§ 2. Hlavné zložky eukaryotickej bunky
Eukaryotické bunky (obr. 8 a 9) sú organizované oveľa zložitejšie ako prokaryotické. Sú veľmi rôznorodé vo veľkosti (od niekoľkých mikrometrov po niekoľko centimetrov), v tvare a v štruktúrnych znakoch (obr. 10).
Ryža. 8. Štruktúra eukaryotickej bunky. Zovšeobecnená schéma
Ryža. 9. Štruktúra bunky podľa elektrónovej mikroskopie
Ryža. 10. Rôzne eukaryotické bunky: 1 – epitelové; 2 – krv (e – erytroitída, / – leukitída); 3 – chrupavka; 4 – kosti; 5 – hladké svalstvo; 6 – spojivové tkanivo; 7 – nervové bunky; 8 – priečne pruhované svalové vlákno
Všeobecná organizácia a prítomnosť základných komponentov sú však vo všetkých eukaryotických bunkách rovnaké (obr. 11).
Ryža. 11. Eukaryotická bunka (schéma)
Plazmalema (vonkajšia bunková membrána). Základom plazmatickej membrány, podobne ako iných membrán v bunkách (napríklad mitochondrie, plastidy atď.), je vrstva lipidov, ktorá má dva rady molekúl (obr. 12). Keďže molekuly lipidov sú polárne (jeden pól je hydrofilný, t. j. priťahovaný vodou a druhý je hydrofóbny, t. j. odpudzovaný vodou), sú usporiadané v určitom poradí. Hydrofilné konce molekúl jednej vrstvy smerujú do vodného prostredia - do cytoplazmy bunky a druhá vrstva - smerom von z bunky - do medzibunkovej látky (u mnohobunkových organizmov) alebo do vodného prostredia (u jednobunkových organizmov). ).
Ryža. 12. Štruktúra bunkovej membrány podľa modelu fluidnej mozaiky. Proteín a glykoproteíny sú ponorené do dvojitej vrstvy lipidových molekúl, pričom ich hydrofilné konce (kruhy) smerujú von a ich hydrofóbne konce (vlnky) smerujú hlboko do membrány.
Proteínové molekuly sú mozaikovo uložené v bimolekulárnej vrstve lipidov. Na vonkajšej strane živočíšnej bunky sú molekuly polysacharidov pripojené k lipidom a proteínovým molekulám plazmalemy, čím sa vytvárajú glykolipidy a glykoproteíny.
Tento agregát tvorí vrstvu glykokalyx. V spojení s ním funkcia receptora plazmatické membrány (pozri nižšie); môže akumulovať aj rôzne látky využívané bunkou. Glykokalyx navyše zvyšuje mechanickú stabilitu plazmalemy.
V bunkách rastlín a húb sa nachádza aj bunková stena, ktorá hrá podpornú a ochrannú úlohu. V rastlinách pozostáva z celulózy a v hubách z chitínu.
Vonkajšia bunková membrána vykonáva množstvo funkcií, vrátane:
♦ mechanický(nosné, tvarotvorné);
♦ bariérová doprava(selektívna priepustnosť pre rôzne látky: vstup potrebných látok do bunky a odstránenie nepotrebných a škodlivých látok);
♦ receptor(stanovenie rôznych chemikálií nachádzajúcich sa v tesnej blízkosti bunky; vnímanie signálov vo forme hormónov; rozpoznávanie „cudzieho“ proteínu bunkami imunitného systému atď.).
Výmena látok medzi bunkou a prostredím sa uskutočňuje rôznymi spôsobmi - pasívne a aktívne.
Molekuly vody a rôznych iónov pasívne (v dôsledku difúzie, osmózy), bez toho, aby bunka spotrebovala energiu, vstupujú cez špeciálne póry - sú to pasívna doprava. Makromolekuly, ako sú bielkoviny, polysacharidy, dokonca aj celé bunky, sú dodávané napr fagocytóza A pinocytóza so spotrebou energie - aktívny transport.
Fagocytózou dochádza k pohlcovaniu celých buniek alebo veľkých častíc (myslite napríklad na výživu u améb alebo fagocytózu ochrannými krvinkami baktérií). Počas pinocytózy sa absorbujú malé častice alebo kvapôčky tekutej látky. Spoločné pre oba procesy je, že absorbované látky sú obklopené invaginatívnou vonkajšou membránou za vzniku vakuoly, ktorá sa potom presúva hlboko do cytoplazmy bunky.
Exocytóza je proces (ktorý je tiež aktívnym transportom) opačným smerom k fagocytóze a pinocytóze (obr. 13). S jeho pomocou možno odstrániť nestrávené zvyšky potravy v prvokoch alebo biologicky aktívne látky vytvorené v sekrečnej bunke.
Cytoplazma. Cytoplazma je obsah bunky ohraničený plazmalemou, s výnimkou jadra. Obsahuje základná látka (hyaloplazma), organely A inklúzie.
Hyaloplazma- viskózna kvapalina, ktorá môže byť v stave oboch Zola(tekutý), príp gél(rôsolovité).
V prípade potreby je cytoplazma schopná reverzibilného prechodu z jedného stavu do druhého. Napríklad počas pohybu améboidov (pamätajte na časť „Protozoa“ z kurzu zoológie) počas tvorby pseudopodu dochádza k rýchlym prechodom cytoplazmy z gélu na sol a naopak. Je to spôsobené prítomnosťou veľkého počtu molekúl vláknitého proteínu v cytoplazme aktina. Keď sa navzájom spoja a vytvoria trojrozmernú sieť, cytoplazma je v gélovom stave a keď sa sieť rozpadne, je v stave sólu.
Hyaloplazma obsahuje rôzne látky – enzýmy, bielkoviny, sacharidy, tuky a iné, organické a minerálne. Prebiehajú tu rôzne chemické procesy – rozklad látok, ich syntéza a modifikácie (zmeny).
Organoidy. Ide o trvalé zložky bunky s určitou štruktúrou a funkciami, ktoré sa nachádzajú v jej cytoplazme. V ďalšom budeme hovoriť o organoidy na všeobecné použitie, sú vlastné všetkým typom buniek všetkých eukaryotov. Sú spojené so zabezpečením životne dôležitých funkcií tých druhých. Organoidy na špeciálne účely nachádza sa len v bunkách určitého (vysoko špecializovaného) typu – napríklad myofibrily vo svalových bunkách.
Organely všeobecného účelu majú rovnakú štruktúru bez ohľadu na to, ktorým bunkám a organizmom patria. Ale medzi nimi sú skupiny s membránou (endoplazmatické retikulum, Golgiho aparát, mitochondrie, plastidy, lyzozómy, vakuoly), ako aj nemembránové ( ribozómy, bunkové centrum)štruktúru.
Endoplazmatické retikulum (ER). EPS pozostáva z membrán a je to zložitý rozvetvený systém tubulov a cisterien, ktorý preniká celou cytoplazmou bunky (obr. 14). Existujú dva typy EPS - hrubý A hladké. Ribozómy sú pripojené na drsné membrány (zo strany cytoplazmy), ale nie sú prítomné na hladkých membránach.
Ryža. 14. Endoplazmatické retikulum
Endoplazmatické retikulum plní v eukaryotickej bunke množstvo dôležitých funkcií:
♦ ohraničujúce(rozdelenie vnútorného objemu bunky do rôznych reakčných priestorov);
♦ účasť na syntéze organických látok(ribozómy sú umiestnené na membránach drsného ER a komplexy enzýmov sú umiestnené na hladkých, čo zabezpečuje syntézu lipidov, sacharidov atď.);
♦ účasť na tvorbe prvkov Golgiho aparátu, lyzozómov;
♦ transport látok.
Golgiho aparát. Golgiho aparát (AG) je systém tankov(ploché vakuoly) a bubliny(vezikuly), umiestnené v tesnej blízkosti bunkového jadra, ktoré vznikajú vďaka EPS v dôsledku oddeľovania jeho malých fragmentov (obr. 15). Pri splynutí týchto fragmentov vznikajú nové cisterny Golgiho aparátu, pričom sa z EPS transportujú rôzne látky, ktoré sa podieľajú na zostavovaní komplexných organických zlúčenín (bielkoviny + sacharidy, proteíny + lipidy atď.), ktoré sa odstraňujú pomoc AG mimo bunky. Tieto biologicky aktívne látky sú buď z bunky odstránené (pomocou sekrečných vakuol exocytózou), alebo sú súčasťou lyzozómov (pozri nižšie) tvorených AG.
Ryža. 15. Golgiho aparát:
Golgiho aparát vykonáva tieto funkcie:
♦ syntéza biologicky aktívne látky produkované bunkou;
♦ sekrécia (odstránenie z bunky) rôznych látok(hormóny, enzýmy, látky, z ktorých sa buduje bunková stena a pod.);
♦ účasť na tvorbe lyzozómov.
Mitochondrie. Všetky typy eukaryotických buniek majú mitochondrie (obr. 16). Vyzerajú buď ako zaoblené telá alebo tyče, menej často - vlákna. Ich veľkosť sa pohybuje od 1 do 7 mikrónov. Počet mitochondrií v bunke sa pohybuje od niekoľkých stoviek až po desiatky tisíc (u veľkých prvokov).
Ryža. 16. Mitochondrie. Hore - mitochondrie (a) v močových kanáloch, viditeľné vo svetelnom mikroskope. Nižšie je uvedený trojrozmerný model mitochondriálnej organizácie: 1 – cristae; 2 – vonkajšia membrána; 3 – vnútorná membrána; 4 – matica
Mitochondrie tvoria dve membrány - externé A interný, medzi ktorými sa nachádza medzimembránový priestor. Vnútorná membrána tvorí mnoho invaginácií - cristae, čo sú buď platničky alebo rúrky. Táto organizácia poskytuje veľkú plochu vnútornej membrány. Obsahuje enzýmy, ktoré zabezpečujú premenu energie obsiahnutej v organických látkach (sacharidy, lipidy) na energiu ATP, potrebnú pre život bunky. Funkciou mitochondrií je teda participovať na energie bunkové procesy. Preto je veľké množstvo mitochondrií vlastné napríklad svalovým bunkám, ktoré vykonávajú veľa práce.
Plastidy. V rastlinných bunkách sa nachádzajú špeciálne organely - plastidy, ktoré majú často vretenovitý alebo zaoblený tvar, niekedy zložitejší. Existujú tri typy plastidov – chloroplasty (obr. 17), chromoplasty a leukoplasty.
Chloroplasty líšia sa zelenou farbou, čo je spôsobené pigmentom - chlorofyl, zabezpečenie procesu fotosyntéza, teda syntéza organických látok z vody (H 2 O) a oxidu uhličitého (CO 2) pomocou energie slnečného žiarenia. Chloroplasty sa nachádzajú najmä v bunkách listov (vo vyšších rastlinách). Tvoria ich dve membrány umiestnené paralelne k sebe, obklopujúce obsah chloroplastov - stroma. Vnútorná membrána tvorí početné sploštené vaky - tylakoidy, ktoré sú naskladané (ako kopa mincí) – zrná - a ležať v stróme. Práve tylakoidy obsahujú chlorofyl.
Chromoplasty určujú žltú, oranžovú a červenú farbu mnohých kvetov a plodov, v bunkách ktorých sú prítomné vo veľkých množstvách. Hlavné pigmenty v ich zložení sú karotény. Funkčným účelom chromoplastov je prilákať zvieratá farbou, zabezpečiť opelenie kvetov a rozptýlenie semien.
Ryža. 17. Plastidy: a – chloroplasty v bunkách listu Elodea viditeľné vo svetelnom mikroskope; b – schéma vnútornej štruktúry chloroplastu s granami, čo sú stohy plochých vačkov umiestnených kolmo na povrch chloroplastu; c – podrobnejší diagram zobrazujúci anastomózne trubice spájajúce jednotlivé komory ventilátora
Leukoplasty sú bezfarebné plastidy obsiahnuté v bunkách podzemných častí rastlín (napríklad v hľuzách zemiakov), semenách a jadre stoniek. V leukoplastoch sa škrob tvorí najmä z glukózy a hromadí sa v zásobných orgánoch rastlín.
Plastidy jedného typu sa môžu transformovať na iný. Napríklad, keď listy na jeseň zmenia farbu, chloroplasty sa premenia na chromoplasty.
lyzozómy. Tieto organely vyzerajú ako vezikuly obklopené membránou s priemerom až 2 mikróny. Obsahujú niekoľko desiatok enzýmov, ktoré štiepia bielkoviny, nukleové kyseliny, polysacharidy a lipidy. Funkciou lyzozómov je podieľať sa na procesoch vnútrobunkového rozkladu komplexných organických zlúčenín (napríklad živín alebo látok „spotrebovaných“ bunkových zložiek). Lyzozómy sa spájajú s fagocytárnymi (alebo pinocytickými) vakuolami a vytvárajú tráviacu vakuolu.
K tvorbe lyzozómov dochádza v dôsledku pučania z cisterien Golgiho aparátu.
Ribozómy. Ribozómy (obr. 18) sú prítomné v bunkách eukaryotov aj prokaryot, keďže plnia dôležitú funkciu v biosyntéza bielkovín(pozri kapitolu 5). Každá bunka obsahuje desiatky, stovky tisíc (až niekoľko miliónov) týchto malých okrúhlych organel.
Ryža. 18. Schéma štruktúry ribozómu usadeného na membráne endoplazmatického retikula: 1 – malá podjednotka; 2 – tRNA; 3 – aminoacyl-tRNA; 4 – aminokyselina; 5 – veľká podjednotka; 6 – membrána endoplazmatického retikula; 7 – syntetizovaný polypeptidový reťazec
Ribozóm pozostáva z dvoch nerovnakých podjednotiek (častí). Tvoria sa oddelene a kombinujú, „zahŕňajúc“ messenger RNA, počas syntézy proteínovej molekuly. Ribozómy obsahujú rôzne proteíny a ribozomálne RNA.
Bunkové inklúzie. Toto je názov pre nestále zložky v bunke, prítomné v hlavnej substancii cytoplazmy vo forme zŕn, granúl alebo kvapiek. Inklúzie môžu alebo nemusia byť obklopené membránou.
Funkčne existujú tri typy inklúzií: rezervné živiny(škrob, glykogén, tuky, bielkoviny), sekrečné inklúzie(látky charakteristické pre žľazové bunky, nimi produkované - hormóny žliaz s vnútornou sekréciou atď.) a zahrnutie špeciálneho účelu(vo vysoko špecializovaných bunkách, napr. hemoglobín v červených krvinkách).
§ 3. Organizácia bunkového jadra. Chromozómy
Bunkové jadro (pozri obr. 8 a 9) je v živote bunky nanajvýš dôležité, pretože slúži ako úložisko dedičných informácií obsiahnutých v chromozómoch (pozri nižšie).
Jadro je ohraničené jadrovým obalom, ktorý oddeľuje jeho obsah (karyoplazma) z cytoplazmy. Škrupina pozostáva z dvoch membrán oddelených medzerou. Oba sú preniknuté početnými pórmi, vďaka ktorým je možná výmena látok medzi jadrom a cytoplazmou. V bunkovom jadre väčšiny eukaryotov je od 1 do 7 jadierka. Sú s nimi spojené procesy syntézy RNA a tRNA.
Hlavné zložky jadra - chromozómov vytvorený z molekuly DNA a rôznych proteínov. Vo svetelnom mikroskope sú zreteľne viditeľné iba v období delenia buniek (mitóza, meióza). V nedeliacej sa bunke vyzerajú chromozómy ako dlhé tenké vlákna rozmiestnené po celom objeme jadra.
Počas delenia buniek tvoria chromozomálne vlákna husté špirály, v dôsledku čoho sa stávajú viditeľnými (pomocou bežného mikroskopu) vo forme tyčiniek, „vláskov“. Celý objem genetickej informácie je distribuovaný medzi chromozómy jadra. V procese ich štúdia boli identifikované nasledujúce vzory:
♦ v jadrách somatických buniek (t.j. nepohlavných telových bunkách) obsahujú všetci jedinci toho istého druhu rovnaký počet chromozómov, ktoré tvoria súbor chromozómov(obr. 19);
Ryža. 19. Chromozómy rôznych druhov rastlín a živočíchov, zobrazené v rovnakej mierke: 1,2 – améba; 3,4 – rozsievky; 5–8, 18,19 – zelené riasy; 9 – muchovník; 10 – lipa; 11.–12. – Drosophila; 13 – losos; 14 – skerda (čeľaď Asteraceae); 15 – rastlina z čeľade aroidov; 16 – motýľ corydalis; 17 – hmyz z čeľade kobylkových; 20 – vodný chrobák; 21 – ploštica kvetná; 22 – Ambystóm obojživelníkov; 23 – aloe (čeľaď ľaliovitých)
♦ každý druh je charakterizovaný vlastnou chromozómovou sadou podľa ich počtu (napr. človek má 46 chromozómov, mucha drozofilka 8, škrkavka 4, rak 196, kôň 66 a kukurica 104);
♦ chromozómy v jadrách somatických buniek možno zoskupiť do párov tzv homológne chromozómy na základe ich podobnosti (v štruktúre a funkcii);
♦ v jadrách zárodočných buniek (gamét) obsahuje každý pár homológnych chromozómov iba jeden, t.j. celkový súbor chromozómov je polovičný oproti somatickým bunkám;
♦ jedna sada chromozómov v zárodočných bunkách sa nazýva haploidný a označuje sa písmenom n av somatickom - diploidný(2n).
Z vyššie uvedeného je zrejmé, že každý pár homológnych chromozómov vzniká spojením otcovských a materských chromozómov počas oplodnenia, teda splynutím pohlavných buniek (gamét). Naopak, pri tvorbe zárodočných buniek sa z každého páru homológnych chromozómov dostane do gaméty len jeden.
Chromozómy rôzne homológne páry sa líšia veľkosťou a tvarom (obr. 20 a 21).
Ryža. 20. Štruktúra a typy chromozómov: a – vzhľad 1 – centroméra; 2 – krátke rameno; 3 – dlhé rameno); vnútorná štruktúra toho istého chromozómu (1 – centroméra; – molekuly DNA); c – typy chromozómov (1 – jednoramenný; rôznoramenný; 3 – rovnoramenné: X – rameno, V – centroméra)
Ryža. 21. Chromozóm pozostáva z DNA a proteínov. Molekula DNA sa replikuje. Dva identické dvojité závitnice DNA zostávajú spojené v oblasti centroméry. Tieto kópie sa neskôr počas delenia buniek stanú samostatnými chromozómami
V tele chromozómov sú primárna konstrikcia (nazývaná centroméra), ku ktorému sú pripevnené nite štiepne vretená. Rozdeľuje chromozóm na dva rameno Chromozómy môžu byť rovnako ramenné, nerovnaké alebo jednoramenné.
Kapitola 5. Metabolizmus
§ 1. Metabolizmus ako jednota asimilácie a disimilácie
Všetky bunky a živé organizmy sú otvorené systémy, to znamená, že sú v stave neustálej výmeny energií a látok s okolím. V neživej prírode existujú otvorené systémy, ale ich existencia je kvalitatívne odlišná od živých organizmov. Zvážte tento príklad: horiaci kus prírodnej síry je v stave výmeny s prostredím. Pri horení sa absorbuje O 2 a uvoľňuje sa SO 2 a energia (vo forme tepla). Avšak v tomto prípade sa kúsok síry ako fyzického tela zničí a stratí svoju primárnu štruktúru.
Pre živé organizmy sa výmena s prostredím ukazuje ako podmienka zachovania a udržania ich štruktúrnej organizácie prostredníctvom sebaobnovy všetkých látok a zložiek, z ktorých sa skladajú.
Metabolizmus (metabolizmus) je súbor procesov prebiehajúcich v živých organizmoch (spotreba, premena, akumulácia a uvoľňovanie látok a energie), ktoré zabezpečujú ich životnú činnosť, vývoj, rast, rozmnožovanie. Počas metabolického procesu sa molekuly, ktoré tvoria bunky, rozkladajú a syntetizujú; obnova bunkových štruktúr a medzibunkovej hmoty.
Metabolizmus je založený na vzájomne prepojených procesoch asimilácia(anabolizmus) a disimilácia(katabolizmus). Pri asimilácii (výmena plastov) sa z jednoduchých syntetizujú zložité látky. Práve vďaka tomu vznikajú v bunke všetky organické látky potrebné na stavbu jej štruktúrnych zložiek, enzýmových systémov atď. Asimilácia prebieha vždy s výdajom energie.
Pri disimilácii (energetickom metabolizme) sa zložité organické látky štiepia na jednoduchšie alebo anorganické. V tomto prípade sa uvoľňuje energia, ktorú bunka vynakladá na vykonávanie rôznych procesov, ktoré zabezpečujú jej životne dôležitú činnosť (syntéza a transport látok, mechanická práca atď.).
Všetky živé organizmy možno rozdeliť do dvoch skupín: autotrofy A heterotrofy, ktoré sa líšia zdrojmi energie a potrebnými látkami na zabezpečenie ich životných funkcií.
Autotrofy– organizmy, ktoré syntetizujú organické zlúčeniny z anorganických látok pomocou energie slnečného žiarenia (napr fototrofy– rastliny, sinice) alebo energia získaná oxidáciou minerálnych (anorganických) látok (ako napr chemotrofy– sírne baktérie, baktérie železa atď.). V dôsledku toho sú schopné samostatne vytvárať látky potrebné pre ich životnú činnosť.
§ 2. Disimilácia v anaeróbnych a aeróbnych organizmoch
Organizmy možno rozdeliť do dvoch skupín a podľa povahy disimilácie - aeróby A anaeróbov. Aeróby (z gréčtiny. démon– vzduch) vyžadujú pre život voľný kyslík. V anaeróboch (gr. ac– negatívna častica) nie je potrebné. V nich sa disimilácia uskutočňuje fermentáciou – bezkyslíkatým, enzymatickým rozkladom organickej hmoty za vzniku jednoduchších organických látok a uvoľňovaním energie. Napríklad:
♦ mliečna fermentácia:
C6H1206 + 2H3P04 + 2ADP -> 2PH + 2ATP + 2H20;
♦ alkoholové kvasenie:
C6H1206 + 2PH + 2ADP -> 2C2H50H + 2C02 + 2ATP + 2H20.
Látky vznikajúce pri kvasení sú organické a preto obsahujú ešte veľa energie.
Ryža. 22. Vzťahy medzi asimiláciou a disimiláciou v autotrofných a heterotrofných organizmoch
V aeróbnych organizmoch dochádza pri procese dýchania v mitochondriách k úplnému rozkladu organických látok (pomocou O 2) na energeticky chudobné konečné produkty CO 2 a H 2 O a uvoľňuje sa podstatne väčšie množstvo energie:
C 6 H 12 0 6 (glukóza) + 0 2 > 6C0 2 + 6H 2 0 + energia (vďaka ktorej sa syntetizuje 38 molekúl ATP).
Uvažujme vo forme zovšeobecnených schém metabolizmus autotrofných a heterotrofných aeróbnych organizmov (obr. 22).
Asimilácia. Jeho najdôležitejšie procesy sú fotosyntéza A biosyntéza bielkovín.
Spojenie organizmu s prostredím je z fyzikálno-chemického hľadiska otvoreným systémom, teda systémom, kde prebiehajú biochemické procesy. Východiskové látky pochádzajú z prostredia a látky, ktoré tiež priebežne vznikajú, sú vynášané von. Rovnováha medzi rýchlosťou a koncentráciou produktov viacsmerných reakcií v tele je podmienená, imaginárna, pretože príjem a odstraňovanie látok sa nezastaví. Nepretržité prepojenie s prostredím nám umožňuje považovať živý organizmus za otvorený systém.
Pre všetky živé bunky je zdrojom energie Slnko. Rastlinné bunky zachytávajú energiu zo slnečného žiarenia pomocou chlorofylu a využívajú ju na asimilačné reakcie počas procesu fotosyntézy. Bunky zvierat, húb a baktérií využívajú slnečnú energiu nepriamo, pri rozklade organických látok syntetizovaných pozemskými rastlinami.
Niektoré z bunkových živín sa počas bunkového dýchania odbúravajú, čím dodávajú energiu potrebnú pre rôzne typy bunkovej aktivity. Tento proces prebieha v organelách nazývaných mitochondrie. Mitochondrie pozostávajú z dvoch membrán: vonkajšej, ktorá oddeľuje organelu od cytoplazmy, a vnútornej, ktorá tvorí početné záhyby. Hlavným produktom dýchania je ATP. Opúšťa mitochondrie a používa sa ako zdroj energie pre mnohé chemické reakcie v cytoplazme a bunkovej membráne. Ak je pre bunkové dýchanie potrebný kyslík, potom sa dýchanie nazýva aeróbne, ale ak k reakciám dochádza v neprítomnosti kyslíka, hovoríme o anaeróbnom dýchaní.
Pre akýkoľvek typ práce vykonávanej v bunke sa energia využíva v jedinej forme – vo forme energie z fosfátových väzieb ATP. ATP je ľahko mobilná zlúčenina. K tvorbe ATP dochádza na vnútornej membráne mitochondrií. ATP sa syntetizuje vo všetkých bunkách počas dýchania v dôsledku energie oxidácie sacharidov, tukov a iných organických látok. V zelených rastlinných bunkách sa hlavné množstvo ATP syntetizuje v chloroplastoch vďaka slnečnej energii. Počas fotosyntézy produkujú mnohonásobne viac ATP ako mitochondrie. ATP sa rozkladá pretrhnutím väzieb fosfor-kyslík a uvoľnením energie. K tomu dochádza pôsobením enzýmu ATPázy pri hydrolýze ATP – pridaním vody s elimináciou molekuly kyseliny fosforečnej. Výsledkom je, že ATP sa premieňa na ADP a ak sa odštiepia dve molekuly kyseliny fosforečnej, potom na AMP. Eliminačná reakcia každej grammolekuly kyseliny je sprevádzaná uvoľnením 40 kJ. Ide o veľmi veľký energetický výdaj, a preto sa väzby fosfor-kyslík v ATP zvyčajne nazývajú makroergické (vysokoenergetické).
Použitie ATP v plastických výmenných reakciách sa uskutočňuje ich spojením s hydrolýzou ATP. Molekuly rôznych látok sa nabijú energiou pripojením fosforovej skupiny uvoľnenej pri hydrolýze z molekuly ATP, teda fosforyláciou.
Zvláštnosťou fosfátových derivátov je, že nemôžu opustiť bunku, hoci ich „vybité“ formy voľne prechádzajú cez membránu. Vďaka tomu zostávajú fosforylované molekuly v bunke, kým sa nevyužijú v príslušných reakciách.
Opačný proces premeny ADP na ATP nastáva pridaním molekuly kyseliny fosforečnej k ADP, čím sa uvoľní voda a absorbuje sa veľké množstvo energie.
ATP je teda univerzálnym a priamym zdrojom energie pre bunkovú činnosť. To vytvára jediný bunkový fond energie a umožňuje ju prerozdeľovať a prepravovať z jednej oblasti bunky do druhej.
Prenos fosfátovej skupiny hrá dôležitú úlohu pri chemických reakciách, ako je zostavovanie makromolekúl z monomérov. Napríklad aminokyseliny môžu byť spojené do peptidov až po predchádzajúcej fosforylácii. Mechanické procesy kontrakcie alebo pohybu, transport rozpustenej látky proti koncentračnému gradientu a ďalšie procesy zahŕňajú spotrebu energie uloženej v ATP.
Proces energetického metabolizmu možno znázorniť nasledovne. Vysokomolekulárne organické látky v cytoplazme sa enzymaticky hydrolýzou premieňajú na jednoduchšie, z ktorých sa skladajú: bielkoviny - na aminokyseliny, poly- a disacharidy - na monosacharidy (+ glukóza), tuky na glycerol a mastné kyseliny. Neprebiehajú žiadne oxidačné procesy, uvoľňuje sa málo energie, ktorá sa nevyužíva a prechádza do tepelnej formy. Väčšina buniek používa najskôr sacharidy. Polysacharidy (škrob v rastlinách a glykogén u zvierat) sa hydrolyzujú na glukózu. Oxidácia glukózy prebieha v troch fázach: glykolýza, oxidačná dekarboxylácia (Krebsov cyklus – cyklus kyseliny citrónovej) a oxidačná fosforylácia (respiračný reťazec). V cytoplazme prebieha glykolýza, v dôsledku ktorej sa jedna molekula glukózy rozdelí na dve molekuly kyseliny pyrohroznovej s uvoľnením dvoch molekúl ATP. V neprítomnosti kyslíka sa kyselina pyrohroznová premieňa buď na etanol (fermentácia) alebo na kyselinu mliečnu (anaeróbne dýchanie).
Keď sa v živočíšnych bunkách vyskytne glykolýza, molekula glukózy so šiestimi uhlíkmi sa rozpadne na dve molekuly kyseliny mliečnej. Tento proces je viacstupňový. Vykonáva sa postupne 13 enzýmami. Pri alkoholovej fermentácii vznikajú z molekuly glukózy dve molekuly etanolu a dve molekuly CO2.
Glykolýza je spoločná fáza pre anaeróbne a aeróbne dýchanie, ostatné dve sa vyskytujú iba za aeróbnych podmienok. Proces bezkyslíkatej oxidácie, pri ktorej sa uvoľňuje a využíva len časť energie metabolitov, je pre anaeróbne organizmy konečný. V prítomnosti kyslíka kyselina pyrohroznová prechádza do mitochondrií, kde sa v dôsledku množstva sekvenčných reakcií úplne aeróbne oxiduje na H2O a CO2 za súčasnej fosforylácie ADP na ATP. V tomto prípade sú dve molekuly ATP produkované glykolýzou, dve Krebsovým cyklom a 34 dýchacím reťazcom. Čistý výťažok pri úplnej oxidácii jednej molekuly glukózy na H2O a CO2 je 38 molekúl.
V aeróbnych organizmoch sa teda konečný rozklad organických látok uskutočňuje ich oxidáciou vzdušným kyslíkom na jednoduché anorganické látky: CO2 a H2O. Tento proces prebieha na mitochondriách. V tomto prípade sa uvoľní maximálne množstvo voľnej energie, ktorej významná časť je rezervovaná v molekulách ATP. Je ľahké vidieť, že aeróbna oxidácia poskytuje bunke voľnú energiu v najväčšej miere.
V dôsledku katabolizmu sa v bunke hromadia energeticky bohaté molekuly ATP a do vonkajšieho prostredia sa uvoľňuje CO2 a prebytočná voda.
Molekuly cukru, ktoré nie sú potrebné na dýchanie, môžu byť uložené v bunke. Nadbytočné lipidy sa buď odbúrajú, po čom sa produkty ich rozpadu dostávajú do mitochondrií ako substrát pre dýchanie, alebo sa ukladajú ako zásoby v cytoplazme vo forme tukových kvapôčok. Proteíny sa tvoria z aminokyselín vstupujúcich do bunky. Syntéza bielkovín prebieha v organelách nazývaných ribozómy. Každý ribozóm pozostáva z dvoch podčastíc – veľkej a malej: obe podčastice zahŕňajú molekuly proteínov a molekuly RNA.
Ribozómy sú často pripojené k špeciálnemu membránovému systému, ktorý pozostáva z cisterien a vezikúl – takzvané endoplazmatické retikulum (ER); v bunkách, ktoré produkujú veľa bielkovín, je endoplazmatické retikulum často veľmi dobre vyvinuté a pokryté ribozómami. Niektoré enzýmy sú účinné iba vtedy, ak sú pripojené k membráne. Nachádza sa tu väčšina enzýmov podieľajúcich sa na syntéze lipidov. Endoplazmatické retikulum je teda ako druh bunkového pracovného stola.
Okrem toho ER rozdeľuje cytoplazmu do oddelených kompartmentov, t.j. oddeľuje rôzne chemické procesy prebiehajúce súčasne v cytoplazme, a tým znižuje pravdepodobnosť, že tieto procesy budú navzájom interferovať.
Produkty produkované danou bunkou sa často používajú mimo bunky. V takýchto prípadoch proteíny syntetizované na ribozómoch prechádzajú cez membrány endoplazmatického retikula a sú zabalené do membránových vezikúl, ktoré sa okolo nich vytvoria, ktoré sa potom oddelia od ER. Tieto vezikuly, sploštené a naskladané na seba, ako naukladané palacinky, tvoria charakteristickú štruktúru nazývanú Golgiho komplex alebo Golgiho aparát. Počas pobytu v Golgiho aparáte bielkoviny prechádzajú určitými zmenami. Keď príde čas, aby opustili bunku, membránové vezikuly sa spoja s bunkovou membránou a vyprázdnia sa, čím sa ich obsah vyleje, t. j. exocytózou dôjde k sekrécii.
Golgiho aparát produkuje aj lyzozómy – membránové vaky obsahujúce tráviace enzýmy. Zistenie, ako bunka vytvára, balí a vyváža určité proteíny a ako „vie“, ktoré proteíny by si mala nechať pre seba, je jedným z najfascinujúcejších odvetví modernej cytológie.
Membrány akejkoľvek bunky sa neustále pohybujú a menia. ER membrány sa pomaly pohybujú v celej bunke. Jednotlivé úseky týchto membrán sa oddeľujú a vytvárajú vezikuly, ktoré sa dočasne stávajú súčasťou Golgiho aparátu a následne procesom exocytózy splývajú s bunkovou membránou.
Neskôr sa materiál membrány vráti do cytoplazmy, kde sa opäť použije.
Výmena látok vstupujúcich do bunky alebo ňou uvoľňovaných von, ako aj výmena rôznych signálov s mikro- a makroprostredím prebieha cez vonkajšiu membránu bunky. Ako je známe, bunková membrána je lipidová dvojvrstva, do ktorej sú vložené rôzne proteínové molekuly, ktoré fungujú ako špecializované receptory, iónové kanály, zariadenia, ktoré aktívne transportujú alebo odstraňujú rôzne chemikálie, medzibunkové kontakty atď. V zdravých eukaryotických bunkách sú fosfolipidy distribuované v membrána asymetricky: vonkajší povrch pozostáva zo sfingomyelínu a fosfatidylcholínu, vnútorný povrch - z fosfatidylserínu a fosfatidyletanolamínu. Udržanie takejto asymetrie vyžaduje výdaj energie. Preto sa pri poškodení bunky, infekcii alebo energetickom hladovaní vonkajší povrch membrány obohatí o pre ňu nezvyčajné fosfolipidy, ktoré sa pre ostatné bunky a enzýmy stávajú signálom o poškodení buniek so zodpovedajúcou reakciou na to. Najdôležitejšiu úlohu zohráva rozpustná forma fosfolipázy A2, ktorá rozkladá kyselinu arachidónovú a z vyššie uvedených fosfolipidov vytvára lyzoformy. Kyselina arachidónová je limitujúcim článkom pre tvorbu zápalových mediátorov, ako sú eikozanoidy, a ochranné molekuly - pentraxíny (C-reaktívny proteín (CRP), prekurzory amyloidných proteínov) - sú naviazané na lyzoformy v membráne, po ktorých nasleduje aktivácia komplementu systému pozdĺž klasickej dráhy a deštrukcie buniek.
Štruktúra membrány pomáha zachovať vlastnosti vnútorného prostredia bunky, jej odlišnosti od vonkajšieho prostredia. To je zabezpečené selektívnou permeabilitou bunkovej membrány a existenciou aktívnych transportných mechanizmov v nej. Ich narušenie v dôsledku priameho poškodenia, napríklad tetrodotoxínom, ouabaínom, tetraetylamóniom, alebo v prípade nedostatočného zásobovania príslušnými „pumpami“ energiou vedie k narušeniu elektrolytového zloženia bunky, zmenám v jej metabolizme, narušeniu špecifických funkcií - kontrakcia, vedenie excitačných impulzov atď. Narušenie bunkových iónových kanálov (vápnik, sodík, draslík a chlorid) u ľudí môže byť geneticky podmienené aj mutáciami v génoch zodpovedných za štruktúru týchto kanálov. Takzvané kanálopatie spôsobujú dedičné ochorenia nervového, svalového a tráviaceho systému. Nadmerný vstup vody do bunky môže viesť k jej prasknutiu – cytolýze – v dôsledku perforácie membrány pri aktivácii komplementu alebo napadnutia cytotoxickými lymfocytmi a bunkami prirodzených zabíjačov.
Bunková membrána má v sebe zabudované mnohé receptory – štruktúry, ktoré po spojení so zodpovedajúcimi špecifickými signálnymi molekulami (ligandmi) prenášajú signál vo vnútri bunky. K tomu dochádza prostredníctvom rôznych regulačných kaskád pozostávajúcich z enzymaticky aktívnych molekúl, ktoré sú postupne aktivované a v konečnom dôsledku prispievajú k implementácii rôznych bunkových programov, ako je rast a proliferácia, diferenciácia, pohyblivosť, starnutie a bunková smrť. Regulačné kaskády sú pomerne početné, ale ich počet ešte nie je úplne stanovený. Systém receptorov a regulačných kaskád s nimi spojených existuje aj vo vnútri bunky; vytvárajú špecifickú regulačnú sieť s bodmi koncentrácie, distribúcie a výberu ďalšej signálnej cesty v závislosti od funkčného stavu bunky, štádia jej vývoja a súčasného pôsobenia signálov z iných receptorov. Výsledkom toho môže byť inhibícia alebo zosilnenie signálu, ktorý ho nasmeruje pozdĺž inej regulačnej dráhy. Receptorový aparát aj dráhy prenosu signálu cez regulačné kaskády, napríklad do jadra, môžu byť narušené v dôsledku genetického defektu, ktorý sa vyskytuje ako vrodený defekt na úrovni organizmu alebo v dôsledku somatickej mutácie v konkrétnom type bunky. Tieto mechanizmy môžu byť poškodené infekčnými agens, toxíny a tiež sa môžu meniť v procese starnutia. Konečným štádiom toho môže byť narušenie funkcií bunky, procesov jej proliferácie a diferenciácie.
Na povrchu buniek sa nachádzajú aj molekuly, ktoré hrajú dôležitú úlohu v procesoch medzibunkovej interakcie. Môžu to byť bunkové adhézne proteíny, histokompatibilné antigény, tkanivovo špecifické, diferencujúce antigény atď. Zmeny v zložení týchto molekúl spôsobujú narušenie medzibunkových interakcií a môžu spôsobiť aktiváciu vhodných mechanizmov na elimináciu takýchto buniek, pretože predstavujú tzv. určité nebezpečenstvo pre integritu tela ako rezervoár infekcie, najmä vírusovej, alebo ako potenciálne iniciátory rastu nádoru.
Porušenie zásobovania energiou bunky
Zdrojom energie v bunke je potrava, po ktorej rozklade sa energia uvoľňuje na finálne látky. Hlavným miestom výroby energie sú mitochondrie, v ktorých dochádza k oxidácii látok pomocou enzýmov dýchacieho reťazca. Oxidácia je hlavným dodávateľom energie, keďže v dôsledku glykolýzy sa z rovnakého množstva oxidačných substrátov (glukózy) v porovnaní s oxidáciou neuvoľní viac ako 5 % energie. Asi 60% energie uvoľnenej pri oxidácii sa akumuluje oxidačnou fosforyláciou vo vysokoenergetických fosfátoch (ATP, kreatínfosfát), zvyšok sa rozptýli ako teplo. Následne sú vysokoenergetické fosfáty bunkou využívané na procesy, ako je chod pumpy, syntéza, delenie, pohyb, sekrécia atď. Existujú tri mechanizmy, ktorých poškodenie môže spôsobiť narušenie zásobovania bunkou energiou: prvým je mechanizmus syntézy enzýmov energetického metabolizmu, druhý je mechanizmus oxidatívnej fosforylácie, tretí je mechanizmus využitia energie.
Porušenie transportu elektrónov v mitochondriálnom dýchacom reťazci alebo rozpojenie oxidácie a fosforylácie ADP so stratou protónového potenciálu, hnacej sily pre tvorbu ATP, vedie k oslabeniu oxidatívnej fosforylácie takým spôsobom, že väčšina energie sa rozptýli vo forme tepla a počet vysokoenergetických zlúčenín klesá. Rozpojenie oxidácie a fosforylácie pod vplyvom adrenalínu využívajú bunky homeotermických organizmov na zvýšenie tvorby tepla pri udržiavaní stálej telesnej teploty pri ochladzovaní alebo jej zvyšovaní pri horúčke. Pri tyreotoxikóze sa pozorujú významné zmeny v mitochondriálnej štruktúre a energetickom metabolizme. Tieto zmeny sú spočiatku reverzibilné, ale po určitom bode sa stanú nezvratnými: mitochondrie sa fragmentujú, rozpadajú alebo napučia, strácajú kryštály, menia sa na vakuoly a nakoniec akumulujú látky ako hyalín, feritín, vápnik, lipofuscín. U pacientov so skorbutom sa mitochondrie spájajú a vytvárajú chondriosféry, pravdepodobne v dôsledku poškodenia membrány peroxidovými zlúčeninami. K výraznému poškodeniu mitochondrií dochádza vplyvom ionizujúceho žiarenia pri premene normálnej bunky na malígnu.
Mitochondrie sú silným depotom vápnikových iónov, kde je jeho koncentrácia o niekoľko rádov vyššia ako v cytoplazme. Keď sú mitochondrie poškodené, vápnik vstupuje do cytoplazmy, čo spôsobuje aktiváciu proteináz s poškodením vnútrobunkových štruktúr a narušením funkcií príslušnej bunky, napríklad kalciové kontraktúry alebo dokonca „vápnikovú smrť“ v neurónoch. V dôsledku narušenia funkčnej kapacity mitochondrií prudko narastá tvorba voľných radikálových peroxidových zlúčenín, ktoré majú veľmi vysokú reaktivitu, a preto poškodzujú dôležité zložky bunky – nukleové kyseliny, proteíny a lipidy. Tento jav sa pozoruje počas takzvaného oxidačného stresu a môže mať negatívne dôsledky na existenciu bunky. Poškodenie vonkajšej membrány mitochondrií je teda sprevádzané uvoľňovaním látok obsiahnutých v medzimembránovom priestore do cytoplazmy, predovšetkým cytochrómu C a niektorých ďalších biologicky aktívnych látok, ktoré spúšťajú reťazové reakcie spôsobujúce programovanú bunkovú smrť – apoptózu. Poškodzovaním mitochondriálnej DNA reakcie voľných radikálov skresľujú genetickú informáciu potrebnú na tvorbu určitých enzýmov dýchacieho reťazca, ktoré sú produkované špecificky v mitochondriách. To vedie k ešte väčšiemu narušeniu oxidačných procesov. Vo všeobecnosti je vlastný genetický aparát mitochondrií v porovnaní s genetickým aparátom jadra menej chránený pred škodlivými vplyvmi, ktoré môžu zmeniť genetickú informáciu v ňom zakódovanú. Výsledkom je, že dysfunkcia mitochondrií sa vyskytuje počas celého života, napríklad počas procesu starnutia, počas malígnej transformácie bunky, ako aj na pozadí dedičných mitochondriálnych ochorení spojených s mutáciou mitochondriálnej DNA vo vajíčku. V súčasnosti je popísaných viac ako 50 mitochondriálnych mutácií, ktoré spôsobujú dedičné degeneratívne ochorenia nervového a svalového systému. Prenášajú sa na dieťa výlučne od matky, pretože mitochondrie spermií nie sú súčasťou zygoty, a teda ani nového organizmu.
Porušenie uchovávania a prenosu genetickej informácie
Bunkové jadro obsahuje väčšinu genetickej informácie a tým zabezpečuje jeho normálne fungovanie. Prostredníctvom selektívnej génovej expresie koordinuje bunkovú aktivitu počas interfázy, ukladá genetickú informáciu a obnovuje a prenáša genetický materiál počas delenia buniek. K replikácii DNA a transkripcii RNA dochádza v jadre. DNA môžu poškodiť rôzne patogénne faktory, ako je ultrafialové a ionizujúce žiarenie, oxidácia voľnými radikálmi, chemikálie, vírusy. Odhaduje sa, že každá bunka teplokrvného živočícha trvá 1 deň. stratí viac ako 10 000 báz. Tu by sme mali pridať porušenia pri kopírovaní počas delenia. Ak by tieto poškodenia pretrvávali, bunka by nebola schopná prežiť. Ochrana spočíva v existencii výkonných opravných systémov, ako je ultrafialová endonukleáza, opravné replikačné a rekombinačné opravné systémy, ktoré nahrádzajú poškodenie DNA. Genetické defekty v reparačných systémoch spôsobujú rozvoj chorôb spôsobených zvýšenou citlivosťou na faktory, ktoré poškodzujú DNA. Ide o xeroderma pigmentosum, ako aj niektoré syndrómy zrýchleného starnutia, sprevádzané zvýšeným sklonom k vzniku zhubných nádorov.
Systém regulácie procesov replikácie DNA, transkripcie messenger RNA (mRNA) a translácie genetickej informácie z nukleových kyselín do štruktúry proteínov je pomerne zložitý a viacúrovňový. Okrem regulačných kaskád spúšťajúcich pôsobenie transkripčných faktorov s celkovým počtom nad 3000, ktoré aktivujú určité gény, existuje aj viacúrovňový regulačný systém sprostredkovaný malými molekulami RNA (interferujúca RNA; RNAi). Ľudský genóm, ktorý pozostáva z približne 3 miliárd purínových a pyrimidínových báz, obsahuje iba 2 % štrukturálnych génov zodpovedných za syntézu proteínov. Zvyšok zabezpečuje syntézu regulačných RNA, ktoré súčasne s transkripčnými faktormi aktivujú alebo blokujú prácu štruktúrnych génov na úrovni DNA v chromozómoch alebo ovplyvňujú procesy translácie messenger RNA (mRNA) pri tvorbe molekuly polypeptidu v cytoplazme. K porušeniu genetickej informácie môže dochádzať ako na úrovni štruktúrnych génov, tak aj na úrovni regulačnej časti DNA s príslušnými prejavmi v podobe rôznych dedičných ochorení.
V poslednej dobe sa veľká pozornosť venuje zmenám v genetickom materiáli, ktoré sa vyskytujú počas individuálneho vývoja organizmu a sú spojené s inhibíciou alebo aktiváciou určitých úsekov DNA a chromozómov v dôsledku ich metylácie, acetylácie a fosforylácie. Tieto zmeny pretrvávajú dlho, niekedy počas celého života organizmu od embryogenézy až po starobu, a nazývajú sa epigenomická dedičnosť.
Proliferácii buniek so zmenenou genetickou informáciou bránia aj systémy (faktory), ktoré riadia mitotický cyklus. Interagujú s cyklín-dependentnými proteínkinázami a ich katalytickými podjednotkami – cyklínmi – a blokujú bunku v prechode celým mitotickým cyklom, pričom zastavujú delenie na hranici medzi presyntetickou a syntetickou fázou (blok G1/S), kým sa nedokončí oprava DNA, a ak to nie je možné, iniciujú bunky programovanej smrti. Medzi tieto faktory patrí gén p53, ktorého mutácia spôsobuje stratu kontroly nad proliferáciou transformovaných buniek; pozoruje sa takmer u 50 % ľudských rakovín. Druhý kontrolný bod mitotického cyklu je na hranici G2/M. Tu je správna distribúcia chromozomálneho materiálu medzi dcérskymi bunkami v mitóze alebo meióze riadená pomocou súboru mechanizmov, ktoré riadia bunkové vretienko, centrum a centroméry (kinetochory). Neúčinnosť týchto mechanizmov vedie k narušeniu distribúcie chromozómov alebo ich častí, čo sa prejavuje absenciou akéhokoľvek chromozómu v jednej z dcérskych buniek (aneuploidia), prítomnosťou nadbytočného chromozómu (polyploidia), oddelením a. časť chromozómu (delécia) a jeho presun na iný chromozóm (translokácia) . Takéto procesy sú veľmi často pozorované počas proliferácie malígnych degenerovaných a transformovaných buniek. Ak sa to stane počas meiózy so zárodočnými bunkami, vedie to buď k smrti plodu v ranom štádiu embryonálneho vývoja, alebo k narodeniu organizmu s chromozomálnym ochorením.
Nekontrolovaná bunková proliferácia počas rastu nádoru nastáva v dôsledku mutácií v génoch, ktoré riadia bunkovú proliferáciu a nazývajú sa onkogény. Spomedzi viac ako 70 v súčasnosti známych onkogénov väčšina patrí ku komponentom regulácie rastu buniek, niektoré sú reprezentované transkripčnými faktormi, ktoré regulujú aktivitu génov, ako aj faktormi, ktoré inhibujú delenie a rast buniek. Ďalším faktorom obmedzujúcim nadmernú expanziu (šírenie) proliferujúcich buniek je skracovanie koncov chromozómov – telomér, ktoré sa v dôsledku čisto stérickej interakcie nedokážu plne replikovať, preto sa po každom delení bunky teloméry skracujú o určitú časť základov. Proliferujúce bunky dospelého organizmu tak po určitom počte delení (zvyčajne od 20 do 100 v závislosti od typu organizmu a jeho veku) vyčerpajú dĺžku telomér a ďalšia replikácia chromozómov sa zastaví. Tento jav sa nevyskytuje v epiteli spermií, enterocytoch a embryonálnych bunkách v dôsledku prítomnosti enzýmu telomerázy, ktorý obnovuje dĺžku telomér po každom delení. Vo väčšine buniek dospelých organizmov je telomeráza blokovaná, ale, žiaľ, je aktivovaná v nádorových bunkách.
Spojenie medzi jadrom a cytoplazmou a transport látok v oboch smeroch sa uskutočňuje cez póry v jadrovej membráne za účasti špeciálnych transportných systémov, ktoré spotrebúvajú energiu. Takto sa do jadra transportujú energetické a plastické látky, signálne molekuly (transkripčné faktory). Reverzný tok prenáša do cytoplazmy molekuly mRNA a transferovej RNA (tRNA), ribozómy potrebné na syntézu proteínov v bunke. Rovnaký spôsob transportu látok je vlastný aj vírusom, najmä HIV. Prenášajú svoj genetický materiál do jadra hostiteľskej bunky s jeho ďalším začlenením do hostiteľského genómu a prenosom novovytvorenej vírusovej RNA do cytoplazmy na ďalšiu syntézu proteínov nových vírusových častíc.
Porušenie procesov syntézy
Procesy syntézy bielkovín prebiehajú v cisternách endoplazmatického retikula, ktoré sú úzko spojené s pórmi v jadrovej membráne, cez ktoré vstupujú do endoplazmatického retikula ribozómy, tRNA a mRNA. Tu prebieha syntéza polypeptidových reťazcov, ktoré následne získajú svoju finálnu podobu v agranulárnom endoplazmatickom retikule a lamelárnom komplexe (Golgiho komplex), kde prechádzajú posttranslačnou modifikáciou a spájajú sa s molekulami sacharidov a lipidov. Novovzniknuté proteínové molekuly nezostávajú na mieste syntézy, ale komplexným regulovaným procesom tzv proteinkinéza, sú aktívne prenesené do tej izolovanej časti bunky, kde budú vykonávať svoju zamýšľanú funkciu. V tomto prípade je veľmi dôležitým krokom štruktúrovanie prenesenej molekuly do vhodnej priestorovej konfigurácie schopnej vykonávať svoju vlastnú funkciu. K tomuto štruktúrovaniu dochádza pomocou špeciálnych enzýmov alebo na matrici špecializovaných molekúl proteínov - chaperónov, ktoré pomáhajú molekule proteínu, novovzniknutej alebo zmenenej vonkajším vplyvom, získať správnu trojrozmernú štruktúru. V prípade nepriaznivého účinku na bunku, kedy existuje možnosť narušenia štruktúry molekúl proteínov (napríklad pri zvýšení telesnej teploty, infekčnom procese, intoxikácii), sa koncentrácia chaperónov v bunke zvyšuje. ostro. Preto sa takéto molekuly nazývajú aj stresové bielkoviny, alebo proteíny tepelného šoku. Porušenie štruktúrovania molekuly proteínu vedie k tvorbe chemicky inertných konglomerátov, ktoré sa ukladajú v bunke alebo mimo nej pri amyloidóze, Alzheimerovej chorobe a pod. V prípade, že primárna štruktúra nastane nesprávne, všetky nasledujúce molekuly budú tiež chybné. Táto situácia nastáva pri takzvaných priónových ochoreniach (skrapia u oviec, besných kráv, kuru, Creutzfeldt-Jakobova choroba u ľudí), kedy defekt jedného z membránových proteínov nervovej bunky spôsobí následné nahromadenie inertných hmôt vo vnútri bunky. a narušenie jeho životných funkcií.
Narušenie procesov syntézy v bunke môže nastať v rôznych štádiách: transkripcia RNA v jadre, translácia polypeptidov v ribozómoch, posttranslačná modifikácia, hypermetylácia a glykozylácia béžovej molekuly, transport a distribúcia proteínov v bunke a ich odstránenie smerom von. V tomto prípade je možné pozorovať zvýšenie alebo zníženie počtu ribozómov, rozpad polyribozómov, expanziu cisterien granulárneho endoplazmatického retikula, stratu ribozómov a tvorbu vezikúl a vakuol. Pri otrave bledou potápkou sa teda poškodí enzým RNA polymeráza, ktorý naruší transkripciu. Difterický toxín inaktiváciou elongačného faktora narúša translačné procesy, čo spôsobuje poškodenie myokardu. Príčinou narušenia syntézy niektorých špecifických proteínových molekúl môžu byť infekčné agens. Napríklad herpetické vírusy inhibujú syntézu a expresiu molekúl MHC antigénu, čo im umožňuje čiastočne sa vyhnúť imunitnej kontrole bacilov moru - syntéze mediátorov akútneho zápalu; Výskyt nezvyčajných bielkovín môže zastaviť ich ďalšie štiepenie a viesť k akumulácii inertného alebo dokonca toxického materiálu. To môže byť do určitej miery uľahčené narušením hnilobných procesov.
Narušenie hnilobných procesov
Súčasne so syntézou proteínu v bunke neustále prebieha jeho rozklad. Za normálnych podmienok to má dôležitý regulačný a formačný význam, napríklad pri aktivácii inaktívnych foriem enzýmov, proteínových hormónov a proteínov mitotického cyklu. Normálny rast a vývoj buniek si vyžaduje presne riadenú rovnováhu medzi syntézou a degradáciou proteínov a organel. V procese syntézy proteínov sa však v dôsledku chýb v prevádzke syntetizačného aparátu, abnormálnej štruktúre molekuly proteínu a jej poškodenia chemickými a bakteriálnymi činidlami neustále vytvára pomerne veľké množstvo defektných molekúl. Podľa niektorých odhadov ich podiel predstavuje asi tretinu všetkých syntetizovaných bielkovín.
Cicavčie bunky majú niekoľko hlavných Spôsoby ničenia bielkovín: cez lyzozomálne proteázy (pentidhydrolázy), kalcium-dependentné proteinázy (endopeptidázy) a proteazómový systém. Okrem toho existujú aj špecializované proteinázy, ako sú kaspázy. Hlavnou organelou, v ktorej dochádza v eukaryotických bunkách k degradácii látok, je lyzozóm, ktorý obsahuje početné hydrolytické enzýmy. V dôsledku procesov endocytózy a rôznych typov autofágie v lyzozómoch a fagolyzozómoch sú zničené defektné molekuly proteínov aj celé organely: poškodené mitochondrie, úseky plazmatickej membrány, niektoré extracelulárne proteíny a obsah sekrečných granúl.
Dôležitým mechanizmom degradácie proteínov je proteazóm, multikatalytická proteinázová štruktúra komplexnej štruktúry lokalizovaná v cytosóle, jadre, endoplazmatickom retikule a na bunkovej membráne. Tento enzýmový systém je zodpovedný za rozklad poškodených bielkovín, ako aj zdravých bielkovín, ktoré musia byť odstránené pre normálnu funkciu buniek. V tomto prípade sa proteíny, ktoré sa majú zničiť, predbežne kombinujú so špecifickým polypeptidom, ubikvitínom. V proteazómoch však môžu byť čiastočne zničené aj neubikvitinované proteíny. Rozklad proteínových molekúl v proteazómoch na krátke polypeptidy (spracovanie) s ich následnou prezentáciou spolu s molekulami MHC I. typu je dôležitým článkom v imunitnej kontrole antigénnej homeostázy v organizme. Keď je funkcia proteazómu oslabená, hromadia sa poškodené a nepotrebné proteíny, čo sprevádza starnutie buniek. Porušenie degradácie proteínov závislých od cyklínu vedie k narušeniu bunkového delenia, degradácii sekrečných proteínov - k rozvoju cystofibrózy. Naopak, zvýšenie funkcie proteazómu sprevádza vyčerpanie organizmu (AIDS, rakovina).
Pri geneticky podmienených poruchách odbúravania bielkovín nie je organizmus životaschopný a odumiera v skorých štádiách embryogenézy. Ak je štiepenie tukov alebo sacharidov narušené, dochádza k chorobám zo skladovania (tezaurismóza). V tomto prípade sa vo vnútri bunky hromadí nadmerné množstvo niektorých látok alebo produktov ich neúplného rozkladu – lipidov, polysacharidov, čo výrazne poškodzuje funkciu bunky. Najčastejšie sa to pozoruje v pečeňových epiteliálnych bunkách (hepatocytoch), neurónoch, fibroblastoch a makrofagocytoch.
Získané poruchy procesov rozkladu látok môžu vzniknúť v dôsledku patologických procesov (napríklad degenerácia bielkovín, tukov, sacharidov a pigmentov) a sú sprevádzané tvorbou neobvyklých látok. Poruchy v systéme lyzozomálnej proteolýzy vedú k zníženej adaptácii pri hladovaní alebo zvýšenom strese a k výskytu niektorých endokrinných dysfunkcií - zníženej hladiny inzulínu, tyreoglobulínu, cytokínov a ich receptorov. Zhoršená degradácia bielkovín spomaľuje rýchlosť hojenia rán, spôsobuje rozvoj aterosklerózy a ovplyvňuje imunitnú odpoveď. Pri hypoxii zmeny intracelulárneho pH, radiačné poškodenie, charakterizované zvýšenou peroxidáciou membránových lipidov, ako aj vplyvom lyzozomotropných látok - bakteriálne endotoxíny, metabolity toxických húb (sporofusarín), kryštály oxidu kremičitého - stabilita lyzozómovej membrány zmeny, aktivované lyzozomálne enzýmy sa uvoľňujú do cytoplazmy, čo spôsobuje deštrukciu bunkových štruktúr a jej smrť.
Kapitola 1
ZÁKLADY FYZIOLÓGIE BUNIEK
I. Dudel
Plazmatická membrána . Živočíšne bunky sú ohraničené plazmatickou membránou (obrázok 1.1). Jeho štruktúre, ktorá je veľmi podobná stavbe mnohých vnútrobunkových membrán, sa budeme venovať trochu podrobnejšie. Hlavná matrica membrány pozostáva z lipidy, hlavne fosfatidylcholín. Tieto lipidy pozostávajú z hydrofilnej hlavovej skupiny, ku ktorej sú pripojené dlhé hydrofóbne uhľovodíkové reťazce. Vo vode takéto lipidy spontánne vytvárajú dvojvrstvový film s hrúbkou 4–5 nm, v ktorom hydrofilné skupiny smerujú k vodnému médiu a hydrofóbne uhľovodíkové reťazce sú usporiadané v dvoch radoch a tvoria bezvodú lipidovú fázu. Bunkové membrány sú lipidové dvojvrstvy tohto typu a obsahujú glykolipidy, cholesterol a fosfolipidy (obr. 1.2). Hydrofilnú časť glykolipidov tvoria oligosacharidy. Glykolipidy sa vždy nachádzajú na vonkajšom povrchu plazmatickej membrány, pričom oligosacharidová časť molekuly je orientovaná ako vlas ponorený do prostredia. Molekuly cholesterolu, rozptýlené medzi fosfolipidmi v takmer rovnakých množstvách, stabilizujú membránu. Distribúcia rôznych lipidov vo vnútornej a vonkajšej vrstve membrány nie je rovnaká a aj v rámci jednej vrstvy sú oblasti, v ktorých sa koncentrujú určité typy lipidov. Toto nerovnomerné rozdelenie
Ryža. 1.1. Schematický nákres bunky zobrazujúci najdôležitejšie organely
má pravdepodobne nejaký, zatiaľ nejasný, funkčný význam.
Hlavnými funkčnými prvkami zabudovanými v relatívne inertnej lipidovej matrici membrány sú veveričky(obr. 1.2). Proteín podľa hmotnosti predstavuje 25 až 75 % v rôznych membránach, ale keďže molekuly proteínov sú oveľa väčšie ako molekuly lipidov, 50 % hmotnosti zodpovedá pomeru 1 molekuly proteínu k 50 molekulám lipidu. Niektoré proteíny prenikajú membránou z jej vonkajšieho na vnútorný povrch, zatiaľ čo iné sú fixované v jednej vrstve. Molekuly proteínov sú zvyčajne orientované tak, že ich hydrofóbne skupiny sú uložené v lipidovej membráne a polárne hydrofilné skupiny na povrchu membrány sú uložené vo vodnej fáze. Mnohé proteíny na vonkajšom povrchu membrány sú glykoproteíny; ich hydrofilné sacharidové skupiny čelia extracelulárnemu prostrediu.
Membránové systémy intracelulárnych organel .
Približne polovicu objemu bunky zaberajú organely izolované z cytosolu membránami. Celkový povrch membrán intracelulárnych organel je najmenej 10-krát väčší ako povrch plazmatickej membrány. Najpoužívanejší membránový systém je endoplazmatické retikulum, predstavujúce sieť

Ryža. 1.2.Schematické znázornenie plazmatickej membrány. Proteíny sú zabudované vo fosfolipidovej dvojvrstve, pričom niektoré z nich prenikajú do dvojvrstvy, zatiaľ čo iné sú ukotvené iba vo vonkajšej alebo vnútornej vrstve.
vysoko stočené rúrky alebo vakové predĺžené štruktúry; veľké oblasti endoplazmatického retikula sú posiate ribozómami; takéto retikulum sa nazýva zrnité alebo drsné (obr. 1.1). Golgiho aparát pozostáva aj z membránovo viazaných lamiel, z ktorých sa odlamujú vezikuly alebo vezikuly (obr. 1.1). Lyzozómy a peroxizómy- Sú to malé špecializované vezikuly. Vo všetkých týchto rôznych organelách obsahuje membrána a priestor, ktorý zahŕňa, špecifické súbory enzýmov; Vo vnútri organel sa hromadia špeciálne metabolické produkty, ktoré slúžia na vykonávanie rôznych funkcií organel.
JadroA mitochondrie sa líšia tým, že každá z týchto organel je obklopená dvoma membránami. Jadro je zodpovedné za kinetickú kontrolu metabolizmu; zložená vnútorná membrána mitochondrií je miestom oxidačného metabolizmu; tu sa v dôsledku oxidácie pyruvátu alebo mastných kyselín syntetizuje vysokoenergetická zlúčenina adenozíntrifosfát (ATP alebo ATP).
Cytoskelet . Cytoplazma obklopujúca organely nemôže byť v žiadnom prípade považovaná za amorfnú; je prestúpená cytoskeletálnou sieťou. Cytoskelet pozostáva z mikrotubulov, aktínových filament a intermediárnych filament (obr. 1.1). Mikrotubuly majú vonkajší priemer približne 25 nm; vznikajú ako bežný polymér v dôsledku zostavovania molekúl tubulínového proteínu. Aktínové vlákna - kontraktilné vlákna, ktoré sa nachádzajú v blízkej membránovej vrstve a v celej bunke, sa zúčastňujú hlavne procesov spojených s pohybom. Medziľahlé vlákna pozostávajú z blokov rôzneho chemického zloženia v rôznych typoch buniek; tvoria rôzne spojenia medzi dvoma ďalšími vyššie uvedenými cytoskeletálnymi prvkami. S cytoskeletom sú spojené aj organely a plazmatická membrána, ktorá nielen udržiava tvar bunky a polohu organel v nej, ale určuje aj zmeny tvaru bunky a jej pohyblivosti.
Cytosol . Približne polovicu objemu bunky zaberá cytozol. Keďže obsahuje približne 20 % hmotnosti bielkovín, ide skôr o gél ako o vodný roztok. Malé molekuly vrátane organických a anorganických ióny, rozpustený vo vodnej fáze. Medzi bunkou a prostredím (extracelulárny priestor) dochádza k výmene iónov; o týchto metabolických procesoch sa bude diskutovať v ďalšej časti. Koncentrácia iónov v extracelulárnom priestore je udržiavaná so značnou presnosťou na konštantnej úrovni; intracelulárna koncentrácia každého iónu má tiež špecifickú hladinu, odlišnú od hladiny mimo bunky (tabuľka 1.1). Najrozšírenejším katiónom v extracelulárnom prostredí je Na+ v bunke je jeho koncentrácia viac ako 10-krát nižšia. Naopak, koncentrácia K + je najvyššia vo vnútri bunky mimo bunky je nižšia o viac ako rád. Najväčší gradient medzi extracelulárnymi a intracelulárnymi koncentráciami existuje pre Ca 2+, koncentrácia voľných iónov vo vnútri bunky je najmenej 10 000-krát nižšia ako mimo nej. Nie všetky ióny sú rozpustené v cytosóle, niektoré z nich sú adsorbované na proteíny alebo uložené v organelách. Napríklad v prípade Ca 2+ sú viazané ióny oveľa početnejšie ako voľné. Väčšina cytosolických proteínov sú enzýmy, za účasti ktorých sa uskutočňuje mnoho procesov intermediárneho metabolizmu: glykolýza a glukoneogenéza, syntéza alebo deštrukcia aminokyselín, syntéza proteínov na ribozómoch (obr. 1.1). Cytosol obsahuje aj lipidové kvapôčky a glykogénové granule, ktoré slúžia ako zásoby dôležitých molekúl.
Tabuľka 1.1.Intra- a extracelulárne koncentrácie iónov vo svalových bunkách homeotermických zvierat. A – – „bunkové anióny s vysokou molekulovou hmotnosťou“
|
Intracelulárna koncentrácia |
Extracelulárna koncentrácia |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca 2+ |
10 –7 10 –8 mmol/l |
Ca 2+ |
2 mmol/l |
|
C l – |
4 mmol/l |
C1 – |
120 mmol/l |
|
HCO 3 – |
8 mmol/l |
HCO 3 – |
27 mmol/l |
|
A – |
155 mmol/l |
Iní katiónov |
5 mmol/l |
|
Pokojový potenciál –90 mV |
|||
1.2. Výmena látok medzi bunkou a prostredím
Stručne sme opísali štruktúru bunky, aby sme tento opis mohli použiť na zopakovanie základov bunkovej fyziológie. Bunku nemožno v žiadnom prípade považovať za statickú entitu, pretože medzi rôznymi vnútrobunkovými kompartmentmi, ako aj medzi kompartmentmi a prostredím, prebieha neustála výmena látok. Bunkové štruktúry sú v dynamickej rovnováhe a interakcie buniek medzi sebou a s vonkajším prostredím sú nevyhnutnou podmienkou pre udržanie života fungujúceho organizmu. V tejto kapitole sa pozrieme na základné mechanizmy takejto výmeny. V nasledujúcich kapitolách budú tieto mechanizmy uvažované tak, ako sa vzťahujú na nervovú bunku a jej funkcie;
avšak rovnaké mechanizmy sú základom fungovania všetkých ostatných orgánov.
Difúzia.Najjednoduchším procesom pohybu hmoty je difúzia. V roztokoch (alebo plynoch) sa atómy a molekuly voľne pohybujú a rozdiely v koncentráciách sa vyrovnávajú difúziou. Uvažujme dva objemy naplnené kvapalinou alebo plynom (obr. 1.3), v ktorých majú látky koncentrácie c 1 a c 2 a oddelené vrstvou s povrchom A a hrúbkou d. Prúdenie hmoty m za čas t popísané Prvý Fickov zákon difúzie:
dm/ dt= DA/ d ( C 1 –С 2)=DA/ dD C(1)
kde D je difúzny koeficient, konštantný pre danú látku, rozpúšťadlo a teplotu. Vo všeobecnejšej forme pre rozdiel v koncentrácii dc vo vzdialenosti dx
dm/dt= –D A dc/dx,(2)
prietok cez sekciu A je úmerný koncentračnému gradientu dc/dx . V rovnici sa objaví znamienko mínus, pretože zmena koncentrácie v smere x je záporná.
Difúzia je najdôležitejší proces, vďaka ktorému sa väčšina molekúl vo vodných roztokoch pohybuje na krátke vzdialenosti. To platí aj pre ich pohyb v bunke, pokiaľ difúzii nebránia membrány. Mnoho látok môže voľne difundovať cez lipidové membrány, najmä voda a rozpustené plyny ako O 2 a CO 2 . Rozpustný v tukoch

Ryža. 1.3.Kvantitatívna difúzna schéma. Tieto dva priestory sú oddelené vrstvou hrúbkyda oblasť A. C – vysoká koncentrácia častíc v ľavej časti objemu, C: – nízka koncentrácia častíc v pravej diely, ružový povrch– koncentračný gradient v difúznej vrstve. Difúzny prietok dm/dt – viď rovnica (1)
látky tiež dobre difundujú cez membrány; To platí aj pre polárne molekuly pomerne malej veľkosti, ako je etanol a močovina, zatiaľ čo cukry prechádzajú cez lipidovú vrstvu ťažko. Súčasne sú lipidové vrstvy prakticky nepriepustné pre nabité molekuly, vrátane anorganických iónov. Pre neelektrolyty sa difúzna rovnica (1) zvyčajne transformuje spojením charakteristík membrány a difúznej látky do jednej parametrická priepustnosť (P):
dm/dt=PAD c.(3)
Na obr. 1,4 sa porovnávajú permeability (P) lipidovej membrány pre rôzne molekuly.
Difúzia cez membránové póry . Plazmatická membrána (a ďalšie bunkové membrány) je priepustná nielen pre látky difundujúce cez lipidovú vrstvu, ale aj pre mnohé ióny, cukry, aminokyseliny a nukleotidy. Tieto látky prechádzajú cez membránu vytvorenými pórmi transportné proteíny, ponorený do membrány. Vo vnútri takýchto proteínov je kanál naplnený vodou s priemerom menším ako 1 nm, cez ktorý môžu difundovať malé molekuly. Pohybujú sa pozdĺž koncentračného gradientu a ak nesú náboj, potom ich pohyb kanálmi je tiež regulovaný membránovým potenciálom. Membránové kanály sú relatívne selektívne

Ryža. 1.4.Priepustnosť umelých lipidových dvojvrstiev pre rôzne látky
vo vzťahu k typu molekúl, ktoré nimi môžu prechádzať, existujú napríklad draslíkové, sodíkové a vápnikové kanály, z ktorých každý je nepriepustný pre takmer akýkoľvek ión okrem špecifického. Takéto selektívnosť je určený nábojom alebo štruktúrou väzobných miest v stenách kanála, čo uľahčuje transport špecifickej molekuly a zabraňuje prenikaniu iných látok cez kanál (obr. 1,5, A) .
Za správaním membránové iónové kanályľahko pozorovateľné, pretože prúd vznikajúci pri pohybe iónov možno merať aj pre jeden kanál. Ukazuje sa, že kanály spontánne a s vysokou frekvenciou menia svoj stav z otvoreného na uzavretý. Draslíkový kanál je charakterizovaný prúdovými impulzmi s amplitúdou asi 2 pA (2 10 –12 A) a trvaním niekoľkých milisekúnd (pozri obr. 2.12, s. 37) [3]. Počas tohto obdobia ním prejdú desaťtisíce iónov. Prechod proteínov z jednej konformácie do druhej sa študuje pomocou rôntgenovej difrakcie, Mössbauerovej spektroskopie a nukleárnej magnetickej rezonancie (NMR). Proteíny sú teda veľmi dynamické, pohyblivé štruktúry a kanál prechádzajúci proteínom nie je len tuhá trubica naplnená vodou (obr. 1.5, A), ale labyrint rýchlo sa pohybujúcich molekulárnych skupín a nábojov. Táto dynamická charakteristika kanála sa odráža v energetický profil kanála, znázornené na obr. 1.5, B. Os x tu predstavuje dĺžku kanála od vonkajšieho roztoku s koncentráciou iónov C 0 a potenciálom 0 k vnútornému roztoku s koncentráciou C 1 a potenciálom E. Ordináta

Ryža. 1.5.A. Schéma proteínu tvoriaceho draslíkový kanál uložený v lipidovej dvojvrstve plazmatickej membrány. Na „stene“ kanála sú pripevnené štyri záporné náboje. B. Schematický energetický profil kanála znázorneného na obr. A. Zvislá os ukazuje hodnoty kinetickej energie potrebnej na prechod kanála; pozdĺž osi x – vzdialenosť medzi vnútorným a vonkajším povrchom membrány. Energetické minimá zodpovedajú väzbovým miestam kladne nabitých iónov s fixnými zápornými nábojmi v stene kanála. Energetické maximá zodpovedajú difúznym prekážkam v kanáli. Predpokladá sa, že konformácia kanálového proteínu spontánne osciluje; možnosti energetického profilu sú zobrazené plnými a prerušovanými čiarami; tieto oscilácie výrazne uľahčujú viazanie iónov pri prekonávaní energetickej bariéry (modifikované)
sú prezentované hladiny iónovej energie na väzbových miestach kanála; vrchol v grafe predstavuje bariéru permeability, ktorú musí energia iónov prekročiť, aby prenikla kanálom, a „ponor“ v grafe predstavuje relatívne stabilný stav (väzba). Napriek prekážke energetického vrcholu môže ión preniknúť kanálom, ak sa energetický profil spontánne zmení; ión sa tak môže náhle ocitnúť „na druhej strane“ energetického vrcholu a môže pokračovať v pohybe do bunky. V závislosti od náboja, veľkosti a stupňa hydratácie iónu a jeho schopnosti viazať sa na štruktúry steny kanála sa energetický profil kanála líši pre rôzne ióny, čo môže vysvetľovať selektivitu jednotlivých typov kanálov.
Difúzna rovnováha iónov . Difúzia rôznych iónov cez membránové kanály by mala viesť k eliminácii rozdielov v koncentráciách medzi extra- a intracelulárnym prostredím. Ako však možno vidieť z tabuľky. 1.1, takéto rozdiely pretrvávajú, preto nejaké musia byť rovnováha medzi difúziou a inými transportnými procesmi cez membránu. Nasledujúce dve časti sa zaoberajú spôsobmi, akými sa táto rovnováha dosahuje. V prípade iónov je difúzna rovnováha ovplyvnená ich nábojom. Difúzia nenabitých molekúl je zabezpečená rozdielom koncentrácií dc a keď sa koncentrácie vyrovnajú, samotný transport sa zastaví. Nabité častice sú navyše ovplyvnené elektrickým poľom. Napríklad, keď ión draslíka opustí bunku pozdĺž jej koncentračného gradientu, nesie jediný kladný náboj. Vnútrobunkové prostredie sa tak stáva viac negatívne nabité, čo vedie k rozdielu potenciálov cez membránu. Vnútrobunkový záporný náboj bráni novým draselným iónom opustiť bunku a tie ióny, ktoré bunku opustia, budú ďalej zvyšovať náboj na membráne. Tok draselných iónov sa zastaví, keď pôsobenie elektrického poľa kompenzuje difúzny tlak v dôsledku rozdielu koncentrácie. Ióny naďalej prechádzajú cez membránu, ale v rovnakom množstve v oboch smeroch. Preto pre daný rozdiel v koncentráciách iónov na membráne existuje rovnovážny potenciál E ión pri ktorej sa zastaví tok iónov cez membránu. Rovnovážny potenciál možno ľahko určiť pomocou Nernstove rovnice:
Eión= RT/ zF* lnC von/ C in(4)
kde R – plynová konštanta, T – absolútna teplota, z – valencia iónov (záporné pre anióny) C von - koncentrácia extracelulárnych iónov, C in - intracelulárna koncentrácia iónov, F Faradayovo číslo. Ak do rovnice dosadíme konštanty, potom pri telesnej teplote (T = 310 K) je rovnovážny potenciál pre ióny draslíka E K sa rovná:
Ek= –61 mB denník /(5)
Ak [ K + out ]/[ K + in ] = 39, ako vyplýva z tabuľky. 1.1 potom
Ek= –61 m B log 39= –97 mV.
Skutočne sa zistilo, že všetky bunky majú membránový potenciál; vo svalových bunkách cicavcov je jeho hladina asi -90 mV. V závislosti od podmienok a relatívnych koncentrácií iónov môžu mať bunky membránový potenciál v rozsahu od -40 do -120 mV. Pre bunku v príklade vyššie (tabuľka 1.1) oddychový potenciál, približne -90 mV, znamená, že toky draselných iónov cez membránové kanály sú približne v rovnováhe. To nie je prekvapujúce, pretože v pokojovej membráne je s najväčšou pravdepodobnosťou otvorený stav draslíkových kanálov, t.j. membrána je najviac priepustná pre ióny draslíka. Membránový potenciál je však určený tokmi iných iónov.
Ľahkosť, s akou môžu nenabité častice difundovať cez membránu, je kvantifikovaná v rovnici (3). Priepustnosť pre nabité častice je opísaná trochu zložitejšou rovnicou:
P= m RT/ dF(6)
Kde m- pohyblivosť iónov v membráne, d - hrúbka membrány, R, T a F – známe termodynamické konštanty. Takto stanovené hodnoty permeability pre rôzne ióny možno použiť na výpočet membránového potenciálu Em , keď ióny draslíka, sodíka a chlóru prechádzajú cez membránu súčasne (s permeabilitou P K, P Na a P Cl v uvedenom poradí). Predpokladá sa, že potenciál v membráne rovnomerne klesá, takže intenzita poľa je konštantná. V tomto prípade platí Goldmanova rovnica alebo rovnica konštantného poľa :
Em= RT/F*ln(PK + PNa + PCl)/ (PK + PNa + PCl)(7)
Pre väčšinu bunkových membrán P K približne 30-krát vyšší ako P Na (pozri tiež časť 1.3). Relatívna hodnota P Cl sa veľmi líši; pre mnohé membrány P Cl malý v porovnaní s R K , avšak pre iné (napríklad v kostrových svaloch) P Cl , výrazne vyššia ako P K.
Aktívny transport, sodíkové čerpadlo . Predchádzajúca časť popisuje pasívnu difúziu iónov a výsledný membránový potenciál pri daných intra- a extracelulárnych koncentráciách iónov. V dôsledku tohto procesu sa však koncentrácia iónov vo vnútri bunky automaticky nestabilizuje, pretože membrána
potenciál je o niečo elektronegatívny ako EK, a veľa v porovnaní s E Na (približne +60 mV). V dôsledku difúzie sa intracelulárne koncentrácie iónov, aspoň draslíka a sodíka, musia vyrovnať s extracelulárnymi. Stabilita iónového gradientu sa dosahuje aktívnym transportom: membránové proteíny transportujú ióny cez membránu proti elektrickým a/alebo koncentračným gradientom, pričom na to spotrebúvajú metabolickú energiu. Najdôležitejším procesom aktívneho transportu je práca Na/K – čerpadlo, ktoré existuje takmer vo všetkých bunkách;
pumpa pumpuje sodíkové ióny von z bunky a súčasne pumpuje draselné ióny do bunky. To zaisťuje nízku intracelulárnu koncentráciu sodných iónov a vysokú koncentráciu draselných iónov (tabuľka 1.1). Koncentračný gradient sodíkových iónov na membráne má špecifické funkcie súvisiace s prenosom informácií vo forme elektrických impulzov (pozri časť 2.2), ako aj so zachovaním ďalších aktívnych transportných mechanizmov a reguláciou objemu buniek (pozri nižšie). Preto nie je prekvapujúce, že viac ako 1/3 energie spotrebovanej článkom sa minie na pumpu Na/K a v niektorých najaktívnejších článkoch až 70 % energie minie na jej prevádzku.
Transportný proteín Na/K je ATPáza. Na vnútornom povrchu membrány štiepi ATP na ADP a fosfát (obr. 1.6). Energia jednej molekuly ATP sa využíva na transport troch iónov sodíka z bunky a súčasne dvoch iónov draslíka do bunky, t.j. celkovo sa z bunky odstráni jeden kladný náboj v jednom cykle. Na/K čerpadlo je teda elektrogénne(vytvára elektrický prúd cez membránu), čo vedie k zvýšeniu elektronegativity membránového potenciálu približne o 10 mV. Transportný proteín vykonáva túto operáciu vysokou rýchlosťou: od 150 do 600 sodíkových iónov za sekundu. Aminokyselinová sekvencia transportného proteínu je známa, ale mechanizmus tohto komplexného metabolického transportu ešte nie je jasný. Tento proces je opísaný pomocou energetických profilov prenosu sodíkových alebo draselných iónov proteínmi (obr. 1.5,5). Podľa povahy zmien v týchto profiloch, spojených s neustálymi zmenami v konformácii transportného proteínu (proces vyžadujúci energiu), je možné posúdiť stechiometriu výmeny: dva draselné ióny sa vymenia za tri sodíkové ióny.
Na/K čerpadlo, rovnaké ako izolované Na+ /K+-dependentná membránová ATPáza, špecificky inhibovaná srdcovým glykozidom ouabaínom (strofantínom). Keďže prevádzka Na/K pumpy je viacstupňová chemická reakcia, je ako všetky chemické reakcie do značnej miery závislá od teploty, ktorá

Ryža. 1.6.Schéma Na/K pumpy-ATPázy (ponorenej v lipidovej dvojvrstve plazmatickej membrány), ktorá v jednom cykle odoberá z bunky tri ióny Na + proti potenciálu a koncentračným gradientom a privádza do bunky dva ióny K + . Počas tohto procesu sa jedna molekula ATP rozloží na ADP a fosfát. Diagram ukazuje ATPázu ako dimér pozostávajúci z veľkej (funkčnej) a malej podjednotky; v membráne existuje ako tetramér tvorený dvoma veľkými a dvoma malými podjednotkami
znázornené na obr. 1.7. Tu je znázornený tok sodíkových iónov zo svalových buniek vzhľadom na čas; toto je prakticky ekvivalentné toku sodíkových iónov sprostredkovanom prevádzkou Na/K pumpy, pretože pasívny tok sodíkových iónov proti koncentračným a potenciálnym gradientom je extrémne malý. Ak sa liek ochladí približne o 18°C, tok sodíkových iónov z bunky sa rýchlo 15-násobne zníži a ihneď po zahriatí sa vráti na pôvodnú úroveň. Tento pokles toku sodíkových iónov z bunky je niekoľkonásobne väčší, ako by zodpovedalo teplotnej závislosti difúzneho procesu alebo jednoduchej chemickej reakcie. Podobný efekt sa pozoruje pri vyčerpaní metabolickej energie v dôsledku otravy dinitrofenolom (DNP) (obr. 1.7,5). Následne je tok sodíkových iónov z bunky zabezpečený energeticky závislou reakciou – aktívnou pumpou. Ďalšou charakteristikou čerpadla, spolu s významnou teplotnou a energetickou závislosťou, je prítomnosť úrovne nasýtenia (ako všetky ostatné chemické reakcie); to znamená, že rýchlosť pumpy sa nemôže zvyšovať donekonečna so zvyšujúcou sa koncentráciou transportovaných iónov (obr. 1.8). Naproti tomu tok pasívne difundujúcej látky sa zvyšuje úmerne s rozdielom koncentrácií v súlade so zákonom difúzie (rovnice 1 a 2).

Ryža. 1.7. A, B. Aktívna doprava Na + . os Y: tok rádioaktívneho 24 Na + z bunky (imp./min). Abscisová os:čas od začiatku experimentu. A. Bunka sa ochladí z 18,3 °C na 0,5 °C; tok Na+ z bunky počas tohto obdobia je inhibovaný. B. Potlačenie toku Na + z bunky dinitrofenolom (DNP) v koncentrácii 0,2 mmol/l (upravené)
Plazmatická membrána obsahuje okrem Na/K pumpy ešte aspoň jednu pumpu – vápnik; táto pumpa pumpuje ióny vápnika (Ca 2+) von z bunky a podieľa sa na udržiavaní ich vnútrobunkovej koncentrácie na extrémne nízkej úrovni (tabuľka 1.1). Kalciová pumpa je prítomná vo veľmi vysokej hustote v sarkoplazmatickom retikule svalových buniek, ktoré akumulujú ióny vápnika v dôsledku rozkladu molekúl ATP (pozri kapitolu 4).
Vplyv Na/K pumpy na membránový potenciál a objem buniek . Na obr. Obrázok 1.9 ukazuje rôzne zložky membránového prúdu a ukazuje intracelulárne koncentrácie iónov, ktoré

Ryža. 1.8.Vzťah medzi rýchlosťou transportu molekúl a ich koncentráciou (na vstupe do kanála alebo na väzobnom mieste pumpy) počas difúzie cez kanál alebo počas čerpacieho transportu. Ten sa pri vysokých koncentráciách nasýti (maximálna rýchlosť, Vmax ) hodnota na osi x zodpovedajúca polovici maximálnej rýchlosti čerpadla ( Vmax /2), je rovnovážna koncentrácia TO m

Ryža. 1.9.Diagram znázorňujúci koncentrácie Na+ , K+ a Cl – vnútri a mimo bunky a cesty pre prenikanie týchto iónov cez bunkovú membránu (cez špecifické iónové kanály alebo pomocou Na/K pumpy. Pri daných koncentračných gradientoch rovnovážne potenciály E Na, E K a E С l – sa rovná uvedeným, membránový potenciál Em = – 90 mV
zabezpečiť ich existenciu. Cez draslíkové kanály sa pozoruje vonkajší prúd draslíkových iónov, pretože membránový potenciál je o niečo elektropozitívnejší ako rovnovážny potenciál draslíkových iónov. Celková vodivosť sodíkových kanálov je oveľa nižšia ako vodivosť draslíkových kanálov, t.j. sodíkové kanály sú pri pokojovom potenciáli otvorené oveľa menej často ako draslíkové kanály; avšak približne rovnaký počet sodíkových iónov vstupuje do bunky ako draselné ióny opúšťajú bunku, pretože na difúziu sodíkových iónov do bunky sú potrebné veľké koncentračné a potenciálne gradienty. Na/K pumpa poskytuje ideálnu kompenzáciu pasívnych difúznych prúdov, pretože transportuje sodíkové ióny von z bunky a draselné ióny do nej. Čerpadlo je teda elektrogénne v dôsledku rozdielu v počte nábojov prenášaných do a z článku, ktorý pri normálnej rýchlosti svojej činnosti vytvára membránový potenciál približne 10 mV je elektronegatívny, ako keby bol produkovaný samotnými pasívnymi tokmi iónov (pozri rovnicu 7). V dôsledku toho sa membránový potenciál približuje rovnovážnemu potenciálu draslíka, čo znižuje únik iónov draslíka. Na činnosť/K pumpa je regulovaná intracelulárnou koncentráciou sodíkových iónov. Rýchlosť pumpy sa spomaľuje, keď sa koncentrácia sodíkových iónov, ktoré sa majú z bunky odstraňovať, znižuje (obr. 1.8), takže prevádzka pumpy a tok sodíkových iónov do bunky sa navzájom vyrovnávajú, pričom sa udržiava vnútrobunková koncentrácia sodíka. iónov na úrovni približne 10 mmol/l.
Na udržanie rovnováhy medzi čerpacími a pasívnymi membránovými prúdmi je potrebných oveľa viac molekúl pumpy Na/K ako kanálových proteínov pre ióny draslíka a sodíka. Keď je kanál otvorený, prejdú ním desaťtisíce iónov za niekoľko milisekúnd (pozri vyššie) a keďže sa kanál zvyčajne otvára niekoľkokrát za sekundu, celkovo ním počas tejto doby prejde viac ako 105 iónov. Jeden proteín pumpy presunie niekoľko stoviek iónov sodíka za sekundu, takže plazmatická membrána musí obsahovať asi 1000-krát viac molekúl pumpy ako molekúl kanálov. Merania kanálových prúdov v pokoji ukázali prítomnosť priemerne jedného draslíkového a jedného sodíkového otvoreného kanála na 1 μm 2 membrány; Z toho vyplýva, že v rovnakom priestore by malo byť prítomných asi 1000 molekúl Na/K pumpy, t.j. vzdialenosť medzi nimi je v priemere 34 nm; Priemer proteínu pumpy ako kanálového proteínu je 8–10 nm. Membrána je teda pomerne husto nasýtená čerpacími molekulami
Skutočnosť, že tok sodných iónov do bunky a iónov draslíka von z bunky je kompenzovaný prevádzkou pumpy, má ďalší dôsledok, ktorý spočíva v udržiavaní stabilného osmotického tlaku a konštantný objem. Vo vnútri bunky je vysoká koncentrácia veľkých aniónov, hlavne proteínov (A - v tabuľke 1.1), ktoré nie sú schopné preniknúť cez membránu (alebo cez ňu prenikajú veľmi pomaly), a preto sú pevnou zložkou vo vnútri bunky. Na vyrovnanie náboja týchto aniónov je potrebný rovnaký počet katiónov. Pôsobením Na/K pumpy sú tieto katióny hlavne draselné ióny. K výraznému zvýšeniu intracelulárnej koncentrácie iónov mohlo dôjsť až pri zvýšení koncentrácie aniónov v dôsledku toku C1 - pozdĺž koncentračného gradientu do bunky (tabuľka 1.1), ale membránový potenciál tomu bráni. Prichádzajúci prúd Cl – pozorované len dovtedy, kým sa nedosiahne rovnovážny potenciál pre ióny chlóru; toto sa pozoruje, keď je gradient iónov chlóru takmer opačný ako gradient iónov draslíka, pretože ióny chlóru sú negatívne nabité (Rovnica 4). Tak sa vytvorí nízka intracelulárna koncentrácia iónov chlóru, čo zodpovedá nízkej extracelulárnej koncentrácii draselných iónov. Výsledkom je obmedzenie celkového počtu iónov v bunke. Ak membránový potenciál klesne pri zablokovaní Na/K pumpy, napríklad počas anoxie, potom sa zníži rovnovážny potenciál pre ióny chlóru a zodpovedajúcim spôsobom sa zvýši intracelulárna koncentrácia chloridových iónov. Po obnovení rovnováhy nábojov vstupujú do bunky aj draselné ióny; celková koncentrácia iónov v bunke sa zvyšuje, čo zvyšuje osmotický tlak; to tlačí vodu do bunky. Bunka napučí. Tento opuch je pozorovaný in vivo v podmienkach nedostatku energie.
Koncentračný gradient Na + ako hnacia sila membránového transportu . Význam Na/K pumpy pre bunku nie je obmedzený na stabilizáciu normálnych gradientov K+ a Na+ na membráne. Energia uložená v membránovom gradiente Na+ , sa často používa na uľahčenie membránového transportu iných látok. Napríklad na obr. 1.10 zobrazuje „simport“ Na+ a molekuly cukru do bunky. Membránový transportný proteín transportuje molekulu cukru do bunky aj proti koncentračnému gradientu, pričom súčasne sa Na+ pohybuje pozdĺž koncentračného a potenciálneho gradientu, čím poskytuje energiu pre transport cukrov. Tento transport cukrov je úplne závislý od existencie vysokého gradientu Na+ ; ak intracelulárna koncentrácia Na+ sa výrazne zvýši, transport cukrov sa zastaví. Pre rôzne c akharov, existujú rôzne systémy importu. Transport aminokyselín do bunky podobnej transportu c akharov znázornený na obr. 1,10; zabezpečuje to aj spád Na+ , Existuje najmenej päť rôznych symportných systémov, z ktorých každý sa špecializuje na jednu skupinu príbuzných aminokyselín.
Okrem systémov simport existujú aj „antiportérov“. Jeden z nich napríklad prenesie jeden vápenatý ión von z bunky v jednom cykle výmenou za tri prichádzajúce sodíkové ióny (obr. 1.10). Energia pre Doprava cca 2+ sa tvorí v dôsledku vstupu troch sodíkových iónov pozdĺž koncentračného a potenciálneho gradientu. Táto energia je dostatočná (pri pokojovom potenciáli) na udržanie vysokého gradientu vápnikových iónov (od menej ako 10–7 mol/l vo vnútri bunky po približne 2 mmol/l mimo bunky).
Endo- a exocytóza . Pre určité látky, ktoré vstupujú do bunky alebo musia byť vylúčené

Ryža. 1.10.Proteíny uložené v lipidovej dvojvrstve membrány sprostredkúvajú symport glukózy a Na+ do bunky, ako aj Ca2+/Na+ –antiport, pri ktorom je hnacou silou gradient Na + na bunkovej membráne
z nej nie sú žiadne transportné kanály; Medzi tieto látky patria napríklad bielkoviny a cholesterol. Môžu prechádzať cez plazmatickú membránu do vezikuly, alebo bubliny, prostredníctvom endo- a exocytózy. Na obr. Obrázok 1.11 ukazuje hlavné mechanizmy týchto procesov. Pri exocytóze tvoria určité organely (pozri nižšie) vezikuly naplnené látkou, ktorú je potrebné z bunky odstrániť, ako sú hormóny alebo extracelulárne enzýmy. Keď sa takéto vezikuly dostanú k plazmatickej membráne, ich lipidová membrána sa s ňou spojí a umožní tak obsahu uniknúť do vonkajšieho prostredia. V opačnom procese, endocytóze, sa plazmatická membrána invaginuje, vytvorí jamku, ktorá sa potom prehĺbi a uzavrie, čím sa vytvorí intracelulárna vezikula naplnená extracelulárnou tekutinou a niektorými makromolekulami. Na zabezpečenie tejto membránovej fúzie a uzavretia vezikuly pôsobia kontraktilné elementy cytoskeletu v spojení so samotnými membránami (pozri nižšie). Endocytóza nie vždy jednoducho zahŕňa zachytenie extracelulárneho prostredia do bunky. Bunková membrána často obsahuje špecifické receptory pre makromolekuly, ako je inzulín alebo antigény, často organizované do špecializovaných skupín. Potom, čo sa tieto makromolekuly naviažu na svoje receptory, nastane endocytóza v oblasti membrány obklopujúcej receptor a makromolekula je selektívne transportovaná do bunky (obr. 1.12, B).
Endo- a exocytóza prebieha v bunkách nepretržite. Množstvo obratu membránového materiálu je významné; do 1 hodiny absorbuje makrofág dvojnásobok povrchu svojej cytoplazmatickej membrány vo forme vezikúl. Vo väčšine buniek nedochádza k obratu membránového materiálu tak intenzívne, ale aj tak by mal byť významný.

Ryža. 1.11.Exocytóza a endocytóza. Hore: intracelulárna vezikula sa spája s lipidovou dvojvrstvou plazmatickej membrány a otvára sa do extracelulárneho priestoru. Tento proces sa nazýva exocytóza. Na spodku: plazmatická membrána invaginuje na malej ploche a uvoľňuje vezikula naplnená extracelulárnym materiálom. Tento proces sa nazýva endocytóza
Endo- a exocytóza nie sú len procesy transportu látok cez bunkovú membránu, ale aj procesy výmeny membrán - štruktúrnych zložiek samotnej bunky. Predmetom úvahy v tejto časti sú ďalšie podobné transportné procesy v bunke a jej organelách.
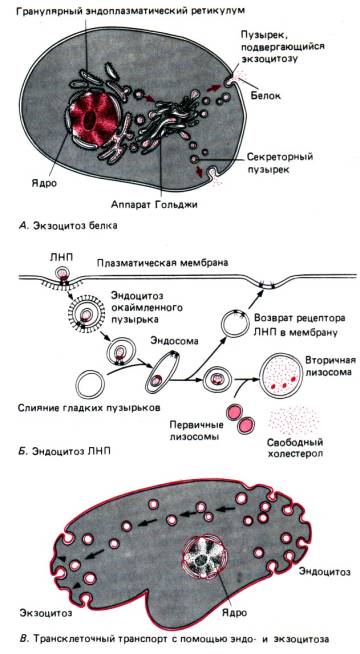
Ryža. 1.12. A–B. Schéma procesov vrátane exo- a endocytózy. A. Proteín syntetizovaný v granulárnom endoplazmatickom retikule je transportovaný cez Golgiho aparát do plazmatickej membrány, kde je vylučovaný exocytózou. B. Cholesterol naviazaný na častice LDL (low-density lipoprotein) sa prichytí na plazmatickú membránu, indukuje tvorbu endocytického vezikula v tejto oblasti membrány a je transportovaný do lyzozómov, kde sa uvoľňuje. IN. Extracelulárny materiál zachytený počas endocytózy (na obrázku napravo), transportované cez bunku vo vezikulách alebo vezikulách a uvoľňované prostredníctvom exocytózy (na obrázku vľavo)
Difúzia . Prirodzene, v cytosóle je rozdiel v koncentráciách eliminovaný difúziou; to isté platí pre tekutiny obsiahnuté v organelách. Vďaka vysokej koncentrácii rozpustených bielkovín je tu difúzia oveľa pomalšia ako vo vode. Lipidové membrány – okolo buniek a vnútri organel – sú dvojrozmerné tekutiny, v ktorých dochádza k difúzii. Lipidy v membránovej dvojvrstve difundujú vo svojej vlastnej vrstve a zriedka sa pohybujú z jednej do druhej. Proteíny v nich ponorené sú tiež celkom mobilné; rotujú okolo osi kolmej na membránu alebo difundujú laterálne s veľmi odlišnými difúznymi konštantami, 2–10 000-krát pomalšie ako fosfolipidy. Ak sa teda niektoré proteíny pohybujú voľne v lipidovej vrstve a rovnakou rýchlosťou ako samotné molekuly lipidov, potom sú iné ukotvené, t.j. celkom pevne spojené s cytoskeletom. V membráne sú „trvalé“ agregáty špecifických proteínov, napríklad pre- a postsynaptické štruktúry nervových buniek. Voľne sa pohybujúce proteíny možno demonštrovať ich naviazaním na fluorescenčné farbivá, ktorých luminiscencia je vyvolaná krátkym osvetlením malej plochy membrány. Takéto experimenty ukazujú, že za menej ako 1 minútu sú proteíny naviazané na farbivo rovnomerne distribuované cez membránu na vzdialenosti až 10 μm.
Aktívny transport v membránach organel .
Aktívne transportné procesy, ktoré zohrávajú dôležitú úlohu vo fungovaní plazmatickej membrány, sa vyskytujú aj vo vnútri bunky, v membránach organel. Špecifické obsahy rôznych organel sa vytvárajú čiastočne internou syntézou a čiastočne aktívnym transportom z cytosólu. Jedným príkladom posledne menovaného je vyššie uvedená Ca2+ pumpa v sarkoplazmatickom retikule svalových buniek. Zaujímavé je najmä to, že v prípade syntézy ATP v mitochondriách platí opačný princíp, čo sa deje v ATPázových pumpách plazmatickej membrány (obr. 1.6). Počas syntézy ATP vedie oxidačný metabolizmus k vytvoreniu strmého gradientu H+ na vnútorných membránach. Tento gradient je hnacou silou pre opačný proces čerpacieho cyklu aktívneho transportu molekúl: H+ ióny sa pohybujú cez membránu pozdĺž gradientu a energia uvoľnená ako výsledok poskytuje syntézu ATP z ADP a fosfátu. Výsledný ATP zase poskytuje energiu bunke vrátane aktívneho transportu.
Transport vo vezikulách . Bunka má veľké množstvo organel a pridružených vezikúl (obr. 1.1). Tieto organely a najmä vezikuly sú v neustálom pohybe a transportujú svoj obsah do iných organel alebo do plazmatickej membrány. Vezikuly môžu tiež migrovať z bunkovej membrány do organel, ako pri endocytóze.
Proces sekrécia bielkovín znázornené na obr. 1,12, A. Proteín sa syntetizuje v blízkosti bunkového jadra na ribozómoch spojených s endoplazmatickým retikulom (takzvané granulárne alebo drsné endoplazmatické retikulum); Keď je proteín v endoplazmatickom retikule, je zabalený do transportných vezikúl, ktoré sa oddelia od organely a migrujú do Golgiho aparátu. Tu sa spájajú s cisternami Golgiho aparátu, kde dochádza k modifikácii proteínu (t.j. premene na glykoproteín). Na koncoch cisterien sa vezikuly opäť oddelia. Sekrečné vezikuly nesúce modifikovaný proteín sa presúvajú k plazmatickej membráne a uvoľňujú svoj obsah exocytózou.
Ďalší príklad transportnej dráhy v bunke je znázornený na obr. 1,12, B; Ide o absorpciu cholesterolu bunkou. Cholesterol transportovaný v krvi sa viaže hlavne na bielkoviny, ako sú častice "lipoproteín s nízkou hustotou"(LDL). Takéto častice sa viažu na špecifické membránové oblasti obsahujúce LDL receptory, kde dochádza k endocytóze a LDL je transportovaný do bunky v „ohraničených“ vezikulách. Tieto vezikuly sa spoja a vytvoria endozómy a počas tohto procesu strácajú svoje „lemovanie“. Endozómy sa zase spájajú s primárnymi lyzozómami, obsahujúcimi prevažne hydrolytické enzýmy, a vytvárajú sekundárne väčšie lyzozómy. V nich sa cholesterol uvoľňuje z častíc LDL a difunduje do cytosólu, kde sa stáva dostupným napríklad pre syntézu lipidových membrán. Vezikuly, ktoré neobsahujú LDL, sú tiež oddelené od endozómov, ktoré sa špeciálnym spôsobom pohybujú k plazmatickej membráne a spájajú sa s ňou, čím vracajú materiál membrány a pravdepodobne aj receptory pre LDL. Od okamihu, keď sa častica LDL naviaže na membránu, kým sa cholesterol uvoľní zo sekundárneho lyzozómu, uplynie 10-15 minút. Poruchy viazania a vstrebávania LDL, teda zásobovania bunky cholesterolom, zohrávajú rozhodujúcu úlohu pri vzniku vážneho a rozšíreného ochorenia, aterosklerózy („kôrnatenie“ tepien).
Existuje mnoho ďalších dopravných ciest podobných tým, ktoré sú znázornené na obr. 1.11 a 1.12, A, pomocou ktorých sa v bunke pohybujú špecifické vezikuly. Nie je presne známe, ako sa pohybujú, ale na tomto procese sa pravdepodobne podieľajú cytoskeletálne prvky. Vezikuly sa môžu posúvať pozdĺž mikrotubulov, pričom energiu na pohyb zjavne poskytuje proteín spojený s vezikulami, ATPáza (pozri nižšie). Zostáva úplne nejasné, koľko rôznych vezikúl, pohybujúcich sa jeden po druhom vo všetkých smeroch, dosiahne svoj cieľ. Je zrejmé, že ich treba „označiť“ spôsobom, ktorý dopravný systém rozpozná a prevedie na cielený pohyb.
Transport tvorbou a deštrukciou organel . Endo- a exocytózu sme doteraz považovali za procesy transportu obsahu vezikúl. Ďalší aspekt týchto procesov spočíva v tom, že cielené odstraňovanie plazmatickej membrány na jednej časti povrchu bunky endocytózou a naopak jej pridávanie na inú exocytózou posúva veľké plochy membrány (obr. 1.12.E), čo dáva bunke príležitosť napríklad vytvoriť výrastok alebo presunúť sa.
Podobné prestavby sú typické aj pre cytoskelet, najmä pre mikrofilamenty a mikrotubuly (obr. 1.1). Mikrovlákna pozostávať predovšetkým z F-aktínový proteín, ktorý je schopný zostaviť sa do vláknitých zväzkov v dôsledku polymerizácie monoméru z cytosólu. Zväzky sú polarizované, to znamená, že často rastú len na jednom konci, pričom akumulujú nové molekuly aktínu, zatiaľ čo druhý koniec je inertný alebo tu dochádza k rozkladu. Vďaka tomuto polarizovanému rastu sa mikrofilamenty efektívne pohybujú a štruktúra ich siete sa môže meniť. Prechod aktínu z depolymerizovaného stavu (sol) do organizovaného stavu (gél) môže nastať veľmi rýchlo vplyvom iných proteínov alebo zmien koncentrácie iónov (pozri nižšie). Existujú aj proteíny, ktoré spôsobujú rozpad aktínových filamentov za vzniku krátkych fragmentov. Tenké výbežky mnohých buniek – filopódie – obsahujú centrálny zväzok aktínu (obr. 1.1) a rôzne pohyby filopódií sú pravdepodobne dôsledkom aktínových prechodov: polymerizácia – depolymerizácia.
Mikrotubulytiež často podstupujú podobné pohyby. Mechanizmus týchto pohybov je podobný - polymerizácia tubulínu z cytosolu tak, že jeden koniec mikrotubulu rastie, zatiaľ čo druhý sa buď nemení, alebo tam dochádza k demontáži. Mikrotubuly sa teda vhodným pridaním alebo odstránením materiálu môžu pohybovať v cytosóle.
Aktívne pohyby cytoskeletu . Zmeny v cytoskeletálnych štruktúrach môžu nastať v dôsledku aktívnych pohybov a preskupení opísaných vyššie. V mnohých prípadoch je pohyb mikrotubulov a aktínových filamentov poháňaný kontraktilnými proteínmi, ktoré viažu filamenty alebo tubuly a môžu ich vzájomne pohybovať. Veveričky myozín a dyneín prítomný v cytosóle všetkých buniek v relatívne vysokých koncentráciách; sú to prvky, ktoré premieňajú energiu na pohyb v špecializovaných bunkách (svaloch) a organelách (cilia). Vo svalových bunkách tvorí myozín hrubé vlákna orientované rovnobežne s aktínovými vláknami. Molekula myozínu sa svojou „hlavou“ pripojí k aktínovému vláknu a pomocou energie ATP vytesňuje myozín pozdĺž molekuly aktínu. Myozín sa potom oddelí od aktínu. Kombinácia mnohých takýchto cyklov spojenie-odpojenie vedie k makroskopickým kontrakcia svalových vlákien(kapitola 4). Dyneín hrá podobnú úlohu pri pohybe mikrotubulov počas mihalníc (obr. 1.1). V cytoplazme nešpecializovaných buniek myozín a dyneín netvoria pravidelné vlákna, ale vo väčšine prípadov malé skupiny molekúl. Dokonca aj v takýchto malých agregátoch sú schopné pohybovať aktínovými vláknami alebo mikrotubulami. Ryža. Obrázok 1.13 znázorňuje tento proces, keď sú opačne polarizované molekuly myozínu tiež pripojené k dvom aktínovým vláknam, polarizovaným v rôznych smeroch. Myozínové hlavové skupiny sa ohýbajú smerom k chvostu molekuly, pričom pri tom spotrebúvajú ATP a dve aktínové vlákna sa pohybujú opačným smerom, po čom sa myozín od nich oddelí. Pohyby tohto druhu, pri ktorých sa energia ATP premieňa na mechanickú prácu, môžu zmeniť tvar cytoskeletu a následne bunky a tiež zabezpečiť transport organel spojených s cytoskeletom.
Procesy intracelulárneho transportu možno najzreteľnejšie demonštrovať na axóne nervovej bunky. Transport Axónov je tu podrobne diskutovaný na ilustráciu udalostí, ktoré sa pravdepodobne vyskytnú podobným spôsobom vo väčšine buniek. Axón, ktorý má priemer len niekoľko mikrónov, môže dosiahnuť dĺžku jeden meter alebo viac a pohyb proteínov difúziou z jadra k distálnemu koncu axónu by trval roky. Už dlho je známe, že keď ktorákoľvek časť axónu podstúpi konstrikciu, časť axónu umiestnená proximálne sa roztiahne. Vyzerá to tak, že odstredivé prúdenie je zablokované v axóne. Takéto prietokovo rýchly transport axónov demonštrovať pohybom rádioaktívnych markerov, ako v experimente znázornenom na obr. 1.14. Rádioaktívne značený leucín bol injikovaný do dorzálneho koreňového ganglia a potom bola meraná rádioaktivita v sedacom nerve vo vzdialenosti 166 mm od tiel neurónových buniek od 2. do 10. hodiny. Počas 10 hodín sa vrchol rádioaktivity v mieste vpichu mierne zmenil. Ale vlna rádioaktivity sa šírila pozdĺž axónu konštantnou rýchlosťou asi 34 mm za 2 hodiny alebo 410 mm/deň. Ukázalo sa, že vo všetkých neurónoch homeotermických zvierat prebieha rýchly axonálny transport rovnakou rýchlosťou a nie sú pozorované žiadne výrazné rozdiely medzi tenkými, nemyelinizovanými vláknami a najhrubšími axónmi, ako aj medzi motorickými a senzorickými vláknami. Typ rádioaktívneho markera tiež neovplyvňuje rýchlosť rýchleho axonálneho transportu; markery môžu slúžiť ako rôzne rádioaktívne

Ryža. 1.13.Nesvalový myozínový komplex sa s určitou orientáciou môže viazať na aktínové vlákna rôznej polarity a pomocou energie ATP ich vzájomne premiestňovať
molekuly, ako sú rôzne aminokyseliny, ktoré sú zahrnuté v proteínoch bunkového tela neurónu. Ak analyzujeme periférnu časť nervu, aby sme určili povahu nosičov rádioaktivity, ktoré sa sem transportujú, potom sa takéto nosiče nachádzajú najmä v proteínovej frakcii, ale aj v zložení mediátorov a voľných aminokyselín. S vedomím, že vlastnosti týchto látok sú rôzne a veľkosti ich molekúl sú obzvlášť odlišné, môžeme konštantnú rýchlosť transportu vysvetliť len transportným mechanizmom spoločným pre všetky z nich.
Popísané vyššie rýchly transport axónov je anterográdna, t.j. smerujúce preč od tela bunky. Ukázalo sa, že niektoré látky prechádzajú z periférie do bunkového tela pomocou retrográdna doprava. Napríklad acetylcholínesteráza je transportovaná týmto smerom rýchlosťou 2-krát nižšou ako rýchlosť rýchleho axonálneho transportu. V neuroanatómii často používaný marker, chrenová peroxidáza, sa tiež pohybuje retrográdnym transportom. Retrográdny transport pravdepodobne hrá dôležitú úlohu v regulácii syntézy proteínov v bunkovom tele. Niekoľko dní po prerezaní axónu sa v tele bunky pozoruje chromatolýza, čo naznačuje narušenie syntézy proteínov. Čas potrebný na chromatolýzu koreluje s trvaním retrográdneho transportu z miesta transekcie axónu do tela bunky. Tento výsledok tiež naznačuje vysvetlenie tejto poruchy – je narušený prenos „signálnej látky“, ktorá reguluje syntézu bielkovín, z periférie. Je zrejmé, že hlavné "vozidlá" slúžia na rýchle axonálne
Ryža. 1.14.Experiment demonštrujúci rýchly axonálny transport v senzorických vláknach mačacieho ischiatického nervu. Tríciom značený leucín sa vstrekuje do dorzálneho koreňového ganglia a rádioaktivita v gangliových a senzorických vláknach sa meria 2, 4, 6, 8 a 10 hodín po injekcii. (spodná časť obrázku). Autor: os x Vynesie sa vzdialenosť od ganglia k oblasti sedacieho nervu, kde sa meranie vykonáva. Na zvislej osi iba pre hornú a dolnú krivku je rádioaktivita (pulzy/min) vynesená na logaritmickej stupnici. "Vlna" zvýšenej rádioaktivity (šípky) pohybuje sa rýchlosťou 410 mm/deň (pozdĺž)
doprava sú vezikuly (vezikuly) a organely, ako sú mitochondrie, ktoré obsahujú látky, ktoré je potrebné transportovať. Pohyb najväčších vezikúl alebo mitochondrií možno pozorovať pomocou mikroskopu in vivo . Takéto častice robia krátke, rýchle pohyby jedným smerom, zastavia sa, často sa pohybujú mierne dozadu alebo do strany, znova sa zastavia a potom trhnú v hlavnom smere. 410 mm/deň zodpovedá priemernej rýchlosti anterográdneho pohybu približne 5 μm/s; rýchlosť každého jednotlivého pohybu musí byť preto oveľa vyššia, a ak vezmeme do úvahy veľkosti organel, filamentov a mikrotubulov, potom sú tieto pohyby skutočne veľmi rýchle. Rýchly axonálny transport vyžaduje značnú koncentráciu ATP. Rýchly axonálny transport blokujú aj jedy ako kolchicín, ktorý narúša mikrotubuly. Z toho vyplýva, že v procese transportu, ktorý uvažujeme, sa vezikuly a organely pohybujú pozdĺž mikrotubulov a aktínových filamentov; tento pohyb zabezpečujú malé agregáty molekúl dyneínu a myozínu, pôsobiace tak, ako je znázornené na obr. 1,13 pomocou energie ATP.
Môže sa na ňom podieľať aj rýchly transport axónov patologické procesy. Niektoré neurotropné vírusy (napríklad herpes alebo poliovírusy) vstupujú do axónu na periférii a pohybujú sa retrográdnym transportom do tela neurónu, kde sa množia a prejavujú svoje toxické účinky. Tetanový toxín, proteín produkovaný baktériami, ktoré sa dostávajú do tela pri poškodení kože, je zachytený nervovými zakončeniami a transportovaný do tela neurónov, kde spôsobuje charakteristické svalové kŕče. Sú známe prípady toxických účinkov na samotný transport axónov, napríklad vystavenie priemyselnému rozpúšťadlu akrylamidu. Okrem toho sa predpokladá, že patogenéza nedostatku vitamínov beriberi a alkoholickej polyneuropatie zahŕňa narušenie rýchleho axonálneho transportu.
Okrem rýchleho axonálneho transportu v bunke je aj dosť intenzívny pomalý transport axónov. Tubulín sa pohybuje pozdĺž axónu rýchlosťou asi 1 mm/deň a aktín sa pohybuje rýchlejšie – až 5 mm/deň. Iné proteíny migrujú s týmito cytoskeletálnymi komponentmi; napríklad sa zdá, že enzýmy sú spojené s aktínom alebo tubulínom. Rýchlosti pohybu tubulínu a aktínu sú zhruba konzistentné s rýchlosťami rastu zistenými pre mechanizmus opísaný vyššie, kde sú molekuly začlenené do aktívneho konca mikrotubulu alebo mikrofilamentu. Preto tento mechanizmus môže byť základom pomalého axonálneho transportu. Rýchlosť pomalého axonálneho transportu tiež približne zodpovedá rýchlosti rastu axónov, čo zjavne naznačuje obmedzenia uložené štruktúrou cytoskeletu na druhý proces.
Na záver tejto časti treba zdôrazniť, že bunky v žiadnom prípade nie sú statické štruktúry, ako sa to napríklad ukazuje na fotografiách z elektrónového mikroskopu. Plazmatická membrána a hlavne organely sú v neustálom rýchlom pohybe a neustálej reštrukturalizácii; preto sú schopné fungovať. Ďalej to nie sú jednoduché komory, v ktorých prebiehajú chemické reakcie, ale vysoko organizované konglomeráty membrán a vlákien, v ktorých reakcie prebiehajú v optimálne organizovanom slede.
1.4. Regulácia bunkových funkcií
Udržiavanie individuálnej bunky ako funkčnej jednotky je primárne regulované jadrom; štúdium takýchto regulačných mechanizmov je predmetom bunkovej biológie a biochémie. Zároveň bunky musia upravovať svoje funkcie v súlade s podmienkami prostredia a potrebami iných buniek tela, t. j. slúžia ako objekty funkčnej regulácie. Nižšie stručne zvážime, ako tieto regulačné vplyvy pôsobia na plazmatickú membránu a ako sa dostávajú do intracelulárnych organel.
Regulačné účinky na bunkovú membránu
Membránový potenciál . V mnohých prípadoch sa regulácia bunkových funkcií uskutočňuje zmenou membránového potenciálu. Lokálne zmeny potenciálu sú možné, keď: 1) cez membránu preteká prúd zo susednej oblasti bunky alebo generovaný inou bunkou; 2) mení sa extracelulárna koncentrácia iónov (často [K + ] von ); 3) membránové iónové kanály sa otvoria. Zmeny membránového potenciálu môžu ovplyvniť konformáciu membránových proteínov, čo spôsobí najmä otvorenie alebo zatvorenie kanálov. Ako je uvedené vyššie, fungovanie niektorých membránových čerpadiel závisí od membránového potenciálu. Nervové bunky sú špecializované na vnímanie zmien membránového potenciálu ako informácie, ktorú treba spracovať a preniesť (pozri kapitolu 2).
Extracelulárne regulačné látky . Najdôležitejším regulačným mechanizmom zahŕňajúcim extracelulárne látky je ich interakcia so špecifickými receptormi na plazmatickej membráne alebo vo vnútri bunky. Medzi tieto látky patria synaptické prenášače, ktoré prenášajú informácie medzi nervovými bunkami, miestnymi činiteľmi a látkami, ktoré cirkulujú v krvi a dostávajú sa do všetkých buniek tela, ako sú hormóny a antigény. Synaptické vysielače sú malé molekuly uvoľnené z nervových zakončení na synapsii;
keď dosiahnu plazmatickú membránu susednej, postsynaptickej bunky, spúšťajú elektrické signály alebo iné regulačné mechanizmy. Táto problematika je podrobne rozobratá v kap. 3.
Miestne chemické látky často vylučované špecializovanými bunkami. Voľne difundujú v extracelulárnom priestore, ale ich pôsobenie je obmedzené na malú skupinu buniek v dôsledku rýchlej deštrukcie týchto látok, či už spontánne alebo pôsobením enzýmov. Jedným príkladom uvoľňovania takýchto činidiel je uvoľňovanie histamínžírne bunky v dôsledku poškodenia alebo imunitnej odpovede. Histamín spôsobuje relaxáciu buniek hladkého svalstva ciev, zvyšuje permeabilitu cievneho endotelu a stimuluje senzorické nervové zakončenia, ktoré sprostredkúvajú pocit svrbenia. Ďalšie lokálne chemické látky sú uvoľňované mnohými inými bunkami. Typickými lokálnymi agentmi sú prostaglandíny, tvoria skupinu približne 20 derivátov mastných kyselín. Sú uvoľňované nepretržite zo široko distribuovaných buniek, ale pôsobia len lokálne, pretože sú rýchlo zničené membránovými fosfolipázami. Rôzne prostaglandíny majú široké spektrum účinku: môžu vyvolať kontrakciu buniek hladkého svalstva, spôsobiť agregáciu krvných doštičiek (trombocytov) alebo potlačiť vývoj žltého telieska vo vaječníkoch.
Obsluhujú ďalší miestni agenti rastové faktory. Najznámejší je nervový rastový faktor (NGF) pre sympatické neuróny, ktorý je nevyhnutný pre rast a prežitie týchto neurónov počas vývoja in vivo alebo v bunkovej kultúre. Cieľové bunky pre túto triedu neurónov zjavne vylučujú NGF a tým zabezpečujú správnu inerváciu. Pri vytváraní orgánov si bunky často potrebujú „nájsť cestu“ k cieľovým bunkám, ktoré môžu byť umiestnené v značnej vzdialenosti. Preto musí existovať veľa špecializovaných rastových faktorov podobných NGF.
Hormóny a antigény prenášané krvou do všetkých buniek. Antigény spúšťajú imunitnú odpoveď buniek nesúcich špecifické protilátky. Antigény sú však spravidla cudzorodé látky, ktoré sa v reagujúcom organizme nevytvárajú (podrobnejšie v kapitole 18). Niektoré hormóny, ako je inzulín alebo tyroxín, ovplyvňujú širokú škálu typov buniek, zatiaľ čo iné, ako napríklad pohlavné hormóny, ovplyvňujú iba špecifický typ buniek. Hormóny sú buď peptidy, ktorých pôsobenie sa spúšťa väzbou na receptor na bunkovej membráne, alebo steroidy a tyroxín, ktoré difundujú cez lipidovú membránu a viažu sa na intracelulárne receptory. Steroidné hormóny sa viažu na jadrový chromatín, čo vedie k transkripcii určitých génov. Výsledné proteíny spôsobujú zmeny bunkových funkcií, čo je špecifický účinok hormónov. Otázky súvisiace s uvoľňovaním a pôsobením hormónov sú podrobne diskutované v kapitole. 17.
Intracelulárna komunikácia zahŕňajúca druhých poslov
Vyššie opísané regulačné funkcie zahŕňajú účinky na bunkovú membránu. Informácie prijaté bunkovou membránou musia často spôsobiť reakciu organel a sú do nich prenášané rôznymi látkami známymi ako druhí poslovia (na rozdiel od prvých poslov, ktoré prichádzajú do bunky z vonkajších zdrojov). Štúdium druhých mediátorov sa rýchlo rozvíja a neexistuje žiadna záruka, že súčasná úroveň pochopenia problému bude dostatočne úplná. Tu sa dotkneme troch dobre preštudovaných mediátorov: Ca2+, cAMP a inozitoltrifosfát.
Vápnik.Najjednoduchším intracelulárnym mediátorom je ión Ca2+. Jeho voľná koncentrácia v pokojovej bunke je veľmi nízka a dosahuje 10_–8–10–7 mol/l. Môže vstúpiť do bunky cez špecifické membránové kanály, keď sú v otvorenom stave, napríklad keď sa zmení membránový potenciál (pozri kapitolu 2). Výsledné zvýšenie koncentrácie Ca 2+ spúšťa dôležité reakcie v bunke, ako je kontrakcia myofibríl, ktorá je základom svalovej kontrakcie (pozri kapitolu 4), alebo uvoľnenie vezikúl obsahujúcich neurotransmitery z nervových zakončení (pozri kapitolu 3). . Obidve reakcie vyžadujú koncentráciu Ca2+ približne 10 –5 mol/l. Ca 2+, ktorý má regulačný účinok, sa môže uvoľňovať aj z vnútrobunkových zásob, ako je endoplazmatické retikulum. Uvoľňovanie Ca 2+ z depa si vyžaduje účasť ďalších sprostredkovateľov (pozri napr. obr. 1.16).
Cyklický adenozínmonofosfát, cAMP. Nedávno bolo dokázané, že cyklický adenozínmonofosfát (cAMP), derivát hlavného energetického zdroja tela, ATP, je dôležitým druhým poslom. Komplexný reťazec reakcií znázornený na obr. 1,15, začína receptorom R s na vonkajšom povrchu plazmatickej membrány, ktorá môže slúžiť ako miesto špecifickej väzby pre rôzne mediátory a hormóny. Po naviazaní na špecifickú „stimulačnú“ molekulu R s mení svoju konformáciu; tieto zmeny ovplyvňujú proteín G s na vnútornom povrchu membrány takým spôsobom, že je možná aktivácia membrány intracelulárnym guanozíntrifosfátom (GTP). Aktivovaný proteín G s , naopak stimuluje enzým na vnútornom povrchu membrány, adenylátcyklázu (AC), ktorá katalyzuje tvorbu cAMP z ATP. Vo vode rozpustný cAMP je mediátor, ktorý prenáša účinok

Ryža. 1.15.Reťazec reakcií zahŕňajúci intracelulárny mediátor cAMP (cyklický adenozínmonofosfát). Excitačné alebo inhibičné externé signály aktivujú membránové receptory R s alebo Ri . Tieto receptory regulujú proces väzby G – proteíny s intracelulárnym GTP (guanozíntrifosfát), čím stimulujú alebo inhibujú intracelulárnu adenylátcyklázu (AC). Amplifikačný enzým AC premieňa adenozíntrifosfát (ATP) na cAMP, ktorý sa potom fosfodiesterázou štiepi na AMP. Voľný cAMP difunduje do bunky a aktivuje adenylátkinázu (A-kinázu), pričom uvoľní jej katalytickú podjednotku C, ktorá katalyzuje fosforyláciu intracelulárnych proteínov, t.j. tvorí konečný efekt extracelulárneho stimulu. Diagram tiež ukazuje farmakologické lieky a toxíny, ktoré spúšťajú (+) alebo inhibujú (-) určité reakcie (modifikované)
stimulácia extracelulárnych receptorov R s k vnútorným štruktúram bunky.
Súbežne so stimulačným reťazcom reakcií zahŕňajúcich R s je možná väzba inhibičných mediátorov a hormónov na zodpovedajúci receptor RI ktorý opäť prostredníctvom GTP-aktivovaného proteínu G , inhibuje AC a tým aj produkciu cAMP. Difúziou do bunky cAMP reaguje s adenylátkinázou (A-kinázou); tým sa uvoľní podjednotka C, ktorá katalyzuje fosforyláciu proteínu P. Táto fosforylácia premieňa proteíny na aktívnu formu a tie teraz môžu uplatniť svoje špecifické regulačné účinky (napríklad spôsobiť degradáciu glykogénu). Tento komplexný regulačný systém je mimoriadne účinný, pretože konečným výsledkom je fosforylácia mnohých proteínov, t.j. regulačný signál prechádza obvodom s vysokým ziskom. Vonkajšie mediátory, ktoré sa viažu na receptory Rs a Rj špecifické pre každú z nich, mimoriadne rôznorodé. Adrenalín, spojenie s Rs alebo Ri podieľa sa na regulácii metabolizmu lipidov a glykogénu, ako aj na posilnení kontrakcie srdcového svalu a ďalších reakciách (pozri kapitolu 19). hormón stimulujúci štítnu žľazu, aktivácia R s stimuluje uvoľňovanie hormónu tyroxínu štítnou žľazou a prostaglandín I inhibuje agregáciu krvných doštičiek. Inhibičné účinky, vrátane adrenalínu, sprostredkované cez RI sa prejavujú v spomalení lipolýzy. teda Systém cAMP je multifunkčný vnútrobunkový regulačný systém, ktoré môžu byť presne kontrolované extracelulárnymi stimulačnými a inhibičnými signálnymi látkami.
Inozitol fosfát "IF" h " Intracelulárny systém druhého posla, inozitol fosfát, bol objavený len nedávno (obr. 1.16). V tomto prípade neexistuje žiadna inhibičná dráha, ale existuje podobnosť so systémom cAMP, v ktorom sa účinok stimulácie R receptora prenáša na GTP-aktivovaný G proteín na vnútornom povrchu membrány. V ďalšom štádiu sa obvyklý membránový lipid fosfatidylinozitol (PI), ktorý predtým prijal dve ďalšie fosfátové skupiny, premení na PI-difosfát (PIP 2), ktorý sa štiepi aktivovanou fosfodiesterázou (PDE) na inozitol trifosfát(IFz) a lipid diacylglycerol(DAG). Inozitoltrifosfát je vo vode rozpustný druhý posol, ktorý difunduje do cytosólu. Pôsobí predovšetkým uvoľňovaním Ca 2+ z endoplazmatického retikula. Ca2+ zase pôsobí ako posol, ako je opísané vyššie; napríklad aktivuje Ca2+-dependentnú fosfokinázu, ktorá fosforyluje enzýmy. Signál nesie aj lipidová podjednotka DAG (obr. 1.16), ktorá v lipidovej fáze plazmatickej membrány difunduje do C-kinázy umiestnenej na jej vnútornom povrchu, ktorá sa aktivuje za účasti fosfatidylserínu ako kofaktora. Potom C-kináza spustí fosforyláciu proteínov a prevedie ich na aktívnu formu.
Intracelulárny systém druhého posla IF3 môže byť tiež riadený rôznymi vonkajšími mediátormi a hormónmi, vrátane acetylcholínu, serotonínu, vazopresínu a hormónu stimulujúceho štítnu žľazu; podobne ako systém cAMP sa vyznačuje rôznymi intracelulárnymi účinkami. Je možné, že tento systém je tiež aktivovaný svetlom vo vizuálnom receptore oka a zohráva ústrednú úlohu vo fototransdukcii (pozri kapitolu 11). Prvýkrát v individuálnom vývoji organizmu je receptor IG systému aktivovaný spermiami, v dôsledku čoho sa IG podieľa na regulačných reakciách sprevádzajúcich oplodnenie vajíčka.
Systémy cAMP a IFz-DAG sú vysoko účinné biologické zosilňovače. Oni

Ryža. 1.16.Reťazec reakcií zahŕňajúci intracelulárny mediátor IPG (inozitoltrifosfát). Rovnako ako v systéme cAMP je extracelulárny signál sprostredkovaný proteínomG, ktorý v tomto prípade aktivuje fosfodiesterázu (PDE). Tento enzým rozkladá fosfatidylinozíndifosfát (PIP 2 ) v plazmatickej membráne pred IF h a diacylglycerol (DAG); AK h difunduje do cytoplazmy. Tu spôsobuje uvoľňovanie Ca 2+ z endoplazmatického retikula; zvýšenie koncentrácie Ca 2+ v cytoplazme ([Ca 2+] i ) aktivuje proteínkinázu, ktorá fosforyluje a tým aktivuje enzýmy. Ďalší produkt, DAG, zostáva v membráne a aktivuje proteínkinázu C (fosfatidylserínový kofaktor, PS). Proteínkináza C tiež fosforyluje enzýmy, ktoré sprostredkovávajú špecifické akcie spojené so stimuláciou vonkajších receptorov R . Vetvy reťazca reakcií zahŕňajúce IF h a DAG môžu byť aktivované nezávisle ionomycínom a forbolesterom (modifikované)
konvertujú reakciu medzi transmiterom a receptorom vonkajšej membrány na fosforyláciu rôznych intracelulárnych proteínov, ktoré potom môžu ovplyvniť rôzne bunkové funkcie. Jedným z významných aspektov problému je, že pokiaľ je dnes známe, existujú iba tieto dva úzko súvisiace regulačné systémy tohto typu, ktoré používajú početné vonkajšie mediátory na reguláciu rôznych vnútrobunkových procesov. Zároveň tieto regulačné systémy, vrátane Ca 2+, spolu úzko interagujú, čo im umožňuje vykonávať jemnú reguláciu bunkových funkcií.
Návody a návody
1. Alberts INBray D., Lewis J., Raff M., Roberts TOWatson J.D.Molecular Biology of the Cell, New York a Londýn, Garland Publishing Inc., 1983.
2. Czihak G., Longer H., Ziegler H.(eds.). Biológia. Berlín, Heidelberg, New York, Springer, 1983.
3. Hille IN. Iónové kanály excitabilných membrán. Sunderland, Massachusetts, Sinauer Assoc., 1984.
4. Hoppe W., Lohmann W.. Marki H., Ziegler H.(eds.). Biofyzika. Berlín, Heidelberg, New York, Springer, 1984.
5. Jungermann TO., Mahler H.Biochemia. Berlín, Heidelberg, New York, Springer, 1980.
6. Kandel E. R., Schwartz– J. H.,(eds.). Princípy neurálnej vedy, New York, Amsterdam, Oxford, Elsevier, 1985.
7. Schiebler T. H., Schmidt W.Anatomic des Menschen. Berlín, Heidelberg, New York, Tokio, Springer, 1983.
Pôvodné články a recenzie
8. Berridge M.J. Molekulárny základ komunikácie v bunke, Sci. Amer, 253, 124-134 (1985).
9. Berridge M. J., Irvine R. F. Inozitoltrifosfát, nový druhý posol v transdukcii bunkového signálu. Nature, 312, 315 321 (1984).
10. Bretscher M.S. Molekuly bunkovej membrány, Sci. Amer., 253, 124-134 (1985).
11. Daut J.Živá bunka ako stroj na prenos energie. Minimálny model metabolizmu myokardu, Biochem. et Biophys. Acta, 895, 41-62 (1987).
12. Hodgkin A.L., Katz IN. Vplyv sodíkových iónov na elektrickú aktivitu obrovského axónu chobotnice. J. Physiol. (Lond.), 108, 37 – 77 (1949).
13. Hodgkin A.L., Keynes R.D. Aktívny transport katiónov v obrovských axónoch z Sépia a Loligo, J. Physiol. (Lond.), 128, 28 – 42 (1955).
14. Dlhšie P. Iónové kanály s konformačnými podstavami, Biophys. J., 47, 581-590 (1985).
15. Ochs S., Worth P.M. Exoplazmatický transport v normálnych a patologických systémoch. In: Fyziológia a patológia axónov, S.G. Waxmann, Ed. New York, Raven Press, 1978.
súhrn ďalších prezentácií„Metódy vyučovania biológie“ - Školská zoológia. Oboznámenie študentov s využitím vedeckých zoologických údajov. Morálna výchova. Dodatočné požehnanie kurníku. Výber metód. Životné procesy. Akvarijné ryby. Výživa. Environmentálna výchova. Materialita životných procesov. Negatívne výsledky. Pozor študenti. Povinná forma. Pri pohľade na malé zvieratá. Ciele a ciele biológie. Príbeh.
„Problémové učenie sa na hodinách biológie“ - Vedomosti. Nové učebnice. Cesta k riešeniu. Problém. Semináre. čo je to úloha? Albrecht Dürer. Problémové učenie na hodinách biológie. Neštandardné lekcie. Čo znamená problémové učenie. Kvalita života. Biológia ako akademický predmet. Otázka. Lekcia riešenia problémov. Znížený záujem o predmet. Laboratórne triedy založené na problémoch.
„Kritické myslenie na hodinách biológie“ - Technológia „kritického myslenia“. Využitie technológie „rozvoja kritického myslenia“. Stôl na lekciu. Motivácia k štúdiu. Ekosystémy. Význam „rozvoja kritického myslenia“. Známky technológie. Technológia RKM. Štruktúra lekcie. Hlavné smery. História techniky. Pedagogické technológie. Pravidlá technológie. Úlohy z biológie. Fotosyntéza. Techniky používané v rôznych fázach lekcie.
„Lekcie biológie s interaktívnou tabuľou“ - Elektronické učebnice. Výhody pre študentov. Interaktívna tabuľa pomáha sprostredkovať informácie každému študentovi. Didaktické úlohy. Riešenie biologických problémov. Výhody práce s interaktívnymi tabuľami. Práca s prezentáciami. Pracujte na porovnávaní predmetov. Pohybujúce sa predmety. Používanie tabuliek. Používanie interaktívnej tabule v procese výučby školákov. Výhody pre učiteľov.
„Prístup systémovej aktivity v biológii“ - Otázky zo seminára. Metóda aktivity. Dryopithecus. Mimozemská cesta ľudského pôvodu. lyzozómy. Chemická organizácia. Gymnospermy. Metabolizmus. Analyzátory. Systémovo-činnostný prístup k vyučovaniu biológie. Chromozómy. Cytoplazma. Slepota. Dĺžka ucha. Klasifikácia osoby. Kostra cicavca. Cesty ľudskej evolúcie. Mitóza. Povrchový komplex. Problematická otázka. Nucleolus. Jadrový obal.
„Počítač v biológii“ - Spoločná aktivita študentov. Rodiny krytosemenných rastlín. Interaktívny tréning. Modely učenia. Príklad hodnotiaceho systému. Otázky inštrukčnej karty. Príklad karty s pokynmi. Výskumníci. Mikroskupiny. Interaktívne vzdelávacie technológie. Kolotoč. Interaktívne vzdelávacie technológie. Interaktívne prístupy na hodinách biológie. Skupinová forma práce. Úlohy pre skupiny „výskumníkov“.
