Primārā struktūra - aminoskābju secība polipeptīdu ķēdē. Olbaltumvielu molekulā, mainot stingru (peptīdu saiti) un elastīgu (α-oglekļa atoms) posmus, veidojas kompakts ķēdes izvietojums telpā.
Akobori metode ir izmantot fenilhidrazīns. Fenilhidrazīns sarauj peptīdu saites proteīnos un saistās ar visām aminoskābēm, izņemot C-galu. Turpmākā hromatogrāfiskā analīze ļauj atpazīt C-gala aminoskābi proteīnā.
Svarīga ir primārās struktūras izpēte vispārējā bioloģiskā un medicīniskā nozīme:
- primārā struktūra nosaka turpmākās olbaltumvielu struktūras.
- proteīna primārās struktūras zināšanas ir nepieciešamas proteīnu mākslīgai sintēzei.
- primārā struktūra nosaka sugas specifiku, piemēram, insulīna proteīnā, parasti dažādās dzīvnieku sugās un cilvēkos molekulas vidū, parasti notiek 3 ekvivalentu aminoskābju aizstāšana.
- primārās struktūras izmaiņas var izraisīt daudzas slimības, piemēram, sirpjveida šūnu anēmiju, kurā hemoglobīnā β ķēdē 6. pozīcijā glutamīnskābi aizstāj ar valīnu. Šī aizstāšana ar nevienlīdzīgu aminoskābi izraisa hemoglobīna darbības traucējumus un sirpjveida sarkano asins šūnu parādīšanos.
Sekundārā struktūra - regulāri atkārtojas polipeptīdu ķēdes izvietojuma modelis telpā. Visbiežāk olbaltumvielās ir sastopami 2 veidu sekundārās struktūras: α - spirāle un β - struktūra.
α - spirāle 1951. gadā to pētīja L. Paulings, izmantojot rentgenstaru difrakcijas metodi. Tā ir labās puses spirālveida struktūra, kuras vienā pagriezienā iederas 3,6 aminoskābes. Spirāles solis (attālums starp blakus esošajiem pagriezieniem) ir 0,54 n.m. α-spirāle ir fiksēta ar ūdeņraža saitēm, kuras ir noslēgtas starp peptīdu saitēm, kuras veido katra 4. aminoskābe. Sekundārā α - struktūra spontāni salokās, un to nosaka proteīna primārā struktūra. Spirālveida struktūrā sakārtoto reģionu īpatsvars dažādos proteīnos atšķiras. Piemēram, hemoglobīnā un mioglobīnā dominē α - strukturālā kroka, kas samazina proteīna molekulas izmēru 4 reizes.
β-struktūra ir “akordeona” forma, un to stabilizē ūdeņraža saites starp vienas polipeptīdu ķēdes attālām sekcijām vai starp vairākām olbaltumvielu molekulām. Ir paralēlas β struktūras, kurās N un C gali atbilst viens otram, un antiparalēlas struktūras. Olbaltumvielu, kas galvenokārt satur β struktūras, piemērs ir imūnglobulīni.
Sekundārā struktūra tiek pētīta ar rentgenstaru difrakcijas analīzi un pētot ultravioleto staru absorbciju olbaltumvielās (jo lielāks α - struktūru īpatsvars, jo lielāka absorbcija).
Sekundārā struktūra tiek iznīcināta denaturācijas laikā.
Terciārā struktūra - ar polipeptīdu ķēdes telpiskā izkārtojuma forma, kas raksturīga katram proteīnam. Šī struktūra veidojas spontāni, un to nosaka primārā struktūra. Terciārā struktūra ievērojami palielina proteīna kompaktumu par desmitiem. Terciārās struktūras veidošanā ir iesaistītas nekovalentās saites (hidrofobās, jonu) un kovalentās (disulfīda) saites.
Terciārā struktūra nosaka olbaltumvielu bioloģisko aktivitāti un fizikāli ķīmiskās īpašības. Ja tiek izjaukta terciārā struktūra, proteīns zaudē savu bioloģisko aktivitāti.
Terciārās struktūras izpētes metodes ir rentgenstaru difrakcijas analīze un atsevišķu aminoskābju radikāļu ķīmiskās aktivitātes noteikšana proteīnā. Pirmo reizi mioglobīna proteīna terciāro struktūru pētīja J. Kendrew (1957). M. Perutz (1959) pētīja hemoglobīna struktūru.
Olbaltumvielu terciārajā struktūrā ietilpst α - spirālveida, β - salocītas struktūras, β - cilpas (kurās polipeptīdu ķēde ir saliekta par 180 0) un tā sauktā nesakārtotā spole. Piemēram, insulīna proteīns satur 57% α - spirālveida apgabalus, 6% β - salocītu struktūru, 10% molekulas ir sakārtotas β - cilpu veidā un 27% molekulas ir nesakārtota spole.
Primārā, sekundārā, terciārā kopums ir uzbūve proteīna molekula. Mūža (dzimtā) konformācija veidojas spontāni, un tās veidošanos sauc par locīšanu. Olbaltumvielu konformācija ir ļoti nestabila un veidojas, piedaloties īpašiem proteīniem - pavadoņi(pavadoņi). Šaperoni spēj saistīties ar daļēji denaturētiem proteīniem, kas atrodas nestabilā stāvoklī, un atjauno to dabisko konformāciju. Šaperonus klasificē pēc molekulmasas (60 - 100 cd.). Visvairāk pētīti ir Sh-60, Sh-70 un Sh-90. Piemēram, Sh-70 mijiedarbojas ar proteīniem, kas bagāti ar hidrofobiem radikāļiem, un aizsargā tos no augstas temperatūras denaturācijas. Kopumā chaperoni aizsargā galvenās ķermeņa olbaltumvielas, novērš denaturāciju un veicina konformācijas veidošanos, atvieglo denaturēto proteīnu transportēšanu lizosomās un piedalās proteīnu sintēzes procesā.
Pēc uzbūves visas olbaltumvielas iedala trīs grupās:
- fibrilārie proteīni: kolagēns, elastīns, fibroīns.
- Globulārie proteīni: hemoglobīns, albumīns, globulīns.
- Jauktie proteīni: miozīns.
Terciārā struktūra ir raksturīga visiem proteīniem. Tikai oligomēriem proteīniem, kas satur vairākas apakšvienības, protomērus, ir kvartāra struktūra. Protomērs tiek uzskatīts par atsevišķu polipeptīdu ķēdi, apakšvienība ir oligomēra proteīna funkcionāli aktīvā daļa. Apakšvienība var saturēt vienu protomēru vai vairākus.
Kvartāra struktūra - apakšvienību skaits un relatīvais izvietojums oligomēru proteīnos. Tikai oligomēriem proteīniem, kas satur vairākas apakšvienības, protomērus, ir kvartāra struktūra. Protomērs tiek uzskatīts par atsevišķu polipeptīdu ķēdi, apakšvienība ir oligomēra proteīna funkcionāli aktīvā daļa. Apakšvienība var saturēt vienu protomēru vai vairākus protomērus.
Kvartārās struktūras veidošanās ietver vājas nekovalentās saites (hidrofobās, jonu, ūdeņraža saites). Olbaltumvielu kvartārā struktūra veidojas spontāni un viegli sadalās denaturējot. Atsevišķas oligomēra proteīna apakšvienības mijiedarbojas viena ar otru, kas izraisa izmaiņas atsevišķu protomēru terciārajā struktūrā. Šo parādību sauc par kooperatīvām izmaiņām protomēra konformācijā, un to parasti pavada olbaltumvielu aktivitātes palielināšanās.
Oligomēriem proteīniem ir vairākas iezīmes salīdzinājumā ar monomēru proteīniem.
- Tiem ir ļoti kompakts iepakojums un salīdzinoši maza saskarnes virsma, tāpēc, atrodoties intracelulāri, tie piesaista mazāk ūdens
- To darbība tiek regulēta organismā. Protomēri parasti ir neaktīvi, bet oligomērie proteīni ir daudz aktīvāki.
- Ja viena veida protomēri ir iesaistīti oligomēra proteīna sintēzē, tas ietaupa ģenētisko materiālu (vairāki identiski protomēri tiek “apzīmogoti” uz īsu DNS daļu)
- Tie ir funkcionāli vairāk pielāgoti ķermeņa apstākļiem.
Oligomēru proteīnu funkcionalitāti ilustrē, salīdzinot proteīnus hemoglobīnu un mioglobīnu, kas ir iesaistīti skābekļa transportēšanā uz audiem. Eritrocītu hemoglobīns ir oligomērs proteīns, kas ietver 4 polipeptīdu ķēdes. Muskuļu mioglobīns ir monomērs proteīns, kas ietver 1 polipeptīda ķēdi. Mioglobīna skābekļa piesātinājuma līkne norāda uz tā tiešu atkarību no skābekļa koncentrācijas. Hemoglobīnam skābekļa piesātinājuma līkne ir S-veida. Tas ir saistīts ar pakāpeniskām secīgām izmaiņām katra no 4 hemoglobīna sastāvā esošā protomēra struktūrā (konformācijā), kā rezultātā krasi palielinās hemoglobīna afinitāte pret skābekli. Šis hemoglobīna piesātinājuma raksturs ar skābekli krasi palielina tā skābekļa ietilpību salīdzinājumā ar mioglobīnu.
Īpašu vietu olbaltumvielu vidū ieņem domēna proteīni .
Domēni ir strukturāli un funkcionāli atsevišķas vienas polipeptīdu ķēdes sadaļas. Domēni var būt atbildīgi par proteīna mijiedarbību ar dažādām vielām - ligandiem (mazmolekulārām vielām, DNS, RNS, polisaharīdiem utt. Domēna proteīnu piemēri ir seruma albumīns, imūnglobulīni un daži enzīmi (aizkuņģa dziedzera tripsīns).
Pateicoties proteīnu augstajai selektivitātei, tos var apvienot kompleksos, kurus visbiežāk sauc par multienzīmu kompleksiem – tās ir vairāku enzīmu strukturālas asociācijas, kas katalizē sarežģītā ķīmiskā procesa atsevišķus posmus. Piemērs: piruvāta dehidrogenāzes komplekss (PDC), trīs veidu enzīmu komplekss, kas katalizē pirovīnskābes (PVA) oksidāciju.
Ir iespējams specifiski kombinēt ne tikai atsevišķus proteīnus, bet arī olbaltumvielas ar lipīdiem (taukiem) šūnu membrānu veidošanās laikā un olbaltumvielas ar nukleīnskābēm hromatīna veidošanās laikā.
Olbaltumvielu fizikāli ķīmiskās īpašības.
Tos lielā mērā nosaka proteīna molekulas konformācija (olbaltumvielas primārā – terciārā struktūra). Olbaltumvielu fizikāli ķīmiskās īpašības parādās šķīdumos.
Šķīdība olbaltumvielas atšķiras atkarībā no olbaltumvielām.
Kopumā olbaltumvielu šķīdība ir augsta, taču tā atšķiras starp dažādiem proteīnu veidiem. To ietekmē šādi faktori:
- proteīna molekulas forma (globulārie proteīni ir labāk šķīstoši nekā fibrilārie proteīni)
- proteīna aminoskābju radikāļu raksturs, polāro nepolāro radikāļu attiecība (jo vairāk polāro hidrofilo radikāļu proteīnā, jo labāka tā šķīdība)
- šķīdinātāja īpašības, sāļu klātbūtne. Zema sāļu koncentrācija (KCL, NaCl) dažkārt palielina olbaltumvielu šķīdību. Piemēram, albumīni labāk šķīst tīrā destilētā ūdenī, globulīnus izšķīdina tikai 10% sāļu (KCL, NaCl) klātbūtnē. Saistaudu proteīni kolagēns un elastīns nešķīst ne ūdenī, ne sāls šķīdumos.
Molekulārā masa olbaltumvielas ir diezgan lielas, svārstās no 6 000 līdz 1 000 000, piemēram, hemoglobīna molekulmasa ir 68 000, albumīna ir 100 000, ribonukleāze ir aptuveni 14 000, miozīna molekulmasa ir 500 000.
Olbaltumvielu molārās masas noteikšanas metodes jābūt maigam un nedrīkst iznīcināt olbaltumvielu molekulas. Piemēram, ebulioskopiskā metode, kuras pamatā ir šķīdumu viršanas temperatūras mērīšana, nav piemērojama olbaltumvielām. Visprecīzākās metodes proteīnu molekulmasas noteikšanai ir ultracentrifugēšanas metode un rentgenstaru difrakcijas metode.
Ultracentrifugēšanas metode(sedimentācija) ir balstīta uz dažādu molekulmasu olbaltumvielu sedimentācijas ātruma izmaiņām, rotējot olbaltumvielu šķīdumus lielā ātrumā. Ar šo metodi atrasto olbaltumvielu molekulmasu apzīmē ar Svedberga vienību (S = 10 -13 c.)
Rentgenstaru difrakcijas metodeļauj aprēķināt molekulmasu, analizējot vairākus proteīna molekulas rentgena attēlus.
Elektroforēzes metode balstās uz olbaltumvielu kustības ātruma atkarību nemainīgā elektriskajā laukā no proteīna molekulmasas (elektroforētiskā mobilitāte ir lielāka olbaltumvielām ar mazāku molekulmasu)
Hromatogrāfijas metode ir balstīta uz dažādu proteīnu atšķirīgo pārvietošanās ātrumu caur molekulāro gēla "sietiem".
Lielas molekulas, kas ir lielākas par gela poru izmēru, iziet cauri gēlam ātrāk nekā mazākas proteīna molekulas, kas saglabājas gēla graudos.
Elektronu mikroskopiskā metode veic, salīdzinot proteīna molekulas izmēru ar zināmas masas references paraugiem.
Ķīmiskās metodes saistīta ar olbaltumvielu ķīmiskā sastāva īpašībām
Olbaltumvielu molekulu forma savādāk. Olbaltumvielu molekulas var būt fibrilāras vai lodveida formas. Fibrilārajiem proteīniem ir pavedienam līdzīga molekulāra forma. Tie parasti nešķīst ūdenī un atšķaidītos sāls šķīdumos. Fibrilārajos proteīnos ietilpst galvenie saistaudu strukturālie proteīni: kolagēns, keratīns, elastīns. Globulārajos proteīnos polipeptīdu ķēdes ir cieši savītas kompaktās sfēriskās struktūrās. Lielākā daļa lodveida proteīnu labi šķīst ūdenī un vājos sāls šķīdumos. Globulārie proteīni ietver fermentus, antivielas, albumīnus un hemoglobīnu. Dažiem proteīniem ir starpposma molekulas, kas satur gan pavedieniem līdzīgus, gan sfēriskus reģionus. Šādu proteīnu piemērs ir muskuļu proteīns miozīns, kas šķīst sāls šķīdumos.
Olbaltumvielu molekulu izmēri ir diapazonā no 1 līdz 100 nm, tuvu koloidālo daļiņu izmēriem. Šī iemesla dēļ olbaltumvielu šķīdumiem ir gan patieso šķīdumu, gan koloidālo šķīdumu īpašības.
Daudzas olbaltumvielu šķīdumu molekulārās kinētiskās īpašības ir līdzīgas koloidālo šķīdumu īpašības .
- Lēns olbaltumvielu difūzijas ātrums, kas nepieciešams to apmaiņai.
- Olbaltumvielu nespēja iziet cauri puscaurlaidīgām membrānām. Nodalījumos ar augstu olbaltumvielu koncentrāciju rodas pārmērīgs hidrostatiskais spiediens, jo ūdens molekulas virzās vienvirziena cauri puscaurlaidīgajai membrānai uz augstu proteīna koncentrāciju. Olbaltumvielu radīto pārmērīgo spiedienu sauc par onkotisko spiedienu. Tas ir svarīgs faktors, kas nosaka ūdens kustību starp audiem, asinīm un zarnām.
- Olbaltumvielu augstā viskozitāte ir saistīta ar dažādu lielu olbaltumvielu molekulu starpmolekulāro mijiedarbību. Paaugstināta asins viskozitāte, jo īpaši, palielina slodzi uz sirds muskuli.
- Daži proteīni spēj veidot želejas, kas palielina olbaltumvielu (piemēram, kolagēna) izturību.
Olbaltumvielu optiskās īpašības nosaka olbaltumvielu molekulu lielums, aminoskābju radikāļu struktūra olbaltumvielās, peptīdu saišu un alfa-spirālveida reģionu klātbūtne olbaltumvielās.
- Olbaltumvielu šķīdumiem ir gaismas laušanas (refrakcijas) un gaismas izkliedes ietekme.Šīs īpašības ir saistītas ar proteīnu molekulu lielo izmēru, kas ir samērojams ar spektra redzamās daļas viļņa garumu. Šajā gadījumā īsie zilie stari ir izkliedēti lielākā mērā nekā garāki sarkanie stari. Refrakcijas pakāpe ir proporcionāla proteīna šķīduma koncentrācijai.
- Olbaltumvielu šķīdumi absorbē ultravioletos starus diapazonā no 190-230 nm peptīdu saišu klātbūtnes dēļ un diapazonā no 260-280 nm sakarā ar ciklisko aminoskābju klātbūtni olbaltumvielās. UV absorbcijas pakāpe ir proporcionāla olbaltumvielu koncentrācijai šķīdumā.
- Olbaltumvielu šķīdumi var pagriezt polarizētās gaismas plakni, kas ir saistīts ar proteīnā esošo aminoskābju optisko aktivitāti un alfa-spirālveida sekciju klātbūtni tajā. Pastāv tieša saikne starp gaismas polarizāciju un olbaltumvielu koncentrāciju šķīdumā.
Olbaltumvielām, kas ir molekulāri šķīdumi, ir patieso risinājumu īpašības . Tā kā olbaltumvielu šķīdumi ir patiesi risinājumi, tie ir ļoti stabili.
1. MODULIS PROTEĪNU STRUKTŪRA, ĪPAŠĪBAS UN FUNKCIJAS
1. MODULIS PROTEĪNU STRUKTŪRA, ĪPAŠĪBAS UN FUNKCIJAS
Moduļa struktūra | Tēmas |
Moduļu vienība 1 | 1.1. Olbaltumvielu strukturālā organizācija. Olbaltumvielu dabiskās konformācijas veidošanās posmi 1.2. Olbaltumvielu funkcionēšanas pamati. Zāles kā ligandi, kas ietekmē olbaltumvielu darbību 1.3. Olbaltumvielu denaturācija un to spontānas renaturācijas iespēja |
Moduļu vienība 2 | 1.4. Oligomēru proteīnu struktūras un darbības iezīmes, izmantojot hemoglobīna piemēru 1.5. Dabiskās olbaltumvielu konformācijas uzturēšana šūnu apstākļos 1.6. Olbaltumvielu daudzveidība. Olbaltumvielu ģimenes, izmantojot imūnglobulīnu piemēru 1.7. Olbaltumvielu fizikāli ķīmiskās īpašības un to atdalīšanas metodes |
Moduļu vienība 1 MONOMERISKO PROTEĪNU STRUKTURĀLĀ ORGANIZĀCIJA UN TO FUNKCIJAS PAMATI
Mācību mērķi Spēt:
1. Izmantot zināšanas par proteīnu strukturālajām iezīmēm un proteīnu funkciju atkarību no to struktūras, lai izprastu iedzimtu un iegūto proteinopātiju attīstības mehānismus.
2. Izskaidrot dažu medikamentu kā ligandu terapeitiskās iedarbības mehānismus, kas mijiedarbojas ar olbaltumvielām un maina to aktivitāti.
3. Izmantot zināšanas par proteīnu struktūru un konformācijas labilitāti, lai izprastu to strukturālo un funkcionālo nestabilitāti un tendenci uz denaturāciju mainīgos apstākļos.
4. Izskaidrot denaturējošo līdzekļu izmantošanu kā medicīnisko materiālu un instrumentu sterilizācijas līdzekli, kā arī antiseptiskus līdzekļus.
Zināt:
1. Olbaltumvielu strukturālās organizācijas līmeņi.
2. Olbaltumvielu primārās struktūras nozīme, kas nosaka to strukturālo un funkcionālo daudzveidību.
3. Olbaltumvielu aktīvā centra veidošanās mehānisms un tā specifiskā mijiedarbība ar ligandu, kas ir proteīnu funkcionēšanas pamatā.
4. Eksogēnu ligandu (zāles, toksīni, indes) ietekmes uz proteīnu uzbūvi un funkcionālo aktivitāti piemēri.
5. Olbaltumvielu denaturācijas cēloņi un sekas, denaturāciju izraisošie faktori.
6. Denaturējošo faktoru izmantošanas piemēri medicīnā kā antiseptiķi un līdzekļi medicīnas instrumentu sterilizācijai.
TĒMA 1.1. PROTEĪNU STRUKTURĀLĀ ORGANIZĀCIJA. DZIMTĀS VEIDOJAS POSMI
PROTEĪNU KONFORMĀCIJAS
Olbaltumvielas ir polimēru molekulas, kuru monomēros ir tikai 20 α-aminoskābes. Aminoskābju kombināciju komplektu un secību proteīnā nosaka gēnu struktūra indivīdu DNS. Katrs proteīns atbilstoši savai specifiskajai struktūrai veic savu funkciju. Dotā organisma olbaltumvielu kopums nosaka tā fenotipiskās īpašības, kā arī iedzimtu slimību klātbūtni vai noslieci uz to attīstību.
1. Aminoskābes, kas veido olbaltumvielas. Peptīdu saite. Olbaltumvielas ir polimēri, kas veidoti no monomēriem - 20 α-aminoskābēm, kuru vispārējā formula ir
Aminoskābes atšķiras pēc α-oglekļa atomam piesaistīto radikāļu struktūras, lieluma un fizikāli ķīmiskajām īpašībām. Aminoskābju funkcionālās grupas nosaka dažādu α-aminoskābju īpašību raksturlielumus. α-aminoskābēs atrodamos radikāļus var iedalīt vairākās grupās:
prolīns, Atšķirībā no pārējiem 19 proteīna monomēriem, tā nav aminoskābe, bet gan iminoskābe, kas ir saistīta gan ar α-oglekļa atomu, gan imino grupu
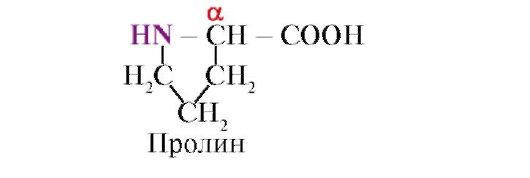 Aminoskābēm ir atšķirīga šķīdība ūdenī. Tas ir saistīts ar radikāļu spēju mijiedarboties ar ūdeni (hidrātu).
Aminoskābēm ir atšķirīga šķīdība ūdenī. Tas ir saistīts ar radikāļu spēju mijiedarboties ar ūdeni (hidrātu).
UZ hidrofils ietver radikāļus, kas satur anjonu, katjonu un polāru neuzlādētas funkcionālās grupas.
UZ hidrofobs ietver radikāļus, kas satur metilgrupas, alifātiskās ķēdes vai gredzenus.
2. Peptīdu saites savieno aminoskābes, veidojot peptīdus. Peptīdu sintēzes laikā vienas aminoskābes α-karboksilgrupa mijiedarbojas ar citas aminoskābes α-aminogrupu, veidojot peptīdu saite:
Olbaltumvielas ir polipeptīdi, t.i. α-aminoskābju lineāri polimēri, kas savienoti ar peptīdu saiti (1.1. att.)
 Rīsi. 1.1. Termini, ko izmanto, lai aprakstītu peptīdu struktūru
Rīsi. 1.1. Termini, ko izmanto, lai aprakstītu peptīdu struktūru
Tiek saukti aminoskābju monomēri, kas veido polipeptīdus aminoskābju atlikumi. Atkārtotu grupu ķēde - NH-CH-CO- veidlapas peptīdu mugurkauls. Aminoskābes atlikumu, kurā ir brīva α-aminogrupa, sauc par N-galu, un to, kurā ir brīva α-karboksilgrupa, sauc par C-galu. Peptīdi tiek rakstīti un lasīti no N-gala līdz C-galam.
Peptīdu saite, ko veido prolīna imino grupa, atšķiras no citām peptīdu saitēm: peptīdu grupas slāpekļa atomam trūkst ūdeņraža,
tā vietā ir saite ar radikāli, kā rezultātā viena gredzena puse tiek iekļauta peptīda mugurkaulā:
 Peptīdi atšķiras pēc aminoskābju sastāva, aminoskābju skaita un aminoskābju savienojuma secības, piemēram, Ser-Ala-Glu-Gis un His-Glu-Ala-Ser ir divi dažādi peptīdi.
Peptīdi atšķiras pēc aminoskābju sastāva, aminoskābju skaita un aminoskābju savienojuma secības, piemēram, Ser-Ala-Glu-Gis un His-Glu-Ala-Ser ir divi dažādi peptīdi.
Peptīdu saites ir ļoti spēcīgas, un to ķīmiskai neenzimātiskajai hidrolīzei ir nepieciešami skarbi apstākļi: analizējamo proteīnu 24 stundas hidrolizē koncentrētā sālsskābē aptuveni 110° temperatūrā. Dzīvā šūnā peptīdu saites var tikt pārtrauktas proteolītiskie enzīmi, sauca proteāzes vai peptīdu hidrolāzes.
3. Olbaltumvielu primārā struktūra. Aminoskābju atlikumi dažādu proteīnu peptīdu ķēdēs nemainās nejauši, bet ir sakārtoti noteiktā secībā. Tiek saukta aminoskābju atlikumu lineārā secība vai pārmaiņu secība polipeptīdu ķēdē proteīna primārā struktūra.
Katra atsevišķa proteīna primārā struktūra ir kodēta DNS molekulā (reģionā, ko sauc par gēnu) un tiek realizēta transkripcijas (informācijas kopēšana uz mRNS) un translācijas (proteīna primārās struktūras sintēzes) laikā. Līdz ar to atsevišķa cilvēka proteīnu primārā struktūra ir no vecākiem uz bērniem iedzimta pārmantotā informācija, kas nosaka konkrētā organisma proteīnu strukturālās īpatnības, no kurām atkarīga esošo proteīnu darbība (1.2. att.).
 Rīsi. 1.2. Attiecība starp genotipu un indivīda organismā sintezēto olbaltumvielu konformāciju
Rīsi. 1.2. Attiecība starp genotipu un indivīda organismā sintezēto olbaltumvielu konformāciju
Katrs no aptuveni 100 000 atsevišķo olbaltumvielu cilvēka organismā ir unikāla primārā struktūra. Viena un tā paša veida olbaltumvielu (piemēram, albumīna) molekulām ir tāda pati aminoskābju atlikumu maiņa, kas atšķir albumīnu no jebkura cita atsevišķa proteīna.
Aminoskābju atlikumu secību peptīdu ķēdē var uzskatīt par informācijas reģistrēšanas veidu. Šī informācija nosaka lineārās peptīdu ķēdes telpisko izvietojumu kompaktākā trīsdimensiju struktūrā, ko sauc uzbūve vāvere. Tiek saukts funkcionāli aktīva proteīna konformācijas veidošanās process locīšana
4. Olbaltumvielu konformācija. Brīva rotācija peptīdu mugurkaulā ir iespējama starp peptīdu grupas slāpekļa atomu un blakus esošo α-oglekļa atomu, kā arī starp α-oglekļa atomu un karbonilgrupas oglekli. Aminoskābju atlikumu funkcionālo grupu mijiedarbības dēļ proteīnu primārā struktūra var iegūt sarežģītākas telpiskās struktūras. Globulārajos proteīnos ir divi galvenie peptīdu ķēžu konformācijas locīšanas līmeņi: sekundārais Un terciārā struktūra.
Olbaltumvielu sekundārā struktūra ir telpiska struktūra, kas veidojas ūdeņraža saišu veidošanās rezultātā starp peptīda mugurkaula funkcionālajām grupām -C=O un -NH-. Šajā gadījumā peptīdu ķēde var iegūt regulāras divu veidu struktūras: α-spirāles Un β-struktūras.
IN α-spirāles veidojas ūdeņraža saites starp karbonilgrupas skābekļa atomu un no tās 4. aminoskābes amīda slāpekļa ūdeņradi; aminoskābju atlikumu sānu ķēdes
atrodas gar spirāles perifēriju, nepiedaloties sekundārās struktūras veidošanā (1.3. att.).
Lielapjoma radikāļi jeb radikāļi ar vienādiem lādiņiem novērš α-spirāles veidošanos. Prolīna atlikums, kuram ir gredzena struktūra, pārtrauc α-spirāli, jo ūdeņraža trūkuma dēļ peptīdu ķēdes slāpekļa atomā nav iespējams izveidot ūdeņraža saiti. Saite starp slāpekli un α-oglekļa atomu ir daļa no prolīna gredzena, tāpēc peptīda mugurkauls šajā vietā kļūst saliekts.
β-struktūra veidojas starp vienas polipeptīdu ķēdes peptīdu mugurkaula lineārajiem reģioniem, tādējādi veidojot salocītas struktūras. Var veidoties polipeptīdu ķēdes vai to daļas paralēli vai antiparalēlas β-struktūras. Pirmajā gadījumā mijiedarbojošo peptīdu ķēžu N- un C-gals sakrīt, bet otrajā tiem ir pretējs virziens (1.4. att.).
Rīsi. 1.3. Olbaltumvielu sekundārā struktūra - α-spirāle
 Rīsi. 1.4. Paralēlas un antiparalēlas β-loksnes struktūras
Rīsi. 1.4. Paralēlas un antiparalēlas β-loksnes struktūras
β-struktūras ir apzīmētas ar platām bultiņām: A - Antiparalēla β-struktūra. B - Paralēlas β-loksnes struktūras
Dažos proteīnos β-struktūras var veidoties ūdeņraža saišu veidošanās dēļ starp dažādu polipeptīdu ķēžu peptīdu mugurkaula atomiem.
Atrodas arī olbaltumvielās apgabali ar neregulāru sekundāru struktūra, kas ietver polipeptīda mugurkaula līkumus, cilpas un pagriezienus. Tie bieži atrodas vietās, kur mainās peptīdu ķēdes virziens, piemēram, kad veidojas paralēla β-loksnes struktūra.
Pamatojoties uz α-spirāļu un β-struktūru klātbūtni, globulāros proteīnus var iedalīt četrās kategorijās.
Rīsi. 1.5. Mioglobīna (A) un hemoglobīna β-ķēdes (B) sekundārā struktūra, kas satur astoņas α-spirāles

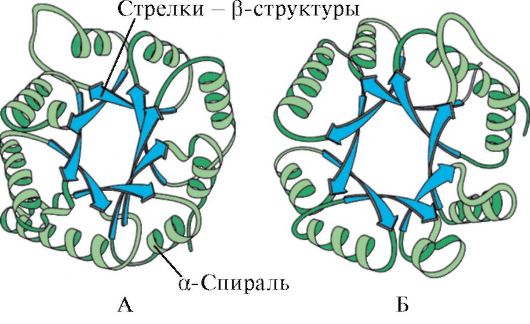 Rīsi. 1.6. Triosefosfāta izomerāzes un piruvāta kināzes domēna sekundārā struktūra
Rīsi. 1.6. Triosefosfāta izomerāzes un piruvāta kināzes domēna sekundārā struktūra
 Rīsi. 1.7. Imūnglobulīna (A) un enzīma superoksīda dismutāzes (B) konstantā domēna sekundārā struktūra
Rīsi. 1.7. Imūnglobulīna (A) un enzīma superoksīda dismutāzes (B) konstantā domēna sekundārā struktūra
IN ceturtā kategorija ietvēra proteīnus, kas satur nelielu daudzumu regulāru sekundāro struktūru. Šie proteīni ietver mazus ar cisteīnu bagātus proteīnus vai metaloproteīnus.
Olbaltumvielu terciārā struktūra- konformācijas veids, kas veidojas mijiedarbības rezultātā starp aminoskābju radikāļiem, kas peptīdu ķēdē var atrasties ievērojamā attālumā viens no otra. Lielākā daļa olbaltumvielu veido telpisku struktūru, kas atgādina lodītes (globulārie proteīni).
Tā kā hidrofobajiem aminoskābju radikāļiem ir tendence apvienoties caur t.s hidrofobās mijiedarbības un starpmolekulāriem van der Vālsa spēkiem, proteīna lodītes iekšpusē veidojas blīvs hidrofobs kodols. Hidrofilie jonizētie un nejonizētie radikāļi galvenokārt atrodas uz proteīna virsmas un nosaka tā šķīdību ūdenī.
 Rīsi. 1.8. Saišu veidi, kas rodas starp aminoskābju radikāļiem proteīna terciārās struktūras veidošanās laikā
Rīsi. 1.8. Saišu veidi, kas rodas starp aminoskābju radikāļiem proteīna terciārās struktūras veidošanās laikā
1 - jonu saite- notiek starp pozitīvi un negatīvi lādētām funkcionālajām grupām;
2 - ūdeņraža saite- notiek starp hidrofilu neuzlādētu grupu un jebkuru citu hidrofilu grupu;
3 - hidrofobās mijiedarbības- rodas starp hidrofobiem radikāļiem;
4 - disulfīda saite- veidojas cisteīna atlikumu SH grupu oksidācijas un to savstarpējās mijiedarbības dēļ
Hidrofilās aminoskābju atliekas, kas atrodas hidrofobajā kodolā, var mijiedarboties savā starpā, izmantojot jonu Un ūdeņraža saites(1.8. att.).
Jonu un ūdeņraža saites, kā arī hidrofobās mijiedarbības ir vājas: to enerģija nav daudz lielāka par molekulu termiskās kustības enerģiju istabas temperatūrā. Proteīna konformācija tiek uzturēta, veidojot daudzas šādas vājas saites. Tā kā atomi, kas veido proteīnu, atrodas pastāvīgā kustībā, ir iespējams pārraut dažas vājās saites un veidot citas, kas izraisa nelielu atsevišķu polipeptīdu ķēdes posmu kustību. Šo olbaltumvielu īpašību mainīt konformāciju dažu vāju saišu pārraušanas un citu vāju saišu veidošanās rezultātā sauc konformācijas labilitāte.
Cilvēka ķermenī ir sistēmas, kas atbalsta homeostāze- iekšējās vides noturība noteiktās veselam organismam pieņemamās robežās. Homeostāzes apstākļos nelielas konformācijas izmaiņas neizjauc proteīnu kopējo struktūru un darbību. Par proteīna funkcionāli aktīvo konformāciju sauc dzimtā uzbūve. Izmaiņas iekšējā vidē (piemēram, glikozes, Ca jonu, protonu uc koncentrācija) izraisa izmaiņas konformācijā un olbaltumvielu funkciju traucējumus.
Dažu proteīnu terciārā struktūra ir stabilizēta disulfīda saites, veidojas divu atlikumu -SH grupu mijiedarbības dēļ
 Rīsi. 1.9. Disulfīda saites veidošanās proteīna molekulā
Rīsi. 1.9. Disulfīda saites veidošanās proteīna molekulā
cisteīns (1.9. att.). Lielākajai daļai intracelulāro proteīnu terciārajā struktūrā nav kovalento disulfīda saišu. To klātbūtne ir raksturīga šūnas izdalītajām olbaltumvielām, kas nodrošina to lielāku stabilitāti ārpusšūnu apstākļos. Tādējādi disulfīda saites atrodas insulīna un imūnglobulīnu molekulās.
Insulīns- proteīna hormons, kas sintezēts aizkuņģa dziedzera β-šūnās un izdalīts asinīs, reaģējot uz glikozes koncentrācijas palielināšanos asinīs. Insulīna struktūrā ir divas disulfīda saites, kas savieno polipeptīda A un B ķēdes, un viena disulfīdsaite A ķēdes ietvaros (1.10. att.).
 Rīsi. 1.10. Disulfīda saites insulīna struktūrā
Rīsi. 1.10. Disulfīda saites insulīna struktūrā
5. Olbaltumvielu virssekundārā struktūra. Proteīnās ar atšķirīgu primāro struktūru un funkcijām tie dažkārt tiek atklāti līdzīgas kombinācijas un sekundāro struktūru relatīvās pozīcijas, kuras sauc par virssekundāro struktūru. Tas ieņem starpposmu starp sekundāro un terciāro struktūru, jo tā ir specifiska sekundārās struktūras elementu kombinācija proteīna terciārās struktūras veidošanā. Virssekundārajām struktūrām ir specifiski nosaukumi, piemēram, “α-spirāle-pagrieziens-a-spirāle”, “leicīna rāvējslēdzējs”, “cinka pirksti” utt. Šādas virssekundārās struktūras ir raksturīgas DNS saistošiem proteīniem.
"Leicīna rāvējslēdzējs."Šāda veida supersekundārā struktūra tiek izmantota, lai savienotu kopā divus proteīnus. Mijiedarbojošo olbaltumvielu virsmā ir α-spirāles reģioni, kas satur vismaz četras leicīna atliekas. Leicīna atlikumi α-spirālē atrodas sešu aminoskābju attālumā viens no otra. Tā kā katrs α-spirāles pagrieziens satur 3,6 aminoskābju atlikumus, leicīna radikāļi atrodas uz katra otrā pagrieziena virsmas. Viena proteīna α-spirāles leicīna atliekas var mijiedarboties ar cita proteīna leicīna atliekām (hidrofobās mijiedarbības), savienojot tās kopā (1.11. att.). Daudzi DNS saistošie proteīni darbojas oligomēru kompleksos, kur atsevišķas apakšvienības ir savstarpēji saistītas ar "leicīna rāvējslēdzējiem".
 Rīsi. 1.11. "Leicīna rāvējslēdzējs" starp divu proteīnu α-spirālveida reģioniem
Rīsi. 1.11. "Leicīna rāvējslēdzējs" starp divu proteīnu α-spirālveida reģioniem
Šādu proteīnu piemērs ir histoni. Histoni- kodolproteīni, kas satur lielu skaitu pozitīvi lādētu aminoskābju - arginīnu un lizīnu (līdz 80%). Histona molekulas tiek apvienotas oligomēros kompleksos, kas satur astoņus monomērus, izmantojot "leicīna rāvējslēdzējus", neskatoties uz šo molekulu ievērojamo homonīmu lādiņu.
"Cinka pirksts"- DNS saistošiem proteīniem raksturīgs supersekundārās struktūras variants, uz proteīna virsmas ir iegarena fragmenta forma un satur apmēram 20 aminoskābju atlikumus (1.12. att.). “Izstieptā pirksta” formu atbalsta cinka atoms, kas saistīts ar četriem aminoskābju radikāļiem – diviem cisteīna atlikumiem un diviem histidīna atlikumiem. Dažos gadījumos histidīna atlikumu vietā ir cisteīna atliekas. Divi cieši guļoši cisteīna atlikumi ir atdalīti no pārējiem diviem Gisili atlikumiem ar Cys secību, kas sastāv no aptuveni 12 aminoskābju atlikumiem. Šis proteīna reģions veido α-spirāli, kuras radikāļi var specifiski saistīties ar DNS galvenās rievas regulējošajiem reģioniem. Individuāla saistīšanās specifika
 Rīsi. 1.12. DNS saistošo proteīnu reģiona primārā struktūra, kas veido "cinka pirksta" struktūru (burti norāda aminoskābes, kas veido šo struktūru)
Rīsi. 1.12. DNS saistošo proteīnu reģiona primārā struktūra, kas veido "cinka pirksta" struktūru (burti norāda aminoskābes, kas veido šo struktūru)
Regulējošais DNS saistošais proteīns ir atkarīgs no aminoskābju atlikumu secības, kas atrodas cinka pirksta reģionā. Šādas struktūras jo īpaši satur steroīdu hormonu receptorus, kas iesaistīti transkripcijas regulēšanā (informācijas nolasīšana no DNS uz RNS).
TĒMA 1.2. PROTEĪNU DARBĪBAS PAMATI. NARKOTIKAS KĀ LIGANDAS, KAS IETEKMĒ PROTEĪNA FUNKCIJU
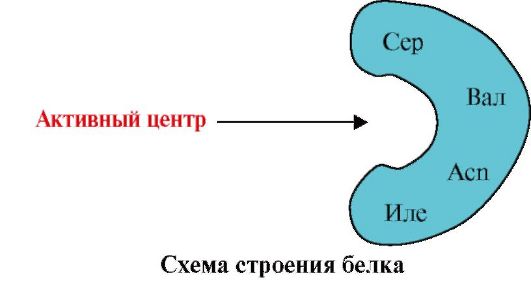
1. Olbaltumvielas aktīvais centrs un tā mijiedarbība ar ligandu. Terciārās struktūras veidošanās laikā uz funkcionāli aktīva proteīna virsmas, parasti padziļinājumā, veidojas reģions, ko veido aminoskābju radikāļi, kas primārajā struktūrā atrodas tālu viens no otra. Šo reģionu, kuram ir unikāla struktūra noteiktam proteīnam un kas spēj specifiski mijiedarboties ar noteiktu molekulu vai līdzīgu molekulu grupu, sauc par proteīna-ligandu saistīšanas vietu vai aktīvo vietu. Ligandi ir molekulas, kas mijiedarbojas ar olbaltumvielām.
Augsta specifika Olbaltumvielas mijiedarbību ar ligandu nodrošina aktīvā centra struktūras komplementaritāte ar liganda struktūru.
Papildināmība- tā ir mijiedarbojošo virsmu telpiskā un ķīmiskā atbilstība. Aktīvajam centram ne tikai telpiski jāatbilst tajā iekļautajam ligandam, bet arī jāveido saitēm (jonu, ūdeņraža un hidrofobās mijiedarbības) starp aktīvajā centrā iekļauto radikāļu funkcionālajām grupām un ligandu, kas satur ligandu. aktīvajā centrā (1.13. att.).
 Rīsi. 1.13. Papildu proteīna mijiedarbība ar ligandu
Rīsi. 1.13. Papildu proteīna mijiedarbība ar ligandu
Dažiem ligandiem, kas pievienoti proteīna aktīvajam centram, ir papildu loma olbaltumvielu darbībā. Šādus ligandus sauc par kofaktoriem, un proteīnus, kas satur daļu, kas nav olbaltumvielas, sauc kompleksi proteīni(pretstatā vienkāršiem proteīniem, kas sastāv tikai no proteīna daļas). Neolbaltumvielu daļu, kas ir cieši savienota ar proteīnu, sauc protezēšanas grupa. Piemēram, mioglobīns, hemoglobīns un citohromi satur protēžu grupu, hēmu, kas satur dzelzs jonu, kas ir cieši piesaistīts aktīvajam centram. Kompleksos proteīnus, kas satur hēmu, sauc par hemoproteīniem.
Ja olbaltumvielām ir piesaistīti specifiski ligandi, izpaužas šo proteīnu funkcija. Tādējādi albumīns, vissvarīgākais asins plazmas proteīns, demonstrē savu transportēšanas funkciju, piesaistot aktīvajam centram hidrofobus ligandus, piemēram, taukskābes, bilirubīnu, dažas zāles utt. (1.14. att.)
Ligandi, kas mijiedarbojas ar peptīdu ķēdes trīsdimensiju struktūru, var būt ne tikai zemmolekulārās organiskās un neorganiskās molekulas, bet arī makromolekulas:
DNS (piemēri ar DNS saistošiem proteīniem, kas apspriesti iepriekš);
Polisaharīdi;
 Rīsi. 1.14. Saikne starp genotipu un fenotipu
Rīsi. 1.14. Saikne starp genotipu un fenotipu
Unikālā cilvēka proteīnu primārā struktūra, kas kodēta DNS molekulā, šūnās tiek realizēta unikālas uzbūves, aktīvā centra struktūras un proteīna funkciju veidā.
Šādos gadījumos proteīns atpazīst specifisku liganda reģionu, kas ir samērīgs un komplementārs saistīšanās vietai. Tādējādi uz hepatocītu virsmas atrodas hormona insulīna receptoru proteīni, kam ir arī proteīna struktūra. Insulīna mijiedarbība ar receptoru izraisa izmaiņas tā konformācijā un signalizācijas sistēmu aktivāciju, kā rezultātā pēc ēdienreizēm hepatocītos tiek uzglabātas barības vielas.
Tādējādi Olbaltumvielu funkcionēšana balstās uz proteīna aktīvā centra specifisko mijiedarbību ar ligandu.
2. Domēna struktūra un tās nozīme proteīnu funkcionēšanā. Globulāro proteīnu garās polipeptīdu ķēdes bieži salokās vairākos kompaktos, relatīvi neatkarīgos reģionos. Viņiem ir neatkarīga terciārā struktūra, kas atgādina globulāro proteīnu struktūru, un tos sauc domēni. Pateicoties proteīnu domēna struktūrai, to terciārā struktūra ir vieglāk veidojama.
Domēna proteīnos ligandu saistīšanās vietas bieži atrodas starp domēniem. Tādējādi tripsīns ir proteolītisks enzīms, ko ražo aizkuņģa dziedzera eksokrīnā daļa un ir nepieciešams pārtikas olbaltumvielu sagremošanai. Tam ir divu domēnu struktūra, un tripsīna saistīšanās centrs ar tā ligandu - pārtikas proteīnu - atrodas rievā starp abiem domēniem. Aktīvajā centrā tiek radīti apstākļi, kas nepieciešami konkrētas pārtikas proteīna vietas efektīvai saistīšanai un tās peptīdu saišu hidrolīzei.
Dažādi domēni proteīnā var pārvietoties viens pret otru, aktīvajam centram mijiedarbojoties ar ligandu (1.15. att.).
Heksokināze- enzīms, kas katalizē glikozes fosforilāciju, izmantojot ATP. Fermenta aktīvā vieta atrodas spraugā starp diviem domēniem. Heksokināzei saistoties ar glikozi, to aptverošie domēni aizveras un substrāts tiek iesprostots, kur notiek fosforilēšanās (sk. 1.15. att.).
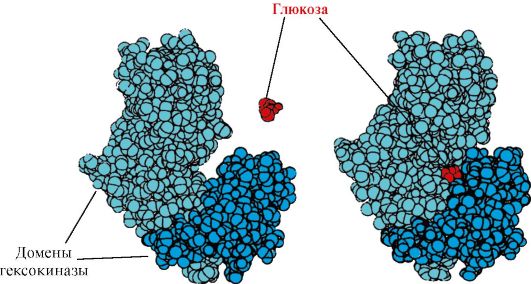 Rīsi. 1.15. Heksokināzes domēnu saistīšanās ar glikozi
Rīsi. 1.15. Heksokināzes domēnu saistīšanās ar glikozi
Dažos proteīnos domēni veic neatkarīgas funkcijas, saistoties ar dažādiem ligandiem. Šādas olbaltumvielas sauc par daudzfunkcionālām.
3. Narkotikas ir ligandi, kas ietekmē olbaltumvielu darbību. Olbaltumvielu mijiedarbība ar ligandiem ir specifiska. Taču proteīna un tā aktīvā centra konformācijas labilitātes dēļ ir iespējams izvēlēties citu vielu, kas varētu arī mijiedarboties ar proteīnu aktīvajā centrā vai citā molekulas daļā.
Vielu, kas pēc struktūras līdzīga dabiskajam ligandam, sauc liganda strukturālais analogs vai nedabisks ligands. Tas arī mijiedarbojas ar proteīnu aktīvajā vietā. Liganda strukturālais analogs var uzlabot proteīna funkciju (agonists), un samaziniet to (antagonists). Ligands un tā strukturālie analogi konkurē viens ar otru par saistīšanos ar proteīnu tajā pašā vietā. Šādas vielas sauc konkurētspējīgi modulatori olbaltumvielu funkciju (regulatori). Daudzas zāles darbojas kā olbaltumvielu inhibitori. Dažus no tiem iegūst, ķīmiski modificējot dabiskos ligandus. Olbaltumvielu funkciju inhibitori var būt zāles un indes.
Atropīns ir konkurējošs M-holīnerģisko receptoru inhibitors. Acetilholīns ir neirotransmiters nervu impulsu pārraidīšanai caur holīnerģiskām sinapsēm. Lai veiktu ierosmi, acetilholīnam, kas izdalās sinaptiskajā spraugā, ir jāsadarbojas ar postsinaptiskās membrānas receptoru proteīnu. Atrasti divi veidi holīnerģiskie receptori:
M receptors papildus acetilholīnam tas selektīvi mijiedarbojas ar muskarīnu (mušmires toksīnu). M - holīnerģiskie receptori atrodas uz gludajiem muskuļiem un, mijiedarbojoties ar acetilholīnu, izraisa to kontrakciju;
H receptorsīpaši saistās ar nikotīnu. N-holīnerģiskie receptori ir atrodami šķērssvītrotu skeleta muskuļu sinapsēs.
Specifisks inhibitors M-holīnerģiskie receptori ir atropīns. Tas ir atrodams belladonna un henbane augos.
 Atropīnam ir acetilholīnam pēc uzbūves un to telpiskā izvietojuma līdzīgas funkcionālās grupas, tāpēc tas ir konkurējošs M-holīnerģisko receptoru inhibitors. Ņemot vērā, ka acetilholīna saistīšanās ar M-holīnerģiskiem receptoriem izraisa gludo muskuļu kontrakciju, atropīnu lieto kā zāles, kas mazina to spazmas. (spazmolītisks līdzeklis). Tādējādi ir zināms, ka atropīnu lieto acu muskuļu atslābināšanai, apskatot dibenu, kā arī spazmu mazināšanai kuņģa-zarnu trakta kolikas laikā. M-holīnerģiskie receptori atrodas arī centrālajā nervu sistēmā (CNS), tāpēc lielas atropīna devas var izraisīt nevēlamu centrālās nervu sistēmas reakciju: motoru un garīgu uzbudinājumu, halucinācijas, krampjus.
Atropīnam ir acetilholīnam pēc uzbūves un to telpiskā izvietojuma līdzīgas funkcionālās grupas, tāpēc tas ir konkurējošs M-holīnerģisko receptoru inhibitors. Ņemot vērā, ka acetilholīna saistīšanās ar M-holīnerģiskiem receptoriem izraisa gludo muskuļu kontrakciju, atropīnu lieto kā zāles, kas mazina to spazmas. (spazmolītisks līdzeklis). Tādējādi ir zināms, ka atropīnu lieto acu muskuļu atslābināšanai, apskatot dibenu, kā arī spazmu mazināšanai kuņģa-zarnu trakta kolikas laikā. M-holīnerģiskie receptori atrodas arī centrālajā nervu sistēmā (CNS), tāpēc lielas atropīna devas var izraisīt nevēlamu centrālās nervu sistēmas reakciju: motoru un garīgu uzbudinājumu, halucinācijas, krampjus.
Ditilīns ir konkurējošs H-holīnerģisko receptoru agonists, kas inhibē neiromuskulāro sinapsu darbību.
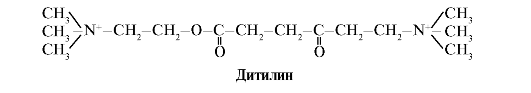 Skeleta muskuļu neiromuskulārās sinapses satur H-holīnerģiskos receptorus. To mijiedarbība ar acetilholīnu izraisa muskuļu kontrakcijas. Dažu ķirurģisku operāciju laikā, kā arī endoskopiskajos pētījumos tiek izmantotas zāles, kas izraisa skeleta muskuļu relaksāciju (muskuļu relaksanti). Tie ietver ditilīnu, kas ir acetilholīna strukturāls analogs. Tas piesaistās H-holīnerģiskiem receptoriem, bet atšķirībā no acetilholīna to ļoti lēni iznīcina enzīms acetilholīnesterāze. Ilgstošas jonu kanālu atvēršanas un pastāvīgas membrānas depolarizācijas rezultātā tiek traucēta nervu impulsu vadīšana un notiek muskuļu relaksācija. Sākotnēji šīs īpašības tika atklātas kurares indē, tāpēc šādas zāles sauc kūrei līdzīgs.
Skeleta muskuļu neiromuskulārās sinapses satur H-holīnerģiskos receptorus. To mijiedarbība ar acetilholīnu izraisa muskuļu kontrakcijas. Dažu ķirurģisku operāciju laikā, kā arī endoskopiskajos pētījumos tiek izmantotas zāles, kas izraisa skeleta muskuļu relaksāciju (muskuļu relaksanti). Tie ietver ditilīnu, kas ir acetilholīna strukturāls analogs. Tas piesaistās H-holīnerģiskiem receptoriem, bet atšķirībā no acetilholīna to ļoti lēni iznīcina enzīms acetilholīnesterāze. Ilgstošas jonu kanālu atvēršanas un pastāvīgas membrānas depolarizācijas rezultātā tiek traucēta nervu impulsu vadīšana un notiek muskuļu relaksācija. Sākotnēji šīs īpašības tika atklātas kurares indē, tāpēc šādas zāles sauc kūrei līdzīgs.
TĒMA 1.3. PROTEĪNU DENATURĀCIJA UN TO SPONTĀNĀS RENATIVĀCIJAS IESPĒJA
1. Tā kā proteīnu dabiskā konformācija tiek saglabāta vājas mijiedarbības dēļ, izmaiņas olbaltumvielu apkārtējās vides sastāvā un īpašībās, ķīmisko reaģentu un fizikālo faktoru iedarbība izraisa izmaiņas to konformācijā (konformācijas labilitātes īpašība). Liela skaita saišu pārraušana noved pie dabiskās konformācijas iznīcināšanas un olbaltumvielu denaturācijas.
Olbaltumvielu denaturācija- tā ir to dabiskās konformācijas iznīcināšana denaturējošu vielu ietekmē, ko izraisa vāju saišu pārrāvums, kas stabilizē proteīna telpisko struktūru. Denaturāciju pavada proteīna unikālās trīsdimensiju struktūras un aktīvā centra iznīcināšana un bioloģiskās aktivitātes zudums (1.16. att.).
Visas viena proteīna denaturētās molekulas iegūst nejaušu konformāciju, kas atšķiras no citām tā paša proteīna molekulām. Aminoskābju radikāļi, kas veido aktīvo centru, izrādās telpiski attālināti viens no otra, t.i. tiek iznīcināta proteīna specifiskā saistīšanās vieta ar ligandu. Denaturācijas laikā olbaltumvielu primārā struktūra paliek nemainīga.
Denaturējošo vielu pielietojums bioloģiskajos pētījumos un medicīnā. Bioķīmiskajos pētījumos pirms zemas molekulmasas savienojumu noteikšanas bioloģiskajā materiālā olbaltumvielas parasti vispirms izņem no šķīduma. Visbiežāk šim nolūkam izmanto trihloretiķskābi (TCA). Pēc TCA pievienošanas šķīdumam denaturētie proteīni izgulsnējas un ir viegli atdalāmi filtrējot (1.1. tabula).
Medicīnā denaturējošus līdzekļus bieži izmanto medicīnisko instrumentu un materiālu sterilizēšanai autoklāvos (denaturējošais līdzeklis ir augsta temperatūra) un kā antiseptiskus līdzekļus (spirts, fenols, hloramīns) piesārņotu virsmu apstrādei, kas satur patogēnu mikrofloru.
2. Spontāna proteīna reaktivācija- proteīnu primārās struktūras, konformācijas un funkcijas determinisma pierādījums. Atsevišķi proteīni ir viena gēna produkti, kuriem ir identiska aminoskābju secība un kas šūnā iegūst tādu pašu konformāciju. Pamatsecinājums, ka proteīna primārajā struktūrā jau ir informācija par tā konformāciju un funkciju, tika izdarīts, pamatojoties uz dažu proteīnu (īpaši ribonukleāzes un mioglobīna) spēju spontāni renatitēties - atjaunot to dabisko konformāciju pēc denaturācijas.
Telpisko proteīnu struktūru veidošanās tiek veikta ar pašsavienošanās metodi - spontānu procesu, kurā polipeptīdu ķēdei, kurai ir unikāla primārā struktūra, ir tendence pieņemt konformāciju ar zemāko brīvo enerģiju šķīdumā. Eksperimentā ar enzīmu ribonukleāzi tika aprakstīta spēja renativēt olbaltumvielas, kas saglabā savu primāro struktūru pēc denaturācijas.
Ribonukleāze ir enzīms, kas sadala saites starp atsevišķiem nukleotīdiem RNS molekulā. Šim lodveida proteīnam ir viena polipeptīda ķēde, kuras terciāro struktūru stabilizē daudzas vājas un četras disulfīda saites.
Ribonukleāzes apstrāde ar urīnvielu, kas sarauj ūdeņraža saites molekulā, un reducētāju, kas pārtrauc disulfīda saites, izraisa fermenta denaturāciju un tā aktivitātes zudumu.
Denaturējošo vielu noņemšana ar dialīzi noved pie proteīna konformācijas un funkcijas atjaunošanas, t.i. uz atdzimšanu. (1.17. att.).
 Rīsi. 1.17. Ribonukleāzes denaturācija un renativācija
Rīsi. 1.17. Ribonukleāzes denaturācija un renativācija
A - ribonukleāzes dabiskā konformācija, kuras terciārajā struktūrā ir četras disulfīda saites; B - denaturēta ribonukleāzes molekula;
B - reaktivēta ribonukleāzes molekula ar atjaunotu struktūru un funkciju
1. Aizpildiet tabulu 1.2.
1.2. tabula. Aminoskābju klasifikācija pēc radikāļu polaritātes
2. Uzrakstiet tetrapeptīda formulu:
Asp — Pro — Fen — Liz
a) izcelt peptīda atkārtojošās grupas, kas veido peptīda mugurkaulu, un mainīgās grupas, ko attēlo aminoskābju radikāļi;
b) marķē N- un C-galus;
c) izcelt peptīdu saites;
d) uzrakstiet citu peptīdu, kas sastāv no tām pašām aminoskābēm;
e) saskaitīt iespējamos tetrapeptīda variantus ar līdzīgu aminoskābju sastāvu.
3. Izskaidrojiet proteīnu primārās struktūras lomu, izmantojot divu strukturāli līdzīgu un evolucionāri tuvu zīdītāju neirohipofīzes peptīdu hormonu - oksitocīna un vazopresīna - salīdzinošās analīzes piemēru (1.3. tabula).
1.3. tabula. Oksitocīna un vazopresīna uzbūve un funkcijas
 Priekš šī:
Priekš šī:
a) salīdzināt divu peptīdu sastāvu un aminoskābju secību;
b) atrast abu peptīdu primārās struktūras līdzību un to bioloģiskās iedarbības līdzību;
c) atrast atšķirības divu peptīdu struktūrā un atšķirības to funkcijās;
d) izdarīt secinājumu par peptīdu primārās struktūras ietekmi uz to funkcijām.
4. Aprakstiet lodveida proteīnu konformācijas veidošanās galvenos posmus (sekundārās, terciārās struktūras, virssekundārās struktūras jēdziens). Norādiet saišu veidus, kas iesaistīti olbaltumvielu struktūru veidošanā. Kuri aminoskābju radikāļi var piedalīties hidrofobās mijiedarbības, jonu, ūdeņraža saišu veidošanā.
Sniedziet piemērus.
5. Definējiet jēdzienu "olbaltumvielu konformācijas labilitāte", norādiet tā pastāvēšanas iemeslus un nozīmi.
6. Paplašiniet šādas frāzes nozīmi: “Olbaltumvielu funkcionēšanas pamatā ir to specifiskā mijiedarbība ar ligandu”, izmantojot terminus un skaidrojot to nozīmi: proteīna konformācija, aktīvais centrs, ligands, komplementaritāte, proteīna funkcija.
7. Izmantojot vienu piemēru, paskaidrojiet, kas ir domēni un kāda ir to loma olbaltumvielu darbībā.
PAŠKONTROLES UZDEVUMI
1. Match.
Funkcionālā grupa aminoskābju radikāļos:
A. Karboksilgrupa B. Hidroksilgrupa C Guanidīna grupa D. Tiola grupa E. Aminogrupa
2. Izvēlieties pareizās atbildes.
Aminoskābes ar polāriem neuzlādētiem radikāļiem ir:
A. Cis B. Asn
B. Glu G. Trīs
3. Izvēlieties pareizās atbildes.
Aminoskābju radikāļi:
A. Nodrošināt primārās struktūras specifiku B. Piedalīties terciārās struktūras veidošanā
B. Atrodas uz proteīna virsmas, tie ietekmē tā šķīdību D. Veido aktīvo centru
D. Piedalīties peptīdu saišu veidošanā
4. Izvēlieties pareizās atbildes.
Starp aminoskābju radikāļiem var veidoties hidrofobiska mijiedarbība:
A. Tre Lay B. Pro Three
B. Met Ile G. Tir Ala D. Val Fen
5. Izvēlieties pareizās atbildes.
Starp aminoskābju radikāļiem var veidoties jonu saites:
A. Gln Asp B. Apr Liz
B. Liza Glu G. Gis Asp D. Asn Apr
6. Izvēlieties pareizās atbildes.
Ūdeņraža saites var veidoties starp aminoskābju radikāļiem:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Match.
Proteīna struktūras veidošanā iesaistītās saites veids:
A. Primārā struktūra B. Sekundārā struktūra
B. Terciārā struktūra
D. Virssekundārā struktūra E. Konformācija.
1. Ūdeņraža saites starp peptīdu mugurkaula atomiem
2. Vājas saites starp aminoskābju radikāļu funkcionālajām grupām
3. Saites starp aminoskābju α-amino un α-karboksilgrupām
8. Izvēlieties pareizās atbildes. Tripsīns:
A. Proteolītiskais enzīms B. Satur divus domēnus
B. Hidrolizē cieti
D. Aktīvā vieta atrodas starp domēniem. D. Sastāv no divām polipeptīdu ķēdēm.
9. Izvēlieties pareizās atbildes. Atropīns:
A. Neirotransmiters
B. Acetilholīna strukturālais analogs
B. Mijiedarbojas ar H-holīnerģiskiem receptoriem
D. Nostiprina nervu impulsu vadīšanu caur holīnerģiskām sinapsēm
D. M-holīnerģisko receptoru konkurētspējīgs inhibitors
10. Izvēlieties pareizos apgalvojumus. Olbaltumvielās:
A. Primārā struktūra satur informāciju par tās aktīvās vietnes struktūru
B. Aktīvais centrs veidojas primārās struktūras līmenī
B. Konformācija ir stingri fiksēta ar kovalentām saitēm
D. Aktīvā vieta var mijiedarboties ar līdzīgu ligandu grupu
proteīnu D konformācijas labilitātes dēļ. Izmaiņas vidē var ietekmēt aktīvās vielas afinitāti
centrs uz ligandu
1. 1-B, 2-G, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
PAMATA NOTEIKUMI UN JĒDZIENI
1. Olbaltumvielas, polipeptīds, aminoskābes
2. Primārās, sekundārās, terciārās olbaltumvielu struktūras
3. Konformācija, native proteīna konformācija
4. Kovalentās un vājās saites olbaltumvielās
5. Konformācijas labilitāte
6. Olbaltumvielu aktīvā vieta
7. Ligands
8. Olbaltumvielu locīšana
9. Ligandu strukturālie analogi
10. Domēna proteīni
11. Vienkāršas un sarežģītas olbaltumvielas
12. Olbaltumvielu denaturēšana, denaturējošie līdzekļi
13. Olbaltumvielu reaktivācija
Atrisināt problēmas
"Olbaltumvielu strukturālā organizācija un to funkcionēšanas pamati"
1. Proteīna – hemoglobīna A (HbA) galvenā funkcija ir skābekļa transportēšana uz audiem. Cilvēku populācijā ir zināmas vairākas šī proteīna formas ar mainītām īpašībām un funkcijām - tā sauktie patoloģiskie hemoglobīni. Piemēram, hemoglobīnam S, kas atrodams pacientu ar sirpjveida šūnu anēmiju (HbS) sarkanajās asins šūnās, ir zema šķīdība zemā skābekļa daļējā spiediena apstākļos (kā tas ir venozās asinīs). Tas noved pie šī proteīna agregātu veidošanās. Olbaltumviela zaudē savu funkciju, izgulsnējas, un sarkanās asins šūnas kļūst neregulāras formas (dažas no tām veido sirpjveida formu) un tiek iznīcinātas ātrāk nekā parasti liesā. Tā rezultātā attīstās sirpjveida šūnu anēmija.
Vienīgā atšķirība HbA primārajā struktūrā tika konstatēta hemoglobīna β-ķēdes N-gala reģionā. Salīdziniet β-virknes N-gala reģionus un parādiet, kā izmaiņas proteīna primārajā struktūrā ietekmē tā īpašības un funkcijas.
 Priekš šī:
Priekš šī:
a) uzrakstiet aminoskābju formulas, ar kurām atšķiras HbA, un salīdziniet šo aminoskābju īpašības (polaritāti, lādiņu).
b) izdarīt secinājumu par šķīdības samazināšanās un skābekļa transportēšanas audos traucējumu iemeslu.
2. Attēlā parādīta proteīna struktūras diagramma, kurai ir saistīšanas centrs ar ligandu (aktīvais centrs). Paskaidrojiet, kāpēc proteīns ir selektīvs ligandu izvēlē. Priekš šī:
a) atcerieties, kas ir proteīna aktīvais centrs, un apsveriet attēlā redzamā proteīna aktīvā centra struktūru;
b) uzrakstiet aminoskābju radikāļu formulas, kas veido aktīvo centru;
c) uzzīmēt ligandu, kas varētu īpaši mijiedarboties ar proteīna aktīvo vietu. Norādiet uz tā funkcionālās grupas, kas var veidot saites ar aminoskābju radikāļiem, kas veido aktīvo centru;
d) norāda saišu veidus, kas rodas starp aktīvā centra ligandu un aminoskābju radikāļiem;
e) izskaidrot, uz ko balstās proteīna-ligandu mijiedarbības specifika.
 3.
Attēlā parādīta proteīna aktīvā vieta un vairāki ligandi.
3.
Attēlā parādīta proteīna aktīvā vieta un vairāki ligandi.
Nosakiet, kurš ligands visticamāk mijiedarbosies ar proteīna aktīvo vietu un kāpēc.
 Kāda veida saites rodas proteīna-ligandu kompleksa veidošanās laikā?
Kāda veida saites rodas proteīna-ligandu kompleksa veidošanās laikā?
4. Dabisko olbaltumvielu ligandu strukturālos analogus var izmantot kā zāles proteīnu aktivitātes modificēšanai.
Acetilholīns ir ierosmes pārraides mediators neiromuskulārās sinapsēs. Kad acetilholīns mijiedarbojas ar proteīniem - skeleta muskuļu postsinaptiskās membrānas receptoriem, atveras jonu kanāli un notiek muskuļu kontrakcija. Ditilīns ir zāles, ko lieto dažās operācijās, lai atslābinātu muskuļus, jo tas traucē nervu impulsu pārraidi caur neiromuskulārām sinapsēm. Izskaidrojiet ditilīna kā muskuļu relaksanta darbības mehānismu. Priekš šī:
a) uzrakstīt acetilholīna un ditilīna formulas un salīdzināt to struktūras;
b) apraksta ditilīna relaksējošās iedarbības mehānismu.
5. Dažu slimību gadījumā pacienta ķermeņa temperatūra paaugstinās, kas tiek uzskatīta par ķermeņa aizsargreakciju. Tomēr augsta temperatūra kaitē ķermeņa olbaltumvielām. Paskaidrojiet, kāpēc temperatūrā virs 40 °C tiek traucēta olbaltumvielu darbība un rodas draudi cilvēku dzīvībai. Lai to izdarītu, atcerieties:
1) Olbaltumvielu struktūra un saites, kas notur tās struktūru dabiskajā konformācijā;
2) Kā mainās olbaltumvielu struktūra un funkcija, paaugstinoties temperatūrai?;
3) Kas ir homeostāze un kāpēc tā ir svarīga cilvēka veselības saglabāšanai.
Moduļu vienība 2 OLIGOMERISMA PROTEĪNI KĀ REGULĒJOŠĀS IETEKMES MĒRĶI. PROTEĪNU STRUKTURĀLĀ UN FUNKCIONĀLĀ DAUDZVEIDĪBA. PROTEĪNU ATŠĶIRŠANAS UN ATTĪRĪŠANAS METODES
Mācību mērķi Spēt:
1. Izmantot zināšanas par oligomēru proteīnu uzbūves īpatnībām un funkcijām, lai izprastu to funkciju regulēšanas adaptīvos mehānismus.
2. Izskaidrot šaperonu lomu proteīna konformācijas sintēzē un uzturēšanā šūnu apstākļos.
3. Izskaidrojiet dzīvības izpausmju daudzveidību ar organismā sintezēto olbaltumvielu struktūru un funkciju dažādību.
4. Analizēt olbaltumvielu struktūras un to funkciju saistību, izmantojot radniecīgu hemoproteīnu - mioglobīna un hemoglobīna, kā arī piecu imūnglobulīnu saimes proteīnu klašu pārstāvju salīdzināšanas piemērus.
5. Pielietot zināšanas par olbaltumvielu fizikālo un ķīmisko īpašību īpatnībām, lai izvēlētos metodes to attīrīšanai no citiem proteīniem un piemaisījumiem.
6. Interpretēt asins plazmas proteīnu kvantitatīvā un kvalitatīvā sastāva rezultātus, lai apstiprinātu vai precizētu klīnisko diagnozi.
Zināt:
1. Oligomēro proteīnu struktūras iezīmes un adaptīvie mehānismi to funkciju regulēšanai, izmantojot hemoglobīna piemēru.
2. Šaperonu struktūra un funkcijas un to nozīme proteīnu dabiskās konformācijas uzturēšanā šūnu apstākļos.
3. Principi proteīnu apvienošanai ģimenēs, pamatojoties uz to uzbūves un funkciju līdzību, izmantojot imūnglobulīnu piemēru.
4. Proteīnu atdalīšanas metodes, pamatojoties uz to fizikāli ķīmisko īpašību īpašībām.
5. Asins plazmas elektroforēze kā proteīnu kvalitatīvā un kvantitatīvā sastāva novērtēšanas metode.
TĒMA 1.4. OLIGOMĒRU PROTEĪNU STRUKTŪRAS UN FUNKCIJAS ĪPAŠĪBAS, IZMANTOJOT HEMOGLOBĪNA PIEMĒRU
1. Daudzi proteīni satur vairākas polipeptīdu ķēdes. Tādus proteīnus sauc oligomērs, un atsevišķas ķēdes - protomēri. Oligomēru proteīnu protomērus savieno daudzas vājas nekovalentas saites (hidrofobās, jonu, ūdeņraža saites). Mijiedarbība
protomēri tiek veikti, pateicoties komplementaritāte to saskares virsmas.
Oligomēru proteīnu protomēru skaits var būt ļoti dažāds: hemoglobīns satur 4 protomērus, enzīms aspartātaminotransferāze ir 12 protomērus, bet tabakas mozaīkas vīrusa proteīns satur 2120 protomērus, kas savienoti ar nekovalentām saitēm. Līdz ar to oligomēru proteīniem var būt ļoti liela molekulmasa.
Viena protomēra mijiedarbību ar citiem var uzskatīt par īpašu proteīna-ligandu mijiedarbības gadījumu, jo katrs protomērs kalpo kā ligands citiem protomēriem. Protomēru skaitu un savienošanas metodi proteīnā sauc kvartāra proteīna struktūra.
Olbaltumvielas var saturēt vienādas vai atšķirīgas struktūras protomērus, piemēram, homodimēri ir proteīni, kas satur divus identiskus protomērus, un heterodimēri ir proteīni, kas satur divus dažādus protomērus.
Ja proteīni satur dažādus protomērus, tad uz tiem var veidoties saistīšanās centri ar dažādiem ligandiem, kas atšķiras pēc struktūras. Kad ligands saistās ar aktīvo vietu, izpaužas šī proteīna funkcija. Centru, kas atrodas uz cita protomēra, sauc par allosterisko (atšķirībā no aktīvā). Sazināšanās allosteriskais ligands vai efektors, tas veic regulējošu funkciju (1.18. att.). Allosteriskā centra mijiedarbība ar efektoru izraisa konformācijas izmaiņas visa oligomēra proteīna struktūrā tā konformācijas labilitātes dēļ. Tas ietekmē aktīvās vietas afinitāti pret konkrētu ligandu un regulē šī proteīna darbību. Visu protomēru konformācijas un funkcijas izmaiņas oligomēra proteīna mijiedarbības laikā ar vismaz vienu ligandu sauc par kooperatīvām konformācijas izmaiņām. Tiek saukti efektori, kas uzlabo olbaltumvielu darbību aktivatori, un efektori, kas kavē tā darbību - inhibitori.
Tādējādi oligomēriem proteīniem, kā arī olbaltumvielām ar domēna struktūru, salīdzinājumā ar monomēra proteīniem ir jauna īpašība - spēja allosteriski regulēt funkcijas (regulācija, pievienojot proteīnam dažādus ligandus). To var redzēt, salīdzinot divu cieši saistītu kompleksu proteīnu, mioglobīna un hemoglobīna, struktūras un funkcijas.
 Rīsi. 1.18. Dimēra proteīna struktūras shēma
Rīsi. 1.18. Dimēra proteīna struktūras shēma
2. Telpisko struktūru veidošanās un mioglobīna funkcionēšana.
Mioglobīns (Mb) ir sarkanajos muskuļos atrodams proteīns, kura galvenā funkcija ir radīt intensīvam muskuļu darbam nepieciešamās O 2 rezerves. Mb ir komplekss proteīns, kas satur proteīna daļu - apoMb un neproteīna daļu - hemu. ApoMB primārā struktūra nosaka tā kompakto lodveida uzbūvi un aktīvā centra struktūru, pie kura ir piesaistīta mioglobīna neproteīna daļa – hēms. Skābeklis, kas nāk no asinīm uz muskuļiem, saistās ar Fe+ 2 hēmām mioglobīnā. Mb ir monomērs proteīns, kam ir ļoti augsta afinitāte pret O 2, tāpēc skābekļa izdalīšanās ar mioglobīnu notiek tikai intensīva muskuļu darba laikā, kad krasi samazinās O 2 daļējais spiediens.
Mv konformācijas veidošanās. Sarkanajos muskuļos uz ribosomām translācijas laikā tiek sintezēta MB primārā struktūra, ko attēlo īpaša 153 aminoskābju atlikumu secība. Mb sekundārajā struktūrā ir astoņas α-spirāles, ko sauc ar latīņu burtiem no A līdz H, starp kurām ir nespirāli reģioni. Mb terciārajai struktūrai ir kompaktas lodītes forma, kuras padziļinājumā aktīvais centrs atrodas starp F un E α-spirālēm (1.19. att.).
 Rīsi. 1.19. Mioglobīna struktūra
Rīsi. 1.19. Mioglobīna struktūra
3. MV aktīvā centra uzbūves un funkcionēšanas īpatnības. Mb aktīvo centru galvenokārt veido hidrofobās aminoskābju radikāļi, kas primārajā struktūrā ir plaši izvietoti viens no otra (piemēram, Tri 3 9 un Fen 138) Slikti šķīst ūdenī ligandi - hēms un O 2 - piesaistās aktīvajam centram. Hēms ir specifisks apoMB ligands (1.20. att.), kura pamatu veido četri pirola gredzeni, kas savienoti ar metiltiltiņiem; centrā atrodas Fe+ 2 atoms, kas savienots ar pirola gredzenu slāpekļa atomiem ar četrām koordinācijas saitēm. Mb aktīvajā centrā papildus hidrofobajiem aminoskābju radikāļiem ir arī divu aminoskābju atlikumi ar hidrofiliem radikāļiem - Gis E 7(Gis 64) un ĢIS F 8(Viņa 93) (1.21. att.).
 Rīsi. 1.20. Hēma struktūra - mioglobīna un hemoglobīna neolbaltumvielu daļa
Rīsi. 1.20. Hēma struktūra - mioglobīna un hemoglobīna neolbaltumvielu daļa
 Rīsi. 1.21. Hēma un O2 atrašanās vieta apomioglobīna un hemoglobīna protomēru aktīvajā vietā
Rīsi. 1.21. Hēma un O2 atrašanās vieta apomioglobīna un hemoglobīna protomēru aktīvajā vietā
Hēms ir kovalenti saistīts ar Viņa F8 caur dzelzs atomu. O 2 piestiprinās pie dzelzs otrā pusē hēma plaknei. Viņa E 7 ir nepieciešams pareizai O 2 orientācijai un atvieglo skābekļa pievienošanu Fe + 2 hēmam
ĢIS F 8 veido koordinācijas saiti ar Fe+ 2 un stingri fiksē hēmu aktīvajā centrā. Gis E 7 nepieciešams pareizai orientācijai cita liganda aktīvajā centrā - O 2 tā mijiedarbības laikā ar Fe + 2 hēmu. Hēma mikrovide rada apstākļus spēcīgai, bet atgriezeniskai O 2 saistīšanai ar Fe + 2 un novērš ūdens iekļūšanu hidrofobajā aktīvajā vietā, kas var izraisīt tā oksidēšanos līdz Fe + 3.
Mb monomēra struktūra un tā aktīvais centrs nosaka proteīna augsto afinitāti pret O 2.
4. Hb oligomēra struktūra un Hb afinitātes regulēšana pret O 2 ligandiem. Cilvēka hemoglobīns- olbaltumvielu saime, piemēram, mioglobīns, kas saistīta ar kompleksiem proteīniem (hemoproteīniem). Tiem ir tetramēra struktūra un tie satur divas α-ķēdes, bet atšķiras pēc pārējo divu polipeptīdu ķēžu (2α-, 2x-ķēdes) struktūras. Otrās polipeptīdu ķēdes struktūra nosaka šo Hb formu funkcionēšanas īpašības. Apmēram 98% hemoglobīna pieaugušā sarkano asins šūnu sastāvā ir hemoglobīns A(2α-, 2p-ķēdes).
Augļa attīstības laikā darbojas divi galvenie hemoglobīna veidi: embrija Hb(2α, 2ε), kas sastopams augļa attīstības sākumposmā, un hemoglobīns F (augļa)- (2α, 2γ), kas aizvieto agrīnu augļa hemoglobīnu sestajā intrauterīnās attīstības mēnesī un tikai pēc dzimšanas tiek aizstāts ar Hb A.
HB A ir proteīns, kas saistīts ar mioglobīnu (MB), kas atrodams pieaugušo sarkanajās asins šūnās. Tā atsevišķo protomēru struktūra ir līdzīga mioglobīna struktūrai. Mioglobīna un hemoglobīna protomēru sekundārās un terciārās struktūras ir ļoti līdzīgas, neskatoties uz to, ka to polipeptīdu ķēžu primārajā struktūrā tikai 24 aminoskābju atlikumi ir identiski (hemoglobīna protomēru sekundārā struktūra, tāpat kā mioglobīns, satur astoņas α-spirāles, apzīmē ar latīņu burtiem no A līdz H , un terciārajai struktūrai ir kompaktas lodītes forma). Bet atšķirībā no mioglobīna hemoglobīnam ir oligomēra struktūra, kas sastāv no četrām polipeptīdu ķēdēm, kas savienotas ar nekovalentām saitēm (1.22. attēls).
Katrs Hb protomērs ir saistīts ar ne-olbaltumvielu daļu - hemu un blakus esošajiem protomēriem. Hb proteīna daļas savienojums ar hēmu ir līdzīgs mioglobīna savienojumam: proteīna aktīvajā centrā hēma hidrofobās daļas ieskauj hidrofobās aminoskābju radikāļi, izņemot His F 8 un His E 7, kas atrodas abās hēma plaknes pusēs un spēlē līdzīgu lomu proteīna funkcionēšanā un saistīšanā ar skābekli (skat. mioglobīna uzbūvi).
 Rīsi. 1.22. Hemoglobīna oligomēriskā struktūra
Rīsi. 1.22. Hemoglobīna oligomēriskā struktūra
Turklāt, Gis E 7 veic svarīgu papildu loma Nv darbībā. Brīvajam hēmam ir 25 000 reižu lielāka afinitāte pret CO nekā pret O2. CO veidojas nelielos daudzumos organismā, un, ņemot vērā tā augsto afinitāti pret hēmu, tas var traucēt šūnu dzīvībai nepieciešamā O 2 transportēšanu. Tomēr hemoglobīna sastāvā hēma afinitāte pret oglekļa monoksīdu pārsniedz afinitāti pret O 2 tikai 200 reizes, jo aktīvajā centrā atrodas His E 7. Atlikusī šīs aminoskābes daļa rada optimālus apstākļus hēma saistīšanai ar O 2 un vājina hēma mijiedarbību ar CO.
5. HB galvenā funkcija ir O2 transportēšana no plaušām uz audiem. Atšķirībā no monomēra mioglobīna, kam ir ļoti augsta afinitāte pret O2 un kas veic skābekļa uzglabāšanas funkciju sarkanajos muskuļos, hemoglobīna oligomērā struktūra nodrošina:
1) ātra HB piesātināšana ar skābekli plaušās;
2) HB spēja izdalīt skābekli audos pie salīdzinoši augsta O 2 daļējā spiediena (20-40 mm Hg);
3) iespēja regulēt Hb afinitāti pret O 2.
6. Kooperatīvās izmaiņas hemoglobīna protomēru konformācijā paātrina O 2 saistīšanos plaušās un izdalīšanos audos. Plaušās augstais O 2 daļējais spiediens veicina tā saistīšanos ar Hb četru protomēru (2α un 2β) aktīvajā vietā. Katra protomēra aktīvais centrs, tāpat kā mioglobīnā, atrodas starp divām α-spirālēm (F un E) hidrofobā kabatā. Tas satur neolbaltumvielu daļu - hēmu, kas pievienota proteīna daļai ar daudzām vājām hidrofobām mijiedarbībām un vienu spēcīgu saiti starp Fe 2 + hēmu un His F 8 (sk. 1.21. att.).
Deoksihemoglobīnā, pateicoties šai saitei ar His F 8, Fe 2 + atoms izvirzās no hēma plaknes histidīna virzienā. O 2 saistīšanās ar Fe 2 + notiek hēma otrā pusē His E 7 reģionā, izmantojot vienu brīvu koordinācijas saiti. Viņa E 7 nodrošina optimālus apstākļus O 2 saistīšanai ar hema dzelzi.
O 2 pievienošana viena protomēra Fe + 2 atomam izraisa tā pārvietošanos hēma plaknē, kam seko ar to saistītais histidīna atlikums
 Rīsi. 1.23. Hemoglobīna protomēra konformācijas izmaiņas, kombinējot ar O 2
Rīsi. 1.23. Hemoglobīna protomēra konformācijas izmaiņas, kombinējot ar O 2
Tas izraisa izmaiņas visu polipeptīdu ķēžu konformācijā to konformācijas labilitātes dēļ. Citu ķēžu konformācijas maiņa atvieglo to mijiedarbību ar nākamajām O 2 molekulām.
Ceturtā O 2 molekula hemoglobīnam piesaistās 300 reizes vieglāk nekā pirmā (1.24. att.).
 Rīsi. 1.24. Kooperatīvas izmaiņas hemoglobīna protomēru konformācijā tās mijiedarbības laikā ar O2
Rīsi. 1.24. Kooperatīvas izmaiņas hemoglobīna protomēru konformācijā tās mijiedarbības laikā ar O2
Audos katra nākamā O 2 molekula tiek atdalīta vieglāk nekā iepriekšējā, arī protomēru konformācijas kooperatīvo izmaiņu dēļ.
7. CO 2 un H+, kas veidojas organisko vielu katabolisma laikā, samazina hemoglobīna afinitāti pret O 2 proporcionāli to koncentrācijai. Šūnu funkcionēšanai nepieciešamo enerģiju galvenokārt ražo mitohondrijās organisko vielu oksidēšanās laikā, izmantojot O 2, ko no plaušām piegādā hemoglobīns. Organisko vielu oksidēšanās rezultātā veidojas to sadalīšanās galaprodukti: CO 2 un K 2 O, kuru daudzums ir proporcionāls notiekošo oksidācijas procesu intensitātei.
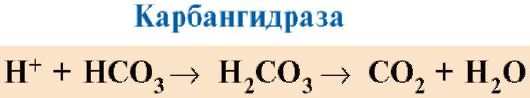
CO 2 izkliedējas no šūnām asinīs un iekļūst sarkanajās asins šūnās, kur enzīma karbanhidrāzes ietekmē tiek pārveidots par ogļskābi. Šī vājā skābe sadalās protonu un bikarbonāta jonos.
H+ spēj pievienoties Viņa radikāļiem 14 6 hemoglobīna α- un β-ķēdēs, t.i. apgabalos, kas atrodas tālu no hema. Hemoglobīna protonēšana samazina tā afinitāti pret O 2, veicina O 2 izvadīšanu no oksiHb, deoksiHb veidošanos un palielina audu piegādi ar skābekli proporcionāli izveidoto protonu skaitam (1.25. att.).
Atbrīvotā skābekļa daudzuma palielināšanos atkarībā no H+ koncentrācijas palielināšanās sarkanajās asins šūnās sauc par Bora efektu (nosaukts dāņu fiziologa Kristiana Bora vārdā, kurš pirmo reizi atklāja šo efektu).
Plaušās augsts skābekļa parciālais spiediens veicina tā saistīšanos ar dezoksiHb, kas samazina proteīna afinitāti pret H+. Ogļskābes iedarbībā atbrīvotie protoni reaģē ar bikarbonātiem, veidojot CO 2 un H 2 O
 Rīsi. 1.25. Hb afinitātes pret O 2 atkarība no CO 2 un protonu koncentrācijas (Bora efekts):
Rīsi. 1.25. Hb afinitātes pret O 2 atkarība no CO 2 un protonu koncentrācijas (Bora efekts):
A- CO 2 un H+ koncentrācijas ietekme uz O 2 izdalīšanos no kompleksa ar HB (Bora efekts); B- deoksihemoglobīna piesātināšana ar skābekli plaušās, CO 2 veidošanās un izdalīšanās.
Iegūtais CO 2 nonāk alveolārajā telpā un tiek izvadīts ar izelpoto gaisu. Tādējādi hemoglobīna izdalītā skābekļa daudzumu audos regulē organisko vielu katabolisma produkti: jo intensīvāk notiek vielu sadalīšanās, piemēram, fizisko slodzi, jo augstāka ir CO 2 un H + koncentrācija un vairāk skābekļa. audi saņem Hb afinitātes pret O 2 samazināšanās rezultātā.
8. Hb afinitātes pret O2 allosteriskā regulēšana ar ligandu - 2,3-bisfosfoglicerātu. Eritrocītos hemoglobīna allosteriskais ligands 2,3-bisfosfoglicerāts (2,3-BPG) tiek sintezēts no glikozes oksidācijas produkta - 1,3-bisfosfoglicerāta. Normālos apstākļos 2,3-BPG koncentrācija ir augsta un salīdzināma ar Hb koncentrāciju. 2,3-BPG ir spēcīgs negatīvs lādiņš -5.
 Bisfosfoglicerāts audu kapilāros, saistoties ar deoksihemoglobīnu, palielina skābekļa izdalīšanos audos, samazinot Hb afinitāti pret O 2.
Bisfosfoglicerāts audu kapilāros, saistoties ar deoksihemoglobīnu, palielina skābekļa izdalīšanos audos, samazinot Hb afinitāti pret O 2.
Tetrameriskās hemoglobīna molekulas centrā ir dobums. To veido visu četru protomēru aminoskābju atlikumi (sk. 1.22. att.). Audu kapilāros Hb protonēšana (Bora efekts) noved pie saites starp hema dzelzi un O2 pārrāvumu. Molekulā
deoksihemoglobīns, salīdzinot ar oksihemoglobīnu, parādās papildu jonu saites, kas savieno protomērus, kā rezultātā palielinās centrālās dobuma izmēri salīdzinājumā ar oksihemoglobīnu. Centrālais dobums ir vieta, kur 2,3-BPG pieķeras hemoglobīnam. Centrālā dobuma lieluma atšķirības dēļ 2,3-BPG var pievienoties tikai deoksihemoglobīnam.
2,3-BPG mijiedarbojas ar hemoglobīnu vietā, kas atrodas tālu no proteīna aktīvajiem centriem un pieder allostērisks(regulējošie) ligandi, un Hb centrālais dobums ir allosteriskais centrs. 2,3-BPG ir spēcīgs negatīvs lādiņš un mijiedarbojas ar piecām pozitīvi lādētām divu Hb β-ķēžu grupām: Val N-gala α-aminogrupu un Lys 82 His 143 radikāļiem (1.26. att.).
 Rīsi. 1.26. BPG deoksihemoglobīna centrālajā dobumā
Rīsi. 1.26. BPG deoksihemoglobīna centrālajā dobumā
BPG saistās ar trim pozitīvi lādētām grupām katrā β-virknē.
Audu kapilāros iegūtais deoksihemoglobīns mijiedarbojas ar 2,3-BPG un starp pozitīvi lādētajiem β-ķēžu radikāļiem un negatīvi lādēto ligandu veidojas jonu saites, kas maina proteīna konformāciju un samazina Hb afinitāti pret O2. . Hb afinitātes samazināšanās pret O 2 veicina efektīvāku O 2 izdalīšanos audos.
Plaušās pie augsta parciālā spiediena skābeklis mijiedarbojas ar Hb, savienojoties ar hema dzelzi; šajā gadījumā mainās proteīna konformācija, samazinās centrālais dobums un 2,3-BPG tiek izspiests no allosteriskā centra
Tādējādi oligomēriem proteīniem ir jaunas īpašības salīdzinājumā ar monomēru proteīniem. Ligandu piestiprināšana vietās
telpiski attālināti viens no otra (allostēriski), var izraisīt konformācijas izmaiņas visā proteīna molekulā. Sakarā ar mijiedarbību ar regulējošajiem ligandiem notiek izmaiņas proteīna molekulas konformācijā un funkcijas pielāgošanās vides izmaiņām.
TĒMA 1.5. PROTEĪNU ATTIECĪBĀS KONFORMĀCIJAS UZTURĒŠANA ŠŪNU APSTĀKĻOS
Šūnās polipeptīdu ķēžu sintēzes laikā to transportēšana caur membrānām uz attiecīgajām šūnas daļām, locīšanas procesā (native konformācijas veidošanās) un oligomēru proteīnu montāžas laikā, kā arī to funkcionēšanas laikā, starpprodukts. , proteīna struktūrā rodas agregācijai pakļautas, nestabilas konformācijas. Hidrofobie radikāļi, kas parasti slēpjas proteīna molekulas iekšienē dabiskajā konformācijā, parādās uz virsmas nestabilā konformācijā un mēdz apvienoties ar citu proteīnu grupām, kas slikti šķīst ūdenī. Visu zināmo organismu šūnās ir atrastas īpašas olbaltumvielas, kas nodrošina optimālu šūnu proteīnu locīšanu, stabilizē to dabisko konformāciju funkcionēšanas laikā un, galvenais, saglabā intracelulāro proteīnu struktūru un funkcijas, ja tiek traucēta homeostāze. Šos proteīnus sauc "pavadītāji" kas franču valodā nozīmē "aukle".
1. Molekulārie šaperoni un to nozīme olbaltumvielu denaturācijas novēršanā.
Šaperoni (CH) tiek klasificēti pēc to apakšvienību masas. Augstas molekulmasas šaperonu masa ir no 60 līdz 110 kDa. No tām visvairāk pētītas trīs klases: Sh-60, Sh-70 un Sh-90. Katrā klasē ietilpst saistītu proteīnu saime. Tādējādi Sh-70 satur proteīnus ar molekulmasu no 66 līdz 78 kDa. Šaperonu ar zemu molekulmasu molekulmasa ir no 40 līdz 15 kDa.
Starp pavadoņiem ir konstitutīvs olbaltumvielas, kuru augstā bazālā sintēze nav atkarīga no stresa ietekmes uz ķermeņa šūnām, un inducējams, kuru sintēze normālos apstākļos ir vāja, bet stresa apstākļos strauji palielinās. Inducējamos šaperonus sauc arī par "karstuma šoka proteīniem", jo tie pirmo reizi tika atklāti šūnās, kas pakļautas augstām temperatūrām. Šūnās proteīnu augstās koncentrācijas dēļ daļēji denaturētu proteīnu spontāna reaktivācija ir sarežģīta. Sh-70 var novērst denaturācijas sākšanos un palīdzēt atjaunot proteīnu dabisko konformāciju. Molekulārie šaperoni-70- ļoti konservēta proteīnu klase, kas atrodama visās šūnas daļās: citoplazmā, kodolā, endoplazmatiskajā retikulumā, mitohondrijās. Atsevišķas polipeptīda ķēdes Ш-70 karboksilgalā ir apgabals, kas ir rieva, kas spēj mijiedarboties ar peptīdiem garumā
no 7 līdz 9 aminoskābju atlikumiem, kas bagātināti ar hidrofobiem radikāļiem. Šādi apgabali globulārajos proteīnos rodas aptuveni ik pēc 16 aminoskābēm. Sh-70 spēj aizsargāt proteīnus no temperatūras inaktivācijas un atjaunot daļēji denaturētu proteīnu konformāciju un aktivitāti.
2. Šaperonu loma olbaltumvielu locīšanas procesā. Proteīna sintēzes laikā uz ribosomas polipeptīda N-gala reģions tiek sintezēts pirms C-gala. Lai izveidotu dabisko konformāciju, ir nepieciešama pilnīga proteīna aminoskābju secība. Olbaltumvielu sintēzes procesā šaperoni-70, pateicoties sava aktīvā centra struktūrai, spēj aizvērt polipeptīda apgabalus, kas ir pakļauti agregācijai, bagātināti ar hidrofobajiem aminoskābju radikāļiem, līdz sintēze ir pabeigta (1.27. attēls, A). ).
 Rīsi. 1.27. Šaperonu līdzdalība olbaltumvielu locīšanas procesā
Rīsi. 1.27. Šaperonu līdzdalība olbaltumvielu locīšanas procesā
A - chaperonu-70 līdzdalība hidrofobās mijiedarbības novēršanā starp sintezētā polipeptīda sekcijām; B - proteīna dabiskās konformācijas veidošanās chaperona kompleksā
Daudzi lielmolekulārie proteīni, kuriem ir sarežģīta uzbūve, piemēram, domēna struktūra, salokās īpašā telpā, ko veido Sh-60. Ш-60 darbojas kā oligomēru komplekss, kas sastāv no 14 apakšvienībām. Tie veido divus dobus gredzenus, no kuriem katrs sastāv no septiņām apakšvienībām, šie gredzeni ir savienoti viens ar otru. Katra Sh-60 apakšvienība sastāv no trim domēniem: apikālā (apikālā), bagātināta ar hidrofobiem radikāļiem, kas vērsti pret gredzena dobumu, starpposma un ekvatoriālā (1.28. att.).
 Rīsi. 1.28. Šaperonīna kompleksa struktūra, kas sastāv no 14 Ш-60
Rīsi. 1.28. Šaperonīna kompleksa struktūra, kas sastāv no 14 Ш-60
A - sānskats; B - skats no augšas
Sintezēti proteīni, kuru virsmā ir nesalocītām molekulām raksturīgi elementi, jo īpaši hidrofobi radikāļi, nonāk šaperona gredzenu dobumā. Konkrētajā šo dobumu vidē tiek meklētas iespējamās konformācijas, līdz tiek atrasta vienīgā enerģētiski vislabvēlīgākā (1.27. att., B). Konformāciju veidošanos un olbaltumvielu izdalīšanos pavada ATP hidrolīze ekvatoriālajā reģionā. Parasti šādai šaperona locīšanai ir nepieciešams ievērojams enerģijas daudzums.
Papildus dalībai proteīnu trīsdimensiju struktūras veidošanā un daļēji denaturētu proteīnu renativācijā, šaperoni ir nepieciešami arī tādu fundamentālu procesu norisei kā oligomēru proteīnu montāža, denaturēto proteīnu atpazīšana un transportēšana lizosomās, proteīnu transportēšana cauri membrānām un līdzdalība proteīnu kompleksu aktivitātes regulēšanā.
TĒMA 1.6. PROTEĪNU DAŽĀDI. PROTEĪNU ĢIMENES: IMUNOGLOBULĪNU PIEMĒRS
1. Olbaltumvielām ir izšķiroša nozīme atsevišķu šūnu un visa daudzšūnu organisma dzīvē, un to funkcijas ir pārsteidzoši daudzveidīgas. To nosaka proteīnu primārās struktūras un konformāciju īpašības, unikālā aktīvā centra struktūra un spēja saistīt specifiskus ligandus.
Tikai ļoti neliela daļa no visiem iespējamiem peptīdu ķēžu variantiem var pieņemt stabilu telpisko struktūru; vairums
no tiem var būt daudzas konformācijas ar aptuveni vienādu Gibsa enerģiju, bet ar dažādām īpašībām. Lielāko daļu zināmo proteīnu primārā struktūra, kas atlasīta bioloģiskās evolūcijas ceļā, nodrošina vienas no konformācijām izcilu stabilitāti, kas nosaka šī proteīna funkcionēšanas īpašības.
2. Olbaltumvielu saimes. Vienā un tajā pašā bioloģiskajā sugā aminoskābju atlikumu aizstāšana var izraisīt dažādu proteīnu parādīšanos, kas veic saistītas funkcijas un kurām ir homologas aminoskābju sekvences. Šādiem radniecīgiem proteīniem ir pārsteidzoši līdzīgas konformācijas: α-spirāļu un/vai β-struktūru skaits un relatīvais novietojums, lielākā daļa polipeptīdu ķēžu pagriezienu un līkumu ir līdzīgi vai identiski. Olbaltumvielas ar homologiem polipeptīdu ķēdes reģioniem, līdzīgu konformāciju un saistītām funkcijām klasificē proteīnu ģimenēs. Olbaltumvielu ģimeņu piemēri: serīna proteināzes, imūnglobulīnu saime, mioglobīnu saime.
Serīna proteināzes- olbaltumvielu saime, kas veic proteolītisko enzīmu funkciju. Tajos ietilpst gremošanas enzīmi – himotripsīns, tripsīns, elastāze un daudzi asinsreces faktori. Šīm olbaltumvielām ir identiskas aminoskābes 40% to pozīcijās un ļoti līdzīga uzbūve (1.29. att.).

Rīsi. 1.29. Elastāzes (A) un himotripsīna (B) telpiskās struktūras
Dažas aminoskābju aizvietošanas ir izraisījušas izmaiņas šo proteīnu substrāta specifikā un funkcionālās daudzveidības rašanos ģimenē.
3. Imūnglobulīnu saime. Imūnsistēmas darbībā liela nozīme ir imūnglobulīnu superģimenes olbaltumvielām, kas ietver trīs proteīnu ģimenes:
Antivielas (imūnglobulīni);
T-limfocītu receptori;
Galvenā histokompatibilitātes kompleksa olbaltumvielas - MHC 1. un 2. klase (Major Histocompatibility Complex).
Visām šīm olbaltumvielām ir domēna struktūra, tās sastāv no homologiem imūnsistēmai līdzīgiem domēniem un veic līdzīgas funkcijas: tās mijiedarbojas ar svešām struktūrām, vai nu izšķīdušas asinīs, limfā vai starpšūnu šķidrumā (antivielas), vai atrodas uz šūnu virsmas (pašu vai ārzemju).
4. Antivielas- specifiskas olbaltumvielas, ko ražo B limfocīti, reaģējot uz svešas struktūras iekļūšanu organismā, ko sauc antigēns.
Antivielu struktūras iezīmes
Vienkāršākās antivielu molekulas sastāv no četrām polipeptīdu ķēdēm: divām identiskām vieglajām - L, kas satur apmēram 220 aminoskābes, un divas identiskas smagas - H, kas sastāv no 440-700 aminoskābēm. Visas četras ķēdes antivielu molekulā ir saistītas ar daudzām nekovalentām saitēm un četrām disulfīda saitēm (1.30. att.).
Antivielu vieglās ķēdes sastāv no diviem domēniem: mainīgā domēna (VL), kas atrodas polipeptīdu ķēdes N-gala reģionā, un konstantā domēna (CL), kas atrodas C-galā. Smagajām ķēdēm parasti ir četri domēni: viens mainīgais (VH), kas atrodas N-galā, un trīs nemainīgie domēni (CH1, CH2, CH3) (sk. 1.30. att.). Katram imūnglobulīna domēnam ir β-loksnes virsbūve, kurā divi cisteīna atlikumi ir saistīti ar disulfīda saiti.
Starp diviem pastāvīgajiem domēniem CH1 un CH2 atrodas reģions, kurā ir liels skaits prolīna atlikumu, kas novērš sekundārās struktūras veidošanos un blakus esošo H-ķēžu mijiedarbību šajā segmentā. Šis eņģes reģions nodrošina antivielas molekulas elastību. Starp smagās un vieglās ķēdes mainīgajiem domēniem ir divas identiskas antigēnu saistīšanās vietas (aktīvās vietas antigēnu saistīšanai), tāpēc šādas antivielas bieži sauc. bivalenti. Antigēna saistīšanā ar antivielu ir iesaistīta nevis visa abu ķēžu mainīgo reģionu aminoskābju secība, bet tikai 20-30 aminoskābes, kas atrodas katras ķēdes hipermainīgajos reģionos. Tieši šie reģioni nosaka katra antivielu veida unikālo spēju mijiedarboties ar atbilstošo komplementāro antigēnu.
Antivielas ir viena no ķermeņa aizsardzības līnijām pret svešiem organismiem. To funkcionēšanu var iedalīt divos posmos: pirmais posms ir antigēna atpazīšana un saistīšanās uz svešu organismu virsmas, kas iespējama, pateicoties antigēnu saistošo vietu klātbūtnei antivielu struktūrā; otrais posms ir antigēna inaktivācijas un iznīcināšanas procesa uzsākšana. Otrā posma specifika ir atkarīga no antivielu klases. Ir piecas smago ķēžu klases, kas viena no otras atšķiras ar konstanto domēnu struktūru: α, δ, ε, γ un μ, pēc kurām izšķir piecas imūnglobulīnu klases: A, D, E, G un M.
Smago ķēžu strukturālās iezīmes piešķir smago ķēžu viras reģioniem un C-gala reģioniem katrai klasei raksturīgu konformāciju. Pēc tam, kad antigēns saistās ar antivielu, konformācijas izmaiņas konstantajos domēnos nosaka antigēna noņemšanas ceļu.
 Rīsi. 1. 30. IgG domēna struktūra
Rīsi. 1. 30. IgG domēna struktūra
Imūnglobulīni M
Imūnglobulīniem M ir divas formas.
Monomēra forma- 1. klases antivielas, ko ražo, attīstot B limfocītus. Pēc tam daudzas B šūnas pāriet uz citu klašu antivielu ražošanu, bet ar to pašu antigēnu saistīšanas vietu. IgM ir iestrādāts membrānā un darbojas kā antigēna atpazīšanas receptors. IgM integrācija šūnu membrānā ir iespējama, jo reģiona astes daļā ir 25 hidrofobās aminoskābju atliekas.
IgM sekrēcijas forma satur piecas monomēru apakšvienības, kas savstarpēji saistītas ar disulfīda saitēm, un papildu polipeptīda J ķēdi (1.31. att.). Šīs formas monomēru smagās ķēdes nesatur hidrofobu. Pentamēram ir 10 antigēnu saistīšanās vietas, un tāpēc tas ir efektīvs, lai atpazītu un noņemtu antigēnu, kas pirmais nonāk organismā. IgM sekrēcijas forma ir galvenā antivielu klase, kas primārās imūnās atbildes laikā izdalās asinīs. IgM saistīšanās ar antigēnu maina IgM konformāciju un izraisa tā saistīšanos ar komplementa sistēmas pirmo proteīna komponentu (komplementa sistēma ir proteīnu kopums, kas iesaistīts antigēna iznīcināšanā) un šīs sistēmas aktivāciju. Ja antigēns atrodas uz mikroorganisma virsmas, komplementa sistēma izraisa šūnas membrānas integritātes traucējumus un baktērijas šūnas nāvi.
Imūnglobulīni G
Kvantitatīvi šīs klases imūnglobulīni dominē asinīs (75% no visiem Ig). IgG - monomēri, galvenā antivielu klase, kas izdalās asinīs sekundārās imūnās atbildes laikā. Pēc IgG mijiedarbības ar mikroorganismu virsmas antigēniem antigēna-antivielu komplekss spēj saistīt un aktivizēt komplementa sistēmas proteīnus vai var mijiedarboties ar specifiskiem makrofāgu un neitrofilu receptoriem. Mijiedarbība ar fagocītiem noved
 Rīsi. 1.31. IgM sekrēcijas formas struktūra
Rīsi. 1.31. IgM sekrēcijas formas struktūra
uz antigēnu-antivielu kompleksu uzsūkšanos un to iznīcināšanu šūnu fagosomās. IgG ir vienīgā antivielu klase, kas spēj iekļūt placentas barjerā un nodrošināt augļa intrauterīnu aizsardzību pret infekcijām.
Imūnglobulīni A
Galvenā antivielu klase, kas atrodas izdalījumos (pienā, siekalās, elpceļu un zarnu trakta izdalījumos). IgA izdalās galvenokārt dimēra formā, kur monomēri ir saistīti viens ar otru caur papildu J ķēdi (1.32. att.).
IgA nesadarbojas ar komplementa sistēmu un fagocītiskajām šūnām, bet, saistoties ar mikroorganismiem, antivielas novērš to piesaisti epitēlija šūnām un iekļūšanu organismā.
Imūnglobulīni E
Imūnglobulīnus E attēlo monomēri, kuros smagās ε-ķēdes, tāpat kā imūnglobulīnu M μ-ķēdes, satur vienu mainīgu un četrus nemainīgus domēnus. Pēc sekrēcijas IgE saistās ar to
 Rīsi. 1.32. IgA struktūra
Rīsi. 1.32. IgA struktūra
C-gala reģioni ar atbilstošiem receptoriem uz tuklo šūnu un bazofilu virsmas. Rezultātā tie kļūst par antigēnu receptoriem uz šo šūnu virsmas (1.33. att.).
 Rīsi. 1.33. IgE mijiedarbība ar antigēnu uz tuklās šūnas virsmas
Rīsi. 1.33. IgE mijiedarbība ar antigēnu uz tuklās šūnas virsmas
Pēc tam, kad antigēns pievienojas attiecīgajām IgE antigēnu saistošajām vietām, šūnas saņem signālu, lai izdalītos bioloģiski aktīvas vielas (histamīns, serotonīns), kas lielā mērā ir atbildīgas par iekaisuma reakcijas attīstību un alerģisku reakciju izpausmēm, piemēram, astma, nātrene, siena drudzis.
Imūnglobulīni D
Imūnglobulīni D ir atrodami ļoti mazos daudzumos serumā, tie ir monomēri. Smagajām δ ķēdēm ir viens mainīgs un trīs nemainīgi domēni. IgD darbojas kā B limfocītu receptori, citas funkcijas joprojām nav zināmas. Specifisku antigēnu mijiedarbība ar receptoriem uz B-limfocītu (IgD) virsmas noved pie šo signālu pārnešanas šūnā un mehānismu aktivizēšanas, kas nodrošina konkrētā limfocītu klona proliferāciju.
TĒMA 1.7. PROTEĪNU FIZIKĀLĀS UN ĶĪMISKĀS ĪPAŠĪBAS UN TO ATdalīšanas metodes
1. Atsevišķi proteīni atšķiras pēc fizikālajām un ķīmiskajām īpašībām:
molekulu forma;
Molekulārais svars;
Kopējais lādiņš, kura lielums ir atkarīgs no aminoskābju anjonu un katjonu grupu attiecības;
Polāro un nepolāro aminoskābju radikāļu attiecība uz molekulu virsmas;
Izturības pakāpes pret dažādiem denaturējošiem līdzekļiem.
2. Olbaltumvielu šķīdība ir atkarīga par iepriekš uzskaitīto olbaltumvielu īpašībām, kā arī par barotnes sastāvu, kurā proteīns ir izšķīdināts (pH vērtības, sāls sastāvs, temperatūra, citu organisko vielu klātbūtne, kas var mijiedarboties ar proteīnu). Olbaltumvielu molekulu lādiņa daudzums ir viens no faktoriem, kas ietekmē to šķīdību. Kad lādiņš izoelektriskajā punktā tiek zaudēts, olbaltumvielas vieglāk agregējas un izgulsnējas. Tas ir īpaši raksturīgi denaturētiem proteīniem, kuru virspusē parādās hidrofobās aminoskābju radikāļi.
Uz olbaltumvielu molekulas virsmas ir gan pozitīvi, gan negatīvi lādēti aminoskābju radikāļi. Šo grupu skaits un līdz ar to arī kopējais olbaltumvielu lādiņš ir atkarīgs no barotnes pH, t.i. H+ - un OH - grupu koncentrāciju attiecība. Skābā vidē H+ koncentrācijas palielināšanās noved pie karboksilgrupu -COO - + H+ > -COOH disociācijas nomākšanas un olbaltumvielu negatīvā lādiņa samazināšanās. Sārmainā vidē liekā OH saistīšanās ar protoniem, kas veidojas aminogrupu -NH 3 + + OH - - NH 2 + H 2 O disociācijas laikā ar ūdens veidošanos, samazina olbaltumvielu pozitīvo lādiņu. . Tiek saukta pH vērtība, pie kuras proteīnam ir nulles lādiņš izoelektriskais punkts (IEP). IET pozitīvi un negatīvi lādēto grupu skaits ir vienāds, t.i. proteīns ir izoelektriskā stāvoklī.
3. Atsevišķu olbaltumvielu atdalīšana.Ķermeņa struktūras un darbības iezīmes ir atkarīgas no tajā sintezēto olbaltumvielu kopuma. Olbaltumvielu struktūras un īpašību izpēte nav iespējama bez to izolēšanas no šūnas un attīrīšanas no citām olbaltumvielām un organiskajām molekulām. Atsevišķu proteīnu izolācijas un attīrīšanas posmi:
Šūnu iznīcināšana pētāmos audus un iegūstot homogenātu.
Homogenāta sadalīšana frakcijās centrifugējot, iegūstot kodolu, mitohondriju, citozola vai citu frakciju, kas satur vēlamo proteīnu.
Selektīva termiskā denaturācija- īslaicīga proteīna šķīduma karsēšana, kuras laikā var noņemt dažus denaturēto olbaltumvielu piemaisījumus (ja proteīns ir relatīvi karstumizturīgs).
Izsālīšana. Dažādi proteīni šķīdumā izgulsnējas dažādās sāls koncentrācijās. Pakāpeniski palielinot sāls koncentrāciju, ir iespējams iegūt vairākas atsevišķas frakcijas, kurās vienā no tām dominē izolēta proteīna saturs. Amonija sulfātu visbiežāk izmanto olbaltumvielu frakcionēšanai. Olbaltumvielas ar vismazāko šķīdību izgulsnējas zemā sāls koncentrācijā.
Gēla filtrēšana- metode molekulu sijāšanai caur uzbriedušām Sephadex granulām (trīsdimensiju dekstrāna polisaharīdu ķēdēm ar porām). Ātrums, ar kādu olbaltumvielas iziet cauri kolonnai, kas piepildīta ar Sephadex, būs atkarīgs no to molekulmasas: jo mazāka ir olbaltumvielu molekulu masa, jo vieglāk tās iekļūst granulās un paliek tur ilgāk, jo ātrāk tās izplūst no tās kolonna.
Ultracentrifugēšana- metode, kas ietver olbaltumvielu ievietošanu centrifūgas mēģenē ultracentrifūgas rotorā. Kad rotors griežas, olbaltumvielu sedimentācijas ātrums ir proporcionāls to molekulmasai: smagāku proteīnu frakcijas atrodas tuvāk mēģenes apakšai, vieglākas - tuvāk virsmai.
Elektroforēze- metode, kuras pamatā ir olbaltumvielu kustības ātruma atšķirības elektriskā laukā. Šī vērtība ir proporcionāla olbaltumvielu lādiņam. Olbaltumvielu elektroforēzi veic uz papīra (šajā gadījumā olbaltumvielu kustības ātrums ir proporcionāls tikai to lādiņam) vai poliakrilamīda gēlā ar noteiktu poru izmēru (olbaltumvielu kustības ātrums ir proporcionāls to lādiņam un molekulmasai) .
Jonu apmaiņas hromatogrāfija- frakcionēšanas metode, kuras pamatā ir jonizētu proteīnu grupu saistīšana ar pretēji lādētām jonu apmaiņas sveķu grupām (nešķīstošiem polimēru materiāliem). Olbaltumvielu saistīšanās spēks ar sveķiem ir proporcionāls proteīna lādiņam. Proteīnus, kas adsorbēti uz jonu apmaiņas polimēru, var nomazgāt, palielinoties NaCl šķīdumu koncentrācijai; jo mazāks ir proteīna lādiņš, jo mazāka ir NaCl koncentrācija, kas nepieciešama, lai nomazgātu ar sveķu jonu grupām saistīto proteīnu.
Afinitātes hromatogrāfija- visspecifiskākā metode atsevišķu proteīnu izolēšanai. Proteīna ligands ir kovalenti piesaistīts inertam polimēram. Kad proteīna šķīdumu izlaiž caur kolonnu ar polimēru, uz kolonnas tiek adsorbēts tikai konkrētajam ligandam specifiskais proteīns, jo proteīns komplementāri saistās ar ligandu.
Dialīze- metode, ko izmanto zemas molekulmasas savienojumu atdalīšanai no izolēta proteīna šķīduma. Metodes pamatā ir olbaltumvielu nespēja iziet cauri puscaurlaidīgai membrānai, atšķirībā no zemas molekulmasas vielām. To izmanto proteīnu attīrīšanai no mazmolekulāriem piemaisījumiem, piemēram, sāļiem pēc izsālīšanas.
UZDEVUMI ĀRPUSSKODU DARBAM
1. Aizpildiet tabulu. 1.4.
1.4. tabula. Radniecīgo proteīnu - mioglobīna un hemoglobīna - struktūras un funkciju salīdzinošā analīze
a) atcerieties Mb un Hb aktīvā centra struktūru. Kāda loma ir hidrofobajiem aminoskābju radikāļiem šo proteīnu aktīvo centru veidošanā? Aprakstiet Mb un Hb aktīvā centra uzbūvi un ligandu piesaistes mehānismus tam. Kādu lomu spēlē His F 8 un His E 7 atlikumi Mv iHv aktīvā centra funkcionēšanā?
b) kādas jaunas īpašības, salīdzinot ar monomēru mioglobīnu, ir cieši saistītajam oligomēriskajam proteīnam hemoglobīnam? Izskaidrojiet kooperatīvo izmaiņu lomu protomēru konformācijā hemoglobīna molekulā, CO 2 un protonu koncentrācijas ietekmi uz hemoglobīna afinitāti pret skābekli, kā arī 2,3-BPG lomu Hb funkcijas allosteriskajā regulēšanā. .
2. Raksturojiet molekulāros šaperonus, pievēršot uzmanību to struktūras un funkciju attiecībām.
3. Kādi proteīni ir sagrupēti ģimenēs? Izmantojot imūnglobulīnu saimes piemēru, identificējiet līdzīgas šīs ģimenes proteīnu struktūras iezīmes un saistītās funkcijas.
4. Bioķīmiskiem un medicīniskiem nolūkiem bieži ir nepieciešami attīrīti atsevišķi proteīni. Paskaidrojiet, uz kādām olbaltumvielu fizikāli ķīmiskajām īpašībām balstās to atdalīšanas un attīrīšanas metodes.
PAŠKONTROLES UZDEVUMI
1. Izvēlieties pareizās atbildes.
Hemoglobīna funkcijas:
A. O 2 transportēšana no plaušām uz audiem B. H + transportēšana no audiem uz plaušām
B. Konstanta asins pH uzturēšana D. CO 2 transportēšana no plaušām uz audiem
D. CO 2 transportēšana no audiem uz plaušām
2. Izvēlieties pareizās atbildes. Ligandsα -protomērs Hb ir: A. Hēma
B. Skābeklis
B. CO G. 2,3-BPG
D. β-protomērs
3. Izvēlieties pareizās atbildes.
Hemoglobīns pretstatā mioglobīnam:
A. Ir kvartāra struktūra
B. Sekundāro struktūru attēlo tikai α-spirāles
B. Pieder pie kompleksiem proteīniem
D. Mijiedarbojas ar allosterisko ligandu D. Kovalenti saistīts ar hemu
4. Izvēlieties pareizās atbildes.
Hb afinitāte pret O2 samazinās:
A. Kad tiek pievienota viena O 2 molekula B. Kad tiek noņemta viena O 2 molekula
B. Mijiedarbojoties ar 2,3-BPG
D. Pievienojot protomēriem H + D. Kad 2,3-BPG koncentrācija samazinās
5. Match.
HB tipus raksturo:
A. Deoksi formā tas veido fibrilārus agregātus B. Satur divas α- un divas δ-ķēdes
B. Dominējošā Hb forma pieaugušo eritrocītos D. Satur hēmu ar Fe+ 3 aktīvajā centrā
D. Satur divas α- un divas γ-ķēdes 1. HbA 2.
6. Match.
Hb ligandi:
A. Saistās ar Hb allosteriskajā centrā
B. Tam ir ļoti augsta afinitāte pret Hb aktīvo vietu
B. Savienojot, tas palielina Hb afinitāti pret O 2 G. Oksidē Fe+ 2 līdz Fe+ 3
D. Veido kovalento saiti ar hisF8
7. Izvēlieties pareizās atbildes.
Pavadoņi:
A. Olbaltumvielas, kas atrodas visās šūnas daļās
B. Sintēze palielinās stresa apstākļos
B. Piedalīties denaturēto proteīnu hidrolīzē
D. Piedalīties proteīnu dabiskās konformācijas uzturēšanā
D. Izveidot organellas, kurās veidojas proteīnu konformācija
8. Sērkociņš. Imūnglobulīni:
A. Sekrēcijas forma ir pentamēra.
B. Ig klase, kas iekļūst placentas barjerā
B. Ig - tuklo šūnu receptors
D. Galvenā Ig klase, kas atrodas epitēlija šūnu izdalījumos. D. B-limfocītu receptors, kura aktivizēšana nodrošina šūnu proliferāciju
9. Izvēlieties pareizās atbildes.
Imūnglobulīni E:
A. Ražo makrofāgi B. Viņiem ir smagas ε ķēdes.
B. Iegults T-limfocītu membrānā
D. Darbojas kā membrānas antigēnu receptori uz tuklo šūnām un bazofīliem
D. Atbildīgs par alerģiskām reakcijām
10. Izvēlieties pareizās atbildes.
Olbaltumvielu atdalīšanas metode balstās uz to molekulmasas atšķirībām:
A. Gēla filtrēšana
B. Ultracentrifugēšana
B. Poliakrilamīda gēla elektroforēze D. Jonu apmaiņas hromatogrāfija
D. Afinitātes hromatogrāfija
11. Izvēlies pareizo atbildi.
Olbaltumvielu atdalīšanas metode balstās uz atšķirībām to šķīdībā ūdenī:
A. Gēla filtrēšana B. Izsālīšana
B. Jonu apmaiņas hromatogrāfija D. Afinitātes hromatogrāfija
D. Elektroforēze poliakrilamīda gēlā
ATBILŽU STANDARTI UZ “PAŠKONTROLES UZDEVUMIEM”
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
PAMATA NOTEIKUMI UN JĒDZIENI
1. Oligomērie proteīni, protomērs, olbaltumvielu ceturtdaļējā struktūra
2. Kooperatīvas izmaiņas protomēra konformācijā
3. Bora efekts
4. Olbaltumvielu funkciju allosteriskā regulēšana, allosteriskais centrs un allosteriskais efektors
5. Molekulārie šaperoni, karstuma šoka proteīni
6. Olbaltumvielu grupas (serīna proteāzes, imūnglobulīni)
7. IgM-, G-, E-, A-struktūras un funkcijas attiecības
8. Olbaltumvielu kopējais lādiņš, olbaltumvielu izoelektriskais punkts
9. Elektroforēze
10. Izsālīšana
11. Gēla filtrēšana
12. Jonu apmaiņas hromatogrāfija
13. Ultracentrifugēšana
14. Afinitātes hromatogrāfija
15. Asins plazmas proteīnu elektroforēze
UZDEVUMI KLASES DARBAM
1. Salīdziniet hemoglobīna (Hb) un mioglobīna (Mb) piesātinājuma pakāpju ar skābekli atkarību no tā daļējā spiediena audos
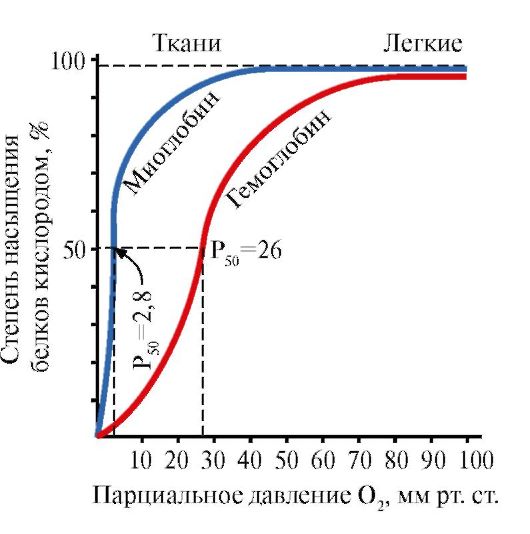 Rīsi. 1.34. Piesātinājuma atkarība Mv unNHskābeklis no tā daļējā spiediena
Rīsi. 1.34. Piesātinājuma atkarība Mv unNHskābeklis no tā daļējā spiediena
Lūdzu, ņemiet vērā, ka olbaltumvielu skābekļa piesātinājuma līkņu forma ir atšķirīga: mioglobīnam - hiperbola, hemoglobīnam - sigmoīda forma.
1. salīdziniet skābekļa parciālā spiediena vērtības, pie kurām Mb un Hb ir piesātināti ar O 2 par 50%. Kuram no šiem proteīniem ir lielāka afinitāte pret O 2?
2. Kādas Mb struktūras īpatnības nosaka tā augsto afinitāti pret O 2?
3. Kādas HB strukturālās īpašības ļauj tam atbrīvot O2 miera stāvoklī esošo audu kapilāros (pie salīdzinoši augsta O2 daļējā spiediena) un strauji palielināt šo izdalīšanos strādājošajos muskuļos? Kāda oligomēru proteīnu īpašība nodrošina šo efektu?
4. Aprēķiniet, kādu O 2 (%) skābekļa hemoglobīna daudzumu dod atpūtai un strādājošiem muskuļiem?
5. Izdarīt secinājumus par olbaltumvielu struktūras saistību ar tā funkciju.
2. Hemoglobīna izdalītā skābekļa daudzums kapilāros ir atkarīgs no katabolisko procesu intensitātes audos (Bora efekts). Kā izmaiņas audu metabolismā regulē Hb afinitāti pret O2? CO 2 un H+ ietekme uz Hb afinitāti pret O 2
 1. aprakstiet Bora efektu.
1. aprakstiet Bora efektu.
2. kādā virzienā notiek diagrammā redzamais process:
a) plaušu kapilāros;
b) audu kapilāros?
3. Kāda ir Bora efekta fizioloģiskā nozīme?
4. Kāpēc Hb mijiedarbība ar H+ vietās, kas atrodas tālu no hema, maina proteīna afinitāti pret O 2?
3. Hb afinitāte pret O2 ir atkarīga no tā liganda - 2,3-bisfosfoglicerāta, kas ir Hb afinitātes pret O2 allosteriskais regulators, koncentrācijas. Kāpēc ligandu mijiedarbība vietā, kas atrodas tālu no aktīvās vietas, ietekmē proteīna darbību? Kā 2,3-BPG regulē Hb afinitāti pret O2? Lai atrisinātu problēmu, atbildiet uz šādiem jautājumiem:
1. kur un no kā tiek sintezēts 2,3-bisfosfoglicerāts (2,3-BPG)? Uzrakstiet tās formulu, norādiet šīs molekulas lādiņu.
2. Ar kuru hemoglobīna formu (oksi vai deoksi) BPG mijiedarbojas un kāpēc? Kurā Hb molekulas daļā notiek mijiedarbība?
3. kādā virzienā notiek diagrammā redzamais process?
a) audu kapilāros;
b) plaušu kapilāros?
4. kur kompleksa koncentrācijai jābūt lielākai
Nv-2,3-BFG:
a) muskuļu kapilāros miera stāvoklī,
b) strādājošo muskuļu kapilāros (ar tādu pašu BPG koncentrāciju eritrocītos)?
5. Kā mainīsies HB afinitāte pret skābekli, cilvēkam pielāgojoties augstkalnu apstākļiem, ja palielinās BPG koncentrācija eritrocītos? Kāda ir šīs parādības fizioloģiskā nozīme?
4. 2,3-BPG iznīcināšana konservētu asiņu uzglabāšanas laikā pasliktina HB funkcijas. Kā mainīsies HB afinitāte pret O 2 konservētās asinīs, ja 2,3-BPG koncentrācija eritrocītos var samazināties no 8 līdz 0,5 mmol/l. Vai ir iespējams pārliet šādas asinis smagi slimiem pacientiem, ja 2,3-BPG koncentrācija tiek atjaunota ne agrāk kā pēc trim dienām? Vai ir iespējams atjaunot sarkano asins šūnu funkcijas, pievienojot asinīm 2,3-BPG?
5. Atcerieties vienkāršāko imūnglobulīna molekulu struktūru. Kādu lomu imūnglobulīni spēlē imūnsistēmā? Kāpēc Igs bieži tiek saukts par bivalentiem? Kā Ig struktūra ir saistīta ar to funkcijām? (Aprakstiet, izmantojot imūnglobulīnu klases piemēru.)
Olbaltumvielu fizikāli ķīmiskās īpašības un to atdalīšanas metodes.
6. Kā proteīna neto lādiņš ietekmē tā šķīdību?
a) nosaka peptīda kopējo lādiņu pie pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) kā mainīsies šī peptīda lādiņš pie pH >7, pH<7, рН <<7?
c) kāds ir proteīna izoelektriskais punkts (IEP) un kādā vidē tas atrodas?
Šī peptīda IET?
d) pie kāda pH vērtības tiks novērota šī peptīda zemākā šķīdība.
7. Kāpēc rūgušpiens, atšķirībā no svaiga piena, vārot “sabiezē” (t.i., piena proteīns kazeīns izgulsnējas)? Svaigā pienā kazeīna molekulām ir negatīvs lādiņš.
8. Gēla filtrēšanu izmanto atsevišķu proteīnu atdalīšanai. Maisījums, kas satur proteīnus A, B, C ar molekulmasu attiecīgi 160 000, 80 000 un 60 000, tika analizēts ar gēlfiltrāciju (1.35. att.). Uzbriedušās gēla granulas ir caurlaidīgas olbaltumvielām, kuru molekulmasa ir mazāka par 70 000. Kāds princips ir šīs atdalīšanas metodes pamatā? Kurš grafiks pareizi atspoguļo frakcionēšanas rezultātus? Norādiet secību, kādā proteīni A, B un C tiek atbrīvoti no kolonnas.
 Rīsi. 1.35. Gēla filtrācijas izmantošana proteīnu atdalīšanai
Rīsi. 1.35. Gēla filtrācijas izmantošana proteīnu atdalīšanai
9. Attēlā 1.36, A parāda elektroforēzes diagrammu uz papīra par veselīga cilvēka asins seruma proteīniem. Proteīna frakciju relatīvie daudzumi, kas iegūti, izmantojot šo metodi, ir: albumīni 54-58%, α1-globulīni 6-7%, α2-globulīni 8-9%, β-globulīni 13%, γ-globulīni 11-12%.
 Rīsi. 1.36. Vesela cilvēka (A) un pacienta (B) asins plazmas proteīnu elektroforēze uz papīra
Rīsi. 1.36. Vesela cilvēka (A) un pacienta (B) asins plazmas proteīnu elektroforēze uz papīra
I - γ-globulīni; II - β-globulīni; III -α 2 -globulīns; IV -α 2 -globulīns; V - albumīni
Daudzas slimības pavada kvantitatīvās izmaiņas seruma olbaltumvielu sastāvā (disproteinēmija). Šo izmaiņu raksturs tiek ņemts vērā, veicot diagnozi un novērtējot slimības smagumu un stadiju.
Izmantojot tabulā norādītos datus. 1.5, izdariet minējumu par slimību, ko raksturo elektroforētiskais profils, kas parādīts attēlā. 1.36.
1.5. tabula. Seruma proteīnu koncentrācijas izmaiņas patoloģijā
Olbaltumvielas ir organiskas vielas. Šiem lielmolekulāriem savienojumiem ir raksturīgs noteikts sastāvs, un pēc hidrolīzes tie sadalās aminoskābēs. Olbaltumvielu molekulas var būt dažādās formās, daudzas no tām sastāv no vairākām polipeptīdu ķēdēm. Informācija par proteīna uzbūvi tiek kodēta DNS, un proteīna molekulu sintēzes procesu sauc par translāciju.
Olbaltumvielu ķīmiskais sastāvs
Vidēji proteīns satur:
- 52% oglekļa;
- 7% ūdeņraža;
- 12% slāpekļa;
- 21% skābekļa;
- 3% sēra.
Olbaltumvielu molekulas ir polimēri. Lai izprastu to uzbūvi, ir jāzina, kas ir to monomēri – aminoskābes.
Aminoskābes
Tos parasti iedala divās kategorijās: pastāvīgi sastopamas un reizēm sastopamas. Pirmie ietver 18 un vēl 2 amīdus: asparagīnskābi un glutamīnskābi. Dažreiz tiek atrastas tikai trīs skābes.
Šīs skābes var klasificēt dažādi: pēc sānu ķēžu rakstura vai to radikāļu lādiņa tās var dalīt arī ar CN un COOH grupu skaitu.
Primārā proteīna struktūra
Aminoskābju maiņas secība olbaltumvielu ķēdē nosaka tās turpmākos organizācijas līmeņus, īpašības un funkcijas. Galvenais starp monomēriem ir peptīds. Tas veidojas, iegūstot ūdeņradi no vienas aminoskābes un OH grupu no citas.
Olbaltumvielu molekulas pirmais organizācijas līmenis ir tajā esošo aminoskābju secība, vienkārši ķēde, kas nosaka olbaltumvielu molekulu struktūru. Tas sastāv no “skeleta”, kam ir regulāra struktūra. Šī ir atkārtotā secība -NH-CH-CO-. Atsevišķas sānu ķēdes attēlo aminoskābju radikāļi (R), to īpašības nosaka proteīna struktūras sastāvu.
Pat ja olbaltumvielu molekulu struktūra ir vienāda, tās var atšķirties pēc īpašībām tikai tāpēc, ka to monomēriem ķēdē ir atšķirīga secība. Aminoskābju secību proteīnā nosaka gēni, un tā nosaka proteīnam noteiktas bioloģiskās funkcijas. Monomēru secība molekulās, kas atbild par vienu un to pašu funkciju, dažādās sugās bieži ir līdzīga. Šādas molekulas ir identiskas vai līdzīgas pēc organizācijas un pilda vienas un tās pašas funkcijas dažāda veida organismos – homologos proteīnos. Nākotnes molekulu struktūra, īpašības un funkcijas tiek noteiktas jau aminoskābju ķēdes sintēzes stadijā.
Dažas kopīgas iezīmes
Olbaltumvielu struktūra ir pētīta ilgu laiku, un to primārās struktūras analīze ļāva izdarīt dažus vispārinājumus. Lielākam proteīnu skaitam raksturīgs visu divdesmit aminoskābju klātbūtne, no kurām īpaši daudz ir glicīna, alanīna, glutamīna un maz triptofāna, arginīna, metionīna un histidīna. Vienīgie izņēmumi ir dažas olbaltumvielu grupas, piemēram, histoni. Tie ir nepieciešami DNS iepakošanai un satur daudz histidīna.
Jebkāda veida organismu kustību (muskuļu darbs, protoplazmas kustība šūnā, skropstu mirgošana vienšūņos utt.) veic olbaltumvielas. Olbaltumvielu struktūra ļauj tiem pārvietoties un veidot šķiedras un gredzenus.

Transporta funkcija ir tāda, ka daudzas vielas tiek transportētas cauri šūnu membrānai ar īpašiem nesējproteīniem.
Šo polimēru hormonālā loma ir uzreiz skaidra: vairāki hormoni pēc struktūras ir proteīni, piemēram, insulīns, oksitocīns.
Rezerves funkciju nosaka tas, ka olbaltumvielas spēj veidot nogulsnes. Piemēram, olu valgumīns, piena kazeīns, augu sēklu proteīni – tie uzglabā lielu daudzumu barības vielu.
Visas cīpslas, locītavu locītavas, skeleta kaulus un nagus veido olbaltumvielas, kas mūs noved pie to nākamās funkcijas - atbalsta.
Olbaltumvielu molekulas ir receptori, kas selektīvi atpazīst noteiktas vielas. Glikoproteīni un lektīni ir īpaši pazīstami ar šo lomu.
Svarīgākie imunitātes faktori ir antivielas, un to izcelsme ir olbaltumvielas. Piemēram, asins recēšanas procesa pamatā ir izmaiņas proteīna fibrinogēnā. Barības vada un kuņģa iekšējās sienas ir pārklātas ar gļotādu proteīnu - licīnu - aizsargkārtu. Toksīni arī pēc izcelsmes ir olbaltumvielas. Ādas pamats, kas aizsargā dzīvnieku ķermeni, ir kolagēns. Visas šīs olbaltumvielu funkcijas ir aizsargājošas.

Nu, pēdējā funkcija ir regulējoša. Ir olbaltumvielas, kas kontrolē genoma darbību. Tas ir, tie regulē transkripciju un tulkošanu.
Neatkarīgi no tā, cik svarīga loma ir olbaltumvielām, proteīnu struktūru zinātnieki atklāja diezgan sen. Un tagad viņi atklāj jaunus veidus, kā izmantot šīs zināšanas.
2. biļete. 1. Lipīdu rakstura būtiskie uztura faktori. Daži lipīdi cilvēka organismā netiek sintezēti, tāpēc tie ir būtiski uztura faktori. Tie ietver taukskābes ar divām vai vairākām dubultsaitēm (poliēns) - neaizstājamās taukskābes. Dažas no šīm skābēm ir substrāti vietējo hormonu - eikozanoīdu sintēzei (8.10. tēma).
Taukos šķīstošie vitamīni veic dažādas funkcijas: A vitamīns piedalās redzes, kā arī šūnu augšanas un diferenciācijas procesā; ir pierādīta tā spēja kavēt noteiktu veidu audzēju augšanu; K vitamīns piedalās asins sarecēšanā; D vitamīns piedalās kalcija metabolisma regulēšanā; E vitamīns- antioksidants, kavē brīvo radikāļu veidošanos un tādējādi novērš šūnu bojājumus lipīdu peroksidācijas rezultātā.
Dokuments
2. Olbaltumvielu struktūra un strukturālās organizācijas līmeņi
Ir četri proteīnu strukturālās organizācijas līmeņi: primārais, sekundārais, terciārais un kvartārais. Katram līmenim ir savas īpašības.
Primārā proteīna struktūra
Olbaltumvielu primārā struktūra ir lineāra aminoskābju polipeptīdu ķēde, kas savienota viena ar otru ar peptīdu saitēm. Primārā struktūra ir vienkāršākais proteīna molekulas strukturālās organizācijas līmenis. Augstu stabilitāti tai piešķir kovalentās peptīdu saites starp vienas aminoskābes α-aminogrupu un citas aminoskābes α-karboksilgrupu. [rādīt].
Ja prolīna vai hidroksiprolīna imino grupa ir iesaistīta peptīdu saites veidošanā, tad tai ir cita forma [rādīt].
Kad šūnās veidojas peptīdu saites, vispirms tiek aktivizēta vienas aminoskābes karboksilgrupa, un pēc tam tā apvienojas ar citas aminoskābes aminogrupu. Polipeptīdu laboratoriskā sintēze tiek veikta aptuveni tādā pašā veidā.
Peptīdu saite ir atkārtots polipeptīdu ķēdes fragments. Tam ir vairākas pazīmes, kas ietekmē ne tikai primārās struktūras formu, bet arī augstākus polipeptīdu ķēdes organizācijas līmeņus:

koplanaritāte - visi peptīdu grupā iekļautie atomi atrodas vienā plaknē;
spēja pastāvēt divās rezonanses formās (keto vai enola formā);
aizvietotāju transpozīcija attiecībā pret C-N saiti;
spēja veidot ūdeņraža saites, un katra no peptīdu grupām var veidot divas ūdeņraža saites ar citām grupām, ieskaitot peptīdu.
Izņēmums ir peptīdu grupas, kas ietver prolīna vai hidroksiprolīna aminogrupu. Tie spēj veidot tikai vienu ūdeņraža saiti (skatīt iepriekš). Tas ietekmē proteīna sekundārās struktūras veidošanos. Polipeptīdu ķēde apgabalā, kur atrodas prolīns vai hidroksiprolīns, viegli izliecas, jo to, kā parasti, nenotur otrā ūdeņraža saite.
Peptīdu un polipeptīdu nomenklatūra. Peptīdu nosaukumu veido to aminoskābju nosaukumi. Divas aminoskābes veido dipeptīdu, trīs veido tripeptīdu, četras veido tetrapeptīdu utt. Katram jebkura garuma peptīdam vai polipeptīda ķēdei ir N-gala aminoskābe, kas satur brīvu aminogrupu, un C-gala aminoskābe, kas satur brīvu karboksilgrupu. grupai. Nosaucot polipeptīdus, visas aminoskābes tiek uzskaitītas secīgi, sākot ar N-galu, aizstājot to nosaukumos, izņemot C-galu, sufiksu -in ar -il (jo peptīdu aminoskābēm vairs nav karboksilgrupa, bet karbonilgrupa). Piemēram, nosaukums, kas parādīts attēlā. 1 tripeptīds - leuc dūņas fenilalāns dūņas treons iekšā.
Olbaltumvielu primārās struktūras iezīmes. Polipeptīdu ķēdes mugurkaulā stingras struktūras (plakanas peptīdu grupas) mijas ar relatīvi mobiliem reģioniem (-CHR), kas spēj griezties ap saitēm. Šādas polipeptīdu ķēdes struktūras iezīmes ietekmē tās telpisko izvietojumu.
Olbaltumvielu sekundārā struktūra
Sekundārā struktūra ir veids, kā polipeptīdu ķēdi salocīt sakārtotā struktūrā, veidojot ūdeņraža saites starp vienas ķēdes peptīdu grupām vai blakus esošajām polipeptīdu ķēdēm. Atbilstoši to konfigurācijai sekundārās struktūras iedala spirālveida (α-spirāle) un slāņaini salocītās (β-struktūra un krusteniskā β-forma).


α-spirāle. Tas ir sekundāras proteīna struktūras veids, kas izskatās kā regulāra spirāle, kas veidojas starppeptīdu ūdeņraža saitēm vienā polipeptīdu ķēdē. α-spirāles struktūras modeli (2. att.), kurā ņemtas vērā visas peptīdu saites īpašības, ierosināja Polings un Korijs. Galvenās α-spirāles iezīmes:
polipeptīdu ķēdes spirālveida konfigurācija ar spirālveida simetriju;
ūdeņraža saišu veidošanās starp katras pirmās un ceturtās aminoskābes atlikuma peptīdu grupām;
spirālveida pagriezienu regularitāte;
visu aminoskābju atlikumu ekvivalence α-spirālē neatkarīgi no to sānu radikāļu struktūras;
aminoskābju sānu radikāļi nepiedalās α-spirāles veidošanā.
Ārēji α-spirāle izskatās kā nedaudz izstiepta elektriskās plīts spirāle. Ūdeņraža saišu regularitāte starp pirmo un ceturto peptīdu grupu nosaka polipeptīdu ķēdes pagriezienu regularitāti. Viena pagrieziena augstums jeb α-spirāles solis ir 0,54 nm; tajā ir 3,6 aminoskābju atlikumi, t.i., katrs aminoskābes atlikums pārvietojas pa asi (vienas aminoskābes atlikuma augstums) par 0,15 nm (0,54:3,6 = 0,15 nm), kas ļauj runāt par visu aminoskābju atlikumu ekvivalenci. α-spirālē. α-spirāles regularitātes periods ir 5 apgriezieni vai 18 aminoskābju atlikumi; viena perioda garums ir 2,7 nm. Rīsi. 3. Pauling-Corey a-helix modelis
β-struktūra. Tas ir sekundāras struktūras veids, kam ir nedaudz izliekta polipeptīdu ķēdes konfigurācija un ko veido starppeptīdu ūdeņraža saites atsevišķās vienas polipeptīdu ķēdes vai blakus esošo polipeptīdu ķēdes sekcijās. To sauc arī par slāņainu kroku struktūru. Ir β-struktūru šķirnes. Ierobežotos slāņu reģionus, ko veido viena proteīna polipeptīda ķēde, sauc par krustenisko β formu (īsu β struktūru). Starp polipeptīdu ķēdes cilpu peptīdu grupām veidojas ūdeņraža saites krusteniskā β formā. Cits tips - pilnīga β-struktūra - ir raksturīga visai polipeptīdu ķēdei, kurai ir iegarena forma un ko notur interpeptīdu ūdeņraža saites starp blakus esošajām paralēlajām polipeptīdu ķēdēm (3. att.). Šī struktūra atgādina akordeona plēšas. Turklāt ir iespējami β-struktūru varianti: tos var veidot paralēlas ķēdes (polipeptīdu ķēžu N-gala gali ir vērsti vienā virzienā) un antiparalēli (N-gala gali ir vērsti dažādos virzienos). Viena slāņa sānu radikāļi ir novietoti starp cita slāņa sānu radikāļiem.
Olbaltumvielās ūdeņraža saišu pārkārtošanās dēļ ir iespējamas pārejas no α-struktūrām uz β-struktūrām un atpakaļ. Regulāru starppeptīdu ūdeņraža saišu vietā gar ķēdi (pateicoties tam, polipeptīdu ķēde ir savīta spirālē), spirālveida posmi atritinās un ūdeņraža saites noslēdzas starp iegarenajiem polipeptīdu ķēžu fragmentiem. Šī pāreja ir atrodama keratīnā, matu proteīnā. Mazgājot matus ar sārmainiem mazgāšanas līdzekļiem, β-keratīna spirālveida struktūra viegli tiek iznīcināta un tas pārvēršas α-keratīnā (cirtaini mati iztaisnojas).
Proteīnu regulāro sekundāro struktūru (α-spirāles un β-struktūru) iznīcināšanu pēc analoģijas ar kristāla kušanu sauc par polipeptīdu “kušanu”. Šajā gadījumā ūdeņraža saites tiek pārtrauktas, un polipeptīdu ķēdes veidojas nejauša mudžeklī. Līdz ar to sekundāro struktūru stabilitāti nosaka interpeptīdu ūdeņraža saites. Citu veidu saites tajā gandrīz nepiedalās, izņemot disulfīda saites gar polipeptīdu ķēdi cisteīna atlikumu vietās. Īsie peptīdi tiek slēgti ciklos disulfīda saišu dēļ. Daudzi proteīni satur gan α-spirāles reģionus, gan β-struktūras. Gandrīz nav dabisko olbaltumvielu, kas sastāv no 100% α-spirāles (izņēmums ir paramiozīns, muskuļu proteīns, kas ir 96-100% α-spirāles), savukārt sintētiskajiem polipeptīdiem ir 100% spirāle.
Citām olbaltumvielām ir dažāda saritināšanās pakāpe. Augsts α-spirālveida struktūru biežums tiek novērots paramiozīnā, mioglobīnā un hemoglobīnā. Turpretim tripsīnā, ribonukleāzē, ievērojama polipeptīdu ķēdes daļa ir salocīta slāņainās β-struktūrās. Atbalstaudu olbaltumvielām: keratīnam (matu, vilnas olbaltumvielām), kolagēnam (cīpslu, ādas olbaltumvielām), fibroīnam (dabiskā zīda olbaltumvielām) ir polipeptīdu ķēžu β-konfigurācija. Proteīnu polipeptīdu ķēžu dažādā spirāles pakāpe norāda uz to, ka acīmredzami pastāv spēki, kas daļēji izjauc spirāli vai “pārtrauc” regulāro polipeptīdu ķēdes locīšanu. Iemesls tam ir proteīna polipeptīdu ķēdes kompaktāka locīšana noteiktā tilpumā, t.i., terciārā struktūrā.
Olbaltumvielu terciārā struktūra
Olbaltumvielu terciārā struktūra ir veids, kā polipeptīdu ķēde tiek izkārtota telpā. Pamatojoties uz to terciārās struktūras formu, olbaltumvielas galvenokārt iedala lodveida un fibrilārajos. Globulārajiem proteīniem visbiežāk ir elipsoīda forma, un fibrilāriem (vītņveidīgiem) proteīniem ir iegarena forma (stieņa vai vārpstas forma).
Tomēr proteīnu terciārās struktūras konfigurācija vēl nedod pamatu domāt, ka fibrilārajiem proteīniem ir tikai β-struktūra, bet globulārajiem proteīniem ir α-spirālveida struktūra. Ir fibrilāri proteīni, kuriem ir spirālveida, nevis slāņaina, salocīta sekundārā struktūra. Piemēram, α-keratīns un paramiozīns (gliemju obturatora muskuļa proteīns), tropomiozīni (skeleta muskuļu olbaltumvielas) pieder pie fibrilāriem proteīniem (tam ir stieņa forma), un to sekundārā struktūra ir α-spirāle; turpretim globulārie proteīni var saturēt lielu skaitu β-struktūru.
Lineāras polipeptīdu ķēdes spiralizācija samazina tās izmēru aptuveni 4 reizes; un iepakošana terciārajā struktūrā padara to desmitiem reižu kompaktāku nekā sākotnējā ķēde.
Saites, kas stabilizē proteīna terciāro struktūru. Saitēm starp aminoskābju sānu radikāļiem ir nozīme terciārās struktūras stabilizācijā. Šos savienojumus var iedalīt:
stiprs (kovalents) [rādīt].
vājš (polārais un van der Vāls) [rādīt].
Daudzas saites starp aminoskābju sānu radikāļiem nosaka proteīna molekulas telpisko konfigurāciju.
Olbaltumvielu terciārās struktūras organizācijas iezīmes. Polipeptīdu ķēdes terciārās struktūras konformāciju nosaka tajā iekļauto aminoskābju sānu radikāļu īpašības (kurām nav manāmas ietekmes uz primāro un sekundāro struktūru veidošanos) un mikrovide, t.i. vidi. Salocīta proteīna polipeptīdu ķēde mēdz iegūt enerģētiski labvēlīgu formu, ko raksturo brīvās enerģijas minimums. Tāpēc nepolāras R-grupas, “izvairoties no ūdens”, it kā veido proteīna terciārās struktūras iekšējo daļu, kurā atrodas galvenā polipeptīdu ķēdes hidrofobo atlikumu daļa. Proteīna globulas centrā gandrīz nav ūdens molekulu. Aminoskābes polārās (hidrofilās) R grupas atrodas šī hidrofobā kodola ārpusē, un tās ieskauj ūdens molekulas. Polipeptīdu ķēde ir sarežģīti saliekta trīsdimensiju telpā. Kad tas saliecas, tiek traucēta sekundārā spirālveida konformācija. Ķēde “pārtrūkst” vājās vietās, kur atrodas prolīns vai hidroksiprolīns, jo šīs aminoskābes ķēdē ir mobilākas, veidojot tikai vienu ūdeņraža saiti ar citām peptīdu grupām. Vēl viena līkuma vieta ir glicīns, kuram ir neliela R grupa (ūdeņradis). Tāpēc citu aminoskābju R-grupas, ja tās ir sakrautas, mēdz aizņemt brīvo vietu glicīna atrašanās vietā. Vairākas aminoskābes - alanīns, leicīns, glutamāts, histidīns - veicina stabilu spirālveida struktūru saglabāšanu proteīnā, un tādas kā metionīns, valīns, izoleicīns, asparagīnskābe veicina β-struktūru veidošanos. Olbaltumvielu molekulā ar terciāru konfigurāciju ir reģioni α-spirāles (spirālveida), β-struktūru (slāņaini) un nejaušas spoles formā. Tikai pareizais proteīna telpiskais izvietojums padara to aktīvu; tā pārkāpums izraisa izmaiņas olbaltumvielu īpašībās un bioloģiskās aktivitātes zudumu.
Kvartārā proteīna struktūra
Olbaltumvielām, kas sastāv no vienas polipeptīdu ķēdes, ir tikai terciārā struktūra. Tajos ietilpst mioglobīns - muskuļu audu proteīns, kas iesaistīts skābekļa saistīšanā, virkne enzīmu (lizocīms, pepsīns, tripsīns utt.). Tomēr daži proteīni ir veidoti no vairākām polipeptīdu ķēdēm, no kurām katrai ir terciārā struktūra. Šādām olbaltumvielām ir ieviests ceturkšņa struktūras jēdziens, kas ir vairāku polipeptīdu ķēžu ar terciāru struktūru organizēšana vienā funkcionālā proteīna molekulā. Tādu proteīnu ar kvartāru struktūru sauc par oligomēru, bet tā polipeptīdu ķēdes ar terciāro struktūru sauc par protomēriem vai apakšvienībām (4. att.).

Kvartārajā organizācijas līmenī olbaltumvielas saglabā terciārās struktūras pamatkonfigurāciju (globulāru vai fibrilāru). Piemēram, hemoglobīns ir proteīns ar kvartāru struktūru un sastāv no četrām apakšvienībām. Katra no apakšvienībām ir lodveida proteīns, un kopumā hemoglobīnam ir arī lodveida konfigurācija. Matu un vilnas proteīniem – keratīniem, kas pēc terciārās struktūras saistīti ar fibrilāriem proteīniem, ir fibrilāra konformācija un ceturtdaļīga struktūra.
Olbaltumvielu kvartārās struktūras stabilizācija. Visas olbaltumvielas, kurām ir kvartāra struktūra, tiek izolētas atsevišķu makromolekulu veidā, kas nesadalās apakšvienībās. Kontakti starp apakšvienību virsmām ir iespējami tikai aminoskābju atlikumu polāro grupu dēļ, jo katras polipeptīdu ķēdes terciārās struktūras veidošanās laikā nepolāro aminoskābju sānu radikāļi (kas veido lielāko daļu visas proteinogēnās aminoskābes) ir paslēptas apakšvienības iekšpusē. Starp to polārajām grupām veidojas daudzas jonu (sāls), ūdeņraža un dažos gadījumos disulfīda saites, kas stingri notur apakšvienības organizēta kompleksa veidā. Vielu, kas sarauj ūdeņraža saites, vai vielu, kas samazina disulfīda tiltus, izmantošana izraisa protomēru sadalīšanos un proteīna ceturtdaļas struktūras iznīcināšanu. Tabulā 1 apkopoti dati par saitēm, kas stabilizē dažādus proteīna molekulas organizācijas līmeņus [rādīt].
Dažu fibrilāro proteīnu strukturālās organizācijas iezīmes
Fibrilāro proteīnu strukturālajai organizācijai ir vairākas iezīmes, salīdzinot ar globulārajiem proteīniem. Šīs īpašības var redzēt keratīna, fibroīna un kolagēna piemērā. Keratīni pastāv α- un β-konformācijās. α-Keratīniem un fibroīnam ir slāņaini salocīta sekundārā struktūra, tomēr keratīnā ķēdes ir paralēlas, bet fibroīnā tās ir pretparalēlas (sk. 3. att.); Turklāt keratīns satur starpķēžu disulfīda saites, savukārt fibroīnā to nav. Disulfīda saišu pārrāvums izraisa polipeptīdu ķēžu atdalīšanu keratīnos. Gluži pretēji, maksimālā disulfīda saišu skaita veidošanās keratīnos, iedarbojoties ar oksidētājiem, rada spēcīgu telpisko struktūru. Kopumā fibrilārajos proteīnos, atšķirībā no globulārajiem proteīniem, dažreiz ir grūti stingri nošķirt dažādus organizācijas līmeņus. Ja pieņemam (kā globulāram proteīnam), ka terciārā struktūra jāveido, izliekot telpā vienu polipeptīdu ķēdi, bet ceturtdaļējā struktūra vairākām ķēdēm, tad fibrilārajos proteīnos jau sekundārās struktūras veidošanās laikā ir iesaistītas vairākas polipeptīdu ķēdes. . Tipisks fibrilāra proteīna piemērs ir kolagēns, kas ir viens no visbiežāk sastopamajiem proteīniem cilvēka organismā (apmēram 1/3 no visu olbaltumvielu masas). Tas ir atrodams audos, kuriem ir augsta izturība un zema stiepjamība (kaulos, cīpslās, ādā, zobos utt.). Kolagēnā trešā daļa aminoskābju atlikumu ir glicīns, un apmēram ceturtā daļa vai nedaudz vairāk ir prolīns vai hidroksiprolīns.

Izolētā kolagēna polipeptīda ķēde (primārā struktūra) izskatās kā pārrauta līnija. Tas satur apmēram 1000 aminoskābes, un tā molekulmasa ir aptuveni 10 5 (5. att., a, b). Polipeptīdu ķēde ir veidota no atkārtotas aminoskābju trio (tripleta) ar šādu sastāvu: gly-A-B, kur A un B ir jebkuras aminoskābes, kas nav glicīns (visbiežāk prolīns un hidroksiprolīns). Kolagēna polipeptīdu ķēdes (vai α-ķēdes) sekundāro un terciāro struktūru veidošanās laikā (5. att., c un d) nevar radīt tipiskas α-spirāles ar spirālveida simetriju. Prolīns, hidroksiprolīns un glicīns (antihelikālās aminoskābes) to traucē. Tāpēc trīs α-ķēdes veido it kā savītas spirāles, piemēram, trīs pavedieni, kas aptinās ap cilindru. Trīs spirālveida α ķēdes veido atkārtotu kolagēna struktūru, ko sauc par tropokolagēnu (5.d attēls). Tropokolagēns savā organizācijā ir kolagēna terciārā struktūra. Plakanie prolīna un hidroksiprolīna gredzeni, kas regulāri mainās gar ķēdi, piešķir tai stingrību, tāpat kā starpķēžu saites starp tropokolagēna α-ķēdēm (tāpēc kolagēns ir izturīgs pret stiepšanos). Tropokolagēns būtībā ir kolagēna fibrilu apakšvienība. Tropokolagēna apakšvienību ieklāšana kolagēna kvartārajā struktūrā notiek pakāpeniski (5.e att.).
Kolagēna struktūru stabilizācija notiek starpķēžu ūdeņraža, jonu un van der Vālsa saišu un neliela skaita kovalento saišu dēļ.
Kolagēna α-ķēdēm ir dažādas ķīmiskās struktūras. Ir dažāda veida α 1 ķēdes (I, II, III, IV) un α 2 ķēdes. Atkarībā no tā, kuras α 1 un α 2 ķēdes ir iesaistītas tropokolagēna trīspavedienu spirāles veidošanā, izšķir četrus kolagēna veidus:
pirmais veids - divas α 1 (I) un viena α 2 ķēde;
otrais veids - trīs α 1 (II) ķēdes;
trešais tips - trīs α 1 (III) ķēdes;
ceturtais tips - trīs α 1 (IV) ķēdes.
Visbiežāk sastopamais kolagēns ir pirmā tipa: tas atrodas kaulaudos, ādā, cīpslās; Otrā tipa kolagēns ir atrodams skrimšļa audos utt. Viena veida audos var būt dažādi kolagēna veidi.
Sakārtotā kolagēna struktūru agregācija, to stingrība un inertums nodrošina kolagēna šķiedru augsto izturību. Kolagēna proteīni satur arī ogļhidrātu komponentus, t.i., tie ir proteīna-ogļhidrātu kompleksi.
Kolagēns ir ārpusšūnu proteīns, ko veido saistaudu šūnas, kas atrodas visos orgānos. Tāpēc ar kolagēna bojājumiem (vai tā veidošanās traucējumiem) rodas vairāki orgānu saistaudu atbalsta funkciju pārkāpumi.
Alfa polipeptīdu ķēde beidzas ar aminoskābju kombināciju valīns-leicīns, bet beta polipeptīdu ķēde beidzas ar kombināciju valīns-histidīns-leicīns. Alfa un beta polipeptīdu ķēdes hemoglobīna molekulā nav sakārtotas lineāri, tā ir primārā struktūra. Sakarā ar intramolekulāro spēku esamību, polipeptīdu ķēdes ir savītas alfa-spirāles spirāles formā (sekundārā struktūra), kas raksturīga olbaltumvielām. Pati alfa-spirāles spirāle telpiski izliecas katrai alfa un beta polipeptīdu ķēdei, veidojot olveida formas pinumus (terciārā struktūra). Atsevišķas polipeptīdu ķēžu alfa-spirāles spirāles daļas apzīmētas ar latīņu burtiem no A līdz H. Visas četras terciārās izliektās alfa un beta polipeptīdu ķēdes atrodas telpiski noteiktās attiecībās - ceturtdaļējā struktūrā. Tos savieno nevis reālas ķīmiskas saites, bet gan starpmolekulārie spēki.
Izrādījās, ka cilvēkiem ir trīs galvenie normālā hemoglobīna veidi: embrionālais – U, augļa – F un pieaugušo hemoglobīns – A. HbU (nosaukts pēc vārda uterus sākuma burta) rodas embrijā no 7 līdz 12 dzīves nedēļām, tad tas pazūd un parādās augļa hemoglobīns, kas pēc trešā mēneša ir galvenais augļa hemoglobīns. Pēc tam pēc angļu vārda “adult” sākuma burta pakāpeniski parādās parasts pieaugušo hemoglobīns, ko sauc par HbA. Augļa hemoglobīna daudzums pakāpeniski samazinās, tā ka dzimšanas brīdī 80% hemoglobīna ir HbA un tikai 20% ir HbF. Pēc piedzimšanas augļa hemoglobīna līmenis turpina samazināties un 2–3 dzīves gados ir tikai 1–2%. Tikpat daudz augļa hemoglobīna ir pieaugušam cilvēkam. HbF daudzums, kas pārsniedz 2%, tiek uzskatīts par patoloģisku pieaugušajiem un bērniem, kas vecāki par 3 gadiem.
Papildus parastajiem hemoglobīna veidiem pašlaik ir zināmi vairāk nekā 50 patoloģiskie varianti. Tie vispirms tika nosaukti ar latīņu burtiem. Hemoglobīna tipu apzīmējumā burta B nav, jo sākotnēji tas apzīmēja HbS.
Hemoglobīns (Hb)- hromoproteīns, kas atrodas sarkanajās asins šūnās un ir iesaistīts skābekļa transportēšanā uz audiem. Hemoglobīnu pieaugušajiem sauc par hemoglobīnu A (Hb A). Tās molekulmasa ir aptuveni 65 000 Da. Hb A molekulai ir kvartāra struktūra, un tā ietver četras apakšvienības - polipeptīdu ķēdes (apzīmētas ar α1, α2, β1 un β2, no kurām katra ir saistīta ar hēmu.
Atcerieties, ka hemoglobīns ir allosterisks proteīns, tā molekulas var atgriezeniski mainīties no vienas konformācijas uz otru. Tas maina proteīna afinitāti pret ligandiem. Konformāciju ar vismazāko afinitāti pret ligandu sauc par sasprindzinājumu jeb T-konformāciju. Konformāciju ar vislielāko afinitāti pret ligandu sauc par atslābinātu vai R-konformāciju.
Hemoglobīna molekulas R- un T-konformācijas atrodas dinamiskā līdzsvara stāvoklī:
Dažādi vides faktori var novirzīt šo līdzsvaru vienā vai otrā virzienā. Allosteriskie regulatori, kas ietekmē Hb afinitāti pret O2, ir: 1) skābeklis; 2) H+ koncentrācija (vidēja pH); 3) oglekļa dioksīds (CO2); 4) 2,3-difosfoglicerāts (DPG). Skābekļa molekulas piesaiste vienai no hemoglobīna apakšvienībām veicina saspringtas konformācijas pāreju uz relaksētu un palielina afinitāti pret skābekli citām tās pašas hemoglobīna molekulas apakšvienībām. Šo parādību sauc par sadarbības efektu. Hemoglobīna saistīšanās ar skābekli sarežģīto raksturu atspoguļo hemoglobīna O2 piesātinājuma līkne, kurai ir S forma (3.1. attēls).

3.1.attēls. Mioglobīna (1) un hemoglobīna (2) skābekļa piesātinājuma līknes.
Molekulārās formas hemoglobīns atšķiras viens no otra polipeptīdu ķēžu struktūrā. Piemērs šādai hemoglobīna daudzveidībai, kas pastāv fizioloģiskos apstākļos, ir augļa hemoglobīns (HbF), kas atrodas asinīs cilvēka embrija attīstības stadijā. Atšķirībā no HbA, tā molekulā ir 2 α- un 2 γ-ķēdes (tas ir, β-ķēdes tiek aizstātas ar γ-ķēdēm). Šādam hemoglobīnam ir lielāka afinitāte pret skābekli. Tas ļauj embrijam caur placentu saņemt skābekli no mātes asinīm. Drīz pēc piedzimšanas HbF bērna asinīs tiek aizstāts ar HbA.
Patoloģiska vai patoloģiska hemoglobīna piemērs ir jau minētais (sk. 2.4.) hemoglobīns S, kas konstatēts pacientiem ar sirpjveida šūnu anēmiju. Kā jūs jau zināt, tas atšķiras no hemoglobīna A, aizstājot glutamātu ar valīnu β-ķēdēs. Šī aminoskābju aizstāšana izraisa HbS šķīdības samazināšanos ūdenī un tā afinitātes pret O2 samazināšanos.
Olbaltumvielu ķīmisko struktūru attēlo alfa aminoskābes, kas savienotas ķēdē caur peptīdu saiti. Dzīvos organismos sastāvu nosaka ģenētiskais kods. Sintēzes procesā vairumā gadījumu tiek izmantotas 20 standarta tipa aminoskābes. To daudzās kombinācijas veido olbaltumvielu molekulas ar visdažādākajām īpašībām. Aminoskābju atlikumi bieži tiek pakļauti pēctranslācijas modifikācijām. Tās var rasties, pirms proteīns sāk pildīt savas funkcijas, un tās darbības laikā šūnā. Dzīvos organismos vairākas molekulas bieži veido sarežģītus kompleksus. Piemērs ir fotosintētiskā asociācija.
Savienojumu mērķis
Olbaltumvielas tiek uzskatītas par svarīgu cilvēku un dzīvnieku uztura sastāvdaļu, jo to organisms nespēj sintezēt visas nepieciešamās aminoskābes. Dažiem no tiem vajadzētu būt ar olbaltumvielu pārtiku. Galvenie savienojumu avoti ir gaļa, rieksti, piens, zivis un graudi. Mazākā mērā olbaltumvielas ir dārzeņos, sēnēs un ogās. Gremošanas laikā ar fermentu palīdzību patērētās olbaltumvielas tiek sadalītas aminoskābēs. Tie jau tiek izmantoti pašu olbaltumvielu biosintēzē organismā vai tiek tālāk sadalīti, lai iegūtu enerģiju.

Vēsturiska atsauce
Insulīna proteīna struktūras secību vispirms noteica Frederijs Sengers. Par savu darbu viņš saņēma Nobela prēmiju 1958. gadā. Sanger izmantoja sekvencēšanas metodi. Izmantojot rentgenstaru difrakciju, pēc tam (1950. gadu beigās) tika iegūtas mioglobīna un hemoglobīna trīsdimensiju struktūras. Darbu veica Džons Kendrū un Makss Perucs.
Olbaltumvielu molekulu struktūra
Tas ietver lineāros polimērus. Tie savukārt sastāv no alfa aminoskābju atlikumiem, kas ir monomēri. Turklāt proteīna struktūrā var būt neaminoskābju komponenti un modificētas aminoskābju atliekas. Apzīmējot sastāvdaļas, tiek izmantoti 1 vai 3 burtu saīsinājumi. Savienojumu, kas satur no diviem līdz vairākiem desmitiem atlieku, bieži sauc par "polipeptīdu". Vienas aminoskābes alfa-karboksilgrupas mijiedarbības rezultātā ar citas aminoskābes alfa-aminogrupu rodas saites (proteīna struktūras veidošanās laikā). Atkarībā no tā, kura aminoskābju atlikuma grupa ir brīva, izšķir savienojuma C- un N-galus: -COOH vai -NH 2 . Proteīnu sintēzes procesā uz ribosomas pirmais gala atlikums parasti ir metionīna atlikums; nākamie ir pievienoti iepriekšējo C-galam.

Organizācijas līmeņi
Tos ierosināja Lindrems-Langs. Neskatoties uz to, ka šis sadalījums tiek uzskatīts par nedaudz novecojušu, tas joprojām tiek izmantots. Tika ierosināts izdalīt četrus savienojuma organizācijas līmeņus. Olbaltumvielu molekulas primāro struktūru nosaka ģenētiskais kods un gēna īpašības. Augstāku līmeni raksturo veidošanās olbaltumvielu locīšanas laikā. Olbaltumvielu telpisko struktūru kopumā nosaka aminoskābju ķēde. Tomēr tas ir diezgan nestabils. To var ietekmēt ārēji faktori. Šajā sakarā pareizāk ir runāt par savienojuma uzbūvi, kas ir vislabvēlīgākā un enerģētiski vēlamā.
1. līmenis
To attēlo polipeptīdu ķēdes aminoskābju atlikumu secība. Parasti to apraksta, izmantojot viena vai trīs burtu apzīmējumus. Olbaltumvielu primāro struktūru raksturo stabilas aminoskābju atlieku kombinācijas. Viņi veic noteiktus uzdevumus. Šādi "konservatīvie motīvi" saglabājas sugu evolūcijas laikā. Tos bieži var izmantot, lai prognozētu nezināma proteīna problēmu. Novērtējot līdzības pakāpi (homoloģiju) dažādu organismu aminoskābju ķēdēs, iespējams noteikt evolūcijas attālumu, kas veidojas starp taksoniem, kas veido šos organismus. Olbaltumvielu primāro struktūru nosaka ar sekvencēšanu vai tās mRNS sākotnējo kompleksu, izmantojot ģenētisko kodu tabulu.

Ķēdes sekcijas lokāla pasūtīšana
Tas ir nākamais organizācijas līmenis - olbaltumvielu sekundārā struktūra. Ir vairāki tā veidi. Polipeptīdu ķēdes daļas lokālo sakārtošanu stabilizē ūdeņraža saites. Populārākie veidi ir:

Telpiskā struktūra
Olbaltumvielu terciārā struktūra ietver iepriekšējā līmeņa elementus. Tos stabilizē dažāda veida mijiedarbība. Hidrofobās saites ir ārkārtīgi svarīgas. Stabilizācija ietver:
- Kovalentā mijiedarbība.
- Jonu saites veidojas starp aminoskābju sānu grupām, kurām ir pretējs lādiņš.
- Ūdeņraža mijiedarbība.
- Hidrofobās saites. Mijiedarbības procesā ar apkārtējiem elementiem H 2 O proteīns salocās tā, ka sānu nepolārās aminoskābju grupas tiek izolētas no ūdens šķīduma. Hidrofilās grupas (polāras) parādās uz molekulas virsmas.
Olbaltumvielu terciāro struktūru nosaka ar magnētiskās (kodolrezonanses) metodēm, noteikta veida mikroskopiju un citām metodēm.
Ieklāšanas princips
Pētījumi liecina, ka ir ērti noteikt vēl vienu līmeni starp 2. un 3. līmeni. To sauc par "arhitektūru", "dēšanas motīvu". To nosaka sekundārās struktūras komponentu (beta virknes un alfa spirāles) relatīvais novietojums kompaktas globulas - proteīna domēna - robežās. Tas var pastāvēt neatkarīgi vai tikt iekļauts lielākā proteīnā kopā ar citiem līdzīgiem proteīniem. Konstatēts, ka stila motīvi ir diezgan konservatīvi. Tie ir atrodami proteīnos, kuriem nav ne evolucionāru, ne funkcionālu attiecību. Arhitektūras definīcija ir racionālas (fiziskās) klasifikācijas pamats.

Domēna organizācija
Vairākām polipeptīdu ķēdēm savstarpēji izkārtojoties viena proteīna kompleksa ietvaros, veidojas proteīnu kvartārā struktūra. Elementi, kas to veido, veidojas atsevišķi uz ribosomām. Tikai pēc sintēzes pabeigšanas šī proteīna struktūra sāk veidoties. Tas var saturēt gan dažādas, gan identiskas polipeptīdu ķēdes. Olbaltumvielu kvartārā struktūra stabilizējas, pateicoties tām pašām mijiedarbībām kā iepriekšējā līmenī. Daži kompleksi var ietvert vairākus desmitus proteīnu.
Olbaltumvielu struktūra: aizsardzības uzdevumi
Citoskeleta polipeptīdi, kas kaut kādā veidā darbojas kā pastiprinājums, piešķir daudzām organellām formu un piedalās to izmaiņās. Strukturālie proteīni nodrošina ķermeņa aizsardzību. Piemēram, kolagēns ir šāds proteīns. Tas veido pamatu saistaudu starpšūnu vielā. Keratīnam ir arī aizsargājoša funkcija. Tas veido ragu, spalvu, matu un citu epidermas atvasinājumu pamatu. Kad olbaltumvielas saistās ar toksīniem, daudzos gadījumos notiek detoksikācija. Tādā veidā tiek izpildīts ķermeņa ķīmiskās aizsardzības uzdevums. Aknu enzīmiem ir īpaši liela nozīme toksīnu neitralizēšanas procesā cilvēka organismā. Viņi spēj sadalīt indes vai pārvērst tos šķīstošā formā. Tas atvieglo to ātrāku izvadīšanu no ķermeņa. Asinīs un citos ķermeņa šķidrumos esošās olbaltumvielas nodrošina imūno aizsardzību, izraisot reakciju gan uz patogēnu uzbrukumu, gan ievainojumu. Imūnglobulīni (antivielas un komplementa sistēmas sastāvdaļas) spēj neitralizēt baktērijas, svešas olbaltumvielas un vīrusus.
Regulēšanas mehānisms
Olbaltumvielu molekulas, kas nedarbojas ne kā enerģijas avots, ne kā būvmateriāls, kontrolē daudzus intracelulārus procesus. Tādējādi, pateicoties tiem, tiek regulēta tulkošana, transkripcija, sagriešana un citu polipeptīdu aktivitāte. Regulēšanas mehānisms ir balstīts uz enzīmu aktivitāti vai izpaužas specifiskas saistīšanās dēļ ar citām molekulām. Piemēram, transkripcijas faktori, aktivatora polipeptīdi un represoru proteīni spēj kontrolēt gēnu transkripcijas intensitāti. To darot, tie mijiedarbojas ar gēnu regulējošām sekvencēm. Vissvarīgākā loma intracelulāro procesu gaitas kontrolēšanā tiek piešķirta proteīnu fosfatāzēm un proteīnkināzēm. Šie fermenti izraisa vai kavē citu proteīnu aktivitāti, pievienojot vai noņemot no tiem fosfātu grupas.

Signāla uzdevums
To bieži apvieno ar regulēšanas funkciju. Tas ir saistīts ar faktu, ka daudzi intracelulāri, kā arī ārpusšūnu polipeptīdi var pārraidīt signālus. Šī spēja piemīt augšanas faktoriem, citokīniem, hormoniem un citiem savienojumiem. Steroīdi tiek transportēti caur asinīm. Hormona mijiedarbība ar receptoru darbojas kā signāls, kas izraisa šūnu reakciju. Steroīdi kontrolē savienojumu saturu asinīs un šūnās, vairošanos, augšanu un citus procesus. Piemērs ir insulīns. Tas regulē glikozes līmeni. Šūnu mijiedarbība tiek veikta, izmantojot signālu proteīnu savienojumus, kas tiek pārraidīti caur starpšūnu vielu.
Elementu transportēšana
Šķīstošajiem proteīniem, kas iesaistīti mazu molekulu kustībā, ir augsta afinitāte pret substrātu, kas atrodas paaugstinātā koncentrācijā. Viņiem ir arī iespēja to viegli atbrīvot vietās, kur tā saturs ir zems. Piemērs ir transporta proteīna hemoglobīns. Tas pārvieto skābekli no plaušām uz citiem audiem, un no tiem pārnes oglekļa dioksīdu. Daži membrānas proteīni ir iesaistīti arī mazu molekulu transportēšanā caur šūnu sienām, mainot tās. Citoplazmas lipīdu slānis ir ūdensnecaurlaidīgs. Tas novērš lādētu vai polāru molekulu difūziju. Membrānas transporta savienojumi parasti tiek sadalīti nesējos un kanālos.
Rezerves savienojumi
Šīs olbaltumvielas veido tā sauktās rezerves. Tie uzkrājas, piemēram, augu sēklās un dzīvnieku olās. Šādas olbaltumvielas darbojas kā rezerves vielas un enerģijas avots. Dažus savienojumus organisms izmanto kā aminoskābju rezervuāru. Tie savukārt ir vielmaiņas regulēšanā iesaistīto aktīvo vielu prekursori.
Šūnu receptori
Šādas olbaltumvielas var atrasties vai nu tieši citoplazmā, vai iestrādātas sienā. Viena savienojuma daļa saņem signālu. Parasti tā ir ķīmiska viela un dažos gadījumos mehāniska iedarbība (piemēram, stiepšanās), gaismas un citi stimuli. Signāla iedarbības procesā uz noteiktu molekulas fragmentu - polipeptīdu receptoru - sākas tā konformācijas izmaiņas. Tie izraisa izmaiņas pārējās daļas konformācijā, kas pārraida stimulu uz citām šūnas sastāvdaļām. Signālu var nosūtīt dažādos veidos. Daži receptori spēj katalizēt ķīmisku reakciju, bet citi darbojas kā jonu kanāli, kas aizveras vai atveras stimula ietekmē. Daži savienojumi šūnā īpaši saista kurjermolekulas.
Motoru polipeptīdi
Ir vesela proteīnu klase, kas nodrošina ķermeņa kustību. Motorolbaltumvielas ir iesaistītas muskuļu kontrakcijā, šūnu kustībā un flagellas un skropstu darbībā. Tie nodrošina arī virziena un aktīvo transportu. Kinezīni un dyneīni transportē molekulas pa mikrotubulām, izmantojot ATP hidrolīzi kā enerģijas avotu. Pēdējie pārvieto organellus un citus elementus centrosomu virzienā no perifērajām šūnu zonām. Kinezīni pārvietojas pretējā virzienā. Dineīni ir atbildīgi arī par flagellas un skropstu darbību.
