5. មុខងារបទប្បញ្ញត្តិ. ប្រូតេអ៊ីនអនុវត្តមុខងារនៃសារធាតុផ្តល់សញ្ញា - អរម៉ូនមួយចំនួន អ៊ីស្តូអ័រម៉ូន និងសារធាតុបញ្ជូនសរសៃប្រសាទ គឺជាអ្នកទទួលសម្រាប់សញ្ញានៃសារធាតុនៃរចនាសម្ព័ន្ធណាមួយ និងធានានូវការបញ្ជូនសញ្ញាបន្ថែមទៀតនៅក្នុងខ្សែសង្វាក់សញ្ញាជីវគីមីនៃកោសិកា។ ឧទាហរណ៏រួមមានអ័រម៉ូនលូតលាស់ somatotropin អរម៉ូនអាំងស៊ុយលីន H- និង M-cholinergic receptors ។
6. មុខងារម៉ូទ័រ. ដោយមានជំនួយពីប្រូតេអ៊ីនដំណើរការនៃការកន្ត្រាក់និងចលនាជីវសាស្រ្តផ្សេងទៀតត្រូវបានអនុវត្ត។ ឧទាហរណ៏រួមមាន tubulin, actin និង myosin ។
7. មុខងារទំនេរ. រុក្ខជាតិមានប្រូតេអ៊ីនបម្រុង ដែលជាសារធាតុចិញ្ចឹមដ៏មានតម្លៃនៅក្នុងរាងកាយសត្វ ប្រូតេអ៊ីនសាច់ដុំបម្រើជាសារធាតុចិញ្ចឹមបម្រុងដែលត្រូវបានប្រមូលផ្តុំនៅពេលចាំបាច់។
ប្រូតេអ៊ីនត្រូវបានកំណត់លក្ខណៈដោយវត្តមាននៃកម្រិតជាច្រើននៃអង្គការរចនាសម្ព័ន្ធ។
រចនាសម្ព័ន្ធបឋមប្រូតេអ៊ីនគឺជាលំដាប់នៃសំណល់អាស៊ីតអាមីណូនៅក្នុងខ្សែសង្វាក់ polypeptide ។ ចំណង peptide គឺជាចំណង carboxamide រវាងក្រុមα-carboxyl នៃអាស៊ីតអាមីណូមួយ និងក្រុមα-amino នៃអាស៊ីតអាមីណូមួយទៀត។
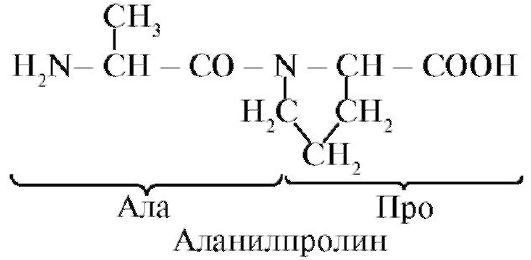
alanylphenylalanylcysteylproline
U ទំ ចំណង eptideមានលក្ខណៈពិសេសជាច្រើន៖
ក) វាមានស្ថេរភាពខ្លាំង ហើយដូច្នេះវាមានទីតាំងនៅក្នុងយន្តហោះតែមួយ - planar; ការបង្វិលជុំវិញចំណង C-N ត្រូវការថាមពលច្រើន ហើយពិបាក។
ខ) ចំណង -CO-NH- មានតួអក្សរពិសេស វាតូចជាងធម្មតា ប៉ុន្តែធំជាងទ្វេ ពោលគឺមាន keto-enol tautomerism៖

គ) សារធាតុជំនួសដែលទាក់ទងទៅនឹងចំណង peptide គឺនៅក្នុង ត្រេកត្រអាល។-ទីតាំង;
ឃ) ឆ្អឹងខ្នង peptide ត្រូវបានហ៊ុំព័ទ្ធដោយខ្សែសង្វាក់ចំហៀងនៃធម្មជាតិផ្សេងៗ អន្តរកម្មជាមួយម៉ូលេគុលសារធាតុរំលាយជុំវិញនោះ ក្រុម carboxyl សេរី និងអាមីណូត្រូវបាន ionized បង្កើតជាមជ្ឈមណ្ឌល cationic និង anionic នៃម៉ូលេគុលប្រូតេអ៊ីន។ អាស្រ័យលើសមាមាត្ររបស់វា ម៉ូលេគុលប្រូតេអ៊ីនទទួលបានបន្ទុកវិជ្ជមាន ឬអវិជ្ជមានសរុប ហើយក៏ត្រូវបានកំណត់លក្ខណៈដោយតម្លៃ pH មួយឬផ្សេងទៀតនៃបរិស្ថាននៅពេលដែលវាឈានដល់ចំណុច isoelectric នៃប្រូតេអ៊ីន។ រ៉ាឌីកាល់បង្កើតជាស្ពានអំបិល អេធើរ និង disulfide នៅខាងក្នុងម៉ូលេគុលប្រូតេអ៊ីន ហើយក៏កំណត់ជួរនៃប្រតិកម្មលក្ខណៈនៃប្រូតេអ៊ីនផងដែរ។
បច្ចុប្បន្នយល់ព្រមពិចារណាប៉ូលីមែរដែលមានសំណល់អាស៊ីតអាមីណូ 100 ឬច្រើនជាងនេះ ជាប្រូតេអ៊ីន ប៉ូលីភីបទីត - ប៉ូលីមែរដែលមានសំណល់អាស៊ីតអាមីណូពី 50-100 អាស៊ីតអាមីណូ peptides ទម្ងន់ម៉ូលេគុលទាប - ប៉ូលីមែរដែលមានសំណល់អាស៊ីតអាមីណូតិចជាង 50 ។
ខ្លះ ទំងន់ម៉ូលេគុលទាប peptides ដើរតួនាទីជីវសាស្រ្តឯករាជ្យ។ ឧទាហរណ៍មួយចំនួននៃ peptides ទាំងនេះ៖
Glutathione - γ-glu-cis-gly - មួយ។មួយនៃ peptides intracellular រីករាលដាលបំផុត ចូលរួមក្នុងដំណើរការ redox នៅក្នុងកោសិកា និងការផ្ទេរអាស៊ីតអាមីណូឆ្លងកាត់ភ្នាសជីវសាស្រ្ត។
Carnosine - β-ala-his - peptide,ដែលមាននៅក្នុងសាច់ដុំរបស់សត្វ, លុបបំបាត់ផលិតផលនៃការបំបែក lipid peroxide, ពន្លឿនដំណើរការនៃការបំបែកកាបូអ៊ីដ្រាតនៅក្នុងសាច់ដុំនិងត្រូវបានចូលរួមនៅក្នុងការរំលាយអាហារថាមពលនៅក្នុងសាច់ដុំក្នុងទម្រង់ជាផូស្វ័រ។
Vasopressin គឺជាអរម៉ូននៃក្រពេញក្រោយនៃក្រពេញ pituitary ដែលពាក់ព័ន្ធនឹងការគ្រប់គ្រងការរំលាយអាហារទឹកនៅក្នុងរាងកាយ៖

ថ្នាំ Phalloidin- សារធាតុពុលរុយ agaric polypeptide ក្នុងកំហាប់ធ្វេសប្រហែសបណ្តាលឱ្យរាងកាយស្លាប់ដោយសារការបញ្ចេញអង់ស៊ីមនិងប៉ូតាស្យូមអ៊ីយ៉ុងពីកោសិកា៖

ថ្នាំ Gramicidin - អង់ទីប៊ីយ៉ូទិកធ្វើសកម្មភាពលើបាក់តេរីក្រាមវិជ្ជមានជាច្រើន ផ្លាស់ប្តូរភាពជ្រាបចូលនៃភ្នាសជីវសាស្រ្តសម្រាប់សមាសធាតុទម្ងន់ម៉ូលេគុលទាប និងបណ្តាលឱ្យស្លាប់កោសិកា៖

ម៉េត-enkephalin - tyr-gly-gly-phen-met - peptide សំយោគនៅក្នុងសរសៃប្រសាទនិងកាត់បន្ថយការឈឺចាប់។
រចនាសម្ព័ន្ធទីពីរនៃប្រូតេអ៊ីនគឺជារចនាសម្ព័ន្ធលំហដែលបង្កើតឡើងជាលទ្ធផលនៃអន្តរកម្មរវាងក្រុមមុខងារនៃឆ្អឹងខ្នង peptide ។
ខ្សែសង្វាក់ peptide មានក្រុម CO និង NH ជាច្រើននៃចំណង peptide ដែលនីមួយៗមានសក្តានុពលក្នុងការចូលរួមក្នុងការបង្កើតចំណងអ៊ីដ្រូសែន។ មានរចនាសម្ព័ន្ធសំខាន់ៗពីរប្រភេទដែលអនុញ្ញាតឱ្យវាកើតឡើង៖ α-helix ដែលខ្សែសង្វាក់ត្រូវបានចងដូចខ្សែទូរស័ព្ទ និងរចនាសម្ព័ន្ធ β-បត់ ដែលផ្នែកពន្លូតនៃខ្សែសង្វាក់មួយឬច្រើនត្រូវបានដាក់នៅម្ខាង។ ចំហៀង។ រចនាសម្ព័ន្ធទាំងពីរនេះមានស្ថេរភាពណាស់។
α-helix ត្រូវបានកំណត់លក្ខណៈដោយការវេចខ្ចប់ក្រាស់បំផុតនៃខ្សែសង្វាក់ polypeptide រមួលសម្រាប់វេននីមួយៗនៃ helix ខាងស្តាំមានសំណល់អាស៊ីតអាមីណូ 3.6 ដែលជារ៉ាឌីកាល់ដែលតែងតែដឹកនាំទៅខាងក្រៅ និងថយក្រោយបន្តិច ពោលគឺទៅកាន់ការចាប់ផ្តើមនៃខ្សែសង្វាក់ polypeptide ។
លក្ខណៈសំខាន់ៗនៃ α-helix៖
1) α-helix ត្រូវបានស្ថេរភាពដោយចំណងអ៊ីដ្រូសែនរវាងអាតូមអ៊ីដ្រូសែននៅអាសូតនៃក្រុម peptide និងកាបូនអ៊ីដ្រាតអុកស៊ីហ៊្សែននៃសំណល់ដែលមានទីតាំងបួននៅតាមបណ្តោយខ្សែសង្វាក់;
2) ក្រុម peptide ទាំងអស់ចូលរួមក្នុងការបង្កើតចំណងអ៊ីដ្រូសែននេះធានានូវស្ថេរភាពអតិបរមានៃα-helix;
3) អាតូមអាសូតនិងអុកស៊ីហ៊្សែនទាំងអស់នៃក្រុម peptide ត្រូវបានចូលរួមនៅក្នុងការបង្កើតចំណងអ៊ីដ្រូសែនដែលកាត់បន្ថយយ៉ាងខ្លាំងនូវ hydrophilicity នៃតំបន់α-helical និងបង្កើន hydrophobicity របស់ពួកគេ;
4) α-helix ត្រូវបានបង្កើតឡើងដោយឯកឯងនិងជាការអនុលោមតាមស្ថេរភាពបំផុតនៃខ្សែសង្វាក់ polypeptide ដែលត្រូវគ្នាទៅនឹងថាមពលឥតគិតថ្លៃអប្បបរមា;
5) នៅក្នុងខ្សែសង្វាក់ polypeptide នៃអាស៊ីត L-amino, helix ដៃស្តាំ, ជាធម្មតាត្រូវបានរកឃើញនៅក្នុងប្រូតេអ៊ីន, មានស្ថេរភាពជាងអ្នកឆ្វេង។
លទ្ធភាពនៃការបង្កើត α-helixកំណត់ដោយរចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីន។ អាស៊ីតអាមីណូមួយចំនួនរារាំងឆ្អឹងខ្នង peptide ពីការបង្វិល។ ឧទាហរណ៍ ក្រុម carboxyl ដែលនៅជាប់គ្នានៃ glutamate និង aspartate ច្រានគ្នាទៅវិញទៅមក ដែលការពារការបង្កើតចំណងអ៊ីដ្រូសែននៅក្នុង α-helix ។ សម្រាប់ហេតុផលដូចគ្នា ការបង្កើតខ្សែសង្វាក់គឺពិបាកនៅកន្លែងដែលសំណល់ lysine និង arginine ដែលត្រូវបានចោទប្រកាន់ជាវិជ្ជមានមានទីតាំងនៅជិតគ្នាទៅវិញទៅមក។ ទោះជាយ៉ាងណាក៏ដោយ proline ដើរតួយ៉ាងធំបំផុតក្នុងការរំខាន α-helix ។ ទីមួយនៅក្នុង proline អាតូមអាសូតគឺជាផ្នែកមួយនៃចិញ្ចៀនរឹងដែលការពារការបង្វិលជុំវិញចំណង N-C ហើយទីពីរ proline មិនបង្កើតជាចំណងអ៊ីដ្រូសែនដោយសារតែអវត្តមាននៃអ៊ីដ្រូសែននៅអាតូមអាសូត។
β-សន្លឹកគឺជារចនាសម្ព័ន្ធស្រទាប់បង្កើតឡើងដោយចំណងអ៊ីដ្រូសែនរវាងបំណែក peptide ដែលរៀបចំតាមលីនេអ៊ែរ។ សង្វាក់ទាំងពីរអាចឯករាជ្យ ឬជាកម្មសិទ្ធិរបស់ម៉ូលេគុល polypeptide ដូចគ្នា។ ប្រសិនបើច្រវាក់ត្រូវបានតម្រង់ទិសក្នុងទិសដៅដូចគ្នានោះ រចនាសម្ព័ន្ធ β បែបនេះត្រូវបានគេហៅថាប៉ារ៉ាឡែល។ ក្នុងករណីនៃទិសដៅច្រវ៉ាក់ផ្ទុយ នោះគឺនៅពេលដែល N-terminus នៃខ្សែសង្វាក់មួយស្របគ្នាជាមួយនឹង C-terminus នៃខ្សែសង្វាក់មួយទៀត រចនាសម្ព័ន្ធ β ត្រូវបានគេហៅថា antiparallel ។ សន្លឹកប្រឆាំងប៉ារ៉ាឡែលដែលមានស្ពានអ៊ីដ្រូសែនស្ទើរតែលីនេអ៊ែរគឺមានភាពស្វាហាប់ជាង។


ប៉ារ៉ាឡែល β-សន្លឹក antiparallel β-សន្លឹក
មិនដូច α-helixឆ្អែតជាមួយនឹងចំណងអ៊ីដ្រូសែន ផ្នែកនីមួយៗនៃខ្សែសង្វាក់ β-សន្លឹកគឺបើកចំហសម្រាប់ការបង្កើតចំណងអ៊ីដ្រូសែនបន្ថែម។ រ៉ាឌីកាល់ចំហៀងនៃអាស៊ីតអាមីណូត្រូវបានតម្រង់ទិសស្ទើរតែកាត់កែងទៅនឹងប្លង់នៃសន្លឹក ឆ្លាស់គ្នាឡើងលើចុះក្រោម។
នៅក្នុងតំបន់ទាំងនោះដែលជាខ្សែសង្វាក់ peptideពត់យ៉ាងខ្លាំង ជាញឹកញាប់មាន β-loop ។ នេះគឺជាបំណែកខ្លីមួយ ដែលសំណល់អាស៊ីតអាមីណូចំនួន 4 ត្រូវបានបត់ដោយ 180° ហើយត្រូវបានរក្សាលំនឹងដោយស្ពានអ៊ីដ្រូសែនមួយរវាងសំណល់ទីមួយ និងទីបួន។ រ៉ាឌីកាល់អាស៊ីតអាមីណូដ៏ធំរំខានដល់ការបង្កើត β-loop ដូច្នេះវាច្រើនតែរួមបញ្ចូលអាស៊ីតអាមីណូតូចបំផុត glycine ។
រចនាសម្ព័ន្ធអនុវិទ្យាល័យប្រូតេអ៊ីន- នេះគឺជាលំដាប់ជាក់លាក់មួយចំនួននៃការជំនួសនៃរចនាសម្ព័ន្ធបន្ទាប់បន្សំ។ ដែនមួយត្រូវបានយល់ថាជាផ្នែកដាច់ដោយឡែកនៃម៉ូលេគុលប្រូតេអ៊ីនដែលមានកម្រិតជាក់លាក់នៃស្វ័យភាពនៃរចនាសម្ព័ន្ធ និងមុខងារ។ ឥឡូវនេះ ដែនត្រូវបានចាត់ទុកថាជាធាតុគ្រឹះនៃរចនាសម្ព័ន្ធនៃម៉ូលេគុលប្រូតេអ៊ីន ហើយទំនាក់ទំនង និងធម្មជាតិនៃការរៀបចំនៃ α-helices និង β-sheets ផ្តល់បន្ថែមសម្រាប់ការយល់ដឹងអំពីការវិវត្តន៍នៃម៉ូលេគុលប្រូតេអ៊ីន និងទំនាក់ទំនង phylogenetic ជាងការប្រៀបធៀបនៃរចនាសម្ព័ន្ធបឋម។
ភារកិច្ចចម្បងនៃការវិវត្តន៍គឺការរចនាប្រូតេអ៊ីនថ្មីកាន់តែច្រើនឡើង។ មានឱកាសគ្មានដែនកំណត់នៃការសំយោគដោយចៃដន្យនូវលំដាប់អាស៊ីតអាមីណូដែលនឹងបំពេញលក្ខខណ្ឌនៃការវេចខ្ចប់ និងធានានូវការបំពេញមុខងារមុខងារ។ ដូច្នេះវាជារឿងធម្មតាក្នុងការស្វែងរកប្រូតេអ៊ីនដែលមានមុខងារខុសៗគ្នា ប៉ុន្តែមានរចនាសម្ព័ន្ធស្រដៀងគ្នា ដែលពួកវាហាក់ដូចជាមានបុព្វបុរសធម្មតា ឬមានការវិវត្តពីគ្នាទៅវិញទៅមក។ វាហាក់បីដូចជាការវិវត្តន៍ នៅពេលដែលប្រឈមមុខនឹងតម្រូវការក្នុងការដោះស្រាយបញ្ហាជាក់លាក់មួយ ចូលចិត្តមិនរចនាប្រូតេអ៊ីនសម្រាប់គោលបំណងនេះតាំងពីដំបូងឡើយ ប៉ុន្តែដើម្បីសម្របរចនាសម្ព័ន្ធដែលបានបង្កើតឡើងយ៉ាងល្អសម្រាប់គោលបំណងនេះ ដោយសម្របខ្លួនវាសម្រាប់គោលបំណងថ្មី។
ឧទាហរណ៍មួយចំនួននៃរចនាសម្ព័ន្ធ suprasecondary ម្តងហើយម្តងទៀត:
1) αα' - ប្រូតេអ៊ីនដែលមានតែα-helices (myoglobin, hemoglobin);
2) ββ' - ប្រូតេអ៊ីនដែលមានតែរចនាសម្ព័ន្ធβ (immunoglobulins, superoxide dismutase);
3) βαβ' - រចនាសម្ព័ន្ធ β-barrel ស្រទាប់ β នីមួយៗមានទីតាំងនៅខាងក្នុងធុង ហើយត្រូវបានភ្ជាប់ទៅនឹងα-helix ដែលមានទីតាំងនៅលើផ្ទៃនៃម៉ូលេគុល (triose phosphoisomerase, lactate dehydrogenase);
4) "ម្រាមដៃស័ង្កសី" - បំណែកប្រូតេអ៊ីនដែលមានសំណល់អាស៊ីតអាមីណូចំនួន 20 អាតូមស័ង្កសីត្រូវបានភ្ជាប់ទៅនឹងសំណល់ cysteine ពីរនិងសំណល់ histidine ពីរដែលបណ្តាលឱ្យមានការបង្កើត "ម្រាមដៃ" នៃសំណល់អាស៊ីតអាមីណូប្រហែល 12 ដែល អាចភ្ជាប់ទៅនឹងតំបន់បទប្បញ្ញត្តិនៃម៉ូលេគុល DNA;
5) "leucine zipper" - ប្រូតេអ៊ីនអន្តរកម្មមានតំបន់ α-helical ដែលមានសំណល់ leucine យ៉ាងហោចណាស់ 4 ពួកវាស្ថិតនៅ 6 អាស៊ីតអាមីណូដាច់ពីគ្នា ពោលគឺពួកវាស្ថិតនៅលើផ្ទៃនៃរាល់វេនទីពីរ ហើយអាចបង្កើតចំណង hydrophobic ជាមួយនឹងសំណល់ leucine ។ ប្រូតេអ៊ីនមួយទៀត។ ជាឧទាហរណ៍ ដោយមានជំនួយពីខ្សែរ៉ូត leucine ម៉ូលេគុលនៃប្រូតេអ៊ីនអ៊ីស្តូនមូលដ្ឋានខ្លាំងអាចស្មុគស្មាញ ដោយយកឈ្នះលើបន្ទុកវិជ្ជមាន។
រចនាសម្ព័ន្ធទីបីនៃប្រូតេអ៊ីន- នេះគឺជាការរៀបចំលំហនៃម៉ូលេគុលប្រូតេអ៊ីន មានស្ថេរភាពដោយចំណងរវាងរ៉ាឌីកាល់ចំហៀងនៃអាស៊ីតអាមីណូ។
ប្រភេទនៃចំណងដែលរក្សាស្ថេរភាពរចនាសម្ព័ន្ធទីបីនៃប្រូតេអ៊ីន៖

អន្តរកម្មអ៊ីដ្រូសែន អ៊ីដ្រូសែន អ៊ីដ្រូសែន ឌីស៊ុលហ្វីត អេឡិចត្រូស្ទិក ភ្ជាប់ចំណងអន្តរកម្ម
អាស្រ័យលើការបត់រចនាសម្ព័ន្ធទីបីនៃប្រូតេអ៊ីនអាចត្រូវបានចាត់ថ្នាក់ជាពីរប្រភេទសំខាន់ - fibrillar និង globular ។
ប្រូតេអ៊ីន Fibrillar- ម៉ូលេគុលដ៏វែងដូចខ្សែស្រលាយមិនរលាយក្នុងទឹក ខ្សែសង្វាក់ polypeptide ដែលត្រូវបានពន្លូតតាមអ័ក្សមួយ។ ទាំងនេះជាចម្បង រចនាសម្ព័ន្ធ និងប្រូតេអ៊ីន។ ឧទាហរណ៍មួយចំនួននៃប្រូតេអ៊ីន fibrillar ទូទៅបំផុត:
1. α-ខេរ៉ាទីន។ សំយោគដោយកោសិកាអេពីដេមី។ ពួកវាមានទំងន់ស្ទើរតែទាំងអស់នៃសក់ រោម រោម ស្នែង ក្រចក ក្រញ៉ាំជើង ជញ្ជីង ចប និងសំបកអណ្តើក ក៏ដូចជាផ្នែកសំខាន់នៃទម្ងន់នៃស្រទាប់ខាងក្រៅនៃស្បែក។ នេះគឺជាក្រុមគ្រួសារទាំងមូលនៃប្រូតេអ៊ីន; ពួកគេគឺស្រដៀងគ្នានៅក្នុងសមាសភាពអាស៊ីតអាមីណូ, មានសំណល់ cysteine ជាច្រើននិងមានការរៀបចំ spatial ដូចគ្នានៃខ្សែសង្វាក់ polypeptide ។
នៅក្នុងកោសិកាសក់ខ្សែសង្វាក់ polypeptide នៃ keratinត្រូវបានរៀបចំជាលើកដំបូងទៅជាសរសៃ ដែលរចនាសម្ព័ន្ធទាំងនោះត្រូវបានបង្កើតឡើងដូចជាខ្សែពួរ ឬខ្សែរមួល ហើយទីបំផុតបំពេញចន្លោះទាំងមូលនៃក្រឡា។ កោសិកាសក់ប្រែជាសំប៉ែត ហើយចុងក្រោយស្លាប់ ហើយជញ្ជាំងកោសិកាបង្កើតជាបំពង់ដែលហៅថា cuticle នៅជុំវិញសក់នីមួយៗ។ នៅក្នុង α-keratin ខ្សែសង្វាក់ polypeptide មានរាងជា α-helix ដែលបត់ជុំវិញគ្នាទៅវិញទៅមកចូលទៅក្នុងខ្សែបីស្នូលជាមួយនឹងការបង្កើតចំណងឆ្លង disulfide ។
សំណល់ស្ថានីយ N មានទីតាំងនៅនៅម្ខាង (ស្របគ្នា) ។ Keratin គឺមិនរលាយក្នុងទឹកដោយសារតែភាពលេចធ្លោនៃអាស៊ីតអាមីណូនៅក្នុងសមាសភាពរបស់វាជាមួយនឹងរ៉ាឌីកាល់ចំហៀងដែលមិនមែនជាប៉ូលដែលប្រឈមមុខនឹងដំណាក់កាល aqueous ។ ក្នុងអំឡុងពេល perm ដំណើរការដូចខាងក្រោមកើតឡើង: ដំបូងស្ពាន disulfide ត្រូវបានបំផ្លាញដោយការកាត់បន្ថយជាមួយនឹង thiols ហើយបន្ទាប់មកនៅពេលដែលសក់ត្រូវបានផ្តល់រូបរាងដែលត្រូវការវាត្រូវបានស្ងួតដោយកំដៅខណៈពេលដែលដោយសារតែការកត់សុីជាមួយនឹងអុកស៊ីសែនបរិយាកាសស្ពាន disulfide ថ្មីត្រូវបានបង្កើតឡើង។ ដែលរក្សារូបរាងស្ទីលម៉ូដសក់។
2. β-Keratins. ទាំងនេះរួមមានសរសៃសូត្រ និងសរសៃពីងពាង។ ពួកវាជាស្រទាប់ប្រឆាំងប៉ារ៉ាឡែល β-pleated ជាមួយនឹងភាពលេចធ្លោនៃ glycine, alanine និង serine នៅក្នុងសមាសភាព។
3. ខូឡាជេន។ ប្រូតេអ៊ីនទូទៅបំផុតនៅក្នុងសត្វខ្ពស់និងប្រូតេអ៊ីន fibrillar សំខាន់នៃជាលិកាភ្ជាប់។ ខូឡាជេនត្រូវបានសំយោគនៅក្នុង fibroblasts និង chondrocytes - កោសិកាជាលិកាភ្ជាប់ឯកទេសដែលបន្ទាប់មកវាត្រូវបានបណ្តេញចេញ។ សរសៃ Collagen មាននៅក្នុងស្បែក សរសៃពួរ ឆ្អឹងខ្ចី និងឆ្អឹង។ ពួកវាមិនលាតសន្ធឹងទេគឺខ្លាំងជាងខ្សែដែក ហើយសរសៃ collagen ត្រូវបានកំណត់លក្ខណៈដោយខ្សែឆ្លងកាត់។
ពេលស្ងោរក្នុងទឹកមានសរសៃ Collagen ដែលមិនអាចរំលាយបាន និងមិនអាចរំលាយបានត្រូវបានបំប្លែងទៅជា gelatin ដោយ hydrolysis នៃចំណង covalent មួយចំនួន។ ខូឡាជេនមាន 35% glycine, 11% alanine, 21% proline និង 4-hydroxyproline (អាស៊ីតអាមីណូតែមួយគត់សម្រាប់ collagen និង elastin) ។ សមាសភាពនេះកំណត់តម្លៃអាហារូបត្ថម្ភទាបនៃ gelatin ជាប្រូតេអ៊ីនអាហារ។ Collagen fibrils ត្រូវបានផ្សំឡើងដោយអនុក្រុម polypeptide ដែលត្រូវបានគេហៅថា tropocollagen ។ អនុក្រុមទាំងនេះត្រូវបានរៀបចំតាមសរសៃក្នុងទម្រង់ជាបាច់ប៉ារ៉ាឡែលក្នុងទម្រង់ក្បាលទៅកន្ទុយ។ ការផ្លាស់ទីលំនៅនៃក្បាលផ្តល់នូវលក្ខណៈឆ្លងកាត់ striations ។ ការចាត់ទុកជាមោឃៈនៅក្នុងរចនាសម្ព័ន្ធនេះ បើចាំបាច់អាចបម្រើជាកន្លែងសម្រាប់ដាក់គ្រីស្តាល់នៃ hydroxyapatite Ca 5 (OH)(PO 4) 3 ដែលដើរតួនាទីយ៉ាងសំខាន់ក្នុងការជីកយករ៉ែឆ្អឹង។


អនុក្រុម Tropocollagen រួមមាននៃខ្សែសង្វាក់ polypeptide បីត្រូវបានចងយ៉ាងតឹងចូលទៅក្នុងខ្សែពួរបីដែលខុសពី α- និង β-keratins ។ នៅក្នុង collagens មួយចំនួន ខ្សែសង្វាក់ទាំងបីមានលំដាប់អាស៊ីតអាមីណូដូចគ្នា ខណៈពេលដែលខ្សែសង្វាក់ផ្សេងទៀតមានតែពីរខ្សែដូចគ្នា ហើយខ្សែទីបីគឺខុសគ្នា។ ខ្សែសង្វាក់ polypeptide នៃ tropocollagen បង្កើតបានជា helix ដៃឆ្វេង ដោយមានសំណល់អាស៊ីតអាមីណូបីប៉ុណ្ណោះក្នុងមួយវេន ដោយសារការពត់ខ្សែសង្វាក់ដែលបណ្តាលមកពី proline និង hydroxyproline ។ ខ្សែសង្វាក់ទាំងបីត្រូវបានភ្ជាប់ទៅគ្នាទៅវិញទៅមក បន្ថែមពីលើចំណងអ៊ីដ្រូសែន ដោយចំណងប្រភេទ covalent ដែលបង្កើតឡើងរវាងសំណល់ lysine ពីរដែលស្ថិតនៅក្នុងខ្សែសង្វាក់ជាប់គ្នា៖

នៅពេលដែលយើងកាន់តែចាស់ការកើនឡើងចំនួននៃតំណភ្ជាប់ឆ្លងកាត់ត្រូវបានបង្កើតឡើងនៅក្នុង និងរវាងផ្នែករង tropocollagen ដែលធ្វើឱ្យសរសៃ collagen កាន់តែរឹង និងផុយ ហើយនេះផ្លាស់ប្តូរលក្ខណៈសម្បត្តិមេកានិចនៃឆ្អឹងខ្ចី និងសរសៃពួរ ធ្វើឱ្យឆ្អឹងកាន់តែផុយ និងកាត់បន្ថយភាពថ្លានៃកញ្ចក់ភ្នែក។
4. អ៊ីឡាស្ទីន។ មាននៅក្នុងជាលិកាយឺតពណ៌លឿងនៃសរសៃចងនិងស្រទាប់យឺតនៃជាលិកាភ្ជាប់នៅក្នុងជញ្ជាំងនៃសរសៃឈាមធំ។ ផ្នែករងសំខាន់នៃសរសៃ elastin គឺ tropoelastin ។ Elastin សម្បូរទៅដោយ glycine និង alanine មានផ្ទុក lysine ច្រើន និង proline តិចតួច។ ផ្នែកតំរៀបស្លឹកនៃ elastin លាតសន្ធឹងនៅពេលដែលភាពតានតឹងត្រូវបានអនុវត្តប៉ុន្តែត្រលប់ទៅប្រវែងដើមវិញនៅពេលដែលបន្ទុកត្រូវបានដកចេញ។ សំណល់ lysine នៃខ្សែសង្វាក់បួនផ្សេងគ្នាបង្កើតជាចំណង covalent ជាមួយគ្នា ហើយអនុញ្ញាតឱ្យ elastin លាតសន្ធឹងបញ្ច្រាស់គ្រប់ទិសទី។
ប្រូតេអ៊ីន Globular- ប្រូតេអ៊ីន ដែលជាខ្សែសង្វាក់ polypeptide ដែលត្រូវបានបត់ចូលទៅក្នុងរាងពងក្រពើ មានសមត្ថភាពបំពេញមុខងារជាច្រើនប្រភេទ។
រចនាសម្ព័ន្ធទីបីនៃប្រូតេអ៊ីន globularវាងាយស្រួលបំផុតក្នុងការពិចារណាដោយប្រើឧទាហរណ៍នៃ myoglobin ។ Myoglobin គឺជាប្រូតេអ៊ីនដែលភ្ជាប់អុកស៊ីហ្សែនតិចតួចដែលមាននៅក្នុងកោសិកាសាច់ដុំ។ វាផ្ទុកអុកស៊ីហ្សែនដែលចងភ្ជាប់ និងជំរុញការផ្ទេររបស់វាទៅ mitochondria ។ ម៉ូលេគុល myoglobin មានខ្សែសង្វាក់ polypeptide មួយ និងក្រុម hemogroup (heme) - ស្មុគស្មាញនៃ protoporphyrin ដែលមានជាតិដែក។
លក្ខណៈសម្បត្តិមូលដ្ឋាន myoglobin:
ក) ម៉ូលេគុល myoglobin គឺបង្រួមខ្លាំង ដែលមានតែម៉ូលេគុលទឹកចំនួន 4 ប៉ុណ្ណោះដែលអាចដាក់នៅខាងក្នុងវាបាន។
ខ) សំណល់អាស៊ីដអាមីណូប៉ូឡាទាំងអស់ លើកលែងតែពីរ មានទីតាំងនៅលើផ្ទៃខាងក្រៅនៃម៉ូលេគុល ហើយពួកវាទាំងអស់ស្ថិតនៅក្នុងស្ថានភាពដែលមានជាតិសំណើម។
គ) ភាគច្រើននៃសំណល់អាស៊ីតអាមីណូ hydrophobic មានទីតាំងនៅខាងក្នុងម៉ូលេគុល myoglobin ហើយដូច្នេះត្រូវបានការពារពីការប៉ះពាល់ជាមួយទឹក។
ឃ) សំណល់ proline នីមួយៗនៅក្នុងម៉ូលេគុល myoglobin មានទីតាំងនៅកន្លែងពត់នៃខ្សែសង្វាក់ polypeptide serine, threonine និង asparagine មានទីតាំងនៅកន្លែងពត់ផ្សេងទៀត ដោយសារអាស៊ីតអាមីណូបែបនេះការពារការបង្កើត α-helix ប្រសិនបើពួកវា។ មានទីតាំងនៅជាប់គ្នា;
ង) ក្រុម heme រាបស្មើស្ថិតនៅក្នុងបែហោងធ្មែញ (ហោប៉ៅ) នៅជិតផ្ទៃនៃម៉ូលេគុលអាតូមដែកមានចំណងសំរបសំរួលពីរដែលដឹកនាំកាត់កែងទៅនឹងយន្តហោះ heme មួយក្នុងចំណោមពួកវាត្រូវបានភ្ជាប់ទៅនឹងសំណល់អ៊ីស្ទីឌីន 93 និងមួយទៀតបម្រើដើម្បីចង។ ម៉ូលេគុលអុកស៊ីសែន។
ចាប់ផ្តើមជាមួយនឹងរចនាសម្ព័ន្ធទីបីនៃប្រូតេអ៊ីនក្លាយជាសមត្ថភាពក្នុងការអនុវត្តមុខងារជីវសាស្រ្តដែលមានដើមរបស់វា។ មូលដ្ឋាននៃដំណើរការនៃប្រូតេអ៊ីនគឺថានៅពេលដែលរចនាសម្ព័ន្ធទីបីត្រូវបានដាក់នៅលើផ្ទៃនៃប្រូតេអ៊ីននោះតំបន់ត្រូវបានបង្កើតឡើងដែលអាចភ្ជាប់ម៉ូលេគុលផ្សេងទៀតដែលហៅថា ligands ។ ភាពជាក់លាក់ខ្ពស់នៃអន្តរកម្មនៃប្រូតេអ៊ីនជាមួយ ligand ត្រូវបានធានាដោយការបំពេញបន្ថែមនៃរចនាសម្ព័ន្ធនៃមជ្ឈមណ្ឌលសកម្មទៅនឹងរចនាសម្ព័ន្ធនៃ ligand នេះ។ ការបំពេញបន្ថែមគឺជាការឆ្លើយឆ្លងតាមលំហ និងគីមីនៃផ្ទៃអន្តរកម្ម។ សម្រាប់ប្រូតេអ៊ីនភាគច្រើនរចនាសម្ព័ន្ធទីបីគឺជាកម្រិតអតិបរមានៃការបត់។
រចនាសម្ព័ន្ធប្រូតេអ៊ីន Quaternary- លក្ខណៈនៃប្រូតេអ៊ីនដែលមានខ្សែសង្វាក់ polypeptide ពីរ ឬច្រើនដែលតភ្ជាប់គ្នាទៅវិញទៅមកដោយចំណងដែលមិនមែនជាកូវ៉ាលេន ភាគច្រើនជាអេឡិចត្រូស្តាត និងអ៊ីដ្រូសែន។ ភាគច្រើនជាញឹកញាប់ ប្រូតេអ៊ីនមានអនុរងពីរ ឬបួន ច្រើនជាង បួនរង ជាធម្មតាមានប្រូតេអ៊ីន។
ប្រូតេអ៊ីនដែលមានរចនាសម្ព័ន្ធ quaternaryជារឿយៗត្រូវបានគេហៅថា oligomeric ។ មានប្រូតេអ៊ីន homomeric និង heteromeric ។ ប្រូតេអ៊ីន Homomeric រួមមានប្រូតេអ៊ីនដែលផ្នែករងទាំងអស់មានរចនាសម្ព័ន្ធដូចគ្នា ឧទាហរណ៍ អង់ស៊ីម catalase មានបួនរងដូចគ្នាបេះបិទ។ ប្រូតេអ៊ីន Heteromeric មានផ្នែករងផ្សេងៗគ្នា ឧទាហរណ៍ អង់ស៊ីម RNA polymerase មានផ្នែករងផ្សេងគ្នាតាមលំដាប់ចំនួនប្រាំ ដែលបំពេញមុខងារផ្សេងៗគ្នា។
អន្តរកម្មនៃអនុឯកតាតែមួយជាមួយនឹង ligand ជាក់លាក់មួយបណ្តាលឱ្យមានការផ្លាស់ប្តូរអនុលោមភាពនៅក្នុងប្រូតេអ៊ីន oligomeric ទាំងមូល និងផ្លាស់ប្តូរភាពស្និទ្ធស្នាលនៃអនុរងផ្សេងទៀតសម្រាប់ ligands;
រចនាសម្ព័ន្ធ quaternary នៃប្រូតេអ៊ីនអាចត្រូវបានពិនិត្យដោយប្រើឧទាហរណ៍អេម៉ូក្លូប៊ីន។ មានខ្សែសង្វាក់ polypeptide ចំនួនបួន និងក្រុមសិប្បនិមិត្ត heme ចំនួនបួន ដែលក្នុងនោះអាតូមដែកស្ថិតនៅក្នុងទម្រង់ Fe 2+ ។ ផ្នែកប្រូតេអ៊ីននៃម៉ូលេគុល - globin - មាន α-chains និង β-chains ពីរដែលមានរហូតដល់ទៅ 70% α-helices ។ ខ្សែសង្វាក់នីមួយៗនៃខ្សែសង្វាក់ទាំងបួនមានរចនាសម្ព័ន្ធទីបីលក្ខណៈ ហើយក្រុមឈាមមួយត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងខ្សែសង្វាក់នីមួយៗ។ hemes នៃច្រវាក់ផ្សេងៗគ្នាមានទីតាំងនៅឆ្ងាយពីគ្នាទៅវិញទៅមក ហើយមានមុំទំនោរខុសៗគ្នា។ ទំនាក់ទំនងផ្ទាល់មួយចំនួនត្រូវបានបង្កើតឡើងរវាងខ្សែសង្វាក់α-ខ្សែសង្វាក់ពីរ និងខ្សែសង្វាក់ β-ពីរ ខណៈពេលដែលទំនាក់ទំនងជាច្រើននៃប្រភេទ α 1 β 1 និង α 2 β 2 ដែលបង្កើតឡើងដោយរ៉ាឌីកាល់ hydrophobic កើតឡើងរវាងខ្សែសង្វាក់ α និង β ។ រវាង α 1 β 1 និង α 2 β 2 ឆានែលមួយនៅសល់។
មិនដូច myoglobinអេម៉ូក្លូប៊ីន លក្ខណៈភាពស្និទ្ធស្នាលទាបសម្រាប់អុកស៊ីសែន ដែលអនុញ្ញាតឱ្យវានៅសម្ពាធផ្នែកទាបនៃអុកស៊ីសែនដែលមាននៅក្នុងជាលិកា ដើម្បីផ្តល់ឱ្យពួកគេនូវផ្នែកសំខាន់នៃអុកស៊ីសែនដែលចង។ អុកស៊ីសែនត្រូវបានចងយ៉ាងងាយស្រួលដោយជាតិដែកអេម៉ូក្លូប៊ីននៅតម្លៃ pH ខ្ពស់និងកំហាប់ CO 2 ទាប លក្ខណៈនៃ alveoli នៃសួត; ការបញ្ចេញអុកស៊ីសែនពីអេម៉ូក្លូប៊ីនត្រូវបានអនុគ្រោះដោយតម្លៃ pH ទាប និងការប្រមូលផ្តុំខ្ពស់នៃ CO 2 លក្ខណៈនៃជាលិកា។
បន្ថែមពីលើអុកស៊ីសែន អេម៉ូក្លូប៊ីនផ្ទុកអ៊ីយ៉ុងអ៊ីដ្រូសែនដែលភ្ជាប់ទៅនឹងសំណល់ histidine នៅក្នុងខ្សែសង្វាក់។ អេម៉ូក្លូប៊ីនក៏ផ្ទុកកាបូនឌីអុកស៊ីតដែលភ្ជាប់ទៅនឹងក្រុមអាមីណូស្ថានីយនៃខ្សែសង្វាក់ polypeptide នីមួយៗដែលបណ្តាលឱ្យមានការបង្កើត carbaminohemoglobin:

INកោសិកាឈាមពណ៌ក្រហម ក្នុងកំហាប់ខ្ពស់គួរសមសារធាតុ 2,3-diphosphoglycerate (DPG) មានវត្តមាន មាតិការបស់វាកើនឡើងជាមួយនឹងការកើនឡើងដល់កម្ពស់ខ្ពស់ និងអំឡុងពេល hypoxia សម្របសម្រួលការបញ្ចេញអុកស៊ីសែនពីអេម៉ូក្លូប៊ីននៅក្នុងជាលិកា។ DPG ស្ថិតនៅក្នុងឆានែលរវាង α 1 β 1 និង α 2 β 2 ធ្វើអន្តរកម្មជាមួយក្រុម β-chains ដែលកខ្វក់ជាវិជ្ជមាន។ នៅពេលដែលអេម៉ូក្លូប៊ីនភ្ជាប់អុកស៊ីសែន DPG ត្រូវបានបង្ខំឱ្យចេញពីបែហោងធ្មែញ។ កោសិកាឈាមក្រហមរបស់បក្សីមួយចំនួនមិនមាន DPG ទេប៉ុន្តែ inositol hexa-phosphate ដែលកាត់បន្ថយភាពស្និទ្ធស្នាលនៃអេម៉ូក្លូប៊ីនសម្រាប់អុកស៊ីសែន។
 2,3-Diphosphoglycerate (DPG)
2,3-Diphosphoglycerate (DPG)
HbA - អេម៉ូក្លូប៊ីនមនុស្សពេញវ័យធម្មតា។, HbF - អេម៉ូក្លូប៊ីនគភ៌ មានភាពស្និទ្ធស្នាលកាន់តែខ្លាំងចំពោះ O 2, HbS - អេម៉ូក្លូប៊ីនក្នុងភាពស្លេកស្លាំងកោសិការ។ ភាពស្លេកស្លាំងកោសិកា Sickle គឺជាជំងឺតំណពូជធ្ងន់ធ្ងរដែលបណ្តាលមកពីភាពមិនប្រក្រតីនៃហ្សែននៃអេម៉ូក្លូប៊ីន។ នៅក្នុងឈាមរបស់មនុស្សឈឺ មានចំនួនកោសិកាឈាមក្រហមស្តើងៗច្រើនខុសពីធម្មតា ដែលដំបូងគេងាយដាច់ និងទីពីរស្ទះសរសៃឈាម។
នៅកម្រិតម៉ូលេគុលអេម៉ូក្លូប៊ីនអេសគឺខុសគ្នាពីអេម៉ូក្លូប៊ីន A មានសំណល់អាស៊ីតអាមីណូមួយនៅទីតាំងទី 6 នៃខ្សែសង្វាក់ β ដែលជំនួសឱ្យសំណល់អាស៊ីត glutamic មានវ៉ាលីន។ ដូច្នេះ អេម៉ូក្លូប៊ីន S មានបន្ទុកអវិជ្ជមានតិចជាងពីរ រូបរាងនៃវ៉ាលីននាំឱ្យរូបរាងនៃទំនាក់ទំនងអ៊ីដ្រូហ្វូប "ស្អិត" នៅលើផ្ទៃនៃម៉ូលេគុល ជាលទ្ធផល ក្នុងអំឡុងពេល deoxygenation ម៉ូលេគុល deoxyhemoglobin S ជាប់គ្នា និងបង្កើតជាមិនរលាយ យូរមិនធម្មតា។ ការប្រមូលផ្តុំដូចខ្សែស្រឡាយ ដែលនាំឱ្យខូចទ្រង់ទ្រាយកោសិកាឈាមក្រហម។
មិនមានហេតុផលដើម្បីគិតថាមានការគ្រប់គ្រងហ្សែនដោយឯករាជ្យលើការបង្កើតកម្រិតនៃអង្គការរចនាសម្ព័ន្ធប្រូតេអ៊ីនខាងលើបឋមនោះទេ ចាប់តាំងពីរចនាសម្ព័ន្ធបឋមកំណត់អនុវិទ្យាល័យ ទីបី និងត្រីមាស (ប្រសិនបើមាន)។ ការអនុលោមតាមប្រភពដើមនៃប្រូតេអ៊ីនគឺជារចនាសម្ព័ន្ធដែលមានស្ថេរភាពបំផុតតាមទែម៉ូឌីណាមិកក្រោមលក្ខខណ្ឌដែលបានផ្តល់ឱ្យ។
បាឋកថា ៦
មានលក្ខណៈសម្បត្តិរាងកាយ គីមី និងជីវសាស្រ្តនៃប្រូតេអ៊ីន។
លក្ខណៈសម្បត្តិរាងកាយរបស់ប្រូតេអ៊ីនវត្តមាននៃទម្ងន់ម៉ូលេគុល birefringence (ការផ្លាស់ប្តូរលក្ខណៈអុបទិកនៃសូលុយស្យុងប្រូតេអ៊ីនក្នុងចលនាធៀបនឹងដំណោះស្រាយនៅពេលសម្រាក) ដោយសារតែរូបរាងមិនរាងស្វ៊ែរនៃប្រូតេអ៊ីន ការចល័តនៅក្នុងវាលអគ្គិសនីដោយសារតែការចោទប្រកាន់នៃម៉ូលេគុលប្រូតេអ៊ីន។ . លើសពីនេះ ប្រូតេអ៊ីនត្រូវបានកំណត់លក្ខណៈដោយលក្ខណៈសម្បត្តិអុបទិក មានសមត្ថភាពក្នុងការបង្វិលប្លង់ប៉ូលនៃពន្លឺ បញ្ចេញកាំរស្មីពន្លឺដោយសារតែទំហំធំនៃភាគល្អិតប្រូតេអ៊ីន និងស្រូបយកកាំរស្មីអ៊ុលត្រាវីយូឡេ។
មួយនៃលក្ខណៈសម្បត្តិរូបវន្តប្រូតេអ៊ីនគឺជាសមត្ថភាពក្នុងការស្រូបយកលើផ្ទៃ ហើយជួនកាលចាប់យកម៉ូលេគុល សមាសធាតុសរីរាង្គម៉ូលេគុលទាប និងអ៊ីយ៉ុងនៅខាងក្នុង។
លក្ខណៈសម្បត្តិគីមីនៃប្រូតេអ៊ីនគឺខុសគ្នាភាពចម្រុះពិសេស ចាប់តាំងពីប្រូតេអ៊ីនត្រូវបានកំណត់លក្ខណៈដោយប្រតិកម្មទាំងអស់នៃរ៉ាឌីកាល់អាស៊ីតអាមីណូ និងត្រូវបានកំណត់លក្ខណៈដោយប្រតិកម្មនៃអ៊ីដ្រូលីស៊ីនៃចំណង peptide ។
មានចំនួនសំខាន់នៃក្រុមអាស៊ីត និងមូលដ្ឋាន, ប្រូតេអ៊ីនបង្ហាញពីលក្ខណៈសម្បត្តិ amphoteric ។ មិនដូចអាស៊ីតអាមីណូឥតគិតថ្លៃ លក្ខណៈសម្បត្តិអាស៊ីតមូលដ្ឋាននៃប្រូតេអ៊ីនត្រូវបានកំណត់ដោយក្រុម α-amino និង α-carboxy ដែលពាក់ព័ន្ធនឹងការបង្កើតចំណង peptide នោះទេប៉ុន្តែដោយរ៉ាឌីកាល់ចោទប្រកាន់នៃសំណល់អាស៊ីតអាមីណូ។ លក្ខណៈសម្បត្តិសំខាន់នៃប្រូតេអ៊ីនត្រូវបានកំណត់ដោយសំណល់ arginine, lysine និង histidine ។ លក្ខណៈសម្បត្តិអាស៊ីតគឺដោយសារតែសំណល់អាស៊ីត aspartic និង glutamic ។
ខ្សែកោង titration ប្រូតេអ៊ីនគឺគ្រប់គ្រាន់ពិបាកបកស្រាយ ដោយសារប្រូតេអ៊ីនណាមួយមានក្រុម titratable ច្រើនពេក មានអន្តរកម្មអេឡិចត្រូស្ទិចរវាងក្រុមអ៊ីយ៉ូដនៃប្រូតេអ៊ីន ហើយ pK នៃក្រុម titratable នីមួយៗត្រូវបានជះឥទ្ធិពលដោយសំណល់ hydrophobic និងចំណងអ៊ីដ្រូសែននៅក្បែរនោះ។ ការអនុវត្តជាក់ស្តែងដ៏អស្ចារ្យបំផុតគឺចំណុច isoelectric នៃប្រូតេអ៊ីន - តម្លៃ pH ដែលបន្ទុកសរុបនៃប្រូតេអ៊ីនគឺសូន្យ។ នៅចំណុចអ៊ីសូអេឡិចត្រិច ប្រូតេអ៊ីនគឺអសកម្មអតិបរិមា មិនផ្លាស់ទីក្នុងវាលអគ្គីសនី និងមានសំបកជាតិទឹកស្តើងបំផុត។
ប្រូតេអ៊ីនបង្ហាញពីលក្ខណៈទប់ស្កាត់ប៉ុន្តែសមត្ថភាពសតិបណ្ដោះអាសន្នរបស់ពួកគេមិនសំខាន់ទេ។ ករណីលើកលែងគឺប្រូតេអ៊ីនដែលមានសំណល់អ៊ីស្ទីឌីនច្រើន។ ឧទាហរណ៍ អេម៉ូក្លូប៊ីនដែលមាននៅក្នុង erythrocytes ដោយសារតែមាតិកាខ្ពស់នៃសំណល់ histidine មានសមត្ថភាពផ្ទុកយ៉ាងសំខាន់នៅ pH ប្រហែល 7 ដែលមានសារៈសំខាន់ខ្លាំងណាស់សម្រាប់តួនាទីដែល erythrocytes ដើរតួក្នុងការដឹកជញ្ជូនអុកស៊ីសែន និងកាបូនឌីអុកស៊ីតនៅក្នុង ឈាម។
ប្រូតេអ៊ីនត្រូវបានកំណត់លក្ខណៈដោយការរលាយក្នុងទឹក។ហើយតាមទស្សនៈរូបវន្ត ពួកវាបង្កើតបានជាដំណោះស្រាយម៉ូលេគុលពិត។ ទោះជាយ៉ាងណាក៏ដោយដំណោះស្រាយប្រូតេអ៊ីនត្រូវបានកំណត់លក្ខណៈដោយលក្ខណៈសម្បត្តិកូឡាជែនមួយចំនួន: ឥទ្ធិពល Tendahl (បាតុភូតខ្ចាត់ខ្ចាយពន្លឺ) អសមត្ថភាពក្នុងការឆ្លងកាត់ភ្នាសពាក់កណ្តាលដែលអាចជ្រាបចូលបាន viscosity ខ្ពស់និងការបង្កើតជែល។
ការរលាយប្រូតេអ៊ីនគឺពឹងផ្អែកខ្លាំងនៅលើកំហាប់នៃអំបិល នោះគឺនៅលើកម្លាំងអ៊ីយ៉ុងនៃដំណោះស្រាយ។ នៅក្នុងទឹកចម្រោះ ប្រូតេអ៊ីនភាគច្រើនមិនសូវរលាយ ប៉ុន្តែភាពរលាយរបស់វាកើនឡើងនៅពេលដែលកម្លាំងអ៊ីយ៉ុងកើនឡើង។ ក្នុងពេលជាមួយគ្នានេះ ចំនួនកើនឡើងនៃអ៊ីយ៉ុងអសរីរាង្គដែលមានជាតិទឹក ភ្ជាប់ទៅនឹងផ្ទៃប្រូតេអ៊ីន ហើយដោយហេតុនេះកម្រិតនៃការប្រមូលផ្តុំរបស់វាថយចុះ។ នៅកម្លាំងអ៊ីយ៉ុងខ្ពស់ អ៊ីយ៉ុងអំបិលយកសែលជាតិទឹកចេញពីម៉ូលេគុលប្រូតេអ៊ីន ដែលនាំទៅដល់ការប្រមូលផ្តុំ និងទឹកភ្លៀងនៃប្រូតេអ៊ីន (បាតុភូតអំបិល)។ ដោយប្រើភាពខុសគ្នានៃការរលាយ វាអាចបំបែកល្បាយនៃប្រូតេអ៊ីនដោយប្រើអំបិលធម្មតា។
ក្នុងចំណោមលក្ខណៈសម្បត្តិជីវសាស្រ្តនៃប្រូតេអ៊ីនជាចម្បងរួមបញ្ចូលសកម្មភាពកាតាលីកររបស់ពួកគេ។ លក្ខណៈសម្បត្តិជីវសាស្រ្តដ៏សំខាន់មួយទៀតនៃប្រូតេអ៊ីនគឺសកម្មភាពអ័រម៉ូនរបស់ពួកគេ ពោលគឺសមត្ថភាពក្នុងការជះឥទ្ធិពលដល់ក្រុមប្រតិកម្មទាំងមូលនៅក្នុងរាងកាយ។ ប្រូតេអ៊ីនមួយចំនួនមានលក្ខណៈសម្បត្តិពុល សកម្មភាពបង្កជំងឺ មុខងារការពារ និងទទួល និងទទួលខុសត្រូវចំពោះបាតុភូតនៃការជាប់កោសិកា។
លក្ខណៈសម្បត្តិជីវសាស្រ្តតែមួយគត់នៃប្រូតេអ៊ីន- ការប្រែពណ៌។ ប្រូតេអ៊ីននៅក្នុងស្ថានភាពធម្មជាតិរបស់ពួកគេត្រូវបានគេហៅថាដើមកំណើត។ Denaturation គឺជាការបំផ្លិចបំផ្លាញនៃរចនាសម្ព័ន្ធ spatial នៃប្រូតេអ៊ីនក្រោមសកម្មភាពរបស់ភ្នាក់ងារ denaturing ។ រចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនមិនត្រូវបានខូចខាតក្នុងអំឡុងពេល denaturation ប៉ុន្តែសកម្មភាពជីវសាស្រ្តរបស់ពួកគេត្រូវបានបាត់បង់ ក៏ដូចជាការរលាយ ការចល័ត electrophoretic និងប្រតិកម្មមួយចំនួនផ្សេងទៀត។ នៅពេលដែល denatured រ៉ាឌីកាល់អាស៊ីតអាមីណូដែលបង្កើតជាមជ្ឈមណ្ឌលសកម្មនៃប្រូតេអ៊ីនគឺនៅឆ្ងាយពីគ្នាទៅវិញទៅមក ពោលគឺមជ្ឈមណ្ឌលភ្ជាប់ជាក់លាក់នៃប្រូតេអ៊ីនជាមួយ ligand ត្រូវបានបំផ្លាញ។ រ៉ាឌីកាល់ Hydrophobic ដែលជាធម្មតាមានទីតាំងនៅក្នុងស្នូល hydrophobic នៃប្រូតេអ៊ីន globular នៅពេលដែល denaturation បញ្ចប់នៅលើផ្ទៃនៃម៉ូលេគុល ដោយហេតុនេះបង្កើតលក្ខខណ្ឌសម្រាប់ការប្រមូលផ្តុំប្រូតេអ៊ីនដែល precipitate ។
សារធាតុ និងលក្ខខណ្ឌដែលបណ្តាលឱ្យខូចទ្រង់ទ្រាយប្រូតេអ៊ីន៖
សីតុណ្ហភាពលើសពី 60 អង្សាសេ - ការបំផ្លាញចំណងខ្សោយនៅក្នុងប្រូតេអ៊ីន;
អាស៊ីតនិងអាល់កាឡាំង - ការផ្លាស់ប្តូរអ៊ីយ៉ូដនៃក្រុមអ៊ីយ៉ុង, ការបំបែកចំណងអ៊ីយ៉ុងនិងអ៊ីដ្រូសែន,
អ៊ុយ - ការបំផ្លិចបំផ្លាញនៃចំណងអ៊ីដ្រូសែន intramolecular ដែលជាលទ្ធផលនៃការបង្កើតចំណងអ៊ីដ្រូសែនជាមួយអ៊ុយ។
ជាតិអាល់កុល phenol chloramine - ការបំផ្លាញចំណង hydrophobic និងអ៊ីដ្រូសែន;
អំបិលនៃលោហធាតុធ្ងន់ - ការបង្កើតអំបិលមិនរលាយនៃប្រូតេអ៊ីនជាមួយអ៊ីយ៉ុងដែកធ្ងន់។
នៅពេលដែលភ្នាក់ងារ denaturing ត្រូវបានយកចេញ ការបង្កើតឡើងវិញគឺអាចធ្វើទៅបាន ចាប់តាំងពីខ្សែសង្វាក់ peptide មានទំនោរក្នុងការទទួលយកការអនុលោមតាមថាមពលឥតគិតថ្លៃទាបបំផុតនៅក្នុងដំណោះស្រាយ។
នៅក្រោមលក្ខខណ្ឌកោសិកាប្រូតេអ៊ីនអាច denature spontaneously ទោះបីជាក្នុងអត្រាទាបជាងនៅសីតុណ្ហភាពខ្ពស់។ ការបង្កើតឡើងវិញដោយឯកឯងនៃប្រូតេអ៊ីនក្នុងកោសិកាគឺពិបាកព្រោះដោយសារកំហាប់ខ្ពស់វាមានប្រូបាបខ្ពស់នៃការប្រមូលផ្តុំម៉ូលេគុលដែលមិនប្រក្រតីដោយផ្នែក។
កោសិកាមានប្រូតេអ៊ីន- មេអំបៅម៉ូលេគុលដែលមានសមត្ថភាពចងភ្ជាប់ទៅនឹងប្រូតេអ៊ីនដែលខូចដោយផ្នែកដែលស្ថិតក្នុងស្ថានភាពមិនស្ថិតស្ថេរ ងាយនឹងប្រមូលផ្តុំ និងស្ដារឡើងវិញនូវទម្រង់ដើមរបស់វា។ ដំបូង ប្រូតេអ៊ីនទាំងនេះត្រូវបានគេរកឃើញថាជាប្រូតេអ៊ីនឆក់កំដៅ ចាប់តាំងពីការសំយោគរបស់ពួកគេបានកើនឡើងនៅពេលដែលកោសិកាត្រូវបានប៉ះពាល់នឹងភាពតានតឹង ឧទាហរណ៍នៅពេលដែលសីតុណ្ហភាពកើនឡើង។ Chaperones ត្រូវបានចាត់ថ្នាក់តាមម៉ាស់នៃអនុក្រុមរបស់វា៖ hsp-60, hsp-70 និង hsp-90 ។ ថ្នាក់នីមួយៗរួមមានក្រុមគ្រួសារនៃប្រូតេអ៊ីនដែលពាក់ព័ន្ធ។
អ្នកបួសម៉ូលេគុល ( hsp-70)ថ្នាក់អភិរក្សខ្ពស់នៃប្រូតេអ៊ីនដែលរកឃើញនៅគ្រប់ផ្នែកទាំងអស់នៃកោសិកា៖ ស៊ីតូប្លាសម នុយក្លីស អេនដូប្លាសមិច រីទីគូល មីតូខនឌ្រី។ នៅ C-terminus នៃខ្សែសង្វាក់ polypeptide តែមួយ hsp-70 មានតំបន់មួយដែលជាចង្អូរដែលមានសមត្ថភាពធ្វើអន្តរកម្មជាមួយនឹងសំណល់អាស៊ីតអាមីណូ peptides 7-9 យូរ ដែលសំបូរទៅដោយរ៉ាឌីកាល់ hydrophobic ។ តំបន់បែបនេះនៅក្នុងប្រូតេអ៊ីន globular កើតឡើងប្រហែលរៀងរាល់ 16 អាមីណូអាស៊ីត។ Hsp-70 អាចការពារប្រូតេអ៊ីនពីការអសកម្មនៃសីតុណ្ហភាព និងស្ដារឡើងវិញនូវការអនុលោមភាព និងសកម្មភាពនៃប្រូតេអ៊ីនដែលមិនមានលក្ខណៈធម្មជាតិ។
Chaperones-60 (hsp-60)ចូលរួមក្នុងការបង្កើតរចនាសម្ព័ន្ធទីបីនៃប្រូតេអ៊ីន។ Hsp-60 មានមុខងារជាប្រូតេអ៊ីន oligomeric ដែលមាន 14 អនុរង។ Hsp-60 បង្កើតជាចិញ្ចៀនពីរដែលចិញ្ចៀននីមួយៗមាន 7 អនុរងដែលតភ្ជាប់គ្នាទៅវិញទៅមក។
ឯកតារងនីមួយៗមានបីដែន៖
ដែន apical មានចំនួននៃសំណល់អាស៊ីតអាមីណូ hydrophobic ប្រឈមមុខនឹងខាងក្នុងនៃបែហោងធ្មែញដែលបង្កើតឡើងដោយអនុរង;
ដែនអេក្វាទ័រមានសកម្មភាព ATPase ហើយចាំបាច់សម្រាប់ការបញ្ចេញប្រូតេអ៊ីនពីស្មុគស្មាញ chaperonin;
ដែនកម្រិតមធ្យមភ្ជាប់ដែន apical និងអេក្វាទ័រ។
ប្រូតេអ៊ីនដែលមានបំណែកនៅលើផ្ទៃរបស់វា។សំបូរទៅដោយអាស៊ីតអាមីណូ hydrophobic ចូលទៅក្នុងបែហោងធ្មែញនៃស្មុគស្មាញ chaperonin ។ នៅក្នុងបរិយាកាសជាក់លាក់នៃបែហោងធ្មែញនេះ នៅក្នុងលក្ខខណ្ឌនៃភាពឯកោពីម៉ូលេគុលផ្សេងទៀតនៃ cytosol កោសិកា ការជ្រើសរើសនៃការអនុលោមតាមប្រូតេអ៊ីនដែលអាចធ្វើបានកើតឡើងរហូតដល់ការអនុលោមភាពអំណោយផលខ្លាំងជាងត្រូវបានរកឃើញ។ ការបង្កើតដែលពឹងផ្អែកលើ chaperone នៃការអនុលោមតាមដើមត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការចំណាយនៃចំនួនថាមពលដ៏សំខាន់ដែលជាប្រភពនៃ ATP ។
ម៉ូឌុល 1 រចនាសម្ព័ន្ធ លក្ខណៈសម្បត្តិ និងមុខងារនៃប្រូតេអ៊ីន
ម៉ូឌុល 1 រចនាសម្ព័ន្ធ លក្ខណៈសម្បត្តិ និងមុខងារនៃប្រូតេអ៊ីន
រចនាសម្ព័ន្ធម៉ូឌុល | ស្បែក |
ឯកតាម៉ូឌុល 1 | ១.១. ការរៀបចំរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីន។ ដំណាក់កាលនៃការបង្កើតការអនុលោមតាមប្រូតេអ៊ីនដើម ១.២. មូលដ្ឋានគ្រឹះនៃដំណើរការប្រូតេអ៊ីន។ ថ្នាំដូចជា ligands ប៉ះពាល់ដល់មុខងារប្រូតេអ៊ីន ១.៣. Denaturation នៃប្រូតេអ៊ីន និងលទ្ធភាពនៃការ renativation spontaneous របស់ពួកគេ។ |
ឯកតាម៉ូឌុល 2 | ១.៤. លក្ខណៈពិសេសនៃរចនាសម្ព័ន្ធនិងមុខងារនៃប្រូតេអ៊ីន oligomeric ដោយប្រើឧទាហរណ៍នៃអេម៉ូក្លូប៊ីន ១.៥. ការថែរក្សាការអនុលោមតាមប្រូតេអ៊ីនដើមក្រោមលក្ខខណ្ឌកោសិកា ១.៦. ភាពខុសគ្នានៃប្រូតេអ៊ីន។ គ្រួសារប្រូតេអ៊ីនដោយប្រើឧទាហរណ៍នៃ immunoglobulins ១.៧. លក្ខណៈសម្បត្តិរូបវិទ្យានៃប្រូតេអ៊ីននិងវិធីសាស្រ្តសម្រាប់ការបំបែករបស់ពួកគេ។ |
ឯកតាម៉ូឌុល 1 ការរៀបចំរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីនម៉ូណូម៉ិច និងមូលដ្ឋានគ្រឹះនៃមុខងាររបស់ពួកគេ
គោលបំណងនៃការសិក្សាអាច៖
1. ប្រើប្រាស់ចំណេះដឹងអំពីលក្ខណៈរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីន និងការពឹងផ្អែកនៃមុខងារប្រូតេអ៊ីនលើរចនាសម្ព័ន្ធរបស់វា ដើម្បីយល់ពីយន្តការនៃការអភិវឌ្ឍនៃជំងឺប្រូតេអ៊ីនពីតំណពូជ និងដែលទទួលបាន។
2. ពន្យល់ពីយន្តការនៃសកម្មភាពព្យាបាលរបស់ថ្នាំមួយចំនួនជាលីហ្គែនដែលមានអន្តរកម្មជាមួយប្រូតេអ៊ីន និងផ្លាស់ប្តូរសកម្មភាពរបស់វា។
3. ប្រើប្រាស់ចំនេះដឹងអំពីរចនាសម្ព័ន្ធ និងការអនុលោមតាមលក្ខខណ្ឌនៃប្រូតេអ៊ីន ដើម្បីយល់ពីអស្ថិរភាពនៃរចនាសម្ព័ន្ធ និងមុខងារ និងទំនោរទៅរកការប្រែពណ៌នៅក្រោមលក្ខខណ្ឌផ្លាស់ប្តូរ។
4. ពន្យល់ពីការប្រើប្រាស់ភ្នាក់ងារ denaturing ជាមធ្យោបាយសម្រាប់ការក្រៀវសម្ភារៈ និងឧបករណ៍វេជ្ជសាស្ត្រ ក៏ដូចជាថ្នាំសំលាប់មេរោគ។
ដឹង៖
1. កម្រិតនៃការរៀបចំរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីន។
2. សារៈសំខាន់នៃរចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនដែលកំណត់ភាពចម្រុះនៃរចនាសម្ព័ន្ធនិងមុខងាររបស់ពួកគេ។
3. យន្តការនៃការបង្កើតមជ្ឈមណ្ឌលសកម្មនៅក្នុងប្រូតេអ៊ីន និងអន្តរកម្មជាក់លាក់របស់វាជាមួយ ligand ដែលបង្កប់នូវមុខងាររបស់ប្រូតេអ៊ីន។
4. ឧទាហរណ៏នៃឥទ្ធិពលនៃ ligands exogenous (ថ្នាំ, ជាតិពុល, សារធាតុពុល) លើការអនុលោមតាមនិងសកម្មភាពមុខងារនៃប្រូតេអ៊ីន។
5. មូលហេតុនិងផលវិបាកនៃប្រូតេអ៊ីន denaturation កត្តាដែលបណ្តាលឱ្យ denaturation ។
6. ឧទាហរណ៍នៃការប្រើប្រាស់កត្តា denaturing ក្នុងឱសថជាថ្នាំសំលាប់មេរោគ និងមធ្យោបាយសម្រាប់ការក្រៀវឧបករណ៍វេជ្ជសាស្ត្រ។
ប្រធានបទ ១.១. ការរៀបចំរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីន។ ដំណាក់កាលនៃការបង្កើតជនជាតិដើម
ការអនុលោមតាមប្រូតេអ៊ីន
ប្រូតេអ៊ីនគឺជាម៉ូលេគុលវត្ថុធាតុ polymer ដែលម៉ូណូមឺរគឺមានតែអាស៊ីតអាមីណូ 20 ប៉ុណ្ណោះ។ សំណុំនិងលំដាប់នៃការរួមបញ្ចូលគ្នានៃអាស៊ីតអាមីណូនៅក្នុងប្រូតេអ៊ីនមួយត្រូវបានកំណត់ដោយរចនាសម្ព័ន្ធនៃហ្សែននៅក្នុង DNA របស់បុគ្គល។ ប្រូតេអ៊ីននីមួយៗស្របតាមរចនាសម្ព័ន្ធជាក់លាក់របស់វាអនុវត្តមុខងារផ្ទាល់ខ្លួនរបស់វា។ សំណុំនៃប្រូតេអ៊ីននៃសារពាង្គកាយដែលបានផ្តល់ឱ្យកំណត់លក្ខណៈ phenotypic របស់វាក៏ដូចជាវត្តមាននៃជំងឺតំណពូជឬ predisposition ចំពោះការអភិវឌ្ឍរបស់វា។
1. អាស៊ីតអាមីណូដែលបង្កើតជាប្រូតេអ៊ីន។ ចំណង Peptide ។ប្រូតេអ៊ីនគឺជាសារធាតុប៉ូលីម៊ែរដែលបង្កើតឡើងពីម៉ូណូមឺរ - អាស៊ីតអាមីណូ 20 ដែលរូបមន្តទូទៅគឺ
អាស៊ីតអាមីណូមានភាពខុសប្លែកគ្នានៅក្នុងរចនាសម្ព័ន្ធ ទំហំ និងលក្ខណៈសម្បត្តិរូបវិទ្យានៃរ៉ាឌីកាល់ដែលភ្ជាប់ទៅនឹងអាតូម α-កាបូន។ ក្រុមមុខងារនៃអាស៊ីតអាមីណូកំណត់លក្ខណៈនៃលក្ខណៈសម្បត្តិនៃអាស៊ីតអាមីណូផ្សេងៗគ្នា។ រ៉ាឌីកាល់ដែលមាននៅក្នុងអាស៊ីតអាមីណូអាចបែងចែកជាក្រុមជាច្រើន៖
ប្រូលីនមិនដូចប្រូតេអ៊ីន monomers ផ្សេងទៀត 19 វាមិនមែនជាអាស៊ីតអាមីណូទេប៉ុន្តែជាអាស៊ីត imino រ៉ាឌីកាល់នៅក្នុង proline ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងអាតូមα-carbon និងក្រុម imino
 អាស៊ីតអាមីណូប្រែប្រួលក្នុងភាពរលាយក្នុងទឹក។នេះគឺដោយសារតែសមត្ថភាពនៃរ៉ាឌីកាល់ដើម្បីធ្វើអន្តរកម្មជាមួយនឹងទឹក (hydrate) ។
អាស៊ីតអាមីណូប្រែប្រួលក្នុងភាពរលាយក្នុងទឹក។នេះគឺដោយសារតែសមត្ថភាពនៃរ៉ាឌីកាល់ដើម្បីធ្វើអន្តរកម្មជាមួយនឹងទឹក (hydrate) ។
TO hydrophilicរួមបញ្ចូលរ៉ាឌីកាល់ដែលមានក្រុមមុខងារ anionic, cationic និង polar uncharged ។
TO hydrophobicរួមបញ្ចូលរ៉ាឌីកាល់ដែលមានក្រុមមេទីល ខ្សែសង្វាក់ aliphatic ឬចិញ្ចៀន។
2. ចំណង Peptide ភ្ជាប់អាស៊ីតអាមីណូដើម្បីបង្កើតជា peptides ។កំឡុងពេលសំយោគ peptide ក្រុម α-carboxyl នៃអាស៊ីតអាមីណូមួយធ្វើអន្តរកម្មជាមួយក្រុម α-amino នៃអាស៊ីតអាមីណូមួយទៀតដើម្បីបង្កើត ចំណង peptide៖
ប្រូតេអ៊ីនគឺជា polypeptides, i.e. ប៉ូលីមែរលីនេអ៊ែរនៃអាស៊ីតអាមីណូដែលតភ្ជាប់ដោយចំណង peptide (រូបភាព 1.1 ។ )
 អង្ករ។ ១.១. ពាក្យដែលប្រើដើម្បីពិពណ៌នាអំពីរចនាសម្ព័ន្ធនៃ peptides
អង្ករ។ ១.១. ពាក្យដែលប្រើដើម្បីពិពណ៌នាអំពីរចនាសម្ព័ន្ធនៃ peptides
monomers នៃអាស៊ីតអាមីណូដែលបង្កើតជា polypeptides ត្រូវបានគេហៅថា សំណល់អាស៊ីតអាមីណូ។ខ្សែសង្វាក់នៃក្រុមដដែលៗ - NH-CH-CO- ទម្រង់ ឆ្អឹងខ្នង peptide ។សំណល់អាស៊ីតអាមីណូដែលមានក្រុម α-amino ឥតគិតថ្លៃត្រូវបានគេហៅថា N-terminal ហើយមួយដែលមានក្រុម α-carboxyl ឥតគិតថ្លៃត្រូវបានគេហៅថា C-terminal ។ Peptides ត្រូវបានសរសេរ និងអានពី N-terminus ទៅ C-terminus ។
ចំណង peptide ដែលបង្កើតឡើងដោយក្រុម imino នៃ proline ខុសពីចំណង peptide ផ្សេងទៀត៖ អាតូមអាសូតនៃក្រុម peptide ខ្វះអ៊ីដ្រូសែន
ផ្ទុយទៅវិញ មានចំណងជាមួយនឹងរ៉ាឌីកាល់ ដែលជាលទ្ធផលដែលផ្នែកម្ខាងនៃចិញ្ចៀនត្រូវបានរួមបញ្ចូលនៅក្នុងឆ្អឹងខ្នង peptide:
 Peptides ខុសគ្នានៅក្នុងសមាសភាពអាស៊ីតអាមីណូ ចំនួនអាស៊ីតអាមីណូ និងលំដាប់នៃការតភ្ជាប់អាស៊ីតអាមីណូ ឧទាហរណ៍ Ser-Ala-Glu-Gis និង His-Glu-Ala-Ser គឺជា peptides ពីរផ្សេងគ្នា។
Peptides ខុសគ្នានៅក្នុងសមាសភាពអាស៊ីតអាមីណូ ចំនួនអាស៊ីតអាមីណូ និងលំដាប់នៃការតភ្ជាប់អាស៊ីតអាមីណូ ឧទាហរណ៍ Ser-Ala-Glu-Gis និង His-Glu-Ala-Ser គឺជា peptides ពីរផ្សេងគ្នា។
ចំណង Peptide គឺខ្លាំង ហើយអ៊ីដ្រូលីស៊ីសដែលមិនមែនជាអង់ស៊ីមគីមីរបស់ពួកវាទាមទារលក្ខខណ្ឌដ៏អាក្រក់៖ ប្រូតេអ៊ីនដែលកំពុងត្រូវបានវិភាគត្រូវបានអ៊ីដ្រូលីហ្សីនក្នុងទឹកអាស៊ីតអ៊ីដ្រូក្លរីកប្រមូលផ្តុំនៅសីតុណ្ហភាពប្រហែល 110° រយៈពេល 24 ម៉ោង។ នៅក្នុងកោសិការស់ ចំណង peptide អាចត្រូវបានបំបែកដោយ អង់ស៊ីម proteolytic,ហៅ ប្រូតេអ៊ីនឬ peptide hydrolases ។
3. រចនាសម្ព័ន្ធបឋមនៃប្រូតេអ៊ីន។សំណល់អាស៊ីតអាមីណូនៅក្នុងខ្សែសង្វាក់ peptide នៃប្រូតេអ៊ីនផ្សេងៗគ្នាមិនឆ្លាស់គ្នាដោយចៃដន្យទេ ប៉ុន្តែត្រូវបានរៀបចំតាមលំដាប់ជាក់លាក់មួយ។ លំដាប់លីនេអ៊ែរ ឬលំដាប់ជំនួសនៃសំណល់អាស៊ីតអាមីណូនៅក្នុងខ្សែសង្វាក់ polypeptide ត្រូវបានគេហៅថា រចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីន។
រចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនបុគ្គលនីមួយៗត្រូវបានអ៊ិនកូដនៅក្នុងម៉ូលេគុល DNA (ក្នុងតំបន់ហៅថាហ្សែន) ហើយត្រូវបានដឹងក្នុងអំឡុងពេលប្រតិចារិក (ចម្លងព័ត៌មានទៅ mRNA) និងការបកប្រែ (ការសំយោគរចនាសម្ព័ន្ធបឋមនៃប្រូតេអ៊ីន) ។ អាស្រ័យហេតុនេះ រចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនរបស់មនុស្សម្នាក់ៗ គឺជាព័ត៌មានដែលចម្លងតាមតំណពូជពីឪពុកម្តាយទៅកូន ដែលកំណត់លក្ខណៈរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីននៃសារពាង្គកាយដែលបានផ្តល់ឱ្យ ដែលមុខងារនៃប្រូតេអ៊ីនដែលមានស្រាប់អាស្រ័យ (រូបភាព 1.2 ។ ) ។
 អង្ករ។ ១.២. ទំនាក់ទំនងរវាង genotype និងការអនុលោមតាមប្រូតេអ៊ីនដែលត្រូវបានសំយោគនៅក្នុងខ្លួនរបស់បុគ្គល
អង្ករ។ ១.២. ទំនាក់ទំនងរវាង genotype និងការអនុលោមតាមប្រូតេអ៊ីនដែលត្រូវបានសំយោគនៅក្នុងខ្លួនរបស់បុគ្គល
ប្រូតេអ៊ីននីមួយៗមានប្រហែល 100,000 នៅក្នុងខ្លួនមនុស្ស ប្លែករចនាសម្ព័ន្ធបឋម។ ម៉ូលេគុលនៃប្រភេទប្រូតេអ៊ីនដូចគ្នា (ឧទាហរណ៍ អាល់ប៊ុយមីន) មានការផ្លាស់ប្តូរដូចគ្នានៃសំណល់អាស៊ីតអាមីណូ ដែលបែងចែកអាល់ប៊ុមប៊ីនពីប្រូតេអ៊ីននីមួយៗ។
លំដាប់នៃសំណល់អាស៊ីតអាមីណូនៅក្នុងខ្សែសង្វាក់ peptide អាចត្រូវបានចាត់ទុកថាជាទម្រង់នៃការកត់ត្រាព័ត៌មាន។ ព័ត៌មាននេះកំណត់ការរៀបចំលំហនៃខ្សែសង្វាក់ peptide លីនេអ៊ែរ ទៅជារចនាសម្ព័ន្ធបីវិមាត្រដែលតូចជាងមុនហៅថា ការអនុលោមតាមកំប្រុក។ ដំណើរការនៃការបង្កើតការអនុលោមតាមប្រូតេអ៊ីនដែលមានមុខងារត្រូវបានគេហៅថា បត់
4. ការអនុលោមតាមប្រូតេអ៊ីន។ការបង្វិលដោយឥតគិតថ្លៃនៅក្នុងឆ្អឹងខ្នង peptide គឺអាចធ្វើទៅបានរវាងអាតូមអាសូតនៃក្រុម peptide និងអាតូមα-carbon ដែលនៅជិតខាង ក៏ដូចជារវាងអាតូមα-carbon និងកាបូននៃក្រុម carbonyl ។ ដោយសារតែអន្តរកម្មនៃក្រុមមុខងារនៃសំណល់អាស៊ីតអាមីណូ រចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនអាចទទួលបានរចនាសម្ព័ន្ធលំហដែលស្មុគស្មាញជាង។ នៅក្នុងប្រូតេអ៊ីន globular មានកម្រិតសំខាន់ពីរនៃការបត់នៃការអនុលោមតាមខ្សែសង្វាក់ peptide: អនុវិទ្យាល័យនិង រចនាសម្ព័ន្ធទីបី។
រចនាសម្ព័ន្ធបន្ទាប់បន្សំនៃប្រូតេអ៊ីនគឺជារចនាសម្ព័ន្ធលំហដែលបង្កើតឡើងជាលទ្ធផលនៃការបង្កើតចំណងអ៊ីដ្រូសែនរវាងក្រុមមុខងារ -C = O និង -NH- នៃឆ្អឹងខ្នង peptide ។ ក្នុងករណីនេះខ្សែសង្វាក់ peptide អាចទទួលបានរចនាសម្ព័ន្ធធម្មតានៃពីរប្រភេទ: α-helicesនិង β-រចនាសម្ព័ន្ធ។
IN α-helicesចំណងអ៊ីដ្រូសែនត្រូវបានបង្កើតឡើងរវាងអាតូមអុកស៊ីសែននៃក្រុម carbonyl និងអ៊ីដ្រូសែននៃអាមីតអាសូតនៃអាស៊ីតអាមីណូទី 4 ពីវា; ខ្សែសង្វាក់ចំហៀងនៃសំណល់អាស៊ីតអាមីណូ
មានទីតាំងនៅតាមបរិវេណនៃវង់ដោយមិនចូលរួមក្នុងការបង្កើតរចនាសម្ព័ន្ធបន្ទាប់បន្សំ (រូបភាព 1.3 ។ ) ។
រ៉ាឌីកាល់ច្រើន ឬរ៉ាឌីកាល់ដែលផ្ទុកបន្ទុកស្មើគ្នា ការពារការបង្កើត α-helix ។ សំណល់ប្រូលីនដែលមានរចនាសម្ព័ន្ធចិញ្ចៀនរំខាន α-helix ចាប់តាំងពីដោយសារតែកង្វះអ៊ីដ្រូសែននៅអាតូមអាសូតនៅក្នុងខ្សែសង្វាក់ peptide វាមិនអាចទៅរួចទេក្នុងការបង្កើតចំណងអ៊ីដ្រូសែន។ ចំណងរវាងអាសូត និងអាតូម α-កាបូនគឺជាផ្នែកមួយនៃរង្វង់ប្រូលីន ដូច្នេះឆ្អឹងខ្នង peptide ប្រែជាកោងនៅចំណុចនេះ។
β-រចនាសម្ព័ន្ធត្រូវបានបង្កើតឡើងរវាងតំបន់លីនេអ៊ែរនៃឆ្អឹងខ្នង peptide នៃខ្សែសង្វាក់ polypeptide មួយដោយហេតុនេះបង្កើតបានជារចនាសម្ព័ន្ធបត់។ ខ្សែសង្វាក់ Polypeptide ឬផ្នែករបស់វាអាចបង្កើតបាន។ ប៉ារ៉ាឡែលឬ រចនាសម្ព័ន្ធប្រឆាំងប៉ារ៉ាឡែល β ។ក្នុងករណីទីមួយ N- និង C-termini នៃខ្សែសង្វាក់ peptide អន្តរកម្មស្របគ្នា ហើយនៅក្នុងទីពីរពួកគេមានទិសដៅផ្ទុយ (រូបភាព 1.4) ។
អង្ករ។ ១.៣. រចនាសម្ព័ន្ធទីពីរនៃប្រូតេអ៊ីន - α-helix
 អង្ករ។ ១.៤. រចនាសម្ព័ន្ធសន្លឹកប៉ារ៉ាឡែល និងប្រឆាំងប៉ារ៉ាឡែល
អង្ករ។ ១.៤. រចនាសម្ព័ន្ធសន្លឹកប៉ារ៉ាឡែល និងប្រឆាំងប៉ារ៉ាឡែល
រចនាសម្ព័ន្ធ β ត្រូវបានចង្អុលបង្ហាញដោយព្រួញធំទូលាយ: A - រចនាសម្ព័ន្ធ β-ប្រឆាំងប៉ារ៉ាឡែល។ ខ - រចនាសម្ព័ន្ធសន្លឹកប៉ារ៉ាឡែល
នៅក្នុងប្រូតេអ៊ីនមួយចំនួន រចនាសម្ព័ន្ធ β អាចត្រូវបានបង្កើតឡើងដោយសារតែការបង្កើតចំណងអ៊ីដ្រូសែនរវាងអាតូមនៃឆ្អឹងខ្នង peptide នៃខ្សែសង្វាក់ polypeptide ផ្សេងៗគ្នា។
ត្រូវបានរកឃើញផងដែរនៅក្នុងប្រូតេអ៊ីន តំបន់ដែលមានអនុវិទ្យាល័យមិនទៀងទាត់រចនាសម្ព័ន្ធដែលរួមមានពត់ រង្វិលជុំ និងវេននៃឆ្អឹងខ្នង polypeptide ។ ជារឿយៗពួកវាមានទីតាំងនៅកន្លែងដែលទិសដៅនៃខ្សែសង្វាក់ peptide ផ្លាស់ប្តូរឧទាហរណ៍នៅពេលដែលរចនាសម្ព័ន្ធសន្លឹកបេតាប៉ារ៉ាឡែលត្រូវបានបង្កើតឡើង។
ដោយផ្អែកលើវត្តមានរបស់ α-helices និង β-structures ប្រូតេអ៊ីន globular អាចត្រូវបានបែងចែកជា 4 ប្រភេទ។
អង្ករ។ ១.៥. រចនាសម្ព័ន្ធបន្ទាប់បន្សំនៃ myoglobin (A) និង hemoglobin β-chain (B) ដែលមាន α-helices ចំនួនប្រាំបី

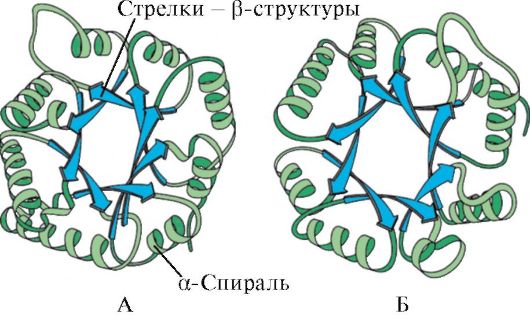 អង្ករ។ ១.៦. រចនាសម្ព័ន្ធបន្ទាប់បន្សំនៃ triosephosphate isomerase និងដែន pyruvate kinase
អង្ករ។ ១.៦. រចនាសម្ព័ន្ធបន្ទាប់បន្សំនៃ triosephosphate isomerase និងដែន pyruvate kinase
 អង្ករ។ ១.៧. រចនាសម្ព័ន្ធបន្ទាប់បន្សំនៃដែនថេរនៃ immunoglobulin (A) និងអង់ស៊ីម superoxide dismutase (B)
អង្ករ។ ១.៧. រចនាសម្ព័ន្ធបន្ទាប់បន្សំនៃដែនថេរនៃ immunoglobulin (A) និងអង់ស៊ីម superoxide dismutase (B)
IN ប្រភេទទីបួនរួមបញ្ចូលប្រូតេអ៊ីនដែលមានបរិមាណតិចតួចនៃរចនាសម្ព័ន្ធបន្ទាប់បន្សំធម្មតា។ ប្រូតេអ៊ីនទាំងនេះរួមមានប្រូតេអ៊ីនដែលមានជាតិ cysteine តូច ឬ metalloproteins ។
រចនាសម្ព័ន្ធទីបីនៃប្រូតេអ៊ីន- ប្រភេទនៃការអនុលោមតាមទម្រង់ដែលត្រូវបានបង្កើតឡើងដោយសារតែអន្តរកម្មរវាងរ៉ាឌីកាល់អាស៊ីតអាមីណូដែលអាចមានទីតាំងនៅចម្ងាយសន្ធឹកសន្ធាប់ពីគ្នាទៅវិញទៅមកនៅក្នុងខ្សែសង្វាក់ peptide ។ ប្រូតេអ៊ីនភាគច្រើនបង្កើតជារចនាសម្ព័ន្ធលំហដែលស្រដៀងនឹង globule (ប្រូតេអ៊ីន globular) ។
ចាប់តាំងពីរ៉ាឌីកាល់អាស៊ីតអាមីណូ hydrophobic មានទំនោរបញ្ចូលគ្នាតាមរយៈអ្វីដែលគេហៅថា អន្តរកម្ម hydrophobicនិងកម្លាំង van der Waals អន្តរម៉ូលេគុល ស្នូល hydrophobic ក្រាស់ត្រូវបានបង្កើតឡើងនៅខាងក្នុង globule ប្រូតេអ៊ីន។ រ៉ាឌីកាល់អ៊ីយ៉ូដ hydrophilic និងមិនអ៊ីយ៉ូដ មានទីតាំងនៅលើផ្ទៃប្រូតេអ៊ីន និងកំណត់ភាពរលាយរបស់វានៅក្នុងទឹក។
 អង្ករ។ ១.៨. ប្រភេទនៃចំណងដែលកើតឡើងរវាងរ៉ាឌីកាល់អាស៊ីតអាមីណូអំឡុងពេលបង្កើតរចនាសម្ព័ន្ធទីបីនៃប្រូតេអ៊ីន
អង្ករ។ ១.៨. ប្រភេទនៃចំណងដែលកើតឡើងរវាងរ៉ាឌីកាល់អាស៊ីតអាមីណូអំឡុងពេលបង្កើតរចនាសម្ព័ន្ធទីបីនៃប្រូតេអ៊ីន
1 - ចំណងអ៊ីយ៉ូដ- កើតឡើងរវាងក្រុមមុខងារវិជ្ជមាន និងអវិជ្ជមាន។
2 - ចំណងអ៊ីដ្រូសែន- កើតឡើងរវាងក្រុម hydrophilic uncharged និងក្រុម hydrophilic ផ្សេងទៀតណាមួយ;
3 - អន្តរកម្ម hydrophobic- កើតឡើងរវាងរ៉ាឌីកាល់ hydrophobic;
4 - ចំណង disulfide- បង្កើតឡើងដោយសារតែការកត់សុីនៃក្រុម SH នៃសំណល់ cysteine និងអន្តរកម្មរបស់ពួកគេជាមួយគ្នាទៅវិញទៅមក
សំណល់អាស៊ីតអាមីណូ hydrophilic ដែលមានទីតាំងនៅខាងក្នុងស្នូល hydrophobic អាចធ្វើអន្តរកម្មជាមួយគ្នាដោយប្រើ អ៊ីយ៉ុងនិង ចំណងអ៊ីដ្រូសែន(រូបភាព 1.8) ។
ចំណងអ៊ីយ៉ុង និងអ៊ីដ្រូសែន ក៏ដូចជាអន្តរកម្ម hydrophobic គឺខ្សោយ៖ ថាមពលរបស់ពួកគេមិនខ្ពស់ជាងថាមពលនៃចលនាកម្ដៅនៃម៉ូលេគុលនៅសីតុណ្ហភាពបន្ទប់នោះទេ។ ការអនុលោមតាមប្រូតេអ៊ីនត្រូវបានរក្សាដោយការបង្កើតចំណងខ្សោយបែបនេះជាច្រើន។ ដោយសារអាតូមដែលបង្កើតជាប្រូតេអ៊ីនមានចលនាថេរ វាអាចទៅរួចក្នុងការបំបែកចំណងខ្សោយមួយចំនួន និងបង្កើតជាធាតុផ្សេងទៀត ដែលនាំឱ្យមានចលនាបន្តិចបន្តួចនៃផ្នែកនីមួយៗនៃខ្សែសង្វាក់ polypeptide ។ ទ្រព្យសម្បត្តិនៃប្រូតេអ៊ីននេះដើម្បីផ្លាស់ប្តូរការអនុលោមតាមលទ្ធផលនៃការបំបែកមួយចំនួននិងបង្កើតចំណងខ្សោយផ្សេងទៀតត្រូវបានគេហៅថា lability អនុលោមភាព។
រាងកាយរបស់មនុស្សមានប្រព័ន្ធដែលគាំទ្រ homeostasis- ភាពស្ថិតស្ថេរនៃបរិយាកាសខាងក្នុងក្នុងដែនកំណត់ដែលអាចទទួលយកបានសម្រាប់រាងកាយដែលមានសុខភាពល្អ។ នៅក្រោមលក្ខខណ្ឌ homeostasis ការផ្លាស់ប្តូរតិចតួចក្នុងការអនុលោមភាពមិនរំខានដល់រចនាសម្ព័ន្ធនិងមុខងារទាំងមូលនៃប្រូតេអ៊ីនទេ។ ការអនុលោមតាមមុខងារនៃប្រូតេអ៊ីនត្រូវបានគេហៅថា ការអនុលោមតាមជនជាតិដើម។ការផ្លាស់ប្តូរបរិយាកាសខាងក្នុង (ឧទាហរណ៍ កំហាប់គ្លុយកូស Ca អ៊ីយ៉ុង ប្រូតុង។
រចនាសម្ព័ន្ធទីបីនៃប្រូតេអ៊ីនមួយចំនួនមានស្ថេរភាព ចំណង disulfide,បង្កើតឡើងដោយសារតែអន្តរកម្មនៃក្រុម -SH នៃសំណល់ពីរ
 អង្ករ។ ១.៩. ការបង្កើតចំណង disulfide នៅក្នុងម៉ូលេគុលប្រូតេអ៊ីន
អង្ករ។ ១.៩. ការបង្កើតចំណង disulfide នៅក្នុងម៉ូលេគុលប្រូតេអ៊ីន
cysteine (រូបភាព 1.9) ។ ប្រូតេអ៊ីនក្នុងកោសិកាភាគច្រើនមិនមានចំណង disulfide covalent នៅក្នុងរចនាសម្ព័ន្ធទីបីរបស់ពួកគេទេ។ វត្តមានរបស់ពួកគេគឺជាលក្ខណៈនៃប្រូតេអ៊ីនដែលលាក់ដោយកោសិកាដែលធានានូវស្ថេរភាពកាន់តែច្រើនរបស់ពួកគេនៅក្នុងលក្ខខណ្ឌក្រៅកោសិកា។ ដូច្នេះចំណង disulfide មាននៅក្នុងម៉ូលេគុលនៃអាំងស៊ុយលីន និង immunoglobulins ។
អាំងស៊ុយលីន- អ័រម៉ូនប្រូតេអុីនដែលសំយោគនៅក្នុងកោសិកា β-កោសិកានៃលំពែង ហើយសម្ងាត់ចូលទៅក្នុងឈាម ដើម្បីឆ្លើយតបទៅនឹងការកើនឡើងនៃកំហាប់គ្លុយកូសក្នុងឈាម។ នៅក្នុងរចនាសម្ព័ន្ធនៃអាំងស៊ុយលីន មានចំណង disulfide ពីរដែលតភ្ជាប់ខ្សែសង្វាក់ polypeptide A និង B និងចំណង disulfide មួយនៅក្នុងខ្សែសង្វាក់ A (រូបភាព 1.10) ។
 អង្ករ។ ១.១០. ចំណង disulfide នៅក្នុងរចនាសម្ព័ន្ធនៃអាំងស៊ុយលីន
អង្ករ។ ១.១០. ចំណង disulfide នៅក្នុងរចនាសម្ព័ន្ធនៃអាំងស៊ុយលីន
5. រចនាសម្ព័ន្ធអនុវិទ្យាល័យនៃប្រូតេអ៊ីន។នៅក្នុងប្រូតេអ៊ីនដែលមានរចនាសម្ព័ន្ធ និងមុខងារចម្បងខុសៗគ្នា ជួនកាលត្រូវបានរកឃើញ បន្សំស្រដៀងគ្នានិងទីតាំងដែលទាក់ទងនៃរចនាសម្ព័ន្ធបន្ទាប់បន្សំ,ដែលត្រូវបានគេហៅថារចនាសម្ព័ន្ធអនុវិទ្យាល័យ។ វាកាន់កាប់ទីតាំងមធ្យមរវាងរចនាសម្ព័ន្ធបន្ទាប់បន្សំ និងទីបី ដោយសារវាគឺជាការរួមបញ្ចូលគ្នាជាក់លាក់នៃធាតុនៃរចនាសម្ព័ន្ធបន្ទាប់បន្សំក្នុងការបង្កើតរចនាសម្ព័ន្ធទីបីនៃប្រូតេអ៊ីន។ រចនាសម្ព័ន្ធអនុវិទ្យាល័យមានឈ្មោះជាក់លាក់ដូចជា “α-helix-turn-a-helix” “leucine zipper” “zinc fingers” ជាដើម។ រចនាសម្ព័ន្ធអនុវិទ្យាល័យបែបនេះគឺជាលក្ខណៈនៃប្រូតេអ៊ីនដែលភ្ជាប់ DNA ។
"ខ្សែរ៉ូត Leucine" ។ប្រភេទនៃរចនាសម្ព័ន្ធអនុវិទ្យាល័យនេះត្រូវបានប្រើដើម្បីភ្ជាប់ប្រូតេអ៊ីនពីរជាមួយគ្នា។ នៅលើផ្ទៃនៃប្រូតេអ៊ីនអន្តរកម្មមានតំបន់ α-helical ដែលមានសំណល់ leucine យ៉ាងតិចបួន។ សំណល់ Leucine នៅក្នុង α-helix ស្ថិតនៅដាច់ពីគ្នានៃអាស៊ីតអាមីណូចំនួនប្រាំមួយ។ ចាប់តាំងពីវេននីមួយៗនៃ α-helix មានសំណល់អាស៊ីតអាមីណូ 3.6 រ៉ាឌីកាល់ leucine មានទីតាំងនៅលើផ្ទៃនៃរាល់វេនទីពីរ។ សំណល់ Leucine នៃα-helix នៃប្រូតេអ៊ីនមួយអាចធ្វើអន្តរកម្មជាមួយនឹងសំណល់ leucine នៃប្រូតេអ៊ីនមួយផ្សេងទៀត (អន្តរកម្ម hydrophobic) ដោយភ្ជាប់ពួកវាជាមួយគ្នា (រូបភាព 1.11 ។ ) ។ ប្រូតេអ៊ីនភ្ជាប់ DNA ជាច្រើនមានមុខងារនៅក្នុងស្មុគស្មាញ oligomeric ដែលផ្នែករងនីមួយៗត្រូវបានភ្ជាប់ទៅគ្នាទៅវិញទៅមកដោយ "leucine zippers" ។
 អង្ករ។ ១.១១. "Leucine zipper" រវាងតំបន់α-helical នៃប្រូតេអ៊ីនពីរ
អង្ករ។ ១.១១. "Leucine zipper" រវាងតំបន់α-helical នៃប្រូតេអ៊ីនពីរ
ឧទាហរណ៏នៃប្រូតេអ៊ីនបែបនេះគឺជាអ៊ីស្តូន។ អ៊ីស្តូន- ប្រូតេអ៊ីននុយក្លេអ៊ែរដែលមានអាស៊ីតអាមីណូមួយចំនួនធំដែលមានបន្ទុកវិជ្ជមាន - arginine និង lysine (រហូតដល់ 80%) ។ ម៉ូលេគុល Histone ត្រូវបានផ្សំចូលទៅក្នុងស្មុគស្មាញ oligomeric ដែលមាន monomers ចំនួនប្រាំបីដោយប្រើ "leucine zippers" ទោះបីជាមានការចោទប្រកាន់ដូចគ្នាយ៉ាងសំខាន់នៃម៉ូលេគុលទាំងនេះក៏ដោយ។
"ម្រាមដៃស័ង្កសី"- បំរែបំរួលនៃរចនាសម្ព័ន្ធអនុវិទ្យាល័យ លក្ខណៈនៃប្រូតេអ៊ីនភ្ជាប់ DNA មានទម្រង់ជាបំណែកពន្លូតនៅលើផ្ទៃប្រូតេអ៊ីន និងមានសំណល់អាស៊ីតអាមីណូប្រហែល 20 (រូបភាព 1.12) ។ រូបរាង "ម្រាមដៃពង្រីក" ត្រូវបានគាំទ្រដោយអាតូមស័ង្កសីដែលភ្ជាប់ទៅនឹងរ៉ាឌីកាល់អាស៊ីតអាមីណូចំនួនបួន - សំណល់ cysteine ពីរនិងសំណល់ histidine ពីរ។ ក្នុងករណីខ្លះជំនួសឱ្យសំណល់ histidine មានសំណល់ cysteine ។ សំណល់ cysteine ដែលស្ថិតនៅជិតគ្នាពីរត្រូវបានបំបែកពីសំណល់ Gisili ពីរផ្សេងទៀតដោយលំដាប់ Cys ដែលមានសំណល់អាស៊ីដអាមីណូប្រហែល 12 ។ តំបន់នៃប្រូតេអ៊ីននេះបង្កើតបានជា α-helix ដែលជារ៉ាឌីកាល់ដែលអាចភ្ជាប់ជាពិសេសទៅនឹងតំបន់បទប្បញ្ញត្តិនៃចង្អូរសំខាន់នៃ DNA ។ ភាពជាក់លាក់នៃការចងបុគ្គល
 អង្ករ។
អង្ករ។
១.១២. រចនាសម្ព័ន្ធចម្បងនៃតំបន់នៃប្រូតេអ៊ីនភ្ជាប់ DNA ដែលបង្កើតជារចនាសម្ព័ន្ធ "ម្រាមដៃស័ង្កសី" (អក្សរបង្ហាញពីអាស៊ីតអាមីណូដែលបង្កើតរចនាសម្ព័ន្ធនេះ)
និយតកម្មនៃប្រូតេអ៊ីនចង DNA អាស្រ័យលើលំដាប់នៃសំណល់អាស៊ីតអាមីណូដែលមានទីតាំងនៅតំបន់ម្រាមដៃស័ង្កសី។ រចនាសម្ព័ន្ធបែបនេះមានជាពិសេសអ្នកទទួលសម្រាប់អរម៉ូនស្តេរ៉ូអ៊ីតដែលពាក់ព័ន្ធនឹងបទប្បញ្ញត្តិនៃការចម្លង (អានព័ត៌មានពី DNA ទៅ RNA) ។
ប្រធានបទ 1.2 ។ មូលដ្ឋានគ្រឹះនៃដំណើរការប្រូតេអ៊ីន។ គ្រឿងញៀនដែលប៉ះពាល់ដល់មុខងារប្រូតេអ៊ីន 1. មជ្ឈមណ្ឌលសកម្មនៃប្រូតេអ៊ីននិងអន្តរកម្មរបស់វាជាមួយ ligand ។
ក្នុងអំឡុងពេលនៃការបង្កើតរចនាសម្ព័ន្ធទីបី តំបន់មួយត្រូវបានបង្កើតឡើងនៅលើផ្ទៃនៃប្រូតេអ៊ីនដែលមានមុខងារជាធម្មតានៅក្នុងកន្លែងសម្រាកដែលបង្កើតឡើងដោយរ៉ាឌីកាល់អាស៊ីតអាមីណូដែលនៅឆ្ងាយពីគ្នាទៅវិញទៅមកនៅក្នុងរចនាសម្ព័ន្ធបឋម។ តំបន់នេះដែលមានរចនាសម្ព័ន្ធពិសេសសម្រាប់ប្រូតេអ៊ីនដែលបានផ្តល់ឱ្យហើយមានសមត្ថភាពធ្វើអន្តរកម្មជាពិសេសជាមួយម៉ូលេគុលជាក់លាក់មួយឬក្រុមនៃម៉ូលេគុលស្រដៀងគ្នាត្រូវបានគេហៅថាកន្លែងភ្ជាប់ប្រូតេអ៊ីន-ligand ឬកន្លែងសកម្ម។ Ligands គឺជាម៉ូលេគុលដែលមានអន្តរកម្មជាមួយប្រូតេអ៊ីន។ភាពជាក់លាក់ខ្ពស់។
អន្តរកម្មនៃប្រូតេអ៊ីនជាមួយ ligand ត្រូវបានធានាដោយការបំពេញបន្ថែមនៃរចនាសម្ព័ន្ធនៃមជ្ឈមណ្ឌលសកម្មទៅនឹងរចនាសម្ព័ន្ធនៃ ligand នេះ។ការបំពេញបន្ថែម
 - នេះគឺជាការឆ្លើយឆ្លងតាមលំហ និងគីមីនៃផ្ទៃអន្តរកម្ម។ មជ្ឈមណ្ឌលសកម្មត្រូវតែមិនត្រឹមតែត្រូវគ្នាជាលក្ខណៈលំហទៅនឹង ligand ដែលរួមបញ្ចូលនៅក្នុងវាប៉ុណ្ណោះទេ ប៉ុន្តែក៏មានចំណង (អន្តរកម្មអ៊ីយ៉ុង អ៊ីដ្រូសែន និងអ៊ីដ្រូហ្វូប៊ីក) ត្រូវតែត្រូវបានបង្កើតឡើងរវាងក្រុមមុខងារនៃរ៉ាឌីកាល់ដែលរួមបញ្ចូលនៅក្នុងមជ្ឈមណ្ឌលសកម្ម និងលីហ្គែន ដែលផ្ទុកលីហ្គែន។ នៅក្នុងមជ្ឈមណ្ឌលសកម្ម (រូបភាព 1.13) ។
- នេះគឺជាការឆ្លើយឆ្លងតាមលំហ និងគីមីនៃផ្ទៃអន្តរកម្ម។ មជ្ឈមណ្ឌលសកម្មត្រូវតែមិនត្រឹមតែត្រូវគ្នាជាលក្ខណៈលំហទៅនឹង ligand ដែលរួមបញ្ចូលនៅក្នុងវាប៉ុណ្ណោះទេ ប៉ុន្តែក៏មានចំណង (អន្តរកម្មអ៊ីយ៉ុង អ៊ីដ្រូសែន និងអ៊ីដ្រូហ្វូប៊ីក) ត្រូវតែត្រូវបានបង្កើតឡើងរវាងក្រុមមុខងារនៃរ៉ាឌីកាល់ដែលរួមបញ្ចូលនៅក្នុងមជ្ឈមណ្ឌលសកម្ម និងលីហ្គែន ដែលផ្ទុកលីហ្គែន។ នៅក្នុងមជ្ឈមណ្ឌលសកម្ម (រូបភាព 1.13) ។
លីហ្គែនមួយចំនួននៅពេលដែលភ្ជាប់ទៅនឹងកណ្តាលសកម្មនៃប្រូតេអ៊ីនដើរតួនាទីជំនួយក្នុងដំណើរការនៃប្រូតេអ៊ីន។ លីហ្គែនបែបនេះត្រូវបានគេហៅថា cofactors ហើយប្រូតេអ៊ីនដែលមានផ្នែកដែលមិនមែនជាប្រូតេអ៊ីនត្រូវបានគេហៅថា ប្រូតេអ៊ីនស្មុគស្មាញ(ផ្ទុយទៅនឹងប្រូតេអ៊ីនសាមញ្ញដែលមានតែផ្នែកប្រូតេអ៊ីន) ។ ផ្នែកដែលមិនមែនជាប្រូតេអ៊ីនដែលភ្ជាប់យ៉ាងរឹងមាំទៅនឹងប្រូតេអ៊ីនត្រូវបានគេហៅថា ក្រុមសិប្បនិម្មិត។ឧទហរណ៍ myoglobin, hemoglobin និង cytochromes មានក្រុមសិប្បនិម្មិតមួយ heme ដែលមានអ៊ីយ៉ុងដែកដែលភ្ជាប់យ៉ាងរឹងមាំទៅនឹងមជ្ឈមណ្ឌលសកម្ម។ ប្រូតេអ៊ីនស្មុគស្មាញដែលមាន heme ត្រូវបានគេហៅថា hemoproteins ។
នៅពេលដែល ligands ជាក់លាក់ត្រូវបានភ្ជាប់ទៅនឹងប្រូតេអ៊ីនមុខងារនៃប្រូតេអ៊ីនទាំងនេះត្រូវបានបង្ហាញ។ ដូច្នេះ អាល់ប៊ុយមីន ដែលជាប្រូតេអ៊ីនដ៏សំខាន់បំផុតនៅក្នុងប្លាស្មាឈាម បង្ហាញមុខងារដឹកជញ្ជូនរបស់វា ដោយភ្ជាប់អ៊ីដ្រូហ្វិកលីហ្គែន ដូចជាអាស៊ីតខ្លាញ់ ប៊ីលីរុយប៊ីន ថ្នាំមួយចំនួនជាដើម ទៅមជ្ឈមណ្ឌលសកម្ម (រូបភាព 1.14)។
Ligands អន្តរកម្មជាមួយរចនាសម្ព័ន្ធបីវិមាត្រនៃខ្សែសង្វាក់ peptide អាចមិនត្រឹមតែជាម៉ូលេគុលសរីរាង្គ និងអសរីរាង្គទាបប៉ុណ្ណោះទេ ថែមទាំងម៉ាក្រូម៉ូលេគុលផងដែរ៖
DNA (ឧទាហរណ៍ជាមួយប្រូតេអ៊ីនភ្ជាប់ DNA ដែលបានពិភាក្សាខាងលើ);
ប៉ូលីសាខ័រ;
 អង្ករ។ ១.១៤. ទំនាក់ទំនងរវាង genotype និង phenotype
អង្ករ។ ១.១៤. ទំនាក់ទំនងរវាង genotype និង phenotype
រចនាសម្ព័ន្ធចម្បងតែមួយគត់នៃប្រូតេអ៊ីនរបស់មនុស្សដែលត្រូវបានអ៊ិនកូដនៅក្នុងម៉ូលេគុល DNA ត្រូវបានដឹងនៅក្នុងកោសិកាក្នុងទម្រង់នៃការអនុលោមភាពតែមួយគត់ រចនាសម្ព័ន្ធកណ្តាលសកម្ម និងមុខងារប្រូតេអ៊ីន។
នៅក្នុងករណីទាំងនេះ ប្រូតេអ៊ីនទទួលស្គាល់តំបន់ជាក់លាក់មួយនៃ ligand ដែលមានលក្ខណៈសមស្រប និងបំពេញបន្ថែមទៅនឹងកន្លែងចង។ ដូច្នេះនៅលើផ្ទៃនៃ hepatocytes មានប្រូតេអ៊ីន receptor សម្រាប់អរម៉ូនអាំងស៊ុយលីនដែលមានរចនាសម្ព័ន្ធប្រូតេអ៊ីនផងដែរ។ អន្តរកម្មនៃអាំងស៊ុយលីនជាមួយអ្នកទទួលបណ្តាលឱ្យមានការផ្លាស់ប្តូរនៅក្នុងការអនុលោមភាពនិងការធ្វើឱ្យសកម្មនៃប្រព័ន្ធសញ្ញារបស់វាដែលនាំឱ្យមានការផ្ទុកសារធាតុចិញ្ចឹមនៅក្នុង hepatocytes បន្ទាប់ពីអាហារ។
ដូច្នេះ ដំណើរការនៃប្រូតេអ៊ីនគឺផ្អែកលើអន្តរកម្មជាក់លាក់នៃមជ្ឈមណ្ឌលសកម្មនៃប្រូតេអ៊ីនជាមួយ ligand ។
2. រចនាសម្ព័ន្ធដែន និងតួនាទីរបស់វាក្នុងដំណើរការនៃប្រូតេអ៊ីន។ខ្សែសង្វាក់ polypeptide វែងនៃប្រូតេអ៊ីន globular ច្រើនតែបត់ចូលទៅក្នុងតំបន់បង្រួម និងឯករាជ្យមួយចំនួន។ ពួកវាមានរចនាសម្ព័ន្ធទី 3 ឯករាជ្យដែលនឹកឃើញដល់ប្រូតេអ៊ីនរាងមូល ហើយត្រូវបានគេហៅថា ដែន។សូមអរគុណដល់រចនាសម្ព័ន្ធដែននៃប្រូតេអ៊ីន រចនាសម្ព័ន្ធទីបីរបស់ពួកគេកាន់តែងាយស្រួលក្នុងការបង្កើត។
នៅក្នុង domain proteins ទីតាំងចង ligand ជាញឹកញាប់ស្ថិតនៅចន្លោះដែន។ ដូច្នេះ trypsin គឺជាអង់ស៊ីម proteolytic ដែលត្រូវបានផលិតដោយផ្នែក exocrine នៃលំពែង និងចាំបាច់សម្រាប់ការរំលាយអាហារប្រូតេអ៊ីនអាហារ។ វាមានរចនាសម្ព័ន្ធដែនពីរ ហើយចំណុចកណ្តាលនៃការចងនៃ trypsin ជាមួយ ligand របស់វា - ប្រូតេអ៊ីនអាហារ - ស្ថិតនៅក្នុងចង្អូររវាងដែនទាំងពីរ។ នៅក្នុងមជ្ឈមណ្ឌលសកម្មលក្ខខណ្ឌចាំបាច់សម្រាប់ការផ្សារភ្ជាប់ប្រកបដោយប្រសិទ្ធភាពនៃទីតាំងជាក់លាក់នៃប្រូតេអ៊ីនអាហារនិងអ៊ីដ្រូលីស៊ីនៃចំណង peptide របស់វាត្រូវបានបង្កើតឡើង។
ដែនផ្សេងគ្នានៅក្នុងប្រូតេអ៊ីនអាចផ្លាស់ទីទាក់ទងគ្នាទៅវិញទៅមកនៅពេលដែលមជ្ឈមណ្ឌលសកម្មមានអន្តរកម្មជាមួយ ligand (រូបភាព 1.15) ។
Hexokinase- អង់ស៊ីមដែលជំរុញ phosphorylation នៃជាតិស្ករដោយប្រើ ATP ។ ទីតាំងសកម្មនៃអង់ស៊ីមនេះ ស្ថិតនៅត្រង់ចំនុចប្រសព្វរវាងដែនទាំងពីរ។ នៅពេលដែល hexokinase ភ្ជាប់ទៅនឹងគ្លុយកូស ដែនជុំវិញវានៅជិត ហើយស្រទាប់ខាងក្រោមនឹងជាប់ ដែលផូស្វ័រកើតឡើង (សូមមើលរូប 1.15)។
 អង្ករ។ ១.១៥. ការភ្ជាប់ដែន hexokinase ទៅនឹងគ្លុយកូស
អង្ករ។ ១.១៥. ការភ្ជាប់ដែន hexokinase ទៅនឹងគ្លុយកូស
នៅក្នុងប្រូតេអ៊ីនមួយចំនួន ដែនអនុវត្តមុខងារឯករាជ្យដោយការចងភ្ជាប់ទៅនឹង ligands ផ្សេងៗ។ ប្រូតេអ៊ីនបែបនេះត្រូវបានគេហៅថាពហុមុខងារ។
3. គ្រឿងញៀនគឺជាលីហ្គែនដែលប៉ះពាល់ដល់មុខងាររបស់ប្រូតេអ៊ីន។អន្តរកម្មនៃប្រូតេអ៊ីនជាមួយ ligands គឺជាក់លាក់។ ទោះបីជាយ៉ាងណាក៏ដោយ ដោយសារតែភាពស៊ីសង្វាក់គ្នានៃប្រូតេអ៊ីន និងមជ្ឈមណ្ឌលសកម្មរបស់វា វាអាចជ្រើសរើសសារធាតុមួយផ្សេងទៀតដែលអាចធ្វើអន្តរកម្មជាមួយប្រូតេអ៊ីននៅក្នុងមជ្ឈមណ្ឌលសកម្ម ឬផ្នែកផ្សេងទៀតនៃម៉ូលេគុលផងដែរ។
សារធាតុដែលស្រដៀងគ្នានៅក្នុងរចនាសម្ព័ន្ធទៅនឹង ligand ធម្មជាតិត្រូវបានគេហៅថា analogue រចនាសម្ព័ន្ធនៃ ligandឬលីហ្គែនដែលមិនមែនជាធម្មជាតិ។ វាក៏មានអន្តរកម្មជាមួយប្រូតេអ៊ីននៅកន្លែងសកម្មផងដែរ។ analogue រចនាសម្ព័ន្ធនៃ ligand អាចបង្កើនមុខងារប្រូតេអ៊ីន (agonist),និងកាត់បន្ថយវា។ (អ្នកប្រឆាំង) ។លីហ្គែន និងអាណាឡូកនៃរចនាសម្ព័ន្ធរបស់វាប្រកួតប្រជែងគ្នាទៅវិញទៅមកសម្រាប់ការផ្សារភ្ជាប់ទៅនឹងប្រូតេអ៊ីននៅកន្លែងតែមួយ។ សារធាតុបែបនេះត្រូវបានគេហៅថា ម៉ូឌុលប្រកួតប្រជែង(និយតករ) នៃមុខងារប្រូតេអ៊ីន។ ថ្នាំជាច្រើនដើរតួជាអ្នកទប់ស្កាត់ប្រូតេអ៊ីន។ ពួកគេមួយចំនួនត្រូវបានទទួលដោយការកែប្រែគីមីនៃ ligands ធម្មជាតិ។ សារធាតុរារាំងមុខងារប្រូតេអ៊ីនអាចជាថ្នាំ និងសារធាតុពុល។
Atropine គឺជាអ្នកទប់ស្កាត់ការប្រកួតប្រជែងនៃអ្នកទទួល M-cholinergic ។ Acetylcholine គឺជាសារធាតុបញ្ជូនសរសៃប្រសាទសម្រាប់ការបញ្ជូនសរសៃប្រសាទតាមរយៈ cholinergic synapses ។ ដើម្បីអនុវត្តការរំភើបចិត្ត acetylcholine ដែលត្រូវបានបញ្ចេញទៅក្នុងប្រហោង synaptic ត្រូវតែមានអន្តរកម្មជាមួយនឹងប្រូតេអ៊ីន receptor នៃភ្នាស postsynaptic ។ រកឃើញពីរប្រភេទ អ្នកទទួល cholinergic៖
M អ្នកទទួលបន្ថែមពីលើ acetylcholine វាជ្រើសរើសអន្តរកម្មជាមួយ muscarine (fly agaric toxin) ។ M - cholinergic receptors មានវត្តមាននៅលើសាច់ដុំរលោងហើយនៅពេលដែលមានអន្តរកម្មជាមួយ acetylcholine បណ្តាលឱ្យមានការកន្ត្រាក់របស់ពួកគេ;
អ្នកទទួល Hជាពិសេសភ្ជាប់ទៅនឹងជាតិនីកូទីន។ អ្នកទទួល N-cholinergic ត្រូវបានរកឃើញនៅ synapses នៃសាច់ដុំគ្រោងឆ្អឹង។
ថ្នាំទប់ស្កាត់ជាក់លាក់ អ្នកទទួល M-cholinergicគឺ atropine ។ វាត្រូវបានរកឃើញនៅក្នុងរុក្ខជាតិ belladonna និង henbane ។
 Atropine មានក្រុមមុខងារស្រដៀងគ្នានៅក្នុងរចនាសម្ព័ន្ធទៅនឹង acetylcholine និងការរៀបចំលំហរបស់ពួកគេ ដូច្នេះវាគឺជាអ្នកទប់ស្កាត់ការប្រកួតប្រជែងនៃអ្នកទទួល M-cholinergic ។ ដោយពិចារណាថាការភ្ជាប់នៃ acetylcholine ទៅនឹងអ្នកទទួល M-cholinergic បណ្តាលឱ្យមានការកន្ត្រាក់នៃសាច់ដុំរលោង, atropine ត្រូវបានគេប្រើជាថ្នាំដែលបន្ថយការស្ពឹករបស់ពួកគេ។ (ថ្នាំ antispasmodic) ។ដូច្នេះវាត្រូវបានគេដឹងថាប្រើ atropine ដើម្បីបន្ធូរសាច់ដុំភ្នែកនៅពេលមើល fundus ក៏ដូចជាដើម្បីបំបាត់ការកន្ត្រាក់អំឡុងពេល colic ក្រពះពោះវៀន។ អ្នកទទួល M-cholinergic ក៏មានវត្តមាននៅក្នុងប្រព័ន្ធសរសៃប្រសាទកណ្តាល (CNS) ដូច្នេះកម្រិតធំនៃ atropine អាចបណ្តាលឱ្យមានប្រតិកម្មដែលមិនចង់បានពីប្រព័ន្ធសរសៃប្រសាទកណ្តាល: ការរំជើបរំជួលផ្លូវចិត្ត ការយល់ច្រឡំ ការប្រកាច់។
Atropine មានក្រុមមុខងារស្រដៀងគ្នានៅក្នុងរចនាសម្ព័ន្ធទៅនឹង acetylcholine និងការរៀបចំលំហរបស់ពួកគេ ដូច្នេះវាគឺជាអ្នកទប់ស្កាត់ការប្រកួតប្រជែងនៃអ្នកទទួល M-cholinergic ។ ដោយពិចារណាថាការភ្ជាប់នៃ acetylcholine ទៅនឹងអ្នកទទួល M-cholinergic បណ្តាលឱ្យមានការកន្ត្រាក់នៃសាច់ដុំរលោង, atropine ត្រូវបានគេប្រើជាថ្នាំដែលបន្ថយការស្ពឹករបស់ពួកគេ។ (ថ្នាំ antispasmodic) ។ដូច្នេះវាត្រូវបានគេដឹងថាប្រើ atropine ដើម្បីបន្ធូរសាច់ដុំភ្នែកនៅពេលមើល fundus ក៏ដូចជាដើម្បីបំបាត់ការកន្ត្រាក់អំឡុងពេល colic ក្រពះពោះវៀន។ អ្នកទទួល M-cholinergic ក៏មានវត្តមាននៅក្នុងប្រព័ន្ធសរសៃប្រសាទកណ្តាល (CNS) ដូច្នេះកម្រិតធំនៃ atropine អាចបណ្តាលឱ្យមានប្រតិកម្មដែលមិនចង់បានពីប្រព័ន្ធសរសៃប្រសាទកណ្តាល: ការរំជើបរំជួលផ្លូវចិត្ត ការយល់ច្រឡំ ការប្រកាច់។
Ditilin គឺជា agonist ប្រកួតប្រជែងនៃអ្នកទទួល H-cholinergic ដែលរារាំងមុខងារនៃសរសៃប្រសាទសាច់ដុំ។
 ការសំយោគសរសៃប្រសាទនៃសាច់ដុំគ្រោងឆ្អឹងមានអ្នកទទួល H-cholinergic ។ អន្តរកម្មរបស់ពួកគេជាមួយ acetylcholine នាំឱ្យមានការកន្ត្រាក់សាច់ដុំ។ ក្នុងអំឡុងពេលប្រតិបត្តិការវះកាត់មួយចំនួន ក៏ដូចជានៅក្នុងការសិក្សា endoscopic ថ្នាំត្រូវបានគេប្រើដែលបណ្តាលឱ្យសម្រាកសាច់ដុំគ្រោងឆ្អឹង។ (ថ្នាំបន្ធូរសាច់ដុំ) ។ទាំងនេះរួមមាន dithiline ដែលជា analogue រចនាសម្ព័ន្ធនៃ acetylcholine ។ វាភ្ជាប់ទៅនឹង H-cholinergic receptors ប៉ុន្តែមិនដូច acetylcholine វាត្រូវបានបំផ្លាញយឺត ៗ ដោយអង់ស៊ីម acetylcholinesterase ។ ជាលទ្ធផលនៃការបើកយូរនៃឆានែលអ៊ីយ៉ុងនិងការ depolarization ជាប់លាប់នៃភ្នាសការដឹកនាំនៃសរសៃប្រសាទត្រូវបានរំខានហើយការសម្រាកសាច់ដុំកើតឡើង។ ដំបូង លក្ខណៈសម្បត្តិទាំងនេះត្រូវបានរកឃើញនៅក្នុងថ្នាំពុល curare ដែលជាមូលហេតុដែលថ្នាំបែបនេះត្រូវបានគេហៅថា ដូច curare ។
ការសំយោគសរសៃប្រសាទនៃសាច់ដុំគ្រោងឆ្អឹងមានអ្នកទទួល H-cholinergic ។ អន្តរកម្មរបស់ពួកគេជាមួយ acetylcholine នាំឱ្យមានការកន្ត្រាក់សាច់ដុំ។ ក្នុងអំឡុងពេលប្រតិបត្តិការវះកាត់មួយចំនួន ក៏ដូចជានៅក្នុងការសិក្សា endoscopic ថ្នាំត្រូវបានគេប្រើដែលបណ្តាលឱ្យសម្រាកសាច់ដុំគ្រោងឆ្អឹង។ (ថ្នាំបន្ធូរសាច់ដុំ) ។ទាំងនេះរួមមាន dithiline ដែលជា analogue រចនាសម្ព័ន្ធនៃ acetylcholine ។ វាភ្ជាប់ទៅនឹង H-cholinergic receptors ប៉ុន្តែមិនដូច acetylcholine វាត្រូវបានបំផ្លាញយឺត ៗ ដោយអង់ស៊ីម acetylcholinesterase ។ ជាលទ្ធផលនៃការបើកយូរនៃឆានែលអ៊ីយ៉ុងនិងការ depolarization ជាប់លាប់នៃភ្នាសការដឹកនាំនៃសរសៃប្រសាទត្រូវបានរំខានហើយការសម្រាកសាច់ដុំកើតឡើង។ ដំបូង លក្ខណៈសម្បត្តិទាំងនេះត្រូវបានរកឃើញនៅក្នុងថ្នាំពុល curare ដែលជាមូលហេតុដែលថ្នាំបែបនេះត្រូវបានគេហៅថា ដូច curare ។
ប្រធានបទ 1.3 ។ ភាពចាស់នៃប្រូតេអ៊ីន និងលទ្ធភាពនៃការបង្កើតឡើងវិញដោយឯកឯងរបស់ពួកគេ
1. ចាប់តាំងពីការអនុលោមតាមប្រភពដើមនៃប្រូតេអ៊ីនត្រូវបានរក្សាដោយសារតែអន្តរកម្មខ្សោយ ការផ្លាស់ប្តូរសមាសភាព និងលក្ខណៈសម្បត្តិនៃបរិស្ថានជុំវិញប្រូតេអ៊ីន ការប៉ះពាល់នឹងសារធាតុគីមី និងកត្តារាងកាយបណ្តាលឱ្យមានការផ្លាស់ប្តូរការអនុលោមតាមរបស់វា (ទ្រព្យសម្បត្តិនៃការអនុលោមតាមលក្ខខណ្ឌ) ។ ការបំបែកចំណងមួយចំនួនធំនាំទៅដល់ការបំផ្លិចបំផ្លាញនៃការអនុលោមតាមប្រភពដើម និងការបង្ខូចទ្រង់ទ្រាយនៃប្រូតេអ៊ីន។
ការប្រែពណ៌នៃប្រូតេអ៊ីន- នេះគឺជាការបំផ្លិចបំផ្លាញនៃការអនុលោមតាមដើមរបស់ពួកគេក្រោមឥទ្ធិពលនៃភ្នាក់ងារ denaturing ដែលបណ្តាលមកពីការដាច់នៃចំណងខ្សោយដែលធ្វើឱ្យមានស្ថេរភាពនៃរចនាសម្ព័ន្ធ spatial នៃប្រូតេអ៊ីន។ Denaturation ត្រូវបានអមដោយការបំផ្លិចបំផ្លាញនៃរចនាសម្ព័ន្ធបីវិមាត្រតែមួយគត់ និងមជ្ឈមណ្ឌលសកម្មនៃប្រូតេអ៊ីន និងការបាត់បង់សកម្មភាពជីវសាស្រ្តរបស់វា (រូបភាព 1.16) ។
រាល់ម៉ូលេគុលដែលមិនមានលក្ខណៈធម្មតានៃប្រូតេអ៊ីនមួយទទួលបានទម្រង់ចៃដន្យដែលខុសពីម៉ូលេគុលផ្សេងទៀតនៃប្រូតេអ៊ីនដូចគ្នា។ រ៉ាឌីកាល់អាស៊ីតអាមីណូដែលបង្កើតជាមជ្ឈមណ្ឌលសកម្ម ប្រែទៅជាឆ្ងាយពីគ្នាទៅវិញទៅមក ពោលគឺឧ។ កន្លែងភ្ជាប់ជាក់លាក់នៃប្រូតេអ៊ីនជាមួយ ligand ត្រូវបានបំផ្លាញ។ ក្នុងអំឡុងពេល denaturation រចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីននៅតែមិនផ្លាស់ប្តូរ។
ការប្រើប្រាស់ភ្នាក់ងារ denaturing ក្នុងការស្រាវជ្រាវជីវសាស្រ្ត និងឱសថ។នៅក្នុងការសិក្សាជីវគីមី មុនពេលកំណត់សមាសធាតុទម្ងន់ម៉ូលេគុលទាបនៅក្នុងសម្ភារៈជីវសាស្រ្ត ប្រូតេអ៊ីនជាធម្មតាត្រូវបានដកចេញពីដំណោះស្រាយជាមុនសិន។ អាស៊ីត Trichloroacetic (TCA) ត្រូវបានគេប្រើញឹកញាប់បំផុតសម្រាប់គោលបំណងនេះ។ បន្ទាប់ពីបន្ថែម TCA ទៅក្នុងដំណោះស្រាយ ប្រូតេអ៊ីន denatured precipitate និងត្រូវបានយកចេញបានយ៉ាងងាយស្រួលដោយការច្រោះ (តារាង 1.1 ។ )
នៅក្នុងវេជ្ជសាស្ត្រ ភ្នាក់ងារបន្សាបជាតិពុលត្រូវបានគេប្រើជាញឹកញាប់ដើម្បីសម្លាប់ឧបករណ៍វេជ្ជសាស្រ្ត និងសម្ភារៈនៅក្នុង autoclaves (ភ្នាក់ងារ denaturing មានសីតុណ្ហភាពខ្ពស់) និងជាថ្នាំសំលាប់មេរោគ (ជាតិអាល់កុល phenol chloramine) សម្រាប់ព្យាបាលផ្ទៃកខ្វក់ដែលមាន microflora បង្កជំងឺ។
2. ប្រតិកម្មប្រូតេអ៊ីនដោយឯកឯង- ភស្តុតាងនៃការកំណត់រចនាសម្ព័ន្ធបឋម ការអនុលោមភាព និងមុខងារនៃប្រូតេអ៊ីន។ ប្រូតេអ៊ីនបុគ្គលគឺជាផលិតផលនៃហ្សែនមួយដែលមានលំដាប់អាស៊ីតអាមីណូដូចគ្នាបេះបិទ និងទទួលបានទម្រង់ដូចគ្នានៅក្នុងកោសិកា។ ការសន្និដ្ឋានជាមូលដ្ឋានដែលរចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនមានព័ត៌មានអំពីការអនុលោមតាមនិងមុខងាររបស់វារួចហើយត្រូវបានធ្វើឡើងដោយផ្អែកលើសមត្ថភាពនៃប្រូតេអ៊ីនមួយចំនួន (ជាពិសេស ribonuclease និង myoglobin) ក្នុងការបង្កើតឡើងវិញដោយឯកឯង - ស្ដារឡើងវិញនូវទម្រង់ដើមរបស់ពួកគេបន្ទាប់ពីការប្រែពណ៌។
ការបង្កើតរចនាសម្ព័ន្ធប្រូតេអ៊ីន spatial ត្រូវបានអនុវត្តដោយវិធីសាស្រ្តនៃការជួបប្រជុំគ្នាដោយខ្លួនឯង - ដំណើរការដោយឯកឯងដែលខ្សែសង្វាក់ polypeptide ដែលមានរចនាសម្ព័ន្ធបឋមតែមួយគត់មានទំនោរទៅនឹងការអនុលោមតាមថាមពលដោយឥតគិតថ្លៃទាបបំផុតនៅក្នុងដំណោះស្រាយ។ សមត្ថភាពក្នុងការបង្កើតឡើងវិញនូវប្រូតេអ៊ីនដែលរក្សារចនាសម្ព័ន្ធចម្បងរបស់ពួកគេបន្ទាប់ពីការ denaturation ត្រូវបានពិពណ៌នានៅក្នុងការពិសោធន៍ជាមួយអង់ស៊ីម ribonuclease ។
Ribonuclease គឺជាអង់ស៊ីមដែលបំបែកចំណងរវាងនុយក្លេអូទីតនីមួយៗនៅក្នុងម៉ូលេគុល RNA ។ ប្រូតេអ៊ីន globular នេះមានខ្សែសង្វាក់ polypeptide មួយដែលជារចនាសម្ព័ន្ធទីបីដែលត្រូវបានធ្វើឱ្យមានស្ថេរភាពដោយចំណងខ្សោយជាច្រើននិង disulfide បួន។
ការព្យាបាល ribonuclease ជាមួយអ៊ុយដែលបំបែកចំណងអ៊ីដ្រូសែននៅក្នុងម៉ូលេគុល និងភ្នាក់ងារកាត់បន្ថយដែលបំបែកចំណង disulfide នាំឱ្យ denaturation នៃអង់ស៊ីម និងការបាត់បង់សកម្មភាពរបស់វា។
ការយកចេញនៃភ្នាក់ងារ denaturing ដោយការលាងឈាមនាំឱ្យមានការស្ដារឡើងវិញនៃការអនុលោមតាមប្រូតេអ៊ីននិងមុខងារ, i.e. ដើម្បីកើតជាថ្មី។ (រូបភាព 1.17) ។
 អង្ករ។ ១.១៧. ការប្រែពណ៌ និងការបង្កើតឡើងវិញនៃ ribonuclease
អង្ករ។ ១.១៧. ការប្រែពណ៌ និងការបង្កើតឡើងវិញនៃ ribonuclease
ក - ការអនុលោមតាមប្រភពដើមនៃ ribonuclease នៅក្នុងរចនាសម្ព័ន្ធទីបីដែលមានចំណង disulfide បួន; ខ - ម៉ូលេគុល ribonuclease denatured;
ខ - ម៉ូលេគុល ribonuclease បានធ្វើឱ្យសកម្មឡើងវិញជាមួយនឹងរចនាសម្ព័ន្ធនិងមុខងារដែលបានស្ដារឡើងវិញ
1. បំពេញតារាង 1.2 ។
តារាង 1.2 ។ ការចាត់ថ្នាក់នៃអាស៊ីតអាមីណូយោងទៅតាមប៉ូលនៃរ៉ាឌីកាល់
2. សរសេររូបមន្តនៃ tetrapeptide៖
Asp - Pro - Fen - Liz
ក) បន្លិចក្រុមដដែលៗនៅក្នុង peptide ដែលបង្កើតជាឆ្អឹងខ្នង peptide និងក្រុមអថេរតំណាងដោយរ៉ាឌីកាល់អាស៊ីតអាមីណូ។
ខ) ដាក់ស្លាក N- និង C-termini;
គ) បន្លិចចំណង peptide;
ឃ) សរសេរ peptide មួយផ្សេងទៀតដែលមានអាស៊ីតអាមីណូដូចគ្នា;
ង) រាប់ចំនួននៃបំរែបំរួលដែលអាចកើតមាននៃ tetrapeptide ដែលមានសមាសធាតុអាស៊ីតអាមីណូស្រដៀងគ្នា។
3. ពន្យល់ពីតួនាទីនៃរចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីន ដោយប្រើឧទាហរណ៍នៃការវិភាគប្រៀបធៀបនៃរចនាសម្ព័ន្ធស្រដៀងគ្នា និងការវិវត្តនៃអរម៉ូន peptide ជិតស្និទ្ធនៃថនិកសត្វ neurohypophysis - អុកស៊ីតូស៊ីន និង vasopressin (តារាង 1.3) ។
តារាង 1.3 ។ រចនាសម្ព័ន្ធនិងមុខងារនៃអុកស៊ីតូស៊ីននិង vasopressin
 សម្រាប់ការនេះ:
សម្រាប់ការនេះ:
ក) ប្រៀបធៀបសមាសភាពនិងលំដាប់អាស៊ីតអាមីណូនៃ peptides ពីរ;
ខ) ស្វែងរកភាពស្រដៀងគ្នានៃរចនាសម្ព័ន្ធចម្បងនៃ peptides ទាំងពីរនិងភាពស្រដៀងគ្នានៃសកម្មភាពជីវសាស្រ្តរបស់ពួកគេ;
គ) រកឃើញភាពខុសគ្នានៅក្នុងរចនាសម្ព័ន្ធនៃ peptides ពីរនិងភាពខុសគ្នានៅក្នុងមុខងាររបស់ពួកគេ;
ឃ) ធ្វើការសន្និដ្ឋានអំពីឥទ្ធិពលនៃរចនាសម្ព័ន្ធចម្បងនៃ peptides លើមុខងាររបស់វា។
4. ពិពណ៌នាអំពីដំណាក់កាលសំខាន់នៃការបង្កើតការអនុលោមតាមប្រូតេអ៊ីន globular (រចនាសម្ព័ន្ធបន្ទាប់បន្សំ, ទីបី, គំនិតនៃរចនាសម្ព័ន្ធអនុវិទ្យាល័យ) ។ ចង្អុលបង្ហាញប្រភេទនៃចំណងដែលពាក់ព័ន្ធនឹងការបង្កើតរចនាសម្ព័ន្ធប្រូតេអ៊ីន។ ដែលរ៉ាឌីកាល់អាស៊ីតអាមីណូអាចចូលរួមក្នុងការបង្កើតអន្តរកម្ម hydrophobic, អ៊ីយ៉ុង, ចំណងអ៊ីដ្រូសែន។
ផ្តល់ឧទាហរណ៍។
5. កំណត់គំនិតនៃ "ភាពទន់ខ្សោយនៃប្រូតេអ៊ីន" បង្ហាញពីហេតុផលសម្រាប់អត្ថិភាព និងសារៈសំខាន់របស់វា។
6. ពង្រីកអត្ថន័យនៃឃ្លាខាងក្រោម៖ "មុខងារនៃប្រូតេអ៊ីនគឺផ្អែកលើអន្តរកម្មជាក់លាក់របស់វាជាមួយ ligand" ដោយប្រើពាក្យ និងពន្យល់អត្ថន័យរបស់វា៖ ការអនុលោមតាមប្រូតេអ៊ីន មជ្ឈមណ្ឌលសកម្ម លីហ្គែន ការបំពេញបន្ថែម មុខងារប្រូតេអ៊ីន។
7. ដោយប្រើឧទាហរណ៍មួយ ពន្យល់ពីអ្វីដែលដែនមាន និងតួនាទីរបស់វានៅក្នុងដំណើរការនៃប្រូតេអ៊ីន។
ភារកិច្ចគ្រប់គ្រងខ្លួនឯង
1. ការប្រកួត។
ក្រុមមុខងារនៅក្នុងរ៉ាឌីកាល់អាស៊ីតអាមីណូ៖
A. ក្រុម Carboxyl B. Hydroxyl ក្រុម C Guanidine ក្រុម D. Thiol ក្រុម E. Amino ក្រុម
2. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។
អាស៊ីតអាមីណូដែលមានរ៉ាឌីកាល់គ្មានប៉ូលគឺ៖
A. Cis B. Asn
B. Glu G. បី
3. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។
រ៉ាឌីកាល់អាស៊ីតអាមីណូ៖
A. ផ្តល់ភាពជាក់លាក់នៃរចនាសម្ព័ន្ធបឋម B. ចូលរួមក្នុងការបង្កើតរចនាសម្ព័ន្ធទីបី
B. មានទីតាំងនៅលើផ្ទៃប្រូតេអ៊ីន ពួកវាមានឥទ្ធិពលលើការរលាយរបស់វា D. បង្កើតជាមជ្ឈមណ្ឌលសកម្ម
ឃ. ចូលរួមក្នុងការបង្កើតចំណង peptide
4. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។
អន្តរកម្ម Hydrophobic អាចបង្កើតរវាងរ៉ាឌីកាល់អាស៊ីតអាមីណូ៖
A. Tre Lay B. Pro បី
B. បានជួប Ile G. Tir Ala D. Val Fen
5. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។
ចំណងអ៊ីយ៉ុងអាចបង្កើតរវាងរ៉ាឌីកាល់អាស៊ីតអាមីណូ៖
A. Gln Asp B. Apr Liz
B. Liz Glu G. Gis Asp D. Asn Apr
6. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។
ចំណងអ៊ីដ្រូសែនអាចបង្កើតបានរវាងរ៉ាឌីកាល់អាស៊ីតអាមីណូ៖
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. ការប្រកួត។
ប្រភេទនៃចំណងពាក់ព័ន្ធនឹងការបង្កើតរចនាសម្ព័ន្ធប្រូតេអ៊ីន៖
A. រចនាសម្ព័ន្ធបឋម B. រចនាសម្ព័ន្ធបន្ទាប់បន្សំ
ខ. រចនាសម្ព័ន្ធថ្នាក់ឧត្តមសិក្សា
ឃ. រចនាសម្ព័ន្ធអនុវិទ្យាល័យ E. អនុលោមភាព។
1. ចំណងអ៊ីដ្រូសែនរវាងអាតូមនៃឆ្អឹងខ្នង peptide
2. ចំណងខ្សោយរវាងក្រុមមុខងារនៃរ៉ាឌីកាល់អាស៊ីតអាមីណូ
3. ចំណងរវាងក្រុម α-amino និង α-carboxyl នៃអាស៊ីតអាមីណូ
8. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។ ទ្រីបស៊ីន៖
A. អង់ស៊ីម Proteolytic B. មានដែនពីរ
ខ. Hydrolyzes ម្សៅ
ឃ. គេហទំព័រសកម្មមានទីតាំងនៅចន្លោះដែន។ ឃ. មានខ្សែសង្វាក់ polypeptide ពីរ។
9. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។ Atropine៖
A. សារធាតុបញ្ជូនសរសៃប្រសាទ
ខ. រចនាសម្ព័ន្ធ analogue នៃ acetylcholine
B. អន្តរកម្មជាមួយអ្នកទទួល H-cholinergic
D. ពង្រឹងការដឹកនាំនៃសរសៃប្រសាទតាមរយៈ cholinergic synapses
ឃ. អ្នករារាំងការប្រកួតប្រជែងនៃអ្នកទទួល M-cholinergic
10. ជ្រើសរើសសេចក្តីថ្លែងការណ៍ត្រឹមត្រូវ។ នៅក្នុងប្រូតេអ៊ីន៖
A. រចនាសម្ព័ន្ធចម្បងមានព័ត៌មានអំពីរចនាសម្ព័ន្ធនៃគេហទំព័រសកម្មរបស់វា។
ខ.មជ្ឈមណ្ឌលសកម្មត្រូវបានបង្កើតឡើងនៅកម្រិតនៃរចនាសម្ព័ន្ធបឋម
ខ. ការអនុលោមភាពត្រូវបានជួសជុលយ៉ាងតឹងរ៉ឹងដោយចំណង covalent
D. គេហទំព័រសកម្មអាចធ្វើអន្តរកម្មជាមួយក្រុមនៃ ligands ស្រដៀងគ្នា
ដោយសារការអនុលោមភាពនៃប្រូតេអ៊ីន D. ការផ្លាស់ប្តូរនៅក្នុងបរិស្ថានអាចប៉ះពាល់ដល់ភាពស្និទ្ធស្នាលនៃសកម្ម
កណ្តាលទៅ ligand
1. 1-B, 2-G, 3-B ។
3. A, B, C, D ។
7. 1-B, 2-D, 3-A ។
8. A, B, C, D ។
លក្ខខណ្ឌមូលដ្ឋាន និងគោលគំនិត
1. ប្រូតេអ៊ីន polypeptide អាស៊ីតអាមីណូ
2. រចនាសម្ព័ន្ធប្រូតេអ៊ីនបឋម, អនុវិទ្យាល័យ, ទីបី
3. ការអនុលោមតាមប្រូតេអ៊ីនកំណើត
4. ចំណង covalent និងខ្សោយនៅក្នុងប្រូតេអ៊ីន
5. ការអនុលោមភាព lability
6. កន្លែងសកម្មប្រូតេអ៊ីន
7. លីហ្គែន
8. ផ្នត់ប្រូតេអ៊ីន
9. analogues រចនាសម្ព័ន្ធនៃ ligands
10. ប្រូតេអ៊ីនដែន
11. ប្រូតេអ៊ីនសាមញ្ញនិងស្មុគស្មាញ
12. ប្រូតេអ៊ីន denaturation ភ្នាក់ងារ denaturing
13. ការធ្វើឱ្យសកម្មនៃប្រូតេអ៊ីន
ដោះស្រាយបញ្ហា
"ការរៀបចំរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីននិងមូលដ្ឋាននៃមុខងាររបស់ពួកគេ"
1. មុខងារសំខាន់នៃប្រូតេអ៊ីន - អេម៉ូក្លូប៊ីន A (HbA) គឺជាការដឹកជញ្ជូនអុកស៊ីសែនទៅជាលិកា។ នៅក្នុងមនុស្ស ទម្រង់ជាច្រើននៃប្រូតេអ៊ីននេះត្រូវបានគេស្គាល់ជាមួយនឹងលក្ខណៈសម្បត្តិ និងមុខងារផ្លាស់ប្តូរ - អ្វីដែលគេហៅថា អេម៉ូក្លូប៊ីនមិនធម្មតា។ ឧទាហរណ៍ អេម៉ូក្លូប៊ីន S ដែលត្រូវបានរកឃើញនៅក្នុងកោសិកាឈាមក្រហមរបស់អ្នកជំងឺដែលមានជំងឺកោសិកាឈឺ (HbS) ត្រូវបានគេរកឃើញថាមានភាពរលាយទាបក្រោមលក្ខខណ្ឌនៃសម្ពាធផ្នែកទាបនៃអុកស៊ីសែន (ដូចករណីនៅក្នុងសរសៃឈាមវ៉ែន)។ នេះនាំឱ្យមានការបង្កើតការប្រមូលផ្តុំនៃប្រូតេអ៊ីននេះ។ ប្រូតេអ៊ីនបាត់បង់មុខងាររបស់វា precipitates ហើយកោសិកាឈាមក្រហមប្រែជារាងមិនទៀងទាត់ (ពួកវាខ្លះបង្កើតជារាងពងក្រពើ) ហើយត្រូវបានបំផ្លាញលឿនជាងធម្មតានៅក្នុងលំពែង។ ជាលទ្ធផល ភាពស្លេកស្លាំងក្នុងកោសិកាមានការរីកចម្រើន។
ភាពខុសគ្នាតែមួយគត់នៅក្នុងរចនាសម្ព័ន្ធបឋមនៃ HbA ត្រូវបានរកឃើញនៅក្នុងតំបន់ N-terminal នៃ hemoglobin β-chain ។ ប្រៀបធៀបតំបន់ N-terminal នៃβ-strand និងបង្ហាញពីរបៀបដែលការផ្លាស់ប្តូររចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនប៉ះពាល់ដល់លក្ខណៈសម្បត្តិ និងមុខងាររបស់វា។
 សម្រាប់ការនេះ:
សម្រាប់ការនេះ:
ក) សរសេររូបមន្តនៃអាស៊ីតអាមីណូដែល HbA ខុសគ្នា និងប្រៀបធៀបលក្ខណៈសម្បត្តិនៃអាស៊ីតអាមីណូទាំងនេះ (ប៉ូល, បន្ទុក) ។
ខ) ធ្វើការសន្និដ្ឋានអំពីមូលហេតុនៃការថយចុះនៃការរលាយ និងការរំខាននៃការដឹកជញ្ជូនអុកស៊ីសែនទៅក្នុងជាលិកា។
2. តួលេខបង្ហាញពីដ្យាក្រាមនៃរចនាសម្ព័ន្ធប្រូតេអ៊ីនដែលមានមជ្ឈមណ្ឌលភ្ជាប់ជាមួយ ligand (មជ្ឈមណ្ឌលសកម្ម) ។ ពន្យល់ពីមូលហេតុដែលប្រូតេអ៊ីនត្រូវបានជ្រើសរើសនៅក្នុងជម្រើសរបស់វានៃ ligand ។ សម្រាប់ការនេះ:
ក) ចងចាំនូវអ្វីដែលជាមជ្ឈមណ្ឌលសកម្មនៃប្រូតេអ៊ីន ហើយពិចារណាពីរចនាសម្ព័ន្ធនៃមជ្ឈមណ្ឌលសកម្មនៃប្រូតេអ៊ីនដែលបង្ហាញក្នុងរូប។
ខ) សរសេររូបមន្តនៃរ៉ាឌីកាល់អាស៊ីតអាមីណូដែលបង្កើតជាមជ្ឈមណ្ឌលសកម្ម។
គ) គូរ ligand ដែលអាចធ្វើអន្តរកម្មជាពិសេសជាមួយទីតាំងសកម្មនៃប្រូតេអ៊ីន។ ចង្អុលបង្ហាញនៅលើវាក្រុមមុខងារដែលអាចបង្កើតចំណងជាមួយរ៉ាឌីកាល់អាស៊ីតអាមីណូដែលបង្កើតជាមជ្ឈមណ្ឌលសកម្ម;
ឃ) បង្ហាញពីប្រភេទនៃចំណងដែលកើតឡើងរវាង ligand និងរ៉ាឌីកាល់អាស៊ីតអាមីណូនៃមជ្ឈមណ្ឌលសកម្ម;
ង) ពន្យល់ពីអ្វីដែលជាក់លាក់នៃអន្តរកម្មប្រូតេអ៊ីន-លីហ្គែនគឺផ្អែកលើ។
 3.
តួលេខបង្ហាញពីទីតាំងសកម្មនៃប្រូតេអ៊ីន និងលីហ្គែនមួយចំនួន។
3.
តួលេខបង្ហាញពីទីតាំងសកម្មនៃប្រូតេអ៊ីន និងលីហ្គែនមួយចំនួន។
កំណត់ថាតើ ligand មួយណាទំនងជាមានអន្តរកម្មជាមួយទីតាំងសកម្មនៃប្រូតេអ៊ីន ហើយហេតុអ្វី។
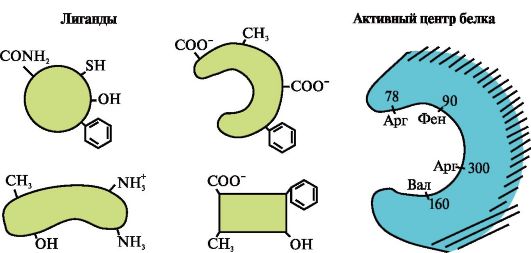 តើមូលបត្របំណុលប្រភេទណាខ្លះដែលកើតឡើងកំឡុងពេលបង្កើតស្មុគស្មាញប្រូតេអ៊ីន-លីហ្គែន?
តើមូលបត្របំណុលប្រភេទណាខ្លះដែលកើតឡើងកំឡុងពេលបង្កើតស្មុគស្មាញប្រូតេអ៊ីន-លីហ្គែន?
4. analogues រចនាសម្ព័ន្ធនៃ ligands ប្រូតេអ៊ីនធម្មជាតិអាចត្រូវបានប្រើជាថ្នាំដើម្បីកែប្រែសកម្មភាពនៃប្រូតេអ៊ីន។
Acetylcholine គឺជាអ្នកសម្រុះសម្រួលនៃការបញ្ជូនរំភើបនៅ synapses neuromuscular ។ នៅពេលដែល acetylcholine ធ្វើអន្តរកម្មជាមួយប្រូតេអ៊ីន - អ្នកទទួលភ្នាស postsynaptic នៃសាច់ដុំគ្រោងឆ្អឹង បណ្តាញអ៊ីយ៉ុងបើក ហើយការកន្ត្រាក់សាច់ដុំកើតឡើង។ Ditilin គឺជាថ្នាំដែលប្រើក្នុងប្រតិបត្តិការមួយចំនួនដើម្បីបន្ធូរសាច់ដុំព្រោះវារំខានដល់ការបញ្ជូនសរសៃប្រសាទតាមរយៈសរសៃប្រសាទសាច់ដុំ។ ពន្យល់ពីយន្តការនៃសកម្មភាពរបស់ឌីទីលីនជាថ្នាំបន្ធូរសាច់ដុំ។ សម្រាប់ការនេះ:
ក) សរសេររូបមន្តនៃ acetylcholine និង dithiline ហើយប្រៀបធៀបរចនាសម្ព័ន្ធរបស់ពួកគេ;
ខ) ពិពណ៌នាអំពីយន្តការនៃឥទ្ធិពលបន្ធូរបន្ថយនៃឌីទីលីន។
5. នៅក្នុងជំងឺមួយចំនួនសីតុណ្ហភាពរាងកាយរបស់អ្នកជំងឺកើនឡើងដែលត្រូវបានគេចាត់ទុកថាជាប្រតិកម្មការពារនៃរាងកាយ។ ទោះជាយ៉ាងណាក៏ដោយ សីតុណ្ហភាពខ្ពស់គឺធ្វើឱ្យខូចដល់ប្រូតេអ៊ីនរាងកាយ។ ពន្យល់ពីមូលហេតុដែលនៅសីតុណ្ហភាពលើសពី 40 °C មុខងាររបស់ប្រូតេអ៊ីនត្រូវបានរំខាន ហើយការគំរាមកំហែងដល់ជីវិតមនុស្សកើតឡើង។ ដើម្បីធ្វើដូចនេះសូមចាំថា:
1) រចនាសម្ព័ន្ធនៃប្រូតេអ៊ីននិងចំណងដែលរក្សារចនាសម្ព័ន្ធរបស់វានៅក្នុងការអនុលោមតាមប្រភពដើម;
2) តើរចនាសម្ព័ន្ធ និងមុខងាររបស់ប្រូតេអ៊ីនប្រែប្រួលយ៉ាងដូចម្តេច ជាមួយនឹងការកើនឡើងសីតុណ្ហភាព?
3) តើ homeostasis ជាអ្វី ហើយហេតុអ្វីបានជាវាសំខាន់សម្រាប់ការថែរក្សាសុខភាពមនុស្ស។
ឯកតាម៉ូឌុល 2 OLIGOMERICK PROTEINS ជាគោលដៅនៃឥទ្ធិពលនិយតកម្ម។ ភាពចម្រុះនៃរចនាសម្ព័ន្ធ និងមុខងារនៃប្រូតេអ៊ីន។ វិធីសាស្រ្តបំបែក និងការបន្សុតប្រូតេអ៊ីន
គោលបំណងនៃការសិក្សាអាច៖
1. ប្រើចំណេះដឹងអំពីលក្ខណៈពិសេសនៃរចនាសម្ព័ន្ធនិងមុខងារនៃប្រូតេអ៊ីន oligomeric ដើម្បីយល់ពីយន្តការសម្របខ្លួននៃបទប្បញ្ញត្តិនៃមុខងាររបស់វា។
2. ពន្យល់ពីតួនាទីរបស់ chaperones ក្នុងការសំយោគ និងថែរក្សាការអនុលោមតាមប្រូតេអ៊ីនក្រោមលក្ខខណ្ឌកោសិកា។
3. ពន្យល់ពីភាពខុសគ្នានៃការបង្ហាញនៃជីវិតដោយភាពខុសគ្នានៃរចនាសម្ព័ន្ធនិងមុខងារនៃប្រូតេអ៊ីនដែលត្រូវបានសំយោគនៅក្នុងរាងកាយ។
4. វិភាគទំនាក់ទំនងរវាងរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីននិងមុខងាររបស់វាដោយប្រើឧទាហរណ៍នៃការប្រៀបធៀបនៃ hemoproteins ពាក់ព័ន្ធ - myoglobin និង hemoglobin ក៏ដូចជាតំណាងនៃក្រុមប្រូតេអ៊ីនទាំងប្រាំនៃគ្រួសារ immunoglobulin ។
5. អនុវត្តចំណេះដឹងអំពីលក្ខណៈពិសេសនៃលក្ខណៈសម្បត្តិរូបវន្ត និងគីមីនៃប្រូតេអ៊ីន ដើម្បីជ្រើសរើសវិធីសាស្រ្តសម្រាប់ការបន្សុតរបស់ពួកគេពីប្រូតេអ៊ីន និងសារធាតុមិនបរិសុទ្ធផ្សេងទៀត។
6. បកស្រាយលទ្ធផលនៃសមាសភាពបរិមាណនិងគុណភាពនៃប្រូតេអ៊ីនប្លាស្មាឈាមដើម្បីបញ្ជាក់ឬបញ្ជាក់ការធ្វើរោគវិនិច្ឆ័យគ្លីនិក។
ដឹង៖
1. លក្ខណៈពិសេសនៃរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីន oligomeric និងយន្តការសម្របខ្លួនសម្រាប់ធ្វើនិយតកម្មមុខងាររបស់ពួកគេដោយប្រើឧទាហរណ៍នៃអេម៉ូក្លូប៊ីន។
2. រចនាសម្ព័ននិងមុខងាររបស់ chaperones និងសារៈសំខាន់របស់ពួកគេសម្រាប់ការរក្សាការអនុលោមតាមប្រភពដើមនៃប្រូតេអ៊ីននៅក្រោមលក្ខខណ្ឌកោសិកា។
3. គោលការណ៍នៃការបញ្ចូលគ្នានៃប្រូតេអ៊ីនចូលទៅក្នុងគ្រួសារដោយផ្អែកលើភាពស្រដៀងគ្នានៃការអនុលោមភាពនិងមុខងាររបស់ពួកគេដោយប្រើឧទាហរណ៍នៃ immunoglobulins ។
4. វិធីសាស្រ្តបំបែកប្រូតេអ៊ីនដោយផ្អែកលើលក្ខណៈនៃលក្ខណៈសម្បត្តិរូបវិទ្យារបស់ពួកគេ។
5. Electrophoresis នៃប្លាស្មាឈាម ជាវិធីសាស្រ្តសម្រាប់វាយតម្លៃសមាសភាពគុណភាព និងបរិមាណនៃប្រូតេអ៊ីន។
ប្រធានបទ 1.4 ។ លក្ខណៈពិសេសនៃរចនាសម្ព័ន្ធនិងមុខងារនៃប្រូតេអ៊ីន OLIGOMERIC ដោយប្រើឧទាហរណ៍នៃ hemoglobin
1. ប្រូតេអ៊ីនជាច្រើនមានខ្សែសង្វាក់ polypeptide ជាច្រើន។ ប្រូតេអ៊ីនបែបនេះត្រូវបានគេហៅថា អូលីហ្គោមិកនិងខ្សែសង្វាក់បុគ្គល - protomers ។ Protomers នៅក្នុងប្រូតេអ៊ីន oligomeric ត្រូវបានតភ្ជាប់ដោយចំណងដែលមិនមែនជាកូវ៉ាលេនខ្សោយជាច្រើន (hydrophobic, ionic, hydrogen) ។ អន្តរកម្ម
protomers ត្រូវបានអនុវត្តដោយអរគុណ ការបំពេញបន្ថែមផ្ទៃទំនាក់ទំនងរបស់ពួកគេ។
ចំនួន protomers នៅក្នុងប្រូតេអ៊ីន oligomeric អាចប្រែប្រួលយ៉ាងខ្លាំង៖ អេម៉ូក្លូប៊ីនមាន 4 protomers អង់ស៊ីម aspartate aminotransferase មាន 12 protomers និងប្រូតេអ៊ីនថ្នាំជក់ mosaic virus មាន protomers 2120 ដែលតភ្ជាប់ដោយចំណងមិនមែន covalent ។ ដូច្នេះ ប្រូតេអ៊ីន oligomeric អាចមានទម្ងន់ម៉ូលេគុលខ្ពស់ណាស់។
អន្តរកម្មរបស់ protomer មួយជាមួយអ្នកដទៃអាចត្រូវបានចាត់ទុកថាជាករណីពិសេសនៃអន្តរកម្មប្រូតេអ៊ីន-ligand ចាប់តាំងពី protomer នីមួយៗដើរតួជា ligand សម្រាប់ protomers ផ្សេងទៀត។ ចំនួននិងវិធីសាស្រ្តនៃការចូលរួម protomers នៅក្នុងប្រូតេអ៊ីនត្រូវបានគេហៅថា រចនាសម្ព័ន្ធប្រូតេអ៊ីន quaternary ។
ប្រូតេអ៊ីនអាចមាន protomers នៃរចនាសម្ព័ន្ធដូចគ្នា ឬផ្សេងគ្នា ឧទាហរណ៍ homodimers គឺជាប្រូតេអ៊ីនដែលមាន protomers ដូចគ្នាពីរ ហើយ heterodimers គឺជាប្រូតេអ៊ីនដែលមាន protomers ពីរផ្សេងគ្នា។
ប្រសិនបើប្រូតេអ៊ីនមាន protomers ផ្សេងគ្នា នោះមជ្ឈមណ្ឌលចងជាមួយនឹង ligands ផ្សេងគ្នាដែលរចនាសម្ព័ន្ធខុសគ្នាអាចត្រូវបានបង្កើតឡើងនៅលើពួកវា។ នៅពេលដែល ligand ភ្ជាប់ទៅកន្លែងសកម្ម មុខងារនៃប្រូតេអ៊ីននេះត្រូវបានបង្ហាញ។ មជ្ឈមណ្ឌលដែលមានទីតាំងនៅលើ protomer ផ្សេងគ្នាត្រូវបានគេហៅថា allosteric (ខុសពីកន្លែងសកម្ម) ។ ទំនាក់ទំនង allosteric ligand ឬ effector,វាអនុវត្តមុខងារនិយតកម្ម (រូបភាព 1.18) ។ អន្តរកម្មនៃមជ្ឈមណ្ឌល allosteric ជាមួយ effector បណ្តាលឱ្យមានការផ្លាស់ប្តូរទម្រង់នៅក្នុងរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីន oligomeric ទាំងមូលដោយសារតែភាពមិនអនុលោមតាមទម្រង់របស់វា។ វាប៉ះពាល់ដល់ភាពស្និទ្ធស្នាលនៃគេហទំព័រសកម្មសម្រាប់ ligand ជាក់លាក់មួយ និងគ្រប់គ្រងមុខងារនៃប្រូតេអ៊ីននោះ។ ការផ្លាស់ប្តូរនៅក្នុងការអនុលោមតាម និងមុខងាររបស់ protomers ទាំងអស់ក្នុងអំឡុងពេលអន្តរកម្មនៃប្រូតេអ៊ីន oligomeric ជាមួយ ligand យ៉ាងហោចណាស់មួយត្រូវបានគេហៅថា cooperative conformational change។ ឥទ្ធិពលដែលបង្កើនមុខងារប្រូតេអ៊ីនត្រូវបានគេហៅថា សកម្មភាព,និងឥទ្ធិពលដែលរារាំងមុខងាររបស់វា - ថ្នាំទប់ស្កាត់។
ដូច្នេះប្រូតេអ៊ីន oligomeric ក៏ដូចជាប្រូតេអ៊ីនដែលមានរចនាសម្ព័ន្ធដែនមានទ្រព្យសម្បត្តិថ្មីបើប្រៀបធៀបទៅនឹងប្រូតេអ៊ីន monomeric - សមត្ថភាពក្នុងការគ្រប់គ្រងមុខងារ allosterically (បទប្បញ្ញត្តិដោយការភ្ជាប់ ligands ផ្សេងគ្នាទៅនឹងប្រូតេអ៊ីន) ។ នេះអាចត្រូវបានគេមើលឃើញដោយការប្រៀបធៀបរចនាសម្ព័ន្ធនិងមុខងារនៃប្រូតេអ៊ីនស្មុគស្មាញពីរដែលទាក់ទងយ៉ាងជិតស្និទ្ធគឺ myoglobin និង hemoglobin ។
 អង្ករ។ ១.១៨. គ្រោងការណ៍នៃរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីន dimeric មួយ។
អង្ករ។ ១.១៨. គ្រោងការណ៍នៃរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីន dimeric មួយ។
2. ការបង្កើតរចនាសម្ព័ន្ធលំហ និងដំណើរការនៃ myoglobin ។
Myoglobin (Mb) គឺជាប្រូតេអ៊ីនដែលមាននៅក្នុងសាច់ដុំក្រហម ដែលមុខងារសំខាន់គឺបង្កើតទុនបម្រុង O 2 ដែលចាំបាច់សម្រាប់ការងារសាច់ដុំខ្លាំង។ Mb គឺជាប្រូតេអ៊ីនស្មុគស្មាញដែលមានផ្នែកប្រូតេអ៊ីន - apoMb និងផ្នែកដែលមិនមានប្រូតេអ៊ីន - heme ។ រចនាសម្ព័ន្ធចម្បងនៃ apoMB កំណត់ការអនុលោមតាមរាងពងក្រពើតូចរបស់វា និងរចនាសម្ព័ន្ធនៃមជ្ឈមណ្ឌលសកម្ម ដែលផ្នែកដែលមិនមែនជាប្រូតេអ៊ីននៃ myoglobin, heme ត្រូវបានភ្ជាប់។ អុកស៊ីសែនចេញមកពីឈាមទៅសាច់ដុំភ្ជាប់ទៅនឹង Fe + 2 hemes នៅក្នុង myoglobin ។ Mb គឺជាប្រូតេអ៊ីន monomeric ដែលមានទំនាក់ទំនងខ្ពស់សម្រាប់ O 2 ដូច្នេះការបញ្ចេញអុកស៊ីសែនដោយ myoglobin កើតឡើងតែក្នុងអំឡុងពេលការងារសាច់ដុំខ្លាំងនៅពេលដែលសម្ពាធផ្នែកនៃ O 2 ថយចុះយ៉ាងខ្លាំង។
ការបង្កើតទម្រង់ Mv ។នៅក្នុងសាច់ដុំក្រហមនៅលើ ribosomes កំឡុងពេលបកប្រែ រចនាសម្ព័ន្ធចម្បងនៃ MB ត្រូវបានសំយោគ ដែលតំណាងដោយលំដាប់ជាក់លាក់នៃសំណល់អាស៊ីតអាមីណូចំនួន 153 ។ រចនាសម្ព័ន្ធបន្ទាប់បន្សំនៃ Mb មាន α-helices ចំនួនប្រាំបី ដែលហៅថាជាអក្សរឡាតាំងពី A ដល់ H ដែលចន្លោះនោះមានតំបន់ដែលមិនមែនជា helical ។ រចនាសម្ព័នទីបីនៃ Mb មានទម្រង់ជា globule បង្រួម នៅក្នុងការសម្រាកដែលមជ្ឈមណ្ឌលសកម្មស្ថិតនៅចន្លោះ F និង E α-helices (រូបភាព 1.19) ។
 អង្ករ។ ១.១៩. រចនាសម្ព័ន្ធ Myoglobin
អង្ករ។ ១.១៩. រចនាសម្ព័ន្ធ Myoglobin
3. លក្ខណៈពិសេសនៃរចនាសម្ព័ន្ធនិងមុខងារនៃមជ្ឈមណ្ឌលសកម្មនៃ MV ។មជ្ឈមណ្ឌលសកម្មនៃ Mb ត្រូវបានបង្កើតឡើងជាចម្បងដោយរ៉ាឌីកាល់អាស៊ីតអាមីណូ hydrophobic ដែលមានគម្លាតយ៉ាងទូលំទូលាយពីគ្នាទៅវិញទៅមកនៅក្នុងរចនាសម្ព័ន្ធបឋម (ឧទាហរណ៍ Tri 3 9 និង Fen 138) រលាយមិនល្អនៅក្នុងលីហ្គែនទឹក - heme និង O 2 - ភ្ជាប់ទៅនឹងមជ្ឈមណ្ឌលសកម្ម។ Heme គឺជា ligand ជាក់លាក់នៃ apoMB (Fig ។ 1.20), មូលដ្ឋាននៃការដែលត្រូវបានបង្កើតឡើងដោយចិញ្ចៀន pyrrole បួនតភ្ជាប់ដោយស្ពានមេទីល; នៅចំកណ្តាលមានអាតូម Fe + 2 ភ្ជាប់ទៅនឹងអាតូមអាសូតនៃចិញ្ចៀន pyrrole ដោយចំណងសំរបសំរួលចំនួនបួន។ នៅក្នុងមជ្ឈមណ្ឌលសកម្មនៃ Mb បន្ថែមពីលើរ៉ាឌីកាល់អាស៊ីតអាមីណូ hydrophobic វាក៏មានសំណល់នៃអាស៊ីតអាមីណូពីរដែលមានរ៉ាឌីកាល់ hydrophilic - Gis E ៧(ជីស ៦៤) និង GIS F ៨(៩៣ របស់ទ្រង់) (រូប ១.២១)។
 អង្ករ។ ១.២០. រចនាសម្ព័ន្ធនៃ heme - ផ្នែកដែលមិនមែនជាប្រូតេអ៊ីននៃ myoglobin និង hemoglobin
អង្ករ។ ១.២០. រចនាសម្ព័ន្ធនៃ heme - ផ្នែកដែលមិនមែនជាប្រូតេអ៊ីននៃ myoglobin និង hemoglobin
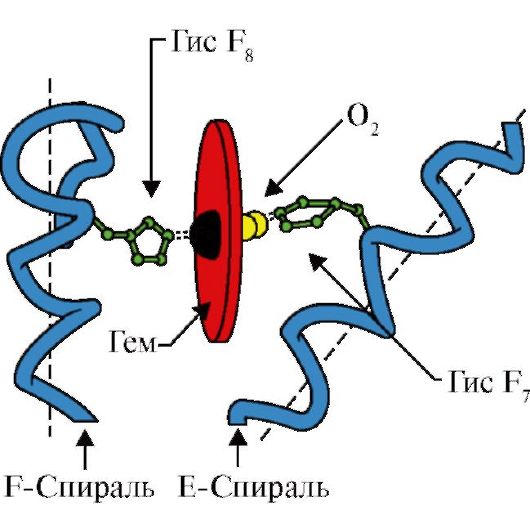 អង្ករ។ ១.២១. ទីតាំងនៃ heme និង O2 នៅក្នុងទីតាំងសកម្មនៃ apomyoglobin និង hemoglobin protomers
អង្ករ។ ១.២១. ទីតាំងនៃ heme និង O2 នៅក្នុងទីតាំងសកម្មនៃ apomyoglobin និង hemoglobin protomers
Heme ត្រូវបានចងភ្ជាប់ជាមួយ F8 របស់គាត់តាមរយៈអាតូមដែក។ O 2 ភ្ជាប់ទៅនឹងដែកនៅផ្នែកម្ខាងទៀតនៃយន្តហោះ heme ។ E 7 របស់គាត់គឺចាំបាច់សម្រាប់ការតំរង់ទិសត្រឹមត្រូវនៃ O 2 និងសម្របសម្រួលការបន្ថែមអុកស៊ីសែនទៅ Fe + 2 heme
GIS F ៨បង្កើតចំណងសំរបសំរួលជាមួយ Fe+ 2 ហើយជួសជុលយ៉ាងរឹងមាំនូវ heme នៅក្នុងមជ្ឈមណ្ឌលសកម្ម។ Gis E ៧ចាំបាច់សម្រាប់ការតំរង់ទិសត្រឹមត្រូវនៅក្នុងមជ្ឈមណ្ឌលសកម្មនៃ ligand មួយផ្សេងទៀត - O 2 កំឡុងពេលអន្តរកម្មរបស់វាជាមួយ Fe + 2 heme ។ microenvironment នៃ heme បង្កើតលក្ខខណ្ឌសម្រាប់ការចងដ៏រឹងមាំ ប៉ុន្តែអាចបញ្ច្រាស់បាននៃ O 2 ទៅ Fe + 2 និងការពារទឹកពីការចូលទៅក្នុងកន្លែងសកម្ម hydrophobic ដែលអាចនាំឱ្យមានអុកស៊ីតកម្មរបស់វាទៅ Fe + 3 ។
រចនាសម្ព័ន្ធ monomeric នៃ Mb និងមជ្ឈមណ្ឌលសកម្មរបស់វាកំណត់ទំនាក់ទំនងខ្ពស់នៃប្រូតេអ៊ីនសម្រាប់ O 2 ។
4. រចនាសម្ព័ន្ធ Oligomeric នៃ Hb និងបទប្បញ្ញត្តិនៃទំនាក់ទំនងនៃ Hb ទៅ O 2 ligands ។ អេម៉ូក្លូប៊ីនរបស់មនុស្ស- ក្រុមគ្រួសារនៃប្រូតេអ៊ីនដូចជា myoglobin ទាក់ទងទៅនឹងប្រូតេអ៊ីនស្មុគស្មាញ (hemoproteins) ។ ពួកវាមានរចនាសម្ព័ន្ធ tetrameric និងមានខ្សែសង្វាក់ α ពីរ ប៉ុន្តែមានភាពខុសប្លែកគ្នានៅក្នុងរចនាសម្ព័ន្ធនៃខ្សែសង្វាក់ polypeptide ពីរផ្សេងទៀត (2α-, 2x-chains) ។ រចនាសម្ព័ន្ធនៃខ្សែសង្វាក់ polypeptide ទីពីរកំណត់លក្ខណៈពិសេសនៃដំណើរការនៃទម្រង់ Hb ទាំងនេះ។ ប្រហែល 98% នៃអេម៉ូក្លូប៊ីននៅក្នុងកោសិកាឈាមក្រហមរបស់មនុស្សពេញវ័យគឺ អេម៉ូក្លូប៊ីន A(2α-, 2p-chains) ។
ក្នុងអំឡុងពេលនៃការលូតលាស់គភ៌ អេម៉ូក្លូប៊ីនមានមុខងារសំខាន់ពីរប្រភេទ៖ អំប្រ៊ីយ៉ុង Hb(2α, 2ε) ដែលត្រូវបានរកឃើញនៅក្នុងដំណាក់កាលដំបូងនៃការអភិវឌ្ឍន៍គភ៌ និង អេម៉ូក្លូប៊ីន F (ទារក)- (2α, 2γ) ដែលជំនួសអេម៉ូក្លូប៊ីនរបស់គភ៌ដំបូងក្នុងខែទីប្រាំមួយនៃការវិវឌ្ឍន៍នៃស្បូន ហើយបន្ទាប់ពីកំណើតត្រូវបានជំនួសដោយ Hb A ។
HB A គឺជាប្រូតេអ៊ីនដែលទាក់ទងទៅនឹង myoglobin (MB) ដែលមាននៅក្នុងកោសិកាឈាមក្រហមរបស់មនុស្សពេញវ័យ។ រចនាសម្ព័ន្ធនៃ protomers បុគ្គលរបស់វាគឺស្រដៀងទៅនឹង myoglobin ។ រចនាសម្ព័ន្ធបន្ទាប់បន្សំ និងទីបីនៃមេម៉ូក្លូប៊ីន និងមេម៉ូក្លូប៊ីនគឺស្រដៀងគ្នាខ្លាំងណាស់ ទោះបីជាការពិតដែលថានៅក្នុងរចនាសម្ព័ន្ធចម្បងនៃខ្សែសង្វាក់ polypeptide របស់ពួកគេមានតែសំណល់អាស៊ីតអាមីណូចំនួន 24 គឺដូចគ្នាបេះបិទ (រចនាសម្ព័ន្ធបន្ទាប់បន្សំនៃអេម៉ូក្លូប៊ីនប្រូតូមឺរ ដូចជា myoglobin មាន α-helices ចំនួនប្រាំបី។ កំណត់ដោយអក្សរឡាតាំងពី A ដល់ H ហើយរចនាសម្ព័ន្ធទីបីមានទម្រង់ជារាងពងក្រពើ) ។ ប៉ុន្តែមិនដូច myoglobin ទេ អេម៉ូក្លូប៊ីនមានរចនាសម្ព័ន្ធ oligomeric ដែលរួមមានខ្សែសង្វាក់ polypeptide បួនដែលតភ្ជាប់ដោយចំណងមិនមែនកូវ៉ាលេន (រូបភាព 1.22) ។
Hb protomer នីមួយៗត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងផ្នែកដែលមិនមែនជាប្រូតេអ៊ីន - heme និង protomers ជិតខាង។ ការតភ្ជាប់នៃផ្នែកប្រូតេអ៊ីននៃ Hb ជាមួយ heme គឺស្រដៀងគ្នាទៅនឹង myoglobin: នៅក្នុងមជ្ឈមណ្ឌលសកម្មនៃប្រូតេអ៊ីនផ្នែក hydrophobic នៃ heme ត្រូវបានហ៊ុំព័ទ្ធដោយរ៉ាឌីកាល់អាស៊ីតអាមីណូ hydrophobic លើកលែងតែ F 8 និង E 7 របស់គាត់ ដែលមានទីតាំងនៅសងខាងនៃយន្តហោះ heme និងដើរតួនាទីស្រដៀងគ្នាក្នុងដំណើរការនៃប្រូតេអ៊ីន និងការភ្ជាប់របស់វាជាមួយអុកស៊ីសែន (សូមមើលរចនាសម្ព័ន្ធនៃ myoglobin) ។
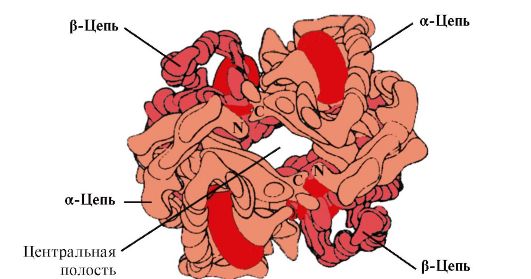 អង្ករ។ ១.២២. រចនាសម្ព័ន្ធ oligomeric នៃអេម៉ូក្លូប៊ីន
អង្ករ។ ១.២២. រចនាសម្ព័ន្ធ oligomeric នៃអេម៉ូក្លូប៊ីន
ក្រៅពីនេះ Gis E ៧អនុវត្តសំខាន់ តួនាទីបន្ថែមនៅក្នុងមុខងាររបស់ Nv ។ heme ឥតគិតថ្លៃមានទំនាក់ទំនងខ្ពស់ជាង 25,000 ដងសម្រាប់ CO ជាង O2 ។ CO ត្រូវបានបង្កើតឡើងក្នុងបរិមាណតិចតួចនៅក្នុងរាងកាយ ហើយដោយសារភាពស្និទ្ធស្នាលខ្ពស់របស់វាសម្រាប់ heme វាអាចរំខានដល់ការដឹកជញ្ជូន O 2 ដែលចាំបាច់សម្រាប់ជីវិតកោសិកា។ ទោះជាយ៉ាងណាក៏ដោយនៅក្នុងសមាសភាពនៃអេម៉ូក្លូប៊ីនភាពស្និទ្ធស្នាលរបស់ heme សម្រាប់កាបូនម៉ូណូអុកស៊ីតលើសពីភាពស្និទ្ធស្នាលសម្រាប់ O 2 ត្រឹមតែ 200 ដងប៉ុណ្ណោះដោយសារតែវត្តមានរបស់ E 7 របស់គាត់នៅក្នុងមជ្ឈមណ្ឌលសកម្ម។ អាស៊ីតអាមីណូដែលនៅសេសសល់នេះបង្កើតលក្ខខណ្ឌល្អប្រសើរបំផុតសម្រាប់ការផ្សារភ្ជាប់នៃ heme ទៅ O 2 និងធ្វើឱ្យអន្តរកម្មរបស់ heme ជាមួយ CO ចុះខ្សោយ។
5. មុខងារសំខាន់របស់ HB គឺការដឹកជញ្ជូន O2 ពីសួតទៅជាលិកា។មិនដូច monomeric myoglobin ដែលមានភាពស្និទ្ធស្នាលខ្ពស់សម្រាប់ O2 និងអនុវត្តមុខងារនៃការរក្សាទុកអុកស៊ីសែននៅក្នុងសាច់ដុំក្រហម រចនាសម្ព័ន្ធ oligomeric នៃអេម៉ូក្លូប៊ីនផ្តល់នូវ:
1) តិត្ថិភាពយ៉ាងឆាប់រហ័សនៃ HB ជាមួយនឹងអុកស៊ីសែននៅក្នុងសួត;
2) សមត្ថភាពរបស់ HB ក្នុងការបញ្ចេញអុកស៊ីសែននៅក្នុងជាលិកានៅសម្ពាធផ្នែកខ្ពស់នៃ O 2 (20-40 mm Hg);
3) លទ្ធភាពនៃការគ្រប់គ្រងភាពស្និទ្ធស្នាលនៃ Hb ទៅ O 2 ។
6. ការផ្លាស់ប្តូរសហករណ៍ក្នុងការអនុលោមតាមអេម៉ូក្លូប៊ីន protomers បង្កើនល្បឿននៃការភ្ជាប់ O 2 នៅក្នុងសួតនិងការបញ្ចេញរបស់វាចូលទៅក្នុងជាលិកា។ នៅក្នុងសួត សម្ពាធផ្នែកខ្ពស់នៃ O 2 ជំរុញការភ្ជាប់របស់វាទៅនឹង Hb នៅកន្លែងសកម្មនៃ protomers បួន (2α និង 2β) ។ មជ្ឈមណ្ឌលសកម្មនៃ protomer នីមួយៗ ដូចជានៅក្នុង myoglobin ស្ថិតនៅចន្លោះ α-helices ពីរ (F និង E) នៅក្នុងហោប៉ៅ hydrophobic ។ វាមានផ្នែកដែលមិនមែនជាប្រូតេអ៊ីន - heme ដែលភ្ជាប់ទៅនឹងផ្នែកប្រូតេអ៊ីនដោយអន្តរកម្ម hydrophobic ខ្សោយជាច្រើន និងចំណងដ៏រឹងមាំមួយរវាង Fe 2 + heme និង F 8 របស់គាត់ (សូមមើលរូប 1.21) ។
នៅក្នុង deoxyhemoglobin ដោយសារតែចំណងនេះជាមួយ F 8 របស់គាត់ អាតូម Fe 2 + លាតសន្ធឹងពីយន្តហោះ heme ឆ្ពោះទៅរក histidine ។ ការចង O 2 ទៅ Fe 2 + កើតឡើងនៅផ្នែកម្ខាងទៀតនៃ heme ក្នុងតំបន់ E 7 របស់គាត់ ដោយប្រើចំណងការសម្របសម្រួលដោយឥតគិតថ្លៃតែមួយ។ អ៊ី 7 របស់គាត់ផ្តល់នូវលក្ខខណ្ឌដ៏ល្អប្រសើរសម្រាប់ការភ្ជាប់ O 2 ទៅនឹងជាតិដែក។
ការបន្ថែម O 2 ទៅអាតូម Fe + 2 នៃ protomer មួយបណ្តាលឱ្យចលនារបស់វាចូលទៅក្នុងយន្តហោះ heme បន្ទាប់មកដោយសំណល់ histidine ដែលជាប់ទាក់ទងនឹងវា
 អង្ករ។ ១.២៣. ការផ្លាស់ប្តូរការអនុលោមតាមអេម៉ូក្លូប៊ីនប្រូតូមឺរនៅពេលផ្សំជាមួយ O 2
អង្ករ។ ១.២៣. ការផ្លាស់ប្តូរការអនុលោមតាមអេម៉ូក្លូប៊ីនប្រូតូមឺរនៅពេលផ្សំជាមួយ O 2
នេះនាំឱ្យមានការផ្លាស់ប្តូរនៅក្នុងការអនុលោមតាមខ្សែសង្វាក់ polypeptide ទាំងអស់ដោយសារតែ lability អនុលោមតាមរបស់ពួកគេ។ ការផ្លាស់ប្តូរការអនុលោមតាមខ្សែសង្វាក់ផ្សេងទៀតសម្របសម្រួលអន្តរកម្មរបស់ពួកគេជាមួយម៉ូលេគុល O 2 ជាបន្តបន្ទាប់។
ម៉ូលេគុល O 2 ទី 4 ភ្ជាប់ទៅនឹងអេម៉ូក្លូប៊ីនងាយស្រួលជាង 300 ដង (រូបភាព 1.24) ។
 អង្ករ។ ១.២៤. ការផ្លាស់ប្តូរសហករណ៍ក្នុងការអនុលោមតាមអេម៉ូក្លូប៊ីន protomers កំឡុងពេលអន្តរកម្មរបស់វាជាមួយ O2
អង្ករ។ ១.២៤. ការផ្លាស់ប្តូរសហករណ៍ក្នុងការអនុលោមតាមអេម៉ូក្លូប៊ីន protomers កំឡុងពេលអន្តរកម្មរបស់វាជាមួយ O2
នៅក្នុងជាលិកា ម៉ូលេគុល O 2 ជាបន្តបន្ទាប់នីមួយៗត្រូវបានបំបែកចេញបានយ៉ាងងាយជាងប្រភេទមុន ផងដែរ ដោយសារតែការផ្លាស់ប្តូរសហករណ៍ក្នុងការអនុលោមតាម protomers ។
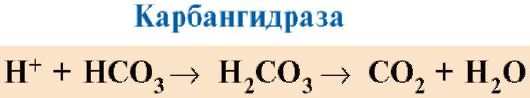
7. CO 2 និង H+ ដែលបង្កើតឡើងក្នុងអំឡុងពេល catabolism នៃសារធាតុសរីរាង្គ កាត់បន្ថយភាពស្និទ្ធស្នាលនៃអេម៉ូក្លូប៊ីនសម្រាប់ O 2 តាមសមាមាត្រទៅនឹងកំហាប់របស់វា។ ថាមពលដែលត្រូវការសម្រាប់ដំណើរការកោសិកាត្រូវបានផលិតជាចម្បងនៅក្នុង mitochondria កំឡុងពេលអុកស៊ីតកម្មនៃសារធាតុសរីរាង្គដោយប្រើ O 2 បញ្ជូនពីសួតដោយអេម៉ូក្លូប៊ីន។ ជាលទ្ធផលនៃការកត់សុីនៃសារធាតុសរីរាង្គផលិតផលចុងក្រោយនៃការរលួយរបស់ពួកគេត្រូវបានបង្កើតឡើង: CO 2 និង K 2 O ដែលជាបរិមាណសមាមាត្រទៅនឹងអាំងតង់ស៊ីតេនៃដំណើរការអុកស៊ីតកម្មដែលកំពុងដំណើរការ។
CO 2 សាយភាយចេញពីកោសិកាទៅក្នុងឈាម ហើយជ្រាបចូលទៅក្នុងកោសិកាឈាមក្រហម ដែលនៅក្រោមសកម្មភាពនៃអង់ស៊ីម carbanhydrase វាត្រូវបានបំប្លែងទៅជាអាស៊ីតកាបូន។ អាស៊ីតខ្សោយនេះបំបែកទៅជាប្រូតុង និងអ៊ីយ៉ុង bicarbonate ។
H+ មានសមត្ថភាពចូលរួមជាមួយរ៉ាឌីកាល់របស់ទ្រង់ 14 6 នៅក្នុងខ្សែសង្វាក់ α- និង β- នៃអេម៉ូក្លូប៊ីន ពោលគឺឧ។ នៅក្នុងតំបន់ដែលឆ្ងាយពី heme ។ ប្រូតុងនៃអេម៉ូក្លូប៊ីនកាត់បន្ថយភាពស្និទ្ធស្នាលរបស់វាចំពោះ O 2 ជំរុញការដក O 2 ចេញពី oxyHb ការបង្កើត deoxyHb និងបង្កើនការផ្គត់ផ្គង់អុកស៊ីសែនទៅជាលិកាក្នុងសមាមាត្រទៅនឹងចំនួនប្រូតុងដែលបានបង្កើតឡើង (រូបភាព 1.25) ។
ការកើនឡើងនៃបរិមាណអុកស៊ីសែនដែលបញ្ចេញដោយអាស្រ័យលើការកើនឡើងនៃកំហាប់ H+ នៅក្នុងកោសិកាឈាមក្រហមត្រូវបានគេហៅថាឥទ្ធិពល Bohr (ដាក់ឈ្មោះតាមសរីរវិទ្យាជនជាតិដាណឺម៉ាក Christian Bohr ដែលបានរកឃើញឥទ្ធិពលនេះជាលើកដំបូង) ។
នៅក្នុងសួត សម្ពាធផ្នែកខ្ពស់នៃអុកស៊ីសែនជំរុញការភ្ជាប់របស់វាទៅនឹង deoxyHb ដែលកាត់បន្ថយភាពស្និទ្ធស្នាលរបស់ប្រូតេអ៊ីនសម្រាប់ H + ។ ប្រូតុងដែលបញ្ចេញក្រោមសកម្មភាពនៃអាស៊ីតកាបូនិកមានប្រតិកម្មជាមួយនឹងប៊ីកាបូណាតដើម្បីបង្កើតជា CO 2 និង H 2 O
 អង្ករ។ ១.២៥. ការពឹងផ្អែកលើភាពស្និទ្ធស្នាលរបស់ Hb សម្រាប់ O 2 លើកំហាប់ CO 2 និងប្រូតុង (ឥទ្ធិពល Bohr):
អង្ករ។ ១.២៥. ការពឹងផ្អែកលើភាពស្និទ្ធស្នាលរបស់ Hb សម្រាប់ O 2 លើកំហាប់ CO 2 និងប្រូតុង (ឥទ្ធិពល Bohr):
ក- ឥទ្ធិពលនៃកំហាប់ CO 2 និង H + លើការបញ្ចេញ O 2 ពីស្មុគស្មាញជាមួយ HB (ឥទ្ធិពល Bohr); ខ- អុកស៊ីសែននៃ deoxyhemoglobin នៅក្នុងសួត ការបង្កើត និងការបញ្ចេញ CO 2 ។
លទ្ធផល CO 2 ចូលទៅក្នុងលំហ alveolar ហើយត្រូវបានដកចេញដោយខ្យល់ដែលដកដង្ហើមចេញ។ ដូច្នេះបរិមាណអុកស៊ីហ៊្សែនដែលបញ្ចេញដោយអេម៉ូក្លូប៊ីននៅក្នុងជាលិកាត្រូវបានគ្រប់គ្រងដោយផលិតផលនៃ catabolism នៃសារធាតុសរីរាង្គ៖ ការបំបែកសារធាតុកាន់តែខ្លាំងឧទាហរណ៍អំឡុងពេលហាត់ប្រាណ កំហាប់ CO 2 និង H + កាន់តែខ្ពស់ និងអុកស៊ីសែនកាន់តែច្រើន។ ជាលិកាទទួលបានជាលទ្ធផលនៃការថយចុះនៃទំនាក់ទំនង Hb សម្រាប់ O 2 ។
8. បទប្បញ្ញត្តិ Allosteric នៃភាពស្និទ្ធស្នាលនៃ Hb សម្រាប់ O2 ដោយ ligand - 2,3-bisphosphoglycerate ។នៅក្នុង erythrocytes, allosteric ligand នៃ hemoglobin, 2,3-bisphosphoglycerate (2,3-BPG) ត្រូវបានសំយោគពីផលិតផលនៃការកត់សុីគ្លុយកូស - 1,3-bisphosphoglycerate ។ នៅក្រោមលក្ខខណ្ឌធម្មតាកំហាប់នៃ 2,3-BPG គឺខ្ពស់ហើយអាចប្រៀបធៀបទៅនឹងកំហាប់ Hb ។ 2,3-BPG មានបន្ទុកអវិជ្ជមានខ្លាំងនៃ -5 ។
 Bisphosphoglycerate នៅក្នុងជាលិកា capillaries ភ្ជាប់ទៅនឹង deoxyhemoglobin បង្កើនការបញ្ចេញអុកស៊ីសែនទៅក្នុងជាលិកាកាត់បន្ថយភាពស្និទ្ធស្នាលនៃ Hb សម្រាប់ O 2 ។
Bisphosphoglycerate នៅក្នុងជាលិកា capillaries ភ្ជាប់ទៅនឹង deoxyhemoglobin បង្កើនការបញ្ចេញអុកស៊ីសែនទៅក្នុងជាលិកាកាត់បន្ថយភាពស្និទ្ធស្នាលនៃ Hb សម្រាប់ O 2 ។
នៅចំកណ្តាលនៃម៉ូលេគុល tetrameric hemoglobin គឺជាបែហោងធ្មែញ។ វាត្រូវបានបង្កើតឡើងដោយសំណល់អាស៊ីតអាមីណូនៃ protomers ទាំងបួន (សូមមើលរូប 1.22) ។ នៅក្នុងជាលិកា capillaries, protonation នៃ Hb (ឥទ្ធិពល Bohr) នាំឱ្យមានការដាច់នៃចំណងរវាងជាតិដែក heme និង O2 ។ នៅក្នុងម៉ូលេគុលមួយ។
deoxyhemoglobin បើប្រៀបធៀបទៅនឹង oxyhemoglobin ចំណងអ៊ីយ៉ុងបន្ថែមលេចឡើងដែលភ្ជាប់ protomers ដែលជាលទ្ធផលដែលវិមាត្រនៃបែហោងធ្មែញកណ្តាលកើនឡើងបើប្រៀបធៀបទៅនឹង oxyhemoglobin ។ បែហោងធ្មែញកណ្តាលគឺជាកន្លែងភ្ជាប់នៃ 2,3-BPG ទៅនឹងអេម៉ូក្លូប៊ីន។ ដោយសារតែភាពខុសគ្នានៃទំហំនៃបែហោងធ្មែញកណ្តាល 2,3-BPG អាចភ្ជាប់ទៅនឹង deoxyhemoglobin ប៉ុណ្ណោះ។
2,3-BPG ធ្វើអន្តរកម្មជាមួយអេម៉ូក្លូប៊ីននៅក្នុងកន្លែងដាច់ស្រយាលពីមជ្ឈមណ្ឌលសកម្មនៃប្រូតេអ៊ីន និងជាកម្មសិទ្ធិរបស់ allosteric(និយតកម្ម) ligands និងបែហោងកណ្តាលនៃ Hb គឺ មជ្ឈមណ្ឌល allosteric ។ 2,3-BPG មានបន្ទុកអវិជ្ជមានខ្លាំង និងធ្វើអន្តរកម្មជាមួយក្រុមដែលមានបន្ទុកវិជ្ជមានចំនួន 5 នៃក្រុម β-chains ពីរនៃ Hb: ក្រុម N-terminal α-amino group នៃ Val និង Lys 82 រ៉ាឌីកាល់ 143 របស់គាត់ (រូបភាព 1.26) ។
 អង្ករ។ ១.២៦. BPG នៅក្នុងបែហោងធ្មែញកណ្តាលនៃ deoxyhemoglobin
អង្ករ។ ១.២៦. BPG នៅក្នុងបែហោងធ្មែញកណ្តាលនៃ deoxyhemoglobin
BPG ភ្ជាប់ទៅនឹងក្រុមដែលមានការចោទប្រកាន់ជាវិជ្ជមានចំនួនបីនៅលើខ្សែβ-strand នីមួយៗ។
នៅក្នុងជាលិកា capillaries លទ្ធផល deoxyhemoglobin អន្តរកម្មជាមួយ 2,3-BPG និងចំណងអ៊ីយ៉ុងត្រូវបានបង្កើតឡើងរវាងរ៉ាឌីកាល់ដែលមានបន្ទុកវិជ្ជមាននៃខ្សែβ-chains និង ligand ចោទប្រកាន់អវិជ្ជមានដែលផ្លាស់ប្តូរការអនុលោមតាមប្រូតេអ៊ីននិងកាត់បន្ថយទំនាក់ទំនងនៃ Hb សម្រាប់ O2 ។ . ការថយចុះនៃភាពស្និទ្ធស្នាលរបស់ Hb សម្រាប់ O 2 ជំរុញឱ្យមានការបញ្ចេញ O 2 កាន់តែមានប្រសិទ្ធភាពទៅក្នុងជាលិកា។
នៅក្នុងសួត, នៅសម្ពាធផ្នែកខ្ពស់, អុកស៊ីសែនធ្វើអន្តរកម្មជាមួយ Hb, ចូលរួមជាមួយជាតិដែក heme; ក្នុងករណីនេះការអនុលោមតាមប្រូតេអ៊ីនផ្លាស់ប្តូរបែហោងធ្មែញកណ្តាលថយចុះហើយ 2,3-BPG ត្រូវបានផ្លាស់ទីលំនៅពីមជ្ឈមណ្ឌល allosteric ។
ដូច្នេះប្រូតេអ៊ីន oligomeric មានលក្ខណៈសម្បត្តិថ្មីបើប្រៀបធៀបទៅនឹងប្រូតេអ៊ីន monomeric ។ ការភ្ជាប់នៃ ligands នៅកន្លែង
គម្លាតឆ្ងាយពីគ្នាទៅវិញទៅមក (allosteric) អាចបណ្តាលឱ្យមានការផ្លាស់ប្តូរទម្រង់នៅក្នុងម៉ូលេគុលប្រូតេអ៊ីនទាំងមូល។ ដោយសារតែអន្តរកម្មជាមួយ ligands និយតកម្ម ការផ្លាស់ប្តូរនៅក្នុងការអនុលោមភាព និងការសម្របខ្លួននៃមុខងារនៃម៉ូលេគុលប្រូតេអ៊ីនទៅនឹងការផ្លាស់ប្តូរបរិស្ថានកើតឡើង។
ប្រធានបទ 1.5 ។ ការថែរក្សាការអនុលោមតាមធម្មជាតិនៃប្រូតេអ៊ីនក្រោមលក្ខខណ្ឌកោសិកា
នៅក្នុងកោសិកា កំឡុងពេលសំយោគខ្សែសង្វាក់ polypeptide ការដឹកជញ្ជូនរបស់ពួកគេតាមរយៈភ្នាសទៅផ្នែកដែលត្រូវគ្នានៃកោសិកា កំឡុងពេលដំណើរការបត់ (ការបង្កើតទម្រង់ដើម) និងកំឡុងពេលប្រមូលផ្តុំប្រូតេអ៊ីន oligomeric ក៏ដូចជាអំឡុងពេលដំណើរការរបស់វា កម្រិតមធ្យម។ , ងាយនឹងប្រមូលផ្តុំ, ការអនុលោមភាពមិនស្ថិតស្ថេរកើតឡើងនៅក្នុងរចនាសម្ព័ន្ធប្រូតេអ៊ីន។ រ៉ាឌីកាល់ Hydrophobic ដែលជាធម្មតាលាក់នៅខាងក្នុងម៉ូលេគុលប្រូតេអ៊ីននៅក្នុងការអនុលោមតាមប្រភពដើម លេចឡើងនៅលើផ្ទៃក្នុងទម្រង់មិនស្ថិតស្ថេរ ហើយមានទំនោរទៅបញ្ចូលគ្នាជាមួយក្រុមប្រូតេអ៊ីនផ្សេងទៀតដែលមិនសូវរលាយក្នុងទឹក។ នៅក្នុងកោសិកានៃសារពាង្គកាយដែលគេស្គាល់ទាំងអស់ ប្រូតេអ៊ីនពិសេសត្រូវបានគេរកឃើញដែលធានាបាននូវការផ្នត់ដ៏ល្អប្រសើរនៃប្រូតេអ៊ីនកោសិកា រក្សាលំនឹងការអនុលោមតាមដើមរបស់វាក្នុងអំឡុងពេលដំណើរការ ហើយសំខាន់បំផុតគឺរក្សារចនាសម្ព័ន្ធ និងមុខងារនៃប្រូតេអ៊ីនក្នុងកោសិកានៅពេលដែល homeostasis ត្រូវបានរំខាន។ ប្រូតេអ៊ីនទាំងនេះត្រូវបានគេហៅថា "អ្នកបួស"ដែលមានន័យថា "មេដោះ" ជាភាសាបារាំង។
1. មេអំបៅម៉ូលេគុល និងតួនាទីរបស់ពួកគេក្នុងការទប់ស្កាត់ការខូចទ្រង់ទ្រាយប្រូតេអ៊ីន។
Chaperones (CH) ត្រូវបានចាត់ថ្នាក់តាមម៉ាស់នៃអនុក្រុមរបស់វា។ ទម្ងន់ម៉ូលេគុលខ្ពស់មានម៉ាស់ពី 60 ទៅ 110 kDa ។ ក្នុងចំណោមពួកគេ ថ្នាក់បីត្រូវបានសិក្សាច្រើនបំផុតគឺ Sh-60, Sh-70 និង Sh-90 ។ ថ្នាក់នីមួយៗរួមមានក្រុមគ្រួសារនៃប្រូតេអ៊ីនដែលពាក់ព័ន្ធ។ ដូច្នេះ Sh-70 មានប្រូតេអ៊ីនដែលមានទម្ងន់ម៉ូលេគុលពី 66 ទៅ 78 kDa ។ ទម្ងន់ម៉ូលេគុលទាបមានទម្ងន់ម៉ូលេគុលពី 40 ទៅ 15 kDa ។
ក្នុងចំណោមចៅក្រមមាន ធម្មនុញ្ញប្រូតេអ៊ីន, ការសំយោគ basal ខ្ពស់ដែលមិនអាស្រ័យលើឥទ្ធិពលស្ត្រេសលើកោសិការបស់រាងកាយ, និង មិនអាចទទួលយកបាន,ការសំយោគដែលនៅក្រោមលក្ខខណ្ឌធម្មតាគឺខ្សោយ ប៉ុន្តែកើនឡើងយ៉ាងខ្លាំងនៅក្រោមភាពតានតឹង។ Chaperones inducible ត្រូវបានគេហៅថា "ប្រូតេអ៊ីនឆក់កំដៅ" ផងដែរព្រោះវាត្រូវបានរកឃើញដំបូងនៅក្នុងកោសិកាដែលប៉ះពាល់នឹងសីតុណ្ហភាពខ្ពស់។ នៅក្នុងកោសិកា ដោយសារកំហាប់ខ្ពស់នៃប្រូតេអ៊ីន ការធ្វើឱ្យសកម្មឡើងវិញដោយឯកឯងនៃប្រូតេអ៊ីនដែលខូចដោយផ្នែកគឺពិបាក។ Sh-70 អាចការពារការចាប់ផ្តើមនៃ denaturation និងជួយស្ដារការអនុលោមតាមប្រភពដើមនៃប្រូតេអ៊ីន។ ម៉ូលេគុលអ្នកបួស-៧០- ថ្នាក់អភិរក្សខ្ពស់នៃប្រូតេអ៊ីនដែលរកឃើញនៅគ្រប់ផ្នែកទាំងអស់នៃកោសិកា៖ cytoplasm, nucleus, endoplasmic reticulum, mitochondria ។ នៅចុង carboxyl នៃខ្សែសង្វាក់ polypeptide តែមួយШ-70 មានតំបន់មួយដែលជាចង្អូរដែលមានសមត្ថភាពធ្វើអន្តរកម្មជាមួយ peptides នៃប្រវែង
ពី 7 ទៅ 9 សំណល់អាស៊ីតអាមីណូដែលសំបូរទៅដោយរ៉ាឌីកាល់ hydrophobic ។ តំបន់បែបនេះនៅក្នុងប្រូតេអ៊ីន globular កើតឡើងប្រហែលរៀងរាល់ 16 អាមីណូអាស៊ីត។ Sh-70 មានសមត្ថភាពការពារប្រូតេអ៊ីនពីភាពអសកម្មនៃសីតុណ្ហភាព និងស្ដារឡើងវិញនូវការអនុលោមភាព និងសកម្មភាពនៃប្រូតេអ៊ីនដែលមិនមានលក្ខណៈធម្មជាតិ។
2. តួនាទីរបស់ chaperones ក្នុងការបត់ប្រូតេអ៊ីន។កំឡុងពេលសំយោគប្រូតេអ៊ីននៅលើ ribosome តំបន់ N-terminal នៃ polypeptide ត្រូវបានសំយោគមុនពេល C-terminal មួយ។ ដើម្បីបង្កើតការអនុលោមតាមប្រភពដើម លំដាប់អាស៊ីតអាមីណូពេញលេញនៃប្រូតេអ៊ីនត្រូវបានទាមទារ។ នៅក្នុងដំណើរការនៃការសំយោគប្រូតេអ៊ីន Chaperones-70 ដោយសារតែរចនាសម្ព័ន្ធនៃមជ្ឈមណ្ឌលសកម្មរបស់ពួកគេអាចបិទតំបន់នៃ polypeptide ដែលងាយនឹងប្រមូលផ្តុំដែលសំបូរទៅដោយរ៉ាឌីកាល់អាស៊ីតអាមីណូ hydrophobic រហូតដល់ការសំយោគត្រូវបានបញ្ចប់ (រូបភាព 1.27, A ។ )
 អង្ករ។ ១.២៧. ការចូលរួមរបស់ chaperones ក្នុងការបត់ប្រូតេអ៊ីន
អង្ករ។ ១.២៧. ការចូលរួមរបស់ chaperones ក្នុងការបត់ប្រូតេអ៊ីន
ក - ការចូលរួមរបស់ chaperones-70 ក្នុងការទប់ស្កាត់អន្តរកម្ម hydrophobic រវាងផ្នែកនៃ polypeptide សំយោគ; ខ - ការបង្កើតការអនុលោមតាមប្រភពដើមនៃប្រូតេអ៊ីននៅក្នុងស្មុគស្មាញ chaperone
ប្រូតេអ៊ីនម៉ូលេគុលខ្ពស់ជាច្រើនដែលមានទម្រង់ស្មុគស្មាញ ដូចជារចនាសម្ព័ន្ធដែនបត់ក្នុងចន្លោះពិសេសដែលបង្កើតឡើងដោយ Sh-60។ Ш-60ដំណើរការជាស្មុគស្មាញ oligomeric ដែលមាន 14 អនុរង។ ពួកវាបង្កើតជារង្វង់ប្រហោងចំនួនពីរ ដែលនីមួយៗមានផ្នែករងចំនួនប្រាំពីរ ចិញ្ចៀនទាំងនេះត្រូវបានតភ្ជាប់ទៅគ្នាទៅវិញទៅមក។ អនុឯកតា Sh-60 នីមួយៗមានដែនចំនួនបី៖ apical (apical) សំបូរទៅដោយរ៉ាឌីកាល់ hydrophobic ប្រឈមមុខនឹងបែហោងធ្មែញនៃចិញ្ចៀន កម្រិតមធ្យម និងអេក្វាទ័រ (រូបភាព 1.28) ។
 អង្ករ។ ១.២៨. រចនាសម្ព័ន្ធនៃស្មុគស្មាញ chaperonin មាន 14 Ш-60
អង្ករ។ ១.២៨. រចនាសម្ព័ន្ធនៃស្មុគស្មាញ chaperonin មាន 14 Ш-60
ក - ទិដ្ឋភាពចំហៀង; ខ - ទិដ្ឋភាពកំពូល
ប្រូតេអ៊ីនសំយោគដែលមានធាតុនៅលើផ្ទៃលក្ខណៈនៃម៉ូលេគុលដែលលាតត្រដាង ជាពិសេសរ៉ាឌីកាល់ hydrophobic ចូលទៅក្នុងបែហោងធ្មែញនៃចិញ្ចៀនរបស់ chaperone ។ នៅក្នុងបរិយាកាសជាក់លាក់នៃបែហោងធ្មែញទាំងនេះ ការអនុលោមតាមដែលអាចកើតមានត្រូវបានរាប់បញ្ចូលរហូតទាល់តែរកឃើញតែមួយគត់ដែលមានថាមពលអំណោយផលបំផុត (រូបភាព 1.27, ខ) ។ ការបង្កើតនៃការអនុលោមតាមនិងការបញ្ចេញប្រូតេអ៊ីនត្រូវបានអមដោយ ATP hydrolysis នៅក្នុងតំបន់អេក្វាទ័រ។ ជាធម្មតា ផ្នត់ដែលពឹងផ្អែកលើ chaperone បែបនេះត្រូវការថាមពលយ៉ាងច្រើន។
បន្ថែមពីលើការចូលរួមក្នុងការបង្កើតរចនាសម្ព័ន្ធបីវិមាត្រនៃប្រូតេអ៊ីន និងការបង្កើតឡើងវិញនៃប្រូតេអ៊ីនដែលខូចដោយផ្នែកនោះ អ្នកដឹកនាំក៏ចាំបាច់សម្រាប់ការកើតឡើងនៃដំណើរការជាមូលដ្ឋានដូចជាការប្រមូលផ្តុំប្រូតេអ៊ីន oligomeric ការទទួលស្គាល់ និងការដឹកជញ្ជូនប្រូតេអ៊ីន denatured ទៅជា lysosomes ។ ការដឹកជញ្ជូនប្រូតេអ៊ីនឆ្លងកាត់ភ្នាស និងការចូលរួមក្នុងបទប្បញ្ញត្តិនៃសកម្មភាពនៃស្មុគស្មាញប្រូតេអ៊ីន។
ប្រធានបទ 1.6 ។ ភាពចម្រុះនៃប្រូតេអ៊ីន។ គ្រួសារប្រូតេអ៊ីន៖ ឧទាហរណ៍នៃ Immunoglobulins
1. ប្រូតេអ៊ីនដើរតួនាទីយ៉ាងសំខាន់ក្នុងជីវិតរបស់កោសិកានីមួយៗ និងសារពាង្គកាយពហុកោសិកាទាំងមូល ហើយមុខងាររបស់វាមានភាពចម្រុះគួរឱ្យភ្ញាក់ផ្អើល។ នេះត្រូវបានកំណត់ដោយលក្ខណៈនៃរចនាសម្ព័ន្ធបឋមនិងការអនុលោមតាមប្រូតេអ៊ីនរចនាសម្ព័ន្ធតែមួយគត់នៃមជ្ឈមណ្ឌលសកម្មនិងសមត្ថភាពក្នុងការចងលីហ្គែនជាក់លាក់។
មានតែប្រភាគតូចបំផុតនៃបំរែបំរួលដែលអាចធ្វើបានទាំងអស់នៃខ្សែសង្វាក់ peptide អាចទទួលយករចនាសម្ព័ន្ធលំហដែលមានស្ថេរភាព។ ភាគច្រើន
ពួកគេអាចទទួលយកការអនុលោមភាពជាច្រើនជាមួយនឹងថាមពល Gibbs ប្រហែលដូចគ្នា ប៉ុន្តែមានលក្ខណៈសម្បត្តិខុសៗគ្នា។ រចនាសម្ព័នចម្បងនៃប្រូតេអ៊ីនដែលគេស្គាល់ភាគច្រើនដែលត្រូវបានជ្រើសរើសដោយការវិវត្តន៍ជីវសាស្រ្តធានានូវស្ថេរភាពពិសេសនៃការអនុលោមតាមលក្ខណៈណាមួយដែលកំណត់លក្ខណៈនៃដំណើរការនៃប្រូតេអ៊ីននេះ។
2. គ្រួសារប្រូតេអ៊ីន។នៅក្នុងប្រភេទជីវសាស្រ្តដូចគ្នា ការជំនួសសំណល់អាស៊ីតអាមីណូអាចនាំទៅដល់ការលេចចេញនូវប្រូតេអ៊ីនផ្សេងៗគ្នា ដែលបំពេញមុខងារដែលពាក់ព័ន្ធ និងមានលំដាប់អាស៊ីតអាមីណូដូចគ្នា។ ប្រូតេអ៊ីនដែលពាក់ព័ន្ធបែបនេះមានការអនុលោមភាពស្រដៀងគ្នាខ្លាំង៖ ចំនួន និងទីតាំងទាក់ទងនៃ α-helices និង/ឬ β-structures ភាគច្រើននៃវេន និងពត់នៃខ្សែសង្វាក់ polypeptide គឺស្រដៀងគ្នា ឬដូចគ្នាបេះបិទ។ ប្រូតេអ៊ីនដែលមានតំបន់ដូចគ្នានៃខ្សែសង្វាក់ polypeptide ការអនុលោមភាពស្រដៀងគ្នា និងមុខងារពាក់ព័ន្ធត្រូវបានចាត់ថ្នាក់ទៅជាក្រុមប្រូតេអ៊ីន។ ឧទាហរណ៍នៃគ្រួសារប្រូតេអ៊ីន៖ ប្រូតេអ៊ីន serine, គ្រួសារ immunoglobulin, គ្រួសារ myoglobin ។
ប្រូតេអ៊ីន Serine- ក្រុមគ្រួសារនៃប្រូតេអ៊ីនដែលអនុវត្តមុខងារនៃអង់ស៊ីម proteolytic ។ ទាំងនេះរួមមានអង់ស៊ីមរំលាយអាហារ - chymotrypsin, trypsin, elastase និងកត្តាកំណកឈាមជាច្រើន។ ប្រូតេអ៊ីនទាំងនេះមានអាស៊ីដអាមីណូដូចគ្នាបេះបិទក្នុង 40% នៃមុខតំណែងរបស់វា និងមានលក្ខណៈស្រដៀងគ្នាខ្លាំង (រូបភាព 1.29)។

អង្ករ។ ១.២៩. រចនាសម្ព័ន្ធលំហនៃ elastase (A) និង chymotrypsin (B)
ការជំនួសអាស៊ីតអាមីណូមួយចំនួនបាននាំឱ្យមានការផ្លាស់ប្តូរនៅក្នុងភាពជាក់លាក់នៃស្រទាប់ខាងក្រោមនៃប្រូតេអ៊ីនទាំងនេះ និងការលេចឡើងនៃភាពចម្រុះមុខងារនៅក្នុងគ្រួសារ។
3. គ្រួសារ Immunoglobulin ។នៅក្នុងដំណើរការនៃប្រព័ន្ធភាពស៊ាំ ប្រូតេអ៊ីននៃក្រុម immunoglobulin superfamily ដើរតួនាទីយ៉ាងធំ ដែលរួមមានប្រូតេអ៊ីនបីគ្រួសារ៖
អង្គបដិប្រាណ (immunoglobulins);
អ្នកទទួល T-lymphocyte;
ប្រូតេអ៊ីននៃស្មុគ្រស្មាញនៃភាពឆបគ្នាប្រវតិ្តសាស្រ្តដ៏សំខាន់ - ថ្នាក់ MHC ថ្នាក់ 1 និង 2 (ស្មុគស្មាញ Histocompatibility ដ៏សំខាន់) ។
ប្រូតេអ៊ីនទាំងអស់នេះមានរចនាសម្ព័ន្ធដែន មានដែនស្រដៀងគ្នានៃប្រព័ន្ធភាពស៊ាំ និងអនុវត្តមុខងារស្រដៀងគ្នា៖ ពួកវាមានអន្តរកម្មជាមួយរចនាសម្ព័ន្ធបរទេស រំលាយនៅក្នុងឈាម ទឹករងៃ ឬសារធាតុរាវអន្តរកោសិកា (អង្គបដិប្រាណ) ឬមានទីតាំងនៅលើផ្ទៃនៃកោសិកា (ផ្ទាល់ខ្លួន ឬ បរទេស)។
4. អង្គបដិប្រាណ- ប្រូតេអ៊ីនជាក់លាក់ដែលផលិតដោយ B lymphocytes ក្នុងការឆ្លើយតបទៅនឹងការបញ្ចូលរចនាសម្ព័ន្ធបរទេសចូលទៅក្នុងខ្លួនគេហៅថា អង់ទីហ្សែន។
លក្ខណៈពិសេសនៃរចនាសម្ព័ន្ធនៃអង្គបដិប្រាណ
ម៉ូលេគុលអង្គបដិប្រាណសាមញ្ញបំផុតមានខ្សែសង្វាក់ polypeptide ចំនួនបួន៖ ពន្លឺដូចគ្នាបេះបិទចំនួនពីរ - L ដែលមានអាស៊ីតអាមីណូប្រហែល 220 និងប្រភេទធ្ងន់ដូចគ្នា - H ដែលមានអាស៊ីតអាមីណូ 440-700 ។ ខ្សែសង្វាក់ទាំងបួននៅក្នុងម៉ូលេគុលអង្គបដិបក្ខត្រូវបានតភ្ជាប់ដោយចំណងដែលមិនមែនជាកូវ៉ាឡង់ជាច្រើន និងចំណង disulfide បួន (រូបភាព 1.30) ។
ខ្សែសង្វាក់ពន្លឺអង្គបដិប្រាណមានដែនពីរ៖ ដែនអថេរ (VL) ដែលមានទីតាំងនៅតំបន់ N នៃខ្សែសង្វាក់ polypeptide និងដែនថេរ (CL) ដែលមានទីតាំងនៅ C-terminus ។ ខ្សែសង្វាក់ធ្ងន់ជាធម្មតាមានដែនចំនួនបួន៖ អថេរមួយ (VH) ដែលមានទីតាំងនៅ N-terminus និងដែនថេរចំនួនបី (CH1, CH2, CH3) (សូមមើលរូប 1.30)។ ដែន immunoglobulin នីមួយៗមាន β-sheet superstructure ដែលក្នុងនោះសំណល់ cysteine ពីរត្រូវបានភ្ជាប់ដោយចំណង disulfide ។
រវាងដែនថេរទាំងពីរ CH1 និង CH2 មានតំបន់ដែលមានសំណល់ប្រូលីនមួយចំនួនធំ ដែលការពារការបង្កើតរចនាសម្ព័ន្ធបន្ទាប់បន្សំ និងអន្តរកម្មនៃខ្សែសង្វាក់ H ជិតខាងនៅក្នុងផ្នែកនេះ។ តំបន់ hinge នេះផ្តល់នូវភាពបត់បែននៃម៉ូលេគុលអង្គបដិប្រាណ។ រវាងដែនអថេរនៃខ្សែសង្វាក់ធ្ងន់ និងស្រាល មានកន្លែងភ្ជាប់អង់ទីហ្សែនដូចគ្នាបេះបិទ (កន្លែងសកម្មសម្រាប់ភ្ជាប់អង់ទីហ្សែន) ដូច្នេះអង្គបដិប្រាណបែបនេះច្រើនតែត្រូវបានគេហៅថា bivalents ។មិនមែនលំដាប់អាស៊ីតអាមីណូទាំងមូលនៃតំបន់អថេរនៃសង្វាក់ទាំងពីរពាក់ព័ន្ធនឹងការចងអង់ទីហ្សែនទៅនឹងអង្គបដិបក្ខនោះទេ ប៉ុន្តែមានតែអាស៊ីដអាមីណូ 20-30 ដែលស្ថិតនៅក្នុងតំបន់អថេរខ្ពស់នៃសង្វាក់នីមួយៗ។ វាគឺជាតំបន់ទាំងនេះដែលកំណត់សមត្ថភាពពិសេសនៃប្រភេទអង់ទីករនីមួយៗដើម្បីធ្វើអន្តរកម្មជាមួយអង់ទីហ្សែនបំពេញបន្ថែមដែលត្រូវគ្នា។
អង្គបដិប្រាណគឺជាខ្សែការពារមួយរបស់រាងកាយប្រឆាំងនឹងការឈ្លានពានរបស់សារពាង្គកាយបរទេស។ មុខងាររបស់ពួកវាអាចបែងចែកជាពីរដំណាក់កាល៖ ដំណាក់កាលដំបូងគឺការទទួលស្គាល់ និងការចងអង់ទីហ្សែននៅលើផ្ទៃនៃសារពាង្គកាយបរទេស ដែលអាចធ្វើទៅបានដោយសារតែវត្តមាននៃកន្លែងភ្ជាប់អង់ទីហ្សែននៅក្នុងរចនាសម្ព័ន្ធអង្គបដិប្រាណ។ ដំណាក់កាលទីពីរគឺការចាប់ផ្តើមនៃដំណើរការអសកម្ម និងការបំផ្លាញអង់ទីហ្សែន។ ភាពជាក់លាក់នៃដំណាក់កាលទីពីរគឺអាស្រ័យលើថ្នាក់នៃអង្គបដិប្រាណ។ មានខ្សែសង្វាក់ធ្ងន់ចំនួនប្រាំដែលខុសគ្នាពីគ្នាទៅវិញទៅមកនៅក្នុងរចនាសម្ព័ន្ធនៃដែនថេរ: α, δ, ε, γ និង μ យោងទៅតាមថ្នាក់នៃ immunoglobulins ប្រាំប្រភេទត្រូវបានសម្គាល់: A, D, E, G និង M ។
លក្ខណៈពិសេសនៃរចនាសម្ព័ន្ធនៃខ្សែសង្វាក់ធ្ងន់ផ្តល់ឱ្យតំបន់ hinge និងតំបន់ C-terminal នៃខ្សែសង្វាក់ធ្ងន់នូវលក្ខណៈនៃការអនុលោមតាមថ្នាក់នីមួយៗ។ បន្ទាប់ពីអង់ទីហ្សែនភ្ជាប់ទៅនឹងអង្គបដិប្រាណ ការផ្លាស់ប្តូរទម្រង់នៅក្នុងដែនថេរកំណត់ផ្លូវសម្រាប់ការដកអង់ទីហ្សែនចេញ។
 អង្ករ។ 1. 30. រចនាសម្ព័ន្ធដែននៃ IgG
អង្ករ។ 1. 30. រចនាសម្ព័ន្ធដែននៃ IgG
Immunoglobulins M
Immunoglobulins M មានពីរទម្រង់។
ទម្រង់ Monomeric- អង្គបដិប្រាណថ្នាក់ទី 1 ផលិតដោយការបង្កើត B lymphocytes ។ ក្រោយមក កោសិកា B ជាច្រើនប្តូរទៅផលិតអង្គបដិប្រាណផ្សេងទៀត ប៉ុន្តែមានកន្លែងភ្ជាប់អង់ទីហ្សែនដូចគ្នា។ IgM ត្រូវបានបង្កប់នៅក្នុងភ្នាស និងដើរតួជាអ្នកទទួលការទទួលស្គាល់អង់ទីហ្សែន។ ការរួមបញ្ចូល IgM ទៅក្នុងភ្នាសកោសិកាគឺអាចធ្វើទៅបានដោយសារតែវត្តមាននៃសំណល់អាស៊ីតអាមីណូ hydrophobic 25 នៅក្នុងផ្នែកកន្ទុយនៃតំបន់។
ទម្រង់សម្ងាត់នៃ IgMមានអនុផ្នែក monomeric ចំនួនប្រាំដែលភ្ជាប់គ្នាទៅវិញទៅមកដោយចំណង disulfide និងខ្សែសង្វាក់ polypeptide J បន្ថែម (រូបភាព 1.31) ។ ខ្សែសង្វាក់ធ្ងន់នៃ monomers នៃទម្រង់នេះមិនមានកន្ទុយ hydrophobic ទេ។ Pentamer មានកន្លែងភ្ជាប់អង់ទីហ្សែនចំនួន 10 ហើយដូច្នេះមានប្រសិទ្ធភាពក្នុងការទទួលស្គាល់ និងដកអង់ទីហ្សែនដែលចូលទៅក្នុងខ្លួនដំបូង។ ទម្រង់សំងាត់នៃ IgM គឺជាថ្នាក់ចម្បងនៃអង្គបដិបក្ខដែលសម្ងាត់ចូលទៅក្នុងឈាមកំឡុងពេលការឆ្លើយតបនៃប្រព័ន្ធភាពស៊ាំបឋម។ ការចង IgM ទៅនឹងអង់ទីហ្សែនផ្លាស់ប្តូរការអនុលោមតាម IgM និងជំរុញការភ្ជាប់របស់វាទៅនឹងសមាសធាតុប្រូតេអ៊ីនដំបូងនៃប្រព័ន្ធបំពេញ (ប្រព័ន្ធបំពេញបន្ថែមគឺជាសំណុំនៃប្រូតេអ៊ីនដែលពាក់ព័ន្ធនឹងការបំផ្លាញអង់ទីហ្សែន) និងការធ្វើឱ្យប្រព័ន្ធនេះសកម្ម។ ប្រសិនបើអង់ទីហ្សែនមានទីតាំងនៅលើផ្ទៃនៃអតិសុខុមប្រាណ ប្រព័ន្ធបំពេញបន្ថែមបណ្តាលឱ្យមានការរំខានដល់ភាពសុចរិតនៃភ្នាសកោសិកា និងការស្លាប់នៃកោសិកាបាក់តេរី។
Immunoglobulins G
តាមបរិមាណ ថ្នាក់នៃសារធាតុ immunoglobulins នេះគ្របដណ្តប់លើឈាម (75% នៃ Igs ទាំងអស់) ។ IgG - monomers ដែលជាថ្នាក់សំខាន់នៃអង្គបដិបក្ខដែលសម្ងាត់ចូលទៅក្នុងឈាមក្នុងអំឡុងពេលការឆ្លើយតបនៃប្រព័ន្ធភាពស៊ាំបន្ទាប់បន្សំ។ បន្ទាប់ពីអន្តរកម្មនៃ IgG ជាមួយអង់ទីហ្សែនផ្ទៃនៃអតិសុខុមប្រាណ ស្មុគស្មាញអង់ទីហ្សែន-អង្គបដិប្រាណអាចចង និងធ្វើឱ្យប្រូតេអ៊ីននៃប្រព័ន្ធបំពេញបន្ថែម ឬអាចធ្វើអន្តរកម្មជាមួយអ្នកទទួលជាក់លាក់នៃ macrophages និងនឺត្រូហ្វីល។ អន្តរកម្មជាមួយ phagocytes នាំឱ្យមាន
 អង្ករ។ ១.៣១. រចនាសម្ព័ន្ធនៃទម្រង់សម្ងាត់នៃ IgM
អង្ករ។ ១.៣១. រចនាសម្ព័ន្ធនៃទម្រង់សម្ងាត់នៃ IgM
ការស្រូបយកអង់ទីហ្សែន - អង់ទីករស្មុគស្មាញ និងការបំផ្លាញរបស់វានៅក្នុង phagosomes កោសិកា។ IgG គឺជាប្រភេទអង្គបដិប្រាណតែមួយគត់ដែលអាចជ្រាបចូលទៅក្នុងរបាំងសុក និងផ្តល់ការការពារក្នុងស្បូនរបស់ទារកពីការឆ្លង។
Immunoglobulins A
ថ្នាក់សំខាន់នៃអង្គបដិប្រាណដែលមានវត្តមាននៅក្នុងសំងាត់ (ទឹកដោះគោ ទឹកមាត់ អាថ៌កំបាំងនៃផ្លូវដង្ហើម និងពោះវៀន)។ IgA ត្រូវបានលាក់កំបាំងជាចម្បងនៅក្នុងទម្រង់ឌីមឺរិច ដែលម៉ូណូម័រត្រូវបានភ្ជាប់ទៅគ្នាទៅវិញទៅមកតាមរយៈខ្សែសង្វាក់ J បន្ថែម (រូបភាព 1.32) ។
IgA មិនមានអន្តរកម្មជាមួយប្រព័ន្ធបំពេញបន្ថែម និងកោសិកា phagocytic ទេ ប៉ុន្តែដោយការផ្សារភ្ជាប់ទៅនឹងអតិសុខុមប្រាណ អង្គបដិបក្ខការពារការភ្ជាប់របស់វាទៅនឹងកោសិកា epithelial និងការជ្រៀតចូលទៅក្នុងខ្លួន។
Immunoglobulins E
Immunoglobulins E ត្រូវបានតំណាងដោយ monomers ដែលក្នុងនោះ ε-chains ធ្ងន់មានដូចជា μ-chains នៃ immunoglobulins M ដែលជាអថេរមួយ និងដែនថេរចំនួនបួន។ បន្ទាប់ពីការសំងាត់ IgE ភ្ជាប់ជាមួយរបស់វា។
 អង្ករ។ ១.៣២. រចនាសម្ព័ន្ធ IgA
អង្ករ។ ១.៣២. រចនាសម្ព័ន្ធ IgA
តំបន់ C-terminal ជាមួយនឹងការទទួលដែលត្រូវគ្នានៅលើផ្ទៃនៃកោសិកា mast និង basophils ។ ជាលទ្ធផល ពួកវាក្លាយទៅជាអ្នកទទួលសម្រាប់អង់ទីហ្សែននៅលើផ្ទៃនៃកោសិកាទាំងនេះ (រូបភាព 1.33)។
 អង្ករ។ ១.៣៣. អន្តរកម្មនៃ IgE ជាមួយអង់ទីហ្សែននៅលើផ្ទៃនៃកោសិកាមេ
អង្ករ។ ១.៣៣. អន្តរកម្មនៃ IgE ជាមួយអង់ទីហ្សែននៅលើផ្ទៃនៃកោសិកាមេ
បន្ទាប់ពីអង់ទីហ្សែនភ្ជាប់ទៅនឹងកន្លែងភ្ជាប់អង់ទីហ្សែនដែលត្រូវគ្នានៃ IgE កោសិកាទទួលបានសញ្ញាមួយដើម្បីបញ្ចេញសារធាតុសកម្មជីវសាស្រ្ត (អ៊ីស្តាមីន សេរ៉ូតូនីន) ដែលទទួលខុសត្រូវយ៉ាងធំធេងចំពោះការវិវត្តនៃប្រតិកម្មរលាក និងការបង្ហាញប្រតិកម្មអាលែហ្សីដូចជា ជំងឺហឺត urticaria គ្រុនក្តៅហៃ។
Immunoglobulins D
Immunoglobulins D ត្រូវបានរកឃើញក្នុងបរិមាណតិចតួចនៅក្នុងសេរ៉ូម។ ខ្សែសង្វាក់ δ ធ្ងន់មានអថេរមួយ និងដែនថេរចំនួនបី។ IgDs ដើរតួជាអ្នកទទួលសម្រាប់ B lymphocytes មុខងារផ្សេងទៀតនៅតែមិនស្គាល់។ អន្តរកម្មនៃអង់ទីហ្សែនជាក់លាក់ជាមួយអ្នកទទួលនៅលើផ្ទៃនៃ B-lymphocytes (IgD) នាំទៅដល់ការបញ្ជូនសញ្ញាទាំងនេះទៅក្នុងកោសិកា និងការធ្វើឱ្យសកម្មនៃយន្តការដែលធានាដល់ការរីកសាយនៃក្លូន lymphocyte ដែលបានផ្តល់ឱ្យ។
ប្រធានបទ 1.7 ។ លក្ខណៈសម្បត្តិរូបវិទ្យា និងគីមីនៃប្រូតេអ៊ីន និងវិធីសាស្រ្តសម្រាប់ការបំបែករបស់ពួកគេ
1. ប្រូតេអ៊ីនបុគ្គលមានភាពខុសប្លែកគ្នានៅក្នុងលក្ខណៈរូបវន្ត និងគីមី៖
រូបរាងនៃម៉ូលេគុល;
ទម្ងន់ម៉ូលេគុល;
បន្ទុកសរុប, ទំហំដែលអាស្រ័យលើសមាមាត្រនៃក្រុម anionic និង cationic នៃអាស៊ីតអាមីណូ;
សមាមាត្រនៃរ៉ាឌីកាល់អាស៊ីតអាមីណូប៉ូលនិងមិនប៉ូលនៅលើផ្ទៃនៃម៉ូលេគុល;
កម្រិតនៃភាពធន់ទ្រាំទៅនឹងភ្នាក់ងារ denaturing ផ្សេងៗ។
2. ភាពរលាយនៃប្រូតេអ៊ីនអាស្រ័យនៅលើលក្ខណៈសម្បត្តិនៃប្រូតេអ៊ីនដែលបានរាយខាងលើ ក៏ដូចជាលើសមាសភាពនៃឧបករណ៍ផ្ទុកដែលប្រូតេអ៊ីនត្រូវបានរំលាយ (តម្លៃ pH សមាសភាពអំបិល សីតុណ្ហភាព វត្តមានសារធាតុសរីរាង្គផ្សេងទៀតដែលអាចមានអន្តរកម្មជាមួយប្រូតេអ៊ីន)។ បរិមាណនៃការចោទប្រកាន់នៃម៉ូលេគុលប្រូតេអ៊ីនគឺជាកត្តាមួយដែលប៉ះពាល់ដល់ការរលាយរបស់វា។ នៅពេលដែលការចោទប្រកាន់នៅចំណុច isoelectric ត្រូវបានបាត់បង់ ប្រូតេអ៊ីនកាន់តែងាយស្រួលប្រមូលផ្តុំ និង precipitate ។ នេះជាតួយ៉ាងពិសេសសម្រាប់ប្រូតេអ៊ីនមិនប្រក្រតី ដែលរ៉ាឌីកាល់អាស៊ីតអាមីណូ hydrophobic លេចឡើងលើផ្ទៃ។
នៅលើផ្ទៃនៃម៉ូលេគុលប្រូតេអ៊ីន មានទាំងរ៉ាឌីកាល់អាស៊ីតអាមីណូ វិជ្ជមាន និងអវិជ្ជមាន។ ចំនួននៃក្រុមទាំងនេះ ហើយដូច្នេះការចោទប្រកាន់សរុបនៃប្រូតេអ៊ីនគឺអាស្រ័យលើ pH នៃមធ្យម ពោលគឺឧ។ សមាមាត្រនៃការប្រមូលផ្តុំនៃក្រុម H + - និង OH - ក្រុម។ នៅក្នុងបរិយាកាសអាសុីតការកើនឡើងនៃកំហាប់ H+ នាំឱ្យមានការបង្ក្រាបការបំបែកក្រុម carboxyl -COO - + H+ > - COOH និងការថយចុះនៃបន្ទុកអវិជ្ជមាននៃប្រូតេអ៊ីន។ នៅក្នុងបរិយាកាសអាល់កាឡាំងការចងនៃ OH លើស - ដោយប្រូតុងដែលបានបង្កើតឡើងក្នុងអំឡុងពេលនៃការបំបែកក្រុមអាមីណូ -NH 3 + + OH - - NH 2 + H 2 O ជាមួយនឹងការបង្កើតទឹកនាំឱ្យមានការថយចុះនៃបន្ទុកវិជ្ជមាននៃប្រូតេអ៊ីន។ . តម្លៃ pH ដែលប្រូតេអ៊ីនមានបន្ទុកសូន្យសុទ្ធត្រូវបានគេហៅថា ចំណុច isoelectric (IEP) ។នៅក្នុង IET ចំនួនក្រុមគិតវិជ្ជមាន និងអវិជ្ជមានគឺដូចគ្នា ពោលគឺឧ។ ប្រូតេអ៊ីនស្ថិតនៅក្នុងស្ថានភាព isoelectric ។
3. ការបំបែកប្រូតេអ៊ីនបុគ្គល។លក្ខណៈពិសេសនៃរចនាសម្ព័ន្ធនិងដំណើរការនៃរាងកាយអាស្រ័យលើសំណុំនៃប្រូតេអ៊ីនដែលត្រូវបានសំយោគនៅក្នុងវា។ ការសិក្សាអំពីរចនាសម្ព័ន្ធ និងលក្ខណៈសម្បត្តិនៃប្រូតេអ៊ីនគឺមិនអាចទៅរួចទេ បើគ្មានការញែកពួកវាចេញពីកោសិកា និងបន្សុទ្ធវាពីប្រូតេអ៊ីន និងម៉ូលេគុលសរីរាង្គផ្សេងទៀត។ ដំណាក់កាលនៃភាពឯកោ និងការបន្សុទ្ធប្រូតេអ៊ីនបុគ្គល៖
ការបំផ្លាញកោសិកាជាលិកាដែលកំពុងសិក្សា និងទទួលបានភាពដូចគ្នា
ការបំបែកភាពដូចគ្នាទៅជាប្រភាគដោយ centrifugation ទទួលបាននុយក្លេអ៊ែរ mitochondrial cytosolic ឬប្រភាគផ្សេងទៀតដែលមានប្រូតេអ៊ីនដែលចង់បាន។
ការប្រែពណ៌កំដៅជ្រើសរើស- កំដៅរយៈពេលខ្លីនៃដំណោះស្រាយប្រូតេអ៊ីន ក្នុងអំឡុងពេលដែលភាពមិនបរិសុទ្ធនៃប្រូតេអ៊ីន denatured មួយចំនួនអាចត្រូវបានយកចេញ (ប្រសិនបើប្រូតេអ៊ីនមានស្ថេរភាពកំដៅ) ។
អំបិលចេញ។ប្រូតេអ៊ីនផ្សេងគ្នា precipitate នៅកំហាប់អំបិលផ្សេងគ្នានៅក្នុងដំណោះស្រាយ។ ដោយការបង្កើនកំហាប់អំបិលបន្តិចម្តងៗ វាអាចទទួលបានប្រភាគដាច់ដោយឡែកមួយចំនួនជាមួយនឹងមាតិកាលេចធ្លោនៃប្រូតេអ៊ីនដាច់ដោយឡែកនៅក្នុងមួយក្នុងចំណោមពួកវា។ អាម៉ូញ៉ូមស៊ុលហ្វាតត្រូវបានគេប្រើញឹកញាប់បំផុតសម្រាប់ការបំបែកប្រូតេអ៊ីន។ ប្រូតេអ៊ីនដែលមានជាតិរលាយតិចបំផុត precipitate នៅកំហាប់អំបិលទាប។
ការច្រោះជែល- វិធីសាស្រ្តនៃការបំបែកម៉ូលេគុលតាមរយៈគ្រាប់ Sephadex ហើម (ខ្សែសង្វាក់ polysaccharide បីវិមាត្រនៃ dextran មានរន្ធញើស) ។ ល្បឿនដែលប្រូតេអ៊ីនឆ្លងកាត់ជួរឈរដែលពោរពេញទៅដោយ Sephadex នឹងអាស្រ័យលើទម្ងន់ម៉ូលេគុលរបស់ពួកគេ៖ ម៉ាស់ម៉ូលេគុលប្រូតេអ៊ីនកាន់តែតូច ពួកវាកាន់តែងាយស្រួលជ្រាបចូលទៅក្នុង granules និងស្នាក់នៅទីនោះបានយូរ ម៉ាស់កាន់តែធំ ពួកវាកាន់តែលឿនជាងមុន ជួរឈរ។
Ultracentrifugation- វិធីសាស្រ្តដែលពាក់ព័ន្ធនឹងការដាក់ប្រូតេអ៊ីននៅក្នុងបំពង់ centrifuge ចូលទៅក្នុង rotor នៃ ultracentrifuge មួយ។ នៅពេលដែល rotor បង្វិលអត្រា sedimentation នៃប្រូតេអ៊ីនគឺសមាមាត្រទៅនឹងទម្ងន់ម៉ូលេគុលរបស់ពួកគេ: ប្រភាគនៃប្រូតេអ៊ីនធ្ងន់ជាងមានទីតាំងនៅជិតបាតនៃបំពង់សាកល្បង, ស្រាលជាង - ខិតទៅជិតផ្ទៃ។
អេឡិចត្រូហ្វីស- វិធីសាស្រ្តផ្អែកលើភាពខុសគ្នានៃល្បឿននៃចលនារបស់ប្រូតេអ៊ីននៅក្នុងវាលអគ្គិសនី។ តម្លៃនេះគឺសមាមាត្រទៅនឹងបន្ទុកនៃប្រូតេអ៊ីន។ Electrophoresis នៃប្រូតេអ៊ីនត្រូវបានអនុវត្តនៅលើក្រដាស (ក្នុងករណីនេះល្បឿននៃចលនាប្រូតេអ៊ីនគឺសមាមាត្រទៅនឹងបន្ទុករបស់វាប៉ុណ្ណោះ) ឬនៅក្នុងជែល polyacrylamide ដែលមានទំហំរន្ធញើសជាក់លាក់ (ល្បឿននៃចលនាប្រូតេអ៊ីនគឺសមាមាត្រទៅនឹងបន្ទុកនិងទម្ងន់ម៉ូលេគុលរបស់វា) ។ .
ការផ្លាស់ប្តូរអ៊ីយ៉ុងក្រូម៉ូសូម- វិធីសាស្រ្តប្រភាគផ្អែកលើការចងនៃក្រុមប្រូតេអ៊ីនដែលមានអ៊ីយ៉ូដជាមួយនឹងក្រុមដែលមានបន្ទុកផ្ទុយគ្នានៃជ័រផ្លាស់ប្តូរអ៊ីយ៉ុង (វត្ថុធាតុ polymeric មិនរលាយ) ។ កម្លាំងនៃការភ្ជាប់ប្រូតេអ៊ីនទៅនឹងជ័រគឺសមាមាត្រទៅនឹងបន្ទុកនៃប្រូតេអ៊ីន។ ប្រូតេអ៊ីនដែលត្រូវបានស្រូបយកទៅវត្ថុធាតុ polymer ផ្លាស់ប្តូរអ៊ីយ៉ុងអាចត្រូវបានទឹកនាំទៅដោយការកើនឡើងនៃកំហាប់នៃដំណោះស្រាយ NaCl ។ ការចោទប្រកាន់ប្រូតេអ៊ីនកាន់តែទាប កំហាប់នៃ NaCl កាន់តែទាបដែលត្រូវការដើម្បីលាងសម្អាតប្រូតេអ៊ីនដែលភ្ជាប់ទៅនឹងក្រុមអ៊ីយ៉ុងនៃជ័រ។
ភាពស្និទ្ធស្នាល chromatography- វិធីសាស្រ្តជាក់លាក់បំផុតសម្រាប់ញែកប្រូតេអ៊ីនបុគ្គល លីហ្គែននៃប្រូតេអ៊ីនមួយត្រូវបានភ្ជាប់ជាមួយវត្ថុធាតុ polymer inert ។ នៅពេលដែលដំណោះស្រាយប្រូតេអ៊ីនត្រូវបានឆ្លងកាត់ជួរឈរជាមួយវត្ថុធាតុ polymer មានតែប្រូតេអ៊ីនជាក់លាក់សម្រាប់ ligand ដែលបានផ្តល់ឱ្យប៉ុណ្ណោះដែលត្រូវបានស្រូបយកនៅលើជួរឈរដោយសារតែការភ្ជាប់ប្រូតេអ៊ីនបំពេញបន្ថែមទៅនឹង ligand ។
ការលាងឈាម- វិធីសាស្រ្តប្រើដើម្បីយកសមាសធាតុទម្ងន់ម៉ូលេគុលទាបចេញពីដំណោះស្រាយនៃប្រូតេអ៊ីនដាច់ដោយឡែក។ វិធីសាស្រ្តនេះគឺផ្អែកលើអសមត្ថភាពនៃប្រូតេអ៊ីនដើម្បីឆ្លងកាត់ភ្នាសពាក់កណ្តាល permeable មិនដូចសារធាតុទម្ងន់ម៉ូលេគុលទាប។ វាត្រូវបានគេប្រើដើម្បីបន្សុទ្ធប្រូតេអ៊ីនពីភាពមិនបរិសុទ្ធនៃម៉ូលេគុលទាប ឧទាហរណ៍ អំបិលបន្ទាប់ពីអំបិលចេញ។
កិច្ចការសម្រាប់ការងារក្រៅម៉ោង
1. បំពេញតារាង។ ១.៤.
តារាង 1.4 ។ ការវិភាគប្រៀបធៀបនៃរចនាសម្ព័ន្ធនិងមុខងារនៃប្រូតេអ៊ីនដែលពាក់ព័ន្ធ - myoglobin និង hemoglobin
ក) ចងចាំរចនាសម្ព័ន្ធនៃមជ្ឈមណ្ឌលសកម្មនៃ Mb និង Hb ។ តើរ៉ាឌីកាល់អាស៊ីតអាមីណូ hydrophobic មានតួនាទីអ្វីនៅក្នុងការបង្កើតមជ្ឈមណ្ឌលសកម្មនៃប្រូតេអ៊ីនទាំងនេះ? ពិពណ៌នាអំពីរចនាសម្ព័ន្ធនៃមជ្ឈមណ្ឌលសកម្មនៃ Mb និង Hb និងយន្តការនៃការភ្ជាប់នៃ ligands ទៅវា។ តើសំណល់ F 8 និង E 7 របស់គាត់មានតួនាទីអ្វីនៅក្នុងដំណើរការនៃមជ្ឈមណ្ឌលសកម្មនៃ Mv iHv?
ខ) តើលក្ខណៈសម្បត្តិថ្មីអ្វីខ្លះ បើប្រៀបធៀបទៅនឹង myoglobin monomeric តើប្រូតេអ៊ីន oligomeric, hemoglobin មានទំនាក់ទំនងយ៉ាងជិតស្និទ្ធដែរឬទេ? ពន្យល់ពីតួនាទីនៃការផ្លាស់ប្តូរសហករណ៍ក្នុងការអនុលោមតាមប្រូតូមឺរនៅក្នុងម៉ូលេគុលអេម៉ូក្លូប៊ីន ឥទ្ធិពលនៃ CO 2 និងការប្រមូលផ្តុំប្រូតុងលើភាពស្និទ្ធស្នាលនៃអេម៉ូក្លូប៊ីនសម្រាប់អុកស៊ីសែន ក៏ដូចជាតួនាទីរបស់ 2,3-BPG ក្នុងបទប្បញ្ញត្តិ allosteric នៃមុខងារ Hb .
2. កំណត់លក្ខណៈរបស់ chaperones ម៉ូលេគុលដោយយកចិត្តទុកដាក់លើទំនាក់ទំនងរវាងរចនាសម្ព័ន្ធនិងមុខងាររបស់វា។
3. តើប្រូតេអ៊ីនអ្វីខ្លះត្រូវបានដាក់ជាក្រុម? ដោយប្រើឧទាហរណ៍នៃគ្រួសារ immunoglobulin កំណត់លក្ខណៈរចនាសម្ព័ន្ធស្រដៀងគ្នានិងមុខងារដែលទាក់ទងនៃប្រូតេអ៊ីននៃគ្រួសារនេះ។
4. ប្រូតេអ៊ីនបុគ្គលដែលបន្សុតត្រូវបានទាមទារជាញឹកញាប់សម្រាប់គោលបំណងជីវគីមី និងឱសថ។ ពន្យល់អំពីលក្ខណៈសម្បត្តិរូបវិទ្យានៃប្រូតេអ៊ីន វិធីសាស្ត្រដែលប្រើសម្រាប់ការបំបែក និងការបន្សុតរបស់វាត្រូវបានផ្អែកលើ។
ភារកិច្ចគ្រប់គ្រងខ្លួនឯង
1. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។
មុខងាររបស់អេម៉ូក្លូប៊ីន៖
A. ការដឹកជញ្ជូន O 2 ពីសួតទៅជាលិកា B. ការដឹកជញ្ជូន H + ពីជាលិកាទៅសួត
B. រក្សា pH ឈាមថេរ D. ការដឹកជញ្ជូន CO 2 ពីសួតទៅជាលិកា
ឃ. ការដឹកជញ្ជូន CO 2 ពីជាលិកាទៅសួត
2. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។ លីហ្គែនα -protomer Hb គឺ៖ A. ហេម
ខ.អុកស៊ីហ្សែន
B. CO G. 2,3-BPG
D. β-Protomer
3. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។
អេម៉ូក្លូប៊ីនប្រឆាំងនឹង myoglobin៖
A. មានរចនាសម្ព័ន្ធបួនជ្រុង
B. រចនាសម្ព័ន្ធបន្ទាប់បន្សំត្រូវបានតំណាងដោយ α-helices ប៉ុណ្ណោះ។
ខ.ជាកម្មសិទ្ធិរបស់ប្រូតេអ៊ីនស្មុគស្មាញ
D. អន្តរកម្មជាមួយ ligand allosteric D. Covalently ចងភ្ជាប់ទៅនឹង heme
4. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។
ភាពស្និទ្ធស្នាលរបស់ Hb សម្រាប់ O2 ថយចុះ៖
A. នៅពេលដែលម៉ូលេគុល O 2 មួយត្រូវបានបន្ថែម B. នៅពេលដែលម៉ូលេគុល O 2 មួយត្រូវបានដកចេញ
ខ.នៅពេលធ្វើអន្តរកម្មជាមួយ 2,3-BPG
D. នៅពេលដែលភ្ជាប់ទៅនឹង protomers H + D. នៅពេលដែលកំហាប់នៃ 2,3-BPG ថយចុះ
5. ការប្រកួត។
ប្រភេទ HB ត្រូវបានកំណត់លក្ខណៈដោយ៖
A. ក្នុងទម្រង់ deoxy វាបង្កើតជា fibrillar aggregates B. មាន α- និង δ-chains ពីរ
B. ទម្រង់លេចធ្លោនៃ Hb នៅក្នុង erythrocytes មនុស្សពេញវ័យ D. មានផ្ទុក heme ជាមួយ Fe+ 3 នៅក្នុងមជ្ឈមណ្ឌលសកម្ម
ឃ.មាន α- និងខ្សែសង្វាក់ γ ពីរ 1. HbA 2 ។
6. ការប្រកួត។
លីហ្គែន Hb៖
A. ភ្ជាប់ទៅ Hb នៅក្នុងមជ្ឈមណ្ឌល allosteric
B. មានភាពស្និទ្ធស្នាលខ្ពស់សម្រាប់គេហទំព័រសកម្មរបស់ Hb
ខ. តាមរយៈការភ្ជាប់ វាបង្កើនភាពស្និទ្ធស្នាលរបស់ Hb សម្រាប់ O 2 G. Oxidizes Fe+ 2 ទៅ Fe+ 3
D. បង្កើតចំណងកូវ៉ាឡេនជាមួយ F8 របស់គាត់។
7. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។
អ្នកដឹកនាំ៖
ក. ប្រូតេអ៊ីនមាននៅគ្រប់ផ្នែកទាំងអស់នៃកោសិកា
ខ.ការសំយោគកើនឡើងក្រោមភាពតានតឹង
ខ. ចូលរួមក្នុង hydrolysis នៃប្រូតេអ៊ីន denatured
ឃ. ចូលរួមក្នុងការរក្សាការអនុលោមតាមប្រភពដើមនៃប្រូតេអ៊ីន
ឃ. បង្កើតសរីរាង្គដែលការអនុលោមតាមប្រូតេអ៊ីនត្រូវបានបង្កើតឡើង
8. ការប្រកួត។ Immunoglobulins៖
A. ទម្រង់សម្ងាត់គឺ pentameric ។
B. Class Ig ដែលជ្រាបចូលទៅក្នុងរបាំងសុក
B. Ig - mast cell receptor
ឃ. ថ្នាក់សំខាន់នៃ Ig មានវត្តមាននៅក្នុងអាថ៌កំបាំងនៃកោសិកា epithelial ។ D. B-lymphocyte receptor ដែលជាការធ្វើឱ្យសកម្មដែលធានានូវការរីកសាយកោសិកា
9. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។
Immunoglobulins អ៊ី៖
A. ផលិតដោយ macrophages B. ពួកវាមានច្រវាក់εធ្ងន់។
ខ.បានបង្កប់នៅក្នុងភ្នាសនៃ T-lymphocytes
D. ដើរតួជាអ្នកទទួលភ្នាសអង់ទីហ្សែននៅលើកោសិកា mast និង basophils
D. ទទួលខុសត្រូវចំពោះប្រតិកម្មអាលែហ្សី
10. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។
វិធីសាស្រ្តបំបែកប្រូតេអ៊ីនគឺផ្អែកលើភាពខុសគ្នានៃទម្ងន់ម៉ូលេគុលរបស់ពួកគេ៖
A. ការច្រោះជែល
ខ. អ៊ុលត្រាហ្សេហ្វហ្គាស
B. Polyacrylamide gel electrophoresis D. Ion exchange chromatography
D. Affinity chromatography
11. ជ្រើសរើសចម្លើយដែលត្រឹមត្រូវ។
វិធីសាស្រ្តបំបែកប្រូតេអ៊ីនគឺផ្អែកលើភាពខុសគ្នានៃភាពរលាយរបស់វានៅក្នុងទឹក៖
A. Gel filtration B. Salting out
ខ. ការផ្លាស់ប្តូរអ៊ីយ៉ុងក្រូម៉ាតូក្រាម ឃ
D. Electrophoresis ក្នុង polyacrylamide gel
ស្តង់ដារនៃចម្លើយចំពោះ "កិច្ចការគ្រប់គ្រងខ្លួនឯង"
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
លក្ខខណ្ឌមូលដ្ឋាន និងគោលគំនិត
1. ប្រូតេអ៊ីន Oligomeric, protomer, រចនាសម្ព័ន្ធ quaternary នៃប្រូតេអ៊ីន
2. ការផ្លាស់ប្តូរសហករណ៍នៅក្នុងការអនុលោមតាម protomer
3. ឥទ្ធិពល Bohr
4. បទប្បញ្ញត្តិ Allosteric នៃមុខងារប្រូតេអ៊ីន មជ្ឈមណ្ឌល allosteric និង allosteric effector
5. ម៉ូលេគុល chaperones ប្រូតេអ៊ីនឆក់កំដៅ
6. ក្រុមគ្រួសារប្រូតេអ៊ីន (ប្រូតេអ៊ីន serine, immunoglobulins)
7. IgM-, G-, E-, A-structure-function ទំនាក់ទំនង
8. បន្ទុកសរុបនៃប្រូតេអ៊ីន ចំណុច isoelectric នៃប្រូតេអ៊ីន
9. Electrophoresis
10. អំបិលចេញ
11. ការច្រោះជែល
12. ការផ្លាស់ប្តូរ chromatography អ៊ីយ៉ុង
13. Ultracentrifugation
14. Affinity chromatography
15. Electrophoresis នៃប្រូតេអ៊ីនប្លាស្មាឈាម
កិច្ចការសម្រាប់ការងារក្នុងថ្នាក់រៀន
1. ប្រៀបធៀបភាពអាស្រ័យនៃកម្រិតនៃការតិត្ថិភាពនៃអេម៉ូក្លូប៊ីន (Hb) និង myoglobin (Mb) ជាមួយនឹងអុកស៊ីសែននៅលើសម្ពាធផ្នែករបស់វានៅក្នុងជាលិកា។
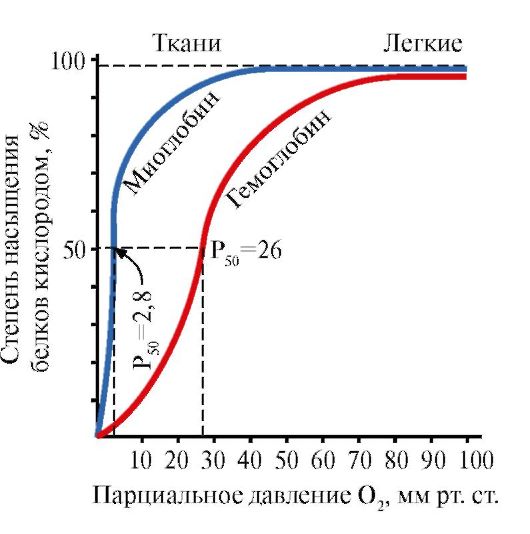 អង្ករ។ ១.៣៤. ការអាស្រ័យនៃតិត្ថិភាព Mv និងNHអុកស៊ីសែនពីសម្ពាធផ្នែករបស់វា។
អង្ករ។ ១.៣៤. ការអាស្រ័យនៃតិត្ថិភាព Mv និងNHអុកស៊ីសែនពីសម្ពាធផ្នែករបស់វា។
សូមចំណាំថារូបរាងនៃខ្សែកោងតិត្ថិភាពអុកស៊ីហ៊្សែនរបស់ប្រូតេអ៊ីនគឺខុសគ្នា: សម្រាប់ myoglobin - អ៊ីពែបូឡាសម្រាប់អេម៉ូក្លូប៊ី - រាង sigmoid ។
1. ប្រៀបធៀបតម្លៃនៃសម្ពាធផ្នែកនៃអុកស៊ីសែនដែល Mb និង Hb ត្រូវបានឆ្អែតជាមួយ O 2 ដោយ 50% ។ តើប្រូតេអ៊ីនមួយណាដែលមានទំនាក់ទំនងល្អជាងសម្រាប់ O 2?
2. តើលក្ខណៈរចនាសម្ព័ន្ធអ្វីខ្លះរបស់ Mb កំណត់ភាពស្និទ្ធស្នាលខ្ពស់របស់វាសម្រាប់ O 2?
3. តើលក្ខណៈរចនាសម្ព័ន្ធអ្វីខ្លះនៃ HB អនុញ្ញាតឱ្យវាបញ្ចេញ O2 នៅក្នុង capillaries នៃជាលិកាសម្រាក (នៅសម្ពាធផ្នែកខ្លះនៃ O2) និងបង្កើនការបញ្ចេញនេះយ៉ាងខ្លាំងនៅក្នុងសាច់ដុំធ្វើការ? តើអ្វីទៅជាទ្រព្យសម្បត្តិនៃប្រូតេអ៊ីន oligomeric ផ្តល់នូវឥទ្ធិពលនេះ?
4. គណនាបរិមាណ O 2 (គិតជា%) អេម៉ូក្លូប៊ីនអុកស៊ីហ្សែនផ្តល់ដល់សាច់ដុំសម្រាក និងធ្វើការ?
5. ធ្វើការសន្និដ្ឋានអំពីទំនាក់ទំនងរវាងរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីន និងមុខងាររបស់វា។
2. បរិមាណអុកស៊ីសែនដែលបញ្ចេញដោយអេម៉ូក្លូប៊ីននៅក្នុងសរសៃឈាមគឺអាស្រ័យលើអាំងតង់ស៊ីតេនៃដំណើរការ catabolic នៅក្នុងជាលិកា (ឥទ្ធិពល Bohr) ។ តើការផ្លាស់ប្តូរការរំលាយអាហារជាលិកាគ្រប់គ្រងទំនាក់ទំនងនៃ Hb សម្រាប់ O2 យ៉ាងដូចម្តេច? ឥទ្ធិពលនៃ CO 2 និង H+ លើទំនាក់ទំនងនៃ Hb សម្រាប់ O 2
 1. ពិពណ៌នាអំពីឥទ្ធិពល Bohr ។
1. ពិពណ៌នាអំពីឥទ្ធិពល Bohr ។
2. តើដំណើរការដែលបង្ហាញក្នុងដ្យាក្រាមដំណើរការក្នុងទិសដៅអ្វី៖
ក) នៅក្នុង capillaries នៃសួត;
ខ) នៅក្នុងជាលិកា capillaries?
3. តើអ្វីទៅជាសារៈសំខាន់ខាងសរីរវិទ្យានៃឥទ្ធិពល Bohr?
4. ហេតុអ្វីបានជាអន្តរកម្មនៃ Hb ជាមួយ H+ នៅកន្លែងដាច់ស្រយាលពី heme ផ្លាស់ប្តូរភាពស្និទ្ធស្នាលនៃប្រូតេអ៊ីនសម្រាប់ O 2?
3. ភាពស្និទ្ធស្នាលនៃ Hb សម្រាប់ O2 អាស្រ័យលើកំហាប់នៃ ligand របស់វា - 2,3-bisphosphoglycerate ដែលជានិយតករ allosteric នៃភាពស្និទ្ធស្នាលនៃ Hb សម្រាប់ O2 ។ ហេតុអ្វីបានជាអន្តរកម្ម ligand នៅកន្លែងដែលឆ្ងាយពីកន្លែងសកម្មប៉ះពាល់ដល់មុខងារប្រូតេអ៊ីន? តើ 2,3-BPG គ្រប់គ្រងភាពស្និទ្ធស្នាលរបស់ Hb សម្រាប់ O2 យ៉ាងដូចម្តេច? ដើម្បីដោះស្រាយបញ្ហា សូមឆ្លើយសំណួរខាងក្រោម៖
1. តើ 2.3-bisphosphoglycerate (2,3-BPG) សំយោគមកពីណា? សរសេររូបមន្តរបស់វា បង្ហាញពីបន្ទុកនៃម៉ូលេគុលនេះ។
2. តើទម្រង់នៃអេម៉ូក្លូប៊ីន (អុកស៊ីដ ឬឌីអុកស៊ីត) BPG មានអន្តរកម្មជាមួយនិងហេតុអ្វី? តើអន្តរកម្មកើតឡើងនៅក្នុងផ្នែកណានៃម៉ូលេគុល Hb?
3. តើដំណើរការដែលបង្ហាញក្នុងដ្យាក្រាមកើតឡើងក្នុងទិសដៅអ្វី?
ក) នៅក្នុង capillaries ជាលិកា;
ខ) នៅក្នុង capillaries នៃសួត?
4. កន្លែងដែលកំហាប់នៃស្មុគស្មាញគួរតែខ្ពស់ជាង
Nv-2,3-BFG៖
ក) នៅក្នុង capillaries នៃសាច់ដុំនៅពេលសម្រាក,
ខ) នៅក្នុង capillaries នៃសាច់ដុំធ្វើការ (ផ្តល់កំហាប់ដូចគ្នានៃ BPG នៅក្នុង erythrocytes)?
5. តើភាពស្និទ្ធស្នាលរបស់ HB សម្រាប់អុកស៊ីហ្សែននឹងផ្លាស់ប្តូរយ៉ាងដូចម្តេច នៅពេលដែលមនុស្សម្នាក់សម្របខ្លួនទៅនឹងលក្ខខណ្ឌកម្ពស់ខ្ពស់ ប្រសិនបើកំហាប់ BPG នៅក្នុងអេរីត្រូស៊ីតកើនឡើង? តើអ្វីជាសារៈសំខាន់ខាងសរីរវិទ្យានៃបាតុភូតនេះ?
4. ការបំផ្លិចបំផ្លាញនៃ 2,3-BPG ក្នុងអំឡុងពេលផ្ទុកឈាមដែលបានបម្រុងទុកធ្វើឱ្យខូចមុខងាររបស់ HB ។ តើភាពស្និទ្ធស្នាលនៃ HB សម្រាប់ O 2 នឹងផ្លាស់ប្តូរយ៉ាងដូចម្តេចនៅក្នុងឈាមដែលបានបម្រុងទុកប្រសិនបើកំហាប់នៃ 2,3-BPG នៅក្នុងអេរីត្រូស៊ីតអាចថយចុះពី 8 ទៅ 0,5 មីល្លីលីត្រ / លីត្រ។ តើវាអាចទៅរួចទេក្នុងការបញ្ជូនឈាមទៅអ្នកជំងឺធ្ងន់ធ្ងរ ប្រសិនបើកំហាប់ 2,3-BPG ត្រូវបានស្តារឡើងវិញមិនលឿនជាងបីថ្ងៃ? តើវាអាចទៅរួចទេក្នុងការស្តារមុខងារកោសិកាឈាមក្រហមដោយបន្ថែម 2,3-BPG ទៅក្នុងឈាម?
5. ចងចាំរចនាសម្ព័ន្ធនៃម៉ូលេគុល immunoglobulin សាមញ្ញបំផុត។ តើ immunoglobulins មានតួនាទីអ្វីខ្លះនៅក្នុងប្រព័ន្ធភាពស៊ាំ? ហេតុអ្វីបានជា Igs ត្រូវបានគេហៅថា bivalents? តើរចនាសម្ព័ន្ធរបស់ Igs ទាក់ទងនឹងមុខងាររបស់វាយ៉ាងដូចម្តេច? (ពិពណ៌នាដោយប្រើឧទាហរណ៍នៃថ្នាក់នៃ immunoglobulins ។ )
លក្ខណៈសម្បត្តិរូបវិទ្យានៃប្រូតេអ៊ីននិងវិធីសាស្រ្តនៃការបំបែករបស់ពួកគេ។
6. តើបន្ទុកសុទ្ធនៃប្រូតេអ៊ីនប៉ះពាល់ដល់ការរលាយរបស់វាយ៉ាងដូចម្តេច?
ក) កំណត់បន្ទុកសរុបនៃ peptide នៅ pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
ខ) តើការចោទប្រកាន់នៃ peptide នេះនឹងផ្លាស់ប្តូរយ៉ាងដូចម្តេចនៅ pH > 7, pH<7, рН <<7?
គ) តើអ្វីជាចំណុចអ៊ីសូអេឡិចត្រិចនៃប្រូតេអ៊ីន (IEP) ហើយតើវាស្ថិតនៅក្នុងបរិយាកាសអ្វី?
IET នៃ peptide នេះ?
ឃ) នៅតម្លៃ pH ណាដែលការរលាយតិចបំផុតនៃ peptide នេះនឹងត្រូវបានអង្កេត។
7. ហេតុអ្វីបានជាទឹកដោះគោជូរមិនដូចទឹកដោះគោស្រស់ “ខាញ់” នៅពេលស្ងោរ (ឧទាហរណ៍ ប្រូតេអ៊ីនទឹកដោះគោ ខេស៊ីន ធ្លាក់ទឹក)? នៅក្នុងទឹកដោះគោស្រស់ ម៉ូលេគុល casein មានបន្ទុកអវិជ្ជមាន។
8. ការច្រោះជែលត្រូវបានប្រើដើម្បីបំបែកប្រូតេអ៊ីនបុគ្គល។ ល្បាយដែលមានប្រូតេអ៊ីន A, B, C ដែលមានទម្ងន់ម៉ូលេគុលស្មើនឹង 160,000, 80,000 និង 60,000 រៀងគ្នា ត្រូវបានវិភាគដោយការច្រោះជែល (រូបភាព 1.35)។ គ្រាប់ជែលដែលហើមអាចជ្រាបចូលបានទៅនឹងប្រូតេអ៊ីនដែលមានទម្ងន់ម៉ូលេគុលតិចជាង 70,000 តើគោលការណ៍អ្វីខ្លះដែលបញ្ជាក់ពីវិធីសាស្ត្របំបែកនេះ? តើក្រាហ្វមួយណាដែលឆ្លុះបញ្ចាំងយ៉ាងត្រឹមត្រូវនូវលទ្ធផលនៃប្រភាគ? ចង្អុលបង្ហាញលំដាប់ដែលប្រូតេអ៊ីន A, B, និង C ត្រូវបានបញ្ចេញចេញពីជួរឈរ។
 អង្ករ។ ១.៣៥. ការប្រើតម្រងជែលដើម្បីបំបែកប្រូតេអ៊ីន
អង្ករ។ ១.៣៥. ការប្រើតម្រងជែលដើម្បីបំបែកប្រូតេអ៊ីន
9. នៅក្នុងរូបភព។ 1.36, A បង្ហាញដ្យាក្រាមនៃ electrophoresis នៅលើក្រដាសនៃប្រូតេអ៊ីនសេរ៉ូមឈាមពីមនុស្សដែលមានសុខភាពល្អ។ បរិមាណដែលទាក់ទងនៃប្រភាគប្រូតេអ៊ីនដែលទទួលបានដោយប្រើវិធីនេះគឺ៖ អាល់ប៊ុយមីន 54-58%, α 1 -globulins 6-7%, α 2 -globulins 8-9%, β-globulins 13%, γ-globulins 11-12% ។
 អង្ករ។ 1.36 Electrophoresis នៅលើក្រដាសនៃប្រូតេអ៊ីនប្លាស្មាឈាមរបស់មនុស្សដែលមានសុខភាពល្អ (A) និងអ្នកជំងឺ (B)
អង្ករ។ 1.36 Electrophoresis នៅលើក្រដាសនៃប្រូតេអ៊ីនប្លាស្មាឈាមរបស់មនុស្សដែលមានសុខភាពល្អ (A) និងអ្នកជំងឺ (B)
ខ្ញុំ - γ-globulins; II - β-globulins; III -α 2 -globulin; IV -α 2 -globulin; វី - អាល់ប៊ុមប៊ីន
ជំងឺជាច្រើនត្រូវបានអមដោយការផ្លាស់ប្តូរបរិមាណនៅក្នុងសមាសភាពនៃប្រូតេអ៊ីនសេរ៉ូម (dysproteinemia) ។ ធម្មជាតិនៃការផ្លាស់ប្តូរទាំងនេះត្រូវបានយកមកពិចារណានៅពេលធ្វើការធ្វើរោគវិនិច្ឆ័យ និងវាយតម្លៃពីភាពធ្ងន់ធ្ងរ និងដំណាក់កាលនៃជំងឺ។
ការប្រើប្រាស់ទិន្នន័យដែលបានផ្តល់ឱ្យក្នុងតារាង។ 1.5 ធ្វើការទាយអំពីជំងឺនេះ ដែលត្រូវបានកំណត់ដោយទម្រង់ electrophoretic ដែលបង្ហាញក្នុងរូប។ ១.៣៦.
តារាង 1.5 ។ ការផ្លាស់ប្តូរនៅក្នុងការប្រមូលផ្តុំនៃប្រូតេអ៊ីនសេរ៉ូមនៅក្នុងរោគវិទ្យា
កំប្រុក- សមាសធាតុសរីរាង្គទម្ងន់ម៉ូលេគុលខ្ពស់ដែលមានសំណល់អាស៊ីតអាមីណូ។
IN សមាសភាពប្រូតេអ៊ីនរួមមានកាបូន អ៊ីដ្រូសែន អាសូត អុកស៊ីហ្សែន ស្ពាន់ធ័រ។ ប្រូតេអ៊ីនខ្លះបង្កើតជាស្មុគស្មាញជាមួយម៉ូលេគុលផ្សេងទៀតដែលមានផូស្វ័រ ជាតិដែក ស័ង្កសី និងទង់ដែង។
ប្រូតេអ៊ីនមានទម្ងន់ម៉ូលេគុលធំ៖ ស៊ុតអាល់ប៊ុមមីន - ៣៦,០០០, អេម៉ូក្លូប៊ីន - ១៥២,០០០, មីយ៉ូស៊ីន - ៥០០,០០០ សម្រាប់ការប្រៀបធៀប៖ ទម្ងន់ម៉ូលេគុលនៃជាតិអាល់កុលគឺ ៤៦ អាស៊ីតអាសេទិក - ៦០ បេនហ្សេន - ៧៨ ។
សមាសធាតុអាស៊ីតអាមីណូនៃប្រូតេអ៊ីន
កំប្រុក- ប៉ូលីម៊ែរមិនតាមកាលកំណត់, ម៉ូណូមឺរដែលមាន អាស៊ីតអាមីណូ. ជាធម្មតា អាស៊ីតអាមីណូ 20 ប្រភេទ ត្រូវបានគេហៅថា monomers ប្រូតេអ៊ីន ទោះបីជាជាង 170 ក្នុងចំណោមពួកវាត្រូវបានរកឃើញនៅក្នុងកោសិកា និងជាលិកាក៏ដោយ។
អាស្រ័យលើថាតើអាស៊ីតអាមីណូអាចត្រូវបានសំយោគនៅក្នុងរាងកាយរបស់មនុស្ស និងសត្វផ្សេងទៀត ពួកគេត្រូវបានសម្គាល់: អាស៊ីតអាមីណូដែលមិនសំខាន់- អាចត្រូវបានសំយោគ; អាស៊ីតអាមីណូសំខាន់ៗ- មិនអាចសំយោគបានទេ។ អាស៊ីតអាមីណូសំខាន់ៗត្រូវតែផ្គត់ផ្គង់ដល់រាងកាយតាមរយៈអាហារ។ រុក្ខជាតិសំយោគអាស៊ីតអាមីណូគ្រប់ប្រភេទ។
អាស្រ័យលើសមាសធាតុអាស៊ីតអាមីណូ។ ប្រូតេអ៊ីនគឺ: ពេញលេញ- មានសំណុំអាស៊ីតអាមីណូទាំងមូល; ខូច- អាស៊ីតអាមីណូមួយចំនួនត្រូវបានបាត់នៅក្នុងសមាសភាពរបស់វា។ ប្រសិនបើប្រូតេអ៊ីនមានតែអាស៊ីតអាមីណូទេនោះគេហៅថា សាមញ្ញ. ប្រសិនបើប្រូតេអ៊ីនមាន បន្ថែមពីលើអាស៊ីតអាមីណូ ដែលជាសមាសធាតុអាស៊ីតអាមីណូ (ក្រុមសិប្បនិម្មិត) ពួកគេត្រូវបានគេហៅថា ស្មុគស្មាញ. ក្រុមសិប្បនិម្មិតអាចត្រូវបានតំណាងដោយលោហៈ (metalloproteins) កាបូអ៊ីដ្រាត (glycoproteins) lipid (lipoproteins) អាស៊ីត nucleic (nucleoproteins) ។
ទាំងអស់។ អាស៊ីតអាមីណូមាន: 1) ក្រុម carboxyl (-COOH), 2) ក្រុមអាមីណូ (-NH 2), 3) រ៉ាឌីកាល់ឬក្រុម R (នៅសល់នៃម៉ូលេគុល) ។ រចនាសម្ព័ន្ធនៃរ៉ាឌីកាល់គឺខុសគ្នាសម្រាប់ប្រភេទផ្សេងគ្នានៃអាស៊ីតអាមីណូ។ អាស្រ័យលើចំនួនក្រុមអាមីណូ និងក្រុម carboxyl ដែលរួមបញ្ចូលក្នុងសមាសភាពនៃអាស៊ីតអាមីណូ ពួកគេត្រូវបានសម្គាល់៖ អាស៊ីតអាមីណូអព្យាក្រឹតមានក្រុម carboxyl មួយនិងក្រុមអាមីណូមួយ; អាស៊ីតអាមីណូមូលដ្ឋានមានក្រុមអាមីណូច្រើនជាងមួយ; អាស៊ីតអាមីណូអាស៊ីតមានក្រុម carboxyl ច្រើនជាងមួយ។
អាស៊ីតអាមីណូគឺ សមាសធាតុ amphotericចាប់តាំងពីនៅក្នុងដំណោះស្រាយ ពួកគេអាចដើរតួជាអាស៊ីត និងមូលដ្ឋាន។ នៅក្នុងដំណោះស្រាយ aqueous អាស៊ីតអាមីណូមាននៅក្នុងទម្រង់អ៊ីយ៉ុងផ្សេងៗគ្នា។
ចំណង Peptide
ប៉េទីត- សារធាតុសរីរាង្គដែលមានសំណល់អាស៊ីតអាមីណូតភ្ជាប់ដោយចំណង peptide ។
ការបង្កើត peptides កើតឡើងជាលទ្ធផលនៃប្រតិកម្ម condensation នៃអាស៊ីតអាមីណូ។ នៅពេលដែលក្រុមអាមីណូនៃអាស៊ីតអាមីណូមួយធ្វើអន្តរកម្មជាមួយក្រុម carboxyl នៃក្រុមមួយទៀត ចំណងអាសូត-កាបូន covalent កើតឡើងរវាងពួកវា ដែលត្រូវបានគេហៅថា peptide. អាស្រ័យលើចំនួននៃសំណល់អាស៊ីតអាមីណូដែលរួមបញ្ចូលនៅក្នុង peptide មាន dipeptides, tripeptides, tetrapeptidesល។ ការបង្កើតចំណង peptide អាចត្រូវបានធ្វើម្តងទៀតច្រើនដង។ នេះនាំឱ្យមានការបង្កើត polypeptides. នៅចុងម្ខាងនៃ peptide មានក្រុមអាមីណូឥតគិតថ្លៃ (ហៅថា N-terminus) ហើយនៅម្ខាងទៀតមានក្រុម carboxyl សេរី (ហៅថា C-terminus)។

អង្គការលំហនៃម៉ូលេគុលប្រូតេអ៊ីន
ដំណើរការនៃមុខងារជាក់លាក់មួយចំនួនដោយប្រូតេអ៊ីនអាស្រ័យលើការកំណត់រចនាសម្ព័ន្ធនៃម៉ូលេគុលរបស់វាលើសពីនេះទៀត វាមិនអំណោយផលខ្លាំងសម្រាប់កោសិកាក្នុងការរក្សាប្រូតេអ៊ីនក្នុងទម្រង់ជាខ្សែសង្វាក់ ដូច្នេះហើយខ្សែសង្វាក់ polypeptide ឆ្លងកាត់ការបត់។ រចនាសម្ព័ន្ធបីវិមាត្រជាក់លាក់ ឬទម្រង់។ មាន 4 កម្រិត អង្គការ spatial នៃប្រូតេអ៊ីន.
រចនាសម្ព័ន្ធប្រូតេអ៊ីនបឋម- លំដាប់នៃការរៀបចំសំណល់អាស៊ីតអាមីណូនៅក្នុងខ្សែសង្វាក់ polypeptide ដែលបង្កើតជាម៉ូលេគុលប្រូតេអ៊ីន។ ចំណងរវាងអាស៊ីតអាមីណូគឺជាចំណង peptide ។

ប្រសិនបើម៉ូលេគុលប្រូតេអ៊ីនមានសំណល់អាស៊ីដអាមីណូចំនួន 10 ប៉ុណ្ណោះ នោះចំនួននៃបំរែបំរួលតាមទ្រឹស្តីនៃម៉ូលេគុលប្រូតេអ៊ីនដែលខុសគ្នាតាមលំដាប់នៃការផ្លាស់ប្តូរអាស៊ីតអាមីណូគឺ 10 20 ។ មានអាស៊ីតអាមីណូចំនួន 20 អ្នកអាចបង្កើតការផ្សំចម្រុះកាន់តែច្រើនពីពួកវា។ ប្រហែលមួយម៉ឺនប្រូតេអ៊ីនផ្សេងៗគ្នាត្រូវបានគេរកឃើញនៅក្នុងខ្លួនមនុស្ស ដែលខុសគ្នាទាំងពីគ្នាទៅវិញទៅមក និងពីប្រូតេអ៊ីននៃសារពាង្គកាយផ្សេងទៀត។
វាគឺជារចនាសម្ព័ន្ធចម្បងនៃម៉ូលេគុលប្រូតេអ៊ីនដែលកំណត់លក្ខណៈសម្បត្តិនៃម៉ូលេគុលប្រូតេអ៊ីន និងការកំណត់រចនាសម្ព័ន្ធលំហរបស់វា។ ការជំនួសអាស៊ីតអាមីណូតែមួយជាមួយខ្សែសង្វាក់ polypeptide នាំឱ្យមានការផ្លាស់ប្តូរលក្ខណៈសម្បត្តិ និងមុខងាររបស់ប្រូតេអ៊ីន។ ឧទាហរណ៍ ការជំនួសអាស៊ីតអាមីណូ glutamic ទីប្រាំមួយជាមួយ valine ក្នុងβ-subunit នៃ hemoglobin នាំឱ្យការពិតដែលថាម៉ូលេគុល hemoglobin ទាំងមូលមិនអាចអនុវត្តមុខងារចម្បងរបស់វា - ការដឹកជញ្ជូនអុកស៊ីសែន; ក្នុងករណីបែបនេះ មនុស្សនោះកើតជំងឺម្យ៉ាងដែលហៅថាជំងឺស្លេកស្លាំងកោសិកាជំងឺ។
រចនាសម្ព័ន្ធបន្ទាប់បន្សំ- បានបញ្ជាឱ្យបត់ខ្សែសង្វាក់ polypeptide ចូលទៅក្នុងវង់មួយ (មើលទៅដូចជានិទាឃរដូវពង្រីក) ។ វេននៃ helix ត្រូវបានពង្រឹងដោយចំណងអ៊ីដ្រូសែនដែលកើតឡើងរវាងក្រុម carboxyl និងក្រុមអាមីណូ។ ស្ទើរតែគ្រប់ក្រុម CO និង NH ចូលរួមក្នុងការបង្កើតចំណងអ៊ីដ្រូសែន។ ពួកវាខ្សោយជាងសារធាតុ peptide ប៉ុន្តែ ធ្វើម្តងទៀតច្រើនដង ផ្តល់ស្ថេរភាព និងភាពរឹងដល់ការកំណត់រចនាសម្ព័ន្ធនេះ។ នៅកម្រិតនៃរចនាសម្ព័ន្ធបន្ទាប់បន្សំមានប្រូតេអ៊ីន: fibroin (សូត្រ, បណ្តាញពីងពាង), keratin (សក់, ក្រចក), collagen (សរសៃពួរ) ។



រចនាសម្ព័ន្ធទីបី- ការវេចខ្ចប់ខ្សែសង្វាក់ polypeptide ចូលទៅក្នុង globules ដែលបណ្តាលមកពីការបង្កើតចំណងគីមី (អ៊ីដ្រូសែន អ៊ីយ៉ុង ឌីស៊ុលហ្វីត) និងការបង្កើតអន្តរកម្ម hydrophobic រវាងរ៉ាឌីកាល់នៃសំណល់អាស៊ីតអាមីណូ។ តួនាទីសំខាន់ក្នុងការបង្កើតរចនាសម្ព័ន្ធទីបីត្រូវបានលេងដោយអន្តរកម្ម hydrophilic-hydrophobic ។ នៅក្នុងដំណោះស្រាយ aqueous រ៉ាឌីកាល់ hydrophobic មានទំនោរលាក់ខ្លួនពីទឹក ដោយដាក់ជាក្រុមនៅក្នុង globule ខណៈពេលដែលរ៉ាឌីកាល់ hydrophilic ដែលជាលទ្ធផលនៃជាតិទឹក (អន្តរកម្មជាមួយ dipoles ទឹក) មានទំនោរលេចឡើងនៅលើផ្ទៃនៃម៉ូលេគុល។ នៅក្នុងប្រូតេអ៊ីនមួយចំនួនរចនាសម្ព័ន្ធទីបីត្រូវបានធ្វើឱ្យមានស្ថេរភាពដោយចំណង disulfide covalent ដែលបង្កើតឡើងរវាងអាតូមស្ពាន់ធ័រនៃសំណល់ cysteine ពីរ។ នៅកម្រិតរចនាសម្ព័ន្ធទីបី មានអង់ស៊ីម អង្គបដិប្រាណ និងអរម៉ូនមួយចំនួន។

រចនាសម្ព័ន្ធបួនជ្រុងលក្ខណៈនៃប្រូតេអ៊ីនស្មុគស្មាញដែលម៉ូលេគុលត្រូវបានបង្កើតឡើងដោយ globules ពីរឬច្រើន។ ឯកតារងត្រូវបានផ្ទុកនៅក្នុងម៉ូលេគុលដោយអន្តរកម្មអ៊ីយ៉ូដ អ៊ីដ្រូហ្វូប៊ីក និងអេឡិចត្រូស្ទិក។ ជួនកាលក្នុងអំឡុងពេលនៃការបង្កើតរចនាសម្ព័ន្ធ quaternary ចំណង disulfide កើតឡើងរវាងអនុរង។ ប្រូតេអ៊ីនដែលត្រូវបានសិក្សាច្រើនបំផុតដែលមានរចនាសម្ព័ន្ធ quaternary គឺ អេម៉ូក្លូប៊ីន. វាត្រូវបានបង្កើតឡើងដោយ α-subunits ពីរ (សំណល់អាស៊ីតអាមីណូ 141) និង β-subunits ពីរ (សំណល់អាស៊ីតអាមីណូ 146) ។ ភ្ជាប់ជាមួយផ្នែករងនីមួយៗគឺជាម៉ូលេគុល heme ដែលមានជាតិដែក។
ប្រសិនបើហេតុផលមួយចំនួន ការអនុលោមតាមលំហនៃប្រូតេអ៊ីនខុសពីធម្មតា ប្រូតេអ៊ីនមិនអាចបំពេញមុខងាររបស់វាបានទេ។ ឧទាហរណ៍ មូលហេតុនៃ "ជំងឺគោឆ្កួត" (ជម្ងឺរលាកស្រោមខួរ) គឺជាការមិនប្រក្រតីនៃ prions ដែលជាប្រូតេអ៊ីនផ្ទៃនៃកោសិកាប្រសាទ។
លក្ខណៈសម្បត្តិនៃប្រូតេអ៊ីន
សមាសធាតុអាស៊ីតអាមីណូនិងរចនាសម្ព័ន្ធនៃម៉ូលេគុលប្រូតេអ៊ីនកំណត់វា។ លក្ខណៈសម្បត្តិ. ប្រូតេអ៊ីនរួមបញ្ចូលគ្នានូវលក្ខណៈសម្បត្តិជាមូលដ្ឋាន និងអាស៊ីត ដែលកំណត់ដោយរ៉ាឌីកាល់អាស៊ីតអាមីណូ៖ អាស៊ីតអាមីណូដែលមានជាតិអាស៊ីតកាន់តែច្រើននៅក្នុងប្រូតេអ៊ីន លក្ខណៈសម្បត្តិអាស៊ីតរបស់វាកាន់តែច្បាស់។ សមត្ថភាពក្នុងការបរិច្ចាគនិងបន្ថែម H + ត្រូវបានកំណត់ លក្ខណៈសម្បត្តិនៃប្រូតេអ៊ីន; សារធាតុទប់ស្កាត់ដ៏មានឥទ្ធិពលបំផុតមួយគឺ អេម៉ូក្លូប៊ីននៅក្នុងកោសិកាឈាមក្រហម ដែលរក្សា pH ឈាមក្នុងកម្រិតថេរ។ មានប្រូតេអ៊ីនរលាយ (fibrinogen) ហើយមានប្រូតេអ៊ីនមិនរលាយដែលអនុវត្តមុខងារមេកានិច (fibroin, keratin, collagen) ។ មានប្រូតេអ៊ីនដែលមានសកម្មភាពគីមី (អង់ស៊ីម) មានប្រូតេអ៊ីនអសកម្មគីមីដែលមានភាពធន់នឹងលក្ខខណ្ឌបរិស្ថានផ្សេងៗ និងប្រភេទដែលមិនស្ថិតស្ថេរខ្លាំង។
កត្តាខាងក្រៅ (កំដៅ វិទ្យុសកម្មអ៊ុលត្រាវីយូឡេ លោហធាតុធ្ងន់ និងអំបិលរបស់វា ការផ្លាស់ប្តូរ pH វិទ្យុសកម្ម ការខ្សោះជាតិទឹក)
អាចបណ្តាលឱ្យមានការរំខានដល់ការរៀបចំរចនាសម្ព័ន្ធនៃម៉ូលេគុលប្រូតេអ៊ីន។ ដំណើរការនៃការបាត់បង់ការអនុលោមតាមបីវិមាត្រដែលមាននៅក្នុងម៉ូលេគុលប្រូតេអ៊ីនដែលបានផ្តល់ឱ្យត្រូវបានគេហៅថា ការប្រែពណ៌. មូលហេតុនៃការ denaturation គឺការបំបែកចំណងដែលធ្វើអោយមានស្ថេរភាពនៃរចនាសម្ព័ន្ធប្រូតេអ៊ីនជាក់លាក់មួយ។ ដំបូង ចំណងដែលខ្សោយបំផុតត្រូវបានខូច ហើយនៅពេលដែលលក្ខខណ្ឌកាន់តែតឹងរ៉ឹង សូម្បីតែអ្នកដែលខ្លាំងជាងក៏ត្រូវខូចដែរ។ ដូច្នេះដំបូង quaternary បន្ទាប់មករចនាសម្ព័ន្ធទីបីនិងទីពីរត្រូវបានបាត់បង់។ ការផ្លាស់ប្តូរនៅក្នុងការកំណត់រចនាសម្ព័ន្ធលំហនាំទៅរកការផ្លាស់ប្តូរលក្ខណៈសម្បត្តិនៃប្រូតេអ៊ីន ហើយជាលទ្ធផល ធ្វើឱ្យវាមិនអាចសម្រាប់ប្រូតេអ៊ីនដើម្បីបំពេញមុខងារជីវសាស្រ្តដែលមានដើមរបស់វា។ ប្រសិនបើ denaturation មិនត្រូវបានអមដោយការបំផ្លាញរចនាសម្ព័ន្ធបឋមនោះវាអាចជា អាចបញ្ច្រាស់បាន។ក្នុងករណីនេះ ការងើបឡើងវិញដោយខ្លួនឯងនៃលក្ខណៈនៃការអនុលោមតាមប្រូតេអ៊ីនកើតឡើង។ ឧទាហរណ៍ ប្រូតេអ៊ីនទទួលភ្នាសឆ្លងកាត់ការប្រែពណ៌បែបនេះ។ ដំណើរការនៃការស្តាររចនាសម្ព័ន្ធប្រូតេអ៊ីនបន្ទាប់ពីការ denaturation ត្រូវបានគេហៅថា ការកែប្រែឡើងវិញ. ប្រសិនបើការស្ដារឡើងវិញនៃការកំណត់រចនាសម្ព័ន្ធលំហនៃប្រូតេអ៊ីនគឺមិនអាចទៅរួចនោះ denaturation ត្រូវបានគេហៅថា មិនអាចត្រឡប់វិញបាន។.
មុខងារនៃប្រូតេអ៊ីន
| មុខងារ | ឧទាហរណ៍និងការពន្យល់ |
|---|---|
| សំណង់ | ប្រូតេអ៊ីនត្រូវបានចូលរួមនៅក្នុងការបង្កើតរចនាសម្ព័ន្ធកោសិកានិងកោសិកាខាងក្រៅ: ពួកគេជាផ្នែកមួយនៃភ្នាសកោសិកា (lipoproteins, glycoproteins), សក់ (keratin), សរសៃពួរ (collagen) ជាដើម។ |
| ការដឹកជញ្ជូន | អេម៉ូក្លូប៊ីនប្រូតេអ៊ីនក្នុងឈាមភ្ជាប់អុកស៊ីសែន និងដឹកជញ្ជូនវាពីសួតទៅគ្រប់ជាលិកា និងសរីរាង្គ ហើយពីពួកវាផ្ទេរកាបូនឌីអុកស៊ីតទៅសួត។ សមាសភាពនៃភ្នាសកោសិការួមមានប្រូតេអ៊ីនពិសេសដែលធានានូវការផ្ទេរសកម្ម និងជ្រើសរើសយ៉ាងតឹងរ៉ឹងនៃសារធាតុ និងអ៊ីយ៉ុងមួយចំនួនពីកោសិកាទៅកាន់បរិយាកាសខាងក្រៅ និងខាងក្រោយ។ |
| បទប្បញ្ញត្តិ | អរម៉ូនប្រូតេអ៊ីនចូលរួមក្នុងបទប្បញ្ញត្តិនៃដំណើរការមេតាប៉ូលីស។ ឧទាហរណ៍ អ័រម៉ូនអាំងស៊ុយលីនគ្រប់គ្រងកម្រិតជាតិស្ករក្នុងឈាម ជំរុញការសំយោគ glycogen និងបង្កើនការបង្កើតខ្លាញ់ពីកាបូអ៊ីដ្រាត។ |
| ការពារ | ដើម្បីឆ្លើយតបទៅនឹងការជ្រៀតចូលនៃប្រូតេអ៊ីនបរទេសឬអតិសុខុមប្រាណ (អង់ទីហ្សែន) ចូលទៅក្នុងរាងកាយប្រូតេអ៊ីនពិសេសត្រូវបានបង្កើតឡើង - អង្គបដិប្រាណដែលអាចចងនិងបន្សាបពួកវា។ Fibrin ដែលបង្កើតឡើងពីសារធាតុ fibrinogen ជួយបញ្ឈប់ការហូរឈាម។ |
| ម៉ូទ័រ | ប្រូតេអ៊ីន contractile actin និង myosin ផ្តល់នូវការកន្ត្រាក់សាច់ដុំនៅក្នុងសត្វពហុកោសិកា។ |
| សញ្ញា | បង្កើតឡើងនៅក្នុងភ្នាសផ្ទៃនៃកោសិកាគឺជាម៉ូលេគុលប្រូតេអ៊ីនដែលមានសមត្ថភាពផ្លាស់ប្តូររចនាសម្ព័ន្ធទីបីរបស់ពួកគេក្នុងការឆ្លើយតបទៅនឹងកត្តាបរិស្ថាន ដូច្នេះការទទួលសញ្ញាពីបរិយាកាសខាងក្រៅ និងការបញ្ជូនពាក្យបញ្ជាទៅកាន់កោសិកា។ |
| ការផ្ទុក | នៅក្នុងរាងកាយរបស់សត្វ, ប្រូតេអ៊ីន, ជាក្បួន, មិនត្រូវបានរក្សាទុក, ជាមួយនឹងការលើកលែងនៃ albumin ស៊ុតនិង casein ទឹកដោះគោ។ ប៉ុន្តែអរគុណចំពោះប្រូតេអ៊ីនសារធាតុមួយចំនួនអាចត្រូវបានរក្សាទុកនៅក្នុងរាងកាយឧទាហរណ៍ក្នុងអំឡុងពេលបំបែកអេម៉ូក្លូប៊ីនជាតិដែកមិនត្រូវបានយកចេញពីរាងកាយនោះទេប៉ុន្តែត្រូវបានរក្សាទុកបង្កើតស្មុគស្មាញជាមួយប្រូតេអ៊ីន ferritin ។ |
| ថាមពល | នៅពេលដែលប្រូតេអ៊ីន 1 ក្រាមបំបែកទៅជាផលិតផលចុងក្រោយ 17.6 kJ ត្រូវបានបញ្ចេញ។ ដំបូងប្រូតេអ៊ីនបំបែកទៅជាអាស៊ីតអាមីណូហើយបន្ទាប់មកចូលទៅក្នុងផលិតផលចុងក្រោយ - ទឹកកាបូនឌីអុកស៊ីតនិងអាម៉ូញាក់។ ទោះជាយ៉ាងណាក៏ដោយ ប្រូតេអ៊ីនត្រូវបានប្រើជាប្រភពថាមពល លុះត្រាតែប្រភពផ្សេងទៀត (កាបូអ៊ីដ្រាត និងខ្លាញ់) ត្រូវបានប្រើប្រាស់។ |
| កាតាលីករ | មុខងារសំខាន់បំផុតមួយនៃប្រូតេអ៊ីន។ ផ្តល់ដោយប្រូតេអ៊ីន - អង់ស៊ីមដែលពន្លឿនប្រតិកម្មជីវគីមីដែលកើតឡើងនៅក្នុងកោសិកា។ ឧទាហរណ៍ ribulose biphosphate carboxylase ជំរុញការជួសជុល CO 2 កំឡុងពេលធ្វើរស្មីសំយោគ។ |
អង់ស៊ីម
អង់ស៊ីម, ឬ អង់ស៊ីមគឺជាប្រភេទប្រូតេអ៊ីនពិសេស ដែលជាកាតាលីករជីវសាស្រ្ត។ អរគុណចំពោះអង់ស៊ីម ប្រតិកម្មជីវគីមីកើតឡើងក្នុងល្បឿនដ៏អស្ចារ្យ។ ល្បឿននៃប្រតិកម្មអង់ស៊ីមគឺរាប់ម៉ឺនដង (ហើយជួនកាលរាប់លាន) ខ្ពស់ជាងល្បឿននៃប្រតិកម្មដែលកើតឡើងដោយមានការចូលរួមពីកាតាលីករអសរីរាង្គ។ សារធាតុដែលអង់ស៊ីមធ្វើសកម្មភាពត្រូវបានគេហៅថា ស្រទាប់ខាងក្រោម.
អង់ស៊ីមគឺជាប្រូតេអ៊ីនរាងមូល, លក្ខណៈរចនាសម្ព័ន្ធអង់ស៊ីមអាចបែងចែកជាពីរក្រុម៖ សាមញ្ញ និងស្មុគស្មាញ។ អង់ស៊ីមសាមញ្ញគឺជាប្រូតេអ៊ីនសាមញ្ញ, i.e. មានតែអាស៊ីតអាមីណូប៉ុណ្ណោះ។ អង់ស៊ីមស្មុគស្មាញគឺជាប្រូតេអ៊ីនស្មុគស្មាញ i.e. បន្ថែមពីលើផ្នែកប្រូតេអ៊ីន ពួកវាមានក្រុមនៃធម្មជាតិដែលមិនមានប្រូតេអ៊ីន - cofactor. អង់ស៊ីមមួយចំនួនប្រើវីតាមីនជា cofactors ។ ម៉ូលេគុលអង់ស៊ីមមានផ្នែកពិសេសមួយហៅថាមជ្ឈមណ្ឌលសកម្ម។ មជ្ឈមណ្ឌលសកម្ម- ផ្នែកតូចមួយនៃអង់ស៊ីម (ពីបីទៅដប់ពីរសំណល់អាស៊ីតអាមីណូ) ដែលការចងនៃស្រទាប់ខាងក្រោម ឬស្រទាប់ខាងក្រោមកើតឡើងដើម្បីបង្កើតជាស្មុគស្មាញអង់ស៊ីម-ស្រទាប់ខាងក្រោម។ នៅពេលបញ្ចប់ប្រតិកម្ម អង់ស៊ីមស្រទាប់ខាងក្រោមនឹងបំបែកទៅជាអង់ស៊ីម និងផលិតផលប្រតិកម្ម។ អង់ស៊ីមមួយចំនួនមាន (លើកលែងតែសកម្ម) មជ្ឈមណ្ឌល allosteric- តំបន់ដែលនិយតករល្បឿនអង់ស៊ីមត្រូវបានភ្ជាប់ ( អង់ស៊ីម allosteric).

ប្រតិកម្មនៃកាតាលីករអង់ស៊ីមត្រូវបានកំណត់លក្ខណៈដោយ៖ 1) ប្រសិទ្ធភាពខ្ពស់ 2) ការជ្រើសរើសយ៉ាងតឹងរ៉ឹង និងទិសដៅនៃសកម្មភាព 3) ភាពជាក់លាក់នៃស្រទាប់ខាងក្រោម 4) បទប្បញ្ញត្តិល្អ និងច្បាស់លាស់។ ភាពជាក់លាក់នៃស្រទាប់ខាងក្រោម និងប្រតិកម្មនៃប្រតិកម្ម catalysis enzymatic ត្រូវបានពន្យល់ដោយសម្មតិកម្មរបស់ E. Fischer (1890) និង D. Koshland (1959)។
E. Fisher (សម្មតិកម្មចាក់សោសោ)បានស្នើថា ការកំណត់រចនាសម្ព័ន្ធលំហនៃមជ្ឈមណ្ឌលសកម្មនៃអង់ស៊ីម និងស្រទាប់ខាងក្រោមត្រូវតែត្រូវគ្នាយ៉ាងពិតប្រាកដចំពោះគ្នាទៅវិញទៅមក។ ស្រទាប់ខាងក្រោមត្រូវបានប្រៀបធៀបទៅនឹង "កូនសោ" ដែលជាអង់ស៊ីមទៅនឹង "សោ" ។
D. Koshland (សម្មតិកម្មស្រោមដៃ)បានផ្តល់យោបល់ថាការឆ្លើយឆ្លងគ្នារវាងរចនាសម្ព័ន្ធនៃស្រទាប់ខាងក្រោម និងមជ្ឈមណ្ឌលសកម្មនៃអង់ស៊ីមត្រូវបានបង្កើតឡើងតែនៅពេលនៃអន្តរកម្មរបស់ពួកគេជាមួយគ្នាទៅវិញទៅមកប៉ុណ្ណោះ។ សម្មតិកម្មនេះត្រូវបានគេហៅថាផងដែរ។ សម្មតិកម្មការឆ្លើយឆ្លងដែលជំរុញ.
អត្រានៃប្រតិកម្មអង់ស៊ីមអាស្រ័យលើ៖ 1) សីតុណ្ហភាព 2) កំហាប់អង់ស៊ីម 3) កំហាប់ស្រទាប់ខាងក្រោម 4) pH ។ វាគួរតែត្រូវបានសង្កត់ធ្ងន់ថាចាប់តាំងពីអង់ស៊ីមគឺជាប្រូតេអ៊ីនសកម្មភាពរបស់ពួកគេគឺខ្ពស់បំផុតនៅក្រោមលក្ខខណ្ឌធម្មតាខាងសរីរវិទ្យា។
អង់ស៊ីមភាគច្រើនអាចដំណើរការបានតែនៅសីតុណ្ហភាពចន្លោះពី ០ ដល់ ៤០ អង្សាសេ។ នៅក្នុងដែនកំណត់ទាំងនេះ អត្រាប្រតិកម្មកើនឡើងប្រហែល 2 ដងជាមួយនឹងការកើនឡើងសីតុណ្ហភាព 10 អង្សាសេ។ នៅសីតុណ្ហភាពលើសពី 40 អង្សាសេ ប្រូតេអ៊ីនឆ្លងកាត់ការប្រែពណ៌ ហើយសកម្មភាពអង់ស៊ីមថយចុះ។ នៅសីតុណ្ហភាពជិតដល់ត្រជាក់ អង់ស៊ីមត្រូវបានអសកម្ម។
នៅពេលដែលបរិមាណនៃស្រទាប់ខាងក្រោមកើនឡើង អត្រានៃប្រតិកម្មអង់ស៊ីមកើនឡើងរហូតដល់ចំនួនម៉ូលេគុលស្រទាប់ខាងក្រោមស្មើនឹងចំនួនម៉ូលេគុលអង់ស៊ីម។ ជាមួយនឹងការកើនឡើងបន្ថែមទៀតនៅក្នុងបរិមាណនៃស្រទាប់ខាងក្រោមល្បឿននឹងមិនកើនឡើងទេព្រោះមជ្ឈមណ្ឌលសកម្មនៃអង់ស៊ីមត្រូវបានឆ្អែត។ ការកើនឡើងនៃកំហាប់អង់ស៊ីមនាំអោយមានសកម្មភាពកាតាលីករកើនឡើង ចាប់តាំងពីចំនួនម៉ូលេគុលស្រទាប់ខាងក្រោមកាន់តែធំឆ្លងកាត់ការផ្លាស់ប្តូរក្នុងមួយឯកតាពេលវេលា។

សម្រាប់អង់ស៊ីមនីមួយៗមានតម្លៃ pH ដ៏ល្អប្រសើរដែលវាបង្ហាញសកម្មភាពអតិបរមា (pepsin - 2.0, salivary amylase - 6.8, pancreatic lipase - 9.0)។ នៅតម្លៃ pH ខ្ពស់ ឬទាប សកម្មភាពអង់ស៊ីមថយចុះ។ ជាមួយនឹងការផ្លាស់ប្តូរ pH ភ្លាមៗ អង់ស៊ីម denatures ។
ល្បឿននៃអង់ស៊ីម allosteric ត្រូវបានគ្រប់គ្រងដោយសារធាតុដែលភ្ជាប់ទៅនឹងមជ្ឈមណ្ឌល allosteric ។ ប្រសិនបើសារធាតុទាំងនេះបង្កើនល្បឿនប្រតិកម្មនោះគេហៅថា ភ្នាក់ងារសកម្មប្រសិនបើពួកគេបន្ថយល្បឿន - ថ្នាំទប់ស្កាត់.
ការចាត់ថ្នាក់នៃអង់ស៊ីម
ដោយផ្អែកលើប្រភេទនៃការផ្លាស់ប្តូរគីមីដែលពួកគេធ្វើកាតាលីករ អង់ស៊ីមត្រូវបានបែងចែកជា 6 ថ្នាក់៖
- អុកស៊ីដង់តេស(ការផ្ទេរអាតូមអ៊ីដ្រូសែន អុកស៊ីសែន ឬអេឡិចត្រុងពីសារធាតុមួយទៅសារធាតុមួយទៀត - dehydrogenase)
- ការផ្ទេរ(ការផ្ទេរមេទីល អាសុីល ផូស្វាត ឬក្រុមអាមីណូពីសារធាតុមួយទៅសារធាតុមួយទៀត - transaminase)
- អ៊ីដ្រូឡាស(ប្រតិកម្មអ៊ីដ្រូលីស៊ីសដែលផលិតផលពីរត្រូវបានបង្កើតឡើងពីស្រទាប់ខាងក្រោម - អាមីឡាស lipase)
- លីយ៉ាស(ការបន្ថែមមិនមែនអ៊ីដ្រូលីកទៅស្រទាប់ខាងក្រោម ឬការផ្ដាច់នៃក្រុមអាតូមពីវា ក្នុងករណីនេះចំណង C-C, C-N, C-O, C-S អាចត្រូវបានបំបែក - decarboxylase),
- អ៊ីសូមេរ៉ាស(ការរៀបចំឡើងវិញតាមម៉ូលេគុល - isomerase),
- លីហ្គាស(ការតភ្ជាប់នៃម៉ូលេគុលពីរដែលជាលទ្ធផលនៃការបង្កើតចំណង C-C, C-N, C-O, C-S - សំយោគ) ។
ថ្នាក់ត្រូវបានបែងចែកទៅជាថ្នាក់រង និងថ្នាក់រង។ នៅក្នុងការចាត់ថ្នាក់អន្តរជាតិបច្ចុប្បន្ន អង់ស៊ីមនីមួយៗមានលេខកូដជាក់លាក់ ដែលមានបួនលេខដែលបំបែកដោយចំនុច។ លេខទីមួយគឺថ្នាក់ ទីពីរគឺ subclass ទីបីគឺ subsubclass ទីបួនគឺជាលេខស៊េរីនៃអង់ស៊ីមក្នុង subclass នេះ ឧទាហរណ៍ arginase code គឺ 3.5.3.1។
ទៅ ការបង្រៀនលេខ 2"រចនាសម្ព័ន្ធ និងមុខងារនៃកាបូអ៊ីដ្រាត និងខ្លាញ់"
ទៅ ការបង្រៀនលេខ ៤"រចនាសម្ព័ន្ធ និងមុខងារនៃអាស៊ីតនុយក្លេអ៊ីក ATP"
ប្រូតេអ៊ីន (ប្រូតេអ៊ីន) គឺជាសមាសធាតុប៉ូលីមែរម៉ូលេគុលខ្ពស់នៃធម្មជាតិ peptide (អាស៊ីត polyheteroamino) ។
រចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនគឺជាលំដាប់នៃសំណល់អាស៊ីតអាមីណូជំនួសនៅក្នុងខ្សែសង្វាក់ polypeptide (PPC) ។
រចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនគឺ កូវ៉ាឡេនរចនាសម្ព័ន្ធ, ចាប់តាំងពីវាត្រូវបានផ្អែកលើ peptideទំនាក់ទំនងរវាងក្រុម a-amino និង a-carboxyl នៃអាស៊ីតអាមីណូ។ ជាលទ្ធផលខ្សែសង្វាក់ polypeptide មិនមានសាខាទេ។
គ្រោងឆ្អឹង (ឆ្អឹងខ្នង, គ្រោងឆ្អឹង) នៃខ្សែសង្វាក់ polypeptide មានធាតុផ្សំរចនាសម្ព័ន្ធដដែលៗជាប្រចាំ។
ខ្សែសង្វាក់ polypeptide មាន vectoriality ទិសដៅនៃខ្សែសង្វាក់គឺពី N-terminus (ការចាប់ផ្តើមនៃខ្សែសង្វាក់) ទៅ C-terminus (ចុងបញ្ចប់នៃខ្សែសង្វាក់) N-terminus គឺជាចុងបញ្ចប់ដែលក្រុម a-amino ឥតគិតថ្លៃ។ មានទីតាំងនៅ។ C-terminus គឺជាចុងបញ្ចប់ដែលមានក្រុម a-carboxyl ឥតគិតថ្លៃ។ លំដាប់អាស៊ីតអាមីណូនៃប្រូតេអ៊ីនត្រូវបានកំណត់ដោយចាប់ផ្តើមពី N-terminus ដោយប្រើឈ្មោះអក្សរកាត់អាមីណូបីអក្សរ ឧទាហរណ៍៖ gly-ala-cis-pro ។ ការកំណត់អក្សរមួយសម្រាប់សំណល់អាស៊ីតអាមីណូនៅក្នុងប្រូតេអ៊ីនក៏អាចត្រូវបានប្រើផងដែរ។
N- និង C-termini នៃប្រូតេអ៊ីនអាចត្រូវបានកែប្រែ។ ក្រុមអាមីណូនៅ N-terminus អាចត្រូវបាន acetylated, formylated ឬ methylated ។ នៅក្នុងប្រូតេអ៊ីនមួយចំនួនសំណល់ N-terminal គឺជា pyrrolidone carbonate (pyroglutamate) ដែលមិនមានក្រុមអាមីណូឥតគិតថ្លៃ។ C-terminus អាចនឹងត្រូវបានរំខាន។ ការកែប្រែស្ថានីយ C គឺកម្រជាងបើប្រៀបធៀបទៅនឹងការកែប្រែស្ថានីយ N ។
មេគុណនៃ polycondensation នៃប្រូតេអ៊ីនប្រែប្រួលក្នុងចន្លោះពី 50 ទៅ 2500 ។ ជាធម្មតា ប្រូតេអ៊ីនមួយមានសំណល់អាស៊ីតអាមីណូពី 100 ទៅ 300 ។ ដោយសារទម្ងន់ម៉ូលេគុលជាមធ្យមនៃសំណល់អាស៊ីតអាមីណូតែមួយគឺប្រហែល 110 Da ទម្ងន់ម៉ូលេគុលនៃប្រូតេអ៊ីនប្រែប្រួលពី 6000 ទៅរាប់លាន Da ។
ប្រូតេអ៊ីននីមួយៗមានរចនាសម្ព័ន្ធបឋមតែមួយគត់។ ប្រូតេអ៊ីនដំបូងដែលរចនាសម្ព័ន្ធចម្បងត្រូវបានបង្កើតឡើងគឺអាំងស៊ុយលីន។ Sanger បានធ្វើដូច្នេះ។ យុទ្ធសាស្ត្ររបស់គាត់មានដូចខាងក្រោម។ ដំបូងគាត់បានបំបែកខ្សែសង្វាក់ polypeptide ពីរហើយបន្ទាប់មកអនុវត្តការបំបែកអង់ស៊ីមជាក់លាក់របស់ពួកគេទៅជា peptides តូចៗដែលមានលំដាប់ត្រួតស៊ីគ្នា។ បន្ទាប់មក សំណល់ស្ថានីយ N ត្រូវបានគេកំណត់អត្តសញ្ញាណដោយប្រើ 1-fluoro-2,4-dinitrobenzene ។ លើសពីនេះទៀតគាត់បានកំណត់សមាសធាតុអាស៊ីតអាមីណូនៃ peptides ហើយនៅទីបំផុតអាចកំណត់រចនាសម្ព័ន្ធរបស់ពួកគេដោយប្រៀបធៀបលំដាប់នៃ peptides ត្រួតស៊ីគ្នា។ ជាទូទៅយុទ្ធសាស្រ្តរបស់ Sanger បានរក្សាសារៈសំខាន់របស់វារហូតមកដល់សព្វថ្ងៃនេះ។ ទោះយ៉ាងណាក៏ដោយ វិធីសាស្រ្តផ្សេងទៀតក៏ត្រូវបានស្នើឡើងផងដែរ។ Edman បានបង្កើតវិធីសាស្រ្តសម្រាប់នីតិវិធីស្វ័យប្រវត្តិសម្រាប់ការបំបែកជាលំដាប់ និងការកំណត់អត្តសញ្ញាណសំណល់អាស៊ីតអាមីណូស្ថានីយ N ។ ការវិភាគការសាយភាយកាំរស្មីអ៊ិចអាចត្រូវបានប្រើដើម្បីឌិគ្រីបរចនាសម្ព័ន្ធបឋម។ លំដាប់នៃសំណល់អាស៊ីតអាមីណូអាចត្រូវបានកំណត់ពីលំដាប់នុយក្លេអូទីតនៃសារ RNA ។
បច្ចុប្បន្ននេះរចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនជាង 2000 ត្រូវបានបង្កើតឡើង។ តាមទ្រឹស្ដីចំនួននៃវ៉ារ្យ៉ង់ផ្សេងគ្នានៃរចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនគឺគ្មានដែនកំណត់។ សូម្បីតែសម្រាប់ polypeptide នៃអាស៊ីតអាមីណូ 20 ផ្សេងគ្នា ចំនួននៃលំដាប់ដែលអាចមានគឺ 20'10 18 ។ នៅក្នុងធម្មជាតិមានជីវិត មានតែប្រភាគតូចមួយនៃលំដាប់ដែលអាចធ្វើបានប៉ុណ្ណោះដែលត្រូវបានដឹង ចំនួនសរុបដែលនៅក្នុងគ្រប់ប្រភេទនៃសារពាង្គកាយមានជីវិតត្រូវបានប៉ាន់ប្រមាណនៅ 10 10 -10 12 ។
រចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនត្រូវបានកំណត់ហ្សែន, i.e. លំដាប់នៃអាស៊ីតអាមីណូនៅក្នុងប្រូតេអ៊ីនត្រូវបានកំណត់ដោយលំដាប់នៃនុយក្លេអូទីតនៅក្នុង ឌីអិនអេ. ការបង្ខូចទ្រង់ទ្រាយនៃ DNA nucleotide នាំទៅដល់ការលេចចេញនូវប្រូតេអ៊ីនមិនធម្មតាជាមួយនឹងលក្ខណៈសម្បត្តិជីវសាស្ត្រដែលបានផ្លាស់ប្តូរ ដែលជាមូលហេតុនៃរោគវិទ្យាម៉ូលេគុល។ ជាពិសេស ភាពស្លេកស្លាំងនៃកោសិកាឈឺគឺបណ្តាលមកពីការផ្លាស់ប្តូរចំណុចនៅក្នុងហ្សែនដែលគ្រប់គ្រងខ្សែសង្វាក់អេម៉ូក្លូប៊ីនប៊ី។ ផលវិបាកនៃការនេះគឺការជំនួសសំណល់ glutamate នៅក្នុងទីតាំងទី 6 នៃ b-chain ជាមួយ valine ។ ការជំនួសនេះនាំឱ្យបាត់បង់បន្ទុកអវិជ្ជមានមួយនៅក្នុងខ្សែសង្វាក់ b នីមួយៗដែលនាំឱ្យមានការផ្លាស់ប្តូរនៃការអនុលោមតាមអេម៉ូក្លូប៊ីន និងការបាត់បង់មុខងារជីវសាស្រ្តរបស់វា។
ប្រូតេអ៊ីន homologous គឺជាប្រូតេអ៊ីនដែលបំពេញមុខងារដូចគ្នានៅក្នុងប្រភេទផ្សេងៗគ្នា។ ឧទាហរណ៍មួយគឺអេម៉ូក្លូប៊ីន៖ នៅក្នុងសត្វឆ្អឹងកងទាំងអស់វាដំណើរការមុខងារដូចគ្នាដែលទាក់ទងនឹងការដឹកជញ្ជូនអុកស៊ីសែន។ ប្រូតេអ៊ីន homologous ត្រូវបានកំណត់លក្ខណៈដោយវត្តមាននៃអាស៊ីតអាមីណូដូចគ្នានៅក្នុងមុខតំណែងជាច្រើន។ ដូចដែលវាបានប្រែក្លាយចំនួននៃសំណល់អាស៊ីតអាមីណូដែលប្រូតេអ៊ីន homologous ខុសគ្នាគឺសមាមាត្រទៅនឹងភាពខុសគ្នាខាងសរីរវិទ្យារវាងប្រភេទសត្វទាំងនេះ។ ឧទាហរណ៍ ម៉ូលេគុល cytochrome C នៃសេះ និងដំបែមានភាពខុសគ្នានៅក្នុងសំណល់អាស៊ីតអាមីណូចំនួន 48 ខណៈដែលម៉ូលេគុលដូចគ្នានៃសាច់មាន់ និងទាខុសគ្នាតែ 2 សំណល់ប៉ុណ្ណោះ។ ចំពោះ cytochromes C ពីសាច់មាន់ និងទួរគី ពួកវាមានលំដាប់អាស៊ីតអាមីណូដូចគ្នាបេះបិទ។ ព័ត៌មានអំពីចំនួនភាពខុសគ្នានៃលំដាប់អាស៊ីតអាមីណូនៃប្រូតេអ៊ីនដូចគ្នាពីប្រភេទផ្សេងៗគ្នា ត្រូវបានប្រើដើម្បីបង្កើតផែនទីវិវត្តន៍ដែលឆ្លុះបញ្ចាំងពីដំណាក់កាលបន្តបន្ទាប់នៃការកើត និងការអភិវឌ្ឍនៃប្រភេទសត្វ និងរុក្ខជាតិផ្សេងៗនៅក្នុងដំណើរការនៃការវិវត្តន៍។
ធម្មទេសនា ៣. រចនាសម្ព័ន្ធប្រូតេអ៊ីន
និយមន័យ៖
ប្រូតេអ៊ីនគឺជាប៉ូលីម៊ែរមិនទៀងទាត់ដែលមានម៉ូណូមឺរ អិល- អាស៊ីតអាមីណូ។
អាស៊ីតអាមីណូ
ស្តេរ៉េអូអ៊ីសូមឺរមានពីរទម្រង់នៅក្នុងធម្មជាតិ៖អិល (ដៃឆ្វេង) និងឃ (dextrorotatory) ។ ក្រៅពីនេះ។អិល -អាស៊ីតអាមីណូដែលបង្កើតជាប្រូតេអ៊ីនមាននៅក្នុងខ្លួនឃ អាស៊ីតអាមីណូដែលមិនត្រូវបានរួមបញ្ចូលនៅក្នុងប្រូតេអ៊ីន។
រូបមន្តទូទៅនៃអាស៊ីតអាមីណូត្រូវបានបង្ហាញក្នុងរូប។
វាជាការពិតសម្រាប់អាស៊ីដអាមីណូ 19 ក្នុងចំណោម 20 ដែលមាននៅក្នុងប្រូតេអ៊ីន។ បន្ថែមពីលើអាស៊ីតអាមីណូទាំង 19 នេះ ប្រូតេអ៊ីនមានផ្ទុកសារធាតុមួយ។ អាស៊ីត imino - ប្រូលីន
អាស៊ីតអាមីណូទាំងអស់មាន α - ក្រុមអាមីណូ។ ដូច្នេះឈ្មោះ - "អាស៊ីតអាមីណូ" ។ នៅក្នុងប្រូលីន - α- អ៊ីមីណូក្រុម។
ការចាត់ថ្នាក់នៃអាស៊ីតអាមីណូដែលបង្កើតជាប្រូតេអ៊ីនតាមគោលការណ៍ប៉ូល (មិនប៉ូល) នៃរ៉ាឌីកាល់។
1. រ៉ាឌីកាល់មិនប៉ូឡា ឬអ៊ីដ្រូហ្វូប៊ីក។
អាលីហ្វាទិច - alanine, valine, leucine, isoleucine ។មានផ្ទុកស្ពាន់ធ័រ methionineក្រអូប - phenylalanine, tryptophan ។អាស៊ីតអ៊ីមីណូ ប្រូលីន
2. ប៉ូលតែរ៉ាឌីកាល់ដែលមិនមានការចោទប្រកាន់។ គ្លីសេរីន។
អាស៊ីត Hydroxyamino - សេរីន, ថូរនីន, ទីរ៉ូស៊ីន។មានក្រុម sulfhydryl ស៊ីស្ទីន។មានក្រុមអាមីដ៖ asparagine, glutamine ។
3. រ៉ាឌីកាល់ចោទប្រកាន់អវិជ្ជមាន។ អាស៊ីត aspartic អាស៊ីត glutamic ។
4. រ៉ាឌីកាល់គិតជាវិជ្ជមាន។ Lysine, arginine, histidine ។
រចនាសម្ព័ន្ធប្រូតេអ៊ីនបឋម
និយមន័យ៖
រចនាសម្ព័ន្ធចម្បងនៃប្រូតេអ៊ីនគឺជាលំដាប់នៃសំណល់អាស៊ីតអាមីណូនៅក្នុងខ្សែសង្វាក់ polypeptide ។
អាស៊ីតអាមីណូត្រូវបានភ្ជាប់គ្នាដើម្បីបង្កើតជា polypeptide ដោយប្រើចំណង covalent (amide) ។
tripeptide ផ្សំឡើងពីអាស៊ីដអាមីណូបីផ្សេងគ្នា មានលទ្ធភាព 3! = 6 រចនាសម្ព័ន្ធបឋមផ្សេងគ្នា។
សារធាតុ oligopeptide ដែលមានអាស៊ីដអាមីណូចំនួន 20 ផ្សេងគ្នាមានភាពចម្រុះនៃរចនាសម្ព័ន្ធបឋមនៃ 20! ដែលមានន័យថា 2x10 18 ។
ភាពចម្រុះនៃរចនាសម្ព័ន្ធបឋមនៃប្រូតេអ៊ីនដែលមានទំហំមធ្យម (ប្រហែល 500 អាស៊ីតអាមីណូ) មានប្រហែល 20,500 វ៉ារ្យ៉ង់រួចហើយ (ប្រសិនបើអាស៊ីតអាមីណូទាំងអស់ត្រូវបានបង្ហាញក្នុងសមាមាត្រស្មើគ្នា) ។
បើកមិនទាន់មាន មិនមែនទេ ហើយនឹងមិនមានមនុស្សពីរនាក់នៅលើផែនដីដែលមានសំណុំប្រូតេអ៊ីនដូចគ្នានោះទេ។
រចនាសម្ព័ន្ធទីពីរនៃប្រូតេអ៊ីន
និយមន័យ៖
រចនាសម្ព័ន្ធបន្ទាប់បន្សំនៃប្រូតេអ៊ីនគឺជារចនាសម្ព័ន្ធលំដាប់នៃខ្សែសង្វាក់ polypeptide ដែលបណ្តាលមកពីចំណងអ៊ីដ្រូសែនរវាង C = O និង ន- ហអាស៊ីតអាមីណូផ្សេងៗគ្នា។
រចនាសម្ព័ន្ធបន្ទាប់បន្សំអាចទៀងទាត់ (α-helix) ឬមិនទៀងទាត់ (រចនាសម្ព័ន្ធβ-សន្លឹក) ។ នៅក្នុង α-helix NH ក្រុម n សំណល់អាស៊ីតអាមីណូទី 1 ធ្វើអន្តរកម្មជាមួយក្រុម C = O នៃសំណល់អាស៊ីតអាមីណូទី (n-4) ។ មានសំណល់អាស៊ីតអាមីណូ 3.6 ក្នុងមួយវេននៃ β-helix ដែលមានអង្កត់ផ្ចិត 10.1 Å។ រយៈពេលអត្តសញ្ញាណនៃ α-helix ធម្មតាគឺ 18 អាមីណូអាស៊ីត (5 វេន) ។ ការរំខាននៃ α-helix ធម្មតាគឺជាចម្បង ប្រូលីន. ឥទ្ធិពលសំខាន់ទីពីរគឺត្រូវបានបញ្ចេញដោយរ៉ាឌីកាល់ដែលមានបន្ទុកស្មើគ្នាដែលមានទីតាំងនៅក្បែរនោះ។
β-folds អាចត្រូវបានបង្កើតឡើងមិនត្រឹមតែដោយតែមួយប៉ុណ្ណោះទេប៉ុន្តែថែមទាំងដោយសារធាតុ polypeptides ដែលនៅជាប់គ្នាដែលជាផ្នែកមួយនៃប្រូតេអ៊ីនមួយ។
អាល់ហ្វាធម្មជាតិសុទ្ធ ឬបេតា - ប្រូតេអ៊ីនមិនមានទេ។
រចនាសម្ព័ន្ធទីបីនៃប្រូតេអ៊ីន
និយមន័យ
រចនាសម្ព័ន្ធទីបីនៃប្រូតេអ៊ីនគឺជាការអនុលោមតាមលំហនៃសារធាតុ polypeptide ដែលមានរចនាសម្ព័ន្ធបន្ទាប់បន្សំ និងត្រូវបានកំណត់ដោយអន្តរកម្មរវាងរ៉ាឌីកាល់។
មានអន្តរកម្មបួនប្រភេទរវាងរ៉ាឌីកាល់។
ប្រភេទនៃអន្តរកម្មរវាងរ៉ាឌីកាល់
1 . កូវ៉ាឡេន ការទំនាក់ទំនងរវាង សល់ ពីរ cysteine (disulfide ស្ពាន)។
2. អន្តរកម្មអ៊ីយ៉ុង (អេឡិចត្រូស្ទិច) រវាងសំណល់អាស៊ីតអាមីណូដែលមានបន្ទុកផ្ទុយគ្នា។ (រ៉ាឌីកាល់បីដែលមានសញ្ញា "+" និងពីរដែលមានសញ្ញា "-") ។
ឧទាហរណ៍ ε-amino ក្រុម lysine ដែលត្រូវបានចោទប្រកាន់ជាវិជ្ជមាន (- NH ៣ +) ត្រូវបានទាក់ទាញដោយក្រុម carboxyl ដែលត្រូវបានចោទប្រកាន់អវិជ្ជមាន - (COO-) glutamic ឬអាស៊ីត aspartic ។
3. ចំណងអ៊ីដ្រូសែន។
អាស៊ីតអាមីណូទាំងអស់ដែលមានក្រុម hydroxyl, amide ឬ carboxyl ត្រូវបានចូលរួម។
4. អន្តរកម្ម hydrophobic . បង្កើតឡើងរវាងរ៉ាឌីកាល់មិនរាងប៉ូលក្នុងបរិយាកាសទឹក។ អាស៊ីតអាមីណូចំនួន 8 ត្រូវបានចូលរួម (ថ្នាក់ដំបូង) ។
រចនាសម្ព័ន្ធទីបីនៃប្រូតេអ៊ីនត្រូវបានកំណត់ទាំងស្រុងដោយរចនាសម្ព័ន្ធចម្បងរបស់វាពោលគឺឧ។ លំដាប់នៃអាស៊ីតអាមីណូដែលនៅក្នុងវេនត្រូវបានកំណត់ដោយកូដហ្សែន។
អន្តរកម្ម Hydrophobic គឺសម្រេចចិត្តដោយសារតែការមិនជ្រើសរើសរបស់ពួកគេ (មិនជាក់លាក់) និងពហុគុណ។
ប្រូតេអ៊ីនភាគច្រើនមានស្នូល hydrophobic ។
រចនាសម្ព័ន្ធប្រូតេអ៊ីន Quaternary
និយមន័យ៖ រចនាសម្ព័ន្ធ quaternary នៃប្រូតេអ៊ីនគឺជាការប្រមូលផ្តុំនៃខ្សែសង្វាក់ polypeptide ពីរឬច្រើនដែលមានរចនាសម្ព័ន្ធទីបីទៅជាសមាសធាតុសំខាន់ oligomeric ។
ចំណងដែលបង្កើត និងរក្សារចនាសម្ព័ន្ធ quaternary គឺដូចគ្នាទៅនឹងរចនាសម្ព័ន្ធទីបី លើកលែងតែ hydrophobic មួយ។
ន ចុងបញ្ចប់នៃខ្សែសង្វាក់បេតាមានអាស៊ីត glutamic ប៉ូល ("-" គិតថ្លៃ) ។ ចំពោះអ្នកជំងឺដែលមានជំងឺស្លេកស្លាំងក្នុងកោសិកា វ៉ាលីនដែលមិនមានប៉ូឡាត្រូវបានប្រើជំនួសវិញ។ក្នុងចំណោមអាស៊ីតអាមីណូ 574 មាន 2 ត្រូវបានជំនួស។
អេម៉ូក្លូប៊ីនបែបនេះបាត់បង់ការរលាយ និងទម្រង់ទឹកភ្លៀង ដែលធ្វើអោយកោសិកាឈាមក្រហមខូចទ្រង់ទ្រាយ។
ភាពស្លេកស្លាំង Sickle cell គឺជាជំងឺហ្សែន។ ហេតុផលគឺការជំនួសនុយក្លេអូទីតតែមួយនៅក្នុងហ្សែនដែលអ៊ិនកូដខ្សែសង្វាក់អេម៉ូក្លូប៊ីនប៊ី។ កុមារដែលជា homozygotes recessive សម្រាប់ allele នេះមិនរស់នៅរហូតដល់ 2 ឆ្នាំទេ។ Heterozygotes មាន 85% ធម្មតា និង 15% កោសិកាឈាមក្រហម។ Homozygotes លេចធ្លោទទួលរងពីជំងឺគ្រុនចាញ់ heterozygotes មិនទទួលរងពីជំងឺគ្រុនចាញ់ទេ។
ប្រូតេអ៊ីន Globular និង fibrillar
95% នៃប្រូតេអ៊ីនមានស្នូល hydrophobic ។ ប្រូតេអ៊ីន fibrillar 5% ។
ភាគច្រើននៃប្រូតេអ៊ីន globular គឺរលាយ។ fibrillar ភាគច្រើនមិនរលាយ (α-keratins - ពួកវាមានទំងន់ស្ទើរតែស្ងួតទាំងមូលនៃសក់រោមចៀមស្នែងរោមក្រចកជញ្ជីងរោម; collagen - ប្រូតេអ៊ីននៃសរសៃពួរឆ្អឹងខ្ចី; fibroin - ប្រូតេអ៊ីនសូត្រ) ។
ប្រូតេអ៊ីន Fibrillar មានសមាមាត្រធំនៃអាស៊ីដអាមីណូដែលត្រូវបានចោទប្រកាន់ច្រើនជាងសារធាតុ globular - ខ្សែសង្វាក់បុគ្គលគឺអាចរលាយបាន ហើយស្មុគស្មាញរបស់ពួកគេគឺមិនប៉ូលនិងមិនរលាយ។
