§ 2. யூகாரியோடிக் கலத்தின் முக்கிய கூறுகள்
யூகாரியோடிக் செல்கள் (படம் 8 மற்றும் 9) புரோகாரியோடிக் செல்களை விட மிகவும் சிக்கலானவை. அவை அளவு (பல மைக்ரோமீட்டர்கள் முதல் பல சென்டிமீட்டர்கள் வரை), வடிவத்தில் மற்றும் கட்டமைப்பு அம்சங்களில் (படம் 10) மிகவும் வேறுபட்டவை.
அரிசி. 8. யூகாரியோடிக் கலத்தின் அமைப்பு. பொதுவான திட்டம்
அரிசி. 9. எலக்ட்ரான் நுண்ணோக்கியின் படி செல் அமைப்பு
அரிசி. 10. பல்வேறு யூகாரியோடிக் செல்கள்: 1 - எபிடெலியல்; 2 - இரத்தம் (இ - எரித்ரோடிஸ், / - லுகிடிஸ்); 3 - குருத்தெலும்பு; 4 - எலும்புகள்; 5 - மென்மையான தசை; 6 - இணைப்பு திசு; 7 - நரம்பு செல்கள்; 8 - ஸ்ட்ரைட்டட் தசை நார்
இருப்பினும், அனைத்து யூகாரியோடிக் செல்களிலும் பொதுவான அமைப்பு மற்றும் அடிப்படை கூறுகளின் இருப்பு ஒரே மாதிரியாக இருக்கும் (படம் 11).
அரிசி. 11. யூகாரியோடிக் செல் (வரைபடம்)
பிளாஸ்மாலெம்மா (வெளிப்புற செல் சவ்வு).பிளாஸ்மா மென்படலத்தின் அடிப்படை, உயிரணுக்களில் உள்ள மற்ற சவ்வுகளைப் போலவே (உதாரணமாக, மைட்டோகாண்ட்ரியா, பிளாஸ்டிட்கள், முதலியன), இரண்டு வரிசை மூலக்கூறுகளைக் கொண்ட கொழுப்பு அடுக்கு ஆகும் (படம் 12). லிப்பிட் மூலக்கூறுகள் துருவமாக இருப்பதால் (ஒரு துருவம் ஹைட்ரோஃபிலிக், அதாவது, தண்ணீரால் ஈர்க்கப்படுகிறது, மற்றொன்று ஹைட்ரோபோபிக், அதாவது தண்ணீரால் விரட்டப்படுகிறது), அவை ஒரு குறிப்பிட்ட வரிசையில் அமைக்கப்பட்டிருக்கும். ஒரு அடுக்கின் மூலக்கூறுகளின் ஹைட்ரோஃபிலிக் முனைகள் அக்வஸ் சூழலை நோக்கி இயக்கப்படுகின்றன - செல்லின் சைட்டோபிளாசம், மற்றும் மற்ற அடுக்கு - கலத்திலிருந்து வெளிப்புறமாக - இடைச்செல்லுலார் பொருள் (பலசெல்லுலர் உயிரினங்களில்) அல்லது அக்வஸ் சூழலை நோக்கி (ஒரு செல்லுலார் உயிரினங்களில் )
அரிசி. 12. திரவ மொசைக் மாதிரியின் படி செல் சவ்வு அமைப்பு. புரதம் மற்றும் கிளைகோபுரோட்டீன்கள் லிப்பிட் மூலக்கூறுகளின் இரட்டை அடுக்கில் மூழ்கியுள்ளன, அவற்றின் ஹைட்ரோஃபிலிக் முனைகள் (வட்டங்கள்) வெளிப்புறமாக இருக்கும், மேலும் அவற்றின் ஹைட்ரோபோபிக் முனைகள் (அலை அலையான கோடுகள்) சவ்வுக்குள் ஆழமாக எதிர்கொள்ளும்.
புரத மூலக்கூறுகள் லிப்பிடுகளின் இரு மூலக்கூறு அடுக்கில் மொசையாக உட்பொதிக்கப்படுகின்றன. விலங்கு உயிரணுவின் வெளிப்புறத்தில், பாலிசாக்கரைடு மூலக்கூறுகள் லிப்பிட்கள் மற்றும் பிளாஸ்மாலெம்மாவின் புரத மூலக்கூறுகளுடன் இணைக்கப்பட்டு, கிளைகோலிப்பிட்கள் மற்றும் கிளைகோபுரோட்டீன்களை உருவாக்குகின்றன.
இந்த மொத்தமானது ஒரு அடுக்கை உருவாக்குகிறது கிளைகோகாலிக்ஸ்.அவருடன் தொடர்புடையவர் ஏற்பி செயல்பாடுபிளாஸ்மா சவ்வுகள் (கீழே காண்க); இது செல் பயன்படுத்தும் பல்வேறு பொருட்களையும் குவிக்கும். கூடுதலாக, கிளைகோகாலிக்ஸ் பிளாஸ்மாலெம்மாவின் இயந்திர நிலைத்தன்மையை அதிகரிக்கிறது.
தாவரங்கள் மற்றும் பூஞ்சைகளின் உயிரணுக்களில் ஒரு செல் சுவர் உள்ளது, அது ஒரு துணை மற்றும் பாதுகாப்பு பாத்திரத்தை வகிக்கிறது. தாவரங்களில் இது செல்லுலோஸைக் கொண்டுள்ளது, மேலும் பூஞ்சைகளில் இது சிட்டினால் ஆனது.
வெளிப்புற செல் சவ்வு பல செயல்பாடுகளை செய்கிறது, அவற்றுள்:
♦ இயந்திரவியல்(ஆதரவு, வடிவம்-கட்டிடம்);
♦ தடை-போக்குவரத்து(வெவ்வேறு பொருட்களுக்கு தேர்ந்தெடுக்கப்பட்ட ஊடுருவல்: தேவையான பொருட்களின் கலத்திற்குள் நுழைதல் மற்றும் தேவையற்ற மற்றும் தீங்கு விளைவிக்கும்வற்றை அகற்றுதல்);
♦ ஏற்பி(செல் அருகாமையில் காணப்படும் பல்வேறு இரசாயனங்கள் தீர்மானித்தல்; ஹார்மோன்கள் வடிவில் சிக்னல்களை உணர்தல்; நோயெதிர்ப்பு மண்டலத்தின் செல்கள் மூலம் "வெளிநாட்டு" புரதத்தை அங்கீகரித்தல், முதலியன).
செல் மற்றும் சுற்றுச்சூழலுக்கு இடையிலான பொருட்களின் பரிமாற்றம் வெவ்வேறு வழிகளில் மேற்கொள்ளப்படுகிறது - செயலற்ற மற்றும் செயலில்.
நீர் மற்றும் பல்வேறு அயனிகளின் மூலக்கூறுகள் செயலற்ற முறையில் (பரவல், சவ்வூடுபரவல் காரணமாக), செல் ஆற்றலைச் செலவழிக்காமல், சிறப்பு துளைகள் வழியாக நுழைகின்றன - இவை செயலற்ற போக்குவரத்து.புரதங்கள், பாலிசாக்கரைடுகள், முழு செல்கள் போன்ற மேக்ரோமிகுலூக்கள் வழங்கப்படுகின்றன பாகோசைடோசிஸ்மற்றும் பினோசைடோசிஸ்ஆற்றல் நுகர்வுடன் - செயலில் போக்குவரத்து.
பாகோசைட்டோசிஸ் மூலம், முழு செல்கள் அல்லது பெரிய துகள்கள் மூழ்கடிக்கப்படுகின்றன (உதாரணமாக, பாக்டீரியாவின் பாதுகாப்பு இரத்த அணுக்கள் மூலம் அமீபாஸ் அல்லது பாகோசைட்டோசிஸில் ஊட்டச்சத்து பற்றி சிந்தியுங்கள்). பினோசைட்டோசிஸின் போது, ஒரு திரவப் பொருளின் சிறிய துகள்கள் அல்லது நீர்த்துளிகள் உறிஞ்சப்படுகின்றன. இரண்டு செயல்முறைகளுக்கும் பொதுவானது என்னவென்றால், உறிஞ்சப்பட்ட பொருட்கள் ஒரு வெற்றிடத்தை உருவாக்குவதற்கு ஊடுருவும் வெளிப்புற சவ்வு மூலம் சூழப்பட்டுள்ளன, பின்னர் அது செல்லின் சைட்டோபிளாஸில் ஆழமாக நகரும்.
எக்சோசைடோசிஸ் என்பது பாகோசைட்டோசிஸ் மற்றும் பினோசைடோசிஸ் (படம் 13) ஆகியவற்றிற்கு எதிர் திசையில் ஒரு செயல்முறையாகும் (செயலில் உள்ள போக்குவரத்து ஆகும்). அதன் உதவியுடன், ஜீரணிக்கப்படாத உணவு புரோட்டோசோவாவில் உள்ளது அல்லது சுரக்கும் கலத்தில் உருவாகும் உயிரியல் ரீதியாக செயல்படும் பொருட்கள் அகற்றப்படலாம்.
சைட்டோபிளாசம்.சைட்டோபிளாசம் என்பது அணுக்கருவைத் தவிர்த்து, பிளாஸ்மாலெம்மாவால் வரையறுக்கப்பட்ட கலத்தின் உள்ளடக்கமாகும். இது கொண்டுள்ளது தரைப் பொருள் (ஹைலோபிளாசம்), உறுப்புகள்மற்றும் சேர்த்தல்கள்.
ஹைலோபிளாஸ்மா- ஒரு பிசுபிசுப்பான திரவம், இரண்டிலும் இருக்கும் திறன் கொண்டது ஜோலா(திரவ), அல்லது ஜெல்(ஜெல்லி போன்றது).
தேவைப்பட்டால், சைட்டோபிளாசம் ஒரு நிலையில் இருந்து மற்றொரு நிலைக்கு மாற்றும் திறன் கொண்டது. எடுத்துக்காட்டாக, அமீபாய்டு இயக்கத்தின் போது (விலங்கியல் பாடத்திலிருந்து "புரோட்டோசோவா" என்ற பிரிவை நினைவில் கொள்ளுங்கள்), ஒரு சூடோபாட் உருவாகும் போது, சைட்டோபிளாசம் ஜெல்லில் இருந்து சோலுக்கு விரைவாக மாறுகிறது மற்றும் நேர்மாறாகவும் நிகழ்கிறது. சைட்டோபிளாஸில் அதிக எண்ணிக்கையிலான இழை புரத மூலக்கூறுகள் இருப்பதால் இது ஏற்படுகிறது ஆக்டினா.ஒரு முப்பரிமாண வலையமைப்பை உருவாக்குவதற்கு அவை ஒன்றோடொன்று இணைக்கப்படும்போது, சைட்டோபிளாசம் ஒரு ஜெல் நிலையில் உள்ளது, மேலும் நெட்வொர்க் சிதைந்தால், அது ஒரு சோல் நிலையில் உள்ளது.
ஹைலோபிளாசம் பல்வேறு பொருட்களைக் கொண்டுள்ளது - நொதிகள், புரதங்கள், கார்போஹைட்ரேட்டுகள், கொழுப்புகள் மற்றும் பிற, கரிம மற்றும் தாது. பல்வேறு இரசாயன செயல்முறைகள் இங்கே நடைபெறுகின்றன - பொருட்களின் முறிவு, அவற்றின் தொகுப்பு மற்றும் மாற்றங்கள் (மாற்றங்கள்).
ஆர்கனாய்டுகள்.இவை ஒரு குறிப்பிட்ட அமைப்பு மற்றும் செயல்பாடுகளைக் கொண்ட ஒரு கலத்தின் நிரந்தர கூறுகள், அதன் சைட்டோபிளாஸில் அமைந்துள்ளன. பின்வருவனவற்றில் நாம் பேசுவோம் பொது நோக்கத்திற்கான ஆர்கனாய்டுகள்,அனைத்து யூகாரியோட்களின் அனைத்து வகையான செல்களிலும் உள்ளார்ந்தவை. அவை பிந்தையவற்றின் முக்கிய செயல்பாடுகளை உறுதி செய்வதோடு தொடர்புடையவை. சிறப்பு நோக்கத்திற்கான ஆர்கனாய்டுகள்ஒரு குறிப்பிட்ட (மிகவும் சிறப்பு வாய்ந்த) வகை உயிரணுக்களில் மட்டுமே காணப்படுகிறது - எடுத்துக்காட்டாக, தசை செல்களில் உள்ள myofibrils.
எந்த உயிரணுக்கள் மற்றும் எந்த உயிரினங்களைச் சேர்ந்தது என்பதைப் பொருட்படுத்தாமல் பொது நோக்கத்திற்கான உறுப்புகள் ஒரே அமைப்பைக் கொண்டுள்ளன. ஆனால் அவற்றில் சவ்வு கொண்ட குழுக்கள் உள்ளன (எண்டோபிளாஸ்மிக் ரெட்டிகுலம், கோல்கி கருவி, மைட்டோகாண்ட்ரியா, பிளாஸ்டிட்கள், லைசோசோம்கள், வெற்றிடங்கள்)அத்துடன் அல்லாத சவ்வு ( ரைபோசோம்கள், செல் மையம்)கட்டமைப்பு.
எண்டோபிளாஸ்மிக் ரெட்டிகுலம் (ER).இபிஎஸ் சவ்வுகளைக் கொண்டுள்ளது மற்றும் இது கலத்தின் முழு சைட்டோபிளாஸத்தையும் ஊடுருவிச் செல்லும் குழாய்கள் மற்றும் சிஸ்டெர்ன்களின் சிக்கலான கிளை அமைப்பு ஆகும் (படம் 14). இரண்டு வகையான EPS உள்ளன - கரடுமுரடானமற்றும் மென்மையான.ரைபோசோம்கள் கரடுமுரடான சவ்வுகளுடன் இணைக்கப்பட்டுள்ளன (சைட்டோபிளாஸ்மிக் பக்கத்திலிருந்து), ஆனால் அவை மென்மையான சவ்வுகளில் இல்லை.
அரிசி. 14. எண்டோபிளாஸ்மிக் ரெட்டிகுலம்
எண்டோபிளாஸ்மிக் ரெட்டிகுலம் யூகாரியோடிக் கலத்தில் பல முக்கியமான செயல்பாடுகளைச் செய்கிறது:
♦ வரையறுத்தல்(செல்லின் உள் அளவை பல்வேறு எதிர்வினை இடைவெளிகளாகப் பிரித்தல்);
♦ கரிம பொருட்களின் தொகுப்பில் பங்கேற்பு(ரைபோசோம்கள் கரடுமுரடான ER இன் சவ்வுகளில் அமைந்துள்ளன, மேலும் என்சைம் வளாகங்கள் மென்மையானவற்றில் அமைந்துள்ளன, லிப்பிடுகள், கார்போஹைட்ரேட்டுகள் போன்றவற்றின் தொகுப்பை உறுதி செய்கின்றன);
♦ கோல்கி எந்திரம், லைசோசோம்களின் கூறுகளை உருவாக்குவதில் பங்கேற்பு;
♦ பொருட்களின் போக்குவரத்து.
கோல்கி எந்திரம்.கோல்கி எந்திரம் (ஏஜி) என்பது ஒரு அமைப்பு தொட்டிகள்(பிளாட் வெற்றிடங்கள்) மற்றும் குமிழ்கள்(வெசிகல்ஸ்), செல் கருவுக்கு அருகாமையில் அமைந்துள்ளது, அதன் சிறிய துண்டுகள் (படம் 15) பிரிக்கப்பட்டதன் விளைவாக EPS காரணமாக உருவாகின்றன. இந்த துண்டுகள் ஒன்றிணைக்கும்போது, கோல்கி எந்திரத்தின் புதிய தொட்டிகள் எழுகின்றன, அதே நேரத்தில் இபிஎஸ்ஸிலிருந்து பல்வேறு பொருட்கள் கொண்டு செல்லப்படுகின்றன, அவை சிக்கலான கரிம சேர்மங்களின் (புரதங்கள் + கார்போஹைட்ரேட்டுகள், புரதங்கள் + லிப்பிடுகள் போன்றவை) அசெம்பிளி செய்வதில் ஈடுபட்டுள்ளன. செல் வெளியே ஏஜி உதவி. உயிரியல் ரீதியாக செயல்படும் இந்த பொருட்கள் செல்லில் இருந்து அகற்றப்படுகின்றன (எக்சோசைடோசிஸ் மூலம் சுரக்கும் வெற்றிடங்களைப் பயன்படுத்தி) அல்லது AG ஆல் உருவாக்கப்பட்ட லைசோசோம்களின் ஒரு பகுதியாகும் (கீழே காண்க).
அரிசி. 15. கோல்கி எந்திரம்:
கோல்கி எந்திரம் பின்வரும் செயல்பாடுகளை செய்கிறது:
♦ தொகுப்புஉயிரணுவால் உற்பத்தி செய்யப்படும் உயிரியல் ரீதியாக செயல்படும் பொருட்கள்;
♦ பல்வேறு பொருட்களின் சுரப்பு (செல்லிலிருந்து அகற்றுதல்).(ஹார்மோன்கள், என்சைம்கள், செல் சுவர் கட்டப்பட்ட பொருட்கள் போன்றவை);
♦ லைசோசோம்களின் உருவாக்கத்தில் பங்கேற்பு.
மைட்டோகாண்ட்ரியா.அனைத்து வகையான யூகாரியோடிக் செல்கள் மைட்டோகாண்ட்ரியாவைக் கொண்டுள்ளன (படம் 16). அவை வட்டமான உடல்கள் அல்லது தண்டுகள் போன்றவை, குறைவாக அடிக்கடி - நூல்கள். அவற்றின் அளவுகள் 1 முதல் 7 மைக்ரான் வரை இருக்கும். ஒரு கலத்தில் உள்ள மைட்டோகாண்ட்ரியாவின் எண்ணிக்கை பல நூறு முதல் பல்லாயிரக்கணக்கான (பெரிய புரோட்டோசோவாவில்) வரை இருக்கும்.
அரிசி. 16. மைட்டோகாண்ட்ரியா. மேலே - மைட்டோகாண்ட்ரியா (அ) சிறுநீர் கால்வாய்களில், ஒளி நுண்ணோக்கியில் தெரியும். மைட்டோகாண்ட்ரியல் அமைப்பின் முப்பரிமாண மாதிரி கீழே உள்ளது: 1 - cristae; 2 - வெளிப்புற சவ்வு; 3 - உள் சவ்வு; 4 - அணி
மைட்டோகாண்ட்ரியன் இரண்டு சவ்வுகளால் உருவாகிறது - வெளிப்புறமற்றும் உள்,இடையே அமைந்துள்ளது சவ்வு இடைவெளி.உள் சவ்வு பல ஊடுருவல்களை உருவாக்குகிறது - கிறிஸ்டே, அவை தட்டுகள் அல்லது குழாய்கள். இந்த அமைப்பு உள் மென்படலத்தின் ஒரு பெரிய பகுதியை வழங்குகிறது. கரிமப் பொருட்களில் (கார்போஹைட்ரேட்டுகள், லிப்பிடுகள்) உள்ள ஆற்றலை ஏடிபி ஆற்றலாக மாற்றுவதை உறுதி செய்யும் என்சைம்கள் இதில் உள்ளன, இது உயிரணுவின் வாழ்க்கைக்கு அவசியம். எனவே, மைட்டோகாண்ட்ரியாவின் செயல்பாடு பங்கேற்பதாகும் ஆற்றல்செல்லுலார் செயல்முறைகள். அதனால்தான் அதிக எண்ணிக்கையிலான மைட்டோகாண்ட்ரியா இயல்பாகவே உள்ளது, எடுத்துக்காட்டாக, நிறைய வேலைகளைச் செய்யும் தசை செல்களில்.
பிளாஸ்டிடுகள்.தாவர உயிரணுக்களில், சிறப்பு உறுப்புகள் காணப்படுகின்றன - பிளாஸ்டிட்கள், அவை பெரும்பாலும் சுழல் வடிவ அல்லது வட்ட வடிவத்தைக் கொண்டுள்ளன, சில நேரங்களில் மிகவும் சிக்கலானவை. மூன்று வகையான பிளாஸ்டிட்கள் உள்ளன - குளோரோபிளாஸ்ட்கள் (படம் 17), குரோமோபிளாஸ்ட்கள் மற்றும் லுகோபிளாஸ்ட்கள்.
குளோரோபிளாஸ்ட்கள்பச்சை நிறத்தில் வேறுபடுகிறது, இது நிறமி காரணமாக உள்ளது - குளோரோபில்செயல்முறையை உறுதி செய்கிறது ஒளிச்சேர்க்கை,அதாவது, சூரிய ஒளியின் ஆற்றலைப் பயன்படுத்தி நீர் (H 2 O) மற்றும் கார்பன் டை ஆக்சைடு (CO 2) ஆகியவற்றிலிருந்து கரிமப் பொருட்களின் தொகுப்பு. குளோரோபிளாஸ்ட்கள் முக்கியமாக இலை செல்களில் (உயர்ந்த தாவரங்களில்) காணப்படுகின்றன. அவை ஒன்றுக்கொன்று இணையாக அமைந்துள்ள இரண்டு சவ்வுகளால் உருவாகின்றன, அவை குளோரோபிளாஸ்ட்களின் உள்ளடக்கங்களைச் சுற்றியுள்ளன - ஸ்ட்ரோமாஉள் சவ்வு பல தட்டையான பைகளை உருவாக்குகிறது - தைலகாய்டுகள்,அடுக்கப்பட்டவை (நாணயங்களின் அடுக்கு போன்றவை) - தானியங்கள் -மற்றும் ஸ்ட்ரோமாவில் பொய். இது குளோரோபில் கொண்ட தைலகாய்டுகள் ஆகும்.
குரோமோபிளாஸ்ட்கள்பல பூக்கள் மற்றும் பழங்களின் மஞ்சள், ஆரஞ்சு மற்றும் சிவப்பு நிறத்தை தீர்மானிக்கவும், அவை அதிக அளவில் இருக்கும் செல்களில் உள்ளன. அவற்றின் கலவையில் முக்கிய நிறமிகள் உள்ளன கரோட்டின்கள்.குரோமோபிளாஸ்ட்களின் செயல்பாட்டு நோக்கம் விலங்குகளை வண்ணத்துடன் ஈர்ப்பது, பூக்களின் மகரந்தச் சேர்க்கை மற்றும் விதைகளின் பரவலை உறுதி செய்வதாகும்.
அரிசி. 17. பிளாஸ்டிட்கள்: ஒரு - எலோடியா இலையின் செல்களில் உள்ள குளோரோபிளாஸ்ட்கள், ஒளி நுண்ணோக்கியில் தெரியும்; b - க்ரானாவுடன் குளோரோபிளாஸ்டின் உள் கட்டமைப்பின் வரைபடம், அவை குளோரோபிளாஸ்டின் மேற்பரப்பில் செங்குத்தாக அமைந்துள்ள தட்டையான பைகளின் அடுக்குகள்; c - தனிப்பட்ட விசிறி அறைகளை இணைக்கும் அனஸ்டோமோசிங் குழாய்களைக் காட்டும் விரிவான வரைபடம்
லுகோபிளாஸ்ட்கள்தாவரங்களின் நிலத்தடி பகுதிகளின் செல்கள் (உதாரணமாக, உருளைக்கிழங்கு கிழங்குகளில்), விதைகள் மற்றும் தண்டுகளின் மையப்பகுதிகளில் நிறமற்ற பிளாஸ்டிட்கள் உள்ளன. லுகோபிளாஸ்ட்களில், ஸ்டார்ச் முக்கியமாக குளுக்கோஸிலிருந்து உருவாகிறது மற்றும் தாவரங்களின் சேமிப்பு உறுப்புகளில் குவிக்கப்படுகிறது.
ஒரு வகை பிளாஸ்டிட்கள் மற்றொரு வகையாக மாறலாம். எடுத்துக்காட்டாக, இலையுதிர் காலத்தில் இலைகள் நிறம் மாறும்போது, குளோரோபிளாஸ்ட்கள் குரோமோபிளாஸ்ட்களாக மாறுகின்றன.
லைசோசோம்கள்.இந்த உறுப்புகள் 2 மைக்ரான் வரை விட்டம் கொண்ட சவ்வால் சூழப்பட்ட வெசிகல்கள் போல இருக்கும். புரதங்கள், நியூக்ளிக் அமிலங்கள், பாலிசாக்கரைடுகள் மற்றும் லிப்பிட்களை உடைக்கும் பல டஜன் நொதிகள் அவற்றில் உள்ளன. லைசோசோம்களின் செயல்பாடு சிக்கலான கரிம சேர்மங்களின் உள்செல்லுலார் முறிவின் செயல்முறைகளில் பங்கேற்பதாகும் (உதாரணமாக, "செலவு" செல்லுலார் கூறுகளின் ஊட்டச்சத்துக்கள் அல்லது பொருட்கள்). லைசோசோம்கள் பாகோசைடிக் (அல்லது பினோசைடிக்) வெற்றிடங்களுடன் இணைந்து செரிமான வெற்றிடத்தை உருவாக்குகின்றன.
லைசோசோம்களின் உருவாக்கம் கோல்கி கருவியின் சிஸ்டெர்னாவிலிருந்து வளரும் காரணமாக ஏற்படுகிறது.
ரைபோசோம்கள்.ரைபோசோம்கள் (படம் 18) யூகாரியோட்டுகள் மற்றும் புரோகாரியோட்டுகள் இரண்டின் செல்களிலும் உள்ளன, ஏனெனில் அவை ஒரு முக்கியமான செயல்பாட்டைச் செய்கின்றன. புரத உயிரியக்கவியல்(அத்தியாயம் 5 பார்க்கவும்). ஒவ்வொரு கலத்திலும் பல்லாயிரக்கணக்கான, நூறாயிரக்கணக்கான (பல மில்லியன்கள் வரை) இந்த சிறிய வட்ட உறுப்புகள் உள்ளன.
அரிசி. 18. எண்டோபிளாஸ்மிக் ரெட்டிகுலத்தின் மென்படலத்தில் அமர்ந்திருக்கும் ரைபோசோமின் கட்டமைப்பின் திட்டம்: 1 - சிறிய துணைக்குழு; 2 - டிஆர்என்ஏ; 3 - அமினோசைல்-டிஆர்என்ஏ; 4 - அமினோ அமிலம்; 5 - பெரிய துணைக்குழு; 6 - எண்டோபிளாஸ்மிக் ரெட்டிகுலம் சவ்வு; 7 - ஒருங்கிணைக்கப்பட்ட பாலிபெப்டைட் சங்கிலி
ரைபோசோம் இரண்டு சமமற்ற துணைக்குழுக்களை (பாகங்கள்) கொண்டுள்ளது. அவை தனித்தனியாக உருவாக்கப்பட்டு, புரத மூலக்கூறின் தொகுப்பின் போது, மெசஞ்சர் ஆர்.என்.ஏ.வை "சூழ்ந்திருக்கும்". ரைபோசோம்களில் பல்வேறு புரதங்கள் மற்றும் ரைபோசோமால் ஆர்என்ஏக்கள் உள்ளன.
செல்லுலார் சேர்த்தல்கள்.தானியங்கள், துகள்கள் அல்லது நீர்த்துளிகள் வடிவில் சைட்டோபிளாஸின் முக்கிய பொருளில் இருக்கும் கலத்தில் உள்ள நிரந்தரமற்ற கூறுகளுக்கு இது பெயர். சேர்த்தல்கள் ஒரு சவ்வினால் சூழப்பட்டிருக்கலாம் அல்லது இல்லாமல் இருக்கலாம்.
செயல்பாட்டு ரீதியாக, மூன்று வகையான சேர்த்தல்கள் உள்ளன: இருப்பு ஊட்டச்சத்துக்கள்(ஸ்டார்ச், கிளைகோஜன், கொழுப்புகள், புரதங்கள்), சுரப்பு சேர்த்தல்கள்(சுரப்பி உயிரணுக்களின் சிறப்பியல்பு பொருட்கள், அவற்றால் உற்பத்தி செய்யப்படுகின்றன - நாளமில்லா சுரப்பிகளின் ஹார்மோன்கள் போன்றவை) மற்றும் சிறப்பு நோக்கத்தைச் சேர்த்தல்(அதிக சிறப்பு வாய்ந்த உயிரணுக்களில், எடுத்துக்காட்டாக சிவப்பு இரத்த அணுக்களில் ஹீமோகுளோபின்).
§ 3. செல் அணுக்கருவின் அமைப்பு. குரோமோசோம்கள்
செல் கரு (படம் 8 மற்றும் 9 ஐப் பார்க்கவும்) செல்லின் வாழ்க்கையில் மிகவும் முக்கியத்துவம் வாய்ந்தது, ஏனெனில் இது குரோமோசோம்களில் உள்ள பரம்பரை தகவல்களின் களஞ்சியமாக செயல்படுகிறது (கீழே காண்க).
அணுக்கரு அதன் உள்ளடக்கங்களை பிரிக்கும் அணுக்கரு உறையால் கட்டப்பட்டுள்ளது (காரியோபிளாசம்)சைட்டோபிளாஸில் இருந்து. ஷெல் ஒரு இடைவெளியால் பிரிக்கப்பட்ட இரண்டு சவ்வுகளைக் கொண்டுள்ளது. அவை இரண்டும் ஏராளமான துளைகளால் ஊடுருவி உள்ளன, இதன் காரணமாக கரு மற்றும் சைட்டோபிளாசம் இடையே பொருட்களின் பரிமாற்றம் சாத்தியமாகும். பெரும்பாலான யூகாரியோட்டுகளின் செல் அணுக்கருவில் 1 முதல் 7 வரை இருக்கும் நியூக்ளியோலி.ஆர்என்ஏ மற்றும் டிஆர்என்ஏ தொகுப்பின் செயல்முறைகள் அவற்றுடன் தொடர்புடையவை.
கர்னலின் முக்கிய கூறுகள் - குரோமோசோம்கள்,டிஎன்ஏ மூலக்கூறு மற்றும் பல்வேறு புரதங்களிலிருந்து உருவாகிறது. ஒளி நுண்ணோக்கியில் அவை செல் பிரிவின் போது மட்டுமே தெளிவாகத் தெரியும் (மைட்டோசிஸ், ஒடுக்கற்பிரிவு).பிரிக்கப்படாத கலத்தில், குரோமோசோம்கள் கருவின் முழு அளவு முழுவதும் விநியோகிக்கப்படும் நீண்ட மெல்லிய நூல்களைப் போல இருக்கும்.
செல் பிரிவின் போது, குரோமோசோமால் இழைகள் அடர்த்தியான சுருள்களை உருவாக்குகின்றன, இதன் விளைவாக அவை தண்டுகள், "ஹேர்பின்கள்" வடிவத்தில் (வழக்கமான நுண்ணோக்கியைப் பயன்படுத்தி) தெரியும். மரபணு தகவல்களின் முழு அளவும் கருவின் குரோமோசோம்களுக்கு இடையில் விநியோகிக்கப்படுகிறது. அவற்றைப் படிக்கும் செயல்பாட்டில், பின்வரும் வடிவங்கள் அடையாளம் காணப்பட்டன:
♦ சோமாடிக் செல்களின் கருக்களில் (அதாவது, உடலின் செல்கள், பாலியல் அல்லாதவை) ஒரே இனத்தைச் சேர்ந்த அனைத்து நபர்களும் ஒரே எண்ணிக்கையிலான குரோமோசோம்களைக் கொண்டுள்ளனர். குரோமோசோம்களின் தொகுப்பு(படம் 19);
அரிசி. 19. வெவ்வேறு வகையான தாவரங்கள் மற்றும் விலங்குகளின் குரோமோசோம்கள், அதே அளவில் சித்தரிக்கப்படுகின்றன: 1,2 - அமீபா; 3,4 - டயட்டம்கள்; 5-8, 18,19 - பச்சை ஆல்கா; 9 - பறக்க agaric; 10 - லிண்டன்; 11-12 - டிரோசோபிலா; 13 - சால்மன்; 14 - skerda (Asteraceae குடும்பம்); 15 - அராய்டு குடும்பத்தைச் சேர்ந்த ஆலை; 16 - கோரிடலிஸ் பட்டாம்பூச்சி; 17 - வெட்டுக்கிளி குடும்பத்தைச் சேர்ந்த பூச்சி; 20 - வாட்டர் ஸ்ட்ரைடர் பிழை; 21 - மலர் பிழை; 22 - ஆம்பிபியன் ஆம்பிஸ்டோமா; 23 - கற்றாழை (லில்லி குடும்பம்)
♦ ஒவ்வொரு இனமும் அவற்றின் எண்ணிக்கைக்கு ஏற்ப அதன் சொந்த குரோமோசோம் தொகுப்பால் வகைப்படுத்தப்படுகிறது (உதாரணமாக, ஒரு நபருக்கு 46 குரோமோசோம்கள் உள்ளன, ஒரு ட்ரோசோபிலா ஈ 8, ஒரு வட்டப்புழு 4, ஒரு நண்டு 196, குதிரைக்கு 66 மற்றும் சோளத்தில் 104 உள்ளன);
♦ சோமாடிக் செல்களின் கருக்களில் உள்ள குரோமோசோம்கள் எனப்படும் ஜோடிகளாக தொகுக்கப்படலாம் ஒரே மாதிரியான குரோமோசோம்கள்அவற்றின் ஒற்றுமையின் அடிப்படையில் (கட்டமைப்பு மற்றும் செயல்பாட்டில்);
♦ கிருமி உயிரணுக்களின் (கேமட்கள்) கருக்களில், ஒவ்வொரு ஜோடி ஹோமோலோகஸ் குரோமோசோம்களிலும் ஒன்று மட்டுமே உள்ளது, அதாவது, குரோமோசோம்களின் மொத்த தொகுப்பு சோமாடிக் செல்களின் பாதி;
♦ கிருமி உயிரணுக்களில் உள்ள குரோமோசோம்களின் ஒரு தொகுப்பு அழைக்கப்படுகிறது ஹாப்ளாய்டுமற்றும் n என்ற எழுத்தால் குறிக்கப்படுகிறது, மற்றும் சோமாடிக் - டிப்ளாய்டு(2n)
மேற்கூறியவற்றிலிருந்து ஒவ்வொரு ஜோடி ஹோமோலோகஸ் குரோமோசோம்களும் கருத்தரிப்பின் போது தந்தை மற்றும் தாய்வழி குரோமோசோம்களின் இணைப்பால் உருவாகின்றன என்பது தெளிவாகிறது, அதாவது பாலின உயிரணுக்களின் (கேமட்கள்) இணைவு. மாறாக, கிருமி உயிரணுக்கள் உருவாகும் போது, ஒவ்வொரு ஜோடி ஹோமோலோகஸ் குரோமோசோம்களிலிருந்தும் ஒன்று மட்டுமே கேமட்டிற்குள் நுழைகிறது.
குரோமோசோம்கள் வெவ்வேறுஒரே மாதிரியான ஜோடிகள் அளவு மற்றும் வடிவத்தில் வேறுபடுகின்றன (படம் 20 மற்றும் 21).
அரிசி. 20. குரோமோசோம்களின் கட்டமைப்பு மற்றும் வகைகள்: a - தோற்றம் 1 - சென்ட்ரோமியர்; 2 - குறுகிய தோள்பட்டை; 3 - நீண்ட தோள்பட்டை); அதே குரோமோசோமின் உள் அமைப்பு (1 - சென்ட்ரோமியர்; - டிஎன்ஏ மூலக்கூறுகள்); c – குரோமோசோம்களின் வகைகள் (1 – ஒற்றை ஆயுதம்; வெவ்வேறு ஆயுதம்; 3 – சம ஆயுதம்: X – கை, V – சென்ட்ரோமியர்)
அரிசி. 21. ஒரு குரோமோசோம் டிஎன்ஏ மற்றும் புரதங்களைக் கொண்டுள்ளது. டிஎன்ஏ மூலக்கூறு நகலெடுக்கப்படுகிறது. ஒரே மாதிரியான இரண்டு டிஎன்ஏ இரட்டை ஹெலிகள் சென்ட்ரோமியர் பகுதியில் இணைக்கப்பட்டுள்ளன. இந்த பிரதிகள் பின்னர் செல் பிரிவின் போது தனி குரோமோசோம்களாக மாறும்
குரோமோசோம்கள் உடலில் உள்ளன முதன்மை சுருக்கம் (சென்ட்ரோமியர் என்று அழைக்கப்படுகிறது),அதில் நூல்கள் இணைக்கப்பட்டுள்ளன பிளவு சுழல்கள்.இது குரோமோசோமை இரண்டாகப் பிரிக்கிறது தோள்பட்டைகுரோமோசோம்கள் சம ஆயுதம், சமமற்ற ஆயுதம் அல்லது ஒற்றை ஆயுதம் கொண்டதாக இருக்கலாம்.
அத்தியாயம் 5. வளர்சிதை மாற்றம்
§ 1. வளர்சிதை மாற்றம் ஒருங்கிணைத்தல் மற்றும் ஒற்றுமையின் ஒற்றுமை
அனைத்து செல்கள் மற்றும் உயிரினங்கள் திறந்த அமைப்புகள், அதாவது, அவை சுற்றுச்சூழலுடன் ஆற்றல் மற்றும் பொருட்களின் நிலையான பரிமாற்ற நிலையில் உள்ளன. உயிரற்ற இயற்கையில் திறந்த அமைப்புகள் உள்ளன, ஆனால் அவற்றின் இருப்பு உயிரினங்களிலிருந்து தரமான முறையில் வேறுபட்டது. இந்த உதாரணத்தைக் கவனியுங்கள்: எரியும் கந்தகத்தின் ஒரு பகுதி சுற்றுச்சூழலுடன் பரிமாறிக்கொள்ளும் நிலையில் உள்ளது. அது எரியும் போது, O 2 உறிஞ்சப்பட்டு, SO 2 மற்றும் ஆற்றல் (வெப்ப வடிவில்) வெளியிடப்படுகிறது. இருப்பினும், இந்த விஷயத்தில், ஒரு உடல் உடலாக கந்தகத்தின் ஒரு பகுதி அழிக்கப்பட்டு அதன் முதன்மை அமைப்பை இழக்கிறது.
உயிரினங்களைப் பொறுத்தவரை, சுற்றுச்சூழலுடனான பரிமாற்றம், அவை கொண்டிருக்கும் அனைத்து பொருட்கள் மற்றும் கூறுகளின் சுய-புதுப்பித்தல் மூலம் அவற்றின் கட்டமைப்பு அமைப்பைப் பாதுகாப்பதற்கும் பராமரிப்பதற்கும் ஒரு நிபந்தனையாக மாறும்.
வளர்சிதை மாற்றம் (வளர்சிதை மாற்றம்) என்பது உயிரினங்களில் நிகழும் செயல்முறைகளின் தொகுப்பாகும் (நுகர்வு, மாற்றம், குவிப்பு மற்றும் பொருட்கள் மற்றும் ஆற்றலின் வெளியீடு) அவற்றின் முக்கிய செயல்பாடு, வளர்ச்சி, வளர்ச்சி, இனப்பெருக்கம் ஆகியவற்றை உறுதி செய்கிறது. வளர்சிதை மாற்ற செயல்பாட்டின் போது, செல்களை உருவாக்கும் மூலக்கூறுகள் உடைந்து ஒருங்கிணைக்கப்படுகின்றன; செல்லுலார் கட்டமைப்புகள் மற்றும் இன்டர்செல்லுலர் பொருள் புதுப்பித்தல்.
வளர்சிதை மாற்றம் ஒன்றோடொன்று இணைக்கப்பட்ட செயல்முறைகளை அடிப்படையாகக் கொண்டது ஒருங்கிணைப்பு(அனபோலிசம்) மற்றும் வேறுபாடு(கேடபாலிசம்). ஒருங்கிணைப்பின் போது (பிளாஸ்டிக் பரிமாற்றம்), சிக்கலான பொருட்கள் எளிமையானவற்றிலிருந்து ஒருங்கிணைக்கப்படுகின்றன. இதற்கு நன்றி, அதன் கட்டமைப்பு கூறுகள், நொதி அமைப்புகள் போன்றவற்றை உருவாக்குவதற்கு தேவையான கலத்தில் உள்ள அனைத்து கரிமப் பொருட்களும் உருவாக்கப்படுகின்றன. ஒருங்கிணைப்பு எப்போதும் ஆற்றல் செலவில் நடைபெறுகிறது.
ஒற்றுமையின் போது (ஆற்றல் வளர்சிதை மாற்றம்), சிக்கலான கரிம பொருட்கள் எளிமையான அல்லது கனிமமாக உடைக்கப்படுகின்றன. இந்த வழக்கில், ஆற்றல் வெளியிடப்படுகிறது, இது செல் அதன் முக்கிய செயல்பாட்டை உறுதி செய்யும் பல்வேறு செயல்முறைகளைச் செய்ய செலவிடப்படுகிறது (பொருட்களின் தொகுப்பு மற்றும் போக்குவரத்து, இயந்திர வேலை போன்றவை).
அனைத்து உயிரினங்களையும் இரண்டு குழுக்களாகப் பிரிக்கலாம்: autotrophsமற்றும் ஹீட்டோரோட்ரோப்கள்,ஆற்றல் மூலங்கள் மற்றும் அவற்றின் முக்கிய செயல்பாடுகளை உறுதிப்படுத்த தேவையான பொருட்களில் வேறுபடுகின்றன.
ஆட்டோட்ரோப்கள்- சூரிய ஒளியின் ஆற்றலைப் பயன்படுத்தி கனிமப் பொருட்களிலிருந்து கரிம சேர்மங்களை ஒருங்கிணைக்கும் உயிரினங்கள் போட்டோட்ரோப்கள்தாவரங்கள், சயனோபாக்டீரியா) அல்லது கனிம (கனிம) பொருட்களின் ஆக்சிஜனேற்றத்திலிருந்து பெறப்பட்ட ஆற்றல் (அதாவது வேதியியல்- சல்பர் பாக்டீரியா, இரும்பு பாக்டீரியா, முதலியன). இதன் விளைவாக, அவர்கள் தங்கள் வாழ்க்கை நடவடிக்கைக்குத் தேவையான பொருட்களை சுயாதீனமாக உருவாக்க முடியும்.
§ 2. காற்றில்லா மற்றும் ஏரோபிக் உயிரினங்களில் வேறுபாடு
உயிரினங்களை இரு குழுக்களாகப் பிரிக்கலாம் மற்றும் ஒற்றுமையின் தன்மைக்கு ஏற்ப - ஏரோப்ஸ்மற்றும் காற்றில்லா.ஏரோப்ஸ் (கிரேக்க மொழியில் இருந்து. பேய்- காற்று) வாழ்க்கைக்கு இலவச ஆக்ஸிஜன் தேவைப்படுகிறது. அனேரோப்ஸில் (கிரேக்கம். ஏசி- எதிர்மறை துகள்) இது தேவையில்லை. அவற்றில், நொதித்தல் - ஆக்ஸிஜன் இல்லாத, கரிமப் பொருட்களின் நொதி முறிவு, எளிமையான கரிமப் பொருட்களின் உருவாக்கம் மற்றும் ஆற்றலின் வெளியீடு ஆகியவற்றால் பிரிக்கப்படுகிறது. உதாரணத்திற்கு:
♦ லாக்டிக் அமில நொதித்தல்:
C 6 H 12 O 6 + 2H 3 PO 4 + 2ADP → 2PH + 2ATP + 2H 2 O;
♦ ஆல்கஹால் நொதித்தல்:
C 6 H 12 O 6 + 2PH + 2ADP → 2C 2 H 5 OH + 2CO 2 + 2ATP + 2H 2 O.
நொதித்தலின் போது உருவாகும் பொருட்கள் கரிமமானவை, எனவே இன்னும் நிறைய ஆற்றலைக் கொண்டிருக்கின்றன.
அரிசி. 22. தன்னியக்க மற்றும் ஹீட்டோரோட்ரோபிக் உயிரினங்களில் ஒருங்கிணைப்பு மற்றும் ஒற்றுமைக்கு இடையிலான உறவுகள்
ஏரோபிக் உயிரினங்களில், மைட்டோகாண்ட்ரியாவில் சுவாசத்தின் போது, கரிமப் பொருட்களின் முழுமையான முறிவு (O 2 ஐப் பயன்படுத்தி) ஆற்றல்-மோசமான இறுதி தயாரிப்புகளான CO 2 மற்றும் H 2 O க்கு ஏற்படுகிறது மற்றும் குறிப்பிடத்தக்க அளவு ஆற்றல் வெளியிடப்படுகிறது:
C 6 H 12 0 6 (குளுக்கோஸ்) + 0 2 > 6C0 2 + 6H 2 0 + ஆற்றல் (இதன் காரணமாக 38 ATP மூலக்கூறுகள் ஒருங்கிணைக்கப்படுகின்றன).
ஆட்டோட்ரோபிக் மற்றும் ஹீட்டோரோட்ரோபிக் ஏரோபிக் உயிரினங்களின் வளர்சிதை மாற்றத்தை பொதுவான திட்டங்களின் வடிவத்தில் கருத்தில் கொள்வோம் (படம் 22).
ஒருங்கிணைப்பு.அதன் மிக முக்கியமான செயல்முறைகள் ஒளிச்சேர்க்கைமற்றும் புரதங்களின் உயிரியக்கவியல்.
சுற்றுச்சூழலுடன் உயிரினத்தின் இணைப்பு, ஒரு இயற்பியல் வேதியியல் பார்வையில், ஒரு திறந்த அமைப்பு, அதாவது, உயிர்வேதியியல் செயல்முறைகள் நடந்து கொண்டிருக்கும் ஒரு அமைப்பு. தொடக்கப் பொருட்கள் சுற்றுச்சூழலில் இருந்து வருகின்றன, மேலும் தொடர்ச்சியாக உருவாகும் பொருட்களும் வெளியே கொண்டு செல்லப்படுகின்றன. உடலில் உள்ள பலதரப்பு எதிர்வினைகளின் தயாரிப்புகளின் வேகம் மற்றும் செறிவு ஆகியவற்றுக்கு இடையேயான சமநிலை நிபந்தனைக்குட்பட்டது, கற்பனையானது, ஏனெனில் உட்கொள்வது மற்றும் பொருட்களை அகற்றுவது நிறுத்தப்படாது. சுற்றுச்சூழலுடனான தொடர்ச்சியான தொடர்பு ஒரு உயிரினத்தை ஒரு திறந்த அமைப்பாகக் கருத அனுமதிக்கிறது.
அனைத்து உயிரணுக்களுக்கும், ஆற்றல் மூலமானது சூரியன். தாவர செல்கள் குளோரோபில் உதவியுடன் சூரிய ஒளியில் இருந்து ஆற்றலைப் பிடிக்கின்றன, ஒளிச்சேர்க்கை செயல்பாட்டின் போது ஒருங்கிணைப்பு எதிர்வினைகளுக்கு அதைப் பயன்படுத்துகின்றன. பூமிக்குரிய தாவரங்களால் தொகுக்கப்பட்ட கரிமப் பொருட்களின் முறிவின் போது விலங்குகள், பூஞ்சை மற்றும் பாக்டீரியாக்களின் செல்கள் சூரிய சக்தியை மறைமுகமாகப் பயன்படுத்துகின்றன.
செல்லுலார் சுவாசத்தின் போது செல்லின் சில ஊட்டச்சத்துக்கள் உடைக்கப்படுகின்றன, இதனால் பல்வேறு வகையான செல்லுலார் செயல்பாட்டிற்கு தேவையான ஆற்றலை வழங்குகிறது. இந்த செயல்முறை மைட்டோகாண்ட்ரியா எனப்படும் உறுப்புகளில் நடைபெறுகிறது. மைட்டோகாண்ட்ரியா இரண்டு சவ்வுகளைக் கொண்டுள்ளது: வெளிப்புறமானது, சைட்டோபிளாஸத்திலிருந்து உறுப்புகளை பிரிக்கிறது, மற்றும் உள் ஒன்று, பல மடிப்புகளை உருவாக்குகிறது. சுவாசத்தின் முக்கிய தயாரிப்பு ஏடிபி ஆகும். இது மைட்டோகாண்ட்ரியாவை விட்டு வெளியேறுகிறது மற்றும் சைட்டோபிளாசம் மற்றும் செல் சவ்வுகளில் பல இரசாயன எதிர்வினைகளுக்கு ஆற்றல் மூலமாகப் பயன்படுத்தப்படுகிறது. செல்லுலார் சுவாசத்திற்கு ஆக்ஸிஜன் தேவைப்பட்டால், சுவாசம் ஏரோபிக் என்று அழைக்கப்படுகிறது, ஆனால் ஆக்ஸிஜன் இல்லாத நிலையில் எதிர்வினைகள் ஏற்பட்டால், நாம் காற்றில்லா சுவாசம் பற்றி பேசுகிறோம்.
ஒரு கலத்தில் செய்யப்படும் எந்தவொரு வேலைக்கும், ஆற்றல் ஒரே வடிவத்தில் பயன்படுத்தப்படுகிறது - ATP இன் பாஸ்பேட் பிணைப்புகளிலிருந்து ஆற்றல் வடிவத்தில். ஏடிபி என்பது எளிதில் செல்லக்கூடிய கலவையாகும். ஏடிபியின் உருவாக்கம் மைட்டோகாண்ட்ரியாவின் உள் மென்படலத்தில் நிகழ்கிறது. கார்போஹைட்ரேட்டுகள், கொழுப்புகள் மற்றும் பிற கரிமப் பொருட்களின் ஆக்சிஜனேற்றத்தின் ஆற்றல் காரணமாக சுவாசத்தின் போது அனைத்து உயிரணுக்களிலும் ஏடிபி ஒருங்கிணைக்கப்படுகிறது. பச்சை தாவர செல்களில், ATP இன் முக்கிய அளவு சூரிய ஆற்றல் காரணமாக குளோரோபிளாஸ்ட்களில் ஒருங்கிணைக்கப்படுகிறது. ஒளிச்சேர்க்கையின் போது, அவை மைட்டோகாண்ட்ரியாவை விட பல மடங்கு அதிக ஏடிபியை உற்பத்தி செய்கின்றன. பாஸ்பரஸ்-ஆக்ஸிஜன் பிணைப்புகளின் முறிவு மற்றும் ஆற்றலின் வெளியீட்டில் ATP சிதைகிறது. ATP இன் நீராற்பகுப்பின் போது ATPase என்ற நொதியின் செயல்பாட்டின் கீழ் இது நிகழ்கிறது - ஒரு பாஸ்போரிக் அமில மூலக்கூறை நீக்குவதன் மூலம் தண்ணீரைச் சேர்ப்பது. இதன் விளைவாக, ஏடிபி ஏடிபியாக மாற்றப்படுகிறது, மேலும் பாஸ்போரிக் அமிலத்தின் இரண்டு மூலக்கூறுகள் பிரிக்கப்பட்டால், பின்னர் ஏஎம்பியாக மாறுகிறது. அமிலத்தின் ஒவ்வொரு கிராம்-மூலக்கூறின் நீக்குதலின் எதிர்வினை 40 kJ வெளியீட்டுடன் சேர்ந்துள்ளது. இது மிகப் பெரிய ஆற்றல் வெளியீடு ஆகும், அதனால்தான் ATP இன் பாஸ்பரஸ்-ஆக்ஸிஜன் பிணைப்புகள் பொதுவாக மேக்ரோர்ஜிஸ்டிக் (உயர் ஆற்றல்) என்று அழைக்கப்படுகின்றன.
பிளாஸ்டிக் பரிமாற்ற எதிர்வினைகளில் ஏடிபியின் பயன்பாடு அவற்றை ஏடிபி ஹைட்ரோலிசிஸுடன் இணைப்பதன் மூலம் மேற்கொள்ளப்படுகிறது. ஏடிபி மூலக்கூறிலிருந்து ஹைட்ரோலிசிஸின் போது வெளியிடப்படும் பாஸ்பரஸ் குழுவை இணைப்பதன் மூலம், அதாவது பாஸ்போரிலேஷன் மூலம் பல்வேறு பொருட்களின் மூலக்கூறுகள் ஆற்றலுடன் சார்ஜ் செய்யப்படுகின்றன.
பாஸ்பேட் வழித்தோன்றல்களின் தனித்தன்மை என்னவென்றால், அவை கலத்தை விட்டு வெளியேற முடியாது, இருப்பினும் அவற்றின் "வெளியேற்ற" வடிவங்கள் சவ்வு வழியாக சுதந்திரமாக செல்கின்றன. இதற்கு நன்றி, பாஸ்போரிலேட்டட் மூலக்கூறுகள் சரியான எதிர்வினைகளில் பயன்படுத்தப்படும் வரை கலத்தில் இருக்கும்.
ஏடிபியை ஏடிபியாக மாற்றும் தலைகீழ் செயல்முறையானது ஏடிபியில் பாஸ்போரிக் அமில மூலக்கூறைச் சேர்ப்பதன் மூலம் நிகழ்கிறது, தண்ணீரை வெளியிடுகிறது மற்றும் அதிக அளவு ஆற்றலை உறிஞ்சுகிறது.
எனவே, ஏடிபி என்பது செல் செயல்பாட்டிற்கான உலகளாவிய மற்றும் நேரடி ஆற்றல் மூலமாகும். இது ஒற்றை செல்லுலார் ஆற்றலை உருவாக்கி, செல்லின் ஒரு பகுதியிலிருந்து மற்றொரு பகுதிக்கு மறுபகிர்வு செய்து கொண்டு செல்வதை சாத்தியமாக்குகிறது.
பாஸ்பேட் குழுவின் பரிமாற்றமானது மோனோமர்களில் இருந்து மேக்ரோமோலிகுல்களின் அசெம்பிளி போன்ற இரசாயன எதிர்வினைகளில் முக்கிய பங்கு வகிக்கிறது. எடுத்துக்காட்டாக, அமினோ அமிலங்கள் முன்பு பாஸ்போரிலேட்டட் செய்யப்பட்ட பின்னரே பெப்டைடுகளாக இணைக்கப்படும். சுருக்கம் அல்லது இயக்கத்தின் இயந்திர செயல்முறைகள், ஒரு செறிவு சாய்வுக்கு எதிராக கரைந்த பொருளின் போக்குவரத்து மற்றும் பிற செயல்முறைகள் ATP இல் சேமிக்கப்பட்ட ஆற்றலின் நுகர்வு ஆகியவற்றை உள்ளடக்கியது.
ஆற்றல் வளர்சிதை மாற்றத்தின் செயல்முறையை பின்வருமாறு குறிப்பிடலாம். சைட்டோபிளாஸில் உள்ள உயர்-மூலக்கூறு கரிமப் பொருட்கள், நீராற்பகுப்பு மூலம், அவை எளிமையானவையாக மாற்றப்படுகின்றன: புரதங்கள் - அமினோ அமிலங்கள், பாலி- மற்றும் டிசாக்கரைடுகள் - மோனோசாக்கரைடுகள் (+ குளுக்கோஸ்), கொழுப்புகள் கிளிசரால் மற்றும் கொழுப்பு அமிலங்கள். ஆக்ஸிஜனேற்ற செயல்முறைகள் எதுவும் இல்லை, சிறிய ஆற்றல் வெளியிடப்படுகிறது, இது பயன்படுத்தப்படாது மற்றும் வெப்ப வடிவத்திற்கு செல்கிறது. பெரும்பாலான செல்கள் முதலில் கார்போஹைட்ரேட்டைப் பயன்படுத்துகின்றன. பாலிசாக்கரைடுகள் (தாவரங்களில் ஸ்டார்ச் மற்றும் விலங்குகளில் கிளைகோஜன்) குளுக்கோஸாக ஹைட்ரோலைஸ் செய்யப்படுகிறது. குளுக்கோஸ் ஆக்சிஜனேற்றம் மூன்று கட்டங்களில் நிகழ்கிறது: கிளைகோலிசிஸ், ஆக்ஸிஜனேற்ற டிகார்பாக்சிலேஷன் (கிரெப்ஸ் சுழற்சி - சிட்ரிக் அமில சுழற்சி) மற்றும் ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷன் (சுவாச சங்கிலி). கிளைகோலிசிஸ், இதன் விளைவாக ஒரு குளுக்கோஸ் மூலக்கூறு பைருவிக் அமிலத்தின் இரண்டு மூலக்கூறுகளாகப் பிரிக்கப்பட்டு ஏடிபியின் இரண்டு மூலக்கூறுகளை வெளியிடுகிறது, இது சைட்டோபிளாஸில் ஏற்படுகிறது. ஆக்ஸிஜன் இல்லாத நிலையில், பைருவிக் அமிலம் எத்தனால் (நொதித்தல்) அல்லது லாக்டிக் அமிலம் (காற்றில்லா சுவாசம்) ஆக மாற்றப்படுகிறது.
விலங்கு உயிரணுக்களில் கிளைகோலிசிஸ் ஏற்படுகையில், குளுக்கோஸின் ஆறு கார்பன் மூலக்கூறு லாக்டிக் அமிலத்தின் இரண்டு மூலக்கூறுகளாக உடைகிறது. இந்த செயல்முறை பல கட்டமாகும். இது 13 என்சைம்களால் தொடர்ச்சியாக மேற்கொள்ளப்படுகிறது. ஆல்கஹால் நொதித்தல் போது, ஒரு குளுக்கோஸ் மூலக்கூறிலிருந்து எத்தனால் இரண்டு மூலக்கூறுகளும் CO2 இன் இரண்டு மூலக்கூறுகளும் உருவாகின்றன.
கிளைகோலிசிஸ் என்பது காற்றில்லா மற்றும் ஏரோபிக் சுவாசத்திற்கு பொதுவான ஒரு கட்டமாகும்; ஆக்ஸிஜன் இல்லாத ஆக்சிஜனேற்றத்தின் செயல்முறை, இதில் வளர்சிதை மாற்றங்களின் ஆற்றலின் ஒரு பகுதி மட்டுமே வெளியிடப்பட்டு பயன்படுத்தப்படுகிறது, இது காற்றில்லா உயிரினங்களுக்கு இறுதியானது. ஆக்ஸிஜன் முன்னிலையில், பைருவிக் அமிலம் மைட்டோகாண்ட்ரியாவுக்குள் செல்கிறது, அங்கு, பல தொடர்ச்சியான எதிர்வினைகளின் விளைவாக, ஏடிபிக்கு ஏடிபிக்கு ஒரே நேரத்தில் பாஸ்போரிலேஷனுடன் H2O மற்றும் CO2 ஆக முழுமையாக ஆக்ஸிஜனேற்றப்படுகிறது. இந்த வழக்கில், இரண்டு ஏடிபி மூலக்கூறுகள் கிளைகோலிசிஸால் உற்பத்தி செய்யப்படுகின்றன, இரண்டு கிரெப்ஸ் சுழற்சியால் மற்றும் 34 சுவாச சங்கிலியால் உற்பத்தி செய்யப்படுகின்றன. ஒரு குளுக்கோஸ் மூலக்கூறின் முழுமையான ஆக்சிஜனேற்றம் H2O மற்றும் CO2 ஆக 38 மூலக்கூறுகள் ஆகும்.
எனவே, ஏரோபிக் உயிரினங்களில், கரிமப் பொருட்களின் இறுதி சிதைவு வளிமண்டல ஆக்ஸிஜனுடன் எளிய கனிம பொருட்களாக ஆக்சிஜனேற்றம் செய்வதன் மூலம் மேற்கொள்ளப்படுகிறது: CO2 மற்றும் H2O. இந்த செயல்முறை மைட்டோகாண்ட்ரியாவின் கிறிஸ்டேயில் நடைபெறுகிறது. இந்த வழக்கில், அதிகபட்ச அளவு இலவச ஆற்றல் வெளியிடப்படுகிறது, இதில் குறிப்பிடத்தக்க பகுதி ஏடிபி மூலக்கூறுகளில் ஒதுக்கப்பட்டுள்ளது. ஏரோபிக் ஆக்சிஜனேற்றம் செல்லுக்கு அதிக அளவு இலவச ஆற்றலை வழங்குகிறது என்பதை எளிதாகக் காணலாம்.
கேடபாலிசத்தின் விளைவாக, ஆற்றல் நிறைந்த ATP மூலக்கூறுகள் கலத்தில் குவிந்து, CO2 மற்றும் அதிகப்படியான நீர் வெளிப்புற சூழலில் வெளியிடப்படுகின்றன.
சுவாசத்திற்குத் தேவையில்லாத சர்க்கரை மூலக்கூறுகள் செல்லில் சேமிக்கப்படும். அதிகப்படியான லிப்பிடுகள் உடைக்கப்படுகின்றன, அதன் பிறகு அவற்றின் முறிவின் தயாரிப்புகள் மைட்டோகாண்ட்ரியாவில் சுவாசத்திற்கான அடி மூலக்கூறாக நுழைகின்றன, அல்லது கொழுப்புத் துளிகள் வடிவில் சைட்டோபிளாஸில் இருப்புகளாக வைக்கப்படுகின்றன. புரதங்கள் செல்லுக்குள் நுழையும் அமினோ அமிலங்களிலிருந்து உருவாக்கப்படுகின்றன. ரைபோசோம்கள் எனப்படும் உறுப்புகளில் புரதத் தொகுப்பு ஏற்படுகிறது. ஒவ்வொரு ரைபோசோமும் இரண்டு துணைத் துகள்களைக் கொண்டுள்ளது - பெரியது மற்றும் சிறியது: இரண்டு துணைத் துகள்களிலும் புரத மூலக்கூறுகள் மற்றும் ஆர்என்ஏ மூலக்கூறுகள் உள்ளன.
ரைபோசோம்கள் பெரும்பாலும் சிஸ்டெர்ன்கள் மற்றும் வெசிகல்ஸ் - எண்டோபிளாஸ்மிக் ரெட்டிகுலம் (ER) எனப்படும் சிறப்பு சவ்வு அமைப்புடன் இணைக்கப்படுகின்றன; அதிக புரதத்தை உற்பத்தி செய்யும் உயிரணுக்களில், எண்டோபிளாஸ்மிக் ரெட்டிகுலம் பெரும்பாலும் நன்கு வளர்ச்சியடைந்து ரைபோசோம்களால் மூடப்பட்டிருக்கும். சில நொதிகள் சவ்வுடன் இணைக்கப்பட்டிருந்தால் மட்டுமே பயனுள்ளதாக இருக்கும். லிப்பிட் தொகுப்பில் ஈடுபட்டுள்ள பெரும்பாலான நொதிகள் இங்கு அமைந்துள்ளன. எனவே, எண்டோபிளாஸ்மிக் ரெட்டிகுலம் ஒரு வகையான செல் வேலைப்பெட்டி போன்றது.
கூடுதலாக, ER சைட்டோபிளாஸை தனித்தனி பெட்டிகளாகப் பிரிக்கிறது, அதாவது, சைட்டோபிளாஸில் ஒரே நேரத்தில் நிகழும் பல்வேறு இரசாயன செயல்முறைகளைப் பிரிக்கிறது, மேலும் இந்த செயல்முறைகள் ஒன்றோடொன்று குறுக்கிடுவதற்கான வாய்ப்பைக் குறைக்கிறது.
கொடுக்கப்பட்ட கலத்தால் உற்பத்தி செய்யப்படும் பொருட்கள் பெரும்பாலும் கலத்திற்கு வெளியே பயன்படுத்தப்படுகின்றன. இதுபோன்ற சந்தர்ப்பங்களில், ரைபோசோம்களில் தொகுக்கப்பட்ட புரதங்கள் எண்டோபிளாஸ்மிக் ரெட்டிகுலத்தின் சவ்வுகளின் வழியாகச் சென்று அவற்றைச் சுற்றி உருவாகும் சவ்வு வெசிகல்களில் தொகுக்கப்படுகின்றன, பின்னர் அவை ER இலிருந்து பிரிக்கப்படுகின்றன. இந்த வெசிகல்ஸ், அடுக்கப்பட்ட அப்பத்தைப் போல, தட்டையான மற்றும் ஒன்றன் மேல் ஒன்றாக அடுக்கி, கோல்கி வளாகம் அல்லது கோல்கி கருவி எனப்படும் ஒரு சிறப்பியல்பு கட்டமைப்பை உருவாக்குகின்றன. கோல்கி கருவியில் தங்கியிருக்கும் போது, புரதங்கள் சில மாற்றங்களுக்கு உட்படுகின்றன. அவர்கள் செல்லை விட்டு வெளியேறும் நேரம் வரும்போது, சவ்வு வெசிகிள்கள் செல் சவ்வுடன் ஒன்றிணைந்து காலியாகி, அவற்றின் உள்ளடக்கங்களை வெளியே கொட்டுகிறது, அதாவது, சுரப்பு எக்சோசைடோசிஸ் மூலம் ஏற்படுகிறது.
கோல்கி எந்திரம் லைசோசோம்களையும் உற்பத்தி செய்கிறது - செரிமான நொதிகளைக் கொண்ட சவ்வுப் பைகள். ஒரு செல் எவ்வாறு சில புரதங்களை உருவாக்குகிறது, பேக்கேஜ் செய்கிறது மற்றும் ஏற்றுமதி செய்கிறது மற்றும் எந்த புரதங்களைத் தனக்குத்தானே வைத்திருக்க வேண்டும் என்பதை அது எவ்வாறு "தெரிகிறது" என்பதைக் கண்டுபிடிப்பது நவீன சைட்டாலஜியின் மிகவும் கவர்ச்சிகரமான கிளைகளில் ஒன்றாகும்.
எந்த செல்லின் சவ்வுகளும் தொடர்ந்து நகர்ந்து மாறிக்கொண்டே இருக்கும். ER சவ்வுகள் செல் முழுவதும் மெதுவாக நகரும். இந்த சவ்வுகளின் தனிப்பட்ட பிரிவுகள் தனித்தனியாக பிரிக்கப்பட்டு வெசிகிள்களை உருவாக்குகின்றன, அவை தற்காலிகமாக கோல்கி கருவியின் ஒரு பகுதியாக மாறும், பின்னர், எக்சோசைடோசிஸ் செயல்முறை மூலம், செல் சவ்வுடன் ஒன்றிணைகின்றன.
பின்னர், சவ்வு பொருள் சைட்டோபிளாஸத்திற்குத் திரும்புகிறது, அங்கு அது மீண்டும் பயன்படுத்தப்படுகிறது.
கலத்திற்குள் நுழையும் அல்லது வெளியில் வெளியிடப்படும் பொருட்களின் பரிமாற்றம், அத்துடன் மைக்ரோ மற்றும் மேக்ரோ சூழலுடன் பல்வேறு சமிக்ஞைகளின் பரிமாற்றம், கலத்தின் வெளிப்புற சவ்வு வழியாக நிகழ்கிறது. அறியப்பட்டபடி, உயிரணு சவ்வு என்பது ஒரு லிப்பிட் பைலேயர் ஆகும், அதில் பல்வேறு புரத மூலக்கூறுகள் உட்பொதிக்கப்பட்டுள்ளன, அவை சிறப்பு ஏற்பிகள், அயன் சேனல்கள், பல்வேறு இரசாயனங்கள், இடைச்செல்லுலார் தொடர்புகள் போன்றவற்றை தீவிரமாக கொண்டு செல்லும் அல்லது அகற்றும் சாதனங்களாக செயல்படுகின்றன. ஆரோக்கியமான யூகாரியோடிக் செல்களில், பாஸ்போலிப்பிட்கள் விநியோகிக்கப்படுகின்றன. சவ்வு சமச்சீரற்றது: வெளிப்புற மேற்பரப்பு ஸ்பிங்கோமைலின் மற்றும் பாஸ்பாடிடைல்கோலின், உள் மேற்பரப்பு - பாஸ்பாடிடைல்செரின் மற்றும் பாஸ்பாடிடைலெத்தனோலமைன் ஆகியவற்றைக் கொண்டுள்ளது. அத்தகைய சமச்சீரற்ற தன்மையை பராமரிக்க ஆற்றல் செலவு தேவைப்படுகிறது. எனவே, உயிரணு சேதம், தொற்று அல்லது ஆற்றல் பட்டினி ஏற்பட்டால், மென்படலத்தின் வெளிப்புற மேற்பரப்பு பாஸ்போலிப்பிட்களால் செறிவூட்டப்படுகிறது, இது அசாதாரணமானது, இது மற்ற செல்கள் மற்றும் என்சைம்களுக்கு ஒரு சமிக்ஞையாக மாறும். பாஸ்போலிபேஸ் A2 இன் கரையக்கூடிய வடிவத்தால் மிக முக்கியமான பங்கு வகிக்கப்படுகிறது, இது அராச்சிடோனிக் அமிலத்தை உடைத்து மேலே குறிப்பிட்ட பாஸ்போலிப்பிட்களிலிருந்து லைசோஃபார்ம்களை உருவாக்குகிறது. அராச்சிடோனிக் அமிலம் ஈகோசனாய்டுகள் போன்ற அழற்சி மத்தியஸ்தர்களை உருவாக்குவதற்கான கட்டுப்படுத்தும் இணைப்பாகும், மேலும் பாதுகாப்பு மூலக்கூறுகள் - பென்ட்ராக்ஸின்கள் (சி-ரியாக்டிவ் புரதம் (சிஆர்பி), அமிலாய்டு புரதங்களின் முன்னோடிகள்) - சவ்வில் உள்ள லைசோஃபார்ம்களுடன் இணைக்கப்படுகின்றன, அதைத் தொடர்ந்து நிரப்பு செயல்படுத்தப்படுகிறது. கிளாசிக்கல் பாதை மற்றும் செல் அழிவுடன் கூடிய அமைப்பு.
மென்படலத்தின் அமைப்பு கலத்தின் உள் சூழலின் பண்புகள், வெளிப்புற சூழலில் இருந்து அதன் வேறுபாடுகளை பாதுகாக்க உதவுகிறது. செல் சவ்வின் தேர்ந்தெடுக்கப்பட்ட ஊடுருவல் மற்றும் அதில் செயலில் போக்குவரத்து வழிமுறைகள் இருப்பதால் இது உறுதி செய்யப்படுகிறது. நேரடி சேதத்தின் விளைவாக அவற்றின் சீர்குலைவு, எடுத்துக்காட்டாக, டெட்ரோடோடாக்சின், ஓவாபைன், டெட்ராஎதிலாமோனியம் அல்லது தொடர்புடைய “பம்ப்களுக்கு” போதுமான ஆற்றல் வழங்கப்படாத நிலையில், கலத்தின் எலக்ட்ரோலைட் கலவையின் இடையூறு, அதன் வளர்சிதை மாற்றத்தில் ஏற்படும் மாற்றங்கள், சீர்குலைவு. குறிப்பிட்ட செயல்பாடுகளின் - சுருக்கம், தூண்டுதல் தூண்டுதல்களின் கடத்தல், முதலியன. மனிதர்களில் செல்லுலார் அயனி சேனல்களின் (கால்சியம், சோடியம், பொட்டாசியம் மற்றும் குளோரைடு) இடையூறுகள் இந்த சேனல்களின் கட்டமைப்பிற்கு காரணமான மரபணுக்களில் உள்ள பிறழ்வுகளால் மரபணு ரீதியாக தீர்மானிக்கப்படலாம். சேனல்லோபதிகள் என்று அழைக்கப்படுபவை நரம்பு, தசை மற்றும் செரிமான அமைப்புகளின் பரம்பரை நோய்களை ஏற்படுத்துகின்றன. கலத்திற்குள் அதிகப்படியான நீர் நுழைவது அதன் சிதைவுக்கு வழிவகுக்கும் - சைட்டோலிசிஸ் - நிரப்பு செயல்படுத்தப்படும்போது சவ்வு துளையிடுதல் அல்லது சைட்டோடாக்ஸிக் லிம்போசைட்டுகள் மற்றும் இயற்கை கொலையாளி செல்கள் தாக்குதலால்.
உயிரணு சவ்வு பல ஏற்பிகளைக் கொண்டுள்ளது - கட்டமைப்புகள், தொடர்புடைய குறிப்பிட்ட சமிக்ஞை மூலக்கூறுகளுடன் (லிகண்ட்ஸ்) இணைந்தால், கலத்திற்குள் ஒரு சமிக்ஞையை அனுப்பும். இது நொதியாக செயல்படும் மூலக்கூறுகளைக் கொண்ட பல்வேறு ஒழுங்குமுறை அடுக்குகள் மூலம் நிகழ்கிறது, அவை தொடர்ச்சியாக செயல்படுத்தப்பட்டு, வளர்ச்சி மற்றும் பெருக்கம், வேறுபாடு, இயக்கம், முதுமை மற்றும் உயிரணு இறப்பு போன்ற பல்வேறு செல்லுலார் திட்டங்களை செயல்படுத்த பங்களிக்கின்றன. ஒழுங்குமுறை அடுக்குகள் பல உள்ளன, ஆனால் அவற்றின் எண்ணிக்கை இன்னும் முழுமையாக தீர்மானிக்கப்படவில்லை. அவற்றுடன் தொடர்புடைய ஏற்பிகள் மற்றும் ஒழுங்குமுறை அடுக்குகளின் அமைப்பும் செல்லுக்குள் உள்ளது; அவை செறிவு, விநியோகம் மற்றும் செல்லின் செயல்பாட்டு நிலை, அதன் வளர்ச்சியின் நிலை மற்றும் பிற ஏற்பிகளிலிருந்து வரும் சமிக்ஞைகளின் ஒரே நேரத்தில் செயல்படும் நிலை ஆகியவற்றைப் பொறுத்து மேலும் சமிக்ஞை பாதையின் தேர்வு புள்ளிகளுடன் ஒரு குறிப்பிட்ட ஒழுங்குமுறை வலையமைப்பை உருவாக்குகின்றன. இதன் விளைவாக சிக்னலைத் தடுப்பது அல்லது வலுப்படுத்துவது, வேறு ஒழுங்குமுறை பாதையில் அதை இயக்குவது. ரிசெப்டர் எந்திரம் மற்றும் சிக்னல் டிரான்ஸ்டக்ஷன் பாதைகள் ரெகுலேட்டரி கேஸ்கேட்கள் வழியாக, எடுத்துக்காட்டாக, உட்கருவுக்கு, மரபணுக் குறைபாட்டின் விளைவாக, உயிரின மட்டத்தில் பிறவி குறைபாடு அல்லது ஒரு குறிப்பிட்ட செல் வகையின் உடலியல் பிறழ்வு காரணமாக சீர்குலைக்கப்படலாம். இந்த வழிமுறைகள் தொற்று முகவர்கள், நச்சுகள் ஆகியவற்றால் சேதமடையலாம், மேலும் வயதான செயல்பாட்டின் போது மாறலாம். இதன் இறுதிக் கட்டம், கலத்தின் செயல்பாடுகள், அதன் பெருக்கம் மற்றும் வேறுபாட்டின் செயல்முறைகளின் இடையூறாக இருக்கலாம்.
உயிரணுக்களின் மேற்பரப்பில் உயிரணுக்களுக்கு இடையேயான தொடர்பு செயல்முறைகளில் முக்கிய பங்கு வகிக்கும் மூலக்கூறுகளும் உள்ளன. இவற்றில் செல் ஒட்டுதல் புரதங்கள், ஹிஸ்டோகாம்பேடிபிலிட்டி ஆன்டிஜென்கள், திசு-குறிப்பிட்ட, வேறுபடுத்தும் ஆன்டிஜென்கள் போன்றவை அடங்கும். இந்த மூலக்கூறுகளின் கலவையில் ஏற்படும் மாற்றங்கள், செல்களுக்கு இடையேயான இடைவினைகளை சீர்குலைக்கும் மற்றும் அத்தகைய செல்களை அகற்றுவதற்கான பொருத்தமான வழிமுறைகளை செயல்படுத்துவதற்கு காரணமாக இருக்கலாம். நோய்த்தொற்றின் நீர்த்தேக்கமாக, குறிப்பாக வைரஸ், அல்லது கட்டி வளர்ச்சியின் சாத்தியமான துவக்கியாக உடலின் ஒருமைப்பாட்டிற்கு சில ஆபத்து.
கலத்தின் ஆற்றல் வழங்கல் மீறல்
கலத்தில் உள்ள ஆற்றலின் ஆதாரம் உணவு, அதன் முறிவுக்குப் பிறகு ஆற்றல் இறுதிப் பொருட்களாக வெளியிடப்படுகிறது. ஆற்றல் உற்பத்தியின் முக்கிய இடம் மைட்டோகாண்ட்ரியா ஆகும், இதில் சுவாச சங்கிலியின் என்சைம்களின் உதவியுடன் பொருட்கள் ஆக்ஸிஜனேற்றப்படுகின்றன. ஆக்ஸிஜனேற்றம் ஆற்றலின் முக்கிய சப்ளையர் ஆகும், ஏனெனில் கிளைகோலிசிஸின் விளைவாக, ஆக்சிஜனேற்றத்துடன் ஒப்பிடும்போது, அதே அளவு ஆக்சிஜனேற்ற அடி மூலக்கூறுகளிலிருந்து (குளுக்கோஸ்) 5% க்கும் அதிகமான ஆற்றல் வெளியிடப்படுவதில்லை. ஆக்ஸிஜனேற்றத்தின் போது வெளியிடப்படும் ஆற்றலில் சுமார் 60% உயர் ஆற்றல் பாஸ்பேட்டுகளில் (ATP, கிரியேட்டின் பாஸ்பேட்) ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷன் மூலம் திரட்டப்படுகிறது, மீதமுள்ளவை வெப்பமாக சிதறடிக்கப்படுகின்றன. பின்னர், உயர் ஆற்றல் பாஸ்பேட்டுகள் பம்ப் செயல்பாடு, தொகுப்பு, பிரிவு, இயக்கம், சுரப்பு போன்ற செயல்முறைகளுக்கு செல்லால் பயன்படுத்தப்படுகின்றன. மூன்று வழிமுறைகள் உள்ளன, இவற்றின் சேதம் செல்லின் ஆற்றல் விநியோகத்தில் இடையூறு ஏற்படுத்தும்: முதலாவது ஆற்றல் வளர்சிதை மாற்ற நொதிகளின் தொகுப்புக்கான வழிமுறை, இரண்டாவது ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷன் பொறிமுறையாகும், மூன்றாவது ஆற்றல் பயன்பாட்டின் பொறிமுறையாகும்.
மைட்டோகாண்ட்ரியல் சுவாசச் சங்கிலியில் எலக்ட்ரான் போக்குவரத்தை சீர்குலைத்தல் அல்லது ஏடிபி ஆக்சிஜனேற்றம் மற்றும் பாஸ்போரிலேஷனை துண்டித்தல், புரோட்டான் திறனை இழப்பது, ஏடிபி உற்பத்திக்கான உந்து சக்தி, ஆக்ஸிஜனேற்ற பாஸ்போரிலேஷன் பலவீனமடைவதற்கு வழிவகுக்கிறது, இதனால் ஆற்றலின் பெரும்பகுதி வெப்பமாக சிதறடிக்கப்படுகிறது. உயர் ஆற்றல் சேர்மங்களின் எண்ணிக்கை குறைகிறது. அட்ரினலின் செல்வாக்கின் கீழ் ஆக்சிஜனேற்றம் மற்றும் பாஸ்போரிலேஷன் ஆகியவை ஹோமியோதெர்மிக் உயிரினங்களின் செல்களால் வெப்ப உற்பத்தியை அதிகரிக்க பயன்படுத்தப்படுகின்றன, அதே நேரத்தில் குளிர்ச்சியின் போது நிலையான உடல் வெப்பநிலையை பராமரிக்கின்றன அல்லது காய்ச்சலின் போது அதை அதிகரிக்கின்றன. மைட்டோகாண்ட்ரியல் அமைப்பு மற்றும் ஆற்றல் வளர்சிதை மாற்றத்தில் குறிப்பிடத்தக்க மாற்றங்கள் தைரோடாக்சிகோசிஸில் காணப்படுகின்றன. இந்த மாற்றங்கள் ஆரம்பத்தில் மீளக்கூடியவை, ஆனால் ஒரு குறிப்பிட்ட கட்டத்திற்குப் பிறகு அவை மீள முடியாததாக மாறும்: மைட்டோகாண்ட்ரியா துண்டு, சிதைவு அல்லது வீக்கம், கிறிஸ்டேயை இழந்து, வெற்றிடங்களாக மாறும், இறுதியில் ஹைலின், ஃபெரிடின், கால்சியம், லிபோஃபுசின் போன்ற பொருட்களைக் குவிக்கும். ஸ்கர்வி நோயாளிகளில், மைட்டோகாண்ட்ரியா உருகி காண்டிரியோஸ்பியர்களை உருவாக்குகிறது, ஒருவேளை பெராக்சைடு சேர்மங்களால் சவ்வு சேதம் ஏற்படலாம். மைட்டோகாண்ட்ரியாவுக்கு குறிப்பிடத்தக்க சேதம் ஒரு சாதாரண உயிரணுவை வீரியம் மிக்க ஒன்றாக மாற்றும் போது அயனியாக்கும் கதிர்வீச்சின் செல்வாக்கின் கீழ் ஏற்படுகிறது.
மைட்டோகாண்ட்ரியா என்பது கால்சியம் அயனிகளின் ஒரு சக்திவாய்ந்த டிப்போ ஆகும், அங்கு அதன் செறிவு சைட்டோபிளாஸில் உள்ளதை விட பல ஆர்டர்கள் அதிகமாக உள்ளது. மைட்டோகாண்ட்ரியா சேதமடையும் போது, கால்சியம் சைட்டோபிளாஸத்தில் நுழைகிறது, இது உள்செல்லுலார் கட்டமைப்புகளுக்கு சேதம் விளைவிக்கும் மற்றும் தொடர்புடைய உயிரணுக்களின் செயல்பாடுகளை சீர்குலைக்கும், எடுத்துக்காட்டாக, கால்சியம் சுருக்கங்கள் அல்லது நியூரான்களில் "கால்சியம் மரணம்". மைட்டோகாண்ட்ரியாவின் செயல்பாட்டுத் திறனை சீர்குலைப்பதன் விளைவாக, ஃப்ரீ ரேடிக்கல் பெராக்சைடு சேர்மங்களின் உருவாக்கம் கூர்மையாக அதிகரிக்கிறது, இது மிக அதிக வினைத்திறனைக் கொண்டுள்ளது, எனவே உயிரணுவின் முக்கிய கூறுகளை சேதப்படுத்துகிறது - நியூக்ளிக் அமிலங்கள், புரதங்கள் மற்றும் லிப்பிடுகள். இந்த நிகழ்வு ஆக்ஸிஜனேற்ற அழுத்தம் என்று அழைக்கப்படும் போது கவனிக்கப்படுகிறது மற்றும் செல் இருப்புக்கு எதிர்மறையான விளைவுகளை ஏற்படுத்தும். எனவே, மைட்டோகாண்ட்ரியாவின் வெளிப்புற சவ்வு சேதமடைவதால், இடைச்சவ்வு இடைவெளியில் உள்ள பொருட்களின் சைட்டோபிளாஸில் வெளியிடப்படுகிறது, முதன்மையாக சைட்டோக்ரோம் சி மற்றும் பிற உயிரியல் ரீதியாக செயல்படும் பொருட்கள், இது திட்டமிடப்பட்ட உயிரணு இறப்பை ஏற்படுத்தும் சங்கிலி எதிர்வினைகளைத் தூண்டுகிறது - அப்போப்டொசிஸ். மைட்டோகாண்ட்ரிய டிஎன்ஏவை சேதப்படுத்துவதன் மூலம், ஃப்ரீ ரேடிக்கல் எதிர்வினைகள் சில சுவாச சங்கிலி நொதிகளை உருவாக்குவதற்குத் தேவையான மரபணு தகவலை சிதைக்கின்றன, அவை குறிப்பாக மைட்டோகாண்ட்ரியாவில் உற்பத்தி செய்யப்படுகின்றன. இது ஆக்ஸிஜனேற்ற செயல்முறைகளில் இன்னும் பெரிய இடையூறுக்கு வழிவகுக்கிறது. பொதுவாக, மைட்டோகாண்ட்ரியாவின் சொந்த மரபணு கருவி, கருவின் மரபணு கருவியுடன் ஒப்பிடுகையில், அதில் குறியிடப்பட்ட மரபணு தகவல்களை மாற்றக்கூடிய தீங்கு விளைவிக்கும் தாக்கங்களிலிருந்து குறைவாகவே பாதுகாக்கப்படுகிறது. இதன் விளைவாக, மைட்டோகாண்ட்ரியாவின் செயலிழப்பு வாழ்நாள் முழுவதும் ஏற்படுகிறது, எடுத்துக்காட்டாக, வயதான செயல்பாட்டின் போது, உயிரணுவின் வீரியம் மிக்க மாற்றத்தின் போது, அத்துடன் முட்டையில் உள்ள மைட்டோகாண்ட்ரியல் டிஎன்ஏவின் பிறழ்வுடன் தொடர்புடைய பரம்பரை மைட்டோகாண்ட்ரியல் நோய்களின் பின்னணிக்கு எதிராக. தற்போது, 50 க்கும் மேற்பட்ட மைட்டோகாண்ட்ரியல் பிறழ்வுகள் விவரிக்கப்பட்டுள்ளன, அவை நரம்பு மற்றும் தசை அமைப்புகளின் பரம்பரை சீரழிவு நோய்களை ஏற்படுத்துகின்றன. அவை தாயிடமிருந்து பிரத்தியேகமாக குழந்தைக்கு பரவுகின்றன, ஏனெனில் விந்தணுவின் மைட்டோகாண்ட்ரியா ஜிகோட்டின் பகுதியாக இல்லை, அதன்படி, புதிய உயிரினம்.
மரபணு தகவல்களின் பாதுகாப்பு மற்றும் பரிமாற்றத்தின் மீறல்
செல் கருவானது பெரும்பாலான மரபணு தகவல்களைக் கொண்டுள்ளது மற்றும் அதன் இயல்பான செயல்பாட்டை உறுதி செய்கிறது. தேர்ந்தெடுக்கப்பட்ட மரபணு வெளிப்பாடு மூலம், இது இடைநிலையின் போது செல் செயல்பாட்டை ஒருங்கிணைக்கிறது, மரபணு தகவலைச் சேமிக்கிறது மற்றும் உயிரணுப் பிரிவின் போது மரபணுப் பொருளை மீண்டும் உருவாக்குகிறது மற்றும் கடத்துகிறது. டிஎன்ஏ பிரதி மற்றும் ஆர்என்ஏ படியெடுத்தல் கருவில் நிகழ்கிறது. புற ஊதா மற்றும் அயனியாக்கும் கதிர்வீச்சு, ஃப்ரீ ரேடிக்கல் ஆக்சிஜனேற்றம், இரசாயனங்கள், வைரஸ்கள் போன்ற பல்வேறு நோய்க்கிருமி காரணிகள் டிஎன்ஏவை சேதப்படுத்தும். சூடான இரத்தம் கொண்ட விலங்கின் ஒவ்வொரு உயிரணுவும் 1 நாள் எடுக்கும் என்று மதிப்பிடப்பட்டுள்ளது. 10,000 க்கும் மேற்பட்ட தளங்களை இழக்கிறது. பிரிவின் போது நகலெடுக்கும் போது மீறல்களைச் சேர்க்க வேண்டும். இந்த சேதங்கள் தொடர்ந்தால், செல் உயிர்வாழ முடியாது. டிஎன்ஏ சேதத்தை மாற்றியமைக்கும் புற ஊதா எண்டோநியூக்லீஸ், ரிப்பேர் ரெப்ளிகேஷன் மற்றும் மறுசீரமைப்பு பழுதுபார்க்கும் அமைப்புகள் போன்ற சக்திவாய்ந்த பழுதுபார்க்கும் அமைப்புகளின் இருப்பில் பாதுகாப்பு உள்ளது. பழுதுபார்க்கும் அமைப்புகளில் உள்ள மரபணு குறைபாடுகள் டிஎன்ஏவை சேதப்படுத்தும் காரணிகளுக்கு அதிகரித்த உணர்திறன் காரணமாக ஏற்படும் நோய்களின் வளர்ச்சியை ஏற்படுத்துகின்றன. இது xeroderma pigmentosum, அத்துடன் சில துரிதப்படுத்தப்பட்ட வயதான நோய்க்குறிகள், வீரியம் மிக்க கட்டிகளை உருவாக்குவதற்கான அதிகரித்த போக்குடன் சேர்ந்து.
டிஎன்ஏ ரெப்ளிகேஷன், மெசஞ்சர் ஆர்என்ஏ (எம்ஆர்என்ஏ) படியெடுத்தல் மற்றும் நியூக்ளிக் அமிலங்களிலிருந்து மரபணு தகவல்களை புரதங்களின் கட்டமைப்பிற்கு மொழிபெயர்த்தல் ஆகியவற்றின் செயல்முறைகளை ஒழுங்குபடுத்தும் அமைப்பு மிகவும் சிக்கலானது மற்றும் பல-நிலையானது. 3000 க்கும் மேற்பட்ட மொத்த எண்ணிக்கையிலான டிரான்ஸ்கிரிப்ஷன் காரணிகளின் செயல்பாட்டைத் தூண்டும் ஒழுங்குமுறை அடுக்குகளுக்கு கூடுதலாக, சில மரபணுக்களை செயல்படுத்துகிறது, சிறிய ஆர்என்ஏ மூலக்கூறுகளால் (ஆர்என்ஏ; ஆர்என்ஏஐ குறுக்கிடுகிறது) மத்தியஸ்தம் செய்யும் பல-நிலை ஒழுங்குமுறை அமைப்பும் உள்ளது. தோராயமாக 3 பில்லியன் பியூரின் மற்றும் பைரிமிடின் அடிப்படைகளைக் கொண்ட மனித மரபணு, புரதத் தொகுப்புக்குக் காரணமான கட்டமைப்பு மரபணுக்களில் 2% மட்டுமே உள்ளது. மீதமுள்ளவை ஒழுங்குமுறை ஆர்என்ஏக்களின் தொகுப்பை வழங்குகின்றன, அவை டிரான்ஸ்கிரிப்ஷன் காரணிகளுடன் ஒரே நேரத்தில், டிஎன்ஏ அளவில் உள்ள கட்டமைப்பு மரபணுக்களின் வேலையை குரோமோசோம்களில் செயல்படுத்துகின்றன அல்லது தடுக்கின்றன அல்லது பாலிபெப்டைட் மூலக்கூறு உருவாகும் போது மெசஞ்சர் ஆர்என்ஏ (எம்ஆர்என்ஏ) மொழிபெயர்ப்பு செயல்முறைகளை பாதிக்கின்றன. சைட்டோபிளாசம். மரபணு தகவல்களின் மீறல் கட்டமைப்பு மரபணுக்களின் மட்டத்திலும், டிஎன்ஏவின் ஒழுங்குமுறைப் பகுதியிலும் பல்வேறு பரம்பரை நோய்களின் வடிவத்தில் தொடர்புடைய வெளிப்பாடுகளுடன் ஏற்படலாம்.
சமீபத்தில், ஒரு உயிரினத்தின் தனிப்பட்ட வளர்ச்சியின் போது ஏற்படும் மரபணுப் பொருட்களில் ஏற்படும் மாற்றங்கள் மற்றும் அவற்றின் மெத்திலேஷன், அசிடைலேஷன் மற்றும் பாஸ்போரிலேஷன் காரணமாக டிஎன்ஏ மற்றும் குரோமோசோம்களின் சில பிரிவுகளைத் தடுப்பது அல்லது செயல்படுத்துவது ஆகியவற்றுடன் அதிக கவனம் ஈர்க்கப்பட்டுள்ளது. இந்த மாற்றங்கள் நீண்ட காலத்திற்கு நீடிக்கின்றன, சில சமயங்களில் கரு உருவாக்கம் முதல் முதுமை வரை உயிரினத்தின் முழு வாழ்நாள் முழுவதும், மேலும் அவை எபிஜெனோமிக் பரம்பரை என்று அழைக்கப்படுகின்றன.
மைட்டோடிக் சுழற்சியைக் கட்டுப்படுத்தும் அமைப்புகளால் (காரணிகள்) மாற்றப்பட்ட மரபணுத் தகவலுடன் செல்களின் பெருக்கம் தடுக்கப்படுகிறது. அவை சைக்ளின் சார்ந்த புரோட்டீன் கைனேஸ்கள் மற்றும் அவற்றின் வினையூக்கி துணைக்குழுக்கள் - சைக்ளின்களுடன் தொடர்பு கொள்கின்றன, மேலும் உயிரணு முழு மைட்டோடிக் சுழற்சியில் செல்வதைத் தடுக்கின்றன, டிஎன்ஏ பழுது முடிவடையும் வரை ப்ரீசிந்தெடிக் மற்றும் செயற்கை கட்டங்களுக்கு (G1/S பிளாக்) இடையேயான எல்லையில் பிரிவை நிறுத்துகின்றன. இது சாத்தியமில்லாத பட்சத்தில், அவை திட்டமிடப்பட்ட இறப்பு செல்களைத் தொடங்குகின்றன. இந்த காரணிகளில் p53 மரபணு அடங்கும், இதன் பிறழ்வு மாற்றப்பட்ட உயிரணுக்களின் பெருக்கத்தின் மீதான கட்டுப்பாட்டை இழக்கிறது; இது கிட்டத்தட்ட 50% மனித புற்றுநோய்களில் காணப்படுகிறது. மைட்டோடிக் சுழற்சியின் இரண்டாவது சோதனைச் சாவடி G2/M எல்லையில் உள்ளது. இங்கே, மைட்டோசிஸ் அல்லது ஒடுக்கற்பிரிவில் உள்ள மகள் செல்களுக்கு இடையே குரோமோசோமால் பொருள் சரியான விநியோகம் செல் சுழல், மையம் மற்றும் சென்ட்ரோமியர்ஸ் (கினெட்டோகோர்ஸ்) ஆகியவற்றைக் கட்டுப்படுத்தும் வழிமுறைகளின் தொகுப்பைப் பயன்படுத்தி கட்டுப்படுத்தப்படுகிறது. இந்த வழிமுறைகளின் பயனற்ற தன்மை குரோமோசோம்கள் அல்லது அவற்றின் பாகங்களின் விநியோகத்தை சீர்குலைக்க வழிவகுக்கிறது, இது மகள் உயிரணுக்களில் ஒன்றில் குரோமோசோம் இல்லாதது (அனீப்ளோயிடி), கூடுதல் குரோமோசோம் (பாலிப்ளோயிடி), பிரித்தல் ஆகியவற்றால் வெளிப்படுகிறது. ஒரு குரோமோசோமின் ஒரு பகுதி (நீக்குதல்) மற்றும் அதை மற்றொரு குரோமோசோமுக்கு மாற்றுதல் (இடமாற்றம்) . இத்தகைய செயல்முறைகள் வீரியம் மிக்க சிதைந்த மற்றும் மாற்றப்பட்ட உயிரணுக்களின் பெருக்கத்தின் போது அடிக்கடி காணப்படுகின்றன. கிருமி உயிரணுக்களுடன் ஒடுக்கற்பிரிவின் போது இது நடந்தால், இது கரு வளர்ச்சியின் ஆரம்ப கட்டத்தில் கருவின் மரணத்திற்கு அல்லது குரோமோசோமால் நோயுடன் ஒரு உயிரினத்தின் பிறப்புக்கு வழிவகுக்கிறது.
கட்டி வளர்ச்சியின் போது கட்டுப்பாடற்ற உயிரணு பெருக்கம், உயிரணு பெருக்கத்தைக் கட்டுப்படுத்தும் மரபணுக்களில் ஏற்படும் பிறழ்வுகளின் விளைவாக ஏற்படுகிறது மற்றும் அவை புற்றுநோய்கள் என்று அழைக்கப்படுகின்றன. தற்போது அறியப்பட்ட 70 க்கும் மேற்பட்ட புற்றுநோய்களில், பெரும்பாலானவை உயிரணு வளர்ச்சி ஒழுங்குமுறையின் கூறுகளைச் சேர்ந்தவை, சில மரபணு செயல்பாட்டைக் கட்டுப்படுத்தும் டிரான்ஸ்கிரிப்ஷன் காரணிகள் மற்றும் செல் பிரிவு மற்றும் வளர்ச்சியைத் தடுக்கும் காரணிகளால் குறிப்பிடப்படுகின்றன. பெருகும் உயிரணுக்களின் அதிகப்படியான விரிவாக்கத்தை (பரவலை) கட்டுப்படுத்தும் மற்றொரு காரணி குரோமோசோம்களின் முனைகளைக் குறைப்பதாகும் - டெலோமியர்ஸ், முற்றிலும் ஸ்டெரிக் தொடர்புகளின் விளைவாக முழுமையாகப் பிரதிபலிக்க முடியாது, எனவே, ஒவ்வொரு செல் பிரிவுக்குப் பிறகு, டெலோமியர்ஸ் சுருக்கப்படுகிறது. தளங்களின் ஒரு குறிப்பிட்ட பகுதி. இவ்வாறு, குறிப்பிட்ட எண்ணிக்கையிலான பிரிவுகளுக்குப் பிறகு (வழக்கமாக உயிரினத்தின் வகை மற்றும் அதன் வயதைப் பொறுத்து 20 முதல் 100 வரை) ஒரு வயதுவந்த உயிரினத்தின் செல்கள் பெருகி டெலோமியர் நீளத்தை வெளியேற்றி, மேலும் குரோமோசோம் பிரதி எடுப்பது நிறுத்தப்படும். டெலோமரேஸ் என்ற நொதி இருப்பதால் இந்த நிகழ்வு விந்தணு எபிட்டிலியம், என்டோரோசைட்டுகள் மற்றும் கரு உயிரணுக்களில் ஏற்படாது, இது ஒவ்வொரு பிரிவிற்குப் பிறகும் டெலோமியர் நீளத்தை மீட்டெடுக்கிறது. வயதுவந்த உயிரினங்களின் பெரும்பாலான உயிரணுக்களில், டெலோமரேஸ் தடுக்கப்படுகிறது, ஆனால், துரதிர்ஷ்டவசமாக, இது கட்டி உயிரணுக்களில் செயல்படுத்தப்படுகிறது.
கருவுக்கும் சைட்டோபிளாஸுக்கும் இடையிலான இணைப்பு மற்றும் இரு திசைகளிலும் உள்ள பொருட்களின் போக்குவரத்து ஆகியவை ஆற்றலை உட்கொள்ளும் சிறப்பு போக்குவரத்து அமைப்புகளின் பங்கேற்புடன் அணு சவ்வில் உள்ள துளைகள் மூலம் மேற்கொள்ளப்படுகின்றன. இந்த வழியில், ஆற்றல் மற்றும் பிளாஸ்டிக் பொருட்கள், சமிக்ஞை மூலக்கூறுகள் (டிரான்ஸ்கிரிப்ஷன் காரணிகள்) கருவுக்கு கொண்டு செல்லப்படுகின்றன. தலைகீழ் ஓட்டம் எம்ஆர்என்ஏ மற்றும் பரிமாற்ற ஆர்என்ஏ (டிஆர்என்ஏ) ஆகியவற்றின் சைட்டோபிளாசம் மூலக்கூறுகளுக்குள் செல்கிறது, கலத்தில் புரதத் தொகுப்புக்குத் தேவையான ரைபோசோம்கள். பொருட்களின் போக்குவரத்தின் அதே பாதை வைரஸ்களிலும் இயல்பாகவே உள்ளது, குறிப்பாக எச்.ஐ.வி. புதிய வைரஸ் துகள்களின் புரோட்டீன்களை மேலும் ஒருங்கிணைக்க சைட்டோபிளாஸத்திற்கு புதிதாக உருவாக்கப்பட்ட வைரஸ் ஆர்என்ஏவை மாற்றுவதன் மூலம் அவை ஹோஸ்ட் ஜெனோமில் மேலும் ஒருங்கிணைக்கப்படுவதன் மூலம் அவற்றின் மரபணுப் பொருளை புரவலன் கலத்தின் கருவிற்கு மாற்றுகின்றன.
தொகுப்பு செயல்முறைகளின் மீறல்
புரோட்டீன் தொகுப்பு செயல்முறைகள் எண்டோபிளாஸ்மிக் ரெட்டிகுலத்தின் தொட்டிகளில் நிகழ்கின்றன, அணு சவ்வில் உள்ள துளைகளுடன் நெருக்கமாக இணைக்கப்பட்டுள்ளன, இதன் மூலம் ரைபோசோம்கள், டிஆர்என்ஏ மற்றும் எம்ஆர்என்ஏ ஆகியவை எண்டோபிளாஸ்மிக் ரெட்டிகுலத்தில் நுழைகின்றன. இங்கே, பாலிபெப்டைட் சங்கிலிகளின் தொகுப்பு மேற்கொள்ளப்படுகிறது, இது பின்னர் அவற்றின் இறுதி வடிவத்தை அக்ரானுலர் எண்டோபிளாஸ்மிக் ரெட்டிகுலம் மற்றும் லேமல்லர் வளாகத்தில் (கோல்கி காம்ப்ளக்ஸ்) பெறுகிறது, அங்கு அவை மொழிபெயர்ப்புக்கு பிந்தைய மாற்றத்திற்கு உட்பட்டு கார்போஹைட்ரேட் மற்றும் லிப்பிட் மூலக்கூறுகளுடன் இணைகின்றன. புதிதாக உருவாக்கப்பட்ட புரத மூலக்கூறுகள் தொகுப்பின் இடத்தில் இருக்காது, ஆனால் சிக்கலான ஒழுங்குபடுத்தப்பட்ட செயல்முறை மூலம் புரோட்டீன்கினேசிஸ், கலத்தின் தனிமைப்படுத்தப்பட்ட பகுதிக்கு தீவிரமாக மாற்றப்படுகின்றன, அங்கு அவை அவற்றின் நோக்கம் கொண்ட செயல்பாட்டைச் செய்யும். இந்த வழக்கில், ஒரு மிக முக்கியமான படி, மாற்றப்பட்ட மூலக்கூறை அதன் உள்ளார்ந்த செயல்பாட்டைச் செய்யக்கூடிய பொருத்தமான இடஞ்சார்ந்த உள்ளமைவாக கட்டமைப்பதாகும். இந்த கட்டமைப்பு சிறப்பு நொதிகளின் உதவியுடன் அல்லது சிறப்பு புரத மூலக்கூறுகளின் மேட்ரிக்ஸில் நிகழ்கிறது - சாப்பரோன்கள், புரத மூலக்கூறு, புதிதாக உருவாக்கப்பட்ட அல்லது வெளிப்புற செல்வாக்கின் காரணமாக மாற்றப்பட்டு, சரியான முப்பரிமாண கட்டமைப்பைப் பெற உதவுகிறது. உயிரணுவில் பாதகமான விளைவு ஏற்பட்டால், புரத மூலக்கூறுகளின் கட்டமைப்பை சீர்குலைக்கும் சாத்தியம் இருக்கும்போது (உதாரணமாக, உடல் வெப்பநிலை அதிகரிப்பு, ஒரு தொற்று செயல்முறை, போதை), கலத்தில் சேப்பரோன்களின் செறிவு அதிகரிக்கிறது. கூர்மையாக. எனவே, அத்தகைய மூலக்கூறுகள் என்றும் அழைக்கப்படுகின்றன அழுத்த புரதங்கள், அல்லது வெப்ப அதிர்ச்சி புரதங்கள். ஒரு புரத மூலக்கூறின் கட்டமைப்பை மீறுவது, அமிலாய்டோசிஸ், அல்சைமர் நோய் போன்றவற்றின் போது செல்லில் அல்லது அதற்கு வெளியே டெபாசிட் செய்யப்படும் வேதியியல் செயலற்ற கூட்டுத்தொகைகளை உருவாக்க வழிவகுக்கிறது. முதன்மை கட்டமைப்பு தவறாக நடந்தால், அனைத்து அடுத்தடுத்த மூலக்கூறுகளும் குறைபாடுடையதாக இருக்கும். இந்த நிலை ப்ரியான் நோய்கள் என்று அழைக்கப்படுபவற்றில் ஏற்படுகிறது (செம்மறியாடு, வெறிபிடித்த மாடுகள், குரு, மனிதர்களில் க்ரூட்ஸ்ஃபெல்ட்-ஜாகோப் நோய்), ஒரு நரம்பு உயிரணுவின் சவ்வு புரதங்களில் ஒன்றில் ஏற்படும் குறைபாடு, உயிரணுக்களுக்குள் மந்த வெகுஜனங்களைத் தொடர்ந்து குவிக்கும் போது. மற்றும் அதன் முக்கிய செயல்பாடுகளின் இடையூறு.
ஒரு கலத்தில் தொகுப்பு செயல்முறைகளின் சீர்குலைவு அதன் பல்வேறு நிலைகளில் நிகழலாம்: கருவில் உள்ள ஆர்என்ஏ படியெடுத்தல், ரைபோசோம்களில் பாலிபெப்டைட்களின் மொழிபெயர்ப்பு, மொழிபெயர்ப்புக்கு பிந்தைய மாற்றம், பீஜ் மூலக்கூறின் ஹைப்பர்மெதிலேஷன் மற்றும் கிளைகோசைலேஷன், கலத்தில் உள்ள புரதங்களின் போக்குவரத்து மற்றும் விநியோகம் மற்றும் அவற்றை அகற்றுதல். வெளியே. இந்த வழக்கில், ரைபோசோம்களின் எண்ணிக்கையில் அதிகரிப்பு அல்லது குறைதல், பாலிரிபோசோம்களின் முறிவு, சிறுமணி எண்டோபிளாஸ்மிக் ரெட்டிகுலத்தின் நீர்த்தேக்கங்களின் விரிவாக்கம், ரைபோசோம்களின் இழப்பு மற்றும் வெசிகல்கள் மற்றும் வெற்றிடங்களின் உருவாக்கம் ஆகியவற்றைக் காணலாம். இவ்வாறு, ஒரு வெளிறிய கிரேப் விஷத்தால், ஆர்என்ஏ பாலிமரேஸ் என்சைம் சேதமடைகிறது, இது டிரான்ஸ்கிரிப்ஷனை சீர்குலைக்கிறது. டிஃப்தீரியா நச்சு, நீட்டிப்பு காரணி செயலிழக்கச் செய்வதன் மூலம், மொழிபெயர்ப்பு செயல்முறைகளை சீர்குலைத்து, மாரடைப்பு சேதத்தை ஏற்படுத்துகிறது. சில குறிப்பிட்ட புரத மூலக்கூறுகளின் தொகுப்பின் இடையூறுக்கான காரணம் தொற்று முகவர்களாக இருக்கலாம். உதாரணமாக, ஹெர்பெஸ் வைரஸ்கள் MHC ஆன்டிஜென் மூலக்கூறுகளின் தொகுப்பு மற்றும் வெளிப்பாட்டைத் தடுக்கின்றன, அவை நோயெதிர்ப்பு கட்டுப்பாட்டை ஓரளவு தவிர்க்க அனுமதிக்கிறது - கடுமையான அழற்சியின் மத்தியஸ்தர்களின் தொகுப்பு; அசாதாரண புரதங்களின் தோற்றம் அவற்றின் மேலும் முறிவை நிறுத்தி மந்தமான அல்லது நச்சுப் பொருட்களின் திரட்சிக்கு வழிவகுக்கும். இது ஒரு குறிப்பிட்ட அளவிற்கு, சிதைவு செயல்முறைகளை சீர்குலைப்பதன் மூலம் எளிதாக்கப்படுகிறது.
சிதைவு செயல்முறைகளின் சீர்குலைவு
உயிரணுவில் புரதத்தின் தொகுப்புடன், அதன் முறிவு தொடர்ந்து நிகழ்கிறது. சாதாரண நிலைமைகளின் கீழ், இது முக்கியமான ஒழுங்குமுறை மற்றும் உருவாக்கும் முக்கியத்துவத்தைக் கொண்டுள்ளது, உதாரணமாக, நொதிகள், புரத ஹார்மோன்கள் மற்றும் மைட்டோடிக் சுழற்சி புரதங்களின் செயலற்ற வடிவங்களை செயல்படுத்தும் போது. இயல்பான உயிரணு வளர்ச்சி மற்றும் வளர்ச்சிக்கு புரதங்கள் மற்றும் உறுப்புகளின் தொகுப்பு மற்றும் சிதைவு ஆகியவற்றுக்கு இடையே நன்றாக கட்டுப்படுத்தப்பட்ட சமநிலை தேவைப்படுகிறது. இருப்பினும், புரதத் தொகுப்பின் செயல்பாட்டில், ஒருங்கிணைக்கும் கருவியின் செயல்பாட்டில் உள்ள பிழைகள், புரத மூலக்கூறின் அசாதாரண கட்டமைப்பு மற்றும் இரசாயன மற்றும் பாக்டீரியா முகவர்களால் அதன் சேதம் ஆகியவற்றால், ஒரு பெரிய எண்ணிக்கையிலான குறைபாடுள்ள மூலக்கூறுகள் தொடர்ந்து உருவாகின்றன. சில மதிப்பீடுகளின்படி, அவற்றின் பங்கு அனைத்து ஒருங்கிணைந்த புரதங்களில் மூன்றில் ஒரு பங்காகும்.
பாலூட்டிகளின் செல்கள் பல முக்கிய அம்சங்களைக் கொண்டுள்ளன புரத அழிவின் வழிகள்:லைசோசோமால் புரோட்டீஸ்கள் (பெண்டைட் ஹைட்ரோலேஸ்கள்), கால்சியம் சார்ந்த புரோட்டினேஸ்கள் (எண்டோபெப்டிடேஸ்கள்) மற்றும் புரோட்டீசோம் அமைப்பு மூலம். கூடுதலாக, காஸ்பேஸ்கள் போன்ற சிறப்பு புரோட்டினேஸ்களும் உள்ளன. யூகாரியோடிக் செல்களில் பொருட்களின் சிதைவு ஏற்படும் முக்கிய உறுப்பு லைசோசோம் ஆகும், இதில் ஏராளமான ஹைட்ரோலைடிக் என்சைம்கள் உள்ளன. லைசோசோம்கள் மற்றும் பாகோலிசோசோம்களில் உள்ள எண்டோசைட்டோசிஸ் மற்றும் பல்வேறு வகையான தன்னியக்க செயல்முறைகள் காரணமாக, குறைபாடுள்ள புரத மூலக்கூறுகள் மற்றும் முழு உறுப்புகளும் அழிக்கப்படுகின்றன: சேதமடைந்த மைட்டோகாண்ட்ரியா, பிளாஸ்மா மென்படலத்தின் பிரிவுகள், சில எக்ஸ்ட்ராசெல்லுலர் புரதங்கள் மற்றும் சுரக்கும் துகள்களின் உள்ளடக்கங்கள்.
புரோட்டீசோம் என்பது புரோட்டீசோம் ஆகும், இது சைட்டோசோல், நியூக்ளியஸ், எண்டோபிளாஸ்மிக் ரெட்டிகுலம் மற்றும் செல் சவ்வு ஆகியவற்றில் உள்ளமைக்கப்பட்ட சிக்கலான கட்டமைப்பின் மல்டிகேடலிடிக் புரோட்டினேஸ் அமைப்பு ஆகும். இந்த நொதி அமைப்பு சேதமடைந்த புரதங்கள் மற்றும் ஆரோக்கியமான புரதங்களை உடைப்பதற்கு பொறுப்பாகும், அவை சாதாரண செல் செயல்பாட்டிற்கு அகற்றப்பட வேண்டும். இந்த வழக்கில், அழிக்கப்பட வேண்டிய புரதங்கள் பூர்வாங்கமாக ஒரு குறிப்பிட்ட பாலிபெப்டைட், ubiquitin உடன் இணைக்கப்படுகின்றன. இருப்பினும், எங்கும் பரவாத புரதங்களும் புரோட்டீசோம்களில் ஓரளவு அழிக்கப்படலாம். புரோட்டீசோம்களில் உள்ள புரத மூலக்கூறுகள் குறுகிய பாலிபெப்டைடுகளாக (செயலாக்கம்) உடைந்து, அவற்றின் அடுத்தடுத்த விளக்கக்காட்சிகளுடன் சேர்ந்து வகை I MHC மூலக்கூறுகள் உடலில் உள்ள ஆன்டிஜெனிக் ஹோமியோஸ்டாசிஸின் நோயெதிர்ப்புக் கட்டுப்பாட்டில் ஒரு முக்கிய இணைப்பாகும். புரோட்டீசோம் செயல்பாடு பலவீனமடையும் போது, சேதமடைந்த மற்றும் தேவையற்ற புரதங்கள் குவிந்து, இது செல் வயதானவுடன் வருகிறது. சைக்ளின் சார்ந்த புரதங்களின் சிதைவின் மீறல் செல் பிரிவின் சீர்குலைவுக்கு வழிவகுக்கிறது, சுரக்கும் புரதங்களின் சிதைவு - சிஸ்டோபிப்ரோசிஸின் வளர்ச்சிக்கு. மாறாக, புரோட்டீசோம் செயல்பாட்டின் அதிகரிப்பு உடலின் குறைபாட்டுடன் (எய்ட்ஸ், புற்றுநோய்) ஏற்படுகிறது.
புரதச் சிதைவின் மரபணு ரீதியாக நிர்ணயிக்கப்பட்ட சீர்குலைவுகளுடன், உயிரினம் சாத்தியமானதாக இல்லை மற்றும் கரு வளர்ச்சியின் ஆரம்ப கட்டங்களில் இறக்கிறது. கொழுப்புகள் அல்லது கார்போஹைட்ரேட்டுகளின் முறிவு சீர்குலைந்தால், சேமிப்பு நோய்கள் (திசௌரிஸ்மோசிஸ்) ஏற்படுகின்றன. இந்த வழக்கில், சில பொருட்கள் அல்லது அவற்றின் முழுமையற்ற முறிவின் தயாரிப்புகளின் அதிகப்படியான அளவு - லிப்பிடுகள், பாலிசாக்கரைடுகள் - கலத்தின் உள்ளே குவிந்து, இது கலத்தின் செயல்பாட்டை கணிசமாக சேதப்படுத்துகிறது. இது பெரும்பாலும் கல்லீரல் எபிடெலியல் செல்கள் (ஹெபடோசைட்டுகள்), நியூரான்கள், ஃபைப்ரோபிளாஸ்ட்கள் மற்றும் மேக்ரோபாகோசைட்டுகளில் காணப்படுகிறது.
நோய்க்குறியியல் செயல்முறைகளின் விளைவாக (உதாரணமாக, புரதம், கொழுப்பு, கார்போஹைட்ரேட் மற்றும் நிறமி சிதைவு) மற்றும் அசாதாரண பொருட்களின் உருவாக்கத்துடன் சேர்ந்து, பொருட்களின் முறிவு செயல்முறைகளின் பெறப்பட்ட கோளாறுகள் ஏற்படலாம். லைசோசோமால் புரோட்டியோலிசிஸ் அமைப்பில் ஏற்படும் இடையூறுகள் உண்ணாவிரதத்தின் போது தழுவல் குறைவதற்கு அல்லது அதிகரித்த மன அழுத்தத்திற்கு வழிவகுக்கும், மேலும் சில நாளமில்லா செயலிழப்புகள் - இன்சுலின், தைரோகுளோபுலின், சைட்டோகைன்கள் மற்றும் அவற்றின் ஏற்பிகளின் அளவு குறைகிறது. பலவீனமான புரதச் சிதைவு காயம் குணப்படுத்தும் விகிதத்தைக் குறைக்கிறது, பெருந்தமனி தடிப்புத் தோல் அழற்சியின் வளர்ச்சியை ஏற்படுத்துகிறது மற்றும் நோய் எதிர்ப்பு சக்தியை பாதிக்கிறது. ஹைபோக்ஸியாவுடன், உள் செல்லுலார் pH இல் ஏற்படும் மாற்றங்கள், கதிர்வீச்சு காயம், சவ்வு லிப்பிட்களின் அதிகரித்த பெராக்ஸைடேஷன், அத்துடன் லைசோசோமோட்ரோபிக் பொருட்களின் செல்வாக்கின் கீழ் - பாக்டீரியா எண்டோடாக்சின்கள், நச்சு பூஞ்சைகளின் வளர்சிதை மாற்றங்கள் (ஸ்போரோஃபுசரின்), சிலிக்கான் ஆக்சைடு படிகங்களின் நிலைத்தன்மை மாற்றங்கள், செயல்படுத்தப்பட்ட லைசோசோமால் என்சைம்கள் சைட்டோபிளாஸில் வெளியிடப்படுகின்றன, இது உயிரணு அமைப்புகளின் அழிவு மற்றும் அதன் மரணத்தை ஏற்படுத்துகிறது.
அத்தியாயம் 1
செல் உடலியலின் அடிப்படைகள்
I. டியூடல்
பிளாஸ்மா சவ்வு . விலங்கு செல்கள் பிளாஸ்மா சவ்வு (படம் 1.1) மூலம் பிணைக்கப்பட்டுள்ளன. அதன் கட்டமைப்பில் நாம் வாழ்வோம், இது பல உள்செல்லுலார் சவ்வுகளின் கட்டமைப்பிற்கு மிகவும் ஒத்திருக்கிறது, இன்னும் கொஞ்சம் விரிவாக. மென்படலத்தின் முக்கிய அணி கொண்டுள்ளது கொழுப்புகள்,முக்கியமாக பாஸ்பாடிடைல்கோலின். இந்த லிப்பிடுகள் ஒரு ஹைட்ரோஃபிலிக் ஹெட் குழுவைக் கொண்டிருக்கின்றன, அதில் நீண்ட ஹைட்ரோபோபிக் ஹைட்ரோகார்பன் சங்கிலிகள் இணைக்கப்பட்டுள்ளன. தண்ணீரில், இத்தகைய கொழுப்புகள் தன்னிச்சையாக 4-5 nm தடிமன் கொண்ட ஒரு இரு அடுக்கு படலத்தை உருவாக்குகின்றன, இதில் ஹைட்ரோஃபிலிக் குழுக்கள் அக்வஸ் மீடியத்தை எதிர்கொள்கின்றன, மேலும் ஹைட்ரோஃபோபிக் ஹைட்ரோகார்பன் சங்கிலிகள் இரண்டு வரிசைகளில் அமைக்கப்பட்டு, நீரற்ற லிப்பிட் கட்டத்தை உருவாக்குகின்றன. உயிரணு சவ்வுகள் இந்த வகையின் லிப்பிட் பிலேயர்ஸ் மற்றும் கிளைகோலிப்பிடுகள், கொழுப்பு மற்றும் பாஸ்போலிப்பிட்கள் (படம் 1.2) ஆகியவற்றைக் கொண்டிருக்கின்றன. கிளைகோலிப்பிட்களின் ஹைட்ரோஃபிலிக் பகுதி ஒலிகோசாக்கரைடுகளால் உருவாகிறது. கிளைகோலிப்பிட்கள் எப்போதும் பிளாஸ்மா மென்படலத்தின் வெளிப்புற மேற்பரப்பில் அமைந்துள்ளன, மூலக்கூறின் ஒலிகோசாக்கரைடு பகுதி சுற்றுச்சூழலில் மூழ்கியிருக்கும் முடியைப் போன்றது. கிட்டத்தட்ட சம அளவுகளில் பாஸ்போலிப்பிட்கள் மத்தியில் சிதறி, கொழுப்பு மூலக்கூறுகள் மென்படலத்தை உறுதிப்படுத்துகின்றன. மென்படலத்தின் உள் மற்றும் வெளிப்புற அடுக்குகளில் உள்ள பல்வேறு லிப்பிட்களின் விநியோகம் ஒரே மாதிரியாக இருக்காது, மேலும் ஒரு அடுக்குக்குள் கூட சில வகையான லிப்பிடுகள் குவிந்திருக்கும் பகுதிகள் உள்ளன. இந்த சீரற்ற விநியோகம்
அரிசி. 1.1. மிக முக்கியமான உறுப்புகளைக் காட்டும் கலத்தின் திட்ட வரைபடம்
ஒருவேளை இன்னும் சில, இன்னும் தெளிவற்ற, செயல்பாட்டு முக்கியத்துவம் உள்ளது.
சவ்வின் ஒப்பீட்டளவில் மந்த கொழுப்பு மேட்ரிக்ஸில் உட்பொதிக்கப்பட்ட முக்கிய செயல்பாட்டு கூறுகள் அணில்கள்(படம் 1.2). எடையின் அடிப்படையில் புரதம் பல்வேறு சவ்வுகளில் 25 முதல் 75% வரை உள்ளது, ஆனால் புரத மூலக்கூறுகள் லிப்பிட் மூலக்கூறுகளை விட பெரியதாக இருப்பதால், எடையில் 50% என்பது 1 புரத மூலக்கூறு மற்றும் 50 லிப்பிட் மூலக்கூறுகளின் விகிதத்திற்கு சமம். சில புரதங்கள் சவ்வை அதன் வெளிப்புறத்திலிருந்து உள் மேற்பரப்புக்கு ஊடுருவிச் செல்கின்றன, மற்றவை ஒரு அடுக்கில் சரி செய்யப்படுகின்றன. புரத மூலக்கூறுகள் வழக்கமாக நோக்குநிலை கொண்டவை, இதனால் அவற்றின் ஹைட்ரோபோபிக் குழுக்கள் லிப்பிட் சவ்வில் உட்பொதிக்கப்படுகின்றன மற்றும் மென்படலத்தின் மேற்பரப்பில் உள்ள துருவ ஹைட்ரோஃபிலிக் குழுக்கள் அக்வஸ் கட்டத்தில் உட்பொதிக்கப்படுகின்றன. மென்படலத்தின் வெளிப்புற மேற்பரப்பில் உள்ள பல புரதங்கள் கிளைகோபுரோட்டீன்கள்; அவற்றின் ஹைட்ரோஃபிலிக் சாக்கரைடு குழுக்கள் புற-செல்லுலார் சூழலை எதிர்கொள்கின்றன.
உள்ளக உறுப்புகளின் சவ்வு அமைப்புகள் .
செல் அளவின் ஏறத்தாழ பாதி சைட்டோசோலில் இருந்து சவ்வுகளால் தனிமைப்படுத்தப்பட்ட உறுப்புகளால் ஆக்கிரமிக்கப்பட்டுள்ளது. உட்புற உறுப்புகளின் சவ்வுகளின் மொத்த பரப்பளவு பிளாஸ்மா மென்படலத்தின் மேற்பரப்பை விட குறைந்தது 10 மடங்கு அதிகமாகும். மிகவும் பரவலாகப் பயன்படுத்தப்படும் சவ்வு அமைப்பு எண்டோபிளாஸ்மிக் ரெட்டிகுலம்,ஒரு பிணையத்தை குறிக்கும்

அரிசி. 1.2பிளாஸ்மா மென்படலத்தின் திட்டவட்டமான பிரதிநிதித்துவம். புரதங்கள் பாஸ்போலிப்பிட் பைலேயரில் பதிக்கப்படுகின்றன, அவற்றில் சில பைலேயரில் ஊடுருவுகின்றன, மற்றவை வெளிப்புற அல்லது உள் அடுக்கில் மட்டுமே நங்கூரமிடப்படுகின்றன.
அதிக சுருண்ட குழாய்கள் அல்லது பை போன்ற நீளமான கட்டமைப்புகள்; எண்டோபிளாஸ்மிக் ரெட்டிகுலத்தின் பெரிய பகுதிகள் ரைபோசோம்களால் பதிக்கப்பட்டுள்ளன; அத்தகைய ரெட்டிகுலம் சிறுமணி அல்லது கடினமான (படம் 1.1) என்று அழைக்கப்படுகிறது. கோல்கி எந்திரம்சவ்வு-பிணைக்கப்பட்ட லேமல்லேகளையும் கொண்டுள்ளது, அதில் இருந்து வெசிகிள்ஸ் அல்லது வெசிகல்ஸ் உடைந்து விடும் (படம் 1.1). லைசோசோம்கள் மற்றும் பெராக்ஸிசோம்கள்- இவை சிறிய சிறப்பு வெசிகல்ஸ். இந்த பல்வேறு உறுப்புகளில், சவ்வு மற்றும் அது சூழ்ந்திருக்கும் இடம் ஆகியவை குறிப்பிட்ட நொதிகளை கொண்டிருக்கின்றன; உறுப்புகளின் உள்ளே, சிறப்பு வளர்சிதை மாற்ற பொருட்கள் குவிந்து, உறுப்புகளின் பல்வேறு செயல்பாடுகளைச் செய்யப் பயன்படுகிறது.
கோர்மற்றும் மைட்டோகாண்ட்ரியாஇந்த உறுப்புகள் ஒவ்வொன்றும் இரண்டு சவ்வுகளால் சூழப்பட்டிருப்பதில் வேறுபடுகின்றன. வளர்சிதை மாற்றத்தின் இயக்கக் கட்டுப்பாட்டிற்கு கருவானது பொறுப்பாகும்; மைட்டோகாண்ட்ரியாவின் மடிந்த உள் சவ்வு ஆக்ஸிஜனேற்ற வளர்சிதை மாற்றத்தின் தளமாகும்; இங்கே, பைருவேட் அல்லது கொழுப்பு அமிலங்களின் ஆக்சிஜனேற்றம் காரணமாக, உயர் ஆற்றல் கலவை அடினோசின் ட்ரைபாஸ்பேட் (ATP, அல்லது ATP) ஒருங்கிணைக்கப்படுகிறது.
சைட்டோஸ்கெலட்டன் . உறுப்புகளைச் சுற்றியுள்ள சைட்டோபிளாசம் எந்த வகையிலும் உருவமற்றதாகக் கருத முடியாது; இது சைட்டோஸ்கெலிட்டல் நெட்வொர்க்கால் ஊடுருவுகிறது. சைட்டோஸ்கெலட்டனில் நுண்குழாய்கள், ஆக்டின் இழைகள் மற்றும் இடைநிலை இழைகள் (படம் 1.1) உள்ளன. நுண்குழாய்கள்சுமார் 25 nm வெளிப்புற விட்டம் கொண்டது; அவை டூபுலின் புரத மூலக்கூறுகளின் தொகுப்பின் விளைவாக வழக்கமான பாலிமர் போல உருவாகின்றன. ஆக்டின் இழைகள்-சுருங்கக்கூடிய இழைகள், சவ்வுக்கு அருகிலுள்ள அடுக்கு மற்றும் செல் முழுவதும் அமைந்துள்ளன, முக்கியமாக இயக்கத்துடன் தொடர்புடைய செயல்முறைகளில் பங்கேற்கின்றன. இடைநிலை இழைகள்பல்வேறு வகையான உயிரணுக்களில் வெவ்வேறு இரசாயன கலவையின் தொகுதிகளைக் கொண்டிருக்கும்; அவை மேலே குறிப்பிடப்பட்ட மற்ற இரண்டு சைட்டோஸ்கெலிட்டல் உறுப்புகளுக்கு இடையே பல்வேறு இணைப்புகளை உருவாக்குகின்றன. உறுப்புகள் மற்றும் பிளாஸ்மா சவ்வு ஆகியவை சைட்டோஸ்கெலட்டனுடன் தொடர்புடையவை, இது செல்லின் வடிவத்தையும் அதில் உள்ள உறுப்புகளின் நிலையையும் பராமரிப்பது மட்டுமல்லாமல், செல்லின் வடிவத்திலும் அதன் இயக்கத்திலும் மாற்றங்களை தீர்மானிக்கிறது.
சைட்டோசோல் . செல் அளவின் பாதி அளவு சைட்டோசோலால் ஆக்கிரமிக்கப்பட்டுள்ளது. எடையில் இது தோராயமாக 20% புரதம் என்பதால், இது நீர்வாழ் கரைசலை விட ஜெல் ஆகும். கரிம மற்றும் கனிம உட்பட சிறிய மூலக்கூறுகள் அயனிகள்,நீர்நிலை கட்டத்தில் கரைக்கப்படுகிறது. செல் மற்றும் சுற்றுச்சூழலுக்கு இடையே அயனிகளின் பரிமாற்றம் நிகழ்கிறது (எக்ஸ்ட்ராசெல்லுலர் ஸ்பேஸ்); இந்த வளர்சிதை மாற்ற செயல்முறைகள் அடுத்த பகுதியில் விவாதிக்கப்படும். புற-செல்லுலார் இடத்தில் உள்ள அயனிகளின் செறிவு ஒரு நிலையான மட்டத்தில் கணிசமான துல்லியத்துடன் பராமரிக்கப்படுகிறது; ஒவ்வொரு அயனியின் உள்ளக செறிவும் ஒரு குறிப்பிட்ட அளவைக் கொண்டுள்ளது, இது கலத்திற்கு வெளியே உள்ளதை விட வேறுபட்டது (அட்டவணை 1.1). புற-செல்லுலார் சூழலில் அதிக அளவில் காணப்படும் கேஷன் ஆகும்நா+ கலத்தில் அதன் செறிவு 10 மடங்கு குறைவாக உள்ளது. மாறாக, செல்லின் உள்ளே K + இன் செறிவு அதிகமாக உள்ளது; புற-செல்லுலார் மற்றும் இன்ட்ராசெல்லுலர் செறிவுகளுக்கு இடையே உள்ள மிகப்பெரிய சாய்வு Ca 2+ க்கு உள்ளது, கலத்தின் உள்ளே இருக்கும் இலவச அயனிகளின் செறிவு வெளியில் இருப்பதை விட குறைந்தது 10,000 மடங்கு குறைவாக உள்ளது. அனைத்து அயனிகளும் சைட்டோசோலில் கரைக்கப்படுவதில்லை; அவற்றில் சில புரதங்களில் உறிஞ்சப்படுகின்றன அல்லது உறுப்புகளில் வைக்கப்படுகின்றன. எடுத்துக்காட்டாக, Ca 2+ விஷயத்தில், கட்டற்ற அயனிகள் கட்டற்ற அயனிகளை விட அதிக எண்ணிக்கையில் உள்ளன. பெரும்பாலான சைட்டோசோலிக் புரதங்கள் என்சைம்கள் ஆகும், இதில் பங்கேற்புடன் இடைநிலை வளர்சிதை மாற்றத்தின் பல செயல்முறைகள் மேற்கொள்ளப்படுகின்றன: கிளைகோலிசிஸ் மற்றும் குளுக்கோனோஜெனீசிஸ், அமினோ அமிலங்களின் தொகுப்பு அல்லது அழிவு, ரைபோசோம்களில் புரத தொகுப்பு (படம் 1.1). சைட்டோசோலில் லிப்பிட் துளிகள் மற்றும் கிளைகோஜன் துகள்கள் உள்ளன, அவை முக்கியமான மூலக்கூறுகளின் இருப்புகளாக செயல்படுகின்றன.
அட்டவணை 1.1.ஹோமியோதெர்மிக் விலங்குகளின் தசை செல்களில் உள் மற்றும் புற-செல்லுலர் அயனி செறிவுகள். ஏ – - "உயர் மூலக்கூறு எடை செல்லுலார் அனான்கள்"
|
செல்லுலார் செறிவு |
எக்ஸ்ட்ராசெல்லுலர் செறிவு |
||
|
நா+ |
12 மிமீல்/லி |
நா+ |
145 மிமீல்/லி |
|
155 மிமீல்/லி |
K+ |
4 மிமீல்/லி |
|
|
Ca 2+ |
10 –7 10 –8 mmol/l |
Ca 2+ |
2 மிமீல்/லி |
|
சி எல் - |
4 மிமீல்/லி |
C1 – |
120 மிமீல்/லி |
|
HCO 3 - |
8 மிமீல்/லி |
HCO 3 - |
27 மிமீல்/லி |
|
A – |
155 மிமீல்/லி |
மற்றவைகள் கேஷன்ஸ் |
5 மிமீல்/லி |
|
ஓய்வு திறன் -90 mV |
|||
1.2 செல் மற்றும் சுற்றுச்சூழலுக்கு இடையில் பொருட்களின் பரிமாற்றம்
செல்லின் கட்டமைப்பை சுருக்கமாக விவரித்துள்ளோம், இதன் மூலம் செல்லுலார் உடலியல் அடிப்படைகளை மதிப்பாய்வு செய்ய இந்த விளக்கத்தைப் பயன்படுத்தலாம். எந்தவொரு சந்தர்ப்பத்திலும் ஒரு கலத்தை ஒரு நிலையான பொருளாகக் கருத முடியாது, ஏனெனில் பல்வேறு உள்செல்லுலார் பெட்டிகளுக்கு இடையில், அதே போல் பெட்டிகள் மற்றும் சுற்றுச்சூழலுக்கு இடையில் பொருட்களின் நிலையான பரிமாற்றம் உள்ளது. செல் கட்டமைப்புகள் டைனமிக் சமநிலையில் உள்ளன, மேலும் செல்கள் ஒருவருக்கொருவர் மற்றும் வெளிப்புற சூழலுடன் தொடர்புகொள்வது செயல்படும் உயிரினத்தின் வாழ்க்கையை பராமரிக்க தேவையான நிபந்தனையாகும். இந்த அத்தியாயத்தில் அத்தகைய பரிமாற்றத்தின் அடிப்படை வழிமுறைகளைப் பார்ப்போம். அடுத்தடுத்த அத்தியாயங்களில், இந்த வழிமுறைகள் நரம்பு செல் மற்றும் அதன் செயல்பாடுகளுக்குப் பயன்படுத்தப்பட்டதாகக் கருதப்படும்;
இருப்பினும், அதே வழிமுறைகள் மற்ற அனைத்து உறுப்புகளின் செயல்பாட்டிற்கும் அடிப்படையாக உள்ளன.
பரவல்.பொருளை நகர்த்துவதற்கான எளிய செயல்முறை பரவல் ஆகும். கரைசல்களில் (அல்லது வாயுக்கள்), அணுக்கள் மற்றும் மூலக்கூறுகள் சுதந்திரமாக நகரும், மேலும் பரவல் காரணமாக செறிவுகளில் உள்ள வேறுபாடுகள் சமநிலையில் இருக்கும். திரவம் அல்லது வாயுவால் நிரப்பப்பட்ட இரண்டு தொகுதிகளைக் கருத்தில் கொள்வோம் (படம் 1.3), இதில் பொருட்கள் செறிவுகளைக் கொண்டுள்ளன. c 1 மற்றும் c 2 மற்றும் மேற்பரப்பு A மற்றும் தடிமன் கொண்ட ஒரு அடுக்கு மூலம் பிரிக்கப்பட்டதுஈ. காலத்தின் போது பொருளின் ஓட்டம் m விவரித்தார் ஃபிக்கின் முதல் பரவல் விதி:
dm/ dt= டிஏ/ ஈ ( சி 1 –சி 2)=டிஏ/ ஈடி சி(1)
இங்கு D என்பது பரவல் குணகம், கொடுக்கப்பட்ட பொருளுக்கு மாறிலி, கரைப்பான் மற்றும் வெப்பநிலை. செறிவு வேறுபாட்டிற்கான பொதுவான வடிவத்தில் dc தொலைவில் dx
dm/dt= –D A dc/dx,(2)
பிரிவு A வழியாக ஓட்டம் செறிவு சாய்வுக்கு விகிதாசாரமாகும் dc/dx . x திசையில் செறிவு மாற்றம் எதிர்மறையாக இருப்பதால் சமன்பாட்டில் கழித்தல் குறி தோன்றும்.
பரவல் என்பது மிக முக்கியமான செயல்முறையாகும், இதன் காரணமாக நீர்வாழ் கரைசல்களில் உள்ள பெரும்பாலான மூலக்கூறுகள் குறுகிய தூரத்திற்கு நகர்கின்றன. சவ்வுகளால் பரவல் தடைபடாததால், கலத்தில் அவற்றின் இயக்கத்திற்கும் இது பொருந்தும். பல பொருட்கள் லிப்பிட் சவ்வுகளில் சுதந்திரமாக பரவலாம், குறிப்பாக நீர் மற்றும் O 2 மற்றும் CO 2 போன்ற கரைந்த வாயுக்கள். கொழுப்பு கரையக்கூடியது

அரிசி. 1.3அளவு பரவல் திட்டம். இரண்டு இடைவெளிகளும் தடிமன் ஒரு அடுக்கு மூலம் பிரிக்கப்படுகின்றனஈமற்றும் பகுதி ஏ. C - தொகுதியின் இடது பகுதியில் உள்ள துகள்களின் அதிக செறிவு, C: - வலதுபுறத்தில் துகள்களின் குறைந்த செறிவு; பாகங்கள், இளஞ்சிவப்பு மேற்பரப்பு- பரவல் அடுக்கில் செறிவு சாய்வு. பரவல் ஓட்டம் dm/dt - பார்க்கவும் சமன்பாடு (1)
பொருட்கள் சவ்வுகள் வழியாக நன்கு பரவுகின்றன; எத்தனால் மற்றும் யூரியா போன்ற சிறிய அளவிலான துருவ மூலக்கூறுகளுக்கும் இது பொருந்தும், அதே நேரத்தில் சர்க்கரைகள் கொழுப்பு அடுக்கு வழியாக சிரமத்துடன் செல்கின்றன. அதே நேரத்தில், கொழுப்பு அடுக்குகள், கனிம அயனிகள் உட்பட, சார்ஜ் செய்யப்பட்ட மூலக்கூறுகளுக்கு நடைமுறையில் ஊடுருவ முடியாதவை. எலக்ட்ரோலைட்டுகள் அல்லாதவற்றுக்கு, பரவல் சமன்பாடு (1) பொதுவாக சவ்வு மற்றும் பரவும் பொருளின் பண்புகளை இணைப்பதன் மூலம் மாற்றப்படுகிறது. அளவுரு-ஊடுருவக்கூடிய தன்மை (P):
dm/dt=P Aடி c.(3)
படத்தில். 1.4 ஒப்பிடப்படுகிறது பல்வேறு மூலக்கூறுகளுக்கான லிப்பிட் சவ்வின் ஊடுருவல் (P).
சவ்வு துளைகள் மூலம் பரவல் . பிளாஸ்மா சவ்வு (மற்றும் பிற உயிரணு சவ்வுகள்) கொழுப்பு அடுக்கு வழியாக பரவும் பொருட்களுக்கு மட்டுமல்ல, பல அயனிகள், சர்க்கரைகள், அமினோ அமிலங்கள் மற்றும் நியூக்ளியோடைட்களுக்கும் ஊடுருவக்கூடியது. இந்த பொருட்கள் உருவாகும் துளைகள் வழியாக சவ்வை கடக்கின்றன போக்குவரத்து புரதங்கள்,படலத்தில் மூழ்கியது. அத்தகைய புரதங்களின் உள்ளே 1 nm க்கும் குறைவான விட்டம் கொண்ட நீர் நிரப்பப்பட்ட சேனல் உள்ளது, இதன் மூலம் சிறிய மூலக்கூறுகள் பரவுகின்றன. அவை செறிவு சாய்வுடன் நகர்கின்றன, மேலும் அவை ஒரு கட்டணத்தை சுமந்தால், சேனல்கள் வழியாக அவற்றின் இயக்கம் சவ்வு ஆற்றலால் கட்டுப்படுத்தப்படுகிறது. மெம்பிரேன் சேனல்கள் ஒப்பீட்டளவில் தேர்ந்தெடுக்கப்பட்டவை

அரிசி. 1.4பல்வேறு பொருட்களுக்கு செயற்கை லிப்பிட் பைலேயர்களின் ஊடுருவல்
அவற்றின் வழியாக செல்லக்கூடிய மூலக்கூறுகளின் வகை தொடர்பாக, பொட்டாசியம், சோடியம் மற்றும் கால்சியம் சேனல்கள் உள்ளன, அவை ஒவ்வொன்றும் ஒரு குறிப்பிட்ட ஒன்றைத் தவிர எந்த அயனிக்கும் ஊடுருவ முடியாதவை. அத்தகைய தேர்ந்தெடுக்கும் திறன்சேனல் சுவர்களில் உள்ள பிணைப்பு தளங்களின் கட்டணம் அல்லது கட்டமைப்பால் தீர்மானிக்கப்படுகிறது, இது ஒரு குறிப்பிட்ட மூலக்கூறின் போக்குவரத்தை எளிதாக்குகிறது மற்றும் சேனல் வழியாக மற்ற பொருட்களின் ஊடுருவலைத் தடுக்கிறது (படம் 1). 1.5, A) .
நடத்தைக்கு பின்னால் சவ்வு அயனி சேனல்கள்கவனிக்க எளிதானது, ஏனெனில் அயனிகளின் இயக்கத்திலிருந்து எழும் மின்னோட்டத்தை ஒரு சேனலுக்கு கூட அளவிட முடியும். சேனல்கள் தன்னிச்சையாகவும் அதிக அதிர்வெண்ணுடனும் தங்கள் நிலையை திறந்த நிலையில் இருந்து மூடியதாக மாற்றுவதாகக் காட்டப்படுகிறது. பொட்டாசியம் சேனல் தற்போதைய பருப்புகளால் வகைப்படுத்தப்படுகிறது, இது சுமார் 2 pA (2 10 –12 A) வீச்சு மற்றும் பல மில்லி விநாடிகளின் கால அளவு (படம் 2.12, ப. 37 ஐப் பார்க்கவும்) [3]. இந்த காலகட்டத்தில், பல்லாயிரக்கணக்கான அயனிகள் அதன் வழியாக செல்கின்றன. புரதங்கள் ஒரு இணக்கத்திலிருந்து மற்றொன்றுக்கு மாறுவது எக்ஸ்ரே டிஃப்ராஃப்ரக்ஷன், மாஸ்பவுர் ஸ்பெக்ட்ரோஸ்கோபி மற்றும் நியூக்ளியர் மேக்னடிக் ரெசோனன்ஸ் (என்எம்ஆர்) மூலம் ஆய்வு செய்யப்படுகிறது. புரோட்டீன்கள் மிகவும் ஆற்றல் வாய்ந்த, மொபைல் கட்டமைப்புகள், மற்றும் புரதம் வழியாக செல்லும் சேனல் ஒரு திடமான, நீர் நிரப்பப்பட்ட குழாய் (படம். 1.5, A), ஆனால் விரைவாக நகரும் மூலக்கூறு குழுக்கள் மற்றும் கட்டணங்களின் ஒரு தளம் ஆகும். சேனலின் இந்த மாறும் பண்பு பிரதிபலிக்கிறது சேனலின் ஆற்றல் விவரக்குறிப்பு,படம் காட்டப்பட்டுள்ளது. 1.5, B. இங்கே, abscissa அச்சு என்பது வெளிப்புறக் கரைசலில் இருந்து அயன் செறிவு C 0 மற்றும் சாத்தியமான 0 செறிவு C 1 மற்றும் சாத்தியமான E உடன் உள்ள உள் தீர்வு வரையிலான சேனலின் நீளத்தைக் குறிக்கிறது. ஆர்டினேட் அச்சு

அரிசி. 1.5A. பிளாஸ்மா மென்படலத்தின் லிப்பிட் பைலேயரில் பதிக்கப்பட்ட பொட்டாசியம் சேனலை உருவாக்கும் புரதத்தின் திட்டம். சேனலின் "சுவரில்" நான்கு எதிர்மறை கட்டணங்கள் நிர்ணயிக்கப்பட்டுள்ளன. பி. படத்தில் காட்டப்பட்டுள்ள சேனலின் திட்ட ஆற்றல் சுயவிவரம். A. ஆர்டினேட் அச்சு, சேனலின் பாதைக்கு தேவையான இயக்க ஆற்றலின் மதிப்புகளைக் காட்டுகிறது; abscissa அச்சில் - மென்படலத்தின் உள் மற்றும் வெளிப்புற மேற்பரப்புகளுக்கு இடையே உள்ள தூரம். ஆற்றல் மினிமா என்பது சேனல் சுவரில் நிலையான எதிர்மறை கட்டணங்களுடன் நேர்மறையாக சார்ஜ் செய்யப்பட்ட அயனிகளின் பிணைப்பு தளங்களுக்கு ஒத்திருக்கிறது. ஆற்றல் அதிகபட்சம் சேனலில் உள்ள பரவல் தடைகளுக்கு ஒத்திருக்கிறது. சேனல் புரதத்தின் இணக்கமானது தன்னிச்சையாக ஊசலாடுவதாகக் கருதப்படுகிறது; ஆற்றல் சுயவிவர விருப்பங்கள் திடமான மற்றும் கோடுகளுடன் காட்டப்பட்டுள்ளன; இந்த ஊசலாட்டங்கள் ஆற்றல் தடையை கடக்கும்போது அயனிகளின் பிணைப்பை பெரிதும் எளிதாக்குகிறது (மாற்றியமைக்கப்பட்டது)
சேனல் பிணைப்பு தளங்களில் அயனி ஆற்றல் நிலைகள் வழங்கப்படுகின்றன; வரைபடத்தில் உள்ள உச்சமானது, சேனலில் ஊடுருவுவதற்கு அயனி ஆற்றல் அதிகமாக இருக்க வேண்டிய ஊடுருவல் தடையை குறிக்கிறது, மேலும் வரைபடத்தில் உள்ள "டிப்" என்பது ஒப்பீட்டளவில் நிலையான நிலையை (பிணைப்பு) குறிக்கிறது. ஆற்றல் உச்சத்தின் தடையாக இருந்தாலும், ஆற்றல் சுயவிவரம் தன்னிச்சையாக மாறினால், அயனி சேனலில் ஊடுருவ முடியும்; அயனி திடீரென ஆற்றல் உச்சத்தின் "மறுபுறத்தில்" தன்னைக் கண்டுபிடித்து, கலத்திற்குள் தொடர்ந்து செல்லலாம். அயனியின் சார்ஜ், அளவு மற்றும் நீரேற்றத்தின் அளவு மற்றும் சேனல் சுவர் கட்டமைப்புகளுடன் பிணைக்கும் திறன் ஆகியவற்றைப் பொறுத்து, சேனலின் ஆற்றல் சுயவிவரம் வெவ்வேறு அயனிகளுக்கு மாறுபடும், இது தனிப்பட்ட சேனல் வகைகளின் தேர்வை விளக்கலாம்.
அயனிகளின் பரவல் சமநிலை . சவ்வு சேனல்கள் மூலம் பல்வேறு அயனிகளின் பரவல் கூடுதல் மற்றும் உள்செல்லுலார் சூழல்களுக்கு இடையே உள்ள செறிவுகளில் உள்ள வேறுபாடுகளை நீக்குவதற்கு வழிவகுக்கும். இருப்பினும், அட்டவணையில் இருந்து பார்க்க முடியும். 1.1, அத்தகைய வேறுபாடுகள் உள்ளன, எனவே, சில இருக்க வேண்டும் சமநிலைசவ்வு முழுவதும் பரவல் மற்றும் பிற போக்குவரத்து செயல்முறைகளுக்கு இடையில். அடுத்த இரண்டு பிரிவுகள் இந்த சமநிலையை அடையும் வழிகளைக் கையாள்கின்றன. அயனிகளின் விஷயத்தில், பரவல் சமநிலை அவற்றின் கட்டணத்தால் பாதிக்கப்படுகிறது. சார்ஜ் செய்யப்படாத மூலக்கூறுகளின் பரவல் செறிவுகளில் உள்ள வேறுபாட்டால் உறுதி செய்யப்படுகிறது dc , மற்றும் செறிவுகள் சமப்படுத்தப்படும் போது, போக்குவரத்து நிறுத்தப்படும். சார்ஜ் செய்யப்பட்ட துகள்கள் கூடுதலாக மின்சார புலத்தால் பாதிக்கப்படுகின்றன. எடுத்துக்காட்டாக, ஒரு பொட்டாசியம் அயனி ஒரு கலத்தை அதன் செறிவு சாய்வுடன் விட்டுச் செல்லும் போது, அது ஒரு நேர்மறை மின்னூட்டத்தைக் கொண்டுள்ளது. இதனால், உள்செல்லுலார் சூழல் மிகவும் எதிர்மறையாக சார்ஜ் செய்யப்படுகிறது, இதன் விளைவாக சவ்வு முழுவதும் சாத்தியமான வேறுபாடு ஏற்படுகிறது. உள்செல்லுலார் நெகடிவ் சார்ஜ் புதிய பொட்டாசியம் அயனிகள் செல்லிலிருந்து வெளியேறுவதைத் தடுக்கிறது, மேலும் செல்லை விட்டு வெளியேறும் அயனிகள் மென்படலத்தின் மீதான கட்டணத்தை மேலும் அதிகரிக்கும். மின்புலத்தின் செயல்பாடு செறிவு வேறுபாடு காரணமாக பரவல் அழுத்தத்தை ஈடுசெய்யும்போது பொட்டாசியம் அயனிகளின் ஓட்டம் நிறுத்தப்படும். அயனிகள் சவ்வு வழியாக தொடர்ந்து செல்கின்றன, ஆனால் இரு திசைகளிலும் சம அளவுகளில். எனவே, சவ்வு மீது அயனி செறிவுகளில் கொடுக்கப்பட்ட வேறுபாட்டிற்கு, உள்ளது சமநிலை சாத்தியம்ஈஅயனி சவ்வு வழியாக அயனிகளின் ஓட்டம் நிறுத்தப்படும். சமநிலை திறனைப் பயன்படுத்தி எளிதாக தீர்மானிக்க முடியும் நெர்ன்ஸ்ட் சமன்பாடுகள்:
ஈஅயனி= ஆர்டி/ zஎஃப்* lnசி அவுட்/ சி இன்(4)
அங்கு ஆர் - வாயு மாறிலி, T - முழுமையான வெப்பநிலை, z - அயன் வேலன்சி (அயனிகளுக்கு எதிர்மறை)சி அவுட் - புற-செல்லுலார் அயனி செறிவு,சி இன் - செல்லுலார் அயனி செறிவு,எஃப் ஃபாரடே எண். நாம் சமன்பாட்டில் மாறிலிகளை மாற்றினால், உடல் வெப்பநிலையில் (T = 310 K) பொட்டாசியம் அயனிகளின் சமநிலை சாத்தியம் E K இதற்கு சமம்:
எக்= –61 மீபி பதிவு /(5)
[ K + out ]/[ K + in என்றால் ] = 39, அட்டவணையில் இருந்து பின்வருமாறு. 1.1 பின்னர்
Ek= –61 m B பதிவு 39= –97 mV.
உண்மையில், எல்லா செல்களும் இருப்பது கண்டறியப்பட்டது சவ்வு திறன்;பாலூட்டிகளின் தசை செல்களில் அதன் நிலை -90 எம்.வி. நிலைமைகள் மற்றும் தொடர்புடைய அயனி செறிவுகளைப் பொறுத்து, செல்கள் -40 முதல் -120 mV வரையிலான சவ்வு திறனைக் கொண்டிருக்கலாம். மேலே உள்ள எடுத்துக்காட்டில் உள்ள கலத்திற்கு (அட்டவணை 1.1) ஓய்வு திறன்,தோராயமாக -90 mV க்கு சமம், சவ்வு சேனல்கள் மூலம் பொட்டாசியம் அயனிகளின் பாய்ச்சல்கள் தோராயமாக சமநிலையில் இருப்பதைக் குறிக்கிறது. இது ஆச்சரியமல்ல, ஏனெனில் ஓய்வு சவ்வுகளில் பொட்டாசியம் சேனல்களின் திறந்த நிலை பெரும்பாலும் உள்ளது, அதாவது. சவ்வு பொட்டாசியம் அயனிகளுக்கு மிகவும் ஊடுருவக்கூடியது. இருப்பினும், சவ்வு திறன் மற்ற அயனிகளின் ஓட்டத்தால் தீர்மானிக்கப்படுகிறது.
மின்னூட்டம் இல்லாத துகள்கள் சவ்வு வழியாக எளிதில் பரவுவது சமன்பாட்டில் (3) அளவிடப்படுகிறது. சார்ஜ் செய்யப்பட்ட துகள்களுக்கு ஊடுருவக்கூடிய தன்மை சற்று சிக்கலான சமன்பாட்டால் விவரிக்கப்படுகிறது:
பி= மீ ஆர்டி/ dF(6)
எங்கே மீ- மென்படலத்தில் அயனி இயக்கம்,ஈ - சவ்வு தடிமன்,ஒரு ஆர், டி மற்றும் எஃப் - அறியப்பட்ட தெர்மோடைனமிக் மாறிலிகள். இந்த வழியில் தீர்மானிக்கப்பட்ட பல்வேறு அயனிகளுக்கான ஊடுருவல் மதிப்புகள் சவ்வு திறனைக் கணக்கிட பயன்படுத்தப்படலாம்எம் , பொட்டாசியம், சோடியம் மற்றும் குளோரின் அயனிகள் சவ்வு வழியாக ஒரே நேரத்தில் செல்லும் போது (ஊடுருவக்கூடிய P உடன் K, P Na மற்றும் P Cl முறையே). மென்படலத்தில் சாத்தியக்கூறு சீராக குறைகிறது என்று கருதப்படுகிறது, அதனால் புல வலிமை நிலையானது. இந்த வழக்கில் அது பொருந்தும் கோல்ட்மேன் சமன்பாடு அல்லது நிலையான புலச் சமன்பாடு :
ஈm= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl )(7)
பெரும்பாலான செல் சவ்வுகளுக்கு பிகே P ஐ விட தோராயமாக 30 மடங்கு அதிகம்நா (பிரிவு 1.3 ஐயும் பார்க்கவும்). ஒப்பீட்டு மதிப்புபி Cl பெரிதும் மாறுபடும்; பல சவ்வுகளுக்குபி Cl R உடன் ஒப்பிடும்போது சிறியதுகே இருப்பினும் மற்றவர்களுக்கு (உதாரணமாக, எலும்பு தசைகளில்)பி Cl , P ஐ விட கணிசமாக அதிகம்கே.
செயலில் போக்குவரத்து, சோடியம் பம்ப் . முந்தைய பகுதி அயனிகளின் செயலற்ற பரவல் மற்றும் கொடுக்கப்பட்ட உள் மற்றும் புற-செல்லுலார் அயனி செறிவுகளின் விளைவாக சவ்வு திறனை விவரிக்கிறது. இருப்பினும், இந்த செயல்முறையின் விளைவாக, செல் உள்ளே உள்ள அயனிகளின் செறிவு தானாகவே நிலைப்படுத்தப்படுவதில்லை, ஏனெனில் சவ்வு
சாத்தியக்கூறுகளை விட சற்றே அதிகமான எலக்ட்ரோநெக்டிவ் EK, மற்றும் ஈ உடன் ஒப்பிடும்போது அதிகம்நா (சுமார் +60 mV). பரவல் காரணமாக, உள்செல்லுலார் அயனி செறிவுகள், குறைந்தபட்சம் பொட்டாசியம் மற்றும் சோடியம், எக்ஸ்ட்ராசெல்லுலர்களுடன் சமப்படுத்தப்பட வேண்டும். அயனி சாய்வின் நிலைத்தன்மை செயலில் உள்ள போக்குவரத்து மூலம் அடையப்படுகிறது: சவ்வு புரதங்கள் மின் மற்றும்/அல்லது செறிவு சாய்வுகளுக்கு எதிராக சவ்வு முழுவதும் அயனிகளை கொண்டு செல்கின்றன, இதற்காக வளர்சிதை மாற்ற ஆற்றலைப் பயன்படுத்துகின்றன. செயலில் போக்குவரத்தின் மிக முக்கியமான செயல்முறை வேலைநா/கே - கிட்டத்தட்ட அனைத்து செல்களிலும் இருக்கும் ஒரு பம்ப்;
பம்ப் சோடியம் அயனிகளை கலத்திலிருந்து வெளியேற்றுகிறது, அதே நேரத்தில் பொட்டாசியம் அயனிகளை செல்லுக்குள் செலுத்துகிறது. இது சோடியம் அயனிகளின் குறைந்த செல்லுலார் செறிவு மற்றும் பொட்டாசியம் அயனிகளின் அதிக செறிவு ஆகியவற்றை உறுதி செய்கிறது (அட்டவணை 1.1). மென்படலத்தில் உள்ள சோடியம் அயனிகளின் செறிவு சாய்வு மின் தூண்டுதல்களின் வடிவத்தில் தகவல் பரிமாற்றம் தொடர்பான குறிப்பிட்ட செயல்பாடுகளைக் கொண்டுள்ளது (பிரிவு 2.2 ஐப் பார்க்கவும்), அத்துடன் பிற செயலில் உள்ள போக்குவரத்து வழிமுறைகளைப் பராமரித்தல் மற்றும் செல் அளவைக் கட்டுப்படுத்துதல் (கீழே காண்க). எனவே, ஒரு கலத்தால் நுகரப்படும் ஆற்றலில் 1/3 க்கும் அதிகமானவை Na/K பம்பில் செலவழிக்கப்படுவதில் ஆச்சரியமில்லை, மேலும் சில செயலில் உள்ள செல்களில் 70% ஆற்றல் வரை அதன் செயல்பாட்டிற்கு செலவிடப்படுகிறது.
Na/K போக்குவரத்து புரதம் ஒரு ATPase ஆகும். சவ்வு உள் மேற்பரப்பில், அது ஏடிபி மற்றும் பாஸ்பேட் (படம். 1.6) ATP உடைக்கிறது. ஒரு ஏடிபி மூலக்கூறின் ஆற்றல், கலத்திலிருந்து மூன்று சோடியம் அயனிகளையும், ஒரே நேரத்தில் இரண்டு பொட்டாசியம் அயனிகளையும் செல்லுக்குள் கொண்டு செல்லப் பயன்படுகிறது, அதாவது மொத்தமாக, ஒரு சுழற்சியில் கலத்திலிருந்து ஒரு நேர்மறை கட்டணம் அகற்றப்படுகிறது. இவ்வாறு, Na/K பம்ப் ஆகும் எலக்ட்ரோஜெனிக்(சவ்வு முழுவதும் ஒரு மின்னோட்டத்தை உருவாக்குகிறது), இது சவ்வு ஆற்றலின் எலக்ட்ரோநெக்டிவிட்டியை தோராயமாக 10 mV ஆக அதிகரிக்க வழிவகுக்கிறது. போக்குவரத்து புரதம் இந்த செயல்பாட்டை அதிக வேகத்தில் செய்கிறது: வினாடிக்கு 150 முதல் 600 சோடியம் அயனிகள். போக்குவரத்து புரதத்தின் அமினோ அமில வரிசை அறியப்படுகிறது, ஆனால் இந்த சிக்கலான வளர்சிதை மாற்ற போக்குவரத்தின் வழிமுறை இன்னும் தெளிவாக இல்லை. இந்த செயல்முறை சோடியம் அல்லது பொட்டாசியம் அயனிகளை புரதங்களால் மாற்றும் ஆற்றல் சுயவிவரங்களைப் பயன்படுத்தி விவரிக்கப்படுகிறது (படம் 1.5,5). இந்த சுயவிவரங்களில் ஏற்படும் மாற்றங்களின் தன்மையால், போக்குவரத்து புரதத்தின் (ஆற்றல் தேவைப்படும் செயல்முறை) இணக்கத்தின் நிலையான மாற்றங்களுடன் தொடர்புடையது, பரிமாற்றத்தின் ஸ்டோச்சியோமெட்ரியை ஒருவர் தீர்மானிக்க முடியும்: இரண்டு பொட்டாசியம் அயனிகள் மூன்று சோடியம் அயனிகளுக்கு பரிமாறிக்கொள்ளப்படுகின்றன.
Na/K பம்ப், தனிமைப்படுத்தப்பட்டதைப் போன்றதுநா+ /K + -சார்ந்த சவ்வு ATPase, குறிப்பாக கார்டியாக் கிளைகோசைட் ouabain (strophanthin) மூலம் தடுக்கப்படுகிறது. Na/K விசையியக்கக் குழாயின் செயல்பாடு பல-நிலை இரசாயன எதிர்வினை என்பதால், அனைத்து இரசாயன எதிர்வினைகளைப் போலவே, இது பெரும்பாலும் வெப்பநிலையைச் சார்ந்தது.

அரிசி. 1.6Na/K பம்ப்-ATPase இன் வரைபடம் (பிளாஸ்மா மென்படலத்தின் லிப்பிட் பிளேயரில் மூழ்கியுள்ளது), இது ஒரு சுழற்சியில் மூன்று Na + அயனிகளை கலத்திலிருந்து சாத்தியமான மற்றும் செறிவு சாய்வுகளுக்கு எதிராக அகற்றி, இரண்டு K அயனிகளை கலத்திற்குள் கொண்டு வருகிறது. + . இந்த செயல்பாட்டின் போது, ஒரு ATP மூலக்கூறு ADP மற்றும் பாஸ்பேட்டாக உடைக்கப்படுகிறது. வரைபடம் ATPase ஐ ஒரு பெரிய (செயல்பாட்டு) மற்றும் சிறிய துணைக்குழுக்களைக் கொண்ட ஒரு டைமராகக் காட்டுகிறது; மென்படலத்தில் இது இரண்டு பெரிய மற்றும் இரண்டு சிறிய துணைக்குழுக்களால் உருவாக்கப்பட்ட டெட்ராமராக உள்ளது
படம் காட்டப்பட்டுள்ளது. 1.7 இங்கே, தசை செல்களில் இருந்து சோடியம் அயனிகளின் ஃப்ளக்ஸ் நேரத்துடன் ஒப்பிடப்படுகிறது; இது Na/K பம்பின் செயல்பாட்டின் மூலம் மத்தியஸ்தம் செய்யப்பட்ட சோடியம் அயனிகளின் ஓட்டத்திற்கு நடைமுறையில் சமமானதாகும், ஏனெனில் செறிவு மற்றும் சாத்தியமான சாய்வுகளுக்கு எதிராக சோடியம் அயனிகளின் செயலற்ற ஓட்டம் மிகவும் சிறியது. மருந்து தோராயமாக 18 டிகிரி செல்சியஸால் குளிர்ந்தால், கலத்திலிருந்து சோடியம் அயனிகளின் ஓட்டம் விரைவாக 15 மடங்கு குறையும், உடனடியாக வெப்பமடைந்த பிறகு அது அதன் அசல் நிலைக்கு மீட்டமைக்கப்படும். கலத்திலிருந்து சோடியம் அயனிகளின் ஓட்டத்தில் ஏற்படும் இந்த குறைவு, பரவல் செயல்முறையின் வெப்பநிலை சார்பு அல்லது ஒரு எளிய இரசாயன எதிர்வினைக்கு ஒத்திருப்பதை விட பல மடங்கு அதிகமாகும். டைனிட்ரோபீனால் (டிஎன்பி) நச்சுத்தன்மையின் விளைவாக வளர்சிதை மாற்ற ஆற்றல் குறையும் போது இதேபோன்ற விளைவு காணப்படுகிறது (படம் 1.7,5). இதன் விளைவாக, கலத்திலிருந்து சோடியம் அயனிகளின் ஓட்டம் ஆற்றல் சார்ந்த எதிர்வினை மூலம் உறுதி செய்யப்படுகிறது - செயலில் உள்ள பம்ப். பம்பின் மற்றொரு சிறப்பியல்பு, குறிப்பிடத்தக்க வெப்பநிலை மற்றும் ஆற்றல் சார்பு ஆகியவற்றுடன், ஒரு செறிவூட்டல் நிலை (மற்ற அனைத்து இரசாயன எதிர்வினைகள் போன்றது); கடத்தப்பட்ட அயனிகளின் செறிவு அதிகரிப்பதால் பம்பின் வேகம் காலவரையின்றி அதிகரிக்க முடியாது என்பதே இதன் பொருள் (படம் 1.8). இதற்கு நேர்மாறாக, ஒரு செயலற்ற முறையில் பரவும் பொருளின் ஃப்ளக்ஸ் பரவல் விதியின்படி செறிவு வேறுபாட்டின் விகிதத்தில் அதிகரிக்கிறது (சமன்பாடுகள் 1 மற்றும் 2).

அரிசி. 1.7 ஏ, பி.செயலில் நா போக்குவரத்து + Y அச்சு:கலத்திலிருந்து கதிரியக்க 24 Na + பாய்ச்சல் (imp./min). அப்சிஸ்ஸா அச்சு:சோதனை தொடங்கிய நேரம். ஏ.செல் 18.3 °C முதல் 0.5 °C வரை குளிர்விக்கப்படுகிறது; ஓட்டம்நா+ இந்த காலகட்டத்தில் செல்லில் இருந்து தடுக்கப்படுகிறது. பி. 0.2 mmol/l (மாற்றியமைக்கப்பட்ட) செறிவில் டைனிட்ரோபீனால் (DNP) உடன் செல்லிலிருந்து Na + ஃப்ளக்ஸை அடக்குதல்
Na/K பம்புடன் கூடுதலாக, பிளாஸ்மா சவ்வு குறைந்தது ஒரு பம்பைக் கொண்டுள்ளது - கால்சியம்;இந்த பம்ப் கால்சியம் அயனிகளை (Ca 2+) கலத்திலிருந்து வெளியேற்றுகிறது மற்றும் அவற்றின் உள்ளக செறிவை மிகக் குறைந்த அளவில் பராமரிப்பதில் ஈடுபட்டுள்ளது (அட்டவணை 1.1). கால்சியம் பம்ப் தசை செல்களின் சர்கோபிளாஸ்மிக் ரெட்டிகுலத்தில் மிக அதிக அடர்த்தியில் உள்ளது, இது ஏடிபி மூலக்கூறுகளின் முறிவின் விளைவாக கால்சியம் அயனிகளைக் குவிக்கிறது (அத்தியாயம் 4 ஐப் பார்க்கவும்).
சவ்வு திறன் மற்றும் செல் அளவு மீது Na/K பம்பின் விளைவு . படத்தில். படம் 1.9 சவ்வு மின்னோட்டத்தின் பல்வேறு கூறுகளைக் காட்டுகிறது மற்றும் அயனிகளின் உள்ளக செறிவுகளைக் காட்டுகிறது

அரிசி. 1.8மூலக்கூறுகளின் போக்குவரத்து விகிதம் மற்றும் அவற்றின் செறிவு (சேனலின் நுழைவாயிலில் அல்லது பம்பின் பிணைப்பு தளத்தில்) சேனல் வழியாக பரவும் போது அல்லது உந்தி போக்குவரத்தின் போது இடையேயான உறவு. பிந்தையது அதிக செறிவுகளில் நிறைவுற்றது (அதிகபட்ச வேகம், Vmax x-அச்சின் மதிப்பு அதிகபட்ச பம்ப் வேகத்தில் பாதிக்கு ( Vmax /2), சமநிலை செறிவு ஆகும் TO மீ

அரிசி. 1.9Na+ செறிவுகளைக் காட்டும் வரைபடம் , K+ மற்றும் Cl - கலத்தின் உள்ளேயும் வெளியேயும் செல் சவ்வு வழியாக இந்த அயனிகளின் ஊடுருவலுக்கான பாதைகள் (குறிப்பிட்ட அயனி சேனல்கள் அல்லது Na/K பம்ப் மூலம். கொடுக்கப்பட்ட செறிவு சாய்வு, சமநிலை சாத்தியங்கள் E Na, E K மற்றும் E Сl – சுட்டிக்காட்டப்பட்டவற்றுக்கு சமம், சவ்வு திறன்எம் = – 90 எம்.வி
அவர்களின் இருப்பை உறுதி. பொட்டாசியம் அயனிகளின் வெளிப்புற மின்னோட்டம் பொட்டாசியம் சேனல்கள் மூலம் காணப்படுகிறது, ஏனெனில் சவ்வு திறன் பொட்டாசியம் அயனிகளுக்கான சமநிலை ஆற்றலை விட சற்றே கூடுதலான எலக்ட்ரோபாசிட்டிவ் ஆகும். சோடியம் சேனல்களின் ஒட்டுமொத்த கடத்துத்திறன் பொட்டாசியம் சேனல்களை விட மிகவும் குறைவாக உள்ளது, அதாவது. சோடியம் சேனல்கள் பொட்டாசியம் சேனல்களை விட மிகவும் குறைவாகவே திறந்திருக்கும். இருப்பினும், பொட்டாசியம் அயனிகள் செல்லில் இருந்து வெளியேறும் போது தோராயமாக அதே எண்ணிக்கையிலான சோடியம் அயனிகள் செல்லுக்குள் நுழைகின்றன, ஏனெனில் சோடியம் அயனிகள் செல்லில் பரவுவதற்கு பெரிய செறிவு மற்றும் சாத்தியமான சாய்வுகள் தேவைப்படுகின்றன. Na/K பம்ப் செயலற்ற பரவல் நீரோட்டங்களுக்கு சிறந்த இழப்பீட்டை வழங்குகிறது, ஏனெனில் அது செல் மற்றும் பொட்டாசியம் அயனிகளை அதற்குள் இருந்து சோடியம் அயனிகளை கொண்டு செல்கிறது. இவ்வாறு, பம்ப் ஆனது கலத்திற்கு உள்ளேயும் வெளியேயும் மாற்றப்படும் கட்டணங்களின் எண்ணிக்கையில் உள்ள வேறுபாட்டால் எலக்ட்ரோஜெனிக் ஆகும், இது சாதாரண வேகத்தில் அதன் செயல்பாட்டின் போது தோராயமாக 10 சவ்வு திறனை உருவாக்குகிறது. mV ஆனது செயலற்ற அயனிப் பாய்வுகளால் மட்டும் உற்பத்தி செய்யப்பட்டதை விட அதிக எலக்ட்ரோநெக்டிவ் ஆகும் (சமன்பாடு 7 ஐப் பார்க்கவும்). இதன் விளைவாக, சவ்வு திறன் பொட்டாசியம் சமநிலை ஆற்றலை அணுகுகிறது, இது பொட்டாசியம் அயனிகளின் கசிவைக் குறைக்கிறது. நா செயல்பாடு/K பம்ப் சோடியம் அயனிகளின் உள்ளக செறிவு மூலம் கட்டுப்படுத்தப்படுகிறது. கலத்திலிருந்து அகற்றப்படும் சோடியம் அயனிகளின் செறிவு குறைவதால் பம்பின் வேகம் குறைகிறது (படம். 1.8), இதனால் பம்ப் செயல்பாடும், கலத்திற்குள் சோடியம் அயனிகளின் ஓட்டமும் ஒன்றையொன்று சமநிலைப்படுத்தி, சோடியத்தின் உள்செறிவை பராமரிக்கிறது. அயனிகள் தோராயமாக 10 mmol/L அளவில்.
உந்தி மற்றும் செயலற்ற சவ்வு மின்னோட்டங்களுக்கு இடையே சமநிலையை பராமரிக்க, பொட்டாசியம் மற்றும் சோடியம் அயனிகளுக்கான சேனல் புரதங்களை விட அதிகமான Na/K பம்ப் மூலக்கூறுகள் தேவைப்படுகின்றன. சேனல் திறந்திருக்கும் போது, பல்லாயிரக்கணக்கான அயனிகள் சில மில்லி விநாடிகளில் (மேலே காண்க) கடந்து செல்கின்றன, மேலும் சேனல் வழக்கமாக வினாடிக்கு பல முறை திறக்கப்படுவதால், மொத்தத்தில் 10 5 அயனிகள் இந்த நேரத்தில் கடந்து செல்கின்றன. ஒரு பம்ப் புரதம் வினாடிக்கு பல நூறு சோடியம் அயனிகளை நகர்த்துகிறது, எனவே பிளாஸ்மா சவ்வு சேனல் மூலக்கூறுகளை விட 1000 மடங்கு அதிகமான பம்ப் மூலக்கூறுகளைக் கொண்டிருக்க வேண்டும். ஓய்வு நேரத்தில் சேனல் நீரோட்டங்களின் அளவீடுகள் 1 μm 2 சவ்வுக்கு சராசரியாக ஒரு பொட்டாசியம் மற்றும் ஒரு சோடியம் திறந்த சேனல் இருப்பதைக் காட்டியது; இதிலிருந்து Na/K பம்பின் சுமார் 1000 மூலக்கூறுகள் ஒரே இடத்தில் இருக்க வேண்டும், அதாவது. அவற்றுக்கிடையேயான தூரம் சராசரியாக 34 nm ஆகும்; பம்ப் புரதத்தின் விட்டம், ஒரு சேனல் புரதமாக, 8-10 nm ஆகும். இதனால், சவ்வு உந்தி மூலக்கூறுகளுடன் மிகவும் அடர்த்தியாக நிறைவுற்றது
கலத்திற்குள் சோடியம் அயனிகளின் ஓட்டமும், கலத்திலிருந்து பொட்டாசியம் அயனிகளும் பம்பின் செயல்பாட்டின் மூலம் ஈடுசெய்யப்படுகின்றன, இது ஒரு நிலையான ஆஸ்மோடிக் அழுத்தத்தை பராமரிப்பதில் மற்றொரு விளைவு உள்ளது. நிலையான தொகுதி.கலத்தின் உள்ளே பெரிய அயனிகளின் அதிக செறிவு உள்ளது, முக்கியமாக புரதங்கள் (A - அட்டவணை 1.1 இல்), அவை சவ்வுக்குள் ஊடுருவ முடியாது (அல்லது அதன் வழியாக மிக மெதுவாக ஊடுருவுகின்றன) எனவே செல் உள்ளே ஒரு நிலையான கூறு ஆகும். இந்த அனான்களின் மின்னூட்டத்தை சமநிலைப்படுத்த, சம எண்ணிக்கையிலான கேஷன்கள் தேவை. Na/K பம்பின் செயல்பாட்டின் காரணமாக, இந்த கேஷன்கள் முக்கியமாக பொட்டாசியம் அயனிகளாகும். C1 இன் ஓட்டத்தின் காரணமாக அயனிகளின் செறிவு அதிகரிப்பால் மட்டுமே உயிரணுக்களுக்குள் அயனி செறிவில் குறிப்பிடத்தக்க அதிகரிப்பு நிகழலாம் - செறிவு சாய்வுடன் கலத்திற்குள் (அட்டவணை 1.1), ஆனால் சவ்வு திறன் இதை எதிர்க்கிறது. உள்வரும் மின்னோட்டம் Cl - குளோரின் அயனிகளுக்கான சமநிலை சாத்தியம் அடையும் வரை மட்டுமே கவனிக்கப்படுகிறது; குளோரின் அயனிகள் எதிர்மறையாக சார்ஜ் செய்யப்படுவதால், குளோரின் அயனி சாய்வு பொட்டாசியம் அயன் சாய்வுக்கு கிட்டத்தட்ட எதிர்மாறாக இருக்கும் போது இது கவனிக்கப்படுகிறது (சமன்பாடு 4). இதனால், பொட்டாசியம் அயனிகளின் குறைந்த புறச் செறிவுடன் தொடர்புடைய குளோரின் அயனிகளின் குறைந்த செல்லுலார் செறிவு நிறுவப்பட்டது. இதன் விளைவாக கலத்தில் உள்ள மொத்த அயனிகளின் எண்ணிக்கை வரம்பு. Na/K பம்ப் தடுக்கப்படும் போது சவ்வு சாத்தியக்கூறு வீழ்ச்சியடைந்தால், உதாரணமாக அனாக்ஸியாவின் போது, குளோரின் அயனிகளுக்கான சமநிலை சாத்தியம் குறைகிறது, மேலும் குளோரைடு அயனிகளின் உள்ளக செறிவு அதற்கேற்ப அதிகரிக்கிறது. கட்டணங்களின் சமநிலையை மீட்டெடுப்பது, பொட்டாசியம் அயனிகளும் செல்லுக்குள் நுழைகின்றன; கலத்தில் உள்ள அயனிகளின் மொத்த செறிவு அதிகரிக்கிறது, இது ஆஸ்மோடிக் அழுத்தத்தை அதிகரிக்கிறது; இது செல்லுக்குள் தண்ணீரை செலுத்துகிறது. செல் வீங்குகிறது. இந்த வீக்கம் கவனிக்கப்படுகிறதுஉயிருள்ள ஆற்றல் இல்லாத நிலையில்.
செறிவு சாய்வு நா + சவ்வு போக்குவரத்துக்கான உந்து சக்தியாக . கலத்திற்கான Na/K பம்பின் முக்கியத்துவம், மென்படலத்தில் உள்ள சாதாரண K + மற்றும் Na + சாய்வுகளின் நிலைப்படுத்தலுடன் மட்டுப்படுத்தப்படவில்லை. சவ்வு சாய்வில் சேமிக்கப்படும் ஆற்றல்நா+ , மற்ற பொருட்களின் சவ்வு போக்குவரத்தை எளிதாக்க பெரும்பாலும் பயன்படுத்தப்படுகிறது. உதாரணமாக, படத்தில். 1.10 "இறக்குமதி" காட்டுகிறதுநா+ மற்றும் செல்லில் சர்க்கரை மூலக்கூறுகள். சவ்வு போக்குவரத்து புரதம் சர்க்கரை மூலக்கூறை ஒரு செறிவு சாய்வுக்கு எதிராக கூட செல்லுக்குள் கொண்டு செல்கிறது, அதே நேரத்தில் Na + செறிவு மற்றும் சாத்தியமான சாய்வு வழியாக நகர்கிறது, ஆற்றலை வழங்குகிறது. சர்க்கரைகளின் போக்குவரத்து.சர்க்கரைகளின் இந்த போக்குவரத்து முற்றிலும் உயர் சாய்வு இருப்பதைப் பொறுத்ததுநா+ ; செல்களுக்குள் செறிவு இருந்தால்நா+ கணிசமாக அதிகரிக்கிறது, சர்க்கரைகளின் போக்குவரத்து நிறுத்தப்படும். பல்வேறு c akharov, பல்வேறு இறக்குமதி அமைப்புகள் உள்ளன. அமினோ அமிலங்களின் போக்குவரத்துபோக்குவரத்துக்கு ஒத்த ஒரு கலத்திற்குள் c அகரோவ் படம் காட்டப்பட்டுள்ளது. 1.10; இது சாய்வு மூலம் வழங்கப்படுகிறதுநா+ , குறைந்தது ஐந்து வெவ்வேறு சிம்போர்ட் அமைப்புகள் உள்ளன, ஒவ்வொன்றும் தொடர்புடைய அமினோ அமிலங்களின் ஒரு குழுவிற்கு சிறப்பு.
இறக்குமதி அமைப்புகளுக்கு கூடுதலாக, உள்ளன "எதிர்ப்பு போர்ட்டர்கள்".அவற்றில் ஒன்று, எடுத்துக்காட்டாக, ஒரு சுழற்சியில் மூன்று உள்வரும் சோடியம் அயனிகளுக்கு ஈடாக ஒரு கால்சியம் அயனியை கலத்திலிருந்து மாற்றுகிறது (படம் 1.10). ஆற்றல் Ca 2+ போக்குவரத்துசெறிவு மற்றும் சாத்தியமான சாய்வு ஆகியவற்றுடன் மூன்று சோடியம் அயனிகளின் நுழைவு காரணமாக உருவாகிறது. இந்த ஆற்றல் அதிக கால்சியம் அயனி சாய்வு (செல்லுக்குள் 10-7 mol/L க்கும் குறைவாக இருந்து செல்லுக்கு வெளியே தோராயமாக 2 mmol/L வரை) பராமரிக்க போதுமானது (ஓய்வெடுக்கும் திறனில்).
எண்டோ- மற்றும் எக்சோசைடோசிஸ் . செல்லுக்குள் நுழையும் அல்லது வெளியேற்றப்பட வேண்டிய சில பொருட்களுக்கு

அரிசி. 1.10மென்படலத்தின் லிப்பிட் பிலேயரில் பதிக்கப்பட்ட புரதங்கள் குளுக்கோஸ் மற்றும் Na + ஆகியவற்றின் அறிகுறியை கலத்திற்குள் மத்தியஸ்தம் செய்கின்றன. Ca2+/Na+ -ஆன்டிபோர்ட், இதில் உந்துவிசை செல் சவ்வு மீது Na + சாய்வு ஆகும்
அதிலிருந்து, போக்குவரத்து சேனல்கள் இல்லை; இந்த பொருட்களில், எடுத்துக்காட்டாக, புரதங்கள் மற்றும் கொலஸ்ட்ரால் ஆகியவை அடங்கும். அவை பிளாஸ்மா சவ்வு வழியாக செல்ல முடியும் கொப்புளங்கள்,அல்லது குமிழ்கள்,எண்டோ- மற்றும் எக்சோசைடோசிஸ் மூலம். படத்தில். இந்த செயல்முறைகளின் முக்கிய வழிமுறைகளை படம் 1.11 காட்டுகிறது. எக்சோசைட்டோசிஸில், சில உறுப்புகள் (கீழே காண்க) ஹார்மோன்கள் அல்லது எக்ஸ்ட்ராசெல்லுலர் என்சைம்கள் போன்ற கலத்திலிருந்து அகற்றப்பட வேண்டிய ஒரு பொருளால் நிரப்பப்பட்ட வெசிகிள்களை உருவாக்குகின்றன. இத்தகைய கொப்புளங்கள் பிளாஸ்மா மென்படலத்தை அடையும் போது, அவற்றின் கொழுப்பு சவ்வு அதனுடன் இணைகிறது, இதனால் உள்ளடக்கங்கள் வெளிப்புற சூழலில் வெளியேற அனுமதிக்கிறது. எதிர் செயல்பாட்டில், எண்டோசைட்டோசிஸ், பிளாஸ்மா சவ்வு ஊடுருவி, ஒரு குழியை உருவாக்குகிறது, பின்னர் அது ஆழமடைந்து மூடுகிறது, எக்ஸ்ட்ராசெல்லுலர் திரவம் மற்றும் சில மேக்ரோமாலிகுல்களால் நிரப்பப்பட்ட ஒரு உள்செல்லுலார் வெசிகிளை உருவாக்குகிறது. இந்த சவ்வு இணைவு மற்றும் வெசிகலின் மூடுதலை உறுதிப்படுத்த, சைட்டோஸ்கெலட்டனின் சுருக்க கூறுகள் சவ்வுகளுடன் இணைந்து செயல்படுகின்றன (கீழே காண்க). எண்டோசைட்டோசிஸ் என்பது செல்களுக்குள் உள்ள புற-செல்லுலார் சூழலைக் கைப்பற்றுவதை எப்போதும் உள்ளடக்குவதில்லை. உயிரணு சவ்வு பெரும்பாலும் இன்சுலின் அல்லது ஆன்டிஜென்கள் போன்ற மேக்ரோமிகுல்களுக்கான குறிப்பிட்ட ஏற்பிகளைக் கொண்டுள்ளது, அவை பெரும்பாலும் சிறப்பு குழுக்களாக ஒழுங்கமைக்கப்படுகின்றன. இந்த மேக்ரோமிகுலூக்கள் அவற்றின் ஏற்பிகளுடன் பிணைக்கப்பட்ட பிறகு, ஏற்பியைச் சுற்றியுள்ள சவ்வு பகுதியில் எண்டோசைட்டோசிஸ் ஏற்படுகிறது, மேலும் மேக்ரோமாலிகுல் தேர்ந்தெடுக்கப்பட்ட முறையில் செல்லுக்குள் கொண்டு செல்லப்படுகிறது (படம் 1.12, பி).
உயிரணுக்களில் எண்டோ- மற்றும் எக்சோசைடோசிஸ் தொடர்ந்து நிகழ்கிறது. சவ்வு பொருள் விற்றுமுதல் அளவு குறிப்பிடத்தக்கது; 1 மணி நேரத்திற்குள், மேக்ரோபேஜ் அதன் சைட்டோபிளாஸ்மிக் மென்படலத்தின் மேற்பரப்பை விட இருமடங்கு வெசிகல்ஸ் வடிவில் உறிஞ்சுகிறது. பெரும்பாலான உயிரணுக்களில், சவ்வு பொருளின் விற்றுமுதல் தீவிரமாக நிகழவில்லை, ஆனால் அது இன்னும் குறிப்பிடத்தக்கதாக இருக்க வேண்டும்.

அரிசி. 1.11.எக்சோசைடோசிஸ் மற்றும் எண்டோசைடோசிஸ். மேலே:செல்லுலார் வெசிகல் பிளாஸ்மா மென்படலத்தின் லிப்பிட் பைலேயருடன் இணைகிறது மற்றும் எக்ஸ்ட்ராசெல்லுலர் இடைவெளியில் திறக்கிறது. இந்த செயல்முறை எக்சோசைடோசிஸ் என்று அழைக்கப்படுகிறது. கீழே:பிளாஸ்மா சவ்வு ஒரு சிறிய பகுதியில் ஊடுருவி வெளிசெல்லுலார் பொருட்களால் நிரப்பப்பட்ட ஒரு வெசிகிளை வெளியிடுகிறது. இந்த செயல்முறை எண்டோசைடோசிஸ் என்று அழைக்கப்படுகிறது
1.3 கலத்திற்குள் உள்ள பொருட்களின் போக்குவரத்து
எண்டோ- மற்றும் எக்சோசைடோசிஸ் என்பது உயிரணு சவ்வு முழுவதும் பொருட்களின் போக்குவரத்து செயல்முறைகள் மட்டுமல்ல, சவ்வுகளின் பரிமாற்ற செயல்முறைகளும் ஆகும் - உயிரணுவின் கட்டமைப்பு கூறுகள். இந்த பிரிவில் கருத்தில் கொள்ள வேண்டிய பொருள் செல் மற்றும் அதன் உறுப்புகளில் உள்ள மற்ற ஒத்த போக்குவரத்து செயல்முறைகள் ஆகும்.
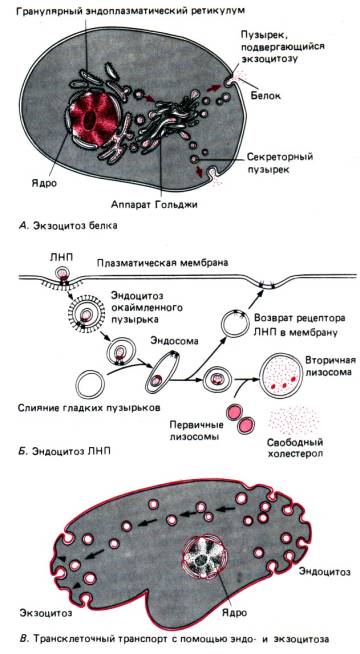
அரிசி. 1.12. ஏ-பி.எக்ஸோ- மற்றும் எண்டோசைடோசிஸ் உள்ளிட்ட செயல்முறைகளின் திட்டம். ஏ.சிறுமணி எண்டோபிளாஸ்மிக் ரெட்டிகுலத்தில் தொகுக்கப்பட்ட புரதம் கோல்கி கருவி மூலம் பிளாஸ்மா மென்படலத்திற்கு கொண்டு செல்லப்படுகிறது, அங்கு அது எக்சோசைடோசிஸ் மூலம் சுரக்கப்படுகிறது. பி.எல்டிஎல் (குறைந்த அடர்த்தி கொழுப்புப்புரதம்) துகள்களுடன் பிணைக்கப்பட்ட கொலஸ்ட்ரால் பிளாஸ்மா மென்படலத்துடன் இணைகிறது, சவ்வின் இந்த பகுதியில் ஒரு எண்டோசைடிக் வெசிகல் உருவாவதைத் தூண்டுகிறது மற்றும் லைசோசோம்களுக்கு கொண்டு செல்லப்படுகிறது, அங்கு அது வெளியிடப்படுகிறது. INஎண்டோசைட்டோசிஸின் போது கைப்பற்றப்பட்ட எக்ஸ்ட்ராசெல்லுலர் பொருள் (படத்தில் வலதுபுறம்),வெசிகிள்ஸ் அல்லது வெசிகல்களில் செல் வழியாக கொண்டு செல்லப்பட்டு, எக்சோசைடோசிஸ் மூலம் வெளியிடப்படுகிறது (படத்தில் இடது)
பரவல் . இயற்கையாகவே, சைட்டோசோலில் பரவல் காரணமாக செறிவு வேறுபாடு நீக்கப்படுகிறது; உறுப்புகளில் உள்ள திரவங்களுக்கும் இது பொருந்தும். கரைந்த புரதத்தின் அதிக செறிவு காரணமாக, நீரை விட இங்கு பரவல் மிகவும் மெதுவாக உள்ளது. லிப்பிட் சவ்வுகள்-செல்களைச் சுற்றியுள்ள மற்றும் உறுப்புகளுக்குள்-இரு பரிமாண திரவங்கள், இதில் பரவல் ஏற்படுகிறது. சவ்வு பைலேயரில் உள்ள லிப்பிடுகள் அவற்றின் சொந்த அடுக்குக்குள் பரவுகின்றன, அரிதாகவே ஒன்றிலிருந்து மற்றொன்றுக்கு நகரும். அவற்றில் மூழ்கியிருக்கும் புரதங்களும் மிகவும் மொபைல் ஆகும்; அவை மென்படலத்திற்கு செங்குத்தாக ஒரு அச்சில் சுழல்கின்றன அல்லது மிகவும் வேறுபட்ட பரவல் மாறிலிகளுடன் பக்கவாட்டில் பரவுகின்றன, பாஸ்போலிப்பிட்களை விட 2–10,000 மடங்கு மெதுவாக. எனவே, சில புரதங்கள் லிப்பிட் அடுக்கில் சுதந்திரமாகவும், கொழுப்பு மூலக்கூறுகளின் அதே வேகத்திலும் நகர்ந்தால், மற்றவை நங்கூரமிடப்படுகின்றன, அதாவது. சைட்டோஸ்கெலட்டனுடன் மிகவும் உறுதியாக இணைக்கப்பட்டுள்ளது. மென்படலத்தில் குறிப்பிட்ட புரதங்களின் "நிரந்தர" திரட்டுகள் உள்ளன, எடுத்துக்காட்டாக, நரம்பு செல்களின் முன் மற்றும் போஸ்டினாப்டிக் கட்டமைப்புகள். சுதந்திரமாக நகரும் புரதங்களை ஃப்ளோரசன்ட் சாயங்களுடன் பிணைப்பதன் மூலம் நிரூபிக்க முடியும், இதன் ஒளிர்வு சவ்வின் ஒரு சிறிய பகுதியை சுருக்கமாக ஒளிரச் செய்வதன் மூலம் தூண்டப்படுகிறது. இத்தகைய சோதனைகள் 1 நிமிடத்திற்கும் குறைவான நேரத்தில், சாயத்துடன் பிணைக்கப்பட்ட புரதங்கள் 10 மைக்ரான் தொலைவில் சவ்வு முழுவதும் ஒரே மாதிரியாக விநியோகிக்கப்படுகின்றன என்பதைக் காட்டுகின்றன.
உறுப்பு சவ்வுகளில் செயலில் போக்குவரத்து .
பிளாஸ்மா மென்படலத்தின் செயல்பாட்டில் முக்கிய பங்கு வகிக்கும் செயலில் போக்குவரத்து செயல்முறைகள், செல் உள்ளே, உறுப்புகளின் சவ்வுகளில் நிகழ்கின்றன. பல்வேறு உறுப்புகளின் குறிப்பிட்ட உள்ளடக்கங்கள் ஓரளவு உள் தொகுப்பு காரணமாகவும், ஓரளவு சைட்டோசோலில் இருந்து செயல்படும் போக்குவரத்து காரணமாகவும் உருவாக்கப்படுகின்றன. பிந்தையவற்றின் ஒரு எடுத்துக்காட்டு, தசை செல்களின் சர்கோபிளாஸ்மிக் ரெட்டிகுலத்தில் மேலே குறிப்பிடப்பட்ட Ca 2+ பம்ப் ஆகும். மைட்டோகாண்ட்ரியாவில் ஏடிபி தொகுப்பின் விஷயத்தில், பிளாஸ்மா மென்படலத்தின் ஏடிபேஸ் பம்புகளில் (படம் 1.6) என்ன நிகழ்கிறது என்பதற்கு எதிர் கொள்கை பொருந்தும் என்பது மிகவும் சுவாரஸ்யமானது. ATP தொகுப்பின் போது, ஆக்ஸிஜனேற்ற வளர்சிதை மாற்றம் செங்குத்தான சாய்வு உருவாவதற்கு வழிவகுக்கிறது. H+ உள் சவ்வுகளில். இந்த சாய்வு மூலக்கூறுகளின் சுறுசுறுப்பான போக்குவரத்தின் உந்தி சுழற்சியின் தலைகீழ் செயல்முறைக்கான உந்து சக்தியாகும்: H + அயனிகள் சவ்வு முழுவதும் சாய்வு வழியாக நகர்கின்றன, இதன் விளைவாக வெளியிடப்படும் ஆற்றல் ADP மற்றும் பாஸ்பேட்டிலிருந்து ATP இன் தொகுப்பை வழங்குகிறது. இதன் விளைவாக வரும் ஏடிபி, செயலில் உள்ள போக்குவரத்து உட்பட கலத்திற்கு ஆற்றலை வழங்குகிறது.
வெசிகல்களில் போக்குவரத்து . செல் அதிக எண்ணிக்கையிலான உறுப்புகள் மற்றும் அதனுடன் தொடர்புடைய வெசிகல்ஸ் (படம் 1.1). இந்த உறுப்புகள் மற்றும் குறிப்பாக வெசிகல்கள் நிலையான இயக்கத்தில் உள்ளன, அவற்றின் உள்ளடக்கங்களை மற்ற உறுப்புகளுக்கு அல்லது பிளாஸ்மா சவ்வுக்கு கொண்டு செல்கின்றன. எண்டோசைட்டோசிஸைப் போலவே, வெசிகல்ஸ் செல் சவ்விலிருந்து உறுப்புகளுக்கு இடம்பெயரலாம்.
செயல்முறை புரதச் சுரப்புபடம் காட்டப்பட்டுள்ளது. 1.12, ஏ.எண்டோபிளாஸ்மிக் ரெட்டிகுலத்துடன் (கிரானுலர், அல்லது கரடுமுரடான, எண்டோபிளாஸ்மிக் ரெட்டிகுலம் என்று அழைக்கப்படுபவை) தொடர்புடைய ரைபோசோம்களில் செல் கருவுக்கு அருகில் புரதம் ஒருங்கிணைக்கப்படுகிறது; எண்டோபிளாஸ்மிக் ரெட்டிகுலத்தில் ஒருமுறை, புரதம் போக்குவரத்து வெசிகல்களில் தொகுக்கப்படுகிறது, அவை உறுப்புகளிலிருந்து பிரிந்து கோல்கி கருவிக்கு இடம்பெயர்கின்றன. இங்கே அவை கோல்கி கருவியின் தொட்டிகளுடன் ஒன்றிணைகின்றன, அங்கு புரதம் மாற்றியமைக்கப்படுகிறது (அதாவது, கிளைகோபுரோட்டீனாக மாற்றப்படுகிறது). சிஸ்டெர்னாவின் முனைகளில், கொப்புளங்கள் மீண்டும் பிரிக்கப்படுகின்றன. மாற்றியமைக்கப்பட்ட புரதத்தைச் சுமந்து செல்லும் சுரப்பு வெசிகிள்கள் பிளாஸ்மா சவ்வுக்கு நகர்ந்து எக்சோசைடோசிஸ் மூலம் அவற்றின் உள்ளடக்கங்களை வெளியிடுகின்றன.
ஒரு கலத்தில் போக்குவரத்து பாதையின் மற்றொரு உதாரணம் படம். 1.12, பி; இது உயிரணு மூலம் கொலஸ்ட்ராலை உறிஞ்சுவதாகும். இரத்தத்தில் கடத்தப்படும் கொலஸ்ட்ரால் முக்கியமாக துகள்கள் போன்ற புரதங்களுடன் பிணைக்கப்பட்டுள்ளது "குறைந்த அடர்த்தி கொழுப்புப்புரதம்"(எல்டிஎல்). இத்தகைய துகள்கள் எல்டிஎல் ஏற்பிகளைக் கொண்ட குறிப்பிட்ட சவ்வுப் பகுதிகளுடன் இணைகின்றன, அங்கு எண்டோசைட்டோசிஸ் ஏற்படுகிறது மற்றும் எல்டிஎல் "எல்லை" வெசிகல்களில் செல்லுக்குள் கொண்டு செல்லப்படுகிறது. இந்த வெசிகிள்கள் ஒன்றிணைந்து எண்டோசோம்களை உருவாக்குகின்றன மற்றும் இந்த செயல்பாட்டின் போது அவற்றின் "விளிம்புகளை" இழக்கின்றன. எண்டோசோம்கள், முதன்மையான லைசோசோம்களுடன் ஒன்றிணைகின்றன, இதில் முக்கியமாக ஹைட்ரோலைடிக் என்சைம்கள் உள்ளன, மேலும் இரண்டாம் நிலை, பெரிய லைசோசோம்களை உருவாக்குகின்றன. அவற்றில், எல்டிஎல் துகள்களிலிருந்து கொலஸ்ட்ரால் வெளியிடப்பட்டு சைட்டோசோலில் பரவுகிறது, எடுத்துக்காட்டாக, லிப்பிட் சவ்வுகளின் தொகுப்புக்காக அது கிடைக்கிறது. எல்டிஎல் இல்லாத வெசிகல்களும் எண்டோசோம்களிலிருந்து பிரிக்கப்படுகின்றன, அவை பிளாஸ்மா சவ்வுக்கு ஒரு சிறப்பு வழியில் நகர்ந்து அதனுடன் ஒன்றிணைந்து, சவ்வு பொருள் மற்றும், அநேகமாக, எல்டிஎல் ஏற்பிகளைத் திரும்பப் பெறுகின்றன. எல்டிஎல் துகள் சவ்வுடன் பிணைந்த தருணத்திலிருந்து இரண்டாம் நிலை லைசோசோமில் இருந்து கொலஸ்ட்ரால் வெளியிடப்படும் வரை, 10-15 நிமிடங்கள் கடந்து செல்கின்றன. எல்.டி.எல் பிணைப்பு மற்றும் உறிஞ்சுதலில் உள்ள குறைபாடுகள், அதாவது, உயிரணுவிற்கு கொலஸ்ட்ரால் வழங்குவதில், ஒரு தீவிரமான மற்றும் பரவலான நோய், பெருந்தமனி தடிப்புத் தோல் அழற்சி (தமனிகளின் "கடினப்படுத்துதல்") வளர்ச்சியில் ஒரு தீர்க்கமான பாத்திரத்தை வகிக்கிறது.
படத்தில் காட்டப்பட்டுள்ளதைப் போன்ற பல போக்குவரத்து வழிகள் உள்ளன. 1.11 மற்றும் 1.12, A, இதன் உதவியுடன் குறிப்பிட்ட வெசிகிள்கள் கலத்தில் நகரும். அவை எவ்வாறு நகர்கின்றன என்பது சரியாகத் தெரியவில்லை, ஆனால் சைட்டோஸ்கெலிட்டல் கூறுகள் இந்த செயல்பாட்டில் ஈடுபட்டிருக்கலாம். நுண்குழாய்கள் வழியாக நுண்குழாய்கள் சறுக்கக்கூடும், இதில் இயக்கத்திற்கான ஆற்றல், ATPase (கீழே காண்க) வெசிகல்களுடன் தொடர்புடைய புரதத்தால் வழங்கப்படுகிறது. அனைத்து திசைகளிலும் ஒன்றன் பின் ஒன்றாக நகர்ந்து எத்தனை வெவ்வேறு வெசிகிள்கள் தங்கள் இலக்கை அடைகின்றன என்பது முற்றிலும் தெளிவாக இல்லை. போக்குவரத்து அமைப்பால் அங்கீகரிக்கப்பட்டு இலக்கு இயக்கமாக மாற்றப்படும் வகையில் அவை வெளிப்படையாக "குறியிடப்பட வேண்டும்".
உறுப்புகளின் உருவாக்கம் மற்றும் அழிவின் மூலம் போக்குவரத்து . இதுவரை, எண்டோ- மற்றும் எக்சோசைடோசிஸ் ஆகியவை வெசிகிள்களின் உள்ளடக்கங்களைக் கொண்டு செல்லும் செயல்முறைகளாகக் கருதினோம். இந்த செயல்முறைகளின் மற்றொரு அம்சம் உள்ளது, இது எண்டோசைட்டோசிஸ் மூலம் செல் மேற்பரப்பின் ஒரு பகுதியில் பிளாஸ்மா மென்படலத்தை இலக்காக அகற்றுவது மற்றும் அதற்கு மாறாக, எக்சோசைடோசிஸ் மூலம் மற்றொன்றைச் சேர்ப்பது சவ்வின் குறிப்பிடத்தக்க பகுதிகளை நகர்த்துகிறது (படம் 1.12.E), கலத்திற்கு வாய்ப்பளிக்கிறது, எடுத்துக்காட்டாக, ஒரு வளர்ச்சி அல்லது நகர்வை உருவாக்குகிறது.
இதேபோன்ற மறுசீரமைப்புகள் சைட்டோஸ்கெலட்டனுக்கும் பொதுவானவை, குறிப்பாக மைக்ரோஃபிலமென்ட்ஸ் மற்றும் மைக்ரோடூபுல்களுக்கு (படம் 1.1). நுண் இழைகள்முதன்மையாக கொண்டது எஃப்-ஆக்டின் புரதம்,சைட்டோசோலில் இருந்து மோனோமரின் பாலிமரைசேஷனின் விளைவாக நார்ச்சத்து மூட்டைகளாக இணைக்கும் திறன் கொண்டது. மூட்டைகள் துருவப்படுத்தப்படுகின்றன, அதாவது, அவை பெரும்பாலும் ஒரு முனையில் மட்டுமே வளர்கின்றன, புதிய ஆக்டின் மூலக்கூறுகளைக் குவிக்கின்றன, மறுமுனை செயலற்றதாகவோ அல்லது பிரித்தெடுப்பதாகவோ இங்கே நிகழ்கிறது. இந்த துருவப்படுத்தப்பட்ட வளர்ச்சியின் காரணமாக, மைக்ரோஃபிலமென்ட்கள் திறமையாக நகரும் மற்றும் அவற்றின் நெட்வொர்க்கின் அமைப்பு மாறலாம். ஆக்டின் ஒரு டிபோலிமரைஸ் செய்யப்பட்ட நிலையிலிருந்து (சோல்) ஒரு ஒழுங்கமைக்கப்பட்ட நிலைக்கு (ஜெல்) மாறுவது மற்ற புரதங்களின் செல்வாக்கின் கீழ் அல்லது அயனி செறிவு மாற்றங்களின் கீழ் மிக விரைவாக நிகழலாம் (கீழே காண்க). ஆக்டின் இழைகள் உடைந்து குறுகிய துண்டுகளை உருவாக்கும் புரதங்களும் உள்ளன. பல செல்களின் மெல்லிய கணிப்புகள் - ஃபிலோபோடியா - ஆக்டின் மைய மூட்டை (படம் 1.1) கொண்டிருக்கும், மேலும் ஃபிலோபோடியாவின் பல்வேறு இயக்கங்கள் ஆக்டின் மாற்றங்களால் இருக்கலாம்: பாலிமரைசேஷன் - டிபோலிமரைசேஷன்.
நுண்குழாய்கள்அடிக்கடி இதே போன்ற இயக்கங்களுக்கு உட்படுகிறது. இந்த இயக்கங்களின் பொறிமுறையானது ஒத்ததாகும் - சைட்டோசோலில் இருந்து டூபுலின் பாலிமரைசேஷன், மைக்ரோடூபூலின் முனைகளில் ஒன்று வளரும், மற்றொன்று மாறாது, அல்லது பிரித்தெடுத்தல் அங்கு நிகழ்கிறது. இவ்வாறு, ஒரு நுண்குழாய், சரியான சேர்த்தல் அல்லது பொருளை அகற்றுவதன் மூலம், சைட்டோசோல் முழுவதும் நகர முடியும்.
சைட்டோஸ்கெலட்டனின் செயலில் இயக்கங்கள் . மேலே விவரிக்கப்பட்ட செயலில் உள்ள இயக்கங்கள் மற்றும் மறுசீரமைப்புகளின் விளைவாக சைட்டோஸ்கெலிட்டல் கட்டமைப்புகளில் மாற்றங்கள் ஏற்படலாம். பல சமயங்களில், நுண்குழாய்கள் மற்றும் ஆக்டின் இழைகளின் இயக்கம் சுருங்கும் புரதங்களால் இயக்கப்படுகிறது, அவை இழைகள் அல்லது குழாய்களை பிணைக்கின்றன மற்றும் அவற்றை ஒன்றுடன் ஒன்று நகர்த்தலாம். அணில்கள் myosin மற்றும் dyneinஒப்பீட்டளவில் அதிக செறிவுகளில் அனைத்து செல்களின் சைட்டோசோலில் உள்ளது; அவை சிறப்பு செல்கள் (தசை) மற்றும் உறுப்புகளில் (சிலியா) ஆற்றலை இயக்கமாக மாற்றும் கூறுகள். தசை செல்களில், மயோசின் ஆக்டின் இழைகளுக்கு இணையாக தடிமனான இழைகளை உருவாக்குகிறது. "தலை" கொண்ட மயோசின் மூலக்கூறு ஆக்டின் இழையுடன் இணைகிறது மற்றும் ஏடிபியின் ஆற்றலைப் பயன்படுத்தி, ஆக்டின் மூலக்கூறுடன் மயோசினை இடமாற்றம் செய்கிறது.மயோசின் பின்னர் ஆக்டினிலிருந்து பிரிகிறது. இதுபோன்ற பல இணைப்பு-துண்டிப்பு சுழற்சிகளின் கலவையானது மேக்ரோஸ்கோபிக்கு வழிவகுக்கிறது தசை நார்களின் சுருக்கம்(அத்தியாயம் 4). சிலியாவின் போது நுண்குழாய்களின் இயக்கத்தில் டைனீன் இதேபோன்ற பாத்திரத்தை வகிக்கிறது (படம் 1.1). சிறப்பு இல்லாத உயிரணுக்களின் சைட்டோபிளாஸில், மயோசின் மற்றும் டைனீன் வழக்கமான இழைகளை உருவாக்குவதில்லை, ஆனால் பெரும்பாலான சந்தர்ப்பங்களில் மூலக்கூறுகளின் சிறிய குழுக்கள். அத்தகைய சிறிய தொகுப்புகளில் கூட, அவை ஆக்டின் இழைகள் அல்லது நுண்குழாய்களை நகர்த்தும் திறன் கொண்டவை. அரிசி. படம் 1.13 இந்த செயல்முறையை விளக்குகிறது, எதிரெதிர் துருவப்படுத்தப்பட்ட மயோசின் மூலக்கூறுகளும் இரண்டு ஆக்டின் இழைகளுடன் இணைக்கப்பட்டு, வெவ்வேறு திசைகளில் துருவப்படுத்தப்படுகின்றன. மயோசின் தலைக்குழுக்கள் மூலக்கூறின் வால் நோக்கி வளைந்து, செயல்பாட்டில் ATP ஐ உட்கொள்கின்றன, மேலும் இரண்டு ஆக்டின் இழைகளும் எதிர் திசையில் நகரும், அதன் பிறகு மயோசின் அவற்றிலிருந்து பிரிகிறது. இந்த வகையான இயக்கங்கள், ATP ஆற்றல் இயந்திர வேலையாக மாற்றப்படும் போது, சைட்டோஸ்கெலட்டனின் வடிவத்தை மாற்றலாம், அதன் விளைவாக, செல், மேலும் சைட்டோஸ்கெலட்டனுடன் தொடர்புடைய உறுப்புகளின் போக்குவரத்தையும் வழங்குகிறது.
உள்செல்லுலார் போக்குவரத்தின் செயல்முறைகள் ஒரு நரம்பு கலத்தின் அச்சில் மிகத் தெளிவாக நிரூபிக்கப்படலாம். ஆக்சன் போக்குவரத்துபெரும்பாலான செல்களில் இதே போன்ற வழிகளில் நிகழக்கூடிய நிகழ்வுகளை விளக்குவதற்கு இங்கு விரிவாக விவாதிக்கப்படுகிறது. ஒரு சில மைக்ரான்கள் விட்டம் கொண்ட ஒரு ஆக்சன் ஒரு மீட்டர் அல்லது அதற்கு மேற்பட்ட நீளத்தை எட்டும், மேலும் அணுக்கருவிலிருந்து தொலைதூர முனை வரை பரவுவதன் மூலம் புரதங்களின் இயக்கம் பல ஆண்டுகள் எடுக்கும். நரம்பிழையின் எந்தப் பகுதியும் சுருங்கும் போது, ஆக்சானின் பகுதியானது அருகாமையில் விரிவடைகிறது என்பது நீண்ட காலமாக அறியப்படுகிறது. ஆக்சனில் மையவிலக்கு ஓட்டம் தடுக்கப்பட்டது போல் தெரிகிறது. அத்தகைய ஓட்டம்-வேகமான ஆக்சன் போக்குவரத்து முடியும்படத்தில் காட்டப்பட்டுள்ள பரிசோதனையில், கதிரியக்க குறிப்பான்களின் இயக்கம் மூலம் நிரூபிக்கப்படும். 1.14. ரேடியோலேபல் செய்யப்பட்ட லியூசின் முதுகு வேர் கேங்க்லியனில் செலுத்தப்பட்டது, பின்னர் 2வது முதல் 10வது மணிநேரம் வரை நரம்பணு செல் உடல்களில் இருந்து 166 மிமீ தொலைவில் உள்ள சியாட்டிக் நரம்பில் கதிரியக்கத்தன்மை அளவிடப்பட்டது. 10 மணி நேரத்திற்கும் மேலாக, உட்செலுத்தப்பட்ட இடத்தில் கதிரியக்கத்தின் உச்சநிலை சற்று மாறியது. ஆனால் கதிரியக்க அலையானது 2 மணி நேரத்தில் சுமார் 34 மிமீ அல்லது ஒரு நாளைக்கு 410 மிமீ என்ற நிலையான வேகத்தில் ஆக்ஸானுடன் பரவுகிறது. ஹோமியோதெர்மிக் விலங்குகளின் அனைத்து நியூரான்களிலும், வேகமான அச்சுப் போக்குவரத்து ஒரே வேகத்தில் நிகழ்கிறது, மேலும் மெல்லிய, அன்மைலினேட்டட் இழைகள் மற்றும் தடிமனான ஆக்சான்கள் மற்றும் மோட்டார் மற்றும் உணர்ச்சி இழைகளுக்கு இடையில் குறிப்பிடத்தக்க வேறுபாடுகள் எதுவும் காணப்படவில்லை. கதிரியக்க குறிப்பான் வகை வேகமான அச்சுப் போக்குவரத்தின் வீதத்தையும் பாதிக்காது; குறிப்பான்கள் பலவிதமான கதிரியக்கமாக செயல்பட முடியும்

அரிசி. 1.13.தசை அல்லாத மயோசின் வளாகம், ஒரு குறிப்பிட்ட நோக்குநிலையுடன், வெவ்வேறு துருவமுனைப்பின் ஆக்டின் இழைகளுடன் பிணைக்க முடியும், மேலும் ஏடிபியின் ஆற்றலைப் பயன்படுத்தி, அவற்றை ஒன்றுக்கொன்று மாற்றியமைக்க முடியும்.
நியூரானின் செல் உடலின் புரதங்களில் சேர்க்கப்பட்டுள்ள பல்வேறு அமினோ அமிலங்கள் போன்ற மூலக்கூறுகள். இங்கு கொண்டு செல்லப்படும் கதிரியக்கத்தின் கேரியர்களின் தன்மையை தீர்மானிக்க நரம்பின் புறப் பகுதியை பகுப்பாய்வு செய்தால், அத்தகைய கேரியர்கள் முக்கியமாக புரதப் பகுதியிலும், மத்தியஸ்தர்கள் மற்றும் இலவச அமினோ அமிலங்களின் கலவையிலும் காணப்படுகின்றன. இந்த பொருட்களின் பண்புகள் வேறுபட்டவை மற்றும் அவற்றின் மூலக்கூறுகளின் அளவுகள் குறிப்பாக வேறுபட்டவை என்பதை அறிந்தால், அவை அனைத்திற்கும் பொதுவான போக்குவரத்து பொறிமுறையால் மட்டுமே நிலையான போக்குவரத்தின் வேகத்தை விளக்க முடியும்.
மேலே விவரிக்கப்பட்ட வேகமான ஆக்சன் போக்குவரத்துஇருக்கிறது முன்னோடி,அதாவது, செல் உடலில் இருந்து இயக்கப்பட்டது. சில பொருட்கள் சுற்றளவில் இருந்து செல் உடலைப் பயன்படுத்தி செல்கின்றன என்று நிரூபிக்கப்பட்டுள்ளது பிற்போக்கு போக்குவரத்து.எடுத்துக்காட்டாக, அசிடைல்கொலினெஸ்டரேஸ் இந்த திசையில் வேகமான அச்சுப் போக்குவரத்தின் வேகத்தை விட 2 மடங்கு குறைவான வேகத்தில் கொண்டு செல்லப்படுகிறது. நியூரோஅனாடமியில் அடிக்கடி பயன்படுத்தப்படும் குறிப்பானான, குதிரைவாலி பெராக்ஸிடேஸ், பிற்போக்கு போக்குவரத்தின் மூலமும் நகரும். பிற்போக்கு போக்குவரத்து ஒருவேளை உயிரணு உடலில் புரதத் தொகுப்பைக் கட்டுப்படுத்துவதில் முக்கிய பங்கு வகிக்கிறது. ஆக்சன் வெட்டப்பட்ட சில நாட்களுக்குப் பிறகு, செல் உடலில் குரோமடோலிசிஸ் காணப்படுகிறது, இது புரதத் தொகுப்பின் சீர்குலைவைக் குறிக்கிறது. குரோமடோலிசிஸுக்குத் தேவைப்படும் நேரம், ஆக்சன் பரிமாற்றத்தின் தளத்திலிருந்து செல் உடலுக்குப் பிற்போக்கு போக்குவரத்தின் காலத்துடன் தொடர்புடையது. இந்த முடிவு இந்த கோளாறுக்கான விளக்கத்தையும் பரிந்துரைக்கிறது - புரதத் தொகுப்பைக் கட்டுப்படுத்தும் "சிக்னல் பொருளின்" சுற்றளவில் இருந்து பரவுவது சீர்குலைந்துள்ளது. முக்கிய "வாகனங்கள்" வேகமான அச்சுக்குப் பயன்படுத்தப்படுகின்றன என்பது வெளிப்படையானது
அரிசி. 1.14.பூனை சியாட்டிக் நரம்பின் உணர்திறன் இழைகளில் விரைவான அச்சுப் போக்குவரத்தை நிரூபிக்கும் சோதனை. டிரிடியம்-லேபிளிடப்பட்ட லியூசின் முதுகு வேர் கேங்க்லியனுக்குள் செலுத்தப்படுகிறது மற்றும் கேங்க்லியனில் உள்ள கதிரியக்கத்தன்மை மற்றும் உணர்ச்சி இழைகள் உட்செலுத்தப்பட்ட 2, 4, 6, 8 மற்றும் 10 மணிநேரங்களில் அளவிடப்படுகிறது. (படத்தின் கீழே).மூலம் x-அச்சுகேங்க்லியனில் இருந்து சியாட்டிக் நரம்பின் பகுதிகளுக்கு அளவீடு செய்யப்படும் தூரம் திட்டமிடப்பட்டுள்ளது. மேல் மற்றும் கீழ் வளைவுகளுக்கு மட்டும் ஆர்டினேட் அச்சில், கதிரியக்கம் (துடிப்புகள்/நிமிடம்) மடக்கை அளவில் வரையப்படுகிறது. அதிகரித்த கதிரியக்கத்தின் "அலை" (அம்புகள்) 410 மிமீ/நாள் வேகத்தில் நகரும் (உடன்)
போக்குவரத்து ஆகும் வெசிகல்ஸ் (வெசிகல்ஸ்) மற்றும் உறுப்புகள்,மைட்டோகாண்ட்ரியா போன்றவை, கடத்தப்பட வேண்டிய பொருட்களைக் கொண்டிருக்கும். நுண்ணோக்கியைப் பயன்படுத்தி மிகப்பெரிய வெசிகிள்ஸ் அல்லது மைட்டோகாண்ட்ரியாவின் இயக்கத்தைக் காணலாம்உயிருள்ள . இத்தகைய துகள்கள் ஒரு திசையில் குறுகிய, விரைவான இயக்கங்களை உருவாக்குகின்றன, நிறுத்துகின்றன, அடிக்கடி சற்று பின்னோக்கி அல்லது பக்கமாக நகர்கின்றன, மீண்டும் நிறுத்துகின்றன, பின்னர் முக்கிய திசையில் இழுக்கப்படுகின்றன. 410 மிமீ/நாள் என்பது ஏறத்தாழ 5 மைக்ரான்/வி ஆண்டிரோகிரேட் இயக்கத்தின் சராசரி வேகத்தை ஒத்துள்ளது; எனவே, ஒவ்வொரு தனிப்பட்ட இயக்கத்தின் வேகமும் மிக அதிகமாக இருக்க வேண்டும், மேலும் உறுப்புகள், இழைகள் மற்றும் நுண்குழாய்களின் அளவுகளை நாம் கணக்கில் எடுத்துக் கொண்டால், இந்த இயக்கங்கள் உண்மையில் மிக வேகமாக இருக்கும். வேகமான அச்சுப் போக்குவரத்துக்கு ஏடிபியின் குறிப்பிடத்தக்க செறிவு தேவைப்படுகிறது. நுண்குழாய்களை சீர்குலைக்கும் கொல்கிசின் போன்ற விஷங்கள், வேகமான அச்சுப் போக்குவரத்தையும் தடுக்கின்றன. இதிலிருந்து நாம் கருதும் போக்குவரத்துச் செயல்பாட்டில், வெசிகல்கள் மற்றும் உறுப்புகள் நுண்குழாய்கள் மற்றும் ஆக்டின் இழைகளுடன் நகர்கின்றன; இந்த இயக்கம் டைனைன் மற்றும் மயோசின் மூலக்கூறுகளின் சிறிய திரட்டுகளால் வழங்கப்படுகிறது, படத்தில் காட்டப்பட்டுள்ளபடி செயல்படுகிறது. 1.13, ஏடிபி ஆற்றலைப் பயன்படுத்துதல்.
வேகமான ஆக்சன் போக்குவரத்தும் இதில் ஈடுபடலாம் நோயியல் செயல்முறைகள்.சில நியூரோட்ரோபிக் வைரஸ்கள் (உதாரணமாக, ஹெர்பெஸ் அல்லது போலியோ வைரஸ்கள்) சுற்றளவில் உள்ள ஆக்சனுக்குள் நுழைந்து, பிற்போக்கு போக்குவரத்தின் மூலம் நியூரானின் உடலுக்குள் செல்கின்றன, அங்கு அவை பெருகி, அவற்றின் நச்சு விளைவுகளை ஏற்படுத்துகின்றன. டெட்டனஸ் டாக்சின், தோல் சேதமடையும் போது உடலில் நுழையும் பாக்டீரியாவால் உற்பத்தி செய்யப்படும் புரதம், நரம்பு முனைகளால் கைப்பற்றப்பட்டு நியூரானின் உடலுக்கு கொண்டு செல்லப்படுகிறது, அங்கு அது சிறப்பியல்பு தசைப்பிடிப்புகளை ஏற்படுத்துகிறது. ஆக்ஸான் போக்குவரத்தில் நச்சு விளைவுகளின் அறியப்பட்ட நிகழ்வுகள் உள்ளன, எடுத்துக்காட்டாக, தொழில்துறை கரைப்பான் அக்ரிலாமைடுக்கு வெளிப்பாடு. கூடுதலாக, வைட்டமின் குறைபாடு பெரிபெரி மற்றும் ஆல்கஹால் பாலிநியூரோபதியின் நோய்க்கிருமி உருவாக்கம் வேகமான அச்சுப் போக்குவரத்தை மீறுவதாக நம்பப்படுகிறது.
கலத்தில் வேகமான அச்சுப் போக்குவரத்துக்கு கூடுதலாக, மிகவும் தீவிரமானது மெதுவான ஆக்சன் போக்குவரத்து.டூபுலின் 1 மிமீ/நாள் வேகத்தில் ஆக்ஸனுடன் நகர்கிறது, மேலும் ஆக்டின் வேகமாக நகரும் - 5 மிமீ/நாள் வரை. மற்ற புரதங்கள் இந்த சைட்டோஸ்கெலிட்டல் கூறுகளுடன் இடம்பெயர்கின்றன; எடுத்துக்காட்டாக, என்சைம்கள் ஆக்டின் அல்லது டூபுலின் உடன் தொடர்புடையதாகத் தோன்றுகிறது. டூபுலின் மற்றும் ஆக்டினின் இயக்கத்தின் விகிதங்கள் முன்பு விவரிக்கப்பட்ட பொறிமுறையின் வளர்ச்சி விகிதங்களுடன் தோராயமாக ஒத்துப்போகின்றன, அங்கு மூலக்கூறுகள் ஒரு நுண்குழாய் அல்லது மைக்ரோஃபிலமென்ட்டின் செயலில் உள்ள முனையில் இணைக்கப்படுகின்றன. எனவே, இந்த பொறிமுறையானது மெதுவான அச்சுப் போக்குவரத்திற்கு அடிபணியலாம். மெதுவான அச்சுப் போக்குவரத்தின் வீதமும் தோராயமாக ஆக்சன் வளர்ச்சி விகிதத்துடன் ஒத்துப்போகிறது, இது இரண்டாவது செயல்பாட்டில் சைட்டோஸ்கெலட்டனின் கட்டமைப்பால் விதிக்கப்பட்ட கட்டுப்பாடுகளை வெளிப்படையாகக் குறிக்கிறது.
இந்த பிரிவின் முடிவில், செல்கள் எந்த வகையிலும் நிலையான கட்டமைப்புகள் அல்ல என்பதை வலியுறுத்த வேண்டும், எடுத்துக்காட்டாக, எலக்ட்ரான் நுண்ணிய புகைப்படங்களில் அவை தோன்றும். பிளாஸ்மா சவ்வுமற்றும் குறிப்பாக உறுப்புகள் நிலையான விரைவான இயக்கம் மற்றும் நிலையான மறுசீரமைப்பு;அதனால்தான் அவர்களால் செயல்பட முடிகிறது. மேலும், இவை இரசாயன எதிர்வினைகள் நடைபெறும் எளிய அறைகள் அல்ல, ஆனால் மிகவும் ஒழுங்கமைக்கப்பட்ட சவ்வுகள் மற்றும் இழைகள்,இதில் எதிர்வினைகள் உகந்த முறையில் ஒழுங்கமைக்கப்பட்ட வரிசையில் நிகழும்.
1.4 செல்லுலார் செயல்பாடுகளை ஒழுங்குபடுத்துதல்
ஒரு செயல்பாட்டு அலகு என தனிப்பட்ட செல் பராமரிப்பு முதன்மையாக கருவால் கட்டுப்படுத்தப்படுகிறது; இத்தகைய ஒழுங்குமுறை வழிமுறைகளின் ஆய்வு செல் உயிரியல் மற்றும் உயிர் வேதியியல் பாடமாகும். அதே நேரத்தில், செல்கள் சுற்றுச்சூழல் நிலைமைகள் மற்றும் உடலின் பிற உயிரணுக்களின் தேவைகளுக்கு ஏற்ப அவற்றின் செயல்பாடுகளை மாற்றியமைக்க வேண்டும், அதாவது அவை செயல்பாட்டு ஒழுங்குமுறையின் பொருள்களாக செயல்படுகின்றன. இந்த ஒழுங்குமுறை தாக்கங்கள் பிளாஸ்மா மென்படலத்தில் எவ்வாறு செயல்படுகின்றன மற்றும் அவை உள்செல்லுலார் உறுப்புகளை எவ்வாறு அடைகின்றன என்பதை கீழே சுருக்கமாகக் கருதுவோம்.
செல் சவ்வு மீது ஒழுங்குமுறை விளைவுகள்
சவ்வு திறன் . பல சந்தர்ப்பங்களில், செல்லுலார் செயல்பாடுகளின் கட்டுப்பாடு சவ்வு திறனை மாற்றுவதன் மூலம் மேற்கொள்ளப்படுகிறது. சாத்தியமான உள்ளூர் மாற்றங்கள் சாத்தியமாகும்: 1) கலத்தின் அருகிலுள்ள பகுதியிலிருந்து மின்னோட்டம் அல்லது மற்றொரு கலத்தால் உருவாக்கப்படுவது சவ்வு வழியாக பாய்கிறது; 2) அயனிகளின் வெளிப்புற செறிவு மாறுகிறது (பெரும்பாலும் [K +]வெளியே ); 3) சவ்வு அயன் சேனல்கள் திறக்கப்படுகின்றன. சவ்வு ஆற்றலில் ஏற்படும் மாற்றங்கள் சவ்வு புரதங்களின் இணக்கத்தை பாதிக்கலாம், குறிப்பாக சேனல்கள் திறக்க அல்லது மூடுவதற்கு காரணமாகிறது. மேலே விவரிக்கப்பட்டுள்ளபடி, சில சவ்வு குழாய்களின் செயல்பாடு சவ்வு திறனைப் பொறுத்தது. நரம்பு செல்கள் சவ்வு ஆற்றலில் ஏற்படும் மாற்றங்களை செயலாக்க மற்றும் கடத்தப்பட வேண்டிய தகவலாக உணர சிறப்பு வாய்ந்தது (அத்தியாயம் 2 ஐப் பார்க்கவும்).
புற-செல்லுலர் ஒழுங்குமுறை பொருட்கள் . பிளாஸ்மா சவ்வு அல்லது செல்லின் உள்ளே உள்ள குறிப்பிட்ட ஏற்பிகளுடன் அவற்றின் தொடர்பு என்பது புற-செல்லுலர் பொருட்களை உள்ளடக்கிய மிக முக்கியமான ஒழுங்குமுறை பொறிமுறையாகும். இந்த பொருட்களில் சினாப்டிக் டிரான்ஸ்மிட்டர்கள் அடங்கும், அவை நரம்பு செல்கள், உள்ளூர் முகவர்கள் மற்றும் இரத்தத்தில் சுற்றும் மற்றும் ஹார்மோன்கள் மற்றும் ஆன்டிஜென்கள் போன்ற உடலின் அனைத்து செல்களையும் அடையும் பொருட்களுக்கு இடையே தகவல்களை அனுப்புகின்றன. சினாப்டிக் டிரான்ஸ்மிட்டர்கள்சினாப்ஸில் உள்ள நரம்பு முடிவுகளிலிருந்து வெளியிடப்படும் சிறிய மூலக்கூறுகள்;
அவை அண்டை, போஸ்ட்னாப்டிக் கலத்தின் பிளாஸ்மா மென்படலத்தை அடையும் போது, அவை மின் சமிக்ஞைகள் அல்லது பிற ஒழுங்குமுறை வழிமுறைகளைத் தூண்டுகின்றன. இந்த பிரச்சினை அத்தியாயத்தில் விரிவாக விவாதிக்கப்படுகிறது. 3.
உள்ளூர் இரசாயன முகவர்கள் பெரும்பாலும் சிறப்பு செல்கள் மூலம் சுரக்கப்படுகிறது. அவை புற-செல்லுலார் இடத்தில் சுதந்திரமாக பரவுகின்றன, ஆனால் இந்த பொருட்களின் விரைவான அழிவு காரணமாக, தன்னிச்சையாக அல்லது என்சைம்களின் செயல்பாட்டின் கீழ் அவற்றின் செயல்பாடு ஒரு சிறிய குழு செல்களுக்கு மட்டுமே. அத்தகைய முகவர்களை விடுவிப்பதற்கான ஒரு எடுத்துக்காட்டு வெளியீடு ஹிஸ்டமின்சேதம் அல்லது நோய் எதிர்ப்பு சக்தி காரணமாக மாஸ்ட் செல்கள். ஹிஸ்டமைன் வாஸ்குலர் மென்மையான தசை செல்களை தளர்த்துகிறது, வாஸ்குலர் எண்டோடெலியத்தின் ஊடுருவலை அதிகரிக்கிறது மற்றும் அரிப்பு உணர்வை மத்தியஸ்தம் செய்யும் உணர்ச்சி நரம்பு முடிவுகளை தூண்டுகிறது. பிற உள்ளூர் இரசாயன முகவர்கள் பல செல்களால் வெளியிடப்படுகின்றன. வழக்கமான உள்ளூர் முகவர்கள் புரோஸ்டாக்லாண்டின்கள்,தோராயமாக 20 கொழுப்பு அமில வழித்தோன்றல்களின் குழுவை உருவாக்குகிறது. அவை பரவலாக விநியோகிக்கப்பட்ட உயிரணுக்களிலிருந்து தொடர்ச்சியாக வெளியிடப்படுகின்றன, ஆனால் அவை உள்நாட்டில் மட்டுமே செயல்படுகின்றன, ஏனெனில் அவை சவ்வு பாஸ்போலிபேஸ்களால் விரைவாக அழிக்கப்படுகின்றன. பல்வேறு புரோஸ்டாக்லாண்டின்கள் பரந்த அளவிலான செயல்பாட்டைக் கொண்டுள்ளன: அவை மென்மையான தசை செல்கள் சுருங்குவதைத் தூண்டலாம், இரத்த பிளேட்லெட்டுகளை (பிளேட்லெட்டுகள்) திரட்டலாம் அல்லது கருப்பையில் கார்பஸ் லியூடியத்தின் வளர்ச்சியை அடக்கலாம்.
பிற உள்ளூர் முகவர்கள் சேவை செய்கிறார்கள் வளர்ச்சி காரணிகள்.நன்கு அறியப்பட்ட நரம்பு வளர்ச்சி காரணி (NGF) அனுதாப நியூரான்களுக்கானது, இது வளர்ச்சியின் போது இந்த நியூரான்களின் வளர்ச்சி மற்றும் உயிர்வாழ்வதற்கு அவசியம்.உயிருள்ள அல்லது செல் கலாச்சாரத்தில். வெளிப்படையாக, இந்த வகை நியூரான்களுக்கான இலக்கு செல்கள் NGF ஐ சுரக்கின்றன மற்றும் அதன் மூலம் சரியான கண்டுபிடிப்பை உறுதி செய்கின்றன. உறுப்புகளை உருவாக்கும் போது, செல்கள் பெரும்பாலும் கணிசமான தொலைவில் அமைந்துள்ள செல்களை இலக்காகக் கொள்ள "தங்கள் வழியைக் கண்டுபிடிக்க" வேண்டும். அதன்படி, NGF போன்ற பல சிறப்பு வளர்ச்சி காரணிகள் இருக்க வேண்டும்.
ஹார்மோன்கள் மற்றும் ஆன்டிஜென்கள் அனைத்து செல்களுக்கும் இரத்தம் கொண்டு செல்லப்படுகிறது. ஆன்டிஜென்கள் குறிப்பிட்ட ஆன்டிபாடிகளைக் கொண்ட செல்களிலிருந்து நோயெதிர்ப்பு மறுமொழியைத் தூண்டுகின்றன. இருப்பினும், ஆன்டிஜென்கள், ஒரு விதியாக, எதிர்வினை உயிரினத்தில் உற்பத்தி செய்யப்படாத வெளிநாட்டு பொருட்கள் (மேலும் விவரங்களுக்கு, அத்தியாயம் 18 ஐப் பார்க்கவும்). இன்சுலின் அல்லது தைராக்ஸின் போன்ற சில ஹார்மோன்கள் பல்வேறு வகையான செல் வகைகளை பாதிக்கின்றன, மற்றவை, பாலியல் ஹார்மோன்கள் போன்றவை, ஒரு குறிப்பிட்ட வகை செல்களை மட்டுமே பாதிக்கின்றன. ஹார்மோன்கள் பெப்டைடுகள் ஆகும், அதன் செயல் உயிரணு சவ்வில் உள்ள ஏற்பியுடன் பிணைப்பதன் மூலம் தூண்டப்படுகிறது, அல்லது ஸ்டெராய்டுகள் மற்றும் தைராக்சின், அவை லிப்பிட் சவ்வு வழியாக பரவி உள்செல்லுலர் ஏற்பிகளுடன் பிணைக்கப்படுகின்றன. ஸ்டீராய்டு ஹார்மோன்கள் நியூக்ளியர் குரோமாடினுடன் பிணைக்கப்படுகின்றன, இதன் விளைவாக சில மரபணுக்களின் படியெடுத்தல் ஏற்படுகிறது. இதன் விளைவாக புரதங்கள் செல்லுலார் செயல்பாடுகளில் மாற்றங்களை ஏற்படுத்துகின்றன, இது ஹார்மோன்களின் குறிப்பிட்ட விளைவு ஆகும். ஹார்மோன்களின் வெளியீடு மற்றும் செயல்பாடு தொடர்பான பிரச்சினைகள் அத்தியாயத்தில் விரிவாக விவாதிக்கப்பட்டுள்ளன. 17.
இரண்டாவது தூதர்களை உள்ளடக்கிய செல்களுக்குள் தொடர்பு
மேலே விவரிக்கப்பட்ட ஒழுங்குமுறை செயல்பாடுகளில் செல் சவ்வு மீதான விளைவுகள் அடங்கும். உயிரணு சவ்வு மூலம் பெறப்பட்ட தகவல்கள் பெரும்பாலும் உறுப்புகளை வினைபுரியச் செய்ய வேண்டும் மற்றும் இரண்டாவது தூதர்கள் எனப்படும் பல்வேறு பொருட்களால் அவற்றுக்கு மாற்றப்படும் (முதல் தூதுவர்கள், வெளிப்புற மூலங்களிலிருந்து கலத்திற்கு வரும்). இரண்டாவது மத்தியஸ்தர்களின் ஆய்வு வேகமாக உருவாகி வருகிறது, மேலும் சிக்கலைப் பற்றிய தற்போதைய புரிதல் போதுமானதாக இருக்கும் என்பதற்கு எந்த உத்தரவாதமும் இல்லை. இங்கே நாம் நன்கு படித்த மூன்று மத்தியஸ்தர்களைத் தொடுவோம்: Ca 2+ , cAMP மற்றும் inositol triphosphate.
கால்சியம்.எளிமையான உள்செல்லுலார் மத்தியஸ்தர் Ca 2+ அயன் ஆகும். ஓய்வெடுக்கும் கலத்தில் அதன் இலவச செறிவு மிகவும் குறைவாக உள்ளது மற்றும் 10_–8–10–7 mol/l ஆக இருக்கும். குறிப்பிட்ட சவ்வு சேனல்கள் திறந்த நிலையில் இருக்கும் போது, எடுத்துக்காட்டாக, சவ்வு சாத்தியம் மாறும்போது (அத்தியாயம் 2 ஐப் பார்க்கவும்) செல்லுக்குள் நுழைய முடியும். இதன் விளைவாக Ca 2+ செறிவு அதிகரிப்பு, தசைச் சுருக்கத்தின் அடிப்படையான myofibrils சுருங்குதல் (அத்தியாயம் 4) அல்லது நரம்பு நுனிகளில் இருந்து நரம்பியக்கடத்திகளைக் கொண்ட வெசிகல்களின் வெளியீடு போன்ற முக்கியமான எதிர்வினைகளை உயிரணுவில் தூண்டுகிறது (அத்தியாயம் 3 ஐப் பார்க்கவும்) . இரண்டு எதிர்வினைகளுக்கும் தோராயமாக 10 –5 mol/l என்ற Ca 2+ செறிவு தேவைப்படுகிறது. ஒரு ஒழுங்குமுறை விளைவைக் கொண்ட Ca 2+, எண்டோபிளாஸ்மிக் ரெட்டிகுலம் போன்ற உள்செல்லுலார் கடைகளிலிருந்தும் வெளியிடப்படலாம். டிப்போவில் இருந்து Ca 2+ இன் வெளியீட்டிற்கு மற்ற இடைத்தரகர்களின் பங்கேற்பு தேவைப்படுகிறது (உதாரணமாக, படம் 1.16 ஐப் பார்க்கவும்).
சுழற்சி அடினோசின் மோனோபாஸ்பேட், cAMP. உடலின் முக்கிய ஆற்றல் மூலமான ஏடிபியின் வழித்தோன்றலான சுழற்சி அடினோசின் மோனோபாஸ்பேட் (சிஏஎம்பி) ஒரு முக்கியமான இரண்டாவது தூதுவர் என்பது சமீபத்தில் நிரூபிக்கப்பட்டுள்ளது. படத்தில் காட்டப்பட்டுள்ள எதிர்வினைகளின் சிக்கலான சங்கிலி. 1.15, ஏற்பியுடன் தொடங்குகிறதுஆர் எஸ் பிளாஸ்மா மென்படலத்தின் வெளிப்புற மேற்பரப்பில், இது பல்வேறு மத்தியஸ்தர்கள் மற்றும் ஹார்மோன்களுக்கான குறிப்பிட்ட பிணைப்பின் தளமாக செயல்படும். ஒரு குறிப்பிட்ட "தூண்டுதல்" மூலக்கூறுடன் பிணைக்கப்பட்ட பிறகுஆர் எஸ் அதன் இணக்கத்தை மாற்றுகிறது; இந்த மாற்றங்கள் புரதத்தை பாதிக்கின்றனஜி எஸ் மென்படலத்தின் உள் மேற்பரப்பில், உள்செல்லுலார் குவானோசின் ட்ரைபாஸ்பேட் (ஜிடிபி) மூலம் பிந்தையதை செயல்படுத்துவது சாத்தியமாகும். செயல்படுத்தப்பட்ட புரதம்ஜி எஸ் , இதையொட்டி, சவ்வின் உள் மேற்பரப்பில் உள்ள நொதியைத் தூண்டுகிறது, அடினிலேட் சைக்லேஸ் (ஏசி), இது ஏடிபியிலிருந்து சிஏஎம்பி உருவாவதை ஊக்குவிக்கிறது. நீரில் கரையக்கூடிய cAMP என்பது விளைவை கடத்தும் மத்தியஸ்தர் ஆகும்

அரிசி. 1.15உள்செல்லுலர் மெசஞ்சர் cAMP (சுழற்சி அடினோசின் மோனோபாஸ்பேட்) சம்பந்தப்பட்ட எதிர்வினைகளின் சங்கிலி. தூண்டுதல் அல்லது தடுக்கும் வெளிப்புற சமிக்ஞைகள் சவ்வு ஏற்பிகளை செயல்படுத்துகிறது Rகள் அல்லது ரி . இந்த ஏற்பிகள் பிணைப்பு செயல்முறையை ஒழுங்குபடுத்துகின்றனஜி - உள்செல்லுலார் ஜிடிபி (குவானோசின் ட்ரைபாஸ்பேட்) கொண்ட புரதங்கள், இதன் மூலம் உள்செல்லுலார் அடினிலேட் சைக்லேஸை (ஏசி) தூண்டுகிறது அல்லது தடுக்கிறது. பெருக்கும் என்சைம் AC அடினோசின் ட்ரைபாஸ்பேட்டை (ATP) cAMP ஆக மாற்றுகிறது, பின்னர் இது பாஸ்போடிஸ்டேரேஸ் மூலம் AMP ஆக உடைக்கப்படுகிறது. இலவச cAMP கலத்திற்குள் பரவுகிறது மற்றும் அடினிலேட் கைனேஸை (A-கைனேஸ்) செயல்படுத்துகிறது, அதன் வினையூக்க துணைக்குழு C ஐ வெளியிடுகிறது, இது உள்செல்லுலார் புரதங்களின் பாஸ்போரிலேஷனை ஊக்குவிக்கிறது, அதாவது. எக்ஸ்ட்ராசெல்லுலர் தூண்டுதலின் இறுதி விளைவை உருவாக்குகிறது. சில எதிர்விளைவுகளைத் தூண்டும் (+) அல்லது தடுக்கும் (-) மருந்தியல் மருந்துகள் மற்றும் நச்சுப் பொருட்களையும் வரைபடம் காட்டுகிறது (மாற்றியமைக்கப்பட்டது)
எக்ஸ்ட்ராசெல்லுலர் ஏற்பி தூண்டுதல்ஆர் எஸ் செல்லின் உள் கட்டமைப்புகளுக்கு.
சம்பந்தப்பட்ட எதிர்வினைகளின் தூண்டுதல் சங்கிலிக்கு இணையாகஆர் எஸ் தடுப்பு மத்தியஸ்தர்கள் மற்றும் ஹார்மோன்களை தொடர்புடைய ஏற்பியுடன் பிணைப்பது சாத்தியமாகும்ஆர் ஐ மீண்டும் GTP-செயல்படுத்தப்பட்ட புரதம் மூலம்ஜி , AC ஐ தடுக்கிறது, இதனால் cAMP உற்பத்தி செய்யப்படுகிறது. கலத்திற்குள் பரவுகிறது, cAMP அடினிலேட் கைனேஸுடன் (A-kinase) வினைபுரிகிறது; இது துணைக்குழு C ஐ வெளியிடுகிறது, இது புரதம் P இன் பாஸ்போரிலேஷனை ஊக்குவிக்கிறது.இந்த பாஸ்போரிலேஷன் புரதங்களை செயலில் உள்ள வடிவமாக மாற்றுகிறது, மேலும் அவை இப்போது அவற்றின் குறிப்பிட்ட ஒழுங்குமுறை விளைவுகளைச் செலுத்தலாம் (உதாரணமாக, கிளைகோஜன் சிதைவை ஏற்படுத்தும்). இந்த சிக்கலான ஒழுங்குமுறை அமைப்பு மிகவும் திறமையானது, ஏனெனில் இறுதி முடிவு பல புரதங்களின் பாஸ்போரிலேஷன் ஆகும், அதாவது, ஒழுங்குமுறை சமிக்ஞை சுற்று வழியாக அதிக லாபத்துடன் பயணிக்கிறது. ஏற்பிகளுடன் பிணைக்கும் வெளிப்புற மத்தியஸ்தர்கள்ஆர் எஸ் மற்றும் ஆர் ஐ அவை ஒவ்வொன்றிற்கும் குறிப்பிட்ட, மிகவும் வேறுபட்டவை. அட்ரினலின், உடன் இணைக்கிறதுஆர் எஸ் அல்லது ஆர் ஐ லிப்பிட் மற்றும் கிளைகோஜன் வளர்சிதை மாற்றத்தை ஒழுங்குபடுத்துவதில் பங்கேற்கிறது, அதே போல் இதய தசையின் சுருக்கம் மற்றும் பிற எதிர்விளைவுகளை மேம்படுத்துகிறது (அத்தியாயம் 19 ஐப் பார்க்கவும்). தைராய்டு-தூண்டுதல் ஹார்மோன், செயல்படுத்துகிறது ஆர் எஸ் , தைராய்டு சுரப்பி மூலம் தைராக்ஸின் என்ற ஹார்மோனின் வெளியீட்டைத் தூண்டுகிறது, மேலும் ப்ரோஸ்டாக்லாண்டின் I இரத்த பிளேட்லெட்டுகளின் திரட்டலைத் தடுக்கிறது. அட்ரினலின் உள்ளிட்ட தடுப்பு விளைவுகள் மூலம் மத்தியஸ்தம் செய்யப்பட்டதுஆர் ஐ லிபோலிசிஸை மெதுவாக்குவதில் வெளிப்படுத்தப்படுகிறது. இதனால், cAMP அமைப்பு ஒரு மல்டிஃபங்க்ஸ்னல் இன்ட்ராசெல்லுலர் ரெகுலேட்டரி சிஸ்டம்,எக்ஸ்ட்ராசெல்லுலர் தூண்டுதல் மற்றும் தடுப்பு சமிக்ஞை பொருட்கள் மூலம் துல்லியமாக கட்டுப்படுத்த முடியும்.
இனோசிட்டால் பாஸ்பேட் "IF" ம " இரண்டாவது தூதரின் உள்ளக அமைப்பு, இனோசிட்டால் பாஸ்பேட், சமீபத்தில்தான் கண்டுபிடிக்கப்பட்டது (படம் 1.16). இந்த வழக்கில், தடுப்பு பாதை இல்லை, ஆனால் சிஏஎம்பி அமைப்புக்கு ஒரு ஒற்றுமை உள்ளது, இதில் ஆர் ஏற்பியின் தூண்டுதலின் விளைவு சவ்வின் உள் மேற்பரப்பில் ஜிடிபி-செயல்படுத்தப்பட்ட ஜி புரதத்திற்கு மாற்றப்படுகிறது. அடுத்த கட்டத்தில், வழக்கமான சவ்வு லிப்பிட் பாஸ்பாடிடிலினோசிட்டால் (PI), முன்பு இரண்டு கூடுதல் பாஸ்பேட் குழுக்களைப் பெற்றுள்ளது, இது PI-டைபாஸ்பேட்டாக (PIP 2) மாற்றப்படுகிறது, இது செயல்படுத்தப்பட்ட பாஸ்போடிஸ்டேரேஸ் (PDE) மூலம் பிளவுபடுத்தப்படுகிறது. இனோசிட்டால் ட்ரைபாஸ்பேட்(IFz) மற்றும் லிப்பிட் டயசில்கிளிசரால்(டிஏஜி) இனோசிட்டால் ட்ரைபாஸ்பேட் என்பது நீரில் கரையக்கூடிய இரண்டாவது தூதுவர், இது சைட்டோசோலில் பரவுகிறது. இது முதன்மையாக எண்டோபிளாஸ்மிக் ரெட்டிகுலத்தில் இருந்து Ca 2+ ஐ வெளியிடுவதன் மூலம் செயல்படுகிறது. மேலே விவரிக்கப்பட்டுள்ளபடி Ca 2+ ஒரு தூதராக செயல்படுகிறது; எடுத்துக்காட்டாக, இது Ca 2+ -சார்ந்த பாஸ்போகினேஸை செயல்படுத்துகிறது, இது என்சைம்களை பாஸ்போரிலேட் செய்கிறது. DAG இன் லிப்பிட் சப்யூனிட் (படம். 1.16) சிக்னலைக் கொண்டு செல்கிறது, பிளாஸ்மா மென்படலத்தின் லிப்பிட் கட்டத்தில் அதன் உள் மேற்பரப்பில் அமைந்துள்ள சி-கைனேஸுக்கு பரவுகிறது, இது பாஸ்பாடிடைல்செரின் பங்கேற்புடன் செயல்படுத்தப்படுகிறது. பின்னர் சி-கைனேஸ் புரதங்களின் பாஸ்போரிலேஷனைத் தூண்டுகிறது, அவற்றை செயலில் உள்ள வடிவமாக மாற்றுகிறது.
அசிடைல்கொலின், செரோடோனின், வாசோபிரசின் மற்றும் தைராய்டு-தூண்டுதல் ஹார்மோன் உள்ளிட்ட பல்வேறு வெளிப்புற மத்தியஸ்தர்கள் மற்றும் ஹார்மோன்கள் மூலம் உள்செல்லுலார் இரண்டாவது மெசஞ்சர் அமைப்பு IF 3ஐயும் கட்டுப்படுத்தலாம்; cAMP அமைப்பைப் போலவே, இது பல்வேறு உள்செல்லுலர் விளைவுகளால் வகைப்படுத்தப்படுகிறது. இந்த அமைப்பு கண்ணின் காட்சி ஏற்பியில் ஒளியினால் செயல்படுத்தப்பட்டு ஒளிமாற்றத்தில் முக்கிய பங்கு வகிக்கிறது (அத்தியாயம் 11 ஐப் பார்க்கவும்). உயிரினத்தின் தனிப்பட்ட வளர்ச்சியில் முதன்முறையாக, IG அமைப்பின் ஏற்பி விந்தணுக்களால் செயல்படுத்தப்படுகிறது, இதன் விளைவாக IG முட்டையின் கருத்தரிப்புடன் கூடிய ஒழுங்குமுறை எதிர்வினைகளில் பங்கேற்கிறது.
cAMP மற்றும் IFz-DAG அமைப்புகள் மிகவும் பயனுள்ளதாக இருக்கும் உயிரியல் மேம்படுத்திகள்.அவர்கள்

அரிசி. 1.16.உள்செல்லுலார் மத்தியஸ்தர் ஐபிஜி (இனோசிட்டால் ட்ரைபாஸ்பேட்) சம்பந்தப்பட்ட எதிர்வினைகளின் சங்கிலி. cAMP அமைப்பைப் போலவே, புற-செல்லுலார் சமிக்ஞை ஒரு புரதத்தின் மூலம் மத்தியஸ்தம் செய்யப்படுகிறதுஜி, இந்த வழக்கில் பாஸ்போடிஸ்டேரேஸை (PDE) செயல்படுத்துகிறது. இந்த நொதி பாஸ்பாடிடிலினோசின் டைபாஸ்பேட்டை உடைக்கிறது (PIP 2 ) IF க்கு முன் பிளாஸ்மா மென்படலத்தில்ம மற்றும் டயசில்கிளிசரால் (DAG); IFம சைட்டோபிளாஸில் பரவுகிறது. இங்கே அது Ca வெளியீட்டை ஏற்படுத்துகிறது 2+ எண்டோபிளாஸ்மிக் ரெட்டிகுலத்தில் இருந்து; Ca செறிவு அதிகரிப்பு 2+ சைட்டோபிளாஸில் ([Ca 2+ ]ஐ ) புரோட்டீன் கைனேஸை செயல்படுத்துகிறது, இது பாஸ்போரிலேட்டுகள் மற்றும் அதனால் என்சைம்களை செயல்படுத்துகிறது. மற்றொரு தயாரிப்பு, DAG, மென்படலத்தில் உள்ளது மற்றும் புரோட்டீன் கைனேஸ் சி (பாஸ்பாடிடைல்செரின் கோஃபாக்டர், PS) செயல்படுத்துகிறது. புரோட்டீன் கைனேஸ் சி, வெளிப்புற ஏற்பி தூண்டுதலுடன் தொடர்புடைய குறிப்பிட்ட செயல்களுக்கு மத்தியஸ்தம் செய்யும் என்சைம்களையும் பாஸ்போரிலேட் செய்கிறது.ஆர் . IF ஐ உள்ளடக்கிய எதிர்வினைகளின் சங்கிலியின் கிளைகள்ம மற்றும் DAG ஆனது முறையே அயனோமைசின் மற்றும் போர்போல் எஸ்டர் மூலம் சுயாதீனமாக செயல்படுத்தப்படலாம் (மாற்றியமைக்கப்பட்டது)
ஒரு டிரான்ஸ்மிட்டர் மற்றும் வெளிப்புற சவ்வு ஏற்பிக்கு இடையேயான எதிர்வினையை பல்வேறு செல்களுக்குள் புரதங்களின் பாஸ்போரிலேஷனாக மாற்றுகிறது, இது பல்வேறு செல் செயல்பாடுகளை பாதிக்கலாம். பிரச்சனையின் குறிப்பிடத்தக்க அம்சங்களில் ஒன்று, இன்று அறியப்பட்ட வரையில், இந்த வகையின் இந்த இரண்டு நெருங்கிய தொடர்புடைய ஒழுங்குமுறை அமைப்புகள் மட்டுமே உள்ளன, இது பல்வேறு உள்செல்லுலார் செயல்முறைகளை ஒழுங்குபடுத்துவதற்கு ஏராளமான வெளிப்புற மத்தியஸ்தர்களால் பயன்படுத்தப்படுகிறது. அதே நேரத்தில், Ca 2+ உள்ளிட்ட இந்த ஒழுங்குமுறை அமைப்புகள், ஒருவருக்கொருவர் நெருக்கமாக தொடர்பு கொள்கின்றன, இது செல்லுலார் செயல்பாடுகளை நன்றாக ஒழுங்குபடுத்த அனுமதிக்கிறது.
பயிற்சிகள் மற்றும் வழிகாட்டிகள்
1. ஆல்பர்ட்ஸ் IN., ப்ரே டி., லூயிஸ் ஜே., ராஃப் எம்., ராபர்ட்ஸ் TO., வாட்சன் ஜே.டி.செல்களின் மூலக்கூறு உயிரியல், நியூயார்க் மற்றும் லண்டன், கார்லேண்ட் பப்ளிஷிங் இன்க்., 1983.
2. சிஹாக் ஜி., லாங்கர் எச்., ஜீக்லர் எச்.(பதிப்பு.). உயிரியல். பெர்லின், ஹைடெல்பெர்க், நியூயார்க், ஸ்பிரிங்கர், 1983.
3. ஹில்லே IN. உற்சாகமான சவ்வுகளின் அயனி சேனல்கள். சுந்தர்லேண்ட், மாஸ்., சினௌர் அசோக்., 1984.
4. ஹோப் டபிள்யூ., லோஹ்மன் டபிள்யூ.. மார்கி எச்., ஜீக்லர் எச்.(பதிப்பு.). உயிர் இயற்பியல். பெர்லின், ஹைடெல்பெர்க், நியூயார்க், ஸ்பிரிங்கர், 1984.
5. ஜங்கர்மேன் TO., மஹ்லர் எச்.உயிர்வேதியியல். பெர்லின், ஹைடெல்பெர்க், நியூயார்க், ஸ்பிரிங்கர், 1980.
6. காண்டல் ஈ.ஆர்., ஸ்வார்ட்ஸ்–ஜே.எச்.,(பதிப்பு.). நரம்பியல் அறிவியலின் கோட்பாடுகள், நியூயார்க், ஆம்ஸ்டர்டாம், ஆக்ஸ்போர்டு, எல்சேவியர், 1985.
7. ஸ்கீப்லர் டி. எச்., ஷ்மிட் டபிள்யூ.உடற்கூறியல் டெஸ் மென்சென். பெர்லின், ஹைடெல்பெர்க், நியூயார்க், டோக்கியோ, ஸ்பிரிங்கர், 1983.
அசல் கட்டுரைகள் மற்றும் மதிப்புரைகள்
8. பெரிட்ஜ் எம்.ஜே.உயிரணுவிற்குள் தொடர்புகொள்வதற்கான மூலக்கூறு அடிப்படை, அறிவியல். அமர், 253, 124-134 (1985).
9. பெரிட்ஜ் எம். ஜே., இர்வின் ஆர்.எஃப்.இனோசிட்டால் ட்ரைபாஸ்பேட், செல்லுலார் சிக்னல் கடத்தலில் ஒரு புதிய இரண்டாவது தூதுவர். இயற்கை, 312, 315 321 (1984).
10. பிரெட்சர் எம்.எஸ்.செல் சவ்வு மூலக்கூறுகள், Sci. அமர்., 253, 124–134 (1985).
11. டவுட் ஜே.ஆற்றல் கடத்தும் இயந்திரமாக வாழும் செல். மாரடைப்பு வளர்சிதை மாற்றத்தின் குறைந்தபட்ச மாதிரி, பயோகெம். மற்றும் உயிரியல்புகள். ஆக்டா, 895, 41–62 (1987).
12. ஹோட்கின் ஏ.எல்., காட்ஸ் IN. சோடியம் அயனிகளின் விளைவு ஸ்க்விட் ராட்சத ஆக்ஸானின் மின் செயல்பாட்டின் மீது. ஜே. பிசியோல். (லண்டன்.), 108, 37-77 (1949).
13. ஹோட்கின் ஏ.எல்., கெய்ன்ஸ் ஆர்.டி.ராட்சத அச்சுகளில் கேஷன்களின் செயலில் போக்குவரத்து செபியாமற்றும் லோலிகோ,ஜே. பிசியோல். (லண்டன்.), 128, 28-42 (1955).
14. நீண்ட பி.கன்ஃபார்மேஷனல் துணை நிலைகளுடன் கூடிய அயனி சேனல்கள், பயோஃபிஸ். ஜே., 47, 581–590 (1985).
15. ஓக்ஸ் எஸ்., வொர்த் பி.எம்.சாதாரண மற்றும் நோயியல் அமைப்புகளில் எக்ஸோபிளாஸ்மிக் போக்குவரத்து. இல்: ஆக்சன்களின் உடலியல் மற்றும் நோயியல், எஸ்.ஜி. வக்ஸ்மேன்,எட். நியூயார்க், ரேவன் பிரஸ், 1978.
மற்ற விளக்கக்காட்சிகளின் சுருக்கம்"உயிரியலைக் கற்பிக்கும் முறைகள்" - பள்ளி விலங்கியல். அறிவியல் விலங்கியல் தரவுகளைப் பயன்படுத்த மாணவர்களை அறிமுகப்படுத்துதல். தார்மீக கல்வி. கோழிப்பண்ணையின் கூடுதல் ஆசீர்வாதம். முறைகளின் தேர்வு. வாழ்க்கை செயல்முறைகள். மீன் மீன். ஊட்டச்சத்து. சுற்றுச்சூழல் கல்வி. வாழ்க்கை செயல்முறைகளின் பொருள். எதிர்மறையான முடிவுகள். மாணவர்களின் கவனத்திற்கு. கட்டாய வடிவம். சிறிய விலங்குகளைப் பார்ப்பது. உயிரியலின் குறிக்கோள்கள் மற்றும் நோக்கங்கள். கதை.
"உயிரியல் பாடங்களில் சிக்கல் அடிப்படையிலான கற்றல்" - அறிவு. புதிய பாடப்புத்தகங்கள். தீர்வுக்கான பாதை. பிரச்சனை. கருத்தரங்குகள். பணி என்றால் என்ன? ஆல்பிரெக்ட் டியூரர். உயிரியல் பாடங்களில் சிக்கல் அடிப்படையிலான கற்றல். தரமற்ற பாடங்கள். பிரச்சனை அடிப்படையிலான கற்றல் என்றால் என்ன. வாழ்க்கை தரம். உயிரியல் ஒரு கல்விப் பாடமாக. கேள்வி. சிக்கல் தீர்க்கும் பாடம். பாடத்தில் ஆர்வம் குறைந்தது. பிரச்சனை அடிப்படையிலான ஆய்வக வகுப்புகள்.
"உயிரியல் பாடங்களில் விமர்சன சிந்தனை" - "விமர்சன சிந்தனை" தொழில்நுட்பம். "விமர்சன சிந்தனையின் வளர்ச்சி" தொழில்நுட்பத்தைப் பயன்படுத்துதல். பாடத்திற்கான அட்டவணை. படிக்க உந்துதல். சுற்றுச்சூழல் அமைப்புகள். "விமர்சன சிந்தனையை வளர்ப்பது" என்பதன் பொருள். தொழில்நுட்பத்தின் அறிகுறிகள். RKM தொழில்நுட்பம். பாடத்தின் அமைப்பு. முக்கிய திசைகள். தொழில்நுட்பத்தின் வரலாறு. கல்வியியல் தொழில்நுட்பங்கள். தொழில்நுட்ப விதிகள். உயிரியல் பணிகள். ஒளிச்சேர்க்கை. பாடத்தின் வெவ்வேறு கட்டங்களில் பயன்படுத்தப்படும் நுட்பங்கள்.
"இன்டராக்டிவ் ஒயிட்போர்டுடன் உயிரியல் பாடங்கள்" - மின்னணு பாடப்புத்தகங்கள். மாணவர்களுக்கு நன்மைகள். ஊடாடும் ஒயிட் போர்டு ஒவ்வொரு மாணவருக்கும் தகவலை தெரிவிக்க உதவுகிறது. டிடாக்டிக் பணிகள். உயிரியல் சிக்கல்களைத் தீர்ப்பது. ஊடாடும் ஒயிட்போர்டுகளுடன் வேலை செய்வதன் நன்மைகள். விளக்கக்காட்சிகளுடன் பணிபுரிதல். பொருட்களை ஒப்பிடுவதில் வேலை செய்யுங்கள். நகரும் பொருள்கள். விரிதாள்களைப் பயன்படுத்துதல். பள்ளி மாணவர்களுக்கு கற்பிக்கும் செயல்பாட்டில் ஊடாடும் வெள்ளை பலகையைப் பயன்படுத்துதல். ஆசிரியர்களுக்கு நன்மைகள்.
“உயிரியலில் சிஸ்டம்-செயல்பாட்டு அணுகுமுறை” - கருத்தரங்கு கேள்விகள். செயல்பாட்டு முறை. டிரையோபிதேகஸ். மனித தோற்றத்தின் வேற்று கிரக பாதை. லைசோசோம்கள். இரசாயன அமைப்பு. ஜிம்னோஸ்பெர்ம்கள். வளர்சிதை மாற்றம். பகுப்பாய்விகள். உயிரியலைக் கற்பிப்பதற்கான முறைமை-செயல்பாட்டு அணுகுமுறை. குரோமோசோம்கள். சைட்டோபிளாசம். குருட்டுத்தன்மை. காது நீளம். ஒரு நபரின் வகைப்பாடு. ஒரு பாலூட்டியின் எலும்புக்கூடு. மனித பரிணாமத்தின் பாதைகள். மைடோசிஸ். மேலோட்டமான வளாகம். பிரச்சனைக்குரிய கேள்வி. நியூக்ளியோலஸ். அணு உறை.
"உயிரியலில் கணினி" - மாணவர்களின் கூட்டு செயல்பாடு. ஆஞ்சியோஸ்பெர்ம் குடும்பங்கள். ஊடாடும் பயிற்சி. கற்றல் மாதிரிகள். மதிப்பீட்டு முறையின் எடுத்துக்காட்டு. அறிவுறுத்தல் அட்டை கேள்விகள். அறிவுறுத்தல் அட்டையின் எடுத்துக்காட்டு. ஆராய்ச்சியாளர்கள். நுண்குழுக்கள். ஊடாடும் கற்றல் தொழில்நுட்பங்கள். கொணர்வி. ஊடாடும் கற்றல் தொழில்நுட்பங்கள். உயிரியல் பாடங்களில் ஊடாடும் அணுகுமுறைகள். வேலையின் குழு வடிவம். "ஆராய்ச்சியாளர்களின்" குழுக்களுக்கான பணிகள்.
