ორგანიზმების უჯრედების პლაზმური მემბრანების მსგავს ფუნქციებს, კერძოდ: გარე გარემოდან მომდინარე ინფორმაციის აღქმას და გადაცემას, იონებისა და ხსნარების ტრანსპორტირებას უჯრედში და მის გარეთ და დაცვას. ციტოპლაზმა გარედან უცხო რეაგენტებისგან. მას შეუძლია შექმნას მეზოსომები - სტრუქტურები, რომლებიც წარმოიქმნება უჯრედში პლაზმალემის ამოღების დროს. ისინი შეიძლება შეიცავდეს ფერმენტებს, რომლებიც მონაწილეობენ პროკარიოტების მეტაბოლურ რეაქციებში, მაგალითად, დნმ-ის რეპლიკაციაში, ცილების სინთეზში.
მეზოსომები ასევე შეიცავს რედოქს ფერმენტებს, ხოლო ფოტოსინთეზი შეიცავს ბაქტერიოქლოროფილს (ბაქტერიებში) და ფიკობილინს (ციანობაქტერიებში).
გარე მემბრანების როლი უჯრედშორის კონტაქტებში
გავაგრძელოთ პასუხის კითხვაზე, თუ რა ფუნქციებს ასრულებს გარე უჯრედის მემბრანა, მოდით ვისაუბროთ მის როლზე მცენარის უჯრედებში.მცენარის უჯრედებში ფორები წარმოიქმნება უჯრედის გარეთა მემბრანის კედლებში, რომლებიც გადადიან ცელულოზის შრეში. მათი მეშვეობით შესაძლებელია უჯრედის ციტოპლაზმის გარედან გასვლა, ასეთ თხელ არხებს პლაზმოდესმატებს უწოდებენ.

მათი წყალობით მეზობელ მცენარეთა უჯრედებს შორის კავშირი ძალიან ძლიერია. ადამიანისა და ცხოველის უჯრედებში მეზობელ უჯრედულ მემბრანებს შორის კონტაქტის ადგილებს დესმოსომა ეწოდება. ისინი დამახასიათებელია ენდოთელური და ეპითელური უჯრედებისთვის და ასევე გვხვდება კარდიომიოციტებში.
პლაზმალემის დამხმარე წარმონაქმნები
იმის გასაგებად, თუ როგორ განსხვავდებიან მცენარეული უჯრედები ცხოველებისგან, ეს ხელს უწყობს მათი პლაზმური მემბრანების სტრუქტურული მახასიათებლების შესწავლას, რაც დამოკიდებულია იმაზე, თუ რა ფუნქციებს ასრულებს გარე უჯრედის მემბრანა. მის ზემოთ ცხოველურ უჯრედებში არის გლიკოკალიქსის ფენა. იგი წარმოიქმნება პოლისაქარიდის მოლეკულებით, რომლებიც დაკავშირებულია გარე უჯრედის მემბრანის ცილებთან და ლიპიდებთან. გლიკოკალიქსის წყალობით ხდება უჯრედებს შორის ადჰეზია (დაწებება), რაც იწვევს ქსოვილების წარმოქმნას, ამიტომ ის მონაწილეობს პლაზმალემის სასიგნალო ფუნქციაში - გარემო სტიმულის ამოცნობაში.
როგორ ხდება გარკვეული ნივთიერებების პასიური ტრანსპორტირება უჯრედის მემბრანებში
როგორც უკვე აღვნიშნეთ, გარე უჯრედის მემბრანა ჩართულია უჯრედსა და გარე გარემოს შორის ნივთიერებების ტრანსპორტირების პროცესში. პლაზმალემის მეშვეობით ტრანსპორტის ორი ტიპი არსებობს: პასიური (დიფუზიური) და აქტიური ტრანსპორტი. პირველი მოიცავს დიფუზიას, გაადვილებულ დიფუზიას და ოსმოსს. ნივთიერებების მოძრაობა კონცენტრაციის გრადიენტის გასწვრივ, პირველ რიგში, დამოკიდებულია მოლეკულების მასაზე და ზომაზე, რომლებიც გადიან უჯრედის მემბრანაში. მაგალითად, მცირე არაპოლარული მოლეკულები ადვილად იხსნება პლაზმალემის შუა ლიპიდურ შრეში, მოძრაობს მასში და მთავრდება ციტოპლაზმაში.

ორგანული ნივთიერებების დიდი მოლეკულები ციტოპლაზმაში შეაღწევს სპეციალური გადამზიდავი ცილების დახმარებით. ისინი სპეციფიკურია სახეობებისთვის და ნაწილაკთან ან იონთან შერწყმისას ისინი პასიურად გადააქვთ მემბრანაში კონცენტრაციის გრადიენტის გასწვრივ (პასიური ტრანსპორტი) ენერგიის დახარჯვის გარეშე. ეს პროცესი საფუძვლად უდევს პლაზმალემის ისეთ თვისებას, როგორიცაა შერჩევითი გამტარიანობა. ამ პროცესში ატფ-ის მოლეკულების ენერგია არ გამოიყენება და უჯრედი ინახავს მას სხვა მეტაბოლური რეაქციებისთვის.
ქიმიური ნაერთების აქტიური ტრანსპორტირება პლაზმალმაში
ვინაიდან გარე უჯრედის მემბრანა უზრუნველყოფს მოლეკულების და იონების გადატანას გარე გარემოდან უჯრედში და უკან, შესაძლებელი ხდება დისიმილაციის პროდუქტების, რომლებიც ტოქსინებია, ამოღება გარედან, ანუ უჯრედშორის სითხეში. ხდება კონცენტრაციის გრადიენტის საწინააღმდეგოდ და მოითხოვს ენერგიის გამოყენებას ATP მოლეკულების სახით. იგი ასევე მოიცავს გადამზიდავ ცილებს, სახელწოდებით ATPases, რომლებიც ასევე ფერმენტებია.

ასეთი ტრანსპორტის მაგალითია ნატრიუმ-კალიუმის ტუმბო (ნატრიუმის იონები ციტოპლაზმიდან გარე გარემოში გადადიან, ხოლო კალიუმის იონები ციტოპლაზმაში ტუმბოს). ნაწლავისა და თირკმელების ეპითელური უჯრედები ამას ახერხებენ. გადაცემის ამ მეთოდის სახეობებია პინოციტოზის და ფაგოციტოზის პროცესები. ამრიგად, შესწავლის შემდეგ, თუ რა ფუნქციებს ასრულებს გარე უჯრედის მემბრანა, შეიძლება დადგინდეს, რომ ჰეტეროტროფულ პროტისტებს, ისევე როგორც უმაღლესი ცხოველური ორგანიზმების უჯრედებს, მაგალითად, ლეიკოციტებს, შეუძლიათ პინო- და ფაგოციტოზი.
ბიოელექტრული პროცესები უჯრედის მემბრანებში
დადგენილია, რომ არსებობს პოტენციური სხვაობა პლაზმალემის გარე ზედაპირს (ის დადებითად არის დამუხტული) და ციტოპლაზმის პარიეტალურ შრეს შორის, რომელიც უარყოფითად არის დამუხტული. მას ეწოდა დასვენების პოტენციალი და ის თანდაყოლილია ყველა ცოცხალ უჯრედში. ნერვულ ქსოვილს კი არა მხოლოდ მოსვენების პოტენციალი აქვს, არამედ შეუძლია სუსტი ბიოდენის გატარება, რასაც აგზნების პროცესს უწოდებენ. ნერვული უჯრედების - ნეირონების გარე გარსები, რომლებიც იღებენ გაღიზიანებას რეცეპტორებისგან, იწყებენ მუხტების შეცვლას: ნატრიუმის იონები მასიურად შედიან უჯრედში და პლაზმალემის ზედაპირი ხდება ელექტროუარყოფითი. ხოლო ციტოპლაზმის პარიეტალური ფენა, კათიონების სიჭარბის გამო, იღებს დადებით მუხტს. ეს განმარტავს, თუ რატომ იტენება ნეირონის გარე უჯრედის მემბრანა, რაც იწვევს ნერვული იმპულსების გამტარობას, რომლებიც საფუძვლად უდევს აგზნების პროცესს.
