§ 2. Mga pangunahing bahagi ng isang eukaryotic cell
Ang mga eukaryotic cell (Larawan 8 at 9) ay mas kumplikado kaysa sa mga prokaryotic. Ang mga ito ay napaka-magkakaibang laki (mula sa ilang micrometer hanggang ilang sentimetro), sa hugis, at sa mga tampok na istruktura (Larawan 10).
kanin. 8. Istraktura ng isang eukaryotic cell. Pangkalahatang pamamaraan
kanin. 9. Estruktura ng cell ayon sa electron microscopy
kanin. 10. Iba't ibang eukaryotic cells: 1 - epithelial; 2 – dugo (e – erythroitis, / – leukitis); 3 - kartilago; 4 – buto; 5 – makinis na kalamnan; 6 - nag-uugnay na tissue; 7 – mga selula ng nerbiyos; 8 – striated na hibla ng kalamnan
Gayunpaman, ang pangkalahatang organisasyon at pagkakaroon ng mga pangunahing sangkap ay pareho sa lahat ng eukaryotic cells (Larawan 11).
kanin. 11. Eukaryotic cell (diagram)
Plasmalemma (panlabas na lamad ng selula). Ang batayan ng lamad ng plasma, tulad ng iba pang mga lamad sa mga selula (halimbawa, mitochondria, plastids, atbp.), Ay isang layer ng mga lipid na may dalawang hanay ng mga molekula (Larawan 12). Dahil ang mga molekula ng lipid ay polar (ang isang poste ay hydrophilic, ibig sabihin, naaakit ng tubig, at ang isa ay hydrophobic, ibig sabihin, tinataboy ng tubig), sila ay nakaayos sa isang tiyak na pagkakasunud-sunod. Ang mga hydrophilic na dulo ng mga molekula ng isang layer ay nakadirekta patungo sa may tubig na kapaligiran - sa cytoplasm ng cell, at ang iba pang layer - palabas mula sa cell - patungo sa intercellular substance (sa mga multicellular na organismo) o ang may tubig na kapaligiran (sa mga unicellular na organismo. ).
kanin. 12. Istraktura ng cell membrane ayon sa fluid mosaic model. Ang mga protina at glycoprotein ay inilulubog sa isang dobleng layer ng mga molekulang lipid, na ang kanilang mga hydrophilic na dulo (mga bilog) ay nakaharap palabas, at ang kanilang mga hydrophobic na dulo (mga kulot na linya) ay nakaharap sa malalim sa lamad
Ang mga molekula ng protina ay mosaically na naka-embed sa isang bimolecular layer ng mga lipid. Sa labas ng selula ng hayop, ang mga molekula ng polysaccharide ay nakakabit sa mga lipid at mga molekula ng protina ng plasmalemma, na bumubuo ng mga glycolipids at glycoproteins.
Ang pinagsama-samang ito ay bumubuo ng isang layer glycocalyx. Nauugnay sa kanya function ng receptor mga lamad ng plasma (tingnan sa ibaba); maaari rin itong makaipon ng iba't ibang sangkap na ginagamit ng cell. Bilang karagdagan, pinahuhusay ng glycocalyx ang mekanikal na katatagan ng plasmalemma.
Sa mga selula ng mga halaman at fungi mayroon ding cell wall na gumaganap ng isang sumusuporta at proteksiyon na papel. Sa mga halaman ito ay binubuo ng selulusa, at sa fungi ito ay gawa sa chitin.
Ang panlabas na lamad ng cell ay gumaganap ng isang bilang ng mga pag-andar, kabilang ang:
♦ mekanikal(pagsuporta, pagbuo ng anyo);
♦ hadlang-transportasyon(pumipili ng pagkamatagusin sa iba't ibang mga sangkap: pagpasok sa cell ng mga kinakailangang sangkap at pag-alis ng mga hindi kailangan at nakakapinsala);
♦ receptor(pagpapasiya ng iba't ibang mga kemikal na matatagpuan malapit sa cell; pang-unawa ng mga signal sa anyo ng mga hormone; pagkilala sa "dayuhang" protina ng mga selula ng immune system, atbp.).
Ang pagpapalitan ng mga sangkap sa pagitan ng cell at ng kapaligiran ay isinasagawa sa iba't ibang paraan - pasibo at aktibo.
Ang mga molekula ng tubig at iba't ibang mga ion ay pasibo (dahil sa pagsasabog, osmosis), nang walang cell na gumugugol ng enerhiya, pumapasok sa pamamagitan ng mga espesyal na pores - ito ay passive na transportasyon. Ang mga macromolecule tulad ng mga protina, polysaccharides, kahit buong mga cell, ay ibinibigay ng phagocytosis At pinocytosis sa pagkonsumo ng enerhiya - aktibong transportasyon.
Sa pamamagitan ng phagocytosis, ang buong mga cell o malalaking particle ay nilamon (halimbawa, isipin ang tungkol sa nutrisyon sa amoebas o phagocytosis ng mga proteksiyon na selula ng dugo ng bakterya). Sa panahon ng pinocytosis, ang mga maliliit na particle o droplet ng isang likidong sangkap ay nasisipsip. Karaniwan sa parehong mga proseso ay ang mga hinihigop na sangkap ay napapalibutan ng isang invaginating na panlabas na lamad upang bumuo ng isang vacuole, na pagkatapos ay gumagalaw nang malalim sa cytoplasm ng cell.
Ang exocytosis ay isang proseso (pagiging aktibong transportasyon din) na kabaligtaran sa direksyon ng phagocytosis at pinocytosis (Larawan 13). Sa tulong nito, ang hindi natutunaw na pagkain ay nananatili sa protozoa o ang mga biologically active substance na nabuo sa secretory cell ay maaaring alisin.
Cytoplasm. Ang cytoplasm ay ang mga nilalaman ng cell, na limitado ng plasmalemma, maliban sa nucleus. Naglalaman ito sangkap sa lupa (hyaloplasm), organelles At mga inklusyon.
Hyaloplasma- isang malapot na likido na maaaring nasa estado ng alinman Zola(likido), o gel(parang halaya).
Kung kinakailangan, ang cytoplasm ay may kakayahang baligtarin ang paglipat mula sa isang estado patungo sa isa pa. Halimbawa, sa panahon ng paggalaw ng amoeboid (tandaan ang seksyong "Protozoa" mula sa kurso ng zoology), sa panahon ng pagbuo ng isang pseudopod, ang mga mabilis na paglipat ng cytoplasm mula sa gel hanggang sol at vice versa ay nangyayari. Ito ay dahil sa pagkakaroon sa cytoplasm ng isang malaking bilang ng mga filamentous na molekula ng protina actina. Kapag kumonekta sila sa isa't isa upang bumuo ng isang three-dimensional na network, ang cytoplasm ay nasa isang gel state, at kapag ang network ay naghiwa-hiwalay, ito ay nasa sol state.
Ang hyaloplasm ay naglalaman ng iba't ibang mga sangkap - mga enzyme, protina, carbohydrates, taba at iba pa, organic at mineral. Ang iba't ibang mga proseso ng kemikal ay nagaganap dito - ang pagkasira ng mga sangkap, ang kanilang synthesis at mga pagbabago (mga pagbabago).
Organoids. Ito ay mga permanenteng bahagi ng isang cell na may isang tiyak na istraktura at pag-andar, na matatagpuan sa cytoplasm nito. Sa mga sumusunod ay pag-uusapan natin pangkalahatang layunin na mga organo, likas sa lahat ng uri ng mga selula ng lahat ng eukaryotes. Ang mga ito ay nauugnay sa pagtiyak ng mahahalagang tungkulin ng huli. Espesyal na Layunin Organoids matatagpuan lamang sa mga cell ng isang tiyak na (highly specialized) na uri - halimbawa, myofibrils sa mga selula ng kalamnan.
Ang mga organelle na may pangkalahatang layunin ay may parehong istraktura anuman ang mga cell at kung aling mga organismo sila nabibilang. Ngunit sa kanila ay may mga grupo na may lamad (endoplasmic reticulum, Golgi apparatus, mitochondria, plastids, lysosomes, vacuoles), pati na rin ang hindi lamad ( ribosom, sentro ng cell) istraktura.
Endoplasmic reticulum (ER). Ang EPS ay binubuo ng mga lamad at isang kumplikadong branched system ng mga tubules at cisterns na tumagos sa buong cytoplasm ng cell (Larawan 14). Mayroong dalawang uri ng EPS - magaspang At makinis. Ang mga ribosome ay nakakabit sa magaspang na lamad (mula sa cytoplasmic side), habang wala sila sa makinis na lamad.
kanin. 14. Endoplasmic reticulum
Ang endoplasmic reticulum ay gumaganap ng ilang mahahalagang tungkulin sa isang eukaryotic cell:
♦ naglilimita(paghahati ng panloob na dami ng cell sa iba't ibang mga puwang ng reaksyon);
♦ pakikilahok sa synthesis ng mga organikong sangkap(Ang mga ribosom ay matatagpuan sa mga lamad ng magaspang na EPS, at sa makinis na mga lamad mayroong mga enzyme complex na nagsisiguro sa synthesis ng mga lipid, carbohydrates, atbp.);
♦ pakikilahok sa pagbuo ng mga elemento ng Golgi apparatus, lysosomes;
♦ transportasyon ng mga sangkap.
Golgi apparatus. Ang Golgi apparatus (AG) ay isang sistema mga tangke(flat vacuoles) at mga bula(vesicles), na matatagpuan malapit sa cell nucleus, na nabuo dahil sa EPS bilang resulta ng paghihiwalay ng mga maliliit na fragment nito (Larawan 15). Kapag ang mga fragment na ito ay pinagsama, ang mga bagong cisterns ng Golgi apparatus ay bumangon, habang ang iba't ibang mga sangkap ay dinadala mula sa EPS, na kasangkot sa pagpupulong ng mga kumplikadong organikong compound (protina + carbohydrates, protina + lipid, atbp.), na inalis kasama ng tulong ng AG sa labas ng cell. Ang mga biologically active substance na ito ay maaaring tinanggal mula sa cell (gamit ang secretory vacuoles sa pamamagitan ng exocytosis) o bahagi ng lysosomes (tingnan sa ibaba) na nabuo ng AG.
kanin. 15. Golgi apparatus:
Ang Golgi apparatus ay gumaganap ng mga sumusunod na function:
♦ synthesis biologically active substances na ginawa ng cell;
♦ pagtatago (pagtanggal mula sa cell) ng iba't ibang mga sangkap(mga hormone, enzyme, mga sangkap kung saan itinayo ang cell wall, atbp.);
♦ pakikilahok sa pagbuo ng mga lysosome.
Mitokondria. Ang lahat ng uri ng eukaryotic cells ay may mitochondria (Larawan 16). Ang mga ito ay mukhang alinman sa mga bilugan na katawan o mga tungkod, mas madalas - mga thread. Ang kanilang mga sukat ay mula 1 hanggang 7 microns. Ang bilang ng mitochondria sa isang cell ay mula sa ilang daan hanggang sampu-sampung libo (sa malaking protozoa).
kanin. 16. Mitokondria. Sa itaas - mitochondria (a) sa mga kanal ng ihi, nakikita sa isang light microscope. Nasa ibaba ang isang three-dimensional na modelo ng mitochondrial organization: 1 – cristae; 2 – panlabas na lamad; 3 – panloob na lamad; 4 – matris
Ang mitochondrion ay nabuo sa pamamagitan ng dalawang lamad - panlabas At panloob, sa pagitan ng kung saan matatagpuan espasyo ng intermembrane. Ang panloob na lamad ay bumubuo ng maraming invaginations - cristae, na alinman sa mga plato o tubo. Ang organisasyong ito ay nagbibigay ng isang malaking lugar ng panloob na lamad. Naglalaman ito ng mga enzyme na nagsisiguro sa conversion ng enerhiya na nilalaman ng mga organikong sangkap (carbohydrates, lipids) sa enerhiya ng ATP, na kinakailangan para sa buhay ng cell. Samakatuwid, ang tungkulin ng mitochondria ay lumahok sa enerhiya mga proseso ng cellular. Iyon ang dahilan kung bakit ang isang malaking bilang ng mitochondria ay likas, halimbawa, sa mga selula ng kalamnan na gumaganap ng maraming trabaho.
Mga plastid. Sa mga selula ng halaman, matatagpuan ang mga espesyal na organel - mga plastid, na kadalasang may hugis ng suliran o bilugan na hugis, kung minsan ay mas kumplikado. Mayroong tatlong uri ng plastids - chloroplasts (Fig. 17), chromoplasts at leucoplasts.
Mga chloroplast naiiba sa berdeng kulay, na dahil sa pigment - chlorophyll, tinitiyak ang proseso potosintesis, ibig sabihin, ang synthesis ng mga organikong sangkap mula sa tubig (H 2 O) at carbon dioxide (CO 2) gamit ang enerhiya ng sikat ng araw. Ang mga chloroplast ay pangunahing matatagpuan sa mga selula ng dahon (sa mas matataas na halaman). Ang mga ito ay nabuo sa pamamagitan ng dalawang lamad na matatagpuan parallel sa bawat isa, na nakapalibot sa mga nilalaman ng chloroplasts - stroma. Ang panloob na lamad ay bumubuo ng maraming mga flat sac - thylakoids, na nakasalansan (tulad ng isang stack ng mga barya) - butil - at humiga sa stroma. Ito ay ang thylakoids na naglalaman ng chlorophyll.
Mga Chromoplast matukoy ang dilaw, orange at pulang kulay ng maraming mga bulaklak at prutas, sa mga selula kung saan sila ay naroroon sa malalaking dami. Ang mga pangunahing pigment sa kanilang komposisyon ay karoten. Ang functional na layunin ng mga chromoplast ay upang maakit ang mga hayop na may kulay, tinitiyak ang polinasyon ng mga bulaklak at pagpapakalat ng mga buto.
kanin. 17. Plastids: a – mga chloroplast sa mga selula ng dahon ng Elodea, nakikita sa isang light microscope; b - diagram ng panloob na istraktura ng chloroplast na may grana, na mga stack ng flat sac na matatagpuan patayo sa ibabaw ng chloroplast; c – isang mas detalyadong diagram na nagpapakita ng mga anastomosing tube na kumukonekta sa mga indibidwal na fan chamber
Mga leukoplast ay walang kulay na mga plastid na nakapaloob sa mga selula ng mga underground na bahagi ng mga halaman (halimbawa, sa mga tubers ng patatas), mga buto at ang core ng mga tangkay. Sa leucoplasts, ang almirol ay pangunahing nabuo mula sa glucose at naipon sa mga organo ng imbakan ng mga halaman.
Ang mga plastid ng isang uri ay maaaring magbago sa isa pa. Halimbawa, kapag ang mga dahon ay nagbabago ng kulay sa taglagas, ang mga chloroplast ay nagiging mga chromoplast.
Mga lysosome. Ang mga organel na ito ay mukhang mga vesicle na napapalibutan ng isang lamad na may diameter na hanggang 2 microns. Naglalaman ang mga ito ng ilang dosenang mga enzyme na sumisira sa mga protina, nucleic acid, polysaccharides at lipid. Ang pag-andar ng mga lysosome ay lumahok sa mga proseso ng intracellular breakdown ng mga kumplikadong organikong compound (halimbawa, mga sustansya o mga sangkap ng "ginugol" na mga bahagi ng cellular). Ang mga lysosome ay nagsasama sa mga phagocytic (o pinocytic) na mga vacuole upang bumuo ng isang digestive vacuole.
Ang pagbuo ng mga lysosome ay nangyayari dahil sa budding mula sa cisternae ng Golgi apparatus.
Mga ribosom. Ang mga ribosom (Larawan 18) ay naroroon sa mga selula ng parehong eukaryotes at prokaryote, dahil gumaganap sila ng mahalagang tungkulin sa biosynthesis ng protina(tingnan ang kabanata 5). Ang bawat cell ay naglalaman ng sampu, daan-daang libo (hanggang sa ilang milyon) ng mga maliliit na bilog na organelle na ito.
kanin. 18. Scheme ng istraktura ng isang ribosome na nakaupo sa lamad ng endoplasmic reticulum: 1 - maliit na subunit; 2 – tRNA; 3 – aminoacyl-tRNA; 4 – amino acid; 5 - malaking subunit; 6 - endoplasmic reticulum lamad; 7 - synthesized polypeptide chain
Ang ribosome ay binubuo ng dalawang hindi pantay na subunits (mga bahagi). Ang mga ito ay nabuo nang hiwalay at pinagsama, "sinasaklaw" ang messenger RNA, sa panahon ng synthesis ng isang molekula ng protina. Ang mga ribosom ay naglalaman ng iba't ibang mga protina at ribosomal RNA.
Mga pagsasama sa cellular. Ito ang pangalang ibinigay sa mga di-permanenteng bahagi sa cell, na nasa pangunahing sangkap ng cytoplasm sa anyo ng mga butil, butil o droplet. Ang mga inklusyon ay maaaring napapalibutan o hindi ng isang lamad.
Sa paggana, mayroong tatlong uri ng mga inklusyon: magreserba ng nutrients(almirol, glycogen, taba, protina), secretory inclusions(mga sangkap na katangian ng mga glandular na selula, na ginawa ng mga ito - mga hormone ng mga glandula ng endocrine, atbp.) at pagsasama ng espesyal na layunin(sa mga highly specialized na mga cell, halimbawa hemoglobin sa mga pulang selula ng dugo).
§ 3. Organisasyon ng cell nucleus. Mga Chromosome
Ang cell nucleus (tingnan ang Fig. 8 at 9) ay pinakamahalaga sa buhay ng cell, dahil ito ay nagsisilbing isang repository ng namamana na impormasyon na nakapaloob sa mga chromosome (tingnan sa ibaba).
Ang nucleus ay napapalibutan ng isang nuclear envelope na naghihiwalay sa mga nilalaman nito (karyoplasma) mula sa cytoplasm. Ang shell ay binubuo ng dalawang lamad na pinaghihiwalay ng isang puwang. Pareho sa kanila ay natatakpan ng maraming mga pores, salamat sa kung saan ang pagpapalitan ng mga sangkap sa pagitan ng nucleus at cytoplasm ay posible. Sa cell nucleus ng karamihan sa mga eukaryote mayroong mula 1 hanggang 7 nucleoli. Ang mga proseso ng RNA at tRNA synthesis ay nauugnay sa kanila.
Pangunahing Mga Bahagi ng Kernel – chromosome, nabuo mula sa isang molekula ng DNA at iba't ibang mga protina. Sa isang light mikroskopyo ay malinaw na nakikita lamang sila sa panahon ng paghahati ng cell (mitosis, meiosis). Sa isang cell na hindi naghahati, ang mga chromosome ay mukhang mahahabang manipis na mga thread na ipinamamahagi sa buong volume ng nucleus.
Sa panahon ng paghahati ng cell, ang mga chromosomal strands ay bumubuo ng mga siksik na spiral, bilang isang resulta kung saan sila ay nakikita (gamit ang isang maginoo na mikroskopyo) sa anyo ng mga rod, "mga hairpins". Ang buong dami ng genetic na impormasyon ay ipinamamahagi sa pagitan ng mga chromosome ng nucleus. Sa proseso ng pag-aaral ng mga ito, natukoy ang mga sumusunod na pattern:
♦ sa nuclei ng mga somatic cells (ibig sabihin, mga cell ng katawan, hindi sekswal) lahat ng indibidwal ng parehong species ay naglalaman ng parehong bilang ng mga chromosome na bumubuo hanay ng mga chromosome(Larawan 19);
kanin. 19. Mga kromosom ng iba't ibang uri ng halaman at hayop, na inilalarawan sa parehong sukat: 1,2 – amoeba; 3,4 - diatoms; 5–8, 18,19 – berdeng algae; 9 - lumipad agaric; 10 - linden; 11–12 – Drosophila; 13 - salmon; 14 – skerda (pamilya ng Asteraceae); 15 - halaman mula sa pamilyang aroid; 16 – corydalis butterfly; 17 – insekto mula sa pamilya ng balang; 20 – water strider bug; 21 – surot ng bulaklak; 22 – amphibian Ambystoma; 23 – aloe (pamilya ng liryo)
♦ ang bawat species ay nailalarawan sa pamamagitan ng sarili nitong chromosome set ayon sa kanilang bilang (halimbawa, ang isang tao ay may 46 chromosome, isang fruit fly ay may 8, isang roundworm ay may 4, isang crayfish ay may 196, isang kabayo ay may 66, at ang mais ay may 104);
♦ Ang mga chromosome sa nuclei ng somatic cells ay maaaring ipangkat sa mga pares na tinatawag na homologous chromosome batay sa kanilang pagkakatulad (sa istraktura at pag-andar);
♦ sa nuclei ng mga cell ng mikrobyo (gametes), ang bawat pares ng homologous chromosome ay naglalaman lamang ng isa, ibig sabihin, ang kabuuang hanay ng mga chromosome ay kalahati ng somatic cells;
♦ tinatawag ang isang solong hanay ng mga chromosome sa mga selula ng mikrobyo haploid at tinutukoy ng titik n, at sa somatic - diploid(2n).
Mula sa itaas ay malinaw na ang bawat pares ng homologous chromosome ay nabuo sa pamamagitan ng unyon ng paternal at maternal chromosome sa panahon ng fertilization, ibig sabihin, ang pagsasanib ng mga sex cell (gametes). Sa kabaligtaran, sa panahon ng pagbuo ng mga cell ng mikrobyo, mula sa bawat pares ng mga homologous chromosome ay isa lamang ang nakakapasok sa gamete.
Mga Chromosome magkaiba magkaiba ang mga homologous na pares sa laki at hugis (Larawan 20 at 21).
kanin. 20. Istraktura at mga uri ng chromosome: a – hitsura 1 – centromere; 2 - maikling balikat; 3 - mahabang balikat); panloob na istraktura ng parehong chromosome (1 – centromere; – mga molekula ng DNA); c – mga uri ng chromosome (1 – single-armed; different-armed; 3 – equal-armed: X – arm, V – centromere)
kanin. 21. Ang chromosome ay binubuo ng DNA at mga protina. Ang molekula ng DNA ay ginagaya. Dalawang magkaparehong DNA double helice ang nananatiling konektado sa sentromere na rehiyon. Ang mga kopyang ito ay nagiging magkahiwalay na chromosome sa paglaon sa panahon ng cell division
Sa katawan ng mga chromosome mayroong ang pangunahing constriction (tinatawag na sentromere), kung saan nakakabit ang mga thread fission spindles. Hinahati nito ang chromosome sa dalawa balikat Ang mga chromosome ay maaaring magkapantay na armado, hindi magkapantay na armado, o single-armed.
Kabanata 5. Metabolismo
§ 1. Metabolismo bilang isang pagkakaisa ng asimilasyon at dissimilation
Ang lahat ng mga cell at buhay na organismo ay bukas na mga sistema, iyon ay, sila ay nasa isang estado ng patuloy na pagpapalitan ng mga enerhiya at mga sangkap sa kapaligiran. May mga bukas na sistema sa walang buhay na kalikasan, ngunit ang kanilang pag-iral ay naiiba sa husay sa mga buhay na organismo. Isaalang-alang ang halimbawang ito: ang isang nasusunog na piraso ng katutubong asupre ay nasa estado ng pakikipagpalitan sa kapaligiran. Kapag ito ay nasusunog, ang O 2 ay nasisipsip, at ang SO 2 at enerhiya (sa anyo ng init) ay inilabas. Gayunpaman, sa kasong ito, ang isang piraso ng asupre bilang isang pisikal na katawan ay nawasak at nawawala ang pangunahing istraktura nito.
Para sa mga buhay na organismo, ang pakikipagpalitan sa kapaligiran ay lumalabas na isang kondisyon para sa pagpapanatili at pagpapanatili ng kanilang istrukturang organisasyon sa pamamagitan ng pag-renew ng sarili ng lahat ng mga sangkap at sangkap kung saan sila binubuo.
Ang metabolismo (metabolismo) ay isang hanay ng mga proseso na nagaganap sa mga buhay na organismo (pagkonsumo, pagbabago, akumulasyon at pagpapalabas ng mga sangkap at enerhiya) na nagsisiguro sa kanilang mahahalagang aktibidad, pag-unlad, paglago, pagpaparami. Sa panahon ng metabolic process, ang mga molecule na bumubuo sa mga cell ay pinaghiwa-hiwalay at na-synthesize; pag-renew ng cellular structures at intercellular substance.
Ang metabolismo ay batay sa magkakaugnay na mga proseso asimilasyon(anabolismo) at dissimilation(katabolismo). Sa panahon ng asimilasyon (plastic exchange), ang mga kumplikadong sangkap ay synthesize mula sa mga simple. Ito ay salamat sa ito na ang lahat ng mga organikong sangkap sa cell na kinakailangan para sa pagtatayo ng mga istrukturang bahagi nito, mga sistema ng enzyme, atbp. Ang asimilasyon ay palaging nagaganap sa paggasta ng enerhiya.
Sa panahon ng dissimilation (energy metabolism), ang mga kumplikadong organikong sangkap ay nahahati sa mas simple o hindi organikong mga bagay. Sa kasong ito, ang enerhiya ay inilabas, na ginugol ng cell upang maisagawa ang iba't ibang mga proseso na matiyak ang mahahalagang aktibidad nito (synthesis at transportasyon ng mga sangkap, mekanikal na gawain, atbp.).
Ang lahat ng mga nabubuhay na organismo ay maaaring nahahati sa dalawang pangkat: mga autotroph At heterotrophs, na naiiba sa mga pinagmumulan ng enerhiya at mga kinakailangang sangkap upang matiyak ang kanilang mahahalagang tungkulin.
Mga autotroph– mga organismo na nag-synthesize ng mga organikong compound mula sa mga di-organikong sangkap gamit ang enerhiya ng sikat ng araw (bilang mga phototroph– halaman, cyanobacteria) o enerhiya na nakuha mula sa oksihenasyon ng mineral (inorganic) na mga sangkap (tulad ng chemotrophs– sulfur bacteria, iron bacteria, atbp.). Dahil dito, nakapag-iisa silang lumikha ng mga sangkap na kinakailangan para sa kanilang aktibidad sa buhay.
§ 2. Dissimilation sa anaerobic at aerobic na mga organismo
Ang mga organismo ay maaaring nahahati sa dalawang pangkat at ayon sa likas na katangian ng dissimilation - aerobes At anaerobes. Aerobes (mula sa Greek. demonyo– hangin) ay nangangailangan ng libreng oxygen habang buhay. Sa anaerobes (Griyego. ac– negatibong butil) hindi kinakailangan. Sa kanila, ang dissimilation ay isinasagawa sa pamamagitan ng pagbuburo - walang oxygen, enzymatic na pagkasira ng organikong bagay na may pagbuo ng mas simpleng mga organikong sangkap at pagpapalabas ng enerhiya. Halimbawa:
♦ pagbuburo ng lactic acid:
C 6 H 12 O 6 + 2H 3 PO 4 + 2ADP → 2PH + 2ATP + 2H 2 O;
♦ alcoholic fermentation:
C 6 H 12 O 6 + 2PH + 2ADP → 2C 2 H 5 OH + 2CO 2 + 2ATP + 2H 2 O.
Ang mga sangkap na nabuo sa panahon ng pagbuburo ay organic at samakatuwid ay naglalaman pa rin ng maraming enerhiya.
kanin. 22. Mga ugnayan sa pagitan ng asimilasyon at dissimilation sa mga autotrophic at heterotrophic na organismo
Sa mga aerobic na organismo, sa panahon ng proseso ng paghinga sa mitochondria, ang isang kumpletong pagkasira ng mga organikong sangkap ay nangyayari (gamit ang O 2) sa mga produktong pangwakas na mahirap sa enerhiya na CO 2 at H 2 O at isang makabuluhang mas malaking halaga ng enerhiya ang inilabas:
C 6 H 12 0 6 (glucose) + 0 2 > 6C0 2 + 6H 2 0 + energy (dahil kung saan 38 ATP molecules ang na-synthesize).
Isaalang-alang natin sa anyo ng mga pangkalahatang iskema ang metabolismo ng mga autotrophic at heterotrophic na aerobic na organismo (Larawan 22).
Asimilasyon. Ang pinakamahalagang proseso nito ay potosintesis At biosynthesis ng mga protina.
Ang koneksyon ng organismo sa kapaligiran, mula sa isang physicochemical point of view, ay isang bukas na sistema, iyon ay, isang sistema kung saan ang mga biochemical na proseso ay nagpapatuloy. Ang mga panimulang sangkap ay nagmumula sa kapaligiran, at ang mga sangkap na patuloy ding nabubuo ay dinadala sa labas. Ang balanse sa pagitan ng bilis at konsentrasyon ng mga produkto ng mga multidirectional na reaksyon sa katawan ay kondisyon, haka-haka, dahil ang paggamit at pag-alis ng mga sangkap ay hindi tumitigil. Ang patuloy na koneksyon sa kapaligiran ay nagpapahintulot sa amin na isaalang-alang ang isang buhay na organismo bilang isang bukas na sistema.
Para sa lahat ng nabubuhay na selula, ang pinagmumulan ng enerhiya ay ang Araw. Ang mga cell ng halaman ay kumukuha ng enerhiya mula sa sikat ng araw sa tulong ng chlorophyll, ginagamit ito para sa mga reaksyon ng asimilasyon sa panahon ng proseso ng photosynthesis. Ang mga selula ng mga hayop, fungi, at bakterya ay gumagamit ng solar energy nang hindi direkta, sa panahon ng pagkasira ng mga organikong sangkap na na-synthesize ng mga halaman sa lupa.
Ang ilan sa mga sustansya ng cell ay nasira sa panahon ng cellular respiration, kaya nagbibigay ng enerhiya na kinakailangan para sa iba't ibang uri ng aktibidad ng cellular. Ang prosesong ito ay nagaganap sa mga organel na tinatawag na mitochondria. Ang Mitochondria ay binubuo ng dalawang lamad: ang panlabas, na naghihiwalay sa organelle mula sa cytoplasm, at ang panloob, na bumubuo ng maraming fold. Ang pangunahing produkto ng paghinga ay ATP. Umalis ito sa mitochondria at ginagamit bilang pinagmumulan ng enerhiya para sa maraming reaksiyong kemikal sa cytoplasm at cell membrane. Kung ang oxygen ay kinakailangan para sa cellular respiration, kung gayon ang paghinga ay tinatawag na aerobic, ngunit kung ang mga reaksyon ay nangyayari sa kawalan ng oxygen, pagkatapos ay nagsasalita tayo ng anaerobic respiration.
Para sa anumang uri ng trabaho na isinagawa sa isang cell, ang enerhiya ay ginagamit sa isa at tanging anyo - sa anyo ng enerhiya mula sa mga phosphate bond ng ATP. Ang ATP ay isang madaling mobile compound. Ang pagbuo ng ATP ay nangyayari sa panloob na lamad ng mitochondria. Ang ATP ay synthesize sa lahat ng mga cell sa panahon ng paghinga dahil sa enerhiya ng oksihenasyon ng carbohydrates, taba at iba pang mga organikong sangkap. Sa berdeng mga selula ng halaman, ang pangunahing halaga ng ATP ay na-synthesize sa mga chloroplast dahil sa solar energy. Sa panahon ng photosynthesis, gumagawa sila ng maraming beses na mas maraming ATP kaysa sa mitochondria. Nabubulok ang ATP sa pagkaputol ng mga bono ng phosphorus-oxygen at paglabas ng enerhiya. Nangyayari ito sa ilalim ng pagkilos ng enzyme ATPase sa panahon ng hydrolysis ng ATP - ang pagdaragdag ng tubig na may pag-aalis ng isang molekula ng phosphoric acid. Bilang resulta, ang ATP ay na-convert sa ADP, at kung ang dalawang molekula ng phosphoric acid ay nahati, pagkatapos ay sa AMP. Ang reaksyon ng pag-aalis ng bawat gramo-molekula ng acid ay sinamahan ng paglabas ng 40 kJ. Ito ay isang napakalaking output ng enerhiya, kaya naman ang phosphorus-oxygen bond ng ATP ay karaniwang tinatawag na macroergistic (high-energy).
Ang paggamit ng ATP sa mga reaksyon ng pagpapalitan ng plastik ay isinasagawa sa pamamagitan ng pagsasama sa kanila ng ATP hydrolysis. Ang mga molekula ng iba't ibang mga sangkap ay sinisingil ng enerhiya sa pamamagitan ng paglakip ng pangkat ng posporus na inilabas sa panahon ng hydrolysis mula sa molekula ng ATP, i.e. sa pamamagitan ng phosphorylation.
Ang kakaiba ng mga derivatives ng pospeyt ay hindi sila maaaring umalis sa cell, kahit na ang kanilang mga "discharged" na mga form ay malayang dumaan sa lamad. Dahil dito, ang mga phosphorylated molecule ay nananatili sa cell hanggang sa magamit sila sa mga naaangkop na reaksyon.
Ang kabaligtaran na proseso ng pag-convert ng ADP sa ATP ay nangyayari sa pamamagitan ng pagdaragdag ng isang molekula ng phosphoric acid sa ADP, pagpapalabas ng tubig at pagsipsip ng malaking halaga ng enerhiya.
Kaya, ang ATP ay isang unibersal at direktang mapagkukunan ng enerhiya para sa aktibidad ng cell. Lumilikha ito ng isang solong cellular pool ng enerhiya at ginagawang posible na muling ipamahagi at dalhin ito mula sa isang lugar ng cell patungo sa isa pa.
Ang paglipat ng pangkat ng pospeyt ay may mahalagang papel sa mga reaksiyong kemikal tulad ng pagpupulong ng mga macromolecule mula sa mga monomer. Halimbawa, ang mga amino acid ay maaaring pagsamahin sa mga peptide lamang pagkatapos ng dating phosphorylated. Ang mga mekanikal na proseso ng pag-urong o paggalaw, transportasyon ng isang dissolved substance laban sa isang gradient ng konsentrasyon at iba pang mga proseso ay kinabibilangan ng pagkonsumo ng enerhiya na nakaimbak sa ATP.
Ang proseso ng metabolismo ng enerhiya ay maaaring kinakatawan bilang mga sumusunod. Ang mga high-molecular na organikong sangkap sa cytoplasm ay enzymatically, sa pamamagitan ng hydrolysis, na na-convert sa mas simple kung saan sila binubuo: mga protina - sa amino acids, poly- at disaccharides - sa monosaccharides (+ glucose), taba sa glycerol at fatty acid. Walang mga proseso ng oxidative, maliit na enerhiya ang inilabas, na hindi ginagamit at napupunta sa thermal form. Karamihan sa mga cell ay gumagamit muna ng carbohydrates. Ang polysaccharides (starch sa mga halaman at glycogen sa mga hayop) ay na-hydrolyzed sa glucose. Ang glucose oxidation ay nangyayari sa tatlong yugto: glycolysis, oxidative decarboxylation (Krebs cycle - citric acid cycle) at oxidative phosphorylation (respiratory chain). Ang Glycolysis, bilang isang resulta kung saan ang isang molekula ng glucose ay nahahati sa dalawang molekula ng pyruvic acid na may paglabas ng dalawang molekula ng ATP, ay nangyayari sa cytoplasm. Sa kawalan ng oxygen, ang pyruvic acid ay na-convert sa alinman sa ethanol (fermentation) o lactic acid (anaerobic respiration).
Kapag ang glycolysis ay nangyayari sa mga selula ng hayop, ang anim na carbon na molekula ng glucose ay nasira sa dalawang molekula ng lactic acid. Ang prosesong ito ay multi-stage. Ito ay isinasagawa nang sunud-sunod sa pamamagitan ng 13 enzymes. Sa panahon ng alcoholic fermentation, dalawang molekula ng ethanol at dalawang molekula ng CO2 ay nabuo mula sa isang molekula ng glucose.
Ang glycolysis ay isang yugto na karaniwan sa anaerobic at aerobic na paghinga; Ang proseso ng oxygen-free na oksihenasyon, kung saan ang bahagi lamang ng enerhiya ng mga metabolite ay inilabas at ginagamit, ay pangwakas para sa mga anaerobic na organismo. Sa pagkakaroon ng oxygen, ang pyruvic acid ay pumasa sa mitochondria, kung saan, bilang isang resulta ng isang bilang ng mga sunud-sunod na reaksyon, ito ay ganap na na-oxidized nang aerobically sa H2O at CO2 na may sabay-sabay na phosphorylation ng ADP sa ATP. Sa kasong ito, ang dalawang molekula ng ATP ay ginawa ng glycolysis, dalawa ng Krebs cycle, at 34 ng respiratory chain. Ang netong ani para sa kumpletong oksihenasyon ng isang molekula ng glucose sa H2O at CO2 ay 38 mga molekula.
Kaya, sa mga aerobic na organismo, ang pangwakas na pagkabulok ng mga organikong sangkap ay isinasagawa sa pamamagitan ng pag-oxidize sa kanila ng atmospheric oxygen sa mga simpleng inorganic na sangkap: CO2 at H2O. Ang prosesong ito ay nagaganap sa cristae ng mitochondria. Sa kasong ito, ang maximum na halaga ng libreng enerhiya ay inilabas, isang makabuluhang bahagi nito ay nakalaan sa mga molekula ng ATP. Madaling makita na ang aerobic oxidation ay nagbibigay ng cell na may libreng enerhiya sa pinakamalaking lawak.
Bilang resulta ng catabolism, ang mga molekulang ATP na mayaman sa enerhiya ay naipon sa cell, at ang CO2 at labis na tubig ay inilalabas sa panlabas na kapaligiran.
Ang mga molekula ng asukal na hindi kailangan para sa paghinga ay maaaring maimbak sa cell. Ang mga labis na lipid ay maaaring masira, pagkatapos kung saan ang mga produkto ng kanilang pagkasira ay pumasok sa mitochondria bilang substrate para sa paghinga, o idineposito bilang mga reserba sa cytoplasm sa anyo ng mga fat droplet. Ang mga protina ay binuo mula sa mga amino acid na pumapasok sa cell. Ang synthesis ng protina ay nangyayari sa mga organel na tinatawag na ribosome. Ang bawat ribosome ay binubuo ng dalawang subparticle - malaki at maliit: ang parehong mga subparticle ay kinabibilangan ng mga molekula ng protina at mga molekula ng RNA.
Ang mga ribosom ay madalas na nakakabit sa isang espesyal na sistema ng lamad na binubuo ng mga cisterns at vesicles - ang tinatawag na endoplasmic reticulum (ER); sa mga selula na gumagawa ng maraming protina, ang endoplasmic reticulum ay kadalasang napakahusay na nabuo at natatakpan ng mga ribosom. Ang ilang mga enzyme ay epektibo lamang kung sila ay nakakabit sa isang lamad. Karamihan sa mga enzyme na kasangkot sa lipid synthesis ay matatagpuan dito. Kaya, ang endoplasmic reticulum ay parang isang uri ng cell workbench.
Bilang karagdagan, hinahati ng ER ang cytoplasm sa magkakahiwalay na mga compartment, ibig sabihin, pinaghihiwalay nito ang iba't ibang mga proseso ng kemikal na nangyayari nang sabay-sabay sa cytoplasm, at sa gayon ay binabawasan ang posibilidad na ang mga prosesong ito ay makagambala sa isa't isa.
Kadalasan ang mga produktong ginawa ng isang naibigay na cell ay ginagamit sa labas ng cell na iyon. Sa ganitong mga kaso, ang mga protina na na-synthesize sa mga ribosome ay dumadaan sa mga lamad ng endoplasmic reticulum at nakabalot sa mga lamad na vesicle na bumubuo sa kanilang paligid, na pagkatapos ay nahiwalay sa ER. Ang mga vesicle na ito, na pinatag at nakasalansan sa ibabaw ng bawat isa, tulad ng mga nakasalansan na pancake, ay bumubuo ng isang katangiang istraktura na tinatawag na Golgi complex, o Golgi apparatus. Sa panahon ng kanilang pananatili sa Golgi apparatus, ang mga protina ay sumasailalim sa ilang mga pagbabago. Kapag dumating ang oras para umalis sila sa cell, ang mga vesicle ng lamad ay sumanib sa lamad ng cell at walang laman, ibinubuhos ang kanilang mga nilalaman, ibig sabihin, ang pagtatago ay nangyayari sa pamamagitan ng exocytosis.
Ang Golgi apparatus ay gumagawa din ng mga lysosome - mga lamad sac na naglalaman ng mga digestive enzymes. Ang pag-alam kung paano gumagawa, nag-iimbak, at nag-e-export ang isang cell ng ilang partikular na protina, at kung paano nito "alam" kung aling mga protina ang dapat nitong panatilihin para sa sarili nito, ay isa sa mga pinakakaakit-akit na sangay ng modernong cytology.
Ang mga lamad ng anumang selula ay patuloy na gumagalaw at nagbabago. Ang mga lamad ng ER ay mabagal na gumagalaw sa buong cell. Ang mga indibidwal na seksyon ng mga lamad na ito ay naghihiwalay at bumubuo ng mga vesicle, na pansamantalang naging bahagi ng Golgi apparatus, at pagkatapos, sa pamamagitan ng proseso ng exocytosis, sumanib sa lamad ng cell.
Nang maglaon, ang materyal na lamad ay ibinalik sa cytoplasm, kung saan muli itong ginagamit.
Ang pagpapalitan ng mga sangkap na pumapasok sa cell o inilabas nito sa labas, pati na rin ang pagpapalitan ng iba't ibang mga signal sa micro- at macroenvironment, ay nangyayari sa pamamagitan ng panlabas na lamad ng cell. Tulad ng nalalaman, ang cell membrane ay isang lipid bilayer kung saan naka-embed ang iba't ibang mga molekula ng protina na kumikilos bilang mga dalubhasang receptor, mga channel ng ion, mga aparato na aktibong nagdadala o nag-aalis ng iba't ibang mga kemikal, intercellular contact, atbp. Sa malusog na mga eukaryotic cell, ang mga phospholipid ay ipinamamahagi sa ang lamad nang walang simetrya: ang panlabas na ibabaw ay binubuo ng sphingomyelin at phosphatidylcholine, ang panloob na ibabaw - ng phosphatidylserine at phosphatidylethanolamine. Ang pagpapanatili ng gayong kawalaan ng simetrya ay nangangailangan ng paggasta ng enerhiya. Samakatuwid, sa kaganapan ng pagkasira ng cell, impeksyon, o pagkagutom sa enerhiya, ang panlabas na ibabaw ng lamad ay pinayaman ng mga phospholipid na hindi pangkaraniwan para dito, na nagiging isang senyas para sa iba pang mga selula at mga enzyme tungkol sa pagkasira ng cell na may kaukulang reaksyon dito. Ang pinakamahalagang papel ay ginampanan ng natutunaw na anyo ng phospholipase A2, na sumisira sa arachidonic acid at lumilikha ng mga lysoform mula sa nabanggit na mga phospholipid. Ang arachidonic acid ay ang limitasyong link para sa paglikha ng mga nagpapaalab na tagapamagitan tulad ng eicosanoids, at mga proteksiyon na molekula - pentraxins (C-reactive protein (CRP), precursors ng amyloid proteins) - ay nakakabit sa mga lysoform sa lamad, na sinusundan ng pag-activate ng pandagdag. sistema sa kahabaan ng klasikal na landas at pagkasira ng cell.
Ang istraktura ng lamad ay nakakatulong na mapanatili ang mga katangian ng panloob na kapaligiran ng cell, ang mga pagkakaiba nito mula sa panlabas na kapaligiran. Tinitiyak ito ng pumipili na pagkamatagusin ng lamad ng cell at ang pagkakaroon ng mga aktibong mekanismo ng transportasyon sa loob nito. Ang kanilang pagkagambala bilang isang resulta ng direktang pinsala, halimbawa, sa pamamagitan ng tetrodotoxin, ouabain, tetraethylammonium, o sa kaso ng hindi sapat na supply ng enerhiya sa kaukulang "mga bomba" ay humahantong sa pagkagambala sa komposisyon ng electrolyte ng cell, mga pagbabago sa metabolismo nito, pagkagambala. ng mga tiyak na pag-andar - pag-urong, pagpapadaloy ng mga impulses ng paggulo, atbp. Ang pagkagambala sa mga channel ng cellular ion (calcium, sodium, potassium at chloride) sa mga tao ay maaari ding genetically na tinutukoy ng mga mutasyon sa mga gene na responsable para sa istraktura ng mga channel na ito. Ang tinatawag na channelopathies ay nagdudulot ng mga namamana na sakit ng nervous, muscular, at digestive system. Ang labis na pagpasok ng tubig sa cell ay maaaring humantong sa pagkalagot nito - cytolysis - dahil sa pagbubutas ng lamad kapag na-activate ang complement o isang pag-atake ng mga cytotoxic lymphocytes at natural killer cells.
Ang cell lamad ay may maraming mga receptor na nakapaloob dito - mga istruktura na, kapag pinagsama sa kaukulang mga tiyak na molekula ng pagbibigay ng senyas (ligands), nagpapadala ng isang senyas sa loob ng cell. Nangyayari ito sa pamamagitan ng iba't ibang mga regulatory cascade na binubuo ng mga enzymatically active molecules na sunud-sunod na isinaaktibo at sa huli ay nag-aambag sa pagpapatupad ng iba't ibang mga cellular program, tulad ng paglaki at paglaganap, pagkakaiba-iba, motility, pagtanda, at pagkamatay ng cell. Ang mga regulatory cascade ay medyo marami, ngunit ang kanilang bilang ay hindi pa ganap na natukoy. Ang sistema ng mga receptor at regulatory cascades na nauugnay sa kanila ay umiiral din sa loob ng cell; lumikha sila ng isang tiyak na network ng regulasyon na may mga punto ng konsentrasyon, pamamahagi at pagpili ng karagdagang landas ng signal depende sa functional na estado ng cell, ang yugto ng pag-unlad nito, at ang sabay-sabay na pagkilos ng mga signal mula sa iba pang mga receptor. Ang resulta nito ay maaaring inhibition o pagpapalakas ng signal, na nagdidirekta nito sa ibang regulatory pathway. Parehong ang receptor apparatus at signal transduction pathway sa pamamagitan ng mga regulatory cascade, halimbawa sa nucleus, ay maaaring maputol bilang resulta ng genetic defect na nangyayari bilang congenital defect sa organismal level o dahil sa somatic mutation sa isang partikular na uri ng cell. Ang mga mekanismong ito ay maaaring mapinsala ng mga nakakahawang ahente, lason, at magbago din sa panahon ng proseso ng pagtanda. Ang huling yugto nito ay maaaring isang pagkagambala sa mga function ng cell, ang mga proseso ng paglaganap at pagkita ng kaibhan nito.
Sa ibabaw ng mga selula mayroon ding mga molekula na may mahalagang papel sa mga proseso ng intercellular interaction. Maaaring kabilang dito ang mga cell adhesion protein, histocompatibility antigens, tissue-specific, differentiating antigens, atbp. Ang mga pagbabago sa komposisyon ng mga molecule na ito ay nagdudulot ng pagkagambala sa intercellular interactions at maaaring maging sanhi ng pag-activate ng mga naaangkop na mekanismo para sa pag-aalis ng naturang mga cell, dahil sila ay nagdudulot ng isang tiyak na panganib sa integridad ng katawan bilang reservoir ng impeksyon, lalo na sa viral, o bilang mga potensyal na nagpasimula ng paglaki ng tumor.
Paglabag sa supply ng enerhiya ng cell
Ang pinagmumulan ng enerhiya sa cell ay pagkain, pagkatapos ng pagkasira kung saan ang enerhiya ay inilabas sa mga huling sangkap. Ang pangunahing lugar ng paggawa ng enerhiya ay mitochondria, kung saan ang mga sangkap ay na-oxidized sa tulong ng mga enzyme ng respiratory chain. Ang oksihenasyon ay ang pangunahing tagapagtustos ng enerhiya, dahil bilang isang resulta ng glycolysis, hindi hihigit sa 5% ng enerhiya ang inilabas mula sa parehong dami ng mga substrate ng oksihenasyon (glucose), kumpara sa oksihenasyon. Humigit-kumulang 60% ng enerhiya na inilabas sa panahon ng oksihenasyon ay naipon ng oxidative phosphorylation sa mga high-energy phosphates (ATP, creatine phosphate), ang natitira ay nawala bilang init. Kasunod nito, ang mga high-energy phosphate ay ginagamit ng cell para sa mga proseso tulad ng pump operation, synthesis, division, movement, secretion, atbp. Mayroong tatlong mekanismo, ang pinsala na maaaring magdulot ng pagkagambala sa supply ng enerhiya ng cell: ang una ay ang mekanismo ng synthesis ng enerhiya metabolismo enzymes, ang pangalawa ay ang mekanismo ng oxidative phosphorylation, ang pangatlo ay ang mekanismo ng paggamit ng enerhiya.
Ang pagkagambala ng transportasyon ng elektron sa mitochondrial respiratory chain o pag-uncoupling ng ADP oxidation at phosphorylation na may pagkawala ng potensyal ng proton, ang puwersang nagtutulak para sa pagbuo ng ATP, ay humahantong sa isang pagpapahina ng oxidative phosphorylation sa paraang ang karamihan sa enerhiya ay nawawala bilang init at bumababa ang bilang ng mga high-energy compound. Ang uncoupling ng oksihenasyon at phosphorylation sa ilalim ng impluwensya ng adrenaline ay ginagamit ng mga selula ng mga homeothermic na organismo upang mapataas ang produksyon ng init habang pinapanatili ang isang pare-parehong temperatura ng katawan sa panahon ng paglamig o pagtaas nito sa panahon ng lagnat. Ang mga makabuluhang pagbabago sa istraktura ng mitochondrial at metabolismo ng enerhiya ay sinusunod sa thyrotoxicosis. Ang mga pagbabagong ito sa una ay nababaligtad, ngunit pagkatapos ng isang tiyak na punto sila ay nagiging hindi na mababawi: mitochondria fragment, nawasak o namamaga, nawawala ang cristae, nagiging vacuoles, at kalaunan ay nag-iipon ng mga sangkap tulad ng hyaline, ferritin, calcium, lipofuscin. Sa mga pasyenteng may scurvy, nagsasama ang mitochondria upang bumuo ng mga chondriosphere, posibleng dahil sa pagkasira ng lamad ng mga compound ng peroxide. Ang makabuluhang pinsala sa mitochondria ay nangyayari sa ilalim ng impluwensya ng ionizing radiation sa panahon ng pagbabago ng isang normal na cell sa isang malignant.
Ang mitochondria ay isang malakas na depot ng mga calcium ions, kung saan ang konsentrasyon nito ay ilang mga order ng magnitude na mas mataas kaysa sa cytoplasm. Kapag nasira ang mitochondria, ang calcium ay pumapasok sa cytoplasm, na nagiging sanhi ng pag-activate ng mga proteinase na may pinsala sa mga istruktura ng intracellular at pagkagambala sa mga pag-andar ng kaukulang cell, halimbawa, mga contracture ng calcium o kahit na "kamatayan ng calcium" sa mga neuron. Bilang resulta ng pagkagambala sa functional na kapasidad ng mitochondria, ang pagbuo ng mga compound ng free radical peroxide ay tumataas nang husto, na may napakataas na reaktibiti at samakatuwid ay nakakapinsala sa mahahalagang bahagi ng cell - nucleic acid, protina at lipid. Ang hindi pangkaraniwang bagay na ito ay sinusunod sa panahon ng tinatawag na oxidative stress at maaaring magkaroon ng negatibong kahihinatnan para sa pagkakaroon ng cell. Kaya, ang pinsala sa panlabas na lamad ng mitochondria ay sinamahan ng paglabas sa cytoplasm ng mga sangkap na nakapaloob sa intermembrane space, lalo na ang cytochrome C at ilang iba pang biologically active substance, na nag-trigger ng mga chain reaction na nagdudulot ng programmed cell death - apoptosis. Sa pamamagitan ng pagkasira ng mitochondrial DNA, ang mga libreng radikal na reaksyon ay pinipihit ang genetic na impormasyon na kinakailangan para sa pagbuo ng ilang mga respiratory chain enzymes, na partikular na ginawa sa mitochondria. Ito ay humahantong sa mas malaking pagkagambala sa mga proseso ng oxidative. Sa pangkalahatan, ang sariling genetic apparatus ng mitochondria, kumpara sa genetic apparatus ng nucleus, ay hindi gaanong protektado mula sa mga nakakapinsalang impluwensya na maaaring magbago sa genetic na impormasyon na naka-encode dito. Bilang isang resulta, ang dysfunction ng mitochondria ay nangyayari sa buong buhay, halimbawa, sa panahon ng proseso ng pagtanda, sa panahon ng malignant na pagbabagong-anyo ng cell, pati na rin laban sa background ng namamana na mitochondrial na mga sakit na nauugnay sa mutation ng mitochondrial DNA sa itlog. Sa kasalukuyan, higit sa 50 mitochondrial mutations ang inilarawan na nagdudulot ng namamana na mga degenerative na sakit ng nervous at muscular system. Ang mga ito ay ipinadala sa bata ng eksklusibo mula sa ina, dahil ang mitochondria ng tamud ay hindi bahagi ng zygote at, nang naaayon, ang bagong organismo.
Paglabag sa pangangalaga at paghahatid ng genetic na impormasyon
Ang cell nucleus ay naglalaman ng karamihan sa genetic na impormasyon at sa gayon ay tinitiyak ang normal na paggana nito. Sa pamamagitan ng selective gene expression, ito ay nag-coordinate ng aktibidad ng cell sa panahon ng interphase, nag-iimbak ng genetic na impormasyon, at muling nililikha at nagpapadala ng genetic material sa panahon ng cell division. Ang pagtitiklop ng DNA at RNA transcription ay nangyayari sa nucleus. Ang iba't ibang mga pathogenic na kadahilanan, tulad ng ultraviolet at ionizing radiation, libreng radical oxidation, mga kemikal, mga virus, ay maaaring makapinsala sa DNA. Tinatayang tumatagal ng 1 araw ang bawat cell ng isang warm-blooded na hayop. nawalan ng higit sa 10,000 base. Dito dapat tayong magdagdag ng mga paglabag kapag nangongopya sa panahon ng paghahati. Kung magpapatuloy ang mga pinsalang ito, hindi makakaligtas ang selula. Ang proteksyon ay nakasalalay sa pagkakaroon ng makapangyarihang mga sistema ng pag-aayos, tulad ng ultraviolet endonuclease, pagtitiklop ng pagkumpuni at mga sistema ng pag-aayos ng recombination, na pumapalit sa pinsala sa DNA. Ang mga genetic na depekto sa mga sistema ng pag-aayos ay nagdudulot ng pag-unlad ng mga sakit na dulot ng pagtaas ng sensitivity sa mga salik na pumipinsala sa DNA. Ito ay xeroderma pigmentosum, pati na rin ang ilang pinabilis na pag-iipon na mga sindrom, na sinamahan ng mas mataas na posibilidad na magkaroon ng mga malignant na tumor.
Ang sistema para sa pag-regulate ng mga proseso ng pagtitiklop ng DNA, transkripsyon ng messenger RNA (mRNA), at pagsasalin ng genetic na impormasyon mula sa mga nucleic acid sa istruktura ng mga protina ay medyo kumplikado at multi-level. Bilang karagdagan sa mga regulatory cascade na nagpapalitaw ng pagkilos ng mga transcription factor na may kabuuang bilang na higit sa 3000, na nag-a-activate ng ilang partikular na gene, mayroon ding multi-level na sistema ng regulasyon na pinapamagitan ng maliliit na RNA molecule (nakakasagabal sa RNA; RNAi). Ang genome ng tao, na binubuo ng humigit-kumulang 3 bilyong purine at pyrimidine base, ay naglalaman lamang ng 2% ng mga istrukturang gene na responsable para sa synthesis ng protina. Ang natitira ay nagbibigay ng synthesis ng mga regulatory RNA, na, kasabay ng mga salik ng transkripsyon, ay nagpapagana o humaharang sa gawain ng mga istrukturang gene sa antas ng DNA sa mga chromosome o nakakaimpluwensya sa mga proseso ng pagsasalin ng messenger RNA (mRNA) sa panahon ng pagbuo ng isang polypeptide molecule sa ang cytoplasm. Ang paglabag sa genetic na impormasyon ay maaaring mangyari kapwa sa antas ng mga istrukturang gene at ang bahagi ng regulasyon ng DNA na may kaukulang mga pagpapakita sa anyo ng iba't ibang mga namamana na sakit.
Kamakailan, maraming pansin ang naakit sa mga pagbabago sa genetic na materyal na nangyayari sa panahon ng indibidwal na pag-unlad ng isang organismo at nauugnay sa pagsugpo o pag-activate ng ilang mga seksyon ng DNA at chromosome dahil sa kanilang methylation, acetylation at phosphorylation. Ang mga pagbabagong ito ay nagpapatuloy sa mahabang panahon, kung minsan sa buong buhay ng organismo mula sa embryogenesis hanggang sa katandaan, at tinatawag na epigenomic heredity.
Ang paglaganap ng mga cell na may binagong genetic na impormasyon ay pinipigilan din ng mga system (mga kadahilanan) na kumokontrol sa mitotic cycle. Nakikipag-ugnayan sila sa cyclin-dependent protein kinases at sa kanilang mga catalytic subunits - cyclins - at hinaharangan ang cell mula sa pagdaan sa buong mitotic cycle, na humihinto sa paghahati sa hangganan sa pagitan ng presynthetic at synthetic phase (G1/S block) hanggang sa makumpleto ang pag-aayos ng DNA, at kung ito ay imposible, sinimulan nila ang mga naka-program na mga selula ng kamatayan. Kabilang sa mga salik na ito ang p53 gene, ang mutation na nagiging sanhi ng pagkawala ng kontrol sa paglaganap ng mga nabagong selula; ito ay sinusunod sa halos 50% ng mga kanser sa tao. Ang pangalawang checkpoint ng mitotic cycle ay nasa hangganan ng G2/M. Dito, ang tamang pamamahagi ng chromosomal na materyal sa pagitan ng mga cell ng anak na babae sa mitosis o meiosis ay kinokontrol gamit ang isang hanay ng mga mekanismo na kumokontrol sa cell spindle, center at centromeres (kinetochores). Ang hindi pagiging epektibo ng mga mekanismong ito ay humahantong sa pagkagambala sa pamamahagi ng mga chromosome o kanilang mga bahagi, na kung saan ay ipinahayag sa pamamagitan ng kawalan ng anumang chromosome sa isa sa mga cell ng anak na babae (aneuploidy), ang pagkakaroon ng isang karagdagang chromosome (polyploidy), ang paghihiwalay ng isang bahagi ng isang chromosome (pagtanggal) at ang paglipat nito sa isa pang chromosome (pagsasalin) . Ang ganitong mga proseso ay madalas na sinusunod sa panahon ng paglaganap ng malignantly degenerated at transformed cells. Kung nangyari ito sa panahon ng meiosis na may mga selulang mikrobyo, hahantong ito sa pagkamatay ng fetus sa maagang yugto ng pag-unlad ng embryonic, o sa pagsilang ng isang organismo na may sakit na chromosomal.
Ang hindi makontrol na paglaganap ng cell sa panahon ng paglaki ng tumor ay nangyayari bilang resulta ng mga mutasyon sa mga gene na kumokontrol sa paglaganap ng cell at tinatawag na oncogenes. Sa higit sa 70 kasalukuyang kilalang oncogenes, karamihan ay nabibilang sa mga bahagi ng regulasyon ng paglago ng cell, ang ilan ay kinakatawan ng mga salik ng transkripsyon na kumokontrol sa aktibidad ng gene, pati na rin ang mga salik na pumipigil sa paghahati at paglaki ng cell. Ang isa pang kadahilanan na naglilimita sa labis na pagpapalawak (pagkalat) ng mga proliferating na mga cell ay ang pag-ikli ng mga dulo ng chromosome - telomeres, na hindi ganap na gumagaya bilang isang resulta ng purong steric na interaksyon, samakatuwid, pagkatapos ng bawat cell division, ang telomeres ay pinaikli ng isang tiyak na bahagi ng mga base. Kaya, ang paglaganap ng mga selula ng isang pang-adultong organismo pagkatapos ng isang tiyak na bilang ng mga dibisyon (karaniwan ay mula 20 hanggang 100 depende sa uri ng organismo at edad nito) ay nauubos ang haba ng telomere at humihinto ang karagdagang pagtitiklop ng kromosom. Ang phenomenon na ito ay hindi nangyayari sa sperm epithelium, enterocytes at embryonic cells dahil sa pagkakaroon ng enzyme telomerase, na nagpapanumbalik ng haba ng telomere pagkatapos ng bawat dibisyon. Sa karamihan ng mga selula ng mga pang-adultong organismo, ang telomerase ay naharang, ngunit, sa kasamaang-palad, ito ay isinaaktibo sa mga selula ng tumor.
Ang koneksyon sa pagitan ng nucleus at cytoplasm at ang transportasyon ng mga sangkap sa parehong direksyon ay isinasagawa sa pamamagitan ng mga pores sa nuclear membrane na may pakikilahok ng mga espesyal na sistema ng transportasyon na kumonsumo ng enerhiya. Sa ganitong paraan, ang enerhiya at mga plastik na sangkap, ang mga molekula ng senyas (mga salik ng transkripsyon) ay dinadala sa nucleus. Ang reverse flow ay dinadala sa cytoplasm molecules ng mRNA at transfer RNA (tRNA), ribosomes na kailangan para sa synthesis ng protina sa cell. Ang parehong ruta ng transportasyon ng mga sangkap ay likas din sa mga virus, lalo na tulad ng HIV. Inilipat nila ang kanilang genetic na materyal sa nucleus ng host cell kasama ang karagdagang pagsasama nito sa host genome at ang paglipat ng bagong nabuong viral RNA sa cytoplasm para sa karagdagang synthesis ng mga protina ng mga bagong particle ng viral.
Paglabag sa mga proseso ng synthesis
Ang mga proseso ng synthesis ng protina ay nangyayari sa mga cisterns ng endoplasmic reticulum, malapit na konektado sa mga pores sa nuclear membrane, kung saan ang mga ribosome, tRNA at mRNA ay pumapasok sa endoplasmic reticulum. Dito, ang synthesis ng polypeptide chain ay isinasagawa, na kasunod na nakuha ang kanilang pangwakas na anyo sa agranular endoplasmic reticulum at ang lamellar complex (Golgi complex), kung saan sumasailalim sila sa post-translational modification at pinagsama sa mga molekula ng carbohydrate at lipid. Ang mga bagong nabuong molekula ng protina ay hindi nananatili sa lugar ng synthesis, ngunit sa pamamagitan ng isang kumplikadong regulated na proseso na tinatawag protinakinesis, ay aktibong inililipat sa nakahiwalay na bahagi ng cell kung saan gagawin nila ang kanilang nilalayon na function. Sa kasong ito, ang isang napakahalagang hakbang ay ang pag-istruktura ng inilipat na molekula sa isang naaangkop na pagsasaayos ng spatial na may kakayahang magsagawa ng likas na pag-andar nito. Ang istrukturang ito ay nangyayari sa tulong ng mga espesyal na enzyme o sa isang matrix ng mga dalubhasang molekula ng protina - mga chaperone, na tumutulong sa molekula ng protina, na bagong nabuo o nagbago dahil sa panlabas na impluwensya, upang makuha ang tamang three-dimensional na istraktura. Sa kaganapan ng isang masamang epekto sa cell, kapag may posibilidad ng pagkagambala sa istraktura ng mga molekula ng protina (halimbawa, na may pagtaas sa temperatura ng katawan, isang nakakahawang proseso, pagkalasing), ang konsentrasyon ng mga chaperone sa cell ay tumataas. matalas. Samakatuwid, ang mga naturang molekula ay tinatawag din mga protina ng stress, o mga protina ng heat shock. Ang paglabag sa istruktura ng isang molekula ng protina ay humahantong sa pagbuo ng mga chemically inert conglomerates, na idineposito sa cell o sa labas nito sa panahon ng amyloidosis, Alzheimer's disease, atbp. Minsan ang isang pre-structured na katulad na molekula ay maaaring magsilbi bilang isang matrix, at sa ito kaso, kung ang pangunahing structuring ay nangyayari nang hindi tama, ang lahat ng kasunod na mga molekula ay magkakaroon din ng depekto. Ang sitwasyong ito ay nangyayari sa tinatawag na mga sakit sa prion (scrapie sa tupa, rabid cows, kuru, Creutzfeldt-Jakob disease sa mga tao), kapag ang isang depekto sa isa sa mga protina ng lamad ng nerve cell ay nagiging sanhi ng kasunod na akumulasyon ng mga inert na masa sa loob ng cell at pagkagambala sa mahahalagang tungkulin nito.
Ang pagkagambala sa mga proseso ng synthesis sa isang cell ay maaaring mangyari sa iba't ibang yugto nito: RNA transcription sa nucleus, pagsasalin ng polypeptides sa ribosomes, post-translational modification, hypermethylation at glycosylation ng beige molecule, transportasyon at pamamahagi ng mga protina sa cell at ang kanilang pagtanggal sa labas. Sa kasong ito, ang isang tao ay maaaring obserbahan ang isang pagtaas o pagbaba sa bilang ng mga ribosome, ang pagkasira ng polyribosomes, pagpapalawak ng mga cisterns ng butil-butil na endoplasmic reticulum, pagkawala ng mga ribosome, at ang pagbuo ng mga vesicle at vacuoles. Kaya, kapag nalason ng isang maputlang grebe, ang RNA polymerase enzyme ay nasira, na nakakagambala sa transkripsyon. Ang lason ng diphtheria, sa pamamagitan ng pag-inactivate ng elongation factor, ay nakakagambala sa mga proseso ng pagsasalin, na nagiging sanhi ng pinsala sa myocardial. Ang sanhi ng pagkagambala sa synthesis ng ilang partikular na molekula ng protina ay maaaring mga nakakahawang ahente. Halimbawa, ang mga herpes virus ay pumipigil sa synthesis at pagpapahayag ng mga molekula ng MHC antigen, na nagpapahintulot sa kanila na bahagyang maiwasan ang immune control ng bacilli ng salot - ang synthesis ng mga tagapamagitan ng talamak na pamamaga. Ang hitsura ng hindi pangkaraniwang mga protina ay maaaring huminto sa kanilang karagdagang pagkasira at humantong sa akumulasyon ng hindi gumagalaw o kahit na nakakalason na materyal. Maaari itong, sa isang tiyak na lawak, ay mapadali sa pamamagitan ng pagkagambala sa mga proseso ng pagkabulok.
Pagkagambala sa mga proseso ng pagkabulok
Kasabay ng synthesis ng protina sa cell, ang pagkasira nito ay patuloy na nangyayari. Sa ilalim ng normal na mga kondisyon, ito ay may mahalagang regulatory at formative significance, halimbawa, sa panahon ng pag-activate ng mga hindi aktibong anyo ng enzymes, protein hormones, at mitotic cycle proteins. Ang normal na paglaki at pag-unlad ng cell ay nangangailangan ng isang pinong kontroladong balanse sa pagitan ng synthesis at pagkasira ng mga protina at organelles. Gayunpaman, sa proseso ng synthesis ng protina, dahil sa mga pagkakamali sa pagpapatakbo ng synthesizing apparatus, abnormal na pag-istruktura ng molekula ng protina, at pagkasira nito ng mga ahente ng kemikal at bacterial, isang medyo malaking bilang ng mga may sira na molekula ay patuloy na nabuo. Ayon sa ilang mga pagtatantya, ang kanilang bahagi ay halos isang katlo ng lahat ng synthesized na protina.
Ang mga selula ng mammalian ay may ilang pangunahing Mga paraan ng pagkasira ng protina: sa pamamagitan ng lysosomal protease (pentide hydrolases), calcium-dependent proteinases (endopeptidases) at ang proteasome system. Bilang karagdagan, mayroon ding mga espesyal na protina, tulad ng mga caspases. Ang pangunahing organelle kung saan ang pagkasira ng mga sangkap ay nangyayari sa mga eukaryotic cells ay ang lysosome, na naglalaman ng maraming hydrolytic enzymes. Dahil sa mga proseso ng endocytosis at iba't ibang uri ng autophagy sa lysosomes at phagolysosomes, parehong may sira na mga molekula ng protina at buong organelles ay nawasak: nasirang mitochondria, mga seksyon ng lamad ng plasma, ilang mga extracellular na protina, at ang mga nilalaman ng secretory granules.
Ang isang mahalagang mekanismo para sa pagkasira ng protina ay ang proteasome, isang multicatalytic proteinase na istraktura ng kumplikadong istraktura na naisalokal sa cytosol, nucleus, endoplasmic reticulum at sa cell membrane. Ang enzyme system na ito ay may pananagutan sa pagsira sa mga nasirang protina pati na rin sa malusog na mga protina na dapat alisin para sa normal na paggana ng cell. Sa kasong ito, ang mga protina na sisirain ay paunang pinagsama sa isang tiyak na polypeptide, ubiquitin. Gayunpaman, ang mga non-ubiquitinated na protina ay maaari ding bahagyang masira sa mga proteasome. Ang pagkasira ng mga molekula ng protina sa mga proteasome sa maikling polypeptides (pagproseso) kasama ang kanilang kasunod na pagtatanghal kasama ng mga molekula ng type I MHC ay isang mahalagang link sa immune control ng antigenic homeostasis sa katawan. Kapag ang function ng proteasome ay humina, ang mga nasira at hindi kinakailangang mga protina ay nag-iipon, na kasama ng pagtanda ng cell. Ang paglabag sa pagkasira ng mga protina na umaasa sa cyclin ay humahantong sa pagkagambala ng cell division, pagkasira ng mga secretory protein - sa pagbuo ng cystofibrosis. Sa kabaligtaran, ang pagtaas ng function ng proteasome ay kasama ng pag-ubos ng katawan (AIDS, cancer).
Sa genetically determined disorders ng pagkasira ng protina, ang organismo ay hindi mabubuhay at namamatay sa mga unang yugto ng embryogenesis. Kung ang pagkasira ng mga taba o carbohydrates ay nagambala, ang mga sakit sa imbakan (thesaurismosis) ay nangyayari. Sa kasong ito, ang isang labis na halaga ng ilang mga sangkap o produkto ng kanilang hindi kumpletong pagkasira - mga lipid, polysaccharides - ay naipon sa loob ng cell, na makabuluhang nakakapinsala sa pag-andar ng cell. Ito ay madalas na sinusunod sa mga selula ng epithelial ng atay (hepatocytes), mga neuron, fibroblast at macrophagocytes.
Ang mga nakuha na karamdaman ng mga proseso ng pagkasira ng mga sangkap ay maaaring lumitaw bilang isang resulta ng mga proseso ng pathological (halimbawa, protina, taba, karbohidrat at pagkabulok ng pigmentary) at sinamahan ng pagbuo ng mga hindi pangkaraniwang sangkap. Ang mga kaguluhan sa lysosomal proteolysis system ay humantong sa pagbaba ng adaptasyon sa panahon ng pag-aayuno o pagtaas ng stress, at sa paglitaw ng ilang mga endocrine dysfunctions - nabawasan ang mga antas ng insulin, thyroglobulin, cytokines at kanilang mga receptor. Ang kapansanan sa pagkasira ng protina ay nagpapabagal sa bilis ng paggaling ng sugat, nagiging sanhi ng pag-unlad ng atherosclerosis, at nakakaapekto sa immune response. Sa hypoxia, ang mga pagbabago sa intracellular pH, pinsala sa radiation, na nailalarawan sa pamamagitan ng pagtaas ng peroxidation ng mga lipid ng lamad, pati na rin sa ilalim ng impluwensya ng mga lysosomotropic na sangkap - bacterial endotoxins, metabolites ng nakakalason na fungi (sporofusarin), silicon oxide crystals - ang katatagan ng lysosome membrane mga pagbabago, ang mga aktibong lysosomal enzyme ay inilabas sa cytoplasm, na nagiging sanhi ng pagkasira ng mga istruktura ng cell at pagkamatay nito.
Kabanata 1
MGA BATAYAN NG CELL PHYSIOLOGY
I. Dudel
Plasma lamad . Ang mga selula ng hayop ay napapalibutan ng isang lamad ng plasma (Larawan 1.1). Tatalakayin natin ang istraktura nito, na halos kapareho sa istraktura ng maraming mga intracellular membrane, nang mas detalyado. Ang pangunahing matrix ng lamad ay binubuo ng mga lipid, higit sa lahat phosphatidylcholine. Ang mga lipid na ito ay binubuo ng isang hydrophilic head group kung saan nakakabit ang mahabang hydrophobic hydrocarbon chain. Sa tubig, ang mga naturang lipid ay kusang bumubuo ng isang bilayer film na 4-5 nm ang kapal, kung saan ang mga hydrophilic na grupo ay nakaharap sa aqueous medium, at ang mga hydrophobic hydrocarbon chain ay nakaayos sa dalawang hanay, na bumubuo ng isang anhydrous lipid phase. Ang mga cell lamad ay mga lipid bilayer ng ganitong uri at naglalaman ng mga glycolipids, cholesterol at phospholipids (Fig. 1.2). Ang hydrophilic na bahagi ng glycolipids ay nabuo ng oligosaccharides. Ang mga glycolipids ay palaging matatagpuan sa panlabas na ibabaw ng lamad ng plasma, na ang oligosaccharide na bahagi ng molekula ay nakatuon tulad ng isang buhok na nahuhulog sa kapaligiran. Nakakalat sa mga phospholipid sa halos pantay na dami, ang mga molekula ng kolesterol ay nagpapatatag sa lamad. Ang pamamahagi ng iba't ibang mga lipid sa panloob at panlabas na mga layer ng lamad ay hindi pareho, at kahit na sa loob ng isang layer ay may mga lugar kung saan ang ilang mga uri ng mga lipid ay puro. Itong hindi pantay na pamamahagi
kanin. 1.1. Schematic drawing ng isang cell na nagpapakita ng pinakamahalagang organelles
marahil ay may ilang, hindi pa malinaw, functional na kahalagahan.
Ang mga pangunahing functional na elemento na naka-embed sa medyo hindi gumagalaw na lipid matrix ng lamad ay mga ardilya(Larawan 1.2). Ang protina ayon sa timbang ay 25 hanggang 75% sa iba't ibang lamad, ngunit dahil ang mga molekula ng protina ay mas malaki kaysa sa mga molekulang lipid, ang 50% ng timbang ay katumbas ng ratio ng 1 molekula ng protina sa 50 mga molekula ng lipid. Ang ilang mga protina ay tumagos sa lamad mula sa panlabas hanggang sa panloob na ibabaw, habang ang iba ay naayos sa isang layer. Ang mga molekula ng protina ay karaniwang nakatuon upang ang kanilang mga hydrophobic na grupo ay naka-embed sa lipid membrane at ang mga polar hydrophilic group sa ibabaw ng lamad ay naka-embed sa aqueous phase. Maraming mga protina sa panlabas na ibabaw ng lamad ay glycoproteins; ang kanilang hydrophilic saccharide group ay nakaharap sa extracellular na kapaligiran.
Mga sistema ng lamad ng intracellular organelles .
Humigit-kumulang kalahati ng dami ng cell ay inookupahan ng mga organel na nakahiwalay sa cytosol ng mga lamad. Ang kabuuang lugar ng ibabaw ng mga lamad ng intracellular organelles ay hindi bababa sa 10 beses na mas malaki kaysa sa ibabaw ng lamad ng plasma. Ang pinakamalawak na ginagamit na sistema ng lamad ay endoplasmic reticulum, kumakatawan sa isang network

kanin. 1.2.Eskematiko na representasyon ng lamad ng plasma. Ang mga protina ay naka-embed sa phospholipid bilayer, na ang ilan sa mga ito ay tumatagos sa bilayer, habang ang iba ay naka-angkla lamang sa panlabas o panloob na layer.
mataas na convoluted tubes o sac-like na pahabang istruktura; ang malalaking lugar ng endoplasmic reticulum ay may mga ribosome; ang naturang reticulum ay tinatawag na butil-butil o magaspang (Larawan 1.1). Golgi apparatus Binubuo rin ang mga lamellae na nakagapos sa lamad, kung saan napuputol ang mga vesicle o vesicle (Larawan 1.1). Mga lysosome at peroxisome- Ito ay maliliit na dalubhasang vesicle. Sa lahat ng iba't ibang organel na ito, ang lamad at ang espasyong nasasakupan nito ay naglalaman ng mga tiyak na hanay ng mga enzyme; Sa loob ng mga organelles, ang mga espesyal na metabolic na produkto ay naipon, na ginagamit upang isagawa ang iba't ibang mga pag-andar ng mga organelles.
CoreAt mitochondria naiiba sa bawat isa sa mga organel na ito ay napapalibutan ng dalawang lamad. Ang nucleus ay responsable para sa kinetic control ng metabolismo; ang nakatiklop na panloob na lamad ng mitochondria ay ang site ng oxidative metabolism; dito, dahil sa oksihenasyon ng pyruvate o fatty acids, na-synthesize ang high-energy compound na adenosine triphosphate (ATP, o ATP).
Cytoskeleton . Ang cytoplasm na nakapalibot sa mga organel ay hindi maaaring ituring na amorphous sa anumang paraan; ito ay tinagos ng isang cytoskeletal network. Ang cytoskeleton ay binubuo ng mga microtubule, actin filament at intermediate filament (Larawan 1.1). Microtubule magkaroon ng panlabas na diameter na mga 25 nm; sila ay nabuo, tulad ng isang regular na polimer, bilang isang resulta ng pagpupulong ng mga molekula ng protina ng tubulin. Actin filament- Ang mga contractile fibers, na matatagpuan sa malapit na lamad na layer at sa buong cell, ay pangunahing nakikibahagi sa mga prosesong nauugnay sa paggalaw. Mga intermediate na filament binubuo ng mga bloke ng iba't ibang komposisyon ng kemikal sa iba't ibang uri ng mga selula; bumubuo sila ng iba't ibang koneksyon sa pagitan ng dalawang iba pang elemento ng cytoskeletal na binanggit sa itaas. Ang mga organelles at ang plasma membrane ay nauugnay din sa cytoskeleton, na hindi lamang nagpapanatili ng hugis ng cell at ang posisyon ng mga organelles sa loob nito, ngunit tinutukoy din ang mga pagbabago sa hugis ng cell at ang kadaliang kumilos.
Cytosol . Halos kalahati ng dami ng cell ay inookupahan ng cytosol. Dahil ito ay humigit-kumulang 20% na protina ayon sa timbang, ito ay higit pa sa isang gel kaysa sa isang may tubig na solusyon. Maliit na molecule, kabilang ang organic at inorganic mga ion, natunaw sa may tubig na bahagi. Ang pagpapalitan ng mga ion ay nangyayari sa pagitan ng cell at ng kapaligiran (extracellular space); ang mga metabolic na prosesong ito ay tatalakayin sa susunod na seksyon. Ang konsentrasyon ng mga ions sa extracellular space ay pinananatili na may malaking katumpakan sa isang pare-parehong antas; ang intracellular na konsentrasyon ng bawat ion ay mayroon ding tiyak na antas, naiiba sa labas ng selula (Talahanayan 1.1). Ang pinaka-masaganang cation sa extracellular na kapaligiran ay Na+ sa cell ang konsentrasyon nito ay higit sa 10 beses na mas mababa. Sa kabaligtaran, ang konsentrasyon ng K + ay pinakamataas sa loob ng cell sa labas ng cell ito ay mas mababa ng higit sa isang order ng magnitude. Ang pinakamalaking gradient sa pagitan ng extracellular at intracellular na konsentrasyon ay umiiral para sa Ca 2+, ang konsentrasyon ng mga libreng ion sa loob ng cell ay hindi bababa sa 10,000 beses na mas mababa kaysa sa labas nito. Hindi lahat ng mga ion ay natutunaw sa cytosol; Halimbawa, sa kaso ng Ca 2+, ang mga nakagapos na ion ay mas marami kaysa sa mga libre. Karamihan sa mga cytosolic na protina ay mga enzyme, na may partisipasyon kung saan isinasagawa ang maraming mga proseso ng intermediate metabolism: glycolysis at gluconeogenesis, synthesis o pagkasira ng mga amino acid, synthesis ng protina sa ribosomes (Fig. 1.1). Ang cytosol ay naglalaman din ng mga patak ng lipid at mga butil ng glycogen, na nagsisilbing mga reserba ng mahahalagang molekula.
Talahanayan 1.1.Mga konsentrasyon ng intra- at extracellular ion sa mga selula ng kalamnan ng mga homeothermic na hayop. A – - "mataas na molekular na timbang na mga cellular anion"
|
Konsentrasyon sa intracellular |
Extracellular na konsentrasyon |
||
|
Na+ |
12 mmol/l |
Na+ |
145 mmol/l |
|
155 mmol/l |
K+ |
4 mmol/l |
|
|
Ca 2+ |
10 –7 10 –8 mmol/l |
Ca 2+ |
2 mmol/l |
|
C l – |
4 mmol/l |
C1 – |
120 mmol/l |
|
HCO 3 – |
8 mmol/l |
HCO 3 – |
27 mmol/l |
|
A – |
155 mmol/l |
Ang iba mga kasyon |
5 mmol/l |
|
Potensyal sa pagpapahinga –90 mV |
|||
1.2. Pagpapalitan ng mga sangkap sa pagitan ng cell at ng kapaligiran
Maikling inilarawan namin ang istraktura ng cell upang magamit namin ang paglalarawang ito upang suriin ang mga pangunahing kaalaman ng cellular physiology. Sa anumang kaso, ang isang cell ay maaaring ituring na isang static na entity, dahil mayroong patuloy na pagpapalitan ng mga sangkap sa pagitan ng iba't ibang mga intracellular compartment, gayundin sa pagitan ng mga compartment at kapaligiran. Ang mga istruktura ng cell ay nasa dynamic na balanse, at ang mga pakikipag-ugnayan ng mga cell sa isa't isa at sa panlabas na kapaligiran ay isang kinakailangang kondisyon para sa pagpapanatili ng buhay ng isang gumaganang organismo. Sa kabanatang ito, titingnan natin ang mga pangunahing mekanismo ng naturang palitan. Sa mga susunod na kabanata ang mga mekanismong ito ay isasaalang-alang bilang inilapat sa nerve cell at sa mga function nito;
gayunpaman, ang parehong mga mekanismo ay sumasailalim sa paggana ng lahat ng iba pang mga organo.
Pagsasabog.Ang pinakasimpleng proseso ng paglipat ng bagay ay pagsasabog. Sa mga solusyon (o mga gas), ang mga atomo at molekula ay malayang gumagalaw, at ang mga pagkakaiba sa mga konsentrasyon ay balanse dahil sa pagsasabog. Isaalang-alang natin ang dalawang volume na puno ng likido o gas (Larawan 1.3), kung saan ang mga sangkap ay may mga konsentrasyon c 1 at c 2 at pinaghihiwalay ng isang layer na may surface area A at kapal d. Daloy ng bagay m sa panahon t inilarawan Ang unang batas ng pagsasabog ni Fick:
dm/ dt= DA/ d ( C 1 –C 2)=DA/ dD C(1)
kung saan ang D ay ang diffusion coefficient, pare-pareho para sa isang partikular na substance, solvent at temperatura. Sa isang mas pangkalahatang anyo para sa pagkakaiba sa konsentrasyon dc sa layo dx
dm/dt= –D A dc/dx,(2)
ang daloy sa seksyon A ay proporsyonal sa gradient ng konsentrasyon dc/dx . Ang minus sign ay lilitaw sa equation dahil ang pagbabago sa konsentrasyon sa x direksyon ay negatibo.
Ang pagsasabog ay ang pinakamahalagang proseso dahil sa kung saan ang karamihan sa mga molekula sa mga may tubig na solusyon ay gumagalaw sa maikling distansya. Nalalapat din ito sa kanilang paggalaw sa cell hangga't hindi nahahadlangan ng mga lamad ang pagsasabog. Maraming mga substance ang maaaring malayang kumalat sa mga lipid membrane, lalo na ang tubig at mga natunaw na gas tulad ng O 2 at CO 2 . Nalulusaw sa taba

kanin. 1.3.Quantitative diffusion scheme. Ang dalawang puwang ay pinaghihiwalay ng isang layer ng kapaldat lugar A. C; – mataas na konsentrasyon ng mga particle sa kaliwang bahagi ng volume, C: – mababang konsentrasyon ng mga particle sa kanan mga bahagi, kulay rosas na ibabaw– gradient ng konsentrasyon sa layer ng pagsasabog. Daloy ng pagsasabog dm/dt – tingnan equation (1)
ang mga sangkap ay nagkakalat din ng mabuti sa pamamagitan ng mga lamad; Nalalapat din ito sa mga polar molecule na may medyo maliit na sukat, tulad ng ethanol at urea, habang ang mga asukal ay dumaan sa lipid layer nang may kahirapan. Kasabay nito, ang mga layer ng lipid ay halos hindi natatagusan sa mga sisingilin na molekula, kabilang ang kahit na mga inorganic na ion. Para sa mga non-electrolytes, ang diffusion equation (1) ay karaniwang binabago sa pamamagitan ng pagsasama-sama ng mga katangian ng lamad at ng diffusing substance sa isa parameter-permeability (P):
dm/dt=P AD c.(3)
Sa Fig. 1.4 ang inihambing permeability (P) ng lipid membrane para sa iba't ibang molekula.
Pagsasabog sa pamamagitan ng mga pores ng lamad . Ang lamad ng plasma (at iba pang mga lamad ng cell) ay natatagusan hindi lamang sa mga sangkap na kumakalat sa layer ng lipid, kundi pati na rin sa maraming mga ion, asukal, amino acid at nucleotides. Ang mga sangkap na ito ay tumatawid sa lamad sa pamamagitan ng mga pores na nabuo mga protina ng transportasyon, nakalubog sa lamad. Sa loob ng naturang mga protina mayroong isang channel na puno ng tubig na may diameter na mas mababa sa 1 nm, kung saan maaaring magkalat ang maliliit na molekula. Gumagalaw sila sa isang gradient ng konsentrasyon, at kung nagdadala sila ng singil, kung gayon ang kanilang paggalaw sa mga channel ay kinokontrol din ng potensyal ng lamad. Ang mga channel ng lamad ay medyo pumipili

kanin. 1.4.Pagkamatagusin ng mga artipisyal na lipid bilayer sa iba't ibang mga sangkap
may kaugnayan sa uri ng mga molekula na maaaring dumaan sa kanila, mayroong, halimbawa, potassium, sodium at calcium channels, na ang bawat isa ay hindi natatagusan sa halos anumang ion maliban sa isang tiyak. ganyan pagpili ay tinutukoy ng singil o istraktura ng mga nagbubuklod na site sa mga dingding ng channel, na nagpapadali sa transportasyon ng isang tiyak na molekula at pinipigilan ang pagtagos ng iba pang mga sangkap sa pamamagitan ng channel (Fig. 1.5, A) .
Sa likod ng ugali mga channel ng ion ng lamad madaling obserbahan, dahil ang kasalukuyang nagmumula sa paggalaw ng mga ions ay maaaring masukat, kahit na para sa isang solong channel. Ipinakita na ang mga channel ay kusang at may mataas na dalas na nagbabago ng kanilang estado mula bukas hanggang sarado. Ang potassium channel ay nailalarawan sa pamamagitan ng kasalukuyang mga pulso na may amplitude na humigit-kumulang 2 pA (2 10 –12 A) at isang tagal ng ilang millisecond (tingnan ang Fig. 2.12, p. 37) [3]. Sa panahong ito, sampu-sampung libong mga ion ang dumadaan dito. Ang paglipat ng mga protina mula sa isang conform patungo sa isa pa ay pinag-aralan gamit ang X-ray diffraction, Mössbauer spectroscopy at nuclear magnetic resonance (NMR). Sa gayon, ang mga protina ay napaka-dynamic, mobile na istruktura, at ang channel na dumadaan sa protina ay hindi lamang isang matibay, puno ng tubig na tubo (Larawan 1.5, A), ngunit isang labirint ng mabilis na gumagalaw na mga molekular na grupo at mga singil. Ang dynamic na katangian ng channel ay makikita sa profile ng enerhiya ng channel, ipinapakita sa Fig. 1.5, B. Dito, ang abscissa axis ay kumakatawan sa haba ng channel mula sa panlabas na solusyon na may konsentrasyon ng ion C 0 at potensyal na 0 hanggang sa panloob na solusyon na may konsentrasyon C 1 at potensyal na E. Ang ordinate axis

kanin. 1.5.A. Schematic ng isang protina na bumubuo ng potassium channel na naka-embed sa lipid bilayer ng plasma membrane. Apat na negatibong singil ang naayos sa "pader" ng channel. B. Schematic energy profile ng channel na ipinapakita sa Fig. A. Ang ordinate axis ay nagpapakita ng mga halaga ng kinetic energy na kinakailangan para sa pagpasa ng channel; kasama ang abscissa axis - ang distansya sa pagitan ng panloob at panlabas na ibabaw ng lamad. Ang energy minima ay tumutugma sa mga nagbubuklod na site ng mga positibong sisingilin na mga ion na may mga nakapirming negatibong singil sa dingding ng channel. Ang energy maxima ay tumutugma sa diffusion obstacles sa channel. Ang conformation ng channel protein ay ipinapalagay na spontaneously oscillate; ang mga pagpipilian sa profile ng enerhiya ay ipinapakita na may mga solid at dashed na linya; ang mga oscillation na ito ay lubos na nagpapadali sa pagbubuklod ng mga ion kapag nalampasan ang energy barrier (binago)
ang mga antas ng enerhiya ng ion sa mga site na nagbubuklod ng channel ay ipinakita; ang peak sa graph ay kumakatawan sa permeability barrier na dapat lampasan ng enerhiya ng ion upang tumagos sa channel, at ang "dip" sa graph ay kumakatawan sa isang medyo stable na estado (binding). Sa kabila ng balakid ng energy peak, ang ion ay maaaring tumagos sa channel kung ang energy profile ay kusang nagbabago; ang ion ay maaaring biglang mahanap ang sarili nito "sa kabilang panig" ng peak ng enerhiya at maaaring magpatuloy na lumipat sa cell. Depende sa singil, laki at antas ng hydration ng ion at ang kakayahang magbigkis sa mga istruktura ng pader ng channel, ang profile ng enerhiya ng channel ay nag-iiba para sa iba't ibang mga ion, na maaaring ipaliwanag ang pagpili ng mga indibidwal na uri ng channel.
Diffusion equilibrium ng mga ion . Ang pagsasabog ng iba't ibang mga ion sa pamamagitan ng mga channel ng lamad ay dapat na humantong sa pag-aalis ng mga pagkakaiba sa mga konsentrasyon sa pagitan ng mga extra- at intracellular na kapaligiran. Bilang, gayunpaman, ay makikita mula sa talahanayan. 1.1, nananatili ang gayong mga pagkakaiba, samakatuwid, dapat mayroong ilan punto ng balanse sa pagitan ng diffusion at iba pang mga proseso ng transportasyon sa buong lamad. Ang susunod na dalawang seksyon ay tumatalakay sa mga paraan kung saan nakakamit ang ekwilibriyong ito. Sa kaso ng mga ions, ang diffusion equilibrium ay apektado ng kanilang singil. Ang pagsasabog ng mga uncharged na molekula ay sinisiguro ng pagkakaiba sa mga konsentrasyon dc , at kapag ang mga konsentrasyon ay napantayan, ang transportasyon mismo ay hihinto. Ang mga naka-charge na particle ay karagdagang apektado ng electric field. Halimbawa, kapag ang isang potassium ion ay umalis sa isang cell kasama ang gradient ng konsentrasyon nito, nagdadala ito ng isang positibong singil. Kaya, ang intracellular na kapaligiran ay nagiging mas negatibong sisingilin, na nagreresulta sa isang potensyal na pagkakaiba sa buong lamad. Pinipigilan ng intracellular negative charge ang mga bagong potassium ions na umalis sa cell, at ang mga ions na umalis sa cell ay higit pang magpapataas ng charge sa membrane. Ang daloy ng mga potassium ions ay humihinto kapag ang pagkilos ng electric field ay nagbabayad para sa diffusion pressure dahil sa pagkakaiba sa konsentrasyon. Ang mga ion ay patuloy na dumadaan sa lamad, ngunit sa pantay na dami sa magkabilang direksyon. Samakatuwid, para sa isang naibigay na pagkakaiba sa mga konsentrasyon ng ion sa lamad, mayroong potensyal ng ekwilibriyo E ion kung saan huminto ang daloy ng mga ions sa lamad. Ang potensyal ng ekwilibriyo ay madaling matukoy gamit Nernst equation:
Eion= RT/ zF* lnC out/ C sa(4)
kung saan si R – gas constant, T – absolute temperature, z – ion valency (negatibo para sa anion) C out - konsentrasyon ng extracellular ion, C sa - konsentrasyon ng intracellular ion, F Numero ng Faraday. Kung papalitan natin ang mga constant sa equation, pagkatapos ay sa temperatura ng katawan (T = 310 K) ang potensyal ng balanse para sa mga potassium ions E K ay katumbas ng:
Ek= –61 mB log /(5)
Kung [ K + out ]/[ K + in ] = 39, tulad ng sumusunod mula sa talahanayan. 1.1 noon
Ek= –61 m B log 39= –97 mV.
Sa katunayan, ito ay natagpuan na ang lahat ng mga cell ay may lamad potensyal; sa mammalian muscle cells ang level nito ay humigit-kumulang -90 mV. Depende sa mga kondisyon at kamag-anak na konsentrasyon ng ion, ang mga cell ay maaaring magkaroon ng potensyal na lamad mula -40 hanggang -120 mV. Para sa cell sa halimbawa sa itaas (Talahanayan 1.1) potensyal na magpahinga, katumbas ng humigit-kumulang -90 mV, ay nagpapahiwatig na ang mga flux ng potassium ions sa pamamagitan ng mga channel ng lamad ay humigit-kumulang sa equilibrium. Hindi ito nakakagulat, dahil sa isang resting lamad ang bukas na estado ng mga channel ng potasa ay malamang, i.e. ang lamad ay pinakapermeable sa potassium ions. Ang potensyal ng lamad, gayunpaman, ay tinutukoy ng mga daloy ng iba pang mga ion.
Ang kadalian kung saan ang mga uncharged na particle ay maaaring kumalat sa lamad ay binibilang sa equation (3). Pagkamatagusin sa mga sisingilin na particle ay inilarawan ng isang bahagyang mas kumplikadong equation:
P= m RT/ dF(6)
saan m- kadaliang kumilos ng ion sa lamad, d - kapal ng lamad, isang R, T at F – kilalang thermodynamic constants. Ang mga halaga ng pagkamatagusin para sa iba't ibang mga ion na tinutukoy sa ganitong paraan ay maaaring magamit upang kalkulahin ang potensyal ng lamad Em , kapag ang potassium, sodium at chlorine ions ay dumaan sa lamad nang sabay-sabay (na may permeability P K, P Na at P Cl ayon sa pagkakabanggit). Ipinapalagay na ang potensyal ay bumaba nang pantay sa lamad, upang ang lakas ng field ay pare-pareho. Sa kasong ito nalalapat ito Goldman equation, o pare-pareho ang field equation :
Em= R T/ F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl )(7)
Para sa karamihan ng mga lamad ng cell P K humigit-kumulang 30 beses na mas mataas kaysa sa P Na (tingnan din ang seksyon 1.3). Kamag-anak na halaga P Cl malaki ang pagkakaiba-iba; para sa maraming lamad P Cl maliit kumpara kay R K , gayunpaman para sa iba (halimbawa, sa mga kalamnan ng kalansay) P Cl , makabuluhang mas mataas kaysa sa P K.
Aktibong transportasyon, sodium pump . Inilalarawan ng nakaraang seksyon ang passive diffusion ng mga ion at ang nagresultang potensyal ng lamad sa ibinigay na intra- at extracellular na mga konsentrasyon ng ion. Gayunpaman, bilang isang resulta ng prosesong ito, ang konsentrasyon ng mga ions sa loob ng cell ay hindi awtomatikong nagpapatatag, dahil ang lamad
ang potensyal ay bahagyang mas electronegative kaysa EK, at marami kumpara sa E Na (mga +60 mV). Dahil sa pagsasabog, ang mga konsentrasyon ng intracellular ion, hindi bababa sa potassium at sodium, ay dapat na katumbas ng mga extracellular. Ang katatagan ng gradient ng ion ay nakakamit sa pamamagitan ng aktibong transportasyon: ang mga protina ng lamad ay nagdadala ng mga ion sa buong lamad laban sa mga gradient ng kuryente at/o konsentrasyon, na kumukonsumo ng metabolic energy para dito. Ang pinakamahalagang proseso ng aktibong transportasyon ay ang trabaho Na/K – isang bomba na umiiral sa halos lahat ng mga cell;
ang pump ay nagbobomba ng sodium ions palabas ng cell habang sabay-sabay na nagbobomba ng potassium ions sa cell. Tinitiyak nito ang isang mababang intracellular na konsentrasyon ng mga sodium ions at isang mataas na konsentrasyon ng mga potassium ions (Talahanayan 1.1). Ang gradient ng konsentrasyon ng mga sodium ions sa lamad ay may mga tiyak na pag-andar na nauugnay sa paghahatid ng impormasyon sa anyo ng mga electrical impulses (tingnan ang Seksyon 2.2), pati na rin ang pagpapanatili ng iba pang mga aktibong mekanismo ng transportasyon at regulasyon ng dami ng cell (tingnan sa ibaba). Samakatuwid, hindi nakakagulat na higit sa 1/3 ng enerhiya na natupok ng isang cell ay ginugol sa Na / K pump, at sa ilan sa mga pinaka-aktibong mga cell hanggang sa 70% ng enerhiya ay ginugol sa operasyon nito.
Ang Na/K transport protein ay isang ATPase. Sa panloob na ibabaw ng lamad, sinisira nito ang ATP sa ADP at pospeyt (Larawan 1.6). Ang enerhiya ng isang molekula ng ATP ay ginagamit upang maghatid ng tatlong sodium ions mula sa cell at sabay-sabay na dalawang potassium ions sa cell, ibig sabihin, sa kabuuan, isang positibong singil ang inalis mula sa cell sa isang cycle. Kaya, ang Na/K pump ay electrogenic(lumilikha ng electrical current sa buong lamad), na humahantong sa pagtaas ng electronegativity ng potensyal ng lamad ng humigit-kumulang 10 mV. Ginagawa ng transport protein ang operasyong ito sa mataas na bilis: mula 150 hanggang 600 sodium ions bawat segundo. Ang pagkakasunud-sunod ng amino acid ng transport protein ay kilala, ngunit ang mekanismo ng kumplikadong metabolic transport na ito ay hindi pa malinaw. Ang prosesong ito ay inilarawan gamit ang mga profile ng enerhiya ng paglilipat ng sodium o potassium ions ng mga protina (Larawan 1.5,5). Sa likas na katangian ng mga pagbabago sa mga profile na ito, na nauugnay sa patuloy na pagbabago sa conformation ng transport protein (isang proseso na nangangailangan ng enerhiya), maaaring hatulan ng isa ang stoichiometry ng palitan: dalawang potassium ions ay ipinagpapalit para sa tatlong sodium ions.
Na/K pump, katulad ng nakahiwalay Na+ /K + -dependent membrane ATPase, partikular na hinarang ng cardiac glycoside ouabain (strophanthin). Dahil ang operasyon ng Na/K pump ay isang multi-stage na kemikal na reaksyon, ito, tulad ng lahat ng kemikal na reaksyon, ay higit na nakadepende sa temperatura, na

kanin. 1.6.Diagram ng isang Na/K pump-ATPase (nalulubog sa lipid bilayer ng plasma membrane), na sa isang cycle ay nag-aalis ng tatlong Na + ions mula sa cell laban sa mga potensyal at gradient ng konsentrasyon at nagdadala ng dalawang K ion sa cell + . Sa prosesong ito, ang isang molekula ng ATP ay nahahati sa ADP at pospeyt. Ang diagram ay nagpapakita ng ATPase bilang isang dimer na binubuo ng isang malaki (functional) at maliit na mga subunit; sa lamad ito ay umiiral bilang isang tetramer na nabuo ng dalawang malaki at dalawang maliit na subunit
ipinapakita sa Fig. 1.7. Dito, ang pagkilos ng mga sodium ions mula sa mga selula ng kalamnan ay ipinapakita na may kaugnayan sa oras; ito ay halos katumbas ng daloy ng sodium ions na pinapamagitan ng pagpapatakbo ng Na/K pump, dahil ang passive flow ng sodium ions laban sa konsentrasyon at mga potensyal na gradient ay napakaliit. Kung ang gamot ay pinalamig ng humigit-kumulang 18°C, ang daloy ng sodium ions mula sa cell ay mabilis na bababa ng 15 beses, at kaagad pagkatapos ng pag-init ay maibabalik ito sa orihinal na antas nito. Ang pagbaba sa daloy ng mga sodium ions mula sa cell ay ilang beses na mas malaki kaysa sa kung saan ay tumutugma sa pagdepende sa temperatura ng proseso ng pagsasabog o isang simpleng kemikal na reaksyon. Ang isang katulad na epekto ay sinusunod kapag ang metabolic energy ay naubos bilang resulta ng pagkalason ng dinitrophenol (DNP) (Larawan 1.7,5). Dahil dito, ang daloy ng mga sodium ions mula sa cell ay sinisiguro ng isang reaksyon na umaasa sa enerhiya - isang aktibong bomba. Ang isa pang katangian ng bomba, kasama ang makabuluhang pag-asa sa temperatura at enerhiya, ay ang pagkakaroon ng antas ng saturation (tulad ng lahat ng iba pang mga reaksiyong kemikal); ito ay nangangahulugan na ang bilis ng pump ay hindi maaaring tumaas nang walang katiyakan habang ang konsentrasyon ng mga transported ions ay tumataas (Larawan 1.8). Sa kaibahan, ang flux ng isang passively diffusing substance ay tumataas sa proporsyon sa pagkakaiba ng konsentrasyon alinsunod sa batas ng diffusion (equation 1 at 2).

kanin. 1.7. A, B. Aktibo Na transportasyon + . Y axis: flux ng radioactive 24 Na + mula sa cell (imp./min). Abscissa axis: oras mula noong simula ng eksperimento. A. Ang cell ay pinalamig mula 18.3 °C hanggang 0.5 °C; daloy Na+ mula sa cell sa panahong ito ay inhibited. B. Pagpigil sa Na + flux mula sa cell sa pamamagitan ng dinitrophenol (DNP) sa isang konsentrasyon na 0.2 mmol/l (binago)
Bilang karagdagan sa Na/K pump, ang plasma membrane ay naglalaman ng hindi bababa sa isa pang pump - kaltsyum; ang pump na ito ay nagbobomba ng mga calcium ions (Ca 2+) palabas ng cell at kasangkot sa pagpapanatili ng kanilang intracellular na konsentrasyon sa napakababang antas (Talahanayan 1.1). Ang calcium pump ay naroroon sa napakataas na density sa sarcoplasmic reticulum ng mga selula ng kalamnan, na nag-iipon ng mga ion ng calcium bilang resulta ng pagkasira ng mga molekula ng ATP (tingnan ang Kabanata 4).
Epekto ng Na/K pump sa potensyal ng lamad at dami ng cell . Sa Fig. Ipinapakita ng Figure 1.9 ang iba't ibang bahagi ng kasalukuyang lamad at ipinapakita ang mga intracellular na konsentrasyon ng mga ions na

kanin. 1.8.Ang ugnayan sa pagitan ng rate ng transportasyon ng mga molekula at ang kanilang konsentrasyon (sa pasukan sa channel o sa binding site ng pump) sa panahon ng pagsasabog sa pamamagitan ng channel o sa panahon ng pumping transport. Ang huli ay nagiging puspos sa mataas na konsentrasyon (maximum na bilis, Vmax ) na halaga sa x-axis na tumutugma sa kalahati ng maximum na bilis ng bomba ( Vmax /2), ay ang konsentrasyon ng ekwilibriyo SA m

kanin. 1.9.Diagram na nagpapakita ng mga konsentrasyon ng Na+ , K+ at Cl – sa loob at labas ng cell at ang mga daanan para sa pagtagos ng mga ions na ito sa pamamagitan ng cell membrane (sa pamamagitan ng mga partikular na channel ng ion o paggamit ng Na/K pump. Sa ibinigay na mga gradient ng konsentrasyon, mga potensyal na equilibrium E Na, E K at E С l – katumbas ng mga ipinahiwatig, potensyal ng lamad Em = – 90 mV
tiyakin ang kanilang pag-iral. Ang isang panlabas na kasalukuyang ng mga potassium ions ay sinusunod sa pamamagitan ng mga channel ng potassium, dahil ang potensyal ng lamad ay bahagyang mas electropositive kaysa sa potensyal ng balanse para sa mga potassium ions. Ang pangkalahatang conductance ng sodium channels ay mas mababa kaysa sa potassium channels, i.e. ang mga channel ng sodium ay bukas nang hindi gaanong madalas kaysa sa mga channel ng potasa sa potensyal na pahinga; gayunpaman, humigit-kumulang sa parehong bilang ng mga sodium ions ang pumapasok sa cell habang ang mga potassium ions ay lumalabas sa cell, dahil ang malaking konsentrasyon at potensyal na mga gradient ay kinakailangan para sa mga sodium ions na lumaganap sa cell. Ang Na/K pump ay nagbibigay ng perpektong kompensasyon para sa mga passive diffusion currents, dahil inililipat nito ang mga sodium ions palabas ng cell at mga potassium ions papunta dito. Kaya, ang bomba ay electrogenic dahil sa pagkakaiba sa bilang ng mga singil na inilipat sa loob at labas ng cell, na sa normal na bilis ng operasyon nito ay lumilikha ng potensyal na lamad na humigit-kumulang 10 Ang mV ay mas electronegative kaysa kung ito ay ginawa ng mga passive ion fluxes lamang (tingnan ang Equation 7). Bilang resulta, ang potensyal ng lamad ay lumalapit sa potensyal na balanse ng potasa, na binabawasan ang pagtagas ng mga potassium ions. Na aktibidad Ang /K pump ay kinokontrol ng intracellular na konsentrasyon ng mga sodium ions. Ang bilis ng pump ay bumabagal habang ang konsentrasyon ng mga sodium ions na aalisin mula sa cell ay bumababa (Fig. 1.8), upang ang pump operation at ang daloy ng sodium ions sa cell ay balanse sa bawat isa, na pinapanatili ang intracellular na konsentrasyon ng sodium ions sa isang antas ng humigit-kumulang 10 mmol/L.
Upang mapanatili ang equilibrium sa pagitan ng pumping at passive membrane currents, mas maraming Na/K pump molecule ang kailangan kaysa sa channel proteins para sa potassium at sodium ions. Kapag ang channel ay bukas, sampu-sampung libong mga ion ang dumaan dito sa loob ng ilang millisecond (tingnan sa itaas), at dahil ang channel ay karaniwang nagbubukas ng ilang beses bawat segundo, sa kabuuan ay higit sa 10 5 mga ion ang dumaan dito sa panahong ito. Ang isang solong pump protein ay gumagalaw ng ilang daang sodium ions bawat segundo, kaya ang plasma membrane ay dapat maglaman ng humigit-kumulang 1000 beses na mas maraming pump molecule kaysa sa channel molecules. Ang mga sukat ng mga alon ng channel sa pahinga ay nagpakita ng pagkakaroon ng isang average ng isang potasa at isang sodium bukas na channel sa bawat 1 μm 2 lamad; Ito ay sumusunod mula dito na ang tungkol sa 1000 molecule ng Na/K pump ay dapat naroroon sa parehong espasyo, i.e. ang distansya sa pagitan ng mga ito ay nasa average na 34 nm; Ang diameter ng pump protein, bilang isang channel protein, ay 8-10 nm. Kaya, ang lamad ay medyo makapal na puspos ng mga pumping molecule
Ang katotohanan na ang daloy ng sodium ions sa cell, at potassium ions sa labas ng cell, ay binabayaran ng pagpapatakbo ng pump, mayroong isa pang kahihinatnan, na binubuo sa pagpapanatili ng isang matatag na osmotic pressure at pare-pareho ang volume. Sa loob ng cell mayroong isang mataas na konsentrasyon ng malalaking anion, pangunahin ang mga protina (A - sa Talahanayan 1.1), na hindi makapasok sa lamad (o tumagos sa pamamagitan nito nang napakabagal) at samakatuwid ay isang nakapirming bahagi sa loob ng cell. Upang balansehin ang singil ng mga anion na ito, kailangan ng pantay na bilang ng mga cation. Dahil sa pagkilos ng Na/K pump, ang mga cation na ito ay pangunahing mga potassium ions. Ang isang makabuluhang pagtaas sa konsentrasyon ng intracellular ion ay maaari lamang mangyari sa isang pagtaas sa konsentrasyon ng mga anion dahil sa daloy ng C1 - kasama ang gradient ng konsentrasyon sa cell (Talahanayan 1.1), ngunit ang potensyal ng lamad ay humahadlang dito. Papasok na kasalukuyang Cl – sinusunod lamang hanggang sa maabot ang potensyal na ekwilibriyo para sa mga chlorine ions; ito ay sinusunod kapag ang chlorine ion gradient ay halos kabaligtaran sa potassium ion gradient, dahil ang mga chlorine ions ay negatibong sisingilin (Equation 4). Kaya, ang isang mababang intracellular na konsentrasyon ng mga chlorine ions ay itinatag, na tumutugma sa isang mababang extracellular na konsentrasyon ng mga potassium ions. Ang resulta ay isang limitasyon ng kabuuang bilang ng mga ions sa cell. Kung ang potensyal ng lamad ay bumaba kapag ang Na/K na bomba ay naharang, halimbawa sa panahon ng anoxia, kung gayon ang equilibrium na potensyal para sa mga chlorine ions ay bumababa, at ang intracellular na konsentrasyon ng mga chloride ion ay tumataas nang naaayon. Ang pagpapanumbalik ng balanse ng mga singil, ang mga potassium ions ay pumapasok din sa cell; ang kabuuang konsentrasyon ng mga ions sa cell ay tumataas, na nagpapataas ng osmotic pressure; pinipilit nito ang tubig sa cell. Ang selda ay namamaga. Ang pamamaga na ito ay sinusunod sa vivo sa mga kondisyon ng kakulangan ng enerhiya.
Gradient ng konsentrasyon Na + bilang isang puwersang nagtutulak para sa transportasyon ng lamad . Ang kahalagahan ng Na/K pump para sa cell ay hindi limitado sa pag-stabilize ng normal na K + at Na + gradients sa lamad. Enerhiya na nakaimbak sa gradient ng lamad Na+ , ay kadalasang ginagamit upang mapadali ang transportasyon ng lamad ng iba pang mga sangkap. Halimbawa, sa Fig. 1.10 ay nagpapakita ng "simport" Na+ at mga molekula ng asukal sa cell. Dinadala ng membrane transport protein ang molekula ng asukal sa cell kahit na laban sa isang gradient ng konsentrasyon, habang ang Na + ay gumagalaw kasama ang konsentrasyon at potensyal na gradient, na nagbibigay ng enerhiya para sa transportasyon ng mga asukal. Ang transportasyon ng mga asukal na ito ay ganap na nakasalalay sa pagkakaroon ng isang mataas na gradient Na+ ; kung intracellular concentration Na+ tumataas nang malaki, humihinto ang transportasyon ng mga asukal. Para sa iba't-ibang c akharov, mayroong iba't ibang mga sistema ng simport. Transport ng mga amino acid sa isang cell na katulad ng transportasyon c akharov na ipinapakita sa Fig. 1.10; ibinibigay din ito ng gradient Na+ , Mayroong hindi bababa sa limang magkakaibang symport system, ang bawat isa ay dalubhasa para sa isang pangkat ng mga kaugnay na amino acid.
Bilang karagdagan sa mga sistema ng simport, mayroon ding "mga anti-porter". Ang isa sa kanila, halimbawa, sa isang cycle ay naglilipat ng isang calcium ion mula sa cell bilang kapalit ng tatlong papasok na sodium ions (Fig. 1.10). Enerhiya para sa Ca 2+ transportasyon ay nabuo dahil sa pagpasok ng tatlong sodium ions kasama ang isang konsentrasyon at potensyal na gradient. Ang enerhiya na ito ay sapat na (sa potensyal na magpahinga) upang mapanatili ang isang mataas na calcium ion gradient (mula sa mas mababa sa 10-7 mol/L sa loob ng cell hanggang sa humigit-kumulang 2 mmol/L sa labas ng cell).
Endo- at exocytosis . Para sa ilang mga sangkap na pumapasok sa cell o dapat ilabas

kanin. 1.10.Ang mga protina na naka-embed sa lipid bilayer ng lamad ay namamagitan sa symport ng glucose at Na + sa cell, pati na rin Ca2+/Na+ –antiport, kung saan ang puwersang nagtutulak ay ang Na + gradient sa lamad ng cell
mula dito, walang mga channel ng transportasyon; Kasama sa mga sangkap na ito, halimbawa, ang mga protina at kolesterol. Maaari silang dumaan sa lamad ng plasma sa mga vesicle, o mga bula, sa pamamagitan ng endo- at exocytosis. Sa Fig. Ipinapakita ng Figure 1.11 ang mga pangunahing mekanismo ng mga prosesong ito. Sa exocytosis, ang ilang organelles (tingnan sa ibaba) ay bumubuo ng mga vesicle na puno ng isang substance na kailangang alisin sa cell, tulad ng mga hormone o extracellular enzymes. Kapag ang gayong mga vesicle ay umabot sa lamad ng plasma, ang kanilang lipid membrane ay sumasama dito, kaya pinapayagan ang mga nilalaman na makatakas sa panlabas na kapaligiran. Sa kabaligtaran na proseso, ang endocytosis, ang lamad ng plasma ay nag-invaginates, na bumubuo ng isang hukay, na pagkatapos ay lumalalim at nagsasara, na bumubuo ng isang intracellular vesicle na puno ng extracellular fluid at ilang mga macromolecules. Upang matiyak ang pagsasanib ng lamad at pagsasara ng vesicle, ang mga contractile na elemento ng cytoskeleton ay kumikilos kasabay ng mga lamad mismo (tingnan sa ibaba). Ang endocytosis ay hindi palaging kasangkot lamang sa pagkuha ng extracellular na kapaligiran sa cell. Ang lamad ng cell ay kadalasang naglalaman ng mga partikular na receptor para sa mga macromolecule, tulad ng insulin o antigens, na kadalasang nakaayos sa mga espesyal na grupo. Matapos ang mga macromolecule na ito ay magbigkis sa kanilang mga receptor, ang endocytosis ay nangyayari sa lugar ng lamad na nakapalibot sa receptor, at ang macromolecule ay piling dinadala sa cell (Larawan 1.12, B).
Ang endo- at exocytosis ay patuloy na nangyayari sa mga selula. Ang halaga ng paglilipat ng materyal ng lamad ay makabuluhan; sa loob ng 1 oras, ang macrophage ay sumisipsip ng dalawang beses sa ibabaw na lugar ng cytoplasmic membrane nito sa anyo ng mga vesicle. Sa karamihan ng mga cell, ang turnover ng materyal na lamad ay hindi nangyayari nang kasing intensive, ngunit dapat pa rin itong maging makabuluhan.

kanin. 1.11.Exocytosis at endocytosis. pataas: ang intracellular vesicle ay sumasama sa lipid bilayer ng plasma membrane at bumubukas sa extracellular space. Ang prosesong ito ay tinatawag na exocytosis. Sa ilalim: ang plasma membrane ay pumapasok sa isang maliit na lugar at naglalabas ng isang vesicle na puno ng extracellular na materyal. Ang prosesong ito ay tinatawag na endocytosis
1.3. Transport ng mga sangkap sa loob ng cell
Ang endo- at exocytosis ay hindi lamang mga proseso ng transportasyon ng mga sangkap sa buong lamad ng cell, kundi pati na rin ang mga proseso ng pagpapalitan ng mga lamad - ang mga istrukturang bahagi ng cell mismo. Ang paksa ng pagsasaalang-alang sa seksyong ito ay iba pang katulad na mga proseso ng transportasyon sa cell at mga organel nito.
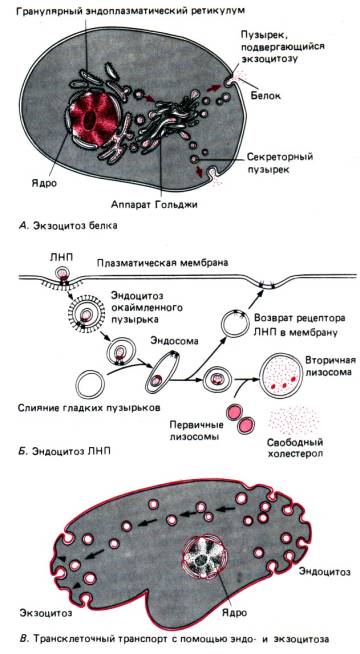
kanin. 1.12. A–B. Scheme ng mga proseso kabilang ang exo- at endocytosis. A. Ang protina na na-synthesize sa granular endoplasmic reticulum ay dinadala sa pamamagitan ng Golgi apparatus sa lamad ng plasma, kung saan ito ay tinatago ng exocytosis. B. Ang kolesterol na nakagapos sa mga particle ng LDL (low-density lipoprotein) ay nakakabit sa lamad ng plasma, nag-uudyok sa pagbuo ng isang endocytic vesicle sa rehiyong ito ng lamad at dinadala sa mga lysosome, kung saan ito ilalabas. SA. Ang extracellular na materyal na nakuha sa panahon ng endocytosis (sa figure sa kanan), dinadala sa pamamagitan ng cell sa mga vesicle, o mga vesicle, at inilabas sa pamamagitan ng exocytosis (sa figure kaliwa)
Pagsasabog . Naturally, sa cytosol ang pagkakaiba sa mga konsentrasyon ay inalis dahil sa pagsasabog; ang parehong ay totoo para sa mga likido na nakapaloob sa mga organelles. Dahil sa mataas na konsentrasyon ng natunaw na protina, ang pagsasabog dito ay mas mabagal kaysa sa tubig. Ang mga lipid membrane—sa paligid ng mga cell at sa loob ng mga organelles—ay mga two-dimensional na likido kung saan nangyayari ang diffusion. Ang mga lipid sa isang bilayer ng lamad ay nagkakalat sa loob ng kanilang sariling layer, bihirang lumipat mula sa isa't isa. Ang mga protina na nahuhulog sa kanila ay medyo mobile din; umiikot ang mga ito sa paligid ng isang axis na patayo sa lamad o nagkakalat sa gilid na may napakakaibang diffusion constants, 2–10,000 beses na mas mabagal kaysa sa mga phospholipid. Kaya, kung ang ilang mga protina ay malayang gumagalaw sa layer ng lipid at sa parehong bilis ng mga molekula ng lipid mismo, kung gayon ang iba ay naka-angkla, i.e. medyo matatag na konektado sa cytoskeleton. Mayroong "permanenteng" aggregates ng mga tiyak na protina sa lamad, halimbawa, pre- at postsynaptic na mga istraktura ng mga nerve cell. Ang malayang paglipat ng mga protina ay maaaring ipakita sa pamamagitan ng pagbubuklod sa kanila sa mga fluorescent dyes, ang luminescence na kung saan ay sapilitan sa pamamagitan ng panandaliang pag-iilaw sa isang maliit na bahagi ng lamad. Ang ganitong mga eksperimento ay nagpapakita na sa mas mababa sa 1 min, ang mga protina na nakagapos sa tina ay pantay na ipinamamahagi sa ibabaw ng lamad sa mga distansya na hanggang 10 μm.
Aktibong transportasyon sa mga lamad ng organelle .
Ang mga aktibong proseso ng transportasyon, na gumaganap ng isang mahalagang papel sa paggana ng lamad ng plasma, ay nangyayari din sa loob ng cell, sa mga lamad ng mga organelles. Ang mga tiyak na nilalaman ng iba't ibang mga organel ay nalikha nang bahagyang dahil sa panloob na synthesis, at bahagyang dahil sa aktibong transportasyon mula sa cytosol. Ang isang halimbawa ng huli ay ang nabanggit na Ca 2+ na bomba sa sarcoplasmic reticulum ng mga selula ng kalamnan. Ito ay lalong kawili-wili na sa kaso ng ATP synthesis sa mitochondria, ang kabaligtaran na prinsipyo ay nalalapat sa kung ano ang nangyayari sa ATPase pumps ng plasma membrane (Fig. 1.6). Sa panahon ng ATP synthesis, ang oxidative metabolism ay humahantong sa pagbuo ng isang matarik na gradient H+ sa mga panloob na lamad. Ang gradient na ito ay ang puwersang nagtutulak para sa baligtad na proseso ng pumping cycle ng aktibong transportasyon ng mga molekula: Ang mga H + ions ay gumagalaw sa lamad kasama ang gradient, at ang enerhiya na inilabas bilang isang resulta ay nagbibigay ng synthesis ng ATP mula sa ADP at pospeyt. Ang nagreresultang ATP, sa turn, ay nagbibigay ng enerhiya sa cell, kabilang ang para sa aktibong transportasyon.
Transport sa mga vesicle . Ang cell ay may malaking bilang ng mga organelles at nauugnay na mga vesicle (Larawan 1.1). Ang mga organel na ito, at lalo na ang mga vesicle, ay patuloy na gumagalaw, dinadala ang kanilang mga nilalaman sa iba pang mga organel o sa lamad ng plasma. Ang mga vesicle ay maaari ding lumipat mula sa lamad ng cell patungo sa mga organel, tulad ng sa endocytosis.
Proseso pagtatago ng protina ipinapakita sa Fig. 1.12, A. Ang protina ay na-synthesize malapit sa cell nucleus sa mga ribosome na nauugnay sa endoplasmic reticulum (ang tinatawag na butil-butil, o magaspang, endoplasmic reticulum); Sa sandaling nasa endoplasmic reticulum, ang protina ay nakabalot sa transport vesicles, na humihiwalay sa organelle at lumipat sa Golgi apparatus. Dito sila sumanib sa cisternae ng Golgi apparatus, kung saan ang protina ay binago (i.e., na-convert sa isang glycoprotein). Sa dulo ng cisternae, muling naghihiwalay ang mga vesicle. Ang mga secretory vesicle na nagdadala ng binagong protina ay lumipat sa lamad ng plasma at naglalabas ng kanilang mga nilalaman sa pamamagitan ng exocytosis.
Ang isa pang halimbawa ng isang landas ng transportasyon sa isang cell ay ipinapakita sa Fig. 1.12, B; Ito ang pagsipsip ng kolesterol ng cell. Ang kolesterol na dinadala sa dugo ay pangunahing nakagapos sa mga protina, tulad ng mga particle "mababang density ng lipoprotein"(LDL). Ang ganitong mga particle ay nakakabit sa mga partikular na lugar ng lamad na naglalaman ng mga receptor ng LDL, kung saan nangyayari ang endocytosis at ang LDL ay dinadala sa cell sa mga "bordered" na vesicle. Ang mga vesicle na ito ay nagsasama upang bumuo ng mga endosom at nawawala ang kanilang "edging" sa panahon ng prosesong ito. Ang mga endosom, naman, ay sumanib sa mga pangunahing lysosome, na naglalaman ng mga hydrolytic enzymes, at bumubuo ng pangalawang, mas malalaking lysosome. Sa kanila, ang kolesterol ay inilabas mula sa mga particle ng LDL at kumakalat sa cytosol, kung saan ito ay magagamit, halimbawa, para sa synthesis ng lipid membranes. Ang mga vesicle na hindi naglalaman ng LDL ay nahihiwalay din sa mga endosom, na gumagalaw sa isang espesyal na paraan sa lamad ng plasma at sumanib dito, bumabalik na materyal ng lamad at, marahil, mga receptor para sa LDL. Mula sa sandaling ang particle ng LDL ay nagbubuklod sa lamad hanggang sa mailabas ang kolesterol mula sa pangalawang lysosome, lumipas ang 10-15 minuto. Ang mga kapansanan sa pagbubuklod at pagsipsip ng LDL, i.e., sa supply ng kolesterol sa cell, ay gumaganap ng isang mapagpasyang papel sa pag-unlad ng isang malubha at laganap na sakit, atherosclerosis ("hardening" ng mga arterya).
Mayroong maraming iba pang mga ruta ng transportasyon na katulad ng ipinapakita sa Fig. 1.11 at 1.12, A, sa tulong ng kung saan ang mga tiyak na vesicle ay gumagalaw sa cell. Hindi alam nang eksakto kung paano sila gumagalaw, ngunit ang mga elemento ng cytoskeletal ay malamang na kasangkot sa prosesong ito. Ang mga vesicle ay maaaring mag-slide sa mga microtubule, kung saan ang enerhiya para sa paggalaw ay maliwanag na ibinibigay ng protina na nauugnay sa mga vesicle, ATPase (tingnan sa ibaba). Ito ay nananatiling ganap na hindi malinaw kung gaano karaming iba't ibang mga vesicle, na gumagalaw nang paisa-isa sa lahat ng direksyon, na nakarating sa kanilang destinasyon. Malinaw na kailangan silang "markahan" sa isang paraan na kinikilala ng sistema ng transportasyon at na-convert sa target na paggalaw.
Transport sa pamamagitan ng pagbuo at pagkasira ng mga organelles . Sa ngayon, isinasaalang-alang namin ang endo- at exocytosis bilang mga proseso ng pagdadala ng mga nilalaman ng mga vesicle. Mayroong isa pang aspeto ng mga prosesong ito, na nakasalalay sa katotohanan na ang naka-target na pag-alis ng lamad ng plasma sa isang bahagi ng ibabaw ng cell sa pamamagitan ng endocytosis at, sa kabaligtaran, ang pagdaragdag nito sa isa pa sa pamamagitan ng exocytosis ay gumagalaw ng malalaking lugar ng lamad (Fig . 1.12.E), na nagbibigay sa cell ng pagkakataon, halimbawa , bumuo ng isang paglaki o paglipat.
Ang mga katulad na muling pagsasaayos ay karaniwan din para sa cytoskeleton, lalo na para sa mga microfilament at microtubule (Larawan 1.1). Mga microfilament pangunahing binubuo ng protina ng F-actin, na may kakayahang mag-assemble sa fibrous bundle bilang resulta ng polymerization ng monomer mula sa cytosol. Ang mga bundle ay polarized, iyon ay, sila ay madalas na lumalaki lamang sa isang dulo, nag-iipon ng mga bagong molekula ng actin, habang ang kabilang dulo ay hindi gumagalaw o disassembly ay nangyayari dito. Dahil sa polarized na paglago na ito, ang mga microfilament ay mahusay na gumagalaw at ang istraktura ng kanilang network ay maaaring magbago. Ang paglipat ng actin mula sa isang depolymerized na estado (sol) sa isang organisadong estado (gel) ay maaaring mangyari nang napakabilis sa ilalim ng impluwensya ng iba pang mga protina o mga pagbabago sa konsentrasyon ng ion (tingnan sa ibaba). Mayroon ding mga protina na nagiging sanhi ng pagkasira ng mga filament ng actin upang bumuo ng mga maikling fragment. Ang mga manipis na projection ng maraming mga cell - filopodia - ay naglalaman ng isang sentral na bundle ng actin (Larawan 1.1), at ang iba't ibang mga paggalaw ng filopodia ay malamang na dahil sa mga paglipat ng actin: polymerization - depolymerization.
Microtubulemadalas ding sumasailalim sa mga katulad na paggalaw. Ang mekanismo ng mga paggalaw na ito ay magkatulad - polymerization ng tubulin mula sa cytosol sa paraan na ang isa sa mga dulo ng microtubule ay lumalaki, habang ang isa ay hindi nagbabago, o ang disassembly ay nangyayari doon. Kaya, ang isang microtubule, sa pamamagitan ng naaangkop na pagdaragdag o pag-alis ng materyal, ay maaaring lumipat sa buong cytosol.
Mga aktibong paggalaw ng cytoskeleton . Maaaring mangyari ang mga pagbabago sa mga istruktura ng cytoskeletal bilang resulta ng parehong aktibong paggalaw at muling pagsasaayos na inilarawan sa itaas. Sa maraming mga kaso, ang paggalaw ng mga microtubule at actin filament ay hinihimok ng mga contractile na protina na nagbubuklod sa mga filament o tubules at maaaring ilipat ang mga ito nang may kaugnayan sa isa't isa. Mga ardilya myosin at dynein naroroon sa cytosol ng lahat ng mga cell sa medyo mataas na konsentrasyon; sila ang mga elementong nagko-convert ng enerhiya sa paggalaw sa mga espesyal na selula (muscle) at organelles (cilia). Sa mga selula ng kalamnan, ang myosin ay bumubuo ng makapal na mga filament na parallel sa actin filament. Ang molekula ng myosin kasama ang "ulo" nito ay nakakabit sa actin filament at, gamit ang enerhiya ng ATP, inilipat ang myosin kasama ang molekula ng actin. Ang Myosin pagkatapos ay humiwalay sa actin. Ang kumbinasyon ng maraming ganoong koneksyon-disconnection cycle ay humahantong sa macroscopic pag-urong ng mga fibers ng kalamnan(kabanata 4). Ang Dynein ay gumaganap ng katulad na papel sa paggalaw ng mga microtubule sa panahon ng cilia (Larawan 1.1). Sa cytoplasm ng mga hindi espesyal na selula, ang myosin at dynein ay hindi bumubuo ng mga regular na hibla, ngunit sa karamihan ng mga kaso ay maliliit na grupo ng mga molekula. Kahit na sa gayong maliliit na pinagsama-samang mga pagsasama-sama, sila ay may kakayahang ilipat ang mga filament ng actin o microtubule. kanin. Ang Figure 1.13 ay naglalarawan ng prosesong ito kapag ang magkasalungat na polarized na mga molekulang myosin ay nakakabit din sa dalawang actin filament, na nakapolarize sa magkaibang direksyon. Ang mga myosin headgroup ay yumuko patungo sa buntot ng molekula, na kumakain ng ATP sa proseso, at ang dalawang actin filament ay gumagalaw sa kabaligtaran na direksyon, pagkatapos ay ang myosin ay humiwalay sa kanila. Ang mga paggalaw ng ganitong uri, kung saan ang enerhiya ng ATP ay na-convert sa mekanikal na gawain, ay maaaring magbago ng hugis ng cytoskeleton at, dahil dito, ang cell, at nagbibigay din ng transportasyon ng mga organel na nauugnay sa cytoskeleton.
Ang mga proseso ng intracellular transport ay pinakamalinaw na maipapakita sa axon ng isang nerve cell. Transportasyon ng axon ay tinalakay dito nang detalyado upang ilarawan ang mga kaganapan na malamang na mangyari sa magkatulad na paraan sa karamihan ng mga cell. Ang isang axon na ilang microns lamang ang diyametro ay maaaring umabot sa haba ng isang metro o higit pa, at ang paggalaw ng mga protina sa pamamagitan ng diffusion mula sa nucleus hanggang sa distal na dulo ng axon ay aabot ng mga taon. Matagal nang alam na kapag ang anumang bahagi ng axon ay sumasailalim sa pagsisikip, ang bahagi ng axon na matatagpuan sa mas malapit ay lumalawak. Mukhang nakaharang ang sentripugal na daloy sa axon. ganyan daloy-mabilis na transportasyon ng axon maipakita sa pamamagitan ng paggalaw ng mga radioactive marker, tulad ng sa eksperimento na ipinapakita sa Fig. 1.14. Ang radiolabeled leucine ay na-injected sa dorsal root ganglion, at pagkatapos ay sinusukat ang radioactivity sa sciatic nerve sa layo na 166 mm mula sa neuronal cell body mula ika-2 hanggang ika-10 oras. Sa paglipas ng 10 oras, bahagyang nagbago ang tugatog ng radyaktibidad sa lugar ng iniksyon. Ngunit ang alon ng radyaktibidad ay kumakalat sa kahabaan ng axon sa pare-parehong bilis na halos 34 mm sa loob ng 2 oras, o 410 mm/araw. Ipinakita na sa lahat ng mga neuron ng mga homeothermic na hayop, ang mabilis na transportasyon ng axonal ay nangyayari sa parehong bilis, at walang kapansin-pansin na mga pagkakaiba ang naobserbahan sa pagitan ng manipis, unmyelinated na mga hibla at ang pinakamakapal na axon, gayundin sa pagitan ng mga motor at sensory fibers. Ang uri ng radioactive marker ay hindi rin nakakaapekto sa rate ng mabilis na axonal transport; Ang mga marker ay maaaring magsilbi bilang iba't ibang radioactive

kanin. 1.13.Ang non-muscle myosin complex, na may isang tiyak na oryentasyon, ay maaaring magbigkis sa mga filament ng actin na may iba't ibang polarity at, gamit ang enerhiya ng ATP, palitan ang mga ito na may kaugnayan sa bawat isa.
mga molekula, tulad ng iba't ibang amino acid, na kasama sa mga protina ng cell body ng neuron. Kung susuriin natin ang peripheral na bahagi ng nerve upang matukoy ang likas na katangian ng mga carrier ng radyaktibidad na dinadala dito, kung gayon ang mga naturang carrier ay matatagpuan pangunahin sa bahagi ng protina, ngunit din sa komposisyon ng mga mediator at libreng amino acid. Alam na ang mga katangian ng mga sangkap na ito ay naiiba at ang mga sukat ng kanilang mga molekula ay partikular na naiiba, maaari nating ipaliwanag ang patuloy na bilis ng transportasyon sa pamamagitan lamang ng isang mekanismo ng transportasyon na karaniwan sa lahat ng mga ito.
Inilarawan sa itaas mabilis na transportasyon ng axon ay anterograde, ibig sabihin, nakadirekta palayo sa cell body. Ipinakita na ang ilang mga sangkap ay lumilipat mula sa paligid patungo sa cell body gamit retrograde na transportasyon. Halimbawa, ang acetylcholinesterase ay dinadala sa direksyong ito sa bilis na 2 beses na mas mabagal kaysa sa bilis ng mabilis na transportasyon ng axonal. Ang isang marker na kadalasang ginagamit sa neuroanatomy, malunggay peroxidase, ay gumagalaw din sa pamamagitan ng retrograde transport. Ang retrograde transport ay malamang na gumaganap ng isang mahalagang papel sa regulasyon ng synthesis ng protina sa katawan ng cell. Ilang araw pagkatapos maputol ang axon, ang chromatolysis ay sinusunod sa katawan ng cell, na nagpapahiwatig ng pagkagambala sa synthesis ng protina. Ang oras na kinakailangan para sa chromatolysis ay nauugnay sa tagal ng retrograde na transportasyon mula sa site ng axon transection sa cell body. Ang resultang ito ay nagmumungkahi din ng paliwanag para sa karamdamang ito - ang paghahatid mula sa paligid ng "signal substance" na kumokontrol sa synthesis ng protina ay nagambala. Ito ay malinaw na ang pangunahing "mga sasakyan" na ginagamit para sa mabilis na axonal
kanin. 1.14.Eksperimento na nagpapakita ng mabilis na axonal transport sa mga sensory fibers ng cat sciatic nerve. Ang leucine na may label na tritium ay itinurok sa dorsal root ganglion at ang radyaktibidad sa ganglion at ang mga sensory fibers ay sinusukat sa 2, 4, 6, 8 at 10 oras pagkatapos ng iniksyon. (ibaba ng larawan). Sa pamamagitan ng x-axis Ang distansya mula sa ganglion hanggang sa mga lugar ng sciatic nerve kung saan ginawa ang pagsukat ay naka-plot. Sa ordinate axis lamang para sa upper at lower curves, ang radioactivity (pulse/min) ay naka-plot sa isang logarithmic scale. "Wave" ng tumaas na radyaktibidad (mga arrow) gumagalaw sa bilis na 410 mm/araw (kasama)
transportasyon ay vesicles (vesicles) at organelles, tulad ng mitochondria, na naglalaman ng mga sangkap na kailangang dalhin. Ang paggalaw ng mga pinakamalaking vesicle o mitochondria ay maaaring obserbahan gamit ang isang mikroskopyo sa vivo . Ang ganitong mga particle ay gumagawa ng maikli, mabilis na paggalaw sa isang direksyon, huminto, madalas na bahagyang paatras o sa gilid, huminto muli, at pagkatapos ay huminto sa pangunahing direksyon. Ang 410 mm/araw ay tumutugma sa isang average na bilis ng anterograde na paggalaw na humigit-kumulang 5 μm/s; ang bilis ng bawat indibidwal na paggalaw ay dapat, samakatuwid, ay mas mataas, at kung isasaalang-alang natin ang mga sukat ng mga organelles, filament at microtubule, kung gayon ang mga paggalaw na ito ay talagang napakabilis. Ang mabilis na transportasyon ng axonal ay nangangailangan ng isang makabuluhang konsentrasyon ng ATP. Ang mga lason tulad ng colchicine, na nakakagambala sa mga microtubule, ay humaharang din sa mabilis na transportasyon ng axonal. Ito ay sumusunod mula dito na sa proseso ng transportasyon na aming isinasaalang-alang, ang mga vesicle at organelles ay gumagalaw kasama ang mga microtubule at actin filament; Ang paggalaw na ito ay ibinibigay ng maliliit na pinagsama-samang mga molekula ng dynein at myosin, na kumikilos tulad ng ipinapakita sa Fig. 1.13, gamit ang ATP energy.
Ang mabilis na transportasyon ng axon ay maaari ding kasangkot sa mga proseso ng pathological. Ang ilang mga neurotropic na virus (halimbawa, herpes o polio virus) ay pumapasok sa axon sa periphery at gumagalaw sa pamamagitan ng retrograde transport sa neuron body, kung saan sila ay dumarami at nagsasagawa ng kanilang mga nakakalason na epekto. Ang tetanus toxin, isang protina na ginawa ng bacteria na pumapasok sa katawan kapag nasira ang balat, ay nakukuha ng nerve endings at dinadala sa neuron body, kung saan ito ay nagdudulot ng mga katangian ng muscle spasms. May mga kilalang kaso ng nakakalason na epekto sa mismong transportasyon ng axon, halimbawa, pagkakalantad sa pang-industriyang solvent acrylamide. Sa karagdagan, ito ay pinaniniwalaan na ang pathogenesis ng bitamina deficiency beriberi at alcoholic polyneuropathy ay nagsasangkot ng isang paglabag sa mabilis na axonal transport.
Bilang karagdagan sa mabilis na transportasyon ng axonal sa cell, mayroon ding medyo matindi mabagal na transportasyon ng axon. Ang tubulin ay gumagalaw sa kahabaan ng axon sa bilis na humigit-kumulang 1 mm/araw, at ang actin ay gumagalaw nang mas mabilis—hanggang 5 mm/araw. Ang iba pang mga protina ay lumilipat kasama ang mga cytoskeletal na sangkap na ito; halimbawa, ang mga enzyme ay lumilitaw na nauugnay sa actin o tubulin. Ang mga rate ng paggalaw ng tubulin at actin ay halos pare-pareho sa mga rate ng paglago na natagpuan para sa mekanismo na inilarawan dati, kung saan ang mga molekula ay isinasama sa aktibong dulo ng isang microtubule o microfilament. Samakatuwid, ang mekanismong ito ay maaaring maging sanhi ng mabagal na transportasyon ng axonal. Ang rate ng mabagal na transportasyon ng axonal ay humigit-kumulang na tumutugma sa rate ng paglaki ng axon, na tila nagpapahiwatig ng mga paghihigpit na ipinataw ng istraktura ng cytoskeleton sa pangalawang proseso.
Sa pagtatapos ng seksyong ito, dapat itong bigyang-diin na ang mga cell ay hindi nangangahulugang mga static na istruktura, na lumilitaw, halimbawa, sa mga mikroskopikong larawan ng elektron. Plasma lamad at lalo na ang mga organelle ay nasa patuloy na mabilis na paggalaw at patuloy na muling pagsasaayos; kaya naman nagagawa nilang gumana. Dagdag pa, ang mga ito ay hindi simpleng mga silid kung saan nagaganap ang mga reaksiyong kemikal, ngunit mataas na organisadong conglomerates ng mga lamad at mga hibla, kung saan ang mga reaksyon ay nangyayari sa isang mahusay na organisadong pagkakasunud-sunod.
1.4. Regulasyon ng mga function ng cellular
Ang pagpapanatili ng indibidwal na cell bilang isang functional unit ay pangunahing kinokontrol ng nucleus; ang pag-aaral ng naturang mga mekanismo ng regulasyon ay ang paksa ng cell biology at biochemistry. Kasabay nito, dapat baguhin ng mga cell ang kanilang mga pag-andar alinsunod sa mga kondisyon sa kapaligiran at mga pangangailangan ng iba pang mga cell ng katawan, ibig sabihin, nagsisilbi silang mga bagay ng functional regulation. Sa ibaba ay maikli nating isasaalang-alang kung paano kumikilos ang mga regulatory influence na ito sa plasma membrane at kung paano sila naabot ang mga intracellular organelles.
Mga epekto sa regulasyon sa lamad ng cell
Lamad potensyal . Sa maraming mga kaso, ang regulasyon ng mga function ng cellular ay isinasagawa sa pamamagitan ng pagbabago ng potensyal ng lamad. Ang mga lokal na pagbabago sa potensyal ay posible kapag: 1) ang kasalukuyang mula sa isang katabing lugar ng cell o nabuo ng isa pang cell ay dumadaloy sa lamad; 2) nagbabago ang extracellular na konsentrasyon ng mga ion (madalas [K + ] palabas ); 3) bukas ang mga channel ng ion ng lamad. Ang mga pagbabago sa potensyal ng lamad ay maaaring makaapekto sa conformation ng mga protina ng lamad, na nagiging sanhi, lalo na, ang mga channel upang magbukas o magsara. Tulad ng inilarawan sa itaas, ang paggana ng ilang mga lamad na bomba ay nakasalalay sa potensyal ng lamad. Ang mga selula ng nerbiyos ay dalubhasa upang makita ang mga pagbabago sa potensyal ng lamad bilang impormasyon na dapat iproseso at ipadala (tingnan ang Kabanata 2).
Extracellular regulatory substance . Ang pinakamahalagang mekanismo ng regulasyon na kinasasangkutan ng mga extracellular substance ay ang kanilang pakikipag-ugnayan sa mga partikular na receptor sa lamad ng plasma o sa loob ng cell. Kasama sa mga sangkap na ito ang mga synaptic transmitters, na nagpapadala ng impormasyon sa pagitan ng mga nerve cell, mga lokal na ahente, at mga sangkap na umiikot sa dugo at umaabot sa lahat ng mga selula ng katawan, tulad ng mga hormone at antigens. Mga synaptic na transmiter ay maliliit na molekula na inilabas mula sa mga nerve ending sa synapse;
kapag naabot nila ang plasma membrane ng isang kalapit, postsynaptic cell, nagti-trigger sila ng mga electrical signal o iba pang mekanismo ng regulasyon. Ang isyung ito ay tinalakay nang detalyado sa Chap. 3.
Mga lokal na ahente ng kemikal madalas na itinago ng mga espesyal na selula. Malayang nagkakalat ang mga ito sa extracellular space, ngunit ang kanilang pagkilos ay limitado sa isang maliit na grupo ng mga selula dahil sa mabilis na pagkasira ng mga sangkap na ito, alinman sa kusang o sa ilalim ng pagkilos ng mga enzyme. Ang isang halimbawa ng pagpapalabas ng naturang mga ahente ay ang pagpapalabas histamine mast cells dahil sa pinsala o immune response. Ang histamine ay nagdudulot ng relaxation ng vascular smooth muscle cells, pinatataas ang permeability ng vascular endothelium at pinasisigla ang sensory nerve endings na nagpapagitna sa pakiramdam ng pangangati. Ang iba pang mga lokal na ahente ng kemikal ay inilalabas ng maraming iba pang mga selula. Ang mga karaniwang lokal na ahente ay prostaglandin, bumubuo ng isang pangkat ng humigit-kumulang 20 derivatives ng fatty acid. Ang mga ito ay patuloy na inilabas mula sa malawak na ipinamamahaging mga selula, ngunit kumikilos lamang nang lokal, dahil mabilis silang nawasak ng mga lamad na phospholipases. Ang iba't ibang mga prostaglandin ay may malawak na spectrum ng pagkilos: maaari silang mag-trigger ng pag-urong ng makinis na mga selula ng kalamnan, maging sanhi ng pagsasama-sama ng mga platelet ng dugo (mga platelet), o sugpuin ang pagbuo ng corpus luteum sa mga ovary.
Naglilingkod ang iba pang lokal na ahente mga kadahilanan ng paglago. Ang pinakakilala ay ang nerve growth factor (NGF) para sa mga sympathetic neuron, na kinakailangan para sa paglaki at kaligtasan ng mga neuron na ito sa panahon ng pag-unlad. sa vivo o sa kultura ng cell. Tila, ang mga target na cell para sa klase ng mga neuron na ito ay nagtatago ng NGF at sa gayon ay tinitiyak ang wastong innervation. Kapag bumubuo ng mga organo, ang mga cell ay madalas na kailangang "hanapin ang kanilang paraan" upang i-target ang mga cell, na maaaring matatagpuan sa malaking distansya. Alinsunod dito, dapat mayroong maraming espesyal na salik ng paglago na katulad ng NGF.
Mga hormone at antigens dinadala ng dugo sa lahat ng mga selula. Ang mga antigen ay nag-trigger ng immune response mula sa mga cell na may mga tiyak na antibodies. Gayunpaman, ang mga antigen, bilang panuntunan, ay mga dayuhang sangkap na hindi ginawa sa tumutugon na organismo (para sa higit pang mga detalye, tingnan ang Kabanata 18). Ang ilang mga hormone, tulad ng insulin o thyroxine, ay nakakaapekto sa iba't ibang uri ng cell, habang ang iba, tulad ng mga sex hormone, ay nakakaapekto lamang sa isang partikular na uri ng cell. Ang mga hormone ay alinman sa mga peptide, na ang pagkilos ay na-trigger sa pamamagitan ng pagbubuklod sa isang receptor sa cell membrane, o mga steroid at thyroxine, na kumakalat sa lipid membrane at nagbubuklod sa mga intracellular receptor. Ang mga steroid na hormone ay nagbubuklod sa nuclear chromatin, na nagreresulta sa transkripsyon ng ilang mga gene. Ang mga resultang protina ay nagdudulot ng mga pagbabago sa mga function ng cellular, na siyang tiyak na epekto ng mga hormone. Ang mga isyu na may kaugnayan sa pagpapalabas at pagkilos ng mga hormone ay tinalakay nang detalyado sa Kabanata. 17.
Intracellular na komunikasyon na kinasasangkutan ng mga pangalawang mensahero
Kasama sa mga function ng regulasyon na inilarawan sa itaas ang mga epekto sa lamad ng cell. Ang impormasyong natatanggap ng cell membrane ay dapat madalas na nagiging sanhi ng mga organel na mag-react at inilipat sa kanila ng iba't ibang mga sangkap na kilala bilang pangalawang mensahero (kumpara sa mga unang mensahero, na dumarating sa cell mula sa mga panlabas na mapagkukunan). Ang pag-aaral ng mga pangalawang tagapamagitan ay mabilis na umuunlad, at walang garantiya na ang kasalukuyang antas ng pag-unawa sa problema ay magiging sapat na kumpleto. Dito ay haharapin natin ang tatlong pinag-aralan na tagapamagitan: Ca 2+ , cAMP at inositol triphosphate.
Kaltsyum.Ang pinakasimpleng intracellular mediator ay ang Ca 2+ ion. Ang libreng konsentrasyon nito sa isang resting cell ay napakababa at umaabot sa 10_–8–10–7 mol/l. Maaari itong pumasok sa cell sa pamamagitan ng mga partikular na channel ng lamad kapag nasa bukas na estado ang mga ito, halimbawa, kapag nagbago ang potensyal ng lamad (tingnan ang Kabanata 2). Ang nagresultang pagtaas sa konsentrasyon ng Ca 2+ ay nag-trigger ng mahahalagang reaksyon sa cell, tulad ng pag-urong ng myofibrils, na siyang batayan ng pag-urong ng kalamnan (tingnan ang Kabanata 4), o ang paglabas ng mga vesicle na naglalaman ng mga neurotransmitter mula sa mga nerve endings (tingnan ang Kabanata 3) . Ang parehong mga reaksyon ay nangangailangan ng konsentrasyon ng Ca 2+ na humigit-kumulang 10 –5 mol/L. Ang Ca 2+, na may epekto sa regulasyon, ay maaari ding ilabas mula sa mga intracellular na tindahan, tulad ng endoplasmic reticulum. Ang paglabas ng Ca 2+ mula sa depot ay nangangailangan ng pakikilahok ng iba pang mga tagapamagitan (tingnan, halimbawa, Fig. 1.16).
Cyclic adenosine monophosphate, cAMP. Kamakailan lamang, napatunayan na ang cyclic adenosine monophosphate (cAMP), isang derivative ng pangunahing pinagmumulan ng enerhiya ng katawan, ang ATP, ay isang mahalagang pangalawang mensahero. Ang kumplikadong kadena ng mga reaksyon na ipinapakita sa Fig. 1.15, nagsisimula sa receptor R s sa panlabas na ibabaw ng lamad ng plasma, na maaaring magsilbi bilang isang tiyak na lugar na nagbubuklod para sa iba't ibang mga tagapamagitan at mga hormone. Pagkatapos magbigkis sa isang tiyak na "stimulating" molecule R s nagbabago ang anyo nito; ang mga pagbabagong ito ay nakakaapekto sa protina G s sa panloob na ibabaw ng lamad sa paraang ang pag-activate ng huli sa pamamagitan ng intracellular guanosine triphosphate (GTP) ay naging posible. Aktibong protina G s , sa turn, ay pinasisigla ang enzyme sa panloob na ibabaw ng lamad, ang adenylate cyclase (AC), na nag-catalyze sa pagbuo ng cAMP mula sa ATP. Ang water-soluble na cAMP ay ang tagapamagitan na nagpapadala ng epekto

kanin. 1.15.Isang hanay ng mga reaksyon na kinasasangkutan ng intracellular messenger cAMP (cyclic adenosine monophosphate). Ang mga excitatory o nagbabawal na mga panlabas na signal ay nagpapagana ng mga receptor ng lamad R s o Ri . Kinokontrol ng mga receptor na ito ang proseso ng pagbubuklod G – mga protina na may intracellular GTP (guanosine triphosphate), sa gayo'y pinasisigla o pinipigilan ang intracellular adenylate cyclase (AC). Ang amplifying enzyme AC ay nagko-convert ng adenosine triphosphate (ATP) sa cAMP, na pagkatapos ay hinahati sa AMP ng phosphodiesterase. Ang libreng cAMP ay kumakalat sa cell at nag-activate ng adenylate kinase (A-kinase), na naglalabas ng catalytic subunit C nito, na nagpapa-catalyze sa phosphorylation ng intracellular protein, i.e. bumubuo ng huling epekto ng isang extracellular stimulus. Ang diagram ay nagpapakita rin ng mga pharmacological na gamot at lason na nagpapalitaw ng (+) o pumipigil (-) ng ilang mga reaksyon (binago)
pagpapasigla ng extracellular receptor R s sa mga panloob na istruktura ng cell.
Kaayon ng stimulatory chain ng mga reaksyong kinasasangkutan R s Ang pagbubuklod ng mga inhibitory mediator at hormones sa kaukulang receptor ay posible R i na muli sa pamamagitan ng GTP-activated protein G , pinipigilan ang AC at sa gayon ay ang produksyon ng cAMP. Ang diffusing sa cell, ang cAMP ay tumutugon sa adenylate kinase (A-kinase); naglalabas ito ng subunit C, na catalyzes ang phosphorylation ng protina P. Ang phosphorylation na ito ay nagko-convert ng mga protina sa isang aktibong anyo, at maaari na nilang isagawa ang kanilang mga partikular na epekto sa regulasyon (halimbawa, maging sanhi ng pagkasira ng glycogen). Ang kumplikadong sistema ng regulasyon ay napakahusay dahil ang resulta ay ang phosphorylation ng maraming mga protina, ibig sabihin, ang signal ng regulasyon ay naglalakbay sa circuit na may mataas na pakinabang. Mga panlabas na tagapamagitan na nagbubuklod sa mga receptor R s at R i tiyak sa bawat isa sa kanila, lubhang magkakaibang. Adrenaline, kumokonekta sa R s o R i nakikilahok sa regulasyon ng metabolismo ng lipid at glycogen, pati na rin sa pagpapahusay ng pag-urong ng kalamnan ng puso at sa iba pang mga reaksyon (tingnan ang Kabanata 19). Ang thyroid-stimulating hormone, pag-activate R s , pinasisigla ang pagpapalabas ng hormone thyroxine ng thyroid gland, at pinipigilan ng prostaglandin I ang pagsasama-sama ng mga platelet ng dugo. Inhibitory effect, kabilang ang adrenaline, mediated through R i ay ipinahayag sa pagbagal ng lipolysis. kaya, Ang cAMP system ay isang multifunctional intracellular regulatory system, na maaaring tumpak na kontrolin ng extracellular stimulatory at inhibitory signaling substances.
Inositol phosphate "KUNG" h " Ang intracellular system ng pangalawang messenger, inositol phosphate, ay natuklasan kamakailan lamang (Larawan 1.16). Sa kasong ito, walang paraan ng pagbabawal, ngunit may pagkakatulad sa sistema ng cAMP, kung saan ang epekto ng pagpapasigla ng R receptor ay inililipat sa GTP-activated G protein sa panloob na ibabaw ng lamad. Sa susunod na yugto, ang karaniwang lamad lipid phosphatidylinositol (PI), na dati nang nakatanggap ng dalawang karagdagang grupo ng pospeyt, ay na-convert sa PI-diphosphate (PIP 2), na na-cleaved ng activated phosphodiesterase (PDE) sa inositol triphosphate(IFz) at lipid diacylglycerol(DAG). Ang Inositol triphosphate ay isang pangalawang messenger na nalulusaw sa tubig na kumakalat sa cytosol. Ito ay pangunahing kumikilos sa pamamagitan ng pagpapakawala ng Ca 2+ mula sa endoplasmic reticulum. Ang Ca 2+ naman ay gumaganap bilang isang mensahero, tulad ng inilarawan sa itaas; halimbawa, pinapagana nito ang Ca 2+ -dependent phosphokinase, na nagpo-phosphorylate ng mga enzyme. Ang lipid subunit ng DAG (Larawan 1.16) ay nagdadala din ng signal, na nagkakalat sa lipid phase ng plasma membrane sa C-kinase na matatagpuan sa panloob na ibabaw nito, na isinaaktibo sa pakikilahok ng phosphatidylserine bilang isang cofactor. Pagkatapos ang C-kinase ay nag-trigger ng phosphorylation ng mga protina, na ginagawang isang aktibong anyo.
Ang intracellular second messenger system na IF 3 ay maaari ding kontrolin ng iba't ibang panlabas na mediator at hormone, kabilang ang acetylcholine, serotonin, vasopressin, at thyroid-stimulating hormone; tulad ng cAMP system, ito ay nailalarawan sa pamamagitan ng iba't ibang intracellular effect. Posible na ang sistemang ito ay isinaaktibo din ng liwanag sa visual na receptor ng mata at gumaganap ng isang pangunahing papel sa phototransduction (tingnan ang Kabanata 11). Sa unang pagkakataon sa indibidwal na pag-unlad ng organismo, ang receptor ng IG system ay isinaaktibo ng tamud, bilang isang resulta kung saan ang IG ay nakikilahok sa mga reaksyon ng regulasyon na kasama ng pagpapabunga ng itlog.
Ang mga sistema ng cAMP at IFz-DAG ay lubos na epektibo mga biological enhancer. sila

kanin. 1.16.Isang chain ng mga reaksyon na kinasasangkutan ng intracellular mediator IPG (inositol triphosphate). Tulad ng sa cAMP system, ang extracellular signal ay pinapamagitan sa pamamagitan ng isang protinaG, na sa kasong ito ay nagpapagana ng phosphodiesterase (PDE). Sinisira ng enzyme na ito ang phosphatidylinosine diphosphate (PIP 2 ) sa lamad ng plasma bago ang IF h at diacylglycerol (DAG); KUNG h diffuses sa cytoplasm. Dito nagiging sanhi ng paglabas ng Ca 2+ mula sa endoplasmic reticulum; pagtaas ng konsentrasyon ng Ca 2+ sa cytoplasm ([Ca 2+ ]i ) pinapagana ang protina kinase, na nagpo-phosphorylate at samakatuwid ay nagpapagana ng mga enzyme. Ang isa pang produkto, DAG, ay nananatili sa lamad at pinapagana ang protina kinase C (phosphatidylserine cofactor, PS). Ang protina kinase C ay nag-phosphorylate din ng mga enzyme na namamagitan sa mga partikular na aksyon na nauugnay sa extrinsic receptor stimulation R . Mga sangay ng chain of reactions na kinasasangkutan ng IF h at ang DAG ay maaaring isaaktibo nang nakapag-iisa sa pamamagitan ng ionomycin at phorbol ester, ayon sa pagkakabanggit (binago)
i-convert ang reaksyon sa pagitan ng isang transmitter at isang panlabas na membrane receptor sa phosphorylation ng iba't ibang intracellular na protina, na maaaring makaimpluwensya sa iba't ibang mga function ng cell. Ang isa sa mga makabuluhang aspeto ng problema ay na, sa abot ng nalalaman ngayon, mayroon lamang itong dalawang malapit na nauugnay na sistema ng regulasyon ng ganitong uri, na ginagamit ng maraming panlabas na tagapamagitan upang ayusin ang iba't ibang mga proseso ng intracellular. Kasabay nito, ang mga regulatory system na ito, kabilang ang Ca 2+, ay malapit na nakikipag-ugnayan sa isa't isa, na nagpapahintulot sa kanila na magsagawa ng mahusay na regulasyon ng mga cellular function.
Mga Tutorial at Gabay
1. Alberts SA., Bray D., Lewis J., Raff M., Roberts SA., Watson J.D.Molecular Biology of the Cell, New York at London, Garland Publishing Inc., 1983.
2. Czihak G., Longer H., Ziegler H.(eds.). Biology. Berlin, Heidelberg, New York, Springer, 1983.
3. Hille SA. Ionic channels ng excitable membranes. Sunderland, Mass., Sinauer Assoc., 1984.
4. Hoppe W., Lohmann W.. Marki H., Ziegler H.(eds.). Biophysics. Berlin, Heidelberg, New York, Springer, 1984.
5. Jungermann SA., Mahler H.Biochemie. Berlin, Heidelberg, New York, Springer, 1980.
6. Kandel E. R., Schwartz– J. H.,(eds.). Mga Prinsipyo ng neural science, New York, Amsterdam, Oxford, Elsevier, 1985.
7. Schiebler T. H., Schmidt W.Anatomic des Menschen. Berlin, Heidelberg, New York, Tokyo, Springer, 1983.
Mga orihinal na artikulo at pagsusuri
8. Berridge M.J. Ang molekular na batayan ng komunikasyon sa loob ng cell, Sci. Amer, 253, 124-134 (1985).
9. Berridge M.J., Irvine R.F. Inositol triphosphate, isang nobelang pangalawang mensahero sa cellular signal transduction. Kalikasan, 312, 315 321 (1984).
10. Bretscher M.S. Ang mga molekula ng lamad ng cell, Sci. Amer., 253, 124–134 (1985).
11. Daut J. Ang buhay na cell bilang isang makinang nagpapalipat-lipat ng enerhiya. Isang minimal na modelo ng myocardial metabolism, Biochem. at Biophys. Acta, 895, 41–62 (1987).
12. Hodgkin A.L., Katz SA. Ang epekto ng sodium ions sa electrical activity ng higanteng axon ng pusit. J. Physiol. (Lond.), 108, 37–77 (1949).
13. Hodgkin A.L., Keynes R.D. Aktibong transportasyon ng mga cation sa mga higanteng axon mula sa Sepia at Loligo, J. Physiol. (Lond.), 128, 28–42 (1955).
14. Mas mahaba ang P. Ionic channels na may conformational substates, Biophys. J., 47, 581–590 (1985).
15. Ochs S., Worth P.M. Exoplasmic transport sa normal at pathologic system. Sa: Physiology at Patolohiya ng Axons, S.G. Waxmann, Ed. New York, Raven Press, 1978.
buod ng iba pang mga presentasyon"Mga paraan ng pagtuturo ng biology" - School zoology. Pagpapakilala sa mga mag-aaral sa paggamit ng siyentipikong zoological data. Edukasyong moral. Karagdagang biyaya ng manukan. Pagpili ng mga pamamaraan. Mga proseso sa buhay. isda sa aquarium. Nutrisyon. Edukasyong Pangkalikasan. Materyalidad ng mga proseso ng buhay. Mga negatibong resulta. Pansin sa mga mag-aaral. Mandatoryong anyo. Nakatingin sa maliliit na hayop. Mga layunin at layunin ng biology. Kwento.
"Pag-aaral na nakabatay sa problema sa mga aralin sa biology" - Kaalaman. Mga bagong aklat-aralin. Ang landas patungo sa solusyon. Problema. Mga seminar. Ano ang isang gawain? Albrecht Durer. Pag-aaral na nakabatay sa problema sa mga aralin sa biology. Mga hindi pamantayang aralin. Ano ang ibig sabihin ng problem-based learning. Ang kalidad ng buhay. Biology bilang isang akademikong asignatura. Tanong. Aralin sa paglutas ng problema. Nabawasan ang interes sa paksa. Mga klase sa laboratoryo batay sa problema.
"Kritikal na pag-iisip sa mga aralin sa biology" - Teknolohiya ng "kritikal na pag-iisip". Gamit ang teknolohiya ng “development of critical thinking”. Talahanayan para sa aralin. Pagganyak sa pag-aaral. Mga ekosistema. Ang kahulugan ng "pagbuo ng kritikal na pag-iisip." Mga palatandaan ng teknolohiya. teknolohiya ng RKM. Istraktura ng aralin. Mga pangunahing direksyon. Kasaysayan ng teknolohiya. Pedagogical na teknolohiya. Mga tuntunin ng teknolohiya. Mga takdang-aralin sa biology. Photosynthesis. Mga pamamaraan na ginamit sa iba't ibang yugto ng aralin.
"Mga aralin sa biology na may interactive na whiteboard" - Mga elektronikong aklat-aralin. Mga benepisyo para sa mga mag-aaral. Ang isang interactive na whiteboard ay tumutulong sa paghahatid ng impormasyon sa bawat mag-aaral. Mga gawaing didactic. Paglutas ng mga biological na problema. Mga kalamangan ng pagtatrabaho sa mga interactive na whiteboard. Paggawa gamit ang mga presentasyon. Magtrabaho sa paghahambing ng mga bagay. Gumagalaw na mga bagay. Paggamit ng mga spreadsheet. Paggamit ng interactive na whiteboard sa proseso ng pagtuturo sa mga mag-aaral. Mga benepisyo para sa mga guro.
"System-activity approach sa biology" - Mga tanong sa seminar. Paraan ng aktibidad. Dryopithecus. Extraterrestrial na landas ng pinagmulan ng tao. Mga lysosome. Organisasyon ng kemikal. Gymnosperms. Metabolismo. Mga Analyzer. System-activity approach sa pagtuturo ng biology. Mga Chromosome. Cytoplasm. Pagkabulag. Haba ng tainga. Pag-uuri ng isang tao. Skeleton ng isang mammal. Mga landas ng ebolusyon ng tao. Mitosis. Mababaw na kumplikado. Problemadong tanong. Nucleolus. Nuklear na sobre.
"Computer sa Biology" - Collaborative na aktibidad ng mga mag-aaral. Angiosperm na pamilya. Interactive na pagsasanay. Mga modelo ng pag-aaral. Isang halimbawa ng sistema ng pagtatasa. Mga tanong sa instruction card. Halimbawa ng instruction card. Mga mananaliksik. Mga microgroup. Mga interactive na teknolohiya sa pag-aaral. Carousel. Mga interactive na teknolohiya sa pag-aaral. Mga interactive na diskarte sa mga aralin sa biology. Pangkatang anyo ng trabaho. Mga gawain para sa mga pangkat ng "mga mananaliksik".
