Eseja
par tēmu: "Reprodukcija"
3. ievads
1. Reprodukcijas veidi 4
1.1. Aseksuāla vairošanās 4
1.2. Seksuālā pavairošana 6
2. Organismu individuālā attīstība 10
2.1. Embrionālais attīstības periods 10
2.2. Pēcdzemdību attīstības periods 13
2.3. Vispārējie attīstības modeļi. Bioģenētiskais likums 15
18. secinājums
Atsauces 18
Ievads
Spēja vairoties, t.i. vienas sugas īpatņu jaunas paaudzes radīšana ir viena no galvenajām dzīvo organismu īpašībām. Reprodukcijas procesā ģenētiskais materiāls tiek pārnests no vecāku paaudzes uz nākamo paaudzi, kas nodrošina ne tikai noteiktas sugas, bet konkrētu vecāku indivīdu īpašību vairošanos. Sugai vairošanās jēga ir aizvietot tās pārstāvjus, kas mirst, kas nodrošina sugas pastāvēšanas nepārtrauktību; turklāt piemērotos apstākļos vairošanās ļauj palielināt kopējo sugu skaitu.
Katram jaunam indivīdam, pirms tas ir spējīgs vairoties, ir jāiziet vairāki augšanas un attīstības posmi. Daži indivīdi mirst pirms reproduktīvās stadijas (vai dzimumbrieduma) plēsēju iznīcināšanas, slimību un dažādu nejaušu notikumu rezultātā; tāpēc suga var izdzīvot tikai ar nosacījumu, ka katra paaudze rada vairāk pēcnācēju nekā bija vecāku indivīdi, kuri piedalījās vairošanās procesā. Populācijas lielums svārstās atkarībā no līdzsvara starp indivīdu vairošanos un izmiršanu. Ir vairākas dažādas izplatīšanas stratēģijas, katrai no kurām ir atšķirīgas priekšrocības un trūkumi; tie visi tiks aprakstīti šajā kopsavilkumā.
1. Reprodukcijas veidi
Ir zināmi dažādi vairošanās veidi, taču tos visus var apvienot divos veidos: seksuālā un aseksuālā.
Seksuālā vairošanās attiecas uz paaudžu maiņu un organismu attīstību no specializētām dzimumšūnām, kas veidojas dzimumdziedzeros. Šajā gadījumā divu dažādu vecāku veidotu dzimumšūnu saplūšanas rezultātā veidojas jauns organisms. Tomēr bezmugurkaulniekiem spermatozoīdi un olšūnas bieži veidojas viena organisma ķermenī. Šo parādību, biseksuālismu, sauc par hermafrodītismu. Ziedošie augi ir arī biseksuāli. Lielākajā daļā segsēklu (ziedošo) augu sugu divdzimuma zieds ietver gan putekšņlapas, kas veido vīrišķās dzimumšūnas (spermas šūnas), gan pīnes, kurās ir olas. Apmēram ceturtdaļā sugu vīrišķie (staminate) un sievišķīgie (pistilāti) ziedi attīstās neatkarīgi, t.i. to ziedi ir viendzimuma. Piemērs ir kaņepes. Dažos augos (kukurūzā, bērzā) gan vīriešu, gan sieviešu ziedi parādās vienam un tam pašam indivīdam.
Attīstās dažas dzīvnieku un augu sugas
neapaugļota olšūna. Šo reprodukcijas veidu sauc par neapstrādātu vai partenoģenētisku.
Bezdzimuma vairošanos raksturo tas, ka jauns indivīds attīstās no neseksuālām, somatiskām (ķermeņa) šūnām.
1.1. Aseksuāla vairošanās
Ar aseksuālu vairošanos jauns organisms var rasties no vienas šūnas vai no vairākām aseksuālām (somatiskām) mātes šūnām. Aseksuālā vairošanās ir saistīta tikai ar vienu no vecākiem. Tā kā šūnas, kas rada meitas organismus, rodas mitozes rezultātā, visi pēcnācēji pēc iedzimtajām īpašībām būs līdzīgi mātes indivīdam.
Rīsi. 1. Zaļās eiglēnas pavairošana
Daudzi vienšūņi (amoeba, zaļā eiglēna u.c.), vienšūnu aļģes (Chlamydomonas) vairojas mitotiski šūnu dalīšanās ceļā (1. att.). Citām vienšūnām dažām zemākajām sēnēm, aļģēm (hlorella), dzīvniekiem, piemēram, malārijas malārijas plazmodija izraisītājiem, raksturīga sporulācija. Šajā gadījumā šūna sadalās lielā skaitā indivīdu, kas ir vienāds ar to kodolu skaitu, kas iepriekš radušies mātes šūnā atkārtotas kodola dalīšanas rezultātā. Sporulēties spēj arī daudzšūnu organismi: tās ir sūnas, augstākās sēnes, daudzšūnu aļģes, pteridofīti un daži citi.
Gan vienšūnu, gan daudzšūnu organismos pumpuru veidošanās ir arī aseksuālās vairošanās metode. Piemēram, rauga sēnītēm un dažiem ciliātiem (sūcošiem ciliātiem), pumpējoties uz mātes šūnas, sākotnēji veidojas neliels bumbulis, kas satur kodolu, pumpuru. Tas aug, sasniedz izmēru, kas ir tuvu mātes ķermenim, un pēc tam atdalās, pārejot uz neatkarīgu eksistenci. Daudzšūnu organismos (saldūdens hidra) nieres sastāv no šūnu grupas no abiem ķermeņa sienas slāņiem. Pumpuri aug, pagarinās, un tā priekšgalā parādās mutes atvere, ko ieskauj taustekļi. Dīgšana beidzas ar nelielas hidras veidošanos, kas pēc tam atdalās no mātes organisma.
Daudzšūnu dzīvniekiem aseksuāla vairošanās notiek tādā pašā veidā (medūzas, annelīdi, plakanie tārpi, adatādaiņi). No katras šādas daļas veidojas pilnvērtīgs indivīds.
Augos ir plaši izplatīta veģetatīvā pavairošana, t.i. ķermeņa daļas spraudeņi, stīgas, bumbuļi. Tādējādi kartupeļi vairojas ar modificētām stumbra pazemes daļām – bumbuļiem. Jasmīnu un vītolu dzinumi un spraudeņi viegli iesakņojas. Vīnogas, jāņogas un ērkšķogas pavairo, izmantojot spraudeņus.
Zemeņu stīgu garie ložņu stublāji veido pumpurus, kas, iesakņojoties, rada jaunu augu. Dažus augus, piemēram, begoniju, var pavairot ar lapu spraudeņiem (lapas lāpstiņu un kātiņu). Lapas apakšpusē, vietās, kur sazarojas lielas vēnas, parādās saknes, augšējā pusē ir pumpuri, un pēc tam dzinumi.
Sakni izmanto arī veģetatīvā pavairošanā. Dārzkopībā avenes, ķiršus, plūmes un rozes pavairo, izmantojot spraudeņus no sānu saknēm. Dālijas vairojas, izmantojot sakņu bumbuļus. Arī stumbra sakneņa pazemes daļas pārveidošana veido jaunus augus. Piemēram, sēt dadzis ar sakneņu palīdzību var iegūt vairāk nekā tūkstoti jaunu īpatņu uz 1 m2 augsnes.
1.2. Seksuālā pavairošana
Seksuālajai pavairošanai ir ļoti lielas evolucionāras priekšrocības salīdzinājumā ar aseksuālo vairošanos. Tas ir saistīts ar faktu, ka pēcnācēju genotips rodas, apvienojot gēnus, kas pieder abiem vecākiem. Tā rezultātā palielinās organismu spēja pielāgoties vides apstākļiem. Tā kā katrā paaudzē tiek veiktas jaunas kombinācijas, jauniem eksistences apstākļiem var pielāgoties daudz lielāks skaits indivīdu nekā aseksuālās vairošanās laikā. Jaunu gēnu kombināciju rašanās nodrošina veiksmīgāku un ātrāku sugas pielāgošanos mainīgajiem vides apstākļiem.
Tādējādi seksuālās vairošanās būtība slēpjas divu dažādu avotu ģenētiskās informācijas pēcteča - vecāku - apvienojumā mantojuma materiālā.
Dzimumšūnas attīstās dzimumdziedzeros: vīriešu spermas, sieviešu olšūnas (vai olšūnas). Pirmajā gadījumā to attīstību sauc par spermatoģenēzi, otrajā - par ooģenēzi (no latīņu ovo - ola).
Dzimumšūnu veidošanās procesā izšķir vairākus posmus. Pirmais posms ir reprodukcijas periods, kurā pirmatnējās dzimumšūnas dalās mitozes ceļā, kā rezultātā palielinās to skaits.
Otrais posms ir izaugsmes periods. Nenobriedušajās vīriešu dzimuma gametās tas nav izteikts. To izmēri nedaudz palielinās. Gluži pretēji, nākotnes olšūnu oocīti palielinās, dažreiz simtiem un biežāk tūkstošiem un pat miljoniem reižu. Ocītu augšana notiek citu ķermeņa šūnu veidotu vielu dēļ. Tādējādi zivīm, abiniekiem un, lielākā mērā, rāpuļiem un putniem olas lielākā daļa ir dzeltenums. Tas tiek sintezēts aknās, īpašā šķīstošā veidā ar asinīm tiek transportēts uz olnīcu, iekļūst augošos oocītos un nogulsnējas tur dzeltenuma plākšņu veidā. Turklāt pašā nākotnes reproduktīvajā šūnā tiek sintezēti daudzi proteīni un liels skaits dažādu RNS: transporta, ribosomu un informatīvo. Dzeltenums ir barības vielu (tauku, olbaltumvielu, ogļhidrātu, vitamīnu u.c.) kopums, kas nepieciešams, lai barotu augošo embriju, un RNS nodrošina proteīnu sintēzi agrīnā attīstības stadijā, kad vēl netiek izmantota pašas postošā informācija.
Nākamais posms, nobriešanas periods jeb mejoze, ir parādīts 2. attēlā. Šūnas, kas nonāk nobriešanas periodā, satur diploīdu hromosomu kopu un jau dubultotu DNS daudzumu.

Rīsi. 2. Dzimumšūnu nobriešana (mejoze)
Mejozes būtība ir tāda, ka katra dzimuma šūna saņem vienu haploīdu hromosomu komplektu. Taču tajā pašā laikā mejoze ir stadija, kuras laikā, kombinējot dažādas mātes un tēva hromosomas, rodas jaunas gēnu kombinācijas, iedzimto tieksmju rekombinācija notiek arī krustošanās rezultātā – sekciju apmaiņa starp homologām hromosomām procesa laikā. par mejozi.
Mejoze ietver divus secīgus dalījumus. Tāpat kā mitozē, katram meiotiskajam dalījumam ir četri posmi: profāze, metafāze, anafāze un telofāze.
Pirmā (I) meiotiskā dalīšana. I fāze sākas ar hromosomu spiralizāciju. Kā jūs atceraties, katra hromosoma sastāv no diviem hromatīdiem, kas savienoti centromērā. Tad homologās hromosomas satuvinās, katrs vienas hromosomas katras hromatīdas punkts tiek apvienots ar citas, homologas hromosomas, atbilstošo hromatīda punktu. Šo homologo hromosomu precīzas un ciešas savienošanas procesu meiozē sauc par konjugāciju. Nākotnē starp šādām hromosomām var notikt krustošanās - identisku vai homologu, t.i., saturošu vienādu gēnu, reģionu apmaiņa. Profāzes beigās starp homologām hromosomām rodas atgrūdoši spēki. Pirmkārt, tie parādās centromēra reģionā un pēc tam citos apgabalos.
I metafāzē hromosomu spiralizācija ir maksimāla. Konjugētās hromosomas atrodas gar ekvatoru, un homologo hromosomu centromēri ir vērsti pret dažādiem šūnas poliem. Uz tiem ir piestiprinātas vārpstas vītnes.
I anafāzē homologo hromosomu rokas beidzot atdalās, un hromosomas pārvietojas uz dažādiem poliem. Līdz ar to no katra homologo hromosomu pāra meitas šūnā nonāk tikai viena. Hromosomu skaits samazinās uz pusi, hromosomu komplekts kļūst haploīds. Tomēr katra hromosoma sastāv no diviem hromatīdiem, t.i., tajā joprojām ir divreiz lielāks DNS daudzums.
I telofāzē uz īsu laiku veidojas kodola apvalks. Starpfāzes laikā starp pirmo un otro mejozes dalījumu DNS reduplikācija nenotiek. Šūnas, kas veidojas pirmās nobriešanas dalīšanas rezultātā, atšķiras pēc tēva un mātes hromosomu sastāva un līdz ar to arī gēnu komplektā.
Piemēram, visas cilvēka šūnas, ieskaitot pirmatnējās dzimumšūnas, satur 46 hromosomas. No tiem 23 saņemti no tēva un 23 no mātes. Kad pēc pirmās meiotiskās dalīšanās veidojas dzimumšūnas, arī spermatocīti un oocīti saņem 23 hromosomas. Tomēr, ņemot vērā tēva un mātes hromosomu atšķirību nejaušību anafāzē I, iegūtās šūnas saņem dažādas vecāku hromosomu kombinācijas. Piemēram, vienā no tām var būt 3 tēva un 20 mātes hromosomas, citai 10 tēva un 13 mātes, trešajai 20 tēva un 3 mātes u.c. Iespējamo kombināciju skaits ir ļoti liels. Ja ņemam vērā arī hromosomu homologo sekciju apmaiņu mejozes pirmās dalīšanas profāzē, tad ir pilnīgi skaidrs, ka katra iegūtā dzimumšūna ir ģenētiski unikāla, jo tai ir savs unikāls gēnu komplekts.
Līdz ar to mejoze ir kombinatīvās genotipiskās variabilitātes pamatā.
Otrais (II) meiotiskais dalījums. Otrā mejozes dalīšanās parasti notiek tāpat kā parastā mitotiskā dalīšanās, ar vienīgo atšķirību, ka dalošā šūna ir haploīda. II anafāzē centromēri, kas savieno māsas hromatīdus katrā hromosomā, sadalās, un hromatīdi, tāpat kā mitozē, no šī brīža kļūst par neatkarīgām hromosomām. Pabeidzot II telofāzi, viss mejozes process beidzas: no sākotnējās primārās dzimumšūnas veidojas četras haploīdas šūnas.
Vīriešiem tie visi tiek pārveidoti par gametām - spermu. Mātītēm nevienmērīgas mejozes dēļ tikai viena šūna ražo dzīvotspējīgu olu. Pārējās trīs meitas šūnas ir daudz mazākas, tās pārvēršas par tā sauktajiem vadošajiem vai reducējošiem ķermeņiem, kas drīz mirst. No bioloģiskā viedokļa tikai vienas olšūnas veidošanās un trīs ģenētiski pilnīgu vadošo ķermeņu nāve ir saistīta ar nepieciešamību vienā šūnā saglabāt visas rezerves barības vielas, kas būs nepieciešamas topošā embrija attīstībai.
Veidošanās periods sastāv no šūnām, kas iegūst noteiktu formu un izmēru, kas atbilst to funkcijai.
Nobriešanas procesā sieviešu dzimumšūnas pārklājas ar membrānām un ir gatavas apaugļošanai tūlīt pēc mejozes pabeigšanas. Daudzos gadījumos, piemēram, rāpuļiem, putniem un zīdītājiem, olu apņemošo šūnu aktivitātes dēļ ap to parādās vairākas papildu membrānas. To funkcija ir aizsargāt olu un augošo embriju no ārējām nelabvēlīgām ietekmēm. Spermas var būt dažāda izmēra un formas.
Spermas funkcija ir piegādāt olšūnai ģenētisko informāciju un stimulēt tās attīstību. Izveidotajā spermā ir mitohondriji, Golgi aparāts, kas izdala fermentus, kas izšķīdina olšūnas membrānu apaugļošanas laikā, tas ir, spermas un olšūnas saplūšanas laikā. Iegūto diploīdu šūnu sauc par zigotu.
2. Organismu individuālā attīstība
Individuālā attīstība jeb ontoģenēze attiecas uz visu indivīda dzīves periodu no brīža, kad spermatozoīds saplūst ar olšūnu un veidojas zigota, līdz organisma nāvei. Ontoģenēzi iedala divos periodos: 1) embrionālā no zigotas veidošanās līdz dzimšanai vai izejai no olšūnas membrānām; 2) pēcdzemdību periodā no izkļūšanas no olšūnas membrānām vai piedzimšanas līdz organisma nāvei.
Zinātni, kas pēta organismu individuālās attīstības modeļus embrionālajā stadijā, sauc par embrioloģiju (no grieķu vārda embrija embrijs).
2.1. Embrionālais attīstības periods
Lielākajā daļā daudzšūnu dzīvnieku, neatkarīgi no to organizācijas sarežģītības, embrija attīstības posmi, kuriem embrijs iziet, ir vienādi. Embrionālajā periodā ir trīs galvenie posmi: šķelšanās, gastrulācija un primārā organoģenēze.
Sadalīšana. Organisma attīstība sākas vienšūnas stadijā. Apaugļota olšūna ir šūna un vienlaikus organisms tās agrīnākajā attīstības stadijā. Atkārtotas dalīšanās rezultātā vienšūnas organisms pārvēršas par daudzšūnu. Diploīdais kodols, kas parādās apaugļošanas laikā, saplūstot spermai un olšūnai, pēc dažām minūtēm sāk dalīties, un līdz ar to dalās citoplazma. Iegūto šūnu izmērs samazinās ar katru dalīšanos, tāpēc dalīšanās procesu sauc par šķelšanos. Sadrumstalošanās periodā šūnu materiāls uzkrājas tālākai attīstībai. Sadrumstalošanās beidzas ar daudzšūnu embrija – blastulas – veidošanos. Blastulai ir dobums, kas piepildīts ar šķidrumu, tā sauktais primārais ķermeņa dobums.
Gadījumos, kad olas citoplazmā ir maz dzeltenuma (kā lancetē) vai salīdzinoši maz (kā vardei), sadrumstalotība ir pilnīga, t.i., ola pilnībā sadalās.
Pretējā gadījumā putniem notiek sadrumstalotības periods. Citoplazma bez dzeltenuma veido tikai 1% no kopējā vistas olas tilpuma; visa atlikušā olas citoplazma un līdz ar to arī zigota ir piepildīta ar dzeltenuma masu. Uzmanīgi aplūkojot vistas olu, uz viena no tās poliem tieši uz dzeltenuma var redzēt nelielu plankumu - blastulu jeb dīgļu disku, kas izveidojies, sasmalcinot bezdzeltenu citoplazmas daļu, kurā atrodas kodols. Šādos gadījumos sasmalcināšanu sauc par nepilnīgu. Nepilnīga sadrumstalotība raksturīga arī dažām zivīm un rāpuļiem.
Visos gadījumos gan lancetē, gan abiniekiem, gan putniem, kā arī citiem dzīvniekiem kopējais šūnu apjoms blastulas stadijā nepārsniedz zigotas tilpumu. Citiem vārdiem sakot, zigotas mitotiskā dalīšanās nav saistīta ar iegūto meitas šūnu augšanu līdz mātes tilpumam, un to izmēri secīgu dalījumu sērijas rezultātā pakāpeniski samazinās. Šī mitotiskās šūnu dalīšanās iezīme šķelšanās laikā tiek novērota apaugļotu olšūnu attīstības laikā visiem dzīvniekiem.
Dažādām dzīvnieku sugām raksturīgas arī dažas citas smalcināšanas pazīmes. Piemēram, visām blastulas šūnām ir diploīds hromosomu komplekts, tās ir identiskas pēc struktūras un atšķiras viena no otras galvenokārt ar tajās esošā dzeltenuma daudzumu. Šādas šūnas, kurām trūkst specializācijas pazīmju noteiktu funkciju veikšanai, sauc par nespecializētām (vai nediferencētām) šūnām. Vēl viena šķelšanās iezīme ir ārkārtīgi īss blastomēru mitotiskais cikls salīdzinājumā ar pieauguša organisma šūnām. Ļoti īsā starpfāzē notiek tikai DNS dublēšanās.
Gastrulācija. Blastula, kas parasti sastāv no liela skaita blastomēru (piemēram, lancetē no 3000 šūnām), attīstības procesā pāriet jaunā stadijā, ko sauc par gastrulu (no grieķu kuņģa kuņģa). Šajā stadijā embrijs sastāv no skaidri atšķiramiem šūnu slāņiem - tā sauktajiem dīgļu slāņiem: ārējais jeb ektoderma (no grieķu ectos - atrodas ārpusē) un iekšējā jeb endoderma (no grieķu entos - atrodas iekšpusē). . Procesu kopumu, kas noved pie gastrulas veidošanās, sauc par gastrulāciju.
Lanceletā gastrulācija tiek veikta, invaginējot vienu no blastulas poliem uz iekšu, pret otru; citiem dzīvniekiem, vai nu atslāņojoties blastulas sieniņai, vai apaudzējot masīvo veģetatīvo polu ar mazām blastulas šūnām. dzīvnieku stabs.
Daudzšūnu dzīvniekiem, izņemot koelenterātus, paralēli gastrulācijai vai, kā lancetē, pēc tās parādās trešā dīgļu slāņa mezoderma (no grieķu valodas mezo, kas atrodas vidū), kas ir šūnu elementu kopums, kas atrodas starp ekto- un endoderma primārā ķermeņa dobuma blastocele. Ar mezodermas parādīšanos embrijs kļūst trīsslāņu.
Tādējādi gastrulācijas procesa būtība ir šūnu masu kustība. Embrija šūnas praktiski dalās un neaug. Tomēr šajā posmā sākas embriju šūnu ģenētiskās informācijas izmantošana, un parādās pirmās diferenciācijas pazīmes.
Diferenciācija jeb diferenciācija ir tās rašanās process un strukturālo un funkcionālo atšķirību palielināšanās starp atsevišķām šūnām un embrija daļām. No morfoloģiskā viedokļa diferenciācija izpaužas vairāku simtu noteiktas struktūras šūnu veidu veidošanā, kas atšķiras viens no otra. No nespecializētām blastula šūnām pamazām izceļas ādas epitēlija šūnas, zarnu epitēlijs, plaušas, parādās nervu un muskuļu šūnas u.c. No bioķīmiskā viedokļa šūnu specializācija slēpjas spējā sintezēt noteiktus proteīnus, kas raksturīgi tikai noteiktam šūnu tipam. Limfocīti sintezē aizsargājošo proteīnu antivielas, muskuļu šūnas saraujas proteīna miozīnu. Katrs šūnu veids ražo savus proteīnus, kas ir unikāli tam. Šūnu bioķīmisko specializāciju nodrošina selektīva, diferenciāla gēnu darbība, t.i., dažādu dīgļu slāņu šūnās sāk darboties atsevišķu orgānu un sistēmu rudimenti, dažādas gēnu grupas.
Dažādām dzīvnieku sugām tie paši dīgļu slāņi rada vienus un tos pašus orgānus un audus. Tas nozīmē, ka tie ir homologi. Tādējādi no ārējā dīgļu slāņa - ektodermas - šūnām posmkājiem veidojas hordati, tostarp zivīm, abiniekiem, rāpuļiem, putniem un zīdītājiem, āda un to atvasinājumi, kā arī nervu sistēma un maņu orgāni. Lielākās daļas dzīvnieku dīgļu slāņu homoloģija ir viens no dzīvnieku pasaules vienotības pierādījumiem.
Organoģenēze. Pēc gastrulācijas pabeigšanas embrijs veido aksiālo orgānu kompleksu: nervu caurulīti, notohordu un zarnu caurulīti. Lanceletā aksiālie orgāni veidojas šādi: ektoderma embrija muguras pusē noliecas pa viduslīniju, pārvēršoties rievā, un ektoderma, kas atrodas pa labi un pa kreisi no tās, sāk augt tās malās. Rieva, nervu sistēmas rudiments, nogrimst zem ektodermas, un tās malas aizveras. Tiek veidota nervu caurule. Pārējā ektoderma ir ādas epitēlija rudiments.
Endodermas muguras daļa, kas atrodas tieši zem nerva rudimenta, ir atdalīta no pārējās endodermas un salocās blīvā auklā - notohordā. No atlikušās endodermas daļas attīstās mezoderma un zarnu epitēlijs. Turpmāka embrionālo šūnu diferenciācija noved pie daudzu dīgļu slāņu - orgānu un audu atvasinājumu rašanās. Dīgļu slāņus veidojošo šūnu specializācijas procesā no ektodermas veidojas nervu sistēma, maņu orgāni, ādas epitēlijs, zobu emalja; no endodermas zarnu epitēlijs, gremošanas dziedzeri aknas un aizkuņģa dziedzeris, žaunu un plaušu epitēlijs; no mezodermas muskuļu audiem, saistaudiem, tai skaitā irdeniem saistaudiem, skrimšļiem un kaulaudiem, asinīm un limfas, kā arī asinsrites sistēmas, nierēm, dzimumdziedzeriem.
2.2. Pēcembrionālais attīstības periods
Piedzimšanas vai organisma atbrīvošanās no olu čaumalām brīdī beidzas embrionālais periods un sākas postembrionālais attīstības periods. Pēcembrionālā attīstība var būt tieša vai pavadīta ar transformāciju (metamorfozi).
Tiešās attīstības laikā (rāpuļiem, putniem, zīdītājiem) no olu čaumalām vai no mātes ķermeņa izdalās maza izmēra organisms, bet jau ir izveidojušies visi pieaugušam dzīvniekam raksturīgie galvenie orgāni. Postembrionālā attīstība šajā gadījumā tiek samazināta galvenokārt līdz augšanai un pubertātei.
Attīstoties ar metamorfozi, no olas izdalās kāpurs, kura struktūra parasti ir vienkāršāka nekā pieaugušam dzīvniekam, ar īpašiem kāpuru orgāniem, kuru pieaugušā stāvoklī nav. Kāpurs barojas, aug, un laika gaitā kāpuru orgāni tiek aizstāti ar orgāniem, kas raksturīgi pieaugušiem indivīdiem. Līdz ar to metamorfozes laikā tiek iznīcināti kāpuru orgāni un parādās pieaugušiem dzīvniekiem raksturīgi orgāni.
Apskatīsim vairākus netiešās pēcembrionālās attīstības piemērus. Ascīdijas kāpuram (cilts Chordata, apakšcilme Larval-Chordata) ir visas galvenās hordātu īpašības: notohords, nervu caurule un žaunu spraugas rīklē. Tas brīvi peld, pēc tam pievienojas kādai cietai virsmai jūras dibenā, kur notiek metamorfoze: izzūd tā aste, notohords un muskuļi, un nervu caurule sadalās atsevišķās šūnās, no kurām lielākā daļa ir fagocitētas. Viss, kas paliek no kāpuru nervu sistēmas, ir šūnu grupa, kas rada nervu gangliju. Pieauguša ascīdiāna ķermeņa uzbūve, kas vada piesaistītu dzīvesveidu, nemaz nelīdzinās parastajām hordātu organizācijas iezīmēm. Tikai zināšanas par ontoģenēzes iezīmēm ļauj noteikt ascīdiešu sistemātisko stāvokli: kāpuru struktūra norāda uz to izcelsmi no hordātiem, kas vadīja brīvu dzīvesveidu. Metamorfozes procesā ascīdi pāriet uz mazkustīgu dzīvesveidu, un tāpēc viņu organizācija tiek vienkāršota.
Abinieku kāpuru forma ir kurkulis, kam raksturīgi žaunu spraugas, sānu līnija, divkameru sirds un viens asinsrites loks. Metamorfozes procesā, kas notiek vairogdziedzera hormona ietekmē, aste atrisinās, parādās ekstremitātes, pazūd sānu līnija, attīstās plaušas un otrs asinsrites loks. Ievērības cienīga ir vairāku kurkuļu un zivju struktūras īpašību līdzība (sānu līnija, sirds un asinsrites sistēmas uzbūve, žaunu spraugas).
Kukaiņu attīstība var kalpot arī par metamorfozes piemēru. Tauriņu kāpuri jeb spāru kāpuri pēc uzbūves, dzīvesveida un dzīvotnes krasi atšķiras no pieaugušiem dzīvniekiem un atgādina viņu senčus, annelīdus.
Pēcdzemdību attīstības periodam ir atšķirīgs ilgums. Piemēram, maijvaboles dzīvo 2-3 gadus kāpuru stāvoklī un no 2-3 stundām līdz 2-3 dienām nobriedušā stāvoklī atkarībā no sugas. Vairumā gadījumu pēcdzemdību periods ir ilgāks. Cilvēkiem tas ietver pubertātes posmu, brieduma posmu un vecuma posmu.
Zīdītājiem un cilvēkiem ir zināma paredzamā dzīves ilguma atkarība no pubertātes un grūtniecības ilguma. Dzīves ilgums parasti pārsniedz
ontoģenēzes pirmsreproduktīvais periods 5-8 reizes.
Pēcembrionālo attīstību pavada augšana. Izšķir nenoteiktu izaugsmi, kas turpinās visu mūžu, un noteiktu izaugsmi, kas ierobežota līdz noteiktam periodam. Nenoteikta augšana tiek novērota kokainajām augu formām, dažiem mīkstmiešiem, mugurkaulniekiem, zivīm un žurkām.
Daudziem dzīvniekiem augšana apstājas neilgi pēc dzimumbrieduma sasniegšanas. Cilvēkiem augšana beidzas 20-25 gadu vecumā.
2.3. Vispārējie attīstības modeļi. Bioģenētiskais likums
Visi daudzšūnu organismi attīstās no apaugļotas olšūnas. Viena veida dzīvniekiem embriju attīstība lielā mērā ir līdzīga. Visos hordātos embrionālajā periodā veidojas aksiālais skeleta notohords, parādās nervu caurule, un rīkles priekšējā daļā veidojas žaunu spraugas. Arī hordātu strukturālais plāns ir tāds pats. Agrīnās attīstības stadijās mugurkaulnieku embriji ir ļoti līdzīgi (3. att.). Šie fakti apstiprina K. Bēra formulētā embrionālās līdzības likuma pamatotību: “Embrijiem jau no agrīnajiem posmiem ir raksturīga noteikta vispārīga līdzība tipa iekšienē.” Dažādu sistemātisku grupu embriju līdzība norāda uz to kopīgo izcelsmi. Pēc tam embriju struktūra atklāj klases, ģints, sugas un, visbeidzot, konkrētajam indivīdam raksturīgās īpašības. Embriju īpašību atšķirības attīstības laikā sauc par embrionālo novirzi un atspoguļo noteiktas sistemātiskas dzīvnieku grupas evolūciju, noteiktas sugas attīstības vēsturi.

Rīsi. 3. Dzimumlīnijas līdzība mugurkaulniekiem: 1 monotrēmi (ehidna), 2 marsupials (ķenguri), 3 artiodaktili (brieži), 4 plēsēji (kaķi), 5 primāti (pērtiķi), 6 - cilvēki
Liela līdzība starp embrijiem agrīnās attīstības stadijās un
atšķirību parādībai vēlākos posmos ir savs izskaidrojums.
Ķermenis attīstības gaitā ir pakļauts mainīgumam.
Mutācijas process ietekmē gēnus, kas nosaka jaunāko embriju strukturālās un vielmaiņas īpatnības. Bet tajās radušās struktūras (senās īpašības, kas raksturīgas tāliem senčiem) spēlē ļoti lielu lomu turpmākās attīstības procesos. Kā norādīts, notohorda primordium izraisa nervu caurules veidošanos, un tās zudums izraisa attīstības pārtraukšanu. Tāpēc izmaiņas agrīnā stadijā parasti izraisa indivīda nepietiekamu attīstību un nāvi. Gluži pretēji, izmaiņas vēlākos posmos, kas ietekmē mazāk nozīmīgas pazīmes, var būt labvēlīgas organismam un šādos gadījumos tiek uztvertas dabiskās atlases ceļā.
Mūsdienu dzīvnieku embrionālajā attīstības periodā to tālajiem senčiem raksturīgo īpašību parādīšanās atspoguļo orgānu struktūras evolūcijas pārmaiņas.
Savā attīstībā organisms iziet vienšūnas stadiju (zigotas stadiju), ko var uzskatīt par primitīvās amēbas filoģenētiskās stadijas atkārtojumu. Visiem mugurkaulniekiem, arī to augstākajiem pārstāvjiem, veidojas notohords, kuru pēc tam aizstāj mugurkauls, un viņu senčiem, spriežot pēc lancetītes, notohords palika uz mūžu. Embrionālās attīstības laikā putniem un zīdītājiem, tostarp cilvēkiem, rīklē parādās žaunu spraugas un atbilstošas starpsienas. Žaunu aparāta daļu veidošanās fakts sauszemes mugurkaulnieku embrijos ir izskaidrojams ar to izcelsmi no zivīm līdzīgiem senčiem, kuri elpoja ar žaunām. Cilvēka embrija sirds uzbūve agrīnā veidošanās periodā atgādina šī orgāna uzbūvi zivīs: tam ir viens ātrijs un viens sirds kambaris. Bezzobu vaļiem embrija periodā attīstās zobi. Šie zobi nešķiļas, tie tiek iznīcināti un izšķīdināti.
Šeit minētie un daudzi citi piemēri norāda uz dziļu saikni starp organismu individuālo attīstību un to vēsturisko attīstību. Šī saikne ir izteikta F. Millera un E. Hekela 19. gadsimtā formulētajā bioģenētiskajā likumā: katra indivīda ontoģenēze (individuālā attīstība) ir īss un straujš tās sugas filoģenēzes (vēsturiskās attīstības) atkārtojums, kurai šis indivīds pieder. .
Secinājums
Pabeidzot darbu pie abstraktā, varam nonākt pie secinājuma, ka spēja vairoties jeb pašreproducēt ir viena no svarīgākajām organiskās dabas īpašībām. Reprodukcija ir īpašība, kas raksturīga visiem dzīviem organismiem bez izņēmuma, no baktērijām līdz zīdītājiem.
Jebkuru dzīvnieku un augu sugu, baktēriju un sēņu pastāvēšana, nepārtrauktība starp vecākiem indivīdiem un viņu pēcnācējiem tiek uzturēta tikai ar vairošanos. Cieši saistīta ar pašvairošanos ir vēl viena dzīvo būtņu īpašība.organismu attīstība. TasTas ir arī raksturīgs visai dzīvībai uz Zemes: mazākajiem vienšūnu organismiem un daudzšūnu augiem un dzīvniekiem.
Bibliogrāfija
Bogens G. Mūsdienu bioloģija. - M.: Mir, 1970.
Green N., Stout W., Taylor D. Biology: 3 sējumos T. 3: tulk. no angļu valodas/red. R. Sopers. - M.: Mir, 1990.
Mamontovs S.G. Bioloģija. Vispārīgi modeļi. M.: Bustards, 2002.
No dzīvniekiem līdz cilvēkiem. M.: Nauka, 1971. gads.
Slyusarev A.A. Bioloģija ar vispārējo ģenētiku. - M.: Medicīna, 1978.
Lekcija Nr.3 Ontoģenēze
1. Gametoģenēze
2. Embrionālais periods
3. Postembrionālais periods
Ontoģenēze– organisma individuālā attīstība no zigotas līdz bioloģiskai nāvei. Proģenēze– periods pirms ontoģenēzes, ietver gametoģenēzi un apaugļošanu. Filoģenēze– sugas evolucionārā attīstība.
Cilvēkiem ir raksturīga seksuāla vairošanās, to raksturo: dzimumdziedzeru, gametu klātbūtne, apaugļošanās process un dzimumdimorfisms.
Gametoģenēze - dzimumšūnu veidošanās process ovogeneze - oocītu nobriešana, spermatoģenēze- spermatozoīdi. Gametoģenēze notiek ķermeņa dzimumdziedzeros. Gametes veidojas no embrionālajām epitēlija šūnām, kas veidojas organisma embrionālajā attīstības periodā.
Veidošanās procesā dzimumšūnas iziet trīs posmus:
1. Reprodukcijas periods (embrionālā epitēlija šūnas dalās mitozes ceļā);
2. Augšanas periods;
3. Nobriešanas periods, šūnas dalās ar mejozi, kā rezultātā veidojas gametas (5. att.).
Rīsi. 5. Dzimumšūnu dalīšanās
Sperma nobriest sēklinieku sēklinieku kanāliņos. Atrodas starp sēklu kanāliņiem intersticiālie audi, kas ražo vīriešu dzimuma hormonu - testosterons. Vīriešu dzimuma hormoni - androgēni, regulē reproduktīvo funkciju, gametoģenēzi un sekundāro seksuālo īpašību veidošanos. Sperma nobriest 70 dienu laikā. Sēklu kanāliņos ir gametas dažādos nobriešanas posmos. 5 ml. Cilvēka sēklas šķidrums satur 12 miljonus spermatozoīdu. Tie ir negatīvi uzlādēti, lādiņš neļauj tiem salipt kopā. Liels skaits vīriešu dzimumšūnu ir bioloģiski lietderīgi, to skaita samazināšanās par 40% padara apaugļošanās procesu neiespējamu. Spermatozoīdi ir mazas, kustīgas šūnas, kas sastāv no galvas, kakla un astes. Uz galvas ir akrosoms, modificēta lizosoma, tajā ir fermenti, kas apaugļošanas laikā izšķīdina olšūnas membrānu. Dzīves ilgums ir no 6 līdz 72 stundām.
Ooģenēze sākas sievietes ķermeņa embrionālajā attīstības periodā. Pirmā mejozes dalījuma profāzes stadijā, kad notiek konjugācija un šķērsošana, iestājas īslaicīga pauze. Turpmāka olšūnas attīstība atsākas meitenes pubertātes laikā. Sieviešu dzimumhormonu – estrogēnu ietekmē ap pirmās kārtas olšūnu veidojas folikuls, šūnu pūslītis, kas aizsargā un baro šūnu.
Kad folikuls nobriest, tas virzās uz olnīcas malu un pēc tam pārsprāgst; šo olšūnas attīstības stadiju sauc par olnīcu vai folikulu; tas ilgst 12 dienas. Folikula plīsumu un olšūnas izdalīšanos no olnīcas vēdera dobumā sauc par ovulāciju.
Pēc ovulācijas ir iespējama apaugļošanās. Sieviešu dzimumšūnu satver olvadu fimbrijas. Ar skropstu epitēlija palīdzību tas iekļūst dzemdē, dzemdes stadija ilgst 12-14 dienas. Līdz tam laikam sieviešu dzimuma hormonu ietekmē dzemdes epitēlijs ir atslābināts. Ja apaugļošanās nenotiek, dzemdes sieniņu gludo muskuļu kontrakcijas rezultātā no organisma izdalās epitēlijs un olšūnas. Šo procesu sauc par menstruāciju un ilgst 3-4 dienas. Tādējādi laiks no folikulu veidošanās sākuma līdz menstruācijas sākumam ir 20-30 dienas, un to sauc par olnīcu-menstruālo ciklu (6. att.).

Rīsi. 6. Ovulācija un olšūnas implantācija dzemdē
Tas iekļauj olnīcu stadija, dzemde un mensis. Olnīcu-menstruālā cikla laikā tiek rekonstruēta visa sievietes reproduktīvā sistēma, mainās hormonu kvalitatīvais un kvantitatīvais sastāvs, mainās veiktspēja, spēks, lokanība un izturība. Organizējot treniņu procesu, tiek ņemtas vērā fizioloģiskās izmaiņas sievietes ķermenī.
Mēslošana- Tā ir olšūnas un spermas saplūšana. Ir trīs mēslošanas posmi:
1) akrosomāla reakcija (20 sekundes) – olas čaumalas izšķīšana ar akrosomu enzīmu palīdzību un spermas iekļūšana olā. Tiek uzskatīts, ka spermatozoīdi, kas rada vislielāko iespiešanās potenciālu (uztraukumu) uz olšūnas membrānas, iekļūst olšūnā;
2) īslaicīga pauze – olšūnā nenotiek redzamas izmaiņas, pastiprinās vielmaiņa;
3) olšūnas un spermas kodolu saplūšana, diploīdā hromosomu komplekta atjaunošana.
Apaugļošanās notiek olvadu augšējā trešdaļā. Apaugļoto olu sauc zigota. Ar zigotas veidošanos sākas cilvēka ontoģenēze.
2. Ontoģenēze cilvēks ietver divus posmus: embrionālais un postembrionālais.
Embrionālā attīstības stadija turpinās no zigotas līdz dzimšanai. Ietver stadijas: zigota, blastula, gastrula, histoģenēze, organoģenēze.
Zigota- atkārtoti sadalās mitozes ceļā, kā rezultātā veidojas daudzšūnu vienslāņa embrijs, blastula. Blastulas izmērs nepalielinās, jo šajā laikā tā atrodas olvados un tai vajadzētu brīvi pārvietoties dzemdē. Sestajā dienā tas nonāk dzemdē un piestiprinās pie tās sienas, iegremdējot atslābinātajā epitēlijā. Šo procesu sauc implantācija. Embrijs turpina attīstīties un kļūst divslāņu, pēc tam trīsslāņu. Šajā attīstības stadijā to sauc gastrula. Rezultātā veidojas trīs dīgļu slāņi: ektoderma, endoderma, mezoderma. Audi un orgāni veidojas no embriju sistēmām.
Tādējādi: blastula- viena slāņa embrijs, gastrula- divu un trīs slāņu embrijs. Histoģenēze– audu atdalīšana no dīgļu slāņiem. Organoģenēze- orgānu likšana. 8 nedēļu vecumā cilvēka embrija masa ir 4 g, izmērs 5 mm. Līdz tam laikam viņš bija izgājis veidošanās procesus un ieguvis cilvēka ķermeņa aprises. Pēc astoņām grūtniecības nedēļām cilvēka embriju sauc par augli.
Cilvēka embrija attīstības laikā, pagaidu iestādes, kas pēc piedzimšanas zaudē savas funkcijas. Pagaidu iestādes ietver: dīgļučaumalas, horions, amnions un placenta, nabassaite. Placenta nodrošina ciešu kontaktu starp bērna un mātes kapilāriem. Attīstošais organisms caur placentu saņem barības vielas, skābekli, antivielas, izvada vielmaiņas produktus. Placenta veic barjerfunkciju, aizsargājot embriju no mikroorganismiem, dažām zālēm un toksīniem (7. att.).

Rīsi. 7. Placenta
1 – dzemde; 2 – amnija; 3 – embrijs; 4 – amnija šķidrums; 5 – placenta; 6 – placentas kapilāri; 7 – vēnas; 8 – artērijas
Bērna orgānu un audu smalkie veidošanās mehānismi var tikt traucēti, saskaroties ar toksiskām, narkotiskām vielām, alkoholu, nikotīnu, kas atrodas mātes asinīs. Cilvēka embrionālās attīstības traucējumus sauc par iedzimtām deformācijām un malformācijām. Tie ietver: “aukslēju šķeltne” (augšējo aukslēju neesamība), “lūpas šķeltne” (augšžokļa kaulu nesaaugšana), polidaktilija (papildu pirksti), sindaktilija (saauguši pirksti). Tiek saukti nelabvēlīgi vides faktori, kas izraisa iedzimtas anomālijas teratogēns(izraisot deformācijas) (8. att.).

Rīsi. 8. Cilvēka organisma attīstību ietekmējošie vides faktori
Cilvēka attīstības embrionālais periods ilgst 36 nedēļas.
3. Tiek saukts periods pēc dzimšanas un pirms bioloģiskās nāves pēcdzemdību periodā. Tas iekļauj nepilngadīgais, pubertāte attīstības un novecošanas periodi.
Nepilngadīgais – periods pirms pubertātes. Puberitāte- pubertātes periods.
Sporta praksē, fizioloģijā, pedagoģijā, psiholoģijā tiek izmantota šāda vecuma periodizācija Cilvēka postembrionālā ontoģenēze.
Laika posmā no 1 līdz 10 dienām bērns tiek izsaukts jaundzimušais;
No 10 dienām līdz 1 gadam - krūtis;
no 1 gada līdz trim – periods Agra bērnība;
no 4 līdz 7 gadiem - pirmā bērnība;
no 8 gadiem līdz 12 meitenēm un līdz 13 zēniem otrā bērnība;
no 12 līdz 15 gadiem meitenēm un no 13 līdz 16 zēniem - pusaudžiem;
17-21 (zēni), 16-20 gadi (meitenes) - pusaudža gados;
20-55 sievietēm un 21-60 vīriešiem periods puberitāte(puberitāte);
no 55 un 60 līdz 70 — gados veci cilvēki;
no 70 līdz 90 veči;
vairāk nekā 90 simtgadnieki.
Cilvēka ontoģenēzē ir jūtīgs un kritisks periodi.
Kritiskie periodi ir raksturīga paaugstināta atsevišķu gēnu un to kompleksu aktivitāte, kas kontrolē jebkādu ķermeņa pazīmju attīstību. Šajos periodos notiek būtiska regulējošo procesu pārstrukturēšana, kvalitatīvs un kvantitatīvs lēciens atsevišķu orgānu un funkcionālo sistēmu attīstībā. Šajos periodos organisms ir visjutīgākais pret vides faktoru ietekmi. Par kritiskajiem ontoģenēzes periodiem tiek uzskatīta implantācija, placentācija, aksiālo orgānu veidošanās, nervu un zarnu caurules, notohordi, sirds veidošanās un citi.
Ja sieviete saslimst ar masaliņām no 3. līdz 9. grūtniecības nedēļai, pastāv risks, ka auglim attīstīsies tādi defekti kā sirds slimības, katarakta un kurlums. Citreiz masaliņas neizraisa augļa anomālijas.
Jutīgi periodi– tie ir periodi, kad tiek samazināta ģenētiskā kontrole un paaugstināta organisma individuālo īpašību jutība pret vides ietekmi, ieskaitot pedagoģisko un koučingu. Tādējādi jutīgais periods dažādu ātruma kvalitātes rādītāju izpausmei iestājas 11-14 gadu vecumā un līdz 15 gadu vecumam tiek sasniegts tā maksimālais līmenis, kad iespējami augsti sportiski sasniegumi. Līdzīga aina ir vērojama ontoģenēzē attiecībā uz veiklības un lokanības īpašību izpausmēm.
Treneriem un skolotājiem, kas strādā fiziskās audzināšanas un sporta jomā, zināšanām par jutīgajiem periodiem ir liela praktiska nozīme, jo jutīgajos periodos vislielākā apmācības efekts.
Tās var iedalīt atkarībā no tā, cik šūnu ir iesaistītas aseksuālās vairošanās procesā: aseksuāla vairošanās, kurā no vienas šūnas rodas meitas paaudze: šūnu dalīšanās vairāku šūnu dalīšanās šizogonija sporulācija sporulācija, kas veidojas vienšūnu raugā...
Kopīgojiet savus darbus sociālajos tīklos
Ja šis darbs jums neder, lapas apakšā ir līdzīgu darbu saraksts. Varat arī izmantot meklēšanas pogu
Organismu vairošanās un attīstība
Plānot
- Reprodukcijas jēdziens un nozīme.
- Reprodukcijas formas un veidi.
- Šūnu cikls. Mitoze. Mejoze.
- Gametu struktūra. Gametoģenēze.
1. Reprodukcijas jēdziens un nozīme
Viena no dzīvo būtņu īpašībām ir diskrētums, tie. jebkurā organizācijas līmenī dzīvo vielu pārstāv elementāras struktūrvienības. Katrs noteiktas sugas indivīds ir mirstīgs, un sugas eksistence tiek saglabāta pavairošana organismiem. Tādējādi dzīves diskrētums paredz tās atražošanu, t.i. reprodukcijas process.
Pavairošana tā ir dzīvo būtņu spēja pavairot savu veidu. Tas nodrošina dzīves nepārtrauktību un nepārtrauktību. Nepārtrauktību nosaka tas, ka vairošanās procesā ģenētiskais materiāls tiek nodots no vecākiem uz pēcnācējiem, kā rezultātā vecāku īpašības tādā vai citādā mērā izpaužas meitas organismos.
Reprodukcijas vērtība:
- Sugas skaita palielināšana vai saglabāšana. Vairošanās dēļ notiek ne tikai vairošanās (t.i., sava veida vairošanās), bet arī dzīvo organismu skaita pieaugums.
- Dzīves nepārtrauktība. Pateicoties vairošanai, vairāk vai mazāk lielas indivīdu grupas (piemēram, populācijas un sugas) var pastāvēt diezgan ilgu laiku, jo to skaita samazināšanos indivīdu dabiskās nāves dēļ kompensē pastāvīga organismu vairošanās un mirušo aizstāšana ar jaundzimušajiem.
- Nepārtrauktība starp paaudzēm. Reprodukcijas procesā ģenētiskā informācija tiek nodota no vecāku paaudzes indivīdiem, nodrošinot konkrētu vecāku un visas sugas, pie kuras pieder organisms, īpašību atražošanu.
- Tiek realizētas tādas dzīvo būtņu īpašības kā iedzimtība un mainīgums.
2. Reprodukcijas formas un veidi
Ir divas galvenās reprodukcijas formas: seksuālā un aseksuālā.
Aseksuāla reprodukcija
Aseksuālās vairošanās laikā no nespecializētām nereproduktīvām ķermeņa somatiskajām šūnām parādās jauns indivīds. Tāpēc aseksuālā vairošanās ietver vienu indivīdu. Aseksuālā vairošanās nodrošina liela skaita identisku īpatņu vairošanos, kas ir izdevīgi konstantos apstākļos dzīvojošām sugām. Mātei identisku pēcnācēju ātra un daudzveidīga vairošanās ir aseksuālās vairošanās bioloģiskā nozīme.
Dabā ir dažādas bezdzimuma pavairošanas iespējas:dalīšanās, sporulācija, sadrumstalotība, pumpuru veidošanās, veģetatīvā pavairošana, klonēšana.
Tos var iedalīt atkarībā no tā, cik šūnu ir iesaistītas aseksuālās reprodukcijas procesā:
- aseksuāla reprodukcija, kurā meitas paaudze rodas no vienas šūnas:
- šūnu dalīšanās
- vairāku šūnu dalīšanās (šizogonija)
- sporulācija (sporulācija)
- pumpuru veidošanās vienšūnu organismos (raugs)
2) bezdzimuma vairošanās, kuras pamatā ir šūnu grupas dalīšanās:
- veģetatīvs
- sadrumstalotība
- daudzšūnu organismu veidošanās (hidra)
Ir arī cita klasifikācija, saskaņā ar kuru bezdzimuma reprodukcijas iespējas ir:
- veģetatīvā pavairošana (t.i., dalīšanās pa mātes ķermeņa daļām):
Divīzija
Daudzkārtēja skaldīšanās (šizogonija)
Budding
Sadrumstalotība
Augu pavairošana ar spraudeņiem, sīpoliem, lapām, sakneņiem
- vairošanās ar sporām, t.i. sporulācija
Sīkāk apskatīsim aseksuālās pavairošanas veidus.
Divīzija. Vienkāršākā aseksuālās vairošanās forma. Ir dažādas sadalīšanas iespējas:
- Sadalīšana ar vienkāršu sašaurināšanos ar divu meitas organismu veidošanos no viena vecāka organisma. Raksturīgs baktērijām un zilaļģēm.
- Sadalīšanās ar mitotisku kodola dalīšanu, kam seko citoplazmas atdalīšana. Raksturīgi vienšūnu organismiem (daudzi vienšūņi - amēba, zaļā eiglēna u.c.; vienšūnas aļģes - hlamidomonas u.c.).
Abos gadījumos tas notiek binārs nodaļa, t.i. divās šūnās. Tomēr ir iespējama arī cita iespēja:
- Daudzkārtēja skaldīšana (šizogonija). Pirmkārt, kodols sadalās atkārtoti un pēc tam citoplazma sadalās daļās. Piemēram, Plasmodium falciparum (malārijas izraisītāja) vairošanās cilvēka eritrocītos. Šajā gadījumā plazmodijai vairākas reizes notiek atkārtota kodola dalīšanās, pēc kuras citoplazma sadalās. Rezultātā 1 plazmodijs rada 12-24 meitas organismus.
Sporu veidošanās (sporulācija). Ļoti plaši izplatīta aseksuālās pavairošanas metode dzīvo organismu vidū un ir sastopama gandrīz visos augos, sēnēs un dažos vienšūņos (piemēram, sporozoju tipa), kā arī prokariotiskajos organismos (daudzās baktērijās, zilaļģēs).
Spora Šī ir šūna, kas pārklāta ar attīstītu aizsargpārklājumu - sporu membrānu, kas ļauj tai izturēt dažādu nelabvēlīgu vides faktoru ietekmi.Mazā izmēra dēļ sporas parasti satur tikai minimālas barības vielu rezerves. Daudzos organismos tā ir galvenā izkliedes vienība, jo liels daudzums izveidoto vieglo sporu tiek brīvi transportēts lielos attālumos, pārvietojoties gaisa masām un ūdens plūsmām.Bieži vien sporas veidojas lielos daudzumos un tām ir niecīgs svars, tāpēc tās vieglāk izplatās vējš, kā arī dzīvnieki, galvenokārt kukaiņi.
Turklāt dažās formās (vienšūņi, sēnes) sporas var darboties kā īpašs dzīves cikla posms, ļaujot tām “pārdzīvot” nelabvēlīgu vides faktoru ietekmi.Stingri sakot, baktēriju sporas nekalpo vairošanai, bet gan izdzīvošanai nelabvēlīgos apstākļos, jo katra baktērija ražo tikai vienu sporu. Baktēriju sporas ir vienas no izturīgākajām: piemēram, tās bieži var izturēt apstrādi ar spēcīgiem dezinfekcijas līdzekļiem un vārīšanu ūdenī.
Daudzos augos sporu veidošanās process(sporoģenēze) tiek veikta īpašās maisiņiem līdzīgās struktūrās, ko sauc par sporangijām. Sporas var būt vai nu kustīgas karogveida aparāta klātbūtnes dēļ (šajā gadījumā tos sauc zoosporas), un nekustīgs, kam liegta iespēja aktīvi kustēties.
Zoosporas tiek novērotas dažās zaļajās aļģēs. Dzīvnieku vidū sporulāciju novēro malārijas plazmodijā un veselā sporozoīdu grupā (vienšūnu organismi).
Budding. Tas ir sastopams gan vienšūnu organismos, piemēram, raugos un dažos ciliātu veidos, gan daudzšūnu organismos, piemēram, koelenterāta tipa (hidrās) pārstāvjiem, kā arī tunikātiem (klases ascīdiem).
Vienšūnu organismiem šī metode sastāv no tuberkulozes (izauguma) veidošanās ar kodolu uz mātes šūnas, kas pēc tam atdalās un kļūst par neatkarīgu organismu.
Daudzšūnu organismiem šī metode sastāv no tā, ka vispirms uz mātes indivīda ķermeņa parādās neliels tuberkulozes izmērs, palielinoties izmēram, pēc tam parādās visu mātes organismam raksturīgo struktūru un orgānu pamati. Pēc tam notiek meitas indivīda atdalīšanās (budrošanās). Pēc tam jaunais, tikko atdalītais organisms aug un sasniedz sākotnējā parauga izmēru.
Rīsi. Topošais organisms koelenterātos (hidrās), 1 pieaugušais organisms, 2 meitas topošais organisms.
Sulīgam augam ir aprakstīta neparasta pumpuru veidošanās forma Bryophyllum - kserofīts, ko bieži audzē kā dekoratīvu telpaugu: miniatūri augi, kas aprīkoti ar mazām saknēm, attīstās gar tā lapu malām; šie "pumpuri" galu galā nokrīt un sāk pastāvēt kā neatkarīgi augi.
Sadrumstalotība indivīda sadalīšana divās vai vairākās daļās, no kurām katra attīstās par jaunu organismu. Sadrumstalošanās notiek, piemēram, pavedienveida aļģēs, piemēram, Spirogyra. Spirogyra pavediens jebkurā vietā var sadalīties divās daļās, no kurām pēc tam veidojas divi organismi.
Fragmentācija tiek novērota arī dažiem zemākiem dzīvniekiem, kuri atšķirībā no augstāk organizētām formām saglabā ievērojamu spēju atjaunoties no salīdzinoši vāji diferencētām šūnām. Piemēram, nemertiešu (primitīvu tārpu grupa, galvenokārt jūras) ķermenis ir īpaši viegli saplēsts daudzās daļās, no kurām katra atjaunošanās rezultātā var radīt jaunu indivīdu. Šajā gadījumā reģenerācija ir normāls un regulēts process; tomēr dažiem dzīvniekiem (piemēram, jūras zvaigznēm) atjaunošana no atsevišķām daļām notiek tikai pēc nejaušas sadrumstalotības. Sadrumstalotību novēro sūkļiem, koelenterātiem (hidrās), medūzām, adatādaiņiem, annelīdiem un plakanajiem tārpiem. Dažreiz šī spēja ir tik labi attīstīta, ka no atsevišķa fragmenta tiek atjaunots vesels indivīds.
Rīsi. . Jūras zvaigznes reģenerācija no viena stara. A, B, C secīgi atjaunošanās posmi
Veģetatīvā pavairošanatā ir vairošanās, kurā no mātes organisma atdalītas daļas attīstās jauns meitas organisms. Šajā gadījumā no mātes īpatņa tiek atdalīta diezgan labi diferencēta daļa, kas tālāk attīstās par neatkarīgu augu. Vai arī augs veido īpašas struktūras, kas īpaši paredzētas veģetatīvai pavairošanai. Piemēram, sīpoli, bumbuļi, bumbuļi, sakneņi, stīgas, pumpuri. Dažas no šīm struktūrām kalpo arī barības vielu uzglabāšanai, ļaujot augam izdzīvot nelabvēlīgos apstākļos, piemēram, aukstumā vai sausumā. Veģetatīvā pavairošana ir raksturīga daudzām augu grupām, sākot no aļģēm un beidzot ar ziedaugiem.
Principā veģetatīvā pavairošana praktiski neatšķiras no sadrumstalotības vai pumpuru veidošanās, taču tradicionāli šis termins tiek attiecināts uz augu organismiem un tikai dažkārt uz dzīvniekiem, pretstatā sadrumstalotībai un pumpuriem.
Klonēšana. Šī ir mākslīga vairošanās metode, kas dabiski nenotiek. Tas ir kļuvis plaši izplatīts tikai pēdējos 30-40 gados un arvien vairāk tiek izmantots ekonomiskiem mērķiem. Ir vairākas īpašas metodes, kas ļauj klonēt dažus augus un dzīvniekus. Klonēt attiecas uz ģenētiski identiskiem pēcnācējiem, kas iegūti no viena indivīda vienas vai otras aseksuālās pavairošanas metodes rezultātā. No šejienes cēlies šīs metodes nosaukums.
Īpaši plaši klonēšanas eksperimenti tiek veikti ar augiem, kas ir saistīts ar to augsto spēju atjaunoties. Atsevišķas šūnas novieto uz barības vielu barotnēm, kur tās sadalās un, izmantojot īpašus paņēmienus, iegūst neorganizētu šūnu masu, vai kalluss. Pēc tam tie izraisa primāro viendabīgo kauliņu diferenciāciju un dažādu audu un orgānu veidošanos un, visbeidzot, veselu augu organismu, kam ir visas tās pašas īpašības kā sākotnējam, no kura tika ņemtas šūnas.
Izmantojot klonēšanas metodi, var iegūt dažādas hibrīda formas. Tādējādi ar enzīmu vai ultraskaņas palīdzību tiek noņemtas augu šūnu šūnu sienas, pēc kurām var saplūst iegūtie “kaili” protoplasti, kā rezultātā veidojas hibrīdšūnas (piemēram, tomātu-kartupeļu vai tabakas-petūnijas hibrīdi). Pēc tam tiek atjaunotas šūnu sienas, veidojas kaluss un pēc tam vesels hibrīds augs.
Dzīvnieku gadījumā izmanto šādu metodi: olas kodolu izņem vai iznīcina, un tā vietā ievieto somatiskās šūnas (piemēram, epitēlija šūnas) kodolu. Nākotnē no šādas olšūnas var iegūt organismu, kas pēc īpašībām ir identisks dzīvnieka kodola donoram. Tādā veidā jūs varat iegūt dažu dzīvnieku spīļo varžu klonus(Xenopus), tritoni (Triturus). Šobrīd ir iegūti pat zīdītāju kloni, piemēram, labi zināmā aita Dollija. Klonēšanas tehnikas izstrādi veic īpaša bioloģijas nozare - biotehnoloģija, kuras uzdevumi neaprobežojas ar to.
Seksuālā reprodukcija
Seksuālā reprodukcijako sauc par paaudžu maiņu un organismu attīstību, kuras pamatā ir specializētu dzimumšūnu saplūšana ( gametas ) un zigotas veidošanos. Gonādos notiek gametu (sieviešu un vīriešu, olšūnu un spermatozoīdu) veidošanās.
Seksuālā vairošanās ir raksturīga lielākajai daļai dzīvo būtņu, tā ir progresīvāka salīdzinājumā ar aseksuālo vairošanos, un tai ir milzīgas ģenētiskas priekšrocības. Dzimumvairošanās vislabāk nodrošina pēcnācēju ģenētisko daudzveidību, jo ir gēnu kombinācija, kas iepriekš piederēja abiem vecākiem. Sugu veidojošo īpatņu genotipu daudzveidība nodrošina iespēju sugai veiksmīgāk un ātrāk pielāgoties mainīgajiem vides apstākļiem.
Apaugļošanas laikā gametas saplūst, veidojot diploīdu zigota no kuras attīstības procesā iegūst nobriedušu organismu. Gametes ir haploīdas – tās satur vienu hromosomu komplektu, zigota ir diploīda, satur dubultu hromosomu komplektu. Šī ir pirmā nākamā organisma šūna.
Gametes parasti ir divu veidu - vīriešu un sieviešu. Tos ražo vīriešu un sieviešu vecāki, ja suga ir divmāju; ( tādi ir daži ziedoši augi, lielākā daļa dzīvnieku un cilvēku) vai viena un tā paša indivīda (farmafrodītisms).
Raksturīgs vīriešu un sieviešu kārtas dzīvniekiemseksuālais dimorfisms- t.i. seksuālās atšķirības struktūrā, izskatā (izmērā, krāsā un citās īpašībās), kā arī uzvedībā. Dzīvniekiem tas sastopams jau zemākajos evolūcijas attīstības posmos, piemēram, apaļajiem helmintiem un posmkājiem, un vislielāko izpausmi sasniedz mugurkaulniekiem, kuros ārējās atšķirības starp tēviņiem un mātītēm ir ļoti izteiksmīgas. To sugu augos, kam raksturīga vīrišķo un mātīšu īpatņu klātbūtne, notiek arī dzimumdimorfisms, taču tas ir ļoti vāji izteikts.
Seksuālās un aseksuālās vairošanās salīdzinājums dots tabulā. 5
Tabula 5
Aseksuālās un seksuālās vairošanās salīdzinājums
|
Aseksuāla reprodukcija |
Seksuālā reprodukcija (izņemot baktērijas) |
|
|
Viens vecāks |
Parasti divi vecāki |
|
|
Gametes nav izveidotas |
Veidojas haploīdas gametas, kuru kodoli saplūst (apaugļošanās), veidojot diploīdu zigotu |
|
|
Nav mejozes |
Kādā dzīves cikla posmā notiek mejoze, kas neļauj hromosomām dubultoties katrā paaudzē. |
|
|
Pēcnācēji ir identiski saviem vecākiem. Vienīgais ģenētiskās variācijas avots ir nejaušas mutācijas |
Pēcnācēji nav identiski vecāku personas. Viņiem ir ģenētiska variācija, kas rodas ģenētiskās rekombinācijas rezultātā. Tas veicina spēcīgāko un piemērotāko indivīdu dabisko atlasi un līdz ar to arī evolūciju. |
|
|
Raksturīgs augiem, dažiem zemākiem dzīvniekiem un mikroorganismiem. Nav sastopams augstākiem dzīvniekiem |
Raksturīgs lielākajai daļai augu un dzīvnieku |
|
|
Bieži vien tas izraisa ātru lielu pēcnācēju skaitu |
Mazāk straujš skaitļu pieaugums |
Seksuālā reprodukcija atkarībā no dzimumšūnu lieluma un funkcijas attiecības var būt trīs iespējas:
Izogāmija (aļģes, vienšūņi). Vīriešu un sieviešu reproduktīvās šūnas ir identiskas pēc izmēra, struktūras un mobilitātes.
Heterogāmija. Šūnas atšķiras pēc izmēra un struktūras.
Oogamy. Šis ir viens no heterogāmijas variantiem, kad olšūna ir liela un nekustīga, un spermatozoīds ir mazāks, tam ir kustības orgāns un ir mobilitāte.
Ir īpašas seksuālās vairošanās formas, piemēram, hermafrodītisms un partenoģenēze.
Hermafrodītisms. Termins "hermafrodītisms" ir grieķu vārdu Hermes (vīriešu skaistuma dievs) un Afrodītes (sieviešu skaistuma dieviete) kombinācija.
Tomēr lielākajā daļā hermafrodīto sugu apaugļošana ietver gametas, kas iegūtas no dažādiem indivīdiem, un tām ir daudz ģenētisku, morfoloģisku un fizioloģisku pielāgojumu, kas novērš pašapaugļošanos un veicina savstarpēju apaugļošanu. Piemēram, daudziem vienšūņiem pašapaugļošanos kavē ģenētiskā nesaderība, daudzos ziedos augos androecium un ginoecium struktūra, kā arī daudziem dzīvniekiem tas, ka olas un spermatozoīdi veidojas vienā un tajā pašā indivīdā dažādos laikos.
Patiesa hermafrodītisma parādība ir sastopama arī organizētākās būtnēs. Jo īpaši tas ir atrodams zīdītājiem. Piemēram, cūkām dažkārt vienā ķermeņa pusē tiek novērota olnīcu attīstība, bet otrā – sēklinieku (sēklinieku) attīstība vai kombinētu struktūru (ovotestes) attīstība, un abos gadījumos notiek sintēze. notiek funkcionāli aktīvo olšūnu un spermatozoīdu daudzums. Šādi dzīvnieki tiek klasificēti kā “vidējais” dzimums, un lielākā daļa vidēja dzimuma īpatņu ir mātītes ar divām XX hromosomām. Līdzīga parādība ir novērota kazām.
Patiess hermafrodītisms rodas arī cilvēkiem, kas izriet no attīstības traucējumiem. Hermafrodītu genotipi ir 46XX vai 46 XY , vairumā gadījumu atsaucoties uz XX (apmēram 60%). Zināms arī viltus hermafrodītisms, kad indivīdiem ir abiem dzimumiem raksturīgi ārējie dzimumorgāni un sekundāras dzimumpazīmes, bet veidojas tikai viena tipa dzimumšūnas – vīrišķās vai sievietes.
Lielākajai daļai ziedošu augu ir hermafrodīti ziedi, kurus parasti sauc par biseksuāliem, jo katrs zieds satur piestiņu un putekšņlapas. Šī iemesla dēļ augļi attīstās no visiem ziediem. Kvieši, ķirši, āboli un daudzas citas augu sugas ir biseksuālas. Līdzās biseksuāļiem evolūcijas gaitā attīstījās augi ar dzimumu atdalīšanu vienas sugas ietvaros, t.i., radās vienmāju un augu divdzimums. Augus, kas satur gan sārtaino (sievišķo), gan stainatisko (vīrišķo) ziedu, sauc par vienmāju. Vienmāju augos augļi attīstās tikai no piestātveida ziediem. Kukurūza ir vienmājugurķi, ķirbi un citi. Turpretim divmāju augi ir augi, kas satur vai nu sēnīšus, vai stainveida ziedus (tās pašas sugas ietvaros). Divmāju augos augļus nes tikai tie, kuriem ir spārnu ziedi (mātītes). Papeles, zemenes un cita veida kokaugi un lakstaugi ir divmāju.
Partenoģenēze (no grieķu val. partenoss jaunava unģenēze dzimšana) ir viena no seksuālās vairošanās modifikācijām, kurā sievietes dzimumšūna attīstās par jaunu indivīdu bez vīrišķās dzimumšūnas apaugļošanās. Partenoģenēzes priekšrocība ir tāda, ka dažos gadījumos tas palielina reprodukcijas ātrumu, un sociālajos kukaiņos tas ļauj regulēt katra veida pēcnācēju skaitu. Partenoģenēze var būt obligāta (obligāta) un fakultatīva (neobligāta).
Piemēram, medus bitē(Apis mellifera) karaliene dēj apaugļotas olas(2п = 32), kurām attīstoties rodas mātītes (mātītes vai strādnieces) un neapaugļotas olas(P = 16), kas ražo tēviņus (dronus), kas ražo spermīnu mitozes, nevis meiozes ceļā.
Rīsi. Partenoģenēzes veidu shēma.
- Šūnu cikls. Mitoze. Mejoze
Šūnu dalīšanās ir organismu vairošanās un individuālās attīstības pamats.
Visi dzīvie organismi sastāv no šūnām. Ķermeņa tipiskās struktūras attīstība, augšana un veidošanās tiek veikta, reproducējot vienu vai oriģinālo šūnu grupu. Dzīves procesā dažas ķermeņa šūnas nolietojas, noveco un iet bojā. Lai saglabātu struktūru un normālu darbību, ķermenim ir jāražo jaunas šūnas, lai aizstātu vecās. Vienīgais veids, kā veidot šūnas, ir sadalīt iepriekšējās.
Šūnu dalīšanās vitāli svarīgs process visiem organismiem. Cilvēka ķermenī, kas sastāv no aptuveni 10 13 šūnām, vairākiem miljoniem no tiem jādalās ik sekundi.
Ir aprakstītas trīs eikariotu šūnu dalīšanas metodes: amitoze (tiešā sadalīšana), mitoze (netiešais dalījums) un mejoze (samazināšanas nodaļa).
Amitoze salīdzinoši reta un maz pētīta šūnu dalīšanās metode. Tas ir aprakstīts novecojošām un patoloģiski izmainītām šūnām. Amitozes gadījumā starpfāzu kodols tiek sadalīts ar sašaurināšanos, un iedzimtā materiāla vienmērīga sadale netiek nodrošināta. Bieži vien kodols sadalās bez turpmākas citoplazmas atdalīšanas un veidojas divkodolu šūnas. Šūna, kas ir pakļauta amitozei, pēc tam nespēj iekļūt normālā mitotiskā ciklā. Tāpēc amitoze parasti rodas nāvei lemtajās šūnās un audos, piemēram, zīdītāju embriju membrānu šūnās un audzēju šūnās.
Mitoze universāla eikariotu šūnu dalīšanas metode. Tās ilgums dzīvnieku šūnās ir aptuveni 1 stunda Mitoze ir nepārtraukts process, ko nosacīti iedala četrās fāzēs: profāze, metafāze, anafāze un telofāze.
Tiek saukta notikumu secība, kas notiek starp dotās šūnas veidošanos un tās sadalīšanos meitas šūnāsšūnu cikls. Šis cikls sastāv no trim galvenajiem posmiem:
1. Starpfāze. Intensīvas sintēzes periods unizaugsmi. Šūna sintezē daudzas vielas, kas nepieciešamas tās augšanai un visu tai raksturīgo funkciju īstenošanai. Starpfāzes laikā notiek DNS replikācija.
2. Mitoze. Šis ir kodola dalīšanās process (kariokinēze), kurā hromatīdi tiek atdalīti viens no otra un pārdalīti kā hromosomas starp meitas šūnām.
3. Citokinēze ir citoplazmas dalīšanās process (citokinēze) starp divām meitas šūnām.
Šūnu cikla ilgums ir atkarīgs no šūnas veida un ārējiem faktoriem, piemēram, temperatūras, barības vielām un skābekļa. Baktēriju šūnas var dalīties ik pēc 20 minūtēm, zarnu epitēlija šūnas - ik pēc 8-10, šūnas sīpola saknes galā - ik pēc 20 stundām, un daudzas nervu sistēmas šūnas nekad nedalās.
Attiecības starp procesiem laika gaitā ir parādītas rīsi.

Rīsi . Šūnu cikla fāzes
Starpfāze sastāv no vairākiem periodiem: G 1, S, G 2.
Periods G 1 sauc par presintētisko. Visvairāk mainīgais ilgums. Šajā laikā šūnā tiek aktivizēti bioloģiskās sintēzes procesi, galvenokārt strukturālie un funkcionālie proteīni. Šūna aug un gatavojas nākamajam periodam. Šajā periodā notiek intensīvi biosintēzes procesi. Mitohondriju, hloroplastu (augos), endoplazmatiskā tīkla, lizosomu, Golgi aparāta, vakuolu un pūslīšu veidošanās. Kodols ražo rRNS, mRNS un tRNS; veidojas ribosomas; šūna sintezē strukturālās un funkcionālās olbaltumvielas. Intensīvs šūnu metabolisms, ko kontrolē fermenti. Šūnu augšana. Vielu veidošanās, kas nomāc vai stimulē nākamās fāzes iestāšanos.
Periods S sauc par sintētisko. Šis ir galvenais mitotiskā cikla periods. Daloties zīdītāju šūnās, tas ilgst apmēram 6 x 10 stundas.Šeit notiek DNS replikācija. Olbaltumvielu molekulu, ko sauc par histoniem, sintēze, kas saistās ar katru DNS virkni. Katra hromosoma pārvēršas par diviem hromatīdiem.
G2 periodu sauc postsintētisks.Tas ir salīdzinoši īss, zīdītāju šūnās apmēram 2 x 5 stundas.Šajā laikā centriolu, mitohondriju un plastidu skaits dubultojas, notiek aktīvi vielmaiņas procesi, uzkrājas olbaltumvielas un enerģija gaidāmajai dalīšanai. Šūna sāk dalīties. Notiek intensīvi biosintēzes procesi. Mitohondriju un hloroplastu sadalījums. Palielinātas enerģijas rezerves. Centriolu replikācija (šūnās, kur tie atrodas) un vārpstas veidošanās sākums
Mitoze nosacīti sadalīts četrās fāzēs: profāze, metafāze, anafāze un telofāze
Profāze . Sākas DNS spiralizācija un pakāpeniski palielinās kodolā. Hromosomas saīsinās, sabiezē, kļūst redzamas un iegūst tipisku bihromatīdu struktūru. Kodols pakāpeniski izzūd. Citoplazmā mikrotubulas ir orientētas ap katru centriolu pāri, veidojot vārpstas centrus. Centrioli pārvietojas uz dažādiem poliem, pa šūnas asi stiepjas mikrotubulas, un sākas ahromatīna vārpstas veidošanās. Kodola apvalks sadalās atsevišķos mazos fragmentos. Hromosomas virzās uz šūnas centru
Metafāze . Hromosomas ir maksimāli spiralizētas un sakārtotas tā, ka to centromēri atrodas vienā plaknē – šūnas ekvatora plaknē. Veidojas metafāzes plāksne.Mitozes vārpstas veidošanās ir pabeigta. Centrioli atrodas pa pāriem pretējos polios, un vārpstas vītnes no dažādiem poliem ir pievienotas katras hromosomas centromēram.
Anafāze . Šī ir īsākā mitozes fāze. Šeit notiek katras hromosomas gareniskā šķelšanās, pavedienu samazināšanāsvārpstas un hromatīdu (meitu hromosomu) diverģence pret šūnas poliem.
Telofāze . Meitas hromosomas, kas sastāv no viena hromatīda, sasniedz šūnas polus. DNS, kas tos veido, sāk despiralizēt, parādās kodols, ap katru meitas hromosomu grupu veidojas kodola membrāna, un ahromatīna vārpstas pavedieni pakāpeniski sadalās. Kodola skaldīšana ir pabeigta.
Sākas citoplazmas dalīšanās(citotomija) un starpsienu veidošanās starp meitas šūnām. Dzīvnieku šūnas veic citotomiju, sašaurinot citoplazmas membrānu. Augos šūnas ekvatora plaknē veidojas membrānas starpsiena, kas aug sāniski, sasniedzot šūnas sieniņu. Rezultātā veidojas divas pilnībā atdalītas meitas šūnas.
Sekosim līdzi iedzimtības materiāla izmaiņasmitotiskā cikla laikā. Galvenie mitotiskā cikla notikumi ir DNS replikācija kas notiek starpfāzē un noved pie iedzimtās informācijas apjoma dubultošanās, unhromatīdu segregācija,notiek mitozes anafāzē un nodrošina vienmērīgu iedzimtas informācijas sadalījumu starp meitas šūnām. Iedzimtais materiāls veic šos procesus, atrodoties dažādās strukturālās formās. Notiek replikatīva sintēzestarpfāzu hromatīns, inkurā DNS molekula atrodas relatīvi despiralizētā stāvoklī. Tiek veikta ģenētiskās informācijas izplatīšanamitotiskās hromosomas,kurā DNS ir maksimāli spiralizēta.
Mitotiskajā ciklā mainās arī iedzimtā materiāla daudzums. Ja hromosomu skaitu haploīdā komplektā apzīmē ar burtu p (diploīdā komplektā attiecīgi 2n), bet DNS molekulu skaitu apzīmē ar burtu ar, tad iespējams izsekot somatiskās šūnas kodola formulas izmaiņām dažādos mitotiskā cikla posmos. Pirms tam S -periods, kad katra hromosoma sastāv no vienas DNS molekulas, kopējais DNS daudzums kodolā atbilst hromosomu skaitam tajā un diploīdas šūnas formulai ir forma 2p2s. Pēc replikācijas, kad katras hromosomas DNS dubultojas, kopējais DNS daudzums kodolā dubultojas un šūnas formula iegūst formu 2n4c. Hromatīdu atdalīšanas rezultātā mitozes anafāzē meitas kodoli saņem diploīdu viena hromatīda hromosomu kopu. Meitas šūnu formula atkal kļūst 2p2s.
Mitozes bioloģiskā nozīmeir tas, ka šīs dalīšanas metodes rezultātā veidojas šūnas ar iedzimtu informāciju, kas kvalitatīvi un kvantitatīvi ir identiska mātes šūnas informācijai. Vienmērīgu iedzimtības materiāla sadalījumu nodrošina DNS replikācijas un hromosomu dubultošanās procesi mitotiskā cikla starpfāzē, kā arī spiralizācija un vienmērīga hromatīdu sadale starp meitas šūnām mitozes laikā. Mitoze nodrošina kariotipa noturības saglabāšanu vairākās šūnu paaudzēs un kalpo kā šūnu mehānisms ķermeņa augšanas un attīstības procesiem, kā arī reģenerācijai un aseksuālai reprodukcijai.
Vairāku vides faktoru darbība var izjaukt normālu mitozes gaitu un izraisīt hromosomu bojājumus, kā arī atsevišķu hromosomu vai visu hromosomu kopu skaita izmaiņas ķermeņa somatiskajās šūnās. Patoloģiskas mitozes var izraisīt vairākas hromosomu slimības. Patoloģiskas mitozes īpaši bieži novēro audzēja šūnās.
Mitozes diagramma:
MITOZE
|
1. Starpfāze To bieži nepareizi sauc par atpūtas stadiju. Starpfāzes ilgums ir atšķirīgs un ir atkarīgs no konkrētās šūnas funkcijas. Šis ir periods, kurā šūna parasti sintezē organellus un palielina izmēru. Nukleoli ir skaidri redzami un aktīvi sintezē ribosomu materiālu. Tieši pirms šūnu dalīšanās tiek replicēta katras hromosomas DNS un histoni. Katru hromosomu tagad attēlo hromatīdu pāris, kas savienoti viens ar otru ar centromēru. Hromosomu viela ir iekrāsota un tiek saukta par hromatīnu, taču pašas šīs struktūras ir grūti saskatāmas. |
|
|
2. Profāze Parasti garākā šūnu dalīšanās fāze. Spiralizācijas un kondensācijas rezultātā hromatīdi saīsinās (līdz 4% no sākotnējā garuma) un sabiezē. Krāsojot, hromatīdi ir skaidri redzami, bet centromēri nav redzami. Dažādos hromatīdu pāros centromērs atrodas atšķirīgi. Dzīvnieku šūnās un zemākajos augos centrioli novirzās uz šūnas pretējiem poliem. No katras centriolas staru veidā izplūst īsi mikrotubuli, kas kopā veido zvaigzni. Nukleoli kļūst mazāki, jo to nukleīnskābe tiek daļēji pārnesta uz noteiktiem hromatīdu pāriem. Profāzes beigās kodola membrāna sadalās un veidojas skaldīšanas vārpsta. |
|
|
3. Metafāze Hromatīdu pāri ar to centromēriem ir pievienoti vārpstas pavedieniem (mikrocaurulēm) un virzās uz augšu un uz leju pa vārpstu, līdz to centromēri sakrīt gar vārpstas ekvatoru perpendikulāri tās asij. |
|
|
4. Anafāze Šis ir ļoti īss posms. Katrs centromērs sadalās divās daļās, un vārpstas pavedieni velk meitas centromērus uz pretējiem poliem. Centromēri velk aiz sevis hromatīdus, kas ir atdalījušies viens no otra, ko tagad sauc par hromosomām. |
|
|
5. Telofāze Hromosomas sasniedz šūnu polus, izstiepjas, izstiepjas, un tās vairs nevar skaidri atšķirt. Vārpstas pavedieni tiek iznīcināti, un centrioli tiek atkārtoti. Ap hromosomām katrā polā veidojas kodola membrāna. Atkal parādās kodols. Telofāzei tūlīt var sekot citokinēze (visas šūnas sadalīšana divās daļās). |
|





Meioze (no grieķu meiosis reducēšana) ir savdabīgs šūnu dalīšanās veids, kā rezultātā hromosomu skaits tajās samazinās uz pusi. Mejoze ir centrālā saite gametoģenēze dzīvniekiem un sporoģenēze augos. Mejoze sastāv no diviem secīgiem dalījumiem, pirms kuriem notiek viena DNS reduplikācija. Visas vielas un enerģija, kas nepieciešama abiem dalījumiem, tiek uzkrāta starpfāzē pirms mejozes es II starpfāze praktiski nav, un sadalījumi ātri seko viens pēc otra. Katrā no meiotiskajām iedalījumiem izšķir tās pašas četras stadijas: profāze, metafāze, anafāze un telofāze, kas raksturīgas mitozei, taču atšķiras pēc vairākām pazīmēm.
Pirmā meiotiskā dalīšanās (mejoze es ) noved pie hromosomu skaita samazināšanās uz pusi, un to sauc par samazināšanos. Rezultātā no vienas diploīdas šūnas(2p 4c) veidojas divas haploīdas šūnas(p 2c) šūnas.
I fāze mejoze ir visilgākā un sarežģītākā. Papildus mitozes profāzei raksturīgajiem DNS spirāles un vārpstas veidošanās procesiem, es Notiek divi ārkārtīgi svarīgi bioloģiski notikumi: konjugācija vai sinapse homologās hromosomas unšķērsojot.
Konjugācija ir homologu hromosomu tuvuma process. Šīs pāra hromosomas veidojas divvērtīgs un tiek saglabāti tā sastāvā ar īpašu proteīnu palīdzību. Tā kā katra hromosoma sastāv no diviem hromatīdiem, divvērtīgā hromatīda ietver četrus hromatīdus un to sauc arī piezīmju grāmatiņa. Producē diploīda šūna P bivalenti. Pēc konjugācijas šūnu formula iegūst formu caurlaide.
Dažās divvērtīgās vietās konjugēto hromosomu hromatīdi krustojas, saplīst un apmainās ar atbilstošām sekcijām. Šo homologo hromosomu fragmentu apmaiņas procesu sauc par šķērsošanu. Tas nodrošina jaunu tēva un mātes gēnu kombināciju veidošanos nākamo gametu hromosomās. Šķērsošana var notikt vairākos apgabalos (vairākkārtēja šķērsošana), nodrošinot augstāku iedzimtās informācijas rekombinācijas pakāpi gametās. Profāzes beigās es palielinās hromosomu spiralizācijas pakāpe, hromatīdi kļūst skaidri atšķirami, vārpstas pavedieni no katra pola pievienojas vienas divvērtīgās hromosomas centromēram. Kodola apvalks tiek iznīcināts, un bivalenti ir vērsti uz šūnas ekvatoriālo plakni.
I metafāzē Mejoze pabeidz vārpstas veidošanos, bivalenti tiek uzstādīti šūnas ekvatoriālajā plaknē. Vārpstas pavedieni no viena pola ir pievienoti katras hromosomas centromēram.
Anafāzē I Mejozē, vārpstas pavedienu ietekmē, homologās hromosomas attālinās viena no otras, virzoties uz šūnas pretējiem poliem. Rezultātā katrā šūnas polā ahaploīds komplektshromosomas, kas satur vienu bihromatīdu hromosomu no katra homologo hromosomu pāra. Anafāzē es dažādu pāru hromosomas, t.i. nehomologās hromosomas uzvedas pilnīgi neatkarīgi viena no otras, nodrošinot dažādu hromosomu veidošanos. kombinācijas tēva un mātes hromosomas nākotnes gametu haploīdajā komplektā. Šādu kombināciju skaits atbilst formulai 2 P, kur p homologo hromosomu pāru skaits. Cilvēkiem šī vērtība ir vienāda ar 2, t.i. 8.4 Cilvēka gametās ir iespējami 10 paternālās un mātes hromosomu kombināciju varianti.
Tātad, homologo hromosomu atšķirības anafāzē es mejoze nodrošina ne tikai hromosomu skaita samazināšanos nākotnes dzimumšūnās, bet arī milzīgu pēdējo daudzveidību dažādu pāru tēva un mātes hromosomu nejaušas kombinācijas dēļ.
I telofāzē Mejoze notiek, veidojoties šūnām, kuru kodolos ir haploīds hromosomu komplekts un divkāršs DNS daudzums, jo katra hromosoma sastāv no diviem hromatīdiem. Šūnām, kas iegūtas pirmās meiotiskās dalīšanas rezultātā, ir formula p2s un pēc īsas starpfāzes viņi sāk nākamo dalījumu.
Otrais meiotiskais dalījums (mejoze II ) norit kā tipiska mitoze (5.3. att.), taču atšķiras ar to, ka tajā nonākušās šūnas satur haploīdu hromosomu kopu. Šīs sadalīšanas rezultātā P bihromatīdu hromosomas (r2c), šķelšanās, veidošanās P vienas hromatīdas hromosomas(ps). Šo iedalījumu sauc vienādojums (vai izlīdzināšana).
Tādējādi pēc diviem secīgiem meiotiskiem dalījumiem no vienas šūnas ar diploīdu divu hromatīdu hromosomu kopu (2x4c) veidojas četras šūnas ar haploīdu vienhromatīdu hromosomu komplektu.(ps).
Meiozes bioloģiskā nozīmesastāv no šūnu veidošanās ar samazinātu hromosomu kopumu un kariotipa noturības saglabāšanu vairākās organismu paaudzēs, kas vairojas. seksuāli transmisīvi. Mejoze kalpo par pamatu kombinētai mainīgumam, nodrošinot gametu ģenētisko daudzveidību, izmantojot tēva un mātes hromosomu šķērsošanas, diverģences un kombinatorikas procesus. Hromosomu struktūras izmaiņas nevienlīdzīgas šķērsošanas dēļ, visu vai atsevišķu hromosomu diverģences pārtraukšana anafāzē I un II meiotiskā dalīšanās izraisa patoloģisku gametu veidošanos un var kalpot par pamatu organisma nāvei vai vairāku hromosomu sindromu attīstībai pēcnācējos.
Diagramma un īsi apraksti par secīgām mejozes stadijām dzīvnieka šūnā.
|
1. Starpfāze Ilgums dažādām sugām ir atšķirīgs. Notiek organellu replikācija un šūnas izmērs palielinās. DNS un histonu replikācija galvenokārt beidzas premeiotiskajā starpfāzē, bet arī daļēji stiepjas līdz profāzes sākumam. Katru hromosomu tagad attēlo hromatīdu pāris, kas savienoti ar centromēru. Hromosomu materiāls ir iekrāsots, bet no visām struktūrām skaidri redzami tikai nukleoli. |
A. Agrīna profāze es |
|
2. Profāze Garākā fāze. To bieži iedala piecos posmos (leptotēma, zigonēma, pahinēma, diplonēma un diakinēze), taču šeit tā tiks uzskatīta par nepārtrauktu hromosomu izmaiņu secību. A . Hromosomas saīsinās un kļūst redzamas kā atsevišķas struktūras. Dažos organismos tie izskatās kā krelles virknes: intensīvi iekrāsota materiāla apgabali - hromomēri - mijas ar nekrāsotām vietām. Hromomēri ir vietas, kur hromosomu materiāls ir ļoti saritināts. |
|
|
B. Homologās hromosomas, kuru izcelsme ir no mātes un tēva gametu kodoliem, tuvojas viena otrai un konjugējas. Šīs hromosomas ir vienāda garuma, to centromēri ieņem vienu un to pašu pozīciju, un tās parasti satur vienādu skaitu gēnu, kas sakārtoti vienā lineārā secībā. Homoloģisko hromosomu hromomēri atrodas blakus. Konjugācijas process var sākties vairākos hromosomu punktos, kas pēc tam tiek savienoti visā garumā (it kā savilkti kopā). Konjugētu homologu hromosomu pārus bieži sauc par bivalentiem. Divvērtīgās vielas saīsina un sabiezē. Šajā gadījumā molekulārā līmenī notiek gan blīvāka iesaiņošana, gan ārēji pamanāma vīšana (spiralizācija). Tagad katra hromosoma ar tās centromēru ir skaidri redzama. |
B. I. fāze |
|
IN. Homoloģiskās hromosomas, kas veido divvērtīgo, ir daļēji atdalītas, it kā atstumjot viena no otras. Tagad jūs varat redzēt, ka katra hromosoma sastāv no diviem hromatīdiem. Hromosomas joprojām ir savienotas viena ar otru vairākos punktos. Šos punktus sauc par chiasmata (no grieķu valodas. chiasma - krusts). Katrā chiasmā pārtraukumu un atkalapvienošanās rezultātā tiek apmainītas hromatīdu sekcijas, kurās ir iesaistīti divi no četriem pavedieniem, kas atrodas katrā chiasmā. Tā rezultātā gēni no vienas hromosomas (piemēram, tēva - A, B, C) tiek saistīti ar gēniem no citas hromosomas (mātes a, b , c), kas rezultātā iegūtajās hromatīdos rada jaunas gēnu kombinācijas. Šo procesu sauc par šķērsošanu. Homologās hromosomas pēc krustošanās neatdalās, jo māsas hromatīdi (abu hromosomu) paliek stingri savienoti līdz anafāzei. |
B. Pārbraukšana profāzes laikā es |
|
G . Homoloģisko hromosomu hromatīdi turpina atgrūst viens otru, un bivalenti iegūst noteiktu konfigurāciju atkarībā no chiasmata skaita. Divvalentiem ar vienu chiasmatu ir krustveida forma, ar diviem chiasmatiem tie ir gredzenveida, un ar trim vai vairāk tie veido cilpas, kas atrodas perpendikulāri viena otrai. Profāzes beigās visas hromosomas ir pilnībā kondensētas un intensīvi iekrāsotas. Šūnā notiek arī citas izmaiņas: centriolu (ja tādas ir) migrācija uz poliem, nukleolu un kodola membrānas iznīcināšana un pēc tam vārpstas pavedienu veidošanās. |
|
|
2. Metafāze Divvērtīgās vielas sarindojas ekvatoriālajā plaknē, veidojot metafāzes plāksni. To centromēri darbojas kā atsevišķas struktūras (lai gan bieži vien šķiet dubultā) un organizē tiem pievienotus vārpstas pavedienus, no kuriem katrs ir vērsts tikai uz vienu no poliem. Šo pavedienu vājā vilkšanas spēka rezultātā katrs bivalents atrodas ekvatora apgabalā, un abi tā centromēri atrodas vienādā attālumā no tā – viens zemāk un otrs augšā. |
D. Vēlīnā metafāze es |
|
3. Anafāze Divi centromēri, kas atrodas katrā bivalentā, vēl nav sadalījušies, bet māsas hromatīdi vairs neatrodas blakus viens otram. Vārpstas pavedieni velk centromērus, no kuriem katrs ir saistīts ar diviem hromatīdiem, pret vārpstas pretējiem poliem. Rezultātā hromosomas tiek sadalītas divos haploīdos komplektos, kas nonāk meitas šūnās. |
E. I anafāze |
|
4. Telofāze Homoloģisko centromēru un saistīto hromatīdu novirze uz pretējiem poliem nozīmē pirmā meiotiskā dalījuma pabeigšanu. Hromosomu skaits vienā komplektā ir kļuvis uz pusi lielāks, bet hromosomas katrā polā sastāv no divām hromatīdām. Sakarā ar krustošanos hiasmātu veidošanās laikā šīs hromatīdas nav ģenētiski identiskas, un otrās meiotiskās dalīšanās laikā tās atdalīsies. Vārpstas un to pavedieni parasti pazūd. Dzīvniekiem un dažiem augiem hromatīdi izplūst, ap tiem katrā polā veidojas kodola membrāna, un iegūtais kodols nonāk starpfāzē. Tad sākas citoplazmas dalīšanās (dzīvniekiem) vai dalošās šūnu sienas veidošanās (augos), kā mitozes gadījumā. Daudzos augos netiek novērota ne telofāze, ne šūnu sieniņu veidošanās, ne starpfāze, un šūna tieši pāriet no anafāzes. I uz II. |
G. Telofāze I dzīvnieka šūnā |







|
II starpfāze Šo posmu parasti novēro tikai dzīvnieku šūnās: tā ilgums atšķiras. S fāze nav, un turpmāka DNS replikācija nenotiek. Otrajā mejozes dalījumā iesaistītie procesi pēc to mehānisma ir līdzīgi tiem, kas notiek mitozē. Tie ietver hromatīdu atdalīšanu abās meitas šūnās, kas izriet no pirmās meiotiskās dalīšanas. Otrais mejozes dalījums no mitozes atšķiras galvenokārt divos veidos: 1) metafāzē II mejoze, māsas hromatīdi bieži ir stipri atdalīti viens no otra; 2) hromosomu skaits ir haploīds. |
|
|
II fāze Šūnās, kas zaudē starpfāzi II , šī posma arī trūkst. Profāzes ilgums II apgriezti proporcionāls telofāzes ilgumam es . Nukleoli un kodola membrānas tiek iznīcinātas, un hromatīdi tiek saīsināti un sabiezināti. Centrioles, ja tādas ir, pārvietojas uz šūnu pretējiem poliem; parādās vārpstas pavedieni. Hromatīdi ir sakārtoti tā, lai to garās asis būtu perpendikulāras pirmās meiotiskās dalīšanas vārpstas asij. |
Z. Profāze P |
|
II metafāze Otrā dalījuma laikā centromēri uzvedas kā dubultās struktūras. Tie organizē vārpstas vītnes, kas vērstas pret abiem poliem, un tādējādi izlīdzina vārpstu pie ekvatora. |
II metafāze |
|
II anafāze Centrs Omērs sadalās, un vārpstas vītnes velk tos uz pretējiem poliem. Centromēri velk pa atdalītajām hromatīdām, kuras tagad sauc par hromosomām. II telofāze Šis posms ir ļoti līdzīgs mitozes telofāzei. Hromosomas izstiepjas, izstiepjas un pēc tam tās ir grūti atšķirt. Vārpstas pavedieni pazūd un centrioli atkārtojas. Ap katru kodolu, kurā tagad ir puse no sākotnējās mātes šūnas (haploīdā) hromosomu skaita, atkal veidojas kodola membrāna. Sekojošas citoplazmas dalīšanās (dzīvniekiem) vai šūnu sienas veidošanās (augos) rezultātā no vienas sākotnējās mātes šūnas tiek iegūtas četras meitas šūnas. |
|


Atšķirības starp mitozes un meiozes stadijām
|
Skatuves |
Mitoze |
Mejoze |
|
Profāze |
Hromomēri nav redzami Homologās hromosomas tiek atdalītas Chiasmata neveidojas Šķērsojumi nenotiek |
Redzams hromomērs Homologās hromosomas ir konjugētas. Veidojas chiasmata. Var notikt krustošanās |
|
Metafāze |
Hromatīdu pāri atrodas pie vārpstas ekvatora Centromēri atrodas vienā plaknē pie vārpstas ekvatora |
Hromatīdu pāri atrodas pie vārpstas ekvatora tikai otrajā mejozes nodaļā Centromēri pirmajā mejozes nodaļā atrodas virs un zem ekvatora vienādos attālumos no tā |
|
Anafāze |
Centromēri sadalās. Hromatīdi atdalās. Atšķirīgās hromatīdas ir identiskas |
Centromēri sadalās tikai meiozes otrajā stadijā. Hromatīdi atdalās meiozes otrās dalīšanās laikā. Pirmajā dalījumā atdalās veselas hromosomas. Atšķirīgās hromosomas var nebūt identiskas krustošanās rezultātā |
|
Telolofāze |
Hromosomu skaits meitas šūnās ir tāds pats kā vecāku šūnās Meitas šūnas satur abas homologās hromosomas (diploīdos) |
Hromosomu skaits meitas šūnās ir uz pusi mazāks nekā vecāku šūnās Meitas šūnas satur tikai vienu no katra homologo hromosomu pāra |
|
Kur notiek šāda veida sadalīšana? |
Iespējama haploīdajās, diploīdās un poliploīdās šūnās Rodas somatisko šūnu un dažu sporu veidošanās laikā, kā arī gametu veidošanās laikā augos, kuros notiek paaudžu maiņa |
Tikai diploīdās un poliploīdās šūnās Gameto- vai sporoģenēzes laikā |
- Gametu struktūra. Gametoģenēze
Seksuālā reprodukcija tiek veikta, izmantojot specializētas dzimumšūnas, ko sauc par gametām. Sieviešu dzimumšūnas sauc olas, tēviņš spermatozoīdi.Gametes no somatiskajām šūnām atšķiras galvenokārt ar to, ka tām ir uz pusi mazāks hromosomu skaits, kā arī zems vielmaiņas procesu līmenis.
Olšūnas salīdzinoši lielas nekustīgas šūnas, parasti apaļas formas; Papildus tipiskām organellām citoplazma satur rezerves barības vielu ieslēgumus dzeltenuma formā ( rīsi .). Olu kodolos veidojas daudzas ribosomu gēnu un mRNS kopijas, nodrošinot topošā embrija dzīvībai svarīgo proteīnu sintēzi. Dažādu organismu olas atšķiras pēc dzeltenuma sadalījuma daudzuma un rakstura tajās. Ir vairāki olu veidi.

Izolecitālssauc par salīdzinoši mazām olām ar nelielu daudzumu vienmērīgi sadalīta dzeltenuma. Kodols tajos atrodas tuvāk centram. Šādas olas ir sastopamas tārpiem, gliemežvākiem un gliemežnīcām, adatādaiņiem un lancetēm.Vidēji telolecitālsStoru un abinieku ikru diametrs ir aptuveni 1,5 × 2 mm, un tie satur vidēji daudz dzeltenuma, kura lielākā daļa ir koncentrēta vienā no poliem (veģetatīvā). Pie pretējā pola (dzīvnieka), kur ir maz dzeltenuma, atrodas olas kodols.
Spēcīgi telolecitālsDažu zivju, rāpuļu, putnu un olnīcu zīdītāju olas satur daudz dzeltenuma, kas aizņem gandrīz visu olas citoplazmas tilpumu. Dzīvnieka polā atrodas dīgļu disks ar aktīvu citoplazmu bez dzeltenuma. Šo olu izmēri ir lieli, 10 15 mm vai vairāk.
Alecital olām praktiski nav dzeltenuma, tās ir mikroskopiski mazas (0,1 × 0,3 mm) un ir raksturīgas placentas zīdītājiem, tostarp cilvēkiem.
Sperma vai spermatozoīdi ir ļoti mazas kustīgas vīriešu dzimuma gametas (piemēram, cilvēka spermas garums ir 50 × 70 µm, bet krokodils 20 µm), ko veido vīriešu dzimumdziedzeri - sēklinieki; to skaits mērāms miljonos. Spermas forma dažādiem dzīvniekiem ir atšķirīga, taču to struktūra ir vienāda, lielākajai daļai no tiem ir galva un kakls. Spermas galvā ir kodols, kas satur haploīdu hromosomu skaitu un ir pārklāts akrosoma. Akrosoma ir īpaša struktūra, modificēts Golgi komplekss, kas satur enzīmus olšūnas membrānas izšķīdināšanai apaugļošanas laikā, ko ierobežo membrāna. Kakls satur daudzus mitohondrijus un divus centriolus. No kakla izaug mikrotubulu veidota aste, kas nodrošina spermatozoīdu kustīgumu. Vidējā daļa ir paplašināta, pateicoties tajā esošajiem daudzajiem mitohondrijiem, kas ir salikti spirālē ap karogiem. Šie mitohondriji nodrošina enerģiju saraušanās mehānismiem, kas nodrošina karogdziedzera kustību. Spermas galvenajai un astes daļai ir flagellai raksturīga struktūra.

Ja paskatās uz cilvēka spermas galvu no augšas, tā šķiet apaļa, un, skatoties no sāniem, tā šķiet saplacināta. Ar karoga kustību vien nepietiek, lai spermatozoīdi pārvietotos attālumā no maksts līdz vietai, kur notiek apaugļošanās. Spermas galvenais lokomotoriskais uzdevums ir spietot ap olšūnu un noteiktā veidā orientēties pirms iekļūšanas oocīta membrānās.
Gametu veidošanās (gametoģenēze).Vīriešu dzimuma epitēlija šūnas Un Sieviešu dzimumdziedzeros notiek virkne secīgu mitotisku un meiotisku dalījumu, ko kopā sauc par gametoģenēzi, kā rezultātā veidojas nobriedušas vīriešu dzimuma gametas (spermatoģenēze) un sieviešu dzimuma gametas (ooģenēze). Abos gadījumos process ir sadalīts trīs fāzēs – reprodukcijas fāzē, augšanas fāzē un nobriešanas fāzē. Reproduktīvā fāze ietver atkārtotus mitotiskus dalījumus, kas izraisa daudzu spermatogoniju vai oogoniju veidošanos. Katrs no tiem iziet augšanas periodu, gatavojoties pirmajam meiotiskajam dalījumam un tai sekojošai citokinēzei. Tad sākas nogatavināšanas fāze, kuras laikā notiek pirmā un otrā meiotiskā dalīšanās Ar sekojoša haploīdu šūnu diferenciācija Un nobriedušu gametu veidošanās.
Spermas attīstība (spermatoģenēze).Spermatozoīdi (spermīns) veidojas vairāku secīgu šūnu dalīšanās rezultātā, ko kopā saucspermatoģenēze,kam seko sarežģīts diferenciācijas process, ko sauc spermioģenēze (20.31. att ). Spermas veidošanās process ilgst aptuveni 70 dienas; uz 1 g sēklinieku svara veidojas 10 7 spermatozoīdu dienā. Sēklu kanāliņu epitēlijs sastāv no ārējā šūnu slāņagerminālais epitēlijsun aptuveni seši šūnu slāņi, kas izveidojušies šī slāņa atkārtotas šūnu dalīšanās rezultātā (20.32. un 20.33. att.); šie slāņi atbilst secīgām spermas attīstības stadijām. Pirmkārt, dīgļu epitēlija šūnu dalīšanās rada daudzas spermatogoniju kas palielinās izmērā un kļūstpirmās kārtas spermatocīti.Pirmā meiotiskā dalīšanās rezultātā šie spermatocīti veido haploīdusotrās kārtas spermatocīti,pēc tam tie iziet otro meiotisko dalīšanos un pārvēršas par spermatīdiem. Starp jaunattīstības šūnu "virknēm" ir lielas Sertoli šūnas, vai trofiskās šūnas,atrodas visā telpā no kanāliņu ārējā slāņa līdz tā lūmenim.

Spermatocīti atrodas daudzās invaginācijās uz Sertoli šūnu sānu virsmām; šeit tie pārvēršas par spermatīdiem un pēc tam pārvietojas uz to Sertoli šūnas malu, kas ir vērsta pret sēklu kanāliņu lūmenu, kur tie nobriest, veidojot spermīns (20.33. att ). Acīmredzot Sertoli šūnas nodrošina mehānisku atbalstu, aizsardzību un barošanu nobriedušai spermai. Visas barības vielas un skābeklis, kas tiek piegādāti jaunattīstības gametām caur asinsvadiem, kas ieskauj sēklu kanāliņus, un vielmaiņas atkritumi, kas nonāk asinīs, iziet cauri Sertoli šūnām. Šīs šūnas arī izdala šķidrumu, ar kuru spermatozoīdi iziet cauri kanāliņiem.
Olu attīstība cilvēkiem (ooģenēze).Atšķirībā no spermatozoīdu veidošanās, kas vīriešiem sākas tikai pubertātes vecumā, olšūnu veidošanās sievietēm sākas pat pirms dzimšanas un katrai dotajai olšūnai beidzas tikai pēc tās apaugļošanas. Ooģenēzes posmi ir parādīti attēlā. 20.36. Augļa attīstības laikā pirmatnējās dzimumšūnas atkārtoti dalās mitozes ceļā, veidojot daudzas lielas šūnas, t.s. oogonija. Oogonia atkal tiek pakļauta mitozei un veidojaspirmās kārtas oocygs,kas paliek profāzes stadijā gandrīz līdz ovulācijai. Pirmās kārtas oocītus ieskauj viens šūnu slānis-granulozes membrāna-un veido tā sauktopirmatnējie folikuli.Sievietes auglis tieši pirms dzimšanas satur apmēram 2 10 6 šie folikuli, bet tikai aptuveni 450 no tiem sasniedz otrās kārtas oocītu stadiju un atstāj olnīcu (ovulāciju). Pirms ovulācijas pirmās kārtas oocīts iziet pirmo meiotisko dalīšanos, veidojot haploīduotrās kārtas oocīts Un pirmais polārais ķermenis.Otrais meiotiskais dalījums sasniedz metafāzes stadiju, bet neturpinās, kamēr oocīts nesaplūst ar spermu. Apaugļošanas laikā otrās kārtas oocīts iziet otro meiotisko dalīšanos, veidojot lielu šūnu - olu un arī otrais polārais ķermenis.Visi polārie ķermeņi ir mazas šūnas; tiem nav nekādas nozīmes ooģenēzē un galu galā tiek iznīcināti.

partenoģenēze
obligāts
neobligāti
Organismu attīstība tikai no neapaugļotas olšūnas.
Veidojas tikai mātītes.
Piemēram, kaukāziešu klinšu ķirzaka
letlets var attīstīties gan bez apaugļošanas, gan ar apaugļošanu
Sieviešu tips
Tēviņi attīstās no neapaugļotām olām
Vīriešu tips
Dažās izogāmās aļģēs
Citi līdzīgi darbi, kas jūs varētu interesēt.vshm> |
|||
| 6644. | Organismu individuālā attīstība (ontoģenēze) | 78,69 KB | |
| Tiek uzskatīts, ka tikai viena spermatozoīda kodola saturs iekļūst dzīvnieku olšūnā. Cilvēku un dažreiz augstāku dzīvnieku gadījumā attīstības periodu pirms dzimšanas bieži sauc par pirmsdzemdību pēc dzimšanas un pēcdzemdību. Lielākajā daļā daudzšūnu dzīvnieku, neatkarīgi no to organizācijas sarežģītības, embrija attīstības posmi, kuriem embrijs iziet, ir vienādi. Sasmalcināšanas raksturs un blastuļu veidi dažādiem mugurkaulniekiem... | |||
| 13714. | Organismu individuālā attīstība un to uzvedība. Ontoģenēze. Dzīves cikls augos un dzīvniekos | 9,96 KB | |
| Ontoģenēzes personības izcelsme ir organisma individuālā attīstība no zigotas veidošanās brīža līdz tā nāvei. Gametes ir dzimumšūnas, kurām ir iedzimta informācija un kurām ir haploīds komplekts. Apaugļošana ir augu vai dzīvnieku vīrišķo un sieviešu reproduktīvo šūnu saplūšanas process, un tas ir seksuālā procesa pamats. Embrionālais dīgtspējas periods ir ontoģenēzes periods no zigotas veidošanās līdz piedzimšanai vai iznākšanai no olšūnas membrānām vai dīgtspējai. | |||
| 10427. | Pavairošana | 6,75 KB | |
| Tā kā organismu vairošanās formas ir ļoti dažādas, tos visus var reducēt līdz diviem galvenajiem veidiem: aseksuāliem un seksuāliem. Bezdzimuma reprodukcijā pēcnācēju vairošanās notiek no viena vecāka, veidojot sporas vai veģetatīvi. Veģetatīvās pavairošanas laikā pēcnācēji rodas no ķermeņa daļām, kas atdalītas no mātes. Veģetatīvās pavairošanas laikā augi saglabā heterozigotitāti daudzu paaudžu garumā. | |||
| 21332. | Ierobežojošie faktori. Organismu pielāgošanās faktoriem | 303,8 KB | |
| Daži dzīvnieki mīl intensīvu karstumu, citi labāk panes mērenu vides temperatūru utt. Turklāt dzīvie organismi tiek iedalīti tajos, kas spēj pastāvēt plašā vai šaurā jebkura vides faktora izmaiņu diapazonā. Ja vides apstākļu ietekme nesasniedz galējās vērtības, dzīvie organismi uz to reaģē ar noteiktām darbībām vai izmaiņām savā stāvoklī, kas galu galā noved pie sugas izdzīvošanas. Pētījuma objekts šajā darbā ir vides faktori, priekšmets ierobežojošie faktori un organismu pielāgošanās... | |||
| 8875. | Vispārēji organismu un vides faktoru mijiedarbības modeļi | 193,58 KB | |
| Augsņu sauszemes un ūdens vides biotiskie faktori Dzīvo organismu bioloģiski aktīvās vielas Antropogēnie faktori Vispārējie organismu un vides faktoru mijiedarbības modeļi Ierobežojošā faktora jēdziens. Lībiga minimālais likums Šelforda likums Antropogēno faktoru ietekmes uz organismu specifika Organismu klasifikācija saistībā ar vides faktoriem 1. Spalvzāles stepju apstākļi reprezentē pilnīgi atšķirīgus abiotisko faktoru režīmus. | |||
| 12700. | Kaitēkļu bioloģiskās īpašības un to apkarošanas pasākumi | 62,79 KB | |
| Īpaši nozīmīgi ražas zudumi rodas nezāļu klātbūtnes rezultātā, kas izvada no augsnes barības vielas un mitrumu, ēno kultivētos augus, kā arī daudzos gadījumos piesārņo produktus ar toksiskām vielām un sēklām, kas izraisa cilvēku un dzīvnieku saindēšanos. Galvenie lauksaimniecības ķīmiskās apstrādes virzieni: mēslošanas līdzekļu, ķīmisko vielu izmantošana, augu aizsardzība pret kaitēkļiem, slimībām un nezālēm, ķīmisko produktu izmantošana lopkopībā, lauksaimniecības produktu konservēšana un... | |||
| 13403. | Vīrusi kā ne-šūnu dzīvības formas. Vīrusa uzbūve, klasifikācija, mijiedarbība ar dažādu organismu šūnām | 12,75 KB | |
| Tie ir bioloģiski objekti, genomi, kas sastāv no nukleīnskābēm DNS vai RNS, kas ražoti dzīvās šūnās, izmantojot to biosintētisko aparātu. Atšķirības starp vīrusiem un citām dzīvības formām: tiem nav šūnu struktūras; 1 veida nukleīnskābes; tikai DNS vai RNS; tiem nav sava metabolisma. Hipotēze par vīrusu izcelsmi: vīrusi radās no normālas šūnas komponentiem, kas izbēga no regulējošo mehānismu kontroles un pārvērtās par neatkarīgu vienību; iespējams, DNS daļā notika virkne ģenētisku izmaiņu, kas... | |||
| 18798. | Augsnes mezofaunas organismu strukturālie un funkcionālie rādītāji Pozim upes ielejas mežos un atklātajos biotopos | 61,54 KB | |
| Tomēr šodien šīs teritorijas attīstība ir vairāku vides problēmu sekas. Jo īpaši ielejā tiek veikti plaši darbi, lai nosusinātu palieņu pļavas, kas tieši ietekmē dabiskos biotopus. | |||
| 19386. | Liberālisma attīstība NVS valstīs | 35,55 KB | |
| Ārējo ekonomisko attiecību evolūcijas procesos izskatāmo izmaiņu kompleksais sistēmiskais raksturs nosaka visas tradicionālo priekšstatu par tautsaimniecības identifikāciju, robežām un suverenitāti aksiomātikas pārskatīšanu un maiņu... | |||
| 8867. | IZGLĪTĪBA UN ATTĪSTĪBA | 162,32 KB | |
| Treniņu un attīstības attiecību problēmas būtība. Pamata pieejas apmācības un attīstības attiecību problēmas risināšanai. Proksimālās attīstības zonas jēdziens L. Treniņu un attīstības attiecību problēmas būtība. | |||
Vairošanās ir organismu īpašība radīt pēcnācējus vai organismu spēja vairoties pašiem. Būdama svarīgākā dzīvo būtņu īpašība, vairošanās nodrošina dzīvības nepārtrauktību un sugu turpināšanos.
Reprodukcijas process ir ārkārtīgi sarežģīts un saistīts ne tikai ar ģenētiskās informācijas nodošanu no vecākiem uz pēcnācējiem, bet arī ar organismu anatomiskajām un fizioloģiskajām īpašībām, to uzvedību un hormonālo kontroli. Organismu vairošanos pavada to augšanas un attīstības procesi.
Dzīvām radībām ir raksturīga ārkārtīgi daudzveidīga reprodukcijas metodes. Tomēr pastāv divas galvenās vairošanās metodes - aseksuālā un seksuālā (16. att.). Aseksuāla vairošanās jeb apomiksis (no grieķu val. aro - bez, mixis - sajaukšana) ir process, kurā piedalās tikai viens vecāks (šūna vai daudzšūnu organisms). Turpretim seksuālā pavairošana ietver divus vecākus, kuriem katram ir sava reproduktīvā sistēma un kuri ražo dzimumšūnas (gametas), kas pēc saplūšanas veido zigotu (apaugļotu olu), kas pēc tam diferencējas par embriju. Līdz ar to seksuālās vairošanās laikā notiek iedzimtu faktoru sajaukums, t.i., process, ko sauc par amfimiksu (no grieķu val. amfi - abās pusēs, mixis - sajaukšana).
Aseksuālā pavairošana
Aseksuāla vairošanās ir raksturīga daudzu sugu organismiem, gan augiem, gan dzīvniekiem. Tas ir atrodams vīrusos, baktērijās, aļģēs, sēnēs, vaskulārajos augos, vienšūņos, sūkļos, koelenterātos, bryozoānos un tunikātos.
Vienkāršākā aseksuālās vairošanās forma ir raksturīga vīrusiem. Viņu reproduktīvais process ir saistīts ar nukleīnskābju molekulām, ar šo molekulu spēju pašam dublēt
un tā pamatā ir relatīvi vāju ūdeņraža saišu specifika starp nukleotīdiem.
Rīsi. 16.Organismu pavairošanas metodes
Saistībā ar citiem organismiem, kas vairojas aseksuāli, izšķir veģetatīvo vairošanos un vairošanos ar sporulāciju.
Veģetatīvā pavairošana ir vairošanās, kurā no mātes organisma atdalītas daļas attīstās jauns organisms. Šis vairošanās veids ir raksturīgs gan vienšūnu, gan daudzšūnu organismiem, taču tiem ir dažādas izpausmes.
Vienšūnu organismos veģetatīvo vairošanos pārstāv tādas formas kā dalīšanās, daudzkārtēja dalīšanās un pumpuru veidošanās. Sadalīšanās ar vienkāršu sašaurināšanos ar divu meitas organismu veidošanos no viena vecāka organisma ir raksturīga baktērijām un zilaļģēm (cianobaktērijām). Gluži pretēji, brūno un zaļo aļģu, kā arī vienšūnu dzīvnieku (sarkodu, flagellatu un ciliātu) vairošanās notiek kodola mitotiski daloties, kam seko citoplazmas sašaurināšanās.
Reproducēšana ar daudzkārtēju skaldīšanu (šizogonija) ietver kodola sadalīšanu, kam seko citoplazmas sadalīšana daļās. Šīs dalīšanas rezultātā no vienas šūnas veidojas vairāki meitas organismi. Vairākkārtējas dalīšanas piemērs ir falciparum plasmodium reprodukcija (P. vivax) cilvēka eritrocītos. Šajā gadījumā plazmodijā atkārtota kodola dalīšanās notiek daudzas reizes bez citokinēzes, kam seko citokinēze. Rezultātā viens plazmodijs rada 12-24 meitas organismus.
Daudzšūnu augu organismos veģetatīvo pavairošanu dalot veic ar spraudeņiem, sīpoliem, lapām un sakneņiem. Bet tā būtībā ir mākslīga pavairošana, ko izmanto lauksaimniecības praksē. Augstāku augu pavairošana mākslīgos apstākļos iespējama arī no vienas šūnas. Organismiem, kas attīstās no vienas šūnas (klona), piemīt visas sākotnējā daudzšūnu organisma īpašības. Šo pavairošanu sauc par klonālo mikropavairošanu. Viens no veģetatīvās pavairošanas veidiem var būt daudzu kultivēto augu potēšana vai pārstādīšana, kas sastāv no pumpura vai dzinuma daļas pārstādīšanas no viena auga uz otru. Protams, šī ir arī pavairošanas metode, kas dabā nenotiek, bet tiek ļoti plaši izmantota lauksaimniecībā.
Daudzšūnu dzīvniekiem veģetatīvā pavairošana notiek, sadalot viņu ķermeni daļās, pēc kurām katra
daļa attīstās par jaunu dzīvnieku. Šāda vairošanās ir raksturīga sūkļiem, koelenterātiem (hidrām), nemerteāniem, plakanajiem tārpiem, adatādaiņiem (zvaigznēm) un dažiem citiem organismiem. Dzīvnieku veģetatīvās reprodukcijas sadrumstalotības veids ir dzīvnieku poliembrionija, kas sastāv no tā, ka noteiktā attīstības stadijā embrijs tiek sadalīts vairākās daļās, no kurām katra attīstās par neatkarīgu organismu. Poliembrionija rodas bruņnešiem. Tomēr pēdējie vairojas seksuāli. Tāpēc poliembrionija drīzāk ir unikāls seksuālās reprodukcijas posms, un poliembrionijas rezultātā iegūtos pēcnācējus pārstāv monozigotiski dvīņi.
Jaunveidošanās sastāv no tuberkulozes (izauguma) veidošanās ar kodolu uz mātes šūnas, kas pēc tam atdalās un kļūst par neatkarīgu organismu. Dīgšana notiek gan vienšūnu augiem, piemēram, raugam, gan vienšūnu dzīvniekiem, piemēram, noteiktām skropstu sugām.
Vairošanās ar sporulāciju ir saistīta ar specializētu šūnu veidošanos - sporas, kas satur kodolu, citoplazmu, ir pārklātas ar blīvu membrānu un spēj ilgstoši pastāvēt nelabvēlīgos apstākļos, kas turklāt veicina to izplatīšanos. Visbiežāk šāda vairošanās notiek baktērijās, aļģēs, sēnēs, sūnās un papardēs. Dažās zaļajās aļģēs no atsevišķām šūnām var veidoties tā sauktās zoosporas.
Dzīvnieku vidū vairošanos sporulācijas ceļā novēro sporozoāniem, jo īpaši falciparum plasmodium.
Daudzu sugu organismos aseksuālā vairošanās var mijas ar dzimumvairošanos.
SEKSUĀLĀ REPRODUKCIJA
Seksuāla vairošanās notiek gan vienšūnu, gan daudzšūnu augos un dzīvniekos.
Kā minēts V un XIII nodaļā, seksuālā pavairošana baktērijās tiek veikta ar konjugāciju, kas kalpo kā seksuālā procesa analogs un ir šo organismu rekombinācijas sistēma, savukārt vienšūņiem seksuālā pavairošana notiek arī konjugācijas vai singāmijas ceļā. un autogāmija.
Daudzšūnu organismos (augos un dzīvniekos) seksuālā vairošanās ir saistīta ar dīgļu vai dzimumšūnu (gametu) veidošanos, apaugļošanos un zigotu veidošanos.
Seksuālā vairošanās ir nozīmīga organismu evolucionāra iegūšana. No otras puses, tas veicina gēnu pārkārtošanos, organismu daudzveidības rašanos un to konkurētspējas paaugstināšanu nepārtraukti mainīgos vides apstākļos.
Vienšūnu organismos seksuālā vairošanās notiek vairākos veidos. Baktērijās seksuālo reprodukciju var analoģizēt ar tajās notiekošo konjugāciju, kas sastāv no plazmīdas vai hromosomu DNS pārnešanas no donora šūnām (satur plazmīdas) uz recipienta šūnām (kas nesatur plazmīdas), kā arī ar baktēriju transdukciju. , kas sastāv no ģenētiskā materiāla pārnešanas no dažām baktēriju šūnām uz citiem fāgiem. Konjugācija ir sastopama arī ciliātos, kur šī procesa laikā notiek kodolu pārnešana no viena indivīda uz otru, kam seko pēdējā sadalīšana.
Daudzšūnu augos un dzīvniekos šūnu seksuālā pavairošana notiek, veidojot sieviešu un vīriešu dzimumšūnas (olas un spermu), pēc tam apaugļojot olšūnu ar spermu un veidojot zigotu. Augos dzimumšūnas tiek ražotas specializētās reproduktīvās struktūrās; dzīvniekiem tās veidojas dzimumdziedzeros, ko sauc par dzimumdziedzeriem (no grieķu valodas. aizgājis - sēklas).
Pastāv būtiska atšķirība starp dzīvnieku somatiskajām un reproduktīvajām šūnām. Tas slēpjas faktā, ka somatiskās šūnas spēj dalīties, tas ir, tās vairojas pašas, un turklāt no tām veidojas dzimumšūnas. Gluži pretēji, dzimumšūnas nedalās, bet “uzsāk” visa organisma vairošanos.
Diploīdas somatiskās šūnas, kurās veidojas vīriešu dzimumšūnas, sauc par spermatogoniju, bet kurās veidojas sieviešu dzimumšūnas - par oogoniju. Vīriešu un sieviešu dzimumšūnu veidošanās (augšanas un diferenciācijas) procesu sauc par gametoģenēzi.
Gametoģenēzes pamatā ir mejoze (no grieķu valodas. mejoze - samazināt), kas ir šūnu kodolu sadalīšanās, ko papildina hromosomu skaita samazināšanās vienā kodolā. Mejoze rodas specializētās dzīvu radību reproduktīvo orgānu šūnās, kas vairojas seksuāli (17. att.). Piemēram, pteridofītos mejoze notiek specializētās sporangu šūnās, kas atrodas uz šo augu lapu apakšējās virsmas un attīstās sporās un pēc tam gametofītos. Pēdējās pastāv atsevišķi, galu galā ražojot vīriešu un sieviešu dzimumšūnas. Ziedošajos augos mejoze notiek specializētās olšūnu šūnās, kas attīstās sporās. Pēdējie ražo gametofītu ar vienu olu. Turklāt šajos augos mejoze notiek arī specializētās putekšņlapu šūnās, kuras arī attīstās sporās, kas galu galā ražo ziedputekšņus ar divām vīriešu dzimuma gametām. Sliekām, kas ir hermafrodīti un satur vīriešu reproduktīvos orgānus vienā ķermeņa segmentā un sieviešu dzimumorgānus citā un kuriem ir raksturīgas
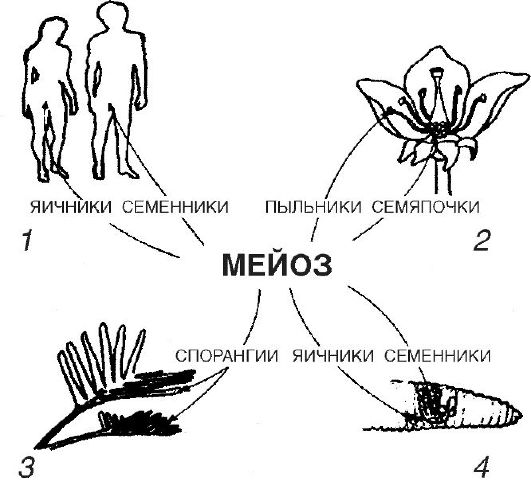
Rīsi. 17. Mejoze dažādos organismos: 1 - Cilvēks; 2 - ziedoši augi; 3 - paparde; 4 - slieka
Viņiem ir iespēja savstarpēji apaugļot dažādus indivīdus; viņiem ir iespēja vienlaikus veikt spermatoģenēzi un oģenēzi.
Mejoze notiek specializētās sēklinieku un olnīcu šūnās, kas ražo attiecīgi vīriešu un sieviešu dzimumšūnas. Ir identificēti proteīni, kas ir mejozes induktori.
Mejozes procesā somatiskajām šūnām (šūnu kodoliem) un nenobriedušām šūnu dīgļiem raksturīgais diploīds hromosomu skaits (2n) mainās uz nobriedušām dzimumšūnām raksturīgo haploīdu skaitu (n). Tātad gametoģenēzes rezultātā dzimumšūnas saņem tikai pusi no somatisko šūnu hromosomām (18. att.).

Rīsi. 18.
Hromosomu uzvedība gametoģenēzes laikā dzīvniekiem ir vienāda gan vīriešiem, gan mātītēm. Tomēr dzimumi atšķiras dažādu mejozes posmu izcelsmes laikā, kas ir īpaši
īpaši pamanāms cilvēkiem. Pēcpubertātes vīriešiem pilns mejozes process tiek pabeigts aptuveni divu mēnešu laikā, savukārt mātītēm pirmais mejozes dalīšanās sākas augļa olnīcā un netiek pabeigts līdz ovulācijas sākumam, kas notiek aptuveni 15 gadu vecumā.
Augstākiem dzīvniekiem tēviņu gadījumā mejozi pavada četru funkcionāli aktīvu gametu veidošanās.
Gluži pretēji, mātītēm katrs otrās kārtas olšūns ražo tikai vienu olu. Pārējie sieviešu meiozes kodolprodukti ir trīs reducējoši ķermeņi, kas nepiedalās vairošanā un deģenerējas.
Mejoze sastāv no divām kodolu daļām. Pirmais meiotiskais kodola dalījums atdala katra homologo hromosomu pāra locekļus pēc tam, kad tie ir savienoti pārī (sinapse) un apmainījušies ar ģenētisko materiālu (šķērsojot). Šīs dalīšanas rezultātā veidojas divi haploīdi kodoli. Otrais meiotiskais dalījums atdala abas hromosomu (hromatīdu) gareniskās puses katrā no šiem kodoliem, veidojot četrus haploīdus kodolus.
Gametoģenēzes procesā notiek arī olšūnu (ovogenēze) un spermatozoīdu (spermatoģenēze) diferenciācija, kas ir priekšnoteikums to funkcijām. Dzīvnieku olas ir daudz lielākas par spermu, parasti ir nekustīgas un satur barojošu materiālu, kas nodrošina embrija attīstību sākotnējā periodā pēc apaugļošanas. Lielākajai daļai dzīvnieku spermai ir karogs, kas nodrošina to kustību neatkarību.
Mejozei ir izcila bioloģiskā nozīme. Pateicoties mejozei, organismu šūnās tiek uzturēts nemainīgs hromosomu skaits neatkarīgi no paaudžu skaita. Tāpēc mejoze saglabā sugu noturību. Visbeidzot, meiozē krustošanās rezultātā notiek gēnu rekombinācija, kas ir viens no evolūcijas faktoriem.
SPERMATOĢĒZE UN OVOĢĒZE
Spermatoģenēze ir nobriedušu vīriešu dzimumšūnu veidošanās process. Spermatozoīdi attīstās vīriešu dzimumdziedzeros (sēkliniekos vai sēkliniekos) no specializētām somatiskajām šūnām (19. att.). Šādas specializētas šūnas darbojas šādi:
ko sauc par pirmatnējām dzimumšūnām, kuras migrē uz sēkliniekiem vīrieša indivīda agrīnajā embrioģenēzes periodā. Līdz ar to pirmatnējās šūnas ir nobriedušu dzimumšūnu priekšteči (prekursori).
Cilvēka sēklinieki sastāv no daudziem kanāliņiem, kuru sienas veido šūnu slāņi, kas atrodas dažādās spermas attīstības stadijās. Kanāliņu ārējo slāni veido lielas šūnas, ko sauc par spermatogoniju. Šīs šūnas satur diploīdu hromosomu komplektu un ir sēklinieku pirmatnējo dzimumšūnu pēcteči. Indivīda pubertātes periodā daļa spermatogoniju pārvietojas uz kanāliņu iekšējo slāni, kur meiozes rezultātā tie attīstās šūnās, ko sauc par pirmās kārtas spermatocītiem (spermatocītiem I), pēc tam par otrās kārtas spermatocītiem. (spermatocīti II) un, visbeidzot,

Rīsi. 19.Spermatoģenēze un ooģenēze
spermatīdos, kas ir haploīdas dzimumšūnas, kas galu galā diferencējas par nobriedušu spermu. Tādējādi vispārīgi var teikt, ka spermatoģenēze tiek uzsākta diploīdās somatiskās šūnās (spermatogonijā), kam seko dzimumšūnu nobriešanas periods, kurā mejozes ceļā notiek divas kodola dalīšanās, izraisot spermatīdu veidošanos. Šis ir cilvēka attēls.
Mejoze (20. att.) spermatoģenēzē notiek vairākos posmos (fāzēs). Starp nodaļām ir divas starpfāzes. Tādējādi meiotisko dalījumu var attēlot kā notikumu virkni, kas seko viens pēc otra, proti: I starpfāze - pirmais meiotiskais dalījums (I agrīnā profāze, I vēlā profāze, I metafāze, I anafāze, I telofāze) - II starpfāze (interokinēze) - otrais meiotiskais dalījums (I fāze, II metafāze, II anafāze, II telofāze). Mejozes process ir ļoti dinamisks, tāpēc mikroskopiskās atšķirības starp dažādiem posmiem atspoguļo nevis pašu stadiju raksturu, bet gan hromosomu īpašības dažādos posmos. I starpfāze

Rīsi. 20.Meiozes fāzes: 1 - pirmais meiotiskais dalījums; 2 - otrais meiotiskais dalījums
ko raksturo fakts, ka tajā notiek hromosomu replikācija (DNS dubultošanās), kas gandrīz pilnībā ir pabeigta agrīnās I fāzes sākumā.
Pirmais meiotiskais dalījums sākas primārajā spermatocītā, un to raksturo ilga profāze, kas sastāv no I un II profāzes, kas pāriet viena otrā. I fāzē ir pieci galvenie posmi - leptonēma, zigonēma, pahinēma, diplonēma un diakinēze.
Leptonēmas stadijā hromosomas kodolā ir attēlotas plānu spirālveida pavedienu veidā, kas satur daudzas tumšas krāsas granulas (hromomērus). Hromomēru un pavedienu šķelšanās netiek atzīmēta, taču tiek uzskatīts, ka hromosomas šajā posmā ir dubultas, t.i., diploīdas. Katra hromosomu pāra homologus visā garumā vieno hromomēri pēc aizdares principa.
Zigonēmas stadiju raksturo sinapses izveidošanās starp homologām hromosomām, kā rezultātā veidojas sapārotas hromosomas (divvērtīgās). Hromosomas X un Y uzvedas nedaudz savādāk nekā autosomas. Tie kondensējas tumšas krāsas heterohromatiskajos ķermeņos, kas savienoti pārī homologu reģionu rezultātā to galos.
Pachinēmas stadijā, kas ir garākā meiozes profāzes stadija, notiek bivalentu kondensācija un katra hromatīda sadalīšanās divās daļās, kā rezultātā katrs divvērtīgais ir sarežģīta spirālveida struktūra, kas sastāv no četrām māsu hromatīdām (tetradēm). Šī posma beigās sākas pārī savienoto divvērtīgo hromosomu atdalīšana. Tagad homologās hromosomas var novērot blakus. Tāpēc atsevišķos preparātos var redzēt četras hromosomas, kas veidojas katra homologa dublēšanās rezultātā, veidojot māsas hromatīdas. Šajā posmā notiek apmaiņa starp homologiem un chiasmata veidošanos.
Diplonēmas stadijā notiek māsu hromatīdu saīsināšana, sabiezēšana un savstarpēja atgrūšanās, kā rezultātā hromatīdi divvērtīgajā gandrīz atdalās. Atdalīšana tiek uzskatīta par nepilnīgu, jo centromērs vēl nav sadalīts katrā hromosomu pārī. Kas attiecas uz bivalentiem, tos dažādās vietās visā garumā notur chiasmata, kas ir struktūras, kas veidojas starp homologiem hromatīdiem, kā rezultātā.
iepriekšējā krustošanās starp sinaptiski savienotiem homologiem. Labos preparātos atkarībā no divvērtīgā garuma var novērot vienu līdz vairākas chiasmatas. Katra šajā posmā novērotā chiasma ir apmaiņas rezultāts, kas notika starp hromatīdiem, kas nav māsas, pachinēmas stadijā. Pastiprinoties bivalentu saspiešanai un atgrūšanai, chiasmata virzās uz hromosomu galiem, t.i., notiek hromosomu terminālizācija. Diplonēmas beigās notiek hromosomu despiralizācija; homologi turpina viens otru atbaidīt.
Diakinēzes stadijā, kas ir līdzīga diplonēmai, turpinās bivalentu saīsināšana un chiasmatas vājinās (samazinās), kā rezultātā veidojas diskrētas vienības hromatīdu veidā (četri). Tūlīt pēc šī posma pabeigšanas notiek kodola membrānas izšķīšana.
I metafāzē bivalenti sasniedz visaugstāko koncentrāciju. Kļūstot ovālas, tie atrodas kodola ekvatoriālajā daļā, kur veido meiotiskās metafāzes I ekvatoriālās plāksnes. Katras divvērtīgās formas nosaka chiasmata skaits un atrašanās vieta. Vīriešiem hiasmātu skaits uz vienu bivalentu I metafāzē parasti ir 1-5. XY bivalents kļūst stieņa formas, pateicoties vienai gala atrašanās vietai.
I anafāzē sākas pretējo centromēru kustība uz šūnas pretējiem poliem. Tā rezultātā tiek atdalītas homologās hromosomas. Katra hromosoma tagad sastāv no diviem hromatīdiem, ko satur centromērs, kas nedalās un paliek neskarts. Tas atšķir mejozes anafāzi I no mitozes anafāzes, kurā centromērs sadalās. Ir svarīgi atzīmēt, ka šķērsošana padara katru hromatīdu ģenētiski atšķirīgu.
Telofāzes I stadijā hromosomas sasniedz polus, kas izbeidz pirmo meiotisko dalījumu. Pēc I telofāzes notiek īsa starpfāze (interkinēze), kurā hromosomas izkliedējas un kļūst difūzas, vai arī I telofāze pāriet tieši otrā meiotiskā dalījuma II fāzē. Nevienā gadījumā DNS replikācija netiek novērota. Pēc pirmās meiotiskās dalīšanās šūnas sauc par otrās kārtas spermatocītiem. Hromosomu skaits katrā šādā šūnā samazinās no 2n līdz 1n, bet DNS saturs vēl nemainās.
Otrais meiotiskais dalījums notiek vairākās fāzēs (II fāze, II metafāze, II anafāze, II telofāze) un ir līdzīga mitotiskajai dalīšanai. II fāzē sekundāro spermatocītu hromosomas paliek polios. II metafāzē katras dubultās hromosomas centromērs sadalās, nodrošinot katrai jaunajai hromosomai savu centromēru. Sākas vārpstas veidošanās, kuras virzienā virzās jaunas hromosomas. II telofāzē beidzas otrā meiotiskā dalīšanās, kā rezultātā katrs otrās kārtas spermatocīts ražo divus spermatīdus, no kuriem pēc tam diferencējas spermatozoīdi. Tāpat kā sekundārajā spermatocītā, hromosomu skaits spermatīdā ir haploīds (1n). Tomēr spermatīdu hromosomas ir vienas, savukārt sekundāro spermatocītu II hromosomas ir dubultas, kas veidotas no diviem hromatīdiem. Līdz ar to katras spermatīda kodolā ir viens nehomologu hromosomu komplekts. Sekundārais meiotiskais dalījums ir mitotiskā tipa dalījums (ekvatoriālais dalījums). Tas atdala dubultās māsas hromatīdus un atšķiras no reducēšanas dalīšanas, kurā tiek atdalītas homologās hromosomas. Vienīgā būtiskā atšķirība no klasiskās mitozes ir tā, ka pastāv haploīds hromosomu kopums.
Tātad pirmās kārtas spermatocītu pirmā meiotiskā dalīšanās noved pie divu sekundāro spermatocītu veidošanās (otrā secība). Abas reducēšanas dalīšanas rezultātā izveidoto struktūru hromatīdas ir māsas hromatīdas. Pēdējie rodas replikācijas rezultātā pirms pirmā meiotiskā dalījuma. Katra sekundārā spermatocīta otrā meiotiskā dalīšanās rezultātā veidojas četri spermatīdi. Tādējādi tipiskā mejozē šūnas dalās divas reizes, bet hromosomas tikai vienu reizi (21. att.).
Spermatoģenēzes pēdējais posms ir saistīts ar diferenciāciju, kas beidzas ar to, ka katrs no relatīvi lielajiem, sfēriski nekustīgajiem spermatīdiem pārvēršas par mazu iegarenu kustīgu spermu.
Lielākajai daļai pieaugušo (seksuāli nobriedušu) vīriešu kārtas dzīvnieku sēkliniekos spermatoģenēze notiek pastāvīgi vai periodiski (sezonāli). Piemēram, kukaiņiem spermatoģenēzes cikla pabeigšana prasa tikai dažas dienas, savukārt zīdītājiem šis cikls ilgst vairākas nedēļas vai pat

Rīsi. 21.Hromosomu sadalījums gametoģenēzes laikā
mēnešus. Pieaugušam cilvēkam spermatoģenēze notiek visu gadu. Primitīvās spermatogonijas attīstības laiks nobriedušā spermā ir aptuveni 74 dienas.
Vīriešu reproduktīvās šūnas, ko ražo dažādu sugu organismi, raksturo mobilitāte un liela izmēra un struktūras dažādība (22. att.). Katrs cilvēka spermatozoīds sastāv no trim sekcijām – galvas, vidusdaļas un astes (23. att.). Spermas galva satur kodolu, kas satur haploīdu hromosomu komplektu.
Galva ir aprīkota ar akrosomu, kas satur lītiskos enzīmus, kas nepieciešami, lai spermas saturs iekļūtu olšūnā. Galvā ir lokalizēti arī divi centrioli - proc-

Rīsi. 22. Spermas formas

Rīsi. 23. Spermas struktūra: A - gaismas mikroskopiskais attēls dažādās plaknēs: 1 - galva, 2 - vidusdaļa, 3 - aste; B - elektronu mikroskopiskā attēla shematiska rekonstrukcija: 1 - kodols, 2 - akrosoma, 3 - centrosoma (proksimālā centriole), 4 - centrālais gredzens, 5 - mitohondriju spirāle,
6 - aksiālā vītne
simāls, kas stimulē spermas apaugļotās olšūnas dalīšanos, un distālais, kas rada astes aksimālo kātu. Spermas vidusdaļa satur astes bazālo ķermeni un mitohondrijus. Spermas asti (procesu) veido iekšējais aksimālais stienis un ārējais apvalks, kam ir citoplazmas izcelsme. Cilvēka spermatozoīdiem ir raksturīga ievērojama kustīgums.
OVOĢĒZES UN OGGLE ŠŪNAS
Olu veidošanās procesu sauc par ooģenēzi. Tās funkcijas ir nodrošināt haploīdu hromosomu kopumu olšūnas kodolā un zigotas uztura vajadzības. Ooģenēze savā izpausmē būtībā ir salīdzināma ar spermatoģenēzi.
Zīdītājiem un cilvēkiem oģenēze sākas pirmsdzemdību periodā (pirms dzimšanas). Oogonijas, kas ir mazas šūnas ar diezgan lielu kodolu un lokalizētas olnīcu folikulās, sāk diferencēties primārajos oocītos. Pēdējie veidojas jau trešajā intrauterīnās attīstības mēnesī, pēc kura tie nonāk pirmās meiotiskās dalīšanas profāzē. Brīdī, kad piedzimst meitene, visi primārie oocīti jau ir pirmās meiotiskās dalīšanās fāzē. Primārie oocīti paliek profāzē līdz sievietes pubertātes sākumam. Viens oocīts satur līdz 100 000 mitohondriju. Kad olnīcu folikuli nobriest pubertātes sākumā, primārajos oocītos atsākas meiotiskā profāze. Pirmā meiotiskā dalīšana katrai jaunattīstības olšūnai tiek pabeigta neilgi pirms šīs olšūnas ovulācijas. Pirmās meiotiskās dalīšanās un citoplazmas nevienmērīgā sadalījuma rezultātā viena iegūtā šūna kļūst par sekundāru oocītu, otra - par polāru (redukcijas) ķermeni.
Sekundārā meiotiskā dalīšanās notiek, kad sekundārais olšūns (attīstošā olšūna) no olnīcas nonāk olvados. Tomēr šī dalīšanās nav pabeigta, kamēr spermatozoīdi iekļūst sekundārajā olšūnā, kas parasti notiek olvados. Kad spermatozoīdi iekļūst sekundārajā olšūnā, tā sadalās, kā rezultātā veidojas olšūna (nobriedusi olšūna) ar kodolu, kas satur vienu komplektu.
no 23 mātes hromosomām. Otra šūna, kas rodas no šī dalījuma, ir otrs polārais ķermenis, kas nav spējīgs tālāk attīstīties. Šajā laikā polārais (redukcijas) ķermenis arī sadalās divās daļās. Tādējādi viena pirmās kārtas oocīta attīstība notiek kopā ar viena ovotīda un trīs redukcijas ķermeņu veidošanos. Olnīcās parasti šādā veidā visu mūžu nobriest 300-400 olšūnu, bet mēnesī nobriest tikai viens oocīts. Olu diferenciācijas laikā veidojas membrānas, un to kodols samazinās.
Dažās dzīvnieku sugās oģenēze notiek ātri un nepārtraukti, un tās rezultātā veidojas liels skaits olu.
Neskatoties uz līdzībām ar spermatoģenēzi, ooģenēzi raksturo dažas specifiskas pazīmes. Primārā olšūna barības materiāls (dzeltenums) nav vienmērīgi sadalīts starp četrām šūnām, kas veidojas meiotiskās dalīšanās rezultātā. Galvenais dzeltenuma daudzums tiek uzglabāts vienā lielā šūnā, savukārt polārajos ķermeņos šīs vielas ir ļoti maz. Dalīšanās rezultātā pirmais un otrais polārais ķermenis saņem tādas pašas hromosomu kopas kā sekundārie oocīti, taču tie nekļūst par dzimumšūnām. Tāpēc olas ir daudz bagātākas ar uzturvielām, salīdzinot ar spermu. Šī atšķirība ir īpaši izteikta attiecībā uz dzīvniekiem, kas dēj olas.
Zīdītāju olām ir ovāla vai nedaudz iegarena forma (24. att.), un tām ir raksturīgas tipiskas šūnu struktūras pazīmes. Tajās ir visas somatiskajām šūnām raksturīgās struktūras, tomēr olšūnas intracelulārā organizācija ir ļoti specifiska un to nosaka tas, ka olšūna ir arī vide, kas nodrošina zigotas attīstību. Viena no olu raksturīgajām iezīmēm ir to membrānu struktūras sarežģītība. Daudziem dzīvniekiem izšķir primārās, sekundārās un terciārās olu membrānas. Primārais apvalks (iekšējais) veidojas oocītu stadijā. Pārstāvot olšūnas virsmas slāni, tam ir sarežģīta struktūra, jo tajā iekļūst blakus esošo folikulu šūnu izaugumi. Sekundāro (vidējo) apvalku pilnībā veido folikulu šūnas, bet terciāro (ārējo) apvalku veido vielas, kas ir olšūnu dziedzeru sekrēcijas produkti, caur kuriem iziet olas. Piemēram, putniem
Olu terciārās membrānas ir olbaltumvielas, apakščaumalas un čaumalu membrānas. Zīdītāju olas raksturo divu membrānu klātbūtne. Olu intracelulāro komponentu struktūra ir specifiska sugu ziņā, un dažreiz tai pat ir individuālas īpašības.

Rīsi. 24. Olu struktūra: 1 - folikulu šūnas; 2 - apvalks; 3 - citoplazma; 4- kodols
MĀLĪBA
Apaugļošana ir vīriešu un sieviešu dzimumšūnu apvienošanas process, kura rezultātā veidojas zigota un pēc tam attīstās jauns organisms. Apaugļošanas procesā zigotā tiek izveidots diploīds hromosomu kopums, kas nosaka šī procesa izcilo bioloģisko nozīmi.
Atkarībā no organismu sugām, kas vairojas seksuāli, izšķir ārējo un iekšējo apaugļošanu. Ārējā apaugļošanās notiek vidē, kurā nonāk vīriešu un sieviešu reproduktīvās šūnas. Piemēram, zivju mēslošana ir ārēja. Viņu izdalītās vīriešu (piena) un sieviešu (kaviāra) reproduktīvās šūnas nonāk ūdenī, kur satiekas un apvienojas.
Iekšējo apaugļošanu nodrošina spermas pārnešana no vīrieša ķermeņa uz sievietes ķermeni. Šāda apaugļošanās notiek zīdītājiem, kuru centrālais punkts ir dzimumšūnu kodolu saplūšana. Tiek uzskatīts, ka vienas spermas saturs iekļūst olšūnā. Apaugļošanās mehānismos
pabeigšanas, daudz kas joprojām ir neskaidrs. Dati par jūras ežu apaugļošanu liecina, ka jau 2 sekundes pēc spermas un olšūnas saskares notiek izmaiņas tās plazmas membrānas elektriskajās īpašībās. Gametu saplūšana notiek 7 sekunžu laikā. Tiek pieņemts, ka tikai viena no daudzajām spermatozoīdiem satura iekļūšana olšūnā ir izskaidrojama ar izmaiņām tās plazmas membrānas elektriskajās īpašībās. Apaugļota olšūna rada zigotu. Ir divi viedokļi par iemesliem, kāpēc spermatozoīdi aktivizē olšūnu metabolismu. Daži uzskata, ka spermas saistīšanās ar ārējiem receptoriem uz šūnu virsmas ir signāls, kas caur membrānu iekļūst olšūnā un aktivizē tur inozitola trifosfātu un kalcija jonus. Citi uzskata, ka spermā ir īpašs ierosinošs faktors.
Pēdējos gados veiktie eksperimentālie sasniegumi liecina, ka zīdītāju, tostarp cilvēku, olu apaugļošana ir iespējama in vitro. Turklāt embrijus, kas izstrādāti in vitro, var implantēt sievietes dzemdē, kur tie tiek tālāk normāli attīstīti. Ir daudz gadījumu, kad piedzimst “mēģenē” bērni.
Atšķirībā no dzīvniekiem, kas vairojas ar zigoģenēzi, daudzi organismi spēj vairoties partenoģenēzes ceļā (no grieķu val. partenoss - jaunava un genos - dzimšana), kas attiecas uz organismu vairošanos no neapaugļotas olšūnas. Ir obligāta un fakultatīva partenoģenēze. Obligātā partenoģenēze ir kļuvusi par galveno noteiktu sugu organismu, piemēram, Kaukāza klinšu ķirzakas, pavairošanas metodi. Šīs sugas dzīvnieki ir tikai mātītes. Gluži pretēji, fakultatīvā partenoģenēze nozīmē, ka olas spēj attīstīties gan bez apaugļošanas, gan pēc apaugļošanas. Savukārt fakultatīvā partenoģenēze ir sieviete un vīrietis. Sieviešu partenoģenēzi bieži novēro bitēm, skudrām un rotiferiem, kurās tēviņi attīstās no neapaugļotām olām. Vīriešu partenoģenēze notiek dažās izogāmās aļģēs.
Partenoģenēze var būt gan dabiska, gan mākslīga (inducēta). Mākslīgās partenoģenēzes mehānisms ir olu kairinājums, izmantojot fizisku vai ķīmisku
ikālie faktori, kas izraisa olšūnu aktivāciju un līdz ar to neapaugļotu olšūnu attīstību. Mākslīgā partenoģenēze ir novērota daudzu sistemātisku grupu dzīvniekiem – adatādaiņiem, tārpiem, mīkstmiešiem un pat zīdītājiem.
Zināmu partenoģenēzes veidu sauc par androgēzi (no grieķu valodas. andros - cilvēks, ģenēze - izcelsme). Ja olšūnā ir inaktivēts kodols un tad tajā iekļūst vairāki spermatozoīdi, tad no šādas olšūnas vīrišķo (spermas) kodolu saplūšanas rezultātā veidojas vīrišķais organisms.
Ir gadījumi, kad partenoģenēze notiek cikliski, atkarībā no sezonas. Piemēram, rotifers, dafnijas un laputis vairojas vasarā partenoģenēzes ceļā, bet rudenī ar olšūnu apaugļošanu un zigotu veidošanos, t.i., ar zigoģenēzi.
Partenoģenēzes un tās formu loma dabā ir neliela, jo tā nenodrošina plašas organismu adaptācijas spējas.
Pretstatā zigoģenēzei un partenoģenēzei pastāv ginoģenēze, kas ir pseidogāmija, kad spermatozoīdi nonāk olšūnā un aktivizē to, bet spermas kodols nesaplūst ar olšūnas kodolu. Šajā gadījumā iegūtie pēcnācēji sastāv tikai no mātītēm. Ginoģenēze dabiski notiek nematodēs un zivīs, taču to var izraisīt arī mākslīgi. Jo īpaši ir aprakstīti mākslīgās ginoģenēzes gadījumi zīdtārpiņiem, zivīm un abiniekiem.
MAIŅI HAPLOIDIJU UN DIPLOĪDIJU.
MAINĪGAS PAAUDZES
Organismiem, kas vairojas seksuāli, to attīstībā ir raksturīgas mainīgas haploīdas un diploīdas fāzes. Daudzos organismos, tostarp zīdītājiem, šī maiņa ir regulāra, un uz to balstās organismu sugu īpašību saglabāšana.
Daudziem organismiem ir raksturīga arī paaudžu maiņa, kad aseksuāli vairojošos īpatņu paaudzes tiek aizstātas ar seksuāli vairojošos indivīdu paaudzēm, veidojoties gametām. To sauc par primāro paaudžu maiņu. Tas ir atrodams sporozoos, flagellates un daudzos augos. Primārā paaudžu maiņa ir regulāra, un tā ir klātesoša
Dabā sastopamais liecina par daudzu organismu filoģenēzē saglabāšanos gan aseksuālās, gan seksuālās vairošanās procesā. Citos organismos notiek seksuālās reprodukcijas maiņa ar partenoģenēzi. To sauc par sekundāro paaudžu maiņu. Piemēram, trematodēs seksuālo reprodukciju regulāri aizstāj partenoģenēze. Šo sekundāro paaudžu maiņu sauc par heterogoniju. Koelenterātos dažos attīstības posmos notiek pāreja no seksuālas vairošanās uz aseksuālu (veģetatīvu). Šo paaudžu sekundārās maiņas formu sauc par metaģenēzi.
SEKSUĀLAIS DIMORFISMS. HERMAFRODĪTISMS
Tēviņiem un mātītēm ir raksturīgas specifiskas fenotipiskas pazīmes. Atšķirības starp mātītēm un tēviņiem to īpašībās sauc par seksuālo dimorfismu. Dzīvniekiem tas notiek jau zemākajos evolūcijas attīstības posmos, piemēram, apaļajos helmintos un posmkājos, un vislielāko izpausmi sasniedz mugurkaulniekiem.
Ja vīriešu un sieviešu reproduktīvās šūnas ražo viens un tas pats indivīds, kuram ir gan vīriešu, gan sieviešu dzimumdziedzeri, tad šo parādību sauc par īstu hermafrodītismu. Tas ir atrodams plakanajos tārpos, annelīdos un moluskos. Plakanajiem tārpiem vīriešu un sieviešu dzimumdziedzeri darbojas visas personas dzīves laikā. Turpretim moluskiem dzimumdziedzeri pārmaiņus ražo olas un spermu.
Patiess hermafrodītisms rodas arī cilvēkiem, kas izriet no attīstības traucējumiem. Hermafrodītu genotipi ir 46 XX vai 46 XY, un vairums gadījumu ir XX (apmēram 60%). XX genotips visbiežāk sastopams negroīdu Āfrikas populāciju hermafrodītos, savukārt XY visbiežāk sastopams japāņu vidū. Abu veidu hermafrodītiem tika novērota tendence uz divpusēju dzimumdziedzeru asimetriju. Starp īstiem hermafrodītiem ir arī hromosomu mozaīkas, kurām dažās somatiskajās šūnās ir XX hromosomu pāris, citās - XY pāris.
Zināms arī viltus hermafrodītisms, kad indivīdiem ir abiem dzimumiem raksturīgi ārējie dzimumorgāni un sekundāras seksuālās īpašības, bet veidojas tikai viena veida dzimumšūnas – vīrišķās vai sievietes.
REPRODUKCIJAS METOŽU IZCELSME
Tiek uzskatīts, ka senākā ir aseksuālā vairošanās, jo īpaši veģetatīvā vairošanās. No pēdējām attīstījās vairošanās ar sporu veidošanos, kuras neapšaubāma priekšrocība ir tā, ka tā sniedz labākas iespējas sugu saglabāšanai un īpaši to izplatībai.
Seksuālā pavairošana ir visefektīvākais organismu pavairošanas veids. Tiek uzskatīts, ka tas attīstījās no aseksuāla, radās apmēram pirms 1 miljarda gadu, un pirmie posmi bija saistīti ar gametu attīstības sarežģījumiem. Primitīvajām gametām bija raksturīga nepietiekama morfoloģiskā diferenciācija, kā rezultātā izogāmija (no grieķu val. isos- vienāds, gamos - laulība), kad dzimumšūnas bija kustīgas izogametes, kas nebija diferencētas vīriešu un sieviešu formās.
Pēc tam attīstījās anizogāmija (no grieķu valodas. anisos- nevienlīdzīgs, gamos - laulība), ko raksturo diferencētu gametu klātbūtne, kas atšķiras tikai pēc izmēra. Vēlākos evolūcijas posmos radās krasas atšķirības gametu mobilitātē, formā un izmērā. Evolūcijas procesā mugurkaulnieki izstrādāja arī vairākas papildu ierīces, kas atvieglo vīriešu spermas pārnešanu sievietes reproduktīvajā traktā un rada apstākļus apaugļotas olšūnas attīstībai. Evolūcijas laikā šīs ierīces attīstījās no ekskrēcijas sistēmas, kas noveda pie uroģenitālās sistēmas veidošanās.
Diploīdais stāvoklis sniedz ārkārtējus ieguvumus organismiem, jo šajā stāvoklī notiek dažādu alēļu uzkrāšanās. Tāpēc seksuālajai pavairošanai ir arī priekšrocība, ko tā nodrošina organismiemOlielāka mainīguma iespēja salīdzinājumā ar aseksuālu, un tam ir izšķiroša nozīme evolūcijā.
Partenoģenēzei ir skaidras reproduktīvās priekšrocības, jo tā rada tikai sieviešu kārtas pēcnācējus. Tomēr tas ir reti. Tiek izmantotas divas hipotēzes, lai izskaidrotu zemo seksuālās reprodukcijas aizstāšanas biežumu ar partenoģenēzi dabiskajās organismu populācijās. Saskaņā ar vienu no tiem (mutāciju akumulatīvais) sekss ir adaptīva adaptācija, jo tas “attīra” genomu no mutācijām, kas atkārtojas laika gaitā, savukārt saskaņā ar citu hipotēzi (ekoloģisku) dzimums ir
Pirmās idejas par izaugsmi un attīstību aizsākās senajā pasaulē. Pat Hipokrāts (460-377 BC) pieņēma, ka olās jau ir pilnībā izveidots organisms, bet ļoti samazinātā formā. Pēc tam šī ideja tika attīstīta par preformacionisma doktrīnu (no lat. preformatio - priekšformēšana), kas īpaši populāra izrādījās 17.-18.gs. Preformacionisma atbalstītāji bija Hārvijs, Malpigi un daudzi citi ievērojami tā laika biologi un ārsti. Preformacionistiem strīdīgs jautājums bija tikai par to, kurās dzimumšūnās organisms bija iepriekš izveidots - sievietes vai vīriešu. Tos, kuri deva priekšroku olām, sauca par ovistiem, un tos, kas lielāku nozīmi piešķīra vīriešu dzimuma šūnām, sauca par dzīvnieku kulistiem. Preformacionisms ir metafiziska doktrīna no sākuma līdz beigām, jo tā noliedza attīstību. Izšķirošo triecienu preformacionismam deva K. Bonets (1720-1793), kurš 1745. gadā atklāja partenoģenēzi, izmantojot piemēru par laputu attīstību no neapaugļotām olām. Pēc tam preformacionisms vairs nevarēja atgūties un sāka zaudēt savu nozīmi.
Senajā pasaulē radās cita doktrīna, kas bija pretēja preformacionismam un vēlāk saņēma nosaukumu epiģenēze (no grieķu valodas. epi- pēc, ģenēze - attīstība). Tāpat kā preformacionisms, arī epiģenēze kļuva plaši izplatīta 17.-18. gadsimtā. Epiģenēzes izplatībā liela nozīme bija K. F. uzskatiem. Vilks (1733-1794), apkopots savā grāmatā “Attīstības teorija” (1759). K.F. Vilks uzskatīja, ka olā nav ne iepriekš izveidota organisma, ne tā daļu un ka tā sastāv no sākotnēji viendabīgas masas. Atšķirībā no preformistiem K.F. Vilks un citi epiģenēzes piekritēji savam laikam bija progresīvi, jo saturēja attīstības ideju. Tomēr vēlāk parādījās jauni momenti. Konkrēti, 1828. gadā K. Bērs publicēja savu darbu “Dzīvnieku attīstības vēsture”, kurā viņš parādīja, ka olšūnas saturs ir neviendabīgs, tas ir, strukturēts, un embrijam attīstoties, struktūras pakāpe palielinās. Tādējādi K. Bērs parādīja gan preformacionisma, gan epiģenēzes nekonsekvenci.
Izaugsme un attīstība ir vissvarīgākās dzīvo būtņu īpašības. Izaugsme ir visa organisma masas palielināšanās šūnu skaita palielināšanās rezultātā, savukārt attīstība ir kvalitatīvas izmaiņas organismā, ko nosaka šūnu diferenciācija un morfoģenēze, nodrošinot progresējošas izmaiņas indivīdos, sākot no olām. un beidzot ar viņu pieaugušo stāvokli.
Ontoģenēze (no grieķu. uz - būtne, ģenēze - attīstība) ir indivīda attīstības vēsture (cikls), sākot ar to dzimumšūnu veidošanos, kas to izraisīja, un beidzot ar tās nāvi. Idejas par ontoģenēzi balstās uz datiem par izaugsmi, attīstību un diferenciāciju. Ontoģenēzes pamatprincipu izpēte ir svarīga cilvēka bioloģijas izpratnei.
Mūsdienu dati par organismu attīstību noraida gan preformacionismu, gan epiģenēzi. Mūsdienu koncepciju ietvaros organisma attīstība tiek saprasta kā process, kurā agrāk izveidojušās struktūras stimulē turpmāko struktūru attīstību. Attīstības process tiek noteikts ģenētiski un ir cieši saistīts ar vidi. Līdz ar to attīstību nosaka iekšējo un ārējo faktoru vienotība.
IZAUGSMES UN ATTĪSTĪBAS VIENOTĪBA
Organisma augšana ir pakāpeniska tā masas palielināšanās un formas maiņa šūnu skaita palielināšanās un to diferenciācijas, audu un orgānu veidošanās, šūnu un audu bioķīmisko izmaiņu rezultātā. Tādējādi augšana ir kvantitatīvu izmaiņu rezultāts šūnu skaita pieauguma (ķermeņa svara) un kvalitatīvu izmaiņu veidā šūnu diferenciācijas un morfoģenēzes veidā. Šūnu diferenciācija ir process, kurā dažas šūnas morfoloģiski, bioķīmiski un funkcionāli atšķiras no citām šūnām. Dažu šūnu reprodukcija un diferenciācija vienmēr ir saskaņota ar citu šūnu augšanu un diferenciāciju. Abi šie procesi notiek visā organisma dzīves ciklā. Tā kā diferencējošās šūnas maina savu formu un šūnu grupas tiek iesaistītas formas izmaiņās, to pavada morfoģenēze, kas ir procesu kopums, kas nosaka šūnu un audu strukturālo organizāciju, kā arī organismu vispārējo morfoloģiju.
Izaugsmi var izmērīt, balstoties uz mērījumu rezultātiem, veidojot ķermeņa izmēra, svara, sausās masas, šūnu skaita, slāpekļa satura un citus rādītājus līknes.
ONTOĢĒZE UN TĀS VEIDI. ONTOĢĒZES PERIODIZĀCIJA
Ontoģenēzi atkarībā no organismu attīstības rakstura iedala tiešā un netiešā, un tāpēc tiek nošķirta tiešā un netiešā attīstība. Tiešā organismu attīstība dabā notiek ne-kūnu un intrauterīnās attīstības veidā, savukārt netiešā attīstība vērojama kāpuru attīstības veidā. Atšķirībā no ontoģenēzes sugu kategorija ir filoģenēze.
Kāpuru attīstība. Šo attīstību saprot kā netiešu attīstību, jo organismu attīstībā ir viens vai vairāki kāpuru posmi. Kāpuru attīstība ir raksturīga kukaiņiem, abiniekiem un adatādaiņiem. Šo dzīvnieku kāpuri vada neatkarīgu dzīvesveidu, pēc tam tiek pakļauti pārvērtībām. Tāpēc šo attīstību sauc arī par attīstību ar metamorfozi (skatīt zemāk).
Ne-kāpuru attīstība. Šī attīstības forma ir raksturīga organismiem, kas attīstās tieši, piemēram, zivīm, rāpuļiem un putniem, kuru olas ir bagātas ar dzeltenumu (barības materiālu). Sakarā ar to ievērojama daļa ontoģenēzes notiek ārējā vidē dētās olās. Embriju vielmaiņu nodrošina provizorisko orgānu attīstība, kas ir embrionālās membrānas (dzeltenuma maisiņš, amnions, alantois).
Intrauterīnā attīstība. Šī attīstība ir raksturīga arī organismiem, kas attīstās tieši, piemēram, zīdītājiem, tostarp cilvēkiem. Tā kā šo organismu olās ir ļoti maz barības vielu, visas embriju dzīvībai svarīgās funkcijas nodrošina mātes ķermenis, veidojot pagaidu orgānus no mātes un embrija audiem, starp kuriem galvenā ir placenta. Evolucionāri intrauterīnā attīstība ir jaunākā forma, taču tā ir visizdevīgākā embrijiem, jo tā efektīvi nodrošina to izdzīvošanu.
Ontoģenēzi iedala proembrionālajā, embrionālajā un postembrionālajā periodā. Cilvēka gadījumā attīstības periodu pirms dzimšanas sauc par pirmsdzemdību jeb pirmsdzemdību, pēc dzimšanas - pēcdzemdību. Attīstošo embriju pirms orgānu rudimentu veidošanās sauc par embriju, pēc orgānu rudimentu veidošanās – par augli.
PROEMBRYONĀLĀ ATTĪSTĪBA
Šis periods organismu individuālajā attīstībā ir saistīts ar gametu veidošanos gametoģenēzes procesā. Vīriešu reproduktīvajām šūnām nav būtisku atšķirību no citām šūnām, savukārt olas atšķiras ar to, ka tajās ir daudz dzeltenuma. Ņemot vērā dzeltenuma daudzumu un izplatību olās, tās iedala trīs veidos:
1) izoletālas olas, kas satur nedaudz dzeltenuma, kas ir vienmērīgi lokalizēts visā šūnā. Šīs olas ražo adatādaiņi (jūras eži), apakšējie akordi (lanceleti) un zīdītāji;
2) telolecitālās olas satur lielu daudzumu dzeltenuma, kas koncentrējas vienā no poliem - veģetatīvā. Šādas olas ražo mīkstmieši, abinieki, rāpuļi un putni. Piemēram, varžu olas sastāv no 50% dzeltenuma, vistu olas (parasti vistu olas) - 95%. Telolecitālo olu otrā polā (dzīvnieka) koncentrējas citoplazma un kodols;
3) centrolecitālie oocīti, kurā ir maz dzeltenuma un tas ieņem centrālo pozīciju. Citoplazma atrodas šādu olu perifērijā. Centrolecitālās olas ražo posmkāji.
Proembrionālo periodu raksturo arī tas, ka šajā periodā gametās notiek vielmaiņas procesi, kas saistīti ar DNS molekulu uzkrāšanos.
EMBRIONĀLĀ ATTĪSTĪBA
Embrioģenēze (no grieķu val. etbryop - dīglis) jeb embrionālais periods sākas ar vīriešu un sieviešu dzimumšūnu saplūšanu, kas atspoguļo olšūnu apaugļošanas procesu.
Organismiem, kam raksturīga intrauterīna attīstība, embrionālais periods beidzas ar piedzimšanu, savukārt organismos, kam raksturīgs kāpuru un ne-kūnu attīstības tips, embrionālais periods beidzas ar organisma atbrīvošanos no attiecīgi olšūnas vai embrija membrānām. Embrionālajā periodā izšķir zigotas, šķelšanās, blastulas, dīgļu slāņu veidošanās, histoģenēzes un organoģenēzes stadijas.
Zigota.Apaugļošana ietver virkni procesu, kuros vīrieša reproduktīvā šūna uzsāk olšūnas attīstību. Olā, ko aktivizē vīriešu dzimuma gameta, notiek vairāki fizikāli un ķīmiski procesi, tostarp pastiprināta proteīnu sintēze. Protoplazmas kustība noved pie olšūnas divpusējas simetrijas izveidošanas. Kodoli saplūst, un tiek atjaunots diploīds hromosomu komplekts. Tādējādi tiek izveidots vienšūnas organisms.
Sadalīšana.Tas atspoguļo zigotas (apaugļotas olšūnas) sākotnējo attīstības periodu, kas sastāv no zigotas sadalīšanas mitozes ceļā. Dalīšanās sākas ar vagas parādīšanos uz olas virsmas. Pirmā vaga noved pie divu šūnu veidošanās - divi blastomēri, otrā - četri blastomēri, trešā - astoņi blastomēri (25. att.). Šūnu grupu, kas veidojas secīgu fragmentāciju rezultātā, sauc par morulu (no latīņu valodas. morum- zīdkoks).
Šīs stadijas bioloģiskā nozīme ir tajā, ka no lielas šūnas, kas ir olšūna, veidojas mazākas šūnas, kurās tiek samazināta citoplazmas un kodola attiecība.
Zigotas sadrumstalotība beidzas ar daudzšūnu struktūras veidošanos, ko sauc par blastulu (no grieķu valodas. blastos - asns). Šī struktūra ir veidota kā vezikula, ko sauc par blastodermu, kas sastāv no viena šūnu slāņa. Tagad šīs šūnas sauc par embrionālām. Blastulas izmērs ir līdzīgs olai. Fragmentācijas periodā palielinās kodolu skaits un kopējais DNS skaits. Tiek sintezēts arī neliels daudzums mRNS un tRNS, savukārt ribosomu RNS vēl nav nosakāms.
Visi dzīvnieki iziet blastula stadiju, taču katram gadījumam ir savas īpatnības. Zīdītājiem dalīšanās notiek nevienmērīgi, tāpēc morulas sastāv no dažāda skaita šūnu. Turklāt daļa šūnu veido struktūru, ko sauc par trofoblastu,

Blastocoel Larva Kurkuķis
formā
Rīsi. 25.Zigotu sadrumstalotība un blastulu veidošanās dažādos organismos: 1 - oriģināla ola; 2 - divi blastomēri; 3 - četri blastomēri; 4 - astoņi blastomēri; 5 - blastula; 6 - pieaugušo forma
kuras šūnas baro embriju un, pateicoties fermentiem, nodrošina tā iekļūšanu dzemdes sieniņā. Vēlāk trofoblastu šūnas atdalās no embrija un veido pūslīšu, kas ir piepildīta ar šķidrumu no dzemdes audiem.
Šīs stadijas bioloģiskā nozīme slēpjas apstāklī, ka no lielas šūnas, piemēram, olšūnas, veidojas mazākas šūnas, kurās tiek samazināta citoplazmas attiecība pret kodolu un kodolam ir jauna citoplazmas vide.
Gastrulācija(no grieķu val kuņģis - kuģa dobums). Tas ir embriju šūnu kustības process pēc blastulas veidošanās, ko pavada divu vai trīs (atkarībā no dzīvnieka veida) embrija slāņu jeb tā saukto dīgļu slāņu veidošanās (26. att.). ).
Izolecitālo olu attīstība (gastrulācija) notiek ar veģetatīvā pola invagināciju (invagināciju) blastulas iekšpusē, kā rezultātā pretējie poli gandrīz saplūst, un blastocoel (blastulas dobums) gandrīz vai pilnībā izzūd. Ārējais
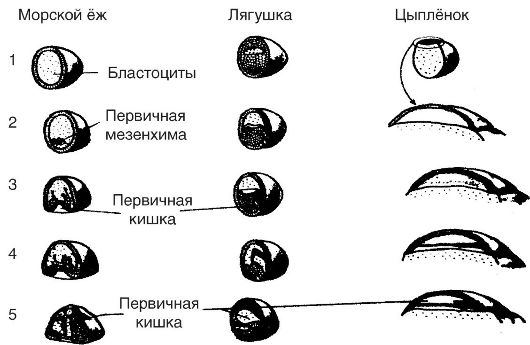
Rīsi. 26.Gastrulācija dažādos organismos: 1 - blastocoel; 2 - primārais mezenhīms; 3, 4, 5 - primārās zarnas
Embrionālo šūnu slāni sauc par ektodermu (no grieķu valodas. ectos -ārā, derma -āda) jeb ārējais dīgļu slānis, savukārt iekšējais ir endoderma (no grieķu valodas. entos - iekšpusē), vai iekšējais dīgļu slānis. Šajā gadījumā izveidoto dobumu sauc par gastrocoel jeb primāro zarnu, kuras ieeju sauc par blastoporu (primāro muti).
Divu dīgļu slāņu attīstība ir raksturīga sūkļiem un koelenterātiem. Tomēr hordātiem gastrulācijas periodā ir raksturīga trešā dīgļu slāņa attīstība - mezoderma (no grieķu val. mezo - vidus), veidojas starp ektodermu un endodermu.
Gastrulācija ir nepieciešams priekšnoteikums turpmākajiem attīstības posmiem, jo tā nogādā šūnas tādā stāvoklī, kas ļauj veidot orgānus. Embrionālais materiāls, kas diferencēts trīs embrionālās anlagās, rada visus jaunattīstības embrija audus un orgānus.
Dīgļu slāņu attīstību (diferenciāciju) pavada dažādu audu un orgānu veidošanās no tiem. Jo īpaši ādas epiderma, nagi un mati, tauku un sviedru dziedzeri, nervu sistēma (smadzenes, muguras smadzenes, gangliji, nervi), maņu orgānu receptoršūnas, acs lēca, mutes epitēlijs. , deguna dobums un tūpļa, zobs -
nav emalja. No endodermas attīstās barības vada, kuņģa, zarnu, žultspūšļa, trahejas, bronhu, plaušu, urīnizvadkanāla, kā arī aknu, aizkuņģa dziedzera, vairogdziedzera, epitēlijķermenīšu un aizkrūts dziedzeru epitēlijs. No mezodermas attīstās gludie muskuļi, skeleta un sirds muskuļi, derma, saistaudi, kauli un skrimšļi, zobu dentīns, asinis un asinsvadi, apzarnis, nieres, sēklinieki un olnīcas. Cilvēkiem smadzenes un muguras smadzenes ir pirmās, kas atdalās. Pēc 2 mēnešiem parādās gandrīz visas ķermeņa struktūras. Embrija savienojums ar vidi tiek veikts caur pagaidu orgāniem. Organoģenēze beidzas embrija perioda beigās. Ja galīgā mutes atvere veidojas primārās mutes vietā (blastopora), tad šos dzīvniekus sauc par protostomiem (tārpiem, mīkstmiešiem, posmkājiem).
Ja galīgā mute veidojas pretējā vietā, tad šos dzīvniekus sauc par deuterostomām (adatādaiņiem, hordātiem).
Lai nodrošinātu embrija saikni ar vidi, tiek izmantoti tā sauktie pagaidu orgāni, kas pastāv īslaicīgi. Atkarībā no olu veida pagaidu orgāni ir dažādas struktūras. Zivīm, rāpuļiem un putniem pagaidu orgāni ietver dzeltenuma maisiņu. Zīdītājiem dzeltenuma maisiņš veidojas embrioģenēzes sākumā, bet neattīstās. Vēlāk tas tiek samazināts. Embrija ārējo membrānu sauc par horionu. Tas ieaug dzemdē. Vietu, kurā notiek vislielākā ieaugšana dzemdē, sauc par placentu. Embrijs ir savienots ar placentu caur nabassaiti jeb nabassaiti, kurā atrodas asinsvadi, kas nodrošina placentas asinsriti. Augļa vielmaiņa tiek nodrošināta caur placentu.
Embrija daļu veidojošā mijiedarbība balstās uz noteiktiem koordinētiem vielmaiņas procesiem. Attīstības modelis ir heterohronija, ar ko saprot dažādu orgānu anlagu veidošanos laika gaitā un atšķirīgu to attīstības intensitāti. Tie orgāni un sistēmas, kurām vajadzētu sākt darboties agrāk, attīstās ātrāk. Piemēram, cilvēkiem augšējo ekstremitāšu rudimenti attīstās ātrāk nekā apakšējās.
Embriji ir ļoti jutīgi pret dažādām ietekmēm. Tāpēc tiek izdalīti kritiskie periodi, t.i., periodi, kad embriji ir visjutīgākie pret kaitīgajiem faktoriem. Cilvēku gadījumā kritiskie embriju ontoģenēzes periodi
ir pirmās dienas pēc apaugļošanas, placentas veidošanās un dzemdību laiks.
Somatisko šūnu kodoli spēj nodrošināt normālu olšūnu attīstību, kas tika noskaidrots eksperimentos par somatisko šūnu kodolu transplantāciju olās, kurās nav kodolu.
Eksperimenti arī parādīja, ka viena blastomēra transformācija no vienas šķirnes aitu 8 un 16 šūnu embrijiem citas šķirnes olas kodolā (pēc pēdējās pārgriešanas uz pusēm) notika kopā ar dzīvotspējīgu embriju attīstību. un jēru piedzimšana.
Kopš Hipokrāta laikiem (5. gadsimtā pirms mūsu ēras) tiek apspriests jautājums par iemesliem, kas izraisa augļa dzimšanu. Jo īpaši pats Hipokrāts ierosināja, ka augļa attīstība sāk savu dzimšanu. Jaunākais angļu pētnieku eksperimentālais darbs, kas veikts ar aitām, ir parādījis, ka aitām atnešanās sākšanos kontrolē komplekss hipotalāms + hipofīze + augļa virsnieru dziedzeri. Hipotalāma kodolu bojājumi, hipofīzes priekšējās daivas vai virsnieru dziedzeru noņemšana paildzina aitu grūsnību. Gluži pretēji, adenokortikotropā hormona (hipofīzes priekšējās daļas sekrēta) vai kortizola (virsnieru dziedzeru sekrēta) ievadīšana aitām saīsina to grūtniecības ilgumu.
Tātad augstāko eikariotu attīstības procesā viena apaugļota zigota šūna turpmākās attīstības laikā mitozes rezultātā rada dažāda veida šūnas - epitēlija, nervu, kaulu, asins šūnas un citas, kurām raksturīgas morfoloģijas un makromolekulārā sastāva daudzveidība. Taču arī dažāda veida šūnām ir raksturīgi, ka tās satur vienus un tos pašus gēnu komplektus, bet ir ļoti specializētas, pildot tikai vienu vai vairākas specifiskas funkcijas, proti, vieni gēni “strādā” šūnās, bet citi ir neaktīvi. Piemēram,

Rīsi. 27. Monozigotisku dvīņu attīstība
tikai sarkanās asins šūnas ir specifiskas hemoglobīna sintēzē un uzglabāšanā. Tādā pašā veidā keratīnu sintezē tikai epidermas šūnas. Tāpēc jau sen ir radušies jautājumi par somatisko šūnu kodolu ģenētisko identitāti un par apaugļotu olšūnu attīstības kontroles mehānismiem kā priekšnoteikumu, lai izprastu šūnu diferenciācijas pamatā esošos mehānismus. Cilmes šūnām ir liela nozīme šūnu diferenciācijā (skatīt zemāk). Eksperimenti ar pelēm ir parādījuši, ka viena veida šūnas spēj pārveidoties par cita veida šūnām. Jo īpaši ir pierādīts, ka diferencētas vai diferencētas aknu šūnas pārvēršas aizkuņģa dziedzera šūnās.
Kopš 50. gadiem. XX gadsimts Daudzās laboratorijās tika veikti eksperimenti par veiksmīgu somatisko šūnu kodolu transplantāciju olās, kurām mākslīgi atņemti pašu kodoli. DNS pētījums no dažādu diferencētu šūnu kodoliem parādīja, ka gandrīz visos gadījumos genomi satur vienādus nukleotīdu pāru secību kopumus. Ir zināmi izņēmumi, kad zīdītāju sarkanās asins šūnas zaudē kodolus pēdējā diferenciācijas stadijā. Bet pa šo laiku baseini
Dažas hemoglobīna mRNS jau ir sintezētas, tāpēc sarkano asins šūnu kodoli vairs nav vajadzīgi. Citi piemēri ietver imūnglobulīnu un T-šūnu gēnus, kas tiek modificēti attīstības laikā.
Viens no galvenajiem posmiem embrionālās ontoģenēzes kontroles mehānismu izpratnes virzienā bija 1960.-1970. gados veikto eksperimentu rezultāti. Angļu pētnieks D. Gurdons, lai noskaidrotu, vai somatisko šūnu kodoliem ir spēja nodrošināt olšūnu tālāku attīstību, ja šie kodoli tiek ievadīti olās, no kurām iepriekš izņemti paši kodoli. Attēlā 28. attēlā parādīta viena no šiem eksperimentiem diagramma, kurā kurkuļa somatisko šūnu kodoli tika pārstādīti varžu olās ar iepriekš izņemtiem kodoliem. Šie eksperimenti parādīja, ka somatisko šūnu kodoli patiešām var nodrošināt olšūnu tālāku attīstību, jo tie spēja apaugļot olas un "piespiest" tās attīstīties tālāk. Tas pierādīja dzīvnieku klonēšanas iespēju.

Rīsi. 28.Eksperimenta shēma par somatisko šūnu kodolu transplantāciju olās ar kodolu (D. Gurdon, 1968)
Vēlāk citi pētnieki veica eksperimentus, kuros viņi parādīja, ka atsevišķu blastomēru pārnešana no vienas šķirnes aitu 8 un 16 dienu embrijiem citas šķirnes aitu olšūnas pusē (pēc pēdējās pārgriešanas uz pusēm) notika kopā ar aitu. dzīvotspējīgu embriju veidošanās ar sekojošu jēru piedzimšanu.
1997. gada sākumā angļu autori konstatēja, ka somatisko šūnu kodolu (pieaugušu aitu embriju šūnas, augļa vai tesmeņa šūnas) ievadīšana mākslīgi atdalītās aitu olās un pēc tam šādi apaugļoto olšūnu implantācija aitas dzemdē notiek kopā ar grūtniecības iestāšanās, kam seko jēru piedzimšana (29. att.). Vienu no jēriem sauca Dollija. 2003. gadā Dollija nomira. Šajā laikā iegūti peļu, govju, trušu, zirgu, žurku un citu dzīvnieku embriji.

Kultūras ievadīšana mātes ķermenī
Rīsi. 29.Somatisko šūnu kodolu transplantācija olās ar kodolu
Šo rezultātu izvērtējums liecina, ka zīdītājus var vairoties aseksuāli, iegūstot pēcnācējus dzīvniekiem, kuru šūnas satur tēva vai mātes izcelsmes kodolmateriālu, atkarībā no donoraitas dzimuma. Šādās šūnās
tikai citoplazma un mitohondriji ir mātes izcelsmes. Šim secinājumam ir ārkārtīgi svarīga vispārēja bioloģiskā nozīme, un tas paplašina mūsu uzskatus par dzīvnieku reproduktīvo potenciālu. Taču svarīgi ir arī piebilst, ka runa ir par ģenētiskām manipulācijām, kuras dabā neeksistē. No otras puses, praktiski šīs ģenētiskās manipulācijas ir tiešs veids, kā klonēt augsti organizētus dzīvniekus ar vēlamām īpašībām, kam ir liela ekonomiska nozīme. Medicīnā šo ceļu var izmantot nākotnē, lai pārvarētu vīriešu neauglību.
Tātad normālai embrija attīstībai nepieciešamā ģenētiskā informācija netiek zaudēta šūnu diferenciācijas laikā tā saukto cilmes šūnu dēļ, kurām ir potenciāls attīstīties par dažāda veida ķermeņa šūnām. Kad cilmes šūnas dalās, katrai jaunajai šūnai ir iespēja vai nu palikt par cilmes šūnu, vai kļūt par šūnu ar specializētāku funkciju (muskuļu šūnas, asins šūnas vai smadzeņu šūnas). Apaugļota olšūna ir totipotenta, jo tā rada dažāda veida šūnas organismā. Totipotentās cilmes šūnas var radīt jebkura veida šūnas, izņemot tās, kas nepieciešamas augļa attīstībai. Cilmes šūnas, kas var radīt dažādus šūnu veidus, parasti sauc multitotipotentsšūnas. Ir konstatēts, ka pieaugušo cilmes šūnas var radīt diferencētas šūnas no embrija nesaistītiem audiem. Somatiskajām šūnām ir arī īpašība, ko sauc par totipotenci, t.i., to genoms satur visu informāciju, ko tās saņēma no apaugļotās olšūnas, kas tās radīja diferenciācijas rezultātā. Šo datu klātbūtne neapšaubāmi nozīmē, ka šūnu diferenciācija ir pakļauta ģenētiskai kontrolei. Cilmes šūnu izpēte ietekmē medicīnu.
Ir konstatēts, ka intensīva proteīnu sintēze pēc apaugļošanas lielākajā daļā eikariotu nav saistīta ar mRNS sintēzi. Ooģenēzes izpēte mugurkaulniekiem, jo īpaši abiniekiem, ir parādījusi, ka intensīva transkripcija notiek pat mejozes I fāzes (īpaši diplonēmas) laikā. Tāpēc gēnu transkripti mRNS vai pro-mRNS molekulu veidā tiek glabāti olās neaktīvā stāvoklī. Ir konstatēts, ka embrionālajās šūnās ir tā sauktā asimetriskā
trīskāršā dalīšanās, kas sastāv no tā, ka embrionālās šūnas dalīšanās rezultātā rodas divas šūnas, no kurām tikai viena pārmanto transkripcijā iesaistītās olbaltumvielas. Tādējādi nevienlīdzīgais transkripcijas faktoru sadalījums starp meitas šūnām izraisa dažādu gēnu kopu ekspresiju tajās pēc dalīšanās, t.i., šūnu diferenciāciju.
Abiniekiem un, iespējams, lielākajai daļai mugurkaulnieku ģenētiskās programmas, kas kontrolē agrīnu attīstību (pirms blastulas stadijas), tiek izveidotas ooģenēzes laikā. Vēlākajos attīstības posmos, kad sākas šūnu diferenciācija (aptuveni no gastrulas stadijas), ir nepieciešamas jaunas programmas gēnu ekspresijai. Tādējādi šūnu diferenciācija ir saistīta ar ģenētiskās informācijas pārprogrammēšanu vienā vai otrā virzienā.
Šūnu diferenciācijas raksturīga iezīme ir tā, ka tā neatgriezeniski noved pie viena vai otra šūnu veida. Šo procesu sauc apņēmība un ir arī ģenētiskā kontrolē, un, kā tagad tiek pieņemts, šūnu diferenciāciju un noteikšanu regulē šūnu mijiedarbība, pamatojoties uz signāliem, ko veic peptīdu augšanas faktori caur tirozīna kināzes receptoriem. Droši vien šādu sistēmu ir daudz. Viens no tiem ir tāds, ka muskuļu un nervu šūnu diferenciāciju regulē neiroregulīni, kas ir membrānas proteīni, kas darbojas caur vienu vai vairākiem tirozīna kināzes receptoriem.
Determinācijas ģenētisko kontroli pierāda arī tā saukto homeiotropo jeb homeotisko mutāciju esamība, kas, kā pierādīts, kukaiņiem izraisa determinācijas izmaiņas konkrētos iztēles diskos. Tā rezultātā dažas ķermeņa daļas attīstās nevietā. Piemēram, Drosofilā mutācijas pārveido antenas diska noteikšanu diskā, kas attīstās par ekstremitātes papildinājumu, kas stiepjas no galvas. Ģints kukaiņiem Oftalmoptera no acs diska var veidoties spārnu struktūras. Pelēm ir pierādīts gēnu kopas (komplekss) Hox esamība, kas sastāv no 38 gēniem un kontrolē ekstremitāšu attīstību.
Jautājums par gēnu aktivitātes regulēšanu embrionālās attīstības laikā ir neatkarīgs. Tiek uzskatīts, ka diferenciācijas laikā gēni darbojas dažādos laikos, kas izpaužas
dažādu mRNS transkripcijā dažādās diferencētās šūnās, t.i., notiek gēnu represijas un derepresijas. Piemēram, jūras ežu blastocītos RNS transkribēto gēnu skaits ir 10%, žurku aknu šūnās arī 10%, liellopu aizkrūts dziedzera šūnās – 15%. Tiek pieņemts, ka nehistona proteīni ir iesaistīti gēnu transkripcijas stāvokļa kontrolē. Šo pieņēmumu apstiprina šādi dati. Kad šūnu hromatīns ir fāzē S pārrakstīts sistēmā in vitro, tad tiek sintezēts tikai histona mRNS, kam seko histoni. Gluži pretēji, ja tiek izmantots šūnu hromatīns no β-fāzes, netiek sintezēts histona mRNS. Kad no β-fāzes hromatīna tiek atdalīti ne-virzuļa proteīni un tiek aizstāti ar nehistona hromosomu proteīniem, kas sintezēti fāze S, tad pēc šāda hromatīna transkripcijas in vitro tiek sintezēts histona mRNS. Turklāt, ja nehistona proteīni nāk no 1 fāzes šūnām un DNS un histoni nāk no S fāzes šūnām, histona mRNS netiek sintezēts. Šie rezultāti liecina, ka hromatīnā esošie nehistona proteīni nosaka spēju transkribēt gēnus, kas kodē histonus Tāpēc uzskatu, ka nehistona hromosomu proteīniem var būt svarīga loma gēnu regulēšanā un ekspresijā eikariotos.
Pieejamie dati liecina, ka olbaltumvielas un steroīdie hormoni ir iesaistīti transkripcijas regulēšanā dzīvniekiem. Olbaltumvielu (insulīna) un steroīdu (estrogona un testosterona) hormoni ir divas signalizācijas sistēmas, ko izmanto starpšūnu sakaros. Augstākiem dzīvniekiem hormoni tiek sintezēti specializētās sekrēcijas šūnās. Nokļūstot asinsritē, tie nonāk audos. Tā kā olbaltumvielu hormonu molekulas ir salīdzinoši lielas, tās neiekļūst šūnās, tāpēc to iedarbību nodrošina mērķa šūnu membrānās lokalizēti receptoru proteīni un cikliskā AMP (cAMP) intracelulārais līmenis. Gluži pretēji, steroīdie hormoni ir mazas molekulas, kā rezultātā tie caur membrānām viegli iekļūst šūnās. Nokļūstot šūnās, tie saistās ar specifiskiem receptoru proteīniem, kas atrodami tikai mērķa šūnu citoplazmā. Tiek apsvērti kompleksi hormonu-olbaltumvielu receptors, Koncentrējoties mērķa šūnu kodolos, tie aktivizē specifisku gēnu transkripciju, mijiedarbojoties ar noteiktiem
mi nehistona proteīni, kas saistās ar specifisku gēnu promotora reģioniem. Līdz ar to kompleksa iesiešana hormons + proteīns (olbaltumvielu receptors) ar nehistona proteīniem atbrīvo promotora reģionus RNS polimerāzes kustībai. Apkopojot datus par embrionālā perioda ģenētisko kontroli organismu ontoģenēzē, varam secināt, ka tā gaitu kontrolē diferenciāla gēnu darbības ieslēgšanās un izslēgšana dažādās šūnās (audios) caur to derepresiju un represijām.
PĒCEMBRYONĀLĀ ATTĪSTĪBA
Pēc organisma piedzimšanas sākas tā pēcdzemdību attīstība (cilvēkam pēcdzemdību), kas dažādos organismos ilgst no vairākām dienām līdz pat simtiem gadu atkarībā no to sugas. Līdz ar to dzīves ilgums ir organismiem raksturīga suga, kas nav atkarīga no to organizācijas līmeņa (skat. zemāk).
Pēcembrionālajā ontoģenēzē tiek izšķirts juvenīlais un pubertātes periods, kā arī vecuma periods, kas beidzas ar nāvi.
Nepilngadīgais periods. Šis periods (no lat. juvenilis- jauni) nosaka laiks no organisma dzimšanas līdz pubertātei. Tas notiek dažādos veidos un ir atkarīgs no organismu ontoģenēzes veida. Šo periodu raksturo tieša vai netieša attīstība.
Attiecībā uz organismiem, kuriem raksturīga tieša attīstība (daudzi bezmugurkaulnieki, zivis, rāpuļi, putni, zīdītāji, cilvēki), no olu čaumalām izšķilušies vai jaundzimušie ir līdzīgi pieaugušām formām, no pēdējiem atšķiras tikai ar mazākiem izmēriem, kā arī kā atsevišķu orgānu nepietiekama attīstība un nepilnīgas ķermeņa proporcijas (30. att.).
Tiešai attīstībai pakļauto organismu augšanas juvenīlajā periodā raksturīga iezīme ir tā, ka palielinās šūnu skaits un izmērs, mainās ķermeņa proporcijas. Cilvēka augšana dažādos viņa ontoģenēzes periodos ir parādīta attēlā. 31. Dažādu cilvēka orgānu augšana ir nevienmērīga. Piemēram, galvas augšana beidzas bērnībā, kājas sasniedz proporcionālu izmēru par aptuveni 10 gadiem. Ārējie dzimumorgāni ļoti ātri aug vecumā no 12 līdz 14 gadiem. Izšķir noteiktu un nenoteiktu izaugsmi. Noteikta izaugsme ir raksturīga organismiem, kas pārtrauc augt noteiktā vecumā,
piemēram, kukaiņi, zīdītāji, cilvēki. Nenoteikta augšana ir raksturīga organismiem, kas aug visu mūžu, piemēram, mīkstmiešiem, zivīm, abiniekiem, rāpuļiem un daudzām augu sugām.

Rīsi. trīsdesmit.Dažādu sugu organismu tieša un netieša attīstība
Netiešās attīstības gadījumā organismos notiek transformācijas, ko sauc par metamorfozēm (no lat. metamorfoze - transformācija).

Rīsi. 31.Izaugsme un attīstība dažādos cilvēka ontoģenēzes periodos
Tie atspoguļo organismu modifikācijas attīstības laikā. Metamorfozes plaši sastopamas koelenterātos (hidrās, medūzās, koraļļu polipi), plakanajos tārpos (fasciola), apaļtārpos (apaļtārpi), mīkstmiešu (austeres, mīdijas, astoņkāji), posmkājos (vēži, upes krabji, omāri, garneles, skorpi, ērces). , kukaiņi) un pat dažos hordātos (tunikāti un abinieki). Šajā gadījumā izšķir pilnīgas un nepilnīgas metamorfozes. Izteiksmīgākās metamorfozes formas tiek novērotas kukaiņiem, kuriem notiek gan nepilnīga, gan pilnīga metamorfoze.
Nepilnīga transformācija ir attīstība, kurā no olu čaumalām iznāk organisms, kura uzbūve ir līdzīga pieauguša organisma uzbūvei, taču tā izmērs ir daudz mazāks. Šādu organismu sauc par kāpuru. Augšanas un attīstības procesā kāpuru izmērs palielinās, bet esošais hitinizētais segums neļauj tālāk palielināt ķermeņa izmērus, kas noved pie molting, t.i., hitinizētā seguma nobiršanas, zem kura ir mīksta kutikula. Pēdējais iztaisnojas, un to papildina dzīvnieka lieluma palielināšanās. Pēc vairākām mešanas reizēm dzīvnieks sasniedz briedumu. Nepilnīga transformācija ir raksturīga, piemēram, blakšu attīstības gadījumā.
Pilnīga metamorfoze ir attīstība, kurā no olu čaumalām izdalās kāpurs, būtiski atšķirīgs
struktūrā no pieaugušiem indivīdiem. Piemēram, tauriņiem un daudziem kukaiņiem kāpuri ir kāpuri. Kāpuri ir pakļauti kausēšanai un var izkausēt vairākas reizes, pēc tam pārvēršoties kūniņās. No pēdējām veidojas pieaugušas formas (imago), kas neatšķiras no sākotnējām.
Mugurkaulniekiem metamorfoze notiek starp abiniekiem un kaulainām zivīm. Kāpuru stadiju raksturo pagaidu orgānu klātbūtne, kas vai nu atkārto senču īpašības, vai arī tiem ir nepārprotami adaptīva nozīme. Piemēram, kurkulim, kas ir vardes kāpuru forma un atkārto sākotnējās formas īpašības, ir raksturīga zivij līdzīga forma, žaunu elpošana un viens asinsrites aplis. Kurkuļu adaptīvās iezīmes ir to piesūcekņi un garās zarnas. Kāpuru formām raksturīgi arī tas, ka, salīdzinot ar pieaugušām formām, tie ir pielāgoti dzīvei pavisam citos apstākļos, ieņemot citu ekoloģisko nišu un citu vietu barības ķēdē. Piemēram, varžu kāpuriem ir žaunu elpošana, bet pieaugušām formām ir plaušu elpošana. Atšķirībā no pieaugušām formām, kas ir plēsēji, varžu kāpuri barojas ar augu pārtiku.
Organismu attīstības notikumu secību bieži sauc par dzīves cikliem, kas var būt vienkārši vai sarežģīti. Vienkāršākie attīstības cikli ir raksturīgi, piemēram, zīdītājiem, kad no apaugļotas olšūnas attīstās organisms, kas atkal ražo oliņas utt.. Sarežģīti bioloģiskie cikli ir cikli dzīvniekiem, kuriem raksturīga attīstība ar metamorfozi. Zināšanām par bioloģiskajiem cikliem ir praktiska nozīme, īpaši gadījumos, kad organismi ir patogēni vai patogēnu nesēji dzīvniekiem un augiem.
Ar metamorfozi saistītā attīstība un diferenciācija ir dabiskās atlases rezultāts, kuras dēļ daudzas kāpuru formas, piemēram, kukaiņu kāpuri un varžu kurkuļi, ir labāk pielāgojušies videi nekā pieaugušas seksuāli nobriedušas formas.
Puberitāte. Šo periodu sauc arī par briedumu, un tas ir saistīts ar organismu dzimumbriedumu. Organismu attīstība šajā periodā sasniedz maksimumu.
Augšanu un attīstību pēcdzemdību periodā lielā mērā ietekmē vides faktori. Augiem noteicošie faktori ir gaisma, mitrums, temperatūra, barības vielu daudzums un kvalitāte augsnē. Dzīvniekiem ļoti svarīga ir pareiza barošana (olbaltumvielu, ogļhidrātu, lipīdu, minerālsāļu, vitamīnu, mikroelementu klātbūtne barībā). Svarīgs ir arī skābeklis, temperatūra, gaisma (D vitamīna sintēze).
Dzīvnieku organismu augšana un individuālā attīstība ir pakļauta neirohumorālajam regulējumam, ko veic humorālie un nervu regulējošie mehānismi. Augos ir atklātas hormoniem līdzīgas aktīvās vielas, ko sauc par fitohormoniem. Pēdējie ietekmē augu organismu dzīvībai svarīgās funkcijas.
Dzīvnieku šūnās to dzīvības procesu laikā tiek sintezētas ķīmiski aktīvas vielas, kas ietekmē dzīvības procesus. Bezmugurkaulnieku un mugurkaulnieku nervu šūnas ražo vielas, ko sauc par neirosekretiem. Endokrīnie jeb iekšējie sekrēcijas dziedzeri izdala arī vielas, ko sauc par hormoniem. Endokrīnie dziedzeri, īpaši tie, kas saistīti ar augšanu un attīstību, tiek regulēti ar neirosekrēciju. Posmkāju augšanas un attīstības regulēšanu ļoti labi ilustrē hormonu ietekme uz kausēšanu. Kāpuru sekrēciju sintēzi šūnās regulē hormoni, kas uzkrājas smadzenēs. Īpašs vēžveidīgo dziedzeris ražo hormonu, kas kavē kausēšanu. Šo hormonu līmenis nosaka kausēšanas biežumu. Kukaiņiem ir noteikta olu nobriešanas un diapauzes hormonālā regulēšana.
Mugurkaulniekiem endokrīnie dziedzeri ir hipofīze, epifīze, vairogdziedzeris, epitēlijķermenīšu dziedzeris, aizkuņģa dziedzeris, virsnieru dziedzeri un dzimumdziedzeri, kas ir cieši saistīti viens ar otru. Mugurkaulnieku hipofīze ražo gonadotropo hormonu, kas stimulē dzimumdziedzeru darbību. Cilvēkiem hipofīzes hormons ietekmē augšanu. Ar trūkumu attīstās pundurisms, ar pārpalikumu - gigantisms. Čiekurveida dziedzeris ražo hormonu, kas ietekmē dzīvnieku seksuālās aktivitātes sezonālās svārstības. Vairogdziedzera hormons ietekmē kukaiņu un abinieku metamorfozi. Zīdītājiem nepietiekama vairogdziedzera attīstība izraisa augšanas aizkavēšanos un dzimumorgānu nepietiekamu attīstību. Cilvēkiem vairogdziedzera defekta dēļ tiek aizkavēta pārkaulošanās un augšana.
(pundurisms), pubertāte nenotiek, garīgā attīstība apstājas (kretinisms). Virsnieru dziedzeri ražo hormonus, kas ietekmē vielmaiņu, šūnu augšanu un diferenciāciju. Dzimumdziedzeri ražo dzimumhormonus, kas nosaka sekundārās seksuālās īpašības. Dzimumdziedzeru noņemšana noved pie neatgriezeniskām vairāku īpašību izmaiņām. Piemēram, kastrētiem gaiļiem ķemmes augšana apstājas un zūd dzimuminstinkts. Kastrēts vīrietis iegūst ārēju līdzību ar sievieti (uz ādas neaug bārda un mati, krūškurvja un iegurņa zonā nogulsnējas tauki, saglabājas balss tembrs utt.).
Augu fitohormoni ir auksīni, citokinīni un giberelīni. Tie regulē šūnu augšanu un dalīšanos, jaunu sakņu veidošanos, ziedu attīstību un citas augu īpašības.
Visos ontoģenēzes periodos organismi spēj atjaunot zaudētās vai bojātās ķermeņa daļas. Šo organismu īpašību sauc reģenerācija, kas var būt fizioloģiski un reparatīvi.
fizioloģiskā reģenerācija - Tā ir zaudēto ķermeņa daļu aizstāšana ķermeņa dzīves laikā. Šāda veida reģenerācijas ir ļoti izplatītas dzīvnieku pasaulē. Piemēram, posmkājiem to attēlo kausēšana, kas saistīta ar augšanu. Rāpuļiem reģenerācija izpaužas kā astes un zvīņu nomaiņa, putniem - spalvas, nagi un piešiem. Zīdītājiem fizioloģiskās atjaunošanās piemērs ir briežu ikgadējā ragu izmešana.
Reparatīvā reģenerācija - Tā ir vardarbīgi atrautās organisma ķermeņa daļas atjaunošana. Šāda veida reģenerācija ir iespējama daudziem dzīvniekiem, taču tās izpausmes ir dažādas. Piemēram, tas ir izplatīts hidrās un ir saistīts ar pēdējo vairošanos, jo viss organisms atjaunojas no daļas. Citos organismos reģenerācija izpaužas kā atsevišķu orgānu spēja atgūties pēc kādas daļas zaudēšanas. Cilvēkiem epitēlija, saistaudu, muskuļu un kaulu audiem ir diezgan augsta reģenerācijas spēja.
Arī daudzu sugu augi spēj atjaunoties.
Datiem par reģenerāciju ir liela nozīme ne tikai bioloģijā. Tos plaši izmanto lauksaimniecībā, medicīnā, īpaši ķirurģijā.
Vecums kā ontoģenēzes stadija. Vecums ir priekšpēdējais dzīvnieku ontoģenēzes posms, un tiek noteikts tā ilgums
kopējais mūža ilgums, kas kalpo kā sugas raksturojums un atšķiras dažādiem dzīvniekiem. Vecums visprecīzāk ir pētīts cilvēkiem.
Ir dažādas cilvēka vecuma definīcijas. Jo īpaši viena no populārākajām definīcijām ir tāda, ka vecums ir secīgu izmaiņu uzkrāšanās, kas pavada organisma vecuma palielināšanos un palielina tā saslimšanas vai nāves iespējamību. Zinātne par cilvēka novecošanu tiek saukta par gerontoloģiju (no grieķu valodas. gerons - vecs vīrs, vecis, logotipi - zinātne). Tās uzdevums ir izpētīt vecuma pārejas modeļus starp briedumu un nāvi.
Zinātniskie pētījumi gerontoloģijā aptver dažādas jomas, sākot no šūnu enzīmu aktivitātes izmaiņu pētījumiem līdz vides stresa psiholoģisko un socioloģisko mazināšanas ietekmes uz vecu cilvēku uzvedību noskaidrošanai.
Cilvēku gadījumā izšķir fizioloģisku vecumu; vecums, kas saistīts ar kalendāro vecumu; un priekšlaicīga novecošana, ko izraisa sociālie faktori un slimības. Saskaņā ar PVO ieteikumiem par vecāka gadagājuma cilvēku jāuzskata apmēram 60-75 gadus vecs, bet vecs - 75 gadi vai vairāk.
Cilvēka vecumam ir raksturīgas vairākas ārējās un iekšējās pazīmes.
No ārējām vecuma pazīmēm visievērojamākā ir kustību gluduma samazināšanās, stājas izmaiņas, ādas elastības, ķermeņa svara, muskuļu tvirtuma un elastības samazināšanās, grumbu parādīšanās uz sejas un citām ķermeņa daļām. ķermenis un zobu zudums. Tā, piemēram, pēc vispārinātiem datiem, cilvēks 30 gadu vecumā zaudē 2 zobus (zaudējuma rezultātā), 40 gadu vecumā - 4 zobus, 50 gadu vecumā - 8 zobus un 60 gadu vecumā - jau 11 zobi. Pirmajā signalizācijas sistēmā notiek manāmas izmaiņas (maņu orgānu asums ir blāvs). Piemēram, maksimālais attālums, kādā veseli cilvēki atšķir noteiktas identiskas skaņas, ir 12 m 20-30 gadu vecumā, 10 m 50 gadu vecumā, 7 m 60 gadu vecumā un tikai 4 m 70 gadu vecumā. otrā signalizācijas sistēma (runas intonācija mainās, balss kļūst blāvi).
Starp iekšējām pazīmēm, pirmkārt, jāmin tādas pazīmes kā orgānu apgrieztā attīstība (involūcija). Samazinās aknu un nieru izmērs, kā arī to skaits
nefronu skaits nierēs (līdz 80 gadu vecumam gandrīz puse), kas samazina nieru funkcionalitāti un ietekmē ūdens-elektrolītu metabolismu. Samazinās asinsvadu elastība, samazinās audu un orgānu asins perfūzija, palielinās perifēro asinsvadu pretestība. Neorganiskie sāļi uzkrājas kaulos, mainās skrimšļi (pārkaļķojas), samazinās orgānu spēja atjaunoties. Šūnās notiek būtiskas izmaiņas, palēninās dalīšanās un to funkcionālā tonusa atjaunošana, samazinās ūdens saturs, samazinās šūnu enzīmu aktivitāte, tiek traucēta asimilācijas un disimilācijas koordinācija. Smadzenēs tiek traucēta olbaltumvielu sintēze, kā rezultātā veidojas patoloģiski proteīni. Paaugstinās šūnu membrānu viskozitāte, tiek traucēta dzimumhormonu sintēze un utilizācija, notiek izmaiņas neironu struktūrā. Strukturālās izmaiņas notiek saistaudu olbaltumvielās un šo audu elastībā. Imunoloģiskās reakcijas ir novājinātas, un palielinās autoimūnu reakciju iespējamība. Endokrīnās sistēmas, jo īpaši dzimumdziedzeru, funkcijas ir samazinātas. Citu pazīmju uzvedība vecumdienās ir parādīta attēlā. 32.

Rīsi. 32.Dažu cilvēka īpašību izmaiņas ar vecumu: 1 - nervu impulsu ātrums; 2 - bazālā metabolisma līmenis; 3 - sirds indekss; 4 - insulīna nieru filtrācijas līmenis; 5 - plaušu plūdmaiņu apjomi; 6 - plazmas plūsmas līmenis nierēs
Vēlme izprast ķermeņa novecošanās būtību ir bijusi jau ilgu laiku. Senajā Grieķijā Hipokrāts uzskatīja, ka novecošana ir saistīta ar pārmērīgu uzturu un nepietiekamu svaigā gaisa iedarbību. Pēc Aristoteļa domām, novecošanos izraisa ķermeņa patērētā siltumenerģija. Pārtikas kā novecošanas faktora nozīmi atzīmēja arī Galēns. Bet ilgu laiku nebija pietiekami daudz zinātnisku datu, lai objektīvi izprastu šo problēmu. Tikai 19. gs. Novecošanas pētījumos ir gūti zināmi panākumi, un ir sākušas parādīties novecošanas teorijas.
Viena no pirmajām slavenākajām cilvēka ķermeņa novecošanas teorijām ir vācu ārsta H. Hufelanda (1762-1836) teorija, kas ilgmūžībā piešķīra lielu nozīmi darba aktivitātei. Esam dzirdējuši viņa teikto, ka ne viens vien sliņķis nenodzīvoja līdz sirmam vecumam. Vēl slavenāks endokrīnā novecošanās teorija, kas cēlies no eksperimentiem, ko pat pagājušā gadsimta vidū veica Bertolds (1849), kurš parādīja, ka sēklinieku pārstādīšana no viena dzīvnieka uz otru notiek kopā ar sekundāru seksuālo īpašību attīstību. Vēlāk franču fiziologs C. Brown-Séquard (1818-1894), pamatojoties uz rezultātiem, injicējot sev sēklinieku ekstraktus, apgalvoja, ka šīs injekcijas rada labvēlīgu un atjaunojošu efektu. 20. gadsimta sākumā. Jau pastāv uzskats, ka vecuma iestāšanās ir saistīta ar endokrīno dziedzeru, jo īpaši dzimumdziedzeru, darbības izzušanu. 20-30 gados. XX gadsimts Pamatojoties uz šo pārliecību, dažādās valstīs ir veiktas daudzas operācijas, lai atjaunotu vecus vai vecus cilvēkus. Piemēram, G. Šteinahs Austrijā sasēja vīriešu spermatozoīdus, kas noveda pie dzimumdziedzeru ārējās sekrēcijas pārtraukšanas un, domājams, zināmā mērā atjaunojās. S.A. Voronovs Francijā pārstādīja sēkliniekus no jauniem dzīvniekiem veciem un no pērtiķiem vīriešiem, bet Tušnovs PSRS atjaunoja gaiļus, injicējot tiem dzimumdziedzeru histolizātus. Visas šīs darbības radīja zināmas sekas, bet tikai īslaicīgas. Pēc šīm ietekmēm novecošanās process turpinājās un vēl intensīvāk.
20. gadsimta sākumā. radās novecošanās mikrobioloģiskā teorija, kura radītājs bija I.I. Mečņikovs, kurš atšķīra fizioloģisku un patoloģisku vecumu. Viņš uzskatīja, ka cilvēka vecums ir patoloģiskas, t.i. priekšlaicīgi. I.I. ideju pamats Mečņikova ortobiozes doktrīna (ortoss -
pareizi, bios - dzīve), saskaņā ar kuru galvenais novecošanās cēlonis ir nervu šūnu bojājumi ar intoksikācijas produktiem, kas veidojas pūšanas rezultātā resnajā zarnā. Attīstot parastā dzīvesveida doktrīnu (higiēnas noteikumu ievērošana, regulārs darbs, atturēšanās no sliktiem ieradumiem), I.I. Mečņikovs arī ierosināja veidu, kā nomākt pūšanas zarnu baktērijas, patērējot raudzētus piena produktus.
30. gados XX gadsimts kļuva plaši izplatīta teorija par centrālās nervu sistēmas (CNS) lomu novecošanā.Šīs teorijas radītājs ir I.P. Pavlovs, kurš noteica centrālās nervu sistēmas integrējošo lomu organismu normālā funkcionēšanā. I.P. sekotāji. Pavlova eksperimenti ar dzīvniekiem parādīja, ka priekšlaicīgu novecošanos izraisa nervu satricinājumi un ilgstoša nervu pārslodze.
Vērts pieminēt ar vecumu saistītu izmaiņu teorija saistaudos, formulēts 30. gados. XX gadsimts A.A. Bogomoļets (1881-1946). Viņš uzskatīja, ka organisma fizioloģisko aktivitāti nodrošina saistaudi (kaulu audi, skrimšļi, cīpslas, saites un šķiedru saistaudi) un ka izmaiņas šūnu koloidālajā stāvoklī, to turgora zudums u.c. nosaka ar vecumu saistītas izmaiņas. organismos. Mūsdienu dati liecina par kalcija uzkrāšanās nozīmi saistaudos, jo tas veicina tā elastības zudumu, kā arī asinsvadu sacietēšanu.
Mūsdienu pieejas novecošanās būtības un mehānismu izpratnei raksturo fizikāli ķīmiskās bioloģijas datu plaša izmantošana un jo īpaši molekulārās ģenētikas sasniegumi. Visizplatītākās mūsdienu idejas par novecošanās mehānismu ir saistītas ar faktu, ka dzīves laikā ķermeņa šūnās uzkrājas somatiskās mutācijas, kuru rezultātā notiek defektīvu olbaltumvielu sintēze vai nelabotas DNS šķērssaites ar olbaltumvielām. Tā kā bojātiem proteīniem ir dezintegrējoša loma šūnu metabolismā, tas noved pie novecošanās. Ir pierādīts, ka kultivētu fibroblastu gadījumā proteīni un mRNS, kas saistīti ar vecām šūnām, nomāc DNS sintēzi jaunos fibroblastos.
Ir arī zināma hipotēze, saskaņā ar kuru novecošana tiek uzskatīta par mitohondriju metabolītu izmaiņu rezultātu ar sekojošu enzīmu disfunkciju.
Cilvēkiem ir pierādīts gēnu esamība, kas nosaka ar novecošanos saistīto iedzimto deģeneratīvo procesu attīstības laiku. Vairāki pētnieki uzskata, ka novecošanās cēlonis ir izmaiņas organisma imunoloģiskajā aizsardzības sistēmā, jo īpaši autoimūnas reakcijas uz ķermeņa struktūrām, kas ir ļoti svarīgas. Visbeidzot, skaidrojot novecošanās mehānismus, eksperti lielu uzmanību pievērš proteīnu bojājumiem, kas saistīti ar brīvo radikāļu veidošanos. Visbeidzot, dažkārt nozīme tiek piešķirta hidrolāzēm, kas izdalās pēc lizosomu sadalīšanās un kas iznīcina šūnas.
Tomēr visaptveroša novecošanas teorija vēl nav izveidota, jo ir skaidrs, ka neviena no šīm teorijām nevar patstāvīgi izskaidrot novecošanās mehānismus.
Nāve.Nāve ir ontoģenēzes pēdējais posms. Nāves jautājums bioloģijā ieņem īpašu vietu, jo nāves sajūta “... ir cilvēka dabā pilnīgi instinktīva un vienmēr ir bijusi viena no cilvēka lielākajām rūpēm” (I. I. Mečņikovs, 1913). Turklāt nāves jautājums bija un ir visu filozofisko un reliģisko mācību uzmanības centrā, lai gan dažādos vēstures laikos nāves filozofija tika pasniegta atšķirīgi. Senajā pasaulē Sokrats un Platons iestājās par dvēseles nemirstību, savukārt Aristotelis noliedza Platona ideju par dvēseles nemirstību un ticēja cilvēka gara nemirstībai, kas turpina dzīvot pēc cilvēka nāves. Arī Cicerons un Seneka atzina turpmāko dzīvi, bet Markuss Aurēlijs nāvi uzskatīja par dabisku parādību, kas būtu jāpieņem bez sūdzībām. 18. gadsimtā Arī I. Kants un I. Fihte (1762-1814) ticēja nākotnes dzīvei, un G. Hēgelis pieturējās pie pārliecības, ka dvēseli uzsūc “absolūta būtne”, lai gan šīs “būtnes” būtība netika atklāta. .
Saskaņā ar visām zināmajām reliģiskajām mācībām cilvēka zemes dzīve turpinās pēc viņa nāves, un cilvēkam nenogurstoši ir jāgatavojas šai nākotnes nāvei. Tomēr dabaszinātnieki un filozofi, kuri neatzīst nemirstību, ticēja un joprojām uzskata, ka nāve ir, kā vairākkārt uzsvēris I. I.. Mečņikovs, organisma dzīves dabiskais iznākums. Tēlaināka nāves definīcija ir tāda, ka tā “...ir skaidra bezjēdzības uzvara pār jēgu, haosa uzvara pār telpu” (V. Solovjovs, 1894).
Zinātniskie pierādījumi liecina, ka vienšūnu organismos (augos un dzīvniekos) nāve ir jānošķir no izbeigšanās.
zināšanas par to esamību. Nāve ir viņu iznīcināšana, savukārt eksistences pārtraukšana ir saistīta ar to sadalīšanu. Līdz ar to vienšūnu organismu trauslumu kompensē to vairošanās. Daudzšūnu augos un dzīvniekos nāve vārda pilnā nozīmē ir organisma dzīves beigas.
Cilvēkiem pubertātes laikā palielinās nāves iespējamība. Jo īpaši attīstītajās valstīs nāves iespējamība pieaug gandrīz eksponenciāli pēc 28 gadu vecuma.
Ir cilvēka klīniskā un bioloģiskā nāve. Klīniskā nāve izpaužas kā samaņas zudums, sirdsdarbības un elpošanas apstāšanās, bet lielākā daļa šūnu un orgānu joprojām ir dzīvi. Notiek šūnu pašatjaunošanās un turpinās zarnu peristaltika. Klīniskā nāve “nesasniedz” bioloģisko nāvi, jo tā ir atgriezeniska, jo klīniskās nāves stāvoklī var “atgriezties” dzīvē. Piemēram, suņi tiek “atgriezti” dzīvē pēc 5-6 minūtēm, cilvēki - pēc 6-7 minūtēm no klīniskās nāves sākuma. Bioloģisko nāvi raksturo tas, ka tā ir neatgriezeniska. Sirdsdarbības un elpošanas apturēšanu pavada pašatjaunošanās procesu pārtraukšana, šūnu nāve un sadalīšanās. Tomēr šūnu nāve nesākas visos orgānos vienlaikus. Pirmkārt, mirst smadzeņu garoza, pēc tam mirst zarnu, plaušu, aknu, muskuļu šūnu un sirds epitēlija šūnas.
Organismu reanimācijas (atdzīvināšanas) pasākumi ir balstīti uz klīniskās nāves ideju, kas mūsdienu medicīnā ir ārkārtīgi svarīga.
MŪŽS
Salīdzinot datus par floras un faunas pārstāvju paredzamo dzīves ilgumu, redzams, ka starp augiem un dzīvniekiem dažādi organismi dzīvo dažādos laikos. Piemēram, zālaugu augi (savvaļas un kultivēti) dzīvo vienu sezonu. Gluži pretēji, to koksnes augiem ir raksturīgs ilgāks mūžs. Piemēram, ķirsis dzīvo 100 gadus, egle - 1000 gadus, ozols -
2000 gadi, priede - līdz 2000-4000 gadiem.
Daudzu sugu zivis dzīvo 55-80 gadus, vardes - 16 gadus, krokodili - 50-60 gadus, dažu sugu putni - līdz 100 gadiem. Zīdītāju dzīves ilgums ir īsāks.
Piemēram, mazie mājlopi dzīvo 20-25 gadus, liellopi - 30 gadus vai vairāk, zirgi - līdz 30 gadiem, suņi - 20 un vairāk gadus, vilki - 15 gadus, lāči - 50 gadus, ziloņi - 100 gadus. Starp zīdītājiem visilgāk dzīvo cilvēki. Daudzi cilvēki dzīvoja līdz 115-120 gadiem vai vairāk.
Saskaņā ar esošajām idejām paredzamais dzīves ilgums ir sugai raksturīga kvantitatīvā īpašība, ko kontrolē genotips. Tiek uzskatīts, ka ģenētiski noteikts dzīves ilgums korelē ar ontoģenēzes periodiem (5. tabula).
5. tabula.Fibroblastu līmeņa atkarība kultivēšanas laikā no donora vecuma
Tiek pieņemts, ka sugas dzīves ilgums ir sugas evolucionārs ieguvums. Kas attiecas uz indivīdu ilgmūžību, to var izskaidrot ar pieņēmumu par noteiktu gēnu kombināciju klātbūtni to genotipos vai nelielu skaitu vai pilnīgu mutāciju neesamību viņu šūnās.
A.A. Bogomolets un I.I. Šmalhauzens aprēķināja, ka cilvēka dabiskajam mūža ilgumam jābūt 120-150 gadiem. Tomēr tikai daži izdzīvo līdz šim vecumam. Tāpēc faktiskais paredzamais dzīves ilgums nesakrīt ar dabisko dzīves ilgumu.
Vidējo dzīves ilgumu ietekmē vairāki faktori (zīdaiņu mirstības samazināšanās, infekciju kontroles efektivitāte, ķirurģijas sasniegumi, uztura un vispārējo dzīves apstākļu uzlabošanās). Galvenie dzīves ilguma samazināšanās iemesli ir mirstība no bada, slimībām un nepietiekama medicīniskā aprūpe. Krievijā pēdējos gados visās Krievijas pilsētās un laukos vērojams negatīvs iedzīvotāju skaits dzimstības samazināšanās un augstās mirstības dēļ.
Medicīnā paredzamais dzīves ilgums ir nācijas veselības rādītājs. PSRS ieņēma pirmo vietu pasaulē veco cilvēku skaita ziņā. Piemēram, uz 1 miljonu iedzīvotāju ir 104 cilvēki vecumā virs 90 gadiem, savukārt Anglijā - 6, Francijā - 7
un ASV - 15 cilvēki. Šobrīd ir vērojamas izmaiņas strādājošo iedzīvotāju robežās, salīdzinot, piemēram, ar 30. gadiem. XX gadsimts Atšķiras arī atšķirība starp pensionēšanās vecumu un cilvēku aktivitātes līmeni. 1982. gadā Vīnē notika Pasaules Asambleja par pasaules iedzīvotāju novecošanas problēmām, kurā tika formulētas prognozes par šo jautājumu līdz 2025. gadam. Tiek pieņemts, ka 60 gadus vecu un vecāku cilvēku skaits, salīdzinot, pieaugs 5 reizes. līdz 1950. gadam, cilvēki, kas vecāki par 80 gadiem - 7 reizes. Saskaņā ar šo starptautisko forumu dažādu tautu, valstu un reģionu iedzīvotāju novecošanas tempi atšķiras.
Geriatrija ir zinātne, kuras uzdevums ir izstrādāt veidus, kā normalizēt novecojoša organisma mainīgās funkcijas. Tomēr mūsdienu medicīnā vēl nav metožu un līdzekļu, kā ietekmēt normālus fizioloģiskos procesus. Tāpēc geriatrijas loma aprobežojas ar tādu slimību ārstēšanu, kas rodas vecumā, un riska faktoru novēršanu (ja iespējams), kas izraisa priekšlaicīgu novecošanos.
DISKUSIJAS JAUTĀJUMI
1. Ko jūs saprotat ar organismu dzimumvairošanos un kāda ir tās bioloģiskā loma?
2. Aprakstiet bezdzimuma vairošanos un nosauciet tās formas.
3. Raksturojiet dzimumprocesa pazīmes vienšūnu un daudzšūnu organismos.
4. Kas ir gametoģenēze?
5. Kāda ir katra dzimumšūnu veida funkcija?
6. Kādus posmus dzimumšūnu attīstībā jūs zināt?
7. Kādas ir spermatoģenēzes un ooģenēzes līdzības un atšķirības?
8. Kas ir mejoze un kāda ir tās bioloģiskā nozīme?
9. Aprakstiet meiozes fāzes.
10. Vai un kā krustošana ietekmē mejozes rezultātus?
11. Aprakstiet apaugļošanas būtību.
12. Kāda ir atšķirība starp zigoģenēzi un partenoģenēzi?
13. Kāda ir haploīdijas un diploīdijas maiņas bioloģiskā loma?
14. Kas ir seksuālais dimorfisms? Ko tu saproti ar hermafrodītismu? Vai un cik bieži ir novēroti hermafrodītisma gadījumi cilvēkiem?
15. Kā jūs iztēlojaties reprodukcijas metožu attīstību?
16. Ko jūs saprotat ar organismu augšanu un attīstību? Kāda ir saistība starp šūnu augšanu un diferenciāciju?
17. Kādi ir šūnu diferenciācijas molekulārie pamati? Ko jūs zināt par cilmes šūnām?
18. Formulējiet ontoģenēzes jēdzienu un nosauciet ontoģenēzes periodus.
19. Kādas ir atšķirības starp tiešo un netiešo attīstību?
20. Kāda ir apaugļošanas ietekme uz olšūnām?
21. Kurā ģenētiskās informācijas realizācijas stadijā tiek veikta gēnu darbības kontrole?
22. Kā no apaugļotas olšūnas veidojas daudzšūnu struktūra?
23. Kā attīstošās šūnas un audi attīstības laikā atšķiras viens no otra?
24. Vai var atjaunot pazaudētas vai bojātas ķermeņa daļas?
25. Kāda ir saikne ontoģenēzē starp vecumu un paredzamo dzīves ilgumu?
26. Formulējiet un nosakiet atšķirību starp dabisko (iespējamo) un faktisko dzīves ilgumu.
27. Kādas organismu novecošanas teorijas jūs zināt?
29. Kādi faktori ietekmē paredzamo dzīves ilgumu?
30. Uz ko balstās organismu klonēšana? Sniedziet organismu klonēšanas piemērus.
31. Vai sperma vai tās saturs iekļūst šūnā?
32. Par ko liecina klonēšanas dati?
33. Preformacionisms un epiģenēze bioloģijas vēsturē. Vai tiem ir zinātniska nozīme?
34. Kādas ir atšķirības starp klīnisko un bioloģisko nāvi?
Reprodukcija ir organismu spēja veidot savu veidu. Vairošanās ir viena no svarīgākajām dzīvības īpašībām, un tā ir iespējama, pateicoties organismu vispārējai spējai radīt pēcnācējus. Tomēr tiešie pēcnācēji ne vienmēr ir līdzīgi saviem vecākiem. Piemēram, no papardes sporām izaug daudzi pēcnācēji, kurus pārstāv dzinumi, kas nav līdzīgi mātes sporu nesošajam augam. Savukārt uz izauguma parādās augs, kas atšķiras no sevis - sporofīts. Šo parādību sauc par paaudžu maiņu.
Īpaša dzimumvairošanās forma ir partenoģenēze jeb jaunava vairošanās, organisma attīstība no neapaugļotas olšūnas. Šī vairošanās forma raksturīga galvenokārt sugām ar īsu dzīves ciklu ar izteiktām sezonālām izmaiņām. Partenoģenēze var būt haploīda vai diploīda. Haploīdā (ģeneratīvā) partenoģenēzē no haploīdas olšūnas attīstās jauns organisms. Iegūtie indivīdi var būt tikai vīrieši, tikai sievietes vai abi. Tas ir atkarīgs no hromosomu dzimuma noteikšanas.
Organisma individuālā attīstība jeb ontoģenēze ir secīgu morfoloģisko, fizioloģisko un bioķīmisko transformāciju kopums, ko organismā veic no tā rašanās brīža līdz nāvei. Ontoģenēzes laikā tiek īstenota iedzimta informācija, ko ķermenis saņēmis no saviem vecākiem. Ontoģenēzē ir divi galvenie periodi – embrionālais un postembrionālais. Embrionālajā stadijā dzīvniekiem veidojas embrijs, kurā veidojas galvenās orgānu sistēmas. Pēcdzemdību periodā attīstības procesi ir pabeigti, iestājas pubertāte, vairošanās, novecošanās un nāve.
