§ 2. Eukaryoottisolun pääkomponentit
Eukaryoottisolut (kuvat 8 ja 9) ovat paljon monimutkaisempia kuin prokaryoottisolut. Ne ovat kooltaan (muutamista mikrometreistä useisiin senttimetreihin), muodoltaan ja rakenteeltaan hyvin erilaisia (kuva 10).
Riisi. 8. Eukaryoottisolun rakenne. Yleinen kaava
Riisi. 9. Solurakenne elektronimikroskopian mukaan
Riisi. 10. Erilaiset eukaryoottisolut: 1 – epiteelisolut; 2 – veri (e – erytriitti, / – leukiitti); 3 – rusto; 4 – luut; 5 – sileät lihakset; 6 – sidekudos; 7 – hermosolut; 8 – poikkijuovainen lihaskuitu
Peruskomponenttien yleinen organisaatio ja läsnäolo ovat kuitenkin samat kaikissa eukaryoottisoluissa (kuvio 11).
Riisi. 11. Eukaryoottisolu (kaavio)
Plasmalemma (ulompi solukalvo). Plasmakalvon perusta, kuten muutkin solujen kalvot (esimerkiksi mitokondriot, plastidit jne.), on lipidien kerros, jossa on kaksi riviä molekyylejä (kuva 12). Koska lipidimolekyylit ovat polaarisia (yksi napa on hydrofiilinen eli vesi vetää puoleensa ja toinen on hydrofobinen eli veden hylkivä), ne on järjestetty tiettyyn järjestykseen. Yhden kerroksen molekyylien hydrofiiliset päät suuntautuvat kohti vesipitoista ympäristöä - solun sytoplasmaan ja toinen kerros - ulospäin solusta - kohti solujenvälistä ainetta (monisoluisissa organismeissa) tai vesiympäristöä (yksisoluisissa organismeissa) ).
Riisi. 12. Solukalvon rakenne nestemosaiikkimallin mukaan. Proteiinit ja glykoproteiinit upotetaan kaksinkertaiseen lipidimolekyylikerrokseen siten, että niiden hydrofiiliset päät (ympyrät) osoittavat ulospäin ja niiden hydrofobiset päät (aaltoviivat) syvälle kalvoon.
Proteiinimolekyylit ovat mosaiikkimaisesti upotettuina bimolekulaariseen lipidien kerrokseen. Eläinsolun ulkopuolella polysakkaridimolekyylit ovat kiinnittyneet plasmalemman lipideihin ja proteiinimolekyyleihin muodostaen glykolipidejä ja glykoproteiineja.
Tämä aggregaatti muodostaa kerroksen glykokaliksi. Liittyy häneen reseptorin toiminta plasmakalvot (katso alla); se voi myös kerätä erilaisia solun käyttämiä aineita. Lisäksi glykokaliksi parantaa plasmalemman mekaanista stabiilisuutta.
Kasvien ja sienten soluissa on myös soluseinä, jolla on tukeva ja suojaava rooli. Kasveissa se koostuu selluloosasta ja sienissä kitiinistä.
Ulompi solukalvo suorittaa useita toimintoja, mukaan lukien:
♦ mekaaninen(tuki, muodonrakennus);
♦ este-kuljetus(selektiivinen läpäisevyys eri aineille: tarvittavien aineiden pääsy soluun ja tarpeettomien ja haitallisten poistaminen);
♦ reseptori(solun välittömässä läheisyydessä olevien erilaisten kemikaalien määrittäminen; hormonien muodossa olevien signaalien havaitseminen; immuunijärjestelmän solujen "vieraan" proteiinin tunnistaminen jne.).
Aineiden vaihto solun ja ympäristön välillä tapahtuu eri tavoin - passiivisesti ja aktiivisesti.
Veden ja erilaisten ionien molekyylit pääsevät passiivisesti (diffuusiosta, osmoosista) ilman, että solu kuluttaa energiaa erityisten huokosten kautta - nämä ovat passiivinen kuljetus. Makromolekyylejä, kuten proteiineja, polysakkarideja, jopa kokonaisia soluja, toimitetaan fagosytoosi Ja pinosytoosi energiankulutuksella - aktiivinen kuljetus.
Fagosytoosin seurauksena kokonaiset solut tai suuret hiukkaset nielaisevat (mieti esimerkiksi ameetojen ravintoa tai bakteerien suojaavien verisolujen fagosytoosia). Pinosytoosin aikana nestemäisen aineen pienet hiukkaset tai pisarat imeytyvät. Molemmille prosesseille on yhteistä, että imeytyviä aineita ympäröi tunkeutuva ulkokalvo muodostaen tyhjiön, joka siirtyy sitten syvälle solun sytoplasmaan.
Eksosytoosi on fagosytoosiin ja pinosytoosiin nähden vastakkainen prosessi (joka on myös aktiivinen kuljetus) (kuva 13). Sen avulla voidaan poistaa sulamattomia ruokajäämiä alkueläimiin tai erityssoluun muodostuneita biologisesti aktiivisia aineita.
Sytoplasma. Sytoplasma on solun sisältö, jota rajoittaa plasmalemma, lukuun ottamatta ydintä. Se sisältää jauhettu aine (hyaloplasma), organellit Ja sulkeumia.
Hyaloplasma- viskoosi neste, joka voi olla jommankumman tilassa Zola(neste), tai geeli(hyytelömäinen).
Tarvittaessa sytoplasma pystyy siirtymään palautuvasti tilasta toiseen. Esimerkiksi ameboidiliikkeen aikana (muista eläintieteen kurssin kohta "Alkueläimet") pseudopodin muodostumisen aikana tapahtuu nopeita sytoplasman siirtymiä geelistä sooliin ja päinvastoin. Tämä johtuu siitä, että sytoplasmassa on suuri määrä filamenttiproteiinimolekyylejä aktina. Kun ne liittyvät toisiinsa muodostaen kolmiulotteisen verkoston, sytoplasma on geelitilassa, ja kun verkko hajoaa, se on soolitilassa.
Hyaloplasma sisältää erilaisia aineita - entsyymejä, proteiineja, hiilihydraatteja, rasvoja ja muita, orgaanisia ja mineraaleja. Täällä tapahtuu erilaisia kemiallisia prosesseja - aineiden hajoamista, niiden synteesiä ja modifikaatioita (muutoksia).
Organoidit. Nämä ovat solun pysyviä komponentteja, joilla on tietty rakenne ja toiminnot ja jotka sijaitsevat sen sytoplasmassa. Seuraavassa puhumme siitä yleiskäyttöiset organoidit, joka on luontainen kaikkien eukaryoottien kaikentyyppisille soluille. Ne liittyvät viimeksi mainitun elintärkeiden toimintojen varmistamiseen. Erikoiskäyttöiset organoidit löytyy vain tietyn (erittäin erikoistuneen) tyypin soluista - esimerkiksi lihassolujen myofibrilleistä.
Yleiskäyttöisillä organelleilla on sama rakenne riippumatta siitä, mihin soluihin ja organismeihin ne kuuluvat. Mutta niiden joukossa on ryhmiä, joissa on kalvo (endoplasminen verkkokalvo, Golgi-laitteisto, mitokondriot, plastidit, lysosomit, vakuolit), sekä ei-kalvo ( ribosomit, solukeskus) rakenne.
Endoplasminen verkkokalvo (ER). EPS koostuu kalvoista ja on monimutkainen haarautunut putkien ja säiliöiden järjestelmä, joka tunkeutuu solun koko sytoplasmaan (kuva 14). EPS:ää on kahta tyyppiä - karkea Ja sileä. Ribosomit ovat kiinnittyneet karkeisiin kalvoihin (sytoplasmapuolelta), kun taas sileillä kalvoilla niitä ei ole.
Riisi. 14. Endoplasminen verkkokalvo
Endoplasmisella retikulumilla on useita tärkeitä toimintoja eukaryoottisolussa:
♦ rajaamalla(solun sisäisen tilavuuden jakaminen erilaisiin reaktiotiloihin);
♦ osallistuminen orgaanisten aineiden synteesiin(ribosomit sijaitsevat karkean EPS:n kalvoilla, ja sileillä kalvoilla on entsyymikomplekseja, jotka varmistavat lipidien, hiilihydraattien jne. synteesin);
♦ osallistuminen Golgi-laitteen elementtien, lysosomien, muodostumiseen;
♦ aineiden kuljetus.
Golgin laite. Golgi-laite (AG) on järjestelmä tankit(litteät vakuolit) ja kuplia(vesikkelit), jotka sijaitsevat solun ytimen välittömässä läheisyydessä, jotka muodostuvat EPS:n vuoksi sen pienten fragmenttien erottumisen seurauksena (kuvio 15). Kun nämä fragmentit sulautuvat yhteen, syntyy uusia Golgi-laitteen vesisäiliöitä, kun taas EPS:stä kulkeutuu erilaisia aineita, jotka osallistuvat monimutkaisten orgaanisten yhdisteiden (proteiinit + hiilihydraatit, proteiinit + lipidit jne.) kokoonpanoon, jotka poistetaan AG:n apua solun ulkopuolella. Nämä biologisesti aktiiviset aineet joko poistetaan solusta (käyttäen eritysvakuoleja eksosytoosin avulla) tai ovat osa AG:n muodostamia lysosomeja (katso alla).
Riisi. 15. Golgi-laite:
Golgi-laite suorittaa seuraavat toiminnot:
♦ synteesi solun tuottamat biologisesti aktiiviset aineet;
♦ erilaisten aineiden eritys (poisto solusta).(hormonit, entsyymit, aineet, joista soluseinä rakennetaan jne.);
♦ osallistuminen lysosomien muodostukseen.
Mitokondriot. Kaikilla eukaryoottisoluilla on mitokondrioita (kuvio 16). Ne näyttävät joko pyöristetyiltä rungoilta tai tangoilta, harvemmin - kierteiltä. Niiden koot vaihtelevat 1-7 mikronia. Mitokondrioiden määrä solussa vaihtelee useista sadaista kymmeniin tuhansiin (suurissa alkueläimissä).
Riisi. 16. Mitokondriot. Yllä - mitokondriot (a) virtsakanavissa, näkyvät valomikroskoopilla. Alla on kolmiulotteinen malli mitokondrioiden järjestäytymisestä: 1 – cristae; 2 – ulkokalvo; 3 – sisäkalvo; 4 – matriisi
Mitokondrio muodostuu kahdesta kalvosta - ulkoinen Ja sisäinen, joiden välissä sijaitsee kalvojen välinen tila. Sisäkalvo muodostaa monia invaginaatioita - cristae, jotka ovat joko levyjä tai putkia. Tämä organisaatio tarjoaa valtavan alueen sisäisestä kalvosta. Se sisältää entsyymejä, jotka varmistavat orgaanisten aineiden (hiilihydraatit, lipidit) sisältämän energian muuntamisen solun elämälle välttämättömäksi ATP-energiaksi. Siksi mitokondrioiden tehtävänä on osallistua energiaa soluprosessit. Siksi suuri määrä mitokondrioita on luonnostaan esimerkiksi lihassoluissa, jotka tekevät paljon työtä.
Plastidit. Kasvisoluissa löytyy erityisiä organelleja - plastideja, joilla on usein karan muotoinen tai pyöristetty muoto, joskus monimutkaisempi. Plastideja on kolmea tyyppiä - kloroplastit (kuva 17), kromoplastit ja leukoplastit.
Kloroplastit eroavat vihreästä väristä, mikä johtuu pigmentistä - klorofylli, prosessin varmistaminen fotosynteesi, eli orgaanisten aineiden synteesi vedestä (H 2 O) ja hiilidioksidista (CO 2) auringonvalon energiaa käyttämällä. Kloroplasteja löytyy pääasiassa lehtisoluista (korkeammissa kasveissa). Ne muodostuvat kahdesta yhdensuuntaisesta kalvosta, jotka ympäröivät kloroplastien sisältöä - stroma. Sisäkalvo muodostaa useita litistettyjä pusseja - tylakoidit, jotka on pinottu (kuten kolikoiden pino) – jyviä - ja makaa stromassa. Tylakoidit sisältävät klorofylliä.
Kromoplastit määrittää monien kukkien ja hedelmien keltaisen, oranssin ja punaisen värin, joiden soluissa niitä on suuria määriä. Niiden koostumuksen pääpigmentit ovat karoteenit. Kromoplastien toiminnallinen tarkoitus on houkutella eläimiä väreihin, mikä varmistaa kukkien pölytyksen ja siementen leviämisen.
Riisi. 17. Plastidit: a – kloroplastit Elodea-lehden soluissa, näkyvät valomikroskoopilla; b – kaavio kloroplastin sisäisestä rakenteesta granoilla, jotka ovat pinoja litteitä pusseja, jotka sijaitsevat kohtisuorassa kloroplastin pintaan nähden; c – yksityiskohtaisempi kaavio, joka esittää yksittäisiä tuuletinkammioita yhdistäviä anastomoosiputkia
Leukoplastit ovat värittömiä plastideja, jotka sisältyvät kasvien maanalaisten osien soluihin (esimerkiksi perunan mukuloihin), siemeniin ja varren ytimeen. Leukoplasteissa tärkkelys muodostuu pääasiassa glukoosista ja kertyy kasvien varastoelimiin.
Yhden tyyppiset plastidit voivat muuttua toiseksi. Esimerkiksi kun lehdet vaihtavat väriä syksyllä, kloroplastit muuttuvat kromoplasteiksi.
Lysosomit. Nämä organellit näyttävät rakkuloilta, joita ympäröi kalvo, jonka halkaisija on enintään 2 mikronia. Ne sisältävät useita kymmeniä entsyymejä, jotka hajottavat proteiineja, nukleiinihappoja, polysakkarideja ja lipidejä. Lysosomien tehtävänä on osallistua monimutkaisten orgaanisten yhdisteiden (esimerkiksi ravinteiden tai "käytettyjen" solukomponenttien aineiden) solunsisäisiin hajoamisprosesseihin. Lysosomit fuusioituvat fagosyyttisten (tai pinosyyttisten) vakuolien kanssa muodostaen ruoansulatusvakuolin.
Lysosomien muodostuminen johtuu Golgi-laitteen säiliöstä silmustumisesta.
Ribosomit. Ribosomeja (kuva 18) on sekä eukaryoottien että prokaryoottien soluissa, koska niillä on tärkeä tehtävä proteiinien biosynteesi(katso luku 5). Jokainen solu sisältää kymmeniä, satoja tuhansia (jopa useita miljoonia) näitä pieniä pyöreitä organelleja.
Riisi. 18. Kaavio endoplasmisen retikulumin kalvolla istuvan ribosomin rakenteesta: 1 – pieni alayksikkö; 2 – tRNA; 3 – aminoasyyli-tRNA; 4 – aminohappo; 5 – suuri alayksikkö; 6 – endoplasminen verkkokalvo; 7 – syntetisoitu polypeptidiketju
Ribosomi koostuu kahdesta epätasaisesta alayksiköstä (osasta). Ne muodostuvat erikseen ja yhdistyvät "sisältäen" lähetti-RNA:n proteiinimolekyylin synteesin aikana. Ribosomit sisältävät erilaisia proteiineja ja ribosomaalisia RNA:ita.
Solujen sulkeumat. Tämä on nimi, joka annetaan solussa oleville ei-pysyville komponenteille, joita esiintyy sytoplasman pääaineessa jyvien, rakeiden tai pisaroiden muodossa. Inkluusio voi olla kalvon ympäröimä tai ei.
Toiminnallisesti sisällytyksiä on kolmen tyyppisiä: vararavinteita(tärkkelys, glykogeeni, rasvat, proteiinit), erittäviä sulkeumia(rauhassoluille tyypilliset, niiden tuottamat aineet - umpierityshormonit jne.) ja erityistarkoituksen sisällyttäminen(erittäin erikoistuneissa soluissa, esimerkiksi hemoglobiini punasoluissa).
§ 3. Soluytimen organisointi. Kromosomit
Solun ydin (katso kuviot 8 ja 9) on äärimmäisen tärkeä solun elämässä, koska se toimii kromosomien sisältämän perinnöllisen informaation varastona (katso alla).
Ydintä rajoittaa ydinvaippa, joka erottaa sen sisällön (karyoplasma) sytoplasmasta. Kuori koostuu kahdesta kalvosta, joita erottaa rako. Molemmissa on lukuisia huokosia, minkä ansiosta aineiden vaihto ytimen ja sytoplasman välillä on mahdollista. Useimpien eukaryoottien soluytimessä on 1-7 nukleolit. Niihin liittyvät RNA- ja tRNA-synteesiprosessit.
Ytimen tärkeimmät komponentit - kromosomit, muodostuu DNA-molekyylistä ja erilaisista proteiineista. Valomikroskoopissa ne näkyvät selvästi vain solunjakautumisen aikana (mitoosi, meioosi). Jakautumattomassa solussa kromosomit näyttävät pitkiltä ohuilta langoilta, jotka ovat jakautuneet koko ytimen tilavuuteen.
Solunjakautumisen aikana kromosomijuosteet muodostavat tiheitä spiraaleja, minkä seurauksena ne tulevat näkyviin (tavanomaisella mikroskoopilla) sauvojen, "hiusneulien" muodossa. Koko geneettisen tiedon määrä on jakautunut ytimen kromosomien kesken. Niitä tutkittaessa tunnistettiin seuraavat mallit:
♦ somaattisten solujen ytimissä (eli kehon solut, ei-seksuaaliset) kaikki saman lajin yksilöt sisältävät saman määrän kromosomeja, jotka muodostavat joukko kromosomeja(kuvio 19);
Riisi. 19. Eri kasvi- ja eläinlajien kromosomit, kuvattuna samassa mittakaavassa: 1,2 – ameba; 3,4 – piilevät; 5–8, 18,19 – viherlevät; 9 - kärpäshelta; 10 - lehmus; 11–12 – Drosophila; 13 - lohi; 14 – skerda (Asteraceae-suku); 15 – aroid-perheen kasvi; 16 - corydalis perhonen; 17 – heinäsirkkasuvun hyönteis; 20 – vesijuoksuvika; 21 – kukkahävikki; 22 – sammakkoeläin Abystoma; 23 - aloe (liljaperhe)
♦ kullekin lajille on ominaista oma kromosomiryhmänsä lukumäärän mukaan (esimerkiksi ihmisellä on 46 kromosomia, hedelmäkärpäsellä 8, sukkulamadolla 4, rapulla 196, hevosella 66 ja maissilla 104);
♦ somaattisten solujen ytimissä olevat kromosomit voidaan ryhmitellä pareiksi nimeltä homologiset kromosomit niiden samankaltaisuuden perusteella (rakenteeltaan ja toiminnaltaan);
♦ sukusolujen (sukusolujen) ytimissä jokainen homologinen kromosomipari sisältää vain yhden, eli kromosomien kokonaismäärä on puolet somaattisten solujen määrästä;
♦ Yksittäistä kromosomisarjaa sukusoluissa kutsutaan haploidi ja on merkitty kirjaimella n, ja somaattisella - diploidi(2n).
Edellä olevasta on selvää, että jokainen homologinen kromosomipari muodostuu isän ja äidin kromosomien yhdistymisestä hedelmöityksen aikana, eli sukupuolisolujen (sukusolujen) fuusiossa. Sitä vastoin sukusolujen muodostumisen aikana jokaisesta homologisesta kromosomiparista vain yksi pääsee sukusoluun.
Kromosomit eri homologiset parit eroavat kooltaan ja muodoltaan (kuvat 20 ja 21).
Riisi. 20. Kromosomien rakenne ja tyypit: a – ulkonäkö 1 – sentromeeri; 2 – lyhyt olkapää; 3 – pitkä olkapää); saman kromosomin sisäinen rakenne (1 – sentromeeri; – DNA-molekyylit); c – kromosomityypit (1 – yksikätinen; erikätinen; 3 – tasakätinen: X – käsivarsi, V – sentromeeri)
Riisi. 21. Kromosomi koostuu DNA:sta ja proteiineista. DNA-molekyyli replikoituu. Kaksi identtistä DNA:n kaksoiskierrettä pysyy kytkettynä sentromeerialueella. Näistä kopioista tulee erillisiä kromosomeja myöhemmin solun jakautumisen aikana
Kehossa kromosomeja on ensisijainen supistuma (kutsutaan sentromeeriksi), johon langat on kiinnitetty fissiokarat. Se jakaa kromosomin kahteen osaan olkapää Kromosomit voivat olla yhtäkätisiä, epätasa-kätisiä tai yksikätisiä.
Luku 5. Aineenvaihdunta
§ 1. Aineenvaihdunta sulautumisen ja hajoamisen yhtenäisyytenä
Kaikki solut ja elävät organismit ovat avoimia järjestelmiä, toisin sanoen ne ovat jatkuvassa energioiden ja aineiden vaihdossa ympäristön kanssa. Elottomassa luonnossa on avoimia järjestelmiä, mutta niiden olemassaolo eroaa laadullisesti elävistä organismeista. Ajatellaanpa tätä esimerkkiä: palava natiivi rikki on vaihtotilassa ympäristön kanssa. Kun se palaa, O 2 imeytyy ja SO 2:ta ja energiaa (lämmön muodossa) vapautuu. Kuitenkin tässä tapauksessa pala rikkiä fyysisenä kappaleena tuhoutuu ja menettää primäärirakenteensa.
Eläville organismeille vaihto ympäristön kanssa on edellytys niiden rakenteellisen organisaation säilyttämiselle ja ylläpitämiselle kaikkien niiden aineiden ja komponenttien itsensä uudistumisen kautta.
Aineenvaihdunta (aineenvaihdunta) on joukko elävissä organismeissa tapahtuvia prosesseja (aineiden ja energian kulutus, muuntaminen, kertyminen ja vapautuminen), jotka varmistavat niiden elintärkeän toiminnan, kehityksen, kasvun ja lisääntymisen. Aineenvaihduntaprosessin aikana solut muodostavat molekyylit hajoavat ja syntetisoidaan; solurakenteiden ja solujen välisen aineen uusiutuminen.
Aineenvaihdunta perustuu toisiinsa yhteydessä oleviin prosesseihin assimilaatiota(anabolia) ja dissimilaatio(katabolismi). Assimilaation (plastisen vaihdon) aikana monimutkaiset aineet syntetisoidaan yksinkertaisista. Tämän ansiosta solussa syntyvät kaikki orgaaniset aineet, jotka ovat välttämättömiä sen rakennekomponenttien, entsyymijärjestelmien jne. rakentamiseen. Assimilaatio tapahtuu aina energiankulutuksella.
Dissimilaation (energia-aineenvaihdunnan) aikana monimutkaiset orgaaniset aineet hajoavat yksinkertaisemmiksi tai epäorgaanisiksi. Tällöin vapautuu energiaa, jonka solu käyttää erilaisten prosessien suorittamiseen, jotka varmistavat sen elintärkeän toiminnan (aineiden synteesi ja kuljetus, mekaaninen työ jne.).
Kaikki elävät organismit voidaan jakaa kahteen ryhmään: autotrofit Ja heterotrofit, jotka eroavat energialähteiden ja elintoimintojensa turvaamiseksi tarvittavien aineiden osalta.
Autotrofit– organismit, jotka syntetisoivat orgaanisia yhdisteitä epäorgaanisista aineista auringonvalon energialla (esim fototrofit– kasvit, syanobakteerit) tai mineraali- (epäorgaanisten) aineiden (esim. kemotrofit– rikkibakteerit, rautabakteerit jne.). Näin ollen he pystyvät itsenäisesti luomaan elämänsä edellyttämiä aineita.
§ 2. Dissimilaatio anaerobisissa ja aerobisissa organismeissa
Organismit voidaan jakaa kahteen ryhmään ja dissimilaation luonteen mukaan - aerobit Ja anaerobit. Aerobes (kreikasta. demoni– ilma) tarvitsevat vapaata happea elämää varten. Anaerobeissa (Kreikka. ac– negatiivinen hiukkanen) se ei ole välttämätöntä. Niissä dissimilaatio suoritetaan fermentoinnilla - orgaanisen aineen hapettomalla, entsymaattisella hajoamisella muodostamalla yksinkertaisempia orgaanisia aineita ja vapauttamalla energiaa. Esimerkiksi:
♦ maitohappokäyminen:
C6H12O6 + 2H3PO4 + 2ADP -> 2PH + 2ATP + 2H20;
♦ alkoholikäyminen:
C6H12O6 + 2PH + 2ADP → 2C2H5OH + 2CO2 + 2ATP + 2H2O.
Käymisen aikana muodostuvat aineet ovat orgaanisia ja sisältävät siksi edelleen paljon energiaa.
Riisi. 22. Assimilaation ja dissimilaation väliset suhteet autotrofisissa ja heterotrofisissa organismeissa
Aerobisissa organismeissa mitokondrioiden hengitysprosessin aikana tapahtuu orgaanisten aineiden täydellinen hajoaminen (O 2:n avulla) energiahuonoiksi lopputuotteiksi CO 2 ja H 2 O ja vapautuu huomattavasti suurempi määrä energiaa:
C 6 H 12 0 6 (glukoosi) + 0 2 > 6C0 2 + 6H 2 0 + energia (jonka ansiosta syntetisoituu 38 ATP-molekyyliä).
Tarkastellaan yleistettyjen kaavioiden muodossa autotrofisten ja heterotrofisten aerobisten organismien aineenvaihduntaa (kuva 22).
Assimilaatio. Sen tärkeimmät prosessit ovat fotosynteesi Ja proteiinien biosynteesi.
Organismin yhteys ympäristöön on fysikaalis-kemiallisesta näkökulmasta avoin järjestelmä, eli järjestelmä, jossa biokemialliset prosessit ovat käynnissä. Lähtöaineet tulevat ympäristöstä, ja myös jatkuvasti muodostuvat aineet kuljetetaan ulos. Tasapaino monisuuntaisten reaktioiden tuotteiden nopeuden ja pitoisuuden välillä kehossa on ehdollinen, kuvitteellinen, koska aineiden saanti ja poistaminen ei pysähdy. Jatkuva yhteys ympäristöön antaa meille mahdollisuuden pitää elävää organismia avoimena järjestelmänä.
Kaikkien elävien solujen energianlähde on aurinko. Kasvisolut vangitsevat energiaa auringonvalosta klorofyllin avulla ja käyttävät sitä assimilaatioreaktioihin fotosynteesin aikana. Eläinten, sienten ja bakteerien solut käyttävät aurinkoenergiaa epäsuorasti maan kasvien syntetisoimien orgaanisten aineiden hajoamisen aikana.
Osa solun ravintoaineista hajoaa soluhengityksen aikana, jolloin saadaan tarvittavaa energiaa erilaisiin solutoimintoihin. Tämä prosessi tapahtuu organelleissa, joita kutsutaan mitokondrioiksi. Mitokondriot koostuvat kahdesta kalvosta: ulompi, joka erottaa organellin sytoplasmasta, ja sisempi, joka muodostaa useita laskoksia. Hengityksen päätuote on ATP. Se poistuu mitokondrioista ja sitä käytetään energialähteenä moniin kemiallisiin reaktioihin sytoplasmassa ja solukalvossa. Jos soluhengitykseen tarvitaan happea, niin hengitystä kutsutaan aerobiseksi, mutta jos reaktioita tapahtuu ilman happea, puhutaan anaerobisesta hengityksestä.
Kaikentyyppiseen solussa suoritettavaan työhön energiaa käytetään yhdessä ja ainoassa muodossa - ATP:n fosfaattisidosten energian muodossa. ATP on helposti liikkuva yhdiste. ATP:n muodostuminen tapahtuu mitokondrioiden sisäkalvolla. ATP syntetisoituu kaikissa soluissa hengityksen aikana hiilihydraattien, rasvojen ja muiden orgaanisten aineiden hapetusenergian vuoksi. Vihreissä kasvisoluissa suurin osa ATP:stä syntetisoituu kloroplasteissa aurinkoenergian vaikutuksesta. Fotosynteesin aikana ne tuottavat monta kertaa enemmän ATP:tä kuin mitokondriot. ATP hajoaa fosfori-happisidosten katkeamisen ja energian vapautumisen myötä. Tämä tapahtuu ATPaasi-entsyymin vaikutuksesta ATP:n hydrolyysin aikana - lisäämällä vettä fosforihappomolekyylin eliminoimalla. Tämän seurauksena ATP muuttuu ADP:ksi, ja jos kaksi fosforihappomolekyyliä hajoaa, sitten AMP:ksi. Kunkin gramman happomolekyylin eliminaatioreaktioon liittyy 40 kJ:n vapautuminen. Tämä on erittäin suuri energiantuotto, minkä vuoksi ATP:n fosfori-happisidoksia kutsutaan yleensä makroergistisiksi (korkeaenergiaisiksi).
ATP:n käyttö plastisissa vaihtoreaktioissa tapahtuu kytkemällä ne ATP-hydrolyysillä. Erilaisten aineiden molekyylit varataan energialla kiinnittämällä ATP-molekyylistä hydrolyysin aikana vapautuva fosforiryhmä eli fosforylaatiolla.
Fosfaattijohdannaisten erityispiirre on, että ne eivät pääse poistumaan solusta, vaikka niiden "purkautuneet" muodot kulkevat vapaasti kalvon läpi. Tämän ansiosta fosforyloidut molekyylit pysyvät solussa, kunnes niitä käytetään sopivissa reaktioissa.
Käänteinen prosessi ADP:n muuntamiseksi ATP:ksi tapahtuu lisäämällä ADP:hen fosforihappomolekyyli, joka vapauttaa vettä ja absorboi suuren määrän energiaa.
Siten ATP on universaali ja suora solutoiminnan energialähde. Tämä luo yhden solun energiaaltaan ja mahdollistaa sen uudelleenjakamisen ja kuljettamisen solun alueelta toiselle.
Fosfaattiryhmän siirrolla on tärkeä rooli kemiallisissa reaktioissa, kuten makromolekyylien kokoamisessa monomeereistä. Esimerkiksi aminohapot voidaan yhdistää peptideiksi vasta, kun ne on aiemmin fosforyloitu. Mekaaniset supistumis- tai liikeprosessit, liuenneen aineen kuljettaminen pitoisuusgradienttia vasten ja muut prosessit sisältävät ATP:hen varastoidun energian kulutuksen.
Energian aineenvaihdunnan prosessi voidaan esittää seuraavasti. Sytoplasman korkeamolekyyliset orgaaniset aineet muunnetaan entsymaattisesti hydrolyysin avulla yksinkertaisemmiksi, joista ne koostuvat: proteiinit - aminohapoiksi, poly- ja disakkaridit - monosakkarideiksi (+ glukoosi), rasvat glyseroliksi ja rasvahapoiksi. Ei ole oksidatiivisia prosesseja, vapautuu vähän energiaa, jota ei käytetä ja menee lämpömuotoon. Useimmat solut käyttävät ensin hiilihydraatteja. Polysakkaridit (tärkkelys kasveissa ja glykogeeni eläimissä) hydrolysoituvat glukoosiksi. Glukoosin hapettuminen tapahtuu kolmessa vaiheessa: glykolyysi, oksidatiivinen dekarboksylaatio (Krebsin sykli - sitruunahapposykli) ja oksidatiivinen fosforylaatio (hengitysketju). Glykolyysi, jonka seurauksena yksi glukoosimolekyyli hajoaa kahdeksi palorypälehappomolekyyliksi vapauttamalla kaksi ATP-molekyyliä, tapahtuu sytoplasmassa. Hapen puuttuessa palorypälehappo muuttuu joko etanoliksi (käyminen) tai maitohapoksi (anaerobinen hengitys).
Kun glykolyysi tapahtuu eläinsoluissa, kuuden hiilen glukoosimolekyyli hajoaa kahdeksi maitohappomolekyyliksi. Tämä prosessi on monivaiheinen. Sen suorittaa peräkkäin 13 entsyymiä. Alkoholikäymisen aikana glukoosimolekyylistä muodostuu kaksi etanolimolekyyliä ja kaksi CO2-molekyyliä.
Glykolyysi on yhteinen vaihe anaerobiselle ja aerobiselle hengitykselle, kaksi muuta tapahtuu vain aerobisissa olosuhteissa. Happivapaa hapetusprosessi, jossa vapautuu ja käytetään vain osa aineenvaihduntatuotteiden energiasta, on lopullinen anaerobisille organismeille. Hapen läsnä ollessa, palorypälehappo siirtyy mitokondrioihin, joissa se hapettuu useiden peräkkäisten reaktioiden seurauksena täysin aerobisesti H2O:ksi ja CO2:ksi samalla, kun ADP fosforyloituu ATP:ksi. Tässä tapauksessa kaksi ATP-molekyyliä tuotetaan glykolyysillä, kaksi Krebsin syklillä ja 34 hengitysketjulla. Nettosaanto yhden glukoosimolekyylin täydelliselle hapetukselle H2O:ksi ja CO2:ksi on 38 molekyyliä.
Siten aerobisissa organismeissa orgaanisten aineiden lopullinen hajoaminen suoritetaan hapettamalla ne ilmakehän hapella yksinkertaisiksi epäorgaanisiksi aineiksi: CO2 ja H2O. Tämä prosessi tapahtuu mitokondrioiden risteyksissä. Tällöin vapautuu maksimimäärä vapaata energiaa, josta merkittävä osa on varattu ATP-molekyyleihin. On helppo nähdä, että aerobinen hapetus antaa solulle suurimmassa määrin vapaata energiaa.
Katabolian seurauksena soluun kerääntyy runsaasti energiaa sisältäviä ATP-molekyylejä ja CO2 ja ylimääräinen vesi vapautuvat ulkoiseen ympäristöön.
Sokerimolekyylejä, joita ei tarvita hengitykseen, voidaan varastoida soluun. Ylimääräiset lipidit joko hajoavat, minkä jälkeen niiden hajoamistuotteet pääsevät mitokondrioihin hengityksen substraattina tai kerääntyvät varastoiksi sytoplasmaan rasvapisaroiden muodossa. Proteiinit rakentuvat soluun tulevista aminohapoista. Proteiinisynteesi tapahtuu organelleissa, joita kutsutaan ribosomeiksi. Jokainen ribosomi koostuu kahdesta osahiukkasesta - suuresta ja pienestä: molemmat alahiukkaset sisältävät proteiinimolekyylejä ja RNA-molekyylejä.
Ribosomit on usein kiinnitetty erityiseen kalvojärjestelmään, joka koostuu säiliöistä ja vesikkeleistä - niin kutsuttuun endoplasmiseen retikulumiin (ER); soluissa, jotka tuottavat paljon proteiinia, endoplasminen retikulumi on usein hyvin kehittynyt ja peitetty ribosomeilla. Jotkut entsyymit ovat tehokkaita vain, jos ne on kiinnitetty kalvoon. Suurin osa lipidisynteesiin osallistuvista entsyymeistä sijaitsee täällä. Siten endoplasminen retikulumi on kuin eräänlainen solutyöpöytä.
Lisäksi ER jakaa sytoplasman erillisiin osastoihin, eli se erottaa erilaisia sytoplasmassa samanaikaisesti tapahtuvia kemiallisia prosesseja ja vähentää siten todennäköisyyttä, että nämä prosessit häiritsevät toisiaan.
Usein tietyn solun tuottamia tuotteita käytetään solun ulkopuolella. Tällaisissa tapauksissa ribosomeihin syntetisoidut proteiinit kulkevat endoplasmisen retikulumin kalvojen läpi ja pakautuvat niiden ympärille muodostuviin kalvorakkuloihin, jotka sitten irrotetaan ER:stä. Nämä litistetyt ja päällekkäin pinottu rakkulat, kuten pinottu pannukakut, muodostavat tyypillisen rakenteen, jota kutsutaan Golgi-kompleksiksi tai Golgi-laitteistoksi. Golgi-laitteessa oleskelunsa aikana proteiinit käyvät läpi tiettyjä muutoksia. Kun niiden aika on poistua solusta, kalvorakkulat sulautuvat solukalvoon ja tyhjennetään kaataen sisältönsä ulos, eli erittyminen tapahtuu eksosytoosin kautta.
Golgi-laitteisto tuottaa myös lysosomeja - kalvopusseja, jotka sisältävät ruoansulatusentsyymejä. Sen selvittäminen, kuinka solu valmistaa, pakkaa ja vie tiettyjä proteiineja ja kuinka se "tietää", mitkä proteiinit sen tulisi säilyttää itselleen, on yksi modernin sytologian kiehtovimmista haaroista.
Minkä tahansa solun kalvot liikkuvat ja muuttuvat jatkuvasti. ER-kalvot liikkuvat hitaasti läpi solun. Näiden kalvojen yksittäiset osat erottuvat ja muodostavat rakkuloita, joista tulee väliaikaisesti osa Golgin laitetta, ja sitten eksosytoosiprosessin kautta sulautuvat solukalvon kanssa.
Myöhemmin kalvomateriaali palautetaan sytoplasmaan, jossa sitä käytetään uudelleen.
Soluun tulevien tai sen ulkopuolelle vapautuvien aineiden vaihto sekä erilaisten signaalien vaihto mikro- ja makroympäristön kanssa tapahtuu solun ulkokalvon kautta. Kuten tiedetään, solukalvo on lipidikaksoiskerros, johon on upotettu erilaisia proteiinimolekyylejä, jotka toimivat erikoistuneina reseptoreina, ionikanavina, erilaisia kemikaaleja aktiivisesti kuljettavina tai poistavina laitteina, solujen väliset kontaktit jne. Terveissä eukaryoottisoluissa fosfolipidit jakautuvat kalvo epäsymmetrisesti: ulkopinta koostuu sfingomyeliinistä ja fosfatidyylikoliinista, sisäpinta fosfatidyyliseriinistä ja fosfatidyylietanoliamiinista. Tällaisen epäsymmetrian ylläpitäminen vaatii energiankulutusta. Siksi soluvaurion, infektion tai energiannälkään sattuessa kalvon ulkopinta rikastuu sille epätavallisilla fosfolipideillä, joista tulee muille soluille ja entsyymeille signaali soluvauriosta vastaavalla reaktiolla. Tärkein rooli on fosfolipaasi A2:n liukoisella muodolla, joka hajottaa arakidonihappoa ja muodostaa lysoformeja edellä mainituista fosfolipideistä. Arakidonihappo on rajoittava linkki tulehdusvälittäjien, kuten eikosanoidien, muodostumiselle, ja suojaavia molekyylejä - pentraksiineja (C-reaktiivinen proteiini (CRP), amyloidiproteiinien esiasteita) - kiinnittyy kalvon lysoformeihin, minkä jälkeen komplementti aktivoituu järjestelmä klassista reittiä pitkin ja solujen tuhoutuminen.
Kalvon rakenne auttaa säilyttämään solun sisäisen ympäristön ominaisuudet, sen erot ulkoiseen ympäristöön. Tämän takaa solukalvon selektiivinen läpäisevyys ja aktiivisten kuljetusmekanismien olemassaolo siinä. Niiden hajoaminen suoran vaurion seurauksena, esimerkiksi tetrodotoksiinin, ouabaiinin, tetraetyyliammoniumin aiheuttamana tai jos vastaavien "pumppujen" ei saa riittävästi energiaa, johtaa solun elektrolyyttikoostumuksen häiriintymiseen, sen aineenvaihdunnan muutoksiin, häiriöihin. spesifisten toimintojen - supistuminen, viritysimpulssien johtuminen jne. Solujen ionikanavien (kalsium, natrium, kalium ja kloridi) häiriö ihmisellä voidaan myös geneettisesti määrittää näiden kanavien rakenteesta vastaavien geenien mutaatioilla. Niin sanotut kanalopatiat aiheuttavat hermoston, lihasten ja ruoansulatusjärjestelmän perinnöllisiä sairauksia. Liiallinen veden pääsy soluun voi johtaa sen repeämiseen - sytolyysiin - johtuen kalvon perforaatiosta komplementin aktivoituessa tai sytotoksisten lymfosyyttien ja luonnollisten tappajasolujen hyökkäyksestä.
Solukalvoon on rakennettu monia reseptoreita - rakenteita, jotka yhdistettynä vastaaviin spesifisiin signalointimolekyyleihin (ligandeihin) välittävät signaalin solun sisällä. Tämä tapahtuu erilaisten säätelykaskadien kautta, jotka koostuvat entsymaattisesti aktiivisista molekyyleistä, jotka aktivoituvat peräkkäin ja lopulta myötävaikuttavat erilaisten soluohjelmien, kuten kasvun ja proliferaation, erilaistumisen, liikkuvuuden, ikääntymisen ja solukuoleman, toteuttamiseen. Sääntelykaskadeja on melko paljon, mutta niiden lukumäärää ei ole vielä täysin määritetty. Reseptorijärjestelmä ja niihin liittyvät säätelykaskadit ovat olemassa myös solun sisällä; ne luovat spesifisen säätelyverkoston keskittymispisteineen, jakaumapisteineen ja lisäsignaalipolun valintaan riippuen solun toiminnallisesta tilasta, sen kehitysvaiheesta ja muiden reseptoreiden signaalien samanaikaisesta vaikutuksesta. Tämän seurauksena voi olla signaalin estyminen tai vahvistuminen, joka ohjaa sen eri säätelyreittiä pitkin. Sekä reseptorilaitteisto että signaalinvälitysreitit säätelykaskadien kautta, esimerkiksi tumaan, voivat häiriintyä johtuen geneettisestä viasta, joka esiintyy synnynnäisenä viana organismitasolla tai johtuen somaattisesta mutaatiosta tietyssä solutyypissä. Nämä mekanismit voivat vaurioitua tartunnanaiheuttajista, toksiineista ja myös muuttua ikääntymisprosessin aikana. Tämän viimeinen vaihe voi olla solun toimintojen, sen lisääntymis- ja erilaistumisprosessien häiriintyminen.
Solujen pinnalla on myös molekyylejä, joilla on tärkeä rooli solujen välisen vuorovaikutuksen prosesseissa. Näitä voivat olla soluadheesioproteiinit, histoyhteensopivuusantigeenit, kudosspesifiset, erilaistuvat antigeenit jne. Muutokset näiden molekyylien koostumuksessa häiritsevät solujen välisiä vuorovaikutuksia ja voivat aktivoida sopivia mekanismeja tällaisten solujen eliminoimiseksi, koska ne aiheuttavat tietty vaara kehon eheydelle infektion, erityisesti virusperäisen, säiliönä tai mahdollisena kasvaimen kasvun käynnistäjänä.
Solun energiansaannin rikkominen
Solun energianlähde on ruoka, jonka hajoamisen jälkeen energiaa vapautuu lopullisiksi aineiksi. Pääasiallinen energiantuotantopaikka on mitokondriot, joissa aineet hapetetaan hengitysketjun entsyymien avulla. Hapetus on pääasiallinen energian toimittaja, koska glykolyysin seurauksena samasta määrästä hapetussubstraatteja (glukoosia) vapautuu enintään 5 % energiasta hapetukseen verrattuna. Noin 60 % hapettumisen aikana vapautuvasta energiasta kertyy oksidatiivisen fosforylaation kautta korkeaenergisiin fosfaatteihin (ATP, kreatiinifosfaatti), loput haihtuvat lämpönä. Myöhemmin solu käyttää korkeaenergisiä fosfaatteja prosesseihin, kuten pumpun toimintaan, synteesiin, jakautumiseen, liikkumiseen, erittymiseen jne. On olemassa kolme mekanismia, joiden vaurioituminen voi aiheuttaa häiriön solun energiansaannissa: ensimmäinen on energia-aineenvaihdunnan entsyymien synteesimekanismi, toinen on oksidatiivisen fosforylaation mekanismi, kolmas on energian käytön mekanismi.
Elektronien kuljetuksen häiriintyminen mitokondrioiden hengitysketjussa tai ADP:n hapettumisen ja fosforylaation irtoaminen protonipotentiaalin, ATP:n muodostumisen liikkeellepaneva voiman kanssa, johtaa oksidatiivisen fosforylaation heikkenemiseen siten, että suurin osa energiasta hajoaa lämpönä ja korkeaenergisten yhdisteiden määrä vähenee. Adrenaliinin vaikutuksesta tapahtuvaa hapettumisen ja fosforylaation irtoamista käytetään homeotermisten organismien soluissa lämmöntuotannon lisäämiseen samalla kun kehon lämpötila pysyy vakiona jäähtymisen aikana tai nostaa sitä kuumeen aikana. Tyreotoksikoosissa havaitaan merkittäviä muutoksia mitokondrioiden rakenteessa ja energia-aineenvaihdunnassa. Nämä muutokset ovat alun perin palautuvia, mutta tietyn pisteen jälkeen ne muuttuvat peruuttamattomiksi: mitokondriot hajoavat, hajoavat tai turpoavat, menettävät ristikot muuttuen tyhjiöiksi ja lopulta kerääntyvät aineita, kuten hyaliinia, ferritiiniä, kalsiumia, lipofuskiinia. Keripukkipotilailla mitokondriot sulautuvat yhteen muodostaen kondriosfäärejä, mikä saattaa johtua peroksidiyhdisteiden aiheuttamasta kalvovauriosta. Merkittäviä mitokondrioiden vaurioita tapahtuu ionisoivan säteilyn vaikutuksesta normaalin solun muuttuessa pahanlaatuiseksi.
Mitokondriot ovat voimakas kalsiumionivarasto, jossa sen pitoisuus on useita suuruusluokkaa suurempi kuin sytoplasmassa. Kun mitokondriot vaurioituvat, kalsium pääsee sytoplasmaan aiheuttaen proteinaasien aktivaatiota vaurioittaen solunsisäisiä rakenteita ja häiritsemällä vastaavan solun toimintoja, esimerkiksi kalsiumkontraktuuria tai jopa "kalsiumkuolemaa" hermosoluissa. Mitokondrioiden toimintakyvyn häiriintymisen seurauksena vapaiden radikaalien peroksidiyhdisteiden muodostuminen lisääntyy jyrkästi, joilla on erittäin korkea reaktiivisuus ja jotka siksi vahingoittavat solun tärkeitä komponentteja - nukleiinihappoja, proteiineja ja lipidejä. Tämä ilmiö havaitaan niin kutsutun oksidatiivisen stressin aikana ja sillä voi olla negatiivisia seurauksia solun olemassaoloon. Siten mitokondrioiden ulkokalvon vaurioitumiseen liittyy kalvojen välisen tilan sisältämien aineiden, ensisijaisesti sytokromi C:n ja joidenkin muiden biologisesti aktiivisten aineiden vapautuminen sytoplasmaan, jotka laukaisevat ketjureaktioita, jotka aiheuttavat ohjelmoitua solukuolemaa - apoptoosia. Vapaiden radikaalien reaktiot vahingoittamalla mitokondrioiden DNA:ta vääristävät geneettistä informaatiota, joka tarvitaan tiettyjen hengitysketjuentsyymien muodostumiseen, joita syntyy erityisesti mitokondrioissa. Tämä johtaa entisestään hapettumisprosessien häiriintymiseen. Yleisesti ottaen mitokondrioiden oma geneettinen laitteisto on ytimen geneettiseen laitteistoon verrattuna vähemmän suojattu haitallisilta vaikutuksilta, jotka voivat muuttaa siihen koodattua geneettistä informaatiota. Seurauksena on, että mitokondrioiden toimintahäiriöitä esiintyy koko elämän ajan, esimerkiksi ikääntymisprosessin aikana, solun pahanlaatuisen transformaation aikana sekä munan mitokondrioiden DNA:n mutaatioon liittyvien perinnöllisten mitokondrioiden sairauksien taustalla. Tällä hetkellä on kuvattu yli 50 mitokondriomutaatiota, jotka aiheuttavat perinnöllisiä hermoston ja lihasten rappeuttavia sairauksia. Ne välittyvät lapselle yksinomaan äidiltä, koska siittiöiden mitokondriot eivät ole osa tsygoottia ja vastaavasti uutta organismia.
Geneettisen tiedon säilyttämisen ja välittämisen rikkominen
Solun ydin sisältää suurimman osan geneettisestä tiedosta ja varmistaa siten sen normaalin toiminnan. Selektiivisen geeniekspression avulla se koordinoi solujen toimintaa interfaasin aikana, tallentaa geneettistä tietoa sekä luo ja välittää geneettistä materiaalia solun jakautumisen aikana. DNA:n replikaatio ja RNA-transkriptio tapahtuvat ytimessä. Erilaiset patogeeniset tekijät, kuten ultravioletti- ja ionisoiva säteily, vapaiden radikaalien hapettuminen, kemikaalit, virukset, voivat vahingoittaa DNA:ta. On arvioitu, että jokainen lämminverisen eläimen solu kestää yhden päivän. menettää yli 10 000 emästä. Tähän pitäisi lisätä rikkomuksia kopioitaessa jaon aikana. Jos nämä vauriot jatkuvat, solu ei pystyisi selviytymään. Suoja on tehokkaiden korjausjärjestelmien olemassaolo, kuten ultraviolettiendonukleaasi, korjausreplikaatio- ja rekombinaatiokorjausjärjestelmät, jotka korvaavat DNA-vaurioita. Korjausjärjestelmien geneettiset viat aiheuttavat sairauksien kehittymistä, joka johtuu lisääntyneestä herkkyydestä DNA:ta vahingoittaville tekijöille. Tämä on xeroderma pigmentosum, samoin kuin jotkin nopeutunut ikääntymisoireyhtymät, joihin liittyy lisääntynyt taipumus kehittää pahanlaatuisia kasvaimia.
Järjestelmä, jolla säädellään DNA:n replikaatiota, lähetti-RNA:n (mRNA) transkriptiota ja geneettisen tiedon translaatiota nukleiinihapoista proteiinien rakenteeseen, on varsin monimutkainen ja monitasoinen. Tiettyjä geenejä aktivoivien, yhteensä yli 3000 transkriptiotekijöiden toiminnan laukaisevien säätelykaskadien lisäksi on olemassa myös monitasoinen säätelyjärjestelmä, jota välittävät pienet RNA-molekyylit (interfering RNA; RNAi). Ihmisen genomi, joka koostuu noin 3 miljardista puriini- ja pyrimidiiniemäksestä, sisältää vain 2 % proteiinisynteesistä vastaavista rakennegeeneistä. Loput tarjoavat säätelevien RNA:iden synteesiä, jotka samanaikaisesti transkriptiotekijöiden kanssa aktivoivat tai estävät rakennegeenien toiminnan DNA-tasolla kromosomeissa tai vaikuttavat lähetti-RNA:n (mRNA) translaatioprosesseihin polypeptidimolekyylin muodostumisen aikana. sytoplasma. Geneettisen tiedon rikkominen voi tapahtua sekä rakennegeenien että DNA:n säätelyosan tasolla vastaavien ilmenemismuotojen kanssa erilaisten perinnöllisten sairauksien muodossa.
Viime aikoina on kiinnitetty paljon huomiota geneettisen materiaalin muutoksiin, joita tapahtuu organismin yksilöllisen kehityksen aikana ja jotka liittyvät tiettyjen DNA-osien ja kromosomien estoon tai aktivoitumiseen niiden metylaatiosta, asetylaatiosta ja fosforylaatiosta johtuen. Nämä muutokset jatkuvat pitkään, joskus koko organismin elinkaaren ajan alkion synnystä vanhuuteen, ja niitä kutsutaan epigenomiseksi perinnöllisyydeksi.
Myös mitoottista kiertoa säätelevät järjestelmät (tekijät) estävät solujen lisääntymisen, joilla on muuttunut geneettinen tieto. Ne ovat vuorovaikutuksessa sykliinistä riippuvaisten proteiinikinaasien ja niiden katalyyttisten alayksiköiden - sykliinien - kanssa ja estävät solua käymästä läpi koko mitoottisen syklin, pysäyttäen jakautumisen presynteettisen ja synteettisen vaiheen rajalla (G1/S-lohko), kunnes DNA:n korjaus on valmis. ja jos tämä on mahdotonta, ne käynnistävät ohjelmoidut kuolemasolut. Näihin tekijöihin kuuluvat p53-geeni, jonka mutaatio aiheuttaa kontrollin menettämisen transformoituneiden solujen lisääntymisen suhteen; sitä havaitaan lähes 50 %:ssa ihmisten syövistä. Mitoosisyklin toinen tarkistuspiste on G2/M rajalla. Tässä kromosomimateriaalin oikeaa jakautumista tytärsolujen välillä mitoosissa tai meioosissa ohjataan käyttämällä mekanismeja, jotka ohjaavat solun karaa, keskustaa ja sentromeereja (kinetokoreja). Näiden mekanismien tehottomuus johtaa kromosomien tai niiden osien jakautumisen häiriintymiseen, mikä ilmenee kromosomien puuttumisena yhdessä tytärsoluista (aneuploidia), ylimääräisen kromosomin läsnäolosta (polyploidia), solujen erottumisesta. osa kromosomista (deleetio) ja sen siirto toiseen kromosomiin (translokaatio) . Tällaisia prosesseja havaitaan hyvin usein pahanlaatuisesti degeneroituneiden ja transformoituneiden solujen lisääntymisen aikana. Jos näin tapahtuu sukusolujen meioosin aikana, se johtaa joko sikiön kuolemaan alkionkehityksen varhaisessa vaiheessa tai kromosomisairaaseen organismin syntymiseen.
Hallitsematon solujen lisääntyminen kasvaimen kasvun aikana tapahtuu mutaatioiden seurauksena geeneissä, jotka säätelevät solujen lisääntymistä ja joita kutsutaan onkogeeneiksi. Yli 70 tällä hetkellä tunnetusta onkogeenista useimmat kuuluvat solujen kasvun säätelyn komponentteihin, joitain edustavat geeniaktiivisuutta säätelevät transkriptiotekijät sekä solun jakautumista ja kasvua estävät tekijät. Toinen lisääntyvien solujen liiallista laajenemista (leviämistä) rajoittava tekijä on kromosomien päiden lyheneminen - telomeerit, jotka eivät pysty täysin replikoitumaan puhtaasti steerisen vuorovaikutuksen seurauksena, joten jokaisen solujakautumisen jälkeen telomeerit lyhenevät tietty osa pohjasta. Siten aikuisen organismin lisääntyvät solut tietyn jakautumismäärän jälkeen (yleensä 20 - 100 organismin tyypistä ja iästä riippuen) kuluttavat telomeeripituuden ja kromosomien lisäreplikaatio pysähtyy. Tätä ilmiötä ei esiinny siittiöiden epiteelissä, enterosyyteissä ja alkiosoluissa johtuen telomeraasientsyymistä, joka palauttaa telomeerin pituuden jokaisen jakautumisen jälkeen. Useimmissa aikuisten organismien soluissa telomeraasi on estetty, mutta valitettavasti se aktivoituu kasvainsoluissa.
Ytimen ja sytoplasman välinen yhteys ja aineiden kuljetus molempiin suuntiin tapahtuu ydinkalvon huokosten kautta erityisten energiaa kuluttavien kuljetusjärjestelmien kanssa. Tällä tavalla ytimeen kuljetetaan energiaa ja muoviaineita, signaalimolekyylejä (transkriptiotekijöitä). Käänteinen virtaus kuljettaa sytoplasmaan mRNA- ja siirto-RNA-molekyylejä (tRNA), ribosomeja, joita tarvitaan proteiinisynteesiin solussa. Sama aineiden kuljetusreitti on ominaista myös viruksille, erityisesti kuten HIV:lle. Ne siirtävät geneettisen materiaalinsa isäntäsolun ytimeen, jolloin se liitetään edelleen isäntägenomiin ja siirretään vasta muodostunut virus-RNA sytoplasmaan uusien viruspartikkelien proteiinien synteesiä varten.
Synteesiprosessien rikkominen
Proteiinin synteesiprosessit tapahtuvat endoplasmisen retikulumin säiliöissä, jotka ovat tiiviisti yhteydessä ydinkalvon huokosiin, joiden kautta ribosomit, tRNA ja mRNA pääsevät endoplasmiseen retikulumiin. Täällä suoritetaan polypeptidiketjujen synteesi, jotka myöhemmin saavat lopullisen muotonsa agranulaarisessa endoplasmisessa retikulumissa ja lamellaarisessa kompleksissa (Golgi-kompleksi), jossa ne käyvät läpi translaation jälkeisen modifikoinnin ja yhdistyvät hiilihydraatti- ja lipidimolekyylien kanssa. Vasta muodostuneet proteiinimolekyylit eivät jää synteesikohtaan, vaan monimutkaisen säädellyn prosessin kautta, ns. proteiinikineesi, siirretään aktiivisesti solun eristettyyn osaan, jossa ne suorittavat aiotun tehtävänsä. Tässä tapauksessa erittäin tärkeä vaihe on siirretyn molekyylin strukturointi sopivaksi avaruudelliseksi konfiguraatioksi, joka pystyy suorittamaan sen luontaisen tehtävän. Tämä strukturoituminen tapahtuu erityisten entsyymien avulla tai erikoistuneiden proteiinimolekyylien - chaperonien - matriisilla, jotka auttavat äskettäin muodostunutta tai ulkoisen vaikutuksen vaikutuksesta muuttunutta proteiinimolekyyliä saamaan oikean kolmiulotteisen rakenteen. Jos soluun kohdistuu haitallinen vaikutus, kun on mahdollista proteiinimolekyylien rakenteen häiriintyminen (esimerkiksi kehon lämpötilan nousun, tarttuvan prosessin, myrkytyksen yhteydessä), chaperonien pitoisuus solussa kasvaa. terävästi. Siksi tällaisia molekyylejä kutsutaan myös stressiproteiinit, tai lämpöshokkiproteiinit. Proteiinimolekyylin rakenteen rikkominen johtaa kemiallisesti inerttien konglomeraattien muodostumiseen, jotka kerrostuvat soluun tai sen ulkopuolelle amyloidoosin, Alzheimerin taudin jne. aikana. Joskus valmiiksi rakennettu samanlainen molekyyli voi toimia matriisina, ja tässä Jos ensisijainen strukturointi tapahtuu väärin, myös kaikki myöhemmät molekyylit ovat viallisia. Tämä tilanne esiintyy niin sanotuissa prionisairauksissa (skrapie lampailla, raivoisat lehmät, kuru, Creutzfeldt-Jakobin tauti ihmisillä), kun jonkin hermosolun kalvoproteiinin vika aiheuttaa myöhemmän inerttien massojen kertymisen solun sisään. ja sen elintoimintojen häiriintyminen.
Solun synteesiprosessien häiriintyminen voi tapahtua sen eri vaiheissa: RNA:n transkriptio tumassa, polypeptidien translaatio ribosomeissa, translaation jälkeinen modifikaatio, beigen molekyylin hypermetylaatio ja glykosylaatio, proteiinien kuljettaminen ja jakautuminen soluun sekä niiden poisto. ulospäin. Tässä tapauksessa voidaan havaita ribosomien lukumäärän lisääntymistä tai vähenemistä, polyribosomien hajoamista, rakeisen endoplasmisen retikulumin vesisäiliöiden laajenemista, ribosomien katoamista ja rakkuloiden ja vakuolien muodostumista. Siten vaalean uikkun myrkytyksessä RNA-polymeraasientsyymi vaurioituu, mikä häiritsee transkriptiota. Kurkkumätätoksiini inaktivoi elongaatiotekijän häiritsee translaatioprosesseja aiheuttaen sydänlihasvaurioita. Syynä joidenkin spesifisten proteiinimolekyylien synteesin häiriintymiseen voivat olla tartunnanaiheuttajat. Esimerkiksi herpesvirukset estävät MHC-antigeenimolekyylien synteesiä ja ilmentymistä, minkä ansiosta ne voivat osittain välttää immuunivasteen torjuntaa - akuutin tulehduksen välittäjien synteesiä. Epätavallisten proteiinien ilmaantuminen voi pysäyttää niiden hajoamisen ja johtaa inertin tai jopa myrkyllisen materiaalin kerääntymiseen. Tätä voi jossain määrin helpottaa hajoamisprosessien häiriintyminen.
Hajoamisprosessien häiriintyminen
Samanaikaisesti solun proteiinisynteesin kanssa sen hajoaminen tapahtuu jatkuvasti. Normaaleissa olosuhteissa tällä on tärkeä säätely- ja muodollinen merkitys esimerkiksi entsyymien, proteiinihormonien ja mitoottisen syklin proteiinien inaktiivisten muotojen aktivoituessa. Normaali solujen kasvu ja kehitys edellyttävät tarkasti hallittua tasapainoa proteiinien ja organellien synteesin ja hajoamisen välillä. Proteiinisynteesiprosessissa synteesilaitteiston toimintavirheiden, proteiinimolekyylin epänormaalin rakenteen ja sen kemiallisten ja bakteerien aiheuttamien vaurioiden vuoksi muodostuu kuitenkin jatkuvasti melko suuri määrä viallisia molekyylejä. Joidenkin arvioiden mukaan niiden osuus on noin kolmannes kaikista syntetisoiduista proteiineista.
Nisäkässoluilla on useita pää proteiinien tuhoamistapoja: lysosomaalisten proteaasien (pentidihydrolaasien), kalsiumista riippuvaisten proteinaasien (endopeptidaasien) ja proteasomijärjestelmän kautta. Lisäksi on olemassa myös erikoistuneita proteinaaseja, kuten kaspaaseja. Tärkein organelli, jossa aineiden hajoaminen tapahtuu eukaryoottisoluissa, on lysosomi, joka sisältää lukuisia hydrolyyttisiä entsyymejä. Lysosomien ja fagolysosomien endosytoosiprosessien ja erityyppisten autofagioiden seurauksena sekä vialliset proteiinimolekyylit että kokonaiset organellit tuhoutuvat: vaurioituneet mitokondriot, plasmakalvon osat, jotkut solunulkoiset proteiinit ja erittyvien rakeiden sisältö.
Tärkeä proteiinien hajoamismekanismi on proteasomi, monikatalyyttinen, monimutkaisen rakenteen omaava proteinaasirakenne, joka sijaitsee sytosolissa, tumassa, endoplasmisessa retikulumissa ja solukalvolla. Tämä entsyymijärjestelmä on vastuussa vaurioituneiden proteiinien sekä terveiden proteiinien hajottamisesta, jotka on poistettava normaalin solun toiminnan kannalta. Tässä tapauksessa tuhottavat proteiinit yhdistetään alustavasti spesifiseen polypeptidiin, ubikvitiiniin. Kuitenkin ei-ubikvitinoituneet proteiinit voivat myös osittain tuhoutua proteasomeissa. Proteasomeissa olevien proteiinimolekyylien hajoaminen lyhyiksi polypeptideiksi (prosessointi) ja niiden esittely yhdessä tyypin I MHC-molekyylien kanssa on tärkeä linkki kehon antigeenisen homeostaasin immuunivalvonnassa. Kun proteasomin toiminta heikkenee, vaurioituneita ja tarpeettomia proteiineja kerääntyy, mikä seuraa solujen ikääntymistä. Sykliinistä riippuvaisten proteiinien hajoamisen rikkominen johtaa solujen jakautumisen häiriintymiseen, erittyvien proteiinien hajoamiseen - kystofibroosin kehittymiseen. Sitä vastoin proteasomien toiminnan lisääntyminen liittyy kehon ehtymiseen (aids, syöpä).
Geneettisesti määrätyillä proteiinien hajoamishäiriöillä organismi ei ole elinkelpoinen ja kuolee alkion synnyn alkuvaiheessa. Jos rasvojen tai hiilihydraattien hajoaminen häiriintyy, esiintyy varastoinnin sairauksia (tesaurismoosi). Tällöin solun sisään kerääntyy liiallinen määrä tiettyjä aineita tai niiden epätäydellisen hajoamisen tuotteita - lipidejä, polysakkarideja, mikä vahingoittaa merkittävästi solun toimintaa. Tämä havaitaan useimmiten maksan epiteelisoluissa (hepatosyyteissä), hermosoluissa, fibroblasteissa ja makrofagosyyteissä.
Aineiden hajoamisprosessien hankitut häiriöt voivat syntyä patologisten prosessien seurauksena (esimerkiksi proteiinin, rasvan, hiilihydraattien ja pigmentin rappeutuminen), ja niihin liittyy epätavallisten aineiden muodostuminen. Häiriöt lysosomaalisessa proteolyysijärjestelmässä johtavat vähentyneeseen sopeutumiseen paaston aikana tai lisääntyneeseen stressiin sekä tiettyjen endokriinisten toimintahäiriöiden esiintymiseen - insuliinin, tyroglobuliinin, sytokiinien ja niiden reseptorien alentumiseen. Heikentynyt proteiinien hajoaminen hidastaa haavan paranemisnopeutta, aiheuttaa ateroskleroosin kehittymistä ja vaikuttaa immuunivasteeseen. Hypoksia, solunsisäisen pH:n muutokset, säteilyvaurio, jolle on ominaista kalvon lipidien lisääntynyt peroksidaatio, sekä lysosomotrooppisten aineiden - bakteerien endotoksiinien, myrkyllisten sienten metaboliittien (sporofusariini), piioksidikiteiden - vaikutuksen alaisena - lysosomikalvon stabiilisuus Muutoksissa aktivoituneita lysosomaalisia entsyymejä vapautuu sytoplasmaan, mikä aiheuttaa solurakenteiden tuhoutumisen ja sen kuoleman.
Luku 1
SOLUFYSIOLOGIAN PERUSTEET
I. Dudel
Plasmakalvo . Eläinsoluja rajoittaa plasmakalvo (kuva 1.1). Pysähdymme sen rakenteeseen, joka on hyvin samanlainen kuin monien solunsisäisten kalvojen rakenne, hieman yksityiskohtaisemmin. Kalvon päämatriisi koostuu lipidit, pääasiassa fosfatidyylikoliinia. Nämä lipidit koostuvat hydrofiilisestä pääryhmästä, johon on kiinnittynyt pitkiä hydrofobisia hiilivetyketjuja. Vedessä tällaiset lipidit muodostavat spontaanisti 4–5 nm paksuisen kaksikerroksisen kalvon, jossa hydrofiiliset ryhmät ovat vesipitoista väliainetta vasten ja hydrofobiset hiilivetyketjut on järjestetty kahteen riviin muodostaen vedettömän lipidifaasin. Solukalvot ovat tämän tyyppisiä lipidikaksoiskerroksia ja sisältävät glykolipidejä, kolesterolia ja fosfolipidejä (kuva 1.2). Glykolipidien hydrofiilinen osa muodostuu oligosakkarideista. Glykolipidit sijaitsevat aina plasmakalvon ulkopinnalla, ja molekyylin oligosakkaridiosa on suunnattu kuin hius, joka on upotettu ympäristöön. Kolesterolimolekyylit ovat hajallaan fosfolipidien joukossa lähes yhtä suuria määriä, ja ne stabiloivat kalvoa. Erilaisten lipidien jakautuminen kalvon sisä- ja ulkokerroksessa ei ole sama, ja jopa yhden kerroksen sisällä on alueita, joihin tietyntyyppiset lipidit ovat keskittyneet. Tämä epätasainen jakautuminen
Riisi. 1.1. Kaaviopiirros solusta, joka näyttää tärkeimmät organellit
Sillä on luultavasti jokin, toistaiseksi epäselvä, toiminnallinen merkitys.
Tärkeimmät toiminnalliset elementit, jotka on upotettu kalvon suhteellisen inerttiin lipidimatriisiin, ovat oravia(Kuva 1.2). Proteiinia on 25-75 painoprosenttia eri kalvoissa, mutta koska proteiinimolekyylit ovat paljon suurempia kuin lipidimolekyylit, 50 painoprosenttia vastaa 1 proteiinimolekyylin suhdetta 50 lipidimolekyyliin. Jotkut proteiinit tunkeutuvat kalvoon sen ulkopuolelta sisäpinnalle, kun taas toiset ovat kiinnittyneet yhteen kerrokseen. Proteiinimolekyylit on yleensä suunnattu siten, että niiden hydrofobiset ryhmät on upotettu lipidikalvoon ja kalvon pinnalla olevat polaariset hydrofiiliset ryhmät ovat upotettuina vesifaasiin. Monet kalvon ulkopinnalla olevat proteiinit ovat glykoproteiineja; niiden hydrofiiliset sakkaridiryhmät kohtaavat solunulkoisen ympäristön.
Solunsisäisten organellien kalvojärjestelmät .
Noin puolet solutilavuudesta ovat soluelimet, jotka on eristetty sytosolista kalvojen avulla. Solunsisäisten organellien kalvojen kokonaispinta-ala on vähintään 10 kertaa suurempi kuin plasmakalvon pinta. Yleisimmin käytetty kalvojärjestelmä on endoplasminen verkkokalvo, edustaa verkkoa

Riisi. 1.2.Plasmakalvon kaaviomainen esitys. Proteiinit on upotettu fosfolipidikaksoiskerrokseen, ja osa niistä tunkeutuu kaksoiskerrokseen, kun taas toiset ovat vain ankkuroituja ulko- tai sisäkerrokseen
erittäin kierretyt putket tai pussimaiset pitkänomaiset rakenteet; suuret alueet endoplasmisesta retikulumista ovat täynnä ribosomeja; tällaista verkkoa kutsutaan rakeiseksi tai karkeaksi (kuva 1.1). Golgin laite koostuu myös kalvoon sitoutuneista lamelleista, joista irtoaa rakkuloita tai rakkuloita (kuva 1.1). Lysosomit ja peroksisomit- Nämä ovat pieniä erikoistuneita rakkuloita. Kaikissa näissä erilaisissa organelleissa kalvo ja sen kattama tila sisältävät erityisiä entsyymijoukkoja; Organellien sisällä kerääntyy erityisiä aineenvaihduntatuotteita, joita käytetään organellien eri toimintojen suorittamiseen.
YdinJa mitokondriot eroavat siinä, että jokaista näistä organelleista ympäröi kaksi kalvoa. Ydin vastaa aineenvaihdunnan kineettisestä hallinnasta; mitokondrioiden laskostunut sisäkalvo on oksidatiivisen aineenvaihdunnan paikka; täällä syntetisoidaan pyruvaatin tai rasvahappojen hapettumisen vuoksi korkeaenerginen yhdiste adenosiinitrifosfaatti (ATP tai ATP).
Sytoskeleton . Organelleja ympäröivää sytoplasmaa ei voida millään tavalla pitää amorfisena; sen läpäisee sytoskeletaalinen verkko. Sytoskeletoni koostuu mikrotubuluksista, aktiinifilamenteista ja välifilamenteista (kuva 1.1). Mikrotubulukset niiden ulkohalkaisija on noin 25 nm; ne muodostuvat tavallisen polymeerin tapaan tubuliiniproteiinimolekyylien kokoonpanon seurauksena. aktiinifilamentit – supistuvat kuidut, jotka sijaitsevat lähellä kalvoa olevaa kerrosta ja koko solua, osallistuvat pääasiassa liikkeeseen liittyviin prosesseihin. Välilangat koostuvat eri kemiallisen koostumuksen lohkoista erityyppisissä soluissa; ne muodostavat erilaisia yhteyksiä kahden muun edellä mainitun solun tukirankaelementin välille. Organellit ja plasmakalvo liittyvät myös sytoskeletoniin, joka ei vain ylläpidä solun muotoa ja soluelinten sijaintia siinä, vaan määrittää myös muutokset solun muodossa ja sen liikkuvuudessa.
Sytosoli . Noin puolet solutilavuudesta on sytosolien käytössä. Koska se sisältää noin 20 painoprosenttia proteiinia, se on enemmän geeliä kuin vesiliuosta. Pienet molekyylit, mukaan lukien orgaaniset ja epäorgaaniset ionit, liuotettuna vesifaasiin. Ionien vaihto tapahtuu solun ja ympäristön välillä (solunulkoinen tila); näitä aineenvaihduntaprosesseja käsitellään seuraavassa osassa. Ionien pitoisuus solunulkoisessa tilassa ylläpidetään huomattavalla tarkkuudella vakiotasolla; kunkin ionin solunsisäisellä pitoisuudella on myös tietty taso, joka on erilainen kuin solun ulkopuolella (taulukko 1.1). Yleisin kationi solunulkoisessa ympäristössä on Na+ solussa sen pitoisuus on yli 10 kertaa pienempi. Päinvastoin, K +:n pitoisuus on korkein solun ulkopuolella, se on pienempi kuin suuruusluokkaa. Suurin gradientti solunulkoisten ja solunsisäisten pitoisuuksien välillä on Ca 2+:lla, vapaiden ionien pitoisuus solun sisällä on vähintään 10 000 kertaa pienempi kuin sen ulkopuolella. Kaikki ionit eivät liukene sytosoliin, osa niistä on adsorboitunut proteiineihin tai kertynyt organelleihin. Esimerkiksi Ca 2+:n tapauksessa sitoutuneita ioneja on paljon enemmän kuin vapaita. Suurin osa sytosolisista proteiineista on entsyymejä, joiden kanssa suoritetaan monia väliaineenvaihdunnan prosesseja: glykolyysi ja glukoneogeneesi, aminohappojen synteesi tai tuhoutuminen, proteiinisynteesi ribosomeissa (kuva 1.1). Sytosoli sisältää myös lipidipisaroita ja glykogeenirakeita, jotka toimivat tärkeiden molekyylien varastoina.
Taulukko 1.1.Solunsisäiset ja ekstrasellulaariset ionipitoisuudet homeotermisten eläinten lihassoluissa. A – - "korkean molekyylipainon omaavat soluanionit"
|
Solunsisäinen pitoisuus |
Solunulkoinen pitoisuus |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca 2+ |
10 -7 10 -8 mmol/l |
Ca 2+ |
2 mmol/l |
|
C l - |
4 mmol/l |
C1 – |
120 mmol/l |
|
HCO 3 – |
8 mmol/l |
HCO 3 – |
27 mmol/l |
|
A – |
155 mmol/l |
muut kationeja |
5 mmol/l |
|
Lepopotentiaali -90 mV |
|||
1.2. Aineiden vaihto solun ja ympäristön välillä
Olemme kuvanneet lyhyesti solun rakenteen, jotta voimme käyttää tätä kuvausta tarkastellaksemme solufysiologian perusteita. Solua ei missään tapauksessa voida pitää staattisena kokonaisuutena, koska eri solunsisäisten osastojen välillä sekä osastojen ja ympäristön välillä tapahtuu jatkuvaa aineiden vaihtoa. Solurakenteet ovat dynaamisessa tasapainossa, ja solujen vuorovaikutus keskenään ja ulkoisen ympäristön kanssa on välttämätön edellytys toimivan organismin elämän ylläpitämiselle. Tässä luvussa tarkastellaan tällaisen vaihdon perusmekanismeja. Seuraavissa luvuissa näitä mekanismeja tarkastellaan hermosoluun ja sen toimintoihin soveltuvina;
kuitenkin samat mekanismit ovat kaikkien muiden elinten toiminnan taustalla.
Diffuusio.Yksinkertaisin aineen liikkumisprosessi on diffuusio. Liuoksissa (tai kaasuissa) atomit ja molekyylit liikkuvat vapaasti ja pitoisuuksien erot tasapainottuvat diffuusion ansiosta. Tarkastellaan kahta nesteellä tai kaasulla täytettyä tilavuutta (kuva 1.3), joissa aineilla on pitoisuuksia c 1 ja c 2 ja erotettu kerroksella, jonka pinta-ala ja paksuus on A d. Aineen virtaus m aikana t kuvattu Fickin ensimmäinen diffuusiolaki:
dm/ dt= DA/ d ( C 1 –C 2)=DA/ dD C(1)
jossa D on diffuusiokerroin, vakio tietylle aineelle, liuottimelle ja lämpötilalle. Yleisemmässä muodossa konsentraatioerolle dc etäisyydellä dx
dm/dt= –D A dc/dx,(2)
virtaus osan A läpi on verrannollinen pitoisuusgradienttiin dc/dx . Miinusmerkki näkyy yhtälössä, koska pitoisuuden muutos x-suunnassa on negatiivinen.
Diffuusio on tärkein prosessi, jonka ansiosta suurin osa vesiliuosten molekyyleistä liikkuu lyhyitä matkoja. Tämä koskee myös niiden liikkumista solussa, mikäli kalvot eivät estä diffuusiota. Monet aineet voivat diffundoitua vapaasti lipidikalvojen läpi, erityisesti vesi ja liuenneet kaasut, kuten O 2 ja CO 2 . Rasvaliukoinen

Riisi. 1.3.Kvantitatiivinen diffuusiokaavio. Nämä kaksi tilaa erotetaan paksuuskerrokselladja alue A. C – suuri hiukkaspitoisuus tilavuuden vasemmassa osassa, C: – pieni hiukkaspitoisuus oikealla osat, vaaleanpunainen pinta– pitoisuusgradientti diffuusiokerroksessa. Diffuusiovirtaus dm/dt – katso yhtälö (1)
aineet myös diffundoituvat hyvin kalvojen läpi; Tämä koskee myös melko pienikokoisia polaarisia molekyylejä, kuten etanolia ja ureaa, kun taas sokerit kulkevat lipidikerroksen läpi vaikeasti. Samaan aikaan lipidikerrokset ovat käytännössä läpäisemättömiä varautuneille molekyyleille, mukaan lukien jopa epäorgaaniset ionit. Ei-elektrolyyttien osalta diffuusioyhtälö (1) muunnetaan yleensä yhdistämällä kalvon ja diffuusioaineen ominaisuudet yhdeksi parametri-läpäisevyys (P):
dm/dt = P AD c.(3)
Kuvassa 1.4 verrataan lipidikalvon läpäisevyys (P) eri molekyyleille.
Diffuusio kalvon huokosten läpi . Plasmakalvo (ja muut solukalvot) ei läpäise ainoastaan lipidikerroksen läpi diffundoituvia aineita, vaan myös monia ioneja, sokereita, aminohappoja ja nukleotideja. Nämä aineet läpäisevät kalvon muodostuneiden huokosten kautta kuljetusproteiinit, upotettu kalvoon. Tällaisten proteiinien sisällä on vedellä täytetty kanava, jonka halkaisija on alle 1 nm, jonka läpi pienet molekyylit voivat diffundoitua. Ne liikkuvat pitoisuusgradienttia pitkin, ja jos ne kantavat varausta, niin niiden liikettä kanavien läpi säätelee myös kalvopotentiaali. Kalvokanavat ovat suhteellisen selektiivisiä

Riisi. 1.4.Keinotekoisten lipidikaksoiskerrosten läpäisevyys eri aineille
niiden läpi kulkevien molekyylien tyypin suhteen on olemassa esimerkiksi kalium-, natrium- ja kalsiumkanavia, joista jokainen on läpäisemätön melkein mille tahansa ionille paitsi tietylle ionille. Sellainen valikoivuus määräytyy kanavan seinämien sitoutumiskohtien varauksen tai rakenteen perusteella, mikä helpottaa tietyn molekyylin kulkeutumista ja estää muiden aineiden tunkeutumisen kanavan läpi (kuva 1). 1,5, A) .
Käyttäytymisen takana kalvon ionikanavia helppo havaita, koska ionien liikkeestä tuleva virta voidaan mitata jopa yhdelle kanavalle. On osoitettu, että kanavat muuttavat tilansa spontaanisti ja suurella taajuudella avoimesta suljettuun. Kaliumkanavalle on tunnusomaista virtapulssit, joiden amplitudi on noin 2 pA (2 10 –12 A) ja joiden kesto on useita millisekunteja (ks. kuva 2.12, s. 37) [3]. Tänä aikana sen läpi kulkee kymmeniä tuhansia ioneja. Proteiinien siirtymistä konformaatiosta toiseen tutkitaan röntgendiffraktiolla, Mössbauer-spektroskopialla ja ydinmagneettisella resonanssilla (NMR). Proteiinit ovat siis hyvin dynaamisia, liikkuvia rakenteita, ja proteiinin läpi kulkeva kanava ei ole vain jäykkä, vedellä täytetty putki (kuva 1.5, A), vaan nopeasti liikkuvien molekyyliryhmien ja varausten labyrintti. Tämä kanavan dynaaminen ominaisuus näkyy kanavan energiaprofiili, esitetty kuvassa. 1.5, B. Tässä abskissa-akseli edustaa kanavan pituutta ulkoisesta liuoksesta, jonka ionipitoisuus on C 0 ja potentiaali 0, sisäiseen liuokseen, jonka pitoisuus on C 1 ja potentiaali E. Ordinaatta-akseli

Riisi. 1.5.A. Kaavio proteiinista, joka muodostaa plasmamembraanin lipidikaksoiskerrokseen upotetun kaliumkanavan. Kanavan "seinään" on kiinnitetty neljä negatiivista varausta. B. Kuvassa 1 esitetyn kanavan kaavamainen energiaprofiili. A. Ordinaattinen akseli näyttää kineettisen energian arvot, jotka tarvitaan kanavan läpikulkuun; pitkin abskissa-akselia - kalvon sisä- ja ulkopinnan välinen etäisyys. Energiaminimit vastaavat positiivisesti varautuneiden ionien sitoutumiskohtia kiinteillä negatiivisilla varauksilla kanavan seinämässä. Energiamaksimit vastaavat kanavan diffuusioesteitä. Kanavaproteiinin konformaation oletetaan värähtelevän spontaanisti; energiaprofiilivaihtoehdot näytetään yhtenäisin ja katkoviivoin; nämä värähtelyt helpottavat suuresti ionien sitoutumista energiaesteen ylittämisessä (muokattu)
ionienergiatasot kanavan sitoutumiskohdissa esitetään; kaavion huippu edustaa permeabiliteettiestettä, joka ionienergian on ylitettävä päästäkseen kanavaan, ja "dip" kuvaajassa edustaa suhteellisen vakaata tilaa (sitoutuminen). Huolimatta energiahuipun esteestä ioni voi tunkeutua kanavaan, jos energiaprofiili muuttuu spontaanisti; ioni voi näin ollen yhtäkkiä löytää itsensä energiahuipun "toiselta puolelta" ja jatkaa liikkumista soluun. Ionin varauksesta, koosta ja hydrataatioasteesta sekä sen kyvystä sitoutua kanavan seinämärakenteisiin riippuen kanavan energiaprofiili vaihtelee eri ioneilla, mikä saattaa selittää yksittäisten kanavatyyppien selektiivisyyden.
Ionien diffuusiotasapaino . Erilaisten ionien diffuusion kalvokanavien läpi pitäisi johtaa solunulkoisen ja solunsisäisen ympäristön välisten pitoisuuksien erojen eliminointiin. Kuten taulukosta kuitenkin näkyy. 1.1, tällaisia eroja on edelleen, joten niitä täytyy olla tasapaino diffuusion ja muiden kalvon läpi kulkevien kuljetusprosessien välillä. Seuraavat kaksi osaa käsittelevät tapoja, joilla tämä tasapaino saavutetaan. Ionien tapauksessa diffuusiotasapainoon vaikuttaa niiden varaus. Varautumattomien molekyylien diffuusio varmistetaan pitoisuuksien erolla DC , ja kun pitoisuudet tasoittuvat, itse kuljetus pysähtyy. Lisäksi sähkökenttä vaikuttaa varautuneisiin hiukkasiin. Esimerkiksi kun kaliumioni poistuu solusta pitoisuusgradienttiaan pitkin, se kantaa yhden positiivisen varauksen. Siten solunsisäinen ympäristö varautuu negatiivisemmin, mikä johtaa potentiaalieroon kalvon poikki. Solunsisäinen negatiivinen varaus estää uusia kaliumioneja poistumasta solusta, ja solusta lähtevät ionit lisäävät kalvon varausta entisestään. Kaliumionien virtaus pysähtyy, kun sähkökentän toiminta kompensoi pitoisuuserosta johtuvaa diffuusiopainetta. Ionit kulkevat edelleen kalvon läpi, mutta yhtä paljon molempiin suuntiin. Siksi tietyllä erolla kalvon ionipitoisuuksissa on olemassa tasapainopotentiaali E ioni jolloin ionien virtaus kalvon läpi pysähtyy. Tasapainopotentiaali voidaan määrittää helposti käyttämällä Nernstin yhtälöt:
Eioni= RT/ zF* lnC ulos/ C sisään(4)
missä R – kaasuvakio, T – absoluuttinen lämpötila, z – ionivalenssi (negatiivinen anioneille) C ulos - solunulkoinen ionipitoisuus, C sisään - solunsisäinen ionipitoisuus, F Faradayn numero. Jos yhtälöön korvataan vakiot, niin kehon lämpötilassa (T = 310 K) kaliumionien tasapainopotentiaali E K on yhtä suuri kuin:
Ek= –61 mB log /(5)
Jos [ K + ulos ]/[ K + sisään ] = 39, kuten taulukosta ilmenee. 1.1 sitten
Ek= –61 m B log 39= –97 mV.
Itse asiassa havaittiin, että kaikilla soluilla on kalvopotentiaali; nisäkkäiden lihassoluissa sen taso on noin -90 mV. Olosuhteista ja suhteellisista ionipitoisuuksista riippuen solujen kalvopotentiaali voi vaihdella -40 - -120 mV. Yllä olevan esimerkin solulle (taulukko 1.1) lepojännite, noin -90 mV, osoittaa, että kalium-ionien vuot kalvokanavien läpi ovat suunnilleen tasapainossa. Tämä ei ole yllättävää, koska lepokalvossa kaliumkanavien avoin tila on todennäköisimmin, ts. kalvo läpäisee parhaiten kaliumioneja. Kalvopotentiaalin määräävät kuitenkin muiden ionien virtaukset.
Se, kuinka helposti varauksettomat hiukkaset voivat diffundoitua kalvon läpi, on kvantifioitu yhtälössä (3). Varautuneiden hiukkasten läpäisevyys kuvataan hieman monimutkaisemmalla yhtälöllä:
P= m RT/ dF(6)
Missä m– ionien liikkuvuus kalvossa, d – kalvon paksuus, R, T ja F – tunnetut termodynaamiset vakiot. Tällä tavalla määritettyjä eri ionien permeabiliteettiarvoja voidaan käyttää kalvopotentiaalin laskemiseen Em , kun kalium-, natrium- ja kloori-ionit kulkevat kalvon läpi samanaikaisesti (läpäisevyydellä P K, P Na ja P Cl vastaavasti). Potentiaalin oletetaan laskevan tasaisesti kalvossa siten, että kentänvoimakkuus on vakio. Tässä tapauksessa se pätee Goldmanin yhtälö tai vakiokenttäyhtälö :
Em = R T/ F * ln (P K + P Na + P Cl )/ (P K + P Na + P Cl ) (7)
Useimmille solukalvoille P K noin 30 kertaa korkeampi kuin P Na (katso myös kohta 1.3). Suhteellinen arvo P Cl vaihtelee suuresti; monille kalvoille P Cl pieni verrattuna R:ään K kuitenkin muille (esimerkiksi luurankolihaksissa) P Cl , huomattavasti korkeampi kuin P K.
Aktiivinen kuljetus, natriumpumppu . Edellisessä osiossa kuvataan ionien passiivista diffuusiota ja siitä johtuvaa kalvopotentiaalia tietyillä intra- ja ekstrasellulaarisilla ionipitoisuuksilla. Tämän prosessin seurauksena ionien pitoisuus solun sisällä ei kuitenkaan stabiloitu automaattisesti, koska kalvo
potentiaali on hieman elektronegatiivisempi kuin EK, ja paljon verrattuna E Na (noin +60 mV). Diffuusiosta johtuen solunsisäiset ionipitoisuudet, ainakin kalium ja natrium, on tasattava solunulkoisten kanssa. Ionigradientin stabiilisuus saavutetaan aktiivisella kuljetuksella: kalvoproteiinit kuljettavat ioneja kalvon läpi sähkö- ja/tai pitoisuusgradientteja vastaan kuluttaen tähän metabolista energiaa. Aktiivikuljetuksen tärkein prosessi on työ Na/K – pumppu, joka on lähes kaikissa soluissa;
pumppu pumppaa natriumioneja ulos kennosta ja samalla pumppaa kaliumioneja kennoon. Tämä varmistaa alhaisen solunsisäisen natriumionipitoisuuden ja korkean kaliumionipitoisuuden (taulukko 1.1). Natrium-ionien pitoisuusgradientilla kalvolla on erityisiä toimintoja, jotka liittyvät tiedon välittämiseen sähköimpulssien muodossa (katso kohta 2.2), sekä muiden aktiivisten kuljetusmekanismien ylläpitoon ja solutilavuuden säätelyyn (katso alla). Siksi ei ole yllättävää, että yli 1/3 kennon kuluttamasta energiasta kuluu Na/K-pumppuun, ja joissakin aktiivisimmista soluista jopa 70 % energiasta kuluu sen toimintaan.
Na/K-kuljetusproteiini on ATPaasi. Kalvon sisäpinnalla se pilkkoo ATP:n ADP:ksi ja fosfaatiksi (kuva 1.6). Yhden ATP-molekyylin energialla siirretään kolme natrium-ionia solusta ja samanaikaisesti kaksi kalium-ionia soluun, eli yhteensä yksi positiivinen varaus poistuu solusta yhdessä syklissä. Siten Na/K-pumppu on sähkögeeninen(luo sähkövirran kalvon läpi), mikä johtaa kalvopotentiaalin elektronegatiivisuuden kasvuun noin 10 mV. Kuljetusproteiini suorittaa tämän toiminnon suurella nopeudella: 150 - 600 natriumionia sekunnissa. Kuljetusproteiinin aminohapposekvenssi tunnetaan, mutta tämän monimutkaisen metabolisen kuljetuksen mekanismi ei ole vielä selvä. Tämä prosessi kuvataan käyttämällä proteiinien natrium- tai kalium-ionien siirtymisen energiaprofiileja (kuvat 1.5,5). Näiden profiilien muutosten luonteen perusteella, jotka liittyvät jatkuviin muutoksiin kuljetusproteiinin konformaatiossa (prosessi, joka vaatii energiaa), voidaan arvioida vaihdon stoikiometriaa: kaksi kalium-ionia vaihtuu kolmeen natrium-ioniin.
Na/K-pumppu, sama kuin eristetty Na+ /K + -riippuvainen kalvon ATPaasi, jota spesifisesti estää sydämen glykosidi ouabaiini (strofantiini). Koska Na/K-pumpun toiminta on monivaiheinen kemiallinen reaktio, se, kuten kaikki kemialliset reaktiot, on suurelta osin riippuvainen lämpötilasta, joka

Riisi. 1.6.Kaavio Na/K-pumppu-ATPaasista (upotettu plasmakalvon lipidikaksoiskerrokseen), joka yhdessä syklissä poistaa kolme Na+-ionia solusta potentiaali- ja pitoisuusgradientteja vastaan ja tuo kaksi K-ionia soluun + . Tämän prosessin aikana yksi ATP-molekyyli hajoaa ADP:ksi ja fosfaatiksi. Kaavio esittää ATPaasin dimeerinä, joka koostuu suuresta (toiminnallisesta) ja pienestä alayksiköstä; kalvossa se esiintyy tetrameerinä, joka muodostuu kahdesta suuresta ja kahdesta pienestä alayksiköstä
esitetty kuvassa. 1.7. Tässä esitetään natriumionien virta lihassoluista suhteessa aikaan; Tämä vastaa käytännössä Na/K-pumpun toiminnan välittämää natriumionien virtausta, koska natriumionien passiivinen virtaus pitoisuutta ja potentiaaligradientteja vastaan on erittäin pieni. Jos lääkettä jäähdytetään noin 18°C, natrium-ionien virtaus solusta vähenee nopeasti 15-kertaiseksi ja palautuu heti lämmityksen jälkeen alkuperäiselle tasolleen. Tämä natriumionien virtauksen väheneminen kennosta on useita kertoja suurempi kuin se, joka vastaisi diffuusioprosessin tai yksinkertaisen kemiallisen reaktion lämpötilariippuvuutta. Samanlainen vaikutus havaitaan, kun aineenvaihduntaenergia on ehtynyt dinitrofenoli (DNP) myrkytyksen seurauksena (Kuva 1.7,5). Näin ollen natriumionien virtaus kennosta varmistetaan energiariippuvaisella reaktiolla - aktiivisella pumpulla. Toinen pumpun ominaisuus, sekä merkittävä lämpötila- ja energiariippuvuus, on kyllästymistaso (kuten kaikki muut kemialliset reaktiot); tämä tarkoittaa, että pumpun nopeus ei voi kasvaa loputtomasti kuljetettujen ionien pitoisuuden kasvaessa (kuva 1.8). Sitä vastoin passiivisesti diffundoituvan aineen virtaus kasvaa suhteessa pitoisuuseroon diffuusiolain mukaisesti (yhtälöt 1 ja 2).

Riisi. 1.7. A, B. Aktiivinen Na-kuljetus + . Y-akseli: radioaktiivisen 24 Na +:n virtaus solusta (imp./min). Abskissa-akseli: aika kokeen alkamisesta. A. Kenno jäähdytetään 18,3 °C:sta 0,5 °C:seen; virtaus Na+ solusta tänä aikana estyy. B. Na + -virtauksen tukahduttaminen solusta dinitrofenolilla (DNP) pitoisuudessa 0,2 mmol/l (muunnettu)
Na/K-pumpun lisäksi plasmakalvossa on ainakin yksi pumppu - kalsium; tämä pumppu pumppaa kalsiumioneja (Ca 2+) ulos solusta ja on mukana pitämässä niiden solunsisäistä pitoisuutta erittäin alhaisella tasolla (taulukko 1.1). Kalsiumpumppua on erittäin tiheänä lihassolujen sarkoplasmisessa retikulumissa, jotka keräävät kalsiumioneja ATP-molekyylien hajoamisen seurauksena (katso luku 4).
Na/K-pumpun vaikutus kalvopotentiaaliin ja solutilavuuteen . Kuvassa Kuva 1.9 esittää kalvovirran eri komponentit ja ionien solunsisäiset pitoisuudet

Riisi. 1.8.Molekyylien kuljetusnopeuden ja niiden pitoisuuden (kanavan sisäänkäynnissä tai pumpun sitoutumiskohdassa) välinen suhde diffuusion aikana kanavan läpi tai pumppauskuljetuksen aikana. Jälkimmäinen kyllästyy korkeissa pitoisuuksissa (maksiminopeus, Vmax ) arvo x-akselilla, joka vastaa puolta pumpun maksiminopeudesta ( Vmax /2), on tasapainopitoisuus TO m

Riisi. 1.9.Kaavio, joka näyttää Na+-pitoisuudet , K+ ja Cl - solun sisällä ja ulkopuolella sekä näiden ionien tunkeutumisreitit solukalvon läpi (spesifisten ionikanavien kautta tai käyttämällä Na/K-pumppua. Tietyillä pitoisuusgradienteilla tasapainopotentiaalit E Na, E K ja E С l – sama kuin osoitetut, kalvopotentiaali Em = – 90 mV
varmistaa niiden olemassaolon. Kaliumionien virtaa ulospäin havaitaan kaliumkanavien kautta, koska kalvopotentiaali on hieman sähköpositiivisempi kuin kalium-ionien tasapainopotentiaali. Natriumkanavien kokonaisjohtavuus on paljon pienempi kuin kaliumkanavien, ts. natriumkanavat ovat auki paljon harvemmin kuin kaliumkanavat lepopotentiaalissa; kuitenkin suunnilleen sama määrä natriumioneja tulee soluun kuin kaliumioneja poistuu solusta, koska natriumionien diffundoituminen soluun vaatii suuria pitoisuuksia ja potentiaaligradientteja. Na/K-pumppu kompensoi ihanteellisesti passiivisia diffuusiovirtoja, koska se kuljettaa natriumioneja ulos solusta ja kaliumioneja siihen. Siten pumppu on sähkögeeninen johtuen kennoon ja sieltä ulos siirrettyjen varausten lukumäärän erosta, joka normaalilla toimintanopeudellaan muodostaa noin 10 kalvopotentiaalin. mV on elektronegatiivisempi kuin jos se olisi tuotettu pelkästään passiivisilla ionivuoilla (katso yhtälö 7). Tämän seurauksena kalvopotentiaali lähestyy kaliumtasapainopotentiaalia, mikä vähentää kalium-ionien vuotoa. No toimintaa/K-pumppua säätelee natriumionien intrasellulaarinen pitoisuus. Pumpun nopeus hidastuu solusta poistettavien natriumionien pitoisuuden pienentyessä (kuva 1.8), jolloin pumpun toiminta ja natrium-ionien virtaus soluun tasapainottavat toisiaan ja ylläpitävät solunsisäistä natriumin pitoisuutta. ioneja noin 10 mmol/l.
Tasapainon ylläpitämiseksi pumppaus- ja passiivisten kalvovirtojen välillä tarvitaan paljon enemmän Na/K-pumppumolekyylejä kuin kalium- ja natriumionien kanavaproteiineja. Kun kanava on auki, sen läpi kulkee kymmeniä tuhansia ioneja muutamassa millisekunnissa (katso yllä), ja koska kanava avautuu yleensä useita kertoja sekunnissa, niin sen läpi kulkee tänä aikana yhteensä yli 105 ionia. Yksittäinen pumppuproteiini siirtää useita satoja natriumioneja sekunnissa, joten plasmakalvon tulee sisältää noin 1000 kertaa enemmän pumppumolekyylejä kuin kanavamolekyylejä. Kanavavirtojen mittaukset levossa osoittivat keskimäärin yhden kalium- ja yhden avoimen natriumkanavan läsnäolon 1 μm 2 -kalvoa kohti; Tästä seuraa, että Na/K-pumpun noin 1000 molekyyliä pitäisi olla samassa tilassa, ts. niiden välinen etäisyys on keskimäärin 34 nm; Pumppuproteiinin halkaisija kanavaproteiinina on 8–10 nm. Siten kalvo on melko tiheästi kyllästetty pumppaavilla molekyyleillä
Se, että natrium-ionien virtaus soluun ja kalium-ionit ulos kennosta kompensoituu pumpun toiminnalla, on toinen seuraus, joka koostuu vakaan osmoottisen paineen ylläpitämisestä ja jatkuva tilavuus. Solun sisällä on suuri pitoisuus suuria anioneja, pääasiassa proteiineja (A - taulukossa 1.1), jotka eivät pysty tunkeutumaan kalvon läpi (tai tunkeutumaan sen läpi hyvin hitaasti) ja ovat siksi kiinteä komponentti solun sisällä. Näiden anionien varauksen tasapainottamiseksi tarvitaan yhtä suuri määrä kationeja. Na/K-pumpun vaikutuksesta nämä kationit ovat pääasiassa kaliumioneja. Merkittävä kasvu solunsisäisessä ionipitoisuudessa voisi tapahtua vain, jos anionien konsentraatio kasvaa C1:n virtauksen johdosta - pitoisuusgradienttia pitkin soluun (taulukko 1.1), mutta kalvopotentiaali vastustaa tätä. Tuleva virta Cl - havaitaan vain, kunnes kloori-ionien tasapainopotentiaali saavutetaan; tämä havaitaan, kun kloori-ionigradientti on melkein vastakkainen kaliumionigradientin kanssa, koska kloori-ionit ovat negatiivisesti varautuneita (yhtälö 4). Siten saadaan aikaan alhainen solunsisäinen kloori-ionien pitoisuus, joka vastaa alhaista solunulkoista kalium-ionien pitoisuutta. Tuloksena on solun ionien kokonaismäärän rajoitus. Jos kalvopotentiaali laskee Na/K-pumpun tukkeutuessa, esimerkiksi anoksian aikana, kloori-ionien tasapainopotentiaali pienenee ja kloridi-ionien solunsisäinen pitoisuus kasvaa vastaavasti. Palauttamalla varaustasapainon kaliumionit tulevat myös soluun; ionien kokonaispitoisuus solussa kasvaa, mikä lisää osmoottista painetta; tämä pakottaa vettä soluun. Solu turpoaa. Tämä turvotus havaitaan in vivo energian puutteen olosuhteissa.
Pitoisuusgradientti Na + kalvokuljetuksen liikkeellepanevana voimana . Na/K-pumpun merkitys solulle ei rajoitu normaalien K+- ja Na+-gradienttien stabilointiin kalvolla. Kalvogradienttiin varastoitunut energia Na+ , käytetään usein helpottamaan muiden aineiden kalvokuljetusta. Esimerkiksi kuvassa Fig. 1.10 näyttää "simport" Na+ ja sokerimolekyylejä soluun. Kalvokuljetusproteiini kuljettaa sokerimolekyylin soluun jopa pitoisuusgradienttia vastaan, kun taas Na + liikkuu samalla pitoisuutta ja potentiaaligradienttia pitkin antaen energiaa sokereiden kuljetus. Tämä sokereiden kuljetus on täysin riippuvainen korkean gradientin olemassaolosta Na+ ; jos solunsisäinen pitoisuus Na+ lisääntyy merkittävästi, sokerien kuljetus pysähtyy. Erilaisille c Akharov, on olemassa erilaisia simpport-järjestelmiä. Aminohappojen kuljetus kuljetusta vastaavaan soluun c kuvassa näkyvä akharov. 1,10; sen tarjoaa myös gradientti Na+ , On olemassa vähintään viisi erilaista symport-järjestelmää, joista jokainen on erikoistunut yhteen sukulaisten aminohappojen ryhmään.
Simport-järjestelmien lisäksi on myös "antiportterit". Yksi niistä esimerkiksi siirtää yhdessä syklissä yhden kalsium-ionin solusta vastineeksi kolmesta sisään tulevasta natrium-ionista (kuva 1.10). Energiaa varten Ca 2+ kuljetus muodostuu kolmen natrium-ionin sisäänpääsyn seurauksena konsentraatio- ja potentiaaligradienttia pitkin. Tämä energia riittää (lepopotentiaalissa) ylläpitämään korkeaa kalsiumionigradienttia (alle 10–7 mol/L solun sisällä noin 2 mmol/l solun ulkopuolella).
Endo- ja eksosytoosi . Tietyille aineille, jotka tulevat soluun tai jotka on poistettava

Riisi. 1.10.Kalvon lipidikaksoiskerrokseen upotetut proteiinit välittävät glukoosin ja Na +:n sympportin soluun sekä Ca2+/Na+ –antiportti, jossa liikkeellepaneva voima on solukalvon Na + -gradientti
siitä ei ole kuljetuskanavia; Näitä aineita ovat esimerkiksi proteiinit ja kolesteroli. Ne voivat kulkeutua plasmakalvon läpi rakkuloita, tai kuplia, endo- ja eksosytoosin kautta. Kuvassa Kuva 1.11 esittää näiden prosessien päämekanismit. Eksosytoosissa tietyt organellit (katso alla) muodostavat rakkuloita, jotka ovat täynnä ainetta, joka on poistettava solusta, kuten hormoneja tai solunulkoisia entsyymejä. Kun tällaiset rakkulat saavuttavat plasmakalvon, niiden lipidikalvo fuusioituu sen kanssa, jolloin sisältö pääsee karkaamaan ulkoiseen ympäristöön. Päinvastaisessa prosessissa, endosytoosissa, plasmakalvo tunkeutuu muodostaen kuopan, joka sitten syvenee ja sulkeutuu muodostaen solunsisäisen rakkulan, joka on täynnä solunulkoista nestettä ja joitain makromolekyylejä. Tämän kalvofuusion ja vesikkelin sulkeutumisen varmistamiseksi sytoskeleton supistuvat elementit toimivat yhdessä itse kalvojen kanssa (katso alla). Endosytoosi ei aina tarkoita pelkästään solunulkoisen ympäristön sieppaamista soluun. Solukalvo sisältää usein spesifisiä reseptoreita makromolekyyleille, kuten insuliinille tai antigeeneille, usein järjestäytyneenä erikoisryhmiin. Kun nämä makromolekyylit sitoutuvat reseptoreihinsa, reseptoria ympäröivällä kalvoalueella tapahtuu endosytoosia ja makromolekyyli kuljetetaan selektiivisesti soluun (kuva 1.12, B).
Endo- ja eksosytoosia esiintyy soluissa jatkuvasti. Kalvomateriaalin vaihdon määrä on merkittävä; 1 tunnin sisällä makrofagi imee vesikkeleinä kaksi kertaa sytoplasmisen kalvonsa pinta-alan. Useimmissa soluissa kalvomateriaalin vaihtuvuus ei tapahdu yhtä intensiivisesti, mutta sen pitäisi silti olla merkittävää.

Riisi. 1.11.Eksosytoosi ja endosytoosi. Ylös: solunsisäinen rakkula fuusioituu plasmakalvon lipidikaksoiskerroksen kanssa ja avautuu solunulkoiseen tilaan. Tätä prosessia kutsutaan eksosytoosiksi. Pohjalla: plasmakalvo tunkeutuu pienelle alueelle ja vapauttaa rakkulan, joka on täynnä solunulkoista materiaalia. Tätä prosessia kutsutaan endosytoosiksi
1.3. Aineiden kuljetus solun sisällä
Endo- ja eksosytoosi eivät ole vain aineiden kuljetusprosesseja solukalvon läpi, vaan myös kalvojen vaihtoprosesseja - itse solun rakennekomponentteja. Tässä osiossa tarkastellaan muita samanlaisia kuljetusprosesseja solussa ja sen organelleissa.
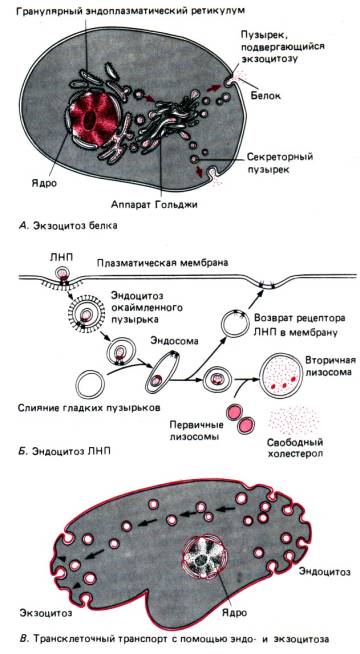
Riisi. 1.12. A–B. Prosessien kaavio, mukaan lukien ekso- ja endosytoosi. A. Rakeisessa endoplasmisessa retikulumissa syntetisoitu proteiini kuljetetaan Golgi-laitteen kautta plasmakalvolle, jossa se erittyy eksosytoosin kautta. B. LDL- (low-density lipoprotein) -partikkeleihin sitoutunut kolesteroli kiinnittyy plasmakalvoon, saa aikaan endosyyttirakkulan muodostumisen kalvon tälle alueelle ja kuljetetaan lysosomeihin, joissa se vapautuu. SISÄÄN. Endosytoosin aikana siepattu solunulkoinen materiaali (kuvassa oikealla), kuljetetaan solun läpi vesikkeleissä tai vesikkeleissä ja vapautuu eksosytoosin kautta (kuvassa vasemmalle)
Diffuusio . Luonnollisesti sytosolissa pitoisuuksien ero eliminoituu diffuusion vuoksi; sama pätee organelleissa oleviin nesteisiin. Liuenneen proteiinin suuren pitoisuuden vuoksi diffuusio on täällä paljon hitaampaa kuin vedessä. Lipidikalvot - solujen ympärillä ja organellien sisällä - ovat kaksiulotteisia nesteitä, joissa diffuusio tapahtuu. Membraanikaksoiskerroksen lipidit diffuusoituvat omassa kerroksessaan siirtyen harvoin toisesta toiseen. Niihin upotetut proteiinit ovat myös melko liikkuvia; ne pyörivät kalvoon nähden kohtisuoran akselin ympäri tai diffuusoituvat sivusuunnassa hyvin erilaisilla diffuusiovakioilla, 2–10 000 kertaa hitaammin kuin fosfolipidit. Joten jos jotkin proteiinit liikkuvat vapaasti lipidikerroksessa ja samalla nopeudella kuin itse lipidimolekyylit, niin toiset ovat ankkuroituja, ts. melko tiukasti kiinni sytoskeletonissa. Kalvossa on tiettyjen proteiinien "pysyviä" aggregaatteja, esimerkiksi hermosolujen pre- ja postsynaptisia rakenteita. Vapaasti liikkuvat proteiinit voidaan osoittaa sitomalla ne fluoresoiviin väriaineisiin, joiden luminesenssi saadaan aikaan valaisemalla lyhyesti kalvon pientä aluetta. Tällaiset kokeet osoittavat, että alle 1 minuutissa väriaineeseen sitoutuneet proteiinit jakautuvat tasaisesti kalvolle jopa 10 μm:n etäisyyksille.
Aktiivinen kuljetus organellikalvoissa .
Aktiiviset kuljetusprosessit, joilla on tärkeä rooli plasmakalvon toiminnassa, tapahtuvat myös solun sisällä, organellien kalvoissa. Erilaisten organellien spesifiset sisällöt syntyvät osittain sisäisen synteesin ja osittain aktiivisen kuljetuksen ansiosta sytosolista. Yksi esimerkki jälkimmäisestä on edellä mainittu Ca 2+ -pumppu lihassolujen sarkoplasmisessa retikulumissa. Erityisen mielenkiintoista on, että mitokondrioiden ATP-synteesin tapauksessa päinvastainen periaate pätee siihen, mitä tapahtuu plasmakalvon ATPaasipumpuissa (kuva 1.6). ATP-synteesin aikana oksidatiivinen aineenvaihdunta johtaa jyrkän gradientin muodostumiseen H+ sisäkalvoilla. Tämä gradientti on liikkeellepaneva voima molekyylien aktiivisen kuljetuksen pumppaussyklin käänteisessä prosessissa: H + -ionit liikkuvat kalvon läpi gradienttia pitkin, ja sen seurauksena vapautuva energia tuottaa ATP:n synteesin ADP:stä ja fosfaatista. Tuloksena oleva ATP puolestaan tarjoaa energiaa solulle, myös aktiiviseen kuljetukseen.
Kuljetus rakkuloissa . Solussa on suuri määrä organelleja ja niihin liittyviä rakkuloita (kuva 1.1). Nämä organellit, ja erityisesti rakkulat, ovat jatkuvassa liikkeessä kuljettaen sisältönsä muihin organelleihin tai plasmakalvoon. Vesikkelit voivat myös siirtyä solukalvolta organelleihin, kuten endosytoosissa.
Käsitellä asiaa proteiinin eritystä esitetty kuvassa. 1.12, A. Proteiini syntetisoidaan lähellä solun ydintä ribosomeissa, jotka liittyvät endoplasmiseen retikulumiin (niin sanottu rakeinen tai karkea endoplasminen verkkokalvo); Endoplasmisessa retikulumissa proteiini pakataan kuljetusrakkuloihin, jotka irtoavat organellesta ja kulkeutuvat Golgin laitteeseen. Täällä ne sulautuvat Golgi-laitteen vesisäiliöihin, joissa proteiini modifioidaan (eli muunnetaan glykoproteiiniksi). Vesikkelien päissä rakkulat erottuvat jälleen. Modifioitua proteiinia kuljettavat eritysrakkulat siirtyvät plasmakalvolle ja vapauttavat sisältönsä eksosytoosin kautta.
Toinen esimerkki solun kuljetusreitistä on esitetty kuvassa. 1,12, B; Tämä on kolesterolin imeytymistä soluihin. Veressä kulkeutuva kolesteroli sitoutuu pääasiassa proteiineihin, kuten hiukkasiin "matalatiheyksinen lipoproteiini"(LDL). Tällaiset hiukkaset kiinnittyvät spesifisille LDL-reseptoreita sisältäville kalvoalueille, joissa tapahtuu endosytoosia ja LDL kuljetetaan soluun "rajoilla olevissa" vesikkeleissä. Nämä vesikkelit sulautuvat muodostaen endosomeja ja menettävät "reunansa" tämän prosessin aikana. Endosomit puolestaan sulautuvat ensisijaisesti hydrolyyttisiä entsyymejä sisältäviin primaarisiin lysosomeihin ja muodostavat sekundaarisia, suurempia lysosomeja. Niissä kolesteroli vapautuu LDL-partikkeleista ja diffundoituu sytosoliin, josta se tulee saataville esimerkiksi lipidikalvojen synteesiä varten. Myös rakkulat, jotka eivät sisällä LDL:ää, erotetaan endosomeista, jotka siirtyvät erityisellä tavalla plasmakalvoon ja sulautuvat siihen palauttaen kalvomateriaalia ja todennäköisesti LDL-reseptoreita. Siitä hetkestä, kun LDL-partikkeli sitoutuu kalvoon, kunnes kolesteroli vapautuu sekundaarisesta lysosomista, kuluu 10-15 minuuttia. LDL:n sitoutumisen ja imeytymisen, eli solun kolesterolin saannin heikkeneminen on ratkaisevassa roolissa vakavan ja laajalle levinneen sairauden, ateroskleroosin (valtimoiden "kovettuminen") kehittymisessä.
On monia muita samanlaisia kuljetusreittejä kuin kuvassa. 1.11 ja 1.12, A, joiden avulla tietyt vesikkelit liikkuvat solussa. Ei tiedetä tarkasti, kuinka ne liikkuvat, mutta luurankosolut ovat todennäköisesti mukana tässä prosessissa. Vesikkelit voivat liukua pitkin mikrotubuluksia, jolloin liikeenergian saa ilmeisesti rakkuloihin liittyvästä proteiinista, ATPaasista (katso alla). On edelleen täysin epäselvää, kuinka monta erilaista rakkulaa, joka liikkuu peräkkäin kaikkiin suuntiin, saavuttaa määränpäänsä. Ne on luonnollisesti "merkittävä" tavalla, jonka liikennejärjestelmä tunnistaa ja muuntaa kohdistetuksi liikkeeksi.
Kuljetus organellien muodostumisen ja tuhoamisen kautta . Tähän mennessä olemme pitäneet endo- ja eksosytoosia vesikkeleiden sisällön kuljetusprosesseina. Näissä prosesseissa on toinen näkökohta, joka perustuu siihen, että plasmakalvon kohdennettu poistaminen solun pinnan yhdestä osasta endosytoosin avulla ja päinvastoin sen lisääminen toiseen osaan eksosytoosilla siirtää suuria alueita kalvosta (kuva 1). 1.12.E), joka antaa solulle mahdollisuuden muodostaa esimerkiksi kasvun tai liikkua.
Samanlaiset uudelleenjärjestelyt ovat tyypillisiä myös sytoskeletille, erityisesti mikrofilamenteille ja mikrotubuleille (kuva 1.1). Mikrofilamentit koostuvat pääasiassa F-aktiiniproteiini, joka pystyy kerääntymään kuitukimppuiksi sytosolista peräisin olevan monomeerin polymeroitumisen seurauksena. Kimput ovat polarisoituneita, eli ne kasvavat usein vain toisesta päästään kerääen uusia aktiinimolekyylejä, kun taas toinen pää on inertti tai purkaminen tapahtuu täällä. Tämän polarisoidun kasvun ansiosta mikrofilamentit liikkuvat tehokkaasti ja niiden verkoston rakenne voi muuttua. Aktiinin siirtyminen depolymeroidusta tilasta (sol) järjestäytyneeseen tilaan (geeli) voi tapahtua hyvin nopeasti muiden proteiinien tai ionipitoisuuden muutosten vaikutuksesta (katso alla). On myös proteiineja, jotka aiheuttavat aktiinifilamenttien hajoamisen muodostaen lyhyitä fragmentteja. Monien solujen ohuet projektiot - filopodia - sisältävät keskeisen aktiinikimpun (kuva 1.1), ja filopodioiden erilaiset liikkeet johtuvat todennäköisesti aktiinin siirtymistä: polymerisaatio - depolymeroituminen.
Mikrotubuluksetmyös usein samankaltaisia liikkeitä. Näiden liikkeiden mekanismi on samanlainen - tubuliinin polymeroituminen sytosolista siten, että yksi mikrotubuluksen päistä kasvaa, kun taas toinen joko ei muutu tai purkaminen tapahtuu siellä. Siten mikrotubulus voi liikkua koko sytosolissa lisäämällä tai poistamalla materiaalia.
Sytoskeleton aktiiviset liikkeet . Muutoksia sytoskeletaalisissa rakenteissa voi tapahtua sekä aktiivisten liikkeiden että edellä kuvattujen uudelleenjärjestelyjen seurauksena. Monissa tapauksissa mikrotubulusten ja aktiinifilamenttien liikettä ohjaavat supistuvat proteiinit, jotka sitovat filamentteja tai tubuluksia ja voivat siirtää niitä suhteessa toisiinsa. Oravat myosiini ja dyneiini läsnä kaikkien solujen sytosolissa suhteellisen korkeina pitoisuuksina; ne ovat elementtejä, jotka muuttavat energian liikkeeksi erikoistuneissa soluissa (lihas) ja organelleissa (värikalvot). Lihassoluissa myosiini muodostaa paksuja filamentteja, jotka ovat suuntautuneet samansuuntaisesti aktiinifilamenttien kanssa. Myosiinimolekyyli "päällään" kiinnittyy aktiinifilamenttiin ja ATP:n energiaa käyttämällä syrjäyttää myosiinin aktiinimolekyyliä pitkin. Myosiini irtoaa sitten aktiinista. Monien tällaisten kytkentä-katkaisujaksojen yhdistelmä johtaa makroskooppiseen lihassäikeiden supistuminen(luku 4). Dyneiinillä on samanlainen rooli mikrotubulusten liikkeessä värien aikana (kuva 1.1). Erikoistumattomien solujen sytoplasmassa myosiini ja dyneiini eivät muodosta säännöllisiä kuituja, vaan useimmissa tapauksissa pieniä molekyyliryhmiä. Jopa niin pienissä aggregaateissa ne pystyvät liikuttamaan aktiinifilamentteja tai mikrotubuluksia. Riisi. Kuva 1.13 havainnollistaa tätä prosessia, kun vastakkaisesti polarisoituneita myosiinimolekyylejä kiinnitetään myös kahteen eri suuntiin polarisoituneeseen aktiinifilamenttiin. Myosiinipääryhmät taipuvat kohti molekyylin häntää kuluttaen prosessissa ATP:tä, ja kaksi aktiinifilamenttia liikkuvat vastakkaiseen suuntaan, minkä jälkeen myosiini irtoaa niistä. Tällaiset liikkeet, joiden aikana ATP-energia muuttuu mekaaniseksi työksi, voivat muuttaa sytoskeleton ja siten myös solun muotoa sekä tarjota solutukirakenteeseen liittyvien organellien kuljetuksen.
Solunsisäisen kuljetuksen prosessit voidaan selkeimmin osoittaa hermosolun aksonilla. Axonin kuljetus käsitellään tässä yksityiskohtaisesti havainnollistamaan tapahtumia, jotka todennäköisesti tapahtuvat samalla tavalla useimmissa soluissa. Vain muutaman mikronin halkaisijaltaan oleva aksoni voi saavuttaa yhden metrin tai enemmän, ja proteiinien liikkuminen diffuusiona ytimestä aksonin distaaliseen päähän kestäisi vuosia. Jo pitkään on tiedetty, että kun jokin aksonin osa joutuu supistumisen läpi, proksimaalisesti sijaitseva aksonin osa laajenee. Näyttää siltä, että keskipakovirtaus on tukossa aksonissa. Sellainen virtaus-nopea aksonikuljetus voi voidaan osoittaa radioaktiivisten merkkien liikkeellä, kuten kuvassa 1 esitetyssä kokeessa. 1.14. Radioaktiivisesti merkittyä leusiinia injektoitiin selkäjuuren ganglioniin, ja sitten radioaktiivisuus mitattiin iskiashermossa 166 mm:n etäisyydeltä hermosolujen kappaleista 2. - 10. tuntiin. Yli 10 tunnin aikana radioaktiivisuuden huippu injektiokohdassa muuttui hieman. Mutta radioaktiivisuusaalto levisi aksonia pitkin vakionopeudella, noin 34 mm 2 tunnissa eli 410 mm/vrk. On osoitettu, että kaikissa homeotermisten eläinten hermosoluissa nopea aksonikuljetus tapahtuu samalla nopeudella, eikä huomattavia eroja ole havaittavissa ohuiden, myelinisoitumattomien kuitujen ja paksuimpien aksonien välillä sekä motoristen ja sensoristen kuitujen välillä. Radioaktiivisen markkerin tyyppi ei myöskään vaikuta nopean aksonikuljetuksen nopeuteen; markkerit voivat toimia erilaisina radioaktiivisina

Riisi. 1.13.Ei-lihaksinen myosiinikompleksi voi tietyllä suuntauksella sitoutua eri polaarisiin aktiinifilamentteihin ja ATP:n energiaa käyttämällä syrjäyttää niitä suhteessa toisiinsa
molekyylejä, kuten erilaisia aminohappoja, jotka sisältyvät neuronin solurungon proteiineihin. Jos analysoimme hermon perifeeristä osaa määrittääksemme tänne kuljetetun radioaktiivisuuden kantajien luonteen, niin tällaisia kantajia löytyy pääasiassa proteiinifraktiosta, mutta myös välittäjien ja vapaiden aminohappojen koostumuksesta. Tietäen, että näiden aineiden ominaisuudet ovat erilaiset ja niiden molekyylien koot ovat erityisen erilaisia, voimme selittää kuljetuksen vakionopeuden vain niille kaikille yhteisellä kuljetusmekanismilla.
Kuvailtu yläpuolella nopea aksonikuljetus On anterogradinen, eli suunnattu poispäin solurungosta. On osoitettu, että jotkut aineet siirtyvät periferialta solurunkoon käyttämällä retrogradinen kuljetus. Esimerkiksi asetyylikoliiniesteraasi kulkeutuu tähän suuntaan nopeudella, joka on 2 kertaa hitaampi kuin nopean aksonikuljetuksen nopeus. Neuroanatomiassa usein käytetty markkeri, piparjuuriperoksidaasi, liikkuu myös retrogradisella kuljetuksella. Retrogradisella kuljetuksella on luultavasti tärkeä rooli proteiinisynteesin säätelyssä solurungossa. Muutama päivä aksonin leikkaamisen jälkeen solurungossa havaitaan kromatolyysiä, mikä viittaa proteiinisynteesin häiriintymiseen. Kromatolyysin vaatima aika korreloi retrogradisen kuljetuksen keston kanssa aksonin transektiokohdasta solurunkoon. Tämä tulos ehdottaa myös selitystä tälle häiriölle - proteiinisynteesiä säätelevän "signaaliaineen" lähetys periferialta katkeaa. On selvää, että tärkeimmät "ajoneuvot" käytetään nopeaan aksonaaliseen
Riisi. 1.14.Koe, joka osoittaa nopean aksonikuljetuksen kissan iskiashermon sensorisissa kuiduissa. Tritiumilla leimattua leusiinia injektoidaan selkäjuuren ganglioniin, ja ganglion ja sensoristen säikeiden radioaktiivisuus mitataan 2, 4, 6, 8 ja 10 tunnin kuluttua injektiosta. (kuvan alareunassa). Tekijä: x-akseli Etäisyys gangliosta iskiashermon alueisiin, joissa mittaus tehdään, piirretään. Ordinaatta-akselilla vain ylemmän ja alemman käyrän radioaktiivisuus (pulssia/min) piirretään logaritmisella asteikolla. Lisääntyneen radioaktiivisuuden "aalto". (nuolet) liikkuu nopeudella 410 mm/vrk (pitki)
kuljetus ovat vesikkelit (vesikkelit) ja organellit, kuten mitokondriot, jotka sisältävät kuljetettavia aineita. Suurimpien rakkuloiden tai mitokondrioiden liikettä voidaan tarkkailla mikroskoopilla in vivo . Tällaiset hiukkaset tekevät lyhyitä, nopeita liikkeitä yhteen suuntaan, pysähtyvät, liikkuvat usein hieman taaksepäin tai sivulle, pysähtyvät uudelleen ja nykivät sitten pääsuuntaan. 410 mm/vrk vastaa noin 5 μm/s anterogradisen liikkeen keskinopeutta; Jokaisen yksittäisen liikkeen nopeuden on siis oltava paljon suurempi, ja jos otamme huomioon organellien, filamenttien ja mikrotubulusten koot, niin nämä liikkeet ovat todellakin erittäin nopeita. Nopea aksonikuljetus vaatii merkittävän ATP-pitoisuuden. Myrkyt, kuten kolkisiini, joka häiritsee mikrotubuluksia, estävät myös nopean aksonikuljetuksen. Tästä seuraa, että tarkastelemassamme kuljetusprosessissa vesikkelit ja organellit liikkuvat mikrotubuluksia ja aktiinifilamentteja pitkin; tämän liikkeen aikaansaavat pienet dyneiini- ja myosiinimolekyylien aggregaatit, jotka toimivat kuvan 1 mukaisesti. 1,13, käyttämällä ATP-energiaa.
Myös nopea aksonikuljetus voi olla mukana patologiset prosessit. Jotkut neurotrooppiset virukset (esimerkiksi herpes- tai poliovirukset) tulevat aksoniin reuna-alueella ja siirtyvät retrogradisen kuljetuksen kautta neuronikehoon, jossa ne lisääntyvät ja aiheuttavat myrkyllisiä vaikutuksia. Jäykkäkouristustoksiini, bakteerien tuottama proteiini, joka tulee kehoon, kun iho on vaurioitunut, vangitaan hermopäätteisiin ja kuljetetaan hermosoluihin, missä se aiheuttaa tyypillisiä lihaskouristuksia. Tiedossa on tapauksia, joissa on myrkyllisiä vaikutuksia itse aksonin kuljetukseen, esimerkiksi altistuminen teolliselle liuottimelle akryyliamidille. Lisäksi uskotaan, että vitamiinipuutoksen beriberin ja alkoholisen polyneuropatian patogeneesiin liittyy nopean aksonikuljetuksen rikkominen.
Nopean aksonikuljetuksen lisäksi solussa on myös melko intensiivistä hidas aksonikuljetus. Tubuliini liikkuu aksonia pitkin nopeudella noin 1 mm/vrk, ja aktiini liikkuu nopeammin – jopa 5 mm/vrk. Muut proteiinit kulkeutuvat näiden sytoskeletaalisten komponenttien mukana; esimerkiksi entsyymit näyttävät liittyvän aktiiniin tai tubuliiniin. Tubuliinin ja aktiinin liikenopeudet ovat suurin piirtein yhdenmukaisia aiemmin kuvatulle mekanismille löydettyjen kasvunopeuksien kanssa, joissa molekyylit liitetään mikrotubuluksen tai mikrofilamentin aktiiviseen päähän. Siksi tämä mekanismi voi olla hitaan aksonikuljetuksen taustalla. Hitaan aksonin kuljetuksen nopeus vastaa myös suunnilleen aksonin kasvunopeutta, mikä ilmeisesti osoittaa rajoituksia, jotka solutukirangan rakenne asettaa toiselle prosessille.
Tämän osion lopuksi on syytä korostaa, että solut eivät suinkaan ole staattisia rakenteita, kuten ne näkyvät esimerkiksi elektronimikroskooppisissa valokuvissa. Plasmakalvo ja erityisesti organellit ovat jatkuvassa nopeassa liikkeessä ja jatkuvassa uudelleenjärjestelyssä; siksi ne pystyvät toimimaan. Lisäksi nämä eivät ole yksinkertaisia kammioita, joissa tapahtuu kemiallisia reaktioita, vaan erittäin organisoituneet kalvojen ja kuitujen konglomeraatit, jossa reaktiot tapahtuvat optimaalisesti järjestetyssä järjestyksessä.
1.4. Solujen toimintojen säätely
Yksittäisen solun ylläpitoa toiminnallisena yksikkönä säätelee ensisijaisesti ydin; tällaisten säätelymekanismien tutkimus on solubiologian ja biokemian aihe. Samalla solujen on muunnettava toimintaansa ympäristöolosuhteiden ja kehon muiden solujen tarpeiden mukaisesti, eli ne toimivat toiminnallisen säätelyn kohteina. Alla tarkastellaan lyhyesti, kuinka nämä säätelyvaikutukset vaikuttavat plasmakalvoon ja kuinka ne saavuttavat solunsisäiset organellet.
Sääntelyvaikutukset solukalvoon
Kalvopotentiaali . Monissa tapauksissa solutoimintojen säätely tapahtuu muuttamalla kalvopotentiaalia. Paikalliset potentiaalin muutokset ovat mahdollisia, kun: 1) kalvon läpi virtaa solun viereiseltä alueelta tai toisen kennon tuottama virta; 2) ionien ekstrasellulaarinen pitoisuus muuttuu (usein [K + ] ulos ); 3) kalvon ionikanavat avautuvat. Muutokset kalvopotentiaalissa voivat vaikuttaa membraaniproteiinien konformaatioon aiheuttaen erityisesti kanavien avautumista tai sulkeutumista. Kuten edellä on kuvattu, joidenkin kalvopumppujen toiminta riippuu kalvopotentiaalista. Hermosolut ovat erikoistuneet havaitsemaan muutoksia kalvopotentiaalissa informaationa, joka on käsiteltävä ja välitettävä (katso luku 2).
Solunulkoiset säätelyaineet . Tärkein solunulkoisten aineiden säätelymekanismi on niiden vuorovaikutus spesifisten reseptorien kanssa plasmakalvolla tai solun sisällä. Näitä aineita ovat synaptiset välittäjät, jotka välittävät tietoa hermosolujen välillä, paikalliset aineet sekä veressä kiertävät ja kaikki kehon solut saavuttavat aineet, kuten hormonit ja antigeenit. Synaptiset lähettimet ovat pieniä molekyylejä, jotka vapautuvat hermopäätteistä synapsissa;
kun ne saavuttavat viereisen postsynaptisen solun plasmakalvon, ne laukaisevat sähköisiä signaaleja tai muita säätelymekanismeja. Tätä asiaa käsitellään yksityiskohtaisesti luvussa. 3.
Paikalliset kemialliset aineet usein erikoistuneet solut erittävät. Ne diffundoituvat vapaasti solunulkoiseen tilaan, mutta niiden toiminta rajoittuu pieneen soluryhmään, koska nämä aineet tuhoutuvat nopeasti joko spontaanisti tai entsyymien vaikutuksesta. Yksi esimerkki tällaisten aineiden vapautumisesta on vapautuminen histamiini syöttösolut vaurion tai immuunivasteen vuoksi. Histamiini rentouttaa verisuonten sileät lihassolut, lisää verisuonten endoteelin läpäisevyyttä ja stimuloi aistihermopäätteitä, jotka välittävät kutinaa. Monet muut solut vapauttavat muita paikallisia kemiallisia aineita. Tyypillisiä paikallisia agentteja ovat prostaglandiinit, muodostaen noin 20 rasvahappojohdannaisen ryhmän. Niitä vapautuu jatkuvasti laajalle levinneistä soluista, mutta ne vaikuttavat vain paikallisesti, koska kalvon fosfolipaasit tuhoavat ne nopeasti. Erilaisilla prostaglandiineilla on laaja vaikutuskirjo: ne voivat laukaista sileiden lihassolujen supistumisen, aiheuttaa verihiutaleiden (verihiutaleiden) aggregaatiota tai tukahduttaa munasarjojen keltaisen kehon kehittymisen.
Muut paikalliset edustajat palvelevat kasvutekijöitä. Tunnetuin on sympaattisten hermosolujen hermokasvutekijä (NGF), joka on välttämätön näiden hermosolujen kasvulle ja selviytymiselle kehityksen aikana. in vivo tai soluviljelmässä. Ilmeisesti tämän neuroniluokan kohdesolut erittävät NGF:ää ja varmistavat siten asianmukaisen hermotuksen. Kun solut muodostavat elimiä, niiden on usein ”löytettävä tiensä” kohdesoluihin, jotka voivat sijaita huomattavien etäisyyksien päässä. Näin ollen täytyy olla monia erikoistuneita kasvutekijöitä, jotka ovat samanlaisia kuin NGF.
Hormonit ja antigeenit veren mukana kaikkiin soluihin. Antigeenit laukaisevat immuunivasteen soluista, joissa on spesifisiä vasta-aineita. Antigeenit ovat kuitenkin pääsääntöisesti vieraita aineita, joita ei tuoteta reagoivassa organismissa (katso lisätietoja luvusta 18). Jotkut hormonit, kuten insuliini tai tyroksiini, vaikuttavat monenlaisiin solutyyppeihin, kun taas toiset, kuten sukupuolihormonit, vaikuttavat vain tiettyyn solutyyppiin. Hormonit ovat joko peptidejä, joiden toiminnan laukaisee sitoutuminen solukalvon reseptoriin, tai steroideja ja tyroksiinia, jotka diffundoituvat lipidikalvon läpi ja sitoutuvat solunsisäisiin reseptoreihin. Steroidihormonit sitoutuvat tuman kromatiiniin, mikä johtaa tiettyjen geenien transkriptioon. Syntyvät proteiinit aiheuttavat muutoksia solujen toiminnassa, mikä on hormonien spesifinen vaikutus. Hormonien vapautumiseen ja toimintaan liittyviä kysymyksiä käsitellään yksityiskohtaisesti luvussa. 17.
Solunsisäinen viestintä, johon osallistuu toisia lähettiläitä
Edellä kuvatut säätelytoiminnot sisältävät vaikutukset solukalvoon. Solukalvon vastaanottaman tiedon on usein saatava organellit reagoimaan, ja ne siirtyvät niihin erilaisten aineiden kautta, jotka tunnetaan nimellä toissijaiset sanansaattajat (toisin kuin ensimmäiset sanansaattajat, jotka tulevat soluun ulkoisista lähteistä). Toisten välittäjien tutkimus kehittyy nopeasti, eikä ole takeita siitä, että ongelman nykyinen ymmärryksen taso on riittävän täydellinen. Tässä käsittelemme kolmea hyvin tutkittua välittäjää: Ca 2+ , cAMP ja inositolitrifosfaatti.
Kalsium.Yksinkertaisin solunsisäinen välittäjäaine on Ca 2+ -ioni. Sen vapaa pitoisuus lepotilassa on hyvin alhainen ja on 10_–8–10–7 mol/l. Se voi päästä soluun tiettyjen kalvokanavien kautta, kun ne ovat avoimessa tilassa, esimerkiksi kalvopotentiaalin muuttuessa (katso luku 2). Tästä johtuva Ca 2+ -pitoisuuden nousu laukaisee solussa tärkeitä reaktioita, kuten lihasten supistumisen perustana olevien myofibrillien supistumisen (katso luku 4) tai välittäjäaineita sisältävien rakkuloiden vapautumisen hermopäätteistä (katso luku 3). . Molemmat reaktiot vaativat Ca 2+ -pitoisuuden noin 10-5 mol/l. Ca 2+, jolla on säätelevä vaikutus, voi vapautua myös solunsisäisistä varastoista, kuten endoplasmisesta retikulumista. Ca 2+:n vapautuminen varastosta edellyttää muiden välittäjien osallistumista (ks. esim. kuva 1.16).
Syklinen adenosiinimonofosfaatti, cAMP. Äskettäin on todistettu, että syklinen adenosiinimonofosfaatti (cAMP), elimistön pääenergianlähteen, ATP:n, johdannainen on tärkeä toinen sanansaattaja. Kuvassa esitetty monimutkainen reaktioketju. 1.15, alkaa reseptorilla R s plasmakalvon ulkopinnalla, joka voi toimia spesifisenä sitoutumispaikkana eri välittäjille ja hormoneille. Sen jälkeen, kun se on sitoutunut tiettyyn "stimuloivaan" molekyyliin R s muuttaa muotoaan; nämä muutokset vaikuttavat proteiiniin G s kalvon sisäpinnalla siten, että jälkimmäisen aktivoituminen solunsisäisellä guanosiinitrifosfaatilla (GTP) tulee mahdolliseksi. Aktivoitu proteiini G s puolestaan stimuloi kalvon sisäpinnalla olevaa entsyymiä, adenylaattisyklaasia (AC), joka katalysoi cAMP:n muodostumista ATP:stä. Vesiliukoinen cAMP on välittäjä, joka välittää vaikutuksen

Riisi. 1.15.Reaktioketju, jossa on mukana solunsisäinen lähetti-cAMP (syklinen adenosiinimonofosfaatti). Eksitatiiviset tai estävät ulkoiset signaalit aktivoivat kalvoreseptoreita R s tai Ri . Nämä reseptorit säätelevät sitoutumisprosessia G – proteiinit, joissa on solunsisäistä GTP:tä (guanosiinitrifosfaattia), mikä stimuloi tai estää solunsisäistä adenylaattisyklaasia (AC). Vahvistava entsyymi AC muuttaa adenosiinitrifosfaatin (ATP) cAMP:ksi, joka sitten hajoaa AMP:ksi fosfodiesteraasin vaikutuksesta. Vapaa cAMP diffundoituu soluun ja aktivoi adenylaattikinaasin (A-kinaasi) vapauttaen sen katalyyttisen alayksikön C, joka katalysoi solunsisäisten proteiinien fosforylaatiota, ts. muodostaa solunulkoisen ärsykkeen lopullisen vaikutuksen. Kaavio näyttää myös farmakologiset lääkkeet ja toksiinit, jotka laukaisevat (+) tai estävät (-) tiettyjä reaktioita (muokattu)
solunulkoisen reseptorin stimulaatio R s solun sisäisiin rakenteisiin.
Rinnakkain stimuloivan reaktioketjun kanssa R s inhiboivien välittäjien ja hormonien sitoutuminen vastaavaan reseptoriin on mahdollista R i joka taas GTP-aktivoidun proteiinin kautta G , estää AC:n ja siten cAMP:n tuotannon. Diffundoituessaan soluun cAMP reagoi adenylaattikinaasin (A-kinaasi) kanssa; tämä vapauttaa alayksikön C, joka katalysoi P-proteiinin fosforylaatiota. Tämä fosforylaatio muuttaa proteiinit aktiiviseen muotoon, ja ne voivat nyt käyttää spesifisiä säätelyvaikutuksiaan (esimerkiksi aiheuttaa glykogeenin hajoamista). Tämä monimutkainen säätelyjärjestelmä on erittäin tehokas, koska lopputuloksena on monien proteiinien fosforylaatio, eli säätelysignaali kulkee piirin läpi suurella vahvistuksella. Ulkoiset välittäjät, jotka sitoutuvat reseptoreihin R s ja R i jokaiselle niille ominaisia, erittäin erilaisia. Adrenaliini, yhteys R s tai R i osallistuu lipidi- ja glykogeeniaineenvaihdunnan säätelyyn sekä sydänlihaksen supistumisen tehostamiseen ja muihin reaktioihin (ks. luku 19). Kilpirauhasta stimuloiva hormoni, aktivointi R s , stimuloi tyroksiinihormonin vapautumista kilpirauhasessa ja prostaglandiini I estää verihiutaleiden aggregaatiota. Estävät vaikutukset, mukaan lukien adrenaliini, välittyvät R i ilmaistaan lipolyysin hidastumisena. Täten, cAMP-järjestelmä on monitoiminen solunsisäinen säätelyjärjestelmä, joita voidaan tarkasti kontrolloida solunulkoisilla stimuloivilla ja inhiboivilla signalointiaineilla.
Inositolifosfaatti "IF" h " Toisen lähettilään, inositolifosfaatin, solunsisäinen järjestelmä löydettiin vasta hiljattain (kuva 1.16). Tässä tapauksessa estoreittiä ei ole, mutta on samankaltaisuutta cAMP-järjestelmän kanssa, jossa R-reseptorin stimulaation vaikutus siirtyy GTP-aktivoituun G-proteiiniin kalvon sisäpinnalla. Seuraavassa vaiheessa tavallinen kalvolipidifosfatidyyli-inositoli (PI), joka on aiemmin saanut kaksi lisäfosfaattiryhmää, muunnetaan PI-difosfaatiksi (PIP 2), joka pilkkoutuu aktivoidun fosfodiesteraasin (PDE) vaikutuksesta. inositolitrifosfaatti(IFz) ja lipidi diasyyliglyseroli(DAG). Inositolitrifosfaatti on vesiliukoinen toinen lähetti, joka diffundoituu sytosoliin. Se toimii ensisijaisesti vapauttamalla Ca 2+:aa endoplasmisesta retikulumista. Ca2+ puolestaan toimii lähettinä, kuten edellä on kuvattu; se esimerkiksi aktivoi Ca 2+ -riippuvaista fosfokinaasia, joka fosforyloi entsyymejä. Myös DAG:n lipidialayksikkö (kuva 1.16) kuljettaa signaalin, diffuusoituen plasmakalvon lipidifaasissa sen sisäpinnalla sijaitsevaan C-kinaasiin, joka aktivoituu fosfatidyyliseriinin osallistuessa kofaktorina. Sitten C-kinaasi laukaisee proteiinien fosforylaation ja muuntaa ne aktiiviseen muotoon.
Solunsisäistä toista lähettijärjestelmää IF3 voidaan myös kontrolloida useilla ulkoisilla välittäjillä ja hormoneilla, mukaan lukien asetyylikoliini, serotoniini, vasopressiini ja kilpirauhasta stimuloiva hormoni; cAMP-järjestelmän tavoin sille on tunnusomaista useat solunsisäiset vaikutukset. On mahdollista, että tämä järjestelmä aktivoituu myös valon vaikutuksesta silmän näköreseptorissa ja sillä on keskeinen rooli valotransduktiossa (katso luku 11). Ensimmäistä kertaa organismin yksilöllisessä kehityksessä siittiö aktivoi IG-järjestelmän reseptorin, minkä seurauksena IG osallistuu munasolun hedelmöittymiseen liittyviin säätelyreaktioihin.
cAMP- ja IFz-DAG-järjestelmät ovat erittäin tehokkaita biologisia tehostajia. Ne

Riisi. 1.16.Reaktioketju, johon liittyy solunsisäinen välittäjä IPG (inositolitrifosfaatti). Kuten cAMP-järjestelmässä, solunulkoinen signaali välittyy proteiinin kauttaG, joka tässä tapauksessa aktivoi fosfodiesteraasin (PDE). Tämä entsyymi hajottaa fosfatidyyli-linosiinidifosfaattia (PIP 2 ) plasmamembraanissa ennen IF:ää h ja diasyyliglyseroli (DAG); JOS h diffundoituu sytoplasmaan. Tässä se aiheuttaa Ca:n vapautumisen 2+ endoplasmisesta retikulumista; Ca-pitoisuuden nousu 2+ sytoplasmassa ([Ca 2+ ]i ) aktivoi proteiinikinaasia, joka fosforyloi ja aktivoi siten entsyymejä. Toinen tuote, DAG, jää kalvoon ja aktivoi proteiinikinaasi C:n (fosfatidyyliseriinikofaktori, PS). Proteiinikinaasi C fosforyloi myös entsyymejä, jotka välittävät spesifisiä vaikutuksia, jotka liittyvät ulkoiseen reseptoristimulaatioon R . IF:n reaktioketjun haarat h ja DAG voidaan aktivoida itsenäisesti ionomysiinillä ja vastaavasti forboliesterillä (muokattu)
muuntaa lähettimen ja ulkokalvoreseptorin välisen reaktion useiden solunsisäisten proteiinien fosforylaatioksi, mikä voi sitten vaikuttaa solun eri toimintoihin. Yksi ongelman merkittävistä puolista on se, että nykyisen tiedon mukaan on olemassa vain nämä kaksi tämän tyyppistä läheisesti liittyvää säätelyjärjestelmää, joita useat ulkoiset välittäjät käyttävät säätelemään erilaisia solunsisäisiä prosesseja. Samaan aikaan nämä säätelyjärjestelmät, mukaan lukien Ca 2+, ovat tiiviissä vuorovaikutuksessa toistensa kanssa, mikä sallii niiden suorittaa solutoimintojen hienosäätelyn.
Oppaat ja oppaat
1. Alberts SISÄÄN., Bray D., Lewis J., Raff M., Roberts TO., Watson J.D.Molecular Biology of the Cell, New York ja Lontoo, Garland Publishing Inc., 1983.
2. Czihak G., Longer H., Ziegler H.(toim.). Biologia. Berliini, Heidelberg, New York, Springer, 1983.
3. Hille SISÄÄN. Herätettävien kalvojen ionikanavat. Sunderland, Mass., Sinauer Assoc., 1984.
4. Hoppe W., Lohmann W.. Marki H., Ziegler H.(toim.). Biofysiikka. Berliini, Heidelberg, New York, Springer, 1984.
5. Jungermann TO., Mahler H.Biochemie. Berliini, Heidelberg, New York, Springer, 1980.
6. Kandel E. R., Schwartz – J. H.,(toim.). Neurotieteen periaatteet, New York, Amsterdam, Oxford, Elsevier, 1985.
7. Schiebler T. H., Schmidt W.Anatomic des Menschen. Berliini, Heidelberg, New York, Tokio, Springer, 1983.
Alkuperäiset artikkelit ja arvostelut
8. Berridge M.J. Solun sisäisen viestinnän molekyyliperusta, Sci. Amer, 253, 124 - 134 (1985).
9. Berridge M. J., Irvine R. F. Inositolitrifosfaatti, uusi toinen lähettiläs solujen signaalinsiirrossa. Nature, 312, 315, 321 (1984).
10. Bretscher M.S. Solukalvon molekyylit, Sei. Amer., 253, 124-134 (1985).
11. Daut J. Elävä solu energiaa muuntavana koneena. Sydänlihaksen aineenvaihdunnan minimimalli, Biochem. et Biophys. Acta, 895, 41-62 (1987).
12. Hodgkin A.L., Katz SISÄÄN. Natrium-ionien vaikutus kalmarin jättimäisen aksonin sähköiseen aktiivisuuteen. J. Physiol. (Lond.), 108, 37–77 (1949).
13. Hodgkin A.L., Keynes R.D. Kationien aktiivinen kuljetus jättimäisissä aksoneissa Seepia ja Loligo, J. Physiol. (Lond.), 128, 28–42 (1955).
14. Pidempi P. Ionikanavat konformaatioalatiloilla, Biophys. J., 47, 581-590 (1985).
15. Ochs S., Worth P.M. Eksoplasminen kuljetus normaaleissa ja patologisissa järjestelmissä. Julkaisussa: Physiology and Pathology of Axons, S.G. Waxmann, Ed. New York, Raven Press, 1978.
yhteenveto muista esityksistä"Biologian opetusmenetelmät" - Koulueläintiede. Opiskelijoiden tutustuttaminen tieteellisen eläintieteellisen tiedon käyttöön. Moraalinen koulutus. Kanakopan lisäsiunaus. Menetelmien valinta. Elämän prosessit. Akvaariokalat. Ravitsemus. Ympäristökasvatus. Elämänprosessien olennaisuus. Negatiiviset tulokset. Huomio opiskelijat. Pakollinen lomake. Pieniä eläimiä katselemassa. Biologian tavoitteet ja tavoitteet. Tarina.
"Ongelmapohjainen oppiminen biologian tunneilla" - Tieto. Uusia oppikirjoja. Polku ratkaisuun. Ongelma. Seminaarit. Mikä on tehtävä? Albrecht Durer. Ongelmalähtöistä oppimista biologian tunneilla. Epätyypilliset oppitunnit. Mitä tarkoittaa ongelmapohjainen oppiminen. Elämän laatu. Biologia akateemisena aineena. Kysymys. Ongelmanratkaisutunti. Vähentynyt kiinnostus aiheeseen. Ongelmapohjaiset laboratoriotunnit.
"Kriittinen ajattelu biologian tunneilla" - "Kriittisen ajattelun" tekniikka. Käyttämällä "kriittisen ajattelun kehittämisen" teknologiaa. Pöytä oppitunnille. Motivaatio opiskella. Ekosysteemit. Sanan "kriittisen ajattelun kehittäminen" merkitys. Tekniikan merkkejä. RKM-tekniikkaa. Oppitunnin rakenne. Pääsuunnat. Tekniikan historia. Pedagogiset tekniikat. Tekniikan säännöt. Biologian tehtävät. Fotosynteesi. Oppitunnin eri vaiheissa käytetyt tekniikat.
"Biologian tunnit interaktiivisella taululla" - Elektroniset oppikirjat. Edut opiskelijoille. Interaktiivinen taulu auttaa välittämään tietoa jokaiselle opiskelijalle. Didaktiset tehtävät. Biologisten ongelmien ratkaiseminen. Interaktiivisten taulujen kanssa työskentelyn edut. Työskentely esitysten kanssa. Työskentele esineiden vertailussa. Liikkuvat esineet. Laskentataulukoiden käyttäminen. Interaktiivisen taulun käyttö koululaisten opetusprosessissa. Edut opettajille.
"System-Activity lähestymistapa biologiassa" - Seminaarikysymykset. Aktiviteettimenetelmä. Dryopithecus. Ihmisestä peräisin oleva maan ulkopuolinen polku. Lysosomit. Kemiallinen organisaatio. Gymnossperms. Aineenvaihdunta. Analysaattorit. Systeemiaktiivinen lähestymistapa biologian opetukseen. Kromosomit. Sytoplasma. Sokeus. Korvan pituus. Henkilön luokitus. Nisäkkään luuranko. Ihmisen evoluution polut. Mitoosi. Pinnallinen kompleksi. Ongelmallinen kysymys. Nucleolus. Ydinvaippa.
"Tietokone biologiassa" - Opiskelijoiden yhteistoiminta. Angiosperm-perheet. Interaktiivinen koulutus. Oppimismalleja. Esimerkki arviointijärjestelmästä. Ohjekorttikysymykset. Esimerkki ohjekortista. Tutkijat. Mikroryhmät. Interaktiiviset oppimistekniikat. Karuselli. Interaktiiviset oppimistekniikat. Interaktiivisia lähestymistapoja biologian tunneilla. Ryhmätyömuoto. Tehtävät "tutkijaryhmille".
