§ 2. اجزای اصلی یک سلول یوکاریوتی
سلول های یوکاریوتی (شکل 8 و 9) بسیار پیچیده تر از سلول های پروکاریوتی سازماندهی شده اند. آنها از نظر اندازه (از چند میکرومتر تا چند سانتی متر)، شکل و ویژگی های ساختاری بسیار متنوع هستند (شکل 10).
برنج. 8. ساختار یک سلول یوکاریوتی. طرح تعمیم یافته
برنج. 9. ساختار سلول بر اساس میکروسکوپ الکترونی
برنج. 10. سلول های یوکاریوتی مختلف: 1 – اپیتلیال. 2 - خون (e - اریترویت، / - لوکیت)؛ 3 – غضروف 4 - استخوان ها 5- عضله صاف 6- بافت همبند 7- سلول های عصبی 8- فیبر ماهیچه ای مخطط
با این حال، سازماندهی کلی و حضور اجزای اساسی در تمام سلول های یوکاریوتی یکسان است (شکل 11).
برنج. 11. سلول یوکاریوتی (نمودار)
پلاسمالما (غشای سلولی خارجی).اساس غشای پلاسمایی مانند سایر غشاهای سلولی (مثلاً میتوکندری، پلاستیدها و غیره) لایه ای از لیپیدها است که دارای دو ردیف مولکول است (شکل 12). از آنجایی که مولکول های چربی قطبی هستند (یک قطب آبدوست است، یعنی توسط آب جذب می شود، و دیگری آبگریز است، یعنی توسط آب دفع می شود)، آنها به ترتیب خاصی مرتب شده اند. انتهای آبدوست مولکول های یک لایه به سمت محیط آبی - به داخل سیتوپلاسم سلول و لایه دیگر - به سمت خارج از سلول - به سمت ماده بین سلولی (در موجودات چند سلولی) یا محیط آبی (در موجودات تک سلولی) هدایت می شود. ).
برنج. 12. ساختار غشای سلولی طبق مدل موزاییک سیال. پروتئین ها و گلیکوپروتئین ها در یک لایه دوگانه از مولکول های چربی غوطه ور می شوند که انتهای آبدوست (دایره ها) آنها به سمت بیرون و انتهای آبگریز (خطوط مواج) آنها به اعماق غشا است.
مولکول های پروتئین به صورت موزاییکی در یک لایه دو مولکولی از لیپیدها قرار می گیرند. در قسمت بیرونی سلول حیوانی، مولکول های پلی ساکارید به لیپیدها و مولکول های پروتئین پلاسمالما متصل می شوند و گلیکولیپیدها و گلیکوپروتئین ها را تشکیل می دهند.
این کل یک لایه را تشکیل می دهد گلیکوکالیکسبا او در ارتباط است عملکرد گیرندهغشاهای پلاسما (به زیر مراجعه کنید)؛ همچنین می تواند مواد مختلفی را که سلول استفاده می کند، جمع کند. علاوه بر این، گلیکوکالیکس پایداری مکانیکی پلاسمالما را افزایش می دهد.
در سلول های گیاهان و قارچ ها نیز دیواره سلولی وجود دارد که نقش حمایتی و محافظتی را ایفا می کند. در گیاهان از سلولز و در قارچ ها از کیتین تشکیل شده است.
غشای سلول خارجی تعدادی از وظایف را انجام می دهد، از جمله:
♦ مکانیکی(حمایت کننده، فرم سازی)؛
♦ مانع-حمل و نقل(نفوذ پذیری انتخابی به مواد مختلف: ورود مواد ضروری به سلول و حذف مواد غیر ضروری و مضر).
♦ گیرنده(تعیین مواد شیمیایی مختلف که در نزدیکی سلول یافت می شوند؛ درک سیگنال ها به شکل هورمون ها؛ تشخیص پروتئین "خارجی" توسط سلول های سیستم ایمنی بدن و غیره).
تبادل مواد بین سلول و محیط به روش های مختلف - غیرفعال و فعال - انجام می شود.
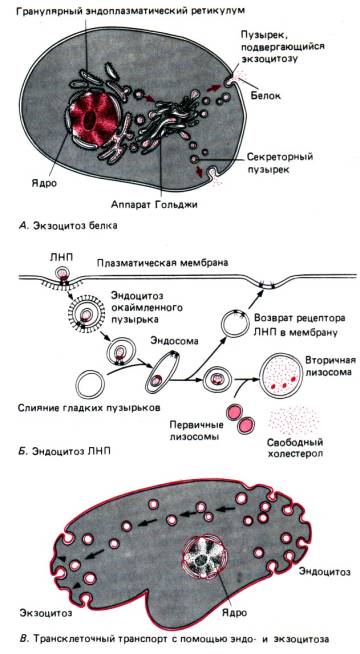
مولکولهای آب و یونهای مختلف بهطور غیرفعال (به دلیل انتشار، اسمز)، بدون اینکه سلول انرژی مصرف کند، از طریق منافذ ویژه وارد میشوند. حمل و نقل غیرفعالماکرومولکول هایی مانند پروتئین ها، پلی ساکاریدها، حتی سلول های کامل، توسط فاگوسیتوزو پینوسیتوزبا مصرف انرژی – حمل و نقل فعال
با فاگوسیتوز، سلولهای کامل یا ذرات بزرگ غرق میشوند (مثلاً به تغذیه در آمیب یا فاگوسیتوز توسط سلولهای خونی محافظ باکتریها فکر کنید). در طی پینوسیتوز، ذرات کوچک یا قطرات یک ماده مایع جذب می شوند. مشترک هر دو فرآیند این است که مواد جذب شده توسط یک غشای بیرونی در حال تهویه احاطه شده و یک واکوئل را تشکیل می دهند که سپس به اعماق سیتوپلاسم سلول حرکت می کند.
اگزوسیتوز فرآیندی است (که همچنین انتقال فعال است) در جهت مخالف فاگوسیتوز و پینوسیتوز (شکل 13). با کمک آن می توان غذای هضم نشده در تک یاخته ها یا مواد فعال بیولوژیکی تشکیل شده در سلول ترشحی را حذف کرد.
سیتوپلاسم.سیتوپلاسم محتویات سلول است که توسط پلاسمالما محدود می شود، به استثنای هسته. آن شامل ماده زمین (هیالوپلاسم)، اندامک هاو اجزاء
هیالوپلاسما- مایع چسبناکی که قادر به قرار گرفتن در حالت هر کدام است زولا(مایع)، یا ژل(ژله مانند).
در صورت لزوم، سیتوپلاسم قادر به انتقال برگشت پذیر از یک حالت به حالت دیگر است. به عنوان مثال، در طول حرکت آمیبوئید (بخش "پروتوزوا" را از درس جانورشناسی به خاطر بسپارید)، در طول تشکیل شبه پاد، انتقال سریع سیتوپلاسم از ژل به سل و بالعکس رخ می دهد. این به دلیل وجود تعداد زیادی مولکول پروتئین رشته ای در سیتوپلاسم است اکتیناهنگامی که آنها به یکدیگر متصل می شوند و یک شبکه سه بعدی تشکیل می دهند، سیتوپلاسم در حالت ژل قرار می گیرد و هنگامی که شبکه متلاشی می شود، در حالت سل قرار می گیرد.
هیالوپلاسم حاوی مواد مختلفی است - آنزیم ها، پروتئین ها، کربوهیدرات ها، چربی ها و غیره، آلی و معدنی. فرآیندهای شیمیایی مختلفی در اینجا اتفاق می افتد - تجزیه مواد، سنتز و تغییرات آنها (تغییر).
ارگانوئیدهااینها اجزای دائمی یک سلول با ساختار و عملکرد خاص هستند که در سیتوپلاسم آن قرار دارند. در ادامه در مورد آن صحبت خواهیم کرد ارگانوئیدهای عمومی،ذاتی در تمام انواع سلول های همه یوکاریوت ها. آنها با اطمینان از عملکردهای حیاتی دومی مرتبط هستند. ارگانوئیدهای خاصفقط در سلول های یک نوع خاص (بسیار تخصصی) یافت می شود - به عنوان مثال، میوفیبریل ها در سلول های عضلانی.
اندامک های هدف کلی بدون توجه به اینکه به کدام سلول و کدام موجودات تعلق دارند، ساختار یکسانی دارند. اما در میان آنها گروه هایی با غشاء وجود دارد (شبکه آندوپلاسمی، دستگاه گلژی، میتوکندری، پلاستیدها، لیزوزوم ها، واکوئل ها)و همچنین غیر غشایی ( ریبوزوم، مرکز سلول)ساختار
شبکه آندوپلاسمی (ER). EPS از غشاها تشکیل شده و یک سیستم منشعب پیچیده از لوله ها و مخازن است که به کل سیتوپلاسم سلول نفوذ می کند (شکل 14). دو نوع EPS وجود دارد - خشنو صاف.ریبوزوم ها به غشاهای ناهموار (از سمت سیتوپلاسمی) متصل هستند، اما روی غشاهای صاف وجود ندارند.
برنج. 14. شبکه آندوپلاسمی
شبکه آندوپلاسمی تعدادی عملکرد مهم را در یک سلول یوکاریوتی انجام می دهد:
♦ تحدید(تقسیم حجم داخلی سلول به فضاهای مختلف واکنش)؛
♦ مشارکت در سنتز مواد آلی(ریبوزوم ها روی غشاهای ER خشن قرار دارند و مجتمع های آنزیمی روی آن های صاف قرار دارند و از سنتز لیپیدها ، کربوهیدرات ها و غیره اطمینان می دهند).
♦ مشارکت در تشکیل عناصر دستگاه گلژی، لیزوزوم ها؛
♦ حمل و نقل مواد
دستگاه گلژی.دستگاه گلژی (AG) یک سیستم است تانک ها(واکوئل های مسطح) و حباب ها(وزیکول ها)، که در مجاورت هسته سلول قرار دارند، که به دلیل EPS در نتیجه جدا شدن قطعات کوچک آن تشکیل می شوند (شکل 15). هنگامی که این قطعات با هم ادغام می شوند، مخازن جدیدی از دستگاه گلژی به وجود می آیند، در حالی که مواد مختلفی از EPS منتقل می شوند که در مونتاژ ترکیبات آلی پیچیده (پروتئین ها + کربوهیدرات ها، پروتئین ها + لیپیدها و غیره) نقش دارند که با حذف آن ها از EPS خارج می شوند. کمک AG در خارج از سلول. این مواد فعال بیولوژیکی یا از سلول حذف می شوند (با استفاده از واکوئل های ترشحی توسط اگزوسیتوز) یا بخشی از لیزوزوم هستند (به زیر مراجعه کنید) که توسط AG تشکیل شده است.
برنج. 15. دستگاه گلژی:
دستگاه گلژی وظایف زیر را انجام می دهد:
♦ سنتزمواد فعال بیولوژیکی تولید شده توسط سلول؛
♦ ترشح (حذف از سلول) مواد مختلف(هورمون ها، آنزیم ها، موادی که دیواره سلولی از آنها ساخته شده و غیره)؛
♦ مشارکت در تشکیل لیزوزوم ها
میتوکندری.همه انواع سلول های یوکاریوتی دارای میتوکندری هستند (شکل 16). آنها مانند بدنه های گرد یا میله ها هستند، کمتر اوقات - موضوعات. اندازه آنها بین 1 تا 7 میکرون است. تعداد میتوکندری ها در یک سلول از چند صد تا ده ها هزار (در تک یاخته های بزرگ) متغیر است.
برنج. 16. میتوکندری. بالا - میتوکندری (a) در مجاری ادرار، قابل مشاهده در میکروسکوپ نوری. در زیر یک مدل سه بعدی از سازمان دهی میتوکندری آمده است: 1 – cristae; 2 - غشای خارجی؛ 3 - غشای داخلی 4 - ماتریس
میتوکندری توسط دو غشاء تشکیل شده است - خارجیو درونی؛ داخلی،که بین آن قرار دارد فضای بین غشاییغشای داخلی بسیاری از فرورفتگی ها را تشکیل می دهد - cristae که یا صفحات یا لوله هستند. این سازمان ناحیه عظیمی از غشای داخلی را فراهم می کند. حاوی آنزیم هایی است که تبدیل انرژی موجود در مواد آلی (کربوهیدرات ها، لیپیدها) را به انرژی ATP که برای زندگی سلول ضروری است، تضمین می کند. بنابراین، وظیفه میتوکندری شرکت در آن است انرژیفرآیندهای سلولی به همین دلیل است که تعداد زیادی میتوکندری ذاتی هستند، به عنوان مثال، در سلول های عضلانی که کارهای زیادی انجام می دهند.
پلاستیدهادر سلول های گیاهی اندامک های خاصی یافت می شود - پلاستیدها که اغلب شکل دوکی شکل یا گرد دارند، گاهی اوقات پیچیده تر. سه نوع پلاستید وجود دارد - کلروپلاست (شکل 17)، کروموپلاست و لوکوپلست.
کلروپلاست هادر رنگ سبز متفاوت است که به دلیل رنگدانه است - کلروفیل،تضمین فرآیند فتوسنتز،به عنوان مثال، سنتز مواد آلی از آب (H2O) و دی اکسید کربن (CO2) با استفاده از انرژی نور خورشید. کلروپلاست ها عمدتاً در سلول های برگ (در گیاهان عالی) یافت می شوند. آنها توسط دو غشاء تشکیل شده اند که به موازات یکدیگر قرار دارند و محتویات کلروپلاست را احاطه کرده اند - استروماغشای داخلی کیسه های مسطح متعددی را تشکیل می دهد - تیلاکوئیدها،که روی هم چیده شده اند (مانند یک پشته سکه) – دانه ها -و در استروما دراز بکشید. این تیلاکوئیدها هستند که حاوی کلروفیل هستند.
کروموپلاست هاتعیین رنگ زرد، نارنجی و قرمز بسیاری از گل ها و میوه ها که در سلول های آنها به مقدار زیاد وجود دارد. رنگدانه های اصلی در ترکیب آنها هستند کاروتن هاهدف عملکردی کروموپلاست ها جذب حیوانات با رنگ، تضمین گرده افشانی گل ها و پراکندگی دانه ها است.
برنج. 17. پلاستیدها: الف - کلروپلاستها در سلولهای برگ Elodea که در میکروسکوپ نوری قابل مشاهده هستند. ب - نمودار ساختار داخلی کلروپلاست با گرانا که پشته هایی از کیسه های مسطح هستند که عمود بر سطح کلروپلاست قرار دارند. ج - یک نمودار دقیق تر که لوله های آناستوموز کننده را نشان می دهد که اتاقک های فن جداگانه را به هم متصل می کند
لکوپلاست هاپلاستیدهای بی رنگ موجود در سلول های قسمت های زیرزمینی گیاهان (به عنوان مثال در غده های سیب زمینی)، دانه ها و هسته ساقه ها هستند. در لوکوپلاست ها، نشاسته عمدتا از گلوکز تشکیل شده و در اندام های ذخیره گیاهان تجمع می یابد.
پلاستیدهای یک نوع می توانند به نوع دیگر تبدیل شوند. به عنوان مثال، هنگامی که برگ ها در پاییز تغییر رنگ می دهند، کلروپلاست ها به کروموپلاست تبدیل می شوند.
لیزوزوم هااین اندامک ها شبیه وزیکول هایی هستند که توسط غشایی با قطر تا 2 میکرون احاطه شده اند. آنها حاوی چندین آنزیم هستند که پروتئین ها، اسیدهای نوکلئیک، پلی ساکاریدها و لیپیدها را تجزیه می کنند. عملکرد لیزوزوم ها شرکت در فرآیندهای تجزیه درون سلولی ترکیبات آلی پیچیده (به عنوان مثال، مواد مغذی یا مواد اجزای سلولی "صرف شده") است. لیزوزوم ها با واکوئل های فاگوسیتیک (یا پینوسیتیک) ترکیب می شوند و یک واکوئل گوارشی تشکیل می دهند.
تشکیل لیزوزوم ها به دلیل جوانه زدن از مخزن دستگاه گلژی رخ می دهد.
ریبوزوم هاریبوزوم ها (شکل 18) در سلول های یوکاریوت ها و پروکاریوت ها وجود دارند، زیرا آنها عملکرد مهمی را در سلول ها انجام می دهند. بیوسنتز پروتئین(به فصل 5 مراجعه کنید). هر سلول حاوی ده ها، صدها هزار (تا چند میلیون) از این اندامک های گرد کوچک است.
برنج. 18. طرح ساختار یک ریبوزوم نشسته بر روی غشای شبکه آندوپلاسمی: 1 - زیر واحد کوچک. 2 - tRNA؛ 3 - aminoacyl-tRNA. 4 - اسید آمینه 5 - زیر واحد بزرگ؛ 6 – غشای شبکه آندوپلاسمی؛ 7- زنجیره پلی پپتیدی سنتز شده
ریبوزوم از دو زیر واحد (قسمت) نابرابر تشکیل شده است. آنها به طور جداگانه تشکیل می شوند و در طول سنتز یک مولکول پروتئین، RNA پیام رسان "دربرگیرنده" ترکیب می شوند. ریبوزوم ها حاوی پروتئین های مختلف و RNA های ریبوزومی هستند.
اجزای سلولیاین نامی است که به اجزای غیر دائمی در سلول داده می شود که در ماده اصلی سیتوپلاسم به شکل دانه ها، دانه ها یا قطرات وجود دارند. آخال ها ممکن است توسط یک غشاء احاطه شوند یا نباشند.
از نظر عملکردی، سه نوع گنجاندن وجود دارد: ذخیره مواد مغذی(نشاسته، گلیکوژن، چربی، پروتئین)، آخال های ترشحی(مواد مشخصه سلول های غده ای تولید شده توسط آنها - هورمون های غدد درون ریز و غیره) و گنجاندن هدف ویژه(در سلول های بسیار تخصصی، به عنوان مثال هموگلوبین در گلبول های قرمز).
§ 3. سازمان هسته سلول. کروموزوم ها
هسته سلول (نگاه کنید به شکل 8 و 9) از اهمیت بالایی در زندگی سلول برخوردار است، زیرا به عنوان مخزن اطلاعات ارثی موجود در کروموزوم ها عمل می کند (نگاه کنید به زیر).
هسته توسط یک پوشش هسته ای محدود شده است که محتویات آن را جدا می کند (کاریوپلاسم)از سیتوپلاسم پوسته از دو غشا تشکیل شده است که با یک شکاف از هم جدا شده اند. هر دوی آنها دارای منافذ متعددی هستند که به لطف آنها تبادل مواد بین هسته و سیتوپلاسم امکان پذیر است. در هسته سلولی اکثر یوکاریوت ها از 1 تا 7 وجود دارد هسته هافرآیندهای سنتز RNA و tRNA با آنها همراه است.
اجزای اصلی هسته – کروموزوم ها،از یک مولکول DNA و پروتئین های مختلف تشکیل شده است. در یک میکروسکوپ نوری آنها فقط در طول دوره تقسیم سلولی به وضوح قابل مشاهده هستند (میتوز، میوز).در یک سلول غیرقابل تقسیم، کروموزوم ها مانند رشته های نازک طولانی هستند که در کل حجم هسته توزیع شده اند.
در طول تقسیم سلولی، رشته های کروموزومی مارپیچ های متراکمی را تشکیل می دهند که در نتیجه آنها (با استفاده از یک میکروسکوپ معمولی) به شکل میله ها، "گیره مو" قابل مشاهده می شوند. کل حجم اطلاعات ژنتیکی بین کروموزوم های هسته توزیع می شود. در فرآیند مطالعه آنها، الگوهای زیر شناسایی شدند:
♦ در هسته سلول های جسمی (یعنی سلول های بدن، غیرجنسی) همه افراد یک گونه حاوی تعداد مشابهی از کروموزوم های تشکیل دهنده هستند. مجموعه ای از کروموزوم ها(شکل 19)؛
برنج. 19. کروموزومهای گونههای مختلف گیاهان و جانوران، که در یک مقیاس نشان داده شدهاند: 1،2 – آمیب. 3،4 - دیاتومها؛ 5-8، 18،19 - جلبک سبز. 9 – فلای آگاریک; 10 - آهک؛ 11-12 - مگس سرکه; 13 - ماهی قزل آلا؛ 14 - skerda (خانواده Asteraceae)؛ 15 - گیاه از خانواده آروید. 16 – پروانه کوریدالیس; 17 - حشره از خانواده ملخ؛ 20 – حشره راهرو آب؛ 21 - حشره گل؛ 22 – آمبیستومای دوزیستان؛ 23 - آلوئه (خانواده زنبق)
♦ هر گونه با توجه به تعداد آنها با مجموعه کروموزوم خود مشخص می شود (به عنوان مثال، یک فرد دارای 46 کروموزوم، مگس مگس سرکه دارای 8 کروموزوم، کرم گرد دارای 4 کروموزوم، خرچنگ دارای 196، اسب دارای 66 کروموزوم و ذرت دارای 104 کروموزوم است).
♦ کروموزوم های موجود در هسته سلول های سوماتیک را می توان به صورت جفت دسته بندی کرد کروموزوم های همولوگبر اساس شباهت آنها (در ساختار و عملکرد)؛
♦ در هسته سلولهای زاینده (گامتها)، هر جفت کروموزوم همولوگ فقط شامل یک کروموزوم است، یعنی کل مجموعه کروموزومها نصف کروموزومهای سوماتیک است.
♦ مجموعه ای از کروموزوم ها در سلول های زایا نامیده می شود هاپلوئیدو با حرف n مشخص می شود و در جسمی - دیپلوئید(2n).
از مطالب فوق واضح است که هر جفت کروموزوم همولوگ از اتحاد کروموزوم های پدری و مادری در طول لقاح، یعنی ادغام سلول های جنسی (گامت ها) تشکیل می شود. برعکس، در طول تشکیل سلولهای زاینده، از هر جفت کروموزوم همولوگ، تنها یکی وارد گامت میشود.
کروموزوم ها ناهمسانجفت های همولوگ در اندازه و شکل متفاوت هستند (شکل 20 و 21).
برنج. 20. ساختار و انواع کروموزومها: الف – ظاهر 1 – سانترومر; 2 - شانه کوتاه 3 - شانه بلند)؛ ساختار داخلی همان کروموزوم (1 - سانترومر؛ - مولکولهای DNA). ج – انواع کروموزوم ها (1 – تک بازو، بازوهای متفاوت؛ 3 – بازوهای مساوی: X – بازو، V – سانترومر)
برنج. 21. یک کروموزوم از DNA و پروتئین تشکیل شده است. مولکول DNA همانند سازی می شود. دو مارپیچ دوگانه DNA یکسان در ناحیه سانترومر متصل می مانند. این کپی ها بعداً در طی تقسیم سلولی به کروموزوم های جداگانه تبدیل می شوند
در بدن کروموزوم ها وجود دارد انقباض اولیه (به نام سانترومر)،که نخ ها به آن وصل شده اند دوک های شکافتکروموزوم را به دو قسمت تقسیم می کند شانهکروموزوم ها می توانند با بازو مساوی، نابرابر بازو یا تک بازو باشند.
فصل 5. متابولیسم
§ 1. متابولیسم به مثابه وحدت جذب و عدم تشبیه
همه سلول ها و موجودات زنده سیستم های باز هستند، یعنی در حالت تبادل دائمی انرژی ها و مواد با محیط هستند. در طبیعت بی جان سیستم های باز وجود دارد، اما وجود آنها از نظر کیفی با موجودات زنده متفاوت است. این مثال را در نظر بگیرید: یک قطعه سوزان گوگرد بومی در حالت تبادل با محیط است. هنگامی که می سوزد، O 2 جذب می شود و SO 2 و انرژی (به شکل گرما) آزاد می شود. اما در این حالت یک قطعه گوگرد به عنوان یک جسم فیزیکی از بین می رود و ساختار اولیه خود را از دست می دهد.
برای موجودات زنده، مبادله با محیط، شرطی برای حفظ و حفظ سازمان ساختاری آنها از طریق خود نوسازی تمام مواد و اجزای تشکیل دهنده آنها است.
متابولیسم (متابولیسم) مجموعه ای از فرآیندهایی است که در موجودات زنده رخ می دهد (مصرف، تبدیل، تجمع و آزادسازی مواد و انرژی) که فعالیت حیاتی، رشد، رشد، تولیدمثل آنها را تضمین می کند. در طی فرآیند متابولیک، مولکول های تشکیل دهنده سلول ها شکسته و سنتز می شوند. تجدید ساختارهای سلولی و مواد بین سلولی.
متابولیسم بر اساس فرآیندهای به هم پیوسته است ادغام(آنابولیسم) و یکسان سازی(کاتابولیسم). در طول جذب (تبادل پلاستیک)، مواد پیچیده از مواد ساده سنتز می شوند. به لطف این است که تمام مواد آلی موجود در سلول برای ساخت اجزای ساختاری آن، سیستم های آنزیمی و غیره ایجاد می شود. جذب همیشه با صرف انرژی انجام می شود.
در طی تجزیه (متابولیسم انرژی)، مواد آلی پیچیده به سادهتر یا غیرآلی تجزیه میشوند. در این حالت انرژی آزاد می شود که توسط سلول صرف انجام فرآیندهای مختلفی می شود که فعالیت حیاتی آن را تضمین می کند (سنتز و حمل و نقل مواد، کار مکانیکی و غیره).
همه موجودات زنده را می توان به دو گروه تقسیم کرد: اتوتروف هاو هتروتروف ها،که در منابع انرژی و مواد لازم برای اطمینان از عملکرد حیاتی آنها متفاوت است.
اتوتروف ها- موجوداتی که با استفاده از انرژی نور خورشید ترکیبات آلی را از مواد معدنی سنتز می کنند (به عنوان مثال فوتوتروف ها- گیاهان، سیانوباکترها) یا انرژی حاصل از اکسیداسیون مواد معدنی (غیر آلی) (مانند شیمی تروف ها- باکتری های گوگرد، باکتری های آهن و غیره). در نتیجه، آنها قادر به ایجاد مستقل مواد مورد نیاز برای فعالیت زندگی خود هستند.
§ 2. تجزیه در موجودات بی هوازی و هوازی
موجودات زنده را می توان به دو گروه تقسیم کرد و با توجه به ماهیت تجزیه - هوازیو بی هوازی هاهوازی (از یونانی. دیو- هوا) برای زندگی به اکسیژن رایگان نیاز دارند. در بی هوازی ها (یونانی. ac– ذره منفی) لازم نیست. در آنها، تجزیه با تخمیر انجام می شود - بدون اکسیژن، تجزیه آنزیمی مواد آلی با تشکیل مواد آلی ساده تر و آزاد شدن انرژی. مثلا:
♦ تخمیر اسید لاکتیک:
C 6 H 12 O 6 + 2H 3 PO 4 + 2ADP → 2PH + 2ATP + 2H 2 O;
♦ تخمیر الکلی:
C 6 H 12 O 6 + 2PH + 2ADP → 2C 2 H 5 OH + 2CO 2 + 2ATP + 2H 2 O.
موادی که در طی تخمیر به وجود می آیند، آلی هستند و بنابراین هنوز هم انرژی زیادی دارند.
برنج. 22. روابط بین جذب و غیر همسان سازی در موجودات اتوتروف و هتروتروف
در موجودات هوازی، در طی فرآیند تنفس در میتوکندری، تجزیه کامل مواد آلی (با استفاده از O 2) به محصولات نهایی کم انرژی CO 2 و H 2 O رخ می دهد و مقدار قابل توجهی انرژی آزاد می شود:
C 6 H 12 0 6 (گلوکز) + 0 2 > 6C0 2 + 6H 2 0 + انرژی (به دلیل آن 38 مولکول ATP سنتز می شود).
اجازه دهید در قالب طرح های تعمیم یافته متابولیسم موجودات هوازی اتوتروف و هتروتروف را در نظر بگیریم (شکل 22).
ادغام.مهمترین فرآیندهای آن هستند فتوسنتزو بیوسنتز پروتئین ها
ارتباط ارگانیسم با محیط، از دیدگاه فیزیکوشیمیایی، یک سیستم باز است، یعنی سیستمی که در آن فرآیندهای بیوشیمیایی در حال انجام است. مواد اولیه از محیط می آیند و موادی که به طور پیوسته نیز تشکیل می شوند به بیرون منتقل می شوند. تعادل بین سرعت و غلظت محصولات واکنش های چند جهته در بدن شرطی، خیالی است، زیرا مصرف و حذف مواد متوقف نمی شود. ارتباط مستمر با محیط به ما این امکان را می دهد که یک موجود زنده را به عنوان یک سیستم باز در نظر بگیریم.
برای تمام سلول های زنده، منبع انرژی خورشید است. سلول های گیاهی انرژی را از نور خورشید با کمک کلروفیل می گیرند و از آن برای واکنش های جذب در طول فرآیند فتوسنتز استفاده می کنند. سلول های حیوانات، قارچ ها و باکتری ها به طور غیرمستقیم از انرژی خورشیدی در طی تجزیه مواد آلی سنتز شده توسط گیاهان زمینی استفاده می کنند.
برخی از مواد مغذی سلول در طی تنفس سلولی شکسته می شوند، بنابراین انرژی لازم برای انواع مختلف فعالیت سلولی را تامین می کنند. این فرآیند در اندامک هایی به نام میتوکندری انجام می شود. میتوکندری از دو غشاء تشکیل شده است: غشا بیرونی که اندامک را از سیتوپلاسم جدا می کند و غشای داخلی که چین های متعددی را تشکیل می دهد. محصول اصلی تنفس ATP است. میتوکندری را ترک می کند و به عنوان منبع انرژی برای بسیاری از واکنش های شیمیایی در سیتوپلاسم و غشای سلولی استفاده می شود. اگر اکسیژن برای تنفس سلولی لازم باشد، تنفس هوازی نامیده می شود، اما اگر واکنش ها در غیاب اکسیژن اتفاق بیفتد، آنگاه از تنفس بی هوازی صحبت می کنیم.
برای هر نوع کاری که در یک سلول انجام می شود، انرژی به یک شکل - به شکل انرژی از پیوندهای فسفات ATP - استفاده می شود. ATP یک ترکیب آسان متحرک است. تشکیل ATP در غشای داخلی میتوکندری اتفاق می افتد. ATP در تمام سلول ها در طول تنفس به دلیل انرژی اکسیداسیون کربوهیدرات ها، چربی ها و سایر مواد آلی سنتز می شود. در سلول های گیاهی سبز، مقدار اصلی ATP در کلروپلاست ها به دلیل انرژی خورشیدی سنتز می شود. در طول فتوسنتز، آنها چندین برابر بیشتر از میتوکندری ATP تولید می کنند. ATP با پاره شدن پیوندهای فسفر-اکسیژن و آزاد شدن انرژی تجزیه می شود. این تحت عمل آنزیم ATPase در طول هیدرولیز ATP رخ می دهد - افزودن آب با حذف یک مولکول اسید فسفریک. در نتیجه، ATP به ADP تبدیل میشود و اگر دو مولکول اسید فسفریک از هم جدا شوند، به AMP تبدیل میشوند. واکنش حذف هر گرم مولکول اسید با آزاد شدن 40 کیلوژول همراه است. این یک خروجی انرژی بسیار بزرگ است، به همین دلیل است که پیوندهای فسفر-اکسیژن ATP معمولاً ماکرو انرژی (پر انرژی) نامیده می شوند.
استفاده از ATP در واکنش های تبادل پلاستیک با جفت کردن آنها با هیدرولیز ATP انجام می شود. مولکول های مواد مختلف با اتصال گروه فسفر آزاد شده در طول هیدرولیز از مولکول ATP، یعنی با فسفوریلاسیون، با انرژی شارژ می شوند.
ویژگی مشتقات فسفات این است که آنها نمی توانند سلول را ترک کنند، اگرچه اشکال "تخلیه شده" آنها آزادانه از غشاء عبور می کند. به لطف این، مولکول های فسفریله در سلول باقی می مانند تا زمانی که در واکنش های مناسب مورد استفاده قرار گیرند.
فرآیند معکوس تبدیل ADP به ATP با افزودن یک مولکول اسید فسفریک به ADP، آزاد شدن آب و جذب مقدار زیادی انرژی اتفاق میافتد.
بنابراین، ATP یک منبع جهانی و مستقیم انرژی برای فعالیت سلولی است. این یک صندوق سلولی واحد از انرژی ایجاد می کند و توزیع مجدد و انتقال آن را از یک ناحیه سلول به ناحیه دیگر امکان پذیر می کند.
انتقال گروه فسفات نقش مهمی در واکنش های شیمیایی مانند جمع آوری ماکرومولکول ها از مونومرها دارد. به عنوان مثال، اسیدهای آمینه را می توان تنها پس از فسفریله شدن به پپتیدها ترکیب کرد. فرآیندهای مکانیکی انقباض یا حرکت، انتقال یک ماده محلول در برابر گرادیان غلظت و سایر فرآیندها شامل مصرف انرژی ذخیره شده در ATP است.
فرآیند متابولیسم انرژی را می توان به صورت زیر نشان داد. مواد آلی با مولکولی بالا در سیتوپلاسم به طور آنزیمی، با هیدرولیز، به مواد سادهتری تبدیل میشوند که از آنها تشکیل میشوند: پروتئینها - به اسیدهای آمینه، پلی و دی ساکاریدها - به مونوساکاریدها (+ گلوکز)، چربیها به گلیسرول و اسیدهای چرب. هیچ فرآیند اکسیداتیو وجود ندارد، انرژی کمی آزاد می شود، که استفاده نمی شود و به شکل حرارتی می رود. اکثر سلول ها ابتدا از کربوهیدرات ها استفاده می کنند. پلی ساکاریدها (نشاسته در گیاهان و گلیکوژن در حیوانات) به گلوکز هیدرولیز می شوند. اکسیداسیون گلوکز در سه مرحله انجام می شود: گلیکولیز، دکربوکسیلاسیون اکسیداتیو (چرخه کربس - چرخه اسید سیتریک) و فسفوریلاسیون اکسیداتیو (زنجیره تنفسی). گلیکولیز که در نتیجه آن یک مولکول گلوکز با آزاد شدن دو مولکول ATP به دو مولکول اسید پیروویک تقسیم می شود، در سیتوپلاسم رخ می دهد. در غیاب اکسیژن، اسید پیروویک یا به اتانول (تخمیر) یا اسید لاکتیک (تنفس بی هوازی) تبدیل می شود.
هنگامی که گلیکولیز در سلول های حیوانی اتفاق می افتد، مولکول شش کربنه گلوکز به دو مولکول اسید لاکتیک تجزیه می شود. این فرآیند چند مرحله ای است. این به طور متوالی توسط 13 آنزیم انجام می شود. در طی تخمیر الکلی، دو مولکول اتانول و دو مولکول CO2 از یک مولکول گلوکز تشکیل می شوند.
گلیکولیز یک فاز مشترک در تنفس بی هوازی و هوازی است. فرآیند اکسیداسیون بدون اکسیژن که در آن تنها بخشی از انرژی متابولیت ها آزاد می شود و مورد استفاده قرار می گیرد، برای موجودات بی هوازی نهایی است. در حضور اکسیژن، اسید پیروویک به میتوکندری می رود، جایی که در نتیجه تعدادی از واکنش های متوالی، به طور کامل هوازی به H2O و CO2 با فسفوریلاسیون همزمان ADP به ATP اکسید می شود. در این حالت، دو مولکول ATP توسط گلیکولیز، دو مولکول توسط چرخه کربس و 34 مولکول توسط زنجیره تنفسی تولید می شوند. بازده خالص برای اکسیداسیون کامل یک مولکول گلوکز به H2O و CO2 38 مولکول است.
بنابراین، در موجودات هوازی، تجزیه نهایی مواد آلی با اکسید کردن آنها با اکسیژن اتمسفر به مواد معدنی ساده انجام می شود: CO2 و H2O. این فرآیند بر روی کریستای میتوکندری انجام می شود. در این حالت حداکثر انرژی آزاد آزاد می شود که بخش قابل توجهی از آن در مولکول های ATP ذخیره می شود. به راحتی می توان دریافت که اکسیداسیون هوازی انرژی آزاد را تا حد زیادی به سلول می دهد.
در نتیجه کاتابولیسم، مولکول های غنی از انرژی ATP در سلول جمع می شوند و CO2 و آب اضافی در محیط خارجی آزاد می شوند.
مولکول های قندی که برای تنفس مورد نیاز نیستند را می توان در سلول ذخیره کرد. لیپیدهای اضافی یا تجزیه می شوند و پس از آن محصولات تجزیه آنها به عنوان بستری برای تنفس وارد میتوکندری می شود یا به صورت ذخایر به شکل قطرات چربی در سیتوپلاسم رسوب می کند. پروتئین ها از اسیدهای آمینه وارد شده به سلول ساخته می شوند. سنتز پروتئین در اندامک هایی به نام ریبوزوم اتفاق می افتد. هر ریبوزوم از دو ذره فرعی تشکیل شده است - بزرگ و کوچک: هر دو ذره فرعی شامل مولکول های پروتئین و مولکول های RNA هستند.
ریبوزوم ها اغلب به یک سیستم غشایی خاص متشکل از مخازن و وزیکول متصل می شوند - به اصطلاح شبکه آندوپلاسمی (ER). در سلول هایی که پروتئین زیادی تولید می کنند، شبکه آندوپلاسمی اغلب به خوبی توسعه یافته و با ریبوزوم پوشیده شده است. برخی از آنزیم ها تنها زمانی موثر هستند که به غشاء متصل شوند. بیشتر آنزیم های دخیل در سنتز لیپیدها در اینجا قرار دارند. بنابراین، شبکه آندوپلاسمی مانند نوعی میز کار سلولی است.
علاوه بر این، ER سیتوپلاسم را به بخشهای جداگانه تقسیم میکند، یعنی فرآیندهای شیمیایی مختلفی را که به طور همزمان در سیتوپلاسم اتفاق میافتند جدا میکند و در نتیجه احتمال تداخل این فرآیندها با یکدیگر را کاهش میدهد.
محصولات تولید شده توسط یک سلول معین اغلب در خارج از سلول استفاده می شوند. در چنین مواردی، پروتئینهای سنتز شده روی ریبوزومها از غشاهای شبکه آندوپلاسمی عبور میکنند و به وزیکولهای غشایی که در اطراف آنها تشکیل میشوند بستهبندی میشوند و سپس از ER جدا میشوند. این وزیکول ها که به صورت مسطح و روی هم چیده شده اند، مانند پنکیک های چیده شده، ساختار مشخصی به نام مجموعه گلژی یا دستگاه گلژی را تشکیل می دهند. در طول اقامت خود در دستگاه گلژی، پروتئین ها دستخوش تغییرات خاصی می شوند. هنگامی که زمان خروج آنها از سلول فرا می رسد، وزیکول های غشایی با غشای سلولی ادغام می شوند و تخلیه می شوند و محتویات آنها بیرون می ریزد، یعنی ترشح با اگزوسیتوز رخ می دهد.
دستگاه گلژی همچنین لیزوزوم ها - کیسه های غشایی حاوی آنزیم های گوارشی را تولید می کند. یافتن اینکه یک سلول چگونه پروتئین های خاصی را می سازد، بسته بندی و صادر می کند، و اینکه چگونه می داند کدام پروتئین ها را باید برای خود نگه دارد، یکی از جذاب ترین شاخه های سیتولوژی مدرن است.
غشاهای هر سلول دائما در حال حرکت و تغییر هستند. غشاهای ER به آرامی در سراسر سلول حرکت می کنند. بخش های منفرد این غشاها جدا شده و وزیکول هایی را تشکیل می دهند که به طور موقت بخشی از دستگاه گلژی می شوند و سپس از طریق فرآیند اگزوسیتوز با غشای سلولی ادغام می شوند.
بعداً، مواد غشایی به سیتوپلاسم بازگردانده می شود و در آنجا دوباره استفاده می شود.
تبادل مواد وارد شده به سلول یا آزاد شده توسط آن در خارج و همچنین تبادل سیگنال های مختلف با محیط میکرو و کلان از طریق غشای بیرونی سلول انجام می شود. همانطور که مشخص است، غشای سلولی یک لایه دولایه لیپیدی است که در آن مولکول های پروتئینی مختلف تعبیه شده است که به عنوان گیرنده های تخصصی، کانال های یونی، دستگاه هایی که به طور فعال مواد شیمیایی مختلف را انتقال یا حذف می کنند، تماس های بین سلولی و غیره عمل می کنند. غشاء به طور نامتقارن: سطح بیرونی از اسفنگومیلین و فسفاتیدیل کولین، سطح داخلی - از فسفاتیدیل سرین و فسفاتیدیل اتانول آمین تشکیل شده است. حفظ چنین عدم تقارن مستلزم صرف انرژی است. بنابراین، در صورت آسیب سلولی، عفونت یا گرسنگی انرژی، سطح بیرونی غشاء با فسفولیپیدهای غیرعادی برای آن غنی می شود که با واکنشی مشابه به آن، سیگنالی برای سایر سلول ها و آنزیم ها در مورد آسیب سلولی می شود. مهمترین نقش را شکل محلول فسفولیپاز A2 ایفا می کند که اسید آراشیدونیک را تجزیه کرده و از فسفولیپیدهای فوق الذکر لیزوفرم ایجاد می کند. اسید آراشیدونیک پیوند محدود کننده ای برای ایجاد واسطه های التهابی مانند ایکوزانوئیدها است و مولکول های محافظ - پنتراکسین ها (پروتئین واکنشی C (CRP)، پیش سازهای پروتئین های آمیلوئید) - به لیزوفرم ها در غشاء متصل می شوند و به دنبال آن مکمل فعال می شود. سیستم در امتداد مسیر کلاسیک و تخریب سلولی.
ساختار غشاء به حفظ ویژگی های محیط داخلی سلول، تفاوت های آن با محیط خارجی کمک می کند. این امر با نفوذ پذیری انتخابی غشای سلولی و وجود مکانیسم های انتقال فعال در آن تضمین می شود. اختلال آنها در نتیجه آسیب مستقیم، به عنوان مثال، توسط تترودوتوکسین، اوابین، تترا اتیل آمونیوم، یا در صورت تامین انرژی ناکافی به "پمپ" مربوطه منجر به اختلال در ترکیب الکترولیت سلول، تغییر در متابولیسم آن، اختلال می شود. از توابع خاص - انقباض، هدایت تکانه های تحریک، و غیره. اختلال در کانال های یونی سلولی (کلسیم، سدیم، پتاسیم و کلرید) در انسان نیز می تواند از نظر ژنتیکی توسط جهش در ژن های مسئول ساختار این کانال ها تعیین شود. به اصطلاح کانالوپاتی باعث بیماری های ارثی سیستم عصبی، عضلانی و گوارشی می شود. ورود بیش از حد آب به سلول می تواند منجر به پارگی آن - سیتولیز - به دلیل سوراخ شدن غشاء در هنگام فعال شدن مکمل یا حمله لنفوسیت های سیتوتوکسیک و سلول های کشنده طبیعی شود.
غشای سلولی گیرنده های زیادی در خود دارد - ساختارهایی که وقتی با مولکول های سیگنال دهی خاص مربوطه (لیگاندها) ترکیب می شوند، سیگنالی را در داخل سلول منتقل می کنند. این امر از طریق آبشارهای تنظیمی مختلف متشکل از مولکول های فعال آنزیمی که به طور متوالی فعال می شوند و در نهایت به اجرای برنامه های سلولی مختلف، مانند رشد و تکثیر، تمایز، تحرک، پیری و مرگ سلولی کمک می کنند، رخ می دهد. آبشارهای نظارتی بسیار زیاد هستند، اما تعداد آنها هنوز به طور کامل مشخص نشده است. سیستم گیرنده ها و آبشارهای تنظیمی مرتبط با آنها نیز در داخل سلول وجود دارد. آنها یک شبکه تنظیمی خاص با نقاط تمرکز، توزیع و انتخاب مسیر سیگنال بعدی را بسته به وضعیت عملکردی سلول، مرحله توسعه آن و عملکرد همزمان سیگنال های گیرنده های دیگر ایجاد می کنند. نتیجه این ممکن است مهار یا تقویت سیگنال، هدایت آن در امتداد یک مسیر تنظیمی متفاوت باشد. هم دستگاه گیرنده و هم مسیرهای انتقال سیگنال از طریق آبشارهای تنظیمی، به عنوان مثال به هسته، می توانند در نتیجه یک نقص ژنتیکی که به عنوان یک نقص مادرزادی در سطح ارگانیسم یا در نتیجه یک جهش جسمی در یک سلول خاص رخ می دهد، مختل شوند. نوع این مکانیسم ها می توانند توسط عوامل عفونی، سموم آسیب ببینند و همچنین در طول فرآیند پیری تغییر کنند. مرحله نهایی این ممکن است اختلال در عملکرد سلول، فرآیندهای تکثیر و تمایز آن باشد.
در سطح سلول ها نیز مولکول هایی وجود دارد که نقش مهمی در فرآیندهای تعامل بین سلولی دارند. اینها ممکن است شامل پروتئین های چسبنده سلولی، آنتی ژن های سازگاری بافتی، آنتی ژن های خاص بافتی، تمایز دهنده و غیره باشد. تغییرات در ترکیب این مولکول ها باعث اختلال در تعاملات بین سلولی می شود و می تواند باعث فعال شدن مکانیسم های مناسب برای از بین بردن چنین سلول هایی شود، زیرا آنها یک خطر خاصی برای یکپارچگی بدن به عنوان مخزن عفونت، به ویژه ویروسی، یا به عنوان آغازگر بالقوه رشد تومور.
نقض منبع انرژی سلول
منبع انرژی در سلول غذا است که پس از تجزیه آن انرژی به مواد نهایی آزاد می شود. محل اصلی تولید انرژی میتوکندری است که در آن مواد با کمک آنزیم های زنجیره تنفسی اکسید می شوند. اکسیداسیون تامین کننده اصلی انرژی است، زیرا در نتیجه گلیکولیز، در مقایسه با اکسیداسیون، بیش از 5٪ انرژی از همان مقدار سوبستراهای اکسیداسیون (گلوکز) آزاد نمی شود. حدود 60 درصد از انرژی آزاد شده در طول اکسیداسیون توسط فسفوریلاسیون اکسیداتیو در فسفات های پرانرژی (ATP، کراتین فسفات) انباشته می شود و بقیه به صورت گرما دفع می شود. متعاقباً، فسفات های پرانرژی توسط سلول برای فرآیندهایی مانند عملکرد پمپ، سنتز، تقسیم، حرکت، ترشح و غیره استفاده می شود. سه مکانیسم وجود دارد که آسیب به آنها می تواند باعث اختلال در تامین انرژی سلول شود: مکانیسم سنتز آنزیم های متابولیسم انرژی، دوم مکانیسم فسفوریلاسیون اکسیداتیو، سوم مکانیسم استفاده از انرژی است.
اختلال در انتقال الکترون در زنجیره تنفسی میتوکندری یا جدا شدن اکسیداسیون و فسفوریلاسیون ADP با از دست دادن پتانسیل پروتون، نیروی محرکه برای تولید ATP، منجر به تضعیف فسفوریلاسیون اکسیداتیو می شود، به گونه ای که بیشتر انرژی به صورت گرما تلف می شود. تعداد ترکیبات پرانرژی کاهش می یابد. جداسازی اکسیداسیون و فسفوریلاسیون تحت تأثیر آدرنالین توسط سلول های موجودات گرمازا برای افزایش تولید گرما در حالی که دمای بدن در هنگام خنک شدن ثابت یا در هنگام تب افزایش می یابد استفاده می شود. تغییرات قابل توجهی در ساختار میتوکندری و متابولیسم انرژی در تیروتوکسیکوز مشاهده می شود. این تغییرات در ابتدا برگشت پذیر هستند، اما پس از یک نقطه معین برگشت ناپذیر می شوند: میتوکندری تکه تکه می شود، متلاشی می شود یا متورم می شود، کریستاها را از دست می دهد، تبدیل به واکوئل می شود و در نهایت موادی مانند هیالین، فریتین، کلسیم، لیپوفوسسین را جمع می کند. در بیماران مبتلا به اسکوربوت، میتوکندری ها به هم می پیوندند و کندریوسفرها را تشکیل می دهند که احتمالاً به دلیل آسیب غشاء توسط ترکیبات پراکسید است. آسیب قابل توجهی به میتوکندری تحت تأثیر تابش یونیزان در طول تبدیل یک سلول طبیعی به یک سلول بدخیم رخ می دهد.
میتوکندری ها انبار قدرتمندی از یون های کلسیم هستند که غلظت آن چندین مرتبه بیشتر از سیتوپلاسم است. هنگامی که میتوکندری آسیب می بیند، کلسیم وارد سیتوپلاسم می شود و باعث فعال شدن پروتئینازها با آسیب به ساختارهای داخل سلولی و اختلال در عملکرد سلول مربوطه می شود، به عنوان مثال، انقباضات کلسیم یا حتی "مرگ کلسیم" در نورون ها. در نتیجه اختلال در ظرفیت عملکردی میتوکندری، تشکیل ترکیبات پراکسید رادیکال آزاد به شدت افزایش می یابد، که واکنش پذیری بسیار بالایی دارند و بنابراین به اجزای مهم سلول - اسیدهای نوکلئیک، پروتئین ها و لیپیدها آسیب می رسانند. این پدیده در زمان به اصطلاح استرس اکسیداتیو مشاهده می شود و می تواند پیامدهای منفی برای وجود سلول داشته باشد. بنابراین، آسیب به غشای خارجی میتوکندری همراه با انتشار در سیتوپلاسم مواد موجود در فضای بین غشایی، در درجه اول سیتوکروم C و برخی دیگر از مواد فعال بیولوژیکی است، که باعث واکنش های زنجیره ای می شود که باعث مرگ برنامه ریزی شده سلول - آپوپتوز می شود. با آسیب رساندن به DNA میتوکندری، واکنش های رادیکال آزاد اطلاعات ژنتیکی لازم برای تشکیل آنزیم های زنجیره تنفسی خاصی را که به طور خاص در میتوکندری تولید می شوند، تحریف می کنند. این منجر به اختلال حتی بیشتر در فرآیندهای اکسیداتیو می شود. به طور کلی، دستگاه ژنتیکی خود میتوکندری، در مقایسه با دستگاه ژنتیکی هسته، کمتر از تأثیرات مضری که می تواند اطلاعات ژنتیکی رمزگذاری شده در آن را تغییر دهد، محافظت می شود. در نتیجه، اختلال عملکرد میتوکندری در طول زندگی رخ می دهد، به عنوان مثال، در طول فرآیند پیری، در طول تبدیل بدخیم سلول، و همچنین در پس زمینه بیماری های میتوکندری ارثی مرتبط با جهش DNA میتوکندری در تخمک. در حال حاضر، بیش از 50 جهش میتوکندریایی توصیف شده است که باعث بیماری های دژنراتیو ارثی سیستم عصبی و عضلانی می شود. آنها منحصراً از مادر به کودک منتقل می شوند، زیرا میتوکندری اسپرم بخشی از زیگوت و بر این اساس، ارگانیسم جدید نیست.
نقض حفظ و انتقال اطلاعات ژنتیکی
هسته سلول حاوی بیشتر اطلاعات ژنتیکی است و در نتیجه عملکرد طبیعی آن را تضمین می کند. از طریق بیان انتخابی ژن، فعالیت سلولی را در طول اینترفاز هماهنگ میکند، اطلاعات ژنتیکی را ذخیره میکند و مواد ژنتیکی را در طول تقسیم سلولی بازسازی و انتقال میدهد. همانندسازی DNA و رونویسی RNA در هسته رخ می دهد. عوامل بیماریزای مختلف مانند اشعه ماوراء بنفش و یونیزان، اکسیداسیون رادیکال های آزاد، مواد شیمیایی، ویروس ها می توانند به DNA آسیب برسانند. تخمین زده می شود که هر سلول یک حیوان خونگرم 1 روز طول می کشد. بیش از 10000 پایگاه را از دست می دهد. در اینجا باید هنگام کپی در حین تقسیم تخلفات را اضافه کنیم. اگر این آسیب ها ادامه پیدا می کرد، سلول نمی توانست زنده بماند. حفاظت در وجود سیستمهای ترمیم قدرتمند مانند اندونوکلئاز فرابنفش، سیستمهای ترمیم تکثیر و نوترکیبی است که جایگزین آسیب DNA میشوند. نقص های ژنتیکی در سیستم های ترمیم باعث ایجاد بیماری هایی می شود که در اثر افزایش حساسیت به عوامل آسیب رساندن به DNA ایجاد می شود. این گزرودرما پیگمنتوزوم و همچنین برخی از سندرم های پیری تسریع شده است که با افزایش تمایل به ایجاد تومورهای بدخیم همراه است.
سیستم تنظیم فرآیندهای همانندسازی DNA، رونویسی RNA پیام رسان (mRNA) و ترجمه اطلاعات ژنتیکی از اسیدهای نوکلئیک به ساختار پروتئین ها کاملاً پیچیده و چند سطحی است. علاوه بر آبشارهای تنظیمی که باعث فعال شدن فاکتورهای رونویسی با تعداد کل بیش از 3000 می شود و ژن های خاصی را فعال می کند، یک سیستم تنظیمی چند سطحی نیز وجود دارد که با واسطه مولکول های RNA کوچک (RNA مداخله گر؛ RNAi) انجام می شود. ژنوم انسان که از حدود 3 میلیارد باز پورین و پیریمیدین تشکیل شده است، تنها حاوی 2 درصد از ژن های ساختاری مسئول سنتز پروتئین است. بقیه، سنتز RNA های تنظیمی را فراهم می کنند، که همزمان با فاکتورهای رونویسی، کار ژن های ساختاری را در سطح DNA در کروموزوم ها فعال یا مسدود می کنند یا بر فرآیندهای ترجمه RNA پیام رسان (mRNA) در طول تشکیل یک مولکول پلی پپتیدی در کروموزوم ها تأثیر می گذارند. سیتوپلاسم نقض اطلاعات ژنتیکی می تواند هم در سطح ژن های ساختاری و هم در بخش تنظیم کننده DNA با تظاهرات مربوطه در قالب بیماری های ارثی مختلف رخ دهد.
اخیراً توجه زیادی به تغییرات در مواد ژنتیکی که در طی رشد فردی یک ارگانیسم رخ می دهد و با مهار یا فعال شدن بخش های خاصی از DNA و کروموزوم ها به دلیل متیلاسیون، استیلاسیون و فسفوریلاسیون آنها همراه است، جلب شده است. این تغییرات برای مدت طولانی و گاهی در تمام طول زندگی ارگانیسم از جنین زایی تا پیری باقی می ماند و به آن وراثت اپی ژنومیک می گویند.
از تکثیر سلول های دارای اطلاعات ژنتیکی تغییر یافته نیز توسط سیستم ها (عوامل) کنترل کننده چرخه میتوزی جلوگیری می شود. آنها با پروتئین کینازهای وابسته به سیکلین و زیرواحدهای کاتالیزوری آنها - سیکلین ها - تعامل دارند و سلول را از گذراندن چرخه کامل میتوزی مسدود می کنند و تقسیم را در مرز بین فازهای پیش سنتزی و مصنوعی (بلوک G1/S) تا پایان ترمیم DNA متوقف می کنند. و اگر این غیرممکن باشد، سلول های مرگ برنامه ریزی شده را راه اندازی می کنند. این عوامل شامل ژن p53 است که جهش آن باعث از دست دادن کنترل بر تکثیر سلول های تبدیل شده می شود. تقریباً در 50 درصد از سرطان های انسانی مشاهده می شود. دومین نقطه بازرسی چرخه میتوزی در مرز G2/M است. در اینجا، توزیع صحیح مواد کروموزومی بین سلولهای دختر در میتوز یا میوز با استفاده از مجموعهای از مکانیسمها کنترل میشود که دوک، مرکز و سانترومرها (kinetochores) سلول را کنترل میکنند. ناکارآمدی این مکانیسم ها منجر به اختلال در توزیع کروموزوم ها یا قسمت های آنها می شود که با عدم وجود کروموزوم در یکی از سلول های دختر (آنئوپلوئیدی)، وجود یک کروموزوم اضافی (پلی پلوئیدی)، جدا شدن یک کروموزوم ظاهر می شود. بخشی از یک کروموزوم (حذف) و انتقال آن به کروموزوم دیگر (انتقال). چنین فرآیندهایی اغلب در طول تکثیر سلول های بدخیم دژنره شده و تبدیل شده مشاهده می شوند. اگر این اتفاق در طول میوز با سلولهای زایا رخ دهد، یا به مرگ جنین در مراحل اولیه رشد جنینی یا تولد ارگانیسمی با بیماری کروموزومی منجر میشود.
تکثیر کنترل نشده سلولی در طول رشد تومور در نتیجه جهش در ژن هایی که تکثیر سلولی را کنترل می کنند رخ می دهد و انکوژن نامیده می شود. در میان بیش از 70 انکوژن در حال حاضر شناخته شده، اکثر آنها به اجزای تنظیم رشد سلولی تعلق دارند، برخی توسط فاکتورهای رونویسی که فعالیت ژن را تنظیم می کنند، و همچنین عواملی که تقسیم و رشد سلولی را مهار می کنند، نشان داده می شوند. یکی دیگر از عوامل محدود کننده انبساط (گسترش) بیش از حد سلول های در حال تکثیر کوتاه شدن انتهای کروموزوم ها - تلومرها است که در نتیجه برهمکنش فضایی صرفاً قادر به تکثیر کامل نیستند، بنابراین، پس از هر تقسیم سلولی، تلومرها به وسیله آنها کوتاه می شوند. قسمت خاصی از پایه ها بنابراین، سلولهای در حال تکثیر یک ارگانیسم بالغ پس از تعداد معینی تقسیم (معمولاً بین 20 تا 100، بسته به نوع ارگانیسم و سن آن) طول تلومر را خسته میکنند و همانندسازی بیشتر کروموزوم متوقف میشود. این پدیده در اپیتلیوم اسپرم، انتروسیت ها و سلول های جنینی به دلیل وجود آنزیم تلومراز که طول تلومر را پس از هر تقسیم ترمیم می کند، رخ نمی دهد. در اکثر سلول های موجودات بالغ، تلومراز مسدود شده است، اما، متأسفانه، در سلول های تومور فعال می شود.
ارتباط بین هسته و سیتوپلاسم و انتقال مواد در هر دو جهت از طریق منافذ در غشای هسته با مشارکت سیستم های حمل و نقل ویژه ای که انرژی مصرف می کنند انجام می شود. به این ترتیب انرژی و مواد پلاستیکی، مولکول های سیگنال دهنده (عوامل رونویسی) به هسته منتقل می شوند. جریان معکوس، مولکولهای mRNA و انتقال RNA (tRNA)، ریبوزومهای لازم برای سنتز پروتئین در سلول را به سیتوپلاسم میبرد. همان مسیر انتقال مواد در ویروس ها، به ویژه مانند HIV نیز ذاتی است. آنها مواد ژنتیکی خود را با ادغام بیشتر آن در ژنوم میزبان و انتقال RNA ویروسی تازه تشکیل شده به سیتوپلاسم برای سنتز بیشتر پروتئین های ذرات ویروسی جدید به هسته سلول میزبان منتقل می کنند.
نقض فرآیندهای سنتز
فرآیندهای سنتز پروتئین در مخازن شبکه آندوپلاسمی رخ می دهد، که از نزدیک با منافذ غشای هسته مرتبط است، که از طریق آن ریبوزوم ها، tRNA و mRNA وارد شبکه آندوپلاسمی می شوند. در اینجا، سنتز زنجیرههای پلی پپتیدی انجام میشود که متعاقباً شکل نهایی خود را در شبکه آندوپلاسمی دانهای و کمپلکس لایهای (کمپلکس گلژی) به دست میآورند، جایی که آنها تحت اصلاح پس از ترجمه قرار میگیرند و با مولکولهای کربوهیدرات و لیپید ترکیب میشوند. مولکول های پروتئینی تازه تشکیل شده در محل سنتز باقی نمی مانند، بلکه از طریق یک فرآیند پیچیده تنظیم شده به نام پروتئین کینز، به طور فعال به آن بخش جدا شده از سلول منتقل می شوند و در آنجا عملکرد مورد نظر خود را انجام می دهند. در این مورد، یک گام بسیار مهم، ساختار مولکول منتقل شده در یک پیکربندی فضایی مناسب است که قادر به انجام عملکرد ذاتی خود باشد. این ساختار با کمک آنزیم های خاص یا روی ماتریکسی از مولکول های پروتئینی تخصصی - چاپرون ها اتفاق می افتد، که به مولکول پروتئین، که به تازگی تشکیل شده یا به دلیل تأثیر خارجی تغییر یافته است، کمک می کند تا ساختار سه بعدی صحیح را به دست آورد. در صورت اثر نامطلوب بر روی سلول، هنگامی که احتمال اختلال در ساختار مولکول های پروتئین وجود دارد (به عنوان مثال، با افزایش دمای بدن، یک فرآیند عفونی، مسمومیت)، غلظت چاپرون ها در سلول افزایش می یابد. به شدت بنابراین به این گونه مولکول ها نیز گفته می شود پروتئین های استرس، یا پروتئین های شوک حرارتی. نقض ساختار یک مولکول پروتئین منجر به تشکیل کنگلومراهای بی اثر شیمیایی می شود که در طول آمیلوئیدوز، بیماری آلزایمر و غیره در سلول یا خارج از آن رسوب می کنند. گاهی اوقات یک مولکول مشابه از پیش ساخته شده می تواند به عنوان ماتریکس عمل کند و در در این صورت، اگر ساختار اولیه اشتباه رخ دهد، تمام مولکول های بعدی نیز معیوب خواهند بود. این وضعیت در بیماری های به اصطلاح پریون (اسکراپی در گوسفند، گاو هار، کورو، بیماری کروتسفلد-ژاکوب در انسان) رخ می دهد، زمانی که نقص در یکی از پروتئین های غشایی سلول عصبی باعث تجمع بعدی توده های بی اثر در داخل سلول می شود. و اختلال در عملکردهای حیاتی آن.
اختلال در فرآیندهای سنتز در یک سلول می تواند در مراحل مختلف آن رخ دهد: رونویسی RNA در هسته، ترجمه پلی پپتیدها در ریبوزوم ها، اصلاح پس از ترجمه، هیپرمتیلاسیون و گلیکوزیلاسیون مولکول بژ، انتقال و توزیع پروتئین ها در سلول و حذف آنها. به بیرون در این حالت می توان افزایش یا کاهش تعداد ریبوزوم ها، تجزیه پلی ریبوزوم ها، انبساط مخازن شبکه آندوپلاسمی دانه ای، از بین رفتن ریبوزوم ها و تشکیل وزیکول ها و واکوئل ها را مشاهده کرد. بنابراین، هنگام مسموم شدن توسط یک خرچنگ رنگ پریده، آنزیم RNA پلیمراز آسیب می بیند، که رونویسی را مختل می کند. سم دیفتری با غیرفعال کردن عامل افزایش طول، فرآیندهای ترجمه را مختل می کند و باعث آسیب میوکارد می شود. علت اختلال در سنتز برخی از مولکول های پروتئینی خاص می تواند عوامل عفونی باشد. به عنوان مثال، ویروس های تبخال سنتز و بیان مولکول های آنتی ژن MHC را مهار می کنند، که به آنها اجازه می دهد تا حدی از کنترل ایمنی باسیل های طاعون جلوگیری کنند - سنتز واسطه های التهاب حاد. ظهور پروتئین های غیر معمول می تواند تجزیه بیشتر آنها را متوقف کند و منجر به تجمع مواد بی اثر یا حتی سمی شود. این می تواند تا حدودی با اختلال در فرآیندهای پوسیدگی تسهیل شود.
اختلال در فرآیندهای پوسیدگی
همزمان با سنتز پروتئین در سلول، تجزیه آن به طور مداوم اتفاق می افتد. در شرایط عادی، این امر اهمیت تنظیمی و شکلدهندهای دارد، به عنوان مثال، در هنگام فعال شدن اشکال غیرفعال آنزیمها، هورمونهای پروتئینی و پروتئینهای چرخه میتوزی. رشد و نمو سلولی طبیعی نیازمند تعادل کنترل شده بین سنتز و تخریب پروتئین ها و اندامک ها است. با این حال، در فرآیند سنتز پروتئین، به دلیل اشتباهات در عملکرد دستگاه سنتز، ساختار غیر طبیعی مولکول پروتئین و آسیب آن توسط عوامل شیمیایی و باکتریایی، تعداد نسبتاً زیادی مولکول معیوب دائماً تشکیل می شود. بر اساس برخی برآوردها، سهم آنها حدود یک سوم کل پروتئین های سنتز شده است.
سلول های پستانداران چندین اصلی دارند روش های تخریب پروتئین:از طریق پروتئازهای لیزوزومی (پنتید هیدرولاز)، پروتئینازهای وابسته به کلسیم (اندوپپتیدازها) و سیستم پروتئازوم. علاوه بر این، پروتئینازهای تخصصی مانند کاسپازها نیز وجود دارد. اندامک اصلی که در آن تجزیه مواد در سلول های یوکاریوتی رخ می دهد، لیزوزوم است که حاوی آنزیم های هیدرولیتیک متعددی است. به دلیل فرآیندهای اندوسیتوز و انواع مختلف اتوفاژی در لیزوزوم ها و فاگولیزوزوم ها، مولکول های پروتئین معیوب و کل اندامک ها از بین می روند: میتوکندری های آسیب دیده، بخش هایی از غشای پلاسما، برخی از پروتئین های خارج سلولی و محتویات گرانول های ترشحی.
یک مکانیسم مهم برای تخریب پروتئین، پروتئازوم است، یک ساختار پروتئیناز چند کاتالیستی با ساختار پیچیده که در سیتوزول، هسته، شبکه آندوپلاسمی و روی غشای سلولی قرار دارد. این سیستم آنزیمی مسئول تجزیه پروتئین های آسیب دیده و همچنین پروتئین های سالم است که برای عملکرد طبیعی سلول باید حذف شوند. در این مورد، پروتئین هایی که باید از بین بروند، ابتدا با یک پلی پپتید خاص، یوبیکوئیتین، ترکیب می شوند. با این حال، پروتئین های غیر یوبی کوئیتین نیز می توانند تا حدی در پروتئازوم ها از بین بروند. تجزیه مولکول های پروتئین در پروتئازوم ها به پلی پپتیدهای کوتاه (در حال پردازش) با ارائه بعدی آنها همراه با مولکول های MHC نوع I یک پیوند مهم در کنترل ایمنی هموستاز آنتی ژنی در بدن است. هنگامی که عملکرد پروتئازوم ضعیف می شود، پروتئین های آسیب دیده و غیر ضروری جمع می شوند که با پیری سلول همراه است. نقض تخریب پروتئین های وابسته به سیکلین منجر به اختلال در تقسیم سلولی، تخریب پروتئین های ترشحی - به توسعه سیستوفیبروز می شود. برعکس، افزایش عملکرد پروتئازوم با تخلیه بدن (ایدز، سرطان) همراه است.
با اختلالات تعیین شده ژنتیکی در تخریب پروتئین، ارگانیسم زنده نیست و در مراحل اولیه جنین زایی می میرد. اگر تجزیه چربی ها یا کربوهیدرات ها مختل شود، بیماری های ذخیره سازی (تزاریسموزیس) رخ می دهد. در این حالت، مقدار زیادی از مواد خاص یا محصولات حاصل از تجزیه ناقص آنها - لیپیدها، پلی ساکاریدها - در داخل سلول تجمع می یابد که به طور قابل توجهی به عملکرد سلول آسیب می رساند. اغلب این در سلول های اپیتلیال کبد (هپاتوسیت ها)، نورون ها، فیبروبلاست ها و ماکروفاگوسیت ها مشاهده می شود.
اختلالات اکتسابی فرآیندهای تجزیه مواد می تواند در نتیجه فرآیندهای پاتولوژیک (به عنوان مثال، پروتئین، چربی، کربوهیدرات و دژنراسیون رنگدانه) ایجاد شود و با تشکیل مواد غیر معمول همراه باشد. اختلالات در سیستم پروتئولیز لیزوزومی منجر به کاهش سازگاری در طول روزه داری یا افزایش استرس و بروز برخی اختلالات غدد درون ریز - کاهش سطح انسولین، تیروگلوبولین، سیتوکین ها و گیرنده های آنها می شود. اختلال در تخریب پروتئین سرعت بهبود زخم را کاهش می دهد، باعث ایجاد آترواسکلروز می شود و بر پاسخ ایمنی تأثیر می گذارد. با هیپوکسی، تغییر در pH داخل سلولی، آسیب ناشی از تشعشع، که با افزایش پراکسیداسیون لیپیدهای غشاء مشخص می شود، و همچنین تحت تأثیر مواد لیزوزوموتروپ - اندوتوکسین های باکتریایی، متابولیت های قارچ های سمی (اسپوروفوسارین)، کریستال های اکسید سیلیکون - پایداری غشای لیزوزوم تغییرات، آنزیم های لیزوزومی فعال شده در سیتوپلاسم آزاد می شوند که باعث تخریب ساختارهای سلولی و مرگ آن می شود.
فصل 1
مبانی فیزیولوژی سلولی
I. دودل
غشای پلاسمایی . سلول های حیوانی توسط یک غشای پلاسمایی محدود شده اند (شکل 1.1). ما در مورد ساختار آن، که بسیار شبیه به ساختار بسیاری از غشاهای درون سلولی است، با جزئیات بیشتری صحبت خواهیم کرد. ماتریس اصلی غشاء شامل لیپیدها،عمدتا فسفاتیدیل کولین این لیپیدها از یک گروه سر آبدوست تشکیل شده اند که زنجیره های هیدروکربنی آبگریز طولانی به آن متصل است. در آب، چنین لیپیدهایی به طور خود به خود یک لایه دولایه به ضخامت 4 تا 5 نانومتر تشکیل می دهند که در آن گروه های آبدوست رو به محیط آبی هستند و زنجیره های هیدروکربنی آبگریز در دو ردیف مرتب شده اند و یک فاز لیپیدی بی آب را تشکیل می دهند. غشاهای سلولی دو لایه لیپیدی از این نوع هستند و حاوی گلیکولیپیدها، کلسترول و فسفولیپیدها هستند (شکل 1.2). بخش آبدوست گلیکولیپیدها توسط الیگوساکاریدها تشکیل می شود. گلیکولیپیدها همیشه در سطح بیرونی غشای پلاسمایی قرار دارند و بخش اولیگوساکارید مولکول مانند مو غوطه ور در محیط است. مولکول های کلسترول که در بین فسفولیپیدها به مقدار تقریباً مساوی پراکنده شده اند، غشا را تثبیت می کنند. توزیع لیپیدهای مختلف در لایه های داخلی و خارجی غشا یکسان نیست و حتی در یک لایه مناطقی وجود دارد که انواع خاصی از لیپیدها در آنها متمرکز شده اند. این توزیع نابرابر
برنج. 1.1. ترسیم شماتیک سلولی که مهم ترین اندامک ها را نشان می دهد
احتمالاً تا حدودی اهمیت عملکردی هنوز نامشخص دارد.
عناصر عملکردی اصلی تعبیه شده در ماتریس لیپیدی نسبتاً بی اثر غشاء عبارتند از سنجاب ها(شکل 1.2). پروتئین بر حسب وزن 25 تا 75 درصد در غشاهای مختلف را تشکیل می دهد، اما از آنجایی که مولکول های پروتئین بسیار بزرگتر از مولکول های لیپید هستند، 50 درصد وزنی معادل نسبت 1 مولکول پروتئین به 50 مولکول چربی است. برخی از پروتئین ها از سطح بیرونی به درونی غشا نفوذ می کنند، در حالی که برخی دیگر در یک لایه ثابت می شوند. مولکول های پروتئین معمولاً به گونه ای جهت گیری می شوند که گروه های آبگریز آنها در غشای لیپیدی و گروه های آب دوست قطبی در سطح غشا در فاز آبی قرار می گیرند. بسیاری از پروتئین های سطح بیرونی غشاء گلیکوپروتئین هستند. گروه ساکارید آبدوست آنها با محیط خارج سلولی روبرو هستند.
سیستم های غشایی اندامک های داخل سلولی .
تقریباً نیمی از حجم سلول توسط اندامک های جدا شده از سیتوزول توسط غشاها اشغال می شود. سطح کل غشاهای اندامک های داخل سلولی حداقل 10 برابر بیشتر از سطح غشای پلاسمایی است. پرکاربردترین سیستم غشایی است شبکه آندوپلاسمی،به نمایندگی از یک شبکه

برنج. 1.2.نمایش شماتیک غشای پلاسمایی پروتئین ها در دولایه فسفولیپیدی تعبیه شده اند و برخی از آنها به لایه دوگانه نفوذ می کنند، در حالی که برخی دیگر فقط به لایه بیرونی یا داخلی متصل می شوند.
لوله های بسیار پیچیده یا ساختارهای دراز کیسه مانند. مناطق وسیعی از شبکه آندوپلاسمی با ریبوزوم پوشانده شده است. چنین شبکه ای دانه ای یا خشن نامیده می شود (شکل 1.1). دستگاه گلژیهمچنین از لاملاهای متصل به غشاء تشکیل شده است که وزیکول ها یا وزیکول ها از آنها جدا می شوند (شکل 1.1). لیزوزوم ها و پراکسی زوم ها- اینها وزیکول های کوچک تخصصی هستند. در تمام این اندامکهای مختلف، غشاء و فضایی که آن را در بر میگیرد شامل مجموعههای خاصی از آنزیمها است. در داخل اندامک ها، محصولات متابولیک ویژه ای تجمع می یابد که برای انجام عملکردهای مختلف اندامک ها استفاده می شود.
هستهو میتوکندریتفاوت آنها در این است که هر یک از این اندامک ها توسط دو غشاء احاطه شده است. هسته مسئول کنترل جنبشی متابولیسم است. غشای داخلی چین خورده میتوکندری محل متابولیسم اکسیداتیو است. در اینجا، به دلیل اکسیداسیون پیروات یا اسیدهای چرب، ترکیب پرانرژی آدنوزین تری فسفات (ATP یا ATP) سنتز می شود.
اسکلت سلولی . سیتوپلاسم احاطه کننده اندامک ها به هیچ وجه نمی تواند آمورف در نظر گرفته شود. توسط یک شبکه اسکلت سلولی نفوذ می کند. اسکلت سلولی از میکروتوبول ها، رشته های اکتین و رشته های میانی تشکیل شده است (شکل 1.1). میکروتوبول هادارای قطر بیرونی حدود 25 نانومتر؛ آنها مانند یک پلیمر معمولی در نتیجه مونتاژ مولکول های پروتئین توبولین تشکیل می شوند. رشته های اکتین –الیاف انقباضی، واقع در لایه نزدیک غشاء و در سراسر سلول، عمدتا در فرآیندهای مرتبط با حرکت شرکت می کنند. رشته های میانیاز بلوک هایی با ترکیب شیمیایی مختلف در انواع مختلف سلول تشکیل شده است. آنها انواع اتصالات را بین دو عنصر اسکلت سلولی دیگر که در بالا ذکر شد تشکیل می دهند. اندامک ها و غشای پلاسمایی نیز با اسکلت سلولی مرتبط هستند که نه تنها شکل سلول و موقعیت اندامک ها را در آن حفظ می کند، بلکه تغییرات شکل سلول و تحرک آن را نیز تعیین می کند.
سیتوزول . حدود نیمی از حجم سلول توسط سیتوزول اشغال شده است. از آنجایی که تقریباً 20 درصد وزن آن پروتئین است، بیشتر یک ژل است تا یک محلول آبی. مولکول های کوچک، از جمله آلی و معدنی یون ها،در فاز آبی حل می شود. تبادل یون ها بین سلول و محیط (فضای خارج سلولی) رخ می دهد. این فرآیندهای متابولیک در بخش بعدی مورد بحث قرار خواهند گرفت. غلظت یون ها در فضای خارج سلولی با دقت قابل توجهی در سطح ثابتی حفظ می شود. غلظت درون سلولی هر یون نیز دارای سطح خاصی است، متفاوت از غلظت خارج از سلول (جدول 1.1). فراوان ترین کاتیون در محیط خارج سلولی است Na+ در سلول غلظت آن بیش از 10 برابر کمتر است. برعکس، غلظت K + در داخل سلول بیشتر از یک مرتبه بزرگتر است. بیشترین گرادیان بین غلظت های خارج سلولی و درون سلولی برای Ca2+ وجود دارد، غلظت یون های آزاد در داخل سلول حداقل 10000 برابر کمتر از خارج از آن است. همه یون ها در سیتوزول حل نمی شوند. به عنوان مثال، در مورد Ca 2+، تعداد یون های محدود بسیار بیشتر از یون های آزاد است. بیشتر پروتئین های سیتوزولی آنزیم هایی هستند که با مشارکت آنها بسیاری از فرآیندهای متابولیسم میانی انجام می شود: گلیکولیز و گلوکونئوژنز، سنتز یا تخریب اسیدهای آمینه، سنتز پروتئین روی ریبوزوم ها (شکل 1.1). سیتوزول همچنین حاوی قطرات لیپید و گرانول های گلیکوژن است که به عنوان ذخایر مولکول های مهم عمل می کنند.
جدول 1.1.غلظت یون داخل و خارج سلولی در سلول های عضلانی حیوانات هوموترمیک آ – – آنیون های سلولی با وزن مولکولی بالا
|
غلظت درون سلولی |
غلظت خارج سلولی |
||
|
Na+ |
12 میلی مول در لیتر |
Na+ |
145 میلی مول در لیتر |
|
155 میلی مول در لیتر |
K+ |
4 میلی مول در لیتر |
|
|
Ca 2+ |
10-7 10-8 میلی مول در لیتر |
Ca 2+ |
2 میلی مول در لیتر |
|
C l - |
4 میلی مول در لیتر |
C1 - |
120 میلی مول در لیتر |
|
HCO 3 - |
8 میلی مول در لیتر |
HCO 3 - |
27 میلی مول در لیتر |
|
آ - |
155 میلی مول در لیتر |
دیگران کاتیون ها |
5 میلی مول در لیتر |
|
پتانسیل استراحت -90 میلی ولت |
|||
1.2. تبادل مواد بین سلول و محیط
ما به طور مختصر ساختار سلول را شرح داده ایم تا بتوانیم از این توضیحات برای بررسی اصول فیزیولوژی سلولی استفاده کنیم. در هیچ موردی نمی توان یک سلول را یک موجود ثابت در نظر گرفت، زیرا تبادل مداوم مواد بین بخش های مختلف درون سلولی و همچنین بین محفظه ها و محیط وجود دارد. ساختارهای سلولی در تعادل دینامیکی هستند و برهمکنش سلول ها با یکدیگر و با محیط خارجی شرط لازم برای حفظ حیات یک موجود زنده است. در این فصل به مکانیسم های اساسی چنین مبادله ای نگاه خواهیم کرد. در فصلهای بعدی، این مکانیسمها در مورد سلول عصبی و عملکردهای آن اعمال میشوند.
با این حال، مکانیسم های یکسانی زیربنای عملکرد همه اندام های دیگر است.
انتشار.ساده ترین فرآیند حرکت ماده انتشار است. در محلول ها (یا گازها)، اتم ها و مولکول ها آزادانه حرکت می کنند و اختلاف غلظت ها به دلیل انتشار متعادل می شود. بیایید دو حجم پر از مایع یا گاز را در نظر بگیریم (شکل 1.3)، که در آنها مواد دارای غلظت هستند.ج 1 و ج 2 و توسط لایه ای با سطح A و ضخامت جدا می شودد جریان ماده m در زمان t شرح داده شده قانون اول انتشار فیک:
dm/ dt= Dآ/ د ( سی 1 –C 2)=Dآ/ دD سی(1)
که در آن D ضریب انتشار، ثابت برای یک ماده، حلال و دما است. به شکل کلی تر برای تفاوت غلظت dc در فاصله dx
dm/dt= –D A dc/dx,(2)
جریان از طریق بخش A با گرادیان غلظت متناسب است dc/dx . علامت منفی در معادله ظاهر می شود زیرا تغییر غلظت در جهت x منفی است.
انتشار مهم ترین فرآیندی است که به دلیل آن بیشتر مولکول های موجود در محلول های آبی در فواصل کوتاه حرکت می کنند. این امر در مورد حرکت آنها در سلول نیز صدق می کند تا جایی که انتشار توسط غشاها مانع نمی شود. بسیاری از مواد می توانند آزادانه از طریق غشاهای لیپیدی، به ویژه آب و گازهای محلول مانند O 2 و CO 2 منتشر شوند. محلول در چربی

برنج. 1.3.طرح انتشار کمی این دو فضا با یک لایه ضخیم از هم جدا شده انددو منطقه آ.ج – غلظت بالای ذرات در قسمت چپ حجم، C: – غلظت کم ذرات در سمت راست قطعات، سطح صورتی- گرادیان غلظت در لایه انتشار. جریان انتشار dm/dt - ببینید معادله 1)
مواد نیز به خوبی از طریق غشاها پخش می شوند. این همچنین در مورد مولکول های قطبی با اندازه نسبتاً کوچک مانند اتانول و اوره صدق می کند، در حالی که قندها به سختی از لایه لیپیدی عبور می کنند. در عین حال، لایههای لیپیدی عملاً در برابر مولکولهای باردار از جمله حتی یونهای معدنی غیرقابل نفوذ هستند. برای غیر الکترولیت ها، معادله انتشار (1) معمولاً با ترکیب ویژگی های غشاء و ماده منتشر کننده به یک تبدیل می شود. نفوذپذیری پارامتر (P):
dm/dt=P AD ج.(3)
در شکل 1.4 مقایسه شده است نفوذپذیری (P) غشای لیپیدی برای مولکول های مختلف.
انتشار از طریق منافذ غشایی . غشای پلاسمایی (و سایر غشاهای سلولی) نه تنها به موادی که از طریق لایه لیپیدی منتشر می شوند، بلکه برای بسیاری از یون ها، قندها، اسیدهای آمینه و نوکلئوتیدها نیز نفوذپذیر است. این مواد از طریق منافذ تشکیل شده از غشاء عبور می کنند پروتئین های انتقال دهنده،غوطه ور در غشاء. در داخل چنین پروتئین هایی یک کانال پر از آب با قطر کمتر از 1 نانومتر وجود دارد که از طریق آن مولکول های کوچک می توانند پخش شوند. آنها در امتداد یک گرادیان غلظت حرکت می کنند و اگر بار حمل کنند، حرکت آنها از طریق کانال ها نیز توسط پتانسیل غشایی تنظیم می شود. کانال های غشایی نسبتا انتخابی هستند

برنج. 1.4.نفوذپذیری دو لایه لیپیدی مصنوعی به مواد مختلف
در رابطه با نوع مولکول هایی که می توانند از آنها عبور کنند، به عنوان مثال، کانال های پتاسیم، سدیم و کلسیم وجود دارد که هر کدام تقریباً به هر یونی غیر از یک یون خاص نفوذ ناپذیر است. چنین گزینش پذیریتوسط بار یا ساختار محل های اتصال در دیواره کانال تعیین می شود، که حمل و نقل یک مولکول خاص را تسهیل می کند و از نفوذ مواد دیگر از طریق کانال جلوگیری می کند. 1.5، الف) .
پشت رفتار کانال های یونی غشاییمشاهده آسان است، زیرا جریان ناشی از حرکت یون ها را می توان حتی برای یک کانال اندازه گیری کرد. نشان داده شده است که کانال ها به طور خود به خود و با فرکانس بالا حالت خود را از باز به بسته تغییر می دهند. کانال پتاسیم توسط پالس های جریان با دامنه حدود 2 pA (2 10-12 A) و مدت زمان چند میلی ثانیه مشخص می شود (شکل 2.12، ص 37 را ببینید) [3]. در این مدت ده ها هزار یون از آن عبور می کنند. انتقال پروتئین ها از یک ترکیب به ترکیب دیگر با استفاده از پراش اشعه ایکس، طیف سنجی Mössbauer و رزونانس مغناطیسی هسته ای (NMR) مورد مطالعه قرار می گیرد. بنابراین پروتئین ها ساختارهای بسیار پویا و متحرکی هستند و کانالی که از پروتئین عبور می کند فقط یک لوله سفت و سخت پر از آب نیست (شکل 1.5، A)، بلکه هزارتویی از گروه ها و بارهای مولکولی به سرعت در حال حرکت است. این ویژگی پویای کانال منعکس شده است مشخصات انرژی کانال،در شکل نشان داده شده است. 1.5، B. در اینجا، محور آبسیسا نشان دهنده طول کانال از محلول خارجی با غلظت یون C 0 و پتانسیل 0 تا محلول داخلی با غلظت C 1 و پتانسیل E است. محور اردینات

برنج. 1.5.الف. شماتیکی از پروتئینی که یک کانال پتاسیمی را در لایه دولایه لیپیدی غشای پلاسمایی تشکیل می دهد. چهار بار منفی روی "دیوار" کانال ثابت شده است. B. نمایه انرژی شماتیک کانال نشان داده شده در شکل. الف) محور ارتین مقادیر انرژی جنبشی مورد نیاز برای عبور از کانال را نشان می دهد. در امتداد محور آبسیسا - فاصله بین سطوح داخلی و خارجی غشاء. حداقل انرژی مربوط به محل اتصال یون های دارای بار مثبت با بارهای منفی ثابت در دیواره کانال است. حداکثر انرژی مربوط به موانع انتشار در کانال است. ترکیب پروتئین کانال به طور خود به خود در حال نوسان است. گزینه های نمایه انرژی با خطوط یکپارچه و چین نشان داده می شوند. این نوسانات اتصال یون ها را هنگام غلبه بر سد انرژی (تغییر شده) بسیار تسهیل می کند.
سطوح انرژی یون در سایت های اتصال کانال ارائه شده است. اوج در نمودار نشان دهنده مانع نفوذپذیری است که انرژی یونی باید از آن فراتر رود تا به کانال نفوذ کند، و "شیب" در نمودار نشان دهنده یک حالت نسبتاً پایدار (بایدینگ) است. با وجود مانع پیک انرژی، اگر پروفایل انرژی خود به خود تغییر کند، یون می تواند به کانال نفوذ کند. بنابراین یون ممکن است به طور ناگهانی خود را "در آن سوی" اوج انرژی پیدا کند و ممکن است به حرکت خود به داخل سلول ادامه دهد. بسته به بار، اندازه و درجه هیدراتاسیون یون و توانایی آن در اتصال به ساختارهای دیواره کانال، مشخصات انرژی کانال برای یونهای مختلف متفاوت است، که ممکن است گزینشپذیری انواع کانالها را توضیح دهد.
تعادل انتشار یون ها . انتشار یون های مختلف از طریق کانال های غشایی باید منجر به حذف تفاوت در غلظت بین محیط های خارج و داخل سلولی شود. همانطور که، با این حال، از جدول قابل مشاهده است. 1.1، چنین تفاوت هایی باقی می ماند، بنابراین، باید برخی از آنها وجود داشته باشد تعادلبین انتشار و سایر فرآیندهای انتقال در سراسر غشاء. دو بخش بعدی به راه هایی می پردازد که از طریق آنها این تعادل حاصل می شود. در مورد یون ها، تعادل انتشار تحت تأثیر بار آنها قرار می گیرد. انتشار مولکول های بدون بار با تفاوت در غلظت ها تضمین می شوددی سی ، و هنگامی که غلظت ها برابر می شوند، خود حمل و نقل متوقف می شود. ذرات باردار علاوه بر این تحت تأثیر میدان الکتریکی قرار می گیرند. به عنوان مثال، هنگامی که یک یون پتاسیم از یک سلول در امتداد گرادیان غلظت خود خارج می شود، حامل یک بار مثبت واحد است. بنابراین، محیط درون سلولی بار منفی بیشتری پیدا می کند و در نتیجه اختلاف پتانسیل در سراسر غشاء ایجاد می شود. بار منفی درون سلولی از خروج یونهای پتاسیم جدید از سلول جلوگیری میکند و یونهایی که سلول را ترک میکنند، بار روی غشاء را بیشتر میکنند. جریان یون های پتاسیم زمانی متوقف می شود که عمل میدان الکتریکی فشار انتشار را به دلیل اختلاف غلظت جبران کند. یون ها همچنان از غشاء عبور می کنند، اما به مقدار مساوی در هر دو جهت. بنابراین، برای یک تفاوت معین در غلظت یون در غشاء، وجود دارد پتانسیل تعادل Eیون که در آن جریان یون ها از طریق غشا متوقف می شود. پتانسیل تعادل را می توان به راحتی با استفاده از آن تعیین کرد معادلات نرنست:
Eیون= آرتی/ zاف* لوگاریتمC بیرون/ سی در(4)
جایی که R - ثابت گاز، T - دمای مطلق، z - ظرفیت یون (منفی برای آنیونها) C بیرون - غلظت یون خارج سلولیسی در - غلظت یون درون سلولیاف شماره فارادی اگر ثابت ها را در معادله جایگزین کنیم، در دمای بدن (T = 310 K) پتانسیل تعادل برای یون های پتاسیم E K برابر است با:
اک= –61 مترB log /(5)
اگر [ K + خارج ]/[ K + داخل ] = 39، مطابق جدول زیر. 1.1 سپس
Ek= –61 m B log 39= –97 mV.
در واقع، مشخص شد که همه سلول ها دارند پتانسیل غشایی؛در سلولهای ماهیچهای پستانداران سطح آن در حدود -90 میلی ولت است. بسته به شرایط و غلظت نسبی یون، سلول ها می توانند پتانسیل غشایی در محدوده 40- تا 120- میلی ولت داشته باشند. برای سلول در مثال بالا (جدول 1.1) پتانسیل استراحت،برابر با 90- میلی ولت، نشان می دهد که شار یون های پتاسیم از طریق کانال های غشایی تقریباً در تعادل هستند. این تعجب آور نیست، زیرا در غشای در حال استراحت، وضعیت باز کانال های پتاسیم به احتمال زیاد، یعنی. غشاء در برابر یون های پتاسیم بیشترین نفوذ را دارد. با این حال، پتانسیل غشاء توسط جریان یون های دیگر تعیین می شود.
سهولت انتشار ذرات بدون بار از طریق غشاء در رابطه (3) تعیین شده است. نفوذپذیری در برابر ذرات باردار با یک معادله کمی پیچیده تر توصیف می شود:
پ= متر آرتی/ dF(6)
جایی که متر- تحرک یون در غشا،د - ضخامت غشاءیک R، T و F - ثابت های ترمودینامیکی شناخته شده مقادیر نفوذپذیری برای یون های مختلف تعیین شده به این روش می تواند برای محاسبه پتانسیل غشا استفاده شود Em هنگامی که یون های پتاسیم، سدیم و کلر به طور همزمان از غشا عبور می کنند (با نفوذپذیری P K، P Na و P Cl به ترتیب). فرض بر این است که پتانسیل به طور یکنواخت در غشا کاهش می یابد، به طوری که قدرت میدان ثابت است. در این مورد اعمال می شود معادله گلدمن یا معادله میدان ثابت :
Em= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl )(7)
برای اکثر غشاهای سلولی Pک تقریبا 30 برابر بیشتر از P Na (به بخش 1.3 نیز مراجعه کنید). ارزش نسبی P Cl بسیار متفاوت است؛ برای بسیاری از غشاها P Cl کوچک در مقایسه با Rک اما برای دیگران (مثلاً در عضلات اسکلتی) P Cl ، به طور قابل توجهی بالاتر از Pک.
حمل و نقل فعال، پمپ سدیم . بخش قبل انتشار غیرفعال یون ها و پتانسیل غشایی حاصل را در غلظت های یون داخل و خارج سلولی مشخص می کند. با این حال، در نتیجه این فرآیند، غلظت یون ها در داخل سلول به طور خودکار تثبیت نمی شود، زیرا غشاء
پتانسیل کمی الکترونگاتیو تر از EK، و بسیار در مقایسه با E Na (حدود +60 میلی ولت). به دلیل انتشار، غلظت یون های درون سلولی، حداقل پتاسیم و سدیم، باید با غلظت های خارج سلولی برابر شود. پایداری گرادیان یونی از طریق انتقال فعال حاصل میشود: پروتئینهای غشایی یونها را در سراسر غشا در برابر گرادیانهای الکتریکی و/یا غلظت حمل میکنند و انرژی متابولیکی را برای این کار مصرف میکنند. مهمترین فرآیند حمل و نقل فعال کار است Na/K - پمپی که تقریباً در تمام سلول ها وجود دارد.
پمپ یون های سدیم را از سلول خارج می کند و همزمان یون های پتاسیم را به داخل سلول پمپ می کند. این امر غلظت کم یونهای سدیم درون سلولی و غلظت بالای یونهای پتاسیم را تضمین میکند (جدول 1.1). گرادیان غلظت یونهای سدیم روی غشاء عملکردهای خاص مربوط به انتقال اطلاعات به شکل تکانههای الکتریکی (به بخش 2.2) و همچنین حفظ سایر مکانیسمهای انتقال فعال و تنظیم حجم سلول (به زیر مراجعه کنید) دارد. بنابراین، جای تعجب نیست که بیش از 1/3 انرژی مصرف شده توسط یک سلول صرف پمپ Na/K شود و در برخی از فعال ترین سلول ها تا 70 درصد انرژی صرف عملکرد آن می شود.
پروتئین انتقال Na/K یک ATPase است. در سطح داخلی غشا، ATP را به ADP و فسفات تجزیه می کند (شکل 1.6). انرژی یک مولکول ATP برای انتقال سه یون سدیم از سلول و همزمان دو یون پتاسیم به داخل سلول استفاده می شود، یعنی در مجموع یک بار مثبت در یک چرخه از سلول حذف می شود. بنابراین، پمپ Na/K است الکتروژنیک(یک جریان الکتریکی در سراسر غشا ایجاد می کند)، که منجر به افزایش الکترونگاتیوی پتانسیل غشا تقریباً 10 میلی ولت می شود. پروتئین انتقال این عمل را با سرعت بالا انجام می دهد: از 150 تا 600 یون سدیم در ثانیه. توالی اسید آمینه پروتئین انتقال شناخته شده است، اما مکانیسم این انتقال متابولیک پیچیده هنوز مشخص نیست. این فرآیند با استفاده از پروفایل های انرژی انتقال یون های سدیم یا پتاسیم توسط پروتئین ها توصیف شده است (شکل 1.5،5). با توجه به ماهیت تغییرات در این پروفایل ها، همراه با تغییرات مداوم در ترکیب پروتئین انتقال (فرآیندی که نیاز به انرژی دارد)، می توان استوکیومتری تبادل را قضاوت کرد: دو یون پتاسیم با سه یون سدیم مبادله می شوند.
پمپ Na/K، همانند ایزوله Na+ /K + ATPase غشایی وابسته، که به طور خاص توسط گلیکوزید قلبی اوابائین (استروفانتین) مهار می شود. از آنجایی که عملکرد پمپ Na/K یک واکنش شیمیایی چند مرحلهای است، مانند تمام واکنشهای شیمیایی، تا حد زیادی به دما وابسته است.

برنج. 1.6.نمودار یک پمپ Na/K-ATPase (غوطهور در دولایه لیپیدی غشای پلاسمایی) که در یک چرخه سه یون Na + را از سلول در برابر شیب پتانسیل و غلظت حذف میکند و دو یون K را به داخل سلول میآورد. + . در طی این فرآیند، یک مولکول ATP به ADP و فسفات تجزیه می شود. نمودار ATPase را به عنوان یک دایمر متشکل از یک زیر واحد بزرگ (عملکردی) و کوچک نشان می دهد. در غشاء به صورت یک تترامر وجود دارد که توسط دو زیر واحد بزرگ و دو زیر واحد کوچک تشکیل شده است
در شکل نشان داده شده است. 1.7. در اینجا، شار یون های سدیم از سلول های ماهیچه ای نسبت به زمان نشان داده شده است. این عملاً معادل جریان یونهای سدیم است که با عملکرد پمپ Na/K انجام میشود، زیرا جریان غیرفعال یونهای سدیم در برابر غلظت و گرادیانهای بالقوه بسیار کم است. اگر دارو تا حدود 18 درجه سانتیگراد خنک شود، جریان یون سدیم از سلول به سرعت 15 برابر کاهش می یابد و بلافاصله پس از حرارت دادن به سطح اولیه خود باز می گردد. این کاهش در جریان یون های سدیم از سلول چندین برابر بیشتر از آن چیزی است که با وابستگی دمایی فرآیند انتشار یا یک واکنش شیمیایی ساده مطابقت دارد. اثر مشابهی زمانی مشاهده می شود که انرژی متابولیک در نتیجه مسمومیت با دینیتروفنول (DNP) کاهش می یابد (شکل 1.7،5). در نتیجه، جریان یون های سدیم از سلول توسط یک واکنش وابسته به انرژی - یک پمپ فعال - تضمین می شود. یکی دیگر از ویژگی های پمپ، همراه با وابستگی قابل توجه به دما و انرژی، وجود سطح اشباع (مانند سایر واکنش های شیمیایی) است. این بدان معنی است که با افزایش غلظت یون های انتقال یافته، سرعت پمپ نمی تواند به طور نامحدود افزایش یابد (شکل 1.8). در مقابل، شار ماده ای که به طور غیرفعال منتشر می شود، متناسب با اختلاف غلظت مطابق با قانون انتشار افزایش می یابد (معادلات 1 و 2).

برنج. 1.7. الف، ب.انتقال فعال سدیم + . محور Y:شار رادیواکتیو 24 Na + از سلول (Imp./min). محور آبسیسا:زمان از شروع آزمایش آ.سلول از 18.3 درجه سانتیگراد تا 0.5 درجه سانتیگراد خنک می شود. جریان Na+ از سلول در این دوره مهار می شود. ب.سرکوب شار Na + از سلول با دینیتروفنول (DNP) در غلظت 0.2 mmol/l (اصلاح شده)
علاوه بر پمپ Na/K، غشای پلاسمایی حاوی حداقل یک پمپ دیگر است - کلسیم؛این پمپ یون های کلسیم (Ca2+) را از سلول خارج می کند و در حفظ غلظت درون سلولی آنها در سطح بسیار پایین نقش دارد (جدول 1.1). پمپ کلسیم با چگالی بسیار بالا در شبکه سارکوپلاسمی سلول های عضلانی وجود دارد که یون های کلسیم را در نتیجه تجزیه مولکول های ATP جمع می کنند (به فصل 4 مراجعه کنید).
اثر پمپ Na/K بر پتانسیل غشا و حجم سلول . در شکل شکل 1.9 اجزای مختلف جریان غشا را نشان می دهد و غلظت یون های درون سلولی را نشان می دهد.

برنج. 1.8.رابطه بین سرعت انتقال مولکول ها و غلظت آنها (در ورودی کانال یا در محل اتصال پمپ) در هنگام انتشار از طریق کانال یا در حین انتقال پمپ. دومی در غلظت های بالا اشباع می شود (حداکثر سرعت، Vmax ) مقدار در محور x مربوط به نیمی از حداکثر سرعت پمپ ( Vmax /2)، غلظت تعادل است به متر

برنج. 1.9.نموداری که غلظت Na+ را نشان می دهد , K+ و Cl – داخل و خارج سلول و مسیرهای نفوذ این یون ها از طریق غشای سلولی (از طریق کانال های یونی خاص یا با استفاده از پمپ Na/K. در شیب غلظت معین، پتانسیل های تعادلی E Na، E K و E С l - برابر با پتانسیل غشایی نشان داده شده است Em = – 90 میلی ولت
وجود آنها را تضمین کند. یک جریان بیرونی یونهای پتاسیم از طریق کانالهای پتاسیم مشاهده میشود، زیرا پتانسیل غشاء کمی الکترومثبتتر از پتانسیل تعادل یونهای پتاسیم است. رسانایی کلی کانال های سدیمی بسیار کمتر از رسانایی کانال های پتاسیم است. کانال های سدیم در حالت استراحت بسیار کمتر از کانال های پتاسیم باز می شوند. با این حال، تقریباً همان تعداد یون های سدیم وارد سلول می شود که یون های پتاسیم از سلول خارج می شوند، زیرا غلظت و گرادیان های بالقوه زیادی برای انتشار یون های سدیم در سلول مورد نیاز است. پمپ Na/K جبرانی ایده آل را برای جریان های انتشار غیرفعال فراهم می کند، زیرا یون های سدیم را از سلول و یون های پتاسیم را به داخل سلول منتقل می کند. بنابراین، پمپ به دلیل تفاوت در تعداد بارهای منتقل شده به داخل و خارج از سلول، الکتروژنیک است که در سرعت عادی کارکرد آن، پتانسیل غشایی تقریباً 10 ایجاد می کند. میلی ولت الکترونگاتیوتر از زمانی است که توسط شارهای یون غیرفعال به تنهایی تولید شود (به معادله 7 مراجعه کنید). در نتیجه پتانسیل غشا به پتانسیل تعادل پتاسیم نزدیک می شود که نشت یون های پتاسیم را کاهش می دهد. فعالیت Naپمپ /K توسط غلظت درون سلولی یون های سدیم تنظیم می شود. سرعت پمپ با کاهش غلظت یونهای سدیمی که باید از سلول خارج شوند کاهش مییابد (شکل 1.8)، به طوری که عملکرد پمپ و جریان یونهای سدیم به داخل سلول، یکدیگر را متعادل میکنند و غلظت سدیم درون سلولی را حفظ میکنند. یون ها در سطح تقریباً 10 میلی مول در لیتر.
برای حفظ تعادل بین پمپاژ و جریان غیرفعال غشاء، مولکول های پمپ Na/K بیشتر از پروتئین های کانالی برای یون های پتاسیم و سدیم مورد نیاز است. هنگامی که کانال باز است، ده ها هزار یون در چند میلی ثانیه از آن عبور می کنند (نگاه کنید به بالا)، و از آنجایی که کانال معمولا چندین بار در ثانیه باز می شود، در مجموع بیش از 105 یون در این مدت از آن عبور می کنند. یک پروتئین پمپ واحد در هر ثانیه چند صد یون سدیم را حرکت می دهد، بنابراین غشای پلاسما باید حدود 1000 برابر بیشتر از مولکول های کانال حاوی مولکول های پمپ باشد. اندازهگیری جریانهای کانال در حالت استراحت، حضور بهطور میانگین یک کانال باز پتاسیم و یک کانال سدیم در هر غشاء 1 میکرومتر را نشان داد. از این نتیجه می شود که حدود 1000 مولکول از پمپ Na/K باید در همان فضا وجود داشته باشد، یعنی. فاصله بین آنها به طور متوسط 34 نانومتر است. قطر پروتئین پمپ، به عنوان یک پروتئین کانال، 8 تا 10 نانومتر است. بنابراین، غشاء کاملاً متراکم با مولکول های پمپاژ اشباع شده است
این واقعیت که جریان یونهای سدیم به داخل سلول و یونهای پتاسیم به بیرون از سلول با عملکرد پمپ جبران میشود، پیامد دیگری نیز وجود دارد که عبارت است از حفظ فشار اسمزی پایدار و حجم ثابتدر داخل سلول غلظت بالایی از آنیونهای بزرگ، عمدتاً پروتئینها (A - در جدول 1.1) وجود دارد که قادر به نفوذ به غشاء نیستند (یا به آرامی از آن نفوذ میکنند) و بنابراین جزء ثابتی در داخل سلول هستند. برای متعادل کردن بار این آنیون ها به تعداد مساوی کاتیون نیاز است. این کاتیون ها به دلیل عملکرد پمپ Na/K عمدتاً یون های پتاسیم هستند. افزایش قابل توجهی در غلظت یون درون سلولی تنها می تواند با افزایش غلظت آنیون ها به دلیل جریان C1 - در امتداد گرادیان غلظت به داخل سلول اتفاق بیفتد (جدول 1.1)، اما پتانسیل غشاء با این امر مقابله می کند. جریان ورودی Cl – فقط تا زمانی که پتانسیل تعادل برای یون های کلر به دست آید مشاهده می شود. این زمانی مشاهده میشود که گرادیان یون کلر تقریباً مخالف گرادیان یون پتاسیم باشد، زیرا یونهای کلر دارای بار منفی هستند (معادله 4). بنابراین، غلظت کم داخل سلولی یون کلر ایجاد می شود که مربوط به غلظت کم یون های پتاسیم خارج سلولی است. نتیجه این است که تعداد کل یون ها در سلول محدود می شود. اگر پتانسیل غشاء در هنگام مسدود شدن پمپ Na/K کاهش یابد، برای مثال در هنگام آنکسی، پتانسیل تعادل برای یونهای کلر کاهش مییابد و غلظت درون سلولی یونهای کلرید بر این اساس افزایش مییابد. با بازیابی تعادل بارها، یون های پتاسیم نیز وارد سلول می شوند. غلظت کل یون ها در سلول افزایش می یابد که باعث افزایش فشار اسمزی می شود. این امر آب را وارد سلول می کند. سلول متورم می شود. این تورم مشاهده می شود in vivo در شرایط کمبود انرژی
گرادیان غلظت Na + به عنوان یک نیروی محرکه برای حمل و نقل غشا . اهمیت پمپ Na/K برای سلول به تثبیت شیب های طبیعی K + و Na + روی غشاء محدود نمی شود. انرژی ذخیره شده در گرادیان غشا Na+ ، اغلب برای تسهیل حمل و نقل غشایی سایر مواد استفاده می شود. به عنوان مثال، در شکل. 1.10 "simport" را نشان می دهد Na+ و مولکول های قند وارد سلول می شوند. پروتئین ناقل غشایی مولکول قند را حتی بر خلاف گرادیان غلظت به داخل سلول منتقل می کند، در حالی که در همان زمان Na + در امتداد غلظت و گرادیان پتانسیل حرکت می کند و انرژی را برای حمل و نقل قندهااین انتقال قندها کاملاً به وجود یک گرادیان بالا بستگی دارد Na+ ; اگر غلظت درون سلولی باشد Na+ به طور قابل توجهی افزایش می یابد، حمل و نقل قند متوقف می شود. برای انواع مختلفج akharov، سیستم های simport مختلفی وجود دارد. انتقال اسید آمینهبه سلولی مشابه انتقالج آخاروف در شکل نشان داده شده است. 1.10; همچنین توسط گرادیان ارائه می شود Na+ , حداقل پنج سیستم سمپورت مختلف وجود دارد که هر کدام برای یک گروه از اسیدهای آمینه مرتبط تخصص دارند.
علاوه بر سیستم های simport، نیز وجود دارد "ضد باربر".برای مثال یکی از آنها در یک چرخه یک یون کلسیم را در ازای سه یون سدیم ورودی از سلول منتقل می کند (شکل 1.10). انرژی برای Ca 2+ حمل و نقلبه دلیل ورود سه یون سدیم در امتداد غلظت و گرادیان پتانسیل تشکیل می شود. این انرژی (در پتانسیل استراحت) برای حفظ گرادیان یون کلسیم بالا (از کمتر از 10-7 مول در لیتر در داخل سلول تا تقریباً 2 میلی مول در لیتر در خارج از سلول) کافی است.
اندو و اگزوسیتوز . برای مواد خاصی که وارد سلول می شوند یا باید دفع شوند

برنج. 1.10.پروتئینهای تعبیهشده در دولایه لیپیدی غشاء، واسطهای برای انتقال گلوکز و Na + به داخل سلول و همچنین Ca2+/Na+ ضد بندر، که در آن نیروی محرکه گرادیان Na + روی غشای سلولی است
از آن، هیچ کانال حمل و نقل وجود ندارد. این مواد شامل پروتئین ها و کلسترول هستند. آنها می توانند از غشای پلاسمایی عبور کنند وزیکول ها،یا حباب ها،از طریق اندوسیتوز و اگزوسیتوز در شکل شکل 1.11 مکانیسم های اصلی این فرآیندها را نشان می دهد. در اگزوسیتوز، اندامکهای خاصی (به زیر مراجعه کنید) وزیکولهایی را تشکیل میدهند که پر از مادهای هستند که باید از سلول خارج شوند، مانند هورمونها یا آنزیمهای خارج سلولی. وقتی چنین وزیکولهایی به غشای پلاسمایی میرسند، غشای لیپیدی آنها با آن ترکیب میشود و به این ترتیب محتویات به محیط خارجی فرار میکنند. در فرآیند مخالف، اندوسیتوز، غشای پلاسمایی داخل میشود و حفرهای را تشکیل میدهد که سپس عمیق و بسته میشود و یک وزیکول درون سلولی پر از مایع خارج سلولی و برخی درشت مولکولها را تشکیل میدهد. برای اطمینان از همجوشی غشاء و بسته شدن وزیکول، عناصر انقباضی اسکلت سلولی در ارتباط با خود غشاها عمل می کنند (به زیر مراجعه کنید). اندوسیتوز همیشه به سادگی شامل گرفتن محیط خارج سلولی به داخل سلول نیست. غشای سلولی اغلب حاوی گیرنده های خاصی برای ماکرومولکول ها، مانند انسولین یا آنتی ژن ها است که اغلب در گروه های تخصصی سازماندهی می شوند. پس از اتصال این ماکرومولکول ها به گیرنده های خود، اندوسیتوز در ناحیه غشای اطراف گیرنده رخ می دهد و ماکرومولکول به طور انتخابی به داخل سلول منتقل می شود (شکل 1.12، B).
اندوسیتوز و اگزوسیتوز به طور مداوم در سلول ها رخ می دهد. مقدار گردش مواد غشایی قابل توجه است. در عرض 1 ساعت، ماکروفاژ دو برابر سطح غشای سیتوپلاسمی خود را به شکل وزیکول جذب می کند. در اکثر سلول ها، گردش مواد غشایی به شدت اتفاق نمی افتد، اما هنوز باید قابل توجه باشد.

برنج. 1.11.اگزوسیتوز و اندوسیتوز. بالا:وزیکول داخل سلولی با دولایه لیپیدی غشای پلاسمایی ترکیب شده و به فضای خارج سلولی باز می شود. این فرآیند اگزوسیتوز نامیده می شود. در پایین:غشای پلاسمایی در یک ناحیه کوچک داخل میشود و یک وزیکول پر از مواد خارج سلولی آزاد میکند. این فرآیند اندوسیتوز نامیده می شود
اندوسیتوز و اگزوسیتوز نه تنها فرآیندهای انتقال مواد از طریق غشای سلولی هستند، بلکه فرآیندهای تبادل غشاها - اجزای ساختاری خود سلول هستند. موضوع مورد بررسی در این بخش، سایر فرآیندهای انتقال مشابه در سلول و اندامک های آن است.
برنج. 1.12. A–B.طرح فرآیندها شامل اگزوسیتوز و اندوسیتوز. آ.پروتئین سنتز شده در شبکه آندوپلاسمی دانه ای از طریق دستگاه گلژی به غشای پلاسمایی منتقل می شود و در آنجا توسط اگزوسیتوز ترشح می شود. ب.کلسترول متصل به ذرات LDL (لیپوپروتئین با چگالی کم) به غشای پلاسمایی متصل می شود و باعث ایجاد یک وزیکول درون سلولی در این ناحیه از غشاء می شود و به لیزوزوم ها منتقل می شود و در آنجا آزاد می شود. که در.مواد خارج سلولی گرفته شده در طول اندوسیتوز (در شکل سمت راست)از طریق سلول در وزیکول ها یا وزیکول ها منتقل می شود و از طریق اگزوسیتوز آزاد می شود (در شکل ترک کرد)
انتشار . به طور طبیعی، در سیتوزول تفاوت در غلظت به دلیل انتشار حذف می شود. همین امر برای مایعات موجود در اندامک ها نیز صادق است. به دلیل غلظت بالای پروتئین محلول، انتشار در اینجا بسیار کندتر از آب است. غشاهای لیپیدی - در اطراف سلول ها و درون اندامک ها - مایعات دو بعدی هستند که در آنها انتشار رخ می دهد. لیپیدها در یک لایه دوگانه غشایی در داخل لایه خود پخش می شوند و به ندرت از یکی به لایه دیگر منتقل می شوند. پروتئین های غوطه ور در آنها نیز کاملاً متحرک هستند. آنها حول محوری عمود بر غشا می چرخند یا به صورت جانبی با ثابت های انتشار بسیار متفاوت منتشر می شوند، 2 تا 10000 برابر کندتر از فسفولیپیدها. بنابراین، اگر برخی از پروتئینها آزادانه در لایه لیپیدی و با سرعتی برابر با خود مولکولهای لیپیدی حرکت کنند، برخی دیگر لنگر میگیرند، یعنی. کاملا محکم به اسکلت سلولی متصل است. تجمعات "دائمی" از پروتئین های خاص در غشاء وجود دارد، به عنوان مثال، ساختارهای پیش و پس سیناپسی سلول های عصبی. حرکت آزادانه پروتئینها را میتوان با اتصال آنها به رنگهای فلورسنت نشان داد که درخشندگی آن با روشن کردن مختصر ناحیه کوچکی از غشا ایجاد میشود. چنین آزمایشهایی نشان میدهد که در کمتر از 1 دقیقه، پروتئینهای متصل به رنگ به طور یکنواخت روی غشاء در فواصل تا 10 میکرومتر توزیع میشوند.
انتقال فعال در غشاهای اندامک .
فرآیندهای انتقال فعال، که نقش حیاتی در عملکرد غشای پلاسمایی دارند، در داخل سلول، در غشای اندامکها نیز رخ میدهند. محتویات خاص اندامک های مختلف تا حدی به دلیل سنتز داخلی و تا حدی به دلیل انتقال فعال از سیتوزول ایجاد می شود. یکی از نمونههای دومی، پمپ Ca2+ فوق الذکر در شبکه سارکوپلاسمی سلولهای عضلانی است. به ویژه جالب است که در مورد سنتز ATP در میتوکندری، اصل مخالف در مورد آنچه در پمپ های ATPase غشای پلاسمایی رخ می دهد اعمال می شود (شکل 1.6). در طول سنتز ATP، متابولیسم اکسیداتیو منجر به تشکیل یک گرادیان تند می شود H+ روی غشاهای داخلی این گرادیان نیروی محرکه برای فرآیند معکوس چرخه پمپاژ انتقال فعال مولکول ها است: یون های H + در سراسر غشاء در امتداد گرادیان حرکت می کنند و انرژی آزاد شده در نتیجه سنتز ATP از ADP و فسفات را فراهم می کند. ATP حاصل به نوبه خود انرژی سلول را از جمله برای انتقال فعال فراهم می کند.
حمل و نقل در وزیکول . سلول دارای تعداد زیادی اندامک و وزیکول های مرتبط است (شکل 1.1). این اندامک ها و به خصوص وزیکول ها در حرکت دائمی هستند و محتویات خود را به سایر اندامک ها یا به غشای پلاسمایی منتقل می کنند. وزیکول ها همچنین می توانند مانند اندوسیتوز از غشای سلولی به اندامک ها مهاجرت کنند.
روند ترشح پروتئیندر شکل نشان داده شده است. 1.12، آ.این پروتئین در نزدیکی هسته سلول روی ریبوزوم های مرتبط با شبکه آندوپلاسمی (به اصطلاح شبکه آندوپلاسمی دانه ای یا خشن) سنتز می شود. هنگامی که پروتئین وارد شبکه آندوپلاسمی شد، به وزیکول های انتقالی بسته بندی می شود که از اندامک جدا شده و به دستگاه گلژی مهاجرت می کنند. در اینجا آنها با مخزن دستگاه گلژی ادغام می شوند، جایی که پروتئین اصلاح می شود (یعنی تبدیل به گلیکوپروتئین می شود). در انتهای سیسترنا، وزیکول ها دوباره جدا می شوند. وزیکول های ترشحی حامل پروتئین اصلاح شده به غشای پلاسمایی حرکت می کنند و محتویات خود را با اگزوسیتوز آزاد می کنند.
نمونه دیگری از مسیر انتقال در یک سلول در شکل 1 نشان داده شده است. 1.12، B; این جذب کلسترول توسط سلول است. کلسترول منتقل شده در خون عمدتاً به پروتئین ها مانند ذرات متصل می شود "لیپوپروتئین با چگالی کم"(LDL). چنین ذرات به نواحی غشایی خاص حاوی گیرنده های LDL متصل می شوند، جایی که اندوسیتوز رخ می دهد و LDL در وزیکول های "مرز" به داخل سلول منتقل می شود. این وزیکولها با هم ترکیب میشوند تا اندوزومها را تشکیل دهند و در طی این فرآیند «لبه» خود را از دست بدهند. اندوزوم ها به نوبه خود با لیزوزوم های اولیه که حاوی آنزیم های هیدرولیتیک هستند ادغام می شوند و لیزوزوم های ثانویه و بزرگتر را تشکیل می دهند. در آنها، کلسترول از ذرات LDL آزاد می شود و به سیتوزول منتشر می شود، جایی که برای سنتز غشاهای لیپیدی در دسترس می شود. وزیکول هایی که حاوی LDL نیستند نیز از اندوزوم ها جدا می شوند که به روش خاصی به غشای پلاسمایی حرکت کرده و با آن ادغام می شوند و مواد غشایی و احتمالاً گیرنده های LDL را برمی گرداند. از لحظه ای که ذره LDL به غشاء متصل می شود تا زمانی که کلسترول از لیزوزوم ثانویه آزاد شود، 10-15 دقیقه می گذرد. اختلال در اتصال و جذب LDL، به عنوان مثال، در تامین کلسترول به سلول، نقش تعیین کننده ای در ایجاد یک بیماری جدی و گسترده، آترواسکلروز ("سخت شدن" شریان ها) دارد.
بسیاری از مسیرهای حمل و نقل دیگر مشابه آنچه در شکل نشان داده شده است وجود دارد. 1.11 و 1.12، A، که با کمک آنها وزیکول های خاصی در سلول حرکت می کنند. دقیقاً مشخص نیست که چگونه حرکت می کنند، اما عناصر اسکلت سلولی احتمالاً در این فرآیند نقش دارند. وزیکولها میتوانند در امتداد میکروتوبولها بلغزند، در این صورت انرژی لازم برای حرکت ظاهراً توسط پروتئین مرتبط با وزیکولها، ATPase تأمین میشود (به زیر مراجعه کنید). کاملاً مشخص نیست که چه تعداد وزیکول مختلف که یکی پس از دیگری در همه جهات حرکت می کنند، به مقصد می رسند. بدیهی است که آنها باید به گونه ای "علامت گذاری" شوند که توسط سیستم حمل و نقل شناسایی شده و به حرکت هدفمند تبدیل شوند.
حمل و نقل با تشکیل و تخریب اندامک ها . تاکنون اندو و اگزوسیتوز را به عنوان فرآیندهای انتقال محتویات وزیکول ها در نظر گرفته ایم. جنبه دیگری از این فرآیندها وجود دارد که در این واقعیت نهفته است که حذف هدفمند غشای پلاسمایی در یک قسمت از سطح سلول توسط اندوسیتوز و برعکس، اضافه شدن آن در قسمت دیگر توسط اگزوسیتوز باعث حرکت مناطق وسیعی از غشاء می شود (شکل 1.12.E)، به سلول این فرصت را می دهد، برای مثال، یک رشد یا حرکت ایجاد کند.
بازآرایی های مشابهی نیز برای اسکلت سلولی، به ویژه برای ریز رشته ها و میکروتوبول ها، معمول است (شکل 1.1). میکروفیلامنت هادر درجه اول از پروتئین F-اکتین،که در نتیجه پلیمریزاسیون مونومر از سیتوزول قادر است در دسته های فیبری جمع شود. بستهها پلاریزه میشوند، یعنی اغلب فقط در یک انتها رشد میکنند و مولکولهای اکتین جدید را جمع میکنند، در حالی که انتهای دیگر خنثی است یا در اینجا جداسازی اتفاق میافتد. با توجه به این رشد قطبی، میکروفیلامنت ها به طور موثر حرکت می کنند و ساختار شبکه آنها می تواند تغییر کند. انتقال اکتین از حالت دپلیمریزه (سول) به حالت سازمان یافته (ژل) می تواند بسیار سریع تحت تأثیر سایر پروتئین ها یا تغییرات در غلظت یون رخ دهد (به زیر مراجعه کنید). همچنین پروتئین هایی وجود دارند که باعث می شوند رشته های اکتین شکسته شوند و قطعات کوتاهی تشکیل دهند. برجستگی های نازک بسیاری از سلول ها - filopodia - حاوی یک دسته مرکزی از اکتین است (شکل 1.1) و حرکات مختلف فیلوپودیا احتمالاً به دلیل انتقال اکتین است: پلیمریزاسیون - دپلیمریزاسیون.
میکروتوبول هاهمچنین اغلب تحت حرکات مشابهی قرار می گیرند. مکانیسم این حرکات مشابه است - پلیمریزاسیون توبولین از سیتوزول به گونه ای که یکی از انتهای میکروتوبول رشد می کند ، در حالی که دیگری یا تغییر نمی کند یا جداسازی آن در آنجا اتفاق می افتد. بنابراین، یک میکروتوبول، با افزودن یا حذف مناسب مواد، می تواند در سراسر سیتوزول حرکت کند.
حرکات فعال اسکلت سلولی . تغییرات در ساختار اسکلت سلولی می تواند در نتیجه هر دو حرکت فعال و بازآرایی که در بالا توضیح داده شد رخ دهد. در بسیاری از موارد، حرکت میکروتوبولها و رشتههای اکتین توسط پروتئینهای انقباضی انجام میشود که رشتهها یا لولهها را به هم متصل میکنند و میتوانند آنها را نسبت به یکدیگر حرکت دهند. سنجاب ها میوزین و دینئینموجود در سیتوزول تمام سلول ها در غلظت های نسبتاً بالا. آنها عناصری هستند که انرژی را در سلول های تخصصی (عضله) و اندامک ها (سیلیا) به حرکت تبدیل می کنند. در سلول های ماهیچه ای، میوزین رشته های ضخیمی را به موازات رشته های اکتین تشکیل می دهد. مولکول میوزین با "سر" خود به رشته اکتین متصل می شود و با استفاده از انرژی ATP، میوزین را در امتداد مولکول اکتین جابجا می کند.سپس میوزین از اکتین جدا می شود. ترکیب بسیاری از چنین چرخه های اتصال-قطع منجر به یک ماکروسکوپی می شود انقباض فیبرهای عضلانی(فصل 4). Dynein نقش مشابهی را در حرکت میکروتوبول ها در طول مژه ایفا می کند (شکل 1.1). در سیتوپلاسم سلول های غیر تخصصی، میوزین و داینئین فیبرهای منظمی را تشکیل نمی دهند، اما در بیشتر موارد گروه های کوچکی از مولکول ها را تشکیل می دهند. حتی در چنین سنگدانه های کوچکی، آنها قادر به حرکت رشته های اکتین یا میکروتوبول ها هستند. برنج. شکل 1.13 این فرآیند را نشان می دهد که مولکول های میوزین قطبی شده مخالف نیز به دو رشته اکتین متصل می شوند که در جهات مختلف قطبی شده اند. گروه های سر میوزین به سمت دم مولکول خم می شوند و در این فرآیند ATP مصرف می کنند و دو رشته اکتین در جهت مخالف حرکت می کنند و پس از آن میوزین از آنها جدا می شود. حرکاتی از این دست که طی آن انرژی ATP به کار مکانیکی تبدیل می شود، می تواند شکل اسکلت سلولی و در نتیجه سلول را تغییر دهد و همچنین انتقال اندامک های مرتبط با اسکلت سلولی را فراهم کند.
فرآیندهای حمل و نقل درون سلولی را می توان به وضوح در آکسون یک سلول عصبی نشان داد. انتقال آکسوندر اینجا به تفصیل مورد بحث قرار گرفته است تا رویدادهایی را نشان دهد که احتمالاً به روش های مشابه در اکثر سلول ها رخ می دهند. یک آکسونی که فقط چند میکرون قطر دارد میتواند به طول یک متر یا بیشتر برسد و حرکت پروتئینها از طریق انتشار از هسته به انتهای انتهایی آکسون سالها طول میکشد. مدتهاست که مشخص شده است که وقتی هر بخشی از آکسون دچار انقباض می شود، بخشی از آکسون که بیشتر در نزدیکی قرار دارد منبسط می شود. به نظر می رسد که جریان گریز از مرکز در آکسون مسدود شده است. چنین جریان سریع انتقال آکسون می تواندبا حرکت نشانگرهای رادیواکتیو مانند آزمایش نشان داده شده در شکل نشان داده می شود. 1.14. لوسین نشاندار شده رادیواکتیو به گانگلیون ریشه پشتی تزریق شد و سپس رادیواکتیویته در عصب سیاتیک در فاصله 166 میلی متری از اجسام سلول های عصبی از ساعت 2 تا 10 اندازه گیری شد. بیش از 10 ساعت، اوج رادیواکتیویته در محل تزریق کمی تغییر کرد. اما موج رادیواکتیویته با سرعت ثابت حدود 34 میلی متر در 2 ساعت یا 410 میلی متر در روز در امتداد آکسون پخش شد. نشان داده شده است که در تمام نورونهای حیوانات هومیوترمیک، انتقال سریع آکسونی با سرعت یکسان انجام میشود و هیچ تفاوت قابلتوجهی بین رشتههای نازک و بدون میلین و ضخیمترین آکسونها و همچنین بین رشتههای حرکتی و حسی مشاهده نمیشود. نوع نشانگر رادیواکتیو نیز بر سرعت انتقال سریع آکسونی تأثیر نمی گذارد. نشانگرها می توانند به عنوان انواع رادیواکتیو عمل کنند

برنج. 1.13.کمپلکس غیر عضلانی میوزین، با جهت گیری خاصی، می تواند به رشته های اکتین با قطبیت های مختلف متصل شود و با استفاده از انرژی ATP، آنها را نسبت به یکدیگر جابجا کند.
مولکول هایی مانند اسیدهای آمینه مختلف که در پروتئین های بدن سلولی نورون قرار دارند. اگر قسمت محیطی عصب را برای تعیین ماهیت حامل های رادیواکتیویته که به اینجا منتقل می شوند تجزیه و تحلیل کنیم، چنین حامل هایی عمدتاً در بخش پروتئین و همچنین در ترکیب واسطه ها و اسیدهای آمینه آزاد یافت می شوند. با دانستن اینکه خواص این مواد متفاوت است و اندازه مولکول های آنها به ویژه متفاوت است، می توانیم سرعت ثابت انتقال را تنها با مکانیسم انتقال مشترک در همه آنها توضیح دهیم.
در بالا توضیح داده شد انتقال سریع آکسوناست قدامی،یعنی به دور از بدن سلولی هدایت می شود. نشان داده شده است که برخی از مواد با استفاده از محیط به بدن سلولی حرکت می کنند حمل و نقل رتروگرادبه عنوان مثال، استیل کولین استراز در این جهت با سرعتی 2 برابر کمتر از سرعت انتقال سریع آکسونی منتقل می شود. یک نشانگر که اغلب در نوروآناتومی استفاده می شود، ترب کوهی پراکسیداز نیز با انتقال رتروگراد حرکت می کند. انتقال رتروگراد احتمالا نقش مهمی در تنظیم سنتز پروتئین در بدن سلولی ایفا می کند. چند روز پس از برش آکسون، کروماتولیز در بدن سلولی مشاهده می شود که نشان دهنده اختلال در سنتز پروتئین است. زمان لازم برای کروماتولیز با مدت زمان انتقال رتروگراد از محل برش آکسون به بدنه سلولی ارتباط دارد. این نتیجه همچنین توضیحی را برای این اختلال نشان می دهد - انتقال از حاشیه "ماده سیگنال" که سنتز پروتئین را تنظیم می کند مختل می شود. واضح است که "وسایل نقلیه" اصلی برای آکسون سریع استفاده می شود
برنج. 1.14.آزمایش انتقال سریع آکسونی در فیبرهای حسی عصب سیاتیک گربه را نشان می دهد. لوسین نشاندار شده با تریتیوم به گانگلیون ریشه پشتی تزریق می شود و رادیواکتیویته در گانگلیون و فیبرهای حسی در 2، 4، 6، 8 و 10 ساعت پس از تزریق اندازه گیری می شود. (پایین تصویر).توسط محور xفاصله گانگلیون تا مناطقی از عصب سیاتیک که در آن اندازه گیری انجام می شود رسم می شود. در محور ارتین فقط برای منحنی های بالا و پایین، رادیواکتیویته (پالس در دقیقه) در مقیاس لگاریتمی رسم می شود. "موج" افزایش رادیواکتیویته (فلش)با سرعت 410 میلی متر در روز حرکت می کند (در طول)
حمل و نقل هستند وزیکول ها (وزیکول ها) و اندامک ها،مانند میتوکندری که حاوی موادی است که باید حمل شوند. حرکت بزرگترین وزیکول ها یا میتوکندری ها را می توان با استفاده از میکروسکوپ مشاهده کرد in vivo . چنین ذراتی حرکات کوتاه و سریعی در یک جهت انجام می دهند، می ایستند، اغلب کمی به عقب یا به پهلو حرکت می کنند، دوباره می ایستند و سپس در جهت اصلی تکان می خورند. 410 میلی متر در روز مربوط به سرعت متوسط حرکت متجانس تقریباً 5 میکرومتر بر ثانیه است. بنابراین، سرعت هر حرکت فردی باید بسیار بیشتر باشد، و اگر اندازه اندامکها، رشتهها و میکروتوبولها را در نظر بگیریم، این حرکات در واقع بسیار سریع هستند. انتقال سریع آکسونی به غلظت قابل توجهی از ATP نیاز دارد. سمومی مانند کلشی سین که میکروتوبول ها را مختل می کند، انتقال سریع آکسونی را نیز مسدود می کند. از این نتیجه می شود که در فرآیند انتقال مورد نظر ما، وزیکول ها و اندامک ها در امتداد میکروتوبول ها و رشته های اکتین حرکت می کنند. این حرکت توسط تودههای کوچکی از مولکولهای دینئین و میوزین ایجاد میشود که همانطور که در شکل نشان داده شده است عمل میکنند. 1.13، با استفاده از انرژی ATP.
حمل و نقل سریع آکسون نیز ممکن است درگیر باشد فرآیندهای پاتولوژیکبرخی از ویروس های نوروتروپیک (به عنوان مثال، ویروس های تبخال یا فلج اطفال) وارد آکسون در محیط می شوند و از طریق انتقال رتروگراد به بدن نورون حرکت می کنند، جایی که تکثیر می شوند و اثرات سمی خود را اعمال می کنند. سم کزاز، پروتئینی است که توسط باکتریها تولید میشود که وقتی پوست آسیب میبیند وارد بدن میشود، توسط پایانههای عصبی گرفته شده و به بدن نورون منتقل میشود، جایی که باعث اسپاسم عضلانی مشخص میشود. موارد شناخته شده ای از اثرات سمی بر انتقال آکسون وجود دارد، به عنوان مثال، قرار گرفتن در معرض حلال صنعتی آکریل آمید. علاوه بر این، اعتقاد بر این است که پاتوژنز بری بری کمبود ویتامین و پلی نوروپاتی الکلی شامل نقض حمل و نقل سریع آکسونی است.
علاوه بر انتقال سریع آکسونی در سلول، بسیار شدید نیز وجود دارد انتقال آکسون آهستهتوبولین در امتداد آکسون با سرعت حدود 1 میلی متر در روز حرکت می کند و اکتین سریعتر حرکت می کند - تا 5 میلی متر در روز. سایر پروتئین ها با این اجزای اسکلت سلولی مهاجرت می کنند. به عنوان مثال، به نظر می رسد آنزیم ها با اکتین یا توبولین مرتبط هستند. سرعت حرکت توبولین و اکتین تقریباً با نرخ رشد یافت شده برای مکانیسمی که قبلاً توضیح داده شد، مطابقت دارد، جایی که مولکولها در انتهای فعال یک میکروتوبول یا میکروفیلامنت گنجانده میشوند. بنابراین، این مکانیسم ممکن است زمینه ساز انتقال آکسونی کند باشد. سرعت انتقال آکسون آهسته نیز تقریباً با سرعت رشد آکسون مطابقت دارد، که ظاهراً نشان دهنده محدودیت های اعمال شده توسط ساختار اسکلت سلولی در فرآیند دوم است.
در پایان این بخش، باید تاکید کرد که سلول ها به هیچ وجه ساختارهای ایستا نیستند، همانطور که مثلاً در عکس های میکروسکوپی الکترونی ظاهر می شوند. غشای پلاسماییو به خصوص اندامک ها در حرکت سریع و دائمی در حال بازسازی هستند.به همین دلیل است که آنها قادر به عملکرد هستند. علاوه بر این، اینها اتاقک های ساده ای نیستند که در آنها واکنش های شیمیایی انجام می شود، بلکه کنگلومراهای بسیار سازمان یافته از غشاها و الیاف،که در آن واکنش ها در یک توالی سازمان یافته بهینه رخ می دهند.
حفظ تک تک سلول به عنوان یک واحد عملکردی در درجه اول توسط هسته تنظیم می شود. مطالعه چنین مکانیسم های تنظیمی موضوع زیست شناسی سلولی و بیوشیمی است. در عین حال، سلول ها باید عملکرد خود را مطابق با شرایط محیطی و نیازهای سایر سلول های بدن تغییر دهند، یعنی به عنوان اهداف تنظیم عملکردی عمل می کنند. در زیر به طور خلاصه بررسی خواهیم کرد که چگونه این تأثیرات تنظیمی بر غشای پلاسمایی عمل می کنند و چگونه به اندامک های درون سلولی می رسند.
اثرات تنظیمی بر روی غشای سلولی
پتانسیل غشایی . در بسیاری از موارد، تنظیم عملکرد سلولی با تغییر پتانسیل غشا انجام می شود. تغییرات محلی در پتانسیل زمانی امکان پذیر است که: 1) جریان از ناحیه مجاور سلول یا تولید شده توسط سلول دیگر از طریق غشاء جریان یابد. 2) غلظت خارج سلولی یون ها تغییر می کند (اغلب [K + ]بیرون ) 3) کانال های یونی غشایی باز می شوند. تغییرات در پتانسیل غشا می تواند بر روی ترکیب پروتئین های غشایی تأثیر بگذارد و به ویژه باعث باز یا بسته شدن کانال ها شود. همانطور که در بالا توضیح داده شد، عملکرد برخی از پمپ های غشایی به پتانسیل غشا بستگی دارد. سلول های عصبی برای درک تغییرات پتانسیل غشاء به عنوان اطلاعاتی که باید پردازش و منتقل شوند، تخصصی هستند (به فصل 2 مراجعه کنید).
مواد تنظیم کننده خارج سلولی . مهمترین مکانیسم تنظیمی مربوط به مواد خارج سلولی، تعامل آنها با گیرنده های خاص روی غشای پلاسمایی یا داخل سلول است. این مواد شامل فرستنده های سیناپسی هستند که اطلاعات را بین سلول های عصبی، عوامل موضعی و موادی که در خون گردش می کنند و به تمام سلول های بدن می رسند مانند هورمون ها و آنتی ژن ها منتقل می کنند. فرستنده های سیناپسیمولکول های کوچکی هستند که از انتهای عصب در سیناپس آزاد می شوند.
هنگامی که آنها به غشای پلاسمایی یک سلول همسایه و پس سیناپسی می رسند، سیگنال های الکتریکی یا سایر مکانیسم های تنظیمی را تحریک می کنند. این موضوع به تفصیل در فصل مورد بحث قرار گرفته است. 3.
عوامل شیمیایی محلی اغلب توسط سلول های تخصصی ترشح می شود. آنها آزادانه در فضای خارج سلولی منتشر می شوند، اما به دلیل تخریب سریع این مواد، خود به خود یا تحت تأثیر آنزیم ها، عمل آنها به گروه کوچکی از سلول ها محدود می شود. یکی از مصادیق انتشار چنین عواملی رهاسازی است هیستامینماست سل ها به دلیل آسیب یا پاسخ ایمنی. هیستامین باعث شل شدن سلول های ماهیچه صاف عروق، افزایش نفوذپذیری اندوتلیوم عروقی و تحریک انتهای عصبی حسی می شود که واسطه احساس خارش است. سایر عوامل شیمیایی محلی توسط بسیاری از سلول های دیگر آزاد می شوند. عوامل محلی معمولی هستند پروستاگلاندین هاگروهی متشکل از 20 مشتق از اسیدهای چرب را تشکیل می دهد. آنها به طور مداوم از سلول های پراکنده آزاد می شوند، اما فقط به صورت موضعی عمل می کنند، زیرا به سرعت توسط فسفولیپازهای غشایی از بین می روند. پروستاگلاندین های مختلف طیف وسیعی از عمل دارند: آنها می توانند باعث انقباض سلول های ماهیچه صاف شوند، باعث تجمع پلاکت های خون (پلاکت ها) شوند یا رشد جسم زرد را در تخمدان ها سرکوب کنند.
سایر عوامل محلی خدمت می کنند عوامل رشد.شناخته شده ترین فاکتور رشد عصبی (NGF) برای نورون های سمپاتیک است که برای رشد و بقای این نورون ها در طول تکامل ضروری است. in vivo یا در کشت سلولی ظاهراً سلولهای هدف این دسته از نورونها NGF ترشح میکنند و در نتیجه عصببندی مناسب را تضمین میکنند. هنگام تشکیل اندامها، سلولها اغلب باید «راه خود» را برای رسیدن به سلولهای هدف پیدا کنند، که میتوانند در فواصل قابل توجهی قرار گیرند. بر این اساس باید فاکتورهای رشد تخصصی زیادی مشابه NGF وجود داشته باشد.
هورمون ها و آنتی ژن ها توسط خون به تمام سلول ها منتقل می شود. آنتی ژن ها باعث ایجاد پاسخ ایمنی از سلول های حاوی آنتی بادی های خاص می شوند. با این حال، آنتی ژن ها، به عنوان یک قاعده، مواد خارجی هستند که در ارگانیسم واکنش دهنده تولید نمی شوند (برای جزئیات بیشتر، به فصل 18 مراجعه کنید). برخی از هورمون ها، مانند انسولین یا تیروکسین، بر طیف گسترده ای از انواع سلول ها تأثیر می گذارند، در حالی که برخی دیگر، مانند هورمون های جنسی، تنها بر نوع خاصی از سلول تأثیر می گذارند. هورمون ها یا پپتیدهایی هستند که عملکرد آنها با اتصال به گیرنده ای در غشای سلولی ایجاد می شود، یا استروئیدها و تیروکسین هستند که از طریق غشای لیپیدی منتشر شده و به گیرنده های داخل سلولی متصل می شوند. هورمون های استروئیدی به کروماتین هسته ای متصل می شوند و در نتیجه ژن های خاصی رونویسی می کنند. پروتئین های به دست آمده باعث تغییراتی در عملکرد سلولی می شوند که اثر خاص هورمون ها است. مسائل مربوط به آزادسازی و عملکرد هورمون ها به تفصیل در فصل مورد بحث قرار گرفته است. 17.
ارتباط درون سلولی شامل پیام رسان های دوم
عملکردهای تنظیمی که در بالا توضیح داده شد شامل تأثیرات روی غشای سلولی است. اطلاعات دریافت شده توسط غشای سلولی اغلب باید باعث واکنش اندامک ها شود و توسط مواد مختلفی که به عنوان پیام رسان دوم شناخته می شوند (برخلاف پیام رسان های اول که از منابع خارجی به سلول می آیند) به آنها منتقل می شود. مطالعه میانجیهای دوم به سرعت در حال پیشرفت است و هیچ تضمینی وجود ندارد که سطح فعلی درک مشکل به اندازه کافی کامل باشد. در اینجا ما به سه واسطه که به خوبی مطالعه شده اند، خواهیم پرداخت: Ca 2+، cAMP و اینوزیتول تری فسفات.
کلسیم.ساده ترین واسطه درون سلولی یون Ca2+ است. غلظت آزاد آن در یک سلول در حال استراحت بسیار کم است و به 10_-8-10-7 mol/l می رسد. می تواند از طریق کانال های غشایی خاص هنگامی که در حالت باز هستند، به عنوان مثال، زمانی که پتانسیل غشاء تغییر می کند، وارد سلول شود (به فصل 2 مراجعه کنید). افزایش حاصل در غلظت Ca2+ باعث واکنش های مهمی در سلول می شود، مانند انقباض میوفیبریل ها، که اساس انقباض عضلانی است (به فصل 4 مراجعه کنید)، یا آزاد شدن وزیکول های حاوی انتقال دهنده های عصبی از پایانه های عصبی (به فصل 3 مراجعه کنید). . هر دو واکنش به غلظت Ca 2 + تقریباً 10-5 mol / L نیاز دارند. Ca 2+ که اثر تنظیمی دارد می تواند از ذخایر داخل سلولی مانند شبکه آندوپلاسمی نیز آزاد شود. آزادسازی Ca 2+ از انبار مستلزم مشارکت سایر واسطه ها است (برای مثال به شکل 1.16 مراجعه کنید).
آدنوزین مونوفسفات حلقوی، cAMP. اخیراً ثابت شده است که آدنوزین مونوفسفات حلقوی (cAMP)، مشتق شده از منبع اصلی انرژی بدن، ATP، یک پیام رسان مهم دوم است. زنجیره پیچیده واکنش نشان داده شده در شکل. 1.15، با گیرنده شروع می شود R s در سطح بیرونی غشای پلاسمایی، که می تواند به عنوان یک محل اتصال خاص برای واسطه ها و هورمون های مختلف عمل کند. پس از اتصال به یک مولکول "محرک" خاص R s ساختار آن را تغییر می دهد. این تغییرات بر پروتئین تأثیر می گذاردجی اس در سطح داخلی غشا به گونه ای است که فعال سازی دومی توسط گوانوزین تری فسفات داخل سلولی (GTP) امکان پذیر می شود. پروتئین فعال شدهجی اس به نوبه خود، آنزیم روی سطح داخلی غشاء، آدنیلات سیکلاز (AC) را تحریک می کند که تشکیل cAMP از ATP را کاتالیز می کند. cAMP محلول در آب واسطه ای است که اثر را منتقل می کند

برنج. 1.15.زنجیره ای از واکنش ها شامل cAMP پیام رسان درون سلولی (آدنوزین مونوفسفات حلقوی). سیگنال های خارجی تحریکی یا مهاری گیرنده های غشایی R را فعال می کننداس یا ری . این گیرنده ها فرآیند اتصال را تنظیم می کنندجی - پروتئین هایی با GTP درون سلولی (گوانوزین تری فسفات) که در نتیجه باعث تحریک یا مهار آدنیلات سیکلاز داخل سلولی (AC) می شود. آنزیم تقویت کننده AC آدنوزین تری فسفات (ATP) را به cAMP تبدیل می کند که سپس توسط فسفودی استراز به AMP تجزیه می شود. cAMP آزاد به داخل سلول منتشر می شود و آدنیلات کیناز (A-kinase) را فعال می کند و زیرواحد کاتالیزوری C خود را آزاد می کند که فسفوریلاسیون پروتئین های داخل سلولی را کاتالیز می کند. اثر نهایی یک محرک خارج سلولی را تشکیل می دهد. این نمودار همچنین داروهای دارویی و سمومی را نشان میدهد که باعث تحریک (+) یا مهار (-) واکنشهای خاص (اصلاح شده) میشوند.
تحریک گیرنده خارج سلولی R s به ساختارهای داخلی سلول
به موازات زنجیره تحریکی از واکنش های شامل R s اتصال واسطه ها و هورمون های بازدارنده به گیرنده مربوطه امکان پذیر است R i که دوباره از طریق پروتئین فعال شده با GTPجی ، AC و در نتیجه تولید cAMP را مهار می کند. با انتشار به داخل سلول، cAMP با آدنیلات کیناز (A-kinase) واکنش می دهد. این زیر واحد C را آزاد می کند که فسفوریلاسیون پروتئین P را کاتالیز می کند.این فسفوریلاسیون پروتئین ها را به شکل فعال تبدیل می کند و آنها اکنون می توانند اثرات تنظیمی خاص خود را اعمال کنند (مثلاً باعث تخریب گلیکوژن شوند). این سیستم پیچیده تنظیمی بسیار کارآمد است زیرا نتیجه نهایی فسفوریلاسیون بسیاری از پروتئین ها است، یعنی سیگنال تنظیمی با بهره بالا از مدار عبور می کند. واسطه های خارجی که به گیرنده ها متصل می شوند R s و R i خاص هر یک از آنها، بسیار متنوع است. آدرنالین، ارتباط با R s یا R i در تنظیم متابولیسم لیپید و گلیکوژن، و همچنین در افزایش انقباض عضله قلب و در سایر واکنش ها شرکت می کند (به فصل 19 مراجعه کنید). هورمون محرک تیروئید، فعال کردن R s ، ترشح هورمون تیروکسین توسط غده تیروئید را تحریک می کند و پروستاگلاندین I از تجمع پلاکت های خون جلوگیری می کند. اثرات مهاری، از جمله آدرنالین، با واسطه R i در کاهش سرعت لیپولیز بیان می شوند. بدین ترتیب، سیستم cAMP یک سیستم تنظیمی درون سلولی چند منظوره است،که می تواند به طور دقیق توسط مواد تحریک کننده و بازدارنده سیگنال خارج سلولی کنترل شود.
اینوزیتول فسفات "IF" ساعت " سیستم درون سلولی پیام رسان دوم، اینوزیتول فسفات، به تازگی کشف شده است (شکل 1.16). در این مورد، هیچ مسیر مهاری وجود ندارد، اما شباهتی با سیستم cAMP وجود دارد، که در آن اثر تحریک گیرنده R به پروتئین G فعال شده با GTP در سطح داخلی غشاء منتقل می شود. در مرحله بعدی، فسفاتیدیل لینوزیتول لیپیدی غشایی معمول (PI)، که قبلاً دو گروه فسفات اضافی دریافت کرده بود، به PI-دی فسفات (PIP 2) تبدیل می شود که توسط فسفودی استراز فعال شده (PDE) به اینوزیتول تری فسفات(IFz) و لیپید دی اسیل گلیسرول(DAG). اینوزیتول تری فسفات یک پیام رسان دوم محلول در آب است که در سیتوزول منتشر می شود. در درجه اول با آزاد کردن Ca 2 + از شبکه آندوپلاسمی عمل می کند. Ca 2+ به نوبه خود به عنوان یک واسطه عمل می کند، همانطور که در بالا توضیح داده شد. به عنوان مثال، فسفوکیناز وابسته به Ca2+ را فعال می کند که آنزیم ها را فسفریله می کند. زیرواحد لیپیدی DAG (شکل 1.16) نیز حامل سیگنال است و در فاز لیپیدی غشای پلاسمایی به C-kinase واقع در سطح داخلی آن منتشر می شود که با مشارکت فسفاتیدیل سرین به عنوان کوفاکتور فعال می شود. سپس C-kinase باعث فسفوریلاسیون پروتئین ها می شود و آنها را به شکل فعال تبدیل می کند.
سیستم پیام رسان دوم داخل سلولی IF 3 همچنین می تواند توسط انواع واسطه ها و هورمون های خارجی از جمله استیل کولین، سروتونین، وازوپرسین و هورمون محرک تیروئید کنترل شود. مانند سیستم cAMP، با انواع اثرات درون سلولی مشخص می شود. این امکان وجود دارد که این سیستم نیز توسط نور در گیرنده بینایی چشم فعال شود و نقش مرکزی را در انتقال نور ایفا کند (به فصل 11 مراجعه کنید). برای اولین بار در رشد فردی ارگانیسم، گیرنده سیستم IG توسط اسپرم فعال می شود، در نتیجه IG در واکنش های تنظیمی همراه با لقاح تخمک شرکت می کند.
سیستم های cAMP و IFz-DAG بسیار موثر هستند تقویت کننده های بیولوژیکیآنها

برنج. 1.16.زنجیره ای از واکنش ها شامل واسطه درون سلولی IPG (اینوزیتول تری فسفات). همانطور که در سیستم cAMP، سیگنال خارج سلولی از طریق یک پروتئین واسطه می شودجی, که در این حالت فسفودی استراز (PDE) را فعال می کند. این آنزیم فسفاتیدیلینوزین دی فسفات (PIP) را تجزیه می کند 2 ) در غشای پلاسمایی قبل از IFساعت و دی اسیل گلیسرول (DAG)؛ اگرساعت در سیتوپلاسم منتشر می شود. در اینجا باعث انتشار Ca می شود 2+ از شبکه آندوپلاسمی؛ افزایش غلظت کلسیم 2+ در سیتوپلاسم ([Ca 2+ ]i ) پروتئین کیناز را فعال می کند که فسفریله و در نتیجه آنزیم ها را فعال می کند. محصول دیگر، DAG، در غشاء باقی می ماند و پروتئین کیناز C (کوفاکتور فسفاتیدیل سرین، PS) را فعال می کند. پروتئین کیناز C همچنین آنزیم هایی را فسفریله می کند که واسطه اعمال خاص مرتبط با تحریک گیرنده بیرونی هستند.آر . شاخه های زنجیره ای از واکنش ها شامل IFساعت و DAG به ترتیب توسط یونومایسین و فوربول استر می توانند به طور مستقل فعال شوند (اصلاح شده)
واکنش بین یک فرستنده و یک گیرنده غشای خارجی را به فسفوریلاسیون انواع پروتئین های درون سلولی تبدیل می کند که سپس می تواند بر عملکردهای مختلف سلول تأثیر بگذارد. یکی از جنبه های مهم مشکل این است که تا آنجا که امروزه شناخته شده است، تنها این دو سیستم تنظیمی نزدیک به هم از این نوع وجود دارد که توسط واسطه های خارجی متعدد برای تنظیم انواع فرآیندهای درون سلولی استفاده می شود. در عین حال، این سیستم های تنظیمی، از جمله Ca 2+، تعامل نزدیکی با یکدیگر دارند که به آنها اجازه می دهد تا تنظیم دقیق عملکردهای سلولی را انجام دهند.
آموزش ها و راهنماها
1. آلبرتز که در.، بری دی.، لوئیس جی.، راف ام.، رابرتز به.، واتسون جی.دی.زیست شناسی مولکولی سلول، نیویورک و لندن، شرکت انتشارات گارلند، 1983.
2. سیهاک جی.، لونگر اچ.، زیگلر اچ.(ویرایشها). زیست شناسی. برلین، هایدلبرگ، نیویورک، اسپرینگر، 1983.
3. هیل که در. کانال های یونی غشاهای تحریک پذیر. ساندرلند، ماساچوست، Sinauer Assoc.، 1984.
4. هوپ دبلیو.، لومان دبلیو.. مارکی اچ.، زیگلر اچ.(ویرایشها). بیوفیزیک برلین، هایدلبرگ، نیویورک، اسپرینگر، 1984.
5. یونگرمن به.، مالر اچ.بیوشیمی. برلین، هایدلبرگ، نیویورک، اسپرینگر، 1980.
6. کندل ای. آر، شوارتز – جی. اچ،(ویرایشها). اصول علوم عصبی، نیویورک، آمستردام، آکسفورد، الزویر، 1985.
7. شیبلر تی. اچ، اشمیت دبلیو.Anatomic des Menschen. برلین، هایدلبرگ، نیویورک، توکیو، اسپرینگر، 1983.
مقالات و نقدهای اصلی
8. بریج ام.جی.اساس مولکولی ارتباط درون سلولی، علم. عامر، 253، 124-134 (1985).
9. بریج ام جی، ایروین آر.اف.اینوزیتول تری فسفات، دومین پیام رسان جدید در انتقال سیگنال سلولی. طبیعت، 312، 315 321 (1984).
10. برتسچر M.S.مولکول های غشای سلولی، علم. Amer., 253, 124-134 (1985).
11. داوت ج.سلول زنده به عنوان یک ماشین انتقال انرژی یک مدل حداقلی از متابولیسم میوکارد، Biochem. et Biophys. Acta, 895, 41-62 (1987).
12. هوچکین ال.ال.، کاتز که در. تاثیر یون های سدیم بر فعالیت الکتریکی آکسون غول پیکر ماهی مرکب. جی. فیزیول. (Land.), 108, 37-77 (1949).
13. Hodgkin A.L., Keynes R.D.انتقال فعال کاتیون ها در آکسون های غول پیکر از سپیاو لولیگو،جی. فیزیول. (Land.), 128, 28-42 (1955).
14. طولانی تر P.کانال های یونی با زیرشاخه های ساختاری، Biophys. J., 47, 581-590 (1985).
15. Ochs S., Worth P.M.انتقال اگزوپلاسمی در سیستم های نرمال و پاتولوژیک در: فیزیولوژی و آسیب شناسی آکسون ها، S.G. واکسمن،اد. نیویورک، انتشارات ریون، 1978.
خلاصه سایر ارائه ها"روش های تدریس زیست شناسی" - جانورشناسی مدرسه. آشنایی دانش آموزان با استفاده از داده های علمی جانورشناسی. تربیت اخلاقی. برکت اضافی مرغداری. انتخاب روش ها فرآیندهای زندگی ماهی آکواریومی. تغذیه. آموزش محیط زیست. مادی بودن فرآیندهای زندگی نتایج منفی قابل توجه دانشجویان فرم اجباری نگاه کردن به حیوانات کوچک اهداف و مقاصد زیست شناسی. داستان.
"یادگیری مبتنی بر مشکل در درس های زیست شناسی" - دانش. کتاب های درسی جدید. مسیر راه حل. مسئله. سمینارها تکلیف چیست؟ آلبرشت دورر. یادگیری مبتنی بر مسئله در درس زیست شناسی. درس های غیر استاندارد منظور از یادگیری مبتنی بر مسئله چیست؟ کیفیت زندگی. زیست شناسی به عنوان یک موضوع دانشگاهی سوال درس حل مسئله. کاهش علاقه به موضوع. کلاس های آزمایشگاهی مبتنی بر مشکل.
"تفکر انتقادی در درس زیست شناسی" - فناوری "تفکر انتقادی". استفاده از فناوری «توسعه تفکر انتقادی». میز برای درس. انگیزه مطالعه. اکوسیستم ها معنی «توسعه تفکر انتقادی». نشانه های تکنولوژی فناوری RKM ساختار درس جهت های اصلی تاریخچه فناوری. فن آوری های آموزشی. قوانین تکنولوژی تکالیف زیست شناسی فتوسنتز. تکنیک های مورد استفاده در مراحل مختلف درس.
"درس های زیست شناسی با تخته سفید تعاملی" - کتاب های درسی الکترونیک. مزایای برای دانش آموزان. یک تخته سفید تعاملی به انتقال اطلاعات به هر دانش آموز کمک می کند. وظایف آموزشی حل مشکلات بیولوژیکی مزایای کار با وایت بردهای تعاملی کار با ارائه ها روی مقایسه اشیاء کار کنید. اجسام متحرک با استفاده از صفحات گسترده استفاده از تخته سفید تعاملی در فرآیند آموزش به دانش آموزان. مزایا برای معلمان
"رویکرد سیستم-فعالیت در زیست شناسی" - سوالات سمینار. روش فعالیت دریوپیتکوس. مسیر فرازمینی منشأ انسانی. لیزوزوم ها سازمان شیمیایی Gymnosperms. متابولیسم. آنالایزرها رویکرد سیستمی-فعالیتی برای آموزش زیست شناسی. کروموزوم ها سیتوپلاسم. نابینایی طول گوش. طبقه بندی یک شخص. اسکلت یک پستاندار. مسیرهای تکامل انسان میتوز مجتمع سطحی. سوال مشکل ساز هسته پاکت هسته ای
"کامپیوتر در زیست شناسی" - فعالیت مشترک دانش آموزان. خانواده های آنژیوسپرم. آموزش تعاملی مدل های یادگیری نمونه ای از سیستم ارزیابی سوالات کارت آموزش نمونه کارت دستورالعمل محققین. میکروگروه ها فناوری های یادگیری تعاملی چرخ فلک. فناوری های یادگیری تعاملی رویکردهای تعاملی در درس زیست شناسی فرم کار گروهی وظایف برای گروه های "محققان".
