Pangunahing istraktura - pagkakasunud-sunod ng mga amino acid sa isang polypeptide chain. Sa isang molekula ng protina, kapag ang alternating rigid (peptide bond) at flexible (α-carbon atom) na mga seksyon, isang compact arrangement ng chain sa espasyo ay nabuo.
Paraan ng Akobori ay gamitin phenylhydrazine. Sinisira ng Phenylhydrazine ang mga peptide bond sa protina at nakakabit sa lahat ng amino acid maliban sa C-terminal. Ginagawang posible ng kasunod na pagsusuri ng chromatographic na makilala ang C-terminal amino acid sa protina.
Ang pag-aaral ng pangunahing istraktura ay mahalaga pangkalahatang biyolohikal at medikal na kahalagahan:
- tinutukoy ng pangunahing istraktura ang mga kasunod na istruktura ng protina.
- Ang kaalaman sa pangunahing istraktura ng isang protina ay kinakailangan para sa artipisyal na synthesis ng mga protina.
- Tinutukoy ng pangunahing istraktura ang pagtitiyak ng mga species, halimbawa, sa protina ng insulin, kadalasan sa gitna ng molekula sa iba't ibang uri ng hayop at tao, nangyayari ang isang kapalit, bilang panuntunan, ng 3 katumbas na mga amino acid.
- Ang mga pagbabago sa pangunahing istraktura ay maaaring humantong sa maraming mga sakit, halimbawa, sickle cell anemia, kung saan sa hemoglobin sa β chain sa posisyon 6, ang glutamic acid ay pinalitan ng valine. Ang pagpapalit na ito ng hindi pantay na amino acid ay humahantong sa pagkagambala sa paggana ng hemoglobin at paglitaw ng hugis-karit na mga pulang selula ng dugo.
Pangalawang istraktura - isang regular na paulit-ulit na pattern ng pag-aayos ng isang polypeptide chain sa kalawakan. Kadalasan, ang 2 uri ng pangalawang istraktura ay matatagpuan sa mga protina: α - helix at β - istraktura.
α - helix noong 1951 ito ay pinag-aralan ni L. Pauling gamit ang X-ray diffraction method. Ito ay isang kanang kamay na helical na istraktura, sa isang pagliko kung saan 3.6 amino acid ang magkasya. Ang spiral pitch (ang distansya sa pagitan ng mga katabing pagliko) ay 0.54 n.m. Ang α-helix ay naayos ng mga hydrogen bond, na sarado sa pagitan ng mga peptide bond na nabuo ng bawat ika-4 na amino acid. Ang pangalawang α - istraktura ay kusang natitiklop at tinutukoy ng pangunahing istraktura ng protina. Ang proporsyon ng mga rehiyon na nakaayos sa isang helical na istraktura ay nag-iiba sa iba't ibang mga protina. Halimbawa, sa hemoglobin at myoglobin, ang α - structural fold ay nangingibabaw, na binabawasan ang laki ng molekula ng protina ng 4 na beses.
β-istruktura ay may hugis ng "accordion" at pinatatag ng hydrogen bonds sa pagitan ng malalayong seksyon ng isang polypeptide chain o sa pagitan ng ilang molekula ng protina. May mga parallel na istruktura ng β, kung saan ang mga dulo ng N at C ay tumutugma sa bawat isa, at mga antiparallel na istruktura. Ang isang halimbawa ng mga protina na nakararami ay naglalaman ng mga istrukturang β ay mga immunoglobulin.
Ang pangalawang istraktura ay pinag-aralan sa pamamagitan ng X-ray diffraction analysis at sa pamamagitan ng pag-aaral ng pagsipsip ng ultraviolet rays ng protina (mas malaki ang proporsyon ng α - structures, mas malaki ang pagsipsip).
Ang pangalawang istraktura ay nawasak sa panahon ng denaturation.
Tertiary structure - may isang anyo ng spatial arrangement ng polypeptide chain na tiyak para sa bawat protina. Ang istrukturang ito ay kusang nabuo at tinutukoy ng pangunahing istraktura. Ang tertiary na istraktura ay makabuluhang pinatataas ang compactness ng protina sa pamamagitan ng sampu. Ang mga non-covalent bond (hydrophobic, ionic) at covalent (disulfide) na mga bono ay kasangkot sa pagbuo ng tersiyaryong istraktura.
Tinutukoy ng tertiary structure ang biological activity at physicochemical properties ng mga protina. Kung ang tertiary na istraktura ay nagambala, ang protina ay nawawala ang biological na aktibidad nito.
Ang mga pamamaraan para sa pag-aaral ng tertiary structure ay X-ray diffraction analysis at pagpapasiya ng kemikal na aktibidad ng mga indibidwal na amino acid radical sa isang protina. Ang tertiary structure ng myoglobin protein ay unang pinag-aralan ni J. Kendrew (1957). Pinag-aralan ni M. Perutz (1959) ang istruktura ng hemoglobin.
Ang tersiyaryong istraktura ng mga protina ay kinabibilangan ng α - helical, β - nakatiklop na mga istraktura, β - mga loop (kung saan ang polypeptide chain ay baluktot ng 180 0) at ang tinatawag na disordered coil. Halimbawa, ang protina ng insulin ay naglalaman ng 57% α - helical regions, 6% β - folded structures, 10% ng molecule ay nakaayos sa anyo ng β - loops at 27% ng molecule ay isang disordered coil.
Ang kabuuan ng pangunahin, pangalawa, tersiyaryo ay pag-aayos molekula ng protina. Ang lifetime (katutubong) conformation ay kusang nabuo at ang pagbuo nito ay tinatawag na folding. Ang conformation ng mga protina ay napaka hindi matatag at nabuo sa pakikilahok ng mga espesyal na protina - mga chaperone(mga kasama). Nagagawa ng mga chaperone na magbigkis sa mga bahagyang denatured na protina na nasa isang hindi matatag na estado at ibalik ang kanilang katutubong conform. Ang mga chaperone ay inuri ayon sa molekular na timbang (60 - 100 cd.). Ang pinaka-pinag-aralan ay Sh-60, Sh-70 at Sh-90. Halimbawa, nakikipag-ugnayan ang Sh-70 sa mga protina na mayaman sa mga hydrophobic radical at pinoprotektahan ang mga ito mula sa denaturation ng mataas na temperatura. Sa pangkalahatan, pinangangalagaan ng mga chaperone ang mga pangunahing protina ng katawan, pinipigilan ang denaturation at itaguyod ang pagbuo ng conformation, pinapadali ang transportasyon ng mga denatured na protina sa mga lysosome, at nakikilahok sa proseso ng synthesis ng protina.
Ayon sa conformation, ang lahat ng mga protina ay nahahati sa tatlong grupo:
- mga protina ng fibrillar: collagen, elastin, fibroin.
- Mga globular na protina: hemoglobin, albumin, globulin.
- Mga pinaghalong protina: myosin.
Ang tersiyaryong istraktura ay likas sa lahat ng mga protina. Ang mga oligomeric na protina lamang, na naglalaman ng ilang mga subunit, mga protomer, ay may istrukturang quaternary. Ang isang protomer ay itinuturing na isang hiwalay na polypeptide chain, ang isang subunit ay ang aktibong bahagi ng isang oligomeric na protina. Ang isang subunit ay maaaring maglaman ng alinman sa isang protomer o ilang.
Quaternary structure - ang bilang at kamag-anak na pag-aayos ng mga subunit sa mga oligomeric na protina. Ang mga oligomeric na protina lamang, na naglalaman ng ilang mga subunit, mga protomer, ay may istrukturang quaternary. Ang isang protomer ay itinuturing na isang hiwalay na polypeptide chain, ang isang subunit ay ang aktibong bahagi ng isang oligomeric na protina. Ang isang subunit ay maaaring maglaman ng isang protomer o maraming protomer.
Ang pagbuo ng quaternary na istraktura ay nagsasangkot ng mahina na non-covalent bond (hydrophobic, ionic, hydrogen). Ang quaternary na istraktura ng mga protina ay kusang nabubuo at madaling masira kapag na-denatured. Ang mga indibidwal na subunit sa isang oligomeric na protina ay nakikipag-ugnayan sa isa't isa, na humahantong sa mga pagbabago sa tertiary na istraktura ng mga indibidwal na protomer. Ang hindi pangkaraniwang bagay na ito ay tinatawag na mga pagbabago sa kooperatiba sa protomer conformation at kadalasang sinasamahan ng pagtaas ng aktibidad ng protina.
Ang mga oligomeric na protina ay may ilang mga tampok kumpara sa mga monomeric na protina.
- Mayroon silang isang napaka-compact na pag-iimpake at isang medyo maliit na ibabaw ng interface, samakatuwid, na matatagpuan sa intracellularly, sila ay nagbubuklod ng mas kaunting tubig
- Ang kanilang aktibidad ay kinokontrol sa katawan. Ang mga protomer ay karaniwang hindi aktibo, ngunit ang mga oligomeric na protina ay mas aktibo.
- Kung ang parehong uri ng mga protomer ay kasangkot sa synthesis ng isang oligomeric na protina, ito ay nagse-save ng genetic material (ilang magkaparehong protomer ay "nakatatak" sa isang maikling seksyon ng DNA)
- Ang mga ito ay functionally mas inangkop sa mga kondisyon ng katawan.
Ang pag-andar ng mga oligomeric na protina ay inilalarawan sa pamamagitan ng paghahambing ng mga protina na hemoglobin at myoglobin, na kasangkot sa transportasyon ng oxygen sa mga tisyu. Ang Hemoglobin ng mga erythrocytes ay isang oligomeric na protina na kinabibilangan ng 4 na polypeptide chain. Ang muscle myoglobin ay isang monomeric na protina na may kasamang 1 polypeptide chain. Ang oxygen saturation curve ng myoglobin ay nagpapahiwatig ng direktang pag-asa nito sa konsentrasyon ng oxygen. Para sa hemoglobin, ang oxygen saturation curve ay S-shaped. Ito ay dahil sa isang unti-unting sunud-sunod na pagbabago sa istraktura (conformation) ng bawat isa sa 4 na protomer sa komposisyon ng hemoglobin, bilang isang resulta kung saan ang affinity ng hemoglobin para sa oxygen ay tumataas nang husto. Ang likas na katangian ng saturation ng hemoglobin na may oxygen ay matalas na pinatataas ang kapasidad ng oxygen nito kumpara sa myoglobin.
Ang isang espesyal na posisyon sa mga protina ay inookupahan ng mga protina ng domain .
Ang mga domain ay hiwalay sa istruktura at functional na mga seksyon ng isang polypeptide chain. Ang mga domain ay maaaring maging responsable para sa pakikipag-ugnayan ng isang protina sa iba't ibang mga sangkap - mga ligand (mga mababang molekular na sangkap, DNA, RNA, polysaccharides, atbp.).
Dahil sa mataas na selectivity ng mga protina, maaari silang pagsamahin sa mga complex, na kung saan ay madalas na tinatawag na multienzyme complex - ito ay mga istrukturang asosasyon ng ilang mga enzyme na nagpapagana ng mga indibidwal na yugto ng isang kumplikadong proseso ng kemikal. Halimbawa: pyruvate dehydrogenase complex (PDC), isang complex ng tatlong uri ng enzymes na nagpapagana sa oksihenasyon ng pyruvic acid (PVA).
Posible na partikular na pagsamahin hindi lamang ang mga indibidwal na protina, kundi pati na rin ang mga protina na may mga lipid (taba) sa panahon ng pagbuo ng mga lamad ng cell, at mga protina na may mga nucleic acid sa panahon ng pagbuo ng chromatin.
Physicochemical properties ng mga protina.
Ang mga ito ay higit na tinutukoy ng conformation ng molekula ng protina (pangunahing - tertiary na istraktura ng protina). Ang mga katangian ng physicochemical ng mga protina ay lumilitaw sa mga solusyon.
Solubility ang mga protina ay nag-iiba mula sa protina hanggang protina.
Sa pangkalahatan, ang solubility ng mga protina ay mataas, ngunit nag-iiba-iba sa iba't ibang uri ng mga protina. Ito ay naiimpluwensyahan ng mga sumusunod na kadahilanan:
- hugis ng molekula ng protina (ang mga globular na protina ay mas natutunaw kaysa sa mga protina ng fibrillar)
- ang likas na katangian ng protina amino acid radical, ang ratio ng mga polar non-polar radical (mas maraming polar hydrophilic radical sa protina, mas mahusay ang solubility nito)
- mga katangian ng solvent, pagkakaroon ng mga asing-gamot. Ang isang mababang konsentrasyon ng mga asin (KCL, NaCl) kung minsan ay nagpapataas ng solubility ng mga protina. Halimbawa, ang mga albumin ay mas mahusay na natutunaw sa purong distilled water, ang mga globulin ay natutunaw lamang sa pagkakaroon ng 10% na mga asing-gamot (KCL, NaCl). Ang connective tissue proteins na collagen at elastin ay hindi matutunaw sa tubig o asin na solusyon.
Molecular mass Ang mga protina ay medyo malaki, mula 6,000 hanggang 1,000,000.
Mga pamamaraan para sa pagtukoy ng molar mass ng mga protina dapat maging banayad at hindi sirain ang mga molekula ng protina. Halimbawa, ang ebullioscopic na paraan, batay sa pagsukat ng boiling point ng mga solusyon, ay hindi naaangkop sa mga protina. Ang pinakatumpak na pamamaraan para sa pagtukoy ng molekular na timbang ng mga protina ay ang ultracentrifugation na paraan at ang X-ray diffraction na paraan.
Paraan ng ultracentrifugation(sedimentation) ay batay sa isang pagbabago sa rate ng sedimentation ng mga protina ng iba't ibang molecular weight kapag umiikot ang mga solusyon sa protina sa mataas na bilis. Ang molekular na timbang ng mga protina na natagpuan sa paraang ito ay itinalaga ng Svedberg unit (S = 10 -13 c.)
Paraan ng X-ray diffraction nagbibigay-daan sa iyo na kalkulahin ang molecular weight sa pamamagitan ng pagsusuri ng maramihang X-ray na imahe ng isang molekula ng protina.
Paraan ng electrophoretic ay batay sa pag-asa ng bilis ng paggalaw ng protina sa isang pare-parehong electric field sa molekular na timbang ng protina (electrophoric mobility ay mas mataas para sa mga protina na may mas mababang molekular na timbang)
Chromatographic na pamamaraan ay batay sa iba't ibang mga rate ng pagpasa ng iba't ibang mga protina sa pamamagitan ng molecular gel "sieves".
Ang mga malalaking molekula na mas malaki kaysa sa laki ng butas ng gel ay dumadaan sa gel nang mas mabilis kaysa sa mas maliliit na molekula ng protina na nananatili sa loob ng mga butil ng gel.
Paraan ng mikroskopiko ng elektron Isinasagawa sa pamamagitan ng paghahambing ng laki ng isang molekula ng protina sa mga sample ng sanggunian ng kilalang masa.
Mga pamamaraan ng kemikal nauugnay sa mga katangian ng kemikal na komposisyon ng mga protina
Hugis ng mga molekula ng protina magkaiba. Ang mga molekula ng protina ay maaaring fibrillar o globular na hugis. Ang mga fibrillar na protina ay may hugis-thread na molekular na hugis. Ang mga ito ay karaniwang hindi matutunaw sa tubig at maghalo ng mga solusyon sa asin. Kasama sa mga protina ng fibrillar ang pangunahing istrukturang protina ng nag-uugnay na tisyu: collagen, keratin, elastin. Sa mga globular na protina, ang mga polypeptide chain ay mahigpit na nakapulupot sa mga compact na spherical na istruktura. Karamihan sa mga globular na protina ay lubos na natutunaw sa tubig at mahinang mga solusyon sa asin. Ang mga globular na protina ay kinabibilangan ng mga enzyme, antibodies, albumin, at hemoglobin. Ang ilang mga protina ay may isang intermediate na uri ng molekula, na naglalaman ng parehong thread-like at spherical na mga rehiyon. Ang isang halimbawa ng naturang mga protina ay ang myosin ng protina ng kalamnan, na natutunaw sa mga solusyon sa asin.
Mga sukat ng mga molekula ng protina ay nasa hanay mula 1 hanggang 100 nm, malapit sa laki ng mga colloidal particle. Dahil dito, ang mga solusyon sa protina ay may mga katangian ng parehong tunay na solusyon at koloidal na solusyon.
Maraming mga molecular kinetic na katangian ng mga solusyon sa protina ay katulad ng mga katangian ng mga colloidal solution .
- Ang mabagal na rate ng pagsasabog ng mga protina na kinakailangan para sa kanilang pagpapalitan.
- Kawalan ng kakayahan ng mga protina na dumaan sa mga semipermeable na lamad. Sa mga compartment na may mataas na konsentrasyon ng protina, ang labis na hydrostatic pressure ay nalikha dahil sa one-way na paggalaw ng mga molekula ng tubig sa pamamagitan ng semi-permeable membrane patungo sa mataas na konsentrasyon ng protina. Ang labis na presyon na nilikha ng mga protina ay tinatawag na oncotic pressure. Ito ay isang mahalagang kadahilanan na tumutukoy sa paggalaw ng tubig sa pagitan ng mga tisyu, dugo, at mga bituka.
- Ang mataas na lagkit ng mga protina ay dahil sa iba't ibang intermolecular na pakikipag-ugnayan ng malalaking molekula ng protina. Ang pagtaas ng lagkit ng dugo, sa partikular, ay nagpapataas ng pagkarga sa kalamnan ng puso.
- Ang ilang mga protina ay nagagawang bumuo ng mga gel, na nagpapataas ng lakas ng mga protina (hal. collagen).
Mga optical na katangian ng mga protina tinutukoy ng laki ng mga molekula ng protina, ang istraktura ng mga radikal na amino acid sa mga protina, ang pagkakaroon ng mga peptide bond at mga alpha-helical na rehiyon sa mga protina.
- Ang mga solusyon sa protina ay mayroon ang epekto ng light refraction (refraction) at light scattering. Ang mga katangiang ito ay dahil sa malaking sukat ng mga molekula ng protina, na katumbas ng haba ng daluyong ng nakikitang bahagi ng spectrum. Ang antas ng repraksyon ay proporsyonal sa konsentrasyon ng solusyon sa protina.
- Mga solusyon sa protina sumipsip ng mga sinag ng ultraviolet sa hanay ng 190-230 nm dahil sa pagkakaroon ng mga peptide bond at sa hanay ng 260-280 nm dahil sa pagkakaroon ng cyclic amino acid sa mga protina. Ang antas ng pagsipsip ng UV ay proporsyonal sa konsentrasyon ng protina sa solusyon.
- Ang mga solusyon sa protina ay maaari paikutin ang eroplano ng polarized light, na dahil sa optical na aktibidad ng mga amino acid na nakapaloob sa protina at ang pagkakaroon ng mga alpha-helical na seksyon sa loob nito. Mayroong direktang kaugnayan sa pagitan ng polariseysyon ng liwanag at ang konsentrasyon ng mga protina sa solusyon.
Ang mga protina, bilang mga molekular na solusyon, ay mayroon katangian ng mga tunay na solusyon . Bilang mga tunay na solusyon, ang mga solusyon sa protina ay lubos na matatag.
MODULE 1 STRUCTURE, PROPERTY AND FUNCTIONS NG PROTEINS
MODULE 1 STRUCTURE, PROPERTY AND FUNCTIONS NG PROTEINS
Istraktura ng module | Mga tema |
Modular na yunit 1 | 1.1. Structural na organisasyon ng mga protina. Mga yugto ng pagbuo ng katutubong conformation ng protina 1.2. Mga pangunahing kaalaman sa paggana ng protina. Mga gamot bilang ligand na nakakaapekto sa paggana ng protina 1.3. Denaturasyon ng mga protina at ang posibilidad ng kanilang kusang pagbabagong-buhay |
Modular na yunit 2 | 1.4. Mga tampok ng istraktura at paggana ng mga oligomeric na protina gamit ang halimbawa ng hemoglobin 1.5. Pagpapanatili ng katutubong conformation ng protina sa ilalim ng mga kondisyon ng cellular 1.6. Iba't ibang mga protina. Mga pamilya ng protina gamit ang halimbawa ng mga immunoglobulin 1.7. Physicochemical properties ng mga protina at mga pamamaraan para sa kanilang paghihiwalay |
Modular unit 1 STRUCTURAL ORGANIZATION NG MONOMERIC PROTEINS AT ANG MGA BASES NG KANILANG PAGGAMIT
Mga layunin sa pagkatuto Magagawang:
1. Gumamit ng kaalaman tungkol sa mga tampok na istruktura ng mga protina at ang pag-asa ng mga function ng protina sa kanilang istraktura upang maunawaan ang mga mekanismo ng pag-unlad ng namamana at nakuha na mga proteinopathies.
2. Ipaliwanag ang mga mekanismo ng therapeutic action ng ilang mga gamot bilang mga ligand na nakikipag-ugnayan sa mga protina at nagbabago sa kanilang aktibidad.
3. Gumamit ng kaalaman tungkol sa istraktura at conformational lability ng mga protina upang maunawaan ang kanilang structural at functional na kawalang-tatag at pagkahilig sa denaturation sa ilalim ng pagbabago ng mga kondisyon.
4. Ipaliwanag ang paggamit ng mga denaturing agent bilang paraan para sa pag-sterilize ng mga medikal na materyales at instrumento, pati na rin bilang antiseptics.
alamin:
1. Mga antas ng istrukturang organisasyon ng mga protina.
2. Ang kahalagahan ng pangunahing istraktura ng mga protina, na tumutukoy sa kanilang pagkakaiba-iba sa istruktura at functional.
3. Ang mekanismo ng pagbuo ng aktibong sentro sa mga protina at ang tiyak na pakikipag-ugnayan nito sa ligand, na sumasailalim sa paggana ng mga protina.
4. Mga halimbawa ng impluwensya ng mga exogenous ligand (mga gamot, lason, lason) sa conformation at functional na aktibidad ng mga protina.
5. Mga sanhi at bunga ng denaturation ng protina, mga salik na nagdudulot ng denaturation.
6. Mga halimbawa ng paggamit ng mga denaturing factor sa gamot bilang antiseptics at paraan para sa pag-sterilize ng mga medikal na instrumento.
PAKSA 1.1. STRUCTURAL ORGANIZATION NG PROTEINS. MGA YUGTO NG KATUTUBONG PAGBUO
MGA KONFORMASYON NG PROTEIN
Ang mga protina ay mga molekulang polimer na ang mga monomer ay 20 α-amino acid lamang. Ang hanay at pagkakasunud-sunod ng kumbinasyon ng mga amino acid sa isang protina ay tinutukoy ng istruktura ng mga gene sa DNA ng mga indibidwal. Ang bawat protina, alinsunod sa tiyak na istraktura nito, ay gumaganap ng sarili nitong function. Ang hanay ng mga protina ng isang naibigay na organismo ay tumutukoy sa mga phenotypic na katangian nito, pati na rin ang pagkakaroon ng mga namamana na sakit o isang predisposisyon sa kanilang pag-unlad.
1. Amino acids na bumubuo sa mga protina. Peptide bond. Ang mga protina ay mga polimer na binuo mula sa mga monomer - 20 α-amino acid, ang pangkalahatang pormula nito ay
Ang mga amino acid ay naiiba sa istraktura, sukat, at physicochemical na katangian ng mga radical na nakakabit sa α-carbon atom. Tinutukoy ng mga functional na grupo ng mga amino acid ang mga katangian ng mga katangian ng iba't ibang mga α-amino acid. Ang mga radical na matatagpuan sa α-amino acids ay maaaring nahahati sa ilang grupo:
proline, Hindi tulad ng iba pang 19 na monomer ng protina, ito ay hindi isang amino acid, ngunit isang imino acid na nauugnay sa parehong α-carbon atom at ang imino group;
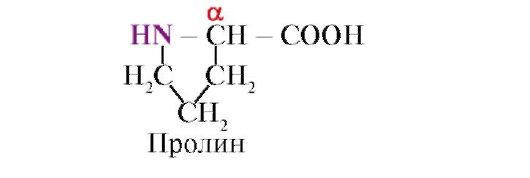 Ang mga amino acid ay nag-iiba sa solubility sa tubig. Ito ay dahil sa kakayahan ng mga radical na makipag-ugnayan sa tubig (hydrate).
Ang mga amino acid ay nag-iiba sa solubility sa tubig. Ito ay dahil sa kakayahan ng mga radical na makipag-ugnayan sa tubig (hydrate).
SA hydrophilic isama ang mga radical na naglalaman ng anionic, cationic at polar uncharged functional group.
SA hydrophobic isama ang mga radical na naglalaman ng mga methyl group, aliphatic chain o ring.
2. Ang mga peptide bond ay nag-uugnay sa mga amino acid upang bumuo ng mga peptide. Sa panahon ng peptide synthesis, ang α-carboxyl group ng isang amino acid ay nakikipag-ugnayan sa α-amino group ng isa pang amino acid upang mabuo peptide bond:
Ang mga protina ay polypeptides, i.e. linear polymers ng α-amino acids na konektado ng isang peptide bond (Fig. 1.1.)
 kanin. 1.1. Mga terminong ginamit upang ilarawan ang istruktura ng mga peptide
kanin. 1.1. Mga terminong ginamit upang ilarawan ang istruktura ng mga peptide
Ang mga monomer ng mga amino acid na bumubuo sa polypeptides ay tinatawag mga residu ng amino acid. Isang hanay ng mga paulit-ulit na grupo - NH-CH-CO- mga form peptide backbone. Ang residue ng amino acid na may libreng α-amino group ay tinatawag na N-terminal, at ang isa na may libreng α-carboxyl group ay tinatawag na C-terminal. Ang mga peptide ay isinusulat at binabasa mula sa N-terminus hanggang C-terminus.
Ang peptide bond na nabuo ng imino group ng proline ay naiiba sa iba pang peptide bond: ang nitrogen atom ng peptide group ay walang hydrogen,
sa halip, mayroong isang bono na may isang radikal, bilang isang resulta kung saan ang isang bahagi ng singsing ay kasama sa peptide backbone:
 Ang mga peptide ay naiiba sa komposisyon ng amino acid, bilang ng mga amino acid at pagkakasunud-sunod ng koneksyon ng amino acid, halimbawa, ang Ser-Ala-Glu-Gis at His-Glu-Ala-Ser ay dalawang magkaibang peptides.
Ang mga peptide ay naiiba sa komposisyon ng amino acid, bilang ng mga amino acid at pagkakasunud-sunod ng koneksyon ng amino acid, halimbawa, ang Ser-Ala-Glu-Gis at His-Glu-Ala-Ser ay dalawang magkaibang peptides.
Ang mga bono ng peptide ay napakalakas, at ang kanilang kemikal na non-enzymatic hydrolysis ay nangangailangan ng malupit na mga kondisyon: ang protina na sinusuri ay hydrolyzed sa concentrated hydrochloric acid sa temperatura na humigit-kumulang 110° sa loob ng 24 na oras. Sa isang buhay na cell, ang mga peptide bond ay maaaring masira ng proteolytic enzymes, tinawag mga protease o peptide hydrolases.
3. Pangunahing istraktura ng mga protina. Ang mga residue ng amino acid sa mga peptide chain ng iba't ibang mga protina ay hindi pumapalit nang random, ngunit nakaayos sa isang tiyak na pagkakasunud-sunod. Ang linear sequence o alternation order ng mga residue ng amino acid sa isang polypeptide chain ay tinatawag pangunahing istraktura ng protina.
Ang pangunahing istraktura ng bawat indibidwal na protina ay naka-encode sa molekula ng DNA (sa isang rehiyon na tinatawag na gene) at natanto sa panahon ng transkripsyon (pagkopya ng impormasyon sa mRNA) at pagsasalin (synthesis ng pangunahing istraktura ng protina). Dahil dito, ang pangunahing istraktura ng mga protina ng isang indibidwal na tao ay ang impormasyong namamana na ipinadala mula sa mga magulang hanggang sa mga bata, na tumutukoy sa mga tampok na istruktura ng mga protina ng isang naibigay na organismo, kung saan nakasalalay ang pag-andar ng umiiral na mga protina (Fig. 1.2.).
 kanin. 1.2. Ang relasyon sa pagitan ng genotype at ang conformation ng mga protina na na-synthesize sa katawan ng indibidwal
kanin. 1.2. Ang relasyon sa pagitan ng genotype at ang conformation ng mga protina na na-synthesize sa katawan ng indibidwal
Ang bawat isa sa humigit-kumulang 100,000 indibidwal na protina sa katawan ng tao ay mayroon kakaiba pangunahing istraktura. Ang mga molekula ng parehong uri ng protina (halimbawa, albumin) ay may parehong paghalili ng mga residue ng amino acid, na nagpapakilala sa albumin mula sa anumang iba pang indibidwal na protina.
Ang pagkakasunud-sunod ng mga residue ng amino acid sa isang peptide chain ay maaaring ituring na isang paraan ng pagtatala ng impormasyon. Tinutukoy ng impormasyong ito ang spatial arrangement ng linear peptide chain sa isang mas compact na three-dimensional na istraktura na tinatawag na pag-aayos ardilya. Ang proseso ng pagbuo ng isang functionally active protein conformation ay tinatawag natitiklop
4. Conformation ng protina. Ang libreng pag-ikot sa peptide backbone ay posible sa pagitan ng nitrogen atom ng peptide group at ng kalapit na α-carbon atom, gayundin sa pagitan ng α-carbon atom at ng carbon ng carbonyl group. Dahil sa pakikipag-ugnayan ng mga functional na grupo ng mga residue ng amino acid, ang pangunahing istraktura ng mga protina ay maaaring makakuha ng mas kumplikadong spatial na istruktura. Sa mga globular na protina, mayroong dalawang pangunahing antas ng pagtitiklop ng conformation ng mga peptide chain: pangalawa At tersiyaryong istraktura.
Pangalawang istraktura ng mga protina ay isang spatial na istraktura na nabuo bilang isang resulta ng pagbuo ng mga bono ng hydrogen sa pagitan ng mga functional na grupo -C=O at -NH- ng peptide backbone. Sa kasong ito, ang peptide chain ay maaaring makakuha ng mga regular na istruktura ng dalawang uri: α-helices At β-istruktura.
SA α-helices ang mga bono ng hydrogen ay nabuo sa pagitan ng oxygen atom ng carbonyl group at ng hydrogen ng amide nitrogen ng ika-4 na amino acid; side chain ng mga residue ng amino acid
ay matatagpuan sa kahabaan ng periphery ng spiral, nang hindi nakikilahok sa pagbuo ng pangalawang istraktura (Larawan 1.3.).
Ang mga bulk radical, o mga radical na may pantay na singil, ay pumipigil sa pagbuo ng isang α-helix. Ang proline residue, na may istraktura ng singsing, ay nakakagambala sa α-helix, dahil dahil sa kakulangan ng hydrogen sa nitrogen atom sa peptide chain, imposibleng bumuo ng hydrogen bond. Ang bono sa pagitan ng nitrogen at ng α-carbon atom ay bahagi ng proline ring, kaya ang peptide backbone ay nagiging baluktot sa puntong ito.
β-Istruktura ay nabuo sa pagitan ng mga linear na rehiyon ng peptide backbone ng isang polypeptide chain, sa gayon ay bumubuo ng mga nakatiklop na istruktura. Maaaring mabuo ang mga polypeptide chain o mga bahagi nito parallel o mga antiparallel β-structure. Sa unang kaso, ang N- at C-termini ng mga nakikipag-ugnay na peptide chain ay nag-tutugma, at sa pangalawa ay mayroon silang kabaligtaran na direksyon (Larawan 1.4).
kanin. 1.3. Pangalawang istraktura ng protina - α-helix
 kanin. 1.4. Parallel at antiparallel β-sheet structures
kanin. 1.4. Parallel at antiparallel β-sheet structures
Ang mga β-structure ay ipinahiwatig ng malalawak na arrow: A - Antiparallel β-structure. B - Parallel β-sheet structures
Sa ilang mga protina, ang mga istruktura ng β ay maaaring mabuo dahil sa pagbuo ng mga bono ng hydrogen sa pagitan ng mga atomo ng peptide backbone ng iba't ibang mga polypeptide chain.
Matatagpuan din sa mga protina mga lugar na may iregular na sekondarya istraktura, na kinabibilangan ng mga liko, mga loop, at mga pagliko ng polypeptide backbone. Madalas silang matatagpuan sa mga lugar kung saan nagbabago ang direksyon ng chain ng peptide, halimbawa, kapag nabuo ang isang parallel na istraktura ng β-sheet.
Batay sa pagkakaroon ng α-helice at β-structure, ang mga globular na protina ay maaaring nahahati sa apat na kategorya.
kanin. 1.5. Pangalawang istraktura ng myoglobin (A) at hemoglobin β-chain (B), na naglalaman ng walong α-helices

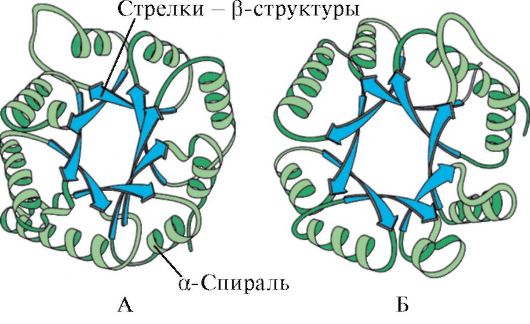 kanin. 1.6. Pangalawang istraktura ng triosephosphate isomerase at pyruvate kinase domain
kanin. 1.6. Pangalawang istraktura ng triosephosphate isomerase at pyruvate kinase domain
 kanin. 1.7. Pangalawang istraktura ng patuloy na domain ng immunoglobulin (A) at ang enzyme superoxide dismutase (B)
kanin. 1.7. Pangalawang istraktura ng patuloy na domain ng immunoglobulin (A) at ang enzyme superoxide dismutase (B)
SA ikaapat na kategorya kasama ang mga protina na naglalaman ng isang maliit na halaga ng mga regular na pangalawang istruktura. Kasama sa mga protina na ito ang maliliit na protinang mayaman sa cysteine o metalloprotein.
Ang istraktura ng tersiyaryo ng protina- isang uri ng conformation na nabuo dahil sa mga pakikipag-ugnayan sa pagitan ng mga amino acid radical, na maaaring matatagpuan sa isang malaking distansya mula sa bawat isa sa peptide chain. Karamihan sa mga protina ay bumubuo ng isang spatial na istraktura na kahawig ng isang globule (globular na protina).
Dahil ang hydrophobic amino acid radicals ay may posibilidad na pagsamahin sa pamamagitan ng tinatawag na hydrophobic na pakikipag-ugnayan at intermolecular van der Waals pwersa, isang siksik na hydrophobic core ay nabuo sa loob ng protina globule. Ang hydrophilic ionized at non-ionized radical ay pangunahing matatagpuan sa ibabaw ng protina at tinutukoy ang solubility nito sa tubig.
 kanin. 1.8. Mga uri ng mga bono na lumitaw sa pagitan ng mga radikal na amino acid sa panahon ng pagbuo ng tersiyaryong istraktura ng isang protina
kanin. 1.8. Mga uri ng mga bono na lumitaw sa pagitan ng mga radikal na amino acid sa panahon ng pagbuo ng tersiyaryong istraktura ng isang protina
1 - ionic bond- nangyayari sa pagitan ng positibo at negatibong sisingilin na mga functional na grupo;
2 - hydrogen bond- nangyayari sa pagitan ng hydrophilic uncharged group at anumang iba pang hydrophilic group;
3 - hydrophobic na pakikipag-ugnayan- bumangon sa pagitan ng mga hydrophobic radical;
4 - disulfide bond- nabuo dahil sa oksihenasyon ng mga pangkat ng SH ng mga labi ng cysteine at ang kanilang pakikipag-ugnayan sa bawat isa
Ang hydrophilic amino acid residues na matatagpuan sa loob ng hydrophobic core ay maaaring makipag-ugnayan sa isa't isa gamit ionic At hydrogen bonds(Larawan 1.8).
Ang mga bono ng ionic at hydrogen, pati na rin ang mga hydrophobic na pakikipag-ugnayan, ay mahina: ang kanilang enerhiya ay hindi mas mataas kaysa sa enerhiya ng thermal motion ng mga molekula sa temperatura ng silid. Ang conformation ng protina ay pinananatili sa pamamagitan ng pagbuo ng maraming tulad ng mahinang mga bono. Dahil ang mga atomo na bumubuo sa isang protina ay patuloy na gumagalaw, posibleng masira ang ilang mahinang mga bono at bumuo ng iba, na humahantong sa bahagyang paggalaw ng mga indibidwal na seksyon ng polypeptide chain. Ang pag-aari na ito ng mga protina upang baguhin ang conformation bilang resulta ng pagsira ng ilan at pagbuo ng iba pang mahinang mga bono ay tinatawag conformational lability.
Ang katawan ng tao ay may mga sistemang sumusuporta homeostasis- katatagan ng panloob na kapaligiran sa loob ng ilang katanggap-tanggap na mga limitasyon para sa isang malusog na katawan. Sa ilalim ng mga kondisyon ng homeostasis, ang mga maliliit na pagbabago sa conformation ay hindi nakakagambala sa pangkalahatang istraktura at pag-andar ng mga protina. Ang functionally active conformation ng isang protina ay tinatawag katutubong anyo. Ang mga pagbabago sa panloob na kapaligiran (halimbawa, ang konsentrasyon ng glucose, Ca ions, protons, atbp.) ay humantong sa mga pagbabago sa conformation at pagkagambala ng mga function ng protina.
Ang tertiary na istraktura ng ilang mga protina ay nagpapatatag disulfide bond, nabuo dahil sa pakikipag-ugnayan ng -SH na mga grupo ng dalawang nalalabi
 kanin. 1.9. Ang pagbuo ng isang disulfide bond sa isang molekula ng protina
kanin. 1.9. Ang pagbuo ng isang disulfide bond sa isang molekula ng protina
cysteine (Larawan 1.9). Karamihan sa mga intracellular na protina ay walang mga covalent disulfide bond sa kanilang tertiary structure. Ang kanilang presensya ay katangian ng mga protina na itinago ng cell, na nagsisiguro sa kanilang higit na katatagan sa mga kondisyon ng extracellular. Kaya, ang mga disulfide bond ay naroroon sa mga molekula ng insulin at immunoglobulin.
Insulin- isang protina na hormone na na-synthesize sa β-cells ng pancreas at itinago sa dugo bilang tugon sa pagtaas ng konsentrasyon ng glucose sa dugo. Sa istruktura ng insulin, mayroong dalawang disulfide bond na nagkokonekta sa polypeptide A at B chain, at isang disulfide bond sa loob ng A chain (Fig. 1.10).
 kanin. 1.10. Mga bono ng disulfide sa istraktura ng insulin
kanin. 1.10. Mga bono ng disulfide sa istraktura ng insulin
5. Supersecondary na istraktura ng mga protina. Sa mga protina na may iba't ibang pangunahing istraktura at pag-andar, kung minsan ay nakikita ang mga ito magkatulad na kumbinasyon at kamag-anak na posisyon ng mga pangalawang istruktura, na tinatawag na supersecondary structure. Sinasakop nito ang isang intermediate na posisyon sa pagitan ng pangalawang at tersiyaryong istruktura, dahil ito ay isang tiyak na kumbinasyon ng mga elemento ng pangalawang istraktura sa pagbuo ng tersiyaryong istraktura ng protina. Ang mga supersecondary na istruktura ay may mga partikular na pangalan, gaya ng "α-helix-turn-a-helix," "leucine zipper," "zinc fingers," atbp. Ang ganitong mga supersecondary na istruktura ay katangian ng mga DNA-binding protein.
"Leucine zipper." Ang ganitong uri ng supersecondary na istraktura ay ginagamit upang pagsamahin ang dalawang protina. Sa ibabaw ng mga nakikipag-ugnay na protina mayroong mga α-helical na rehiyon na naglalaman ng hindi bababa sa apat na nalalabi ng leucine. Ang mga residue ng leucine sa α-helix ay matatagpuan sa pagitan ng anim na amino acid. Dahil ang bawat pagliko ng α-helix ay naglalaman ng 3.6 na residue ng amino acid, ang mga leucine radical ay matatagpuan sa ibabaw ng bawat ikalawang pagliko. Leucine residues ng α-helix ng isang protina ay maaaring makipag-ugnayan sa leucine residues ng isa pang protina (hydrophobic interaksyon), pagkonekta sa kanila nang sama-sama (Fig. 1.11.). Maraming DNA binding protein ang gumagana sa mga oligomeric complex kung saan ang mga indibidwal na subunit ay naka-link sa isa't isa sa pamamagitan ng "leucine zippers."
 kanin. 1.11. "Leucine zipper" sa pagitan ng α-helical na rehiyon ng dalawang protina
kanin. 1.11. "Leucine zipper" sa pagitan ng α-helical na rehiyon ng dalawang protina
Ang isang halimbawa ng naturang mga protina ay mga histone. Mga histone- mga protina ng nukleyar, na naglalaman ng isang malaking bilang ng mga positibong sisingilin na amino acid - arginine at lysine (hanggang sa 80%). Ang mga molekula ng histone ay pinagsama sa mga oligomeric complex na naglalaman ng walong monomer gamit ang "leucine zippers", sa kabila ng makabuluhang homonymous na singil ng mga molekulang ito.
"Zinc finger"- isang variant ng supersecondary na istraktura, katangian ng DNA-binding proteins, ay may anyo ng isang pinahabang fragment sa ibabaw ng protina at naglalaman ng mga 20 amino acid residues (Fig. 1.12). Ang hugis ng "extended finger" ay sinusuportahan ng zinc atom na nakagapos sa apat na amino acid radical - dalawang cysteine residues at dalawang histidine residues. Sa ilang mga kaso, sa halip na mga labi ng histidine, mayroong mga labi ng cysteine. Dalawang malapit na nakalatag na mga labi ng cysteine ay pinaghihiwalay mula sa iba pang dalawang nalalabi sa Gisili sa pamamagitan ng isang pagkakasunud-sunod ng Cys na binubuo ng humigit-kumulang 12 mga residu ng amino acid. Ang rehiyong ito ng protina ay bumubuo ng isang α-helix, ang mga radikal na maaaring partikular na magbigkis sa mga rehiyon ng regulasyon ng pangunahing uka ng DNA. Indibidwal na nagbubuklod na pagtitiyak
 kanin. 1.12. Ang pangunahing istraktura ng rehiyon ng mga protina na nagbubuklod ng DNA na bumubuo sa istraktura ng "zinc finger" (ang mga titik ay nagpapahiwatig ng mga amino acid na bumubuo sa istrukturang ito)
kanin. 1.12. Ang pangunahing istraktura ng rehiyon ng mga protina na nagbubuklod ng DNA na bumubuo sa istraktura ng "zinc finger" (ang mga titik ay nagpapahiwatig ng mga amino acid na bumubuo sa istrukturang ito)
Ang regulatory DNA binding protein ay nakasalalay sa pagkakasunud-sunod ng mga residue ng amino acid na matatagpuan sa rehiyon ng zinc finger. Ang ganitong mga istraktura ay naglalaman, sa partikular, mga receptor para sa mga steroid hormone na kasangkot sa regulasyon ng transkripsyon (pagbabasa ng impormasyon mula sa DNA hanggang RNA).
PAKSA 1.2. MGA BATAYANG PANGKAT NG PROTEIN FUNCTIONING. MGA DROGA BILANG LIGAND NA NAKAKAAPEKTO SA PROTEIN FUNCTION
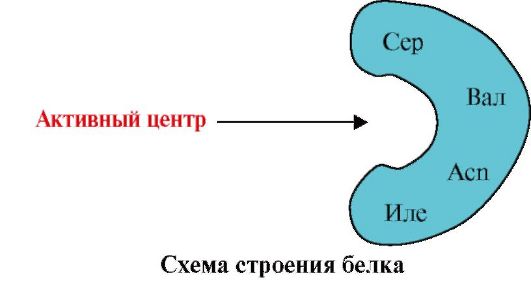
1. Ang aktibong sentro ng protina at ang pakikipag-ugnayan nito sa ligand. Sa panahon ng pagbuo ng tertiary structure, ang isang rehiyon ay nabuo sa ibabaw ng isang functionally active protein, kadalasan sa isang recess, na nabuo ng mga amino acid radical na malayo sa isa't isa sa pangunahing istraktura. Ang rehiyong ito, na may natatanging istraktura para sa isang partikular na protina at may kakayahang partikular na makipag-ugnayan sa isang partikular na molekula o grupo ng mga katulad na molekula, ay tinatawag na protein-ligand binding site o aktibong site. Ang mga ligand ay mga molekula na nakikipag-ugnayan sa mga protina.
Mataas na pagtitiyak Ang pakikipag-ugnayan ng protina sa ligand ay sinisiguro ng complementarity ng istraktura ng aktibong sentro sa istraktura ng ligand.
Complementarity- ito ang spatial at kemikal na pagsusulatan ng mga nakikipag-ugnayan na ibabaw. Ang aktibong sentro ay hindi lamang dapat tumutugma sa spatially sa ligand na kasama dito, ngunit ang mga bono (ionic, hydrogen, at hydrophobic na pakikipag-ugnayan) ay dapat mabuo sa pagitan ng mga functional na grupo ng mga radical na kasama sa aktibong sentro at ang ligand, na humahawak sa ligand. sa aktibong sentro (Larawan 1.13).
 kanin. 1.13. Komplementaryong pakikipag-ugnayan ng protina sa ligand
kanin. 1.13. Komplementaryong pakikipag-ugnayan ng protina sa ligand
Ang ilang mga ligand, kapag nakakabit sa aktibong sentro ng isang protina, ay gumaganap ng isang pantulong na papel sa paggana ng mga protina. Ang ganitong mga ligand ay tinatawag na cofactor, at ang mga protina na naglalaman ng isang bahaging hindi protina ay tinatawag kumplikadong mga protina(kumpara sa mga simpleng protina, na binubuo lamang ng bahagi ng protina). Ang bahaging hindi protina, na mahigpit na konektado sa protina, ay tinatawag grupong prostetik. Halimbawa, ang myoglobin, hemoglobin at cytochromes ay naglalaman ng isang prosthetic group, heme, na naglalaman ng isang iron ion, na mahigpit na nakakabit sa aktibong sentro. Ang mga kumplikadong protina na naglalaman ng heme ay tinatawag na hemoproteins.
Kapag ang mga tiyak na ligand ay nakakabit sa mga protina, ang pag-andar ng mga protina na ito ay ipinahayag. Kaya, ang albumin, ang pinakamahalagang protina sa plasma ng dugo, ay nagpapakita ng transport function nito sa pamamagitan ng paglakip ng mga hydrophobic ligand, tulad ng mga fatty acid, bilirubin, ilang gamot, atbp. sa aktibong sentro (Fig. 1.14)
Ang mga ligand na nakikipag-ugnayan sa tatlong-dimensional na istraktura ng peptide chain ay maaaring hindi lamang mga low-molecular na organic at inorganic na molekula, kundi pati na rin ang mga macromolecule:
DNA (mga halimbawang may DNA-binding protein na tinalakay sa itaas);
Polysaccharides;
 kanin. 1.14. Relasyon sa pagitan ng genotype at phenotype
kanin. 1.14. Relasyon sa pagitan ng genotype at phenotype
Ang natatanging pangunahing istraktura ng mga protina ng tao, na naka-encode sa molekula ng DNA, ay natanto sa mga cell sa anyo ng isang natatanging conformation, aktibong istraktura ng sentro at mga function ng protina.
Sa mga kasong ito, kinikilala ng protina ang isang tiyak na rehiyon ng ligand na katapat at komplementaryo sa binding site. Kaya, sa ibabaw ng mga hepatocytes mayroong mga protina ng receptor para sa hormone insulin, na mayroon ding istraktura ng protina. Ang pakikipag-ugnayan ng insulin sa receptor ay nagdudulot ng pagbabago sa conformation nito at pag-activate ng mga sistema ng pagbibigay ng senyas, na humahantong sa pag-imbak ng mga sustansya sa mga hepatocytes pagkatapos kumain.
kaya, Ang paggana ng mga protina ay batay sa tiyak na pakikipag-ugnayan ng aktibong sentro ng protina sa ligand.
2. Ang istraktura ng domain at ang papel nito sa paggana ng mga protina. Ang mahahabang polypeptide chain ng mga globular na protina ay madalas na natitiklop sa ilang mga compact, medyo independiyenteng mga rehiyon. Mayroon silang isang independiyenteng istraktura ng tersiyaryo, na nakapagpapaalaala sa mga globular na protina, at tinatawag na mga domain. Salamat sa istruktura ng domain ng mga protina, mas madaling mabuo ang kanilang tertiary structure.
Sa mga protina ng domain, ang mga site na nagbubuklod ng ligand ay madalas na matatagpuan sa pagitan ng mga domain. Kaya, ang trypsin ay isang proteolytic enzyme na ginawa ng exocrine na bahagi ng pancreas at kinakailangan para sa panunaw ng mga protina ng pagkain. Mayroon itong dalawang-domain na istraktura, at ang sentro ng pagbubuklod ng trypsin kasama ang ligand nito - protina ng pagkain - ay matatagpuan sa uka sa pagitan ng dalawang domain. Sa aktibong sentro, ang mga kondisyon na kinakailangan para sa epektibong pagbubuklod ng isang tiyak na site ng protina ng pagkain at hydrolysis ng mga peptide bond nito ay nilikha.
Ang iba't ibang mga domain sa isang protina ay maaaring lumipat nang may kaugnayan sa isa't isa kapag ang aktibong sentro ay nakikipag-ugnayan sa ligand (Larawan 1.15).
Hexokinase- isang enzyme na nag-catalyze sa phosphorylation ng glucose gamit ang ATP. Ang aktibong site ng enzyme ay matatagpuan sa lamat sa pagitan ng dalawang domain. Kapag ang hexokinase ay nagbubuklod sa glucose, ang mga domain na nakapaligid dito ay malapit at ang substrate ay nakulong, kung saan nangyayari ang phosphorylation (tingnan ang Fig. 1.15).
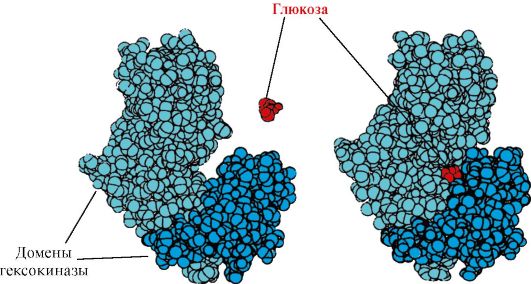 kanin. 1.15. Pagbubuklod ng mga domain ng hexokinase sa glucose
kanin. 1.15. Pagbubuklod ng mga domain ng hexokinase sa glucose
Sa ilang mga protina, ang mga domain ay nagsasagawa ng mga independiyenteng pag-andar sa pamamagitan ng pagbubuklod sa iba't ibang mga ligand. Ang ganitong mga protina ay tinatawag na multifunctional.
3. Ang mga gamot ay mga ligand na nakakaapekto sa paggana ng mga protina. Ang pakikipag-ugnayan ng mga protina sa mga ligand ay tiyak. Gayunpaman, dahil sa conformational lability ng protina at ang aktibong sentro nito, posible na pumili ng isa pang substansiya na maaari ring makipag-ugnayan sa protina sa aktibong sentro o iba pang bahagi ng molekula.
Ang isang sangkap na katulad ng istraktura sa isang natural na ligand ay tinatawag structural analogue ng ligand o isang di-likas na ligand. Nakikipag-ugnayan din ito sa protina sa aktibong site. Ang isang structural analogue ng isang ligand ay maaaring parehong mapahusay ang function ng protina (agonista), at bawasan ito (kalaban). Ang ligand at ang mga istrukturang analogue nito ay nakikipagkumpitensya sa isa't isa para sa pagbubuklod sa protina sa parehong site. Ang mga naturang sangkap ay tinatawag mapagkumpitensyang modulators(mga regulator) ng mga function ng protina. Maraming mga gamot ang kumikilos bilang mga inhibitor ng protina. Ang ilan sa mga ito ay nakuha sa pamamagitan ng kemikal na pagbabago ng mga natural na ligand. Ang mga inhibitor ng mga function ng protina ay maaaring mga gamot at lason.
Ang Atropine ay isang mapagkumpitensyang inhibitor ng M-cholinergic receptors. Ang acetylcholine ay isang neurotransmitter para sa paghahatid ng mga nerve impulses sa pamamagitan ng cholinergic synapses. Upang maisagawa ang paggulo, ang acetylcholine na inilabas sa synaptic cleft ay dapat makipag-ugnayan sa receptor protein ng postsynaptic membrane. Dalawang uri ang natagpuan mga cholinergic receptor:
M receptor bilang karagdagan sa acetylcholine, pumipili itong nakikipag-ugnayan sa muscarine (fly agaric toxin). M - cholinergic receptors ay naroroon sa makinis na kalamnan at, kapag nakikipag-ugnayan sa acetylcholine, nagiging sanhi ng kanilang pag-urong;
H receptor partikular na nagbubuklod sa nikotina. Ang mga N-cholinergic receptor ay matatagpuan sa mga synapses ng striated skeletal muscles.
Tukoy na inhibitor Mga receptor ng M-cholinergic ay atropine. Ito ay matatagpuan sa mga halaman ng belladonna at henbane.
 Ang Atropine ay may mga functional na grupo na katulad ng istraktura sa acetylcholine at ang kanilang spatial arrangement, samakatuwid ito ay isang mapagkumpitensyang inhibitor ng M-cholinergic receptors. Isinasaalang-alang na ang pagbubuklod ng acetylcholine sa M-cholinergic receptors ay nagiging sanhi ng pag-urong ng makinis na mga kalamnan, ang atropine ay ginagamit bilang isang gamot na nagpapagaan ng kanilang spasm. (antispasmodic). Kaya, kilala ang paggamit ng atropine upang i-relax ang mga kalamnan ng mata kapag tinitingnan ang fundus, pati na rin upang mapawi ang mga spasms sa panahon ng gastrointestinal colic. Ang mga M-cholinergic receptor ay naroroon din sa gitnang sistema ng nerbiyos (CNS), kaya ang malalaking dosis ng atropine ay maaaring maging sanhi ng hindi kanais-nais na reaksyon mula sa gitnang sistema ng nerbiyos: motor at mental na pagkabalisa, guni-guni, kombulsyon.
Ang Atropine ay may mga functional na grupo na katulad ng istraktura sa acetylcholine at ang kanilang spatial arrangement, samakatuwid ito ay isang mapagkumpitensyang inhibitor ng M-cholinergic receptors. Isinasaalang-alang na ang pagbubuklod ng acetylcholine sa M-cholinergic receptors ay nagiging sanhi ng pag-urong ng makinis na mga kalamnan, ang atropine ay ginagamit bilang isang gamot na nagpapagaan ng kanilang spasm. (antispasmodic). Kaya, kilala ang paggamit ng atropine upang i-relax ang mga kalamnan ng mata kapag tinitingnan ang fundus, pati na rin upang mapawi ang mga spasms sa panahon ng gastrointestinal colic. Ang mga M-cholinergic receptor ay naroroon din sa gitnang sistema ng nerbiyos (CNS), kaya ang malalaking dosis ng atropine ay maaaring maging sanhi ng hindi kanais-nais na reaksyon mula sa gitnang sistema ng nerbiyos: motor at mental na pagkabalisa, guni-guni, kombulsyon.
Ang Ditilin ay isang mapagkumpitensyang agonist ng H-cholinergic receptors, na pumipigil sa paggana ng neuromuscular synapses.
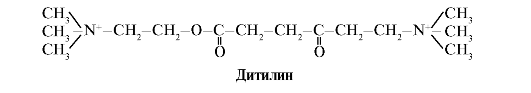 Ang mga neuromuscular synapses ng skeletal muscles ay naglalaman ng H-cholinergic receptors. Ang kanilang pakikipag-ugnayan sa acetylcholine ay humahantong sa mga contraction ng kalamnan. Sa ilang mga operasyon sa operasyon, pati na rin sa mga endoscopic na pag-aaral, ginagamit ang mga gamot na nagdudulot ng pagpapahinga ng mga kalamnan ng kalansay. (mga relaxant ng kalamnan). Kabilang dito ang dithiline, na isang structural analogue ng acetylcholine. Ito ay nakakabit sa H-cholinergic receptors, ngunit hindi katulad ng acetylcholine, ito ay napakabagal na nawasak ng enzyme acetylcholinesterase. Bilang resulta ng matagal na pagbubukas ng mga channel ng ion at patuloy na depolarization ng lamad, ang pagpapadaloy ng mga nerve impulses ay nagambala at nangyayari ang relaxation ng kalamnan. Sa una, ang mga katangiang ito ay natuklasan sa curare poison, kaya naman tinawag ang mga naturang gamot parang curare.
Ang mga neuromuscular synapses ng skeletal muscles ay naglalaman ng H-cholinergic receptors. Ang kanilang pakikipag-ugnayan sa acetylcholine ay humahantong sa mga contraction ng kalamnan. Sa ilang mga operasyon sa operasyon, pati na rin sa mga endoscopic na pag-aaral, ginagamit ang mga gamot na nagdudulot ng pagpapahinga ng mga kalamnan ng kalansay. (mga relaxant ng kalamnan). Kabilang dito ang dithiline, na isang structural analogue ng acetylcholine. Ito ay nakakabit sa H-cholinergic receptors, ngunit hindi katulad ng acetylcholine, ito ay napakabagal na nawasak ng enzyme acetylcholinesterase. Bilang resulta ng matagal na pagbubukas ng mga channel ng ion at patuloy na depolarization ng lamad, ang pagpapadaloy ng mga nerve impulses ay nagambala at nangyayari ang relaxation ng kalamnan. Sa una, ang mga katangiang ito ay natuklasan sa curare poison, kaya naman tinawag ang mga naturang gamot parang curare.
PAKSA 1.3. DENATURASYON NG MGA PROTEIN AT ANG POSIBILIDAD NG KANILANG SPONTANEOUS RENATIVATION
1. Dahil ang katutubong conformation ng mga protina ay pinananatili dahil sa mahinang pakikipag-ugnayan, ang mga pagbabago sa komposisyon at mga katangian ng kapaligiran na nakapalibot sa protina, ang pagkakalantad sa mga kemikal na reagents at pisikal na mga kadahilanan ay nagdudulot ng pagbabago sa kanilang conformation (ang ari-arian ng conformational lability). Ang pagsira sa isang malaking bilang ng mga bono ay humahantong sa pagkasira ng katutubong conformation at denaturation ng mga protina.
Denaturasyon ng mga protina- ito ay ang pagkasira ng kanilang katutubong conformation sa ilalim ng impluwensya ng denaturing agent, sanhi ng pagkalagot ng mahina na mga bono na nagpapatatag sa spatial na istraktura ng protina. Ang denaturation ay sinamahan ng pagkasira ng natatanging three-dimensional na istraktura at aktibong sentro ng protina at ang pagkawala ng biological na aktibidad nito (Larawan 1.16).
Ang lahat ng mga denatured molecule ng isang protina ay nakakakuha ng random na conformation na naiiba sa iba pang molekula ng parehong protina. Ang mga amino acid radical na bumubuo sa aktibong sentro ay lumalabas na spatially na malayo sa isa't isa, i.e. ang tiyak na binding site ng protina na may ligand ay nawasak. Sa panahon ng denaturation, ang pangunahing istraktura ng mga protina ay nananatiling hindi nagbabago.
Application ng denaturing agent sa biological na pananaliksik at gamot. Sa mga biochemical na pag-aaral, bago matukoy ang mababang molekular na timbang na mga compound sa biological na materyal, ang mga protina ay karaniwang inalis muna mula sa solusyon. Ang trichloroacetic acid (TCA) ay kadalasang ginagamit para sa layuning ito. Pagkatapos idagdag ang TCA sa solusyon, ang mga na-denatured na protina ay namuo at madaling maalis sa pamamagitan ng pagsasala (Talahanayan 1.1.)
Sa medisina, ang mga ahente ng denaturing ay kadalasang ginagamit para i-sterilize ang mga medikal na instrumento at materyales sa mga autoclave (ang denaturing agent ay mataas ang temperatura) at bilang antiseptics (alcohol, phenol, chloramine) para sa paggamot sa mga kontaminadong ibabaw na naglalaman ng pathogenic microflora.
2. Kusang pag-activate ng protina- patunay ng determinismo ng pangunahing istraktura, pag-aayos at pag-andar ng mga protina. Ang mga indibidwal na protina ay mga produkto ng isang gene na may magkaparehong pagkakasunud-sunod ng amino acid at nakakakuha ng parehong conformation sa cell. Ang pangunahing konklusyon na ang pangunahing istraktura ng isang protina ay naglalaman na ng impormasyon tungkol sa conform at function nito ay ginawa batay sa kakayahan ng ilang mga protina (sa partikular, ribonuclease at myoglobin) na kusang mag-renativate - ibalik ang kanilang katutubong conformation pagkatapos ng denaturation.
Ang pagbuo ng mga istruktura ng spatial na protina ay isinasagawa sa pamamagitan ng paraan ng self-assembly - isang kusang proseso kung saan ang isang polypeptide chain, na may natatanging pangunahing istraktura, ay may posibilidad na magpatibay ng isang conform na may pinakamababang libreng enerhiya sa solusyon. Ang kakayahang mag-renativate ng mga protina na nagpapanatili ng kanilang pangunahing istraktura pagkatapos ng denaturation ay inilarawan sa isang eksperimento sa enzyme ribonuclease.
Ang Ribonuclease ay isang enzyme na sumisira sa mga bono sa pagitan ng mga indibidwal na nucleotide sa isang molekula ng RNA. Ang globular protein na ito ay may isang polypeptide chain, ang tersiyaryong istraktura nito ay pinatatag ng maraming mahina at apat na disulfide bond.
Ang paggamot sa ribonuclease na may urea, na sumisira sa mga bono ng hydrogen sa molekula, at isang ahente ng pagbabawas, na sumisira sa mga bono ng disulfide, ay humahantong sa denaturation ng enzyme at pagkawala ng aktibidad nito.
Ang pag-alis ng mga denaturing agent sa pamamagitan ng dialysis ay humahantong sa pagpapanumbalik ng conformation at function ng protina, i.e. sa muling pagsilang. (Larawan 1.17).
 kanin. 1.17. Denaturasyon at pagbabagong-buhay ng ribonuclease
kanin. 1.17. Denaturasyon at pagbabagong-buhay ng ribonuclease
A - katutubong conformation ng ribonuclease, sa tersiyaryong istraktura kung saan mayroong apat na disulfide bond; B - denatured ribonuclease molecule;
B - reactivated ribonuclease molecule na may naibalik na istraktura at paggana
1. Punan ang talahanayan 1.2.
Talahanayan 1.2. Pag-uuri ng mga amino acid ayon sa polarity ng mga radical
2. Isulat ang formula ng tetrapeptide:
Asp - Pro - Fen - Liz
a) i-highlight ang mga paulit-ulit na grupo sa peptide na bumubuo sa peptide backbone at ang mga variable na grupo na kinakatawan ng mga amino acid radical;
b) lagyan ng label ang N- at C-termini;
c) i-highlight ang mga peptide bond;
d) magsulat ng isa pang peptide na binubuo ng parehong mga amino acid;
e) bilangin ang bilang ng mga posibleng variant ng isang tetrapeptide na may katulad na komposisyon ng amino acid.
3. Ipaliwanag ang papel ng pangunahing istraktura ng mga protina gamit ang halimbawa ng isang paghahambing na pagsusuri ng dalawang magkatulad na istruktura at ebolusyonaryong malapit na peptide hormone ng mammalian neurohypophysis - oxytocin at vasopressin (Talahanayan 1.3).
Talahanayan 1.3. Istraktura at pag-andar ng oxytocin at vasopressin
 Para dito:
Para dito:
a) ihambing ang komposisyon at pagkakasunud-sunod ng amino acid ng dalawang peptides;
b) hanapin ang pagkakapareho ng pangunahing istraktura ng dalawang peptides at ang pagkakapareho ng kanilang biological na pagkilos;
c) makahanap ng mga pagkakaiba sa istraktura ng dalawang peptides at mga pagkakaiba sa kanilang mga pag-andar;
d) gumuhit ng isang konklusyon tungkol sa impluwensya ng pangunahing istraktura ng mga peptides sa kanilang mga pag-andar.
4. Ilarawan ang mga pangunahing yugto ng pagbuo ng conformation ng mga globular na protina (pangalawa, tertiary na istruktura, ang konsepto ng supersecondary na istraktura). Ipahiwatig ang mga uri ng mga bono na kasangkot sa pagbuo ng mga istruktura ng protina. Aling mga amino acid radical ang maaaring lumahok sa pagbuo ng mga hydrophobic na pakikipag-ugnayan, ionic, hydrogen bond.
Magbigay ng halimbawa.
5. Tukuyin ang konsepto ng "conformational lability ng mga protina", ipahiwatig ang mga dahilan para sa pagkakaroon at kahalagahan nito.
6. Palawakin ang kahulugan ng sumusunod na parirala: "Ang paggana ng mga protina ay batay sa kanilang partikular na pakikipag-ugnayan sa ligand," gamit ang mga termino at ipinapaliwanag ang kanilang kahulugan: protein conformation, active center, ligand, complementarity, protein function.
7. Gamit ang isang halimbawa, ipaliwanag kung ano ang mga domain at kung ano ang kanilang papel sa paggana ng mga protina.
MGA GAWAIN SA PAGKONTROL SA SARILI
1. tugma.
Functional group sa amino acid radical:
A. Carboxyl group B. Hydroxyl group C Guanidine group D. Thiol group E. Amino group
2. Piliin ang tamang sagot.
Ang mga amino acid na may polar uncharged radicals ay:
A. Cis B. Asn
B. Glu G. Tatlo
3. Piliin ang tamang sagot.
Mga radikal na amino acid:
A. Magbigay ng pagtitiyak ng pangunahing istraktura B. Makilahok sa pagbuo ng istrukturang tersiyaryo
B. Matatagpuan sa ibabaw ng protina, naiimpluwensyahan nila ang solubility nito D. Bumuo ng aktibong sentro
D. Makilahok sa pagbuo ng mga peptide bond
4. Piliin ang tamang sagot.
Ang mga hydrophobic na pakikipag-ugnayan ay maaaring mabuo sa pagitan ng mga radikal na amino acid:
A. Tre Lay B. Pro Three
B. Nakilala si Ile G. Tir Ala D. Val Fen
5. Piliin ang tamang sagot.
Ang mga ionic bond ay maaaring mabuo sa pagitan ng mga amino acid radical:
A. Gln Asp B. Abr Liz
B. Liz Glu G. Gis Asp D. Asn Abr
6. Piliin ang tamang sagot.
Ang mga hydrogen bond ay maaaring mabuo sa pagitan ng mga amino acid radical:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. tugma.
Uri ng bono na kasangkot sa pagbuo ng istraktura ng protina:
A. Pangunahing istraktura B. Pangalawang istraktura
B. Tertiary structure
D. Supersecondary na istraktura E. Conformation.
1. Hydrogen bonds sa pagitan ng mga atomo ng peptide backbone
2. Mahinang mga bono sa pagitan ng mga functional na grupo ng mga radikal na amino acid
3. Mga bono sa pagitan ng α-amino at α-carboxyl na grupo ng mga amino acid
8. Piliin ang tamang sagot. Trypsin:
A. Proteolytic enzyme B. Naglalaman ng dalawang domain
B. Nag-hydrolyze ng starch
D. Ang aktibong site ay matatagpuan sa pagitan ng mga domain. D. Binubuo ng dalawang polypeptide chain.
9. Piliin ang tamang sagot. Atropine:
A. Neurotransmitter
B. Structural analogue ng acetylcholine
B. Nakikipag-ugnayan sa mga H-cholinergic receptor
D. Pinapalakas ang pagpapadaloy ng mga nerve impulses sa pamamagitan ng cholinergic synapses
D. Competitive inhibitor ng M-cholinergic receptors
10. Piliin ang mga tamang pahayag. Sa mga protina:
A. Ang pangunahing istraktura ay naglalaman ng impormasyon tungkol sa istraktura ng aktibong site nito
B. Ang aktibong sentro ay nabuo sa antas ng pangunahing istraktura
B. Ang conformation ay mahigpit na naayos sa pamamagitan ng covalent bonds
D. Ang aktibong site ay maaaring makipag-ugnayan sa isang grupo ng mga katulad na ligand
dahil sa conformational lability ng mga protina D. Ang mga pagbabago sa kapaligiran ay maaaring makaapekto sa affinity ng active
sentro sa ligand
1. 1-B, 2-G, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
MGA BATAYANG TERMINO AT KONSEPTO
1. Protina, polypeptide, amino acids
2. Pangunahin, pangalawa, pangatlong mga istruktura ng protina
3. Conformation, native protein conformation
4. Covalent at mahinang mga bono sa protina
5. Conformational lability
6. Protina aktibong site
7. Ligands
8. Pagtitiklop ng protina
9. Structural analogues ng ligands
10. Mga protina ng domain
11. Simple at kumplikadong mga protina
12. Protein denaturation, denaturing agent
13. Muling pag-activate ng mga protina
Lutasin ang mga problema
"Ang istrukturang organisasyon ng mga protina at ang batayan ng kanilang paggana"
1. Ang pangunahing pag-andar ng protina - hemoglobin A (HbA) ay ang transportasyon ng oxygen sa mga tisyu. Sa populasyon ng tao, maraming anyo ng protina na ito ang kilala na may mga binagong katangian at paggana - ang tinatawag na abnormal na hemoglobin. Halimbawa, ang hemoglobin S, na matatagpuan sa mga pulang selula ng dugo ng mga pasyente na may sakit sa sickle cell (HbS), ay natagpuan na may mababang solubility sa ilalim ng mga kondisyon ng mababang bahagyang presyon ng oxygen (tulad ng kaso sa venous blood). Ito ay humahantong sa pagbuo ng mga pinagsama-samang protina na ito. Ang protina ay nawawalan ng paggana, namuo, at ang mga pulang selula ng dugo ay nagiging hindi regular na hugis (ang ilan sa mga ito ay bumubuo ng isang karit na hugis) at nawasak nang mas mabilis kaysa karaniwan sa pali. Bilang resulta, nabubuo ang sickle cell anemia.
Ang tanging pagkakaiba sa pangunahing istraktura ng HbA ay natagpuan sa rehiyon ng N-terminal ng hemoglobin β-chain. Ihambing ang mga rehiyon ng N-terminal ng β-strand at ipakita kung paano nakakaapekto ang mga pagbabago sa pangunahing istruktura ng protina sa mga katangian at paggana nito.
 Para dito:
Para dito:
a) isulat ang mga formula ng mga amino acid kung saan naiiba ang HbA at ihambing ang mga katangian ng mga amino acid na ito (polarity, charge).
b) gumawa ng isang konklusyon tungkol sa dahilan ng pagbaba sa solubility at pagkagambala ng transportasyon ng oxygen sa mga tisyu.
2. Ang figure ay nagpapakita ng isang diagram ng istraktura ng isang protina na may isang nagbubuklod na sentro na may isang ligand (aktibong sentro). Ipaliwanag kung bakit ang protina ay pumipili sa pagpili ng ligand. Para dito:
a) tandaan kung ano ang aktibong sentro ng isang protina at isaalang-alang ang istraktura ng aktibong sentro ng protina na ipinapakita sa figure;
b) isulat ang mga pormula ng mga radikal na amino acid na bumubuo sa aktibong sentro;
c) gumuhit ng ligand na maaaring partikular na makipag-ugnayan sa aktibong site ng protina. Ipahiwatig dito ang mga functional na grupo na maaaring bumuo ng mga bono sa mga amino acid radical na bumubuo sa aktibong sentro;
d) ipahiwatig ang mga uri ng mga bono na lumitaw sa pagitan ng ligand at amino acid radical ng aktibong sentro;
e) ipaliwanag kung saan nakabatay ang pagtitiyak ng interaksyon ng protina-ligand.
 3.
Ipinapakita ng figure ang aktibong site ng protina at ilang ligand.
3.
Ipinapakita ng figure ang aktibong site ng protina at ilang ligand.
Tukuyin kung aling ligand ang pinakamalamang na nakikipag-ugnayan sa aktibong site ng protina at bakit.
 Anong mga uri ng mga bono ang lumitaw sa panahon ng pagbuo ng isang protina-ligand complex?
Anong mga uri ng mga bono ang lumitaw sa panahon ng pagbuo ng isang protina-ligand complex?
4. Ang mga istrukturang analogue ng natural na mga ligand ng protina ay maaaring gamitin bilang mga gamot upang baguhin ang aktibidad ng mga protina.
Ang acetylcholine ay isang tagapamagitan ng paghahatid ng paggulo sa mga neuromuscular synapses. Kapag ang acetylcholine ay nakikipag-ugnayan sa mga protina - mga receptor ng postsynaptic membrane ng mga kalamnan ng kalansay, ang mga channel ng ion ay bubukas at nangyayari ang pag-urong ng kalamnan. Ang Ditilin ay isang gamot na ginagamit sa ilang mga operasyon upang makapagpahinga ng mga kalamnan, dahil nakakagambala ito sa paghahatid ng mga nerve impulses sa pamamagitan ng neuromuscular synapses. Ipaliwanag ang mekanismo ng pagkilos ng ditilin bilang muscle relaxant. Para dito:
a) isulat ang mga formula ng acetylcholine at dithiline at ihambing ang kanilang mga istruktura;
b) ilarawan ang mekanismo ng nakakarelaks na epekto ng ditilin.
5. Sa ilang mga sakit, ang temperatura ng katawan ng pasyente ay tumataas, na itinuturing na isang proteksiyon na reaksyon ng katawan. Gayunpaman, ang mataas na temperatura ay nakakapinsala sa mga protina ng katawan. Ipaliwanag kung bakit sa mga temperaturang higit sa 40 °C ang paggana ng mga protina ay naaabala at may banta sa buhay ng tao. Upang gawin ito, tandaan:
1) Ang istraktura ng mga protina at ang mga bono na humahawak sa istraktura nito sa katutubong conform;
2) Paano nagbabago ang istraktura at paggana ng mga protina sa pagtaas ng temperatura?;
3) Ano ang homeostasis at bakit ito mahalaga sa pagpapanatili ng kalusugan ng tao.
Modular unit 2 OLIGOMERICK PROTEINS BILANG MGA TARGET NG REGULATORY IMPLUENCES. STRUCTURAL AT FUNCTIONAL DIVERSITY NG PROTEINS. MGA PAMAMARAAN PARA SA PAGHIWALAY AT PAGLILID NG MGA PROTEIN
Mga layunin sa pagkatuto Magagawang:
1. Gumamit ng kaalaman tungkol sa mga tampok ng istraktura at mga function ng oligomeric na mga protina upang maunawaan ang mga adaptive na mekanismo ng regulasyon ng kanilang mga function.
2. Ipaliwanag ang papel ng mga chaperone sa synthesis at pagpapanatili ng conformation ng protina sa ilalim ng mga kondisyon ng cellular.
3. Ipaliwanag ang iba't ibang mga manifestations ng buhay sa pamamagitan ng iba't ibang mga istraktura at mga function ng mga protina synthesize sa katawan.
4. Pag-aralan ang kaugnayan sa pagitan ng istraktura ng mga protina at ang kanilang pag-andar gamit ang mga halimbawa ng paghahambing ng mga kaugnay na hemoprotein - myoglobin at hemoglobin, pati na rin ang mga kinatawan ng limang klase ng mga protina ng pamilya ng immunoglobulin.
5. Ilapat ang kaalaman tungkol sa mga kakaibang katangian ng physicochemical properties ng mga protina upang pumili ng mga paraan para sa kanilang paglilinis mula sa iba pang mga protina at mga dumi.
6. Bigyang-kahulugan ang mga resulta ng quantitative at qualitative na komposisyon ng mga protina ng plasma ng dugo upang kumpirmahin o linawin ang klinikal na diagnosis.
alamin:
1. Mga tampok ng istraktura ng mga oligomeric na protina at adaptive na mekanismo para sa pag-regulate ng kanilang mga function gamit ang halimbawa ng hemoglobin.
2. Ang istraktura at mga tungkulin ng mga chaperone at ang kanilang kahalagahan para sa pagpapanatili ng katutubong conformation ng mga protina sa ilalim ng mga kondisyon ng cellular.
3. Mga prinsipyo ng pagsasama-sama ng mga protina sa mga pamilya batay sa pagkakapareho ng kanilang conformation at mga function gamit ang halimbawa ng mga immunoglobulin.
4. Mga pamamaraan para sa paghihiwalay ng mga protina batay sa mga katangian ng kanilang physicochemical properties.
5. Electrophoresis ng plasma ng dugo bilang isang paraan para sa pagtatasa ng qualitative at quantitative na komposisyon ng mga protina.
PAKSA 1.4. MGA TAMPOK NG ISTRUKTURA AT PAGGAMIT NG OLIGOMERIC PROTEINS GAMIT ANG HALIMBAWA NG HEMOGLOBIN
1. Maraming protina ang naglalaman ng ilang polypeptide chain. Ang ganitong mga protina ay tinatawag oligomeric, at mga indibidwal na kadena - mga protomer. Ang mga protomer sa mga oligomeric na protina ay konektado ng maraming mahinang non-covalent bond (hydrophobic, ionic, hydrogen). Pakikipag-ugnayan
ang mga protomer ay isinasagawa salamat sa complementarity kanilang mga contact surface.
Ang bilang ng mga protomer sa mga oligomeric na protina ay maaaring mag-iba nang malaki: ang hemoglobin ay naglalaman ng 4 na protomer, ang enzyme aspartate aminotransferase ay may 12 protomer, at ang tabako na mosaic virus na protina ay naglalaman ng 2120 na mga protomer na konektado ng mga non-covalent bond. Dahil dito, ang mga oligomeric na protina ay maaaring magkaroon ng napakataas na molekular na timbang.
Ang pakikipag-ugnayan ng isang protomer sa iba ay maaaring ituring na isang espesyal na kaso ng pakikipag-ugnayan ng protina-ligand, dahil ang bawat protomer ay nagsisilbing ligand para sa iba pang mga protomer. Ang bilang at paraan ng pagsali sa mga protomer sa isang protina ay tinatawag istraktura ng quaternary na protina.
Ang mga protina ay maaaring maglaman ng mga protomer ng pareho o magkakaibang mga istraktura, halimbawa, ang mga homodimer ay mga protina na naglalaman ng dalawang magkaparehong protomer, at ang mga heterodimer ay mga protina na naglalaman ng dalawang magkaibang protomer.
Kung ang mga protina ay naglalaman ng iba't ibang mga protomer, ang mga nagbubuklod na sentro na may iba't ibang mga ligand na naiiba sa istraktura ay maaaring mabuo sa kanila. Kapag ang isang ligand ay nagbubuklod sa aktibong site, ang pag-andar ng protina na ito ay ipinahayag. Ang isang sentro na matatagpuan sa ibang protomer ay tinatawag na allosteric (iba sa aktibo). Nakikipag-ugnayan allosteric ligand o effector, ito ay gumaganap ng isang regulatory function (Larawan 1.18). Ang pakikipag-ugnayan ng allosteric center sa effector ay nagdudulot ng mga pagbabago sa conformational sa istraktura ng buong oligomeric na protina dahil sa conformational lability nito. Nakakaapekto ito sa affinity ng aktibong site para sa isang partikular na ligand at kinokontrol ang paggana ng protina na iyon. Ang pagbabago sa conformation at function ng lahat ng protomer sa panahon ng interaksyon ng isang oligomeric na protina na may hindi bababa sa isang ligand ay tinatawag na cooperative conformational na pagbabago. Ang mga effector na nagpapahusay ng function ng protina ay tinatawag mga activator, at mga effector na pumipigil sa paggana nito ay mga inhibitor.
Kaya, ang mga oligomeric na protina, pati na rin ang mga protina na may istraktura ng domain, ay may bagong pag-aari kumpara sa mga monomeric na protina - ang kakayahang allosterically regulate ang mga function (regulasyon sa pamamagitan ng paglakip ng iba't ibang mga ligand sa protina). Ito ay makikita sa pamamagitan ng paghahambing ng mga istruktura at pag-andar ng dalawang malapit na nauugnay na kumplikadong protina, myoglobin at hemoglobin.
 kanin. 1.18. Scheme ng istraktura ng isang dimeric na protina
kanin. 1.18. Scheme ng istraktura ng isang dimeric na protina
2. Pagbubuo ng mga spatial na istruktura at paggana ng myoglobin.
Ang Myoglobin (Mb) ay isang protina na matatagpuan sa mga pulang kalamnan, ang pangunahing tungkulin nito ay upang lumikha ng mga reserbang O 2 na kinakailangan para sa matinding trabaho ng kalamnan. Ang Mb ay isang kumplikadong protina na naglalaman ng isang bahagi ng protina - apoMb at isang bahagi na hindi protina - heme. Tinutukoy ng pangunahing istraktura ng apoMB ang compact globular conform nito at ang istraktura ng aktibong sentro, kung saan nakakabit ang non-protein na bahagi ng myoglobin, heme. Ang oxygen na nagmumula sa dugo patungo sa mga kalamnan ay nagbubuklod sa Fe+ 2 hemes sa myoglobin. Ang Mb ay isang monomeric na protina na may napakataas na pagkakaugnay para sa O 2, samakatuwid ang paglabas ng oxygen ng myoglobin ay nangyayari lamang sa panahon ng matinding muscular work, kapag ang bahagyang presyon ng O 2 ay bumababa nang husto.
Pagbuo ng Mv conformation. Sa mga pulang kalamnan, sa mga ribosom sa panahon ng pagsasalin, ang pangunahing istraktura ng MB ay na-synthesize, na kinakatawan ng isang tiyak na pagkakasunud-sunod ng 153 na mga residu ng amino acid. Ang pangalawang istraktura ng Mb ay naglalaman ng walong α-helices, na tinatawag sa Latin na mga letra mula A hanggang H, kung saan may mga non-helical na rehiyon. Ang tertiary na istraktura ng Mb ay may anyo ng isang compact globule, sa recess kung saan ang aktibong sentro ay matatagpuan sa pagitan ng F at E α-helice (Larawan 1.19).
 kanin. 1.19. Istraktura ng myoglobin
kanin. 1.19. Istraktura ng myoglobin
3. Mga tampok ng istraktura at paggana ng aktibong sentro ng MV. Ang aktibong sentro ng Mb ay nakararami na nabubuo ng mga hydrophobic amino acid radical, malawak na may pagitan sa isa't isa sa pangunahing istraktura (halimbawa, Tri 3 9 at Fen 138) Hindi gaanong natutunaw sa mga ligand ng tubig - heme at O 2 - nakakabit sa aktibong sentro. Ang heme ay isang tiyak na ligand ng apoMB (Larawan 1.20), ang batayan nito ay binubuo ng apat na pyrrole ring na konektado ng methenyl bridges; sa gitna ay mayroong Fe+ 2 atom na konektado sa nitrogen atoms ng pyrrole ring sa pamamagitan ng apat na coordination bond. Sa aktibong sentro ng Mb, bilang karagdagan sa mga hydrophobic amino acid radical, mayroon ding mga nalalabi ng dalawang amino acid na may hydrophilic radical - Gis E 7(Gis 64) at GIS F 8(Ang kanyang 93) (Larawan 1.21).
 kanin. 1.20. Ang istraktura ng heme - ang hindi protina na bahagi ng myoglobin at hemoglobin
kanin. 1.20. Ang istraktura ng heme - ang hindi protina na bahagi ng myoglobin at hemoglobin
 kanin. 1.21. Lokasyon ng heme at O2 sa aktibong site ng apomyoglobin at hemoglobin protomer
kanin. 1.21. Lokasyon ng heme at O2 sa aktibong site ng apomyoglobin at hemoglobin protomer
Ang Heme ay covalently na nakatali sa Kanyang F8 sa pamamagitan ng isang iron atom. Ang O 2 ay nakakabit sa bakal sa kabilang panig ng heme plane. Ang kanyang E 7 ay kinakailangan para sa tamang oryentasyon ng O 2 at pinapadali ang pagdaragdag ng oxygen sa Fe + 2 heme
GIS F 8 bumubuo ng isang bono ng koordinasyon sa Fe+ 2 at matatag na inaayos ang heme sa aktibong sentro. Gis E 7 kinakailangan para sa tamang oryentasyon sa aktibong sentro ng isa pang ligand - O 2 sa panahon ng pakikipag-ugnayan nito sa Fe + 2 heme. Ang microenvironment ng heme ay lumilikha ng mga kondisyon para sa malakas ngunit nababaligtad na pagbubuklod ng O 2 hanggang Fe + 2 at pinipigilan ang tubig na makapasok sa hydrophobic active site, na maaaring humantong sa oksihenasyon nito sa Fe + 3.
Ang monomeric na istraktura ng Mb at ang aktibong sentro nito ay tumutukoy sa mataas na pagkakaugnay ng protina para sa O 2.
4. Oligomeric na istraktura ng Hb at regulasyon ng affinity ng Hb sa O 2 ligands. Mga hemoglobin ng tao- isang pamilya ng mga protina, tulad ng myoglobin, na nauugnay sa mga kumplikadong protina (hemoproteins). Mayroon silang tetrameric na istraktura at naglalaman ng dalawang α-chain, ngunit naiiba sa istraktura ng iba pang dalawang polypeptide chain (2α-, 2x-chain). Tinutukoy ng istraktura ng pangalawang polypeptide chain ang mga tampok ng paggana ng mga form na ito ng Hb. Humigit-kumulang 98% ng hemoglobin sa mga pulang selula ng dugo ng isang may sapat na gulang ay hemoglobin A(2α-, 2p-chain).
Sa panahon ng pag-unlad ng pangsanggol, dalawang pangunahing uri ng hemoglobin ang gumagana: embryonic Hb(2α, 2ε), na matatagpuan sa mga unang yugto ng pag-unlad ng pangsanggol, at hemoglobin F (pangsanggol)- (2α, 2γ), na pumapalit sa maagang fetal hemoglobin sa ikaanim na buwan ng intrauterine development at pagkatapos lamang ng kapanganakan ay pinalitan ng Hb A.
Ang HB A ay isang protina na nauugnay sa myoglobin (MB) na matatagpuan sa mga nasa hustong gulang na pulang selula ng dugo ng tao. Ang istraktura ng mga indibidwal na protomer nito ay katulad ng myoglobin. Ang pangalawang at tertiary na mga istruktura ng myoglobin at hemoglobin protomer ay halos magkapareho, sa kabila ng katotohanan na sa pangunahing istraktura ng kanilang mga polypeptide chain ay 24 na residu ng amino acid lamang ang magkapareho (ang pangalawang istraktura ng hemoglobin protomer, tulad ng myoglobin, ay naglalaman ng walong α-helice, itinalaga ng mga Latin na titik mula A hanggang H , at ang istrukturang tersiyaryo ay may anyo ng isang compact globule). Ngunit hindi tulad ng myoglobin, ang hemoglobin ay may oligomeric na istraktura, na binubuo ng apat na polypeptide chain na konektado ng non-covalent bond (Larawan 1.22).
Ang bawat Hb protomer ay nauugnay sa isang hindi protina na bahagi - heme at mga kalapit na protomer. Ang koneksyon ng bahagi ng protina ng Hb na may heme ay katulad ng sa myoglobin: sa aktibong sentro ng protina, ang mga hydrophobic na bahagi ng heme ay napapalibutan ng mga hydrophobic amino acid radical maliban sa His F 8 at His E 7, na matatagpuan sa magkabilang panig ng heme plane at may katulad na papel sa paggana ng protina at ang pagbubuklod nito sa oxygen (tingnan ang istraktura ng myoglobin).
 kanin. 1.22. Oligomeric na istraktura ng hemoglobin
kanin. 1.22. Oligomeric na istraktura ng hemoglobin
Bukod sa, Gis E 7 gumaganap ng isang mahalagang karagdagang tungkulin sa paggana ng Nv. Ang libreng heme ay may 25,000 beses na mas mataas na affinity para sa CO kaysa sa O2. Ang CO ay nabuo sa maliit na dami sa katawan at, dahil sa mataas na pagkakaugnay nito para sa heme, maaari itong makagambala sa transportasyon ng O 2 na kinakailangan para sa buhay ng cell. Gayunpaman, sa komposisyon ng hemoglobin, ang affinity ng heme para sa carbon monoxide ay lumampas sa affinity para sa O 2 ng 200 beses lamang dahil sa pagkakaroon ng His E 7 sa aktibong sentro. Ang natitira sa amino acid na ito ay lumilikha ng pinakamainam na kondisyon para sa pagbubuklod ng heme sa O 2 at pinapahina ang pakikipag-ugnayan ng heme sa CO.
5. Ang pangunahing pag-andar ng HB ay ang transportasyon ng O2 mula sa mga baga patungo sa mga tisyu. Hindi tulad ng monomeric myoglobin, na may napakataas na affinity para sa O2 at gumaganap ng function ng pag-iimbak ng oxygen sa mga pulang kalamnan, ang oligomeric na istraktura ng hemoglobin ay nagbibigay ng:
1) mabilis na saturation ng HB na may oxygen sa mga baga;
2) ang kakayahan ng HB na maglabas ng oxygen sa mga tisyu sa medyo mataas na bahagyang presyon ng O 2 (20-40 mm Hg);
3) ang posibilidad na i-regulate ang affinity ng Hb sa O 2.
6. Ang mga pagbabago sa kooperatiba sa conformation ng hemoglobin protomer ay nagpapabilis sa pagbubuklod ng O 2 sa mga baga at ang paglabas nito sa mga tisyu. Sa baga, ang mataas na bahagyang presyon ng O 2 ay nagtataguyod ng pagbubuklod nito sa Hb sa aktibong site ng apat na protomer (2α at 2β). Ang aktibong sentro ng bawat protomer, tulad ng sa myoglobin, ay matatagpuan sa pagitan ng dalawang α-helice (F at E) sa isang hydrophobic pocket. Naglalaman ito ng bahaging hindi protina - heme, na nakakabit sa bahagi ng protina ng maraming mahinang hydrophobic na interaksyon at isang malakas na bono sa pagitan ng Fe 2 + heme at Kanyang F 8 (tingnan ang Fig. 1.21).
Sa deoxyhemoglobin, dahil sa bond na ito sa Kanyang F 8, ang Fe 2 + atom ay nakausli mula sa heme plane patungo sa histidine. Ang pagbubuklod ng O 2 sa Fe 2 + ay nangyayari sa kabilang panig ng heme sa His E 7 na rehiyon gamit ang isang libreng bono ng koordinasyon. Ang kanyang E 7 ay nagbibigay ng pinakamainam na kondisyon para sa pagbubuklod ng O 2 sa heme iron.
Ang pagdaragdag ng O 2 sa Fe + 2 atom ng isang protomer ay nagiging sanhi ng paggalaw nito sa heme plane, na sinusundan ng histidine residue na nauugnay dito
 kanin. 1.23. Pagbabago sa conformation ng hemoglobin protomer kapag pinagsama sa O 2
kanin. 1.23. Pagbabago sa conformation ng hemoglobin protomer kapag pinagsama sa O 2
Ito ay humahantong sa isang pagbabago sa conformation ng lahat ng polypeptide chain dahil sa kanilang conformational lability. Ang pagbabago ng conformation ng iba pang mga chain ay nagpapadali sa kanilang pakikipag-ugnayan sa mga kasunod na O 2 molecules.
Ang ikaapat na molekula ng O 2 ay nakakabit sa hemoglobin ng 300 beses na mas madali kaysa sa una (Larawan 1.24).
 kanin. 1.24. Mga pagbabago sa kooperatiba sa conformation ng hemoglobin protomer sa panahon ng pakikipag-ugnayan nito sa O2
kanin. 1.24. Mga pagbabago sa kooperatiba sa conformation ng hemoglobin protomer sa panahon ng pakikipag-ugnayan nito sa O2
Sa mga tisyu, ang bawat kasunod na molekula ng O 2 ay mas madaling matanggal kaysa sa nauna, dahil din sa mga pagbabago sa kooperatiba sa conformation ng mga protomer.
7. CO 2 at H+, na nabuo sa panahon ng catabolism ng mga organikong sangkap, binabawasan ang pagkakaugnay ng hemoglobin para sa O 2 sa proporsyon sa kanilang konsentrasyon. Ang enerhiya na kinakailangan para sa paggana ng cell ay pangunahing ginawa sa mitochondria sa panahon ng oksihenasyon ng mga organikong sangkap gamit ang O 2 na inihatid mula sa mga baga ng hemoglobin. Bilang resulta ng oksihenasyon ng mga organikong sangkap, ang mga huling produkto ng kanilang agnas ay nabuo: CO 2 at K 2 O, ang halaga nito ay proporsyonal sa intensity ng patuloy na proseso ng oksihenasyon.
Ang CO 2 ay nagkakalat mula sa mga selula patungo sa dugo at tumagos sa mga pulang selula ng dugo, kung saan, sa ilalim ng pagkilos ng enzyme carbanhydrase, ito ay na-convert sa carbonic acid. Ang mahinang acid na ito ay naghihiwalay sa isang proton at isang bikarbonate ion.
Ang H+ ay may kakayahang sumali sa Kanyang mga radikal 14 6 sa α- at β-chain ng hemoglobin, i.e. sa mga lugar na malayo sa heme. Binabawasan ng protonation ng hemoglobin ang pagkakaugnay nito para sa O 2, itinataguyod ang pag-alis ng O 2 mula sa oxyHb, ang pagbuo ng deoxyHb, at pinatataas ang supply ng oxygen sa mga tisyu sa proporsyon sa bilang ng mga proton na nabuo (Fig. 1.25).
Ang pagtaas sa dami ng inilabas na oxygen depende sa pagtaas ng konsentrasyon ng H+ sa mga pulang selula ng dugo ay tinatawag na Bohr effect (pinangalanan sa Danish na physiologist na si Christian Bohr, na unang nakatuklas ng epektong ito).
Sa baga, ang mataas na partial pressure ng oxygen ay nagtataguyod ng pagbubuklod nito sa deoxyHb, na nagpapababa sa pagkakaugnay ng protina para sa H +. Ang pinakawalan na mga proton sa ilalim ng pagkilos ng carbonic acid ay tumutugon sa mga bikarbonate upang bumuo ng CO 2 at H 2 O
 kanin. 1.25. Pag-asa ng affinity ng Hb para sa O 2 sa konsentrasyon ng CO 2 at mga proton (epekto ng Bohr):
kanin. 1.25. Pag-asa ng affinity ng Hb para sa O 2 sa konsentrasyon ng CO 2 at mga proton (epekto ng Bohr):
A- impluwensya ng CO 2 at H+ na konsentrasyon sa pagpapalabas ng O 2 mula sa complex na may HB (Bohr effect); B- oxygenation ng deoxyhemoglobin sa baga, pagbuo at pagpapalabas ng CO 2.
Ang nagreresultang CO 2 ay pumapasok sa puwang ng alveolar at inaalis kasama ng hanging ibinuga. Kaya, ang dami ng oxygen na inilabas ng hemoglobin sa mga tisyu ay kinokontrol ng mga produkto ng catabolism ng mga organikong sangkap: mas matindi ang pagkasira ng mga sangkap, halimbawa sa pisikal na ehersisyo, mas mataas ang konsentrasyon ng CO 2 at H + at mas maraming oxygen. natatanggap ng mga tisyu bilang isang resulta ng pagbaba sa affinity ng Hb para sa O 2.
8. Allosteric regulation ng affinity ng Hb para sa O2 ng ligand - 2,3-bisphosphoglycerate. Sa erythrocytes, ang allosteric ligand ng hemoglobin, 2,3-bisphosphoglycerate (2,3-BPG), ay synthesize mula sa produkto ng glucose oxidation - 1,3-bisphosphoglycerate. Sa ilalim ng normal na mga kondisyon, ang konsentrasyon ng 2,3-BPG ay mataas at maihahambing sa konsentrasyon ng Hb. Ang 2,3-BPG ay may malakas na negatibong singil na -5.
 Ang bisphosphoglycerate sa mga capillaries ng tissue, na nagbubuklod sa deoxyhemoglobin, ay nagpapataas ng pagpapalabas ng oxygen sa mga tisyu, na binabawasan ang affinity ng Hb para sa O 2.
Ang bisphosphoglycerate sa mga capillaries ng tissue, na nagbubuklod sa deoxyhemoglobin, ay nagpapataas ng pagpapalabas ng oxygen sa mga tisyu, na binabawasan ang affinity ng Hb para sa O 2.
Sa gitna ng molekula ng tetrameric hemoglobin ay isang lukab. Ito ay nabuo sa pamamagitan ng mga residue ng amino acid ng lahat ng apat na protomer (tingnan ang Fig. 1.22). Sa tissue capillaries, ang protonation ng Hb (Bohr effect) ay humahantong sa pagkaputol ng bond sa pagitan ng heme iron at O2. Sa isang molekula
deoxyhemoglobin, kumpara sa oxyhemoglobin, lumilitaw ang mga karagdagang ionic bond na kumokonekta sa mga protomer, bilang isang resulta kung saan ang mga sukat ng gitnang lukab ay tumaas kumpara sa oxyhemoglobin. Ang gitnang lukab ay ang site ng attachment ng 2,3-BPG sa hemoglobin. Dahil sa pagkakaiba sa laki ng gitnang lukab, ang 2,3-BPG ay maaari lamang ilakip sa deoxyhemoglobin.
Ang 2,3-BPG ay nakikipag-ugnayan sa hemoglobin sa isang site na malayo sa mga aktibong sentro ng protina at nabibilang sa allosteric(regulatory) ligand, at ang gitnang lukab ng Hb ay allosteric center. Ang 2,3-BPG ay may malakas na negatibong singil at nakikipag-ugnayan sa limang positibong sisingilin na grupo ng dalawang β-chain ng Hb: ang N-terminal α-amino group ng Val at ang Lys 82 His 143 radical (Fig. 1.26).
 kanin. 1.26. BPG sa gitnang lukab ng deoxyhemoglobin
kanin. 1.26. BPG sa gitnang lukab ng deoxyhemoglobin
Ang BPG ay nagbubuklod sa tatlong pangkat na may positibong charge sa bawat β-strand.
Sa mga tissue capillaries, ang nagreresultang deoxyhemoglobin ay nakikipag-ugnayan sa 2,3-BPG at ang mga ionic bond ay nabuo sa pagitan ng mga positibong sisingilin na radical ng mga β-chain at ang negatibong sisingilin na ligand, na nagbabago sa conformation ng protina at binabawasan ang affinity ng Hb para sa O2 . Ang pagbaba sa affinity ng Hb para sa O 2 ay nagtataguyod ng mas mahusay na paglabas ng O 2 sa tissue.
Sa mga baga, sa mataas na bahagyang presyon, ang oxygen ay nakikipag-ugnayan sa Hb, sumasali sa heme iron; sa kasong ito, ang conformation ng protina ay nagbabago, ang gitnang lukab ay bumababa at ang 2,3-BPG ay inilipat mula sa allosteric center
Kaya, ang mga oligomeric na protina ay may mga bagong katangian kumpara sa mga monomeric na protina. Pagkakabit ng mga ligand sa mga site
spatially na malayo sa isa't isa (allosteric), ay maaaring magdulot ng mga pagbabago sa conformational sa buong molekula ng protina. Dahil sa pakikipag-ugnayan sa mga regulatory ligand, isang pagbabago sa conformation at adaptation ng function ng molekula ng protina sa mga pagbabago sa kapaligiran ay nangyayari.
PAKSA 1.5. MAINTENANCE NG KATUTUBONG PAGSASABUHAY NG MGA PROTEIN SA ILALIM NG MGA KUNDISYON NG CELL
Sa mga cell, sa panahon ng synthesis ng polypeptide chain, ang kanilang transportasyon sa pamamagitan ng mga lamad sa kaukulang mga bahagi ng cell, sa panahon ng proseso ng natitiklop (pagbuo ng katutubong conform) at sa panahon ng pagpupulong ng mga oligomeric na protina, pati na rin sa panahon ng kanilang paggana, intermediate , aggregation-prone, hindi matatag na mga conformation ay lumitaw sa istraktura ng protina. Ang mga hydrophobic radical, kadalasang nakatago sa loob ng molekula ng protina sa native conformation, ay lumilitaw sa ibabaw sa isang hindi matatag na conformation at malamang na pagsamahin sa mga grupo ng iba pang mga protina na hindi gaanong natutunaw sa tubig. Sa mga selula ng lahat ng kilalang organismo, natagpuan ang mga espesyal na protina na nagsisiguro ng pinakamainam na pagtitiklop ng mga protina ng cell, nagpapatatag ng kanilang katutubong conform sa panahon ng paggana at, higit sa lahat, nagpapanatili ng istraktura at pag-andar ng mga intracellular na protina kapag ang homeostasis ay nabalisa. Ang mga protina na ito ay tinatawag "chaperones" na nangangahulugang "yaya" sa Pranses.
1. Molecular chaperones at ang kanilang papel sa pagpigil sa denaturation ng protina.
Ang mga Chaperone (CH) ay inuri ayon sa masa ng kanilang mga subunit. Ang mga chaperone na may mataas na molekular na timbang ay may masa mula 60 hanggang 110 kDa. Kabilang sa mga ito, tatlong klase ang pinaka-pinag-aralan: Sh-60, Sh-70 at Sh-90. Kasama sa bawat klase ang isang pamilya ng mga nauugnay na protina. Kaya, kasama sa Sh-70 ang mga protina na may molekular na timbang mula 66 hanggang 78 kDa. Ang mga chaperone na may mababang timbang na molekular ay may molekular na timbang mula 40 hanggang 15 kDa.
Kabilang sa mga chaperone ay mayroong constitutive protina, ang mataas na basal synthesis na hindi nakadepende sa mga epekto ng stress sa mga selula ng katawan, at inducible, ang synthesis kung saan sa ilalim ng normal na mga kondisyon ay mahina, ngunit tumataas nang husto sa ilalim ng stress. Ang mga inducible chaperone ay tinatawag ding "heat shock proteins" dahil una silang natuklasan sa mga cell na nakalantad sa mataas na temperatura. Sa mga cell, dahil sa mataas na konsentrasyon ng mga protina, ang kusang muling pag-activate ng bahagyang denatured na mga protina ay mahirap. Maaaring pigilan ng Sh-70 ang pagsisimula ng denaturation at makatulong na maibalik ang native conformation ng mga protina. Molecular chaperones-70- isang napaka-conserved na klase ng mga protina na matatagpuan sa lahat ng bahagi ng cell: cytoplasm, nucleus, endoplasmic reticulum, mitochondria. Sa dulo ng carboxyl ng solong polypeptide chain Ш-70 mayroong isang rehiyon na isang uka na may kakayahang makipag-ugnayan sa mga peptide ng haba
mula 7 hanggang 9 na residu ng amino acid na pinayaman ng mga hydrophobic radical. Ang mga nasabing rehiyon sa mga globular na protina ay nangyayari sa humigit-kumulang sa bawat 16 na amino acid. Ang Sh-70 ay may kakayahang protektahan ang mga protina mula sa hindi aktibo na temperatura at ibalik ang conformation at aktibidad ng bahagyang denatured na mga protina.
2. Ang papel ng mga chaperone sa pagtitiklop ng protina. Sa panahon ng synthesis ng protina sa ribosome, ang N-terminal na rehiyon ng polypeptide ay synthesize bago ang C-terminal. Upang mabuo ang katutubong conformation, kinakailangan ang kumpletong pagkakasunud-sunod ng amino acid ng protina. Sa proseso ng synthesis ng protina, ang mga chaperones-70, dahil sa istraktura ng kanilang aktibong sentro, ay maaaring isara ang mga lugar ng polypeptide na madaling kapitan ng pagsasama-sama, na pinayaman sa mga hydrophobic amino acid radical hanggang sa makumpleto ang synthesis (Larawan 1.27, A ).
 kanin. 1.27. Pakikilahok ng mga chaperone sa pagtitiklop ng protina
kanin. 1.27. Pakikilahok ng mga chaperone sa pagtitiklop ng protina
A - pakikilahok ng mga chaperones-70 sa pag-iwas sa mga hydrophobic na pakikipag-ugnayan sa pagitan ng mga seksyon ng synthesized polypeptide; B - pagbuo ng katutubong conformation ng protina sa chaperone complex
Maraming mga high-molecular protein na may kumplikadong conformation, tulad ng isang domain structure, na nakatiklop sa isang espesyal na espasyo na nabuo ng Sh-60. Ш-60 function bilang isang oligomeric complex na binubuo ng 14 subunits. Bumubuo sila ng dalawang guwang na singsing, ang bawat isa ay binubuo ng pitong mga subunit, ang mga singsing na ito ay konektado sa bawat isa. Ang bawat subunit ng Sh-60 ay binubuo ng tatlong domain: apical (apical), pinayaman ng mga hydrophobic radical na nakaharap sa lukab ng singsing, intermediate at equatorial (Fig. 1.28).
 kanin. 1.28. Ang istraktura ng chaperonin complex na binubuo ng 14 Ш-60
kanin. 1.28. Ang istraktura ng chaperonin complex na binubuo ng 14 Ш-60
A - side view; B - tuktok na view
Ang mga synthesized na protina, na may mga elemento sa ibabaw na katangian ng mga nakabukas na molekula, sa partikular na mga hydrophobic radical, ay pumapasok sa lukab ng mga chaperone ring. Sa partikular na kapaligiran ng mga cavity na ito, ang mga posibleng conformation ay hinahanap hanggang ang isa lamang na masigasig na pinaka-kanais-nais ay matagpuan (Larawan 1.27, B). Ang pagbuo ng mga conformation at paglabas ng protina ay sinamahan ng ATP hydrolysis sa rehiyon ng ekwador. Kadalasan, ang naturang chaperone-dependent folding ay nangangailangan ng malaking halaga ng enerhiya.
Bilang karagdagan sa pakikilahok sa pagbuo ng tatlong-dimensional na istraktura ng mga protina at ang pagbabagong-buhay ng bahagyang denatured na mga protina, ang mga chaperone ay kinakailangan din para sa paglitaw ng mga pangunahing proseso tulad ng pagpupulong ng mga oligomeric na protina, pagkilala at transportasyon ng mga denatured na protina sa mga lysosome, transportasyon ng mga protina sa mga lamad, at pakikilahok sa regulasyon ng aktibidad ng mga kumplikadong protina.
PAKSA 1.6. IBA'T IBANG PROTEIN. MGA PAMILYANG PROTEIN: HALIMBAWA NG IMMUNOGLOBULINS
1. Ang mga protina ay gumaganap ng isang mapagpasyang papel sa buhay ng mga indibidwal na selula at ang buong multicellular na organismo, at ang kanilang mga pag-andar ay nakakagulat na magkakaibang. Ito ay tinutukoy ng mga katangian ng pangunahing istraktura at mga conformation ng mga protina, ang natatanging istraktura ng aktibong sentro at ang kakayahang magbigkis ng mga tiyak na ligand.
Isang napakaliit na bahagi lamang ng lahat ng posibleng variant ng mga peptide chain ang maaaring magpatibay ng isang matatag na spatial na istraktura; karamihan
sa mga ito ay maaaring tumagal ng maraming mga conformation na may humigit-kumulang sa parehong Gibbs enerhiya, ngunit may iba't ibang mga katangian. Ang pangunahing istraktura ng karamihan sa mga kilalang protina na pinili ng biological evolution ay nagsisiguro ng pambihirang katatagan ng isa sa mga conformation, na tumutukoy sa mga katangian ng paggana ng protina na ito.
2. Mga pamilyang protina. Sa loob ng parehong biological species, ang mga pagpapalit ng mga residue ng amino acid ay maaaring humantong sa paglitaw ng iba't ibang mga protina na gumaganap ng mga nauugnay na function at may mga homologous na pagkakasunud-sunod ng amino acid. Ang ganitong mga kaugnay na protina ay may kapansin-pansing magkatulad na mga conformation: ang bilang at kamag-anak na posisyon ng α-helice at/o β-structure, karamihan sa mga pagliko at liko ng mga polypeptide chain ay magkapareho o magkapareho. Ang mga protina na may mga homologous na rehiyon ng polypeptide chain, katulad na conformation at mga kaugnay na function ay inuri sa mga pamilya ng protina. Mga halimbawa ng mga pamilya ng protina: serine proteinases, immunoglobulin family, myoglobin family.
Mga serine na protina- isang pamilya ng mga protina na gumaganap ng function ng proteolytic enzymes. Kabilang dito ang digestive enzymes - chymotrypsin, trypsin, elastase at maraming blood clotting factor. Ang mga protina na ito ay may magkaparehong mga amino acid sa 40% ng kanilang mga posisyon at isang katulad na anyo (Larawan 1.29).

kanin. 1.29. Mga spatial na istruktura ng elastase (A) at chymotrypsin (B)
Ang ilang mga pagpapalit ng amino acid ay humantong sa mga pagbabago sa pagtitiyak ng substrate ng mga protina na ito at ang paglitaw ng pagkakaiba-iba ng pagganap sa loob ng pamilya.
3. Pamilya ng immunoglobulin. Sa paggana ng immune system, ang mga protina ng immunoglobulin superfamily ay may malaking papel, na kinabibilangan ng tatlong pamilya ng mga protina:
Antibodies (immunoglobulins);
T-lymphocyte receptors;
Mga protina ng pangunahing histocompatibility complex - MHC klase 1 at 2 (Major Histocompatibility Complex).
Ang lahat ng mga protina na ito ay may istraktura ng domain, binubuo ng mga homologous na immune-like domain at gumaganap ng mga katulad na function: nakikipag-ugnayan sila sa mga dayuhang istruktura, maaaring natunaw sa dugo, lymph o intercellular fluid (antibodies), o matatagpuan sa ibabaw ng mga selula (sariling o dayuhan).
4. Antibodies- mga tiyak na protina na ginawa ng B lymphocytes bilang tugon sa pagpasok ng isang dayuhang istraktura sa katawan, na tinatawag na antigen.
Mga tampok ng istraktura ng mga antibodies
Ang pinakasimpleng molekula ng antibody ay binubuo ng apat na polypeptide chain: dalawang magkaparehong magaan - L, na naglalaman ng mga 220 amino acid, at dalawang magkaparehong mabibigat - H, na binubuo ng 440-700 amino acid. Ang lahat ng apat na kadena sa molekula ng antibody ay konektado ng maraming non-covalent bond at apat na disulfide bond (Larawan 1.30).
Ang mga antibody light chain ay binubuo ng dalawang domain: isang variable domain (VL), na matatagpuan sa N-terminal region ng polypeptide chain, at isang constant domain (CL), na matatagpuan sa C-terminus. Ang mabibigat na chain ay karaniwang may apat na domain: isang variable (VH), na matatagpuan sa N-terminus, at tatlong pare-parehong domain (CH1, CH2, CH3) (tingnan ang Fig. 1.30). Ang bawat immunoglobulin domain ay may β-sheet superstructure kung saan ang dalawang cysteine residues ay pinag-uugnay ng isang disulfide bond.
Sa pagitan ng dalawang pare-parehong mga domain CH1 at CH2 mayroong isang rehiyon na naglalaman ng isang malaking bilang ng mga proline residues, na pumipigil sa pagbuo ng isang pangalawang istraktura at ang pakikipag-ugnayan ng mga kalapit na H-chain sa segment na ito. Ang rehiyon ng bisagra na ito ay nagbibigay ng flexibility ng molekula ng antibody. Sa pagitan ng mga variable na domain ng heavy at light chain ay may dalawang magkaparehong antigen-binding site (aktibong site para sa binding antigens), kaya ang mga naturang antibodies ay madalas na tinatawag bivalents. Hindi ang buong pagkakasunud-sunod ng amino acid ng mga variable na rehiyon ng parehong mga chain ay kasangkot sa pagbubuklod ng antigen sa antibody, ngunit 20-30 amino acid lamang na matatagpuan sa mga hypervariable na rehiyon ng bawat chain. Ang mga rehiyong ito ang tumutukoy sa natatanging kakayahan ng bawat uri ng antibody na makipag-ugnayan sa kaukulang pantulong na antigen.
Ang mga antibodies ay isa sa mga linya ng depensa ng katawan laban sa pagsalakay ng mga dayuhang organismo. Ang kanilang paggana ay maaaring nahahati sa dalawang yugto: ang unang yugto ay ang pagkilala at pagbubuklod ng antigen sa ibabaw ng mga dayuhang organismo, na posible dahil sa pagkakaroon ng mga antigen-binding site sa istraktura ng antibody; ang ikalawang yugto ay ang pagsisimula ng proseso ng inactivation at pagkasira ng antigen. Ang pagtitiyak ng ikalawang yugto ay nakasalalay sa klase ng mga antibodies. Mayroong limang mga klase ng mabibigat na kadena na naiiba sa bawat isa sa istraktura ng mga pare-parehong domain: α, δ, ε, γ at μ, ayon sa kung saan ang limang klase ng mga immunoglobulin ay nakikilala: A, D, E, G at M.
Ang mga tampok na istruktura ng mga mabibigat na kadena ay nagbibigay sa mga rehiyon ng bisagra at mga rehiyon ng C-terminal ng mga mabibigat na kadena ng isang katangian ng conformation ng bawat klase. Matapos ang antigen ay nagbubuklod sa isang antibody, ang mga pagbabago sa conformational sa pare-parehong mga domain ay tumutukoy sa landas para sa pagtanggal ng antigen.
 kanin. 1. 30. Domain structure ng IgG
kanin. 1. 30. Domain structure ng IgG
Immunoglobulins M
Ang mga immunoglobulin M ay may dalawang anyo.
Monomeric na anyo- Unang klase ng mga antibodies na ginawa ng pagbuo ng B lymphocytes. Kasunod nito, maraming mga B cell ang lumipat sa paggawa ng iba pang mga klase ng antibodies, ngunit may parehong antigen-binding site. Ang IgM ay naka-embed sa lamad at gumaganap bilang isang antigen recognition receptor. Ang pagsasama ng IgM sa lamad ng cell ay posible dahil sa pagkakaroon ng 25 hydrophobic amino acid residues sa buntot na bahagi ng rehiyon.
Secretory form ng IgM naglalaman ng limang monomeric subunits na naka-link sa isa't isa sa pamamagitan ng disulfide bond at isang karagdagang polypeptide J chain (Larawan 1.31). Ang mabibigat na kadena ng mga monomer ng form na ito ay hindi naglalaman ng isang hydrophobic tail. Ang pentamer ay may 10 antigen binding site at samakatuwid ay epektibo sa pagkilala at pag-alis ng antigen na unang pumasok sa katawan. Ang secretory form ng IgM ay ang pangunahing klase ng mga antibodies na itinago sa dugo sa panahon ng pangunahing immune response. Ang pagbubuklod ng IgM sa isang antigen ay nagbabago sa konpormasyon ng IgM at naghihikayat sa pagbubuklod nito sa unang bahagi ng protina ng sistemang pandagdag (ang sistema ng pandagdag ay isang hanay ng mga protina na kasangkot sa pagkasira ng antigen) at pag-activate ng sistemang ito. Kung ang antigen ay matatagpuan sa ibabaw ng isang microorganism, ang complement system ay nagdudulot ng pagkagambala sa integridad ng cell lamad at pagkamatay ng bacterial cell.
Mga immunoglobulin G
Sa dami, ang klase ng mga immunoglobulin na ito ay nangingibabaw sa dugo (75% ng lahat ng Igs). IgG - monomer, ang pangunahing klase ng mga antibodies na itinago sa dugo sa panahon ng pangalawang immune response. Matapos ang pakikipag-ugnayan ng IgG sa mga antigen sa ibabaw ng mga microorganism, ang antigen-antibody complex ay nagagawang magbigkis at mag-activate ng mga protina ng complement system o maaaring makipag-ugnayan sa mga partikular na receptor ng macrophage at neutrophils. Ang pakikipag-ugnayan sa mga phagocytes ay humahantong
 kanin. 1.31. Istraktura ng secretory form ng IgM
kanin. 1.31. Istraktura ng secretory form ng IgM
sa pagsipsip ng mga antigen-antibody complex at ang kanilang pagkasira sa mga cell phagosome. Ang IgG ay ang tanging klase ng mga antibodies na nakakapasok sa placental barrier at nagbibigay ng intrauterine na proteksyon ng fetus mula sa mga impeksyon.
Mga immunoglobulin A
Ang pangunahing klase ng mga antibodies na naroroon sa mga pagtatago (gatas, laway, pagtatago ng respiratory tract at intestinal tract). Ang IgA ay pangunahing inilihim sa dimeric na anyo, kung saan ang mga monomer ay naka-link sa isa't isa sa pamamagitan ng karagdagang J chain (Larawan 1.32).
Ang IgA ay hindi nakikipag-ugnayan sa sistema ng pandagdag at mga phagocytic na selula, ngunit sa pamamagitan ng pagbubuklod sa mga mikroorganismo, pinipigilan ng mga antibodies ang kanilang pagkabit sa mga epithelial cells at pagtagos sa katawan.
Mga immunoglobulin E
Ang mga immunoglobulin E ay kinakatawan ng mga monomer kung saan naglalaman ang mabibigat na ε-chain, tulad ng mga μ-chain ng immunoglobulins M, isang variable at apat na pare-parehong domain. Pagkatapos ng pagtatago, ang IgE ay nagbubuklod kasama nito
 kanin. 1.32. Istraktura ng IgA
kanin. 1.32. Istraktura ng IgA
Mga rehiyon ng C-terminal na may kaukulang mga receptor sa ibabaw ng mga mast cell at basophil. Bilang resulta, sila ay nagiging mga receptor para sa mga antigen sa ibabaw ng mga selulang ito (Larawan 1.33).
 kanin. 1.33. Pakikipag-ugnayan ng IgE sa antigen sa ibabaw ng isang mast cell
kanin. 1.33. Pakikipag-ugnayan ng IgE sa antigen sa ibabaw ng isang mast cell
Matapos ang antigen ay nakakabit sa kaukulang antigen-binding na mga site ng IgE, ang mga selula ay tumatanggap ng isang senyas upang i-secrete ang biologically active substances (histamine, serotonin), na higit na responsable para sa pagbuo ng nagpapasiklab na reaksyon at para sa pagpapakita ng mga reaksiyong alerdyi tulad ng hika, urticaria, hay fever.
Mga immunoglobulin D
Ang mga immunoglobulin D ay matatagpuan sa napakaliit na dami sa serum; Ang mabibigat na δ chain ay may isang variable at tatlong pare-parehong domain. Ang mga IgD ay kumikilos bilang mga receptor para sa mga B lymphocyte ay hindi pa rin alam. Ang pakikipag-ugnayan ng mga tiyak na antigens na may mga receptor sa ibabaw ng B-lymphocytes (IgD) ay humahantong sa paghahatid ng mga signal na ito sa cell at ang pag-activate ng mga mekanismo na nagsisiguro sa paglaganap ng isang naibigay na lymphocyte clone.
PAKSA 1.7. PISIKAL AT KEMIKAL NA MGA KATANGIAN NG MGA PROTEIN AT MGA PARAAN PARA SA KANILANG PAGHIWALAY
1. Ang mga indibidwal na protina ay naiiba sa pisikal at kemikal na mga katangian:
Hugis ng mga molekula;
Molekular na timbang;
Ang kabuuang singil, ang magnitude nito ay depende sa ratio ng mga anionic at cationic na grupo ng mga amino acid;
Ang ratio ng polar at non-polar amino acid radicals sa ibabaw ng mga molecule;
Mga antas ng paglaban sa iba't ibang mga ahente ng denaturing.
2. Depende ang solubility ng protina sa mga katangian ng mga protina na nakalista sa itaas, pati na rin sa komposisyon ng daluyan kung saan ang protina ay natunaw (mga halaga ng pH, komposisyon ng asin, temperatura, ang pagkakaroon ng iba pang mga organikong sangkap na maaaring makipag-ugnayan sa protina). Ang halaga ng singil ng mga molekula ng protina ay isa sa mga salik na nakakaapekto sa kanilang solubility. Kapag ang singil sa isoelectric point ay nawala, ang mga protina ay mas madaling pinagsama-sama at namuo. Ito ay partikular na tipikal para sa mga denatured na protina, kung saan lumilitaw ang mga hydrophobic amino acid radical sa ibabaw.
Sa ibabaw ng isang molekula ng protina mayroong parehong positibo at negatibong sisingilin na mga radikal na amino acid. Ang bilang ng mga pangkat na ito, at samakatuwid ang kabuuang singil ng mga protina, ay nakasalalay sa pH ng daluyan, i.e. ratio ng mga konsentrasyon ng H+ - at OH - mga grupo. Sa isang acidic na kapaligiran Ang pagtaas sa konsentrasyon ng H+ ay humahantong sa pagsugpo sa dissociation ng mga carboxyl group -COO - + H+ > - COOH at pagbaba sa negatibong singil ng mga protina. Sa isang alkaline na kapaligiran, ang pagbubuklod ng labis na OH - ng mga proton na nabuo sa panahon ng paghihiwalay ng mga grupo ng amino -NH 3 + + OH - - NH 2 + H 2 O na may pagbuo ng tubig, ay humahantong sa pagbawas sa positibong singil ng mga protina. . Ang halaga ng pH kung saan ang isang protina ay may net zero charge ay tinatawag isoelectric point (IEP). Sa IET, ang bilang ng positibo at negatibong sisingilin na mga grupo ay pareho, i.e. ang protina ay nasa isang isoelectric na estado.
3. Paghihiwalay ng mga indibidwal na protina. Ang mga tampok ng istraktura at paggana ng katawan ay nakasalalay sa hanay ng mga protina na na-synthesize dito. Ang pag-aaral sa istraktura at mga katangian ng mga protina ay imposible nang hindi ihiwalay ang mga ito mula sa selula at nililinis ang mga ito mula sa iba pang mga protina at mga organikong molekula. Mga yugto ng paghihiwalay at paglilinis ng mga indibidwal na protina:
Pagkasira ng cell ang tissue na pinag-aaralan at nakakakuha ng homogenate.
Paghihiwalay ng homogenate sa mga fraction sa pamamagitan ng centrifugation, pagkuha ng nuclear, mitochondrial, cytosolic o iba pang fraction na naglalaman ng gustong protina.
Selective thermal denaturation- panandaliang pag-init ng isang solusyon sa protina, kung saan maaaring alisin ang ilan sa mga na-denatured na dumi ng protina (kung ang protina ay medyo matatag sa init).
Pag-asin. Ang iba't ibang mga protina ay namuo sa iba't ibang mga konsentrasyon ng asin sa solusyon. Sa pamamagitan ng unti-unting pagtaas ng konsentrasyon ng asin, posible na makakuha ng isang bilang ng mga hiwalay na fraction na may isang nangingibabaw na nilalaman ng nakahiwalay na protina sa isa sa mga ito. Ang ammonium sulfate ay kadalasang ginagamit para sa fractionation ng protina. Ang mga protina na may pinakamababang solubility ay namuo sa mababang konsentrasyon ng asin.
Pagsala ng gel- isang paraan ng pagsasala ng mga molekula sa pamamagitan ng namamaga na mga butil ng Sephadex (three-dimensional polysaccharide chain ng dextran na may mga pores). Ang bilis kung saan ang mga protina ay dumaan sa isang column na puno ng Sephadex ay depende sa kanilang molekular na timbang: mas maliit ang masa ng mga molekula ng protina, mas madaling tumagos ang mga ito sa mga butil at manatili doon nang mas mahaba ang masa, mas mabilis ang mga ito mula sa hanay.
Ultracentrifugation- isang paraan na nagsasangkot ng paglalagay ng mga protina sa isang centrifuge tube sa rotor ng isang ultracentrifuge. Kapag umiikot ang rotor, ang sedimentation rate ng mga protina ay proporsyonal sa kanilang molekular na timbang: ang mga praksyon ng mas mabibigat na protina ay matatagpuan mas malapit sa ilalim ng test tube, mas magaan - mas malapit sa ibabaw.
Electrophoresis- isang paraan batay sa mga pagkakaiba sa bilis ng paggalaw ng mga protina sa isang electric field. Ang halagang ito ay proporsyonal sa singil ng mga protina. Ang electrophoresis ng mga protina ay isinasagawa sa papel (sa kasong ito, ang bilis ng paggalaw ng protina ay proporsyonal lamang sa kanilang singil) o sa isang polyacrylamide gel na may isang tiyak na laki ng butas (ang bilis ng paggalaw ng protina ay proporsyonal sa kanilang singil at molekular na timbang) .
Ion exchange chromatography- isang paraan ng fractionation batay sa pagbubuklod ng mga ionized na grupo ng mga protina na may magkasalungat na sisingilin na mga grupo ng mga resin ng palitan ng ion (mga hindi matutunaw na polymeric na materyales). Ang lakas ng pagbubuklod ng protina sa dagta ay proporsyonal sa singil ng protina. Ang mga protina na na-adsorbed sa ion-exchange polymer ay maaaring hugasan sa pagtaas ng mga konsentrasyon ng mga solusyon sa NaCl; mas mababa ang singil ng protina, mas mababa ang konsentrasyon ng NaCl na kinakailangan upang hugasan ang protina na nakagapos sa mga ionic na grupo ng dagta.
Affinity chromatography- ang pinakaspesipikong paraan para sa paghihiwalay ng mga indibidwal na protina Ang isang ligand ng isang protina ay covalently na nakakabit sa isang inert polymer. Kapag ang isang solusyon ng protina ay dumaan sa isang column na may polymer, tanging ang partikular na protina para sa isang ligand ay na-adsorbed sa column dahil sa komplementaryong pagbubuklod ng protina sa ligand.
Dialysis- isang paraan na ginagamit upang alisin ang mababang molekular na timbang na mga compound mula sa isang solusyon ng nakahiwalay na protina. Ang pamamaraan ay batay sa kawalan ng kakayahan ng mga protina na dumaan sa isang semi-permeable na lamad, hindi tulad ng mababang molekular na timbang na mga sangkap. Ito ay ginagamit upang linisin ang mga protina mula sa mga mababang molekular na dumi, halimbawa, mga asing-gamot pagkatapos mag-asin.
MGA TAKDANG ARALIN PARA SA EXTRACURRICULAR WORK
1. Punan ang talahanayan. 1.4.
Talahanayan 1.4. Comparative analysis ng istraktura at pag-andar ng mga kaugnay na protina - myoglobin at hemoglobin
a) tandaan ang istraktura ng aktibong sentro ng Mb at Hb. Ano ang papel ng hydrophobic amino acid radicals sa pagbuo ng mga aktibong sentro ng mga protina na ito? Ilarawan ang istraktura ng aktibong sentro ng Mb at Hb at ang mga mekanismo ng pagkakabit ng mga ligand dito. Ano ang papel na ginagampanan ng His F 8 at His E 7 residues sa paggana ng aktibong sentro ng Mv iHv?
b) anong mga bagong katangian kumpara sa monomeric myoglobin ang mayroon ang malapit na nauugnay na oligomeric protein, hemoglobin? Ipaliwanag ang papel ng mga pagbabago sa kooperatiba sa conformation ng mga protomer sa hemoglobin molecule, ang epekto ng CO 2 at proton concentrations sa affinity ng hemoglobin para sa oxygen, pati na rin ang papel ng 2,3-BPG sa allosteric regulation ng Hb function. .
2. Ilarawan ang mga molekular na chaperone, na binibigyang pansin ang kaugnayan sa pagitan ng kanilang istraktura at pag-andar.
3. Anong mga protina ang napapangkat sa mga pamilya? Gamit ang halimbawa ng pamilya ng immunoglobulin, tukuyin ang mga katulad na katangian ng istruktura at mga kaugnay na function ng mga protina ng pamilyang ito.
4. Ang mga dalisay na indibidwal na protina ay kadalasang kinakailangan para sa biochemical at panggamot na layunin. Ipaliwanag kung anong mga katangian ng physicochemical ng mga protina ang mga pamamaraan na ginamit para sa paghihiwalay at paglilinis ng mga ito.
MGA GAWAIN SA PAGKONTROL SA SARILI
1. Piliin ang tamang sagot.
Mga function ng hemoglobin:
A. Transport ng O 2 mula sa baga papunta sa tissue B. Transport ng H + mula sa tissues papunta sa baga
B. Pagpapanatili ng pare-parehong pH ng dugo D. Pagdala ng CO 2 mula sa baga patungo sa mga tisyu
D. Pagdala ng CO 2 mula sa mga tisyu patungo sa mga baga
2. Piliin ang tamang sagot. Ligandα -protomer Hb ay: A. Heme
B. Oxygen
B. CO G. 2,3-BPG
D. β-Protomer
3. Piliin ang tamang sagot.
Hemoglobin kumpara sa myoglobin:
A. May istrukturang quaternary
B. Ang pangalawang istraktura ay kinakatawan lamang ng mga α-helice
B. Nabibilang sa mga kumplikadong protina
D. Nakikipag-ugnayan sa isang allosteric ligand D. Covalently bound sa heme
4. Piliin ang tamang sagot.
Bumababa ang affinity ng Hb para sa O2:
A. Kapag ang isang O 2 molekula ay idinagdag B. Kapag ang isang O 2 molekula ay tinanggal
B. Kapag nakikipag-ugnayan sa 2,3-BPG
D. Kapag naka-attach sa mga protomer H + D. Kapag bumababa ang konsentrasyon ng 2,3-BPG
5. tugma.
Ang mga uri ng HB ay nailalarawan sa pamamagitan ng:
A. Sa deoxy form ito ay bumubuo ng fibrillar aggregates B. Naglalaman ng dalawang α- at dalawang δ-chain
B. Ang nangingibabaw na anyo ng Hb sa mga adult erythrocytes D. Naglalaman ng heme na may Fe+ 3 sa aktibong sentro
D. Naglalaman ng dalawang α- at dalawang γ-chain 1. HbA 2.
6. tugma.
Hb ligand:
A. Nagbubuklod sa Hb sa allosteric center
B. May napakataas na affinity para sa aktibong site ng Hb
B. Sa pamamagitan ng pagsali, pinapataas nito ang affinity ng Hb para sa O 2 G. Oxidizes Fe+ 2 to Fe+ 3
D. Nakabubuo ng covalent bond sa kanyangF8
7. Piliin ang tamang sagot.
Chaperones:
A. Ang mga protina ay nasa lahat ng bahagi ng selula
B. Tumataas ang synthesis sa ilalim ng stress
B. Makilahok sa hydrolysis ng mga denatured protein
D. Makilahok sa pagpapanatili ng katutubong conformation ng mga protina
D. Lumilikha sila ng mga organel kung saan nabuo ang conformation ng mga protina.
8. Tugma. Mga immunoglobulin:
A. Ang secretory form ay pentameric.
B. Class Ig na tumatagos sa placental barrier
B. Ig - mast cell receptor
D. Ang pangunahing klase ng Ig ay naroroon sa mga pagtatago ng mga epithelial cells. D. B-lymphocyte receptor, ang pag-activate nito ay nagsisiguro sa paglaganap ng cell
9. Piliin ang tamang sagot.
Mga immunoglobulin E:
A. Ginawa ng mga macrophage B. Mayroon silang mabibigat na ε chain.
B. Naka-embed sa lamad ng T-lymphocytes
D. Kumilos bilang mga receptor ng antigen ng lamad sa mga mast cell at basophil
D. Responsable para sa mga reaksiyong alerhiya
10. Piliin ang tamang sagot.
Ang paraan para sa paghihiwalay ng mga protina ay batay sa mga pagkakaiba sa kanilang molekular na timbang:
A. Pagsala ng gel
B. Ultracentrifugation
B. Polyacrylamide gel electrophoresis D. Ion exchange chromatography
D. Affinity chromatography
11. Piliin ang tamang sagot.
Ang paraan para sa paghihiwalay ng mga protina ay batay sa mga pagkakaiba sa kanilang solubility sa tubig:
A. Gel filtration B. Salting out
B. Ion exchange chromatography D. Affinity chromatography
D. Electrophoresis sa polyacrylamide gel
MGA PAMANTAYAN NG MGA SAGOT SA “SELF-CONTROL TASKS”
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
MGA BATAYANG TERMINO AT KONSEPTO
1. Oligomeric na mga protina, protomer, quaternary na istraktura ng mga protina
2. Mga pagbabago sa kooperatiba sa conformation ng protomer
3. Epekto ng Bohr
4. Allosteric na regulasyon ng mga function ng protina, allosteric center at allosteric effector
5. Molecular chaperones, heat shock proteins
6. Mga pamilya ng protina (serine protease, immunoglobulins)
7. IgM-, G-, E-, A-structure-function na relasyon
8. Kabuuang singil ng mga protina, isoelectric point ng mga protina
9. Electrophoresis
10. Pag-aasin
11. Pagsala ng gel
12. Ion exchange chromatography
13. Ultracentrifugation
14. Affinity chromatography
15. Electrophoresis ng mga protina ng plasma ng dugo
MGA GAWAIN PARA SA CLASSROOM WORK
1. Ihambing ang mga dependency ng mga antas ng saturation ng hemoglobin (Hb) at myoglobin (Mb) sa oxygen sa bahagyang presyon nito sa mga tisyu
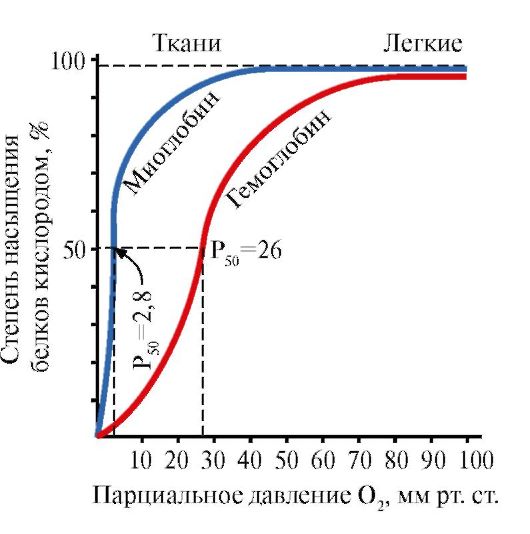 kanin. 1.34. Dependence ng saturation Mv atNHoxygen mula sa bahagyang presyon nito
kanin. 1.34. Dependence ng saturation Mv atNHoxygen mula sa bahagyang presyon nito
Mangyaring tandaan na ang hugis ng mga curves ng saturation ng protina ng oxygen ay iba: para sa myoglobin - isang hyperbola, para sa hemoglobin - isang hugis na sigmoid.
1. ihambing ang mga halaga ng bahagyang presyon ng oxygen kung saan ang Mb at Hb ay puspos ng O 2 ng 50%. Alin sa mga protina na ito ang may mas mataas na affinity para sa O 2?
2. Anong mga istrukturang katangian ng Mb ang tumutukoy sa mataas na pagkakaugnay nito para sa O 2?
3. Anong mga tampok na istruktura ng HB ang nagpapahintulot na ilabas ang O2 sa mga capillary ng mga resting tissue (sa medyo mataas na bahagyang presyon ng O2) at matalas na dagdagan ang paglabas na ito sa mga gumaganang kalamnan? Anong pag-aari ng mga oligomeric na protina ang nagbibigay ng epektong ito?
4. Kalkulahin kung anong halaga ng O 2 (in%) ang ibinibigay ng oxygenated hemoglobin sa mga kalamnan na nagpapahinga at nagtatrabaho?
5. gumawa ng mga konklusyon tungkol sa kaugnayan sa pagitan ng istraktura ng isang protina at ang function nito.
2. Ang dami ng oxygen na inilabas ng hemoglobin sa mga capillary ay nakasalalay sa intensity ng mga proseso ng catabolic sa mga tisyu (Bohr effect). Paano kinokontrol ng mga pagbabago sa metabolismo ng tissue ang affinity ng Hb para sa O2? Epekto ng CO 2 at H+ sa affinity ng Hb para sa O 2
 1. ilarawan ang epekto ng Bohr.
1. ilarawan ang epekto ng Bohr.
2. sa anong direksyon nagpapatuloy ang prosesong ipinapakita sa diagram:
a) sa mga capillary ng baga;
b) sa tissue capillaries?
3. Ano ang pisyolohikal na kahalagahan ng epekto ng Bohr?
4. Bakit binabago ng pakikipag-ugnayan ng Hb sa H+ sa mga site na malayo sa heme ang affinity ng protina para sa O 2?
3. Ang affinity ng Hb para sa O2 ay nakasalalay sa konsentrasyon ng ligand nito - 2,3-bisphosphoglycerate, na isang allosteric regulator ng affinity ng Hb para sa O2. Bakit ang pakikipag-ugnayan ng ligand sa isang site na malayo sa aktibong site ay nakakaapekto sa paggana ng protina? Paano kinokontrol ng 2,3-BPG ang affinity ng Hb para sa O2? Upang malutas ang problema, sagutin ang mga sumusunod na tanong:
1. saan at mula sa ano ang 2.3-bisphosphoglycerate (2,3-BPG) synthesize? Isulat ang formula nito, ipahiwatig ang singil ng molekula na ito.
2. Aling anyo ng hemoglobin (oxy o deoxy) ang nakikipag-ugnayan sa BPG at bakit? Sa anong bahagi ng molekula ng Hb nangyayari ang pakikipag-ugnayan?
3. sa anong direksyon nangyayari ang prosesong ipinapakita sa diagram?
a) sa mga capillary ng tissue;
b) sa mga capillary ng baga?
4. kung saan ang konsentrasyon ng complex ay dapat na mas mataas
Nv-2,3-BFG:
a) sa mga capillary ng mga kalamnan sa pamamahinga,
b) sa mga capillary ng mga gumaganang kalamnan (nagbigay ng parehong konsentrasyon ng BPG sa mga erythrocytes)?
5. Paano magbabago ang affinity ng HB para sa oxygen kapag ang isang tao ay umaangkop sa mga kondisyon ng mataas na altitude, kung ang konsentrasyon ng BPG sa mga erythrocytes ay tumaas? Ano ang pisyolohikal na kahalagahan ng hindi pangkaraniwang bagay na ito?
4. Ang pagkasira ng 2,3-BPG sa panahon ng pag-iimbak ng napanatili na dugo ay nakakapinsala sa mga pag-andar ng HB. Paano magbabago ang affinity ng HB para sa O 2 sa napanatili na dugo kung ang konsentrasyon ng 2,3-BPG sa erythrocytes ay maaaring bumaba mula 8 hanggang 0.5 mmol/l. Posible bang magsalin ng gayong dugo sa mga pasyenteng may malubhang karamdaman kung ang konsentrasyon ng 2,3-BPG ay naibalik nang hindi mas maaga kaysa pagkatapos ng tatlong araw? Posible bang ibalik ang mga function ng pulang selula ng dugo sa pamamagitan ng pagdaragdag ng 2,3-BPG sa dugo?
5. Alalahanin ang istraktura ng pinakasimpleng mga molekula ng immunoglobulin. Ano ang papel na ginagampanan ng mga immunoglobulin sa immune system? Bakit madalas na tinatawag na bivalents ang Igs? Paano nauugnay ang istraktura ng Igs sa kanilang pag-andar? (Ilarawan gamit ang isang halimbawa ng isang klase ng mga immunoglobulin.)
Physicochemical properties ng mga protina at mga pamamaraan para sa kanilang paghihiwalay.
6. Paano nakakaapekto ang netong singil ng isang protina sa solubility nito?
a) matukoy ang kabuuang singil ng peptide sa pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) paano magbabago ang singil ng peptide na ito sa pH >7, pH<7, рН <<7?
c) ano ang isoelectric point ng isang protina (IEP) at sa anong kapaligiran ito namamalagi?
IET ng peptide na ito?
d) sa anong halaga ng pH makikita ang pinakamababang solubility ng peptide na ito.
7. Bakit ang maasim na gatas, hindi tulad ng sariwang gatas, ay "curdle" kapag pinakuluan (ibig sabihin, ang milk protein casein ay namuo)? Sa sariwang gatas, ang mga molekula ng casein ay may negatibong singil.
8. Ang pagsasala ng gel ay ginagamit upang paghiwalayin ang mga indibidwal na protina. Ang isang halo na naglalaman ng mga protina A, B, C na may mga molekular na timbang na katumbas ng 160,000, 80,000 at 60,000, ayon sa pagkakabanggit, ay nasuri sa pamamagitan ng pagsasala ng gel (Larawan 1.35). Ang namamagang mga butil ng gel ay natatagusan ng mga protina na may timbang na molekular na mas mababa sa 70,000 Anong prinsipyo ang pinagbabatayan ng paraan ng paghihiwalay na ito? Aling graph ang wastong sumasalamin sa mga resulta ng fractionation? Ipahiwatig ang pagkakasunud-sunod kung saan ang mga protina A, B, at C ay inilabas mula sa hanay.
 kanin. 1.35. Paggamit ng Gel Filtration sa Paghihiwalay ng Mga Protina
kanin. 1.35. Paggamit ng Gel Filtration sa Paghihiwalay ng Mga Protina
9. Sa Fig. 1.36, A ay nagpapakita ng isang diagram ng electrophoresis sa papel ng mga protina ng serum ng dugo mula sa isang malusog na tao. Ang mga kamag-anak na halaga ng mga fraction ng protina na nakuha gamit ang pamamaraang ito ay: albumin 54-58%, α 1 -globulins 6-7%, α 2 -globulins 8-9%, β-globulins 13%, γ-globulins 11-12% .
 kanin. 1.36 Electrophoresis sa papel ng mga protina ng plasma ng dugo ng isang malusog na tao (A) at isang pasyente (B)
kanin. 1.36 Electrophoresis sa papel ng mga protina ng plasma ng dugo ng isang malusog na tao (A) at isang pasyente (B)
I - γ-globulins; II - β-globulins; III -α 2 -globulin; IV -α 2 -globulin; V - mga albumin
Maraming mga sakit ang sinamahan ng dami ng mga pagbabago sa komposisyon ng mga serum na protina (dysproteinemia). Ang likas na katangian ng mga pagbabagong ito ay isinasaalang-alang kapag gumagawa ng diagnosis at tinatasa ang kalubhaan at yugto ng sakit.
Gamit ang mga datos na ibinigay sa talahanayan. 1.5, gumawa ng isang hula tungkol sa sakit, na kung saan ay nailalarawan sa pamamagitan ng electrophoretic profile na ipinakita sa Fig. 1.36.
Talahanayan 1.5. Mga pagbabago sa konsentrasyon ng mga protina ng serum sa patolohiya
Ang mga protina ay mga organikong sangkap. Ang mga high-molecular compound na ito ay nailalarawan sa pamamagitan ng isang tiyak na komposisyon at, sa hydrolysis, nasira sa mga amino acid. Ang mga molekula ng protina ay maaaring dumating sa maraming iba't ibang anyo, marami sa kanila ay binubuo ng ilang polypeptide chain. Ang impormasyon tungkol sa istraktura ng isang protina ay naka-encode sa DNA, at ang proseso ng synthesis ng mga molekula ng protina ay tinatawag na pagsasalin.
Kemikal na komposisyon ng mga protina
Ang average na protina ay naglalaman ng:
- 52% carbon;
- 7% hydrogen;
- 12% nitrogen;
- 21% oxygen;
- 3% asupre.
Ang mga molekula ng protina ay mga polimer. Upang maunawaan ang kanilang istraktura, kinakailangang malaman kung ano ang kanilang mga monomer - mga amino acid -.
Mga amino acid
Karaniwang nahahati ang mga ito sa dalawang kategorya: patuloy na nangyayari at paminsan-minsang nangyayari. Ang una ay kinabibilangan ng 18 at 2 pang amides: aspartic at glutamic acid. Minsan mayroon lamang tatlong acid na matatagpuan.
Ang mga acid na ito ay maaaring uriin sa iba't ibang paraan: ayon sa likas na katangian ng mga side chain o ang singil ng kanilang mga radical, maaari din silang hatiin sa bilang ng mga grupo ng CN at COOH.
Pangunahing istraktura ng protina
Ang pagkakasunud-sunod ng paghahalili ng mga amino acid sa isang chain ng protina ay tumutukoy sa mga kasunod na antas ng organisasyon, mga katangian at mga pag-andar nito. Ang pangunahing isa sa pagitan ng mga monomer ay peptide. Ito ay nabuo sa pamamagitan ng abstraction ng hydrogen mula sa isang amino acid at ang OH group mula sa isa pa.
Ang unang antas ng organisasyon ng isang molekula ng protina ay ang pagkakasunud-sunod ng mga amino acid sa loob nito, isang kadena lamang na tumutukoy sa istraktura ng mga molekula ng protina. Binubuo ito ng isang "skeleton" na may regular na istraktura. Ito ang paulit-ulit na sequence -NH-CH-CO-. Ang mga indibidwal na side chain ay kinakatawan ng mga amino acid radical (R), ang kanilang mga katangian ay tumutukoy sa komposisyon ng istraktura ng protina.
Kahit na ang istraktura ng mga molekula ng protina ay pareho, maaari silang mag-iba sa mga katangian lamang dahil ang kanilang mga monomer ay may ibang pagkakasunud-sunod sa kadena. Ang pagkakasunud-sunod ng mga amino acid sa isang protina ay tinutukoy ng mga gene at nagdidikta ng ilang biological function sa protina. Ang pagkakasunud-sunod ng mga monomer sa mga molekula na responsable para sa parehong pag-andar ay madalas na magkatulad sa iba't ibang mga species. Ang ganitong mga molekula ay magkapareho o magkatulad sa organisasyon at gumaganap ng parehong mga tungkulin sa iba't ibang uri ng mga organismo - mga homologous na protina. Ang istraktura, mga katangian at pag-andar ng hinaharap na mga molekula ay itinatag na sa yugto ng synthesis ng isang kadena ng mga amino acid.
Ilang karaniwang tampok
Ang istraktura ng mga protina ay pinag-aralan nang mahabang panahon, at ang pagtatasa ng kanilang pangunahing istraktura ay naging posible na gumawa ng ilang mga generalization. Ang isang mas malaking bilang ng mga protina ay nailalarawan sa pagkakaroon ng lahat ng dalawampung amino acid, kung saan mayroong maraming glycine, alanine, glutamine at maliit na tryptophan, arginine, methionine, at histidine. Ang tanging pagbubukod ay ang ilang mga grupo ng mga protina, halimbawa, mga histone. Ang mga ito ay kailangan para sa DNA packaging at naglalaman ng maraming histidine.
Ang anumang uri ng paggalaw ng mga organismo (trabaho ng kalamnan, paggalaw ng protoplasm sa isang cell, pagkutitap ng cilia sa protozoa, atbp.) Ay isinasagawa ng mga protina. Ang istraktura ng mga protina ay nagpapahintulot sa kanila na lumipat, bumuo ng mga hibla at singsing.

Ang pag-andar ng transportasyon ay ang maraming mga sangkap ay dinadala sa buong lamad ng cell sa pamamagitan ng mga espesyal na protina ng carrier.
Ang papel ng hormonal ng mga polymer na ito ay agad na malinaw: ang isang bilang ng mga hormone ay mga protina sa istraktura, halimbawa insulin, oxytocin.
Ang pag-andar ng reserba ay tinutukoy ng katotohanan na ang mga protina ay nakakagawa ng mga deposito. Halimbawa, ang egg valgumin, milk casein, mga protina ng buto ng halaman - nag-iimbak sila ng malaking halaga ng mga sustansya.
Ang lahat ng mga tendon, articular joints, skeletal bones, at hooves ay nabuo ng mga protina, na nagdadala sa atin sa kanilang susunod na function - suporta.
Ang mga molekula ng protina ay mga receptor, na nagsasagawa ng pumipili na pagkilala sa ilang mga sangkap. Ang mga glycoprotein at lectin ay lalo na kilala para sa papel na ito.
Ang pinakamahalagang salik ng kaligtasan sa sakit ay mga antibodies at mga protina sa pinagmulan. Halimbawa, ang proseso ng pamumuo ng dugo ay batay sa mga pagbabago sa fibrinogen protein. Ang mga panloob na dingding ng esophagus at tiyan ay may linya na may proteksiyon na layer ng mauhog na protina - lycins. Ang mga lason ay mga protina din sa pinagmulan. Ang batayan ng balat na nagpoprotekta sa katawan ng mga hayop ay collagen. Ang lahat ng mga function ng protina ay proteksiyon.

Well, ang huling function ay regulasyon. May mga protina na kumokontrol sa paggana ng genome. Ibig sabihin, kinokontrol nila ang transkripsyon at pagsasalin.
Gaano man kahalaga ang papel na ginagampanan ng mga protina, ang istraktura ng mga protina ay nalutas ng mga siyentipiko matagal na ang nakalipas. At ngayon ay nakakatuklas sila ng mga bagong paraan upang magamit ang kaalamang ito.
Ticket 2. 1. Mahahalagang nutritional kadahilanan ng lipid kalikasan. Ang ilang mga lipid ay hindi na-synthesize sa katawan ng tao at samakatuwid ay mahalagang nutritional factor. Kabilang dito ang mga fatty acid na may dalawa o higit pang double bond (polyene) - mahahalagang fatty acid. Ang ilan sa mga acid na ito ay mga substrate para sa synthesis ng mga lokal na hormone - eicosanoids (paksa 8.10).
Mga bitamina na natutunaw sa taba magsagawa ng iba't ibang mga function: bitamina A nakikilahok sa proseso ng pangitain, pati na rin ang paglaki at pagkita ng kaibhan ng cell; ang kakayahang pigilan ang paglaki ng ilang uri ng mga tumor ay napatunayan na; bitamina K nakikilahok sa pamumuo ng dugo; bitamina D nakikilahok sa regulasyon ng metabolismo ng calcium; bitamina E- antioxidant, pinipigilan ang pagbuo ng mga libreng radikal at sa gayon ay kinokontra ang pinsala sa cell bilang resulta ng lipid peroxidation.
Dokumento
2.Istruktura at mga antas ng istrukturang organisasyon ng mga protina
Mayroong apat na antas ng istrukturang organisasyon ng mga protina: pangunahin, pangalawa, tersiyaryo at quaternary. Ang bawat antas ay may sariling katangian.
Pangunahing istraktura ng protina
Ang pangunahing istraktura ng mga protina ay isang linear polypeptide chain ng mga amino acid na konektado ng mga peptide bond. Ang pangunahing istraktura ay ang pinakasimpleng antas ng istrukturang organisasyon ng isang molekula ng protina. Ang mataas na katatagan ay ibinibigay dito ng mga covalent peptide bond sa pagitan ng α-amino group ng isang amino acid at ng α-carboxyl group ng isa pang amino acid. [ipakita].
Kung ang imino group ng proline o hydroxyproline ay kasangkot sa pagbuo ng isang peptide bond, kung gayon mayroon itong ibang anyo [ipakita].
Kapag nabuo ang mga peptide bond sa mga cell, ang carboxyl group ng isang amino acid ay unang naisaaktibo, at pagkatapos ay pinagsama ito sa amino group ng isa pa. Ang synthesis ng laboratoryo ng polypeptides ay isinasagawa sa humigit-kumulang sa parehong paraan.
Ang isang peptide bond ay isang paulit-ulit na fragment ng isang polypeptide chain. Mayroon itong isang bilang ng mga tampok na nakakaapekto hindi lamang sa hugis ng pangunahing istraktura, kundi pati na rin sa mas mataas na antas ng organisasyon ng polypeptide chain:

coplanarity - lahat ng mga atom na kasama sa peptide group ay nasa parehong eroplano;
ang kakayahang umiral sa dalawang anyo ng resonance (keto o enol form);
trans na posisyon ng mga substituent na may kaugnayan sa C-N bond;
ang kakayahang bumuo ng mga bono ng hydrogen, at ang bawat isa sa mga grupo ng peptide ay maaaring bumuo ng dalawang mga bono ng hydrogen sa iba pang mga grupo, kabilang ang mga peptide.
Ang pagbubukod ay mga peptide group na kinasasangkutan ng amino group ng proline o hydroxyproline. Nagagawa lamang nilang bumuo ng isang hydrogen bond (tingnan sa itaas). Nakakaapekto ito sa pagbuo ng pangalawang istraktura ng protina. Ang polypeptide chain sa lugar kung saan matatagpuan ang proline o hydroxyproline ay madaling yumuko, dahil hindi ito hinahawakan, gaya ng dati, ng pangalawang hydrogen bond.
Nomenclature ng peptides at polypeptides. Ang pangalan ng peptides ay binubuo ng mga pangalan ng kanilang constituent amino acids. Dalawang amino acid ang gumagawa ng isang dipeptide, tatlo ang gumagawa ng isang tripeptide, apat ang gumagawa ng isang tetrapeptide, atbp. Ang bawat peptide o polypeptide chain ng anumang haba ay may isang N-terminal amino acid na naglalaman ng isang libreng amino group at isang C-terminal amino acid na naglalaman ng isang libreng carboxyl pangkat. Kapag pinangalanan ang polypeptides, ang lahat ng amino acid ay nakalista nang sunud-sunod, simula sa N-terminal, pinapalitan sa kanilang mga pangalan, maliban sa C-terminal, ang suffix -in na may -yl (dahil ang mga amino acid sa peptides ay wala nang isang pangkat ng carboxyl, ngunit isang carbonyl). Halimbawa, ang pangalan na ipinapakita sa Fig. 1 tripeptide - leuc banlik phenylalane banlik threon sa.
Mga tampok ng pangunahing istraktura ng protina. Sa backbone ng polypeptide chain, ang mga matibay na istruktura (flat peptide groups) ay humalili sa mga relatibong mobile na rehiyon (-CHR), na may kakayahang umikot sa paligid ng mga bono. Ang ganitong mga tampok na istruktura ng polypeptide chain ay nakakaapekto sa spatial arrangement nito.
Pangalawang istraktura ng protina
Ang pangalawang istraktura ay isang paraan ng pagtitiklop ng polypeptide chain sa isang ordered structure dahil sa pagbuo ng hydrogen bonds sa pagitan ng mga peptide group ng parehong chain o katabing polypeptide chain. Ayon sa kanilang pagsasaayos, ang mga pangalawang istruktura ay nahahati sa helical (α-helix) at layered-folded (β-structure at cross-β-form).


α-Helix. Ito ay isang uri ng pangalawang istruktura ng protina na mukhang isang regular na helix, na nabuo dahil sa interpeptide hydrogen bond sa loob ng isang polypeptide chain. Ang modelo ng istraktura ng α-helix (Larawan 2), na isinasaalang-alang ang lahat ng mga katangian ng peptide bond, ay iminungkahi nina Pauling at Corey. Mga pangunahing tampok ng α-helix:
helical configuration ng polypeptide chain na may helical symmetry;
ang pagbuo ng mga bono ng hydrogen sa pagitan ng mga grupo ng peptide ng bawat una at ikaapat na nalalabi sa amino acid;
regularidad ng spiral turns;
ang katumbas ng lahat ng mga residue ng amino acid sa α-helix, anuman ang istraktura ng kanilang mga side radical;
Ang mga side radical ng mga amino acid ay hindi nakikilahok sa pagbuo ng α-helix.
Sa panlabas, ang α-helix ay mukhang isang bahagyang nakaunat na spiral ng isang electric stove. Ang regularidad ng mga bono ng hydrogen sa pagitan ng una at ikaapat na grupo ng peptide ay tumutukoy sa pagiging regular ng mga pagliko ng polypeptide chain. Ang taas ng isang pagliko, o ang pitch ng α-helix, ay 0.54 nm; kabilang dito ang 3.6 na residue ng amino acid, ibig sabihin, ang bawat residue ng amino acid ay gumagalaw sa kahabaan ng axis (ang taas ng isang residue ng amino acid) sa pamamagitan ng 0.15 nm (0.54:3.6 = 0.15 nm), na nagpapahintulot sa amin na pag-usapan ang tungkol sa katumbas ng lahat ng residue ng amino acid sa α-helix. Ang regularity period ng isang α-helix ay 5 turns o 18 amino acid residues; ang haba ng isang panahon ay 2.7 nm. kanin. 3. Pauling-Corey a-helix model
β-Istruktura. Ito ay isang uri ng pangalawang istraktura na may bahagyang kurbadong configuration ng polypeptide chain at nabuo sa pamamagitan ng interpeptide hydrogen bonds sa loob ng mga indibidwal na seksyon ng isang polypeptide chain o katabing polypeptide chain. Tinatawag din itong isang layered-fold na istraktura. May mga uri ng β-structure. Ang limitadong layered na mga rehiyon na nabuo ng isang polypeptide chain ng isang protina ay tinatawag na cross-β form (short β structure). Ang mga hydrogen bond sa cross-β form ay nabuo sa pagitan ng mga peptide group ng mga loop ng polypeptide chain. Ang isa pang uri - ang kumpletong β-istruktura - ay katangian ng buong polypeptide chain, na may pinahabang hugis at hawak ng interpeptide hydrogen bonds sa pagitan ng magkatabing parallel polypeptide chain (Fig. 3). Ang istraktura na ito ay kahawig ng mga bellow ng isang akurdyon. Bukod dito, posible ang mga variant ng β-structure: maaari silang mabuo ng mga parallel chain (ang mga dulo ng N-terminal ng polypeptide chain ay nakadirekta sa parehong direksyon) at antiparallel (ang mga dulo ng N-terminal ay nakadirekta sa iba't ibang direksyon). Ang mga side radical ng isang layer ay inilalagay sa pagitan ng mga side radical ng isa pang layer.
Sa mga protina, ang mga paglipat mula sa α-structure hanggang sa β-structure at likod ay posible dahil sa muling pagsasaayos ng mga hydrogen bond. Sa halip na regular na interpeptide hydrogen bonds sa kahabaan ng chain (salamat sa kung saan ang polypeptide chain ay pinaikot sa isang spiral), ang helical section ay nakakalas at ang hydrogen bond ay malapit sa pagitan ng mga pahabang fragment ng polypeptide chain. Ang paglipat na ito ay matatagpuan sa keratin, ang protina ng buhok. Kapag naghuhugas ng buhok gamit ang mga alkaline detergent, ang helical na istraktura ng β-keratin ay madaling nawasak at ito ay nagiging α-keratin (kulot na buhok ay tumutuwid).
Ang pagkasira ng mga regular na pangalawang istruktura ng mga protina (α-helice at β-structure), sa pamamagitan ng pagkakatulad sa pagtunaw ng isang kristal, ay tinatawag na "pagtunaw" ng mga polypeptides. Sa kasong ito, ang mga bono ng hydrogen ay nasira, at ang mga polypeptide chain ay nasa anyo ng isang random na tangle. Dahil dito, ang katatagan ng pangalawang istruktura ay tinutukoy ng interpeptide hydrogen bond. Ang iba pang mga uri ng mga bono ay halos walang bahagi dito, maliban sa mga disulfide bond sa kahabaan ng polypeptide chain sa mga lokasyon ng cysteine residues. Ang mga maikling peptide ay sarado sa mga cycle dahil sa disulfide bond. Maraming mga protina ang naglalaman ng parehong α-helical na rehiyon at β-structure. Halos walang mga natural na protina na binubuo ng 100% α-helix (ang pagbubukod ay paramyosin, isang protina ng kalamnan na 96-100% α-helix), habang ang mga sintetikong polypeptide ay may 100% na helix.
Ang iba pang mga protina ay may iba't ibang antas ng likid. Ang isang mataas na dalas ng α-helical na mga istruktura ay sinusunod sa paramyosin, myoglobin, at hemoglobin. Sa kaibahan, sa trypsin, isang ribonuclease, isang makabuluhang bahagi ng polypeptide chain ay nakatiklop sa layered β-structures. Mga protina ng sumusuporta sa mga tisyu: keratin (protina ng buhok, lana), collagen (protina ng mga tendon, balat), fibroin (protina ng natural na sutla) ay may β-configuration ng mga polypeptide chain. Ang iba't ibang antas ng helicity ng polypeptide chain ng mga protina ay nagpapahiwatig na, malinaw naman, may mga puwersa na bahagyang nakakagambala sa helicity o "sinisira" ang regular na pagtitiklop ng polypeptide chain. Ang dahilan para dito ay isang mas compact na natitiklop ng protina polypeptide chain sa isang tiyak na dami, ibig sabihin, sa isang tersiyaryong istraktura.
Ang istraktura ng tersiyaryo ng protina
Ang tersiyaryong istraktura ng isang protina ay ang paraan ng pagkakaayos ng polypeptide chain sa espasyo. Batay sa hugis ng kanilang tertiary na istraktura, ang mga protina ay pangunahing nahahati sa globular at fibrillar. Ang mga globular na protina ay kadalasang may hugis na ellipsoid, at ang mga protina ng fibrillar (tulad ng sinulid) ay may pinahabang hugis (hugis ng baras o spindle).
Gayunpaman, ang pagsasaayos ng tertiary na istraktura ng mga protina ay hindi pa nagbibigay ng dahilan upang isipin na ang mga fibrillar na protina ay mayroon lamang β-istraktura, at ang mga globular na protina ay may α-helical na istraktura. May mga fibrillar na protina na may helical, sa halip na layered, nakatiklop na pangalawang istraktura. Halimbawa, ang α-keratin at paramyosin (protina ng obturator muscle ng mollusks), tropomyosin (protein ng skeletal muscles) ay nabibilang sa fibrillar proteins (may hugis ng baras), at ang pangalawang istraktura nito ay α-helix; sa kaibahan, ang mga globular na protina ay maaaring maglaman ng isang malaking bilang ng mga β-istruktura.
Ang spiralization ng isang linear polypeptide chain ay binabawasan ang laki nito ng humigit-kumulang 4 na beses; at ang pag-iimpake sa tertiary structure ay ginagawa itong sampu-sampung beses na mas compact kaysa sa orihinal na chain.
Mga bono na nagpapatatag sa tertiary na istraktura ng isang protina. Ang mga bono sa pagitan ng mga side radical ng mga amino acid ay gumaganap ng isang papel sa pagpapatatag ng tertiary na istraktura. Ang mga koneksyon na ito ay maaaring nahahati sa:
malakas (covalent) [ipakita].
mahina (polar at van der Waals) [ipakita].
Maraming mga bono sa pagitan ng mga side radical ng mga amino acid ang tumutukoy sa spatial na pagsasaayos ng molekula ng protina.
Mga tampok ng samahan ng istrukturang tersiyaryo ng protina. Ang conformation ng tertiary na istraktura ng polypeptide chain ay tinutukoy ng mga katangian ng side radicals ng mga amino acid na kasama dito (na walang kapansin-pansing epekto sa pagbuo ng pangunahin at pangalawang istruktura) at ang microenvironment, i.e., ang kapaligiran. Kapag nakatiklop, ang polypeptide chain ng isang protina ay may posibilidad na magkaroon ng isang energetically favorable form, na nailalarawan sa pamamagitan ng isang minimum na libreng enerhiya. Samakatuwid, ang mga nonpolar R-group, "pag-iwas" sa tubig, ay bumubuo, tulad nito, ang panloob na bahagi ng tertiary na istraktura ng protina, kung saan matatagpuan ang pangunahing bahagi ng hydrophobic residues ng polypeptide chain. Halos walang mga molekula ng tubig sa gitna ng globule ng protina. Ang polar (hydrophilic) R na grupo ng amino acid ay matatagpuan sa labas ng hydrophobic core na ito at napapalibutan ng mga molekula ng tubig. Ang polypeptide chain ay intricately baluktot sa three-dimensional space. Kapag ito ay yumuko, ang pangalawang helical conformation ay nasisira. Ang kadena ay "nasisira" sa mga mahihinang punto kung saan matatagpuan ang proline o hydroxyproline, dahil ang mga amino acid na ito ay mas gumagalaw sa kadena, na bumubuo lamang ng isang hydrogen bond sa iba pang mga peptide group. Ang isa pang lugar ng liko ay ang glycine, na may maliit na R group (hydrogen). Samakatuwid, ang mga R-group ng iba pang mga amino acid, kapag nakasalansan, ay may posibilidad na sumakop sa libreng espasyo sa lokasyon ng glycine. Ang isang bilang ng mga amino acid - alanine, leucine, glutamate, histidine - ay nag-aambag sa pagpapanatili ng matatag na helical na istruktura sa protina, at tulad ng methionine, valine, isoleucine, aspartic acid ay pinapaboran ang pagbuo ng β-structure. Sa isang molekula ng protina na may isang tersiyaryong pagsasaayos, mayroong mga rehiyon sa anyo ng α-helices (helical), β-structure (layered) at isang random na coil. Tanging ang tamang spatial folding ng protina ay ginagawang aktibo ito; ang paglabag nito ay humahantong sa mga pagbabago sa mga katangian ng protina at pagkawala ng biological na aktibidad.
Quaternary na istraktura ng protina
Ang mga protina na binubuo ng isang polypeptide chain ay may tertiary structure lamang. Kabilang dito ang myoglobin - isang protina ng kalamnan tissue na kasangkot sa pagbubuklod ng oxygen, isang bilang ng mga enzymes (lysozyme, pepsin, trypsin, atbp.). Gayunpaman, ang ilang mga protina ay binuo mula sa ilang mga polypeptide chain, na ang bawat isa ay may isang tersiyaryong istraktura. Para sa mga naturang protina, ang konsepto ng quaternary na istraktura ay ipinakilala, na kung saan ay ang organisasyon ng ilang mga polypeptide chain na may isang tertiary na istraktura sa isang solong functional na molekula ng protina. Ang nasabing protina na may istrukturang quaternary ay tinatawag na oligomer, at ang mga polypeptide chain nito na may tertiary na istraktura ay tinatawag na mga protomer o subunits (Larawan 4).

Sa antas ng quaternary ng organisasyon, pinapanatili ng mga protina ang pangunahing pagsasaayos ng istrukturang tersiyaryo (globular o fibrillar). Halimbawa, ang hemoglobin ay isang protina na may istrukturang quaternary at binubuo ng apat na subunits. Ang bawat isa sa mga subunit ay isang globular na protina at, sa pangkalahatan, ang hemoglobin ay mayroon ding globular na pagsasaayos. Mga protina ng buhok at lana - ang mga keratin, na nauugnay sa istrukturang tersiyaryo sa mga protina ng fibrillar, ay may konpormasyon ng fibrillar at istrukturang quaternary.
Pagpapatatag ng istraktura ng quaternary ng protina. Ang lahat ng mga protina na may isang quaternary na istraktura ay nakahiwalay sa anyo ng mga indibidwal na macromolecules na hindi nasira sa mga subunit. Ang mga contact sa pagitan ng mga ibabaw ng mga subunit ay posible lamang dahil sa mga polar group ng mga residue ng amino acid, dahil sa panahon ng pagbuo ng tertiary na istraktura ng bawat isa sa mga polypeptide chain, ang mga side radical ng non-polar amino acids (na bumubuo sa karamihan ng lahat ng proteinogenic amino acids) ay nakatago sa loob ng subunit. Maraming ionic (asin), hydrogen, at sa ilang mga kaso ang disulfide bond ay nabuo sa pagitan ng kanilang mga polar group, na mahigpit na humahawak sa mga subunit sa anyo ng isang organisadong complex. Ang paggamit ng mga sangkap na sumisira sa mga bono ng hydrogen o mga sangkap na nagpapababa sa mga tulay na disulfide ay nagdudulot ng disaggregation ng mga protomer at pagkasira ng quaternary na istraktura ng protina. Sa mesa 1 ay nagbubuod ng data sa mga bono na nagpapatatag ng iba't ibang antas ng organisasyon ng molekula ng protina [ipakita].
Mga tampok ng istrukturang organisasyon ng ilang mga fibrillar na protina
Ang istrukturang organisasyon ng mga fibrillar na protina ay may ilang mga tampok kumpara sa mga globular na protina. Ang mga tampok na ito ay makikita sa halimbawa ng keratin, fibroin at collagen. Ang mga keratin ay umiiral sa α- at β-conformations. Ang α-Keratins at fibroin ay may layered-folded secondary structure, gayunpaman, sa keratin ang mga chain ay parallel, at sa fibroin sila ay antiparallel (tingnan ang Fig. 3); Bilang karagdagan, ang keratin ay naglalaman ng mga interchain na disulfide bond, habang ang fibroin ay wala sa kanila. Ang pagkasira ng mga disulfide bond ay humahantong sa paghihiwalay ng mga polypeptide chain sa keratins. Sa kabaligtaran, ang pagbuo ng maximum na bilang ng mga disulfide bond sa mga keratin sa pamamagitan ng pagkakalantad sa mga ahente ng oxidizing ay lumilikha ng isang malakas na istraktura ng spatial. Sa pangkalahatan, sa mga fibrillar na protina, hindi tulad ng mga globular na protina, kung minsan ay mahirap na mahigpit na makilala sa pagitan ng iba't ibang antas ng organisasyon. Kung tinatanggap natin (tulad ng para sa isang globular na protina) na ang tertiary na istraktura ay dapat mabuo sa pamamagitan ng paglalagay ng isang polypeptide chain sa espasyo, at ang quaternary na istraktura sa pamamagitan ng ilang mga chain, kung gayon sa fibrillar proteins ilang polypeptide chain ang kasangkot na sa panahon ng pagbuo ng pangalawang istraktura. . Ang karaniwang halimbawa ng fibrillar protein ay ang collagen, na isa sa pinakamaraming protina sa katawan ng tao (mga 1/3 ng masa ng lahat ng protina). Ito ay matatagpuan sa mga tisyu na may mataas na lakas at mababang extensibility (buto, tendon, balat, ngipin, atbp.). Sa collagen, ang ikatlong bahagi ng mga residue ng amino acid ay glycine, at halos isang-kapat o bahagyang higit pa ay proline o hydroxyproline.

Ang nakahiwalay na polypeptide chain ng collagen (pangunahing istraktura) ay mukhang isang putol na linya. Naglalaman ito ng humigit-kumulang 1000 amino acid at may molecular weight na humigit-kumulang 10 5 (Larawan 5, a, b). Ang polypeptide chain ay binuo mula sa paulit-ulit na trio ng mga amino acid (triplet) ng sumusunod na komposisyon: gly-A-B, kung saan ang A at B ay anumang amino acid maliban sa glycine (pinaka madalas na proline at hydroxyproline). Ang mga collagen polypeptide chain (o α-chain) sa panahon ng pagbuo ng pangalawang at tertiary na istruktura (Larawan 5, c at d) ay hindi makakagawa ng tipikal na α-helice na may helical symmetry. Ang proline, hydroxyproline at glycine (antihelical amino acids) ay nakakasagabal dito. Samakatuwid, ang tatlong α-chain ay bumubuo, kumbaga, baluktot na mga spiral, tulad ng tatlong mga thread na bumabalot sa isang silindro. Tatlong helical α chain ang bumubuo ng paulit-ulit na istruktura ng collagen na tinatawag na tropocollagen (Larawan 5d). Ang Tropocollagen sa organisasyon nito ay ang tertiary structure ng collagen. Ang mga flat ring ng proline at hydroxyproline na regular na nagpapalit-palit sa kadena ay nagbibigay ng higpit, gayundin ang mga interchain bond sa pagitan ng mga α-chain ng tropocollagen (kaya naman ang collagen ay lumalaban sa pag-unat). Ang Tropocollagen ay mahalagang isang subunit ng collagen fibrils. Ang pagtula ng mga subunit ng tropocollagen sa quaternary na istraktura ng collagen ay nangyayari sa isang hakbang-hakbang na paraan (Larawan 5e).
Ang pagpapatatag ng mga istruktura ng collagen ay nangyayari dahil sa interchain na hydrogen, ionic at van der Waals na mga bono at isang maliit na bilang ng mga covalent bond.
Ang mga α-chain ng collagen ay may iba't ibang istrukturang kemikal. Mayroong iba't ibang uri ng α 1 chain (I, II, III, IV) at α 2 chain. Depende sa kung aling mga α 1 - at α 2 -chain ang kasangkot sa pagbuo ng tatlong-stranded na helix ng tropocollagen, apat na uri ng collagen ang nakikilala:
ang unang uri - dalawang α 1 (I) at isang α 2 chain;
ang pangalawang uri - tatlong α 1 (II) chain;
ikatlong uri - tatlong α 1 (III) chain;
ikaapat na uri - tatlong α 1 (IV) chain.
Ang pinakakaraniwang collagen ay ang unang uri: ito ay matatagpuan sa bone tissue, balat, tendons; Ang collagen ng pangalawang uri ay matatagpuan sa cartilage tissue, atbp. Maaaring may iba't ibang uri ng collagen sa isang uri ng tissue.
Ang iniutos na pagsasama-sama ng mga istruktura ng collagen, ang kanilang katigasan at kawalang-kilos ay tinitiyak ang mataas na lakas ng mga hibla ng collagen. Ang mga protina ng collagen ay naglalaman din ng mga bahagi ng carbohydrate, ibig sabihin, ang mga ito ay mga protina-karbohidrat complex.
Ang collagen ay isang extracellular protein na nabuo sa pamamagitan ng connective tissue cells na matatagpuan sa lahat ng organ. Samakatuwid, na may pinsala sa collagen (o pagkagambala sa pagbuo nito), maraming mga paglabag sa mga sumusuportang function ng connective tissue ng mga organo ang nagaganap.
Ang alpha polypeptide chain ay nagtatapos sa amino acid combination na valine-leucine, at ang beta polypeptide chain ay nagtatapos sa combination na valine-histidine-leucine. Ang mga alpha at beta polypeptide chain sa molekula ng hemoglobin ay hindi nakaayos nang linearly ito ang pangunahing istraktura. Dahil sa pagkakaroon ng intramolecular forces, ang mga polypeptide chain ay pinaikot sa anyo ng isang alpha-helix helix (pangalawang istraktura) na tipikal ng mga protina. Ang alpha-helix helix mismo ay yumuko nang spatially para sa bawat alpha at beta polypeptide chain, na bumubuo ng mga plexuse na hugis ovoid (tertiary structure). Ang mga indibidwal na bahagi ng alpha-helix helices ng polypeptide chain ay minarkahan ng Latin na mga letra mula A hanggang H. Ang lahat ng apat na tertiary curved alpha at beta polypeptide chain ay matatagpuan spatially sa isang tiyak na relasyon - isang quaternary na istraktura. Ang mga ito ay konektado hindi sa pamamagitan ng tunay na mga bono ng kemikal, ngunit sa pamamagitan ng mga puwersa ng intermolecular.
Ito ay naka-out na ang mga tao ay may tatlong pangunahing uri ng normal na hemoglobin: embryonic - U, pangsanggol - F at adult hemoglobin - A. HbU (pinangalanan pagkatapos ng unang titik ng salitang matris) ay nangyayari sa embryo sa pagitan ng 7 at 12 na linggo ng buhay, pagkatapos ito ay nawawala at lumilitaw ang fetal hemoglobin, na pagkatapos ng ikatlong buwan ay ang pangunahing hemoglobin ng fetus. Kasunod nito, unti-unting lumilitaw ang ordinaryong hemoglobin ng may sapat na gulang, na tinatawag na HbA, pagkatapos ng unang titik ng salitang Ingles na "adult". Ang halaga ng fetal hemoglobin ay unti-unting bumababa, upang sa sandali ng kapanganakan, 80% ng hemoglobin ay HbA at 20% lamang ang HbF. Pagkatapos ng kapanganakan, ang fetal hemoglobin ay patuloy na bumababa at sa 2-3 taon ng buhay ito ay 1-2% lamang. Ang parehong halaga ng fetal hemoglobin ay nasa isang may sapat na gulang. Ang halaga ng HbF na higit sa 2% ay itinuturing na pathological para sa isang may sapat na gulang at para sa mga bata na higit sa 3 taong gulang.
Bilang karagdagan sa mga normal na uri ng hemoglobin, higit sa 50 pathological variant ay kasalukuyang kilala. Una silang pinangalanan gamit ang mga letrang Latin. Ang titik B ay wala sa pagtatalaga ng mga uri ng hemoglobin, dahil orihinal itong itinalagang HbS.
Hemoglobin (Hb)- isang chromoprotein na nasa pulang selula ng dugo at kasangkot sa transportasyon ng oxygen sa mga tisyu. Ang hemoglobin sa mga matatanda ay tinatawag na hemoglobin A (Hb A). Ang molekular na timbang nito ay humigit-kumulang 65,000 Da. Ang molekula ng Hb A ay may quaternary na istraktura at may kasamang apat na subunits - polypeptide chain (tinalagang α1, α2, β1 at β2, na ang bawat isa ay nauugnay sa heme.
Tandaan na ang hemoglobin ay isang allosteric na protina ang mga molekula nito ay maaaring magbago mula sa isang conform patungo sa isa pa. Binabago nito ang pagkakaugnay ng protina para sa mga ligand. Ang conformation na may pinakamaliit na affinity para sa ligand ay tinatawag na tense, o T-conformation. Ang conform na may pinakamalaking affinity para sa ligand ay tinatawag na relaxed, o R-conformation.
Ang R- at T-conformation ng molekula ng hemoglobin ay nasa isang estado ng dynamic na equilibrium:
Maaaring ilipat ng iba't ibang salik sa kapaligiran ang balanseng ito sa isang direksyon o sa iba pa. Ang mga allosteric regulator na nakakaapekto sa affinity ng Hb para sa O2 ay: 1) oxygen; 2) konsentrasyon ng H+ (medium pH); 3) carbon dioxide (CO2); 4) 2,3-diphosphoglycerate (DPG). Ang attachment ng isang molekula ng oxygen sa isa sa mga subunit ng hemoglobin ay nagtataguyod ng paglipat ng isang tense conformation sa isang nakakarelaks at pinatataas ang affinity para sa oxygen ng iba pang mga subunit ng parehong molekula ng hemoglobin. Ang phenomenon na ito ay tinatawag na cooperative effect. Ang kumplikadong katangian ng pagbubuklod ng hemoglobin sa oxygen ay makikita ng hemoglobin O2 saturation curve, na may hugis-S (Larawan 3.1).

Larawan 3.1. Mga kurba ng myoglobin (1) at hemoglobin (2) oxygen saturation.
Mga anyo ng molekular Ang hemoglobin ay naiiba sa bawat isa sa istraktura ng mga polypeptide chain. Ang isang halimbawa ng iba't ibang hemoglobin na umiiral sa ilalim ng mga kondisyong pisyolohikal ay fetal hemoglobin (HbF), naroroon sa dugo sa panahon ng embryonic na yugto ng pag-unlad ng tao. Hindi tulad ng HbA, ang molekula nito ay naglalaman ng 2 α- at 2 γ-chain (iyon ay, ang β-chain ay pinalitan ng γ-chain). Ang ganitong hemoglobin ay may mas mataas na pagkakaugnay para sa oxygen. Ito ang nagpapahintulot sa embryo na makatanggap ng oxygen mula sa dugo ng ina sa pamamagitan ng inunan. Sa lalong madaling panahon pagkatapos ng kapanganakan, ang HbF sa dugo ng sanggol ay pinalitan ng HbA.
Ang isang halimbawa ng abnormal o pathological hemoglobin ay ang nabanggit na (tingnan ang 2.4.) hemoglobin S, na matatagpuan sa mga pasyente na may sickle cell anemia. Tulad ng alam mo na, ito ay naiiba sa hemoglobin A sa pamamagitan ng pagpapalit ng glutamate sa valine sa mga β-chain. Ang pagpapalit ng amino acid na ito ay nagdudulot ng pagbaba sa solubility ng HbS sa tubig at pagbaba sa affinity nito para sa O2.
Ang kemikal na istraktura ng mga protina ay kinakatawan ng mga alpha amino acid na konektado sa isang chain sa pamamagitan ng isang peptide bond. Sa mga buhay na organismo, ang komposisyon ay tinutukoy ng genetic code. Sa proseso ng synthesis, sa karamihan ng mga kaso, 20 amino acids ng karaniwang uri ang ginagamit. Ang kanilang maraming kumbinasyon ay bumubuo ng mga molekula ng protina na may malawak na pagkakaiba-iba ng mga katangian. Ang mga residue ng amino acid ay kadalasang napapailalim sa mga pagbabago pagkatapos ng pagsasalin. Maaari silang bumangon bago magsimula ang protina upang maisagawa ang mga function nito, at sa panahon ng aktibidad nito sa cell. Sa mga buhay na organismo, maraming mga molekula ang kadalasang bumubuo ng mga kumplikadong complex. Ang isang halimbawa ay photosynthetic association.
Layunin ng mga koneksyon
Ang mga protina ay itinuturing na isang mahalagang bahagi ng nutrisyon ng tao at hayop dahil sa katotohanan na ang kanilang mga katawan ay hindi maaaring synthesize ang lahat ng kinakailangang mga amino acid. Ang ilan sa kanila ay dapat kasama ng mga pagkaing protina. Ang pangunahing pinagmumulan ng mga compound ay karne, mani, gatas, isda, at butil. Sa isang mas mababang lawak, ang mga protina ay naroroon sa mga gulay, mushroom at berries. Sa panahon ng panunaw sa pamamagitan ng mga enzyme, ang mga natupok na protina ay hinahati sa mga amino acid. Ginagamit na ang mga ito sa biosynthesis ng kanilang sariling mga protina sa katawan o sumasailalim sa karagdagang pagkasira upang makakuha ng enerhiya.

Makasaysayang sanggunian
Ang pagkakasunud-sunod ng istraktura ng protina ng insulin ay unang natukoy ni Frederij Senger. Para sa kanyang trabaho natanggap niya ang Nobel Prize noong 1958. Ginamit ni Sanger ang sequencing method. Gamit ang X-ray diffraction, ang mga three-dimensional na istruktura ng myoglobin at hemoglobin ay kasunod na nakuha (sa huling bahagi ng 1950s). Ang gawain ay isinagawa nina John Kendrew at Max Perutz.
Istraktura ng molekula ng protina
Kabilang dito ang mga linear polymers. Ang mga ito, sa turn, ay binubuo ng alpha amino acid residues, na mga monomer. Bilang karagdagan, ang istraktura ng protina ay maaaring magsama ng mga bahagi ng isang likas na hindi amino acid at binagong mga residu ng amino acid. Kapag nagtatalaga ng mga bahagi, ginagamit ang 1- o 3-titik na pagdadaglat. Ang isang tambalang naglalaman ng dalawa hanggang ilang dosenang residues ay madalas na tinutukoy bilang isang "polypeptide." Bilang resulta ng pakikipag-ugnayan ng alpha-carboxyl group ng isang amino acid sa alpha-amino group ng isa pa, lumilitaw ang mga bono (sa panahon ng pagbuo ng istraktura ng protina). Ang C- at N-terminal na mga dulo ng compound ay nakikilala, depende sa kung aling grupo ng residue ng amino acid ang libre: -COOH o -NH 2 . Sa proseso ng synthesis ng protina sa ribosome, ang unang nalalabi sa terminal ay karaniwang isang residue ng methionine; ang mga kasunod ay nakakabit sa C-terminus ng mga nauna.

Mga antas ng organisasyon
Sila ay iminungkahi ni Lindrem-Lang. Sa kabila ng katotohanan na ang dibisyong ito ay itinuturing na medyo lipas na, ito ay ginagamit pa rin. Iminungkahi na makilala ang apat na antas ng organisasyon ng koneksyon. Ang pangunahing istraktura ng isang molekula ng protina ay tinutukoy ng genetic code at mga katangian ng gene. Ang mas mataas na antas ay nailalarawan sa pamamagitan ng pagbuo sa panahon ng pagtitiklop ng protina. Ang spatial na istraktura ng isang protina ay tinutukoy sa kabuuan ng chain ng amino acid. Gayunpaman, ito ay medyo labile. Maaaring maimpluwensyahan ito ng mga panlabas na salik. Sa pagsasaalang-alang na ito, mas tama na pag-usapan ang tungkol sa conformation ng tambalan na pinaka-kanais-nais at masigasig na mas gusto.
Antas 1
Ito ay kinakatawan ng isang pagkakasunud-sunod ng mga residue ng amino acid ng isang polypeptide chain. Bilang isang tuntunin, ito ay inilarawan gamit ang isa- o tatlong-titik na mga notasyon. Ang pangunahing istraktura ng mga protina ay nailalarawan sa pamamagitan ng matatag na kumbinasyon ng mga residue ng amino acid. Gumagawa sila ng mga tiyak na gawain. Ang ganitong mga "konserbatibong motif" ay nananatiling napanatili sa panahon ng ebolusyon ng mga species. Madalas itong magamit upang mahulaan ang problema ng hindi kilalang protina. Sa pamamagitan ng pagtatasa sa antas ng pagkakatulad (homology) sa mga chain ng amino acid mula sa iba't ibang mga organismo, posibleng matukoy ang evolutionary distance na nabuo sa pagitan ng taxa na bumubuo sa mga organismo na ito. Ang pangunahing istraktura ng mga protina ay tinutukoy sa pamamagitan ng pagkakasunud-sunod o sa pamamagitan ng orihinal na kumplikado ng mRNA nito gamit ang isang genetic code table.

Lokal na pag-order ng isang seksyon ng chain
Ito ang susunod na antas ng organisasyon - ang pangalawang istraktura ng mga protina. Mayroong ilang mga uri nito. Ang lokal na pag-order ng isang bahagi ng isang polypeptide chain ay pinapatatag ng hydrogen bonds. Ang pinakasikat na mga uri ay:

Istraktura ng spatial
Ang tersiyaryong istraktura ng mga protina ay kinabibilangan ng mga elemento ng nakaraang antas. Pinapatatag ang mga ito sa pamamagitan ng iba't ibang uri ng pakikipag-ugnayan. Ang mga hydrophobic bond ay pinakamahalaga. Ang pagpapatatag ay kinabibilangan ng:
- Covalent na pakikipag-ugnayan.
- Ionic bond na nabuo sa pagitan ng amino acid side group na may magkasalungat na singil.
- Mga pakikipag-ugnayan ng hydrogen.
- Mga hydrophobic bond. Sa proseso ng pakikipag-ugnayan sa mga nakapaligid na elemento H 2 O, ang protina ay natitiklop upang ang mga side non-polar amino acid na grupo ay nakahiwalay sa may tubig na solusyon. Ang mga hydrophilic group (polar) ay lumilitaw sa ibabaw ng molekula.
Ang tertiary na istraktura ng mga protina ay tinutukoy ng magnetic (nuclear) resonance method, ilang uri ng microscopy at iba pang mga pamamaraan.
Prinsipyo ng pagtula
Ipinakita ng pananaliksik na madaling matukoy ang isa pang antas sa pagitan ng mga antas 2 at 3. Tinatawag itong "architecture", "laying motif". Ito ay tinutukoy ng kamag-anak na posisyon ng mga bahagi ng pangalawang istraktura (beta strands at alpha helices) sa loob ng mga hangganan ng isang compact globule - ang domain ng protina. Maaari itong umiral nang nakapag-iisa o maisama sa isang mas malaking protina kasama ng iba pang katulad na mga protina. Ito ay itinatag na ang mga motibo sa pag-istilo ay medyo konserbatibo. Ang mga ito ay matatagpuan sa mga protina na walang ebolusyonaryo o functional na mga relasyon. Ang kahulugan ng arkitektura ay ang batayan ng rasyonal (pisikal) na pag-uuri.

Organisasyon ng domain
Sa magkaparehong pag-aayos ng ilang polypeptide chain sa loob ng isang kumplikadong protina, nabuo ang quaternary na istraktura ng mga protina. Ang mga elemento na bumubuo dito ay nabuo nang hiwalay sa mga ribosom. Sa pagtatapos lamang ng synthesis nagsisimulang mabuo ang istrukturang protina na ito. Maaari itong maglaman ng magkaibang at magkaparehong polypeptide chain. Ang quaternary na istraktura ng mga protina ay nagpapatatag dahil sa parehong mga pakikipag-ugnayan tulad ng sa nakaraang antas. Ang ilang mga complex ay maaaring magsama ng ilang dosenang protina.
Istraktura ng protina: mga gawaing proteksiyon
Ang mga polypeptides ng cytoskeleton, na kumikilos sa ilang paraan bilang pampalakas, ay nagbibigay sa maraming organel ng kanilang hugis at nakikilahok sa pagbabago nito. Ang mga istrukturang protina ay nagbibigay ng proteksyon para sa katawan. Halimbawa, ang collagen ay isang protina. Ito ay bumubuo ng batayan sa intercellular substance ng connective tissues. Ang keratin ay mayroon ding proteksiyon na function. Ito ay bumubuo ng batayan ng mga sungay, balahibo, buhok at iba pang mga derivatives ng epidermis. Kapag ang mga protina ay nagbubuklod ng mga lason, sa maraming kaso ay nangyayari ang detoxification. Ito ay kung paano nagagawa ang gawain ng proteksyon ng kemikal ng katawan. Ang mga enzyme ng atay ay may partikular na mahalagang papel sa proseso ng pag-neutralize ng mga lason sa katawan ng tao. Nagagawa nilang masira ang mga lason o i-convert ang mga ito sa natutunaw na anyo. Pinapadali nito ang mas mabilis na transportasyon mula sa katawan. Ang mga protina na nasa dugo at iba pang likido sa katawan ay nagbibigay ng immune defense sa pamamagitan ng pag-trigger ng tugon sa parehong pag-atake ng pathogen at pinsala. Ang mga immunoglobulin (antibodies at mga bahagi ng sistemang pandagdag) ay may kakayahang i-neutralize ang bakterya, mga dayuhang protina at mga virus.
Mekanismo ng regulasyon
Ang mga molekula ng protina, na hindi kumikilos bilang pinagmumulan ng enerhiya o bilang isang materyal na gusali, ay kumokontrol sa maraming proseso ng intracellular. Kaya, dahil sa kanila, ang pagsasalin, transkripsyon, pagpipiraso, at ang aktibidad ng iba pang polypeptides ay kinokontrol. Ang mekanismo ng regulasyon ay batay sa aktibidad ng enzymatic o nagpapakita ng sarili dahil sa tiyak na pagbubuklod sa iba pang mga molekula. Halimbawa, ang transcription factor, activator polypeptides, at repressor proteins ay may kakayahang kontrolin ang intensity ng gene transcription. Sa paggawa nito, nakikipag-ugnayan sila sa mga sequence ng regulasyon ng gene. Ang pinakamahalagang papel sa pagkontrol sa kurso ng mga proseso ng intracellular ay itinalaga sa protina phosphatases at protina kinases. Ang mga enzyme na ito ay nagpapalitaw o humahadlang sa aktibidad ng iba pang mga protina sa pamamagitan ng pagdaragdag o pag-alis ng mga grupo ng pospeyt mula sa kanila.

Gawain ng signal
Madalas itong pinagsama sa function ng regulasyon. Ito ay dahil sa ang katunayan na maraming intracellular, pati na rin ang extracellular, polypeptides ay maaaring magpadala ng mga signal. Ang mga kadahilanan ng paglago, cytokine, hormone at iba pang mga compound ay may ganitong kakayahan. Ang mga steroid ay dinadala sa pamamagitan ng dugo. Ang pakikipag-ugnayan ng hormone sa receptor ay kumikilos bilang isang senyas na nagpapalitaw sa tugon ng cell. Kinokontrol ng mga steroid ang nilalaman ng mga compound sa dugo at mga selula, pagpaparami, paglaki at iba pang mga proseso. Ang isang halimbawa ay insulin. Kinokontrol nito ang mga antas ng glucose. Ang pakikipag-ugnayan ng mga cell ay isinasagawa sa pamamagitan ng signal protein compound na ipinadala sa pamamagitan ng intercellular substance.
Transport ng mga elemento
Ang mga natutunaw na protina na kasangkot sa paggalaw ng maliliit na molekula ay may mataas na pagkakaugnay para sa substrate, na naroroon sa tumaas na konsentrasyon. Mayroon din silang kakayahang ilabas ito nang madali sa mga lugar kung saan mababa ang nilalaman nito. Ang isang halimbawa ay ang transport protein hemoglobin. Inililipat nito ang oxygen mula sa mga baga patungo sa iba pang mga tisyu, at mula sa kanila ay naglilipat ito ng carbon dioxide. Ang ilang mga protina ng lamad ay kasangkot din sa transportasyon ng maliliit na molekula sa pamamagitan ng mga pader ng cell, na binabago ang mga ito. Ang lipid layer ng cytoplasm ay hindi tinatablan ng tubig. Pinipigilan nito ang pagsasabog ng mga sisingilin o polar na molekula. Ang mga koneksyon sa transportasyon ng lamad ay karaniwang nahahati sa mga carrier at channel.
Mga backup na koneksyon
Ang mga protina na ito ay bumubuo ng tinatawag na mga reserba. Nag-iipon sila, halimbawa, sa mga buto ng halaman at mga itlog ng hayop. Ang ganitong mga protina ay kumikilos bilang isang reserbang pinagmumulan ng bagay at enerhiya. Ang ilang mga compound ay ginagamit ng katawan bilang isang amino acid reservoir. Ang mga ito, sa turn, ay mga precursor ng mga aktibong sangkap na kasangkot sa regulasyon ng metabolismo.
Mga cellular receptor
Ang ganitong mga protina ay maaaring matatagpuan nang direkta sa cytoplasm o naka-embed sa dingding. Ang isang bahagi ng koneksyon ay tumatanggap ng signal. Bilang isang patakaran, ito ay isang kemikal na sangkap, at sa ilang mga kaso ay isang mekanikal na epekto (kahabaan, halimbawa), liwanag at iba pang mga stimuli. Sa proseso ng pagkakalantad ng isang signal sa isang tiyak na fragment ng molekula - ang polypeptide receptor - nagsisimula ang mga pagbabago sa conformational nito. Pinipukaw nila ang isang pagbabago sa conformation ng natitirang bahagi na nagpapadala ng stimulus sa iba pang mga bahagi ng cell. Ang pagpapadala ng signal ay maaaring gawin sa iba't ibang paraan. Ang ilang mga receptor ay may kakayahang mag-catalyze ng isang kemikal na reaksyon, habang ang iba ay kumikilos bilang mga channel ng ion na nagsasara o nagbubukas sa ilalim ng impluwensya ng isang stimulus. Ang ilang mga compound ay partikular na nagbubuklod sa mga molekula ng mensahero sa loob ng cell.
Mga polypeptide ng motor
Mayroong isang buong klase ng mga protina na nagbibigay ng paggalaw sa katawan. Ang mga protina ng motor ay kasangkot sa pag-urong ng kalamnan, paggalaw ng cell, at aktibidad ng flagella at cilia. Nagbibigay din sila ng direksyon at aktibong transportasyon. Ang mga kinesin at dynein ay nagdadala ng mga molekula kasama ang mga microtubule gamit ang ATP hydrolysis bilang isang mapagkukunan ng enerhiya. Ang huli ay naglilipat ng mga organel at iba pang elemento patungo sa sentrosom mula sa mga peripheral na cellular na lugar. Ang mga kinesin ay gumagalaw sa kabaligtaran na direksyon. Ang mga dynein ay responsable din para sa aktibidad ng flagella at cilia.
