§ 2. Eikariotu šūnas galvenās sastāvdaļas
Eikariotu šūnas (8. un 9. att.) ir sakārtotas daudz sarežģītāk nekā prokariotu šūnas. Tie ir ļoti dažādi gan pēc izmēra (no vairākiem mikrometriem līdz vairākiem centimetriem), gan pēc formas, gan pēc konstrukcijas īpatnībām (10. att.).
Rīsi. 8. Eikariotu šūnas uzbūve. Vispārināta shēma
Rīsi. 9. Šūnu uzbūve pēc elektronmikroskopijas
Rīsi. 10. Dažādas eikariotu šūnas: 1 – epitēlija; 2 – asinis (e – eritroīts, / – leikīts); 3 – skrimslis; 4 – kauli; 5 – gludie muskuļi; 6 – saistaudi; 7 – nervu šūnas; 8 – šķērssvītrota muskuļu šķiedra
Tomēr vispārējā organizācija un fundamentālo komponentu klātbūtne visās eikariotu šūnās ir vienāda (11. att.).
Rīsi. 11. Eikariotu šūna (diagramma)
Plazmalemma (šūnu ārējā membrāna). Plazmas membrānas pamats, tāpat kā citas šūnu membrānas (piemēram, mitohondriji, plastidi u.c.), ir lipīdu slānis, kurā ir divas molekulu rindas (12. att.). Tā kā lipīdu molekulas ir polāras (viens pols ir hidrofils, t.i., to pievelk ūdens, bet otrs ir hidrofobs, t.i., ūdens atgrūž), tās ir sakārtotas noteiktā secībā. Viena slāņa molekulu hidrofilie gali ir vērsti uz ūdens vidi - šūnas citoplazmā, bet otra slāņa - uz āru no šūnas - uz starpšūnu vielu (daudzšūnu organismos) vai ūdens vidi (vienšūnu organismos). ).
Rīsi. 12. Šūnu membrānas uzbūve pēc šķidruma mozaīkas modeļa. Olbaltumvielas un glikoproteīni ir iegremdēti dubultā lipīdu molekulu slānī ar to hidrofilajiem galiem (apļiem) uz āru un to hidrofobajiem galiem (viļņainām līnijām) dziļi membrānā.
Olbaltumvielu molekulas ir mozaīkas veidā iestrādātas bimolekulārā lipīdu slānī. Dzīvnieka šūnas ārpusē polisaharīdu molekulas ir pievienotas plazmlemmas lipīdiem un olbaltumvielu molekulām, veidojot glikolipīdus un glikoproteīnus.
Šis pildviela veido slāni glikokalikss. Saistīts ar viņu receptoru funkcija plazmas membrānas (skatīt zemāk); tajā var uzkrāties arī dažādas šūnas izmantotās vielas. Turklāt glikokalikss uzlabo plazmalemmas mehānisko stabilitāti.
Augu un sēņu šūnās ir arī šūnu siena, kurai ir atbalsta un aizsargājoša loma. Augos tas sastāv no celulozes, bet sēnēs - no hitīna.
Šūnu ārējā membrāna veic vairākas funkcijas, tostarp:
♦ mehānisks(atbalstīšana, formu veidošana);
♦ barjeru transports(selektīvā caurlaidība dažādām vielām: nepieciešamo vielu iekļūšana šūnā un nevajadzīgo un kaitīgo izvadīšana);
♦ receptoru(dažādu šūnas tiešā tuvumā atrasto ķīmisko vielu noteikšana; signālu uztveršana hormonu veidā; imūnsistēmas šūnu „svešā” proteīna atpazīšana utt.).
Vielu apmaiņa starp šūnu un vidi tiek veikta dažādos veidos - pasīvā un aktīva.
Ūdens un dažādu jonu molekulas pasīvi (difūzijas, osmozes dēļ), šūnai netērējot enerģiju, nonāk caur īpašām porām - tās ir pasīvais transports. Makromolekulas, piemēram, olbaltumvielas, polisaharīdi, pat veselas šūnas, piegādā fagocitoze Un pinocitoze ar enerģijas patēriņu - aktīvais transports.
Ar fagocitozi tiek uzņemtas veselas šūnas vai lielas daļiņas (piemēram, padomājiet par amēbu uzturu vai baktēriju aizsargājošo asins šūnu fagocitozi). Pinocitozes laikā tiek absorbētas nelielas šķidras vielas daļiņas vai pilieni. Abiem procesiem kopīgs ir tas, ka absorbētās vielas ieskauj invaginējoša ārējā membrāna, veidojot vakuolu, kas pēc tam dziļi pārvietojas šūnas citoplazmā.
Eksocitoze ir fagocitozei un pinocitozei pretējs process (arī aktīvs transports) (13. att.). Ar tās palīdzību var izņemt nesagremotas barības atliekas vienšūņos vai izvadīt bioloģiski aktīvās vielas, kas veidojas sekrēcijas šūnā.
Citoplazma. Citoplazma ir šūnas saturs, ko ierobežo plazmalemma, izņemot kodolu. Tas satur zemes viela (hialoplazma), organellas Un ieslēgumi.
Hialoplazma- viskozs šķidrums, kas var būt vienā vai otrā stāvoklī Zola(šķidrums), vai želeja(želejveida).
Ja nepieciešams, citoplazma spēj atgriezeniski pāriet no viena stāvokļa uz otru. Piemēram, amēboīdu kustības laikā (atcerieties sadaļu “Protozoa” no zooloģijas kursa), veidojoties pseidopodam, notiek straujas citoplazmas pārejas no gēla uz solu un otrādi. Tas ir saistīts ar to, ka citoplazmā ir liels skaits pavedienveida olbaltumvielu molekulu aktīna. Kad tie savienojas viens ar otru, veidojot trīsdimensiju tīklu, citoplazma atrodas želejas stāvoklī, un, kad tīkls sadalās, tā atrodas sola stāvoklī.
Hialoplazmā ir dažādas vielas – fermenti, olbaltumvielas, ogļhidrāti, tauki un citas, organiskas un minerālvielas. Šeit notiek dažādi ķīmiskie procesi - vielu sadalīšanās, to sintēze un modifikācijas (izmaiņas).
Organoīdi. Tās ir pastāvīgas šūnas sastāvdaļas ar noteiktu struktūru un funkcijām, kas atrodas tās citoplazmā. Tālāk mēs runāsim par vispārējas nozīmes organoīdi, raksturīgs visu veidu eikariotu šūnām. Tie ir saistīti ar pēdējo dzīvībai svarīgo funkciju nodrošināšanu. Speciālie organoīdi atrodami tikai noteikta (ļoti specializēta) tipa šūnās - piemēram, miofibrils muskuļu šūnās.
Vispārējas nozīmes organellām ir tāda pati struktūra neatkarīgi no tā, kurām šūnām un organismiem tās pieder. Bet starp tiem ir grupas ar membrānu (endoplazmatiskais tīkls, Golgi aparāts, mitohondriji, plastidi, lizosomas, vakuoli), kā arī nemembrānas ( ribosomas, šūnu centrs) struktūra.
Endoplazmatiskais tīkls (ER). EPS sastāv no membrānām un ir sarežģīta sazarota kanāliņu un cisternu sistēma, kas iekļūst visā šūnas citoplazmā (14. att.). Ir divu veidu EPS - raupja Un gluda. Ribosomas ir piestiprinātas pie raupjām membrānām (no citoplazmas puses), bet uz gludajām membrānām tās nav.
Rīsi. 14. Endoplazmatiskais tīklojums
Endoplazmatiskais retikulums eikariotu šūnā veic vairākas svarīgas funkcijas:
♦ norobežojot(šūnas iekšējā tilpuma sadalīšana dažādās reakcijas telpās);
♦ dalība organisko vielu sintēzē(ribosomas atrodas uz raupjā ER membrānām, un enzīmu kompleksi atrodas uz gludajām, nodrošinot lipīdu, ogļhidrātu uc sintēzi);
♦ līdzdalība Golgi aparāta elementu, lizosomu veidošanā;
♦ vielu transportēšana.
Golgi aparāts. Golgi aparāts (AG) ir sistēma tvertnes(plakanās vakuolas) un burbuļi(pūslīši), kas atrodas tiešā šūnas kodola tuvumā, kas veidojas EPS dēļ tās mazo fragmentu atdalīšanas rezultātā (15. att.). Saplūstot šiem fragmentiem, rodas jaunas Golgi aparāta cisternas, savukārt no EPS tiek transportētas dažādas vielas, kas ir iesaistītas sarežģītu organisko savienojumu (olbaltumvielas + ogļhidrāti, olbaltumvielas + lipīdi u.c.) montāžā, kas tiek izvadīti kopā ar AG palīdzība ārpus šūnas. Šīs bioloģiski aktīvās vielas tiek izņemtas no šūnas (izmantojot sekrēcijas vakuolus eksocitozes ceļā), vai arī ir daļa no lizosomām (skatīt zemāk), ko veido AG.
Rīsi. 15. Golgi aparāts:
Golgi aparāts veic šādas funkcijas:
♦ sintēze bioloģiski aktīvās vielas, ko ražo šūna;
♦ dažādu vielu sekrēcija (izņemšana no šūnas).(hormoni, fermenti, vielas, no kurām būvē šūnu sienu u.c.);
♦ dalība lizosomu veidošanā.
Mitohondriji. Visu veidu eikariotu šūnās ir mitohondriji (16. att.). Tie izskatās kā noapaļoti ķermeņi vai stieņi, retāk - pavedieni. To izmēri svārstās no 1 līdz 7 mikroniem. Mitohondriju skaits šūnā svārstās no vairākiem simtiem līdz desmitiem tūkstošu (lielos vienšūņos).
Rīsi. 16.Mitohondriji. Augšā - mitohondriji (a) urīnceļos, redzami gaismas mikroskopā. Zemāk ir mitohondriju organizācijas trīsdimensiju modelis: 1 – cristae; 2 – ārējā membrāna; 3 – iekšējā membrāna; 4 – matrica
Mitohondriju veido divas membrānas - ārējā Un iekšējs, starp kurām atrodas starpmembrānu telpa. Iekšējā membrāna veido daudzas invaginācijas - cristae, kas ir vai nu plāksnes, vai caurules. Šī organizācija nodrošina milzīgu iekšējās membrānas laukumu. Tas satur fermentus, kas nodrošina organiskajās vielās (ogļhidrātos, lipīdos) esošās enerģijas pārvēršanu ATP enerģijā, kas nepieciešama šūnas dzīvībai. Tāpēc mitohondriju funkcija ir piedalīties enerģijušūnu procesi. Tāpēc liels skaits mitohondriju ir raksturīgi, piemēram, muskuļu šūnām, kas veic lielu darbu.
Plastīdi. Augu šūnās atrodamas īpašas organellas - plastidi, kuriem bieži ir vārpstveida vai noapaļota forma, dažreiz sarežģītāka. Ir trīs veidu plastidi - hloroplasti (17. att.), hromoplasti un leikoplasti.
Hloroplasti atšķiras zaļā krāsā, kas ir pigmenta dēļ - hlorofils, procesa nodrošināšana fotosintēze, i., organisko vielu sintēze no ūdens (H 2 O) un oglekļa dioksīda (CO 2), izmantojot saules gaismas enerģiju. Hloroplasti galvenokārt atrodami lapu šūnās (augstākajos augos). Tos veido divas membrānas, kas atrodas paralēli viena otrai un ieskauj hloroplastu saturu - stroma. Iekšējā membrāna veido daudzus saplacinātus maisiņus - tilakoīdi, kas ir sakrautas (kā monētu kaudze) - graudi - un guli stromā. Tie ir tilakoīdi, kas satur hlorofilu.
Hromoplasti nosaka dzelteno, oranžo un sarkano krāsu daudziem ziediem un augļiem, kuru šūnās tie atrodas lielos daudzumos. Galvenie pigmenti to sastāvā ir karotīni. Hromoplastu funkcionālais mērķis ir piesaistīt dzīvniekus ar krāsu, nodrošinot ziedu apputeksnēšanu un sēklu izplatīšanos.
Rīsi. 17. Plastīdi: a – gaismas mikroskopā redzami hloroplasti Elodejas lapas šūnās; b – hloroplasta iekšējās struktūras diagramma ar granām, kas ir plakanu maisiņu krāvumi, kas atrodas perpendikulāri hloroplasta virsmai; c – detalizētāka diagramma, kurā parādītas anastomozes caurules, kas savieno atsevišķas ventilatora kameras
Leikoplasti ir bezkrāsaini plastidi, kas atrodas augu pazemes daļu šūnās (piemēram, kartupeļu bumbuļos), sēklās un stublāju serdē. Leikoplastos ciete galvenokārt veidojas no glikozes un uzkrājas augu uzglabāšanas orgānos.
Viena veida plastidi var pārveidoties par citu. Piemēram, kad rudenī lapas maina krāsu, hloroplasti pārvēršas hromoplastos.
Lizosomas.Šīs organellas izskatās kā pūslīši, ko ieskauj membrāna ar diametru līdz 2 mikroniem. Tie satur vairākus desmitus enzīmu, kas noārda olbaltumvielas, nukleīnskābes, polisaharīdus un lipīdus. Lizosomu funkcija ir piedalīties komplekso organisko savienojumu (piemēram, barības vielu vai “izlietoto” šūnu komponentu) intracelulārā sadalīšanās procesos. Lizosomas saplūst ar fagocītiskām (vai pinocītu) vakuolām, veidojot gremošanas vakuolu.
Lizosomu veidošanās notiek Golgi aparāta cisternu veidošanās dēļ.
Ribosomas. Ribosomas (18. att.) atrodas gan eikariotu, gan prokariotu šūnās, jo tām ir svarīga funkcija olbaltumvielu biosintēze(skat. 5. nodaļu). Katrā šūnā ir desmitiem, simtiem tūkstošu (līdz pat vairākiem miljoniem) šo mazo apaļo organellu.
Rīsi. 18. Uz endoplazmatiskā tīkla membrānas sēdošas ribosomas uzbūves shēma: 1 – mazā apakšvienība; 2 – tRNS; 3 – aminoacil-tRNS; 4 – aminoskābe; 5 – liela apakšvienība; 6 – endoplazmatiskā tīkla membrāna; 7 – sintezēta polipeptīdu ķēde
Ribosoma sastāv no divām nevienlīdzīgām apakšvienībām (daļām). Tie tiek veidoti atsevišķi un proteīna molekulas sintēzes laikā apvienojas, “aptverot” ziņojuma RNS. Ribosomas satur dažādus proteīnus un ribosomu RNS.
Šūnu ieslēgumi. Tas ir nosaukums, kas dots nepastāvīgajiem komponentiem šūnā, kas atrodas citoplazmas galvenajā vielā graudu, granulu vai pilienu veidā. Ieslēgumus var vai nevar ieskauj membrāna.
Funkcionāli ir trīs veidu ieslēgumi: rezerves barības vielas(ciete, glikogēns, tauki, olbaltumvielas), sekrēcijas ieslēgumi(dziedzeru šūnām raksturīgas vielas, ko tās ražo - endokrīno dziedzeru hormoni utt.) un īpaša mērķa iekļaušana(ļoti specializētās šūnās, piemēram, hemoglobīns sarkanajās asins šūnās).
§ 3. Šūnas kodola organizācija. Hromosomas
Šūnas kodolam (sk. 8. un 9. att.) ir ārkārtīgi liela nozīme šūnas dzīvē, jo tas kalpo kā hromosomās ietvertās iedzimtības informācijas krātuve (skatīt zemāk).
Kodols ir ierobežots ar kodola apvalku, kas atdala tā saturu (karioplazma) no citoplazmas. Apvalks sastāv no divām membrānām, kuras atdala sprauga. Abas ir caurstrāvotas ar daudzām porām, pateicoties kurām ir iespējama vielu apmaiņa starp kodolu un citoplazmu. Lielākajā daļā eikariotu šūnu kodolā ir no 1 līdz 7 kodoli. Ar tiem ir saistīti RNS un tRNS sintēzes procesi.
Kodola galvenās sastāvdaļas - hromosomas, veidojas no DNS molekulas un dažādiem proteīniem. Gaismas mikroskopā tie ir skaidri redzami tikai šūnu dalīšanās periodā (mitoze, mejoze). Nedalošā šūnā hromosomas izskatās kā gari plāni pavedieni, kas sadalīti visā kodola tilpumā.
Šūnu dalīšanās laikā hromosomu pavedieni veido blīvas spirāles, kā rezultātā kļūst redzamas (izmantojot parasto mikroskopu) stieņu, “matadatu” veidā. Viss ģenētiskās informācijas apjoms ir sadalīts starp kodola hromosomām. To izpētes procesā tika identificēti šādi modeļi:
♦ somatisko šūnu kodolos (t.i., neseksuālās ķermeņa šūnās) visi vienas sugas indivīdi satur vienādu skaitu hromosomu, kas veido hromosomu komplekts(19. att.);
Rīsi. 19. Dažādu augu un dzīvnieku sugu hromosomas, attēlotas vienā mērogā: 1,2 – amēba; 3,4 – kramaļģes; 5–8, 18,19 – zaļaļģes; 9 – mušmire; 10 – liepa; 11–12 – Drosophila; 13 – lasis; 14 – skerda (Asteraceae dzimta); 15 – augs no aroid dzimtas; 16 – corydalis tauriņš; 17 – kukainis no siseņu dzimtas; 20 – ūdens strider blaktis; 21 – puķu blakts; 22 – abinieku ambistoma; 23 - alveja (liliju ģimene)
♦ katrai sugai ir raksturīgs savs hromosomu komplekts atbilstoši to skaitam (piemēram, cilvēkam ir 46 hromosomas, Drosophila mušai – 8, apaļtārpam – 4, vēžiem – 196, zirgam – 66, kukurūzai – 104);
♦ hromosomas somatisko šūnu kodolos var grupēt pa pāriem, ko sauc homologās hromosomas pamatojoties uz to līdzību (struktūrā un funkcijās);
♦ dzimumšūnu (gametu) kodolos katrs homologo hromosomu pāris satur tikai vienu, t.i., kopējais hromosomu kopums ir uz pusi mazāks nekā somatisko šūnu;
♦ sauc par vienu hromosomu kopu dzimumšūnās haploīds un tiek apzīmēts ar burtu n, un somatiski - diploīds(2n).
No iepriekš minētā ir skaidrs, ka katrs homologo hromosomu pāris veidojas, saplūstot tēva un mātes hromosomām apaugļošanas laikā, t.i., dzimumšūnu (gametu) saplūsmē. Un otrādi, dzimumšūnu veidošanās laikā no katra homologo hromosomu pāra gametē nonāk tikai viena.
Hromosomas savādāk homologie pāri atšķiras pēc izmēra un formas (20. un 21. att.).
Rīsi. 20. Hromosomu uzbūve un veidi: a – izskats 1 – centromērs; 2 – īss plecs; 3 – garais plecs); vienas un tās pašas hromosomas iekšējā struktūra (1 – centromērs; – DNS molekulas); c – hromosomu veidi (1 – vienroku; dažādu roku; 3 – vienādroku: X – roka, V – centromērs)
Rīsi. 21. Hromosoma sastāv no DNS un olbaltumvielām. DNS molekula tiek replicēta. Divas identiskas DNS dubultās spirāles paliek savienotas centromēra reģionā. Šīs kopijas vēlāk šūnu dalīšanās laikā kļūst par atsevišķām hromosomām
Hromosomu ķermenī ir primārais sašaurinājums (saukts par centromēru), pie kuriem ir piestiprināti pavedieni skaldīšanas vārpstas. Tas sadala hromosomu divās daļās plecu Hromosomas var būt ar vienādām rokām, ar nevienlīdzīgām rokām vai ar vienu roku.
5. nodaļa. Metabolisms
§ 1. Metabolisms kā asimilācijas un disimilācijas vienotība
Visas šūnas un dzīvie organismi ir atvērtas sistēmas, tas ir, tie atrodas nepārtrauktas enerģijas un vielu apmaiņas stāvoklī ar vidi. Nedzīvajā dabā ir atvērtas sistēmas, taču to eksistence kvalitatīvi atšķiras no dzīviem organismiem. Apsveriet šo piemēru: degošs vietējā sēra gabals atrodas apmaiņas stāvoklī ar vidi. Kad tas sadedzina, tiek absorbēts O 2, un izdalās SO 2 un enerģija (siltuma veidā). Tomēr šajā gadījumā sēra gabals kā fiziskais ķermenis tiek iznīcināts un zaudē savu primāro struktūru.
Dzīviem organismiem apmaiņa ar vidi izrādās nosacījums to strukturālās organizācijas saglabāšanai un uzturēšanai, pašatjaunojoties visām vielām un sastāvdaļām, no kurām tie sastāv.
Metabolisms (vielmaiņa) ir dzīvajos organismos notiekošu procesu kopums (vielu un enerģijas patēriņš, transformācija, uzkrāšanās un izdalīšanās), kas nodrošina to dzīvības aktivitāti, attīstību, augšanu, vairošanos. Vielmaiņas procesā tiek sadalītas un sintezētas molekulas, kas veido šūnas; šūnu struktūru un starpšūnu vielas atjaunošana.
Metabolisma pamatā ir savstarpēji saistīti procesi asimilācija(anabolisms) un disimilācija(katabolisms). Asimilācijas (plastiskās apmaiņas) laikā sarežģītas vielas tiek sintezētas no vienkāršām. Pateicoties tam, šūnā rodas visas organiskās vielas, kas nepieciešamas tās strukturālo komponentu, enzīmu sistēmu u.c. uzbūvei. Asimilācija vienmēr notiek ar enerģijas patēriņu.
Disimilācijas (enerģijas vielmaiņas) laikā sarežģītās organiskās vielas sadalās vienkāršākos vai neorganiskās. Šajā gadījumā tiek atbrīvota enerģija, ko šūna tērē dažādu procesu veikšanai, kas nodrošina tās dzīvībai svarīgo darbību (vielu sintēze un transportēšana, mehāniskais darbs utt.).
Visus dzīvos organismus var iedalīt divās grupās: autotrofi Un heterotrofi, kas atšķiras ar enerģijas avotiem un to dzīvības funkciju nodrošināšanai nepieciešamajām vielām.
Autotrofi- organismi, kas sintezē organiskos savienojumus no neorganiskām vielām, izmantojot saules gaismas enerģiju (kā fototrofi– augi, zilaļģes) vai enerģija, kas iegūta, oksidējot minerālvielas (neorganiskās) vielas (piemēram, ķīmijtrofi– sēra baktērijas, dzelzs baktērijas utt.). Līdz ar to viņi spēj patstāvīgi radīt savai dzīves darbībai nepieciešamās vielas.
§ 2. Disimilācija anaerobos un aerobos organismos
Organismus var iedalīt divās grupās un pēc disimilācijas veida - aerobi Un anaerobi. Aerobes (no grieķu. dēmons– gaiss) dzīvībai nepieciešams brīvs skābeklis. Anaerobos (grieķu. ac– negatīva daļiņa) tas nav nepieciešams. Tajos tiek veikta disimilācija ar fermentāciju - bezskābekļa, organisko vielu enzīmu sadalīšana ar vienkāršāku organisko vielu veidošanos un enerģijas izdalīšanos. Piemēram:
♦ pienskābes fermentācija:
C6H12O6 + 2H3PO4 + 2ADP → 2PH + 2ATP + 2H2O;
♦ alkoholiskā fermentācija:
C6H12O6 + 2PH + 2ADP → 2C2H5OH + 2CO2 + 2ATP + 2H2O.
Vielas, kas veidojas fermentācijas laikā, ir organiskas un tāpēc joprojām satur daudz enerģijas.
Rīsi. 22. Asimilācijas un disimilācijas attiecības autotrofos un heterotrofos organismos
Aerobos organismos elpošanas procesā mitohondrijās notiek pilnīga organisko vielu sadalīšanās (izmantojot O 2) līdz enerģētiski nabadzīgajiem galaproduktiem CO 2 un H 2 O un izdalās ievērojami lielāks enerģijas daudzums:
C 6 H 12 0 6 (glikoze) + 0 2 > 6C0 2 + 6H 2 0 + enerģija (kuras dēļ tiek sintezētas 38 ATP molekulas).
Apskatīsim vispārinātu shēmu veidā autotrofo un heterotrofo aerobo organismu metabolismu (22. att.).
Asimilācija. Tās svarīgākie procesi ir fotosintēze Un proteīnu biosintēze.
Organisma saistība ar vidi no fizikāli ķīmiskā viedokļa ir atvērta sistēma, tas ir, sistēma, kurā notiek bioķīmiskie procesi. Izejvielas nāk no vides, un vielas, kas arī nepārtraukti veidojas, tiek iznestas ārā. Līdzsvars starp daudzvirzienu reakciju produktu ātrumu un koncentrāciju organismā ir nosacīts, iedomāts, jo vielu uzņemšana un izvadīšana neapstājas. Nepārtraukta saikne ar vidi ļauj uzskatīt dzīvo organismu par atvērtu sistēmu.
Visām dzīvajām šūnām enerģijas avots ir Saule. Augu šūnas ar hlorofila palīdzību uztver enerģiju no saules gaismas, izmantojot to asimilācijas reakcijās fotosintēzes procesā. Dzīvnieku, sēnīšu un baktēriju šūnas saules enerģiju izmanto netieši, sadalot zemes augu sintezētās organiskās vielas.
Daļa šūnu barības vielu sadalās šūnu elpošanas laikā, tādējādi piegādājot dažāda veida šūnu darbībai nepieciešamo enerģiju. Šis process notiek organellās, ko sauc par mitohondrijiem. Mitohondriji sastāv no divām membrānām: ārējās, kas atdala organellu no citoplazmas, un iekšējās, kas veido daudzas krokas. Galvenais elpošanas produkts ir ATP. Tas atstāj mitohondrijus un tiek izmantots kā enerģijas avots daudzām ķīmiskām reakcijām citoplazmā un šūnu membrānā. Ja šūnu elpošanai nepieciešams skābeklis, tad elpošanu sauc par aerobo, bet, ja reakcijas notiek bez skābekļa, tad runājam par anaerobo elpošanu.
Jebkura veida darbam, ko veic šūnā, enerģija tiek izmantota vienā un vienīgajā veidā - enerģijas veidā no ATP fosfātu saitēm. ATP ir viegli mobils savienojums. ATP veidošanās notiek uz mitohondriju iekšējās membrānas. ATP tiek sintezēts visās šūnās elpošanas laikā, pateicoties ogļhidrātu, tauku un citu organisko vielu oksidācijas enerģijai. Zaļo augu šūnās galvenais ATP daudzums saules enerģijas ietekmē tiek sintezēts hloroplastos. Fotosintēzes laikā tie ražo daudzkārt vairāk ATP nekā mitohondriji. ATP sadalās, pārraujot fosfora-skābekļa saites un atbrīvojoties enerģijai. Tas notiek enzīma ATPāzes iedarbībā ATP hidrolīzes laikā - ūdens pievienošana ar fosforskābes molekulas izvadīšanu. Rezultātā ATP tiek pārveidots par ADP, un, ja tiek atdalītas divas fosforskābes molekulas, tad par AMP. Katras grama skābes molekulas eliminācijas reakciju pavada 40 kJ izdalīšanās. Tā ir ļoti liela enerģijas izvade, tāpēc ATP fosfora-skābekļa saites parasti sauc par makroerģistiskām (augstas enerģijas).
ATP izmantošana plastmasas apmaiņas reakcijās tiek veikta, savienojot tās ar ATP hidrolīzi. Dažādu vielu molekulas tiek uzlādētas ar enerģiju, piesaistot no ATP molekulas hidrolīzes laikā izdalīto fosfora grupu, t.i., fosforilējot.
Fosfātu atvasinājumu īpatnība ir tāda, ka tie nevar atstāt šūnu, lai gan to “izlādētās” formas brīvi iziet cauri membrānai. Pateicoties tam, fosforilētās molekulas paliek šūnā, līdz tiek izmantotas atbilstošās reakcijās.
Apgrieztais process ADP pārveidošanai par ATP notiek, pievienojot ADP fosforskābes molekulu, atbrīvojot ūdeni un absorbējot lielu enerģijas daudzumu.
Tādējādi ATP ir universāls un tiešs enerģijas avots šūnu darbībai. Tas rada vienotu šūnu enerģijas fondu un ļauj to pārdalīt un transportēt no vienas šūnas zonas uz citu.
Fosfātu grupas pārnešanai ir svarīga loma ķīmiskās reakcijās, piemēram, makromolekulu montāžā no monomēriem. Piemēram, aminoskābes var apvienot peptīdos tikai pēc iepriekšējas fosforilēšanas. Mehāniskie kontrakcijas vai kustības procesi, izšķīdušās vielas transportēšana pret koncentrācijas gradientu un citi procesi ietver ATP uzkrātās enerģijas patēriņu.
Enerģijas metabolisma procesu var attēlot šādi. Citoplazmā esošās lielmolekulārās organiskās vielas fermentatīvi hidrolīzes ceļā tiek pārveidotas par vienkāršākām, no kurām tās sastāv: olbaltumvielas - aminoskābēs, poli- un disaharīdi - monosaharīdos (+ glikoze), tauki glicerīnā un taukskābēs. Nav oksidatīvo procesu, izdalās maz enerģijas, kas netiek izmantota un nonāk termiskā formā. Lielākā daļa šūnu vispirms izmanto ogļhidrātus. Polisaharīdi (ciete augos un glikogēns dzīvniekiem) tiek hidrolizēti līdz glikozei. Glikozes oksidēšanās notiek trīs fāzēs: glikolīze, oksidatīvā dekarboksilēšana (Krebsa cikls – citronskābes cikls) un oksidatīvā fosforilēšanās (elpošanas ķēde). Citoplazmā notiek glikolīze, kuras rezultātā viena glikozes molekula tiek sadalīta divās pirovīnskābes molekulās, atbrīvojot divas ATP molekulas. Ja nav skābekļa, pirovīnskābe tiek pārveidota par etanolu (fermentācija) vai pienskābi (anaerobā elpošana).
Kad dzīvnieku šūnās notiek glikolīze, sešu oglekļa glikozes molekula sadalās divās pienskābes molekulās. Šis process ir daudzpakāpju. To secīgi veic 13 fermenti. Alkoholiskās fermentācijas laikā no glikozes molekulas veidojas divas etanola molekulas un divas CO2 molekulas.
Glikolīze ir fāze, kas raksturīga anaerobai un aerobai elpošanai, pārējās divas notiek tikai aerobos apstākļos. Bezskābekļa oksidācijas process, kurā tiek atbrīvota un izmantota tikai daļa no metabolītu enerģijas, ir galīgs anaerobiem organismiem. Skābekļa klātbūtnē pirovīnskābe nokļūst mitohondrijās, kur vairāku secīgu reakciju rezultātā tā aerobiski tiek pilnībā oksidēta līdz H2O un CO2, vienlaikus fosforilējot ADP līdz ATP. Šajā gadījumā divas ATP molekulas tiek ražotas ar glikolīzi, divas ar Krebsa ciklu un 34 ar elpošanas ķēdi. Neto iznākums vienas glikozes molekulas pilnīgai oksidēšanai par H2O un CO2 ir 38 molekulas.
Tādējādi aerobos organismos organisko vielu galīgā sadalīšanās tiek veikta, oksidējot tās ar atmosfēras skābekli līdz vienkāršām neorganiskām vielām: CO2 un H2O. Šis process notiek uz mitohondriju kristāla. Šajā gadījumā tiek atbrīvots maksimālais brīvās enerģijas daudzums, no kura ievērojama daļa tiek rezervēta ATP molekulās. Ir viegli redzēt, ka aerobā oksidēšana nodrošina šūnu vislielāko brīvo enerģiju.
Katabolisma rezultātā šūnā uzkrājas ar enerģiju bagātas ATP molekulas, un ārējā vidē izdalās CO2 un liekais ūdens.
Šūnā var uzglabāt cukura molekulas, kas nav vajadzīgas elpošanai. Pārmērīgie lipīdi tiek vai nu sadalīti, pēc tam to sadalīšanās produkti nonāk mitohondrijās kā elpošanas substrāts, vai arī tiek nogulsnēti kā rezerves citoplazmā tauku pilienu veidā. Olbaltumvielas tiek veidotas no aminoskābēm, kas nonāk šūnā. Olbaltumvielu sintēze notiek organellās, ko sauc par ribosomām. Katra ribosoma sastāv no divām apakšdaļiņām – lielām un mazām: abās apakšdaļiņās ir proteīna molekulas un RNS molekulas.
Ribosomas bieži ir pievienotas īpašai membrānu sistēmai, kas sastāv no cisternām un pūslīšiem – tā sauktajam endoplazmatiskajam tīklam (ER); šūnās, kas ražo daudz olbaltumvielu, endoplazmatiskais tīklojums bieži ir ļoti labi attīstīts un pārklāts ar ribosomām. Daži fermenti ir efektīvi tikai tad, ja tie ir pievienoti membrānai. Šeit atrodas lielākā daļa lipīdu sintēzē iesaistīto enzīmu. Tādējādi endoplazmatiskais tīkls ir kā sava veida šūnu darbagalds.
Turklāt ER sadala citoplazmu atsevišķos nodalījumos, t.i., atdala dažādus ķīmiskos procesus, kas vienlaikus notiek citoplazmā, un tādējādi samazina iespējamību, ka šie procesi traucēs viens otru.
Produkti, ko ražo konkrēta šūna, bieži tiek izmantoti ārpus šūnas. Šādos gadījumos uz ribosomām sintezētie proteīni iziet cauri endoplazmatiskā tīkla membrānām un tiek iesaiņoti membrānas pūslīšos, kas veidojas ap tām, kas pēc tam tiek atdalīti no ER. Šīs pūslīši, saplacinot un sakraujoties viens virs otra, līdzīgi kā saliktas pankūkas, veido raksturīgu struktūru, ko sauc par Golgi kompleksu jeb Golgi aparātu. Uzturoties Golgi aparātā, olbaltumvielās notiek noteiktas izmaiņas. Kad pienāk laiks tām iziet no šūnas, membrānas pūslīši saplūst ar šūnas membrānu un tiek iztukšoti, izlejot to saturu, t.i., sekrēcija notiek eksocitozes ceļā.
Golgi aparāts ražo arī lizosomas – membrānas maisiņus, kas satur gremošanas enzīmus. Viena no aizraujošākajām mūsdienu citoloģijas nozarēm ir noskaidrot, kā šūna ražo, iepako un eksportē noteiktas olbaltumvielas un kā tā “zina”, kuras olbaltumvielas tai vajadzētu paturēt sev.
Jebkuras šūnas membrānas pastāvīgi pārvietojas un mainās. ER membrānas lēnām pārvietojas visā šūnā. Atsevišķas šo membrānu sekcijas atdalās un veido pūslīšus, kas uz laiku kļūst par daļu no Golgi aparāta, un pēc tam eksocitozes procesā saplūst ar šūnu membrānu.
Vēlāk membrānas materiāls tiek atgriezts citoplazmā, kur to atkal izmanto.
Vielu apmaiņa, kas nonāk šūnā vai izdalās no tās ārpusē, kā arī dažādu signālu apmaiņa ar mikro- un makrovidi notiek caur šūnas ārējo membrānu. Kā zināms, šūnu membrāna ir lipīdu divslānis, kurā ir iestrādātas dažādas olbaltumvielu molekulas, kas darbojas kā specializēti receptori, jonu kanāli, ierīces, kas aktīvi transportē vai noņem dažādas ķīmiskas vielas, starpšūnu kontakti utt. Veselās eikariotu šūnās fosfolipīdi tiek izplatīti membrāna asimetriski: ārējā virsma sastāv no sfingomielīna un fosfatidilholīna, iekšējā virsma - no fosfatidilserīna un fosfatidiletanolamīna. Lai saglabātu šādu asimetriju, ir nepieciešami enerģijas izdevumi. Tāpēc šūnu bojājumu, infekcijas vai enerģijas bada gadījumā membrānas ārējā virsma tiek bagātināta ar tai neparastiem fosfolipīdiem, kas kļūst par signālu citām šūnām un fermentiem par šūnu bojājumiem ar atbilstošu reakciju uz to. Nozīmīgākā loma ir šķīstošajai fosfolipāzes A2 formai, kas noārda arahidonskābi un veido lizoformas no iepriekšminētajiem fosfolipīdiem. Arahidonskābe ir ierobežojošā saikne iekaisuma mediatoru, piemēram, eikozanoīdu, radīšanai, un aizsargājošās molekulas - pentraksīni (C-reaktīvais proteīns (CRP), amiloīda proteīnu prekursori) - tiek piesaistīti lizoformām membrānā, kam seko komplementa aktivācija. sistēma pa klasisko ceļu un šūnu iznīcināšanu.
Membrānas struktūra palīdz saglabāt šūnas iekšējās vides īpašības, tās atšķirības no ārējās vides. To nodrošina šūnu membrānas selektīvā caurlaidība un aktīvo transporta mehānismu esamība tajā. To izjaukšana tiešu bojājumu rezultātā, piemēram, ar tetrodotoksīnu, ouabaīnu, tetraetilamoniju vai nepietiekamas enerģijas padeves gadījumā attiecīgajiem “sūkņiem” izraisa šūnas elektrolītu sastāva traucējumus, vielmaiņas izmaiņas, traucējumus. specifisku funkciju - kontrakcijas, ierosmes impulsu vadīšanas utt.. Šūnu jonu kanālu (kalcija, nātrija, kālija un hlorīda) darbības traucējumus cilvēkiem var ģenētiski noteikt arī par šo kanālu struktūru atbildīgo gēnu mutācijas. Tā sauktās kanopātijas izraisa iedzimtas nervu, muskuļu un gremošanas sistēmas slimības. Pārmērīga ūdens iekļūšana šūnā var izraisīt tās plīsumu – citolīzi – membrānas perforācijas dēļ, kad tiek aktivizēts komplements, vai citotoksisku limfocītu un dabisko killer šūnu uzbrukumu.
Šūnu membrānā ir iebūvēti daudzi receptori – struktūras, kas, apvienojoties ar attiecīgajām specifiskajām signalizācijas molekulām (ligandiem), pārraida signālu šūnas iekšienē. Tas notiek caur dažādām regulējošām kaskādēm, kas sastāv no fermentatīvi aktīvām molekulām, kuras tiek secīgi aktivizētas un galu galā veicina dažādu šūnu programmu īstenošanu, piemēram, augšanu un proliferāciju, diferenciāciju, kustīgumu, novecošanos un šūnu nāvi. Normatīvo kaskādes ir diezgan daudz, taču to skaits vēl nav pilnībā noteikts. Ar tiem saistītā receptoru un regulējošo kaskāžu sistēma pastāv arī šūnas iekšienē; tie veido specifisku regulējošo tīklu ar koncentrācijas punktiem, sadali un tālākā signāla ceļa izvēli atkarībā no šūnas funkcionālā stāvokļa, attīstības stadijas un vienlaicīgas citu receptoru signālu darbības. Rezultāts var būt signāla kavēšana vai stiprināšana, novirzot to pa citu regulēšanas ceļu. Gan receptoru aparāts, gan signāla transdukcijas ceļi caur regulējošām kaskādēm, piemēram, uz kodolu, var tikt traucēti ģenētiska defekta rezultātā, kas rodas kā iedzimts defekts organisma līmenī vai somatiskas mutācijas dēļ konkrētajā šūnu tipā. Šos mehānismus var sabojāt infekcijas izraisītāji, toksīni, kā arī mainīties novecošanās procesā. Pēdējais posms var būt šūnas funkciju, tās proliferācijas un diferenciācijas procesu pārkāpums.
Uz šūnu virsmas atrodas arī molekulas, kurām ir svarīga loma starpšūnu mijiedarbības procesos. Tie var ietvert šūnu adhēzijas proteīnus, histokompatibilitātes antigēnus, audu specifiskus, diferencējošus antigēnus utt. Izmaiņas šo molekulu sastāvā izraisa starpšūnu mijiedarbības traucējumus un var izraisīt atbilstošu mehānismu aktivizēšanos šādu šūnu eliminācijai, jo tās rada zināmas briesmas organisma integritātei kā infekcijas, īpaši vīrusu, rezervuāram vai kā potenciāliem audzēja augšanas ierosinātājiem.
Šūnas enerģijas piegādes pārkāpums
Enerģijas avots šūnā ir pārtika, pēc kuras sadalīšanās enerģija tiek atbrīvota gala vielās. Galvenā enerģijas ieguves vieta ir mitohondriji, kuros vielas oksidējas ar elpošanas ķēdes enzīmu palīdzību. Oksidācija ir galvenais enerģijas piegādātājs, jo glikolīzes rezultātā no tāda paša daudzuma oksidācijas substrātu (glikozes) izdalās ne vairāk kā 5% enerģijas, salīdzinot ar oksidēšanos. Apmēram 60% no oksidācijas laikā atbrīvotās enerģijas tiek uzkrāta oksidatīvās fosforilēšanās rezultātā augstas enerģijas fosfātos (ATP, kreatīna fosfāts), pārējā daļa tiek izkliedēta kā siltums. Pēc tam šūna izmanto augstas enerģijas fosfātus tādos procesos kā sūkņa darbība, sintēze, dalīšanās, kustība, sekrēcija utt. Ir trīs mehānismi, kuru bojājumi var izraisīt šūnas enerģijas piegādes traucējumus: pirmais ir enerģijas metabolisma enzīmu sintēzes mehānisms, otrais ir oksidatīvās fosforilācijas mehānisms, trešais ir enerģijas izmantošanas mehānisms.
Elektronu transporta pārrāvums mitohondriju elpošanas ķēdē vai ADP oksidācijas un fosforilēšanās atvienošana ar protonu potenciāla zudumu, kas ir ATP ģenerācijas dzinējspēks, noved pie oksidatīvās fosforilācijas pavājināšanās tādā veidā, ka lielākā daļa enerģijas tiek izkliedēta kā siltums un samazinās augstas enerģijas savienojumu skaits. Oksidācijas un fosforilēšanās atsaistīšanu adrenalīna ietekmē homeotermisko organismu šūnas izmanto, lai palielinātu siltuma ražošanu, vienlaikus saglabājot nemainīgu ķermeņa temperatūru dzesēšanas laikā vai paaugstinot to drudža laikā. Tireotoksikozes gadījumā tiek novērotas būtiskas izmaiņas mitohondriju struktūrā un enerģijas metabolismā. Šīs izmaiņas sākotnēji ir atgriezeniskas, bet pēc noteikta brīža kļūst neatgriezeniskas: mitohondriji sadalās, sadalās vai uzbriest, zaudē kristas, pārvēršoties vakuolās, un galu galā uzkrājas tādas vielas kā hialīns, feritīns, kalcijs, lipofuscīns. Pacientiem ar skorbutu mitohondriji saplūst, veidojot hondriosfēras, iespējams, peroksīda savienojumu izraisītu membrānu bojājumu dēļ. Nozīmīgs mitohondriju bojājums rodas jonizējošā starojuma ietekmē normālas šūnas pārveidošanās laikā par ļaundabīgu.
Mitohondriji ir spēcīgs kalcija jonu depo, kur tā koncentrācija ir par vairākām kārtām augstāka nekā citoplazmā. Kad mitohondriji ir bojāti, kalcijs nonāk citoplazmā, izraisot proteināžu aktivāciju ar intracelulāro struktūru bojājumiem un attiecīgās šūnas funkciju traucējumiem, piemēram, kalcija kontraktūras vai pat “kalcija nāvi” neironos. Mitohondriju funkcionālās kapacitātes traucējumu rezultātā strauji palielinās brīvo radikāļu peroksīdu savienojumu veidošanās, kuriem ir ļoti augsta reaktivitāte un tāpēc tie bojā svarīgas šūnas sastāvdaļas - nukleīnskābes, olbaltumvielas un lipīdus. Šī parādība tiek novērota tā sauktā oksidatīvā stresa laikā, un tai var būt negatīvas sekas uz šūnas pastāvēšanu. Tādējādi mitohondriju ārējās membrānas bojājumus pavada starpmembrānu telpā esošo vielu, galvenokārt citohroma C un dažu citu bioloģiski aktīvo vielu izdalīšanās citoplazmā, kas izraisa ķēdes reakcijas, kas izraisa ieprogrammētu šūnu nāvi - apoptozi. Bojājot mitohondriju DNS, brīvo radikāļu reakcijas izkropļo ģenētisko informāciju, kas nepieciešama noteiktu elpošanas ķēdes enzīmu veidošanai, kas tiek ražoti tieši mitohondrijās. Tas noved pie vēl lielākiem oksidatīvo procesu traucējumiem. Kopumā pašu mitohondriju ģenētiskais aparāts, salīdzinot ar kodola ģenētisko aparātu, ir mazāk aizsargāts no kaitīgām ietekmēm, kas var mainīt tajā iekodēto ģenētisko informāciju. Rezultātā mitohondriju disfunkcija notiek visu mūžu, piemēram, novecošanās procesā, šūnas ļaundabīgās transformācijas laikā, kā arī uz iedzimtu mitohondriju slimību fona, kas saistītas ar mitohondriju DNS mutāciju olšūnā. Pašlaik ir aprakstītas vairāk nekā 50 mitohondriju mutācijas, kas izraisa iedzimtas nervu un muskuļu sistēmas deģeneratīvas slimības. Tie tiek nodoti bērnam tikai no mātes, jo spermas mitohondriji nav daļa no zigotas un attiecīgi jaunā organisma.
Ģenētiskās informācijas saglabāšanas un pārsūtīšanas pārkāpums
Šūnas kodols satur lielāko daļu ģenētiskās informācijas un tādējādi nodrošina tā normālu darbību. Izmantojot selektīvu gēnu ekspresiju, tas koordinē šūnu aktivitāti starpfāzes laikā, saglabā ģenētisko informāciju un atjauno un pārraida ģenētisko materiālu šūnu dalīšanās laikā. Kodolā notiek DNS replikācija un RNS transkripcija. Dažādi patogēni faktori, piemēram, ultravioletais un jonizējošais starojums, brīvo radikāļu oksidēšanās, ķīmiskās vielas, vīrusi, var bojāt DNS. Tiek lēsts, ka katrai siltasiņu dzīvnieka šūnai nepieciešama 1 diena. zaudē vairāk nekā 10 000 bāzes. Šeit mums vajadzētu pievienot pārkāpumus kopēšanas laikā sadalīšanas laikā. Ja šie bojājumi saglabātos, šūna nespētu izdzīvot. Aizsardzība slēpjas spēcīgu remontsistēmu, piemēram, ultravioleto endonukleāzes, labošanas replikācijas un rekombinācijas labošanas sistēmu esamībā, kas aizstāj DNS bojājumus. Remontsistēmu ģenētiskie defekti izraisa slimību attīstību, ko izraisa paaugstināta jutība pret faktoriem, kas bojā DNS. Tas ir xeroderma pigmentosum, kā arī daži paātrinātas novecošanas sindromi, ko papildina paaugstināta tendence attīstīties ļaundabīgiem audzējiem.
Sistēma DNS replikācijas, Messenger RNS (mRNS) transkripcijas un ģenētiskās informācijas pārveidošanai no nukleīnskābēm olbaltumvielu struktūrā regulēšanas sistēma ir diezgan sarežģīta un daudzlīmeņu. Papildus regulējošajām kaskādēm, kas izraisa transkripcijas faktoru darbību, kuru kopējais skaits pārsniedz 3000, kas aktivizē noteiktus gēnus, pastāv arī daudzlīmeņu regulēšanas sistēma, ko mediē mazas RNS molekulas (traucējošā RNS; RNAi). Cilvēka genoms, kas sastāv no aptuveni 3 miljardiem purīna un pirimidīna bāzu, satur tikai 2% no strukturālajiem gēniem, kas ir atbildīgi par olbaltumvielu sintēzi. Pārējie nodrošina regulējošo RNS sintēzi, kas vienlaikus ar transkripcijas faktoriem aktivizē vai bloķē strukturālo gēnu darbu DNS līmenī hromosomās vai ietekmē kurjerRNS (mRNS) translācijas procesus polipeptīda molekulas veidošanās laikā. citoplazma. Ģenētiskās informācijas pārkāpums var notikt gan strukturālo gēnu, gan DNS regulējošās daļas līmenī ar atbilstošām izpausmēm dažādu iedzimtu slimību veidā.
Pēdējā laikā liela uzmanība tiek pievērsta izmaiņām ģenētiskajā materiālā, kas notiek organisma individuālās attīstības laikā un ir saistītas ar noteiktu DNS sekciju un hromosomu inhibīciju vai aktivāciju to metilēšanas, acetilēšanas un fosforilēšanās dēļ. Šīs izmaiņas saglabājas ilgu laiku, dažreiz visā organisma dzīves laikā no embrioģenēzes līdz vecumam, un tās sauc par epigenomisko iedzimtību.
Šūnu ar izmainītu ģenētisko informāciju proliferāciju novērš arī sistēmas (faktori), kas kontrolē mitotisko ciklu. Tie mijiedarbojas ar no ciklīna atkarīgām proteīnkināzēm un to katalītiskajām apakšvienībām – ciklīniem – un bloķē šūnu iziet cauri pilnam mitotiskajam ciklam, apturot dalīšanos uz robežas starp presintētisko un sintētisko fāzi (G1/S bloks), līdz DNS remonts ir pabeigts, un, ja tas nav iespējams, viņi ierosina ieprogrammētas nāves šūnas. Šie faktori ietver p53 gēnu, kura mutācijas rezultātā tiek zaudēta kontrole pār transformēto šūnu proliferāciju; to novēro gandrīz 50% cilvēku vēža gadījumu. Otrais mitotiskā cikla kontrolpunkts atrodas pie G2/M robežas. Šeit pareizu hromosomu materiāla sadalījumu starp meitas šūnām mitozē vai mejozē kontrolē, izmantojot mehānismu kopumu, kas kontrolē šūnu vārpstu, centru un centromērus (kinetohorus). Šo mehānismu neefektivitāte izraisa hromosomu vai to daļu sadalījuma traucējumus, kas izpaužas kā hromosomu neesamība vienā no meitas šūnām (aneuploīdija), papildu hromosomas klātbūtne (poliploīdija), hromosomu atdalīšanās. hromosomas daļa (delecija) un tās pārnešana uz citu hromosomu (translokācija) . Šādi procesi ļoti bieži tiek novēroti ļaundabīgi deģenerētu un transformētu šūnu proliferācijas laikā. Ja tas notiek mejozes laikā ar dzimumšūnām, tas noved pie augļa nāves agrīnā embrija attīstības stadijā vai organisma ar hromosomu slimību piedzimšanas.
Nekontrolēta šūnu proliferācija audzēja augšanas laikā notiek gēnu mutāciju rezultātā, kas kontrolē šūnu proliferāciju un tiek saukti par onkogēniem. No vairāk nekā 70 pašlaik zināmajiem onkogēniem lielākā daļa pieder pie šūnu augšanas regulēšanas komponentiem, dažus pārstāv transkripcijas faktori, kas regulē gēnu aktivitāti, kā arī faktori, kas kavē šūnu dalīšanos un augšanu. Vēl viens faktors, kas ierobežo proliferējošo šūnu pārmērīgu paplašināšanos (izplatīšanos), ir hromosomu galu saīsināšanās - telomēri, kuri tīri steriskas mijiedarbības rezultātā nespēj pilnībā vairoties, tāpēc pēc katras šūnu dalīšanās telomēri tiek saīsināti par noteikta bāzu daļa. Tādējādi pieauguša organisma proliferējošās šūnas pēc noteikta dalīšanās skaita (parasti no 20 līdz 100 atkarībā no organisma veida un vecuma) izsmeļ telomēra garumu un tālākā hromosomu replikācija apstājas. Šī parādība nenotiek spermatozoīdu epitēlijā, enterocītos un embrionālajās šūnās enzīma telomerāzes klātbūtnes dēļ, kas atjauno telomēra garumu pēc katras dalīšanās. Lielākajā daļā pieaugušo organismu šūnu telomerāze ir bloķēta, bet diemžēl tā tiek aktivizēta audzēja šūnās.
Savienojums starp kodolu un citoplazmu un vielu transportēšana abos virzienos tiek veikta caur porām kodola membrānā, piedaloties īpašām transporta sistēmām, kas patērē enerģiju. Tādā veidā uz kodolu tiek transportētas enerģijas un plastmasas vielas, signālmolekulas (transkripcijas faktori). Apgrieztā plūsma citoplazmā ienes mRNS molekulas un pārnes RNS (tRNS), ribosomas, kas nepieciešamas olbaltumvielu sintēzei šūnā. Tas pats vielu transportēšanas ceļš ir raksturīgs arī vīrusiem, jo īpaši tādiem kā HIV. Viņi pārnes savu ģenētisko materiālu saimniekšūnas kodolā, to tālāk iekļaujot saimnieka genomā un jaunizveidotās vīrusa RNS pārnesot citoplazmā, lai turpinātu jaunu vīrusu daļiņu proteīnu sintēzi.
Sintēzes procesu pārkāpums
Olbaltumvielu sintēzes procesi notiek endoplazmatiskā tīkla cisternās, kas ir cieši saistītas ar porām kodola membrānā, caur kurām endoplazmatiskajā retikulā nonāk ribosomas, tRNS un mRNS. Šeit tiek veikta polipeptīdu ķēžu sintēze, kas pēc tam iegūst savu galīgo formu agranulārajā endoplazmatiskajā retikulā un lamelārajā kompleksā (Golgi kompleksā), kur tās tiek pakļautas pēctranslācijas modifikācijām un apvienojas ar ogļhidrātu un lipīdu molekulām. Jaunizveidotās olbaltumvielu molekulas nepaliek sintēzes vietā, bet gan sarežģītā regulētā procesā, ko sauc proteīnkinēze, tiek aktīvi pārvietoti uz to izolēto šūnas daļu, kur tie pildīs paredzēto funkciju. Šajā gadījumā ļoti svarīgs solis ir pārnestās molekulas strukturēšana atbilstošā telpiskā konfigurācijā, kas spēj veikt tai raksturīgo funkciju. Šī strukturēšana notiek ar īpašu enzīmu palīdzību vai uz specializētu proteīna molekulu matricas - šaperoniem, kas palīdz jaunizveidotajai vai ārējās ietekmes rezultātā mainītajai proteīna molekulai iegūt pareizu trīsdimensiju struktūru. Nelabvēlīgas ietekmes uz šūnu gadījumā, kad ir iespējami proteīna molekulu struktūras traucējumi (piemēram, ar ķermeņa temperatūras paaugstināšanos, infekcijas procesu, intoksikāciju), palielinās chaperonu koncentrācija šūnā. asi. Tāpēc šādas molekulas sauc arī par stresa proteīni, vai karstuma šoka proteīni. Proteīna molekulas strukturēšanas pārkāpums izraisa ķīmiski inertu konglomerātu veidošanos, kas nogulsnējas šūnā vai ārpus tās amiloidozes, Alcheimera slimības uc laikā. Dažreiz iepriekš strukturēta līdzīga molekula var kalpot kā matrica, un šajā gadījumā. Gadījumā, ja primārā strukturēšana notiek nepareizi, visas nākamās molekulas arī būs bojātas. Šāda situācija rodas tā saukto prionu slimību gadījumā (aitu skrepi slimība, niknās govis, kuru, Kreicfelda-Jakoba slimība cilvēkiem), kad kāda no nervu šūnas membrānas olbaltumvielām defekts izraisa sekojošu inertu masu uzkrāšanos šūnas iekšienē. un tās dzīvībai svarīgo funkciju traucējumi.
Sintēzes procesu traucējumi šūnā var notikt dažādos tās posmos: RNS transkripcija kodolā, polipeptīdu translācija ribosomās, pēctranslācijas modifikācija, smilškrāsas molekulas hipermetilēšana un glikozilācija, olbaltumvielu transportēšana un sadale šūnā un to izvadīšana. uz ārpusi. Šajā gadījumā var novērot ribosomu skaita palielināšanos vai samazināšanos, poliribosomu sadalīšanos, granulētā endoplazmatiskā retikuluma cisternu paplašināšanos, ribosomu zudumu, pūslīšu un vakuolu veidošanos. Tādējādi, saindējoties ar bālu grebu, tiek bojāts RNS polimerāzes enzīms, kas izjauc transkripciju. Difterijas toksīns, inaktivējot pagarinājuma faktoru, izjauc translācijas procesus, izraisot miokarda bojājumus. Dažu specifisku olbaltumvielu molekulu sintēzes traucējumu cēlonis var būt infekcijas izraisītāji. Piemēram, herpes vīrusi kavē MHC antigēnu molekulu sintēzi un ekspresiju, kas ļauj tiem daļēji izvairīties no mēra baciļu imūnās kontroles – akūtu iekaisumu mediatoru sintēzes. Neparastu proteīnu parādīšanās var apturēt to turpmāku sadalīšanos un izraisīt inerta vai pat toksiska materiāla uzkrāšanos. To zināmā mērā var veicināt sabrukšanas procesu traucējumi.
Sabrukšanas procesu traucējumi
Vienlaikus ar olbaltumvielu sintēzi šūnā nepārtraukti notiek tā sadalīšanās. Normālos apstākļos tam ir svarīga regulējoša un veidojoša nozīme, piemēram, aktivizējot neaktīvas enzīmu formas, proteīna hormonus un mitotiskā cikla proteīnus. Normālai šūnu augšanai un attīstībai ir nepieciešams precīzi kontrolēts līdzsvars starp proteīnu un organellu sintēzi un sadalīšanos. Tomēr olbaltumvielu sintēzes procesā sintezējošā aparāta darbības kļūdu, proteīna molekulas patoloģiskas strukturēšanas un ķīmisko un baktēriju izraisītu bojājumu dēļ pastāvīgi veidojas diezgan liels skaits bojātu molekulu. Saskaņā ar dažām aplēsēm to daļa ir aptuveni trešā daļa no visiem sintezētajiem proteīniem.
Zīdītāju šūnām ir vairākas galvenās proteīnu iznīcināšanas veidi: caur lizosomu proteāzēm (pentīdu hidrolāzes), no kalcija atkarīgām proteināzēm (endopeptidāzes) un proteasomu sistēmu. Turklāt ir arī specializētas proteināzes, piemēram, kaspāzes. Galvenā organelle, kurā notiek vielu sadalīšanās eikariotu šūnās, ir lizosoma, kas satur daudzus hidrolītiskos enzīmus. Sakarā ar endocitozes un dažāda veida autofagijas procesiem lizosomās un fagolizosomās tiek iznīcinātas gan defektīvās olbaltumvielu molekulas, gan veseli organoīdi: bojāti mitohondriji, plazmas membrānas sekcijas, daži ārpusšūnu proteīni un sekrēcijas granulu saturs.
Svarīgs proteīnu sadalīšanās mehānisms ir proteasoma, sarežģītas struktūras multikatalītiskā proteināzes struktūra, kas lokalizēta citozolā, kodolā, endoplazmatiskajā retikulumā un uz šūnu membrānas. Šī enzīmu sistēma ir atbildīga par bojāto proteīnu, kā arī veselīgu olbaltumvielu sadalīšanu, kas ir jānoņem normālai šūnu darbībai. Šajā gadījumā iznīcināmās olbaltumvielas sākotnēji tiek kombinētas ar specifisku polipeptīdu, ubikvitīnu. Tomēr proteasomās var daļēji iznīcināt arī neubikvitinātos proteīnus. Olbaltumvielu molekulu sadalīšana proteasomās īsos polipeptīdos (apstrāde) ar to sekojošu prezentāciju kopā ar I tipa MHC molekulām ir svarīga saikne antigēnu homeostāzes imūnā kontrolē organismā. Kad proteasomu funkcija ir novājināta, uzkrājas bojāti un nevajadzīgi proteīni, kas pavada šūnu novecošanos. Ciklinatkarīgo proteīnu sadalīšanās pārkāpums izraisa šūnu dalīšanās traucējumus, sekrēcijas proteīnu degradāciju - cistofibrozes attīstību. Un otrādi, proteasomu funkcijas palielināšanās pavada ķermeņa noplicināšanos (AIDS, vēzis).
Ar ģenētiski noteiktiem olbaltumvielu sadalīšanās traucējumiem organisms nav dzīvotspējīgs un iet bojā agrīnās embrioģenēzes stadijās. Ja tiek traucēta tauku vai ogļhidrātu sadalīšanās, rodas uzkrāšanās slimības (tezaurismoze). Šādā gadījumā šūnas iekšienē uzkrājas pārmērīgs daudzums noteiktu vielu vai to nepilnīgas sadalīšanās produktu - lipīdu, polisaharīdu, kas būtiski bojā šūnas darbību. Visbiežāk tas tiek novērots aknu epitēlija šūnās (hepatocītos), neironos, fibroblastos un makrofagocītos.
Iegūtie vielu sadalīšanās procesu traucējumi var rasties patoloģisku procesu rezultātā (piemēram, olbaltumvielu, tauku, ogļhidrātu un pigmentu deģenerācija), un tos pavada neparastu vielu veidošanās. Lizosomu proteolīzes sistēmas traucējumi izraisa adaptācijas samazināšanos badošanās vai paaugstināta stresa laikā, kā arī noteiktu endokrīno disfunkciju rašanos - insulīna, tiroglobulīna, citokīnu un to receptoru līmeņa pazemināšanos. Proteīna degradācijas traucējumi palēnina brūču dzīšanas ātrumu, izraisa aterosklerozes attīstību un ietekmē imūnreakciju. Ar hipoksiju, intracelulārā pH izmaiņām, radiācijas bojājumiem, kam raksturīga paaugstināta membrānas lipīdu peroksidācija, kā arī lizosomotropo vielu ietekmē - baktēriju endotoksīni, toksisko sēnīšu metabolīti (sporofusarīns), silīcija oksīda kristāli - lizosomu membrānas stabilitāte. izmaiņu rezultātā citoplazmā izdalās aktivētie lizosomu enzīmi, kas izraisa šūnu struktūru iznīcināšanu un to nāvi.
1. nodaļa
ŠŪNU FIZIOLOĢIJAS PAMATI
I. Dudel
Plazmas membrāna . Dzīvnieku šūnas ierobežo plazmas membrāna (1.1. attēls). Mēs pakavēsimies pie tā struktūras, kas ir ļoti līdzīga daudzu intracelulāro membrānu struktūrai, nedaudz sīkāk. Membrānas galvenā matrica sastāv no lipīdi, galvenokārt fosfatidilholīns. Šie lipīdi sastāv no hidrofilas galvas grupas, kurai pievienotas garas hidrofobās ogļūdeņražu ķēdes. Ūdenī šādi lipīdi spontāni veido 4–5 nm biezu divslāņu plēvi, kurā hidrofilās grupas ir vērstas pret ūdens vidi, bet hidrofobās ogļūdeņražu ķēdes ir sakārtotas divās rindās, veidojot bezūdens lipīdu fāzi. Šūnu membrānas ir šāda veida lipīdu divslāņi un satur glikolipīdus, holesterīnu un fosfolipīdus (1.2. att.). Glikolipīdu hidrofilo daļu veido oligosaharīdi. Glikolipīdi vienmēr atrodas uz plazmas membrānas ārējās virsmas, un molekulas oligosaharīda daļa ir orientēta kā vidē iegremdēts mats. Izkaisītas starp fosfolipīdiem gandrīz vienādos daudzumos, holesterīna molekulas stabilizē membrānu. Dažādu lipīdu sadalījums membrānas iekšējā un ārējā slānī nav vienāds, un pat viena slāņa ietvaros ir zonas, kurās koncentrējas noteikta veida lipīdi. Šis nevienmērīgais sadalījums
Rīsi. 1.1. Šūnas shematisks zīmējums, kurā parādīti svarīgākie organoīdi
iespējams, tam ir kāda, pagaidām neskaidra, funkcionāla nozīme.
Galvenie funkcionālie elementi, kas iestrādāti relatīvi inertā membrānas lipīdu matricā, ir vāveres(1.2. att.). Proteīns dažādās membrānās veido no 25 līdz 75% svara, bet, tā kā olbaltumvielu molekulas ir daudz lielākas nekā lipīdu molekulas, 50% pēc svara ir līdzvērtīgi 1 proteīna molekulas attiecībai pret 50 lipīdu molekulām. Daži proteīni iekļūst membrānā no tās ārējās uz iekšējo virsmu, bet citi ir fiksēti vienā slānī. Olbaltumvielu molekulas parasti ir orientētas tā, ka to hidrofobās grupas ir iestrādātas lipīdu membrānā un polārās hidrofilās grupas uz membrānas virsmas ir iestrādātas ūdens fāzē. Daudzi proteīni uz membrānas ārējās virsmas ir glikoproteīni; to hidrofilās saharīdu grupas saskaras ar ārpusšūnu vidi.
Intracelulāro organellu membrānu sistēmas .
Apmēram pusi no šūnu tilpuma aizņem organoīdi, kas izolēti no citozola ar membrānām. Kopējais intracelulāro organellu membrānu virsmas laukums ir vismaz 10 reizes lielāks par plazmas membrānas virsmu. Visplašāk izmantotā membrānas sistēma ir Endoplazmatiskais tīkls, pārstāv tīklu

Rīsi. 1.2.Plazmas membrānas shematisks attēlojums. Proteīni ir iestrādāti fosfolipīdu divslānī, daži no tiem iekļūst divslānī, bet citi ir tikai noenkuroti pie ārējā vai iekšējā slāņa.
ļoti izliektas caurules vai maisiņiem līdzīgas iegarenas struktūras; lieli endoplazmatiskā tīkla laukumi ir izraibināti ar ribosomām; šādu tīklu sauc par graudainu vai raupju (1.1. att.). Golgi aparāts sastāv arī no membrānām saistītām lamelēm, no kurām atdalās pūslīši jeb pūslīši (1.1. att.). Lizosomas un peroksisomas- Tie ir mazi specializēti pūslīši. Visās šajās dažādajās organellās membrāna un tās aptvertā telpa satur īpašus enzīmu komplektus; Organellu iekšpusē uzkrājas īpaši vielmaiņas produkti, ko izmanto dažādu organellu funkciju veikšanai.
KodolsUn mitohondriji atšķiras ar to, ka katru no šīm organellām ieskauj divas membrānas. Kodols ir atbildīgs par metabolisma kinētisko kontroli; salocītā mitohondriju iekšējā membrāna ir oksidatīvā metabolisma vieta; šeit piruvāta vai taukskābju oksidēšanās dēļ tiek sintezēts augstas enerģijas savienojums adenozīna trifosfāts (ATP jeb ATP).
Citoskelets . Citoplazmu, kas ieskauj organellus, nekādā veidā nevar uzskatīt par amorfu; to caurstrāvo citoskeleta tīkls. Citoskelets sastāv no mikrotubulām, aktīna pavedieniem un starppavedieniem (1.1. att.). Mikrotubulasārējais diametrs ir aptuveni 25 nm; tie veidojas, tāpat kā parasts polimērs, tubulīna proteīna molekulu montāžas rezultātā. Aktīna pavedieni - kontraktilās šķiedras, kas atrodas gandrīz membrānas slānī un visā šūnā, galvenokārt piedalās procesos, kas saistīti ar kustību. Starpposma pavedieni sastāv no dažāda ķīmiskā sastāva blokiem dažāda veida šūnās; tie veido dažādus savienojumus starp diviem citiem iepriekš minētajiem citoskeleta elementiem. Organelli un plazmas membrāna ir saistītas arī ar citoskeletu, kas ne tikai uztur šūnas formu un organellu stāvokli tajā, bet arī nosaka izmaiņas šūnas formā un tās kustīgumā.
Citozols . Apmēram pusi no šūnu tilpuma aizņem citozols. Tā kā tajā ir aptuveni 20% olbaltumvielu, tas ir vairāk želeja nekā ūdens šķīdums. Mazas molekulas, ieskaitot organiskās un neorganiskās joni, izšķīdināts ūdens fāzē. Starp šūnu un vidi notiek jonu apmaiņa (ārpusšūnu telpa); šie vielmaiņas procesi tiks apspriesti nākamajā sadaļā. Jonu koncentrācija ārpusšūnu telpā tiek uzturēta ar ievērojamu precizitāti nemainīgā līmenī; arī katra jona intracelulārajai koncentrācijai ir noteikts līmenis, kas atšķiras no ārpus šūnas esošās koncentrācijas (1.1. tabula). Ekstracelulārajā vidē visizplatītākais katjons ir Na+ šūnā tā koncentrācija ir vairāk nekā 10 reizes zemāka. Gluži pretēji, K + koncentrācija ir visaugstākā šūnas iekšpusē, tā ir mazāka par vairāk nekā vienu pakāpi. Vislielākais gradients starp ekstracelulāro un intracelulāro koncentrāciju pastāv Ca 2+, brīvo jonu koncentrācija šūnā ir vismaz 10 000 reižu zemāka nekā ārpus tās. Ne visi joni ir izšķīdināti citozolā, daži no tiem adsorbējas uz olbaltumvielām vai nogulsnējas organellās. Piemēram, Ca 2+ gadījumā saistīto jonu ir daudz vairāk nekā brīvo. Lielākā daļa citozola proteīnu ir fermenti, ar kuru līdzdalību tiek veikti daudzi starpposma metabolisma procesi: glikolīze un glikoneoģenēze, aminoskābju sintēze vai iznīcināšana, proteīnu sintēze uz ribosomām (1.1. att.). Citozols satur arī lipīdu pilienus un glikogēna granulas, kas kalpo kā svarīgu molekulu rezerves.
1.1. tabula.Intra- un ārpusšūnu jonu koncentrācija homeotermisko dzīvnieku muskuļu šūnās. A – - "augstas molekulmasas šūnu anjoni"
|
Intracelulārā koncentrācija |
Ārpusšūnu koncentrācija |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca 2+ |
10 –7 10 –8 mmol/l |
Ca 2+ |
2 mmol/l |
|
C l – |
4 mmol/l |
C1 – |
120 mmol/l |
|
HCO 3 – |
8 mmol/l |
HCO 3 – |
27 mmol/l |
|
A – |
155 mmol/l |
Citi katjoni |
5 mmol/l |
|
Potenciāls miera stāvoklī -90 mV |
|||
1.2. Vielu apmaiņa starp šūnu un vidi
Mēs esam īsi aprakstījuši šūnas struktūru, lai mēs varētu izmantot šo aprakstu, lai pārskatītu šūnu fizioloģijas pamatus. Šūnu nekādā gadījumā nevar uzskatīt par statisku vienību, jo notiek pastāvīga vielu apmaiņa starp dažādiem intracelulāriem nodalījumiem, kā arī starp nodalījumiem un vidi. Šūnu struktūras atrodas dinamiskā līdzsvarā, un šūnu mijiedarbība savā starpā un ar ārējo vidi ir nepieciešams nosacījums funkcionējoša organisma dzīvības uzturēšanai. Šajā nodaļā aplūkosim šādas apmaiņas pamatmehānismus. Turpmākajās nodaļās šie mehānismi tiks aplūkoti, jo tie attiecas uz nervu šūnu un tās funkcijām;
tomēr tie paši mehānismi ir visu pārējo orgānu darbības pamatā.
Difūzija.Vienkāršākais vielas pārvietošanas process ir difūzija. Šķīdumos (vai gāzēs) atomi un molekulas pārvietojas brīvi, un koncentrāciju atšķirības tiek līdzsvarotas difūzijas dēļ. Aplūkosim divus tilpumus, kas pildīti ar šķidrumu vai gāzi (1.3. att.), kuros vielām ir koncentrācijas c 1 un c 2 un atdalītas ar slāni ar virsmas laukumu A un biezumu d. Vielas plūsma m laikā t aprakstīts Fika pirmais difūzijas likums:
dm/ dt= DA/ d ( C 1 –С 2)=DA/ dD C(1)
kur D ir difūzijas koeficients, konstants konkrētai vielai, šķīdinātājam un temperatūrai. Vispārīgākā formā koncentrācijas starpībai dc attālumā dx
dm/dt= –D A dc/dx, (2)
plūsma caur A sekciju ir proporcionāla koncentrācijas gradientam dc/dx . Mīnusa zīme parādās vienādojumā, jo koncentrācijas izmaiņas x virzienā ir negatīvas.
Difūzija ir vissvarīgākais process, kura dēļ lielākā daļa molekulu ūdens šķīdumos pārvietojas nelielos attālumos. Tas attiecas arī uz to kustību šūnā, ciktāl difūziju netraucē membrānas. Daudzas vielas var brīvi izkliedēties caur lipīdu membrānām, īpaši ūdens un izšķīdušās gāzes, piemēram, O 2 un CO 2 . Taukos šķīstošs

Rīsi. 1.3.Kvantitatīvās difūzijas shēma. Abas telpas atdala biezuma slānisdun apgabals A. C – liela daļiņu koncentrācija tilpuma kreisajā daļā, C: – zema daļiņu koncentrācija labajā daļas, rozā virsma– koncentrācijas gradients difūzijas slānī. Difūzijas plūsma dm/dt – sk vienādojums (1)
vielas arī labi izkliedējas caur membrānām; Tas attiecas arī uz diezgan maza izmēra polārajām molekulām, piemēram, etanolu un urīnvielu, savukārt cukuri ar grūtībām iziet cauri lipīdu slānim. Tajā pašā laikā lipīdu slāņi ir praktiski necaurlaidīgi lādētām molekulām, tostarp pat neorganiskiem joniem. Attiecībā uz neelektrolītiem difūzijas vienādojumu (1) parasti pārveido, apvienojot membrānas un izkliedējošās vielas īpašības vienā parametru caurlaidība (P):
dm/dt=P AD c.(3)
Attēlā 1.4 tiek salīdzināti lipīdu membrānas caurlaidība (P) dažādām molekulām.
Difūzija caur membrānas porām . Plazmas membrāna (un citas šūnu membrānas) ir caurlaidīga ne tikai vielām, kas izkliedējas caur lipīdu slāni, bet arī daudziem joniem, cukuriem, aminoskābēm un nukleotīdiem. Šīs vielas šķērso membrānu caur izveidotajām porām transporta proteīni, iegremdēts membrānā. Šādos proteīnos ir ar ūdeni piepildīts kanāls, kura diametrs ir mazāks par 1 nm, pa kuru var izkliedēties mazas molekulas. Tie pārvietojas pa koncentrācijas gradientu, un, ja tie nes lādiņu, tad to kustību pa kanāliem regulē arī membrānas potenciāls. Membrānas kanāli ir salīdzinoši selektīvi

Rīsi. 1.4.Mākslīgo lipīdu divslāņu caurlaidība dažādām vielām
attiecībā uz molekulu veidu, kas var iziet cauri tām, ir, piemēram, kālija, nātrija un kalcija kanāli, no kuriem katrs ir necaurlaidīgs gandrīz jebkuram jonam, izņemot konkrētu. Tādas selektivitāte To nosaka kanāla sieniņu saistīšanās vietu lādiņš vai struktūra, kas atvieglo konkrētas molekulas transportēšanu un novērš citu vielu iekļūšanu caur kanālu (att. 1,5, A) .
Aiz uzvedības membrānas jonu kanāli viegli novērot, jo strāvu, kas rodas no jonu kustības, var izmērīt pat vienam kanālam. Ir parādīts, ka kanāli spontāni un ar augstu frekvenci maina savu stāvokli no atvērta uz slēgtu. Kālija kanālu raksturo strāvas impulsi, kuru amplitūda ir aptuveni 2 pA (2 10 –12 A) un ilgums ir vairākas milisekundes (sk. 2.12. att. 37. lpp.) [3]. Šajā periodā caur to iziet desmitiem tūkstošu jonu. Olbaltumvielu pāreja no vienas konformācijas uz otru tiek pētīta, izmantojot rentgenstaru difrakciju, Mössbauer spektroskopiju un kodolmagnētisko rezonansi (NMR). Tādējādi olbaltumvielas ir ļoti dinamiskas, kustīgas struktūras, un kanāls, kas iet caur proteīnu, nav tikai stingra, ar ūdeni pildīta caurule (1.5. att., A), bet gan ātri kustīgu molekulāro grupu un lādiņu labirints. Šī kanāla dinamiskā īpašība ir atspoguļota kanāla enerģijas profils, attēlā parādīts. 1.5, B. Šeit abscisu ass attēlo kanāla garumu no ārējā šķīduma ar jonu koncentrāciju C 0 un potenciālu 0 līdz iekšējam šķīdumam ar koncentrāciju C 1 un potenciālu E. Ordinātu ass

Rīsi. 1.5.A. Shēma proteīnam, kas veido kālija kanālu, kas iestrādāts plazmas membrānas lipīdu divslānī. Uz kanāla “sienas” ir fiksēti četri negatīvi lādiņi. B. attēlā parādītā kanāla shematiskais enerģijas profils. A. Ordinātu ass parāda kanāla caurbraukšanai nepieciešamās kinētiskās enerģijas vērtības; pa abscisu asi – attālums starp membrānas iekšējo un ārējo virsmu. Enerģijas minimumi atbilst pozitīvi lādētu jonu saistīšanās vietām ar fiksētiem negatīviem lādiņiem kanāla sienā. Enerģijas maksimumi atbilst difūzijas šķēršļiem kanālā. Tiek pieņemts, ka kanāla proteīna konformācija spontāni svārstās; enerģijas profila opcijas ir parādītas ar nepārtrauktām un pārtrauktām līnijām; šīs svārstības ievērojami atvieglo jonu saistīšanos, pārvarot enerģijas barjeru (modificēts)
parādīti jonu enerģijas līmeņi kanālu saistīšanās vietās; maksimums grafikā apzīmē caurlaidības barjeru, kas jāpārsniedz jonu enerģijai, lai iekļūtu kanālā, un diagrammas “iekritums” apzīmē relatīvi stabilu stāvokli (saistīšanās). Neskatoties uz enerģijas maksimuma šķērsli, jons var iekļūt kanālā, ja enerģijas profils spontāni mainās; Tādējādi jons var pēkšņi atrasties enerģijas maksimuma “otrpus” un turpināt pārvietoties šūnā. Atkarībā no jona lādiņa, izmēra un hidratācijas pakāpes un tā spējas saistīties ar kanāla sienu konstrukcijām, kanāla enerģijas profils dažādiem joniem atšķiras, kas var izskaidrot atsevišķu kanālu veidu selektivitāti.
Jonu difūzijas līdzsvars . Dažādu jonu difūzijai caur membrānas kanāliem vajadzētu novērst koncentrāciju atšķirības starp ārpusšūnu un intracelulāro vidi. Tomēr, kā redzams no tabulas. 1.1, šādas atšķirības saglabājas, tāpēc tām ir jābūt līdzsvars starp difūziju un citiem transportēšanas procesiem cauri membrānai. Nākamās divas sadaļas attiecas uz veidiem, kā tiek sasniegts šis līdzsvars. Jonu gadījumā difūzijas līdzsvaru ietekmē to lādiņš. Nelādētu molekulu difūziju nodrošina koncentrāciju atšķirība dc , un, kad koncentrācijas ir izlīdzinātas, pats transports apstājas. Uzlādētās daļiņas papildus ietekmē elektriskais lauks. Piemēram, kad kālija jons atstāj šūnu pa koncentrācijas gradientu, tam ir viens pozitīvs lādiņš. Tādējādi intracelulārā vide kļūst negatīvāk uzlādēta, kā rezultātā membrānā rodas potenciāla atšķirība. Intracelulārais negatīvais lādiņš neļauj jauniem kālija joniem iziet no šūnas, un tie joni, kas atstāj šūnu, vēl vairāk palielinās membrānas lādiņu. Kālija jonu plūsma apstājas, kad elektriskā lauka darbība kompensē difūzijas spiedienu koncentrācijas starpības dēļ. Joni turpina iet cauri membrānai, bet vienādos daudzumos abos virzienos. Tāpēc noteiktai jonu koncentrācijas atšķirībai membrānā pastāv līdzsvara potenciāls E jonu pie kura apstājas jonu plūsma caur membrānu. Līdzsvara potenciālu var viegli noteikt, izmantojot Nernsta vienādojumi:
Ejonu= RT/ zF* lnC ārā/ C iekšā(4)
kur R – gāzes konstante, T – absolūtā temperatūra, z – jonu valence (anjoniem negatīva) C ārā - ekstracelulāro jonu koncentrācija, C iekšā - intracelulāro jonu koncentrācija, F Faradeja numurs. Ja vienādojumā aizvietojam konstantes, tad ķermeņa temperatūrā (T = 310 K) kālija jonu līdzsvara potenciāls E K ir vienāds ar:
Ek= –61 mB žurnāls /(5)
Ja [ K + out ]/[ K + in ] = 39, kā parādīts tabulā. 1.1 tad
Ek= –61 m B log 39= –97 mV.
Patiešām, tika konstatēts, ka visām šūnām ir membrānas potenciāls; zīdītāju muskuļu šūnās tā līmenis ir aptuveni -90 mV. Atkarībā no apstākļiem un relatīvās jonu koncentrācijas šūnām var būt membrānas potenciāls no -40 līdz -120 mV. Šūnai iepriekš minētajā piemērā (1.1. tabula) atpūtas potenciāls, vienāds ar aptuveni -90 mV, norāda, ka kālija jonu plūsmas caur membrānas kanāliem ir aptuveni līdzsvarā. Tas nav pārsteidzoši, jo miera stāvoklī kālija kanālu stāvoklis visticamāk ir atvērts, t.i. membrāna ir viscaurlaidīgākā pret kālija joniem. Tomēr membrānas potenciālu nosaka citu jonu plūsmas.
Vieglums, ar kādu neuzlādētās daļiņas var izkliedēties caur membrānu, ir kvantitatīvi noteikts vienādojumā (3). Lādētu daļiņu caurlaidība ir aprakstīts ar nedaudz sarežģītāku vienādojumu:
P= m RT/ dF(6)
Kur m- jonu mobilitāte membrānā, d - membrānas biezums, a R, T un F – zināmās termodinamiskās konstantes. Šādi noteiktās dažādu jonu caurlaidības vērtības var izmantot, lai aprēķinātu membrānas potenciālu Em , kad kālija, nātrija un hlora joni vienlaikus iziet cauri membrānai (ar caurlaidību P K, P Na un P Cl attiecīgi). Tiek pieņemts, ka potenciāls vienmērīgi samazinās membrānā, lai lauka stiprums būtu nemainīgs. Šajā gadījumā tas attiecas Goldmana vienādojums jeb konstanta lauka vienādojums :
Em= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl ) (7)
Lielākajai daļai šūnu membrānu P K aptuveni 30 reizes lielāks par P Na (skatīt arī 1.3. apakšpunktu). Relatīvā vērtība P Cl ļoti atšķiras; daudzām membrānām P Cl mazs, salīdzinot ar R K tomēr citiem (piemēram, skeleta muskuļos) P Cl , ievērojami augstāks par P K.
Aktīvais transports, nātrija sūknis . Iepriekšējā sadaļā ir aprakstīta jonu pasīvā difūzija un no tā izrietošais membrānas potenciāls noteiktā intra- un ārpusšūnu jonu koncentrācijā. Tomēr šī procesa rezultātā jonu koncentrācija šūnā netiek automātiski stabilizēta, jo membrāna
potenciāls ir nedaudz elektronnegatīvāks nekā EK, un daudz salīdzinot ar E Na (apmēram +60 mV). Difūzijas dēļ intracelulāro jonu koncentrācija, vismaz kālija un nātrija, ir jāsalīdzina ar ārpusšūnu. Jonu gradienta stabilitāte tiek panākta ar aktīvo transportu: membrānas proteīni transportē jonus cauri membrānai pret elektriskiem un/vai koncentrācijas gradientiem, patērējot tam vielmaiņas enerģiju. Vissvarīgākais aktīvā transporta process ir darbs Na/K – sūknis, kas pastāv gandrīz visās šūnās;
sūknis izsūknē nātrija jonus no šūnas, vienlaikus iesūknējot kālija jonus šūnā. Tas nodrošina zemu intracelulāro nātrija jonu koncentrāciju un augstu kālija jonu koncentrāciju (1.1. tabula). Nātrija jonu koncentrācijas gradientam uz membrānas ir specifiskas funkcijas, kas saistītas ar informācijas pārraidi elektrisko impulsu veidā (skat. 2.2. nodaļu), kā arī citu aktīvo transporta mehānismu uzturēšanu un šūnu tilpuma regulēšanu (skat. zemāk). Tāpēc nav pārsteidzoši, ka vairāk nekā 1/3 no šūnas patērētās enerģijas tiek tērēta Na/K sūknim, un dažās no aktīvākajām šūnām tā darbībai tiek tērēta līdz 70% enerģijas.
Na/K transporta proteīns ir ATPāze. Uz membrānas iekšējās virsmas tas sadala ATP ADP un fosfātos (1.6. att.). Vienas ATP molekulas enerģija tiek izmantota trīs nātrija jonu transportēšanai no šūnas un vienlaikus divus kālija jonus šūnā, t.i., kopumā vienā ciklā no šūnas tiek noņemts viens pozitīvais lādiņš. Tādējādi Na/K sūknis ir elektrogēns(rada elektrisko strāvu pāri membrānai), kas izraisa membrānas potenciāla elektronegativitātes palielināšanos par aptuveni 10 mV. Transporta proteīns veic šo darbību lielā ātrumā: no 150 līdz 600 nātrija joniem sekundē. Transporta proteīna aminoskābju secība ir zināma, taču šī sarežģītā vielmaiņas transporta mehānisms vēl nav skaidrs. Šis process ir aprakstīts, izmantojot nātrija vai kālija jonu pārneses enerģijas profilus ar olbaltumvielām (1.5.,5. att.). Pēc šo profilu izmaiņu rakstura, kas saistītas ar pastāvīgām transporta proteīna konformācijas izmaiņām (process, kas prasa enerģiju), var spriest par apmaiņas stehiometriju: divi kālija joni tiek apmainīti pret trim nātrija joniem.
Na/K sūknis, tāds pats kā izolēts Na+ /K + atkarīgā membrānas ATPāze, ko īpaši inhibē sirds glikozīds ouabaīns (strofantīns). Tā kā Na/K sūkņa darbība ir daudzpakāpju ķīmiska reakcija, tā, tāpat kā visas ķīmiskās reakcijas, lielā mērā ir atkarīga no temperatūras, kas

Rīsi. 1.6.Na/K sūkņa-ATPāzes diagramma (iegremdēta plazmas membrānas lipīdu divslānī), kas vienā ciklā no šūnas noņem trīs Na + jonus pret potenciāla un koncentrācijas gradientiem un ienes šūnā divus K jonus. + . Šī procesa laikā viena ATP molekula tiek sadalīta ADP un fosfātā. Diagramma parāda ATPāzi kā dimēru, kas sastāv no lielas (funkcionālas) un mazas apakšvienības; membrānā tas pastāv kā tetramērs, ko veido divas lielas un divas mazas apakšvienības
attēlā parādīts. 1.7. Šeit tiek parādīta nātrija jonu plūsma no muskuļu šūnām attiecībā pret laiku; tas ir praktiski līdzvērtīgs nātrija jonu plūsmai, ko nodrošina Na/K sūkņa darbība, jo nātrija jonu pasīvā plūsma pret koncentrāciju un potenciālajiem gradientiem ir ārkārtīgi maza. Ja zāles atdzesē par aptuveni 18°C, nātrija jonu plūsma no šūnas ātri samazināsies 15 reizes, un uzreiz pēc karsēšanas tā tiks atjaunota sākotnējā līmenī. Šis nātrija jonu plūsmas samazinājums no šūnas ir vairākas reizes lielāks nekā tas, kas atbilstu difūzijas procesa vai vienkāršas ķīmiskas reakcijas atkarībai no temperatūras. Līdzīgs efekts vērojams, ja vielmaiņas enerģija ir izsmelta saindēšanās ar dinitrofenolu (DNP) rezultātā (1.7.,5. att.). Līdz ar to nātrija jonu plūsmu no šūnas nodrošina no enerģijas atkarīga reakcija - aktīvs sūknis. Vēl viena sūkņa īpašība, kā arī ievērojama temperatūras un enerģijas atkarība, ir piesātinājuma līmeņa klātbūtne (tāpat kā visas citas ķīmiskās reakcijas); tas nozīmē, ka sūkņa ātrums nevar pieaugt bezgalīgi, palielinoties transportēto jonu koncentrācijai (1.8. att.). Turpretim pasīvi difundējošas vielas plūsma palielinās proporcionāli koncentrācijas starpībai saskaņā ar difūzijas likumu (1. un 2. vienādojums).

Rīsi. 1.7. A, B. Aktīvs Na transports + . Y ass: radioaktīvā 24 Na + plūsma no šūnas (imp./min). Abscisu ass: laiks kopš eksperimenta sākuma. A.Šūnu atdzesē no 18,3 °C līdz 0,5 °C; plūsma Na+ no šūnas šajā periodā tiek kavēta. B. Na + plūsmas nomākšana no šūnas ar dinitrofenolu (DNP) koncentrācijā 0,2 mmol/l (modificēts)
Papildus Na/K sūknim plazmas membrānā ir vēl vismaz viens sūknis - kalcijs;šis sūknis izsūknē kalcija jonus (Ca 2+) no šūnas un ir iesaistīts to intracelulārās koncentrācijas uzturēšanā ārkārtīgi zemā līmenī (1.1. tabula). Kalcija sūknis ļoti lielā blīvumā atrodas muskuļu šūnu sarkoplazmatiskajā retikulumā, kurā ATP molekulu sadalīšanās rezultātā uzkrājas kalcija joni (sk. 4. nodaļu).
Na / K sūkņa ietekme uz membrānas potenciālu un šūnu tilpumu . Attēlā 1.9. attēlā parādītas dažādas membrānas strāvas sastāvdaļas un parādīta jonu intracelulārā koncentrācija, kas

Rīsi. 1.8.Sakarība starp molekulu transportēšanas ātrumu un to koncentrāciju (pie ieejas kanālā vai sūkņa saistīšanās vietā) difūzijas laikā caur kanālu vai sūknēšanas transportēšanas laikā. Pēdējais kļūst piesātināts augstā koncentrācijā (maksimālais ātrums, Vmax ) vērtība uz x ass, kas atbilst pusei no maksimālā sūkņa ātruma ( Vmax /2), ir līdzsvara koncentrācija UZ m

Rīsi. 1.9.Diagramma, kas parāda Na+ koncentrāciju , K+ un Cl – šūnā un ārpusē un šo jonu iekļūšanas ceļi caur šūnas membrānu (caur specifiskiem jonu kanāliem vai izmantojot Na/K sūkni. Pie dotiem koncentrācijas gradientiem, līdzsvara potenciāliem E Na, E K un E С l – vienāds ar norādītajiem, membrānas potenciāls Em = – 90 mV
nodrošināt to eksistenci. Caur kālija kanāliem tiek novērota kālija jonu ārējā strāva, jo membrānas potenciāls ir nedaudz elektropozitīvāks nekā kālija jonu līdzsvara potenciāls. Nātrija kanālu kopējā vadītspēja ir daudz zemāka nekā kālija kanālu vadītspēja, t.i. nātrija kanāli ir atvērti daudz retāk nekā kālija kanāli miera stāvoklī; taču šūnā nonāk aptuveni tikpat daudz nātrija jonu, cik kālija joni iziet no šūnas, jo ir nepieciešama liela koncentrācija un potenciālie gradienti, lai nātrija joni difundētu šūnā. Na/K sūknis nodrošina ideālu pasīvās difūzijas strāvu kompensāciju, jo tas transportē nātrija jonus no šūnas un kālija jonus tajā. Tādējādi sūknis ir elektrogēns, jo atšķiras lādiņu skaits, kas tiek pārnesti uz šūnu un no tās, kas normālā darbības ātrumā rada membrānas potenciālu aptuveni 10 mV ir elektronnegatīvāks nekā tad, ja to radītu tikai pasīvās jonu plūsmas (sk. 7. vienādojumu). Rezultātā membrānas potenciāls tuvojas kālija līdzsvara potenciālam, kas samazina kālija jonu noplūdi. Na aktivitāte/K sūkni regulē nātrija jonu intracelulārā koncentrācija. Samazinoties no šūnas izvadāmo nātrija jonu koncentrācijai, sūkņa ātrums palēninās (1.8. att.), lai sūkņa darbība un nātrija jonu plūsma šūnā līdzsvarotu viens otru, saglabājot nātrija intracelulāro koncentrāciju. jonu līmenī aptuveni 10 mmol/l.
Lai saglabātu līdzsvaru starp sūknēšanas un pasīvās membrānas strāvām, ir nepieciešams daudz vairāk Na / K sūkņa molekulu nekā kanālu proteīni kālija un nātrija joniem. Kad kanāls ir atvērts, caur to dažās milisekundēs iziet desmitiem tūkstošu jonu (skat. iepriekš), un, tā kā kanāls parasti atveras vairākas reizes sekundē, tad kopumā šajā laikā caur to iziet vairāk nekā 10 5 joni. Viens sūkņa proteīns pārvieto vairākus simtus nātrija jonu sekundē, tāpēc plazmas membrānā jāsatur apmēram 1000 reižu vairāk sūkņa molekulu nekā kanālu molekulās. Kanālu strāvu mērījumi miera stāvoklī uzrādīja vidēji viena kālija un viena nātrija atvērtā kanāla klātbūtni uz 1 μm 2 membrānas; No tā izriet, ka vienā telpā jāatrodas aptuveni 1000 Na/K sūkņa molekulām, t.i. attālums starp tiem ir vidēji 34 nm; Sūkņa proteīna kā kanāla proteīna diametrs ir 8–10 nm. Tādējādi membrāna ir diezgan blīvi piesātināta ar sūknēšanas molekulām
Faktam, ka nātrija jonu ieplūšana šūnā un kālija jonu ieplūšana no šūnas tiek kompensēta ar sūkņa darbību, ir arī citas sekas, kas sastāv no stabila osmotiskā spiediena uzturēšanas un nemainīgs apjoms.Šūnas iekšpusē ir liela lielu anjonu koncentrācija, galvenokārt olbaltumvielas (A - 1.1. tabulā), kas nespēj iekļūt membrānā (vai iekļūt caur to ļoti lēni) un tāpēc ir fiksēta sastāvdaļa šūnas iekšpusē. Lai līdzsvarotu šo anjonu lādiņu, ir nepieciešams vienāds skaits katjonu. Pateicoties Na/K sūkņa darbībai, šie katjoni galvenokārt ir kālija joni. Ievērojams intracelulāro jonu koncentrācijas pieaugums varētu notikt tikai ar anjonu koncentrācijas palielināšanos C1 plūsmas dēļ – pa koncentrācijas gradientu šūnā (1.1. tabula), taču membrānas potenciāls to neitralizē. Ienākošā strāva Cl – novērota tikai līdz tiek sasniegts hlora jonu līdzsvara potenciāls; to novēro, ja hlora jonu gradients ir gandrīz pretējs kālija jonu gradientam, jo hlora joni ir negatīvi lādēti (4. vienādojums). Tādējādi tiek noteikta zema hlora jonu intracelulārā koncentrācija, kas atbilst zemai kālija jonu ekstracelulārajai koncentrācijai. Rezultāts ir kopējā jonu skaita ierobežojums šūnā. Ja, bloķējot Na/K sūkni, piemēram, anoksijas laikā, membrānas potenciāls samazinās, tad hlora jonu līdzsvara potenciāls samazinās, un attiecīgi palielinās hlorīda jonu intracelulārā koncentrācija. Atjaunojot lādiņu līdzsvaru, šūnā nonāk arī kālija joni; palielinās kopējā jonu koncentrācija šūnā, kas palielina osmotisko spiedienu; tas iespiež ūdeni šūnā. Šūna uzbriest. Šis pietūkums tiek novērots in vivo enerģijas trūkuma apstākļos.
Koncentrācijas gradients Na + kā membrānas transportēšanas dzinējspēks . Na/K sūkņa nozīme šūnai neaprobežojas tikai ar normālu K + un Na + gradientu stabilizēšanu uz membrānas. Enerģija, kas uzkrāta membrānas gradientā Na+ , bieži izmanto, lai atvieglotu citu vielu transportēšanu uz membrānām. Piemēram, attēlā. 1.10 parāda “vienkāršs” Na+ un cukura molekulas nonāk šūnā. Membrānas transporta proteīns transportē cukura molekulu šūnā pat pret koncentrācijas gradientu, bet tajā pašā laikā Na + pārvietojas pa koncentrāciju un potenciālo gradientu, nodrošinot enerģiju cukura transportēšana.Šis cukuru transports ir pilnībā atkarīgs no augsta gradienta esamības Na+ ; ja intracelulārā koncentrācija Na+ ievērojami palielinās, cukuru transportēšana apstājas. Dažādiem c akharov, ir dažādas simport sistēmas. Aminoskābju transportēšanašūnā, kas ir līdzīga transportēšanai c akharov parādīts attēlā. 1,10; to nodrošina arī gradients Na+ , Ir vismaz piecas dažādas simport sistēmas, no kurām katra ir specializējusies vienai saistīto aminoskābju grupai.
Papildus vienkāršajām sistēmām ir arī "antiporteri". Viens no tiem, piemēram, vienā ciklā izvada no šūnas vienu kalcija jonu apmaiņā pret trim ienākošajiem nātrija joniem (1.10. att.). Enerģija priekš Ca 2+ transports veidojas trīs nātrija jonu iekļūšanas dēļ pa koncentrāciju un potenciālo gradientu. Šī enerģija ir pietiekama (pie miera potenciāla), lai uzturētu augstu kalcija jonu gradientu (no mazāk nekā 10–7 mol/L šūnā līdz aptuveni 2 mmol/L ārpus šūnas).
Endo- un eksocitoze . Par noteiktām vielām, kas nonāk šūnā vai jāizvada

Rīsi. 1.10.Membrānas lipīdu divslānī iestrādātie proteīni veicina glikozes un Na + iekļūšanu šūnā, kā arī Ca2+/Na+ –antiports, kurā dzinējspēks ir Na + gradients uz šūnas membrānas
no tā nav transporta kanālu; Šīs vielas ietver, piemēram, olbaltumvielas un holesterīnu. Tie var iekļūt caur plazmas membrānu pūslīši, vai burbuļi, caur endo- un eksocitozi. Attēlā 1.11. attēlā parādīti šo procesu galvenie mehānismi. Eksocitozes gadījumā daži organoīdi (skatīt zemāk) veido pūslīšus, kas piepildīti ar vielu, kas ir jāizņem no šūnas, piemēram, hormoniem vai ārpusšūnu enzīmiem. Kad šādi pūslīši sasniedz plazmas membrānu, to lipīdu membrāna saplūst ar to, tādējādi ļaujot saturam izkļūt ārējā vidē. Pretējā procesā, endocitozē, plazmas membrāna invaginējas, veidojot bedri, kas pēc tam padziļinās un aizveras, veidojot intracelulāru pūslīšu, kas piepildīta ar ārpusšūnu šķidrumu un dažām makromolekulām. Lai nodrošinātu šo membrānas saplūšanu un pūslīšu slēgšanu, citoskeleta kontraktilie elementi darbojas kopā ar pašām membrānām (skatīt zemāk). Endocitoze ne vienmēr ietver tikai ārpusšūnu vides uztveršanu šūnā. Šūnu membrāna bieži satur specifiskus makromolekulu receptorus, piemēram, insulīnu vai antigēnus, kas bieži ir sakārtoti specializētās grupās. Pēc šo makromolekulu saistīšanās ar saviem receptoriem membrānas zonā, kas ieskauj receptoru, notiek endocitoze, un makromolekula selektīvi tiek transportēta šūnā (1.12. att., B).
Endo- un eksocitoze šūnās notiek nepārtraukti. Membrānas materiāla apgrozījuma apjoms ir ievērojams; 1 stundas laikā makrofāgs vezikulu veidā absorbē divkāršu savas citoplazmas membrānas virsmas laukumu. Lielākajā daļā šūnu membrānas materiāla aprite nenotiek tik intensīvi, bet tai tomēr vajadzētu būt nozīmīgai.

Rīsi. 1.11.Eksocitoze un endocitoze. Uz augšu: intracelulārā pūslīša saplūst ar plazmas membrānas lipīdu divslāni un atveras ārpusšūnu telpā. Šo procesu sauc par eksocitozi. Apakšā: plazmas membrāna invaginējas nelielā apgabalā un atbrīvo pūslīšus, kas piepildītas ar ārpusšūnu materiālu. Šo procesu sauc par endocitozi
1.3. Vielu transportēšana šūnā
Endo- un eksocitoze ir ne tikai vielu transportēšanas procesi pa šūnas membrānu, bet arī membrānu - pašas šūnas strukturālo komponentu - apmaiņas procesi. Šajā sadaļā tiek aplūkoti citi līdzīgi transporta procesi šūnā un tās organellās.
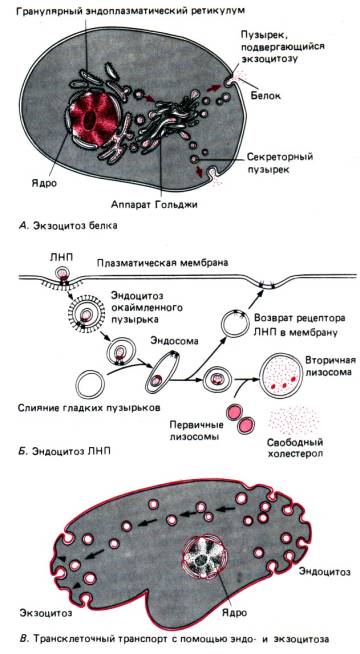
Rīsi. 1.12. A–B. Procesu shēma, ieskaitot ekso- un endocitozi. A. Granulētajā endoplazmatiskajā retikulā sintezētais proteīns caur Golgi aparātu tiek transportēts uz plazmas membrānu, kur tas tiek izdalīts eksocitozes ceļā. B. Holesterīns, kas saistīts ar ZBL (zema blīvuma lipoproteīnu) daļiņām, piesaistās plazmas membrānai, izraisa endocītu pūslīšu veidošanos šajā membrānas reģionā un tiek transportēts uz lizosomām, kur tas tiek atbrīvots. IN. Ekstracelulārs materiāls, kas notverts endocitozes laikā (attēlā labajā pusē), transportē caur šūnu pūslīšos vai pūslīšos un izdalās eksocitozes ceļā (attēlā pa kreisi)
Difūzija . Dabiski, ka citozolā koncentrāciju atšķirības tiek novērstas difūzijas dēļ; tas pats attiecas uz šķidrumiem, kas atrodas organellās. Augstās izšķīdušo olbaltumvielu koncentrācijas dēļ difūzija šeit ir daudz lēnāka nekā ūdenī. Lipīdu membrānas — ap šūnām un organellās — ir divdimensiju šķidrumi, kuros notiek difūzija. Membrānas divslāņu lipīdi izkliedējas savā slānī, reti pārvietojoties no viena uz otru. Arī tajos iegremdētie proteīni ir diezgan kustīgi; tie griežas ap asi, kas ir perpendikulāra membrānai, vai izkliedējas sāniski ar ļoti atšķirīgām difūzijas konstantēm, 2–10 000 reižu lēnāk nekā fosfolipīdi. Tātad, ja dažas olbaltumvielas brīvi pārvietojas lipīdu slānī un tādā pašā ātrumā kā pašas lipīdu molekulas, tad citas ir noenkurotas, t.i. diezgan stingri savienots ar citoskeletu. Membrānā ir “pastāvīgi” specifisku proteīnu agregāti, piemēram, nervu šūnu pre- un postsinaptiskās struktūras. Brīvi kustīgus proteīnus var demonstrēt, saistot tos ar fluorescējošām krāsvielām, kuru luminiscenci inducē, īslaicīgi apgaismojot nelielu membrānas laukumu. Šādi eksperimenti liecina, ka mazāk nekā 1 minūtē ar krāsvielu saistītie proteīni tiek vienmērīgi sadalīti pa membrānu attālumos līdz 10 μm.
Aktīvs transports organellu membrānās .
Aktīvie transporta procesi, kuriem ir būtiska nozīme plazmas membrānas darbībā, notiek arī šūnas iekšienē, organellu membrānās. Dažādu organellu specifiskais saturs veidojas daļēji iekšējās sintēzes dēļ un daļēji aktīvās transportēšanas dēļ no citozola. Viens no pēdējiem piemēriem ir iepriekš minētais Ca 2+ sūknis muskuļu šūnu sarkoplazmatiskajā retikulumā. Īpaši interesanti ir tas, ka ATP sintēzes gadījumā mitohondrijās pretējs princips attiecas uz to, kas notiek plazmas membrānas ATPāzes sūkņos (1.6. att.). ATP sintēzes laikā oksidatīvais metabolisms izraisa stāva gradienta veidošanos H+ uz iekšējām membrānām. Šis gradients ir virzītājspēks molekulu aktīvās transportēšanas sūknēšanas cikla reversajam procesam: H + joni pārvietojas pa membrānu pa gradientu, un rezultātā atbrīvotā enerģija nodrošina ATP sintēzi no ADP un fosfāta. Iegūtais ATP savukārt nodrošina enerģiju šūnai, tostarp aktīvai transportēšanai.
Transports pūslīšos . Šūnā ir liels skaits organellu un ar tām saistīto pūslīšu (1.1. att.). Šīs organellas un jo īpaši pūslīši atrodas pastāvīgā kustībā, transportējot to saturu uz citām organellām vai plazmas membrānu. Vezikulas var arī migrēt no šūnu membrānas uz organellām, piemēram, endocitozē.
Process olbaltumvielu sekrēcija attēlā parādīts. 1.12, A. Proteīns tiek sintezēts netālu no šūnas kodola uz ribosomām, kas saistītas ar endoplazmas tīklu (tā saukto granulēto jeb raupjo endoplazmas tīklu); Nokļūstot endoplazmatiskajā retikulā, proteīns tiek iesaiņots transporta pūslīšos, kas atdalās no organellas un migrē uz Golgi aparātu. Šeit tie saplūst ar Golgi aparāta cisternām, kur proteīns tiek modificēts (t.i., pārveidots par glikoproteīnu). Cisternu galos pūslīši atkal atdalās. Sekretārie pūslīši, kas satur modificēto proteīnu, pārvietojas uz plazmas membrānu un izdala to saturu eksocitozes ceļā.
Cits transporta ceļa piemērs šūnā ir parādīts attēlā. 1,12, B; Tā ir holesterīna uzsūkšanās šūnās. Asinīs transportētais holesterīns galvenokārt saistās ar olbaltumvielām, piemēram, daļiņām "zema blīvuma lipoproteīns"(ZBL). Šādas daļiņas pievienojas specifiskiem membrānas apgabaliem, kas satur ZBL receptorus, kur notiek endocitoze un ZBL tiek transportēts šūnā “apmales” pūslīšos. Šīs pūslīši saplūst, veidojot endosomas, un šī procesa laikā zaudē "apmales". Endosomas savukārt saplūst ar primārajām lizosomām, kurās pārsvarā ir hidrolītiskie enzīmi, un veido sekundāras, lielākas lizosomas. Tajos holesterīns izdalās no ZBL daļiņām un izkliedējas citozolā, kur kļūst pieejams, piemēram, lipīdu membrānu sintēzei. No endosomām tiek atdalītas arī vezikulas, kas nesatur ZBL, kas īpašā veidā pārvietojas uz plazmas membrānu un saplūst ar to, atgriežot membrānas materiālu un, iespējams, arī ZBL receptorus. No brīža, kad ZBL daļiņa saistās ar membrānu, līdz holesterīns tiek atbrīvots no sekundārās lizosomas, paiet 10-15 minūtes. ZBL saistīšanās un uzsūkšanās traucējumiem, t.i., holesterīna apgādē šūnām, ir izšķiroša nozīme nopietnas un plaši izplatītas slimības – aterosklerozes (artēriju “sacietēšanas”) attīstībā.
Ir daudz citu transporta maršrutu, kas ir līdzīgi tiem, kas parādīti attēlā. 1.11 un 1.12, A, ar kuru palīdzību šūnā pārvietojas specifiski pūslīši. Nav precīzi zināms, kā viņi pārvietojas, taču, iespējams, šajā procesā ir iesaistīti citoskeleta elementi. Pūslīši var slīdēt pa mikrotubulām, un tādā gadījumā kustību enerģiju acīmredzot nodrošina ar pūslīšiem saistītais proteīns ATPāze (skatīt zemāk). Paliek pilnīgi neskaidrs, cik dažādu pūslīšu, kustoties viena pēc otras visos virzienos, sasniedz galamērķi. Tie acīmredzot ir “jāatzīmē” tādā veidā, ko atpazīst transporta sistēma un pārvērš mērķtiecīgā kustībā.
Pārvadāšana, veidojot un iznīcinot organellus . Līdz šim endo- un eksocitozi esam uzskatījuši par vezikulu satura transportēšanas procesiem. Ir vēl viens šo procesu aspekts, kas slēpjas faktā, ka plazmas membrānas mērķtiecīga noņemšana no vienas šūnas virsmas daļas ar endocitozes palīdzību un, gluži pretēji, tās pievienošana citai ar eksocitozi, pārvieto lielus membrānas laukumus (att. 1.12.E), dodot šūnai iespēju, piemēram, izveidot izaugumu vai pārvietoties.
Līdzīgi pārkārtojumi ir raksturīgi arī citoskeletam, īpaši mikrofilamentiem un mikrotubulām (1.1. att.). Mikrofilamenti galvenokārt sastāv no F-aktīna proteīns, kas monomēra polimerizācijas rezultātā no citozola spēj salikt šķiedru kūlīšos. Saiņi ir polarizēti, tas ir, bieži vien aug tikai vienā galā, uzkrājot jaunas aktīna molekulas, savukārt otrs gals ir inerts vai šeit notiek izjaukšana. Pateicoties šai polarizētajai augšanai, mikrošķiedras pārvietojas efektīvi un to tīkla struktūra var mainīties. Aktīna pāreja no depolimerizēta stāvokļa (sol) uz organizētu stāvokli (gelu) var notikt ļoti ātri citu proteīnu ietekmē vai jonu koncentrācijas izmaiņu ietekmē (skatīt zemāk). Ir arī proteīni, kas izraisa aktīna pavedienu sadalīšanos, veidojot īsus fragmentus. Daudzu šūnu plānās projekcijas - filopodijas - satur centrālu aktīna kūli (1.1. att.), un, iespējams, dažādas filopodijas kustības ir saistītas ar aktīna pārejām: polimerizāciju - depolimerizāciju.
Mikrotubulasarī bieži iziet līdzīgas kustības. Šo kustību mehānisms ir līdzīgs - tubulīna polimerizācija no citozola tā, ka viens mikrocaurules gals aug, bet otrs vai nu nemainās, vai arī tur notiek izjaukšana. Tādējādi mikrotubulis, atbilstoši pievienojot vai noņemot materiālu, var pārvietoties visā citozolā.
Citoskeleta aktīvās kustības . Izmaiņas citoskeleta struktūrās var rasties gan aktīvu kustību, gan iepriekš aprakstīto pārkārtojumu rezultātā. Daudzos gadījumos mikrotubulu un aktīna pavedienu kustību virza kontraktilie proteīni, kas saista pavedienus vai kanāliņus un var tos pārvietot attiecībā pret otru. Vāveres miozīns un dyneīns atrodas visu šūnu citozolā salīdzinoši augstā koncentrācijā; tie ir elementi, kas pārvērš enerģiju kustībā specializētās šūnās (muskuļos) un organellās (skropstiņos). Muskuļu šūnās miozīns veido biezus pavedienus, kas orientēti paralēli aktīna pavedieniem. Miozīna molekula ar savu "galvu" pievienojas aktīna pavedienam un, izmantojot ATP enerģiju, izspiež miozīnu gar aktīna molekulu. Pēc tam miozīns atdalās no aktīna. Daudzu šādu savienojuma-atvienošanas ciklu kombinācija noved pie makroskopiskām muskuļu šķiedru kontrakcija(4. nodaļa). Dineīnam ir līdzīga loma mikrotubulu kustībā skropstu laikā (1.1. att.). Nespecializētu šūnu citoplazmā miozīns un dyneīns neveido regulāras šķiedras, bet vairumā gadījumu nelielas molekulu grupas. Pat šādos mazos agregātos tie spēj pārvietot aktīna pavedienus vai mikrotubulas. Rīsi. 1.13. attēls ilustrē šo procesu, kad pretēji polarizētas miozīna molekulas ir pievienotas arī diviem aktīna pavedieniem, kas polarizēti dažādos virzienos. Miozīna galvas grupas noliecas pret molekulas asti, procesā patērējot ATP, un divi aktīna pavedieni pārvietojas pretējā virzienā, pēc tam miozīns no tiem atdalās. Šāda veida kustības, kuru laikā ATP enerģija tiek pārvērsta mehāniskā darbā, var mainīt citoskeleta un līdz ar to arī šūnas formu, kā arī nodrošināt ar citoskeletu saistīto organellu transportēšanu.
Intracelulārā transporta procesus visskaidrāk var parādīt uz nervu šūnas aksona. Axon transportsšeit ir detalizēti aplūkots, lai ilustrētu notikumus, kas lielākajā daļā šūnu varētu notikt līdzīgi. Aksons, kura diametrs ir tikai daži mikroni, var sasniegt vienu metru vai vairāk, un proteīnu pārvietošanās difūzijas ceļā no kodola uz aksona distālo galu aizņemtu vairākus gadus. Jau sen ir zināms, ka tad, kad jebkura aksona daļa tiek sašaurināta, tā aksona daļa, kas atrodas vairāk proksimāli, izplešas. Izskatās, ka centrbēdzes plūsma ir bloķēta aksonā. Tādas plūsma-ātrs aksonu transports var to pierāda radioaktīvo marķieru kustība, kā parādīts attēlā. 1.14. Radioaktīvi iezīmēts leicīns tika injicēts muguras saknes ganglijā, un pēc tam no 2. līdz 10. stundai tika mērīta radioaktivitāte sēžas nervā 166 mm attālumā no neironu šūnu ķermeņiem. 10 stundu laikā radioaktivitātes maksimums injekcijas vietā nedaudz mainījās. Bet radioaktivitātes vilnis izplatījās pa aksonu ar nemainīgu ātrumu aptuveni 34 mm 2 stundās jeb 410 mm/dienā. Ir pierādīts, ka visos homeotermisko dzīvnieku neironos ātrs aksonu transports notiek ar vienādu ātrumu, un nav novērojamas ievērojamas atšķirības starp plānām, nemielinizētajām šķiedrām un biezākajiem aksoniem, kā arī starp motorajām un sensorajām šķiedrām. Radioaktīvā marķiera veids arī neietekmē ātrās aksonu transportēšanas ātrumu; marķieri var kalpot kā dažādi radioaktīvi

Rīsi. 1.13.Nemuskuļu miozīna komplekss ar noteiktu orientāciju var saistīties ar dažādas polaritātes aktīna pavedieniem un, izmantojot ATP enerģiju, pārvietot tos vienu pret otru
molekulas, piemēram, dažādas aminoskābes, kas ir iekļautas neirona šūnu ķermeņa proteīnos. Ja analizējam nerva perifēro daļu, lai noteiktu šeit transportēto radioaktivitātes nesēju raksturu, tad šādi nesēji galvenokārt atrodami olbaltumvielu frakcijā, bet arī mediatoru un brīvo aminoskābju sastāvā. Zinot, ka šo vielu īpašības ir dažādas un to molekulu izmēri ir īpaši atšķirīgi, pastāvīgo transportēšanas ātrumu varam izskaidrot tikai ar tām visām kopīgu transporta mehānismu.
Aprakstīts iepriekš ātrs aksonu transports ir anterograde, i., novirzīts prom no šūnas ķermeņa. Ir pierādīts, ka dažas vielas pārvietojas no perifērijas uz šūnu ķermeni, izmantojot retrogrāds transports. Piemēram, acetilholīnesterāze tiek transportēta šajā virzienā ar ātrumu, kas ir 2 reizes mazāks nekā ātras aksonu transportēšanas ātrums. Marķieris, ko bieži izmanto neiroanatomijā, mārrutku peroksidāze, pārvietojas arī ar retrogrādu transportu. Retrogrādajam transportam, iespējams, ir svarīga loma olbaltumvielu sintēzes regulēšanā šūnu ķermenī. Dažas dienas pēc aksona griešanas šūnas ķermenī tiek novērota hromatolīze, kas norāda uz olbaltumvielu sintēzes traucējumiem. Laiks, kas nepieciešams hromatolīzei, korelē ar retrogrādā transportēšanas ilgumu no aksona transekcijas vietas uz šūnas ķermeni. Šis rezultāts arī liecina par šī traucējuma skaidrojumu - tiek traucēta proteīnu sintēzi regulējošās “signālvielas” pārraide no perifērijas. Ir skaidrs, ka galvenie "transportlīdzekļi", ko izmanto ātrai aksonālai
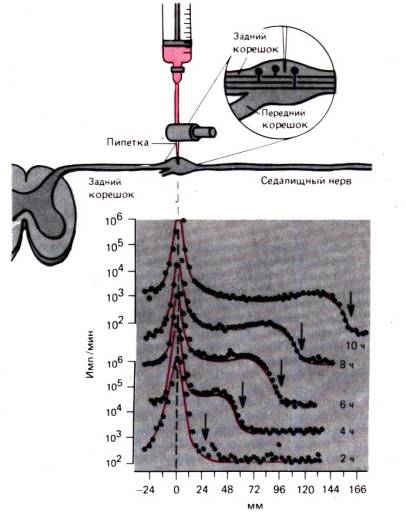
Rīsi. 1.14.Eksperiments, kas demonstrē ātru aksonu transportu kaķa sēžas nerva maņu šķiedrās. Ar tritiju iezīmēto leicīnu injicē muguras saknes ganglijā, un radioaktivitāti ganglijā un sensorajās šķiedrās mēra 2, 4, 6, 8 un 10 stundas pēc injekcijas. (attēla apakšā). Autors x-ass Tiek attēlots attālums no ganglija līdz sēžas nerva vietām, kur tiek veikts mērījums. Uz ordinātu ass tikai augšējai un apakšējai līknei radioaktivitāte (impulsi/min) tiek attēlota logaritmiskā skalā. Paaugstinātas radioaktivitātes "vilnis". (bultiņas) pārvietojas ar ātrumu 410 mm/dienā (garu)
transports ir pūslīši (vezikulas) un organellas, piemēram, mitohondriji, kas satur vielas, kuras nepieciešams transportēt. Lielāko pūslīšu jeb mitohondriju kustību var novērot, izmantojot mikroskopu in vivo . Šādas daļiņas veic īsas, straujas kustības vienā virzienā, apstājas, bieži pārvietojas nedaudz atpakaļ vai uz sāniem, atkal apstājas un pēc tam raustas galvenajā virzienā. 410 mm/dienā atbilst vidējam anterogradas kustības ātrumam aptuveni 5 μm/s; Tāpēc katras atsevišķas kustības ātrumam ir jābūt daudz lielākam, un, ja ņemam vērā organellu, pavedienu un mikrotubulu izmērus, tad šīs kustības patiešām ir ļoti ātras. Ātrai aksonu transportēšanai nepieciešama ievērojama ATP koncentrācija. Indes, piemēram, kolhicīns, kas izjauc mikrotubulas, arī bloķē ātru aksonu transportu. No tā izriet, ka mūsu aplūkotajā transportēšanas procesā pūslīši un organellas pārvietojas pa mikrotubulām un aktīna pavedieniem; šo kustību nodrošina nelieli dyneīna un miozīna molekulu agregāti, kas darbojas, kā parādīts attēlā. 1.13, izmantojot ATP enerģiju.
Var būt iesaistīts arī ātrs aksonu transports patoloģiskie procesi. Daži neirotropiski vīrusi (piemēram, herpes vai poliomielīta vīrusi) iekļūst aksonā perifērijā un ar retrogrādu transportu pārvietojas uz neironu ķermeni, kur vairojas un iedarbojas uz toksisku iedarbību. Stingumkrampju toksīns, proteīns, ko ražo baktērijas, kas nonāk organismā, kad āda ir bojāta, tiek uztverta ar nervu galiem un tiek transportēta uz neironu ķermeni, kur izraisa raksturīgas muskuļu spazmas. Ir zināmi toksiskas ietekmes gadījumi uz pašu aksonu transportēšanu, piemēram, rūpnieciskā šķīdinātāja akrilamīda iedarbība. Turklāt tiek uzskatīts, ka vitamīnu deficīta beriberi un alkoholiskās polineiropātijas patoģenēze ietver ātru aksonu transporta pārkāpumu.
Papildus ātrai aksonu transportēšanai šūnā notiek arī diezgan intensīva lēna aksonu transportēšana. Tubulīns pārvietojas pa aksonu ar ātrumu aptuveni 1 mm / dienā, un aktīns pārvietojas ātrāk - līdz 5 mm / dienā. Citas olbaltumvielas migrē kopā ar šiem citoskeleta komponentiem; piemēram, šķiet, ka fermenti ir saistīti ar aktīnu vai tubulīnu. Tubulīna un aktīna kustības ātrums aptuveni atbilst augšanas ātrumiem, kas konstatēti iepriekš aprakstītajam mehānismam, kur molekulas ir iekļautas mikrotubulas vai mikrofilamenta aktīvajā galā. Tāpēc šis mehānisms var būt lēnas aksonu transporta pamatā. Lēna aksonu transporta ātrums arī aptuveni atbilst aksonu augšanas ātrumam, kas acīmredzot norāda uz ierobežojumiem, ko citoskeleta struktūra uzliek otrajam procesam.
Noslēdzot šo sadaļu, jāuzsver, ka šūnas nekādā gadījumā nav statiskas struktūras, kā tās parādās, piemēram, elektronu mikroskopiskās fotogrāfijās. Plazmas membrāna un jo īpaši organoīdi atrodas pastāvīgā straujā kustībā un pastāvīgā pārstrukturēšanā; tāpēc viņi spēj darboties. Turklāt tās nav vienkāršas kameras, kurās notiek ķīmiskās reakcijas, bet gan augsti organizēti membrānu un šķiedru konglomerāti, kurā reakcijas notiek optimāli organizētā secībā.
Atsevišķas šūnas kā funkcionālas vienības uzturēšanu galvenokārt regulē kodols; šādu regulējošo mehānismu izpēte ir šūnu bioloģijas un bioķīmijas priekšmets. Tajā pašā laikā šūnām ir jāpārveido savas funkcijas atbilstoši vides apstākļiem un citu ķermeņa šūnu vajadzībām, t.i., tās kalpo kā funkcionālās regulēšanas objekti. Tālāk mēs īsi apsvērsim, kā šīs regulējošās ietekmes iedarbojas uz plazmas membrānu un kā tās sasniedz intracelulāros organellus.
Regulējošā ietekme uz šūnu membrānu
Membrānas potenciāls . Daudzos gadījumos šūnu funkciju regulēšana tiek veikta, mainot membrānas potenciālu. Vietējās potenciāla izmaiņas ir iespējamas, ja: 1) caur membrānu plūst strāva no blakus esošās šūnas zonas vai citas šūnas radīta; 2) mainās jonu ekstracelulārā koncentrācija (bieži vien [K + ]ārā ); 3) atveras membrānas jonu kanāli. Membrānas potenciāla izmaiņas var ietekmēt membrānas proteīnu konformāciju, jo īpaši izraisot kanālu atvēršanos vai aizvēršanos. Kā aprakstīts iepriekš, dažu membrānas sūkņu darbība ir atkarīga no membrānas potenciāla. Nervu šūnas ir specializētas, lai uztvertu membrānas potenciāla izmaiņas kā informāciju, kas ir jāapstrādā un jāpārraida (sk. 2. nodaļu).
Ekstracelulārās regulējošās vielas . Vissvarīgākais regulējošais mehānisms, kas ietver ārpusšūnu vielas, ir to mijiedarbība ar specifiskiem receptoriem uz plazmas membrānas vai šūnas iekšpusē. Šīs vielas ietver sinaptiskos raidītājus, kas pārraida informāciju starp nervu šūnām, vietējiem aģentiem un vielām, kas cirkulē asinīs un sasniedz visas ķermeņa šūnas, piemēram, hormonus un antigēnus. Sinaptiskie raidītāji ir mazas molekulas, kas atbrīvotas no nervu galiem sinapsē;
kad tie sasniedz blakus esošās, postsinaptiskās šūnas plazmas membrānu, tie iedarbina elektriskos signālus vai citus regulējošos mehānismus. Šis jautājums ir detalizēti apspriests sadaļā. 3.
Vietējie ķīmiskie aģenti bieži izdala specializētas šūnas. Tie brīvi izkliedējas ārpusšūnu telpā, bet to darbība ir ierobežota ar nelielu šūnu grupu, jo šīs vielas tiek ātri iznīcinātas vai nu spontāni, vai fermentu ietekmē. Viens šādu līdzekļu izdalīšanās piemērs ir atbrīvošanās histamīns tuklo šūnu bojājumu vai imūnās atbildes reakcijas dēļ. Histamīns izraisa asinsvadu gludo muskuļu šūnu atslābināšanos, palielina asinsvadu endotēlija caurlaidību un stimulē sensoro nervu galus, kas ir niezes sajūtas starpnieks. Citas vietējās ķīmiskās vielas izdala daudzas citas šūnas. Tipiski vietējie aģenti ir prostaglandīni, veido aptuveni 20 taukskābju atvasinājumu grupu. Tie tiek atbrīvoti nepārtraukti no plaši izplatītām šūnām, bet darbojas tikai lokāli, jo tos ātri iznīcina membrānas fosfolipāzes. Dažādiem prostaglandīniem ir plašs darbības spektrs: tie var izraisīt gludo muskuļu šūnu kontrakciju, izraisīt asins trombocītu (trombocītu) agregāciju vai nomākt dzeltenā ķermeņa attīstību olnīcās.
Apkalpo citi vietējie aģenti augšanas faktori. Vispazīstamākais ir simpātisko neironu nervu augšanas faktors (NGF), kas nepieciešams šo neironu augšanai un izdzīvošanai attīstības laikā. in vivo vai šūnu kultūrā. Acīmredzot šīs neironu klases mērķa šūnas izdala NGF un tādējādi nodrošina pareizu inervāciju. Veidojot orgānus, šūnām bieži ir “jāatrod ceļš” uz mērķa šūnām, kuras var atrasties ievērojamā attālumā. Attiecīgi ir jābūt daudziem specializētiem augšanas faktoriem, kas līdzīgi NGF.
Hormoni un antigēni ar asinīm pārnes uz visām šūnām. Antigēni izraisa imūnreakciju no šūnām, kurās ir specifiskas antivielas. Tomēr antigēni, kā likums, ir svešas vielas, kuras netiek ražotas reaģējošajā organismā (sīkāku informāciju skatīt 18. nodaļā). Daži hormoni, piemēram, insulīns vai tiroksīns, ietekmē ļoti dažādus šūnu tipus, savukārt citi, piemēram, dzimumhormoni, ietekmē tikai noteiktu šūnu veidu. Hormoni ir vai nu peptīdi, kuru darbību izraisa saistīšanās ar šūnu membrānas receptoriem, vai steroīdi un tiroksīns, kas izkliedējas caur lipīdu membrānu un saistās ar intracelulāriem receptoriem. Steroīdie hormoni saistās ar kodolhromatīnu, kā rezultātā notiek noteiktu gēnu transkripcija. Iegūtie proteīni izraisa izmaiņas šūnu funkcijās, kas ir hormonu specifiskā iedarbība. Jautājumi, kas saistīti ar hormonu izdalīšanos un darbību, ir detalizēti apskatīti nodaļā. 17.
Intracelulāra komunikācija, iesaistot otros sūtņus
Iepriekš aprakstītās regulējošās funkcijas ietver ietekmi uz šūnu membrānu. Informācijai, ko saņem šūnu membrāna, bieži ir jāraisa organoīdu reakcija, un tā tiek pārnesta uz tām ar dažādām vielām, kas pazīstamas kā otrie vēstneši (atšķirībā no pirmajiem vēstnešiem, kas šūnā nonāk no ārējiem avotiem). Otro mediatoru izpēte strauji attīstās, un nav garantijas, ka pašreizējais problēmas izpratnes līmenis būs pietiekami pilnīgs. Šeit mēs pieskarsimies trim labi izpētītiem mediatoriem: Ca 2+ , cAMP un inozitola trifosfāts.
Kalcijs.Vienkāršākais intracelulārais mediators ir Ca 2+ jons. Tā brīvā koncentrācija miera stāvoklī esošā šūnā ir ļoti zema un ir 10_–8–10–7 mol/l. Tas var iekļūt šūnā pa specifiskiem membrānas kanāliem, kad tie atrodas atvērtā stāvoklī, piemēram, mainoties membrānas potenciālam (sk. 2. nodaļu). Iegūtais Ca 2+ koncentrācijas pieaugums šūnā izraisa svarīgas reakcijas, piemēram, miofibrilu kontrakciju, kas ir muskuļu kontrakcijas pamatā (skat. 4. nodaļu), vai neirotransmiteru saturošu pūslīšu izdalīšanos no nervu galiem (sk. 3. nodaļu). . Abām reakcijām nepieciešama Ca 2+ koncentrācija aptuveni 10–5 mol/L. Ca 2+, kam ir regulējoša iedarbība, var izdalīties arī no intracelulārajiem krājumiem, piemēram, no endoplazmatiskā tīkla. Ca 2+ izdalīšanai no depo nepieciešama citu starpnieku līdzdalība (sk., piemēram, 1.16. att.).
Cikliskais adenozīna monofosfāts, cAMP. Nesen ir pierādīts, ka cikliskais adenozīna monofosfāts (cAMP), ķermeņa galvenā enerģijas avota ATP atvasinājums, ir svarīgs otrais vēstnesis. Sarežģītā reakciju ķēde, kas parādīta attēlā. 1.15, sākas ar receptoru R s uz plazmas membrānas ārējās virsmas, kas var kalpot kā dažādu mediatoru un hormonu specifiskas saistīšanās vieta. Pēc saistīšanās ar konkrētu "stimulējošu" molekulu R s maina savu uzbūvi; šīs izmaiņas ietekmē olbaltumvielas G s uz membrānas iekšējās virsmas tādā veidā, ka kļūst iespējama tās aktivizēšana ar intracelulāro guanozīna trifosfātu (GTP). Aktivizēts proteīns G s , savukārt, stimulē membrānas iekšējās virsmas fermentu adenilāta ciklāzi (AC), kas katalizē cAMP veidošanos no ATP. Ūdenī šķīstošs cAMP ir starpnieks, kas pārraida efektu

Rīsi. 1.15.Reakciju ķēde, kas ietver intracelulāro kurjeru cAMP (ciklisko adenozīna monofosfātu). Uzbudinoši vai inhibējoši ārējie signāli aktivizē membrānas receptorus R s vai Ri . Šie receptori regulē saistīšanās procesu G – proteīni ar intracelulāru GTP (guanozīna trifosfātu), tādējādi stimulējot vai inhibējot intracelulāro adenilāta ciklāzi (AC). Pastiprinošais enzīms AC pārvērš adenozīna trifosfātu (ATP) par cAMP, ko pēc tam fosfodiesterāze sadala par AMP. Brīvais cAMP difundē šūnā un aktivizē adenilāta kināzi (A-kināzi), atbrīvojot tās katalītisko apakšvienību C, kas katalizē intracelulāro proteīnu fosforilēšanos, t.i. veido ekstracelulārā stimula galīgo efektu. Diagrammā parādītas arī farmakoloģiskās zāles un toksīni, kas izraisa (+) vai kavē (-) noteiktas reakcijas (modificētas)
ekstracelulāro receptoru stimulēšana R s uz šūnas iekšējām struktūrām.
Paralēli stimulējošai reakciju ķēdei, kas ietver R s ir iespējama inhibējošo mediatoru un hormonu saistīšanās ar atbilstošo receptoru R i kas atkal caur GTP aktivētu proteīnu G , kavē AC un tādējādi cAMP veidošanos. Izkliedējoties šūnā, cAMP reaģē ar adenilāta kināzi (A-kināzi); tas atbrīvo C apakšvienību, kas katalizē proteīna P fosforilēšanos.Šī fosforilēšana pārvērš olbaltumvielas aktīvā formā, un tagad tās var iedarboties uz savu specifisko regulējošo iedarbību (piemēram, izraisīt glikogēna noārdīšanos). Šī sarežģītā regulēšanas sistēma ir ārkārtīgi efektīva, jo gala rezultāts ir daudzu proteīnu fosforilēšanās, t.i., regulējošais signāls pārvietojas pa ķēdi ar lielu pastiprinājumu. Ārējie mediatori, kas saistās ar receptoriem R s un R i katram no tiem raksturīgi, ārkārtīgi daudzveidīgi. Adrenalīns, kas savieno ar R s vai R i piedalās lipīdu un glikogēna metabolisma regulēšanā, kā arī sirds muskuļa kontrakcijas pastiprināšanā un citās reakcijās (skat. 19. nodaļu). Vairogdziedzera stimulējošais hormons, aktivizējot R s , stimulē vairogdziedzera hormona tiroksīna izdalīšanos, un prostaglandīns I kavē asins trombocītu agregāciju. Inhibējošie efekti, tostarp adrenalīns, ko izraisa R i ir izteikti lipolīzes palēnināšanās. Tādējādi cAMP sistēma ir daudzfunkcionāla intracelulāra regulēšanas sistēma, ko var precīzi kontrolēt ar ārpusšūnu stimulējošām un inhibējošām signālu vielām.
Inozīta fosfāts "IF" h " Otrā vēstneša – inozitola fosfāta – intracelulārā sistēma tika atklāta pavisam nesen (1.16. att.). Šajā gadījumā nav inhibējošā ceļa, bet ir līdzība ar cAMP sistēmu, kurā R receptoru stimulācijas efekts tiek pārnests uz GTP aktivētu G proteīnu uz membrānas iekšējās virsmas. Nākamajā posmā parastais membrānas lipīds fosfatidilinozitols (PI), iepriekš saņēmis divas papildu fosfātu grupas, tiek pārveidots par PI-difosfātu (PIP 2), ko aktivētā fosfodiesterāze (PDE) sadala par. inozīta trifosfāts(IFz) un lipīdu diacilglicerīns(DAG). Inozitola trifosfāts ir ūdenī šķīstošs otrs vēstnesis, kas izkliedējas citozolā. Tas darbojas galvenokārt, atbrīvojot Ca 2+ no endoplazmatiskā tīkla. Ca 2+ savukārt darbojas kā vēstnesis, kā aprakstīts iepriekš; piemēram, tas aktivizē Ca 2+ atkarīgo fosfokināzi, kas fosforilē enzīmus. Signālu nes arī DAG lipīdu apakšvienība (1.16. att.), kas plazmas membrānas lipīdu fāzē izkliedējas uz tās iekšējās virsmas esošo C-kināzi, kas tiek aktivizēta, kā kofaktoram piedaloties fosfatidilserīnam. Tad C-kināze izraisa proteīnu fosforilēšanos, pārvēršot tos aktīvā formā.
Intracelulāro otro kurjeru sistēmu IF 3 var kontrolēt arī dažādi ārējie mediatori un hormoni, tostarp acetilholīns, serotonīns, vazopresīns un vairogdziedzeri stimulējošais hormons; tāpat kā cAMP sistēmai, to raksturo dažādi intracelulāri efekti. Iespējams, ka šo sistēmu aktivizē arī gaisma acs vizuālajā receptorā, un tai ir galvenā loma fototransdukcijā (sk. 11. nodaļu). Pirmo reizi organisma individuālajā attīstībā IG sistēmas receptoru aktivizē sperma, kā rezultātā IG piedalās regulējošajās reakcijās, kas pavada olšūnas apaugļošanu.
CAMP un IFz-DAG sistēmas ir ļoti efektīvas bioloģiskie pastiprinātāji. Viņi

Rīsi. 1.16.Reakciju ķēde, kas ietver intracelulāro mediatoru IPG (inositola trifosfātu). Tāpat kā cAMP sistēmā, ekstracelulārais signāls tiek mediēts caur proteīnuG, kas šajā gadījumā aktivizē fosfodiesterāzi (PDE). Šis enzīms sadala fosfatidilinozīndifosfātu (PIP 2 ) plazmas membrānā pirms IF h un diacilglicerīns (DAG); JA h izkliedējas citoplazmā. Šeit tas izraisa Ca izdalīšanos 2+ no endoplazmatiskā tīkla; Ca koncentrācijas palielināšanās 2+ citoplazmā ([Ca 2+ ] i ) aktivizē proteīnkināzi, kas fosforilē un tādējādi aktivizē fermentus. Cits produkts, DAG, paliek membrānā un aktivizē proteīna kināzi C (fosfatidilserīna kofaktors, PS). Proteīnkināze C arī fosforilē enzīmus, kas mediē specifiskas darbības, kas saistītas ar ārējo receptoru stimulāciju R . Reakciju ķēdes atzari, kas ietver IF h un DAG var aktivizēt neatkarīgi ar attiecīgi jonomicīnu un forbola esteri (modificēts)
pārvērš reakciju starp raidītāju un ārējās membrānas receptoru dažādu intracelulāru proteīnu fosforilācijā, kas pēc tam var ietekmēt dažādas šūnu funkcijas. Viens no nozīmīgākajiem problēmas aspektiem ir tas, ka, cik šodien zināms, pastāv tikai šīs divas cieši saistītās šāda veida regulējošās sistēmas, kuras izmanto daudzi ārējie mediatori, lai regulētu dažādus intracelulāros procesus. Tajā pašā laikā šīs regulējošās sistēmas, tostarp Ca 2+, cieši mijiedarbojas viena ar otru, kas ļauj tām veikt precīzu šūnu funkciju regulēšanu.
Pamācības un ceļveži
1. Alberts IN., Bray D., Lewis J., Raff M., Roberts UZ., Vatsons J.D.Šūnu molekulārā bioloģija, Ņujorka un Londona, Garland Publishing Inc., 1983.
2. Czihak G., Longer H., Ziegler H.(red.). Bioloģija. Berlīne, Heidelberga, Ņujorka, Springere, 1983.
3. Hille IN. Uzbudināmo membrānu jonu kanāli. Sanderlenda, Masač., Sinauera asoc., 1984.
4. Hope W., Lohmann W.. Marki H., Ziegler H.(red.). Biofizika. Berlīne, Heidelberga, Ņujorka, Springere, 1984.
5. Džungermanis UZ., Mālers H.Bioķīmija. Berlīne, Heidelberga, Ņujorka, Springere, 1980.
6. Kandels E.R., Švarcs–J.H.,(red.). Neironu zinātnes principi, Ņujorka, Amsterdama, Oksforda, Elsevier, 1985.
7. Šiblers T. H., Šmits V.Anatomic des Menschen. Berlīne, Heidelberga, Ņujorka, Tokija, Springere, 1983.
Oriģinālie raksti un apskati
8. Beridža M.J. Komunikācijas molekulārais pamats šūnā, Sci. Amer, 253, 124-134 (1985).
9. Beridža M. J., Ērvins R. F. Inozitola trifosfāts, jauns otrs vēstnesis šūnu signālu pārraidē. Nature, 312, 315, 321 (1984).
10. Bretscher M.S.Šūnu membrānas molekulas, Sci. Amer., 253, 124–134 (1985).
11. Dauts Dž. Dzīvā šūna kā enerģijas pārvades iekārta. Minimālais miokarda metabolisma modelis, Biochem. et Biophys. Acta, 895, 41–62 (1987).
12. Hodžkins A.L., Katz IN. Nātrija jonu ietekme uz kalmāru milzu aksona elektrisko aktivitāti. J. Physiol. (Lond.), 108, 37–77 (1949).
13. Hodžkins A.L., Keinss R.D. Aktīvā katjonu transportēšana milzu aksonos no Sēpija un Loligo, J. Physiol. (Lond.), 128, 28–42 (1955).
14. Garāks P. Jonu kanāli ar konformācijas apakšstāvokļiem, Biophys. J., 47, 581–590 (1985).
15. Ochs S., Vērts P.M. Eksoplazmatiskais transports normālās un patoloģiskās sistēmās. In: Physiology and Pathology of Axons, S.G. Vaksmans, Ed. Ņujorka, Raven Press, 1978.
citu prezentāciju kopsavilkums“Bioloģijas mācīšanas metodes” - skolas zooloģija. Studentu iepazīstināšana ar zinātnisko zooloģisko datu izmantošanu. Morālā izglītība. Papildu vistu kūts svētība. Metožu izvēle. Dzīves procesi. Akvārija zivis. Uzturs. Vides izglītība. Dzīvības procesu materialitāte. Negatīvie rezultāti. Uzmanību studentiem. Obligātā forma. Skatoties uz maziem dzīvniekiem. Bioloģijas mērķi un uzdevumi. Stāsts.
“Problēmu mācīšanās bioloģijas stundās” - Zināšanas. Jaunas mācību grāmatas. Ceļš uz risinājumu. Problēma. Semināri. Kas ir uzdevums? Albrehts Durers. Problēmmācība bioloģijas stundās. Nestandarta nodarbības. Ko nozīmē problēmbāzēta mācīšanās. Dzīves kvalitāte. Bioloģija kā akadēmisks priekšmets. Jautājums. Problēmu risināšanas nodarbība. Samazināta interese par tēmu. Problēmas laboratorijas nodarbības.
“Kritiskā domāšana bioloģijas stundās” - “Kritiskās domāšanas tehnoloģija”. Izmantojot “kritiskās domāšanas attīstības” tehnoloģiju. Nodarbības galds. Motivācija mācīties. Ekosistēmas. Jēdziena “kritiskās domāšanas attīstīšana” nozīme. Tehnoloģiju pazīmes. RKM tehnoloģija. Nodarbības struktūra. Galvenie virzieni. Tehnoloģiju vēsture. Pedagoģiskās tehnoloģijas. Tehnoloģiju noteikumi. Bioloģijas uzdevumi. Fotosintēze. Dažādos nodarbības posmos izmantotās metodes.
“Bioloģijas stundas ar interaktīvo tāfeli” - Elektroniskās mācību grāmatas. Ieguvumi studentiem. Interaktīvā tāfele palīdz nodot informāciju ikvienam skolēnam. Didaktiskie uzdevumi. Bioloģisko problēmu risināšana. Priekšrocības darbam ar interaktīvajām tāfelēm. Darbs ar prezentācijām. Darbs pie objektu salīdzināšanas. Kustīgie objekti. Izklājlapu izmantošana. Interaktīvās tāfeles izmantošana skolēnu mācību procesā. Ieguvumi skolotājiem.
“Sistēmas darbības pieeja bioloģijā” - Semināra jautājumi. Darbības metode. Dryopithecus. Cilvēka izcelsmes ārpuszemes ceļš. Lizosomas. Ķīmiskā organizācija. Gymnosperms. Vielmaiņa. Analizatori. Sistēmas darbības pieeja bioloģijas mācīšanai. Hromosomas. Citoplazma. Aklums. Ausu garums. Personas klasifikācija. Zīdītāja skelets. Cilvēka evolūcijas ceļi. Mitoze. Virspusējs komplekss. Problemātisks jautājums. Nucleolus. Kodolenerģijas apvalks.
“Dators bioloģijā” - Studentu kopdarbība. Angiospermu ģimenes. Interaktīva apmācība. Mācību modeļi. Vērtēšanas sistēmas piemērs. Instrukciju karšu jautājumi. Instrukciju kartes piemērs. Pētnieki. Mikrogrupas. Interaktīvās mācību tehnoloģijas. Karuselis. Interaktīvās mācību tehnoloģijas. Interaktīvas pieejas bioloģijas stundās. Grupas darba forma. Uzdevumi “pētnieku” grupām.
