អត្ថបទ
លើប្រធានបទ៖ "ការបន្តពូជ"
សេចក្តីផ្តើម ៣
1. ប្រភេទនៃការបន្តពូជ 4
១.១ ការបន្តពូជដោយភេទ ៤
១.២ ការបន្តពូជផ្លូវភេទ ៦
2. ការអភិវឌ្ឍន៍បុគ្គលនៃសារពាង្គកាយ 10
2.1 រយៈពេលនៃការអភិវឌ្ឍន៍អំប្រ៊ីយ៉ុង 10
២.២ រយៈពេលក្រោយការវិវឌ្ឍន៍ ១៣
2.3 គំរូទូទៅនៃការអភិវឌ្ឍន៍។ ច្បាប់ជីវវិទ្យា ១៥
សេចក្តីសន្និដ្ឋាន 18
ឯកសារយោង 18
សេចក្តីផ្តើម
សមត្ថភាពក្នុងការបន្តពូជ, i.e. ការបង្កើតមនុស្សជំនាន់ថ្មីនៃប្រភេទដូចគ្នា គឺជាលក្ខណៈសំខាន់មួយនៃសារពាង្គកាយមានជីវិត។ ក្នុងអំឡុងពេលនៃការបន្តពូជ សម្ភារៈហ្សែនត្រូវបានផ្ទេរពីជំនាន់មេទៅជំនាន់ក្រោយ ដែលធានាដល់ការបន្តពូជនៃលក្ខណៈមិនត្រឹមតែនៃប្រភេទសត្វដែលបានផ្តល់ឱ្យប៉ុណ្ណោះទេ ប៉ុន្តែនៃបុគ្គលមាតាបិតាជាក់លាក់។ សម្រាប់ប្រភេទសត្វ អត្ថន័យនៃការបន្តពូជគឺដើម្បីជំនួសអ្នកតំណាងរបស់វាដែលបានស្លាប់ ដែលធានាដល់ការបន្តនៃអត្ថិភាពនៃប្រភេទសត្វ។ លើសពីនេះទៀតនៅក្រោមលក្ខខណ្ឌសមរម្យ ការបន្តពូជធ្វើឱ្យវាអាចបង្កើនចំនួនសរុបនៃប្រភេទសត្វ។
បុគ្គលថ្មីនីមួយៗ មុននឹងឈានដល់ដំណាក់កាលដែលវាមានសមត្ថភាពបន្តពូជ ត្រូវឆ្លងកាត់ដំណាក់កាលជាច្រើននៃការលូតលាស់ និងការអភិវឌ្ឍន៍។ បុគ្គលមួយចំនួនបានស្លាប់មុនពេលឈានដល់ដំណាក់កាលបន្តពូជ (ឬភាពចាស់ទុំខាងផ្លូវភេទ) ដែលជាលទ្ធផលនៃការបំផ្លាញដោយសត្វមំសាសី ជំងឺ និងព្រឹត្តិការណ៍ចៃដន្យផ្សេងៗ។ ដូច្នេះ ប្រភេទសត្វអាចរស់បានតែក្នុងលក្ខខណ្ឌដែលជំនាន់នីមួយៗបង្កើតកូនចៅច្រើនជាងមានបុគ្គលមេដែលបានចូលរួមក្នុងការបន្តពូជ។ ទំហំប្រជាជនប្រែប្រួលអាស្រ័យលើតុល្យភាពរវាងការបន្តពូជ និងការផុតពូជរបស់បុគ្គល។ មានយុទ្ធសាស្ត្រផ្សព្វផ្សាយផ្សេងៗគ្នាជាច្រើន ដែលនីមួយៗមានគុណសម្បត្តិ និងគុណវិបត្តិខុសៗគ្នា។ ពួកគេទាំងអស់នឹងត្រូវបានពិពណ៌នានៅក្នុងអរូបីនេះ។
1. ប្រភេទនៃការបន្តពូជ
ទម្រង់ផ្សេងៗនៃការបន្តពូជត្រូវបានគេស្គាល់ ប៉ុន្តែពួកវាអាចបញ្ចូលគ្នាជាពីរប្រភេទគឺ ផ្លូវភេទ និងផ្លូវភេទ។
ការបន្តពូជផ្លូវភេទសំដៅទៅលើការផ្លាស់ប្តូរនៃជំនាន់ និងការវិវត្តនៃសារពាង្គកាយពីកោសិកាផ្លូវភេទពិសេសដែលបង្កើតឡើងនៅក្នុង gonads ។ ក្នុងករណីនេះ សារពាង្គកាយថ្មីមានការវិវឌ្ឍន៍ជាលទ្ធផលនៃការលាយបញ្ចូលគ្នានៃកោសិកាមេជីវិតពីរដែលបង្កើតឡើងដោយឪពុកម្តាយផ្សេងគ្នា។ ទោះជាយ៉ាងណាក៏ដោយ នៅក្នុងសត្វដែលមិនមានឆ្អឹងខ្នង មេជីវិតឈ្មោល និងស៊ុតត្រូវបានបង្កើតឡើងជាញឹកញាប់នៅក្នុងរាងកាយនៃសារពាង្គកាយមួយ។ បាតុភូតនេះ ភាពទ្វេភេទ ហៅថា អរិយសច្ច ។ រុក្ខជាតិផ្កាក៏មានលក្ខណៈទ្វេរភេទផងដែរ។ នៅក្នុងប្រភេទភាគច្រើននៃរុក្ខជាតិ angiosperms (ផ្កា) ផ្កា bisexual រួមមានទាំង stamens ដែលបង្កើតកោសិកាផ្លូវភេទបុរស (កោសិកាមេជីវិតឈ្មោល) និង pistils ដែលមានពង។ ប្រហែលមួយភាគបួននៃប្រភេទនេះ ផ្កាឈ្មោល និងផ្កាញី (ផ្កាស្តៅ) អភិវឌ្ឍដោយឯករាជ្យពោលគឺឧ។ ផ្ការបស់ពួកគេគឺមិនមានភេទ។ ឧទាហរណ៏មួយគឺ hemp ។ នៅក្នុងរុក្ខជាតិមួយចំនួន (ពោត, birch) ទាំងផ្កាឈ្មោលនិងស្រីលេចឡើងនៅលើបុគ្គលដូចគ្នា។
ប្រភេទសត្វ និងរុក្ខជាតិមួយចំនួនមានការរីកចម្រើន
ស៊ុតដែលមិនបានបង្កកំណើត។ ប្រភេទនៃការបន្តពូជនេះត្រូវបានគេហៅថា ព្រហ្មចារី ឬ parthenogenetic ។
ការបន្តពូជដោយភេទត្រូវបានកំណត់លក្ខណៈដោយការពិតដែលថាបុគ្គលថ្មីវិវឌ្ឍចេញពីកោសិកា somatic (រាងកាយ) ដែលមិនមែនជាផ្លូវភេទ។
1.1 ការបន្តពូជដោយភេទ
ជាមួយនឹងការបន្តពូជដោយភេទ សរីរាង្គថ្មីមួយអាចកើតចេញពីកោសិកាតែមួយ ឬពីកោសិកា asexual (somatic) ជាច្រើនរបស់ម្តាយ។ ការបន្តពូជដោយភេទដូចគ្នាពាក់ព័ន្ធនឹងឪពុកម្តាយតែមួយ។ ដោយសារកោសិកាដែលបង្កើតឱ្យមានសារពាង្គកាយកូនស្រីកើតឡើងជាលទ្ធផលនៃជំងឺ mitosis កូនចៅទាំងអស់នឹងមានលក្ខណៈស្រដៀងនឹងលក្ខណៈតំណពូជចំពោះបុគ្គលមាតា។
អង្ករ។ 1. ការបន្តពូជនៃ euglena ពណ៌បៃតង
protozoa ជាច្រើន (amoeba, euglena បៃតង។ ផ្សិត unicellular ផ្សេងទៀត algae (chlorella) សត្វ ឧទាហរណ៍ ភ្នាក់ងារមូលហេតុនៃជំងឺគ្រុនចាញ់ plasmodium គ្រុនចាញ់ ត្រូវបានកំណត់លក្ខណៈដោយ sporulation ។ ក្នុងករណីនេះ កោសិកាបំបែកទៅជាបុគ្គលមួយចំនួនធំ ស្មើនឹងចំនួនស្នូលដែលបានបង្កើតឡើងពីមុននៅក្នុងកោសិកាមេ ដែលជាលទ្ធផលនៃការបែងចែកស្នូលរបស់វាម្តងហើយម្តងទៀត។ សារពាង្គកាយពហុកោសិកាក៏មានសមត្ថភាពបង្កើត sporulation ផងដែរ: ទាំងនេះគឺជាស្លែ, ផ្សិតខ្ពស់, សារាយពហុកោសិកា, pteriophytes និងមួយចំនួនផ្សេងទៀត។
ទាំងនៅក្នុងសារពាង្គកាយ unicellular និង multicellular, budding ក៏ជាវិធីសាស្រ្តនៃការបន្តពូជ asexual ផងដែរ។ ឧទាហរណ៍នៅក្នុងផ្សិតផ្សិត និង ciliates មួយចំនួន (ស៊ីលីតដែលបឺត) នៅពេលដែលដុះលើកោសិកាមេ មើមតូចមួយដែលមានស្នូល ពន្លកត្រូវបានបង្កើតឡើងដំបូង។ វាធំឡើង ឈានដល់ទំហំជិតនឹងរាងកាយរបស់ម្តាយ ហើយបន្ទាប់មកបំបែក បន្តទៅអត្ថិភាពឯករាជ្យ។ នៅក្នុងសារពាង្គកាយពហុកោសិកា (hydra ទឹកសាប) តំរងនោមមានក្រុមកោសិកាពីស្រទាប់ទាំងពីរនៃជញ្ជាំងរាងកាយ។ ពន្លកដុះលូតលាស់ វែង ហើយមាត់មួយលេចឡើងនៅចុងខាងមុខរបស់វា ហ៊ុំព័ទ្ធដោយតង់។ ការចេញផ្កាបញ្ចប់ដោយការបង្កើត hydra តូចមួយដែលបន្ទាប់មកបំបែកចេញពីសារពាង្គកាយម្តាយ។
នៅក្នុងសត្វពហុកោសិកា ការបន្តពូជដោយភេទដូចគ្នាកើតឡើងក្នុងវិធីដូចគ្នា (ចាហួយ, អន្ទង់, ដង្កូវនាង, អេកណូឌែម) ។ ពីផ្នែកនីមួយៗ បុគ្គលដែលពោរពេញដោយសមត្ថភាពរីកចម្រើន។
ការបន្តពូជរបស់រុក្ខជាតិគឺរីករាលដាលនៅក្នុងរុក្ខជាតិ, i.e. ការកាត់ផ្នែករាងកាយ, tendrils, មើម។ ដូច្នេះដំឡូងបន្តពូជដោយផ្នែកក្រោមដីដែលបានកែប្រែនៃដើម - មើម។ ផ្កាម្លិះ និង willow និង cuttings យក root បានយ៉ាងងាយស្រួល។ ទំពាំងបាយជូ, currants និង gooseberries ត្រូវបានបន្តពូជដោយប្រើ cuttings ។
ដើមដុះវែងនៃផ្លែស្ត្របឺរី បង្កើតជាពន្លកដុះពន្លក បង្កើតបានជារុក្ខជាតិថ្មី។ រុក្ខជាតិមួយចំនួនដូចជា begonia អាចត្រូវបានបន្តពូជដោយការកាត់ស្លឹក (ស្លឹកស្លឹក និង petiole) ។ នៅផ្នែកខាងក្រោមនៃស្លឹក នៅកន្លែងដែលមានសាខាសរសៃធំៗ ឫសលេចឡើង នៅផ្នែកខាងលើមានពន្លក ហើយបន្ទាប់មកពន្លក។
ឫសក៏ត្រូវបានគេប្រើសម្រាប់ការបន្តពូជលូតលាស់ផងដែរ។ នៅក្នុងការថែសួន ផ្លែ Raspberries, cherries, plums និងផ្កាកុលាបត្រូវបានបន្តពូជដោយប្រើ cuttings ពីឫសនៅពេលក្រោយ។ Dahlias បន្តពូជដោយប្រើមើមឫស។ ការកែប្រែផ្នែកក្រោមដីនៃដើម rhizome ក៏បង្កើតជារុក្ខជាតិថ្មី។ ជាឧទាហរណ៍ ការសាបព្រួសស្មៅដោយប្រើមើមអាចបង្កើតបានមនុស្សថ្មីជាងមួយពាន់នាក់ក្នុង 1 ម 2 នៃដី។
1.2 ការបន្តពូជផ្លូវភេទ
ការបន្តពូជផ្លូវភេទមានគុណសម្បត្តិវិវត្តន៍ដ៏អស្ចារ្យណាស់បើប្រៀបធៀបទៅនឹងការបន្តពូជដោយភេទ។ នេះគឺដោយសារតែការពិតដែលថា genotype នៃ offspring កើតឡើងដោយការរួមបញ្ចូលហ្សែនដែលជាកម្មសិទ្ធិរបស់ឪពុកម្តាយទាំងពីរ។ ជាលទ្ធផលសមត្ថភាពរបស់សារពាង្គកាយក្នុងការសម្របខ្លួនទៅនឹងលក្ខខណ្ឌបរិស្ថានកើនឡើង។ ចាប់តាំងពីការផ្សំថ្មីត្រូវបានអនុវត្តនៅក្នុងជំនាន់នីមួយៗ បុគ្គលមួយចំនួនធំអាចត្រូវបានសម្របខ្លួនទៅនឹងលក្ខខណ្ឌថ្មីនៃអត្ថិភាពជាងអំឡុងពេលបន្តពូជដោយភេទ។ ការលេចឡើងនៃការរួមបញ្ចូលគ្នានៃហ្សែនថ្មីធានាបាននូវការសម្របខ្លួនកាន់តែជោគជ័យ និងឆាប់រហ័សនៃប្រភេទសត្វទៅនឹងការផ្លាស់ប្តូរលក្ខខណ្ឌបរិស្ថាន។
ដូច្នេះខ្លឹមសារនៃការបន្តពូជផ្លូវភេទស្ថិតនៅក្នុងការរួមបញ្ចូលគ្នានៅក្នុងសម្ភារៈតំណពូជនៃកូនចៅនៃពត៌មានហ្សែនពីប្រភពពីរផ្សេងគ្នា - ឪពុកម្តាយ។
កោសិកាផ្លូវភេទមានការរីកចម្រើននៅក្នុង gonads: មេជីវិតឈ្មោល, អូវុលស្ត្រី (ឬស៊ុត) ។ ក្នុងករណីដំបូងការអភិវឌ្ឍន៍របស់ពួកគេត្រូវបានគេហៅថា spermatogenesis នៅក្នុងទីពីរ - oogenesis (ពីឡាតាំង ovo - ស៊ុត) ។
នៅក្នុងដំណើរការនៃការបង្កើតកោសិកាមេរោគ ដំណាក់កាលមួយចំនួនត្រូវបានសម្គាល់។ ដំណាក់កាលទី 1 គឺជាដំណាក់កាលនៃការបន្តពូជ ដែលកោសិកាដំណុះដំបូងត្រូវបានបែងចែកដោយ mitosis ដែលបណ្តាលឱ្យមានការកើនឡើងនៃចំនួនរបស់វា។
ដំណាក់កាលទីពីរគឺជារយៈពេលនៃការលូតលាស់។ នៅក្នុង gametes បុរសមិនទាន់ពេញវ័យវាមិនត្រូវបានគេបញ្ចេញសម្លេងទេ។ ទំហំរបស់ពួកគេកើនឡើងបន្តិច។ ផ្ទុយទៅវិញ ស៊ុតនាពេលអនាគត oocytes កើនឡើងក្នុងទំហំ ជួនកាលរាប់រយ និងច្រើនដងរាប់ពាន់លានដង។ ការលូតលាស់នៃ oocytes ត្រូវបានអនុវត្តដោយសារតែសារធាតុដែលបង្កើតឡើងដោយកោសិកាផ្សេងទៀតនៃរាងកាយ។ ដូច្នេះនៅក្នុងត្រី អំពែរ និងក្នុងវិសាលភាពធំជាងនេះ នៅក្នុងសត្វល្មូន និងសត្វស្លាប ភាគច្រើននៃស៊ុតគឺ yolk ។ វាត្រូវបានសំយោគនៅក្នុងថ្លើម ដឹកជញ្ជូនក្នុងទម្រង់ពិសេសរលាយដោយឈាមទៅកាន់អូវែរ ជ្រាបចូលទៅក្នុង oocytes ដែលកំពុងលូតលាស់ ហើយត្រូវបានដាក់នៅទីនោះក្នុងទម្រង់ជាបន្ទះពណ៌លឿង។ លើសពីនេះទៀតនៅក្នុងកោសិកាបន្តពូជនាពេលអនាគតខ្លួនវាប្រូតេអ៊ីនជាច្រើននិង RNAs មួយចំនួនធំត្រូវបានសំយោគ: ការដឹកជញ្ជូន ribosomal និងព័ត៌មាន។ yolk គឺជាការប្រមូលផ្ដុំនៃសារធាតុចិញ្ចឹម (ខ្លាញ់ ប្រូតេអ៊ីន កាបូអ៊ីដ្រាត វីតាមីន។
ដំណាក់កាលបន្ទាប់ ដំណាក់កាលនៃភាពចាស់ទុំ ឬ meiosis ត្រូវបានបង្ហាញក្នុងរូបភាពទី 2 ។ កោសិកាដែលចូលក្នុងដំណាក់កាលនៃភាពចាស់ទុំមានសំណុំក្រូម៉ូសូម diploid និងចំនួន DNA កើនឡើងទ្វេដងរួចទៅហើយ។

អង្ករ។ 2. ភាពចាស់ទុំនៃកោសិកាមេរោគ (meiosis)
ខ្លឹមសារនៃ meiosis គឺថាកោសិកាផ្លូវភេទនីមួយៗទទួលបានសំណុំក្រូម៉ូសូម haploid តែមួយ។ ទោះបីជាយ៉ាងណាក៏ដោយ ក្នុងពេលជាមួយគ្នានោះ meiosis គឺជាដំណាក់កាលមួយដែលការរួមផ្សំថ្មីនៃហ្សែនត្រូវបានបង្កើតឡើងដោយការបញ្ចូលគ្នារវាងក្រូម៉ូសូមមាតា និងបិតាផ្សេងៗគ្នា ការបញ្ចូលគ្នានៃទំនោរតំណពូជក៏កើតឡើងជាលទ្ធផលនៃការឆ្លងកាត់ - ការផ្លាស់ប្តូរផ្នែករវាងក្រូម៉ូសូមដូចគ្នាក្នុងអំឡុងពេលដំណើរការ។ នៃ meiosis ។
Meiosis ពាក់ព័ន្ធនឹងការបែងចែកពីរជាបន្តបន្ទាប់។ ដូចនៅក្នុង mitosis ការបែងចែក meiotic នីមួយៗមាន 4 ដំណាក់កាល: prophase, metaphase, anaphase និង telophase ។
ទីមួយ (I) ការបែងចែក meiotic ។ Prophase I ចាប់ផ្តើមជាមួយនឹងការបំប្លែងក្រូម៉ូសូម។ ដូចដែលអ្នកចងចាំ ក្រូម៉ូសូមនីមួយៗមានក្រូម៉ាទីតពីរដែលតភ្ជាប់នៅកណ្តាល។ បន្ទាប់មក ក្រូម៉ូសូមដូចគ្នាចូលជិតគ្នា ចំណុចនីមួយៗនៃក្រូម៉ូសូមនីមួយៗនៃក្រូម៉ូសូមមួយត្រូវបានផ្សំជាមួយចំណុចដែលត្រូវគ្នានៃក្រូម៉ូសូមនៃក្រូម៉ូសូមដូចគ្នាមួយទៀត។ ដំណើរការនៃភាពច្បាស់លាស់ និងជិតស្និទ្ធនេះ ប្រមូលផ្តុំនៃក្រូម៉ូសូម homologous នៅក្នុង meiosis ត្រូវបានគេហៅថា conjugation ។ នៅពេលអនាគត ការឆ្លងកាត់អាចកើតឡើងរវាងក្រូម៉ូសូមបែបនេះ ដែលជាការផ្លាស់ប្តូរនៃដូចគ្នា ឬដូចគ្នា ពោលគឺមានហ្សែនដូចគ្នា តំបន់។ ឆ្ពោះទៅចុងបញ្ចប់នៃ prophase កម្លាំងច្រណែនកើតឡើងរវាងក្រូម៉ូសូមដូចគ្នា។ ទីមួយពួកវាលេចឡើងនៅតំបន់កណ្តាល ហើយបន្ទាប់មកនៅតំបន់ផ្សេងទៀត។
នៅក្នុង metaphase I, chromosome spiralization គឺអតិបរមា។ ក្រូម៉ូសូមរួមមានទីតាំងនៅតាមបណ្តោយអេក្វាទ័រ ដោយមានចំណុចកណ្តាលនៃក្រូម៉ូសូមដូចគ្នាប្រឈមមុខនឹងបង្គោលផ្សេងៗនៃកោសិកា។ ខ្សែស្រឡាយ spindle ត្រូវបានភ្ជាប់ជាមួយពួកគេ។
នៅក្នុង anaphase I ដៃនៃក្រូម៉ូសូម homologous ទីបំផុតបំបែកចេញពីគ្នា ហើយក្រូម៉ូសូមផ្លាស់ទីទៅប៉ូលផ្សេងគ្នា។ អាស្រ័យហេតុនេះ ពីគូនៃក្រូម៉ូសូមដូចគ្នា មានតែមួយប៉ុណ្ណោះចូលទៅក្នុងកោសិកាកូនស្រី។ ចំនួនក្រូម៉ូសូមថយចុះពាក់កណ្តាល សំណុំក្រូម៉ូសូមក្លាយជា haploid ។ ទោះយ៉ាងណាក៏ដោយ ក្រូម៉ូសូមនីមួយៗមានក្រូម៉ាតពីរ ពោលគឺវានៅតែផ្ទុក DNA ទ្វេដង។
នៅក្នុង telophase I ស្រោមសំបុត្រនុយក្លេអ៊ែរត្រូវបានបង្កើតឡើងក្នុងរយៈពេលខ្លី។ ក្នុងអំឡុងពេលអន្តរដំណាក់កាលរវាងការបែងចែកទីមួយនិងទីពីរនៃ meiosis ការចម្លង DNA មិនកើតឡើងទេ។ កោសិកាដែលបង្កើតឡើងជាលទ្ធផលនៃការបែងចែកដំបូងនៃភាពចាស់ទុំមានភាពខុសគ្នានៅក្នុងសមាសភាពនៃក្រូម៉ូសូមមាតា និងមាតា ហើយជាលទ្ធផលនៅក្នុងសំណុំនៃហ្សែន។
ជាឧទាហរណ៍ កោសិកាមនុស្សទាំងអស់ រួមទាំងកោសិកាមេជីវិតដំបូង មានក្រូម៉ូសូមចំនួន 46 ។ ក្នុងនោះ ២៣នាក់ទទួលពីឪពុក និង២៣នាក់ពីម្តាយ។ នៅពេលដែលកោសិកាមេរោគត្រូវបានបង្កើតឡើងបន្ទាប់ពីការបែងចែក meiotic ដំបូង spermatocytes និង oocytes ក៏ទទួលបានក្រូម៉ូសូមចំនួន 23 ផងដែរ។ ទោះបីជាយ៉ាងណាក៏ដោយ ដោយសារតែភាពចៃដន្យនៃភាពខុសគ្នានៃក្រូម៉ូសូមមាតា និងបិតានៅក្នុងអាណាផាស I កោសិកាលទ្ធផលទទួលបានភាពខុសគ្នាជាច្រើននៃការរួមបញ្ចូលគ្នានៃក្រូម៉ូសូមមាតាបិតា។ ឧទាហរណ៍ មួយក្នុងចំណោមពួកគេអាចមានក្រូម៉ូសូមមាតា 3 និង 20 មាតាបិតា 10 និងមាតា 13 ផ្សេងទៀត មាតាបិតា 20 និងមាតា 3 ។ល។ ចំនួននៃបន្សំដែលអាចធ្វើបានគឺមានទំហំធំណាស់។ ប្រសិនបើយើងពិចារណាផងដែរអំពីការផ្លាស់ប្តូរផ្នែកដូចគ្នានៃក្រូម៉ូសូមនៅក្នុងដំណាក់កាលនៃការបែងចែកដំបូងនៃ meiosis នោះវាច្បាស់ណាស់ថាកោសិកាមេរោគនីមួយៗមានហ្សែនតែមួយគត់ព្រោះវាផ្ទុកនូវសំណុំហ្សែនតែមួយគត់របស់វា។
ជាលទ្ធផល meiosis គឺជាមូលដ្ឋាននៃភាពប្រែប្រួលហ្សែនរួមបញ្ចូលគ្នា។
ទីពីរ (II) ការបែងចែក meiotic ។ ការបែងចែកទីពីរនៃ meiosis ជាទូទៅដំណើរការតាមរបៀបដូចគ្នានឹងការបែងចែក mitotic ធម្មតា ដោយភាពខុសគ្នាតែមួយគត់គឺថាកោសិកាបែងចែកគឺ haploid ។ នៅក្នុង anaphase II, centromeres ដែលភ្ជាប់ក្រូម៉ូសូមបងស្រីនៅក្នុងការបែងចែកក្រូម៉ូសូមនីមួយៗ ហើយក្រូម៉ូសូមដូចជានៅក្នុង mitosis ចាប់ពីពេលនេះក្លាយជាក្រូម៉ូសូមឯករាជ្យ។ ជាមួយនឹងការបញ្ចប់នៃ telophase II ដំណើរការទាំងមូលនៃ meiosis បានបញ្ចប់: កោសិកា haploid ចំនួនបួនត្រូវបានបង្កើតឡើងពីកោសិកាមេដើមដំបូង។
ចំពោះបុរស ពួកវាទាំងអស់ត្រូវបានបំប្លែងទៅជា gametes - មេជីវិតឈ្មោល។ ចំពោះស្ត្រី ដោយសារតែ meiosis មិនស្មើគ្នា មានតែកោសិកាមួយប៉ុណ្ណោះដែលផលិតស៊ុតដែលអាចសម្រេចបាន។ កោសិកាកូនស្រីបីផ្សេងទៀតមានទំហំតូចជាងនេះ វាបានក្លាយទៅជាអ្វីដែលហៅថាការណែនាំ ឬការកាត់បន្ថយជារាងកាយ ដែលឆាប់ស្លាប់។ តាមទស្សនៈជីវសាស្រ្ត ការបង្កើតស៊ុតតែមួយ និងការស្លាប់នៃសាកសពមគ្គុទ្ទេសក៍ពេញលេញនៃហ្សែនចំនួនបី គឺដោយសារតែតម្រូវការក្នុងការរក្សាទុកនៅក្នុងកោសិកាតែមួយនូវសារធាតុបម្រុងទាំងអស់ដែលនឹងត្រូវការសម្រាប់ការអភិវឌ្ឍនៃអំប្រ៊ីយ៉ុងនាពេលអនាគត។
កំឡុងពេលបង្កើតមានកោសិកាដែលទទួលបាននូវរូបរាង និងទំហំជាក់លាក់មួយដែលត្រូវនឹងមុខងាររបស់វា។
កំឡុងពេលដំណើរការនៃភាពចាស់ទុំ កោសិកាមេជីវិតឈ្មោលត្រូវបានគ្របដណ្ដប់ដោយភ្នាស ហើយត្រៀមខ្លួនសម្រាប់ការបង្កកំណើតភ្លាមៗបន្ទាប់ពីការបញ្ចប់នៃ meiosis ។ ក្នុងករណីជាច្រើន ឧទាហរណ៍នៅក្នុងសត្វល្មូន សត្វស្លាប និងថនិកសត្វ ដោយសារតែសកម្មភាពនៃកោសិកាជុំវិញស៊ុត ភ្នាសបន្ថែមមួយចំនួនបានលេចឡើងនៅជុំវិញវា។ មុខងាររបស់ពួកគេគឺការពារស៊ុត និងបង្កើតអំប្រ៊ីយ៉ុងពីឥទ្ធិពលអវិជ្ជមានពីខាងក្រៅ។ មេជីវិតឈ្មោលអាចមានទំហំ និងរាងខុសៗគ្នា។
មុខងាររបស់មេជីវិតឈ្មោលគឺបញ្ជូនព័ត៌មានហ្សែនទៅស៊ុត និងជំរុញការអភិវឌ្ឍន៍របស់វា។ មេជីវិតឈ្មោលដែលបានបង្កើតឡើងមានផ្ទុកនូវ mitochondria ដែលជាឧបករណ៍ Golgi ដែលបញ្ចេញអង់ស៊ីមដែលរំលាយភ្នាសស៊ុតអំឡុងពេលបង្កកំណើត ពោលគឺកំឡុងពេលការលាយបញ្ចូលគ្នានៃមេជីវិតឈ្មោល និងស៊ុត។ កោសិកា diploid លទ្ធផលត្រូវបានគេហៅថា zygote ។
2. ការអភិវឌ្ឍន៍បុគ្គលនៃសារពាង្គកាយ
ការអភិវឌ្ឍន៍បុគ្គល ឬ ontogenesis សំដៅលើរយៈពេលទាំងមូលនៃជីវិតរបស់បុគ្គល ចាប់ពីពេលដែលមេជីវិតឈ្មោលបញ្ចូលគ្នាជាមួយស៊ុត និងការបង្កើតហ្សីហ្គោតរហូតដល់ការស្លាប់របស់សារពាង្គកាយ។ Ontogenesis ត្រូវបានបែងចែកជាពីរដំណាក់កាល៖ 1) អំប្រ៊ីយ៉ុងពីការបង្កើតហ្សីហ្គោតរហូតដល់កំណើតឬចេញពីភ្នាសស៊ុត។ 2) postembryonic ពីការចេញពីភ្នាសស៊ុតឬពីកំណើតដល់ស្លាប់នៃសារពាង្គកាយ។
វិទ្យាសាស្ត្រដែលសិក្សាពីគំរូនៃការអភិវឌ្ឍន៍បុគ្គលនៃសារពាង្គកាយនៅដំណាក់កាលអំប្រ៊ីយ៉ុងត្រូវបានគេហៅថា អំប្រ៊ីយ៉ុង (មកពីអំប្រ៊ីយ៉ុងក្រិក) ។
2.1 រយៈពេលនៃការអភិវឌ្ឍន៍អំប្រ៊ីយ៉ុង
នៅក្នុងសត្វពហុកោសិកា ដោយមិនគិតពីភាពស្មុគស្មាញនៃអង្គការរបស់ពួកគេ ដំណាក់កាលនៃការអភិវឌ្ឍន៍អំប្រ៊ីយ៉ុងដែលអំប្រ៊ីយ៉ុងឆ្លងកាត់គឺដូចគ្នា។ នៅក្នុងអំប្រ៊ីយ៉ុងមានដំណាក់កាលសំខាន់ៗចំនួនបីគឺៈ ការបំបែក, gastrulation និង organogenesis បឋម។
ការបំបែក។ ការអភិវឌ្ឍនៃសារពាង្គកាយមួយចាប់ផ្តើមនៅដំណាក់កាលកោសិកាតែមួយ។ ស៊ុតបង្កកំណើតគឺជាកោសិកាមួយ ហើយក្នុងពេលជាមួយគ្នានោះសារពាង្គកាយមួយនៅដំណាក់កាលដំបូងនៃការអភិវឌ្ឍន៍របស់វា។ ជាលទ្ធផលនៃការបែងចែកម្តងហើយម្តងទៀត សារពាង្គកាយកោសិកាតែមួយប្រែទៅជាពហុកោសិកា។ ស្នូល diploid ដែលលេចឡើងក្នុងអំឡុងពេលបង្កកំណើតតាមរយៈការលាយបញ្ចូលគ្នានៃមេជីវិតឈ្មោល និងស៊ុតមួយ ចាប់ផ្តើមបែងចែកក្នុងរយៈពេលពីរបីនាទី ហើយ cytoplasm ក៏បែងចែកជាមួយវាផងដែរ។ កោសិកាលទ្ធផលថយចុះក្នុងទំហំជាមួយនឹងការបែងចែកនីមួយៗ ដូច្នេះដំណើរការបែងចែកត្រូវបានគេហៅថា cleavage ។ ក្នុងអំឡុងពេលនៃការបែងចែកសម្ភារៈកោសិកាប្រមូលផ្តុំសម្រាប់ការអភិវឌ្ឍបន្ថែមទៀត។ ការបែកខ្ញែកបញ្ចប់ដោយការបង្កើតអំប្រ៊ីយ៉ុងពហុកោសិកាគឺ ប្លស្លូឡា។ blastula មានបែហោងធ្មែញដែលពោរពេញទៅដោយសារធាតុរាវដែលហៅថាបែហោងធ្មែញរាងកាយបឋម។
ក្នុងករណីដែលមានពណ៌លឿងតិចតួចនៅក្នុង cytoplasm នៃស៊ុត (ដូចនៅក្នុង lancelet) ឬតិចតួច (ដូចនៅក្នុងកង្កែប) ការបំបែកគឺពេញលេញ ពោលគឺស៊ុតបានបែងចែកទាំងស្រុង។
បើមិនដូច្នោះទេរយៈពេលនៃការបែកបាក់កើតឡើងនៅក្នុងបក្សី។ cytoplasm គ្មាន yolk បង្កើតបានតែ 1% នៃបរិមាណសរុបនៃស៊ុតមាន់មួយ; cytoplasm ដែលនៅសេសសល់ទាំងមូលនៃស៊ុត ហើយដូច្នេះ zygote ត្រូវបានបំពេញដោយម៉ាស់នៃ yolk ។ ប្រសិនបើអ្នកក្រឡេកមើលពងមាន់ឱ្យជិត នោះនៅលើបង្គោលមួយរបស់វាដោយផ្ទាល់នៅលើ yolk អ្នកអាចមើលឃើញចំណុចតូចមួយ - blastula ឬឌីសមេរោគដែលបង្កើតឡើងជាលទ្ធផលនៃការកំទេចផ្នែកដែលគ្មានពណ៌លឿងនៃ cytoplasm ដែលមានស្នូល។ ក្នុងករណីបែបនេះការកំទេចត្រូវបានគេហៅថាមិនពេញលេញ។ ការបំបែកមិនពេញលេញក៏ជាលក្ខណៈនៃត្រី និងសត្វល្មូនមួយចំនួនផងដែរ។
នៅក្នុងគ្រប់ករណីទាំងអស់ ទាំងនៅក្នុង lancelet និងនៅក្នុង amphibians និងនៅក្នុងបក្សីក៏ដូចជានៅក្នុងសត្វដទៃទៀត បរិមាណសរុបនៃកោសិកានៅដំណាក់កាល blastula មិនលើសពីបរិមាណនៃ zygote នោះទេ។ ម៉្យាងទៀតការបែងចែក mitotic នៃ zygote មិនត្រូវបានអមដោយការរីកលូតលាស់នៃកោសិកាកូនស្រីលទ្ធផលទៅនឹងបរិមាណរបស់ម្តាយនោះទេ ហើយទំហំរបស់ពួកគេជាលទ្ធផលនៃការបែងចែកជាបន្តបន្ទាប់បន្តបន្ទាប់គ្នាថយចុះ។ លក្ខណៈពិសេសនៃការបែងចែកកោសិកា mitotic ក្នុងអំឡុងពេលបំបែកត្រូវបានគេសង្កេតឃើញក្នុងអំឡុងពេលនៃការបង្កើតស៊ុតបង្កកំណើតនៅក្នុងសត្វទាំងអស់។
លក្ខណៈពិសេសមួយចំនួនផ្សេងទៀតនៃការកំទេចក៏ជាលក្ខណៈនៃប្រភេទសត្វផ្សេងៗផងដែរ។ ឧទាហរណ៍ កោសិកាទាំងអស់នៅក្នុង blastula មានសំណុំក្រូម៉ូសូម diploid គឺដូចគ្នាបេះបិទនៅក្នុងរចនាសម្ព័ន្ធ និងខុសគ្នាពីគ្នាទៅវិញទៅមកជាចម្បងនៅក្នុងបរិមាណនៃ yolk ដែលពួកគេផ្ទុក។ កោសិកាបែបនេះដែលខ្វះសញ្ញានៃឯកទេសដើម្បីបំពេញមុខងារជាក់លាក់ ត្រូវបានគេហៅថាកោសិកាដែលមិនពិសេស (ឬមិនខុសគ្នា)។ លក្ខណៈពិសេសមួយទៀតនៃការបំបែកគឺវដ្តខ្លីបំផុតនៃ blastomeres បើប្រៀបធៀបទៅនឹងកោសិកានៃសារពាង្គកាយពេញវ័យ។ ក្នុងអំឡុងពេលអន្តរដំណាក់កាលខ្លីបំផុត មានតែការចម្លង DNA ប៉ុណ្ណោះដែលកើតឡើង។
ក្រពះ។ blastula តាមក្បួនមួយមាន blastomeres មួយចំនួនធំ (ឧទាហរណ៍នៅក្នុង lancelet ពី 3000 កោសិកា) ក្នុងអំឡុងពេលដំណើរការអភិវឌ្ឍន៍បានឆ្លងទៅដំណាក់កាលថ្មីមួយដែលត្រូវបានគេហៅថា gastrula (ពីក្រពះ gaster ក្រិក) ។ អំប្រ៊ីយ៉ុងនៅដំណាក់កាលនេះមានស្រទាប់កោសិកាដែលអាចបែងចែកបានយ៉ាងច្បាស់ - ដែលគេហៅថាស្រទាប់មេរោគ៖ ខាងក្រៅ ឬ ectoderm (មកពីភាសាក្រិច ectos - មានទីតាំងនៅខាងក្រៅ) និងខាងក្នុង ឬ endoderm (មកពីភាសាក្រិក entos - ដែលមានទីតាំងនៅខាងក្នុង) . សំណុំនៃដំណើរការដែលនាំទៅដល់ការបង្កើត gastrula ត្រូវបានគេហៅថា gastrulation ។
នៅក្នុង lancelet, gastrulation ត្រូវបានអនុវត្តដោយការ invagination នៃបង្គោលមួយនៃ blastula ខាងក្នុង, ឆ្ពោះទៅរកផ្សេងទៀត; នៅក្នុងសត្វផ្សេងទៀត, ដោយការ delamination នៃជញ្ជាំងនៃ blastula ឬដោយការ overgrowing បង្គោលលូតលាស់ដ៏ធំជាមួយនឹងកោសិកាតូចមួយនៃ។ បង្គោលសត្វ។
នៅក្នុងសត្វពហុកោសិកា លើកលែងតែ coelenterates ស្របជាមួយ gastrulation ឬដូចនៅក្នុង lancelet បន្ទាប់ពីវា mesoderm ស្រទាប់ទីបី (មកពីភាសាក្រិច mesos ដែលមានទីតាំងនៅកណ្តាល) លេចឡើងដែលជាសំណុំនៃធាតុកោសិកាដែលស្ថិតនៅចន្លោះ ecto- និង endoderm នៅក្នុងបែហោងធ្មែញរាងកាយបឋម blastocele ។ ជាមួយនឹងរូបរាងនៃ mesoderm អំប្រ៊ីយ៉ុងក្លាយជាបីស្រទាប់។
ដូច្នេះខ្លឹមសារនៃដំណើរការ gastrulation គឺចលនានៃកោសិកា។ កោសិកានៃអំប្រ៊ីយ៉ុងអនុវត្តបានបែងចែក និងមិនលូតលាស់។ ទោះជាយ៉ាងណាក៏ដោយនៅដំណាក់កាលនេះការប្រើប្រាស់ព័ត៌មានហ្សែននៃកោសិកាអំប្រ៊ីយ៉ុងចាប់ផ្តើមហើយសញ្ញាដំបូងនៃភាពខុសគ្នាលេចឡើង។
ភាពខុសគ្នា ឬភាពខុសគ្នា គឺជាដំណើរការនៃការកើតឡើងរបស់វា និងការកើនឡើងនៃភាពខុសគ្នានៃរចនាសម្ព័ន្ធ និងមុខងាររវាងកោសិកានីមួយៗ និងផ្នែកនៃអំប្រ៊ីយ៉ុង។ តាមទស្សនៈ morphological ភាពខុសគ្នាត្រូវបានបង្ហាញនៅក្នុងការបង្កើតកោសិការាប់រយប្រភេទនៃរចនាសម្ព័ន្ធជាក់លាក់ដែលខុសគ្នាពីគ្នាទៅវិញទៅមក។ ពីកោសិកា blastula ដែលមិនពិសេស កោសិកា epithelial នៃស្បែក ភ្នាសពោះវៀន សួត លេចចេញជាបណ្តើរៗ កោសិកាសរសៃប្រសាទ និងសាច់ដុំ។ល។ តាមទស្សនៈគីមីជីវៈ ឯកទេសកោសិកាស្ថិតនៅក្នុងសមត្ថភាពក្នុងការសំយោគប្រូតេអ៊ីនជាក់លាក់ដែលជាលក្ខណៈនៃប្រភេទកោសិកាដែលបានផ្តល់ឱ្យប៉ុណ្ណោះ។ Lymphocytes សំយោគអង់ទីករការពារប្រូតេអ៊ីន កោសិកាសាច់ដុំ contractile protein myosin ។ ប្រភេទនៃកោសិកានីមួយៗផលិតប្រូតេអ៊ីនផ្ទាល់ខ្លួនរបស់វា ដែលមានតែមួយគត់ចំពោះវា។ ឯកទេសជីវគីមីនៃកោសិកាត្រូវបានធានាដោយការជ្រើសរើស សកម្មភាពឌីផេរ៉ង់ស្យែលនៃហ្សែន ពោលគឺនៅក្នុងកោសិកានៃស្រទាប់មេរោគផ្សេងៗគ្នា បំណែកនៃសរីរាង្គ និងប្រព័ន្ធមួយចំនួន ក្រុមហ្សែនផ្សេងៗគ្នាចាប់ផ្តើមដំណើរការ។
នៅក្នុងប្រភេទសត្វផ្សេងៗគ្នា ស្រទាប់មេរោគដូចគ្នាបង្កើតឱ្យមានសរីរាង្គ និងជាលិកាដូចគ្នា។ នេះមានន័យថាពួកគេមានលក្ខណៈដូចគ្នា។ ដូច្នេះពីកោសិកានៃស្រទាប់មេរោគខាងក្រៅ - ectoderm - នៅក្នុង arthropods, chordates រួមទាំងត្រី amphibians សត្វល្មូនបក្សីនិងថនិកសត្វស្បែកនិងនិស្សន្ទវត្ថុរបស់ពួកគេក៏ដូចជាប្រព័ន្ធសរសៃប្រសាទនិងសរីរាង្គអារម្មណ៍ត្រូវបានបង្កើតឡើង។ ភាពដូចគ្នានៃស្រទាប់មេជីវិតរបស់សត្វភាគច្រើន គឺជាភស្តុតាងមួយនៃការរួបរួមនៃពិភពសត្វ។
សរីរាង្គ។ បន្ទាប់ពីការបញ្ចប់នៃ gastrulation អំប្រ៊ីយ៉ុងបង្កើតជាស្មុគ្រស្មាញនៃសរីរាង្គអ័ក្ស: បំពង់សរសៃប្រសាទ, notochord និងបំពង់ពោះវៀន។ នៅក្នុង lancelet សរីរាង្គអ័ក្សត្រូវបានបង្កើតឡើងដូចខាងក្រោម: ectoderm នៅផ្នែកខាងខ្នងនៃអំប្រ៊ីយ៉ុងពត់តាមបណ្តោយបន្ទាត់កណ្តាលប្រែទៅជាចង្អូរហើយ ectoderm ដែលមានទីតាំងនៅខាងស្តាំនិងខាងឆ្វេងរបស់វាចាប់ផ្តើមដុះនៅលើគែមរបស់វា។ ចង្អូរ, ឫសគល់នៃប្រព័ន្ធសរសៃប្រសាទ, លិចនៅក្រោម ectoderm និងគែមរបស់វានៅជិត។ បំពង់សរសៃប្រសាទត្រូវបានបង្កើតឡើង។ នៅសល់នៃ ectoderm គឺជា rudiment នៃ epithelium ស្បែក។
ផ្នែក dorsal នៃ endoderm ដែលមានទីតាំងនៅដោយផ្ទាល់នៅក្រោម rudiment សរសៃប្រសាទត្រូវបានបំបែកចេញពី endoderm ដែលនៅសល់ហើយបត់ចូលទៅក្នុងខ្សែក្រាស់ហៅថា notochord ។ ពីផ្នែកដែលនៅសល់នៃ endoderm, mesoderm និង epithelium ពោះវៀនមានការរីកចម្រើន។ ភាពខុសគ្នាបន្ថែមទៀតនៃកោសិកាអំប្រ៊ីយ៉ុងនាំទៅដល់ការលេចចេញនូវដេរីវេនៃស្រទាប់មេរោគជាច្រើន - សរីរាង្គ និងជាលិកា។ នៅក្នុងដំណើរការនៃឯកទេសនៃកោសិកាដែលបង្កើតស្រទាប់មេរោគ ប្រព័ន្ធប្រសាទ, សរីរាង្គអារម្មណ៍, epithelium ស្បែក, និង enamel ធ្មេញត្រូវបានបង្កើតឡើងពី ectoderm នេះ; ពី epithelium ពោះវៀន endoderm, ក្រពេញរំលាយអាហារថ្លើមនិងលំពែង, epithelium នៃ gills និងសួត; ពីជាលិកាសាច់ដុំ mesoderm ជាលិកាភ្ជាប់រួមទាំងជាលិកាភ្ជាប់រលុងឆ្អឹងខ្ចីនិងជាលិកាឆ្អឹងឈាមនិងកូនកណ្តុរក៏ដូចជាប្រព័ន្ធឈាមរត់តម្រងនោម gonads ។
2.2 រយៈពេល Postembryonic នៃការអភិវឌ្ឍន៍
នៅពេលចាប់កំណើត ឬការបញ្ចេញសារពាង្គកាយចេញពីសំបកស៊ុត រយៈពេលនៃអំប្រ៊ីយ៉ុងនឹងបញ្ចប់ ហើយរយៈពេលក្រោយការវិវឌ្ឍន៍ចាប់ផ្តើម។ ការអភិវឌ្ឍន៍ Postembryonic អាចត្រូវបានដោយផ្ទាល់ឬអមដោយការបំប្លែង (metamorphosis) ។
ក្នុងអំឡុងពេលនៃការអភិវឌ្ឍន៍ដោយផ្ទាល់ (នៅក្នុងសត្វល្មូន សត្វស្លាប ថនិកសត្វ) សារពាង្គកាយដែលមានទំហំតូចផុសចេញពីសំបកស៊ុត ឬពីរាងកាយរបស់ម្តាយ ប៉ុន្តែជាមួយនឹងសរីរាង្គសំខាន់ៗទាំងអស់នៃសត្វពេញវ័យបានបង្កើតឡើងរួចហើយ។ ការអភិវឌ្ឍន៍ Postembryonic ក្នុងករណីនេះត្រូវបានកាត់បន្ថយជាចម្បងទៅនឹងការលូតលាស់និងភាពពេញវ័យ។
កំឡុងពេលវិវត្តន៍ទៅជាមេតាណុល សត្វដង្កូវមួយផុសចេញពីស៊ុត ជាធម្មតាមានរចនាសម្ព័ន្ធសាមញ្ញជាងសត្វពេញវ័យ ដោយមានសរីរាង្គដង្កូវពិសេសដែលអវត្តមានក្នុងស្ថានភាពពេញវ័យ។ ដង្កូវស៊ី លូតលាស់ និងយូរៗទៅសរីរាង្គដង្កូវត្រូវបានជំនួសដោយសរីរាង្គលក្ខណៈរបស់មនុស្សពេញវ័យ។ ជាលទ្ធផលក្នុងអំឡុងពេល metamorphosis សរីរាង្គដង្កូវត្រូវបានបំផ្លាញហើយសរីរាង្គលក្ខណៈរបស់សត្វពេញវ័យលេចឡើង។
ចូរយើងក្រឡេកមើលឧទាហរណ៍មួយចំនួននៃការអភិវឌ្ឍន៍ postembryonic ដោយប្រយោល។ ដង្កូវ ascidian (phylum Chordata, subphylum Larval-Chordata) មានលក្ខណៈសំខាន់ៗទាំងអស់នៃ chordates: notochord បំពង់សរសៃប្រសាទ និង gill slits នៅក្នុង pharynx ។ វាហែលដោយសេរី បន្ទាប់មកភ្ជាប់ទៅនឹងផ្ទៃរឹងមួយចំនួននៅបាតសមុទ្រ ដែលជាកន្លែងការបំប្លែងសារជាតិកើតឡើង៖ កន្ទុយ ដុំពក និងសាច់ដុំបាត់ ហើយបំពង់សរសៃប្រសាទបានបំបែកទៅជាកោសិកានីមួយៗ ដែលភាគច្រើនត្រូវបាន phagocytosed ។ អ្វីទាំងអស់ដែលនៅសេសសល់នៃប្រព័ន្ធសរសៃប្រសាទដង្កូវគឺជាក្រុមនៃកោសិកាដែលបង្កើតឱ្យមានការប្រសាទប្រសាទ។ រចនាសម្ព័នរាងកាយរបស់ ascidian ពេញវ័យដែលដឹកនាំរបៀបរស់នៅដែលភ្ជាប់មកជាមួយ គឺមិនស្រដៀងនឹងលក្ខណៈពិសេសធម្មតានៃការរៀបចំ chordates នោះទេ។ មានតែចំណេះដឹងអំពីលក្ខណៈពិសេសនៃ ontogenesis ប៉ុណ្ណោះដែលធ្វើឱ្យវាអាចកំណត់ទីតាំងជាប្រព័ន្ធនៃ ascidians: រចនាសម្ព័ន្ធរបស់ larvae បង្ហាញពីប្រភពដើមរបស់ពួកគេពី chordates ដែលដឹកនាំរបៀបរស់នៅដោយឥតគិតថ្លៃ។ ក្នុងអំឡុងពេលនៃដំណើរការ metamorphosis, ascidians ប្តូរទៅជារបៀបរស់នៅស្ងប់ស្ងាត់ហើយដូច្នេះអង្គការរបស់ពួកគេត្រូវបានធ្វើឱ្យសាមញ្ញ។
ទម្រង់ដង្កូវរបស់សត្វអំពិលអំពែក គឺជាកូនខ្លា ដែលត្រូវបានកំណត់លក្ខណៈដោយស្នាមប្រេះ បន្ទាត់ក្រោយ បេះដូងបន្ទប់ពីរ និងរង្វង់ឈាមរត់មួយ។ ក្នុងអំឡុងពេលនៃដំណើរការ metamorphosis ដែលកើតឡើងក្រោមឥទិ្ធពលនៃអរម៉ូនទីរ៉ូអ៊ីត, កន្ទុយបានដោះស្រាយ, អវយវៈលេចឡើង, បន្ទាត់ក្រោយបាត់, សួតនិងរង្វង់ទីពីរនៃឈាមរត់។ គួរឱ្យកត់សម្គាល់គឺភាពស្រដៀងគ្នានៃរចនាសម្ព័ន្ធមួយចំនួននៃ tadpoles និងត្រី (បន្ទាត់ក្រោយ, រចនាសម្ព័ន្ធនៃបេះដូងនិងប្រព័ន្ធឈាមរត់, gill slits) ។
ការអភិវឌ្ឍនៃសត្វល្អិតក៏អាចបម្រើជាឧទាហរណ៍នៃការបំប្លែងសារជាតិ។ ដង្កូវមេអំបៅ ឬដង្កូវនាគមានភាពខុសប្លែកគ្នាយ៉ាងខ្លាំងនៅក្នុងរចនាសម្ព័ន របៀបរស់នៅ និងជម្រកពីសត្វពេញវ័យ ហើយស្រដៀងនឹងបុព្វបុរសរបស់ពួកវា។
រយៈពេល postembryonic នៃការអភិវឌ្ឍន៍មានរយៈពេលខុសគ្នា។ ឧទាហរណ៍ សត្វរុយរស់នៅ 2-3 ឆ្នាំក្នុងស្ថានភាពដង្កូវ ហើយពី 2-3 ម៉ោងទៅ 2-3 ថ្ងៃក្នុងសភាពចាស់ទុំ អាស្រ័យលើប្រភេទសត្វ។ ក្នុងករណីភាគច្រើនរយៈពេល postembryonic គឺវែងជាង។ នៅក្នុងមនុស្ស វារួមបញ្ចូលដំណាក់កាលពេញវ័យ ដំណាក់កាលពេញវ័យ និងដំណាក់កាលចាស់។
នៅក្នុងថនិកសត្វ និងមនុស្ស មានការពឹងផ្អែកលើអាយុសង្ឃឹមរស់លើរយៈពេលនៃភាពពេញវ័យ និងការមានផ្ទៃពោះ។ អាយុសង្ឃឹមរស់ជាធម្មតាលើសពី
រយៈពេលមុនពេលបន្តពូជនៃ ontogenesis 5-8 ដង។
ការអភិវឌ្ឍន៍ Postembryonic ត្រូវបានអមដោយការលូតលាស់។ ភាពខុសគ្នាមួយត្រូវបានបង្កើតឡើងរវាងការលូតលាស់មិនកំណត់ ដែលបន្តពេញមួយជីវិត និងការលូតលាស់ច្បាស់លាស់ ត្រូវបានកំណត់ត្រឹមរយៈពេលជាក់លាក់មួយ។ ការលូតលាស់មិនកំណត់ត្រូវបានសង្កេតឃើញនៅក្នុងទម្រង់ឈើនៃរុក្ខជាតិ សត្វមូស សត្វឆ្អឹងខ្នង ត្រី និងកណ្តុរ។
នៅក្នុងសត្វជាច្រើន ការលូតលាស់ឈប់ភ្លាមៗបន្ទាប់ពីឈានដល់ភាពពេញវ័យផ្លូវភេទ។ នៅក្នុងមនុស្ស ការលូតលាស់បញ្ចប់នៅអាយុ 20-25 ឆ្នាំ។
2.3 គំរូទូទៅនៃការអភិវឌ្ឍន៍។ ច្បាប់ជីវហ្សែន
សារពាង្គកាយពហុកោសិកាទាំងអស់វិវត្តន៍ចេញពីស៊ុតបង្កកំណើត។ ការអភិវឌ្ឍនៃអំប្រ៊ីយ៉ុងនៅក្នុងសត្វដែលជាកម្មសិទ្ធិរបស់ប្រភេទដូចគ្នាគឺភាគច្រើនស្រដៀងគ្នា។ នៅក្នុង chordates ទាំងអស់នៅក្នុងរយៈពេលអំប្រ៊ីយ៉ុង គ្រោងឆ្អឹងអ័ក្សត្រូវបានបង្កើតឡើង បំពង់សរសៃប្រសាទលេចឡើង ហើយស្នាមប្រេះត្រូវបានបង្កើតឡើងនៅផ្នែកខាងមុខនៃ pharynx ។ ផែនការរចនាសម្ព័ន្ធនៃ chordates ក៏ដូចគ្នាដែរ។ នៅដំណាក់កាលដំបូងនៃការអភិវឌ្ឍន៍ អំប្រ៊ីយ៉ុងឆ្អឹងខ្នងគឺស្រដៀងគ្នាខ្លាំងណាស់ (រូបភាពទី 3) ។ ការពិតទាំងនេះបញ្ជាក់ពីសុពលភាពនៃច្បាប់នៃភាពស្រដៀងគ្នានៃអំប្រ៊ីយ៉ុងដែលបង្កើតឡើងដោយ K. Baer៖ "ការតាំងបង្ហាញអំប្រ៊ីយ៉ុង តាំងពីដំណាក់កាលដំបូងបំផុត ភាពស្រដៀងគ្នាទូទៅមួយចំនួននៅក្នុងប្រភេទ។" ភាពស្រដៀងគ្នានៃអំប្រ៊ីយ៉ុងនៃក្រុមប្រព័ន្ធផ្សេងៗគ្នាបង្ហាញពីប្រភពដើមទូទៅរបស់ពួកគេ។ ក្រោយមក រចនាសម្ព័ន្ធនៃអំប្រ៊ីយ៉ុងបង្ហាញពីលក្ខណៈនៃថ្នាក់ ហ្សែន ប្រភេទសត្វ និងចុងក្រោយ លក្ខណៈលក្ខណៈនៃបុគ្គលដែលបានផ្តល់ឱ្យ។ ភាពខុសគ្នានៃលក្ខណៈនៃអំប្រ៊ីយ៉ុងក្នុងអំឡុងពេលនៃការអភិវឌ្ឍន៍ត្រូវបានគេហៅថា ភាពខុសគ្នានៃអំប្រ៊ីយ៉ុង និងឆ្លុះបញ្ចាំងពីការវិវត្តនៃក្រុមសត្វដែលមានប្រព័ន្ធជាក់លាក់មួយ ប្រវត្តិនៃការអភិវឌ្ឍន៍នៃប្រភេទសត្វដែលបានផ្តល់ឱ្យ។

អង្ករ។ 3. ភាពស្រដៀងគ្នានៃពូជនៅក្នុងសត្វឆ្អឹងខ្នង៖ 1 monotremes (echidna), 2 marsupials (kangaroos), 3 artiodactyls (សត្វក្តាន់), 4 carnivores (ឆ្មា), 5 primates (ស្វា), 6 - មនុស្ស
ភាពស្រដៀងគ្នាដ៏អស្ចារ្យរវាងអំប្រ៊ីយ៉ុងនៅដំណាក់កាលដំបូងនៃការអភិវឌ្ឍន៍និង
បាតុភូតនៃភាពខុសគ្នានៅដំណាក់កាលក្រោយមានការពន្យល់ផ្ទាល់ខ្លួន។
រាងកាយទទួលរងនូវភាពប្រែប្រួលពេញមួយការអភិវឌ្ឍន៍។
ដំណើរការផ្លាស់ប្តូរប៉ះពាល់ដល់ហ្សែនដែលកំណត់លក្ខណៈរចនាសម្ព័ន្ធ និងការរំលាយអាហាររបស់អំប្រ៊ីយ៉ុងដែលក្មេងជាងគេ។ ប៉ុន្តែរចនាសម្ព័ន្ធដែលកើតឡើងនៅក្នុងពួកគេ (លក្ខណៈបុរាណលក្ខណៈនៃបុព្វបុរសឆ្ងាយ) ដើរតួយ៉ាងសំខាន់ក្នុងដំណើរការនៃការអភិវឌ្ឍន៍បន្ថែមទៀត។ ដូចដែលបានបញ្ជាក់រួចមក primordium notochord ជំរុញឱ្យមានការបង្កើតបំពង់សរសៃប្រសាទ ហើយការបាត់បង់របស់វានាំទៅដល់ការបញ្ចប់នៃការអភិវឌ្ឍន៍។ ដូច្នេះការផ្លាស់ប្តូរនៅក្នុងដំណាក់កាលដំបូងជាធម្មតានាំទៅរកការអភិវឌ្ឍតិចតួច និងការស្លាប់របស់បុគ្គល។ ផ្ទុយទៅវិញ ការផ្លាស់ប្តូរនៅដំណាក់កាលក្រោយៗ ដែលប៉ះពាល់ដល់លក្ខណៈមិនសូវសំខាន់ អាចមានប្រយោជន៍សម្រាប់សារពាង្គកាយ ហើយក្នុងករណីបែបនេះត្រូវបានជ្រើសរើសដោយការជ្រើសរើសធម្មជាតិ។
រូបរាងនៅក្នុងរយៈពេលនៃអំប្រ៊ីយ៉ុងនៃការអភិវឌ្ឍនៃសត្វសម័យទំនើបនៃលក្ខណៈលក្ខណៈនៃបុព្វបុរសឆ្ងាយរបស់ពួកគេឆ្លុះបញ្ចាំងពីការផ្លាស់ប្តូរការវិវត្តនៅក្នុងរចនាសម្ព័ន្ធនៃសរីរាង្គ។
នៅក្នុងការអភិវឌ្ឍន៍របស់វា សារពាង្គកាយឆ្លងកាត់ដំណាក់កាលកោសិកាតែមួយ (ដំណាក់កាលហ្សីហ្គោត) ដែលអាចចាត់ទុកថាជាការផ្ទួននៃដំណាក់កាល phylogenetic នៃ amoeba បុព្វកាល។ នៅក្នុងសត្វឆ្អឹងកងទាំងអស់ រួមទាំងអ្នកតំណាងខ្ពស់បំផុតរបស់ពួកគេ ស្នាមរន្ធមួយត្រូវបានបង្កើតឡើង ដែលបន្ទាប់មកត្រូវបានជំនួសដោយឆ្អឹងខ្នង ហើយនៅក្នុងបុព្វបុរសរបស់ពួកគេ ដោយវិនិច្ឆ័យដោយ lancelet នោះ notochord នៅតែមានសម្រាប់ជីវិត។ ក្នុងអំឡុងពេលនៃការបង្កើតអំប្រ៊ីយ៉ុងនៃបក្សី និងថនិកសត្វ រួមទាំងមនុស្ស ស្នាមប្រេះ និង septa ដែលត្រូវគ្នានឹងលេចឡើងនៅក្នុង pharynx ។ ការពិតនៃការបង្កើតផ្នែកខ្លះនៃប្រដាប់ប្រមាត់នៅក្នុងអំប្រ៊ីយ៉ុងនៃសត្វឆ្អឹងកងនៅលើដីត្រូវបានពន្យល់ដោយប្រភពដើមរបស់វាពីបុព្វបុរសដូចត្រីដែលដកដង្ហើមជាមួយ gills ។ រចនាសម្ព័ននៃបេះដូងនៃអំប្រ៊ីយ៉ុងរបស់មនុស្សនៅក្នុងដំណាក់កាលដំបូងនៃការបង្កើតប្រហាក់ប្រហែលនឹងរចនាសម្ព័ន្ធនៃសរីរាង្គនេះនៅក្នុងត្រី: វាមាន atrium មួយ និង ventricle មួយ។ ត្រីបាឡែនដែលគ្មានធ្មេញបង្កើតធ្មេញក្នុងអំឡុងពេលអំប្រ៊ីយ៉ុង។ ធ្មេញទាំងនេះមិនផ្ទុះទេ វាត្រូវបានបំផ្លាញ និងរលាយ។
ឧទាហរណ៍ដែលបានផ្ដល់ឱ្យនៅទីនេះ និងជាច្រើនផ្សេងទៀតបង្ហាញពីទំនាក់ទំនងយ៉ាងស៊ីជម្រៅរវាងការអភិវឌ្ឍន៍បុគ្គលនៃសារពាង្គកាយ និងការអភិវឌ្ឍន៍ប្រវត្តិសាស្ត្ររបស់វា។ ការតភ្ជាប់នេះត្រូវបានបញ្ជាក់នៅក្នុងច្បាប់ជីវហ្សែនដែលបង្កើតដោយ F. Müller និង E. Haeckel ក្នុងសតវត្សទី 19: ontogenesis (ការអភិវឌ្ឍន៍បុគ្គល) របស់បុគ្គលម្នាក់ៗគឺជាពាក្យដដែលៗខ្លីៗ និងឆាប់រហ័សនៃ phylogeny (ការអភិវឌ្ឍន៍ប្រវត្តិសាស្ត្រ) នៃប្រភេទសត្វដែលបុគ្គលនេះជាកម្មសិទ្ធិ។ .
សេចក្តីសន្និដ្ឋាន
ការបញ្ចប់ការងារលើអរូបី យើងអាចសន្និដ្ឋានបានថា សមត្ថភាពក្នុងការបន្តពូជ ឬបន្តពូជដោយខ្លួនឯង គឺជាលក្ខណៈសំខាន់បំផុតមួយនៃធម្មជាតិសរីរាង្គ។ ការបន្តពូជគឺជាទ្រព្យសម្បត្តិដែលមាននៅក្នុងគ្រប់ភាវៈរស់ទាំងអស់ដោយគ្មានករណីលើកលែង ចាប់ពីបាក់តេរីរហូតដល់ថនិកសត្វ។
អត្ថិភាពនៃប្រភេទសត្វ និងរុក្ខជាតិណាមួយ បាក់តេរី និងផ្សិត ការបន្តរវាងបុគ្គលមេ និងកូនចៅរបស់វាត្រូវបានរក្សាបានតែតាមរយៈការបន្តពូជប៉ុណ្ណោះ។ ទាក់ទងយ៉ាងជិតស្និទ្ធទៅនឹងការបន្តពូជដោយខ្លួនឯងគឺជាទ្រព្យសម្បត្តិមួយផ្សេងទៀតនៃភាវៈរស់។ការអភិវឌ្ឍសារពាង្គកាយ។ វា។វាក៏មាននៅក្នុងជីវិតទាំងអស់នៅលើផែនដីផងដែរ៖ សារពាង្គកាយកោសិកាតែមួយតូចបំផុត និងរុក្ខជាតិ និងសត្វពហុកោសិកា។
គន្ថនិទ្ទេស
Bogen G. ជីវវិទ្យាទំនើប។ - អិមៈ Mir ឆ្នាំ 1970 ។
Green N., Stout W., Taylor D. Biology: ក្នុង 3 ភាគ T. 3: trans ។ ពីភាសាអង់គ្លេស / ed ។ R. Soper ។ - M. : Mir, ឆ្នាំ 1990 ។
Mamontov S.G. ជីវវិទ្យា។ លំនាំទូទៅ។ អិមៈ Bustard, 2002 ។
ពីសត្វទៅមនុស្ស។ M.: Nauka, 1971 ។
Slyusarev A.A. ជីវវិទ្យាជាមួយហ្សែនទូទៅ។ - អិមៈ វេជ្ជសាស្ត្រ ឆ្នាំ ១៩៧៨។
មេរៀនទី 3 Ontogenesis
1. Gametogenesis
2. រយៈពេលអំប្រ៊ីយ៉ុង
3. រយៈពេល Postembryonic
Ontogenesis- ការវិវឌ្ឍន៍បុគ្គលនៃសារពាង្គកាយពី zygote ដល់ការស្លាប់ដោយជីវសាស្រ្ត។ ការបន្តពូជ- រយៈពេលមុនពេល ontogenesis រួមមាន gametogenesis និងការបង្កកំណើត។ រូបវិទ្យា- ការវិវត្តន៍នៃប្រភេទសត្វ។
មនុស្សត្រូវបានកំណត់លក្ខណៈដោយការបន្តពូជផ្លូវភេទ វាត្រូវបានកំណត់លក្ខណៈដោយ: វត្តមានរបស់ gonads, gametes, ដំណើរការនៃការបង្កកំណើត និង dimorphism ផ្លូវភេទ។
Gametogenesis -ដំណើរការនៃការបង្កើតកោសិកាមេរោគ ovogenesis -ភាពចាស់ទុំ oocyte, ការបង្កើតមេជីវិតឈ្មោល។- មេជីវិតឈ្មោល។ Gametogenesis កើតឡើងនៅក្នុង gonads នៃរាងកាយ។ Gametes ត្រូវបានបង្កើតឡើងពីកោសិកា epithelial អំប្រ៊ីយ៉ុងដែលត្រូវបានបង្កើតឡើងក្នុងអំឡុងពេលនៃអំប្រ៊ីយ៉ុងនៃការអភិវឌ្ឍនៃសារពាង្គកាយ។
នៅក្នុងដំណើរការនៃការបង្កើត កោសិកាមេរោគឆ្លងកាត់បីដំណាក់កាល៖
1. រយៈពេលបន្តពូជ (កោសិកានៃ epithelium embryonic បែងចែកដោយ mitosis);
2. រយៈពេលលូតលាស់;
3. រយៈពេលនៃភាពចាស់ទុំកោសិកាបែងចែកដោយ meiosis ដែលជាលទ្ធផលនៅក្នុងការបង្កើត gametes (រូបភាពទី 5) ។
អង្ករ។ 5. ការបែងចែកកោសិកាមេរោគ
មេជីវិតឈ្មោលពេញវ័យនៅក្នុងបំពង់ seminiferous នៃពងស្វាស។ ស្ថិតនៅចន្លោះបំពង់ seminiferous ជាលិកា interstitialផលិតអរម៉ូនភេទបុរស តេស្តូស្តេរ៉ូន. អរម៉ូនភេទបុរស - អង់ដ្រូសែនគ្រប់គ្រងមុខងារបន្តពូជ gametogenesis និងការបង្កើតលក្ខណៈផ្លូវភេទបន្ទាប់បន្សំ។ មេជីវិតឈ្មោលពេញវ័យក្នុងរយៈពេល 70 ថ្ងៃ។ បំពង់ seminiferous មាន gametes នៅដំណាក់កាលផ្សេងគ្នានៃភាពចាស់ទុំ។ ក្នុង 5 មីលីលីត្រ។ សារធាតុរាវ seminal របស់មនុស្សមានមេជីវិតឈ្មោល 12 លាន។ ពួកគេត្រូវបានចោទប្រកាន់អវិជ្ជមាន ការចោទប្រកាន់រារាំងពួកគេមិនឱ្យនៅជាប់គ្នា។ មួយចំនួនធំនៃ gametes បុរសគឺសមស្របតាមជីវសាស្រ្ត ការថយចុះចំនួនរបស់ពួកគេ 40% ធ្វើឱ្យដំណើរការនៃការបង្កកំណើតមិនអាចទៅរួចនោះទេ។ Spermatozoa គឺជាកោសិកាតូចៗដែលមានចលនាដែលមានក្បាល ក និងកន្ទុយ។ នៅលើក្បាលគឺ acrosome, lysosome ដែលត្រូវបានកែប្រែ វាមានអង់ស៊ីមដែលរំលាយភ្នាសនៃស៊ុតអំឡុងពេលបង្កកំណើត។ អាយុកាលជាមធ្យមគឺពី 6 ទៅ 72 ម៉ោង។
Oogenesis ចាប់ផ្តើមនៅក្នុងរយៈពេល embryonic នៃការអភិវឌ្ឍនៃរាងកាយរបស់ស្ត្រី។ នៅដំណាក់កាល prophase នៃការបែងចែកដំបូងនៃ meiosis នៅពេលដែលការភ្ជាប់គ្នានិងការឆ្លងកាត់កើតឡើងការផ្អាកបណ្តោះអាសន្នកើតឡើង។ ការវិវឌ្ឍបន្ថែមទៀតនៃស៊ុតចាប់ផ្តើមឡើងវិញក្នុងអំឡុងពេលពេញវ័យរបស់ក្មេងស្រី។ ក្រោមឥទិ្ធពលនៃអរម៉ូនភេទស្ត្រី - អរម៉ូនអ៊ឹស្ត្រូសែនដែលជាឫសគល់កោសិកាដែលការពារនិងចិញ្ចឹមកោសិកាត្រូវបានបង្កើតឡើងនៅជុំវិញ oocyte លំដាប់ទីមួយ។
នៅពេលដែល follicle ពេញវ័យ វាផ្លាស់ទីទៅគែមនៃ ovary ហើយបន្ទាប់មកបានផ្ទុះឡើង ដំណាក់កាលនៃការបង្កើតស៊ុតនេះត្រូវបានគេហៅថា ovary ឬ follicular វាមានរយៈពេល 12 ថ្ងៃ។ ការដាច់នៃ follicle និងការបញ្ចេញពងចេញពីអូវែរចូលទៅក្នុងប្រហោងពោះត្រូវបានគេហៅថា ovulation ។
បន្ទាប់ពីការបញ្ចេញពងអូវុលការបង្កកំណើតអាចធ្វើទៅបាន។ gamete ស្ត្រីត្រូវបានចាប់យកដោយ fimbriae នៃបំពង់ fallopian ។ ដោយមានជំនួយពី epithelium ciliated វាផ្លាស់ទីទៅក្នុងស្បូន; ដំណាក់កាលស្បូនមានរយៈពេល 12-14 ថ្ងៃ។ នៅពេលនេះ ក្រោមឥទ្ធិពលនៃអរម៉ូនភេទស្ត្រី ក្រពេញស្បូនត្រូវបានបន្ធូរ។ ប្រសិនបើការបង្កកំណើតមិនកើតឡើងទេ epithelium និងស៊ុតត្រូវបានបញ្ចេញចេញពីរាងកាយដែលជាលទ្ធផលនៃការកន្ត្រាក់នៃសាច់ដុំរលោងនៃជញ្ជាំងស្បូន។ ដំណើរការនេះត្រូវបានគេហៅថា mensis និងមានរយៈពេល 3-4 ថ្ងៃ។ ដូច្នេះពេលវេលាពីការចាប់ផ្តើមនៃការបង្កើត follicle ដល់ការចាប់ផ្តើមនៃការមករដូវគឺ 20-30 ថ្ងៃហើយត្រូវបានគេហៅថាវដ្តអូវែរ - មករដូវ។ (រូបភាពទី 6) ។

អង្ករ។ 6. ការបញ្ចេញពងអូវុល និងការផ្សាំស៊ុតចូលទៅក្នុងស្បូន
វារួមបញ្ចូល ដំណាក់កាលអូវែ ស្បូន និងមករដូវ។ក្នុងអំឡុងពេលវដ្តរដូវ អូវែរ ប្រព័ន្ធបន្តពូជទាំងមូលរបស់ស្ត្រីត្រូវបានស្ថាបនាឡើងវិញ សមាសភាពគុណភាព និងបរិមាណនៃអរម៉ូនផ្លាស់ប្តូរ ការសម្តែង កម្លាំង ភាពបត់បែន និងការផ្លាស់ប្តូរការស៊ូទ្រាំ។ ការផ្លាស់ប្តូរសរីរវិទ្យានៅក្នុងរាងកាយស្ត្រីត្រូវបានគេយកមកពិចារណានៅពេលរៀបចំដំណើរការបណ្តុះបណ្តាល។
ការបង្កកំណើត- នេះគឺជាការលាយបញ្ចូលគ្នានៃស៊ុត និងមេជីវិតឈ្មោល។ មានបីដំណាក់កាលនៃការបង្កកំណើត៖
1) ប្រតិកម្ម acrosomal (20 វិនាទី) - ការរំលាយសំបកស៊ុតដោយអង់ស៊ីម acrosome និងការជ្រៀតចូលនៃមេជីវិតឈ្មោលចូលទៅក្នុងស៊ុត។ វាត្រូវបានគេជឿថាមេជីវិតឈ្មោលដែលបង្កើតសក្តានុពលជ្រៀតចូលដ៏អស្ចារ្យបំផុត (ភាពរំភើប) នៅលើភ្នាសស៊ុតជ្រាបចូលទៅក្នុងស៊ុត;
2) ការផ្អាកបណ្តោះអាសន្ន - មិនមានការផ្លាស់ប្តូរដែលអាចមើលឃើញនៅក្នុងស៊ុតទេ ការរំលាយអាហារកើនឡើង។
3) ការលាយបញ្ចូលគ្នានៃស្នូលនៃស៊ុត និងមេជីវិតឈ្មោល ការស្ដារឡើងវិញនូវសំណុំក្រូម៉ូសូម diploid ។
ការបង្កកំណើតកើតឡើងនៅផ្នែកខាងលើទីបីនៃបំពង់ fallopian ។ ស៊ុតបង្កកំណើតត្រូវបានគេហៅថា ហ្សីហ្គោត. ជាមួយនឹងការបង្កើត zygote មួយ ontogenesis របស់មនុស្សចាប់ផ្តើម។
2. Ontogenesisមនុស្សមានពីរដំណាក់កាល៖ អំប្រ៊ីយ៉ុងនិង postembryonic ។
ដំណាក់កាលនៃការអភិវឌ្ឍន៍អំប្រ៊ីយ៉ុងបន្តពីហ្សីហ្គោតរហូតដល់កំណើត។ រួមបញ្ចូលទាំងដំណាក់កាល: zygote, blastula, gastrula, histogenesis, organogenesis ។
ហ្សីហ្គោត- បែងចែកម្តងហើយម្តងទៀតដោយ mitosis ដែលបណ្តាលឱ្យមានការបង្កើតអំប្រ៊ីយ៉ុងស្រទាប់តែមួយពហុកោសិកា - blastula. blastula មិនកើនឡើងក្នុងទំហំទេ ព្រោះនៅពេលនេះវាស្ថិតនៅក្នុងបំពង់ fallopian ហើយគួរតែផ្លាស់ទីដោយសេរីចូលទៅក្នុងស្បូន។ នៅថ្ងៃទីប្រាំមួយ វាចូលទៅក្នុងស្បូន ហើយភ្ជាប់ទៅនឹងជញ្ជាំងរបស់វា ធ្លាក់ចូលទៅក្នុង epithelium ដែលត្រូវបានបន្ធូរ។ ដំណើរការនេះត្រូវបានគេហៅថា ការផ្សាំ. អំប្រ៊ីយ៉ុងបន្តអភិវឌ្ឍ ហើយក្លាយជាពីរ បន្ទាប់មកមានបីស្រទាប់។ នៅដំណាក់កាលនៃការអភិវឌ្ឍន៍នេះត្រូវបានគេហៅថា gastrula. ជាលទ្ធផលស្រទាប់មេរោគចំនួនបីត្រូវបានបង្កើតឡើង: ectoderm, endoderm, mesoderm ។ ជាលិកា និងសរីរាង្គត្រូវបានបង្កើតឡើងពីប្រព័ន្ធអំប្រ៊ីយ៉ុង។
ដូចនេះ៖ blastula- អំប្រ៊ីយ៉ុងស្រទាប់តែមួយ gastrula- អំប្រ៊ីយ៉ុងពីរនិងបីស្រទាប់។ Histogenesis- ការដាក់ជាលិកាពីស្រទាប់មេរោគ។ សរីរាង្គ- ការដាក់សរីរាង្គ។ នៅអាយុ 8 សប្តាហ៍ម៉ាស់អំប្រ៊ីយ៉ុងរបស់មនុស្សគឺ 4 ក្រាមទំហំ 5 ម។ មកដល់ពេលនេះ គាត់បានឆ្លងកាត់ដំណើរការបង្កើត និងទទួលបានវណ្ឌវង្កនៃរាងកាយមនុស្ស។ បន្ទាប់ពីមានផ្ទៃពោះប្រាំបីសប្តាហ៍ អំប្រ៊ីយ៉ុងរបស់មនុស្សត្រូវបានគេហៅថាទារក។
ក្នុងអំឡុងពេលនៃការបង្កើតអំប្រ៊ីយ៉ុងរបស់មនុស្ស។ អាជ្ញាធរបណ្តោះអាសន្ន,ដែលបន្ទាប់ពីកំណើតបាត់បង់មុខងាររបស់ពួកគេ។ អាជ្ញាធរបណ្ដោះអាសន្នរួមមាន៖ មេរោគសំបក, chorion, amnion និងសុក, ទងផ្ចិត. សុកធានានូវទំនាក់ទំនងជិតស្និទ្ធរវាង capillaries របស់កុមារនិងម្តាយ។ សារពាង្គកាយដែលកំពុងអភិវឌ្ឍទទួលបានសារធាតុចិញ្ចឹម អុកស៊ីហ្សែន អង្គបដិប្រាណតាមរយៈសុក និងដកផលិតផលមេតាបូលីសចេញ។ សុកអនុវត្តមុខងាររារាំង ការពារអំប្រ៊ីយ៉ុងពីអតិសុខុមប្រាណ ថ្នាំ និងជាតិពុលមួយចំនួន (រូបភាពទី 7) ។

អង្ករ។ 7. សុក
1 - ស្បូន; 2 - អាម៉ានីន; 3 - អំប្រ៊ីយ៉ុង; 4 - សារធាតុរាវ amniotic; 5 - សុក; 6 - capillaries នៃសុក; 7 - សរសៃឈាមវ៉ែន; 8 - សរសៃឈាម
យន្តការតូចៗនៃការបង្កើតសរីរាង្គ និងជាលិការបស់កុមារអាចត្រូវបានរំខាននៅពេលដែលប៉ះពាល់នឹងជាតិពុល សារធាតុគ្រឿងញៀន ជាតិអាល់កុល ជាតិនីកូទីនដែលមាននៅក្នុងឈាមរបស់ម្តាយ។ ភាពមិនប្រក្រតីនៃការអភិវឌ្ឍន៍អំប្រ៊ីយ៉ុងរបស់មនុស្សត្រូវបានគេហៅថា ខូចទ្រង់ទ្រាយពីកំណើត និងការខូចទ្រង់ទ្រាយ។ ទាំងនេះរួមបញ្ចូលៈ "ក្រអូមមាត់ឆែប" (អវត្ដមាននៃក្រអូមមាត់ខាងលើ), "បបូរមាត់ឆែប" (មិនលាយបញ្ចូលគ្នានៃឆ្អឹង maxillary), polydactyly (ម្រាមដៃបន្ថែម), syndactyly (ម្រាមដៃបញ្ចូលគ្នា) ។ កត្តាបរិស្ថានមិនអំណោយផលដែលបណ្តាលឱ្យខូចទ្រង់ទ្រាយពីកំណើតត្រូវបានគេហៅថា teratogenic(បណ្តាលឱ្យខូចទ្រង់ទ្រាយ) (រូបភាពទី 8) ។

អង្ករ។ 8. កត្តាបរិស្ថានដែលជះឥទ្ធិពលដល់ការអភិវឌ្ឍន៍រាងកាយរបស់មនុស្ស
រយៈពេលអំប្រ៊ីយ៉ុងនៃការអភិវឌ្ឍន៍មនុស្សមានរយៈពេល 36 សប្តាហ៍។
3. រយៈពេលបន្ទាប់ពីកំណើតនិងមុនពេលស្លាប់ជីវសាស្រ្តត្រូវបានគេហៅថា postembryonic ។វារួមបញ្ចូល អនីតិជន, ភាពពេញវ័យរយៈពេលនៃការអភិវឌ្ឍន៍និងភាពចាស់។
អនីតិជន - រយៈពេលមុនពេលពេញវ័យ។ ភាពពេញវ័យ- រយៈពេលនៃភាពពេញវ័យ។
នៅក្នុងការអនុវត្តកីឡា សរីរវិទ្យា គរុកោសល្យ ចិត្តវិទ្យា ការកំណត់អាយុខាងក្រោមត្រូវបានប្រើប្រាស់ postembryonic ontogenesis របស់មនុស្ស។
ក្នុងរយៈពេលពី 1 ទៅ 10 ថ្ងៃកុមារត្រូវបានហៅ ទារកទើបនឹងកើត;
ពី 10 ថ្ងៃទៅ 1 ឆ្នាំ - ទ្រូង;
ពី 1 ឆ្នាំទៅបី - រយៈពេល កុមារភាពដំបូង;
ពី 4 ទៅ 7 ឆ្នាំ - កុមារភាពដំបូង;
ចាប់ពីអាយុ 8 ឆ្នាំដល់ 12 ឆ្នាំសម្រាប់ក្មេងស្រី និងរហូតដល់ 13 ឆ្នាំសម្រាប់ក្មេងប្រុស កុមារភាពទីពីរ;
ពី 12 ឆ្នាំដល់ 15 ឆ្នាំសម្រាប់ក្មេងស្រីនិងពី 13 ទៅ 16 ឆ្នាំសម្រាប់ក្មេងប្រុស - ក្មេងជំទង់;
17-21 (ក្មេងប្រុស), 16-20 ឆ្នាំ (ក្មេងស្រី) - វ័យជំទង់;
20-55 សម្រាប់ស្ត្រី និង 21-60 សម្រាប់បុរសមករដូវ ភាពពេញវ័យ(ភាពពេញវ័យ);
ពី 55 និង 60 ទៅ 70 - មនុស្សចាស់;
ពី 70 ទៅ 90 បុរសចំណាស់;
ច្រើនជាង 90 រាប់រយឆ្នាំ.
នៅក្នុង ontogenesis របស់មនុស្សមាន រសើបនិងរិះគន់រយៈពេល។
រយៈពេលដ៏សំខាន់ត្រូវបានកំណត់លក្ខណៈដោយការកើនឡើងសកម្មភាពនៃហ្សែនបុគ្គល និងស្មុគស្មាញរបស់ពួកគេដែលគ្រប់គ្រងការវិវត្តនៃសញ្ញាណាមួយនៃរាងកាយ។ ក្នុងអំឡុងពេលទាំងនេះមានការរៀបចំឡើងវិញយ៉ាងសំខាន់នៃដំណើរការបទប្បញ្ញត្តិ ការលោតផ្លោះប្រកបដោយគុណភាព និងបរិមាណក្នុងការអភិវឌ្ឍន៍សរីរាង្គ និងប្រព័ន្ធមុខងារនីមួយៗ។ ក្នុងអំឡុងពេលទាំងនេះរាងកាយមានភាពរសើបបំផុតចំពោះឥទ្ធិពលនៃកត្តាបរិស្ថាន។ រយៈពេលសំខាន់នៃ ontogenesis ត្រូវបានចាត់ទុកថា implantation, placentation, ការបង្កើតសរីរាង្គអ័ក្ស, បំពង់សរសៃប្រសាទ និងពោះវៀន, notochords, ការបង្កើតបេះដូង និងផ្សេងទៀត។
ប្រសិនបើស្ត្រីឆ្លងជំងឺស្អូចនៅចន្លោះសប្តាហ៍ទី 3 និងទី 9 នៃការមានផ្ទៃពោះ វាមានហានិភ័យដែលទារកនឹងវិវត្តទៅជាជំងឺបេះដូង ជំងឺភ្នែកឡើងបាយ និងថ្លង់។ នៅពេលផ្សេងទៀត ជំងឺស្អូចមិនបណ្តាលឱ្យខូចទ្រង់ទ្រាយគភ៌ទេ។
រយៈពេលរសើប- ទាំងនេះគឺជារយៈពេលនៃការថយចុះការគ្រប់គ្រងហ្សែន និងការកើនឡើងនូវភាពប្រែប្រួលនៃលក្ខណៈបុគ្គលនៃរាងកាយចំពោះឥទ្ធិពលបរិស្ថាន រួមទាំងគរុកោសល្យ និងការបង្វឹក។ ដូច្នេះរយៈពេលដ៏រសើបសម្រាប់ការបង្ហាញពីសូចនាករផ្សេងៗនៃគុណភាពនៃល្បឿនកើតឡើងនៅអាយុ 11-14 ឆ្នាំហើយនៅអាយុ 15 ឆ្នាំកម្រិតអតិបរមារបស់វាត្រូវបានឈានដល់នៅពេលដែលសមិទ្ធិផលកីឡាខ្ពស់អាចធ្វើទៅបាន។ រូបភាពស្រដៀងគ្នានេះត្រូវបានគេសង្កេតឃើញនៅក្នុង ontogenesis សម្រាប់ការបង្ហាញពីគុណភាពនៃ dexterity និងភាពបត់បែន។
សម្រាប់គ្រូបង្វឹក និងគ្រូបង្រៀនដែលធ្វើការក្នុងវិស័យអប់រំកាយ និងកីឡា ចំណេះដឹងអំពីរយៈពេលដ៏រសើបគឺមានសារៈសំខាន់ជាក់ស្តែងណាស់ ចាប់តាំងពីអំឡុងពេលដ៏រសើបបំផុត ប្រសិទ្ធិភាពបណ្តុះបណ្តាល.
ពួកវាអាចបែងចែកបានអាស្រ័យលើចំនួនកោសិកាដែលជាប់ពាក់ព័ន្ធក្នុងដំណើរការនៃការបន្តពូជដោយភេទ៖ ការបន្តពូជដោយភេទដែលជំនាន់កូនស្រីកើតចេញពីកោសិកាតែមួយ៖ ការបែងចែកកោសិកាច្រើន ការបែងចែកកោសិកា schizogony sporulation sporulation budding នៅក្នុង unicellular yeast...
ចែករំលែកការងាររបស់អ្នកនៅលើបណ្តាញសង្គម
ប្រសិនបើការងារនេះមិនសមនឹងអ្នកទេ នៅផ្នែកខាងក្រោមនៃទំព័រមានបញ្ជីការងារស្រដៀងគ្នា។ អ្នកក៏អាចប្រើប៊ូតុងស្វែងរកផងដែរ។
ការបន្តពូជនិងការអភិវឌ្ឍនៃសារពាង្គកាយ
ផែនការ
- គំនិតនិងអត្ថន័យនៃការបន្តពូជ។
- ទម្រង់និងប្រភេទនៃការបន្តពូជ។
- វដ្តកោសិកា។ មីតូស៊ីស។ ម៉ីអូស៊ីស។
- រចនាសម្ព័ន្ធនៃ gametes ។ Gametogenesis។
1. គំនិត និងអត្ថន័យនៃការបន្តពូជ
លក្ខណៈសម្បត្តិមួយនៃវត្ថុមានជីវិតគឺភាពមិនច្បាស់លាស់, ទាំងនោះ។ នៅកម្រិតណាមួយនៃអង្គការ សារធាតុរស់នៅត្រូវបានតំណាងដោយអង្គភាពរចនាសម្ព័ន្ធបឋម។ បុគ្គលនីមួយៗនៃប្រភេទសត្វដែលបានផ្តល់ឱ្យគឺរមែងស្លាប់ ហើយអត្ថិភាពនៃប្រភេទសត្វត្រូវបានរក្សាការបន្តពូជ សារពាង្គកាយ។ ដូច្នេះភាពមិនច្បាស់លាស់នៃជីវិតសន្មតថាការបន្តពូជរបស់វាពោលគឺឧ។ ដំណើរការបន្តពូជ។
ការបន្តពូជ នេះគឺជាសមត្ថភាពរបស់សត្វមានជីវិតក្នុងការបន្តពូជប្រភេទរបស់វា។ នេះធានានូវភាពបន្តនិងភាពជាប់លាប់នៃជីវិត។ ការបន្តត្រូវបានកំណត់ដោយការពិតដែលថាក្នុងអំឡុងពេលដំណើរការនៃការបន្តពូជសម្ភារៈហ្សែនត្រូវបានផ្ទេរពីឪពុកម្តាយទៅកូនចៅដែលជាលទ្ធផលដែលលក្ខណៈរបស់ឪពុកម្តាយត្រូវបានបង្ហាញដល់កម្រិតមួយឬមួយផ្សេងទៀតនៅក្នុងសារពាង្គកាយកូនស្រី។
តម្លៃបន្តពូជ៖
- ការបង្កើន ឬរក្សាចំនួនប្រភេទសត្វ។ ដោយសារតែការបន្តពូជ មិនត្រឹមតែការបន្តពូជកើតឡើង (ពោលគឺការបន្តពូជនៃប្រភេទរបស់វាផ្ទាល់) ប៉ុន្តែក៏មានការកើនឡើងនៃចំនួនសារពាង្គកាយមានជីវិតផងដែរ។
- ការបន្តជីវិត។ សូមអរគុណដល់ការបន្តពូជ ក្រុមបុគ្គលច្រើន ឬតិច (ឧទាហរណ៍ ចំនួនប្រជាជន និងប្រភេទសត្វ) អាចមានរយៈពេលយូរណាស់ ចាប់តាំងពីការថយចុះនៃចំនួនរបស់ពួកគេដោយសារតែការស្លាប់ធម្មជាតិរបស់បុគ្គលត្រូវបានផ្តល់សំណងដោយការបន្តពូជជាបន្តបន្ទាប់នៃសារពាង្គកាយ និងការបន្តពូជ។ ការជំនួសអ្នកស្លាប់ដោយទារកទើបនឹងកើត។
- ការបន្តរវាងជំនាន់។ ក្នុងអំឡុងពេលដំណើរការបន្តពូជ ព័ត៌មានហ្សែនត្រូវបានបញ្ជូនពីបុគ្គលនៃជំនាន់មាតាបិតា ដែលធានាដល់ការបន្តពូជនៃលក្ខណៈរបស់ឪពុកម្តាយជាក់លាក់ និងប្រភេទសត្វទាំងមូលដែលសារពាង្គកាយជាកម្មសិទ្ធិ។
- លក្ខណៈសម្បត្តិនៃភាវៈមានជីវិតបែបនេះ គឺជាតំណពូជ និងភាពប្រែប្រួលត្រូវបានដឹង។
2. ទម្រង់និងប្រភេទនៃការបន្តពូជ
មានទម្រង់សំខាន់ពីរនៃការបន្តពូជ៖ ផ្លូវភេទ និងផ្លូវភេទ។
ការបន្តពូជផ្លូវភេទ
ក្នុងអំឡុងពេលបន្តពូជដោយភេទ បុគ្គលថ្មីលេចឡើងពីកោសិកា somatic មិនបន្តពូជដែលមិនពិសេសនៃរាងកាយ។ ដូច្នេះ ការបន្តពូជដោយភេទគឺពាក់ព័ន្ធនឹងបុគ្គលម្នាក់។ ការបន្តពូជតាមភេទធានាដល់ការបន្តពូជនៃបុគ្គលដូចគ្នាបេះបិទមួយចំនួនធំ ដែលផ្តល់អត្ថប្រយោជន៍សម្រាប់ប្រភេទសត្វដែលរស់នៅក្នុងស្ថានភាពថេរ។ ការបន្តពូជយ៉ាងឆាប់រហ័ស និងច្រើននៃកូនចៅដូចគ្នាទៅនឹងម្តាយ គឺជាអត្ថន័យជីវសាស្រ្តនៃការបន្តពូជដោយភេទដូចគ្នា។
នៅក្នុងធម្មជាតិ មានជម្រើសផ្សេងគ្នាសម្រាប់ការបន្តពូជដោយភេទដូចគ្នា៖ការបែងចែក, sporulation, ការបែងចែក, budding, ការបន្តពូជលូតលាស់, ការក្លូន។
ពួកវាអាចបែងចែកបានអាស្រ័យលើចំនួនកោសិកាដែលជាប់ពាក់ព័ន្ធក្នុងដំណើរការនៃការបន្តពូជតាមភេទ៖
- ការបន្តពូជដោយភេទដូចគ្នា ដែលជំនាន់កូនស្រីកើតចេញពីកោសិកាតែមួយ៖
- ការបែងចែកកោសិកា
- ការបែងចែកកោសិកាច្រើន (schizogony)
- sporulation (sporulation)
- ការរីកដុះដាលនៅក្នុងសារពាង្គកាយកោសិកា (ផ្សិត)
2) ការបន្តពូជដោយភេទដែលផ្អែកលើការបែងចែកក្រុមនៃកោសិកា៖
- បន្លែ
- ការបែកខ្ញែក
- ការចេញផ្កានៅក្នុងសារពាង្គកាយពហុកោសិកា (hydra)
មានការចាត់ថ្នាក់មួយទៀត យោងទៅតាមជម្រើសសម្រាប់ការបន្តពូជដោយភេទគឺ៖
- ការបន្តពូជលូតលាស់ (ឧទាហរណ៍ការបែងចែកតាមផ្នែកនៃរាងកាយម្តាយ)៖
ការបែងចែក
ការប្រេះស្រាំច្រើន (ជំងឺវិកលចរិក)
ពន្លក
ការបែងចែក
ការបន្តពូជរបស់រុក្ខជាតិដោយការកាប់, អំពូល, ស្លឹក, រមាស
- ការបន្តពូជដោយ spores, i.e. sporulation
សូមក្រឡេកមើលប្រភេទនៃការបន្តពូជដោយភេទដោយលម្អិតបន្ថែមទៀត។
ការបែងចែក។ ទម្រង់សាមញ្ញបំផុតនៃការបន្តពូជផ្លូវភេទ។ មានជម្រើសបែងចែកផ្សេងៗគ្នា៖
- ការបែងចែកដោយការបង្រួមសាមញ្ញជាមួយនឹងការបង្កើតសារពាង្គកាយកូនស្រីពីរពីសារពាង្គកាយមេមួយ។ លក្ខណៈនៃបាក់តេរីនិង cyanobacteria ។
- ការបែងចែកដោយការបែងចែក mitotic នៃស្នូល បន្តដោយការបំបែកនៃ cytoplasm ។ លក្ខណៈនៃសារពាង្គកាយឯកកោសិកា (ប្រូតូហ្សូអាជាច្រើន - អាមីបា អ៊ីហ្គលេណា បៃតង។
ក្នុងករណីទាំងពីរវាកើតឡើងគោលពីរ ការបែងចែក, i.e. ចូលទៅក្នុងកោសិកាពីរ។ ទោះយ៉ាងណាក៏ដោយ ជម្រើសមួយផ្សេងទៀតក៏អាចធ្វើទៅបានដែរ៖
- ការប្រេះស្រាំច្រើន (វិកលចរិក) ។ ទីមួយ ស្នូលបែងចែកម្តងហើយម្តងទៀត ហើយបន្ទាប់មក cytoplasm បែងចែកជាផ្នែកៗ។ ឧទាហរណ៍ ការបន្តពូជរបស់ Plasmodium falciparum (ភ្នាក់ងារបង្កជំងឺគ្រុនចាញ់) នៅក្នុង erythrocytes របស់មនុស្ស។ ក្នុងករណីនេះ plasmodia ឆ្លងកាត់ការបែងចែកនៃស្នូលជាច្រើនដង ក្រោយមក cytoplasm បែងចែក។ ជាលទ្ធផល 1 plasmodium ផ្តល់នូវសារពាង្គកាយកូនស្រី 12-24 ។
ការបង្កើតស្ព័រ (sporulation). វិធីសាស្រ្តរីករាលដាលយ៉ាងខ្លាំងនៃការបន្តពូជដោយភេទក្នុងចំនោមសារពាង្គកាយមានជីវិត ហើយត្រូវបានរកឃើញនៅក្នុងរុក្ខជាតិស្ទើរតែទាំងអស់ ផ្សិត និងប្រូតូហ្សូអាមួយចំនួន (ឧទាហរណ៍ប្រភេទ sporozoan) ក៏ដូចជាសារពាង្គកាយ prokaryotic (បាក់តេរីជាច្រើន សារាយខៀវបៃតង)។
ស្ព័រ នេះគឺជាកោសិកាដែលគ្របដណ្ដប់ដោយគម្របការពារដែលត្រូវបានអភិវឌ្ឍ - ភ្នាស spores ដែលអនុញ្ញាតឱ្យវាទប់ទល់នឹងឥទ្ធិពលនៃកត្តាបរិស្ថានដែលមិនអំណោយផលផ្សេងៗ។ដោយសារទំហំតូចរបស់វា ស្ព័រជាធម្មតាមានផ្ទុកសារធាតុចិញ្ចឹមតិចតួចប៉ុណ្ណោះ។. នៅក្នុងសារពាង្គកាយជាច្រើន វាគឺជាឯកតាសំខាន់នៃការបែកខ្ចាត់ខ្ចាយ ចាប់តាំងពីចំនួនដ៏ច្រើននៃ spores ពន្លឺត្រូវបានបង្កើតឡើងត្រូវបានដឹកជញ្ជូនដោយសេរីលើចម្ងាយដ៏សំខាន់ដោយចលនានៃម៉ាស់ខ្យល់ និងលំហូរទឹក។ជាញឹកញយ ស្ព័រត្រូវបានបង្កើតឡើងក្នុងបរិមាណច្រើន និងមានទម្ងន់ស្រាល ដែលធ្វើឲ្យពួកវាងាយស្រួលក្នុងការសាយភាយដោយខ្យល់ ក៏ដូចជាដោយសត្វ ភាគច្រើនជាសត្វល្អិត។
លើសពីនេះទៀតនៅក្នុងទម្រង់មួយចំនួន (ប្រូតូហ្សូអាផ្សិត) ស្ព័រអាចដើរតួជាដំណាក់កាលពិសេសនៃវដ្តជីវិតដែលអនុញ្ញាតឱ្យពួកគេ "រស់រានមានជីវិត" ផលប៉ះពាល់នៃកត្តាបរិស្ថានដែលមិនអំណោយផល។spores បាក់តេរី និយាយយ៉ាងតឹងរ៉ឹងមិនបម្រើសម្រាប់ការបន្តពូជទេប៉ុន្តែដើម្បីរស់រានមានជីវិតក្រោមលក្ខខណ្ឌមិនអំណោយផលព្រោះបាក់តេរីនីមួយៗបង្កើតបានតែ spores មួយ។ ស្ព័របាក់តេរីគឺស្ថិតក្នុងចំណោមធន់ទ្រាំបំផុត៖ ឧទាហរណ៍ ពួកវាអាចទប់ទល់នឹងការព្យាបាលជាមួយថ្នាំសំលាប់មេរោគដ៏ខ្លាំង និងការដាំឱ្យពុះក្នុងទឹក។
នៅក្នុងរុក្ខជាតិជាច្រើនដំណើរការនៃការបង្កើត spores(sporogenesis) ត្រូវបានអនុវត្តនៅក្នុងរចនាសម្ព័ន្ធដូចថង់ពិសេសដែលហៅថា sporangia ។ Spores អាចជាចលនាដោយសារតែវត្តមាននៃឧបករណ៍ flagellar (ក្នុងករណីនេះពួកគេត្រូវបានគេហៅថាសួនសត្វ), និងគ្មានចលនា បាត់បង់សមត្ថភាពក្នុងការធ្វើចលនាយ៉ាងសកម្ម។
Zoospores ត្រូវបានគេសង្កេតឃើញនៅក្នុងសារាយពណ៌បៃតងមួយចំនួន។ ក្នុងចំនោមសត្វពាហនៈ sporulation ត្រូវបានគេសង្កេតឃើញនៅក្នុង plasmodium គ្រុនចាញ់ និងក្រុមទាំងមូលនៃ sporozoans (សារពាង្គកាយឯកតា)។
ពន្លក។ វាត្រូវបានគេរកឃើញទាំងនៅក្នុងសារពាង្គកាយឯកតា ឧទាហរណ៍នៅក្នុងផ្សិត និងប្រភេទមួយចំនួននៃ ciliates និងនៅក្នុងសារពាង្គកាយពហុកោសិកា ឧទាហរណ៍នៅក្នុងតំណាងនៃប្រភេទ coelenterate (hydra) ក៏ដូចជានៅក្នុង tunicates (ថ្នាក់ ascidians) ។
ចំពោះសារពាង្គកាយឯកតា វិធីសាស្ត្រនេះមាននៅក្នុងការបង្កើតមើម (ការរីកដុះដាល) ជាមួយនឹងស្នូលនៅលើកោសិកាមេ ដែលបន្ទាប់មកបំបែក និងក្លាយជាសារពាង្គកាយឯករាជ្យ។
សម្រាប់សារពាង្គកាយពហុកោសិកា វិធីសាស្រ្តនេះមាននៅក្នុងការពិតដែលថាដំបូងមើមតូចមួយលេចឡើងនៅលើរាងកាយរបស់មាតាបុគ្គលដែលមានទំហំកើនឡើងបន្ទាប់មកបំណែកនៃរចនាសម្ព័ន្ធនិងសរីរាង្គទាំងអស់ដែលជាលក្ខណៈនៃសារពាង្គកាយមាតាលេចឡើង។ បន្ទាប់មកការបែកគ្នា (ពន្លក) នៃកូនស្រីបុគ្គលកើតឡើង។ បន្ទាប់ពីនេះ សារពាង្គកាយវ័យក្មេងដែលទើបនឹងបំបែកបានលូតលាស់ និងឈានដល់ទំហំនៃគំរូដើម។
អង្ករ។ ពន្លកនៅក្នុង coelenterates (hydra), 1 សារពាង្គកាយពេញវ័យ, 2 កូនស្រី budding សារពាង្គកាយ។
ទម្រង់មិនធម្មតានៃពន្លកត្រូវបានពិពណ៌នានៅក្នុងរុក្ខជាតិទឹកដម Bryophyllum - xerophyte ដែលជារឿយៗត្រូវបានដាំដុះជារុក្ខជាតិលម្អក្នុងផ្ទះ៖ រុក្ខជាតិតូចៗដែលបំពាក់ដោយឫសតូចៗ ដុះតាមគែមស្លឹករបស់វា “ពន្លក” ទាំងនេះនៅទីបំផុតបានរលំហើយចាប់ផ្តើមមានជារុក្ខជាតិឯករាជ្យ។
ការបែងចែក ការបែងចែកបុគ្គលទៅជាពីរ ឬច្រើនផ្នែក ដែលផ្នែកនីមួយៗវិវត្តទៅជាសារពាង្គកាយថ្មី។ ការបែកខ្ញែកកើតឡើងជាឧទាហរណ៍នៅក្នុងសារាយដែលមានសរសៃដូចជា Spirogyra ។ សរសៃ Spirogyra អាចបំបែកជាពីរផ្នែកនៅកន្លែងណាមួយ ដែលសារពាង្គកាយពីរត្រូវបានបង្កើតឡើងជាបន្តបន្ទាប់។
ការបែកខ្ញែកក៏ត្រូវបានគេសង្កេតឃើញផងដែរនៅក្នុងសត្វទាបមួយចំនួន ដែលមិនដូចទម្រង់ដែលមានការរៀបចំខ្ពស់នោះទេ រក្សានូវសមត្ថភាពសំខាន់ក្នុងការបង្កើតឡើងវិញពីកោសិកាដែលមានលក្ខណៈខុសគ្នាតិចតួច។ ឧទាហរណ៍ រាងកាយរបស់ nemerteans (ក្រុមនៃពពួក Worm បុព្វកាល ជាចម្បងនៅក្នុងសមុទ្រ) ត្រូវបានរហែកយ៉ាងងាយជាផ្នែកជាច្រើន ដែលផ្នែកនីមួយៗអាចបង្កើតឱ្យមានបុគ្គលថ្មីជាលទ្ធផលនៃការបង្កើតឡើងវិញ។ ក្នុងករណីនេះ ការបង្កើតឡើងវិញគឺជាដំណើរការធម្មតា និងគ្រប់គ្រង។ ទោះយ៉ាងណាក៏ដោយនៅក្នុងសត្វមួយចំនួន (ឧទាហរណ៍ត្រីផ្កាយ) ការស្ដារឡើងវិញពីផ្នែកនីមួយៗកើតឡើងតែបន្ទាប់ពីការបំបែកដោយចៃដន្យប៉ុណ្ណោះ។ ការបែកខ្ញែកត្រូវបានគេសង្កេតឃើញនៅក្នុងអេប៉ុង coelenterates (hydras), jellyfish, echinoderms, annelids និង flatworms ។ ជួនកាលសមត្ថភាពនេះត្រូវបានអភិវឌ្ឍយ៉ាងល្អដែលបុគ្គលទាំងមូលត្រូវបានស្ដារឡើងវិញពីបំណែកដាច់ដោយឡែកមួយ។
អង្ករ។ . ការបង្កើតឡើងវិញនៃត្រីផ្កាយពីកាំរស្មីមួយ។ A, B, C ដំណាក់កាលបន្តបន្ទាប់នៃការបង្កើតឡើងវិញ
ការបន្តពូជលូតលាស់នេះគឺជាការបន្តពូជ ដែលនៅក្នុងសារពាង្គកាយកូនស្រីថ្មីបានវិវត្តន៍ចេញពីផ្នែកដែលបំបែកចេញពីសារពាង្គកាយម្តាយ។ ក្នុងករណីនេះ ផ្នែកដែលមានភាពខុសប្លែកគ្នាយ៉ាងល្អត្រូវបានបំបែកចេញពីគំរូម្តាយ ដែលវាវិវត្តទៅជារុក្ខជាតិឯករាជ្យ។ ឬរុក្ខជាតិបង្កើតរចនាសម្ព័ន្ធពិសេសដែលត្រូវបានរចនាឡើងជាពិសេសសម្រាប់ការបន្តពូជលូតលាស់។ ឧទាហរណ៍អំពូល, មើម, corms, rhizomes, tendrils, buds ។ រចនាសម្ព័ន្ធទាំងនេះមួយចំនួនក៏បម្រើក្នុងការរក្សាទុកសារធាតុចិញ្ចឹមផងដែរ ដែលអនុញ្ញាតឱ្យរុក្ខជាតិអាចរស់រានបានរយៈពេលនៃលក្ខខណ្ឌមិនអំណោយផលដូចជាត្រជាក់ ឬគ្រោះរាំងស្ងួត។ ការបន្តពូជរបស់រុក្ខជាតិគឺជាលក្ខណៈនៃក្រុមរុក្ខជាតិជាច្រើន ចាប់ពីសារាយរហូតដល់រុក្ខជាតិផ្កា។
ជាគោលការណ៍ ការបន្តពូជរបស់រុក្ខជាតិគឺអនុវត្តមិនខុសពីការបំបែក ឬពន្លកទេ ប៉ុន្តែជាប្រពៃណី ពាក្យនេះត្រូវបានអនុវត្តចំពោះសារពាង្គកាយរុក្ខជាតិ ហើយពេលខ្លះចំពោះសត្វប៉ុណ្ណោះ ផ្ទុយពីការបែកខ្ញែក និងការពន្លក។
ក្លូន។ នេះគឺជាវិធីសាស្រ្តនៃការបន្តពូជសិប្បនិម្មិតដែលមិនកើតឡើងដោយធម្មជាតិ។ វាបានរីករាលដាលតែក្នុងរយៈពេល 30-40 ឆ្នាំចុងក្រោយនេះ ហើយត្រូវបានប្រើប្រាស់កាន់តែខ្លាំងឡើងសម្រាប់គោលបំណងសេដ្ឋកិច្ច។ មានបច្ចេកទេសពិសេសមួយចំនួនដែលអនុញ្ញាតឱ្យអ្នកក្លូនរុក្ខជាតិ និងសត្វមួយចំនួន។ក្លូន សំដៅទៅលើពូជពង្សដែលដូចគ្នាបេះបិទទៅនឹងហ្សែនដែលទទួលបានពីបុគ្គលម្នាក់ដែលជាលទ្ធផលនៃវិធីសាស្រ្តមួយឬមួយផ្សេងទៀតនៃការបន្តពូជដោយភេទដូចគ្នា។ នេះគឺជាកន្លែងដែលឈ្មោះនៃវិធីសាស្រ្តនេះមកពី។
ការពិសោធន៍ក្លូនត្រូវបានអនុវត្តយ៉ាងទូលំទូលាយលើរុក្ខជាតិ ដែលដោយសារតែសមត្ថភាពខ្ពស់ក្នុងការបង្កើតឡើងវិញ។ កោសិកានីមួយៗត្រូវបានដាក់នៅលើប្រព័ន្ធផ្សព្វផ្សាយសារធាតុចិញ្ចឹមដែលពួកគេបែងចែក ហើយដោយប្រើបច្ចេកទេសពិសេស ម៉ាស់កោសិកាដែលមិនមានរចនាសម្ព័ន្ធត្រូវបានទទួល ឬ callus ។ បន្ទាប់មកពួកវាបណ្តាលឱ្យមានភាពខុសគ្នានៃ calli ដូចគ្នាបឋម និងការបង្កើតជាលិកា និងសរីរាង្គផ្សេងៗ ហើយទីបំផុតសារពាង្គកាយរុក្ខជាតិទាំងមូលដែលមានលក្ខណៈសម្បត្តិដូចគ្នាទៅនឹងកោសិកាដើមដែលកោសិកាត្រូវបានយក។
ដោយប្រើវិធីសាស្ត្រក្លូន ទម្រង់កូនកាត់ផ្សេងៗអាចទទួលបាន។ ដូច្នេះ ដោយមានជំនួយពីអង់ស៊ីម ឬអ៊ុលត្រាសោន ជញ្ជាំងកោសិកានៃកោសិការុក្ខជាតិត្រូវបានដកចេញ បន្ទាប់ពីនោះ protoplasts "អាក្រាត" អាចបញ្ចូលគ្នា ដែលបណ្តាលឱ្យកោសិកាកូនកាត់ (ឧទាហរណ៍ ប៉េងប៉ោះ-ដំឡូង ឬកូនកាត់ថ្នាំជក់-ប៉េទុយនី) ។ បន្ទាប់ពីនេះជញ្ជាំងកោសិកាត្រូវបានស្ដារឡើងវិញ callus ត្រូវបានបង្កើតឡើងហើយបន្ទាប់មករុក្ខជាតិកូនកាត់ទាំងមូល។
ក្នុងករណីសត្វ វិធីសាស្ត្រខាងក្រោមត្រូវបានប្រើប្រាស់៖ ស្នូលនៃស៊ុតត្រូវបានដកចេញ ឬបំផ្លាញ ហើយស្នូលនៃកោសិកា somatic (ឧទាហរណ៍កោសិកា epithelial) ត្រូវបានដាក់នៅកន្លែងរបស់វា។ នៅពេលអនាគត ពីស៊ុតបែបនេះ សារពាង្គកាយមួយអាចទទួលបាន ដែលមានលក្ខណៈដូចគ្នាបេះបិទទៅនឹងម្ចាស់ជំនួយស្នូលសត្វ។ នៅក្នុងវិធីនេះ អ្នកអាចទទួលបានក្លូនរបស់សត្វមួយចំនួន កង្កែបតោង(Xenopus), newts (Triturus) ។ បច្ចុប្បន្ននេះសូម្បីតែក្លូននៃថនិកសត្វក៏ត្រូវបានគេទទួលបានដែរ ដូចជាចៀមល្បីឈ្មោះ Dolly ។ ការអភិវឌ្ឍនៃបច្ចេកទេសក្លូនត្រូវបានអនុវត្តដោយសាខាពិសេសនៃជីវវិទ្យា - ជីវបច្ចេកវិទ្យាដែលភារកិច្ចមិនត្រូវបានកំណត់ចំពោះរឿងនេះទេ។
ការបន្តពូជផ្លូវភេទ
ការបន្តពូជផ្លូវភេទហៅថាការផ្លាស់ប្តូរនៃជំនាន់ និងការអភិវឌ្ឍន៍នៃសារពាង្គកាយដោយផ្អែកលើការលាយបញ្ចូលគ្នានៃកោសិកាមេរោគឯកទេស ( gametes ) និងការបង្កើតហ្សីហ្គោត។ ការបង្កើត gametes (ភេទស្រី និងបុរស ស៊ុត និងមេជីវិតឈ្មោល) កើតឡើងនៅក្នុង gonads ។
ការបន្តពូជផ្លូវភេទគឺជាតួយ៉ាងសម្រាប់សត្វមានជីវិតភាគច្រើន មានភាពជឿនលឿនជាងបើប្រៀបធៀបទៅនឹងការបន្តពូជដោយភេទ និងមានគុណសម្បត្តិហ្សែនដ៏ធំសម្បើម។ ការបន្តពូជផ្លូវភេទធានាបាននូវភាពចម្រុះនៃហ្សែននៃកូនចៅ ពីព្រោះ មានការបញ្ចូលគ្នានៃហ្សែនដែលពីមុនជាកម្មសិទ្ធិរបស់ឪពុកម្តាយទាំងពីរ។ ភាពចម្រុះនៃប្រភេទហ្សែនរបស់បុគ្គលដែលបង្កើតជាប្រភេទសត្វផ្តល់នូវលទ្ធភាពនៃការសម្របខ្លួនបានជោគជ័យ និងឆាប់រហ័សនៃប្រភេទសត្វទៅនឹងការផ្លាស់ប្តូរលក្ខខណ្ឌបរិស្ថាន។
កំឡុងពេលបង្កកំណើត gametes បញ្ចូលគ្នាដើម្បីបង្កើតជា diploidហ្សីហ្គោត ពីនោះនៅក្នុងដំណើរការនៃការអភិវឌ្ឍសារពាង្គកាយចាស់ទុំត្រូវបានទទួល។ Gametes គឺ haploid - ពួកវាមានសំណុំក្រូម៉ូសូមមួយ zygote គឺ diploid ដែលមានសំណុំក្រូម៉ូសូមទ្វេ។ នេះគឺជាកោសិកាដំបូងនៃសារពាង្គកាយនាពេលអនាគត។
Gametes ជាធម្មតាមានពីរប្រភេទ - បុរសនិងស្ត្រី។ ពួកវាត្រូវបានផលិតដោយឪពុកម្តាយខាងប្រុសនិងស្រីប្រសិនបើប្រភេទនេះមាន dioecious; ( ទាំងនេះគឺជារុក្ខជាតិផ្កាមួយចំនួន សត្វ និងមនុស្សភាគច្រើន) ឬដោយបុគ្គលដូចគ្នា (harmaphroditism) ។
លក្ខណៈសម្រាប់សត្វឈ្មោលនិងញីdimorphism ផ្លូវភេទ- i.e. ភាពខុសគ្នាខាងផ្លូវភេទនៅក្នុងរចនាសម្ព័ន្ធ រូបរាង (ទំហំ ពណ៌ និងលក្ខណៈសម្បត្តិផ្សេងទៀត) ក៏ដូចជាអាកប្បកិរិយា។ នៅក្នុងសត្វ វាកើតឡើងរួចទៅហើយនៅដំណាក់កាលទាបនៃការអភិវឌ្ឍន៍វិវត្តន៍ ឧទាហរណ៍នៅក្នុង helminths ជុំ និង arthropods ហើយឈានដល់ការបញ្ចេញមតិដ៏អស្ចារ្យបំផុតរបស់វានៅក្នុងសត្វឆ្អឹងខ្នង ដែលភាពខុសគ្នាខាងក្រៅរវាងបុរស និងស្ត្រីគឺបង្ហាញយ៉ាងខ្លាំង។ នៅក្នុងរុក្ខជាតិនៃប្រភេទសត្វទាំងនោះដែលត្រូវបានកំណត់លក្ខណៈដោយវត្តមានរបស់បុគ្គលប្រុស និងស្រី ភាពខុសប្រក្រតីផ្លូវភេទក៏កើតឡើងដែរ ប៉ុន្តែវាត្រូវបានបង្ហាញតិចតួចណាស់។
ការប្រៀបធៀបនៃការបន្តពូជផ្លូវភេទ និងផ្លូវភេទត្រូវបានផ្តល់ឱ្យក្នុងតារាង។ ៥
តុ ៥
ការប្រៀបធៀបការបន្តពូជផ្លូវភេទ និងផ្លូវភេទ
|
ការបន្តពូជផ្លូវភេទ |
ការបន្តពូជផ្លូវភេទ (មិនរាប់បញ្ចូលបាក់តេរី) |
|
|
ឪពុកម្តាយម្នាក់ |
ជាធម្មតាឪពុកម្តាយពីរនាក់ |
|
|
Gametes មិនត្រូវបានបង្កើតឡើងទេ។ |
Haploid gametes ត្រូវបានបង្កើតឡើងដែលជាស្នូលដែលហ្វុយស៊ីប (ការបង្កកំណើត) ដើម្បីបង្កើតជាហ្សីកូតឌីផូអ៊ីត |
|
|
គ្មាន meiosis |
នៅដំណាក់កាលខ្លះនៃវដ្តជីវិត meiosis កើតឡើងដែលរារាំងក្រូម៉ូសូមមិនឱ្យកើនឡើងទ្វេដងក្នុងជំនាន់នីមួយៗ។ |
|
|
កូនចៅគឺដូចគ្នាបេះបិទទៅនឹងឪពុកម្តាយរបស់ពួកគេ។. ប្រភពតែមួយគត់នៃការប្រែប្រួលហ្សែនគឺការផ្លាស់ប្តូរដោយចៃដន្យ |
កូនចៅមិនដូចគ្នាទេ។ បុគ្គលមាតាបិតា។ ពួកវាបង្ហាញពីការប្រែប្រួលហ្សែនដែលបណ្តាលមកពីការផ្សំហ្សែនឡើងវិញ. នេះជំរុញការជ្រើសរើសធម្មជាតិនៃបុគ្គលដែលខ្លាំង និងសមបំផុត ហើយជាលទ្ធផល ការវិវត្តន៍។ |
|
|
លក្ខណៈរុក្ខជាតិ សត្វទាបមួយចំនួន និងអតិសុខុមប្រាណ។ មិនត្រូវបានរកឃើញនៅក្នុងសត្វខ្ពស់ជាង |
លក្ខណៈរុក្ខជាតិ និងសត្វភាគច្រើន |
|
|
ជារឿយៗនាំឱ្យមានការបង្កើតយ៉ាងឆាប់រហ័សនៃកូនចៅមួយចំនួនធំ |
តិចជាងការកើនឡើងយ៉ាងឆាប់រហ័សនៃចំនួន |
ការបន្តពូជផ្លូវភេទ អាស្រ័យលើសមាមាត្រនៃកោសិកាមេជីវិតក្នុងទំហំ និងមុខងារអាចមានបីជម្រើស៖
អ៊ីសូហ្គាមី (សារាយ, ប្រូហ្សូអា) ។ កោសិកាបន្តពូជបុរស និងស្ត្រីគឺដូចគ្នាបេះបិទក្នុងទំហំ រចនាសម្ព័ន្ធ និងការចល័ត។
Heterogamy ។ កោសិកាមានទំហំ និងរចនាសម្ព័ន្ធខុសគ្នា។
Oogamy នេះគឺជាវ៉ារ្យ៉ង់មួយនៃ heterogamy នៅពេលដែលស៊ុតមានទំហំធំ និងមិនអាចចល័តបាន ហើយមេជីវិតឈ្មោលមានទំហំតូចជាង មានសរីរាង្គនៃចលនា និងមានភាពចល័ត។
មានទម្រង់ពិសេសនៃការបន្តពូជផ្លូវភេទ ដូចជា hermaphroditism និង parthenogenesis ។
Hermaphroditism ។ ពាក្យ "hermaphroditism" គឺជាការរួមបញ្ចូលគ្នានៃឈ្មោះក្រិក Hermes (ព្រះនៃភាពស្រស់ស្អាតបុរស) និង Aphrodite (នាគរាជនៃភាពស្រស់ស្អាតស្ត្រី) ។
ទោះជាយ៉ាងណាក៏ដោយ នៅក្នុងប្រភេទសត្វ hermaphroditic ភាគច្រើន ការបង្កកំណើតពាក់ព័ន្ធនឹង gametes ដែលបានមកពីបុគ្គលផ្សេងៗគ្នា ហើយពួកវាមានការសម្របសម្រួលហ្សែន រូបវិទ្យា និងសរីរវិទ្យាជាច្រើនដែលការពារការបង្កកំណើតដោយខ្លួនឯង និងអនុគ្រោះដល់ការបង្កកំណើតឆ្លង។ ឧទាហរណ៍ ការបង្កកំណើតដោយខ្លួនឯងនៃប្រូតូហ្សូអាជាច្រើនត្រូវបានរារាំងដោយភាពមិនស៊ីគ្នានៃហ្សែន នៅក្នុងរុក្ខជាតិផ្កាជាច្រើនដោយរចនាសម្ព័ន្ធរបស់ androecium និង gynoecium និងនៅក្នុងសត្វជាច្រើនដោយការពិតដែលថាស៊ុត និងមេជីវិតឈ្មោលត្រូវបានបង្កើតឡើងនៅក្នុងបុគ្គលតែមួយនៅពេលផ្សេងគ្នា។
បាតុភូតនៃ hermaphroditism ពិតត្រូវបានរកឃើញផងដែរនៅក្នុងសត្វដែលមានរចនាសម្ព័ន្ធកាន់តែច្រើន។ ជាពិសេសវាត្រូវបានរកឃើញនៅក្នុងថនិកសត្វ។ ឧទាហរណ៍នៅក្នុងសត្វជ្រូក ការវិវត្តនៃអូវែជួនកាលត្រូវបានគេសង្កេតឃើញនៅផ្នែកម្ខាងនៃរាងកាយ និងការវិវឌ្ឍន៍នៃពងស្វាស (ពងស្វាស) នៅម្ខាងទៀត ឬការអភិវឌ្ឍនៃរចនាសម្ព័ន្ធរួមបញ្ចូលគ្នា (ovotestis) ហើយក្នុងករណីទាំងពីរនេះ ការសំយោគមុខងារ។ ស៊ុតនិងមេជីវិតឈ្មោលសកម្មកើតឡើង។ សត្វបែបនេះត្រូវបានចាត់ថ្នាក់ជាការរួមភេទ "កម្រិតមធ្យម" ដោយភាគច្រើននៃបុគ្គលនៃប្រភេទផ្លូវភេទកម្រិតមធ្យមគឺជាស្ត្រីដែលមានក្រូម៉ូសូម XX ពីរ។ បាតុភូតស្រដៀងគ្នានេះត្រូវបានកត់សម្គាល់នៅក្នុងពពែ។
Hermaphroditism ពិតក៏កើតមានចំពោះមនុស្សដែរ ដែលបណ្តាលមកពីការវិវត្តន៍ខុសប្រក្រតី។ ហ្សែននៃពពួក hermaphrodites គឺ 46XX ឬ 46 XY ជាមួយនឹងករណីភាគច្រើនសំដៅទៅលើ XX (ប្រហែល 60%) ។ ជំងឺក្លនលូនក្លែងក្លាយ ត្រូវបានគេស្គាល់ផងដែរ នៅពេលដែលបុគ្គលម្នាក់ៗមានប្រដាប់បន្តពូជខាងក្រៅ និងលក្ខណៈផ្លូវភេទបន្ទាប់បន្សំ ដែលជាលក្ខណៈនៃភេទទាំងពីរ ប៉ុន្តែផលិតកោសិកាមេរោគនៃប្រភេទតែមួយ គឺបុរស ឬស្ត្រី។
រុក្ខជាតិផ្កាភាគច្រើនមានផ្កា hermaphroditic ដែលជាធម្មតាត្រូវបានគេហៅថា bisexual ដោយសារតែផ្កានីមួយៗមាន pistil និង stamens ។ សម្រាប់ហេតុផលនេះផ្លែឈើអភិវឌ្ឍពីផ្កាទាំងអស់។ ស្រូវសាលី ផ្លែ cherry ផ្លែប៉ោម និងប្រភេទរុក្ខជាតិជាច្រើនទៀតគឺជាសត្វទ្វេរភេទ។ បន្ថែមពីលើ bisexuals ក្នុងអំឡុងពេលវិវត្តន៍ រុក្ខជាតិជាមួយនឹងការបំបែកភេទនៅក្នុងប្រភេទដូចគ្នាបានបង្កើតឡើង ពោលគឺ monoecious និង dioecy នៃរុក្ខជាតិកើតឡើង។ រុក្ខជាតិដែលមានទាំងផ្កា pistillate (ស្រី) និង staminate (បុរស) ត្រូវបានគេហៅថា monoecious ។ នៅក្នុងរុក្ខជាតិ monoecious ផ្លែឈើអភិវឌ្ឍតែពីផ្កា pistillate ។ ពោតគឺ monoeciousត្រសក់ ល្ពៅ និងអ្នកដទៃ។ ផ្ទុយទៅវិញរុក្ខជាតិ dioecious គឺជារុក្ខជាតិដែលមានផ្កា pistillate ឬ staminate (នៅក្នុងប្រភេទដូចគ្នា) ។ នៅក្នុងរុក្ខជាតិ dioecious មានតែផ្កាដែលមានផ្កា pistillate (ស្រី) ប៉ុណ្ណោះដែលបង្កើតផល។ Poplar, strawberry និងប្រភេទផ្សេងទៀតនៃរុក្ខជាតិ woody និង herbaceous គឺ dioecious ។
Parthenogenesis (មកពីភាសាក្រិក។ parthenos ព្រហ្មចារី និងហ្សែន កំណើត) គឺជាការកែប្រែមួយនៃការបន្តពូជផ្លូវភេទដែល gamete ស្ត្រីវិវត្តទៅជាបុគ្គលថ្មីដោយគ្មានការបង្កកំណើតដោយ gamete បុរស។ អត្ថប្រយោជន៍នៃ parthenogenesis គឺថាក្នុងករណីខ្លះវាបង្កើនអត្រានៃការបន្តពូជហើយនៅក្នុងសត្វល្អិតសង្គមវាអនុញ្ញាតឱ្យអ្នកគ្រប់គ្រងចំនួនកូនចៅនៃប្រភេទនីមួយៗ។ Parthenogenesis អាចជាកាតព្វកិច្ច (ជាកាតព្វកិច្ច) និង facultative (ស្រេចចិត្ត) ។
ឧទាហរណ៍នៅក្នុងសត្វឃ្មុំ(Apis mellifera) ម្ចាស់ក្សត្រីដាក់ពងដែលបង្កកំណើត(2 ភី = 32) ដែលនៅពេលអភិវឌ្ឍ បង្កើតបានជាស្ត្រី (ម្ចាស់ក្សត្រី ឬកម្មករ) និងពងមិនទាន់បង្កកំណើត។(ទំ = 16) ដែលផលិតបុរស (ដ្រូន) ដែលផលិតមេជីវិតឈ្មោលដោយ mitosis ជាជាង meiosis ។
អង្ករ។ គ្រោងការណ៍នៃប្រភេទនៃ parthenogenesis ។
- វដ្តកោសិកា។ មីតូស៊ីស។ ម៉ីអូស៊ីស
ការបែងចែកកោសិកាគឺជាមូលដ្ឋាននៃការបន្តពូជ និងការអភិវឌ្ឍន៍បុគ្គលនៃសារពាង្គកាយ។
ភាវៈរស់ទាំងអស់ត្រូវបានបង្កើតឡើងពីកោសិកា។ ការអភិវឌ្ឍន៍ ការរីកលូតលាស់ និងការបង្កើតរចនាសម្ព័ន្ធធម្មតានៃរាងកាយត្រូវបានអនុវត្តតាមរយៈការបន្តពូជនៃកោសិកាដើមមួយ ឬក្រុម។ នៅក្នុងដំណើរការនៃជីវិត កោសិកាមួយចំនួនរបស់រាងកាយអស់រលីង អាយុ និងស្លាប់។ ដើម្បីរក្សារចនាសម្ព័ន្ធ និងដំណើរការធម្មតា រាងកាយត្រូវតែផលិតកោសិកាថ្មី ដើម្បីជំនួសកោសិកាចាស់។ មធ្យោបាយតែមួយគត់ដើម្បីបង្កើតកោសិកាគឺដោយការបែងចែកមុន ៗ ។
ការបែងចែកកោសិកា ដំណើរការសំខាន់សម្រាប់សារពាង្គកាយទាំងអស់។ នៅក្នុងខ្លួនមនុស្សមានប្រមាណ ១០ 13 កោសិកាជាច្រើនលាននៃពួកគេត្រូវតែបែងចែកជារៀងរាល់វិនាទី។
វិធីសាស្រ្តបីនៃការបែងចែកកោសិកា eukaryotic ត្រូវបានពិពណ៌នា៖អាមីតូស៊ីស (ការបែងចែកដោយផ្ទាល់),មីតូស៊ីស (ការបែងចែកដោយប្រយោល) និង meiosis (ផ្នែកកាត់បន្ថយ) ។
អាមីតូស៊ីស វិធីសាស្រ្តកម្រ និងសិក្សាតិចតួចនៃការបែងចែកកោសិកា។ វាត្រូវបានពិពណ៌នាសម្រាប់ភាពចាស់ និងកោសិកាដែលប្រែប្រួលតាមរោគសាស្ត្រ។ នៅក្នុង amitosis ស្នូល interphase ត្រូវបានបែងចែកដោយការរឹតបន្តឹង ហើយការចែកចាយឯកសណ្ឋាននៃសម្ភារៈតំណពូជមិនត្រូវបានធានាទេ។ ជារឿយៗស្នូលបែងចែកដោយគ្មានការបំបែកជាបន្តបន្ទាប់នៃ cytoplasm និងកោសិកា binucleate ត្រូវបានបង្កើតឡើង។ កោសិកាដែលបានឆ្លងកាត់ amitosis ជាបន្តបន្ទាប់មិនអាចចូលទៅក្នុងវដ្ត mitotic ធម្មតាបានទេ។ ដូច្នេះ amitosis កើតឡើងជាក្បួននៅក្នុងកោសិកា និងជាលិកាដែលនឹងត្រូវស្លាប់ ឧទាហរណ៍នៅក្នុងកោសិកានៃភ្នាសអំប្រ៊ីយ៉ុងនៃថនិកសត្វ និងនៅក្នុងកោសិកាដុំសាច់។
មីតូស៊ីស វិធីសាស្រ្តសកលនៃការបែងចែកកោសិកា eukaryotic ។ រយៈពេលរបស់វានៅក្នុងកោសិកាសត្វគឺប្រហែល 1 ម៉ោង Mitosis គឺជាដំណើរការបន្តដែលជាធម្មតាត្រូវបានបែងចែកទៅជាបួនដំណាក់កាលគឺ prophase, metaphase, anaphase និង telophase ។
លំដាប់នៃព្រឹត្តិការណ៍ដែលកើតឡើងរវាងការបង្កើតកោសិកាដែលបានផ្តល់ឱ្យ និងការបែងចែករបស់វាទៅជាកោសិកាកូនស្រីត្រូវបានគេហៅថាវដ្តកោសិកា. វដ្តនេះមានបីដំណាក់កាលសំខាន់ៗ៖
1. អន្តរដំណាក់កាល។ រយៈពេលនៃការសំយោគខ្លាំងនិងកំណើន។ កោសិកាសំយោគសារធាតុជាច្រើនដែលចាំបាច់សម្រាប់ការលូតលាស់របស់វា និងការអនុវត្តមុខងារទាំងអស់របស់វា។ ក្នុងអំឡុងពេល interphase ការចម្លង DNA កើតឡើង។
2. មីតូស៊ីស។ នេះគឺជាដំណើរការនៃការបែងចែកនុយក្លេអ៊ែរ (karyokinesis) ដែលក្រូម៉ាទីតត្រូវបានបំបែកចេញពីគ្នាទៅវិញទៅមក និងចែកចាយឡើងវិញជាក្រូម៉ូសូមរវាងកោសិកាកូនស្រី។
3. Cytokinesis គឺជាដំណើរការនៃការបែងចែក cytoplasm (cytokinesis) រវាងកោសិកាកូនស្រីពីរ។
រយៈពេលនៃវដ្តកោសិកាអាស្រ័យលើប្រភេទនៃកោសិកា និងលើកត្តាខាងក្រៅដូចជា សីតុណ្ហភាព សារធាតុចិញ្ចឹម និងអុកស៊ីសែន។ កោសិកាបាក់តេរីអាចបែងចែករៀងរាល់ 20 នាទីម្តង កោសិកា epithelial ពោះវៀន - រៀងរាល់ 8-10 កោសិកានៅចុងឫសខ្ទឹមបារាំង - រៀងរាល់ 20 ម៉ោងម្តង ហើយកោសិកាជាច្រើននៃប្រព័ន្ធសរសៃប្រសាទមិនដែលបែងចែកឡើយ។
ទំនាក់ទំនងរវាងដំណើរការតាមពេលវេលាត្រូវបានបង្ហាញនៅក្នុងអង្ករ។

អង្ករ។ ដំណាក់កាលនៃវដ្តកោសិកា
អន្តរដំណាក់កាល រួមមានរយៈពេលជាច្រើន៖ G 1, S, G 2 ។
រយៈពេល G 1 ហៅថា presynthetic. អថេរភាគច្រើនក្នុងរយៈពេល។ នៅពេលនេះដំណើរការសំយោគជីវសាស្រ្តត្រូវបានធ្វើឱ្យសកម្មនៅក្នុងកោសិកាដែលជាចម្បងប្រូតេអ៊ីនរចនាសម្ព័ន្ធនិងមុខងារ។ កោសិកាលូតលាស់ និងរៀបចំសម្រាប់រយៈពេលបន្ទាប់។ ក្នុងអំឡុងពេលនេះ ដំណើរការជីវសំយោគដែលពឹងផ្អែកខ្លាំងកើតឡើង។ ការបង្កើត mitochondria, chloroplasts (នៅក្នុងរុក្ខជាតិ), endoplasmic reticulum, lysosomes, Golgi apparatus, vacuoles និង vesicles ។ nucleolus ផលិត rRNA, mRNA និង tRNA; ribosomes ត្រូវបានបង្កើតឡើង; កោសិកាសំយោគប្រូតេអ៊ីនរចនាសម្ព័ន្ធ និងមុខងារ។ ការរំលាយអាហារកោសិកាខ្លាំងដែលគ្រប់គ្រងដោយអង់ស៊ីម។ ការលូតលាស់កោសិកា។ ការបង្កើតសារធាតុដែលទប់ស្កាត់ឬជំរុញការចាប់ផ្តើមនៃដំណាក់កាលបន្ទាប់។
សម័យ S ហៅថាសំយោគ. នេះគឺជារយៈពេលដ៏សំខាន់នៅក្នុងវដ្ត mitotic ។ ក្នុងការបែងចែកកោសិកាថនិកសត្វ វាមានរយៈពេលប្រហែល 6 x 10 ម៉ោង។ ការចម្លង DNA កើតឡើងនៅទីនេះ។ ការសំយោគម៉ូលេគុលប្រូតេអ៊ីនហៅថា អ៊ីស្តូន ដែលភ្ជាប់ទៅនឹងខ្សែ DNA នីមួយៗ។ ក្រូម៉ូសូមនីមួយៗប្រែទៅជាក្រូម៉ាតពីរ។
រយៈពេល G2 ត្រូវបានគេហៅថា ក្រោយសំយោគ។វាខ្លីណាស់នៅក្នុងកោសិកាថនិកសត្វវាមានប្រហែល 2 x 5 ម៉ោង។ នៅពេលនេះចំនួននៃ centrioles, mitochondria និង plastids កើនឡើងទ្វេដង ដំណើរការមេតាបូលីសសកម្មកើតឡើង ប្រូតេអ៊ីន និងថាមពលត្រូវបានប្រមូលផ្តុំសម្រាប់ការបែងចែកនាពេលខាងមុខ។ កោសិកាចាប់ផ្តើមបែងចែក។ ដំណើរការ biosynthesis ដែលពឹងផ្អែកខ្លាំងកើតឡើង។ ការបែងចែក mitochondria និង chloroplasts ។ ទុនបម្រុងថាមពលកើនឡើង។ ការចម្លងនៃ centrioles (នៅក្នុងកោសិកាទាំងនោះដែលពួកគេមានវត្តមាន) និងការចាប់ផ្តើមនៃការបង្កើត spindle
មីតូស៊ីស បែងចែកតាមលក្ខខណ្ឌជាបួនដំណាក់កាល៖ prophase, metaphase, anaphase និង telophase
ប្រូហ្វាស . DNA spiralization ចាប់ផ្តើម និងកើនឡើងបន្តិចម្តងៗនៅក្នុងស្នូល។ ក្រូម៉ូសូមខ្លី ក្រាស់ អាចមើលឃើញ និងទទួលបានរចនាសម្ព័ន្ធ bichromatid ធម្មតា។ nucleolus បាត់បន្តិចម្តងៗ។ នៅក្នុង cytoplasm, microtubules ត្រូវបានតម្រង់ទិសជុំវិញគូនីមួយៗនៃ centrioles បង្កើតជាមជ្ឈមណ្ឌល spindle ។ Centrioles ផ្លាស់ទីទៅបង្គោលផ្សេងគ្នា microtubules លាតសន្ធឹងតាមអ័ក្សកោសិកា ហើយការបង្កើត spindle achromatin ចាប់ផ្តើម។ ស្រោមសំបុត្រនុយក្លេអ៊ែរបំបែកទៅជាបំណែកតូចៗដាច់ដោយឡែក។ ក្រូម៉ូសូមផ្លាស់ទីទៅកណ្តាលកោសិកា
មេតាហ្វាស . ក្រូម៉ូសូមត្រូវបានបង្រួបបង្រួមជាអតិបរមា និងរៀបចំតាមរបៀបដែលកណ្តាលរបស់ពួកវាស្ថិតនៅក្នុងប្លង់តែមួយ - យន្តហោះនៃអេក្វាទ័រកោសិកា។ បន្ទះ metaphase ត្រូវបានបង្កើតឡើង។ ការបង្កើត spindle mitotic ត្រូវបានបញ្ចប់។ Centrioles មានទីតាំងនៅជាគូនៅប៉ូលទល់មុខ ហើយខ្សែស្រឡាយ spindle ពីប៉ូលផ្សេងគ្នាត្រូវបានភ្ជាប់ទៅនឹង centromere នៃក្រូម៉ូសូមនីមួយៗ។
អាណាផាស . នេះគឺជាដំណាក់កាលខ្លីបំផុតនៃ mitosis ។ នៅទីនេះការបំបែកបណ្តោយនៃក្រូម៉ូសូមនីមួយៗកើតឡើង ការកាត់បន្ថយនៃខ្សែស្រឡាយspindles និងការបង្វែរក្រូម៉ូសូម (ក្រូម៉ូសូមកូនស្រី) ឆ្ពោះទៅរកប៉ូលនៃកោសិកា។
តេឡូហ្វាស . ក្រូម៉ូសូមកូនស្រីដែលមានក្រូម៉ាទីតមួយឈានដល់ប៉ូលនៃកោសិកា។ DNA ដែលផ្សំពួកវាចាប់ផ្តើមស្រងូតស្រងាត់ នុយក្លេអូលូសលេចឡើង ភ្នាសនុយក្លេអ៊ែរត្រូវបានបង្កើតឡើងជុំវិញក្រុមក្រូម៉ូសូមកូនស្រីនីមួយៗ ហើយខ្សែស្រឡាយនៃ achromatin spindle បានបែកបាក់បន្តិចម្តងៗ។ ការបំបែកនុយក្លេអ៊ែរត្រូវបានបញ្ចប់។
ការបែងចែក Cytoplasmic ចាប់ផ្តើម(ការធ្វើស៊ីតូតូមិច) និងការបង្កើត septum រវាងកោសិកាកូនស្រី។ កោសិកាសត្វធ្វើ cytotomy ដោយការបង្រួមនៃភ្នាស cytoplasmic ។ នៅក្នុងរុក្ខជាតិ ភ្នាសភ្នាសត្រូវបានបង្កើតឡើងនៅក្នុងប្លង់នៃកោសិកាអេក្វាទ័រ ដែលលូតលាស់នៅពេលក្រោយ ឈានដល់ជញ្ជាំងកោសិកា។ ជាលទ្ធផលកោសិកាកូនស្រីពីរដែលបំបែកចេញពីគ្នាទាំងស្រុងត្រូវបានបង្កើតឡើង។
តោះតាមដានទាំងអស់គ្នា ការផ្លាស់ប្តូរសម្ភារៈតំណពូជក្នុងអំឡុងពេលវដ្ត mitotic ។ ព្រឹត្តិការណ៍សំខាន់ៗនៃវដ្ត mitotic គឺការចម្លង DNA កើតឡើងនៅក្នុង interphase និងនាំឱ្យមានការកើនឡើងទ្វេដងនៃចំនួនព័ត៌មានតំណពូជ និងការបែងចែក chromatid,កើតឡើងនៅក្នុង anaphase នៃ mitosis និងធានាការចែកចាយឯកសណ្ឋាននៃពត៌មានតំណពូជរវាងកោសិកាកូនស្រី។ សម្ភារៈតំណពូជអនុវត្តដំណើរការទាំងនេះខណៈពេលដែលមានទម្រង់រចនាសម្ព័ន្ធផ្សេងៗគ្នា។ ការសំយោគចម្លងឆ្លងកាត់interphase chromatin, ក្នុងដែលក្នុងនោះម៉ូលេគុល DNA ស្ថិតនៅក្នុងស្ថានភាពធ្លាក់ទឹកចិត្ត។ ការចែកចាយព័ត៌មានហ្សែនត្រូវបានអនុវត្តក្រូម៉ូសូម mitotic,ដែលក្នុងនោះ DNA ត្រូវបានបំភាយជាអតិបរមា។
នៅក្នុងវដ្ត mitotic បរិមាណនៃសម្ភារៈតំណពូជក៏ផ្លាស់ប្តូរផងដែរ។ ប្រសិនបើចំនួនក្រូម៉ូសូមនៅក្នុងសំណុំ haploid ត្រូវបានតាងដោយអក្សរ p (ក្នុងសំណុំ diploid រៀងគ្នា 2n) ហើយចំនួនម៉ូលេគុល DNA ត្រូវបានតាងដោយអក្សរជាមួយ បន្ទាប់មកវាអាចធ្វើទៅបានដើម្បីតាមដានការផ្លាស់ប្តូរនៅក្នុងរូបមន្តនៃស្នូលនៃកោសិកា somatic នៅដំណាក់កាលផ្សេងគ្នានៃវដ្ត mitotic ។ ពីមុនស -កំឡុងពេលក្រូម៉ូសូមនីមួយៗមានម៉ូលេគុល DNA មួយ បរិមាណ DNA សរុបនៅក្នុងស្នូលត្រូវគ្នានឹងចំនួនក្រូម៉ូសូមនៅក្នុងនោះ ហើយរូបមន្តនៃកោសិកា diploid មានទម្រង់ 2p2s ។ បន្ទាប់ពីការចម្លងឡើងវិញ នៅពេលដែល DNA នៃក្រូម៉ូសូមនីមួយៗកើនឡើងទ្វេដង នោះចំនួនសរុបនៃ DNA នៅក្នុងស្នូលកើនឡើងទ្វេដង ហើយរូបមន្តកោសិកាទទួលបានទម្រង់ 2n4c ។ ជាលទ្ធផលនៃការបំបែក chromatid នៅក្នុង anaphase នៃ mitosis ស្នូលកូនស្រីទទួលបានសំណុំ diploid នៃក្រូម៉ូសូមតែមួយក្រូម៉ូសូម។ រូបមន្តនៃកោសិកាកូនស្រីម្តងទៀតក្លាយជា 2p2s ។
សារៈសំខាន់ជីវសាស្រ្តនៃ mitosisគឺថាជាលទ្ធផលនៃវិធីសាស្រ្តនៃការបែងចែកនេះ កោសិកាត្រូវបានបង្កើតឡើងជាមួយនឹងព័ត៌មានតំណពូជ ដែលមានលក្ខណៈគុណភាព និងបរិមាណដូចគ្នាទៅនឹងព័ត៌មាននៃកោសិកាម្តាយ។ ការចែកចាយឯកសណ្ឋាននៃសម្ភារៈតំណពូជត្រូវបានធានាដោយដំណើរការនៃការចម្លង DNA និងក្រូម៉ូសូមកើនឡើងទ្វេដងនៅក្នុងដំណាក់កាលនៃវដ្ត mitotic ក៏ដូចជាដោយការតំរៀបស្លឹក និងការចែកចាយឯកសណ្ឋាននៃ chromatids រវាងកោសិកាកូនស្រីអំឡុងពេល mitosis ។ Mitosis ធានានូវការថែរក្សានូវភាពស្ថិតស្ថេរនៃ karyotype លើការបង្កើតកោសិកាមួយចំនួន និងបម្រើជាយន្តការកោសិកាសម្រាប់ដំណើរការនៃការលូតលាស់ និងការអភិវឌ្ឍនៃរាងកាយ ក៏ដូចជាការបង្កើតឡើងវិញ និងការបន្តពូជដោយភេទដូចគ្នា។
សកម្មភាពនៃកត្តាបរិស្ថានមួយចំនួនអាចបង្អាក់ដំណើរធម្មតានៃ mitosis និងនាំឱ្យមានការខូចខាតដល់ក្រូម៉ូសូម ក៏ដូចជាការផ្លាស់ប្តូរចំនួនក្រូម៉ូសូមនីមួយៗ ឬសំណុំក្រូម៉ូសូមទាំងមូលនៅក្នុងកោសិកា somatic នៃរាងកាយ។ មីតូសរោគសាស្ត្រអាចបណ្តាលឱ្យមានជំងឺក្រូម៉ូសូមមួយចំនួន។ មីតូសរោគត្រូវបានគេសង្កេតឃើញជាពិសេសជាញឹកញាប់នៅក្នុងកោសិកាដុំសាច់។
ដ្យាក្រាម mitosis៖
មីតូស៊ីស
|
1. អន្តរដំណាក់កាល ជារឿយៗវាត្រូវបានគេហៅថាមិនត្រឹមត្រូវ ដំណាក់កាលសម្រាក។ រយៈពេលនៃ interphase ប្រែប្រួល និងអាស្រ័យលើមុខងារនៃក្រឡាដែលបានផ្តល់ឱ្យ។ នេះគឺជាអំឡុងពេលដែលកោសិកាជាធម្មតាសំយោគសរីរាង្គ និងបង្កើនទំហំ។ nucleoli អាចមើលឃើញយ៉ាងច្បាស់ និងសំយោគយ៉ាងសកម្មនូវសារធាតុ ribosomal ។ មុនពេលការបែងចែកកោសិកា DNA និងអ៊ីស្តូននៃក្រូម៉ូសូមនីមួយៗត្រូវបានចម្លង។ ឥឡូវនេះក្រូម៉ូសូមនីមួយៗត្រូវបានតំណាងដោយក្រូម៉ាទីតមួយគូដែលភ្ជាប់គ្នាទៅវិញទៅមកដោយ centromere ។ សារធាតុនៃក្រូម៉ូសូមមានស្នាមប្រឡាក់ ហើយត្រូវបានគេហៅថាក្រូម៉ាទីន ប៉ុន្តែរចនាសម្ព័ន្ធទាំងនេះខ្លួនឯងពិបាកមើលណាស់។ |
|
|
2. Prophase ជាធម្មតាដំណាក់កាលវែងបំផុតនៃការបែងចែកកោសិកា។ Chromatids ខ្លី (រហូតដល់ 4% នៃប្រវែងដើមរបស់ពួកគេ) និងក្រាស់ជាលទ្ធផលនៃការ spiralization និង condensation របស់ពួកគេ។ នៅពេលដែលមានស្នាមប្រឡាក់ ក្រូម៉ាទីតអាចមើលឃើញយ៉ាងច្បាស់ ប៉ុន្តែកណ្តាលមិនអាចមើលឃើញទេ។ នៅក្នុងគូផ្សេងគ្នានៃ chromatids, centromere មានទីតាំងស្ថិតនៅផ្សេងគ្នា។ នៅក្នុងកោសិកាសត្វ និងនៅក្នុងរុក្ខជាតិទាប centrioles បង្វែរទៅបង្គោលផ្ទុយនៃកោសិកា។ ពី centriole នីមួយៗ microtubules ខ្លីៗផុសឡើងក្នុងទម្រង់ជាកាំរស្មី បង្កើតជាផ្កាយ។ នុយក្លេអូលីក្លាយជាតូចជាងមុន ដោយសារអាស៊ីតនុយក្លេអ៊ីករបស់ពួកវាត្រូវបានផ្ទេរដោយផ្នែកទៅគូមួយចំនួននៃក្រូម៉ាទីត។ ឆ្ពោះទៅចុងបញ្ចប់នៃ prophase ភ្នាសនុយក្លេអ៊ែរបានបែកខ្ញែក ហើយ spindle fission ត្រូវបានបង្កើតឡើង។ |
|
|
3. Metaphase គូនៃ chromatids ត្រូវបានភ្ជាប់ដោយ centromeres របស់ពួកគេទៅនឹងសរសៃ spindle (microtubules) ហើយផ្លាស់ទីឡើងលើនិងចុះក្រោមរហូតដល់ centromeres របស់ពួកគេតម្រង់តាមខ្សែអេក្វាទ័រនៃ spindle កាត់កែងទៅនឹងអ័ក្សរបស់វា។ |
|
|
4. អាណាផាស នេះគឺជាដំណាក់កាលខ្លីណាស់។ centromere នីមួយៗបំបែកជាពីរ ហើយសរសៃ spindle ទាញ centromeres កូនស្រីទៅបង្គោលទល់មុខ។ Centromeres ទាញពីក្រោយពួកវានូវក្រូម៉ូសូមដែលបានបំបែកពីគ្នាទៅវិញទៅមក ដែលឥឡូវនេះត្រូវបានគេហៅថាក្រូម៉ូសូម។ |
|
|
5. Telophase ក្រូម៉ូសូមឈានដល់ប៉ូលកោសិកា ដកដង្ហើម ពន្លូត ហើយពួកវាមិនអាចសម្គាល់បានច្បាស់ទៀតទេ។ សរសៃ spindle ត្រូវបានបំផ្លាញ ហើយ centrioles ត្រូវបានចម្លង។ ភ្នាសនុយក្លេអ៊ែរត្រូវបានបង្កើតឡើងជុំវិញក្រូម៉ូសូមនៅបង្គោលនីមួយៗ។ នុយក្លេអូលលេចឡើងម្តងទៀត។ Telophase អាចត្រូវបានអនុវត្តភ្លាមៗដោយ cytokinesis (ការបែងចែកកោសិកាទាំងមូលជាពីរ) ។ |
|





Meiosis (ពីភាសាក្រិក meiosis ការកាត់បន្ថយ) វិធីពិសេសនៃការបែងចែកកោសិកា ដែលនាំទៅដល់ការកាត់ពាក់កណ្តាលនៃចំនួនក្រូម៉ូសូមនៅក្នុងពួកគេ។ Meiosis គឺជាតំណភ្ជាប់កណ្តាល gametogenesis នៅក្នុងសត្វនិង sporogenesis នៅក្នុងរុក្ខជាតិ។ Meiosis មានការបែងចែកបន្តបន្ទាប់គ្នាពីរដែលនាំមុខដោយការចម្លង DNA តែមួយ។ សារធាតុ និងថាមពលទាំងអស់ដែលចាំបាច់សម្រាប់ការបែងចែកទាំងពីរត្រូវបានរក្សាទុកក្នុងកំឡុងពេល interphase មុន meiosis I. ដំណាក់កាលទី II អវត្តមានជាក់ស្តែង ហើយការបែកបាក់គ្នាកើតឡើងយ៉ាងឆាប់រហ័ស។ នៅក្នុងការបែងចែក meiotic នីមួយៗមានបួនដំណាក់កាលដូចគ្នាត្រូវបានសម្គាល់: prophase, metaphase, anaphase និង telophase ដែលជាលក្ខណៈនៃ mitosis ប៉ុន្តែខុសគ្នានៅក្នុងលក្ខណៈមួយចំនួន។
ការបែងចែក meiotic ដំបូង (meiosis I ) នាំឱ្យមានការថយចុះពាក់កណ្តាលនៃចំនួនក្រូម៉ូសូម ហើយត្រូវបានគេហៅថាការកាត់បន្ថយ។ ជាលទ្ធផលពីកោសិកា diploid មួយ។(2 ទំ 4c) កោសិកា haploid ពីរត្រូវបានបង្កើតឡើង(ទំ 2 គ) កោសិកា។
ប្រូហ្វាស I meiosis គឺវែងបំផុតនិងស្មុគស្មាញបំផុត។ បន្ថែមពីលើដំណើរការនៃ DNA helixation និងការបង្កើត spindle ធម្មតានៃ prophase នៃ mitosis, I ព្រឹត្តិការណ៍ជីវសាស្ត្រសំខាន់ពីរកើតឡើង៖ការរួមផ្សំ ឬ synapsis ក្រូម៉ូសូម homologous និងឆ្លងកាត់។
Conjugation គឺជាដំណើរការនៃភាពជិតស្និទ្ធនៃក្រូម៉ូសូម homologous ។ ក្រូម៉ូសូមដែលបានផ្គូផ្គងទាំងនេះបង្កើតបាន។ bivalent និងត្រូវបានរក្សាទុកនៅក្នុងសមាសភាពរបស់វាដោយមានជំនួយពីប្រូតេអ៊ីនពិសេស។ ដោយសារក្រូម៉ូសូមនីមួយៗមានក្រូម៉ាទីតពីរ នោះ bivalent រួមមានក្រូម៉ាទីបួន ហើយត្រូវបានគេហៅថាសៀវភៅកត់ត្រា។ កោសិកា diploid ផលិតទំ bivalents ។ បន្ទាប់ពីការផ្សំ រូបមន្តក្រឡាយកទម្រង់ឆ្លងកាត់។
នៅកន្លែងខ្លះនៃ bivalent, chromatids នៃក្រូម៉ូសូម conjugated ប្រសព្វ បំបែក និងផ្លាស់ប្តូរផ្នែកដែលត្រូវគ្នា។ ដំណើរការនៃការផ្លាស់ប្តូរបំណែកនៃក្រូម៉ូសូម homologous ត្រូវបានគេហៅថាឆ្លងកាត់។ វាធានានូវការបង្កើតការរួមផ្សំថ្មីនៃហ្សែនឪពុក និងមាតានៅក្នុងក្រូម៉ូសូមនៃ gametes នាពេលអនាគត។ ការឆ្លងកាត់អាចកើតឡើងនៅក្នុងតំបន់ជាច្រើន (ការឆ្លងកាត់ច្រើនដង) ផ្តល់នូវកម្រិតខ្ពស់នៃការផ្សំឡើងវិញនៃព័ត៌មានតំណពូជនៅក្នុង gametes ។ នៅចុងបញ្ចប់នៃ prophase I កម្រិតនៃការក្រៀវនៃក្រូម៉ូសូមកើនឡើង ក្រូម៉ូសូមក្លាយទៅជាអាចបែងចែកបានយ៉ាងច្បាស់ ខ្សែស្រឡាយ spindle ពីបង្គោលនីមួយៗត្រូវបានភ្ជាប់ទៅកណ្តាលនៃក្រូម៉ូសូម bivalent មួយ។ ស្រោមសំបុត្រនុយក្លេអ៊ែរត្រូវបានបំផ្លាញ ហើយ bivalents ត្រូវបានតម្រង់ឆ្ពោះទៅរកយន្តហោះអេក្វាទ័រនៃកោសិកា។
នៅក្នុង metaphase I Meiosis បញ្ចប់ការបង្កើត spindle, bivalents ត្រូវបានដំឡើងនៅក្នុងយន្តហោះអេក្វាទ័រនៃកោសិកា។ ខ្សែ spindle ពីបង្គោលមួយត្រូវបានភ្ជាប់ទៅនឹង centromere នៃក្រូម៉ូសូមនីមួយៗ។
នៅក្នុងអាណាហ្វាស I នៅក្នុង meiosis នៅក្រោមសកម្មភាពនៃខ្សែស្រឡាយ spindle ក្រូម៉ូសូម homologous ផ្លាស់ទីឆ្ងាយពីគ្នាទៅវិញទៅមកឆ្ពោះទៅរកប៉ូលទល់មុខនៃកោសិកា។ ជាលទ្ធផលនៅបង្គោលនីមួយៗនៃក្រឡា aសំណុំ haploidក្រូម៉ូសូមដែលមានក្រូម៉ូសូម bichromatid មួយពីគូនៃក្រូម៉ូសូមដូចគ្នា។ នៅក្នុង anaphase I ក្រូម៉ូសូមនៃគូផ្សេងគ្នា, i.e. ក្រូម៉ូសូមដែលមិនមានលក្ខណៈដូចគ្នា មានឥរិយាបទដោយឯករាជ្យទាំងស្រុងពីគ្នាទៅវិញទៅមក ដោយធានានូវការបង្កើតនូវភាពខុសគ្នាដ៏ធំទូលាយនៃបន្សំ ក្រូម៉ូសូមរបស់ឪពុក និងម្តាយនៅក្នុងសំណុំ haploid នៃ gametes នាពេលអនាគត។ ចំនួននៃបន្សំបែបនេះត្រូវគ្នាទៅនឹងរូបមន្ត 2 P, ដែលជាកន្លែងដែលទំ ចំនួនគូនៃក្រូម៉ូសូម homologous ។ នៅក្នុងមនុស្ស តម្លៃនេះគឺស្មើនឹង 2, i.e. 8.4 10 វ៉ារ្យ៉ង់នៃការរួមបញ្ចូលគ្នានៃក្រូម៉ូសូមមាតានិងមាតាគឺអាចធ្វើទៅបាននៅក្នុង gametes របស់មនុស្ស។
ដូច្នេះភាពខុសគ្នានៃក្រូម៉ូសូម homologous នៅក្នុង anaphase I meiosis ធានាមិនត្រឹមតែការកាត់បន្ថយចំនួនក្រូម៉ូសូមនៅក្នុងកោសិកាបន្តពូជនាពេលអនាគតប៉ុណ្ណោះទេ ប៉ុន្តែវាក៏ជាភាពចម្រុះដ៏ធំនៃកោសិកាក្រោយៗទៀតផងដែរ ដោយសារតែការរួមបញ្ចូលគ្នាចៃដន្យនៃក្រូម៉ូសូមមាតា និងបិតានៃគូផ្សេងៗគ្នា។
នៅក្នុង telophase I Meiosis កើតឡើងនៅក្នុងការបង្កើតកោសិកាដែលស្នូលមានសំណុំក្រូម៉ូសូម haploid និងទ្វេដងនៃចំនួន DNA ចាប់តាំងពីក្រូម៉ូសូមនីមួយៗមានក្រូម៉ាតពីរ។ កោសិកាដែលកើតចេញពីការបែងចែក meiotic ដំបូងមានរូបមន្ត p2s ហើយបន្ទាប់ពី interphase ខ្លី ពួកគេចាប់ផ្តើមផ្នែកបន្ទាប់។
ការបែងចែក meiotic ទីពីរ (meiosis II ) ដំណើរការជា mitosis ធម្មតា (រូបភាព 5.3) ប៉ុន្តែខុសគ្នាត្រង់ថា កោសិកាដែលចូលទៅក្នុងវាមានសំណុំក្រូម៉ូសូម haploid ។ ជាលទ្ធផលនៃការបែងចែកនេះ។ទំ ក្រូម៉ូសូម bichromatid (r2c), បំបែក, ទម្រង់ទំ ក្រូម៉ូសូមតែមួយ(ps) ។ ការបែងចែកនេះត្រូវបានគេហៅថាសមីការ (ឬស្មើគ្នា) ។
ដូច្នេះបន្ទាប់ពីការបែងចែក meiotic ពីរបន្តបន្ទាប់គ្នា ពីកោសិកាមួយជាមួយនឹងសំណុំ diploid នៃក្រូម៉ូសូមពីរក្រូម៉ាទីត (2x4c) កោសិកាចំនួនបួនដែលមានសំណុំ haploid នៃក្រូម៉ូសូមតែមួយក្រូម៉ូសូមត្រូវបានបង្កើតឡើង។(ps) ។
សារៈសំខាន់ជីវសាស្រ្តនៃ meiosisមាននៅក្នុងការបង្កើតកោសិកាជាមួយនឹងសំណុំក្រូម៉ូសូមកាត់បន្ថយ និងរក្សាភាពស្ថិតស្ថេរនៃ karyotype ក្នុងសារពាង្គកាយជាច្រើនជំនាន់ដែលបន្តពូជ។ ឆ្លងតាមការរួមភេទ។ Meiosis បម្រើជាមូលដ្ឋានសម្រាប់ភាពប្រែប្រួលរួមបញ្ចូលគ្នា ដោយផ្តល់នូវភាពចម្រុះហ្សែននៃ gametes តាមរយៈដំណើរការនៃការឆ្លងកាត់ ភាពខុសគ្នា និងការរួមបញ្ចូលគ្នានៃក្រូម៉ូសូមមាតា និងបិតា។ ការផ្លាស់ប្តូររចនាសម្ព័ន្ធក្រូម៉ូសូមដោយសារតែការឆ្លងកាត់មិនស្មើគ្នា ការរំខានដល់ការបង្វែរក្រូម៉ូសូមទាំងអស់ ឬបុគ្គលនៅក្នុង anaphaseខ្ញុំ និង II ការបែងចែក meiotic នាំឱ្យមានការបង្កើត gametes មិនធម្មតានិងអាចបម្រើជាមូលដ្ឋានសម្រាប់ការស្លាប់នៃសារពាង្គកាយឬការវិវត្តនៃរោគសញ្ញាក្រូម៉ូសូមមួយចំនួននៅក្នុងកូនចៅ។
ដ្យាក្រាម និងការពិពណ៌នាសង្ខេបនៃដំណាក់កាលបន្តបន្ទាប់នៃ meiosis នៅក្នុងកោសិកាសត្វ។
|
1. អន្តរដំណាក់កាល រយៈពេលខុសគ្នាក្នុងចំណោមប្រភេទផ្សេងៗគ្នា។ ការចម្លងសរីរាង្គកើតឡើង ហើយកោសិកាកើនឡើងក្នុងទំហំ។ ការចម្លង DNA និង histones បញ្ចប់ជាចម្បងនៅក្នុងដំណាក់កាល premeiotic interphase ប៉ុន្តែផ្នែកខ្លះក៏លាតសន្ធឹងដល់ការចាប់ផ្តើមនៃ prophase ផងដែរ។ ឥឡូវនេះក្រូម៉ូសូមនីមួយៗត្រូវបានតំណាងដោយក្រូម៉ាទីតមួយគូដែលតភ្ជាប់ដោយ centromere ។ សារធាតុក្រូម៉ូសូមមានស្នាមប្រឡាក់ ប៉ុន្តែនៃរចនាសម្ព័ន្ធទាំងអស់មានតែ nucleoli ប៉ុណ្ណោះដែលអាចមើលឃើញយ៉ាងច្បាស់។ |
A. ដំណាក់កាលដំបូង I |
|
2. Prophase ដំណាក់កាលវែងបំផុត។ ជារឿយៗវាត្រូវបានបែងចែកទៅជា 5 ដំណាក់កាល (leptotema, zygonema, pachynema, diplonema និង diakinesis) ប៉ុន្តែនៅទីនេះវានឹងត្រូវបានគេចាត់ទុកថាជាលំដាប់បន្តនៃការផ្លាស់ប្តូរក្រូម៉ូសូម។ ក . ក្រូម៉ូសូមខ្លី និងមើលឃើញជារចនាសម្ព័ន្ធដាច់ដោយឡែក។ នៅក្នុងសារពាង្គកាយមួយចំនួន ពួកវាមើលទៅដូចជាខ្សែអង្កាំ៖ តំបន់នៃវត្ថុធាតុដែលមានស្នាមប្រឡាក់ខ្លាំង - ក្រូម៉ូសូម - ឆ្លាស់គ្នាជាមួយនឹងតំបន់ដែលមិនមានស្នាមប្រឡាក់។ Chromomeres គឺជាកន្លែងដែលសម្ភារៈក្រូម៉ូសូមត្រូវបាន coil យ៉ាងខ្លាំង។ |
|
|
ខ. ក្រូម៉ូសូម homologous ដែលកើតចេញពីស្នូលនៃ gametes មាតា និងបិតា ចូលទៅជិតគ្នាទៅវិញទៅមក និងភ្ជាប់គ្នា។ ក្រូម៉ូសូមទាំងនេះមានប្រវែងដូចគ្នា centromeres របស់ពួកគេកាន់កាប់ទីតាំងដូចគ្នា ហើយជាធម្មតាពួកវាមានចំនួនដូចគ្នានៃហ្សែនដែលបានរៀបចំក្នុងលំដាប់លីនេអ៊ែរដូចគ្នា។ ក្រូម៉ូសូមនៃក្រូម៉ូសូម homologous ស្ថិតនៅជាប់គ្នា។ ដំណើរការនៃការភ្ជាប់គ្នាអាចចាប់ផ្តើមនៅចំណុចជាច្រើននៅលើក្រូម៉ូសូមដែលបន្ទាប់មកត្រូវបានភ្ជាប់តាមបណ្តោយប្រវែងទាំងមូល (ដូចជាប្រសិនបើបង្រួមជាមួយគ្នា) ។ គូនៃក្រូម៉ូសូមដូចគ្នាដែលរួមបញ្ចូលគ្នាត្រូវបានគេហៅថា bivalents ។ bivalents ខ្លីនិងក្រាស់។ ក្នុងករណីនេះ ការវេចខ្ចប់ក្រាស់ទាំងពីរកើតឡើងនៅកម្រិតម៉ូលេគុល និងការរមួលដែលអាចមើលឃើញពីខាងក្រៅ (spiralization)។ ឥឡូវនេះក្រូម៉ូសូមនីមួយៗដែលមាន centromere របស់វាអាចមើលឃើញយ៉ាងច្បាស់។ |
ខ.ប្រូហ្វាស I |
|
IN ក្រូម៉ូសូមដូចគ្នាដែលបង្កើតជា bivalent ត្រូវបានបំបែកដោយផ្នែក ដូចជាការរុញច្រានឆ្ងាយពីគ្នាទៅវិញទៅមក។ ឥឡូវនេះអ្នកអាចឃើញថាក្រូម៉ូសូមនីមួយៗមានក្រូម៉ាតពីរ។ ក្រូម៉ូសូមនៅតែភ្ជាប់គ្នាទៅវិញទៅមកនៅចំណុចជាច្រើន។ ចំណុចទាំងនេះត្រូវបានគេហៅថា chiasmata (មកពីភាសាក្រិក។ចៃម៉ា - ឈើឆ្កាង) ។ នៅក្នុង chiasma នីមួយៗ ផ្នែកនៃ chromatids ត្រូវបានផ្លាស់ប្តូរជាលទ្ធផលនៃការសម្រាក និងការជួបជុំគ្នា ដែលនៅក្នុងនោះ 2 នៃ 4 threads មានវត្តមាននៅក្នុង chiasm នីមួយៗត្រូវបានពាក់ព័ន្ធ។ ជាលទ្ធផល ហ្សែនពីក្រូម៉ូសូមមួយ (ឧទាហរណ៍ ឪពុក - A, B, C) ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងហ្សែនពីក្រូម៉ូសូមមួយទៀត (ម្តាយ a,ខ , គ) ដែលនាំឱ្យមានការរួមផ្សំហ្សែនថ្មីនៅក្នុង chromatids លទ្ធផល។ ដំណើរការនេះត្រូវបានគេហៅថាឆ្លងកាត់។ ក្រូម៉ូសូមដូចគ្នាមិនបំបែកចេញពីគ្នាបន្ទាប់ពីឆ្លងកាត់ទេ ចាប់តាំងពីក្រូម៉ូសូមបងស្រី (នៃក្រូម៉ូសូមទាំងពីរ) នៅតែភ្ជាប់គ្នាយ៉ាងរឹងមាំរហូតដល់ anaphase ។ |
ខ. ឆ្លងកាត់អំឡុងពេលប្រូហ្វាស I |
|
ជី . chromatids នៃក្រូម៉ូសូម homologous បន្តវាយគ្នាទៅវិញទៅមក ហើយ bivalents ទទួលបានការកំណត់ជាក់លាក់មួយអាស្រ័យលើចំនួន chiasmata ។ Bivalents ដែលមាន chiasmata មួយមានរាងឈើឆ្កាង ដោយ chiasmata ពីរពួកវាមានរាងជារង្វង់ ហើយមានបី ឬច្រើនពួកវាបង្កើតជារង្វិលជុំដែលកាត់កែងទៅគ្នាទៅវិញទៅមក។ នៅចុងបញ្ចប់នៃ prophase ក្រូម៉ូសូមទាំងអស់ត្រូវបាន condensed ទាំងស្រុង និងមានស្នាមប្រឡាក់ខ្លាំង។ ការផ្លាស់ប្តូរផ្សេងទៀតកើតឡើងនៅក្នុងកោសិកា៖ ការធ្វើចំណាកស្រុកនៃ centrioles (ប្រសិនបើមាន) ទៅកាន់ប៉ូល ការបំផ្លាញ nucleoli និងភ្នាសនុយក្លេអ៊ែរ ហើយបន្ទាប់មកការបង្កើត filaments spindle ។ |
|
|
2. Metaphase bivalents តម្រង់ជួរនៅក្នុងយន្តហោះអេក្វាទ័រ បង្កើតជាបន្ទះ metaphase ។ centromeres របស់ពួកគេមានឥរិយាបទជារចនាសម្ព័ន្ធតែមួយ (ទោះបីជាពួកវាច្រើនតែលេចឡើងទ្វេរដង) និងរៀបចំសរសៃ spindle ដែលភ្ជាប់ទៅនឹងពួកវា ដែលនីមួយៗត្រូវបានតម្រង់ឆ្ពោះទៅរកបង្គោលតែមួយប៉ុណ្ណោះ។ ជាលទ្ធផលនៃកម្លាំងទាញខ្សោយនៃខ្សែស្រឡាយទាំងនេះ ខ្សែសង្វាក់នីមួយៗស្ថិតនៅក្នុងតំបន់អេក្វាទ័រ ហើយមជ្ឈមណ្ឌលទាំងពីររបស់វាស្ថិតនៅចម្ងាយដូចគ្នាពីវា មួយខាងក្រោម និងមួយទៀតនៅខាងលើ។ |
ឃ. ដំណាក់កាលមេតាយឺត I |
|
3. អាណាផាស មជ្ឈមណ្ឌលទាំងពីរដែលមានវត្តមាននៅក្នុង bivalent នីមួយៗមិនទាន់បែងចែកនៅឡើយទេ ប៉ុន្តែ chromatids បងប្អូនស្រីលែងនៅជាប់គ្នាទៀតហើយ។ សរសៃ spindle ទាញ centromeres ដែលនីមួយៗត្រូវបានផ្សារភ្ជាប់ជាមួយនឹង chromatids ពីរឆ្ពោះទៅរកប៉ូលទល់មុខនៃ spindle ។ ជាលទ្ធផលក្រូម៉ូសូមត្រូវបានបែងចែកទៅជាសំណុំ haploid ពីរដែលបញ្ចប់នៅក្នុងកោសិកាកូនស្រី។ |
អ៊ី. អាណាផាស I |
|
4. Telophase ភាពខុសគ្នានៃ centromeres ដូចគ្នា និងក្រូម៉ាទីតដែលជាប់ទាក់ទងគ្នាទៅនឹងប៉ូលទល់មុខ មានន័យថាការបញ្ចប់នៃការបែងចែក meiotic ដំបូង។ ចំនួនក្រូម៉ូសូមក្នុងមួយឈុតបានក្លាយទៅជាធំជាងពាក់កណ្តាល ប៉ុន្តែក្រូម៉ូសូមនៅបង្គោលនីមួយៗមានក្រូម៉ាតពីរ។ ដោយសារតែការឆ្លងកាត់ក្នុងអំឡុងពេលនៃការបង្កើត chiasmata ក្រូម៉ាទីតទាំងនេះមានលក្ខណៈហ្សែនមិនដូចគ្នាបេះបិទ ហើយក្នុងអំឡុងពេលនៃការបែងចែក meiotic ទីពីរពួកគេនឹងបំបែកចេញពីគ្នា។ spindles និងខ្សែស្រឡាយរបស់ពួកគេជាធម្មតាបាត់។ នៅក្នុងសត្វ និងរុក្ខជាតិខ្លះ chromatids despiral ភ្នាសនុយក្លេអ៊ែរបង្កើតនៅជុំវិញពួកវានៅបង្គោលនីមួយៗ ហើយស្នូលលទ្ធផលចូលទៅក្នុង interphase ។ បន្ទាប់មកការបែងចែក cytoplasm ចាប់ផ្តើម (នៅក្នុងសត្វ) ឬការបង្កើតជញ្ជាំងកោសិកាបែងចែក (នៅក្នុងរុក្ខជាតិ) ដូចជានៅក្នុង mitosis ។ នៅក្នុងរុក្ខជាតិជាច្រើន ទាំង telophase ឬការបង្កើតជញ្ជាំងកោសិកា ឬ interphase មិនត្រូវបានគេសង្កេតឃើញទេ ហើយកោសិកាបានផ្លាស់ប្តូរដោយផ្ទាល់ពី anaphase ។ខ្ញុំដើម្បីប្រកាស II ។ |
G. Telophase I នៅក្នុងកោសិកាសត្វ |







|
ដំណាក់កាលទី II ដំណាក់កាលនេះជាធម្មតាត្រូវបានគេសង្កេតឃើញតែនៅក្នុងកោសិកាសត្វប៉ុណ្ណោះ៖ រយៈពេលរបស់វាប្រែប្រួល។ដំណាក់កាល S គឺអវត្តមាន ហើយគ្មានការចម្លង DNA ទៀតកើតឡើងទេ។ ដំណើរការដែលពាក់ព័ន្ធនឹងការបែងចែកទីពីរនៃ meiosis គឺស្រដៀងគ្នានៅក្នុងយន្តការរបស់ពួកគេទៅនឹងអ្វីដែលកើតឡើងនៅក្នុង mitosis ។ ពួកគេពាក់ព័ន្ធនឹងការបំបែក chromatids នៅក្នុងកោសិកាកូនស្រីទាំងពីរដែលបណ្តាលមកពីការបែងចែក meiotic ដំបូង។ ការបែងចែកទីពីរនៃ meiosis ខុសគ្នាពី mitosis ជាចម្បងតាមពីរវិធី: 1) នៅក្នុង metaphase II meiosis, បងស្រី chromatids ជាញឹកញាប់ត្រូវបានបំបែកយ៉ាងខ្លាំងពីគ្នាទៅវិញទៅមក; 2) ចំនួននៃក្រូម៉ូសូមគឺ haploid ។ |
|
|
ដំណាក់កាលទី II នៅក្នុងកោសិកាដែលបាត់បង់អន្តរដំណាក់កាល II ដំណាក់កាលនេះក៏បាត់ដែរ។ រយៈពេលនៃ prophase II សមាមាត្របញ្ច្រាសទៅនឹងរយៈពេលនៃ telophase I . នុយក្លេអូលី និងភ្នាសនុយក្លេអ៊ែរត្រូវបានបំផ្លាញ ហើយក្រូម៉ាទីតត្រូវបានខ្លី និងក្រាស់។ Centrioles ប្រសិនបើមានវត្តមាន ផ្លាស់ទីទៅប៉ូលទល់មុខនៃកោសិកា។ សរសៃ spindle លេចឡើង។ chromatids ត្រូវបានរៀបចំតាមរបៀបដែលអ័ក្សវែងរបស់ពួកគេកាត់កែងទៅនឹងអ័ក្ស spindle នៃការបែងចែក meiotic ដំបូង។ |
Z. Prophase P |
|
Metaphase II ក្នុងអំឡុងពេលនៃការបែងចែកទីពីរ centromeres មានឥរិយាបទដូចជារចនាសម្ព័ន្ធទ្វេ។ ពួកគេរៀបចំខ្សែស្រឡាយ spindle ឆ្ពោះទៅរកបង្គោលទាំងពីរ ហើយដូច្នេះតម្រឹម spindle នៅអេក្វាទ័រ។ |
Metaphase II |
|
អាណាផាស II មជ្ឈមណ្ឌល omer បែងចែក ហើយខ្សែស្រឡាយ spindle ទាញពួកវាទៅបង្គោលទល់មុខ។ Centromeres ទាញតាមក្រូម៉ាទីតដែលបំបែកដែលឥឡូវត្រូវបានគេហៅថាក្រូម៉ូសូម។ Telophase II ដំណាក់កាលនេះគឺស្រដៀងទៅនឹង telophase នៃ mitosis ។ ក្រូម៉ូសូម despiral, stretch ហើយបន្ទាប់មកពិបាកក្នុងការបែងចែក។ សរសៃ spindle បាត់ ហើយ centrioles ចម្លង។ នៅជុំវិញស្នូលនីមួយៗ ដែលឥឡូវនេះមានពាក់កណ្តាលនៃចំនួនក្រូម៉ូសូម (haploid) នៃកោសិកាមេដើម ភ្នាសនុយក្លេអ៊ែរបង្កើតម្តងទៀត។ ជាលទ្ធផលនៃការបែងចែកជាបន្តបន្ទាប់នៃ cytoplasm (នៅក្នុងសត្វ) ឬការបង្កើតជញ្ជាំងកោសិកា (នៅក្នុងរុក្ខជាតិ) កោសិកាកូនស្រីចំនួន 4 ត្រូវបានទទួលពីកោសិកាមេដើមតែមួយ។ |
|


ភាពខុសគ្នារវាងដំណាក់កាលនៃ mitosis និង meiosis
|
ដំណាក់កាល |
មីតូស៊ីស |
ម៉ីអូស៊ីស |
|
ប្រូហ្វាស |
Chromomeres មិនអាចមើលឃើញទេ។ ក្រូម៉ូសូម homologous ត្រូវបានបំបែក Chiasmata មិនត្រូវបានបង្កើតឡើងទេ។ ការឆ្លងកាត់មិនកើតឡើងទេ។ |
Chromomeres អាចមើលឃើញ ក្រូម៉ូសូម homologous ត្រូវបានបញ្ចូលគ្នា។ Chiasmata ត្រូវបានបង្កើតឡើង។ ការឆ្លងកាត់អាចកើតឡើង |
|
មេតាហ្វាស |
គូ Chromatid មានទីតាំងនៅអេក្វាទ័រ spindle Centromeres តម្រង់ជួរនៅក្នុងយន្តហោះដូចគ្នានៅអេក្វាទ័រ spindle |
គូនៃ chromatids មានទីតាំងនៅអេក្វាទ័រ spindle តែនៅក្នុងផ្នែកទីពីរនៃ meiosis ប៉ុណ្ណោះ។ Centromeres នៅក្នុងផ្នែកដំបូងនៃ meiosis មានទីតាំងនៅខាងលើនិងខាងក្រោមអេក្វាទ័រនៅចម្ងាយស្មើគ្នាពីវា |
|
អាណាផាស |
កណ្តាលបែងចែក។ ក្រូម៉ាតដាច់ដោយឡែក។ ក្រូម៉ាទីតខុសគ្នាគឺដូចគ្នាបេះបិទ |
Centromeres បែងចែកតែនៅក្នុងដំណាក់កាលទីពីរនៃ meiosis ប៉ុណ្ណោះ។ chromatids ដាច់ដោយឡែកពីគ្នាក្នុងអំឡុងពេលនៃការបែងចែកទីពីរនៃ meiosis ។ នៅក្នុងផ្នែកទីមួយ ក្រូម៉ូសូមទាំងមូលដាច់ដោយឡែកពីគ្នា។ ការបង្វែរក្រូម៉ូសូមប្រហែលជាមិនដូចគ្នាទេ ដែលជាលទ្ធផលនៃការឆ្លងកាត់ |
|
Telolophase |
ចំនួនក្រូម៉ូសូមនៅក្នុងកោសិកាកូនស្រីគឺដូចគ្នាទៅនឹងកោសិកាមេដែរ។ កោសិកាកូនស្រីមានទាំងក្រូម៉ូសូមដូចគ្នា (ក្នុងឌីផូអ៊ីល) |
ចំនួនក្រូម៉ូសូមនៅក្នុងកោសិកាកូនស្រីគឺពាក់កណ្តាលនៃកោសិកាមេ កោសិកាកូនស្រីមានតែមួយគូនៃក្រូម៉ូសូមដូចគ្នា។ |
|
តើការបែងចែកប្រភេទនេះកើតឡើងនៅឯណា? |
អាចធ្វើទៅបាននៅក្នុងកោសិកា haploid, diploid និង polyploid កើតឡើងក្នុងអំឡុងពេលនៃការបង្កើតកោសិកា somatic និង spores មួយចំនួនក៏ដូចជាក្នុងអំឡុងពេលនៃការបង្កើត gametes នៅក្នុងរុក្ខជាតិដែលការជំនួសនៃជំនាន់កើតឡើង។ |
មានតែនៅក្នុងកោសិកា diploid និង polyploid ប៉ុណ្ណោះ។ កំឡុងពេល gameto- ឬ sporogenesis |
- រចនាសម្ព័ន្ធនៃ gametes ។ Gametogenesis
ការបន្តពូជផ្លូវភេទត្រូវបានអនុវត្តដោយប្រើកោសិកាពិសេសដែលហៅថា gametes ។ gametes ស្ត្រីត្រូវបានគេហៅថាស៊ុត, បុរស មេជីវិតឈ្មោល។Gametes ខុសគ្នាពីកោសិកា somatic ជាចម្បងដោយមានចំនួនក្រូម៉ូសូមពាក់កណ្តាល ក៏ដូចជាកម្រិតទាបនៃដំណើរការមេតាបូលីស។
អូវុល កោសិកាដែលមិនមានចលនាធំ ជាធម្មតាមានរាងមូល។ បន្ថែមពីលើសរីរាង្គធម្មតា cytoplasm ផ្ទុកនូវសារធាតុចិញ្ចឹមបម្រុងក្នុងទម្រង់ជា yolk (អង្ករ .) នៅក្នុងស្នូលនៃស៊ុត ច្បាប់ចម្លងជាច្រើននៃហ្សែន ribosomal និង mRNA ត្រូវបានបង្កើតឡើង ដែលធានាដល់ការសំយោគប្រូតេអ៊ីនសំខាន់ៗនៃអំប្រ៊ីយ៉ុងនាពេលអនាគត។ ស៊ុតនៃសារពាង្គកាយផ្សេងៗគ្នាមានភាពខុសគ្នានៅក្នុងបរិមាណនិងធម្មជាតិនៃការចែកចាយនៃ yolk នៅក្នុងពួកគេ។ មានស៊ុតជាច្រើនប្រភេទ។

ភាពឯកោត្រូវបានគេហៅថាស៊ុតតូចៗដែលមានចំនួនតិចតួចនៃ yolk ចែកចាយស្មើៗគ្នា។ ស្នូលនៅក្នុងពួកវាមានទីតាំងនៅជិតកណ្តាល។ ស៊ុតបែបនេះត្រូវបានរកឃើញនៅក្នុងពពួក Worm, bivalves និង gastropods, echinoderms និង lancelets ។telolecithal ល្មមស៊ុតរបស់ sturgeon និង amphibians មានអង្កត់ផ្ចិតប្រហែល 1.5 × 2 mm និងមានបរិមាណមធ្យមនៃ yolk ដែលភាគច្រើនត្រូវបានប្រមូលផ្តុំនៅបង្គោលមួយ (លូតលាស់) ។ នៅបង្គោលទល់មុខ (សត្វ) ដែលជាកន្លែងដែលមានពណ៌លឿងតិចតួច មានស្នូលនៃស៊ុត។
Telolecithal ខ្លាំងស៊ុតរបស់ត្រីមួយចំនួន សត្វល្មូន សត្វស្លាប និងថនិកសត្វ oviparous មានផ្ទុកនូវ yolk ជាច្រើន ដែលកាន់កាប់ស្ទើរតែបរិមាណទាំងមូលនៃ cytoplasm នៃស៊ុត។ នៅបង្គោលសត្វមានឌីសមេរោគដែលមាន cytoplasm សកម្មដោយគ្មាន yolk ។ ទំហំនៃស៊ុតទាំងនេះមានទំហំធំ 10 15 មមឬច្រើនជាងនេះ។
អាឡិចស៊ីតាល់ ស៊ុតគឺស្ទើរតែគ្មាន yolk មានទំហំតូចតាមមីក្រូទស្សន៍ (0.1 × 0.3 mm) និងជាលក្ខណៈនៃថនិកសត្វសុក រួមទាំងមនុស្សផងដែរ។
មេជីវិតឈ្មោល។ ឬមេជីវិតឈ្មោលគឺជា gametes បុរសចល័តតូចណាស់ (ឧទាហរណ៍មេជីវិតឈ្មោលរបស់មនុស្សមានប្រវែង 50 × 70 µm និងក្រពើ 20 µm) បង្កើតឡើងដោយ gonads បុរស - ពងស្វាស; ចំនួនរបស់ពួកគេគឺរាប់លាន។ រូបរាងរបស់មេជីវិតឈ្មោលមានភាពខុសប្លែកគ្នានៅក្នុងសត្វផ្សេងៗគ្នា ប៉ុន្តែរចនាសម្ព័ន្ធរបស់វាដូចគ្នា ភាគច្រើននៃពួកគេមានក្បាល និងក។ នៅក្នុងក្បាលមេជីវិតឈ្មោល មានស្នូលមួយដែលមានចំនួនក្រូម៉ូសូម haploid និងគ្របដណ្តប់។អាក្រូសូម។ Acrosome គឺជារចនាសម្ព័ន្ធពិសេស ស្មុគស្មាញ Golgi ដែលត្រូវបានកែប្រែ ដែលមានអង់ស៊ីមសម្រាប់រំលាយភ្នាសនៃស៊ុតអំឡុងពេលបង្កកំណើត ដែលត្រូវបានចងភ្ជាប់ដោយភ្នាស។ កមាន mitochondria ជាច្រើន និង centrioles ពីរ។ កន្ទុយដែលបង្កើតឡើងដោយ microtubules ដុះចេញពីកញ្ចឹងក និងធានាឱ្យមានចលនារបស់មេជីវិតឈ្មោល។ ផ្នែកកណ្តាលត្រូវបានពង្រីកដោយសារតែ mitochondria ជាច្រើនដែលវាមាន ប្រមូលផ្តុំគ្នាជាវង់ជុំវិញ flagellum ។ mitochondria ទាំងនេះផ្តល់ថាមពលសម្រាប់យន្តការ contractile ដែលផ្តល់ចលនានៃ flagellum ។ ផ្នែកសំខាន់និងកន្ទុយនៃមេជីវិតឈ្មោលមានលក្ខណៈរចនាសម្ព័ន្ធនៃ flagella ។

ប្រសិនបើអ្នកក្រឡេកមើលក្បាលមេជីវិតឈ្មោលរបស់មនុស្សពីខាងលើ វាមើលទៅរាងមូល ហើយនៅពេលដែលមើលពីចំហៀង វាហាក់ដូចជាសំប៉ែត។ ចលនា Flagellar តែឯងមិនគ្រប់គ្រាន់សម្រាប់មេជីវិតឈ្មោលក្នុងការធ្វើដំណើរចម្ងាយពីទ្វារមាសទៅកន្លែងបង្កកំណើតនោះទេ។ ភារកិច្ចចម្បងរបស់មេជីវិតឈ្មោលគឺត្រូវហែលជុំវិញ oocyte និងតម្រង់ទិសខ្លួនឯងតាមវិធីជាក់លាក់មួយ មុនពេលជ្រៀតចូលភ្នាសរបស់ oocyte ។
ការបង្កើត gametes (gametogenesis) ។កោសិកានៃ epithelium germinal ចំពោះបុរសនិង ហ្គូណាដស្ត្រីឆ្លងកាត់ការបែងចែក mitotic និង meiotic ជាបន្តបន្ទាប់ដែលត្រូវបានគេហៅថាជាសមូហភាព gametogenesis ដែលបណ្តាលឱ្យមានការបង្កើត gametes បុរសពេញវ័យ (spermatogenesis) និង gametes ស្ត្រី (oogenesis) ។ ក្នុងករណីទាំងពីរនេះ ដំណើរការចែកចេញជាបីដំណាក់កាល គឺដំណាក់កាលបន្តពូជ ដំណាក់កាលលូតលាស់ និងដំណាក់កាលពេញវ័យ។ ដំណាក់កាលបន្តពូជពាក់ព័ន្ធនឹងការបែងចែក mitotic ម្តងហើយម្តងទៀតដែលនាំទៅដល់ការបង្កើត spermatogonia ឬ oogonia ជាច្រើន។ ពួកគេម្នាក់ៗឆ្លងកាត់រយៈពេលនៃការលូតលាស់ក្នុងការរៀបចំសម្រាប់ការបែងចែក meiotic ដំបូង និង cytokinesis ជាបន្តបន្ទាប់។ បន្ទាប់មកដំណាក់កាលនៃភាពចាស់ទុំចាប់ផ្តើម ក្នុងអំឡុងពេលដែលការបែងចែក meiotic ទីមួយ និងទីពីរកើតឡើងជាមួយ ភាពខុសគ្នាជាបន្តបន្ទាប់នៃកោសិកា haploidនិង ការបង្កើត gametes ចាស់ទុំ។
ការបង្កើតមេជីវិតឈ្មោល (ការបង្កើតមេជីវិតឈ្មោល) ។មេជីវិតឈ្មោល (មេជីវិតឈ្មោល) ត្រូវបានបង្កើតឡើងជាលទ្ធផលនៃការបែងចែកកោសិកាជាបន្តបន្ទាប់ ដែលហៅថាជាសមូហភាព។ការបង្កើតមេជីវិតឈ្មោល,បន្តដោយដំណើរការខុសគ្នាស្មុគ្រស្មាញដែលហៅថា spermiogenesis (រូបភាព 20.31 ) ដំណើរការនៃការបង្កើតមេជីវិតឈ្មោលត្រូវចំណាយពេលប្រហែល 70 ថ្ងៃ; ក្នុង 1 ក្រាមនៃទំងន់ពងស្វាស 10 ត្រូវបានបង្កើតឡើង 7 មេជីវិតឈ្មោលក្នុងមួយថ្ងៃ។ epithelium នៃ tubule seminiferous មានស្រទាប់ខាងក្រៅនៃកោសិកាepithelium មេរោគនិងប្រហែលប្រាំមួយស្រទាប់នៃកោសិកាបានបង្កើតឡើងជាលទ្ធផលនៃការបែងចែកកោសិកាម្តងហើយម្តងទៀតនៃស្រទាប់នេះ (រូបភាព 20.32 និង 20.33); ស្រទាប់ទាំងនេះត្រូវគ្នាទៅនឹងដំណាក់កាលបន្តបន្ទាប់នៃការអភិវឌ្ឍន៍មេជីវិតឈ្មោល។ ទីមួយ ការបែងចែកកោសិការ epithelial germinal ផ្តល់នូវការកើនឡើងជាច្រើន។ spermatogonia ដែលបង្កើនទំហំនិងក្លាយជាលំដាប់ដំបូង spermatocytes ។ជាលទ្ធផលនៃការបែងចែក meiotic ដំបូង spermatocytes ទាំងនេះបង្កើតបានជា haploidលំដាប់ទីពីរ spermatocytes,បន្ទាប់ពីនោះពួកគេឆ្លងកាត់ការបែងចែក meiotic ទីពីរហើយប្រែទៅជា spermatids ។ រវាង "strands" នៃកោសិកាដែលកំពុងអភិវឌ្ឍមានធំកោសិកា Sertoli ឬ កោសិកា trophic,មានទីតាំងនៅទូទាំងចន្លោះទាំងមូលពីស្រទាប់ខាងក្រៅនៃ tubule ទៅ lumen របស់វា។

Spermatocytes មានទីតាំងនៅក្នុង invaginations ជាច្រើននៅលើផ្ទៃក្រោយនៃកោសិកា Sertoli; នៅទីនេះពួកវាប្រែទៅជា spermatids ហើយបន្ទាប់មកផ្លាស់ទីទៅគែមនៃកោសិកា Sertoli ដែលប្រឈមមុខនឹង lumen នៃ tubule seminiferous ដែលពួកគេមានភាពចាស់ទុំបង្កើតជាមេជីវិតឈ្មោល (រូបភាព 20.33 ) ជាក់ស្តែង កោសិកា Sertoli ផ្តល់ជំនួយមេកានិច ការការពារ និងអាហារូបត្ថម្ភដល់មេជីវិតឈ្មោលដែលចាស់ទុំ។ សារធាតុចិញ្ចឹម និងអុកស៊ីហ៊្សែនទាំងអស់ដែលត្រូវបានបញ្ជូនទៅ gametes ដែលកំពុងលូតលាស់តាមរយៈសរសៃឈាមជុំវិញបំពង់ seminiferous និងកាកសំណល់រំលាយអាហារដែលបញ្ចេញទៅក្នុងឈាមឆ្លងកាត់កោសិកា Sertoli ។ កោសិកាទាំងនេះក៏បញ្ចេញសារធាតុរាវដែលមេជីវិតឈ្មោលឆ្លងកាត់បំពង់។
ការវិវត្តនៃស៊ុតនៅក្នុងមនុស្ស (oogenesis) ។មិនដូចការបង្កើតមេជីវិតឈ្មោលដែលចាប់ផ្តើមលើបុរសតែនៅពេញវ័យទេ ការបង្កើតពងចំពោះស្ត្រីចាប់ផ្តើមសូម្បីតែមុនពេលកើត និងត្រូវបានបញ្ចប់សម្រាប់ស៊ុតនីមួយៗដែលបានផ្តល់ឲ្យតែបន្ទាប់ពីការបង្កកំណើតរបស់វា។ ដំណាក់កាលនៃ oogenesis ត្រូវបានបង្ហាញនៅក្នុងរូបភព។ 20.36. ក្នុងអំឡុងពេលនៃការលូតលាស់គភ៌ កោសិកាមេជីវិតដំបូងបានបែងចែកម្តងហើយម្តងទៀតតាមរយៈ mitosis បង្កើតកោសិកាធំៗជាច្រើនដែលហៅថាអូហ្គោនី។ Oogonia ម្តងទៀតឆ្លងកាត់ mitosis និងទម្រង់ការបញ្ជាទិញដំបូង oocygs,ដែលនៅតែមាននៅដំណាក់កាល prophase ស្ទើរតែរហូតដល់ការបញ្ចេញពងអូវុល។ oocytes លំដាប់ទីមួយត្រូវបានហ៊ុំព័ទ្ធដោយស្រទាប់តែមួយនៃកោសិកា- ភ្នាស granulosa- និងបង្កើតអ្វីដែលគេហៅថាឫសគល់បឋម។ទារកស្ត្រីភ្លាមៗមុនពេលសម្រាលមានប្រហែល 2 10 6 follicles ទាំងនេះ ប៉ុន្តែមានតែប្រហែល 450 ប៉ុណ្ណោះក្នុងចំណោមពួកវាឈានដល់ដំណាក់កាលនៃ oocytes លំដាប់ទីពីរហើយចាកចេញពី ovary (ovulation) ។ មុនពេលបញ្ចេញពងអូវុល oocyte លំដាប់ទីមួយឆ្លងកាត់ការបែងចែក meiotic ដំបូងបង្កើតជា haploidលំដាប់ទីពីរ oocyteនិង រាងកាយប៉ូលដំបូង។ការបែងចែក meiotic ទីពីរឈានដល់ដំណាក់កាល metaphase ប៉ុន្តែមិនបន្តរហូតដល់ oocyte ប្រសព្វជាមួយមេជីវិតឈ្មោល។ ក្នុងអំឡុងពេលនៃការបង្កកំណើត oocyte លំដាប់ទីពីរឆ្លងកាត់ការបែងចែក meiotic ទីពីរបង្កើតជាកោសិកាដ៏ធំមួយ -ស៊ុតនិងផងដែរ។ រាងប៉ូលទីពីរ។សាកសពប៉ូលទាំងអស់គឺជាកោសិកាតូចៗ។ ពួកវាមិនមានតួនាទីនៅក្នុង oogenesis ហើយត្រូវបានបំផ្លាញជាយថាហេតុ។

parthenogenesis
កាតព្វកិច្ច
ស្រេចចិត្ត
ការអភិវឌ្ឍនៃសារពាង្គកាយបានតែពីស៊ុតដែលមិនបានបង្កកំណើត។
មានតែស្ត្រីទេដែលត្រូវបានបង្កើតឡើង។
ឧទហរណ៍ ជីងចក់ថ្ម Caucasian
letlets អាចអភិវឌ្ឍបានទាំងដោយគ្មានការបង្កកំណើត និងជាមួយនឹងការបង្កកំណើត
ប្រភេទស្រី
បុរសកើតចេញពីពងមិនទាន់បង្កកំណើត
ប្រភេទបុរស
នៅក្នុងសារាយ isogamous មួយចំនួន
ការងារស្រដៀងគ្នាផ្សេងទៀតដែលអាចចាប់អារម្មណ៍ you.vshm> |
|||
| 6644. | ការអភិវឌ្ឍបុគ្គលនៃសារពាង្គកាយ (ontogenesis) | 78.69 គីឡូបៃ | |
| វាត្រូវបានគេជឿថាមាតិកានុយក្លេអ៊ែរនៃមេជីវិតឈ្មោលតែមួយជ្រាបចូលទៅក្នុងស៊ុតរបស់សត្វ។ ក្នុងករណីមនុស្ស និងជួនកាលសត្វខ្ពស់ៗ រយៈពេលនៃការអភិវឌ្ឍន៍មុនពេលកើត ជារឿយៗត្រូវបានគេហៅថាមុនពេលសម្រាល និងក្រោយពេលសម្រាល។ នៅក្នុងសត្វពហុកោសិកា ដោយមិនគិតពីភាពស្មុគស្មាញនៃអង្គការរបស់ពួកគេ ដំណាក់កាលនៃការអភិវឌ្ឍន៍អំប្រ៊ីយ៉ុងដែលអំប្រ៊ីយ៉ុងឆ្លងកាត់គឺដូចគ្នា។ ធម្មជាតិនៃការបុក និងប្រភេទនៃ blastulas នៅក្នុងសត្វឆ្អឹងខ្នងផ្សេងៗគ្នា... | |||
| 13714. | ការអភិវឌ្ឍន៍បុគ្គលនៃសារពាង្គកាយ និងអាកប្បកិរិយារបស់វា។ Ontogenesis ។ វដ្តជីវិតនៅក្នុងរុក្ខជាតិនិងសត្វ | 9.96 គីឡូបៃ | |
| ប្រភពដើមនៃបុគ្គលិកលក្ខណៈ Ontogenesis គឺជាការវិវឌ្ឍន៍បុគ្គលនៃសារពាង្គកាយមួយ ចាប់ពីពេលនៃការបង្កើតហ្សីហ្គោត រហូតដល់ការស្លាប់របស់វា។ Gametes គឺជាកោសិកាផ្លូវភេទដែលផ្ទុកព័ត៌មានតំណពូជ និងមានសំណុំ haploid ។ ការបង្កកំណើត គឺជាដំណើរការនៃការលាយបញ្ចូលគ្នានៃកោសិកាបន្តពូជបុរស និងស្ត្រីនៃរុក្ខជាតិ ឬសត្វ ហើយជាមូលដ្ឋាននៃដំណើរការផ្លូវភេទ។ រយៈពេលដំណុះនៃអំប្រ៊ីយ៉ុង គឺជារយៈពេលនៃ ontogenesis ពីការបង្កើត zygote រហូតដល់កំណើត ឬកើតចេញពីភ្នាសស៊ុត ឬដំណុះ។ | |||
| 10427. | ការបន្តពូជ | 6.75 គីឡូបៃ | |
| ជាមួយនឹងទម្រង់ជាច្រើននៃការបន្តពូជរបស់សារពាង្គកាយ ពួកវាទាំងអស់អាចត្រូវបានកាត់បន្ថយទៅជាពីរប្រភេទសំខាន់ៗ៖ ភេទ និងផ្លូវភេទ។ នៅក្នុងការបន្តពូជដោយភេទដូចគ្នា ការបន្តពូជនៃកូនចៅកើតឡើងពីមេមួយតាមរយៈការបង្កើត spores ឬលូតលាស់។ ក្នុងអំឡុងពេលបន្តពូជលូតលាស់ កូនចៅកើតចេញពីផ្នែករាងកាយដែលបំបែកចេញពីម្តាយ។ ក្នុងអំឡុងពេលបន្តពូជលូតលាស់ រុក្ខជាតិរក្សា heterozygosity ជាច្រើនជំនាន់។ | |||
| 21332. | កត្តាកំណត់។ ការសម្របសម្រួលនៃសារពាង្គកាយទៅនឹងកត្តា | 303.8 គីឡូបៃ | |
| សត្វខ្លះចូលចិត្តកំដៅខ្លាំង ខ្លះទៀតទ្រាំនឹងសីតុណ្ហភាពបរិស្ថានកម្រិតមធ្យមបានប្រសើរជាង។ ប្រសិនបើឥទ្ធិពលនៃលក្ខខណ្ឌបរិស្ថានមិនឈានដល់តម្លៃខ្លាំងទេ សារពាង្គកាយមានជីវិតមានប្រតិកម្មចំពោះវាជាមួយនឹងសកម្មភាពជាក់លាក់ ឬការផ្លាស់ប្តូរនៅក្នុងស្ថានភាពរបស់ពួកគេ ដែលនៅទីបំផុតនាំទៅរកការរស់រានមានជីវិតនៃប្រភេទសត្វ។ កម្មវត្ថុនៃការសិក្សាក្នុងការងារនេះគឺកត្តាបរិស្ថាន ប្រធានបទគឺការកំណត់កត្តា និងការសម្របខ្លួនរបស់សារពាង្គកាយទៅ... | |||
| 8875. | គំរូទូទៅនៃអន្តរកម្មរវាងសារពាង្គកាយ និងកត្តាបរិស្ថាន | 193.58 គីឡូបៃ | |
| កត្តាជីវសាស្ត្រនៃបរិស្ថានលើដី និងទឹកនៃដី សារធាតុសកម្មជីវសាស្រ្តនៃសារពាង្គកាយមានជីវិត កត្តាមនុស្សភពផែនដី គំរូទូទៅនៃអន្តរកម្មរវាងសារពាង្គកាយ និងកត្តាបរិស្ថាន គំនិតនៃកត្តាកំណត់មួយ។ ច្បាប់អប្បបរមារបស់ Liebig ច្បាប់ Shelford ជាក់លាក់នៃផលប៉ះពាល់នៃកត្តា anthropogenic លើសារពាង្គកាយ ចំណាត់ថ្នាក់នៃសារពាង្គកាយទាក់ទងនឹងកត្តាបរិស្ថាន 1. លក្ខខណ្ឌនៃស្មៅស្មៅ feather steppes តំណាងឱ្យរបបខុសគ្នាទាំងស្រុងនៃកត្តា abiotic ។ | |||
| 12700. | លក្ខណៈជីវសាស្រ្តនៃសត្វល្អិត និងវិធានការដើម្បីកំចាត់ពួកវា | 62.79 គីឡូបៃ | |
| ការបាត់បង់ដំណាំគួរឱ្យកត់សម្គាល់ជាពិសេសកើតឡើងជាលទ្ធផលនៃវត្តមានស្មៅដែលយកសារធាតុចិញ្ចឹម និងសំណើមចេញពីដី រុក្ខជាតិដាំដុះដែលមានម្លប់ ហើយក្នុងករណីជាច្រើនបំពុលផលិតផលជាមួយសារធាតុពុល និងគ្រាប់ពូជដែលបណ្តាលឱ្យពុលមនុស្ស និងសត្វ។ ទិសដៅសំខាន់នៃគីមីវិទ្យានៃកសិកម្ម៖ ការប្រើប្រាស់ជីគីមី ការការពាររុក្ខជាតិពីសត្វល្អិត ជំងឺ និងស្មៅ ការប្រើប្រាស់ផលិតផលគីមីក្នុងការចិញ្ចឹមសត្វ ផលិតផលកសិកម្មកំប៉ុង និង... | |||
| 13403. | មេរោគជាទម្រង់ជីវិតមិនមែនកោសិកា។ រចនាសម្ព័ន្ធ ការចាត់ថ្នាក់ អន្តរកម្មនៃមេរោគជាមួយកោសិកានៃសារពាង្គកាយផ្សេងៗ | 12.75 គីឡូបៃ | |
| ទាំងនេះគឺជាវត្ថុជីវសាស្រ្ត ហ្សែនដែលផ្សំឡើងដោយអាស៊ីត nucleic DNA ឬ RNA ដែលផលិតនៅក្នុងកោសិការស់នៅដោយប្រើឧបករណ៍ជីវសំយោគរបស់វា។ ភាពខុសគ្នារវាងមេរោគ និងទម្រង់ជីវិតផ្សេងទៀត៖ ពួកវាមិនមានរចនាសម្ព័ន្ធកោសិកា អាស៊ីតនុយក្លេអ៊ីក 1 ប្រភេទ មានតែ DNA ឬ RNA ប៉ុណ្ណោះ ពួកគេមិនមានការរំលាយអាហារផ្ទាល់ខ្លួនទេ។ សម្មតិកម្មនៃប្រភពដើមនៃមេរោគ៖ មេរោគបានកើតចេញពីសមាសធាតុនៃកោសិកាធម្មតាដែលគេចផុតពីការគ្រប់គ្រងនៃយន្តការនិយតកម្ម ហើយប្រែទៅជាអង្គភាពឯករាជ្យមួយ ប្រហែលជាការផ្លាស់ប្តូរហ្សែនជាច្រើនបានកើតឡើងនៅលើផ្នែកមួយនៃ DNA ដែល... | |||
| 18798. | សូចនាកររចនាសម្ព័ន្ធ និងមុខងារនៃសារពាង្គកាយ mesofauna ដីនៅក្នុងព្រៃ និង biotopes បើកចំហនៃជ្រលងទន្លេ Pozim | 61.54 គីឡូបៃ | |
| ទោះជាយ៉ាងណាក៏ដោយសព្វថ្ងៃនេះការអភិវឌ្ឍន៍នៃទឹកដីនេះគឺជាផលវិបាកនៃបញ្ហាបរិស្ថានមួយចំនួន។ ជាពិសេស ការងារកំពុងត្រូវបានអនុវត្តយ៉ាងទូលំទូលាយនៅតាមជ្រលងភ្នំ ដើម្បីបង្ហូរដីវាលទំនាបលិចទឹក ដែលជះឥទ្ធិពលដោយផ្ទាល់ទៅលើជីវតូបធម្មជាតិ។ | |||
| 19386. | ការអភិវឌ្ឍនៃសេរីនិយមនៅក្នុង CIS | 35.55 គីឡូបៃ | |
| លក្ខណៈប្រព័ន្ធស្មុគ្រស្មាញនៃការផ្លាស់ប្តូរដែលកំពុងពិចារណានៅក្នុងដំណើរការនៃការវិវត្តនៃទំនាក់ទំនងសេដ្ឋកិច្ចបរទេសកំណត់ការពិនិត្យឡើងវិញ និងការផ្លាស់ប្តូរនៃ axiomatics ទាំងមូលនៃគំនិតប្រពៃណីអំពីការកំណត់អត្តសញ្ញាណ ព្រំដែន និងអធិបតេយ្យភាពនៃសេដ្ឋកិច្ចជាតិ ... | |||
| 8867. | ការអប់រំ និងការអភិវឌ្ឍន៍ | 162.32 គីឡូបៃ | |
| ខ្លឹមសារនៃបញ្ហានៃទំនាក់ទំនងរវាងការបណ្តុះបណ្តាលនិងការអភិវឌ្ឍន៍។ វិធីសាស្រ្តជាមូលដ្ឋានក្នុងការដោះស្រាយបញ្ហានៃទំនាក់ទំនងរវាងការបណ្តុះបណ្តាលនិងការអភិវឌ្ឍន៍។ គំនិតនៃតំបន់នៃការអភិវឌ្ឍន៍ជិតៗ L. ខ្លឹមសារនៃបញ្ហានៃទំនាក់ទំនងរវាងការបណ្តុះបណ្តាល និងការអភិវឌ្ឍន៍។ | |||
ការបន្តពូជគឺជាកម្មសិទ្ធិរបស់សារពាង្គកាយដើម្បីបង្កើតកូនចៅ ឬសមត្ថភាពរបស់សារពាង្គកាយក្នុងការបន្តពូជខ្លួនឯង។ ក្នុងនាមជាទ្រព្យសម្បត្តិដ៏សំខាន់បំផុតនៃភាវៈរស់ ការបន្តពូជធានានូវភាពបន្តនៃជីវិត និងការបន្តនៃប្រភេទសត្វ។
ដំណើរការនៃការបន្តពូជគឺស្មុគស្មាញខ្លាំងណាស់ ហើយត្រូវបានផ្សារភ្ជាប់មិនត្រឹមតែជាមួយនឹងការផ្ទេរព័ត៌មានហ្សែនពីឪពុកម្តាយទៅកូនចៅប៉ុណ្ណោះទេ ប៉ុន្តែថែមទាំងជាមួយនឹងលក្ខណៈសម្បត្តិកាយវិភាគវិទ្យា និងសរីរវិទ្យានៃសារពាង្គកាយ ជាមួយនឹងអាកប្បកិរិយា និងការគ្រប់គ្រងអ័រម៉ូន។ ការបន្តពូជរបស់សារពាង្គកាយត្រូវបានអមដោយដំណើរការនៃការលូតលាស់ និងការអភិវឌ្ឍន៍របស់វា។
សត្វមានជីវិតត្រូវបានកំណត់លក្ខណៈដោយភាពចម្រុះយ៉ាងខ្លាំងនៅក្នុងវិធីសាស្រ្តនៃការបន្តពូជរបស់ពួកគេ។ ទោះយ៉ាងណាក៏ដោយ មានវិធីសាស្រ្តសំខាន់ពីរនៃការបន្តពូជ - ផ្លូវភេទ និងផ្លូវភេទ (រូបភាព 16) ។ ការបន្តពូជដោយភេទ ឬ apomixis (មកពីភាសាក្រិច។ អារ៉ូ -ដោយគ្មាន ល្បាយ -ការលាយបញ្ចូលគ្នា) គឺជាដំណើរការមួយដែលមានមេតែមួយ (កោសិកា ឬសារពាង្គកាយពហុកោសិកា) ចូលរួម។ ផ្ទុយទៅវិញ ការបន្តពូជផ្លូវភេទពាក់ព័ន្ធនឹងឪពុកម្តាយពីរនាក់ ដែលកូននីមួយៗមានប្រព័ន្ធបន្តពូជផ្ទាល់ខ្លួន និងបង្កើតកោសិកាផ្លូវភេទ (gametes) ដែលបន្ទាប់ពីការលាយបញ្ចូលគ្នា បង្កើតជាហ្សីហ្គោត (ស៊ុតបង្កកំណើត) ដែលបន្ទាប់មកមានភាពខុសគ្នាទៅជាអំប្រ៊ីយ៉ុង។ អាស្រ័យហេតុនេះ ក្នុងអំឡុងពេលបន្តពូជផ្លូវភេទ មានការលាយបញ្ចូលគ្នានៃកត្តាតំណពូជ ពោលគឺដំណើរការមួយហៅថា amphimixis (មកពីភាសាក្រិច។ អំពែ -នៅភាគីទាំងពីរ, ល្បាយ -លាយ) ។
ការបន្តពូជដោយភេទ
ការបន្តពូជដោយភេទជាលក្ខណៈនៃសារពាង្គកាយជាច្រើនប្រភេទ ទាំងរុក្ខជាតិ និងសត្វ។ វាត្រូវបានរកឃើញនៅក្នុងមេរោគ បាក់តេរី សារាយ ផ្សិត រុក្ខជាតិសរសៃឈាម ប្រូតូហ្សូ អេប៉ុង coelenterates bryozoans និង tunicates ។
ទម្រង់ដ៏សាមញ្ញបំផុតនៃការបន្តពូជផ្លូវភេទគឺជាលក្ខណៈនៃមេរោគ។ ដំណើរការបន្តពូជរបស់ពួកគេត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងម៉ូលេគុលអាស៊ីត nucleic ជាមួយនឹងសមត្ថភាពនៃម៉ូលេគុលទាំងនេះក្នុងការចម្លងដោយខ្លួនឯង
និងផ្អែកលើភាពជាក់លាក់នៃចំណងអ៊ីដ្រូសែនខ្សោយរវាងនុយក្លេអូទីត។
អង្ករ។ ១៦.វិធីសាស្រ្តនៃការបន្តពូជនៃសារពាង្គកាយ
ទាក់ទងទៅនឹងសារពាង្គកាយផ្សេងទៀតដែលបន្តពូជដោយភេទ ភាពខុសគ្នាមួយត្រូវបានបង្កើតឡើងរវាងការបន្តពូជលូតលាស់ និងការបន្តពូជដោយ sporulation ។
ការបន្តពូជរបស់សារពាង្គកាយគឺជាការបន្តពូជដែលសារពាង្គកាយថ្មីមួយវិវឌ្ឍន៍ចេញពីផ្នែកដែលបំបែកចេញពីសារពាង្គកាយមាតា។ ប្រភេទនៃការបន្តពូជនេះគឺជាលក្ខណៈនៃសារពាង្គកាយ unicellular និង multicellular ប៉ុន្តែមានការបង្ហាញខុសៗគ្នានៅក្នុងពួកវា។
នៅក្នុងសារពាង្គកាយឯកតា ការបន្តពូជលូតលាស់ត្រូវបានតំណាងដោយទម្រង់ដូចជាការបែងចែក ការបំបែកច្រើន និងការពន្លក។ ការបែងចែកដោយការបង្រួមសាមញ្ញជាមួយនឹងការបង្កើតសារពាង្គកាយកូនស្រីពីរពីសារពាង្គកាយមេតែមួយគឺជាលក្ខណៈនៃបាក់តេរីនិងសារាយខៀវបៃតង (cyanobacteria) ។ ផ្ទុយទៅវិញ ការបន្តពូជដោយការបែងចែកនៃសារាយពណ៌ត្នោត និងបៃតង ក៏ដូចជាសត្វឯកកោ (sarcodes, flagellates និង ciliates) កើតឡើងតាមរយៈការបែងចែក mitotic នៃស្នូលដែលបន្តដោយការបង្រួមនៃ cytoplasm ។
ការបន្តពូជដោយការប្រេះស្រាំច្រើន (schizogony) ពាក់ព័ន្ធនឹងការបែងចែកស្នូល បន្តដោយការបែងចែក cytoplasm ទៅជាផ្នែក។ ជាលទ្ធផលនៃការបែងចែកនេះ សារពាង្គកាយកូនស្រីជាច្រើនត្រូវបានបង្កើតឡើងពីកោសិកាមួយ។ ឧទាហរណ៏នៃការបែងចែកច្រើនគឺការបន្តពូជនៃ falciparum plasmodium (P. vivax)នៅក្នុង erythrocytes របស់មនុស្ស។ ក្នុងករណីនេះនៅក្នុង plasmodia ការបែងចែកនុយក្លេអ៊ែរម្តងហើយម្តងទៀតកើតឡើងច្រើនដងដោយគ្មាន cytokinesis បន្ទាប់មក cytokinesis ។ ជាលទ្ធផល plasmodium មួយផ្តល់នូវសារពាង្គកាយកូនស្រី 12-24 ។
នៅក្នុងសារពាង្គកាយរុក្ខជាតិពហុកោសិកា ការបន្តពូជលូតលាស់ដោយការបែងចែកត្រូវបានអនុវត្តដោយការកាត់ អំពូល ស្លឹក និងមើម។ ប៉ុន្តែនេះជាការបន្តពូជសិប្បនិម្មិតយ៉ាងសំខាន់ដែលប្រើក្នុងការអនុវត្តកសិកម្ម។ ការបន្តពូជនៃរុក្ខជាតិខ្ពស់ជាងក្រោមលក្ខខណ្ឌសិប្បនិម្មិតក៏អាចធ្វើទៅបានផងដែរពីកោសិកាតែមួយ។ សារពាង្គកាយដែលវិវឌ្ឍន៍ចេញពីកោសិកាតែមួយ (ក្លូន) មានលក្ខណៈសម្បត្តិទាំងអស់នៃសារពាង្គកាយពហុកោសិកាដើម។ ការបន្តពូជនេះត្រូវបានគេហៅថា clonal micropropagation ។ ទម្រង់មួយនៃការបន្តពូជលូតលាស់អាចជាការផ្សាំ ឬស្ទូងរុក្ខជាតិដាំដុះជាច្រើន ដែលមានការស្ទូងពន្លក ឬផ្នែកនៃពន្លកពីដើមមួយទៅរុក្ខជាតិមួយទៀត។ ជាការពិតណាស់នេះក៏ជាវិធីសាស្រ្តនៃការបន្តពូជផងដែរ ដែលមិនកើតឡើងនៅក្នុងធម្មជាតិ ប៉ុន្តែត្រូវបានប្រើប្រាស់យ៉ាងទូលំទូលាយក្នុងវិស័យកសិកម្ម។
នៅក្នុងសត្វពហុកោសិកា ការបន្តពូជលូតលាស់កើតឡើងដោយការបំបែករាងកាយរបស់ពួកគេទៅជាផ្នែកៗ ហើយបន្ទាប់មកនីមួយៗ
ផ្នែកមួយវិវត្តទៅជាសត្វថ្មី។ ការបន្តពូជបែបនេះគឺជាតួយ៉ាងសម្រាប់អេប៉ុង, coelenterates (hydras), nemerteans, flatworms, echinoderms (ត្រីផ្កាយ) និងសារពាង្គកាយមួយចំនួនទៀត។ ទម្រង់ជិតស្និទ្ធនៃការបែកខ្ញែកទៅនឹងការបន្តពូជលូតលាស់របស់សត្វគឺពហុវប្បធម៍របស់សត្វដែលមាននៅក្នុងការពិតដែលថានៅដំណាក់កាលជាក់លាក់នៃការអភិវឌ្ឍន៍អំប្រ៊ីយ៉ុងត្រូវបានបែងចែកជាផ្នែកជាច្រើនដែលនីមួយៗវិវត្តទៅជាសារពាង្គកាយឯករាជ្យ។ Polyembryony កើតឡើងនៅក្នុង armadillos ។ ទោះយ៉ាងណាក៏ដោយ ក្រោយមកទៀតបន្តពូជផ្លូវភេទ។ ដូច្នេះហើយ អំប្រ៊ីយ៉ុងគឺជាដំណាក់កាលពិសេសមួយក្នុងការបន្តពូជផ្លូវភេទ ហើយកូនចៅដែលកើតចេញពីពហុស្បូនត្រូវបានតំណាងដោយកូនភ្លោះ monozygotic ។
ការចេញផ្កាមានការបង្កើតមើម (ការរីកដុះដាល) ជាមួយនឹងស្នូលនៅលើកោសិកាមេ ដែលបន្ទាប់មកបំបែក និងក្លាយជាសារពាង្គកាយឯករាជ្យ។ ការដុះពន្លកកើតឡើងទាំងនៅក្នុងរុក្ខជាតិ unicellular ឧទាហរណ៍នៅក្នុង yeast និងនៅក្នុងសត្វ unicellular ឧទាហរណ៍នៅក្នុងប្រភេទមួយចំនួននៃ ciliates ។
ការបន្តពូជដោយ sporulation ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការបង្កើតកោសិកាឯកទេស - spores ដែលមានស្នូល cytoplasm ត្រូវបានគ្របដណ្ដប់ដោយភ្នាសក្រាស់ និងមានសមត្ថភាពអាចរស់នៅបានយូរក្នុងស្ថានភាពមិនអំណោយផល ដែលលើសពីនេះទៀត រួមចំណែកដល់ការបែកខ្ញែករបស់វា។ ភាគច្រើនជាញឹកញាប់ ការបន្តពូជបែបនេះកើតឡើងនៅក្នុងបាក់តេរី សារាយ ផ្សិត ស្លែ និង ferns ។ នៅក្នុងសារាយបៃតងខ្លះ អ្វីដែលគេហៅថា zoospores អាចបង្កើតចេញពីកោសិកានីមួយៗ។
ក្នុងចំណោមសត្វ ការបន្តពូជដោយ sporulation ត្រូវបានគេសង្កេតឃើញនៅក្នុង sporozoans ជាពិសេសនៅក្នុង falciparum plasmodium ។
នៅក្នុងសារពាង្គកាយនៃប្រភេទសត្វជាច្រើន ការបន្តពូជដោយភេទអាចជំនួសដោយការបន្តពូជផ្លូវភេទ។
ការផលិតផ្លូវភេទ
ការបន្តពូជផ្លូវភេទកើតឡើងទាំងនៅក្នុងរុក្ខជាតិ និងសត្វ unicellular និង multicellular ។
ដូចដែលបានកត់សម្គាល់នៅក្នុងជំពូកទី V និង XIII ការបន្តពូជផ្លូវភេទនៅក្នុងបាក់តេរីត្រូវបានអនុវត្តដោយការភ្ជាប់គ្នាដែលបម្រើជា analogue នៃដំណើរការផ្លូវភេទ និងជាប្រព័ន្ធនៃការផ្សំឡើងវិញនៃសារពាង្គកាយទាំងនេះ ខណៈពេលដែលនៅក្នុងប្រូហ្សូអា ការបន្តពូជផ្លូវភេទក៏កើតឡើងដោយការភ្ជាប់គ្នា ឬតាមរយៈ syngamy ។ និង autogamy ។
នៅក្នុងសារពាង្គកាយពហុកោសិកា (រុក្ខជាតិ និងសត្វ) ការបន្តពូជផ្លូវភេទត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការបង្កើតមេរោគ ឬកោសិកាផ្លូវភេទ (gametes) ការបង្កកំណើត និងការបង្កើតហ្សីហ្គោត។
ការបន្តពូជផ្លូវភេទគឺជាការទទួលការវិវត្តន៍ដ៏សំខាន់នៃសារពាង្គកាយ។ ម្យ៉ាងវិញទៀត វារួមចំណែកដល់ការចាត់ថ្នាក់នៃហ្សែន ការកើតនៃភាពចម្រុះនៃសារពាង្គកាយ និងការបង្កើនការប្រកួតប្រជែងរបស់ពួកគេក្នុងការផ្លាស់ប្តូរជាបន្តបន្ទាប់នៃលក្ខខណ្ឌបរិស្ថាន។
នៅក្នុងសារពាង្គកាយកោសិកាតែមួយ ការបន្តពូជផ្លូវភេទកើតឡើងក្នុងទម្រង់ជាច្រើន។ នៅក្នុងបាក់តេរី ការបន្តពូជផ្លូវភេទអាចត្រូវបានធ្វើអាណាឡូកជាមួយនឹងការភ្ជាប់គ្នាដែលកើតឡើងនៅក្នុងពួកវា ដែលមាននៅក្នុងការផ្ទេរ plasmid ឬ chromosomal DNA ពីកោសិកាអ្នកបរិច្ចាគ (ដែលមានផ្ទុក plasmids) ទៅកាន់កោសិកាអ្នកទទួល (មិនមានផ្ទុក plasmids) ក៏ដូចជាជាមួយនឹងការឆ្លងបាក់តេរី។ ដែលមាននៅក្នុងការផ្ទេរសម្ភារៈហ្សែនពីកោសិកាបាក់តេរីមួយចំនួនទៅ phages ផ្សេងទៀត។ ការភ្ជាប់គ្នាត្រូវបានរកឃើញផងដែរនៅក្នុង ciliates ដែលក្នុងនោះក្នុងអំឡុងពេលដំណើរការនេះ ការផ្ទេរស្នូលកើតឡើងពីបុគ្គលមួយទៅមនុស្សម្នាក់ទៀត បន្ទាប់មកដោយការបែងចែកក្រោយ។
នៅក្នុងរុក្ខជាតិ និងសត្វពហុកោសិកា ការបន្តពូជផ្លូវភេទនៃកោសិកាកើតឡើងតាមរយៈការបង្កើតកោសិកាមេជីវិតឈ្មោល (ស៊ុត និងមេជីវិតឈ្មោល) ការបង្កកំណើតជាបន្តបន្ទាប់នៃស៊ុតដោយមេជីវិតឈ្មោល និងការបង្កើតហ្សីហ្គោត។ នៅក្នុងរុក្ខជាតិ កោសិកាផ្លូវភេទត្រូវបានផលិតនៅក្នុងរចនាសម្ព័ន្ធបន្តពូជឯកទេស ហើយនៅក្នុងសត្វ ពួកវាត្រូវបានផលិតជា gonads ដែលហៅថា gonads (មកពីភាសាក្រិច។ បានទៅ -គ្រាប់ពូជ) ។
មានភាពខុសគ្នាដ៏សំខាន់រវាងកោសិកា somatic និងបន្តពូជរបស់សត្វ។ វាស្ថិតនៅក្នុងការពិតដែលថាកោសិកា somatic មានសមត្ថភាពបែងចែក ពោលគឺពួកវាបង្កើតឡើងវិញដោយខ្លួនឯង ហើយលើសពីនេះទៀត កោសិកាមេរោគត្រូវបានបង្កើតឡើងពីពួកវា។ ផ្ទុយទៅវិញ កោសិកាផ្លូវភេទមិនបែងចែកទេ ប៉ុន្តែពួកគេ "ចាប់ផ្តើម" ការបន្តពូជនៃសារពាង្គកាយទាំងមូល។
កោសិកា somatic Diploid ដែលកោសិកាមេជីវិតឈ្មោលត្រូវបានបង្កើតឡើងត្រូវបានគេហៅថា spermatogonia ហើយនៅក្នុងនោះកោសិកាមេជីវិតឈ្មោលត្រូវបានបង្កើតឡើង - oogonia ។ ដំណើរការនៃការបង្កើត (ការលូតលាស់ និងភាពខុសគ្នា) នៃកោសិកាមេជីវិតឈ្មោល និងស្ត្រីត្រូវបានគេហៅថា gametogenesis ។
Gametogenesis គឺផ្អែកលើ meiosis (មកពីភាសាក្រិក។ meiosis -កាត់បន្ថយ) ដែលជាការបែងចែកកាត់បន្ថយនៃស្នូលកោសិកា អមដោយការថយចុះនៃចំនួនក្រូម៉ូសូមក្នុងមួយស្នូល។ Meiosis កើតឡើងនៅក្នុងកោសិកាឯកទេសនៃសរីរាង្គបន្តពូជរបស់សត្វមានជីវិតដែលបន្តពូជផ្លូវភេទ (រូបភាព 17) ។ ឧទាហរណ៍នៅក្នុង pteriophytes, meiosis កើតឡើងនៅក្នុងកោសិកា sporangial ឯកទេសដែលមានទីតាំងនៅលើផ្ទៃខាងក្រោមនៃស្លឹករបស់រុក្ខជាតិទាំងនេះហើយវិវត្តទៅជា spores ហើយបន្ទាប់មកចូលទៅក្នុង gametophytes ។ ក្រោយមកទៀតមានដោយឡែកពីគ្នា ទីបំផុតផលិតមេអំបៅប្រុស និងស្រី។ នៅក្នុងរុក្ខជាតិផ្កា meiosis កើតឡើងនៅក្នុងកោសិកាឯកទេសនៃអូវុលដែលវិវត្តទៅជា spores ។ ក្រោយមកទៀតបង្កើត gametophyte ជាមួយស៊ុតមួយ។ លើសពីនេះទៀតនៅក្នុងរុក្ខជាតិទាំងនេះ meiosis ក៏កើតឡើងនៅក្នុងកោសិកា anther ឯកទេសផងដែរដែលវិវត្តទៅជា spores ដែលទីបំផុតបង្កើត pollen ជាមួយ gametes ឈ្មោលពីរ។ នៅក្នុងដង្កូវនាង ដែលជាពពួក hermaphrodites និងមានសរីរាង្គបន្តពូជរបស់បុរសនៅក្នុងផ្នែកមួយនៃរាងកាយ និងស្ត្រីនៅក្នុងផ្នែកមួយទៀត ហើយដែលត្រូវបានកំណត់លក្ខណៈដោយ
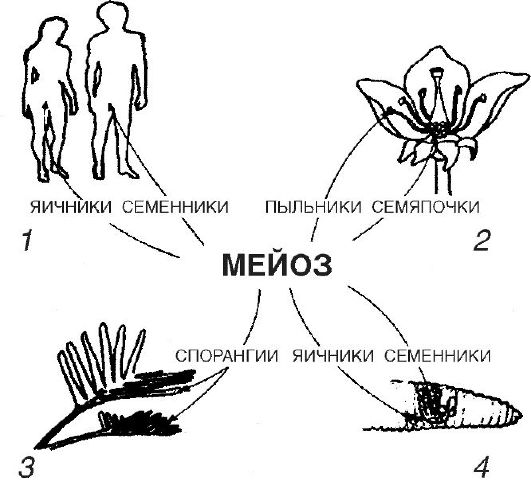
អង្ករ។ ១៧. Meiosis នៅក្នុងសារពាង្គកាយផ្សេងៗគ្នា៖ 1 - មនុស្ស; 2 - រុក្ខជាតិផ្កា; 3 - fern; 4 - ដង្កូវនាង
ពួកគេមានសមត្ថភាពក្នុងការបង្កកំណើតរវាងបុគ្គលផ្សេងៗគ្នា ពួកគេមានសមត្ថភាពធ្វើ spermatogenesis និង oogenesis ក្នុងពេលដំណាលគ្នា។
Meiosis កើតឡើងនៅក្នុងកោសិកាពិសេសនៃពងស្វាស និងអូវែរ ដែលបង្កើត gametes បុរស និងស្ត្រី។ ប្រូតេអ៊ីនដែលជាកត្តាជំរុញនៃ meiosis ត្រូវបានកំណត់អត្តសញ្ញាណ។
ក្នុងអំឡុងពេលដំណើរការនៃ meiosis ចំនួន diploid នៃក្រូម៉ូសូម (2n) ដែលជាលក្ខណៈនៃកោសិកា somatic (ស្នូលកោសិកា) និងកោសិកាមិនទាន់ពេញវ័យបានផ្លាស់ប្តូរទៅជាចំនួន haploid (n) លក្ខណៈនៃកោសិកាដំណុះចាស់ទុំ។ ដូច្នេះជាលទ្ធផលនៃ gametogenesis កោសិកាមេរោគទទួលបានតែពាក់កណ្តាលនៃក្រូម៉ូសូមនៃកោសិកា somatic (រូបភាព 18) ។

អង្ករ។ ១៨.
ឥរិយាបថនៃក្រូម៉ូសូមក្នុងអំឡុងពេល gametogenesis នៅក្នុងសត្វគឺដូចគ្នាទាំងបុរសនិងស្ត្រី។ ទោះជាយ៉ាងណាក៏ដោយ ភេទខុសគ្នានៅក្នុងពេលវេលានៃប្រភពដើមនៃដំណាក់កាលផ្សេងៗគ្នានៃ meiosis ដែលជាពិសេស
ជាពិសេសគួរឱ្យកត់សម្គាល់ចំពោះមនុស្ស។ ចំពោះបុរសក្រោយសម្រាល ដំណើរការពេញលេញនៃ meiosis ត្រូវបានបញ្ចប់ក្នុងរយៈពេលប្រហែល 2 ខែ ខណៈពេលដែលចំពោះស្ត្រី ការបែងចែក meiotic ដំបូងចាប់ផ្តើមនៅក្នុង ovary របស់ទារក ហើយមិនត្រូវបានបញ្ចប់រហូតដល់ការបញ្ចេញពងអូវុលចាប់ផ្តើម ដែលកើតឡើងនៅអាយុប្រហែល 15 ឆ្នាំ។
នៅក្នុងសត្វខ្ពស់ក្នុងករណីបុរស meiosis ត្រូវបានអមដោយការបង្កើត gametes សកម្មចំនួនបួន។
ផ្ទុយទៅវិញ ចំពោះស្ត្រី អូវែលំដាប់ទីពីរនីមួយៗ ផលិតស៊ុតតែមួយប៉ុណ្ណោះ។ ផលិតផលនុយក្លេអ៊ែរផ្សេងទៀតនៃ meiosis ស្ត្រីគឺជាសាកសពកាត់បន្ថយចំនួនបីដែលមិនចូលរួមក្នុងការបន្តពូជនិង degenerate ។
Meiosis មានការបែងចែកនុយក្លេអ៊ែរពីរ។ ការបែងចែកនុយក្លេអ៊ែរ meiotic ដំបូងបំបែកសមាជិកនៃក្រូម៉ូសូមដូចគ្នានីមួយៗបន្ទាប់ពីពួកគេបានផ្គូផ្គងគ្នាទៅវិញទៅមក (សំយោគ) និងផ្លាស់ប្តូរសម្ភារៈហ្សែន (ឆ្លងកាត់) ។ ជាលទ្ធផលនៃការបែងចែកនេះ ស្នូល haploid ពីរត្រូវបានបង្កើតឡើង។ ការបែងចែក meiotic ទីពីរបំបែកផ្នែកបណ្តោយពីរនៃក្រូម៉ូសូម (ក្រូម៉ាទីត) នៅក្នុងស្នូលនីមួយៗដែលបង្កើតបានជាស្នូលបួន haploid ។
ក្នុងអំឡុងពេលនៃដំណើរការ gametogenesis ភាពខុសគ្នានៃស៊ុត (ovogenesis) និង spermatozoa (spermatogenesis) ក៏កើតឡើងដែរ ដែលជាតម្រូវការជាមុនសម្រាប់មុខងាររបស់វា។ ស៊ុតរបស់សត្វមានទំហំធំជាងមេជីវិតឈ្មោល ជាធម្មតាមិនមានចលនា និងមានសារធាតុចិញ្ចឹមដែលធានាដល់ការវិវឌ្ឍន៍នៃអំប្រ៊ីយ៉ុងក្នុងដំណាក់កាលដំបូងបន្ទាប់ពីការបង្កកំណើត។ មេជីវិតឈ្មោលរបស់សត្វភាគច្រើនមាន flagellum ដែលធានានូវឯករាជ្យភាពនៃចលនា។
Meiosis មានសារៈសំខាន់ជីវសាស្រ្តដ៏អស្ចារ្យ។ សូមអរគុណដល់ meiosis ចំនួនថេរនៃក្រូម៉ូសូមត្រូវបានរក្សាទុកនៅក្នុងកោសិកានៃសារពាង្គកាយដោយមិនគិតពីចំនួនជំនាន់។ ដូច្នេះ meiosis រក្សាភាពស្ថិតស្ថេរនៃប្រភេទសត្វ។ ទីបំផុតនៅក្នុង meiosis ដែលជាលទ្ធផលនៃការឆ្លងកាត់ ការផ្សំហ្សែនកើតឡើង ដែលជាកត្តាមួយនៃការវិវត្តន៍។
SPERMATOGENESIS និង OVOGENESIS
Spermatogenesis គឺជាដំណើរការនៃការបង្កើតកោសិកាមេជីវិតឈ្មោលពេញវ័យ។ Spermatozoa វិវត្តនៅក្នុង gonads បុរស (ពងស្វាសឬពងស្វាស) ពីកោសិកា somatic ឯកទេស (រូបភាព 19) ។ កោសិកាឯកទេសបែបនេះមានមុខងារដូចខាងក្រោមៈ
ហៅថាកោសិកាមេជីវិតដំបូង ដែលធ្វើចំណាកស្រុកទៅកាន់ពងស្វាសក្នុងដំណាក់កាលដំបូងនៃការបង្កើតអំប្រ៊ីយ៉ុងរបស់បុរស។ ដូច្នេះ កោសិកាដើមគឺជាអ្នកបង្កើតមុន (មុនគេ) នៃកោសិកាមេជីវិតចាស់។
ពងស្វាសរបស់មនុស្សមានបំពង់ជាច្រើនដែលជញ្ជាំងត្រូវបានបង្កើតឡើងដោយស្រទាប់កោសិកាដែលស្ថិតក្នុងដំណាក់កាលផ្សេងៗគ្នានៃការអភិវឌ្ឍន៍មេជីវិតឈ្មោល។ ស្រទាប់ខាងក្រៅនៃ tubules ត្រូវបានបង្កើតឡើងដោយកោសិកាធំ ៗ ដែលហៅថា spermatogonia ។ កោសិកាទាំងនេះមានសំណុំក្រូម៉ូសូម diploid និងជាកូនចៅនៃកោសិកាមេជីវិតដំបូងនៅក្នុងពងស្វាស។ ក្នុងអំឡុងពេលពេញវ័យរបស់បុគ្គលម្នាក់ ផ្នែកនៃ spermatogonia ផ្លាស់ទីទៅស្រទាប់ខាងក្នុងនៃ tubules ដែលជាលទ្ធផលនៃ meiosis ពួកវាវិវត្តទៅជាកោសិកាដែលហៅថា spermatocytes លំដាប់ទីមួយ (spermatocytes I) បន្ទាប់មកចូលទៅក្នុង spermatocytes លំដាប់ទីពីរ។ (spermatocytes II) និងទីបំផុត

អង្ករ។ ១៩.Spermatogenesis និង oogenesis
ចូលទៅក្នុង spermatids ដែលជាកោសិកា germ haploid ដែលនៅទីបំផុត ខុសគ្នាទៅជាមេជីវិតឈ្មោលពេញវ័យ។ ដូច្នេះក្នុងន័យទូទៅ យើងអាចនិយាយបានថា ការបង្កើតមេជីវិតឈ្មោលត្រូវបានផ្តួចផ្តើមឡើងនៅក្នុងកោសិកា somatic diploid (spermatogonia) បន្តដោយរយៈពេលនៃភាពចាស់នៃកោសិកាមេជីវិតដែលក្នុងនោះការបែងចែកនុយក្លេអ៊ែរពីរកើតឡើងតាមរយៈ meiosis ដែលនាំទៅដល់ការបង្កើត spermatids ។ នេះគឺជារូបភាពរបស់មនុស្ស។
Meiosis (រូបភាពទី 20) នៅក្នុង spermatogenesis កើតឡើងក្នុងដំណាក់កាលជាច្រើន (ដំណាក់កាល) ។ រវាងការបែងចែកមាន interphases ពីរ។ ដូច្នេះការបែងចែក meiotic អាចត្រូវបានតំណាងជាស៊េរីនៃព្រឹត្តិការណ៍ជាបន្តបន្ទាប់គ្នាដូចជាៈ interphase I - ការបែងចែក meiotic ដំបូង (ដំណាក់កាលដំបូង I, យឺត prophase I, metaphase I, anaphase I, telophase I) - interphase II (interokinesis) ។ - ការបែងចែក meiotic ទីពីរ (ដំណាក់កាលទី I, metaphase II, anaphase II, telophase II) ។ ដំណើរការនៃ meiosis គឺមានភាពស្វាហាប់ខ្លាំងណាស់ ដូច្នេះភាពខុសគ្នានៃមីក្រូទស្សន៍រវាងដំណាក់កាលផ្សេងៗគ្នាឆ្លុះបញ្ចាំងពីធម្មជាតិនៃដំណាក់កាលដោយខ្លួនឯង ប៉ុន្តែជាលក្ខណៈសម្បត្តិរបស់ក្រូម៉ូសូមនៅដំណាក់កាលផ្សេងៗគ្នា។ អន្តរដំណាក់កាល I

អង្ករ។ ២០.ដំណាក់កាល Meiosis៖ 1 - ការបែងចែក meiotic ដំបូង; 2 - ការបែងចែក meiotic ទីពីរ
កំណត់លក្ខណៈដោយការពិតដែលថាការចម្លងក្រូម៉ូសូម (DNA ទ្វេដង) កើតឡើងនៅក្នុងវាដែលស្ទើរតែបានបញ្ចប់ទាំងស្រុងដោយការចាប់ផ្តើមនៃដំណាក់កាលដំបូង I.
ការបែងចែក meiotic ដំបូងចាប់ផ្តើមនៅក្នុង spermatocyte បឋមនិងត្រូវបានកំណត់លក្ខណៈដោយ prophase វែងដែលមាន prophase I និង prophase II ផ្លាស់ប្តូរទៅគ្នាទៅវិញទៅមក។ នៅក្នុងដំណាក់កាលទី 1 មាន 5 ដំណាក់កាលសំខាន់ៗ - leptonema, zygonema, pachynema, diplonema និង diakinesis ។
នៅដំណាក់កាល leptonema ក្រូម៉ូសូមនៅក្នុងស្នូលត្រូវបានបង្ហាញជាទម្រង់នៃខ្សែស្រឡាយវង់ស្តើងដែលមានគ្រាប់ពណ៌ងងឹតជាច្រើន (ក្រូម៉ូមេ) ។ ការបំបែកក្រូម៉ូសូម និងអំបោះមិនត្រូវបានគេកត់សំគាល់ទេ ប៉ុន្តែវាត្រូវបានគេជឿថាក្រូម៉ូសូមនៅដំណាក់កាលនេះគឺទ្វេរដង ពោលគឺ diploid ។ Homologues នៃគូក្រូម៉ូសូមនីមួយៗត្រូវបានបង្រួបបង្រួមដោយ chromomeres តាមបណ្តោយប្រវែងរបស់ពួកគេយោងទៅតាមគោលការណ៍ clasp ។
ដំណាក់កាល zygonema ត្រូវបានកំណត់ដោយការបង្កើត synapses រវាងក្រូម៉ូសូម homologous ដែលបណ្តាលឱ្យមានការបង្កើតក្រូម៉ូសូមគូ (bivalents) ។ ក្រូម៉ូសូម X និង Y មានឥរិយាបទខុសគ្នាខ្លះបើប្រៀបធៀបទៅនឹងអូតូសូម។ ពួកវាប្រមូលផ្តុំទៅជារូបធាតុ heterochromatic ពណ៌ងងឹត ផ្គូផ្គងជាលទ្ធផលនៃតំបន់ដូចគ្នានៅខាងចុងរបស់វា។
នៅដំណាក់កាល pachynema ដែលជាដំណាក់កាលវែងបំផុតនៅក្នុង meiotic prophase ការ condensation នៃ bivalents កើតឡើងហើយការបែងចែក chromatid នីមួយៗជាពីរដែលជាលទ្ធផលដែល bivalent នីមួយៗគឺជារចនាសម្ព័ន្ធ helical ស្មុគស្មាញដែលមាន chromatids បងប្អូនស្រីបួន (tetrads) ។ នៅចុងបញ្ចប់នៃដំណាក់កាលនេះ ការបំបែកក្រូម៉ូសូមទ្វេជាគូចាប់ផ្តើម។ ឥឡូវនេះក្រូម៉ូសូម homologous អាចត្រូវបានគេសង្កេតឃើញដោយចំហៀង។ ដូច្នេះ នៅក្នុងការត្រៀមលក្ខណៈមួយចំនួន គេអាចមើលឃើញក្រូម៉ូសូមចំនួនបួន ដែលត្រូវបានបង្កើតឡើងជាលទ្ធផលនៃការចម្លងនៃ homolog នីមួយៗ បង្កើតបានជាក្រូម៉ូសូមបងស្រី។ នៅដំណាក់កាលនេះ ការផ្លាស់ប្តូររវាង homologues និងការបង្កើត chiasmata កើតឡើង។
នៅដំណាក់កាល diplonema ការខ្លី ការឡើងក្រាស់ និងការច្រានទៅវិញទៅមកនៃ chromatids បងស្រីកើតឡើង ជាលទ្ធផលដែល chromatids នៅក្នុង bivalent ត្រូវបានបំបែកស្ទើរតែ។ ការបំបែកត្រូវបានចាត់ទុកថាមិនពេញលេញទេព្រោះ centromere មិនទាន់ត្រូវបានបំបែកនៅក្នុងគូក្រូម៉ូសូមនីមួយៗ។ សម្រាប់ bivalents ពួកគេត្រូវបានប្រារព្ធឡើងនៅកន្លែងផ្សេងៗគ្នាតាមបណ្តោយប្រវែងរបស់ពួកគេដោយ chiasmata ដែលជារចនាសម្ព័ន្ធដែលបង្កើតឡើងរវាង chromatids ដូចគ្នាជាលទ្ធផល។
ការឆ្លងកាត់ពីមុនរវាង homolologs ភ្ជាប់ synaptically ។ នៅក្នុងការត្រៀមលក្ខណៈដ៏ល្អ គេអាចសង្កេតឃើញ chiasmata មួយទៅច្រើន អាស្រ័យលើប្រវែងនៃ bivalent ។ chiasma នីមួយៗដែលបានសង្កេតនៅដំណាក់កាលនេះតំណាងឱ្យលទ្ធផលនៃការផ្លាស់ប្តូរដែលបានកើតឡើងរវាង chromatids ដែលមិនមែនជាបងប្អូនក្នុងដំណាក់កាល pachynema ។ នៅពេលដែលការបង្ហាប់ និងការច្រានចោលនៃ bivalents កាន់តែខ្លាំងនោះ chiasmata ផ្លាស់ទីឆ្ពោះទៅរកចុងបញ្ចប់នៃក្រូម៉ូសូម ពោលគឺការបញ្ចប់នៃក្រូម៉ូសូមកើតឡើង។ នៅចុងបញ្ចប់នៃ diploma, despiralization នៃក្រូម៉ូសូមកើតឡើង; មនុស្សដូចគ្នានៅតែបន្តវាយលុកគ្នាទៅវិញទៅមក។
នៅដំណាក់កាលនៃ diakinesis ដែលស្រដៀងនឹង diplonema ការធ្វើឱ្យខ្លីនៃ bivalents បន្តហើយ chiasmata ចុះខ្សោយ (ថយចុះ) ដែលជាលទ្ធផលដែលឯកតាដាច់ពីគ្នាក្នុងទម្រង់ chromatids (បួន) ត្រូវបានបង្កើតឡើង។ ភ្លាមៗបន្ទាប់ពីការបញ្ចប់នៃដំណាក់កាលនេះការរលាយនៃភ្នាសនុយក្លេអ៊ែរកើតឡើង។
នៅក្នុង metaphase I, bivalents ឈានដល់ការផ្តោតអារម្មណ៍ខ្ពស់បំផុតរបស់ពួកគេ។ ក្លាយជារាងពងក្រពើ ពួកវាមានទីតាំងនៅផ្នែកអេក្វាទ័រនៃស្នូល ដែលពួកគេបង្កើតជាបន្ទះអេក្វាទ័រនៃ meiotic metaphase I. រូបរាងរបស់ bivalent នីមួយៗត្រូវបានកំណត់ដោយចំនួន និងទីតាំងរបស់ chiasmata ។ ចំពោះបុរស ចំនួននៃ chiasmata ក្នុងមួយ bivalent នៅក្នុង metaphase I ជាធម្មតាគឺ 1-5 ។ XY bivalent ក្លាយជារាងជាដំបងដែលជាលទ្ធផលនៃ chiasm ដែលមានទីតាំងនៅស្ថានីយតែមួយ។
នៅក្នុង anaphase I ចលនានៃ centromeres ទល់មុខទៅនឹងប៉ូលទល់មុខនៃកោសិកាចាប់ផ្តើម។ ជាលទ្ធផលក្រូម៉ូសូម homologous ត្រូវបានបំបែក។ ឥឡូវនេះក្រូម៉ូសូមនីមួយៗមានក្រូម៉ាទីតពីរដែលនៅជាប់គ្នាដោយ centromere ដែលមិនបែងចែក និងនៅដដែល។ នេះបែងចែក anaphase I នៃ meiosis ពី anaphase នៃ mitosis ដែលក្នុងនោះ centromere ឆ្លងកាត់ការបែងចែក។ វាជាការសំខាន់ក្នុងការកត់សម្គាល់ថាការឆ្លងកាត់ធ្វើឱ្យ chromatid នីមួយៗមានហ្សែនខុសគ្នា។
នៅដំណាក់កាល telophase I ក្រូម៉ូសូមឈានដល់ប៉ូលដែលបញ្ចប់ការបែងចែក meiotic ដំបូង។ បន្ទាប់ពី telophase I, interphase ខ្លី (interkinesis) កើតឡើងដែលក្រូម៉ូសូម despiral និងក្លាយជា diffuse ឬ telophase I ឆ្លងកាត់ដោយផ្ទាល់ទៅ prophase II នៃការបែងចែក meiotic ទីពីរ។ ក្នុងករណីណាក៏ដោយមិនត្រូវបានគេសង្កេតឃើញការចម្លង DNA ទេ។ បន្ទាប់ពីការបែងចែក meiotic ដំបូង កោសិកាត្រូវបានគេហៅថា spermatocytes លំដាប់ទីពីរ។ ចំនួនក្រូម៉ូសូមនៅក្នុងកោសិកានីមួយៗ ថយចុះពី 2 នទៅ 1n ប៉ុន្តែខ្លឹមសារ DNA មិនទាន់ផ្លាស់ប្តូរទេ។
ការបែងចែក meiotic ទីពីរកើតឡើងក្នុងដំណាក់កាលជាច្រើន (ដំណាក់កាលទី II, metaphase II, anaphase II, telophase II) និងស្រដៀងទៅនឹងការបែងចែក mitotic ។ នៅក្នុងដំណាក់កាលទី II ក្រូម៉ូសូមនៃ spermatocytes ទីពីរនៅតែស្ថិតនៅប៉ូល។ នៅក្នុង metaphase II, centromere នៃក្រូម៉ូសូមទ្វេនីមួយៗបែងចែកដោយផ្តល់នូវក្រូម៉ូសូមថ្មីនីមួយៗជាមួយនឹង centromere របស់វា។ ការបង្កើត spindle ចាប់ផ្តើមឆ្ពោះទៅរកបង្គោលដែលក្រូម៉ូសូមថ្មីផ្លាស់ទី។ នៅក្នុង telophase II ការបែងចែក meiotic ទីពីរបញ្ចប់ជាលទ្ធផលដែល spermatocyte លំដាប់ទីពីរនីមួយៗបង្កើត spermatids ពីរដែល spermatozoa បន្ទាប់មកខុសគ្នា។ ដូចនៅក្នុងមេជីវិតឈ្មោលបន្ទាប់បន្សំ ចំនួនក្រូម៉ូសូមនៅក្នុងមេជីវិតឈ្មោលគឺ haploid (1n) ។ ទោះជាយ៉ាងណាក៏ដោយ ក្រូម៉ូសូមនៃមេជីវិតឈ្មោលគឺនៅលីវ ខណៈពេលដែលក្រូម៉ូសូមនៃមេជីវិតឈ្មោលទីពីរមានទ្វេដង ដែលត្រូវបានបង្កើតឡើងពីក្រូម៉ាទីតពីរ។ ដូច្នេះហើយ ស្នូលនៃមេជីវិតឈ្មោលនីមួយៗមានសំណុំក្រូម៉ូសូមដែលមិនមានលក្ខណៈដូចគ្នា ការបែងចែក meiotic ទីពីរគឺជាការបែងចែកនៃប្រភេទ mitotic (ការបែងចែកអេក្វាទ័រ) ។ វាបំបែកក្រូម៉ាទីតប្អូនស្រីពីរ ហើយខុសពីការបែងចែកកាត់បន្ថយ ដែលក្រូម៉ូសូមដូចគ្នាត្រូវបានបំបែក។ ភាពខុសគ្នាដ៏សំខាន់តែមួយគត់ពី mitosis បុរាណគឺថាមានសំណុំក្រូម៉ូសូម haploid ។
ដូច្នេះការបែងចែក meiotic ដំបូងនៃ spermatocytes លំដាប់ទីមួយនាំឱ្យមានការបង្កើត spermatocytes ទីពីរ (លំដាប់ទីពីរ) ។ chromatids ទាំងពីរនៃរចនាសម្ព័ន្ធដែលបានបង្កើតឡើងជាលទ្ធផលនៃការបែងចែកកាត់បន្ថយគឺជា chromatids បងស្រី។ ក្រោយមកទៀតកើតឡើងជាលទ្ធផលនៃការចម្លងមុនការបែងចែក meiotic ដំបូង។ ការបែងចែកមេជីវិតឈ្មោលទីពីរនៃមេជីវិតឈ្មោលបន្ទាប់បន្សំនីមួយៗ បង្កើតបានជាមេជីវិតឈ្មោលចំនួនបួន។ ដូច្នេះនៅក្នុង meiosis ធម្មតា កោសិកាបែងចែកពីរដង ខណៈដែលក្រូម៉ូសូមបែងចែកតែម្តងគត់ (រូបភាព 21)។
ដំណាក់កាលចុងក្រោយនៅក្នុង spermatogenesis ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងភាពខុសគ្នា ដែលបញ្ចប់ដោយ spermatids រាងស្វ៊ែរដែលមានទំហំធំ និងមិនអាចចល័តបានប្រែទៅជាមេជីវិតឈ្មោលតូចមួយដែលមានរាងវែង។
នៅក្នុងសត្វឈ្មោលពេញវ័យ (ពេញវ័យ) មេជីវិតឈ្មោលកើតឡើងនៅក្នុងពងស្វាសជានិច្ច ឬតាមកាលកំណត់ (តាមរដូវ)។ ឧទាហរណ៍ នៅក្នុងសត្វល្អិត វាត្រូវការពេលតែពីរបីថ្ងៃប៉ុណ្ណោះ ដើម្បីបញ្ចប់វដ្តមេជីវិតឈ្មោល ខណៈពេលដែលនៅក្នុងថនិកសត្វ វដ្តនេះអូសបន្លាយជាច្រើនសប្តាហ៍ ឬសូម្បីតែ

អង្ករ។ ២១.ការចែកចាយក្រូម៉ូសូមក្នុងអំឡុងពេល gametogenesis
ខែ។ នៅក្នុងមនុស្សពេញវ័យ spermatogenesis កើតឡើងពេញមួយឆ្នាំ។ ពេលវេលានៃការអភិវឌ្ឍន៍ spermatogonia បឋមទៅជាមេជីវិតឈ្មោលពេញវ័យគឺប្រហែល 74 ថ្ងៃ។
កោសិកាបន្តពូជរបស់បុរស ដែលផលិតដោយសារពាង្គកាយនៃប្រភេទផ្សេងៗគ្នា ត្រូវបានកំណត់លក្ខណៈដោយការចល័ត និងភាពសម្បូរបែបនៃទំហំ និងរចនាសម្ព័ន្ធ (រូបភាព 22)។ មេជីវិតឈ្មោលរបស់មនុស្សម្នាក់ៗមានបីផ្នែក - ក្បាល ផ្នែកកណ្តាល និងកន្ទុយ (រូបភាព 23) ។ ក្បាលមេជីវិតឈ្មោលមានស្នូលមួយដែលមានសំណុំក្រូម៉ូសូម haploid ។
ក្បាលត្រូវបានបំពាក់ដោយ acrosome ដែលមានអង់ស៊ីម lytic ដែលចាំបាច់សម្រាប់មាតិកាមេជីវិតឈ្មោលចូលទៅក្នុងស៊ុត។ កណ្តាលពីរក៏ត្រូវបានធ្វើមូលដ្ឋានីយកម្មនៅក្នុងក្បាល - proc-

អង្ករ។ ២២. ទម្រង់មេជីវិតឈ្មោល។

អង្ករ។ ២៣. រចនាសម្ព័ន្ធមេជីវិតឈ្មោល៖ ក - រូបភាពមីក្រូទស្សន៍ស្រាលនៅក្នុងយន្តហោះផ្សេងៗគ្នា៖ ១ - ក្បាល ២ - ផ្នែកកណ្តាល ៣ - កន្ទុយ; ខ - ការបង្កើតឡើងវិញតាមគ្រោងការណ៍នៃរូបភាពមីក្រូទស្សន៍អេឡិចត្រុង៖ 1 - ស្នូល, 2 - អាក្រូសូម, 3 - កណ្តាល (ជិតកណ្តាល), 4 - ចិញ្ចៀនកណ្តាល, 5 - helix mitochondrial,
6 - ខ្សែស្រឡាយអ័ក្ស
ស៊ីម៉ាល់ ដែលជំរុញការបែងចែកស៊ុតដែលបង្កកំណើតដោយមេជីវិតឈ្មោល និងផ្នែកខាងចុង ដែលបង្កើតបានជាផ្នែកអ័ក្សនៃកន្ទុយ។ ផ្នែកកណ្តាលនៃមេជីវិតឈ្មោលមានរាងកាយ basal នៃកន្ទុយនិង mitochondria ។ កន្ទុយ (ដំណើរការ) នៃមេជីវិតឈ្មោលត្រូវបានបង្កើតឡើងដោយដំបង aximal ខាងក្នុង និងសំបកខាងក្រៅ ដែលមានដើមកំណើតពី cytoplasmic ។ មេជីវិតឈ្មោលរបស់មនុស្សត្រូវបានកំណត់លក្ខណៈដោយចលនាដ៏សំខាន់។
កោសិកា OVOGENESIS និង OGGLE
ដំណើរការនៃការបង្កើតស៊ុតត្រូវបានគេហៅថា oogenesis ។ មុខងាររបស់វាគឺដើម្បីធានានូវសំណុំក្រូម៉ូសូម haploid នៅក្នុងស្នូលនៃស៊ុត និងតម្រូវការអាហារូបត្ថម្ភរបស់ហ្សីហ្គោត។ Oogenesis នៅក្នុងការបង្ហាញរបស់វាគឺអាចប្រៀបធៀបជាមូលដ្ឋានទៅនឹង spermatogenesis ។
នៅក្នុងថនិកសត្វនិងមនុស្ស oogenesis ចាប់ផ្តើមនៅអំឡុងពេលមានផ្ទៃពោះ (មុនពេលកើត)។ Oogonia ដែលជាកោសិកាតូចៗដែលមានស្នូលធំគួរសម និងបានធ្វើមូលដ្ឋានីយកម្មនៅក្នុងឫសអូវែចាប់ផ្តើមខុសគ្នាទៅជា oocytes បឋម។ ក្រោយមកទៀតត្រូវបានបង្កើតឡើងរួចហើយនៅក្នុងខែទី 3 នៃការអភិវឌ្ឍន៍ពោះវៀនបន្ទាប់ពីនោះពួកគេចូលទៅក្នុងដំណាក់កាលនៃការបែងចែក meiotic ដំបូង។ នៅពេលដែលក្មេងស្រីកើតមក អូវែបឋមទាំងអស់គឺស្ថិតនៅក្នុងដំណាក់កាលនៃការបែងចែក meiotic ដំបូងរួចទៅហើយ។ oocytes បឋមនៅតែស្ថិតក្នុងដំណាក់កាល prophase រហូតដល់ការចាប់ផ្តើមនៃភាពពេញវ័យចំពោះស្ត្រី។ អូវែមួយមានផ្ទុកមីតូឆុនឌ្រីរហូតដល់ 100,000 ។ នៅពេលដែលឫសអូវែមានភាពចាស់ទុំនៅពេលចាប់ផ្តើមនៃភាពពេញវ័យ ប្រូហ្វាស meiotic បន្តនៅក្នុង oocytes បឋម។ ការបែងចែក meiotic ដំបូងសម្រាប់ស៊ុតដែលកំពុងលូតលាស់នីមួយៗត្រូវបានបញ្ចប់ក្នុងរយៈពេលខ្លីមុនពេលនៃការបញ្ចេញពងអូវុលនៃស៊ុតនោះ។ ជាលទ្ធផលនៃការបែងចែក meiotic ដំបូងនិងការចែកចាយមិនស្មើគ្នានៃ cytoplasm កោសិកាលទ្ធផលមួយក្លាយជា oocyte បន្ទាប់បន្សំមួយទៀត - រាងប៉ូល (កាត់បន្ថយ) ។
ការបែងចែក meiotic ទីពីរកើតឡើងនៅពេលដែល oocyte ទីពីរ (ស៊ុតដែលកំពុងលូតលាស់) ឆ្លងកាត់ពីអូវែរចូលទៅក្នុងបំពង់ fallopian ។ ទោះជាយ៉ាងណាក៏ដោយ ការបែងចែកនេះមិនត្រូវបានបញ្ចប់រហូតដល់មេជីវិតឈ្មោលជ្រាបចូលទៅក្នុង oocyte ទីពីរ ដែលជាធម្មតាកើតឡើងនៅក្នុងបំពង់ fallopian ។ នៅពេលដែលមេជីវិតឈ្មោលជ្រាបចូលទៅក្នុង oocyte ទីពីរ ក្រោយមកទៀតបានបែងចែកជាលទ្ធផល បង្កើតជាពងមាន់មួយ (ពងចាស់) ដែលមាន pronucleous ដែលមានសំណុំតែមួយ។
នៃក្រូម៉ូសូមមាតាចំនួន 23 ។ កោសិកាផ្សេងទៀតដែលកើតចេញពីការបែងចែកនេះគឺជារូបកាយប៉ូលទីពីរ មិនអាចអភិវឌ្ឍបន្ថែមទៀតបានទេ។ នៅពេលនេះរាងប៉ូល (កាត់បន្ថយ) ក៏ត្រូវបែងចែកជាពីរ។ ដូច្នេះការអភិវឌ្ឍនៃ oocyte លំដាប់ទីមួយត្រូវបានអមដោយការបង្កើត ovotide មួយនិងសាកសពកាត់បន្ថយបី។ នៅក្នុងអូវែ អូវែរ ៣០០-៤០០ អូវែ ជាធម្មតាមានភាពចាស់ទុំតាមរបៀបនេះពេញមួយជីវិត ប៉ុន្តែមានតែ oocyte មួយប៉ុណ្ណោះដែលចាស់ទុំក្នុងមួយខែ។ ក្នុងអំឡុងពេលនៃភាពខុសគ្នានៃស៊ុតភ្នាសត្រូវបានបង្កើតឡើងហើយស្នូលរបស់វាថយចុះនៅក្នុងទំហំ។
នៅក្នុងប្រភេទសត្វមួយចំនួន oogenesis កើតឡើងយ៉ាងឆាប់រហ័ស និងជាបន្តបន្ទាប់ ហើយជាលទ្ធផលក្នុងការផលិតពងមាន់យ៉ាងច្រើន។
ទោះបីជាមានភាពស្រដៀងគ្នាជាមួយ spermatogenesis ក៏ដោយ oogenesis ត្រូវបានកំណត់ដោយលក្ខណៈពិសេសមួយចំនួន។ សារធាតុអាហារូបត្ថម្ភ (yolk) នៃ oocyte បឋមមិនត្រូវបានចែកចាយស្មើៗគ្នាក្នុងចំណោមកោសិកាទាំងបួនដែលត្រូវបានបង្កើតឡើងជាលទ្ធផលនៃការបែងចែក meiotic ។ បរិមាណសំខាន់នៃ yolk ត្រូវបានរក្សាទុកនៅក្នុងកោសិកាធំមួយ ខណៈដែលសាកសពប៉ូលមានផ្ទុកសារធាតុនេះតិចតួចបំផុត។ ជាលទ្ធផលនៃការបែងចែក សាកសពប៉ូលទី 1 និងទី 2 ទទួលបានសំណុំក្រូម៉ូសូមដូចគ្នាទៅនឹង oocytes ទីពីរ ប៉ុន្តែពួកវាមិនក្លាយជាកោសិកាមេរោគទេ។ ដូច្នេះ ស៊ុតសម្បូរទៅដោយសារធាតុចិញ្ចឹមច្រើនបើធៀបនឹងមេជីវិតឈ្មោល។ ភាពខុសគ្នានេះត្រូវបានប្រកាសជាពិសេសនៅក្នុងករណីនៃសត្វពង។
ស៊ុតរបស់ថនិកសត្វមានរូបរាងរាងពងក្រពើ ឬរាងវែងបន្តិច (រូបភាពទី 24) ហើយត្រូវបានកំណត់ដោយលក្ខណៈធម្មតានៃរចនាសម្ព័ន្ធកោសិកា។ ពួកវាផ្ទុកនូវរចនាសម្ព័ន្ធទាំងអស់នៃកោសិកា somatic ទោះជាយ៉ាងណាក៏ដោយ អង្គការខាងក្នុងនៃកោសិកានៃស៊ុតគឺជាក់លាក់ណាស់ ហើយត្រូវបានកំណត់ដោយការពិតដែលថាស៊ុតក៏ជាបរិស្ថានដែលធានាដល់ការវិវត្តនៃហ្សីហ្គោតផងដែរ។ លក្ខណៈពិសេសមួយនៃលក្ខណៈនៃស៊ុតគឺភាពស្មុគស្មាញនៃរចនាសម្ព័ន្ធនៃភ្នាសរបស់វា។ នៅក្នុងសត្វជាច្រើន ភ្នាសបឋម ទីពីរ និងទីបីនៃស៊ុតត្រូវបានសម្គាល់។ សែលបឋម (ខាងក្នុង) ត្រូវបានបង្កើតឡើងនៅដំណាក់កាល oocyte ។ តំណាងឱ្យស្រទាប់ផ្ទៃនៃ oocyte វាមានរចនាសម្ព័ន្ធស្មុគ្រស្មាញ ដោយសារវាត្រូវបានជ្រាបចូលដោយការរីកធំនៃកោសិកា follicular ដែលនៅជាប់នឹងវា។ សែលទីពីរ (កណ្តាល) ត្រូវបានបង្កើតឡើងដោយកោសិកា follicular ហើយសំបកទីបី (ខាងក្រៅ) ត្រូវបានបង្កើតឡើងដោយសារធាតុដែលជាផលិតផលសម្ងាត់នៃក្រពេញអូវែដែលស៊ុតឆ្លងកាត់។ ឧទាហរណ៍នៅក្នុងបក្សី។
ភ្នាសទីបីនៃស៊ុតគឺជាភ្នាស albumen, subshell និងសែល។ ស៊ុតរបស់ថនិកសត្វត្រូវបានកំណត់លក្ខណៈដោយវត្តមាននៃភ្នាសពីរ។ រចនាសម្ព័ន្ធនៃសមាសធាតុ intracellular នៃស៊ុតគឺជាក់លាក់ទាក់ទងនឹងប្រភេទសត្វ ហើយជួនកាលថែមទាំងមានលក្ខណៈបុគ្គលទៀតផង។

អង្ករ។ ២៤. រចនាសម្ព័ន្ធស៊ុត៖ 1 - កោសិកា follicular; 2 - សែល; 3 - cytoplasm; ៤- ស្នូល
ជីជាតិ
ការបង្កកំណើតគឺជាដំណើរការនៃការរួមបញ្ចូលគ្នារវាង gametes បុរស និងស្ត្រី ដែលនាំទៅដល់ការបង្កើត zygote និងការអភិវឌ្ឍជាបន្តបន្ទាប់នៃសារពាង្គកាយថ្មីមួយ។ ក្នុងអំឡុងពេលនៃការបង្កកំណើត សំណុំក្រូម៉ូសូម diploid ត្រូវបានបង្កើតឡើងនៅក្នុង zygote ដែលកំណត់ពីសារៈសំខាន់ជីវសាស្រ្តដ៏អស្ចារ្យនៃដំណើរការនេះ។
អាស្រ័យលើប្រភេទនៃសារពាង្គកាយដែលបន្តពូជផ្លូវភេទ ការបង្កកំណើតខាងក្រៅ និងខាងក្នុងត្រូវបានសម្គាល់។ ការបង្កកំណើតខាងក្រៅកើតឡើងនៅក្នុងបរិយាកាសដែលកោសិកាបន្តពូជរបស់បុរស និងស្ត្រីចូល។ ឧទាហរណ៍ការបង្កកំណើតនៅក្នុងត្រីគឺខាងក្រៅ។ កោសិកាបន្តពូជរបស់បុរស (ទឹកដោះគោ) និងស្ត្រី (ពងត្រី) ដែលបញ្ចេញដោយពួកវាចូលទៅក្នុងទឹក ដែលពួកគេជួបគ្នា និងរួបរួមគ្នា។
ការបង្កកំណើតខាងក្នុងត្រូវបានធានាដោយការផ្ទេរមេជីវិតឈ្មោលពីរាងកាយបុរសទៅរាងកាយស្ត្រី។ ការបង្កកំណើតបែបនេះកើតឡើងនៅក្នុងថនិកសត្វ ដែលជាចំណុចកណ្តាលនៃការលាយបញ្ចូលគ្នានៃស្នូលនៃកោសិកាមេជីវិត។ វាត្រូវបានគេជឿថាមាតិកានៃមេជីវិតឈ្មោលមួយជ្រាបចូលទៅក្នុងស៊ុត។ នៅក្នុងយន្តការនៃការបង្កកំណើត
ការបញ្ចប់ ភាគច្រើននៅតែមិនច្បាស់លាស់។ ទិន្នន័យស្តីពីការបង្កកំណើតនៅក្នុង urchins សមុទ្របង្ហាញថារួចទៅហើយ 2 វិនាទីបន្ទាប់ពីការទំនាក់ទំនងនៃមេជីវិតឈ្មោលនិងស៊ុតការផ្លាស់ប្តូរកើតឡើងនៅក្នុងលក្ខណៈសម្បត្តិអគ្គិសនីនៃភ្នាសប្លាស្មានៃក្រោយ។ ការលាយបញ្ចូលគ្នានៃ gametes កើតឡើងក្នុងរយៈពេល 7 វិនាទី។ វាត្រូវបានសន្មត់ថាការជ្រៀតចូលនៃមាតិកានៃមេជីវិតឈ្មោលតែមួយគត់ក្នុងចំណោមមេជីវិតឈ្មោលជាច្រើនចូលទៅក្នុងស៊ុតត្រូវបានពន្យល់ដោយការផ្លាស់ប្តូរលក្ខណៈសម្បត្តិអគ្គិសនីនៃភ្នាសប្លាស្មារបស់វា។ ស៊ុតបង្កកំណើតបង្កើតបានជាហ្សីហ្គោត។ មានមតិពីរអំពីហេតុផលសម្រាប់ការធ្វើឱ្យសកម្មនៃការរំលាយអាហារស៊ុតដោយមេជីវិតឈ្មោល។ អ្នកខ្លះជឿថាការភ្ជាប់មេជីវិតឈ្មោលទៅនឹងអ្នកទទួលខាងក្រៅនៅលើផ្ទៃកោសិកាគឺជាសញ្ញាមួយដែលចូលទៅក្នុងស៊ុតតាមរយៈភ្នាស និងធ្វើឱ្យសកម្ម inositol triphosphate និងអ៊ីយ៉ុងកាល់ស្យូមនៅទីនោះ។ អ្នកផ្សេងទៀតជឿថាមេជីវិតឈ្មោលមានកត្តាចាប់ផ្តើមពិសេស។
ការវិវឌ្ឍន៍ពិសោធន៍ដែលបានធ្វើឡើងក្នុងប៉ុន្មានឆ្នាំថ្មីៗនេះបានបង្ហាញថាការបង្កកំណើតនៃស៊ុតរបស់ថនិកសត្វ រួមទាំងមនុស្សគឺអាចធ្វើទៅបាននៅក្នុង vitro ។ ជាងនេះទៅទៀត អំប្រ៊ីយ៉ុងដែលត្រូវបានបង្កើតនៅក្នុង vitro អាចត្រូវបានបញ្ចូលទៅក្នុងស្បូនរបស់ស្ត្រី ដែលពួកគេទទួលបានការអភិវឌ្ឍន៍ធម្មតាបន្ថែមទៀត។ មានករណីជាច្រើននៃកំណើតរបស់កុមារ "បំពង់សាកល្បង" ។
មិនដូចសត្វដែលបន្តពូជដោយ zygogenesis ទេ សារពាង្គកាយជាច្រើនមានសមត្ថភាពបន្តពូជដោយ parthenogenesis (មកពីភាសាក្រិច។ parthenos -ព្រហ្មចារី និង ហ្សែន -កំណើត) ដែលសំដៅលើការបន្តពូជនៃសារពាង្គកាយពីស៊ុតដែលមិនទាន់បង្កកំណើត។ មាន parthenogenesis កាតព្វកិច្ច និង facultative ។ Obligate parthenogenesis បានក្លាយជាវិធីសាស្រ្តសំខាន់នៃការបន្តពូជនៃសារពាង្គកាយនៃប្រភេទសត្វមួយចំនួន ឧទាហរណ៍ ជីងចក់ Caucasian ។ សត្វនៃប្រភេទនេះគឺមានតែស្រីប៉ុណ្ណោះ។ ផ្ទុយទៅវិញ facultative parthenogenesis មានន័យថាស៊ុតអាចអភិវឌ្ឍបានទាំងដោយគ្មានការបង្កកំណើត និងបន្ទាប់ពីការបង្កកំណើត។ នៅក្នុងវេន facultative parthenogenesis គឺស្ត្រី និងបុរស។ រោគស្ត្រី ជារឿយៗត្រូវបានគេសង្កេតឃើញនៅក្នុងសត្វឃ្មុំ ស្រមោច និងសត្វ rotifers ដែលក្នុងនោះបុរសកើតចេញពីស៊ុតមិនទាន់បង្កកំណើត។ បុរស parthenogenesis កើតឡើងនៅក្នុងសារាយ isogamous មួយចំនួន។
Parthenogenesis អាចមានទាំងធម្មជាតិ និងសិប្បនិម្មិត (បង្កឡើង)។ យន្តការនៃ parthenogenesis សិប្បនិម្មិតគឺការរលាកនៃស៊ុតដោយប្រើរូបវិទ្យា ឬគីមី
កត្តា ical ដែលនាំទៅដល់ការធ្វើឱ្យសកម្មនៃស៊ុត និងជាផលវិបាកដល់ការវិវត្តនៃស៊ុតមិនទាន់បង្កកំណើត។ parthenogenesis សិប្បនិម្មិតត្រូវបានគេសង្កេតឃើញនៅក្នុងសត្វនៃក្រុមប្រព័ន្ធជាច្រើន - echinoderms ដង្កូវ mollusks និងសូម្បីតែថនិកសត្វ។
ទម្រង់ដែលគេស្គាល់នៃ parthenogenesis ត្រូវបានគេហៅថា androgenesis (មកពីភាសាក្រិក។ Andros -បុរស ហ្សែន -ប្រភពដើម) ។ ប្រសិនបើស្នូលនៅក្នុងស៊ុតអសកម្ម ហើយបន្ទាប់មកមេជីវិតឈ្មោលជាច្រើនជ្រាបចូលទៅក្នុងវា នោះសារពាង្គកាយបុរសនឹងវិវឌ្ឍន៍ចេញពីស៊ុតបែបនេះ ដែលជាលទ្ធផលនៃការលាយបញ្ចូលគ្នានៃស្នូលនៃមេជីវិតឈ្មោល (មេជីវិតឈ្មោល)។
មានករណីដែល parthenogenesis កើតឡើងជាវដ្ត អាស្រ័យលើរដូវកាល។ ឧទាហរណ៍ rotifers, daphnia និង aphids បន្តពូជនៅរដូវក្តៅដោយ parthenogenesis ហើយនៅរដូវស្លឹកឈើជ្រុះដោយការបង្កកំណើតនៃស៊ុតនិងការបង្កើត zygotes ពោលគឺដោយ zygogenesis ។
តួនាទីនៃ parthenogenesis និងទម្រង់របស់វានៅក្នុងធម្មជាតិគឺតូច ដោយសារវាមិនផ្តល់នូវសមត្ថភាពសម្របខ្លួនយ៉ាងទូលំទូលាយនៃសារពាង្គកាយ។
ផ្ទុយទៅនឹង zygogenesis និង parthenogenesis មាន gynogenesis ដែលជា pseudogamy នៅពេលដែលមេជីវិតឈ្មោលចូលទៅក្នុងស៊ុត និងធ្វើឱ្យវាសកម្ម ប៉ុន្តែស្នូលមេជីវិតឈ្មោលមិនបញ្ចូលគ្នាជាមួយស្នូលនៃស៊ុតនោះទេ។ ក្នុងករណីនេះ កូនចៅដែលមានលទ្ធផលគឺមានតែស្រីប៉ុណ្ណោះ។ Gynogenesis កើតឡើងដោយធម្មជាតិនៅក្នុង nematodes និងត្រី ប៉ុន្តែវាក៏អាចត្រូវបានបង្កឡើងដោយសិប្បនិម្មិតផងដែរ។ ជាពិសេស ករណីនៃរោគស្ត្រីសិប្បនិម្មិតនៅក្នុងដង្កូវនាង ត្រី និងសត្វអំពិលអំពែក ត្រូវបានពិពណ៌នា។
ភាពរីករាយ និងការប្រើប្រាស់ជំនួស។
ការផ្លាស់ប្តូរនៃជំនាន់
សារពាង្គកាយដែលបន្តពូជផ្លូវភេទត្រូវបានកំណត់ដោយដំណាក់កាល haploid និង diploid ឆ្លាស់គ្នាក្នុងការអភិវឌ្ឍន៍របស់វា។ នៅក្នុងសារពាង្គកាយជាច្រើន រួមទាំងថនិកសត្វ ការឆ្លាស់គ្នានេះគឺទៀងទាត់ ហើយការរក្សានូវលក្ខណៈនៃប្រភេទសត្វគឺផ្អែកលើវា។
សារពាង្គកាយជាច្រើនក៏ត្រូវបានកំណត់លក្ខណៈដោយការផ្លាស់ប្តូរនៃជំនាន់នៅពេលដែលជំនាន់នៃបុគ្គលបន្តពូជដោយភេទត្រូវបានជំនួសដោយជំនាន់នៃបុគ្គលបន្តពូជផ្លូវភេទជាមួយនឹងការបង្កើត gametes ។ នេះត្រូវបានគេហៅថាការផ្លាស់ប្តូរបឋមនៃជំនាន់។ វាត្រូវបានរកឃើញនៅក្នុង sporozoans, flagellates និងរុក្ខជាតិជាច្រើន។ ការផ្លាស់ប្តូរបឋមនៃជំនាន់គឺទៀងទាត់ ហើយវាមានវត្តមាន
អ្វីដែលត្រូវបានរកឃើញនៅក្នុងធម្មជាតិបង្ហាញពីការអភិរក្សនៅក្នុង phylogeny នៃសារពាង្គកាយជាច្រើននៃការបន្តពូជទាំងផ្លូវភេទនិងផ្លូវភេទ។ នៅក្នុងសារពាង្គកាយផ្សេងទៀត ការផ្លាស់ប្តូរនៃការបន្តពូជផ្លូវភេទជាមួយ parthenogenesis កើតឡើង។ នេះត្រូវបានគេហៅថាការផ្លាស់ប្តូរជំនាន់បន្ទាប់បន្សំ។ ឧទាហរណ៍នៅក្នុង trematodes ការបន្តពូជផ្លូវភេទត្រូវបានជំនួសជាទៀងទាត់ដោយ parthenogenesis ។ ការជំនួសបន្ទាប់បន្សំនៃជំនាន់នេះត្រូវបានគេហៅថា heterogony ។ នៅក្នុង coelenterates នៅដំណាក់កាលខ្លះនៃការអភិវឌ្ឍន៍ការផ្លាស់ប្តូរកើតឡើងពីការបន្តពូជផ្លូវភេទទៅជា asexual (លូតលាស់) ។ ទម្រង់នៃការជំនួសបន្ទាប់បន្សំនៃជំនាន់នេះត្រូវបានគេហៅថា metagenesis ។
DIMORPHISM ផ្លូវភេទ។ រោគស្ត្រី
បុរសនិងស្ត្រីត្រូវបានកំណត់លក្ខណៈដោយលក្ខណៈ phenotypic ជាក់លាក់។ ភាពខុសគ្នារវាងស្ត្រី និងបុរសនៅក្នុងលក្ខណៈសម្បត្តិរបស់ពួកគេត្រូវបានគេហៅថា dimorphism ផ្លូវភេទ។ នៅក្នុងសត្វវាកើតឡើងរួចហើយនៅដំណាក់កាលទាបនៃការអភិវឌ្ឍន៍ការវិវត្តន៍ ឧទាហរណ៍នៅក្នុង helminths ជុំ និង arthropods ហើយឈានដល់ការបញ្ចេញមតិដ៏អស្ចារ្យបំផុតរបស់វានៅក្នុងសត្វឆ្អឹងខ្នង។
ប្រសិនបើកោសិកាបន្តពូជរបស់បុរស និងស្ត្រី ត្រូវបានផលិតដោយបុគ្គលដូចគ្នា ដែលមានទាំងបុរស និងស្ត្រី នោះបាតុភូតនេះត្រូវបានគេហៅថា hermaphroditism ពិត។ វាត្រូវបានរកឃើញនៅក្នុងពពួកដង្កូវនាង អន្ទង់ និងមូស។ នៅក្នុងពពួកដង្កូវនាង សត្វ gonads ឈ្មោល និងញី ដំណើរការពេញមួយជីវិតរបស់មនុស្សម្នាក់ៗ។ ផ្ទុយទៅវិញ នៅក្នុង mollusks gonads ផលិតស៊ុត និងមេជីវិតឈ្មោលឆ្លាស់គ្នា។
Hermaphroditism ពិតក៏កើតមានចំពោះមនុស្សដែរ ដែលបណ្តាលមកពីការវិវត្តន៍ខុសប្រក្រតី។ ហ្សែននៃពពួក hermaphrodites គឺ 46 XX ឬ 46 XY ដែលករណីភាគច្រើនគឺ XX (ប្រហែល 60%) ។ Genotype XX ត្រូវបានរកឃើញញឹកញាប់បំផុតនៅក្នុង hermaphrodites នៃប្រជាជនអាហ្វ្រិក Negroid ខណៈពេលដែល XY ត្រូវបានរកឃើញញឹកញាប់បំផុតក្នុងចំណោមជនជាតិជប៉ុន។ នៅក្នុង hermaphrodites នៃប្រភេទទាំងពីរ, ទំនោរឆ្ពោះទៅរក asymmetry gonadal ទ្វេភាគីត្រូវបានកត់សម្គាល់។ ក្នុងចំណោម hermaphrodites ពិតប្រាកដក៏មានក្រូម៉ូសូម mosaics ដែលនៅក្នុងកោសិកា somatic មួយចំនួនមានក្រូម៉ូសូម XX គូហើយនៅក្នុងផ្សេងទៀត - គូនៃ XY ។
ជំងឺក្លនលូនក្លែងក្លាយ ត្រូវបានគេស្គាល់ផងដែរ នៅពេលដែលបុគ្គលម្នាក់ៗមានប្រដាប់បន្តពូជខាងក្រៅ និងលក្ខណៈផ្លូវភេទបន្ទាប់បន្សំ ដែលជាលក្ខណៈនៃភេទទាំងពីរ ប៉ុន្តែផលិតកោសិកាមេរោគតែមួយប្រភេទ គឺបុរស ឬស្ត្រី។
ប្រភពដើមនៃវិធីសាស្ត្រផលិតឡើងវិញ
វាត្រូវបានគេជឿថាបុរាណបំផុតគឺការបន្តពូជដោយភេទជាពិសេសការបន្តពូជលូតលាស់។ ពីក្រោយនេះ ការបន្តពូជដោយការបង្កើត spores ត្រូវបានបង្កើតឡើង អត្ថប្រយោជន៍ដែលមិនគួរឱ្យសង្ស័យនោះគឺថាវាផ្តល់ឱកាសកាន់តែប្រសើរឡើងសម្រាប់ការអភិរក្សប្រភេទសត្វ និងជាពិសេសសម្រាប់ការបែកខ្ញែករបស់វា។
ការបន្តពូជផ្លូវភេទគឺជាមធ្យោបាយដ៏មានប្រសិទ្ធភាពបំផុតនៃការបន្តពូជរបស់សារពាង្គកាយ។ វាត្រូវបានគេជឿថាវាវិវត្តន៍ពីការរួមភេទដែលលេចឡើងប្រហែល 1 ពាន់លានឆ្នាំមុនហើយដំណាក់កាលដំបូងត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងភាពស្មុគស្មាញក្នុងការវិវត្តនៃ gametes ។ gametes បុព្វកាលត្រូវបានកំណត់លក្ខណៈដោយភាពខុសគ្នា morphological មិនគ្រប់គ្រាន់ដែលជាលទ្ធផលនៃ isogamy (មកពីភាសាក្រិច។ isos-ស្មើ, ហ្គាម៉ូស -អាពាហ៏ពិពាហ៌) នៅពេលដែលកោសិកាផ្លូវភេទគឺជា អ៊ីសូហ្កាមេត ដែលមិនបែងចែកទៅជាទម្រង់បុរស និងស្ត្រី។
ក្រោយមក anisogamy បានអភិវឌ្ឍ (មកពីភាសាក្រិក។ អានីសូស-មិនស្មើគ្នា ហ្គាម៉ូស -អាពាហ៍ពិពាហ៍) កំណត់លក្ខណៈដោយវត្តមាននៃ gametes ខុសគ្នាដែលខុសគ្នាតែនៅក្នុងទំហំ។ នៅដំណាក់កាលក្រោយៗនៃការវិវត្តន៍ ភាពខុសប្លែកគ្នាយ៉ាងមុតស្រួចបានកើតឡើងនៅក្នុងភាពចល័ត រូបរាង និងទំហំនៃ gametes ។ នៅក្នុងដំណើរការនៃការវិវត្តន៍ សត្វឆ្អឹងខ្នងក៏បានបង្កើតឧបករណ៍បន្ថែមមួយចំនួនដែលជួយសម្រួលដល់ការផ្ទេរមេជីវិតឈ្មោលរបស់បុរសចូលទៅក្នុងប្រដាប់បន្តពូជរបស់ស្ត្រី និងបង្កើតលក្ខខណ្ឌសម្រាប់ការអភិវឌ្ឍនៃស៊ុតបង្កកំណើត។ ក្នុងអំឡុងពេលនៃការវិវត្តន៍ ឧបករណ៍ទាំងនេះបានអភិវឌ្ឍចេញពីប្រព័ន្ធ excretory ដែលនាំឱ្យមានការបង្កើតប្រព័ន្ធ genitourinary ។
រដ្ឋ diploid ផ្តល់អត្ថប្រយោជន៍យ៉ាងខ្លាំងដល់សារពាង្គកាយ ដោយសារតែការប្រមូលផ្តុំនៃ alleles ផ្សេងគ្នាកើតឡើងនៅក្នុងរដ្ឋនេះ។ ដូច្នេះ ការបន្តពូជផ្លូវភេទក៏មានអត្ថប្រយោជន៍ផងដែរ ដែលវាផ្តល់សារពាង្គកាយអូលទ្ធភាពកាន់តែច្រើននៃភាពប្រែប្រួលបើប្រៀបធៀបទៅនឹងភេទដូចគ្នា ហើយនេះដើរតួយ៉ាងសំខាន់ក្នុងការវិវត្តន៍។
Parthenogenesis មានគុណសម្បត្តិបន្តពូជច្បាស់លាស់ ព្រោះវាបង្កើតបានតែពូជស្ត្រីប៉ុណ្ណោះ។ ទោះយ៉ាងណាក៏ដោយវាកម្រណាស់។ សម្មតិកម្មពីរត្រូវបានប្រើដើម្បីពន្យល់ពីប្រេកង់ទាបនៃការជំនួសការបន្តពូជផ្លូវភេទដោយ parthenogenesis នៅក្នុងចំនួនប្រជាជនធម្មជាតិនៃសារពាង្គកាយ។ យោងទៅតាមមួយក្នុងចំណោមពួកគេ (ការបំប្លែងបំរែបំរួល) ការរួមភេទគឺជាការបន្សាំដែលសម្របខ្លួនព្រោះវា "សម្អាត" ហ្សែនពីការផ្លាស់ប្តូរដែលកើតឡើងម្តងហើយម្តងទៀតខណៈពេលដែលយោងទៅតាមសម្មតិកម្មមួយផ្សេងទៀត (អេកូឡូស៊ី) ការរួមភេទគឺ
គំនិតដំបូងអំពីការរីកចម្រើន និងការអភិវឌ្ឍន៍ មានតាំងពីសម័យបុរាណមកម្ល៉េះ។ សូម្បីតែ Hippocrates (460-377 មុនគ។ ស។ បន្ទាប់មកគំនិតនេះត្រូវបានបង្កើតឡើងជាគោលលទ្ធិនៃ preformationism (ពី lat. ទម្រង់មុន - preformation) ដែលបានក្លាយជាការពេញនិយមជាពិសេសនៅក្នុងសតវត្សទី 17-18 ។ អ្នកគាំទ្រនៃ preformationism គឺ Harvey, Malpighi និងអ្នកជីវវិទូនិងគ្រូពេទ្យលេចធ្លោជាច្រើនទៀតនៅសម័យនោះ។ សម្រាប់ preformationists បញ្ហាចម្រូងចម្រាសគឺមានតែនៅក្នុងកោសិកាផ្លូវភេទដែលសារពាង្គកាយត្រូវបាន preform - ស្រីឬបុរស។ អ្នកដែលចូលចិត្តស៊ុតត្រូវបានគេហៅថា ovists ហើយអ្នកដែលយកចិត្តទុកដាក់កាន់តែខ្លាំងចំពោះកោសិកាបន្តពូជរបស់បុរសត្រូវបានគេហៅថាសត្វ។ Preformationism គឺជាគោលលទ្ធិ metaphysical ពីដើមដល់ចប់ ព្រោះវាបដិសេធការអភិវឌ្ឍន៍។ ការប៉ះទង្គិចយ៉ាងមុតមាំចំពោះ preformationism ត្រូវបានដោះស្រាយដោយ C. Bonnet (1720-1793) ដែលបានរកឃើញ parthenogenesis ក្នុងឆ្នាំ 1745 ដោយប្រើឧទាហរណ៍នៃការវិវត្តនៃ aphids ពីស៊ុតដែលមិនទាន់បង្កកំណើត។ បន្ទាប់ពីនេះ ការកែទម្រង់មុនមិនអាចស្តារឡើងវិញបានទៀតទេ ហើយចាប់ផ្តើមបាត់បង់សារៈសំខាន់របស់វា។
នៅសម័យបុរាណ លទ្ធិមួយទៀតបានក្រោកឡើងដែលផ្ទុយនឹង preformationism ហើយក្រោយមកបានទទួលឈ្មោះថា epigenesis (មកពីភាសាក្រិច។ epi-បន្ទាប់ពី, ហ្សែន -ការអភិវឌ្ឍន៍) ។ ដូចជា preformationism ដែរ epigenesis ក៏រីករាលដាលនៅក្នុងសតវត្សទី 17-18 ។ នៅក្នុងការរីករាលដាលនៃ epigenesis ទស្សនៈរបស់ K.F. គឺមានសារៈសំខាន់ដ៏អស្ចារ្យ។ Wolf (1733-1794) សង្ខេបនៅក្នុងសៀវភៅរបស់គាត់ "ទ្រឹស្តីនៃការអភិវឌ្ឍន៍" (1759) ។ K.F. ចចកបានជឿថាស៊ុតមិនមានសារពាង្គកាយដែលបានបង្កើតមុន ឬផ្នែករបស់វាទេ ហើយថាវាមានម៉ាស់ដូចគ្នាដំបូង។ ផ្ទុយទៅនឹង preformists ទស្សនៈរបស់ K.F. ចចក និងអ្នកគាំទ្រដទៃទៀតនៃ epigenesis ត្រូវបានរីកចម្រើនសម្រាប់ពេលវេលារបស់ពួកគេ ដោយសារតែពួកគេមានគំនិតនៃការអភិវឌ្ឍន៍។ ទោះយ៉ាងណាក៏ដោយ ពេលវេលាថ្មីៗបានលេចចេញមក។ ជាពិសេសនៅឆ្នាំ 1828 លោក K. Baer បានបោះពុម្ពផ្សាយស្នាដៃរបស់គាត់ "The History of Animal Development" ដែលក្នុងនោះគាត់បានបង្ហាញថា ខ្លឹមសារនៃស៊ុតមានភាពខុសប្លែកគ្នា ពោលគឺមានរចនាសម្ព័ន្ធ ហើយកម្រិតនៃរចនាសម្ព័ន្ធកើនឡើងនៅពេលដែលអំប្រ៊ីយ៉ុងមានការរីកចម្រើន។ ដូច្នេះ K. Baer បានបង្ហាញពីភាពមិនស៊ីសង្វាក់នៃទាំង preformationism និង epigenesis ។
ការលូតលាស់ និងការអភិវឌ្ឍន៍ គឺជាទ្រព្យសម្បត្តិដ៏សំខាន់បំផុតនៃភាវៈរស់។ ការលូតលាស់គឺជាការកើនឡើងនៃបរិមាណនៃសារពាង្គកាយទាំងមូលដែលជាលទ្ធផលនៃការកើនឡើងនៃចំនួនកោសិកា ខណៈពេលដែលការអភិវឌ្ឍន៍គឺជាការផ្លាស់ប្តូរគុណភាពនៅក្នុងរាងកាយដែលត្រូវបានកំណត់ដោយភាពខុសគ្នានៃកោសិកា និង morphogenesis ធានានូវការផ្លាស់ប្តូររីកចម្រើននៅក្នុងបុគ្គលដោយចាប់ផ្តើមពីស៊ុត។ និងបញ្ចប់ដោយរដ្ឋពេញវ័យរបស់ពួកគេ។
Ontogenesis (មកពីភាសាក្រិក។ នៅលើ -សត្វ, ហ្សែន -ការអភិវឌ្ឍន៍) គឺជាប្រវត្តិ (វដ្ដ) នៃការអភិវឌ្ឍន៍របស់បុគ្គលម្នាក់ ដោយចាប់ផ្តើមពីការបង្កើតកោសិកាមេរោគ ដែលបង្កើតវាឡើង និងបញ្ចប់ដោយការស្លាប់របស់វា។ គំនិតអំពី ontogeny គឺផ្អែកលើទិន្នន័យស្តីពីការលូតលាស់ ការអភិវឌ្ឍន៍ និងភាពខុសគ្នា។ ការសិក្សាអំពីគោលការណ៍ជាមូលដ្ឋាននៃ ontogenesis គឺមានសារៈសំខាន់សម្រាប់ការយល់ដឹងអំពីជីវវិទ្យារបស់មនុស្ស។
ទិន្នន័យទំនើបស្តីពីការអភិវឌ្ឍនៃសារពាង្គកាយបដិសេធទាំង preformationism និង epigenesis ។ នៅក្នុងក្របខ័ណ្ឌនៃគំនិតទំនើប ការអភិវឌ្ឍន៍នៃសារពាង្គកាយមួយត្រូវបានគេយល់ថាជាដំណើរការដែលរចនាសម្ព័ន្ធដែលបានបង្កើតឡើងមុននេះជំរុញឱ្យមានការអភិវឌ្ឍនៃរចនាសម្ព័ន្ធជាបន្តបន្ទាប់។ ដំណើរការអភិវឌ្ឍន៍ត្រូវបានកំណត់តាមហ្សែន និងទាក់ទងយ៉ាងជិតស្និទ្ធជាមួយបរិស្ថាន។ អាស្រ័យហេតុនេះ ការអភិវឌ្ឍន៍ត្រូវបានកំណត់ដោយការរួបរួមនៃកត្តាខាងក្នុង និងខាងក្រៅ។
ឯកភាពនៃកំណើន និងការអភិវឌ្ឍន៍
ការលូតលាស់នៃសារពាង្គកាយមួយគឺជាការកើនឡើងបន្តិចម្តងៗនៃម៉ាស់របស់វា និងការផ្លាស់ប្តូររូបរាងជាលទ្ធផលនៃការកើនឡើងនៃចំនួនកោសិកា និងភាពខុសគ្នារបស់វា ការបង្កើតជាលិកា និងសរីរាង្គ និងការផ្លាស់ប្តូរជីវគីមីនៅក្នុងកោសិកា និងជាលិកា។ ដូច្នេះការលូតលាស់គឺជាលទ្ធផលនៃការផ្លាស់ប្តូរបរិមាណនៅក្នុងទម្រង់នៃការកើនឡើងនៃចំនួនកោសិកា (ទំងន់រាងកាយ) និងការផ្លាស់ប្តូរគុណភាពនៅក្នុងទម្រង់នៃភាពខុសគ្នានៃកោសិកា និង morphogenesis ។ ភាពខុសគ្នានៃកោសិកាគឺជាដំណើរការដែលកោសិកាមួយចំនួនក្លាយជា morphologically, biochemically, និងមុខងារខុសគ្នាពីកោសិកាផ្សេងទៀត។ ការបន្តពូជ និងភាពខុសគ្នានៃកោសិកាមួយចំនួនតែងតែសម្របសម្រួលជាមួយការលូតលាស់ និងភាពខុសគ្នានៃកោសិកាផ្សេងទៀត។ ដំណើរការទាំងពីរនេះកើតឡើងពេញមួយវដ្តជីវិតទាំងមូលនៃសារពាង្គកាយ។ ដោយសារកោសិកាខុសគ្នាផ្លាស់ប្តូររូបរាងរបស់ពួកគេ ហើយក្រុមនៃកោសិកាត្រូវបានចូលរួមនៅក្នុងការផ្លាស់ប្តូររូបរាង នេះត្រូវបានអមដោយ morphogenesis ដែលជាសំណុំនៃដំណើរការដែលកំណត់រចនាសម្ព័ន្ធនៃកោសិកា និងជាលិកា ក៏ដូចជា morphology ទូទៅនៃសារពាង្គកាយ។
ការលូតលាស់អាចត្រូវបានវាស់ដោយការបង្កើតខ្សែកោងនៃទំហំរាងកាយ ទម្ងន់ ម៉ាស់ស្ងួត ចំនួនកោសិកា មាតិកាអាសូត និងសូចនាករផ្សេងទៀតដោយផ្អែកលើលទ្ធផលនៃការវាស់វែង។
ONTOGENESIS និងប្រភេទរបស់វា។ PERIODIZATION នៃ ONTOGENESIS
Ontogenesis អាស្រ័យលើធម្មជាតិនៃការអភិវឌ្ឍនៃសារពាង្គកាយត្រូវបានចាត់ថ្នាក់ទៅជាដោយផ្ទាល់ និងដោយប្រយោល ហើយដូច្នេះភាពខុសគ្នាមួយត្រូវបានធ្វើឡើងរវាងការអភិវឌ្ឍន៍ដោយផ្ទាល់ និងដោយប្រយោល។ ការអភិវឌ្ឍន៍ដោយផ្ទាល់នៃសារពាង្គកាយនៅក្នុងធម្មជាតិកើតឡើងក្នុងទម្រង់នៃការវិវត្តន៍មិនមែនដង្កូវ និងក្នុងស្បូន ចំណែកការអភិវឌ្ឍន៍ដោយប្រយោលត្រូវបានគេសង្កេតឃើញក្នុងទម្រង់នៃការអភិវឌ្ឍន៍ដង្កូវ។ ផ្ទុយទៅនឹង ontogeny ប្រភេទប្រភេទគឺ phylogeny ។
ការអភិវឌ្ឍដង្កូវ។ ការអភិវឌ្ឍន៍នេះត្រូវបានគេយល់ថាជាការអភិវឌ្ឍន៍ដោយប្រយោល ដោយសារសារពាង្គកាយមានដំណាក់កាលដង្កូវមួយ ឬច្រើនក្នុងការអភិវឌ្ឍន៍របស់វា។ ការវិវឌ្ឍន៍របស់ដង្កូវគឺជាតួយ៉ាងសម្រាប់សត្វល្អិត អំពែរ និងអេក្វាឌែម។ ដង្កូវនៃសត្វទាំងនេះដឹកនាំរបៀបរស់នៅឯករាជ្យបន្ទាប់មកឆ្លងកាត់ការផ្លាស់ប្តូរ។ ដូច្នេះការអភិវឌ្ឍន៍នេះត្រូវបានគេហៅថាការអភិវឌ្ឍន៍ជាមួយ metamorphosis (សូមមើលខាងក្រោម) ។
ការអភិវឌ្ឍមិនមែនដង្កូវ។ ទម្រង់នៃការអភិវឌ្ឍន៍នេះគឺជាលក្ខណៈនៃសារពាង្គកាយដែលអភិវឌ្ឍដោយផ្ទាល់ ឧទាហរណ៍ ត្រី សត្វល្មូន និងសត្វស្លាប ដែលពងរបស់វាសម្បូរទៅដោយ yolk (សារធាតុចិញ្ចឹម)។ ដោយសារតែនេះផ្នែកសំខាន់នៃ ontogeny កើតឡើងនៅក្នុងស៊ុតដាក់នៅក្នុងបរិយាកាសខាងក្រៅ។ ការរំលាយអាហាររបស់អំប្រ៊ីយ៉ុងត្រូវបានធានាដោយការអភិវឌ្ឍសរីរាង្គបណ្តោះអាសន្នដែលជាភ្នាសអំប្រ៊ីយ៉ុង (ថង់ yolk, amnion, allantois) ។
ការអភិវឌ្ឍន៍ពោះវៀន។ ការអភិវឌ្ឍន៍នេះក៏ជាលក្ខណៈនៃសារពាង្គកាយដែលអភិវឌ្ឍដោយផ្ទាល់ ដូចជាថនិកសត្វ រួមទាំងមនុស្សផងដែរ។ ដោយសារស៊ុតនៃសារពាង្គកាយទាំងនេះមានសារធាតុចិញ្ចឹមតិចតួចណាស់ មុខងារសំខាន់ៗទាំងអស់របស់អំប្រ៊ីយ៉ុងត្រូវបានផ្តល់ដោយរាងកាយមាតា តាមរយៈការបង្កើតសរីរាង្គបណ្តោះអាសន្នពីជាលិការបស់ម្តាយ និងអំប្រ៊ីយ៉ុង ដែលក្នុងនោះផ្នែកសំខាន់គឺសុក។ តាមការវិវត្តន៍ ការវិវឌ្ឍន៍នៃស្បូនគឺជាទម្រង់ចុងក្រោយបំផុត ប៉ុន្តែវាមានប្រយោជន៍បំផុតសម្រាប់អំប្រ៊ីយ៉ុង ព្រោះវាធានាបាននូវការរស់រានមានជីវិតរបស់ពួកគេ។
Ontogenesis ត្រូវបានបែងចែកទៅជារយៈពេល proembryonic, អំប្រ៊ីយ៉ុង និង postembryonic ។ ក្នុងករណីមនុស្សរយៈពេលនៃការវិវឌ្ឍមុនពេលសម្រាលត្រូវបានគេហៅថាមុនពេលសម្រាល ឬសម្រាលមុនពេលសម្រាលក្រោយសម្រាល។ អំប្រ៊ីយ៉ុងដែលកំពុងលូតលាស់មុនពេលបង្កើត rudiments សរីរាង្គត្រូវបានគេហៅថា អំប្រ៊ីយ៉ុង បន្ទាប់ពីការបង្កើត rudiments សរីរាង្គ - ទារក។
ការអភិវឌ្ឍន៍ PROEMBRYONAL
រយៈពេលនេះនៅក្នុងការអភិវឌ្ឍបុគ្គលនៃសារពាង្គកាយត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការបង្កើត gametes នៅក្នុងដំណើរការនៃការ gametogenesis ។ កោសិកាបន្តពូជរបស់បុរសមិនមានភាពខុសគ្នាខ្លាំងពីកោសិកាផ្សេងទៀតទេ ខណៈពេលដែលស៊ុតមានភាពខុសគ្នាត្រង់ថាវាមានផ្ទុក yolk ច្រើន។ ដោយគិតពីបរិមាណនៃ yolk និងការចែកចាយរបស់វានៅក្នុងស៊ុត ក្រោយមកទៀតត្រូវបានចាត់ថ្នាក់ជាបីប្រភេទ៖
1) ស៊ុត isolecithal, មាន yolk តិចតួច ដែលត្រូវបានធ្វើមូលដ្ឋានីយកម្ម រាបស្មើនៅទូទាំងកោសិកា។ ស៊ុតទាំងនេះត្រូវបានផលិតដោយ echinoderms ( urchins សមុទ្រ), chordates ទាប (lancelets) និងថនិកសត្វ;
2) ស៊ុត telolecithal មានផ្ទុកនូវបរិមាណដ៏ច្រើននៃ yolk ដែលត្រូវបានប្រមូលផ្តុំនៅបង្គោលមួយ - បន្លែ។ ស៊ុតបែបនេះត្រូវបានផលិតដោយ mollusks, amphibians, សត្វល្មូន និងបក្សី។ ឧទាហរណ៍ស៊ុតកង្កែបមាន yolk 50% ស៊ុតមាន់ (ជាទូទៅពងមាន់) - 95% ។ នៅបង្គោលផ្សេងទៀត (សត្វ) នៃស៊ុត telolecithal, cytoplasm និង nucleus ត្រូវបានប្រមូលផ្តុំ;
3) កោសិកាកណ្តាល oocytes, ដែលក្នុងនោះមាន yolk តិចតួចហើយវាកាន់កាប់ទីតាំងកណ្តាល។ cytoplasm ស្ថិតនៅលើបរិវេណនៃស៊ុតបែបនេះ។ ស៊ុត Centrolecithal ត្រូវបានផលិតដោយ arthropods ។
រយៈពេល proembryonic ក៏ត្រូវបានកំណត់លក្ខណៈដោយការពិតដែលថាក្នុងអំឡុងពេលនេះដំណើរការមេតាប៉ូលីសដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការប្រមូលផ្តុំនៃម៉ូលេគុល DNA កើតឡើងនៅក្នុង gametes ។
ការអភិវឌ្ឍន៍អំប្រ៊ីយ៉ុង
Embryogenesis (មកពីភាសាក្រិក។ អេបប្រាយ - germ) ឬរយៈពេលនៃអំប្រ៊ីយ៉ុង ចាប់ផ្តើមជាមួយនឹងការបញ្ចូលគ្នានៃកោសិកាមេជីវិតឈ្មោល និងស្ត្រី ដែលតំណាងឱ្យដំណើរការនៃការបង្កកំណើតរបស់ស៊ុត។
នៅក្នុងសារពាង្គកាយដែលត្រូវបានកំណត់ដោយការវិវឌ្ឍក្នុងស្បូន រយៈពេលនៃអំប្រ៊ីយ៉ុងបញ្ចប់ដោយការចាប់កំណើត ហើយនៅក្នុងសារពាង្គកាយដែលកំណត់លក្ខណៈដោយប្រភេទនៃការលូតលាស់របស់ដង្កូវ និងមិនមែនដង្កូវនោះ រយៈពេលនៃអំប្រ៊ីយ៉ុងបញ្ចប់ដោយការបញ្ចេញសារពាង្គកាយពីស៊ុត ឬភ្នាសអំប្រ៊ីយ៉ុងរៀងៗខ្លួន។ ក្នុងអំឡុងពេលអំប្រ៊ីយ៉ុងដំណាក់កាលនៃ zygote, ការបំបែក, blastula, ការបង្កើតស្រទាប់មេរោគ, histogenesis និង organogenesis ត្រូវបានសម្គាល់។
ហ្សីហ្គោត។ការបង្កកំណើតពាក់ព័ន្ធនឹងដំណើរការជាបន្តបន្ទាប់ដែលកោសិកាបន្តពូជរបស់បុរសចាប់ផ្តើមបង្កើតពងមួយ។ នៅក្នុងស៊ុតដែលត្រូវបានធ្វើឱ្យសកម្មដោយ gamete បុរស ដំណើរការរាងកាយ និងគីមីមួយចំនួនកើតឡើង រួមទាំងការបង្កើនការសំយោគប្រូតេអ៊ីន។ ចលនានៃ protoplasm នាំឱ្យមានការបង្កើតស៊ីមេទ្រីទ្វេភាគីនៃស៊ុត។ នុយក្លេអ៊ែ និងសំណុំក្រូម៉ូសូម diploid ត្រូវបានស្ដារឡើងវិញ។ នេះបង្កើតសារពាង្គកាយកោសិកាតែមួយ។
ការបំបែក។វាតំណាងឱ្យរយៈពេលដំបូងនៃការអភិវឌ្ឍនៃ zygote (ស៊ុតបង្កកំណើត) ដែលមានការបែងចែក zygote តាមរយៈ mitosis ។ ការបែងចែកចាប់ផ្តើមជាមួយនឹងរូបរាងនៃ furrow នៅលើផ្ទៃនៃស៊ុត។ furrow ដំបូងនាំឱ្យមានការបង្កើតកោសិកាពីរ - blastomeres ពីរ, ទីពីរ - blastomeres បួន, ទីបី - ប្រាំបី blastomeres (រូបភាព 25) ។ ក្រុមនៃកោសិកាដែលបានបង្កើតឡើងជាលទ្ធផលនៃការបែងចែកជាបន្តបន្ទាប់ត្រូវបានគេហៅថា morula (មកពីឡាតាំង។ អនុស្សាវរីយ៍- មៀន) ។
សារៈសំខាន់ជីវសាស្រ្តនៃដំណាក់កាលនេះស្ថិតនៅក្នុងការពិតដែលថាពីកោសិកាធំដែលជាស៊ុតកោសិកាតូចៗត្រូវបានបង្កើតឡើងដែលសមាមាត្រនៃ cytoplasm ទៅ nucleus ត្រូវបានកាត់បន្ថយ។
ការបែកខ្ញែកនៃ zygote បញ្ចប់ដោយការបង្កើតរចនាសម្ព័ន្ធពហុកោសិកាហៅថា blastula (មកពីភាសាក្រិក។ blastos -ពន្លក) ។ រចនាសម្ព័ន្ធនេះមានរាងដូច vesicle ហៅថា blastoderm ដែលមានស្រទាប់តែមួយនៃកោសិកា។ ឥឡូវនេះកោសិកាទាំងនេះត្រូវបានគេហៅថាអំប្រ៊ីយ៉ុង។ blastula មានទំហំប្រហាក់ប្រហែលនឹងស៊ុត។ ក្នុងអំឡុងពេលនៃការបែកខ្ញែក ចំនួនស្នូល និងចំនួនសរុបនៃ DNA កើនឡើង។ បរិមាណតិចតួចនៃ mRNA និង tRNA ក៏ត្រូវបានសំយោគផងដែរ ខណៈដែល ribosomal RNA មិនទាន់អាចរកឃើញនៅឡើយ។
សត្វទាំងអស់ឆ្លងកាត់ដំណាក់កាល blastula ប៉ុន្តែករណីនីមួយៗមានលក្ខណៈផ្ទាល់ខ្លួនរបស់វា។ នៅក្នុងថនិកសត្វ ការបែងចែកគឺមិនស្មើគ្នា ដូច្នេះ morulae មានកោសិកាខុសៗគ្នា។ លើសពីនេះទៀតផ្នែកមួយនៃកោសិកាបង្កើតរចនាសម្ព័ន្ធហៅថា trophoblast ។

Blastocoel Larvaកូនទា
ទម្រង់
អង្ករ។ ២៥.ការបែកខ្ញែកនៃ zygotes និងការបង្កើត blastulas នៅក្នុងសារពាង្គកាយផ្សេងៗគ្នា៖ 1 - ស៊ុតដើម; 2 - blastomeres ពីរ; 3 - blastomeres បួន; 4 - blastomeres ប្រាំបី; 5 - blastula; 6 - ទម្រង់មនុស្សពេញវ័យ
កោសិកាដែលចិញ្ចឹមអំប្រ៊ីយ៉ុង ហើយអរគុណដល់អង់ស៊ីម ធានានូវការជ្រៀតចូលរបស់ក្រោយចូលទៅក្នុងជញ្ជាំងស្បូន។ ក្រោយមក កោសិកា trophoblast របូតចេញពីអំប្រ៊ីយ៉ុង ហើយបង្កើតជា vesicle ដែលពោរពេញទៅដោយសារធាតុរាវចេញពីជាលិកាស្បូន។
សារៈសំខាន់ជីវសាស្រ្តនៃដំណាក់កាលនេះស្ថិតនៅក្នុងការពិតដែលថាពីកោសិកាធំដូចជាស៊ុតកោសិកាតូចៗត្រូវបានបង្កើតឡើងដែលសមាមាត្រនៃ cytoplasm ទៅ nucleus ត្រូវបានកាត់បន្ថយ ហើយស្នូលមានបរិយាកាស cytoplasmic ថ្មី។
ក្រពះ(មកពីភាសាក្រិក ក្រពះ -បែហោងធ្មែញនាវា) ។ នេះគឺជាដំណើរការនៃចលនានៃកោសិកាអំប្រ៊ីយ៉ុងបន្ទាប់ពីការកកើតនៃ blastula ដែលត្រូវបានអមដោយការបង្កើតស្រទាប់ពីរឬបី (អាស្រ័យលើប្រភេទសត្វ) នៃស្រទាប់អំប្រ៊ីយ៉ុង ឬហៅថាស្រទាប់មេរោគ (រូបភាពទី 26)។ )
ការអភិវឌ្ឍ (gastrulation) នៃស៊ុត isolecithal កើតឡើងដោយការ invagination (invagination) នៃបង្គោលលូតលាស់នៅខាងក្នុង blastula ដែលជាលទ្ធផលដែលប៉ូលផ្ទុយគ្នាស្ទើរតែបញ្ចូលគ្នា ហើយ blastocoel (បែហោងធ្មែញ blastula) ស្ទើរតែបាត់ឬទាំងស្រុង។ ខាងក្រៅ
អង្ករ។ ២៦.Gastrulation នៅក្នុងសារពាង្គកាយផ្សេងៗ៖ 1 - blastocoel; 2 - mesenchyme បឋម; 3, 4, 5 - ពោះវៀនបឋម
ស្រទាប់នៃកោសិកាអំប្រ៊ីយ៉ុងត្រូវបានគេហៅថា ectoderm (មកពីភាសាក្រិក។ ectos -នៅខាងក្រៅ, ស្បែក -ស្បែក) ឬស្រទាប់មេរោគខាងក្រៅ ចំណែកឯផ្នែកខាងក្នុងគឺជា endoderm (មកពីភាសាក្រិច។ អ៊ីនតូស -ខាងក្នុង) ឬស្រទាប់មេរោគខាងក្នុង។ បែហោងធ្មែញដែលបង្កើតឡើងក្នុងករណីនេះត្រូវបានគេហៅថា gastrocoel ឬពោះវៀនបឋមដែលជាច្រកចូលដែលត្រូវបានគេហៅថា blastopore (មាត់បឋម) ។
ការអភិវឌ្ឍនៃស្រទាប់មេរោគពីរគឺជាលក្ខណៈនៃអេប៉ុង និង coelenterates ។ ទោះជាយ៉ាងណាក៏ដោយ chordates ក្នុងអំឡុងពេល gastrulation ត្រូវបានកំណត់ដោយការអភិវឌ្ឍនៃស្រទាប់មេរោគទីបី - mesoderm (ពីភាសាក្រិក។ មេសូស -កណ្តាល) បង្កើតឡើងរវាង ectoderm និង endoderm ។
Gastrulation គឺជាតម្រូវការជាមុនចាំបាច់សម្រាប់ដំណាក់កាលបន្តបន្ទាប់នៃការអភិវឌ្ឍន៍ ព្រោះវានាំកោសិកាចូលទៅក្នុងទីតាំងដែលធ្វើឱ្យវាអាចបង្កើតសរីរាង្គបាន។ សារធាតុអំប្រ៊ីយ៉ុងដែលបែងចែកទៅជារន្ធអំប្រ៊ីយ៉ុងចំនួន 3 ផ្តល់នូវការកើនឡើងដល់ជាលិកា និងសរីរាង្គទាំងអស់នៃអំប្រ៊ីយ៉ុងដែលកំពុងលូតលាស់។
ការអភិវឌ្ឍន៍ (ភាពខុសគ្នា) នៃស្រទាប់មេរោគត្រូវបានអមដោយការបង្កើតជាលិកា និងសរីរាង្គផ្សេងៗពីពួកគេ។ ជាពិសេស, អេពីដេមីនៃស្បែក, ក្រចកនិងសក់, ក្រពេញ sebaceous និងញើស, ប្រព័ន្ធសរសៃប្រសាទ (ខួរក្បាល, ខួរឆ្អឹងខ្នង, ganglia, សរសៃប្រសាទ), កោសិកាទទួលនៃសរីរាង្គអារម្មណ៍, កញ្ចក់ភ្នែក, epithelium នៃមាត់។ , ប្រហោងច្រមុះ និងរន្ធគូថ, ធ្មេញ -
enamel enamel ។ ពី endoderm នេះ epithelium នៃបំពង់អាហារ, ក្រពះ, ពោះវៀន, ថង់ទឹកប្រមាត់, trachea, ទងសួត, សួត, បង្ហួរនោម, ក៏ដូចជាថ្លើម, លំពែង, ទីរ៉ូអ៊ីត, ក្រពេញ Parathyroid និង thymus អភិវឌ្ឍ។ ពី mesoderm អភិវឌ្ឍសាច់ដុំរលោង, គ្រោងឆ្អឹងនិងសាច់ដុំបេះដូង, dermis, ជាលិកាភ្ជាប់, ឆ្អឹងនិងឆ្អឹងខ្ចី, ធ្មេញធ្មេញ, ឈាមនិងសរសៃឈាម, mesentery, តម្រងនោម, ពងស្វាសនិងអូវែ។ នៅក្នុងមនុស្ស ខួរក្បាល និងខួរឆ្អឹងខ្នង គឺជាមនុស្សដំបូងគេដែលបំបែកចេញពីគ្នា។ បន្ទាប់ពី 2 ខែរចនាសម្ព័ន្ធរាងកាយស្ទើរតែទាំងអស់លេចឡើង។ ការតភ្ជាប់នៃអំប្រ៊ីយ៉ុងជាមួយបរិស្ថានត្រូវបានអនុវត្តតាមរយៈសរីរាង្គបណ្តោះអាសន្ន។ Organogenesis បញ្ចប់នៅចុងបញ្ចប់នៃរយៈពេលអំប្រ៊ីយ៉ុង។ ប្រសិនបើការបើកមាត់ច្បាស់លាស់ត្រូវបានបង្កើតឡើងនៅកន្លែងនៃមាត់បឋម (blastopore) បន្ទាប់មកសត្វទាំងនេះត្រូវបានគេហៅថា protostomes (ដង្កូវនាង mollusks, arthropods) ។
ប្រសិនបើមាត់ច្បាស់លាស់ត្រូវបានបង្កើតឡើងនៅកន្លែងផ្ទុយគ្នានោះសត្វទាំងនេះត្រូវបានគេហៅថា deuterostomes (echinoderms, chordates) ។
ដើម្បីធានាបាននូវការតភ្ជាប់នៃអំប្រ៊ីយ៉ុងជាមួយនឹងបរិស្ថាន អ្វីដែលគេហៅថាសរីរាង្គបណ្តោះអាសន្នត្រូវបានប្រើដែលមានជាបណ្តោះអាសន្ន។ អាស្រ័យលើប្រភេទស៊ុត សរីរាង្គបណ្តោះអាសន្នមានរចនាសម្ព័ន្ធខុសៗគ្នា។ នៅក្នុងត្រី សត្វល្មូន និងសត្វស្លាប សរីរាង្គបណ្តោះអាសន្នរួមមានថង់ yolk ។ នៅក្នុងថនិកសត្វ ថង់ yolk ត្រូវបានបង្កើតឡើងនៅដើមដំបូងនៃ embryogenesis ប៉ុន្តែមិនមានការវិវឌ្ឍន៍ទេ។ ក្រោយមកវាត្រូវបានកាត់បន្ថយ។ ភ្នាសខាងក្រៅនៃអំប្រ៊ីយ៉ុងត្រូវបានគេហៅថា chorion ។ វាដុះចូលទៅក្នុងស្បូន។ កន្លែងនៃការលូតចូលស្បូនខ្លាំងបំផុតត្រូវបានគេហៅថា សុក។ អំប្រ៊ីយ៉ុងត្រូវបានភ្ជាប់ទៅនឹងសុកតាមរយៈទងផ្ចិត ឬទងផ្ចិតដែលមានសរសៃឈាមដែលផ្តល់ចរន្តឈាមសុក។ ការរំលាយអាហាររបស់ទារកត្រូវបានធានាតាមរយៈសុក។
អន្តរកម្មទ្រង់ទ្រាយនៃផ្នែកនៃអំប្រ៊ីយ៉ុងគឺផ្អែកលើដំណើរការមេតាបូលីសដែលបានសម្របសម្រួលជាក់លាក់។ គំរូនៃការអភិវឌ្ឍន៍គឺ heterochrony ដែលត្រូវបានគេយល់ថាជាការបង្កើតផ្សេងគ្នានៃ anlages សរីរាង្គតាមពេលវេលានិងអាំងតង់ស៊ីតេផ្សេងគ្នានៃការអភិវឌ្ឍរបស់ពួកគេ។ សរីរាង្គនិងប្រព័ន្ធដែលគួរចាប់ផ្តើមដំណើរការមុននឹងអភិវឌ្ឍលឿនជាងមុន។ ជាឧទាហរណ៍ ចំពោះមនុស្ស ឫសគល់នៃអវយវៈខាងលើមានការរីកចម្រើនលឿនជាងផ្នែកខាងក្រោម។
អំប្រ៊ីយ៉ុងមានភាពរសើបខ្លាំងចំពោះឥទ្ធិពលផ្សេងៗ។ ដូច្នេះ រយៈពេលដ៏សំខាន់ត្រូវបានសម្គាល់ ពោលគឺរយៈពេលដែលអំប្រ៊ីយ៉ុងមានភាពរសើបបំផុតចំពោះកត្តាបំផ្លាញ។ នៅក្នុងករណីនៃមនុស្ស, រយៈពេលដ៏សំខាន់នៃ ontogenesis អំប្រ៊ីយ៉ុង
គឺជាថ្ងៃដំបូងបន្ទាប់ពីការបង្កកំណើត ពេលវេលានៃការបង្កើតសុក និងការសម្រាលកូន។
ស្នូលនៃកោសិកា somatic មានសមត្ថភាពធានានូវការអភិវឌ្ឍធម្មតានៃស៊ុត ដែលត្រូវបានបញ្ជាក់ឱ្យច្បាស់លាស់នៅក្នុងការពិសោធន៍លើការប្តូរស្នូលកោសិកា somatic ទៅជាស៊ុតដែលខ្វះស្នូល។
ការពិសោធន៍ក៏បានបង្ហាញផងដែរថា ការបំប្លែងអំប្រ៊ីយ៉ុង blastomere តែមួយពី 8- និង 16-cell embryos នៃចៀមនៃពូជមួយចូលទៅក្នុង anucleate ពាក់កណ្តាលនៃស៊ុត (បន្ទាប់ពីកាត់ពាក់កណ្តាលចុងក្រោយ) នៃពូជមួយផ្សេងទៀតត្រូវបានអមដោយការអភិវឌ្ឍន៍នៃអំប្រ៊ីយ៉ុងដែលអាចសម្រេចបាន។ និងកំណើតនៃកូនចៀម។
ចាប់តាំងពីសម័យ Hippocrates (សតវត្សទី 5 មុនគ្រឹស្តសករាជ) សំណួរនៃហេតុផលដែលចាប់ផ្តើមកំណើតនៃទារកត្រូវបានពិភាក្សា។ ជាពិសេស Hippocrates ខ្លួនគាត់ផ្ទាល់បានស្នើថា ការអភិវឌ្ឍន៍គភ៌ចាប់ផ្តើមកំណើតរបស់វា។ ការងារពិសោធន៍ចុងក្រោយបំផុតដោយអ្នកស្រាវជ្រាវជនជាតិអង់គ្លេសដែលអនុវត្តលើចៀម បានបង្ហាញថានៅក្នុងចៀមដែលចាប់ផ្តើមនៃការចិញ្ចឹមចៀមត្រូវបានគ្រប់គ្រងដោយស្មុគស្មាញមួយ។ អ៊ីប៉ូតាឡាមូស + ក្រពេញភីតូរីស + ក្រពេញ Adrenal របស់ទារក។ការខូចខាតដល់ស្នូលអ៊ីប៉ូតាឡាម ការដកយកចេញនូវស្រទាប់ខាងមុខនៃក្រពេញភីតូរីស ឬក្រពេញ Adrenal ពន្យារការមានផ្ទៃពោះរបស់ចៀម។ ផ្ទុយទៅវិញ ការគ្រប់គ្រងអរម៉ូន adenocorticotropic (ការសំងាត់ពីក្រពេញភីតូរីសមុន) ឬ cortisol (ការសំងាត់ពីក្រពេញ Adrenal) ដល់សត្វចៀម កាត់បន្ថយរយៈពេលនៃការមានផ្ទៃពោះ។
ដូច្នេះនៅក្នុងដំណើរការនៃការអភិវឌ្ឍនៃ eukaryotes ខ្ពស់ដែលជាកោសិកាហ្សីហ្គោតជីជាតិតែមួយក្នុងអំឡុងពេលនៃការអភិវឌ្ឍបន្ថែមទៀតដែលជាលទ្ធផលនៃ mitosis ផ្តល់នូវការកើនឡើងដល់កោសិកានៃប្រភេទផ្សេងគ្នា - epithelial, សរសៃប្រសាទ, ឆ្អឹង, កោសិកាឈាមនិងផ្សេងទៀតដែលត្រូវបានកំណត់លក្ខណៈដោយ ភាពខុសគ្នានៃ morphology និងសមាសភាព macromolecular ។ ទោះបីជាយ៉ាងណាក៏ដោយ វាក៏ជាលក្ខណៈនៃកោសិកានៃប្រភេទផ្សេងៗគ្នា ដែលពួកវាមានសំណុំហ្សែនដូចគ្នា ប៉ុន្តែមានឯកទេសខ្ពស់ អនុវត្តមុខងារជាក់លាក់មួយ ឬច្រើន ពោលគឺហ្សែនខ្លះ "ដំណើរការ" នៅក្នុងកោសិកា ខណៈពេលដែលកោសិកាខ្លះទៀតអសកម្ម។ ឧទាហរណ៍,

អង្ករ។ ២៧. ការអភិវឌ្ឍកូនភ្លោះ monozygotic
មានតែកោសិកាឈាមក្រហមប៉ុណ្ណោះដែលជាក់លាក់ក្នុងការសំយោគ និងផ្ទុកអេម៉ូក្លូប៊ីន។ ដូចគ្នាដែរ មានតែកោសិកាអេពីដេមឺមប៉ុណ្ណោះដែលសំយោគ keratin ។ ដូច្នេះសំណួរបានកើតឡើងជាយូរមកហើយអំពីអត្តសញ្ញាណហ្សែននៃស្នូលនៃកោសិកា somatic និងអំពីយន្តការគ្រប់គ្រងនៃការបង្កើតស៊ុតបង្កកំណើត ដែលជាតម្រូវការជាមុនសម្រាប់ការយល់ដឹងអំពីយន្តការនៃភាពខុសគ្នានៃកោសិកា។ កោសិកាដើមមានសារៈសំខាន់ខ្លាំងក្នុងការបែងចែកកោសិកា (សូមមើលខាងក្រោម)។ ការពិសោធន៍លើសត្វកណ្តុរបានបង្ហាញថាកោសិកានៃប្រភេទមួយមានសមត្ថភាពបំប្លែងទៅជាកោសិកានៃប្រភេទមួយផ្សេងទៀត។ ជាពិសេស កោសិកាថ្លើមដែលមានលក្ខណៈខុសប្លែកគ្នា ឬខុសប្លែកគ្នាត្រូវបានបង្ហាញថា បំប្លែងទៅជាកោសិកាលំពែង។
ចាប់តាំងពីទសវត្សរ៍ទី 50 ។ សតវត្សទី XX នៅក្នុងមន្ទីរពិសោធន៍ជាច្រើន ការពិសោធន៍ត្រូវបានអនុវត្តលើការផ្ទេរស្នូលកោសិកា somatic ចូលទៅក្នុងស៊ុតដោយជោគជ័យ ដោយបានដកហូតស្នូលរបស់ពួកគេដោយសិប្បនិម្មិត។ ការសិក្សាអំពី DNA ពីស្នូលនៃកោសិកាផ្សេងគ្នា បានបង្ហាញថា ស្ទើរតែគ្រប់ករណីទាំងអស់ ហ្សែនមានសំណុំដូចគ្នានៃលំដាប់នៃគូនុយក្លេអូទីត។ មានករណីលើកលែងដែលគេស្គាល់ថា កោសិកាឈាមក្រហមរបស់ថនិកសត្វបាត់បង់ស្នូលរបស់ពួកគេក្នុងដំណាក់កាលចុងក្រោយនៃភាពខុសគ្នា។ ប៉ុន្តែនៅពេលនេះអាងហែលទឹក
អេម៉ូក្លូប៊ីន mRNAs មួយចំនួនត្រូវបានសំយោគរួចហើយ ដូច្នេះស្នូលលែងត្រូវការដោយកោសិកាឈាមក្រហមទៀតហើយ។ ឧទាហរណ៍ផ្សេងទៀតរួមមាន immunoglobulin និងហ្សែន T-cell ដែលត្រូវបានកែប្រែកំឡុងពេលអភិវឌ្ឍ។
ដំណាក់កាលសំខាន់មួយក្នុងទិសដៅនៃការយល់ដឹងអំពីយន្តការគ្រប់គ្រងនៃ ontogenesis អំប្រ៊ីយ៉ុងគឺជាលទ្ធផលនៃការពិសោធន៍ដែលបានធ្វើឡើងនៅក្នុងទសវត្សរ៍ឆ្នាំ 1960-1970 ។ អ្នកស្រាវជ្រាវជនជាតិអង់គ្លេស D. Gurdon ដើម្បីស្វែងយល់ថាតើស្នូលនៃកោសិកា somatic មានសមត្ថភាពធានាដល់ការវិវត្តន៍នៃស៊ុតបន្ថែមទៀតដែរឬទេ ប្រសិនបើស្នូលទាំងនេះត្រូវបានបញ្ចូលទៅក្នុងស៊ុតដែលស្នូលរបស់ពួកគេត្រូវបានដកចេញពីមុន។ នៅក្នុងរូបភព។ រូបភាពទី 28 បង្ហាញពីដ្យាក្រាមនៃការពិសោធន៍មួយក្នុងចំណោមការពិសោធន៍ទាំងនេះ ដែលស្នូលនៃកោសិកា somatic tadpole ត្រូវបានប្តូរទៅក្នុងស៊ុតកង្កែបជាមួយនឹងស្នូលដែលបានដកចេញពីមុន។ ការពិសោធន៍ទាំងនេះបានបង្ហាញថា ស្នូលនៃកោសិកា somatic ពិតជាអាចធានាដល់ការវិវឌ្ឍន៍បន្ថែមទៀតនៃស៊ុត ដោយសារតែពួកគេអាចបង្កកំណើតស៊ុត និង "បង្ខំ" ពួកវាឱ្យអភិវឌ្ឍបន្ថែមទៀត។ នេះបានបង្ហាញពីលទ្ធភាពនៃការក្លូនសត្វ។

អង្ករ។ ២៨.គ្រោងការណ៍នៃការពិសោធន៍លើការប្តូរកោសិកា somatic nuclei ចូលទៅក្នុងស៊ុត anucleated (D. Gurdon, 1968)
ក្រោយមក អ្នកស្រាវជ្រាវផ្សេងទៀតបានធ្វើការពិសោធន៍ ដែលពួកគេបានបង្ហាញថា ការផ្ទេរ blastomeres បុគ្គលពី 8- និង 16-day អំប្រ៊ីយ៉ុងចៀមនៃពូជមួយចូលទៅក្នុងពាក់កណ្តាលនៃស៊ុត (បន្ទាប់ពីកាត់ពាក់កណ្តាលចុងក្រោយ) នៃពូជមួយផ្សេងទៀតត្រូវបានអមដោយ ការបង្កើតអំប្រ៊ីយ៉ុងដែលអាចសម្រេចបានជាមួយនឹងកំណើតជាបន្តបន្ទាប់នៃកូនចៀម។
នៅដើមឆ្នាំ 1997 អ្នកនិពន្ធភាសាអង់គ្លេសបានបង្កើតឡើងថា ការដាក់បញ្ចូលកោសិកា somatic nuclei (កោសិកានៃអំប្រ៊ីយ៉ុង ទារកគភ៌ ឬស្បូនរបស់ចៀមពេញវ័យ) ចូលទៅក្នុងស៊ុតចៀមដែលផ្ដាច់ដោយសិប្បនិម្មិត ហើយបន្ទាប់មកការផ្សាំពងដែលបង្កកំណើតទៅក្នុងស្បូនរបស់ចៀមត្រូវបានអមដោយ ការកើតឡើងនៃការមានផ្ទៃពោះតាមពីក្រោយដោយកំណើតនៃកូនចៀម (រូបភាព 29) ។ កូនចៀមមួយក្បាលមានឈ្មោះថា Dolly។ នៅឆ្នាំ 2003 Dolly បានស្លាប់។ ក្នុងអំឡុងពេលនេះ អំប្រ៊ីយ៉ុងរបស់សត្វកណ្ដុរ គោ ទន្សាយ សេះ កណ្តុរ និងសត្វដទៃទៀតត្រូវបានទទួល។

ការណែនាំអំពីវប្បធម៌ចូលទៅក្នុងខ្លួនរបស់ម្តាយ
អង្ករ។ ២៩.ការប្តូរស្នូលកោសិកា somatic ចូលទៅក្នុងស៊ុត anucleated
ការវាយតម្លៃនៃលទ្ធផលទាំងនេះបង្ហាញថា ថនិកសត្វអាចបន្តពូជដោយភេទដូចគ្នា បង្កើតកូនចៅរបស់សត្វដែលកោសិកាមានសារធាតុនុយក្លេអ៊ែរនៃប្រភពដើមពីមាតា ឬមាតា អាស្រ័យលើភេទរបស់ចៀមម្ចាស់ជំនួយ។ នៅក្នុងកោសិកាបែបនេះ
មានតែ cytoplasm និង mitochondria ប៉ុណ្ណោះដែលមានដើមកំណើតពីម្តាយ។ ការសន្និដ្ឋាននេះមានសារៈសំខាន់ខ្លាំងណាស់សម្រាប់ជីវសាស្រ្តទូទៅ និងពង្រីកទស្សនៈរបស់យើងលើសក្តានុពលបន្តពូជរបស់សត្វ។ ប៉ុន្តែវាមានសារៈសំខាន់ផងដែរក្នុងការបន្ថែមថាយើងកំពុងនិយាយអំពីឧបាយកលហ្សែនដែលមិនមាននៅក្នុងធម្មជាតិ។ ម្យ៉ាងវិញទៀត ក្នុងន័យជាក់ស្តែង ឧបាយកលហ្សែនទាំងនេះតំណាងឱ្យវិធីផ្ទាល់នៃការក្លូនសត្វដែលមានការរៀបចំខ្ពស់ជាមួយនឹងលក្ខណៈសម្បត្តិដែលចង់បាន ដែលមានសារៈសំខាន់ខាងសេដ្ឋកិច្ច។ នៅក្នុងពាក្យវេជ្ជសាស្រ្ត ផ្លូវនេះអាចត្រូវបានប្រើប្រាស់នៅពេលអនាគត ដើម្បីយកឈ្នះលើភាពគ្មានកូនរបស់បុរស។
ដូច្នេះព័ត៌មានហ្សែនដែលចាំបាច់សម្រាប់ការអភិវឌ្ឍធម្មតានៃអំប្រ៊ីយ៉ុងមិនត្រូវបានបាត់បង់ក្នុងអំឡុងពេលនៃភាពខុសគ្នានៃកោសិកាដោយសារតែកោសិកាដើមហៅថាកោសិកាដែលមានសក្តានុពលក្នុងការវិវត្តទៅជាប្រភេទផ្សេងៗនៃកោសិការាងកាយ។ នៅពេលដែលកោសិកាដើមបែងចែក កោសិកាថ្មីនីមួយៗមានសក្តានុពលក្នុងការនៅតែជាកោសិកាដើម ឬក្លាយជាកោសិកាដែលមានមុខងារពិសេសជាង (កោសិកាសាច់ដុំ កោសិកាឈាម ឬកោសិកាខួរក្បាល)។ ស៊ុតបង្កកំណើតមានផ្ទុកនូវសារធាតុចិញ្ចឹម ព្រោះវាបង្កើតកោសិកាផ្សេងៗក្នុងរាងកាយ។ កោសិកាដើម Totipotent អាចបង្កើតកោសិកាប្រភេទណាមួយ លើកលែងតែកោសិកាដែលចាំបាច់សម្រាប់ការអភិវឌ្ឍគភ៌។ កោសិកាដើម ដែលអាចបង្កើតកោសិកាជាច្រើនប្រភេទ ត្រូវបានគេហៅថាជាទូទៅ ពហុមុខងារកោសិកា។ វាត្រូវបានបង្កើតឡើងថាកោសិកាដើមពេញវ័យអាចបង្កើតកោសិកាផ្សេងគ្នាពីជាលិកាដែលមិនទាក់ទងអំប្រ៊ីយ៉ុង។ កោសិកា Somatic ក៏មានទ្រព្យសម្បត្តិមួយហៅថា totipotency ពោលគឺហ្សែនរបស់ពួកគេផ្ទុកនូវព័ត៌មានទាំងអស់ដែលពួកគេបានទទួលពីស៊ុតបង្កកំណើត ដែលផ្តល់ឱ្យពួកគេកើនឡើងជាលទ្ធផលនៃភាពខុសគ្នា។ វត្តមាននៃទិន្នន័យទាំងនេះច្បាស់ជាមានន័យថា ភាពខុសគ្នានៃកោសិកាគឺស្ថិតនៅក្រោមការគ្រប់គ្រងហ្សែន។ ការសិក្សាអំពីកោសិកាដើមមានផលប៉ះពាល់ដល់ថ្នាំ។
វាត្រូវបានបង្កើតឡើងថាការសំយោគប្រូតេអ៊ីនដែលពឹងផ្អែកខ្លាំងបន្ទាប់ពីការបង្កកំណើតនៅក្នុង eukaryotes ភាគច្រើនមិនត្រូវបានអមដោយការសំយោគ mRNA ទេ។ ការសិក្សាអំពី oogenesis នៅក្នុងសត្វឆ្អឹងកង ជាពិសេសនៅក្នុង amphibians បានបង្ហាញថាការចម្លងដែលពឹងផ្អែកខ្លាំងកើតឡើងសូម្បីតែក្នុងអំឡុងពេល prophase I (ជាពិសេស diplonema) នៃ meiosis ។ ដូច្នេះ ការចម្លងហ្សែនក្នុងទម្រង់ជា mRNA ឬម៉ូលេគុល pro-mRNA ត្រូវបានរក្សាទុកនៅក្នុងស៊ុតក្នុងស្ថានភាពងងុយគេង។ វាត្រូវបានបង្កើតឡើងថានៅក្នុងកោសិកាអំប្រ៊ីយ៉ុងមានអ្វីដែលគេហៅថា asymmetric
ការបែងចែក ternary ដែលមាននៅក្នុងការពិតដែលថាការបែងចែកនៃកោសិកាអំប្រ៊ីយ៉ុងផ្តល់នូវកោសិកាពីរដែលក្នុងនោះមានតែកោសិកាមួយប៉ុណ្ណោះដែលទទួលមរតកប្រូតេអ៊ីនដែលពាក់ព័ន្ធនឹងការចម្លង។ ដូច្នេះ ការចែកចាយមិនស្មើគ្នានៃកត្តាចម្លងរវាងកោសិកាកូនស្រីនាំទៅរកការបង្ហាញនៃសំណុំហ្សែនផ្សេងៗគ្នានៅក្នុងពួកវាបន្ទាប់ពីការបែងចែក ពោលគឺចំពោះភាពខុសគ្នានៃកោសិកា។
នៅក្នុង amphibians និងប្រហែលជា vertebrates ភាគច្រើន កម្មវិធីហ្សែនដែលគ្រប់គ្រងការអភិវឌ្ឍន៍ដំបូង (មុនដំណាក់កាល blastula) ត្រូវបានបង្កើតឡើងកំឡុងពេល oogenesis ។ ដំណាក់កាលបន្ទាប់នៃការអភិវឌ្ឍន៍ នៅពេលដែលភាពខុសគ្នានៃកោសិកាចាប់ផ្តើម (ប្រហែលពីដំណាក់កាល gastrula) ត្រូវការកម្មវិធីថ្មីសម្រាប់ការបញ្ចេញហ្សែន។ ដូច្នេះ ភាពខុសគ្នានៃកោសិកាត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការសរសេរឡើងវិញនៃព័ត៌មានហ្សែនក្នុងទិសដៅមួយឬមួយផ្សេងទៀត។
លក្ខណៈពិសេសនៃភាពខុសគ្នានៃកោសិកាគឺថាវាមិនអាចត្រឡប់ទៅរកប្រភេទកោសិកាមួយឬមួយផ្សេងទៀត។ ដំណើរការនេះត្រូវបានគេហៅថា ការប្តេជ្ញាចិត្តហើយក៏ស្ថិតក្រោមការគ្រប់គ្រងហ្សែនផងដែរ ហើយដូចដែលត្រូវបានសន្មត់ថា ភាពខុសគ្នានៃកោសិកា និងការប្តេជ្ញាចិត្តត្រូវបានគ្រប់គ្រងដោយអន្តរកម្មនៃកោសិកាដោយផ្អែកលើសញ្ញាដែលធ្វើឡើងដោយកត្តាលូតលាស់ peptide តាមរយៈអ្នកទទួល tyrosine kinase ។ ប្រហែលជាមានប្រព័ន្ធបែបនេះជាច្រើន។ មួយក្នុងចំណោមពួកគេគឺថាភាពខុសគ្នានៃកោសិកាសាច់ដុំនិងសរសៃប្រសាទត្រូវបានគ្រប់គ្រងដោយ neuroregulins ដែលជាប្រូតេអ៊ីនភ្នាសដែលធ្វើសកម្មភាពតាមរយៈអ្នកទទួល tyrosine kinase មួយឬច្រើន។
ការគ្រប់គ្រងហ្សែននៃការប្តេជ្ញាចិត្តក៏ត្រូវបានបង្ហាញដោយអត្ថិភាពនៃការផ្លាស់ប្តូរដែលហៅថា homeiotropic ឬ homeotic mutations ដែលត្រូវបានបង្ហាញនៅក្នុងសត្វល្អិតដើម្បីបង្កឱ្យមានការផ្លាស់ប្តូរក្នុងការកំណត់នៅក្នុងឌីសស្រមើលស្រមៃជាក់លាក់។ ជាលទ្ធផលផ្នែកខ្លះនៃរាងកាយមានការវិវឌ្ឍន៍ចេញពីកន្លែង។ ឧទាហរណ៍នៅក្នុង Drosophila ការផ្លាស់ប្តូរផ្លាស់ប្តូរការកំណត់នៃឌីសអង់តែនទៅជាឌីសដែលវិវត្តទៅជាឧបសម្ព័ន្ធនៃអវយវៈដែលលាតសន្ធឹងពីក្បាល។ នៅក្នុងសត្វល្អិតនៃ genus កែវភ្នែករចនាសម្ព័ន្ធស្លាបអាចវិវត្តចេញពីឌីសភ្នែក។ នៅក្នុងសត្វកណ្តុរ អត្ថិភាពនៃចង្កោមហ្សែន (ស្មុគស្មាញ) Hox ត្រូវបានបង្ហាញដែលមាន 38 ហ្សែន និងគ្រប់គ្រងការវិវត្តនៃអវយវៈ។
បញ្ហានៃបទប្បញ្ញត្តិនៃសកម្មភាពហ្សែនក្នុងអំឡុងពេលនៃការអភិវឌ្ឍន៍អំប្រ៊ីយ៉ុងមានសារៈសំខាន់ដោយឯករាជ្យ។ វាត្រូវបានគេជឿថាក្នុងអំឡុងពេលនៃភាពខុសគ្នាហ្សែនធ្វើសកម្មភាពនៅពេលផ្សេងគ្នាដែលត្រូវបានបង្ហាញ
នៅក្នុងការចម្លងនៅក្នុងកោសិកាផ្សេងគ្នានៃ mRNAs ផ្សេងៗគ្នា ពោលគឺការបង្រ្កាប និងការបង្អត់ហ្សែនកើតឡើង។ ឧទាហរណ៍ចំនួនហ្សែនដែលបានចម្លងទៅជា RNA នៅក្នុង blastocytes urchin សមុទ្រគឺ 10% នៅក្នុងកោសិកាថ្លើមកណ្តុរវាក៏ 10% ហើយនៅក្នុងកោសិកា thymus គោក្របីវាមាន 15% ។ វាត្រូវបានគេសន្មត់ថាប្រូតេអ៊ីនដែលមិនមែនជា histone ត្រូវបានចូលរួមនៅក្នុងការគ្រប់គ្រងស្ថានភាពចម្លងនៃហ្សែន។ ទិន្នន័យខាងក្រោមគាំទ្រការសន្មត់នេះ។ នៅពេលដែលកោសិកាក្រូម៉ាទីនស្ថិតនៅក្នុងដំណាក់កាល សបានចម្លងនៅក្នុងប្រព័ន្ធ នៅក្នុង vitro,បន្ទាប់មកមានតែអ៊ីស្តូន mRNA ប៉ុណ្ណោះដែលត្រូវបានសំយោគ បន្ទាប់មកដោយអ៊ីស្តូន។ ផ្ទុយទៅវិញ នៅពេលដែលក្រូម៉ាទីននៃកោសិកាពីដំណាក់កាល β ត្រូវបានប្រើ គ្មានអ៊ីស្តូន mRNA ត្រូវបានសំយោគទេ។ នៅពេលដែលប្រូតេអ៊ីនដែលមិនមែនជាភីស្តូនត្រូវបានយកចេញពីក្រូម៉ាទីននៃដំណាក់កាល β និងត្រូវបានជំនួសដោយប្រូតេអ៊ីនក្រូម៉ូសូមដែលមិនមែនជាអ៊ីស្តូនដែលត្រូវបានសំយោគនៅក្នុង ដំណាក់កាល សបន្ទាប់មកបន្ទាប់ពីការចម្លងនៃ chromatin បែបនេះ នៅក្នុង vitroអ៊ីស្តូន mRNA ត្រូវបានសំយោគ។ លើសពីនេះទៅទៀត នៅពេលដែលប្រូតេអ៊ីនដែលមិនមែនជាអ៊ីស្តូនមានប្រភពចេញពីកោសិកា 1 ដំណាក់កាល ហើយ DNA និងអ៊ីស្តូនមកពីកោសិកា S-phase នោះគ្មានអ៊ីស្តូន mRNA ត្រូវបានសំយោគទេ។ លទ្ធផលទាំងនេះបង្ហាញថាប្រូតេអ៊ីនដែលមិនមែនជាអ៊ីស្តូនដែលមាននៅក្នុងក្រូម៉ាទីនកំណត់សមត្ថភាពក្នុងការចម្លងហ្សែនដែលអ៊ិនកូដអ៊ីស្តូន។ ដូច្នេះជឿថាប្រូតេអ៊ីនក្រូម៉ូសូមដែលមិនមែនជាអ៊ីស្តូនអាចដើរតួយ៉ាងសំខាន់ក្នុងការគ្រប់គ្រង និងការបញ្ចេញហ្សែននៅក្នុង eukaryotes ។
ទិន្នន័យដែលអាចរកបានបង្ហាញថា ប្រូតេអ៊ីន និងអរម៉ូនស្តេរ៉ូអ៊ីតពាក់ព័ន្ធនឹងបទប្បញ្ញត្តិនៃការចម្លងនៅក្នុងសត្វ។ ប្រូតេអ៊ីន (អាំងស៊ុយលីន) និងអរម៉ូនស្តេរ៉ូអ៊ីត (អេស្ត្រូហ្គន និងតេស្តូស្តេរ៉ូន) គឺជាប្រព័ន្ធសញ្ញាពីរដែលប្រើក្នុងទំនាក់ទំនងអន្តរកោសិកា។ នៅក្នុងសត្វខ្ពស់ អ័រម៉ូនត្រូវបានសំយោគនៅក្នុងកោសិកាសម្ងាត់ពិសេស។ បញ្ចេញចូលទៅក្នុងចរន្តឈាមពួកគេចូលទៅក្នុងជាលិកា។ ចាប់តាំងពីម៉ូលេគុលនៃអរម៉ូនប្រូតេអ៊ីនមានទំហំធំល្មម ពួកវាមិនជ្រាបចូលទៅក្នុងកោសិកាទេ ដូច្នេះឥទ្ធិពលរបស់វាត្រូវបានផ្តល់ដោយប្រូតេអ៊ីន receptor ដែលធ្វើមូលដ្ឋានីយកម្មនៅក្នុងភ្នាសនៃកោសិកាគោលដៅ និងកម្រិត intracellular នៃ cyclic AMP (cAMP)។ ផ្ទុយទៅវិញ អ័រម៉ូនស្តេរ៉ូអ៊ីត គឺជាម៉ូលេគុលតូចៗ ដែលជាលទ្ធផលដែលពួកវាងាយជ្រាបចូលទៅក្នុងកោសិកាតាមរយៈភ្នាស។ នៅពេលដែលនៅខាងក្នុងកោសិកាពួកវាភ្ជាប់ទៅនឹងប្រូតេអ៊ីនទទួលជាក់លាក់ដែលត្រូវបានរកឃើញនៅក្នុង cytoplasm នៃកោសិកាគោលដៅប៉ុណ្ណោះ។ ភាពស្មុគស្មាញត្រូវបានពិចារណា អ័រម៉ូន - អ្នកទទួលប្រូតេអ៊ីន,ដោយផ្តោតលើស្នូលនៃកោសិកាគោលដៅ ពួកវាធ្វើឱ្យដំណើរការចម្លងនៃហ្សែនជាក់លាក់តាមរយៈអន្តរកម្មជាមួយជាក់លាក់
mi ប្រូតេអ៊ីនដែលមិនមែនជាអ៊ីស្តូនដែលភ្ជាប់ទៅនឹងតំបន់ផ្សព្វផ្សាយនៃហ្សែនជាក់លាក់។ ជាលទ្ធផលការចងនៃស្មុគស្មាញ អ័រម៉ូន + ប្រូតេអ៊ីន (អ្នកទទួលប្រូតេអ៊ីន)ជាមួយនឹងប្រូតេអ៊ីនដែលមិនមែនជា histone រំដោះតំបន់ផ្សព្វផ្សាយសម្រាប់ចលនានៃ RNA polymerase ។ ដោយសង្ខេបទិន្នន័យស្តីពីការគ្រប់គ្រងហ្សែននៃរយៈពេលអំប្រ៊ីយ៉ុងនៅក្នុង ontogenesis នៃសារពាង្គកាយ យើងអាចសន្និដ្ឋានថា ដំណើររបស់វាត្រូវបានគ្រប់គ្រងដោយការបើក និងបិទឌីផេរ៉ង់ស្យែលនៃសកម្មភាពនៃហ្សែននៅក្នុងកោសិកាផ្សេងៗគ្នា (ជាលិកា) តាមរយៈការធ្លាក់ទឹកចិត្ត និងការបង្ក្រាបរបស់ពួកគេ។
ការវិវឌ្ឍន៍ក្រោយឆ្លងទន្លេ
បន្ទាប់ពីកំណើតនៃសារពាង្គកាយមួយ ការអភិវឌ្ឍន៍ក្រោយកំណើតរបស់វាចាប់ផ្តើម (ក្រោយសម្រាលសម្រាប់មនុស្ស) ដែលនៅក្នុងសារពាង្គកាយផ្សេងៗគ្នាមានរយៈពេលពីច្រើនថ្ងៃទៅរាប់រយឆ្នាំ អាស្រ័យលើប្រភេទរបស់វា។ អាស្រ័យហេតុនេះ អាយុសង្ឃឹមរស់ គឺជាលក្ខណៈប្រភេទនៃសារពាង្គកាយ ដែលមិនអាស្រ័យលើកម្រិតនៃអង្គការរបស់ពួកគេ (សូមមើលខាងក្រោម)។
នៅក្នុង ontogenesis postembryonic ភាពខុសគ្នាមួយត្រូវបានធ្វើឡើងរវាងអនីតិជន និងអំឡុងពេលពេញវ័យ ក៏ដូចជារយៈពេលនៃភាពចាស់ ដែលបញ្ចប់ដោយការស្លាប់។
រយៈពេលអនីតិជន។ រយៈពេលនេះ (ពីឡាតាំង។ អនីតិជន- វ័យក្មេង) ត្រូវបានកំណត់ដោយពេលវេលាពីកំណើតនៃសារពាង្គកាយរហូតដល់ពេញវ័យ។ វាកើតឡើងតាមវិធីផ្សេងៗគ្នា និងអាស្រ័យលើប្រភេទនៃ ontogenesis នៃសារពាង្គកាយ។ រយៈពេលនេះត្រូវបានកំណត់លក្ខណៈដោយការអភិវឌ្ឍន៍ដោយផ្ទាល់ឬដោយប្រយោល។
នៅក្នុងករណីនៃសារពាង្គកាយដែលត្រូវបានកំណត់ដោយការអភិវឌ្ឍផ្ទាល់ (សត្វឆ្អឹងខ្នងជាច្រើន ត្រី សត្វល្មូន សត្វស្លាប ថនិកសត្វ មនុស្ស) ដែលញាស់ចេញពីសំបកស៊ុត ឬទារកទើបនឹងកើតគឺស្រដៀងទៅនឹងទម្រង់មនុស្សពេញវ័យ ខុសពីប្រភេទក្រោយៗទៀតក្នុងទំហំតូចជាងនេះផងដែរ។ ដូចជាការវិវឌ្ឍន៍នៃសរីរាង្គនីមួយៗ និងសមាមាត្ររាងកាយមិនល្អឥតខ្ចោះ (រូបភាព 30) ។
លក្ខណៈពិសេសលក្ខណៈនៃការលូតលាស់ក្នុងអនីតិជននៃសារពាង្គកាយដែលទទួលរងនូវការអភិវឌ្ឍន៍ដោយផ្ទាល់គឺថាមានការកើនឡើងនៃចំនួន និងទំហំនៃកោសិកា និងសមាមាត្រនៃរាងកាយផ្លាស់ប្តូរ។ ការលូតលាស់របស់មនុស្សក្នុងអំឡុងពេលផ្សេងគ្នានៃ ontogenesis របស់គាត់ត្រូវបានបង្ហាញនៅក្នុងរូបភព។ 31. ការលូតលាស់នៃសរីរាង្គមនុស្សផ្សេងគ្នាគឺមិនស្មើគ្នា។ ឧទាហរណ៍ការលូតលាស់ក្បាលបញ្ចប់ក្នុងវ័យកុមារភាពជើងឈានដល់ទំហំសមាមាត្រប្រហែល 10 ឆ្នាំ។ ប្រដាប់បន្តពូជខាងក្រៅលូតលាស់យ៉ាងឆាប់រហ័សនៅចន្លោះអាយុ 12 ទៅ 14 ឆ្នាំ។ ភាពខុសគ្នាមួយត្រូវបានបង្កើតឡើងរវាងកំណើនច្បាស់លាស់ និងគ្មានកំណត់។ ការលូតលាស់ជាក់លាក់គឺជាលក្ខណៈនៃសារពាង្គកាយដែលឈប់លូតលាស់នៅអាយុជាក់លាក់មួយ
ឧទាហរណ៍ សត្វល្អិត ថនិកសត្វ មនុស្ស។ ការលូតលាស់មិនកំណត់គឺជាលក្ខណៈនៃសារពាង្គកាយដែលលូតលាស់ពេញមួយជីវិតរបស់ពួកគេ ឧទាហរណ៍ មច្ឆា ត្រី អំភ្លី សត្វល្មូន និងប្រភេទរុក្ខជាតិជាច្រើន។

អង្ករ។ សាមសិបការអភិវឌ្ឍដោយផ្ទាល់និងដោយប្រយោលនៃសារពាង្គកាយនៃប្រភេទផ្សេងៗគ្នា
នៅក្នុងករណីនៃការអភិវឌ្ឍន៍ដោយប្រយោល សារពាង្គកាយឆ្លងកាត់ការផ្លាស់ប្តូរដែលហៅថា metamorphoses (ពី lat ។ metamorphosis -ការផ្លាស់ប្តូរ)។

អង្ករ។ ៣១.ការលូតលាស់និងការអភិវឌ្ឍន៍ក្នុងរយៈពេលផ្សេងៗគ្នានៃ ontogenesis របស់មនុស្ស
ពួកវាតំណាងឱ្យការកែប្រែនៃសារពាង្គកាយកំឡុងពេលអភិវឌ្ឍ។ Metamorphoses ត្រូវបានរកឃើញយ៉ាងទូលំទូលាយនៅក្នុង coelenterates (hydra, jellyfish, coral polyps), flatworms (fasciola), roundworms ( roundworms), mollusks (oysters, mussels, octopuses), arthropods (crayfish, ក្តាមទន្លេ, បង្កង, បង្គា, ខ្យាដំរី, ពីងពាង, mites ។ សត្វល្អិត) និងសូម្បីតែនៅក្នុង chordates មួយចំនួន (tunicates និង amphibians) ។ ក្នុងករណីនេះការបំប្លែងពេញលេញនិងមិនពេញលេញត្រូវបានសម្គាល់។ ទម្រង់នៃការបំប្លែងសារជាតិដែលបង្ហាញឱ្យឃើញច្រើនបំផុតត្រូវបានគេសង្កេតឃើញនៅក្នុងសត្វល្អិតដែលឆ្លងកាត់ការបំប្លែងសារជាតិមិនពេញលេញ និងពេញលេញ។
ការបំប្លែងមិនពេញលេញគឺជាការវិវឌ្ឍន៍ដែលសារពាង្គកាយមួយផុសចេញពីសំបកស៊ុត ដែលរចនាសម្ព័ន្ធរបស់វាស្រដៀងទៅនឹងរចនាសម្ព័ន្ធរបស់សារពាង្គកាយពេញវ័យ ប៉ុន្តែទំហំរបស់វាតូចជាងច្រើន។ សារពាង្គកាយបែបនេះត្រូវបានគេហៅថា larva ។ ក្នុងអំឡុងពេលនៃការលូតលាស់ និងការអភិវឌ្ឍន៍ ទំហំនៃដង្កូវកើនឡើង ប៉ុន្តែគម្រប chitinized ដែលមានស្រាប់រារាំងការកើនឡើងបន្ថែមទៀតនៃទំហំរាងកាយ ដែលនាំទៅដល់ការរលាយ ពោលគឺការស្រក់នៃគម្រប chitinized ដែលនៅក្រោមនោះមាន cuticle ទន់។ ក្រោយមកទៀតតម្រង់ចេញហើយនេះត្រូវបានអមដោយការកើនឡើងនៃទំហំនៃសត្វ។ បន្ទាប់ពី moults ជាច្រើនសត្វឈានដល់ភាពពេញវ័យ។ ការផ្លាស់ប្តូរមិនពេញលេញគឺជាតួយ៉ាងឧទាហរណ៍នៅក្នុងករណីនៃការអភិវឌ្ឍនៃ bedbugs ។
ការបំប្លែងសារជាតិពេញលេញគឺជាការវិវឌ្ឍន៍ដែលដង្កូវមួយត្រូវបានបញ្ចេញចេញពីសំបកស៊ុត ដែលខុសគ្នាខ្លាំង
នៅក្នុងរចនាសម្ព័ន្ធពីបុគ្គលពេញវ័យ។ ឧទាហរណ៍នៅក្នុងមេអំបៅ និងសត្វល្អិតជាច្រើនដង្កូវគឺជាដង្កូវនាង។ Caterpillars ទទួលរងនូវការ molting ហើយអាច molt ជាច្រើនដងបន្ទាប់មកប្រែទៅជា pupae ។ ពីក្រោយ ទម្រង់មនុស្សពេញវ័យ (imago) មានការរីកចម្រើន ដែលមិនខុសពីទម្រង់ដើម។
នៅក្នុង vertebrates, metamorphosis កើតឡើងក្នុងចំណោម amphibians និង bony fishes ។ ដំណាក់កាលដង្កូវត្រូវបានកំណត់លក្ខណៈដោយវត្តមាននៃសរីរាង្គបណ្តោះអាសន្នដែលមានលក្ខណៈឡើងវិញនៃបុព្វបុរសឬមានសារៈសំខាន់សម្របខ្លួនយ៉ាងច្បាស់។ ជាឧទាហរណ៍ កូនកង្កែបដែលជាទម្រង់ដង្កូវរបស់កង្កែប ហើយធ្វើឡើងវិញនូវលក្ខណៈនៃទម្រង់ដើម ត្រូវបានកំណត់លក្ខណៈដោយរូបរាងដូចត្រី វត្តមាននៃការដកដង្ហើម Gill និងរង្វង់មួយនៃឈាមរត់។ លក្ខណៈពិសេសនៃការសម្របខ្លួនរបស់ tadpoles គឺជញ្ជក់ និងពោះវៀនវែង។ វាក៏ជាលក្ខណៈនៃទម្រង់ដង្កូវដែរ ដែលបើប្រៀបធៀបទៅនឹងទម្រង់មនុស្សពេញវ័យ ពួកវាសម្របខ្លួនទៅនឹងជីវិតក្នុងលក្ខខណ្ឌខុសគ្នាទាំងស្រុង ដោយកាន់កាប់តំបន់អេកូឡូស៊ីផ្សេងគ្នា និងកន្លែងផ្សេងគ្នានៅក្នុងខ្សែសង្វាក់អាហារ។ ឧទាហរណ៍ ដង្កូវកង្កែបមានដង្ហើមជីល ខណៈទម្រង់មនុស្សពេញវ័យមានដង្ហើមសួត។ មិនដូចទម្រង់មនុស្សពេញវ័យដែលជាសត្វស៊ីសាច់ទេ ដង្កូវកង្កែបចិញ្ចឹមលើអាហាររុក្ខជាតិ។
លំដាប់នៃព្រឹត្តិការណ៍នៅក្នុងការអភិវឌ្ឍនៃសារពាង្គកាយត្រូវបានគេហៅថា វដ្តជីវិត ដែលអាចមានលក្ខណៈសាមញ្ញ ឬស្មុគស្មាញ។ វដ្តនៃការអភិវឌ្ឍន៍ដ៏សាមញ្ញបំផុតគឺជាលក្ខណៈនៃថនិកសត្វ នៅពេលដែលសារពាង្គកាយមួយវិវត្តន៍ចេញពីស៊ុតបង្កកំណើត ដែលបង្កើតពងម្តងទៀត។ល។ វដ្ដជីវសាស្រ្តដ៏ស្មុគស្មាញគឺជាវដ្តនៃសត្វដែលត្រូវបានកំណត់ដោយការវិវឌ្ឍន៍ជាមួយនឹងមេតាណុល។ ចំណេះដឹងអំពីវដ្ដជីវសាស្រ្តមានសារៈសំខាន់ជាក់ស្តែង ជាពិសេសក្នុងករណីដែលសារពាង្គកាយជាភ្នាក់ងារបង្ករោគ ឬជាអ្នកផ្ទុកមេរោគនៅក្នុងសត្វ និងរុក្ខជាតិ។
ការអភិវឌ្ឍន៍ និងភាពខុសគ្នាដែលទាក់ទងនឹងការបំប្លែងសារជាតិ គឺជាលទ្ធផលនៃការជ្រើសរើសធម្មជាតិ ដោយសារទម្រង់ដង្កូវជាច្រើន ដូចជាដង្កូវនាង និងកូនកង្កែប ត្រូវបានសម្របខ្លួនទៅនឹងបរិស្ថានបានប្រសើរជាងទម្រង់ផ្លូវភេទពេញវ័យ។
ភាពពេញវ័យ។ រយៈពេលនេះត្រូវបានគេហៅផងដែរថាមានភាពចាស់ទុំហើយវាត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងភាពចាស់ទុំខាងផ្លូវភេទរបស់សារពាង្គកាយ។ ការអភិវឌ្ឍនៃសារពាង្គកាយក្នុងអំឡុងពេលនេះឈានដល់កម្រិតអតិបរមារបស់វា។
ការលូតលាស់និងការអភិវឌ្ឍន៍ក្នុងដំណាក់កាលក្រោយសម្រាលត្រូវបានជះឥទ្ធិពលយ៉ាងខ្លាំងដោយកត្តាបរិស្ថាន។ ចំពោះរុក្ខជាតិ កត្តាកំណត់គឺពន្លឺ សំណើម សីតុណ្ហភាព បរិមាណ និងគុណភាពនៃសារធាតុចិញ្ចឹមនៅក្នុងដី។ សម្រាប់សត្វ ការចិញ្ចឹមត្រឹមត្រូវគឺមានសារៈសំខាន់បំផុត (វត្តមាននៃប្រូតេអ៊ីន កាបូអ៊ីដ្រាត ខ្លាញ់ អំបិលរ៉ែ វីតាមីន មីក្រូធាតុនៅក្នុងចំណី)។ អុកស៊ីសែន សីតុណ្ហភាព ពន្លឺ (ការសំយោគវីតាមីន D) ក៏សំខាន់ផងដែរ។
ការលូតលាស់ និងការអភិវឌ្ឍន៍បុគ្គលនៃសារពាង្គកាយសត្វគឺស្ថិតនៅក្រោមបទប្បញ្ញត្តិ neurohumoral ដោយយន្តការនិយតកម្មកំប្លែង និងសរសៃប្រសាទ។ សារធាតុសកម្មដូចអ័រម៉ូនហៅថា phytohormones ត្រូវបានរកឃើញនៅក្នុងរុក្ខជាតិ។ ក្រោយមកទៀតប៉ះពាល់ដល់មុខងារសំខាន់នៃសារពាង្គកាយរុក្ខជាតិ។
នៅក្នុងកោសិកាសត្វ ក្នុងអំឡុងពេលដំណើរការជីវិត សារធាតុសកម្មគីមីត្រូវបានសំយោគដែលប៉ះពាល់ដល់ដំណើរការជីវិត។ កោសិកាសរសៃប្រសាទនៃសត្វឆ្អឹងខ្នង និងសត្វឆ្អឹងខ្នងផលិតសារធាតុហៅថា neurosecretes ។ ក្រពេញ endocrine ឬខាងក្នុង ក្រពេញបញ្ចេញសារធាតុម្យ៉ាងហៅថា អ័រម៉ូន។ ក្រពេញ endocrine ជាពិសេសអ្នកដែលទាក់ទងនឹងការលូតលាស់និងការអភិវឌ្ឍន៍ត្រូវបានគ្រប់គ្រងដោយ neurosecretions ។ នៅក្នុង arthropods បទប្បញ្ញត្តិនៃការលូតលាស់និងការអភិវឌ្ឍន៍ត្រូវបានបង្ហាញយ៉ាងល្អដោយឥទ្ធិពលនៃអរម៉ូនលើការរលាយ។ ការសំយោគនៃការសំងាត់របស់ដង្កូវដោយកោសិកាត្រូវបានគ្រប់គ្រងដោយអរម៉ូនដែលប្រមូលផ្តុំនៅក្នុងខួរក្បាល។ ក្រពេញពិសេសនៅក្នុង crustaceans ផលិតអរម៉ូនដែលរារាំងការ molting ។ កម្រិតនៃអរម៉ូនទាំងនេះកំណត់ភាពញឹកញាប់នៃការ molting ។ នៅក្នុងសត្វល្អិត បទបញ្ជាអ័រម៉ូននៃភាពចាស់ទុំនៃស៊ុត និង diapause ត្រូវបានបង្កើតឡើង។
នៅក្នុងសត្វឆ្អឹងខ្នង ក្រពេញ endocrine គឺជាក្រពេញ pituitary gland ក្រពេញ pineal gland ក្រពេញទីរ៉ូអ៊ីត ក្រពេញ Parathyroid លំពែង ក្រពេញ Adrenal និង gonads ដែលមានទំនាក់ទំនងយ៉ាងជិតស្និទ្ធជាមួយគ្នា។ ក្រពេញភីតូរីសនៅក្នុងសត្វឆ្អឹងខ្នងផលិតអរម៉ូន gonadotropic ដែលរំញោចសកម្មភាពរបស់ gonads ។ ចំពោះមនុស្ស អ័រម៉ូនភីតូរីស ប៉ះពាល់ដល់ការលូតលាស់។ ជាមួយនឹងកង្វះ ភាពតឿនឹងវិវឌ្ឍន៍ ភាពធំធាត់មានការរីកចម្រើន។ ក្រពេញ pineal ផលិតអរម៉ូនដែលប៉ះពាល់ដល់ការប្រែប្រួលតាមរដូវនៃសកម្មភាពផ្លូវភេទរបស់សត្វ។ អរម៉ូនទីរ៉ូអ៊ីតមានឥទ្ធិពលលើការបំប្លែងសារជាតិនៃសត្វល្អិត និងសត្វអំពិលអំពែក។ នៅក្នុងថនិកសត្វ ការថយចុះនៃក្រពេញទីរ៉ូអ៊ីត នាំឱ្យមានការយឺតយ៉ាវនៃការលូតលាស់ និងការវិវត្តន៍នៃសរីរាង្គប្រដាប់បន្តពូជ។ ចំពោះមនុស្ស ដោយសារបញ្ហាក្រពេញទីរ៉ូអ៊ីត ដំណើរការ ossification និងការលូតលាស់ត្រូវបានពន្យារពេល។
(ភាពតឿ) ភាពពេញវ័យមិនកើតឡើង ការអភិវឌ្ឍន៍ផ្លូវចិត្តឈប់ (cretinism) ។ ក្រពេញ Adrenal ផលិតអរម៉ូនដែលមានឥទ្ធិពលលើការរំលាយអាហារ ការលូតលាស់ និងភាពខុសគ្នានៃកោសិកា។ gonads ផលិតអរម៉ូនភេទដែលកំណត់លក្ខណៈផ្លូវភេទបន្ទាប់បន្សំ។ ការយកចេញនៃ gonads នាំឱ្យមានការផ្លាស់ប្តូរដែលមិនអាចត្រឡប់វិញបាននៅក្នុងលក្ខណៈមួយចំនួន។ ជាឧទាហរណ៍ នៅក្នុងមាន់រងាវ ការលូតលាស់របស់សិតសក់ឈប់ ហើយសភាវគតិផ្លូវភេទត្រូវបាត់បង់។ បុរសដែលត្រូវកាត់ចេញទទួលបានភាពស្រដៀងគ្នាខាងក្រៅទៅនឹងមនុស្សស្រី (ពុកចង្ការ និងសក់មិនដុះលើស្បែក ខ្លាញ់ត្រូវបានដាក់នៅលើទ្រូង និងអាងត្រគាក សាច់ដុំនៃសំឡេងត្រូវបានរក្សាទុក ។ល។)
phytohormones របស់រុក្ខជាតិគឺ auxins, cytokinins និង gibberellins ។ ពួកវាគ្រប់គ្រងការលូតលាស់ និងការបែងចែកកោសិកា ការបង្កើតឫសថ្មី ការលូតលាស់ផ្កា និងលក្ខណៈសម្បត្តិរុក្ខជាតិផ្សេងៗទៀត។
នៅគ្រប់ដំណាក់កាលនៃ ontogenesis សារពាង្គកាយមានសមត្ថភាពក្នុងការស្តារផ្នែករាងកាយដែលបាត់បង់ឬខូច។ ទ្រព្យសម្បត្តិនៃសារពាង្គកាយនេះត្រូវបានគេហៅថា ការបង្កើតឡើងវិញ,ដែលអាចជាសរីរវិទ្យានិងសំណង។
ការបង្កើតឡើងវិញសរីរវិទ្យា - នេះគឺជាការជំនួសផ្នែករាងកាយដែលបាត់បង់ក្នុងអំឡុងពេលនៃជីវិតរបស់រាងកាយ។ ការបង្កើតឡើងវិញនៃប្រភេទនេះគឺជារឿងធម្មតាណាស់នៅក្នុងពិភពសត្វ។ ឧទាហរណ៍នៅក្នុង arthropods វាត្រូវបានតំណាងដោយ molting ដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការលូតលាស់។ នៅក្នុងសត្វល្មូនការបង្កើតឡើងវិញត្រូវបានបង្ហាញនៅក្នុងការជំនួសកន្ទុយនិងជញ្ជីងនៅក្នុងបក្សី - ស្លាបក្រញ៉ាំជើងនិងស្ពឺ។ នៅក្នុងថនិកសត្វ ឧទាហរណ៏នៃការបង្កើតឡើងវិញខាងសរីរវិទ្យាគឺជាការស្រក់ប្រចាំឆ្នាំនៃសត្វក្តាន់ដោយសត្វក្តាន់។
ការបង្កើតឡើងវិញ - នេះគឺជាការស្ដារឡើងវិញនូវផ្នែករាងកាយនៃសារពាង្គកាយមួយដែលត្រូវបានរហែកយ៉ាងធ្ងន់ធ្ងរ។ ការបង្កើតឡើងវិញនៃប្រភេទនេះគឺអាចធ្វើទៅបាននៅក្នុងសត្វជាច្រើនប៉ុន្តែការបង្ហាញរបស់វាខុសគ្នា។ ឧទាហរណ៍ វាជារឿងធម្មតានៅក្នុង hydras ហើយត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការបន្តពូជនៃក្រោយ ចាប់តាំងពីសារពាង្គកាយទាំងមូលបង្កើតឡើងវិញពីផ្នែកមួយ។ នៅក្នុងសារពាង្គកាយផ្សេងទៀត ការបង្កើតឡើងវិញបង្ហាញរាងដោយខ្លួនឯងក្នុងទម្រង់នៃសមត្ថភាពនៃសរីរាង្គនីមួយៗក្នុងការស្តារឡើងវិញបន្ទាប់ពីការបាត់បង់ផ្នែកណាមួយ។ នៅក្នុងមនុស្ស ជាលិកាភ្ជាប់ សាច់ដុំ និងឆ្អឹង មានសមត្ថភាពបង្កើតឡើងវិញបានខ្ពស់។
រុក្ខជាតិនៃប្រភេទជាច្រើនក៏មានសមត្ថភាពបង្កើតឡើងវិញផងដែរ។
ទិន្នន័យស្តីពីការបង្កើតឡើងវិញមានសារៈសំខាន់ដ៏អស្ចារ្យមិនត្រឹមតែនៅក្នុងជីវវិទ្យាប៉ុណ្ណោះទេ។ ពួកវាត្រូវបានគេប្រើយ៉ាងទូលំទូលាយក្នុងវិស័យកសិកម្ម ថ្នាំពេទ្យ ជាពិសេសការវះកាត់។
អាយុចាស់ជាដំណាក់កាលនៃ ontogenesis ។ អាយុចាស់គឺជាដំណាក់កាលចុងក្រោយនៃ ontogenesis សត្វ ហើយរយៈពេលរបស់វាត្រូវបានកំណត់
អាយុកាលសរុប ដែលបម្រើជាលក្ខណៈប្រភេទ និងប្រែប្រួលក្នុងចំណោមសត្វផ្សេងៗគ្នា។ អាយុចាស់ត្រូវបានសិក្សាយ៉ាងត្រឹមត្រូវបំផុតចំពោះមនុស្ស។
មានភាពខុសគ្នានៃនិយមន័យនៃភាពចាស់របស់មនុស្ស។ ជាពិសេស និយមន័យដ៏ពេញនិយមបំផុតមួយគឺថា អាយុចាស់គឺជាការប្រមូលផ្តុំនៃការផ្លាស់ប្តូរជាបន្តបន្ទាប់ដែលអមជាមួយនឹងការកើនឡើងនៃអាយុនៃសារពាង្គកាយ និងបង្កើនលទ្ធភាពនៃជំងឺឬការស្លាប់របស់វា។ វិទ្យាសាស្រ្តនៃភាពចាស់របស់មនុស្សត្រូវបានគេហៅថា gerontology (មកពីភាសាក្រិក។ ហ្គ្រិន -បុរសចំណាស់, បុរសចំណាស់, ឡូហ្គោ -វិទ្យាសាស្ត្រ) ។ ភារកិច្ចរបស់វាគឺដើម្បីសិក្សាគំរូនៃការផ្លាស់ប្តូរអាយុរវាងភាពចាស់ទុំនិងការស្លាប់។
ការស្រាវជ្រាវវិទ្យាសាស្ត្រនៅក្នុង gerontology ពង្រីកទៅផ្នែកផ្សេងៗ ពីការសិក្សាអំពីការផ្លាស់ប្តូរសកម្មភាពនៃអង់ស៊ីមកោសិកា ដល់ការបំភ្លឺពីឥទ្ធិពលនៃការបន្ធូរបន្ថយផ្លូវចិត្ត និងសង្គមក្នុងភាពតានតឹងផ្នែកបរិស្ថានលើអាកប្បកិរិយារបស់មនុស្សចាស់។
នៅក្នុងករណីនៃមនុស្ស, សរីរវិទ្យាអាយុចាស់ត្រូវបានសម្គាល់; អាយុចាស់ដែលទាក់ទងនឹងអាយុប្រតិទិន; និងភាពចាស់មុនអាយុដែលបណ្តាលមកពីកត្តាសង្គម និងជំងឺ។ អនុលោមតាមអនុសាសន៍របស់អង្គការសុខភាពពិភពលោក មនុស្សចាស់គួរត្រូវបានចាត់ទុកថាមានអាយុប្រហែល 60-75 ឆ្នាំ និងចាស់ - 75 ឆ្នាំឬច្រើនជាងនេះ។
ភាពចាស់របស់មនុស្សត្រូវបានកំណត់លក្ខណៈដោយសញ្ញាខាងក្រៅ និងខាងក្នុងមួយចំនួន។
ក្នុងចំណោមសញ្ញាខាងក្រៅនៃភាពចាស់ ការកត់សម្គាល់បំផុតគឺការថយចុះនៃភាពរលូននៃចលនា ការផ្លាស់ប្តូរឥរិយាបថ ការថយចុះនៃការបត់បែនស្បែក ទម្ងន់រាងកាយ ភាពរឹងមាំ និងការបត់បែននៃសាច់ដុំ រូបរាងនៃស្នាមជ្រួញនៅលើមុខ និងផ្នែកផ្សេងទៀតនៃ រាងកាយនិងការបាត់បង់ធ្មេញ។ ដូច្នេះឧទាហរណ៍យោងទៅតាមទិន្នន័យទូទៅមនុស្សម្នាក់នៅអាយុ 30 ឆ្នាំបាត់បង់ធ្មេញ 2 (ជាលទ្ធផលនៃការបាត់បង់) នៅអាយុ 40 ឆ្នាំ - ធ្មេញ 4 នៅអាយុ 50 ឆ្នាំ - 8 ធ្មេញហើយនៅអាយុ 60 ឆ្នាំ - ធ្មេញ ១១ រួចហើយ។ ប្រព័ន្ធសញ្ញាដំបូងឆ្លងកាត់ការផ្លាស់ប្តូរគួរឱ្យកត់សម្គាល់ (ភាពច្បាស់លាស់នៃសរីរាង្គអារម្មណ៍ត្រូវបានរិល) ។ ជាឧទាហរណ៍ ចម្ងាយអតិបរមាដែលមនុស្សដែលមានសុខភាពល្អអាចសម្គាល់សំឡេងដូចគ្នាបានគឺ 12 ម៉ែត្រនៅអាយុ 20-30 ឆ្នាំ 10 ម៉ែត្រនៅអាយុ 50 ឆ្នាំ 7 ម៉ែត្រនៅអាយុ 60 ឆ្នាំ និងត្រឹមតែ 4 ម៉ែត្រនៅអាយុ 70 ឆ្នាំប៉ុណ្ណោះ។ ប្រព័ន្ធសញ្ញាទីពីរ (បំរែបំរួលបំរែបំរួលការនិយាយ សំលេងក្លាយជារិល)។
ក្នុងចំណោមសញ្ញាខាងក្នុង ជាដំបូងយើងគួរនិយាយអំពីសញ្ញាដូចជាការវិវត្តន៍បញ្ច្រាស់ (ការបញ្ចូល) នៃសរីរាង្គ។ មានការថយចុះនៃទំហំនៃថ្លើមនិងតម្រងនោមក៏ដូចជានៅក្នុងចំនួន
ចំនួននៃ nephrons នៅក្នុងតម្រងនោម (នៅអាយុ 80 ឆ្នាំស្ទើរតែពាក់កណ្តាល) ដែលកាត់បន្ថយមុខងារនៃតម្រងនោមនិងប៉ះពាល់ដល់ការរំលាយអាហារទឹក - អេឡិចត្រូលីត។ ភាពយឺតនៃសរសៃឈាមថយចុះ ការបញ្ចូលឈាមនៃជាលិកា និងសរីរាង្គមានការថយចុះ ហើយភាពធន់នឹងសរសៃឈាមគ្រឿងកុំព្យូទ័រកើនឡើង។ អំបិលអសរីរាង្គកកកុញនៅក្នុងឆ្អឹង ការផ្លាស់ប្តូរឆ្អឹងខ្ចី (calcifies) និងសមត្ថភាពនៃសរីរាង្គក្នុងការបង្កើតឡើងវិញមានការថយចុះ។ ការផ្លាស់ប្តូរសំខាន់ៗកើតឡើងនៅក្នុងកោសិកា ការបែងចែក និងការស្ដារឡើងវិញនៃសម្លេងមុខងាររបស់ពួកគេថយចុះ មាតិកាទឹកថយចុះ សកម្មភាពនៃអង់ស៊ីមកោសិកាថយចុះ ការសម្របសម្រួលរវាងការបង្រួម និងការបំបែកកោសិកាត្រូវបានរំខាន។ នៅក្នុងខួរក្បាលការសំយោគប្រូតេអ៊ីនត្រូវបានរំខានដែលបណ្តាលឱ្យមានការបង្កើតប្រូតេអ៊ីនមិនធម្មតា។ viscosity នៃភ្នាសកោសិកាកើនឡើង ការសំយោគ និងការប្រើប្រាស់អរម៉ូនភេទត្រូវបានរំខាន ហើយការផ្លាស់ប្តូរកើតឡើងនៅក្នុងរចនាសម្ព័ន្ធនៃណឺរ៉ូន។ ការផ្លាស់ប្តូររចនាសម្ព័ន្ធកើតឡើងនៅក្នុងប្រូតេអ៊ីននៃជាលិកាភ្ជាប់និងការបត់បែននៃជាលិកានេះ។ ប្រតិកម្មភាពស៊ាំត្រូវបានចុះខ្សោយ ហើយលទ្ធភាពនៃប្រតិកម្មអូតូអ៊ុយមីនកើនឡើង។ មុខងារនៃប្រព័ន្ធ endocrine ជាពិសេស gonads ត្រូវបានកាត់បន្ថយ។ អាកប្បកិរិយានៃសញ្ញាផ្សេងទៀតនៅក្នុងវ័យចាស់ត្រូវបានបង្ហាញនៅក្នុងរូបភព។ ៣២.

អង្ករ។ ៣២.ការផ្លាស់ប្តូរលក្ខណៈរបស់មនុស្សមួយចំនួនទៅតាមអាយុ៖ 1 - ល្បឿននៃការជំរុញសរសៃប្រសាទ; 2 - កម្រិតនៃការរំលាយអាហារមូលដ្ឋាន; 3 - សន្ទស្សន៍បេះដូង; 4 - កម្រិតនៃការច្រោះតំរងនោមសម្រាប់អាំងស៊ុយលីន; 5 - បរិមាណទឹករលកនៃសួត; 6 - កម្រិតនៃលំហូរប្លាស្មានៅក្នុងតម្រងនោម
បំណងប្រាថ្នាចង់យល់ពីធម្មជាតិនៃភាពចាស់នៃរាងកាយមានតាំងពីយូរយារណាស់មកហើយ។ នៅប្រទេសក្រិចបុរាណ Hippocrates ជឿថាភាពចាស់ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការលើសនៃអាហារ និងការប៉ះពាល់នឹងខ្យល់ស្រស់មិនគ្រប់គ្រាន់។ យោងតាម Aristotle ភាពចាស់គឺបណ្តាលមកពីការប្រើប្រាស់ថាមពលកំដៅដោយរាងកាយ។ សារៈសំខាន់នៃអាហារដែលជាកត្តានៃភាពចាស់ក៏ត្រូវបានកត់សម្គាល់ដោយ Galen ផងដែរ។ ប៉ុន្តែអស់រយៈពេលជាយូរមកហើយមិនមានទិន្នន័យវិទ្យាសាស្ត្រគ្រប់គ្រាន់ដើម្បីយល់ច្បាស់អំពីបញ្ហានេះទេ។ មានតែនៅក្នុងសតវត្សទី 19 ប៉ុណ្ណោះ។ មានការរីកចម្រើនខ្លះក្នុងការសិក្សាអំពីភាពចាស់ ហើយទ្រឹស្ដីនៃភាពចាស់បានចាប់ផ្តើមលេចឡើង។
ទ្រឹស្ដីមួយក្នុងចំណោមទ្រឹស្ដីដ៏ល្បីល្បាញបំផុតដំបូងបង្អស់នៃភាពចាស់នៃរាងកាយរបស់មនុស្សគឺទ្រឹស្តីរបស់វេជ្ជបណ្ឌិតជនជាតិអាឡឺម៉ង់ H. Hufeland (1762-1836) ដែលបានភ្ជាប់សារៈសំខាន់ទៅនឹងសកម្មភាពការងារក្នុងភាពជាប់បានយូរ។ យើងបានឮសំដីរបស់គាត់ថា គ្មានមនុស្សខ្ជិលម្នាក់រស់ដល់ចាស់ទេ។ កាន់តែល្បី ទ្រឹស្តី endocrine នៃភាពចាស់,ដែលមានប្រភពមកពីការពិសោធន៍ដែលបានធ្វើឡើងសូម្បីតែនៅពាក់កណ្តាលសតវត្សមុនចុងក្រោយដោយ Berthold (1849) ដែលបានបង្ហាញថាការប្តូរពងស្វាសពីសត្វមួយទៅសត្វមួយទៀតត្រូវបានអមដោយការវិវត្តនៃលក្ខណៈផ្លូវភេទបន្ទាប់បន្សំ។ ក្រោយមក អ្នកជំនាញខាងសរីរវិទ្យាជនជាតិបារាំង C. Brown-Séquard (1818-1894) ដោយផ្អែកលើលទ្ធផលនៃការចាក់ខ្លួនឯងជាមួយនឹងសារធាតុចម្រាញ់ចេញពីពងស្វាស បានអះអាងថា ការចាក់ថ្នាំទាំងនេះផ្តល់ផលមានប្រយោជន៍ និងធ្វើឱ្យមានថាមពលឡើងវិញ។ នៅដើមសតវត្សទី 20 ។ មានជំនឿរួចហើយថាការចាប់ផ្តើមនៃភាពចាស់ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការផុតពូជនៃសកម្មភាពនៃក្រពេញ endocrine ជាពិសេស gonads ។ នៅទសវត្ស 20-30 ។ សតវត្សទី XX ដោយផ្អែកលើជំនឿនេះ ការវះកាត់ជាច្រើនត្រូវបានធ្វើឡើងនៅក្នុងប្រទេសផ្សេងៗគ្នា ដើម្បីធ្វើអោយមនុស្សចាស់ ឬមនុស្សចាស់មានថាមពលឡើងវិញ។ ជាឧទាហរណ៍ G. Steinach នៅប្រទេសអូទ្រីសបានចងខ្សែមេជីវិតឈ្មោលរបស់បុរស ដែលនាំទៅដល់ការបញ្ឈប់ការសំងាត់ខាងក្រៅនៃ gonads ហើយសន្មតថាធ្វើឱ្យមានភាពរស់រវើកឡើងវិញ។ S.A. Voronov នៅប្រទេសបារាំងបានប្តូរពងស្វាសពីសត្វវ័យក្មេងទៅសត្វចាស់និងពីស្វាទៅបុរសហើយ Tushnov នៅសហភាពសូវៀតបានធ្វើឱ្យសត្វមាន់រស់ឡើងវិញដោយចាក់វាជាមួយ histolysates នៃ gonads ។ ប្រតិបត្តិការទាំងអស់នេះនាំឱ្យមានផលប៉ះពាល់មួយចំនួន ប៉ុន្តែបានត្រឹមតែបណ្តោះអាសន្នប៉ុណ្ណោះ។ បន្ទាប់ពីផលប៉ះពាល់ទាំងនេះ ដំណើរការនៃភាពចាស់បានបន្ត ហើយរឹតតែខ្លាំងថែមទៀត។
នៅដើមសតវត្សទី 20 ។ ក្រោកឡើង ទ្រឹស្តីមីក្រូជីវសាស្រ្តនៃភាពចាស់អ្នកបង្កើតនោះគឺ I.I. Mechnikov ដែលបែងចែករវាងអាយុសរីរវិទ្យានិងរោគសាស្ត្រ។ គាត់ជឿថាអាយុចាស់របស់មនុស្សគឺជារោគសាស្ត្រ, i.e. មុនអាយុ។ មូលដ្ឋាននៃគំនិតរបស់ I.I គោលលទ្ធិរបស់ Mechnikov នៃ orthobiosis (អ័រធូស -
ត្រឹមត្រូវ ជីវវិទ្យា - life) យោងទៅតាមមូលហេតុចម្បងនៃភាពចាស់គឺការខូចខាតដល់កោសិកាសរសៃប្រសាទដោយផលិតផល intoxication ដែលត្រូវបានបង្កើតឡើងជាលទ្ធផលនៃ putrefaction នៅក្នុងពោះវៀនធំ។ ការអភិវឌ្ឍគោលលទ្ធិនៃរបៀបរស់នៅធម្មតា (ការប្រតិបត្តិនៃច្បាប់អនាម័យការងារទៀងទាត់ការជៀសវាងពីទម្លាប់អាក្រក់) I.I. Mechnikov ក៏បានស្នើនូវវិធីមួយដើម្បីទប់ស្កាត់បាក់តេរីពោះវៀនដែលបញ្ចេញចោលដោយការទទួលទានផលិតផលទឹកដោះគោដែលមានជាតិ fermented ។
ក្នុងទសវត្សរ៍ទី 30 សតវត្សទី XX បានរីករាលដាល ទ្រឹស្តីអំពីតួនាទីនៃប្រព័ន្ធសរសៃប្រសាទកណ្តាល (CNS) ក្នុងភាពចាស់។អ្នកបង្កើតទ្រឹស្តីនេះគឺ I.P. Pavlov ដែលបានបង្កើតតួនាទីរួមបញ្ចូលគ្នានៃប្រព័ន្ធសរសៃប្រសាទកណ្តាលក្នុងដំណើរការធម្មតានៃសារពាង្គកាយ។ អ្នកដើរតាម I.P. ការពិសោធន៍របស់ Pavlov លើសត្វបានបង្ហាញថា ភាពចាស់មុនអាយុគឺបណ្តាលមកពីការប៉ះទង្គិចផ្លូវចិត្ត និងភាពតានតឹងខាងសរសៃប្រសាទយូរ។
មានតម្លៃនិយាយ ទ្រឹស្តីនៃការផ្លាស់ប្តូរទាក់ទងនឹងអាយុនៅក្នុងជាលិកាភ្ជាប់,បង្កើតនៅទសវត្សរ៍ទី 30 ។ សតវត្សទី XX A.A. Bogomolets (1881-1946) ។ គាត់ជឿថាសកម្មភាពសរីរវិទ្យានៃរាងកាយត្រូវបានធានាដោយជាលិកាភ្ជាប់ (ជាលិកាឆ្អឹង, ឆ្អឹងខ្ចី, សរសៃពួរ, សរសៃចងនិងជាលិកាភ្ជាប់សរសៃ) ហើយការផ្លាស់ប្តូរនៅក្នុងស្ថានភាពនៃកោសិកា colloidal ការបាត់បង់ turgor របស់ពួកគេជាដើម។ កំណត់ការផ្លាស់ប្តូរទាក់ទងនឹងអាយុ។ នៅក្នុងសារពាង្គកាយ។ ទិន្នន័យទំនើបបង្ហាញពីសារៈសំខាន់នៃការប្រមូលផ្តុំជាតិកាល់ស្យូមនៅក្នុងជាលិកាភ្ជាប់ ព្រោះវារួមចំណែកដល់ការបាត់បង់ការបត់បែនរបស់វា ក៏ដូចជាការឡើងរឹងនៃសរសៃឈាម។
វិធីសាស្រ្តទំនើបក្នុងការយល់ដឹងអំពីខ្លឹមសារ និងយន្តការនៃភាពចាស់ត្រូវបានកំណត់លក្ខណៈដោយការប្រើប្រាស់យ៉ាងទូលំទូលាយនៃទិន្នន័យពីជីវវិទ្យាគីមីសាស្ត្រ និងជាពិសេសសមិទ្ធិផលនៃហ្សែនម៉ូលេគុល។ គំនិតទំនើបសាមញ្ញបំផុតអំពីយន្តការនៃភាពចាស់បានពុះកញ្ជ្រោលរហូតដល់ការពិតដែលថាក្នុងអំឡុងពេលនៃជីវិត ការផ្លាស់ប្តូរ somatic កកកុញនៅក្នុងកោសិកានៃរាងកាយ ដែលជាលទ្ធផលដែលការសំយោគប្រូតេអ៊ីនដែលខូច ឬ DNA ដែលមិនជួសជុលបានឆ្លងកាត់តំណភ្ជាប់ជាមួយប្រូតេអ៊ីនកើតឡើង។ ចាប់តាំងពីប្រូតេអ៊ីនដែលមានបញ្ហាដើរតួនាទីបំបែកនៅក្នុងការរំលាយអាហារកោសិកា នេះនាំទៅរកភាពចាស់។ នៅក្នុងករណីនៃ fibroblasts វប្បធម៌ ប្រូតេអ៊ីន និង mRNAs ដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងកោសិកាចាស់ត្រូវបានបង្ហាញថាទប់ស្កាត់ការសំយោគ DNA នៅក្នុង fibroblasts វ័យក្មេង។
វាក៏មានសម្មតិកម្មដែលគេស្គាល់ផងដែរដែលយោងទៅតាមភាពចាស់ត្រូវបានគេចាត់ទុកថាជាលទ្ធផលនៃការផ្លាស់ប្តូរនៃសារធាតុរំលាយអាហារ mitochondrial ជាមួយនឹងការថយចុះមុខងារនៃអង់ស៊ីមជាបន្តបន្ទាប់។
នៅក្នុងមនុស្សអត្ថិភាពនៃហ្សែនត្រូវបានបង្ហាញដែលកំណត់ពេលវេលានៃការវិវត្តនៃដំណើរការ degenerative តំណពូជដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងភាពចាស់។ អ្នកស្រាវជ្រាវមួយចំនួនជឿថា មូលហេតុនៃភាពចាស់គឺការផ្លាស់ប្តូរប្រព័ន្ធការពារភាពស៊ាំរបស់រាងកាយ ជាពិសេសប្រតិកម្មអូតូអ៊ុយមីនចំពោះរចនាសម្ព័ន្ធរាងកាយដែលមានសារៈសំខាន់យ៉ាងសំខាន់។ ជាចុងក្រោយ នៅពេលពន្យល់អំពីយន្តការនៃភាពចាស់ អ្នកជំនាញយកចិត្តទុកដាក់យ៉ាងខ្លាំងចំពោះការខូចខាតប្រូតេអ៊ីនដែលទាក់ទងនឹងការបង្កើតរ៉ាឌីកាល់សេរី។ ជាចុងក្រោយ ជួនកាលសារៈសំខាន់ត្រូវបានភ្ជាប់ទៅនឹង hydrolases ដែលត្រូវបានបញ្ចេញបន្ទាប់ពីការបំបែកនៃ lysosomes ដែលបំផ្លាញកោសិកា។
ទោះជាយ៉ាងណាក៏ដោយ ទ្រឹស្ដីដ៏ទូលំទូលាយនៃភាពចាស់មិនទាន់ត្រូវបានបង្កើតឡើងនៅឡើយទេ ព្រោះវាច្បាស់ណាស់ថា គ្មានទ្រឹស្តីណាមួយអាចពន្យល់ដោយឯករាជ្យអំពីយន្តការនៃភាពចាស់នោះទេ។
ការស្លាប់។ការស្លាប់គឺជាដំណាក់កាលចុងក្រោយនៃ ontogenesis ។ សំណួរនៃការស្លាប់នៅក្នុងជីវវិទ្យាកាន់កាប់កន្លែងពិសេសមួយ ពីព្រោះអារម្មណ៍នៃការស្លាប់ "... គឺជាសភាវគតិទាំងស្រុងនៅក្នុងធម្មជាតិរបស់មនុស្ស ហើយតែងតែជាកង្វល់ដ៏ធំបំផុតរបស់មនុស្ស" (I.I. Mechnikov, 1913) ។ ជាងនេះទៅទៀត សំណួរនៃការស្លាប់គឺជាចំណុចកណ្តាលនៃការយកចិត្តទុកដាក់នៃការបង្រៀនទស្សនវិជ្ជា និងសាសនាទាំងអស់ ទោះបីជាទស្សនវិជ្ជានៃការស្លាប់ត្រូវបានបង្ហាញខុសគ្នាក្នុងសម័យប្រវត្តិសាស្ត្រផ្សេងៗគ្នាក៏ដោយ។ នៅសម័យបុរាណ សូក្រាត និងផ្លាតូបានឈ្លោះប្រកែកគ្នាអំពីភាពអមតៈនៃព្រលឹង ចំណែកអារីស្តូតបានបដិសេធគំនិតរបស់ផ្លាតូអំពីភាពអមតៈនៃព្រលឹង ហើយជឿលើភាពអមតៈនៃព្រលឹងមនុស្ស ដែលបន្តរស់នៅបន្ទាប់ពីការស្លាប់របស់មនុស្ស។ Cicero និង Seneca ក៏ទទួលស្គាល់ជីវិតនាពេលអនាគតដែរ ប៉ុន្តែ Marcus Aurelius បានចាត់ទុកការស្លាប់ជាបាតុភូតធម្មជាតិដែលគួរទទួលយកដោយគ្មានត្អូញត្អែរ។ នៅសតវត្សទី 18 I. Kant និង I. Fichte (1762-1814) ក៏ជឿលើជីវិតអនាគតដែរ ហើយ G. Hegel បានប្រកាន់ខ្ជាប់នូវជំនឿដែលថាព្រលឹងត្រូវបានស្រូបចូលដោយ "ភាពពេញលេញ" ទោះបីជាធម្មជាតិនៃ "ភាព" នេះមិនត្រូវបានបង្ហាញឱ្យដឹងក៏ដោយ។ .
ស្របតាមការបង្រៀនសាសនាដែលគេស្គាល់ទាំងអស់ ជីវិតនៅលើផែនដីរបស់មនុស្សម្នាក់នៅតែបន្តបន្ទាប់ពីការស្លាប់របស់គាត់ ហើយមនុស្សម្នាក់ត្រូវតែត្រៀមខ្លួនដោយមិនចេះនឿយហត់សម្រាប់ការស្លាប់នាពេលអនាគតនេះ។ ទោះជាយ៉ាងណាក៏ដោយ អ្នកវិទ្យាសាស្ត្រធម្មជាតិ និងទស្សនវិទូដែលមិនទទួលស្គាល់ភាពអមតៈបានជឿ ហើយនៅតែជឿថាការស្លាប់គឺដូចដែល I.I. បានសង្កត់ធ្ងន់ម្តងហើយម្តងទៀត។ Mechnikov ដែលជាលទ្ធផលធម្មជាតិនៃជីវិតរបស់សារពាង្គកាយ។ និយមន័យជាន័យធៀបនៃសេចក្តីស្លាប់គឺថាវា “... គឺជាជ័យជម្នះយ៉ាងច្បាស់លាស់នៃភាពគ្មានន័យលើអត្ថន័យ ភាពវឹកវរលើលំហ” (V. Solovyov, 1894)។
ភ័ស្តុតាងវិទ្យាសាស្ត្របង្ហាញថានៅក្នុងសារពាង្គកាយកោសិកាតែមួយ (រុក្ខជាតិ និងសត្វ) ការស្លាប់គួរតែត្រូវបានសម្គាល់ពីការបញ្ចប់។
ចំណេះដឹងអំពីអត្ថិភាពរបស់ពួកគេ។ សេចក្តីស្លាប់គឺជាការបំផ្លិចបំផ្លាញរបស់ពួកគេ ខណៈពេលដែលការបញ្ចប់នៃអត្ថិភាពត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការបែងចែករបស់ពួកគេ។ អាស្រ័យហេតុនេះ ភាពផុយស្រួយនៃសារពាង្គកាយកោសិកាតែមួយត្រូវបានផ្តល់សំណងដោយការបន្តពូជរបស់វា។ នៅក្នុងពហុកោសិកា និងសត្វ ការស្លាប់គឺនៅក្នុងន័យពេញលេញនៃពាក្យថា ចុងបញ្ចប់នៃជីវិតរបស់សារពាង្គកាយ។
ចំពោះមនុស្ស លទ្ធភាពនៃការស្លាប់កើនឡើងក្នុងអំឡុងពេលពេញវ័យ។ នៅក្នុងប្រទេសអភិវឌ្ឍន៍ ជាពិសេស លទ្ធភាពនៃការស្លាប់កើនឡើងស្ទើរតែជានិទស្សន្តបន្ទាប់ពីអាយុ 28 ឆ្នាំ។
មានការស្លាប់ខាងគ្លីនិក និងជីវសាស្រ្តរបស់មនុស្ស។ ការស្លាប់តាមគ្លីនិកត្រូវបានបង្ហាញក្នុងការបាត់បង់ស្មារតី ការបញ្ឈប់ចង្វាក់បេះដូង និងការដកដង្ហើម ប៉ុន្តែកោសិកា និងសរីរាង្គភាគច្រើននៅតែមានជីវិត។ ការបន្តកោសិកាដោយខ្លួនឯងកើតឡើង ហើយការ peristalsis ពោះវៀននៅតែបន្ត។ ការស្លាប់តាមគ្លីនិកមិន "ឈានដល់" ការស្លាប់ដោយជីវសាស្រ្តទេព្រោះវាអាចត្រឡប់វិញបាន ចាប់តាំងពីនៅក្នុងស្ថានភាពនៃការស្លាប់តាមគ្លីនិក មនុស្សម្នាក់អាច "ត្រឡប់" រស់ឡើងវិញបាន។ ឧទាហរណ៍សត្វឆ្កែត្រូវបាន "ត្រលប់មកវិញ" នូវជីវិតបន្ទាប់ពី 5-6 នាទីមនុស្ស - បន្ទាប់ពី 6-7 នាទីពីការចាប់ផ្តើមនៃការស្លាប់គ្លីនិក។ ការស្លាប់ជីវសាស្រ្តត្រូវបានកំណត់លក្ខណៈដោយការពិតដែលថាវាមិនអាចត្រឡប់វិញបាន។ ការបញ្ឈប់ចង្វាក់បេះដូង និងការដកដង្ហើមត្រូវបានអមដោយការបញ្ចប់នៃដំណើរការបន្តដោយខ្លួនឯង ការស្លាប់ និងការរលួយនៃកោសិកា។ ទោះជាយ៉ាងណាក៏ដោយ ការស្លាប់កោសិកាមិនចាប់ផ្តើមនៅក្នុងសរីរាង្គទាំងអស់ក្នុងពេលតែមួយនោះទេ។ ទីមួយ Cortex ខួរក្បាលស្លាប់ បន្ទាប់មកកោសិកា epithelial នៃពោះវៀន សួត ថ្លើម កោសិកាសាច់ដុំ និងបេះដូងស្លាប់។
វិធានការសម្រាប់ការរស់ឡើងវិញ (ការរស់ឡើងវិញ) នៃសារពាង្គកាយគឺផ្អែកលើគំនិតអំពីការស្លាប់តាមគ្លីនិក ដែលមានសារៈសំខាន់ពិសេសក្នុងវេជ្ជសាស្ត្រទំនើប។
អាយុកាល
ការប្រៀបធៀបទិន្នន័យស្តីពីអាយុសង្ឃឹមរស់របស់អ្នកតំណាងនៃរុក្ខជាតិ និងសត្វបង្ហាញថា ក្នុងចំណោមរុក្ខជាតិ និងសត្វ មានសារពាង្គកាយផ្សេងៗគ្នារស់នៅសម្រាប់ពេលវេលាខុសៗគ្នា។ ឧទាហរណ៍ រុក្ខជាតិស្មៅ (ព្រៃ និងដាំដុះ) រស់នៅបានមួយរដូវ។ ផ្ទុយទៅវិញ រុក្ខជាតិឈើរបស់ពួកគេត្រូវបានកំណត់លក្ខណៈដោយអាយុកាលយូរជាង។ ឧទាហរណ៍ cherry រស់នៅ 100 ឆ្នាំ, spruce - 1000 ឆ្នាំ, ដើមឈើអុក -
2000 ឆ្នាំស្រល់ - រហូតដល់ 2000-4000 ឆ្នាំ។
ត្រីនៃប្រភេទសត្វជាច្រើនរស់នៅ 55-80 ឆ្នាំ, កង្កែប - 16 ឆ្នាំ, ក្រពើ - 50-60 ឆ្នាំ, សត្វស្លាបនៃប្រភេទខ្លះ - រហូតដល់ 100 ឆ្នាំ។ អាយុកាលរបស់ថនិកសត្វគឺខ្លីជាង។
ឧទាហរណ៍សត្វចិញ្ចឹមតូចៗរស់នៅ 20-25 ឆ្នាំគោក្របី - 30 ឆ្នាំឬច្រើនជាងនេះសេះ - រហូតដល់ 30 ឆ្នាំឆ្កែ - 20 ឆ្នាំឬច្រើនជាងនេះចចក - 15 ឆ្នាំខ្លាឃ្មុំ - 50 ឆ្នាំដំរី - 100 ឆ្នាំ។ ក្នុងចំណោមថនិកសត្វ មនុស្សគឺជាសត្វដែលរស់នៅបានយូរជាងគេ។ មនុស្សជាច្រើនរស់នៅរហូតដល់ 115-120 ឆ្នាំឬច្រើនជាងនេះ។
អនុលោមតាមគំនិតដែលមានស្រាប់ អាយុសង្ឃឹមរស់គឺជាលក្ខណៈបរិមាណជាក់លាក់មួយប្រភេទដែលត្រូវគ្រប់គ្រងដោយប្រភេទហ្សែន។ វាត្រូវបានគេជឿថាអាយុកាលដែលកំណត់ដោយហ្សែនទាក់ទងទៅនឹងរយៈពេលនៃ ontogenesis (តារាងទី 5) ។
តារាងទី 5 ។ការពឹងផ្អែកនៃកម្រិតនៃ fibroblasts កំឡុងពេលដាំដុះលើអាយុរបស់អ្នកបរិច្ចាគ
វាត្រូវបានសន្មត់ថាអាយុកាលនៃប្រភេទសត្វគឺជាការទទួលបានវិវឌ្ឍន៍នៃប្រភេទសត្វ។ ចំពោះភាពជាប់បានយូររបស់បុគ្គល វាអាចត្រូវបានពន្យល់ដោយការសន្មត់នៃវត្តមាននៃការរួមផ្សំនៃហ្សែនមួយចំនួននៅក្នុង genotypes របស់ពួកគេ ឬចំនួនតិចតួច ឬអវត្តមានពេញលេញនៃការផ្លាស់ប្តូរនៅក្នុងកោសិការបស់ពួកគេ។
A.A. Bogomolets និង I.I. Schmalhausen បានគណនាថាអាយុកាលធម្មជាតិរបស់មនុស្សគួរតែមានពី 120-150 ឆ្នាំ។ ទោះយ៉ាងណាក៏ដោយ មានតែមនុស្សមួយចំនួនប៉ុណ្ណោះដែលនៅរស់ដល់អាយុនេះ។ ដូច្នេះអាយុសង្ឃឹមរស់ពិតប្រាកដមិនស្របនឹងអាយុសង្ឃឹមរបស់ធម្មជាតិទេ។
អាយុសង្ឃឹមរស់ជាមធ្យមត្រូវបានជះឥទ្ធិពលដោយកត្តាមួយចំនួន (ការថយចុះនៃការស្លាប់របស់ទារក ប្រសិទ្ធភាពនៃការគ្រប់គ្រងការឆ្លងមេរោគ ការជឿនលឿនក្នុងការវះកាត់ ការកែលម្អអាហារូបត្ថម្ភ និងលក្ខខណ្ឌរស់នៅទូទៅ)។ មូលហេតុចម្បងនៃការថយចុះនៃអាយុសង្ឃឹមរស់គឺការស្លាប់ដោយសារភាពអត់ឃ្លាន ជំងឺ និងការថែទាំវេជ្ជសាស្រ្តមិនគ្រប់គ្រាន់។ នៅប្រទេសរុស្ស៊ីក្នុងរយៈពេលប៉ុន្មានឆ្នាំថ្មីៗនេះចំនួនប្រជាជនអវិជ្ជមានត្រូវបានគេសង្កេតឃើញនៅក្នុងទីក្រុងនិងតំបន់ជនបទទាំងអស់នៃប្រទេសរុស្ស៊ីដោយសារតែការថយចុះនៃអត្រាកំណើតនិងអត្រាមរណភាពខ្ពស់។
នៅក្នុងពាក្យវេជ្ជសាស្រ្ត អាយុសង្ឃឹមរស់គឺជាសូចនាករនៃសុខភាពរបស់ប្រទេសមួយ។ ស.ស.យ.ក ជាប់ចំណាត់ថ្នាក់លេខ ១ លើពិភពលោក ទាក់ទងនឹងចំនួនមនុស្សចាស់។ ឧទាហរណ៍ ក្នុងចំណោមប្រជាជន 1 លាននាក់ មានមនុស្ស 104 នាក់ដែលមានអាយុលើសពី 90 ឆ្នាំខណៈពេលដែលនៅប្រទេសអង់គ្លេស - 6 នាក់បារាំង - 7 ។
និងសហរដ្ឋអាមេរិក - 15 នាក់។ បច្ចុប្បន្ននេះមានការផ្លាស់ប្តូរនៅក្នុងព្រំដែននៃចំនួនប្រជាជនធ្វើការធៀបនឹង 30s ។ សតវត្សទី XX គម្លាតរវាងអាយុចូលនិវត្តន៍ និងកម្រិតសកម្មភាពរបស់មនុស្សក៏ខុសគ្នាដែរ។ នៅឆ្នាំ 1982 សន្និបាតពិភពលោកស្តីពីបញ្ហានៃភាពចាស់នៃចំនួនប្រជាជនពិភពលោកត្រូវបានប្រារព្ធឡើងនៅទីក្រុងវីយែនដែលការព្យាករណ៍ត្រូវបានបង្កើតឡើងលើបញ្ហានេះរហូតដល់ឆ្នាំ 2025 ។ គេសន្មត់ថាចំនួនមនុស្សដែលមានអាយុចាប់ពី 60 ឆ្នាំឡើងទៅនឹងកើនឡើង 5 ដងបើប្រៀបធៀប។ ដល់ឆ្នាំ 1950 មនុស្សដែលមានអាយុលើសពី 80 ឆ្នាំ - 7 ដង។ យោងតាមវេទិកាអន្តរជាតិនេះ មានភាពខុសប្លែកគ្នានៅក្នុងអត្រានៃភាពចាស់នៃចំនួនប្រជាជនក្នុងចំណោមប្រជាជន ប្រទេស និងតំបន់ផ្សេងៗគ្នា។
Geriatrics គឺជាវិទ្យាសាស្ត្រមួយដែលមានភារកិច្ចបង្កើតវិធីដើម្បីធ្វើឱ្យមុខងារផ្លាស់ប្តូរនៃរាងកាយដែលមានវ័យចំណាស់។ ទោះជាយ៉ាងណាក៏ដោយ ឱសថទំនើបមិនទាន់មានវិធីសាស្រ្ត និងមធ្យោបាយមានឥទ្ធិពលលើដំណើរការសរីរវិទ្យាធម្មតានៅឡើយ។ ដូច្នេះតួនាទីរបស់ geriatrics ត្រូវបានកំណត់ចំពោះការព្យាបាលនៃជំងឺដែលកើតឡើងក្នុងវ័យចាស់ និងការលុបបំបាត់ (ប្រសិនបើអាចធ្វើទៅបាន) នៃកត្តាហានិភ័យដែលបណ្តាលឱ្យមានភាពចាស់មុនអាយុ។
បញ្ហាសម្រាប់ការពិភាក្សា
1. តើអ្នកយល់យ៉ាងណាចំពោះការបន្តពូជផ្លូវភេទរបស់សារពាង្គកាយ ហើយតើវាមានតួនាទីជីវសាស្រ្តអ្វីខ្លះ?
2. ពិពណ៌នាអំពីការបន្តពូជដោយភេទ និងដាក់ឈ្មោះទម្រង់របស់វា។
3. ពិពណ៌នាអំពីលក្ខណៈពិសេសនៃដំណើរការផ្លូវភេទនៅក្នុងសារពាង្គកាយឯកតា និងពហុកោសិកា។
4. តើ gametogenesis ជាអ្វី?
5. តើប្រភេទ gamete នីមួយៗមានមុខងារអ្វីខ្លះ?
6. តើអ្នកដឹងដំណាក់កាលអ្វីខ្លះក្នុងការអភិវឌ្ឍន៍ gametes?
7. តើភាពស្រដៀងគ្នា និងភាពខុសគ្នារវាង spermatogenesis និង oogenesis គឺជាអ្វី?
8. អ្វីទៅជា meiosis ហើយតើអ្វីទៅជាសារៈសំខាន់ជីវសាស្រ្តរបស់វា?
9. ពិពណ៌នាអំពីដំណាក់កាលនៃ meiosis ។
10. តើការឆ្លងកាត់ប៉ះពាល់ដល់លទ្ធផលនៃ meiosis និងយ៉ាងដូចម្តេច?
11. រៀបរាប់ពីខ្លឹមសារនៃការបង្កកំណើត។
12. តើអ្វីជាភាពខុសគ្នារវាង zygogenesis និង parthenogenesis?
13. តើតួនាទីជីវសាស្រ្តនៃការឆ្លាស់គ្នា haploidy និង diploidy គឺជាអ្វី?
14. តើអ្វីជាភាពខុសប្រក្រតីផ្លូវភេទ? តើអ្នកយល់យ៉ាងណាចំពោះការកើតជំងឺហឺត? តើករណីជំងឺក្លនលូនត្រូវបានគេសង្កេតឃើញលើមនុស្សហើយតើញឹកញាប់ប៉ុណ្ណា?
15. តើអ្នកស្រមៃមើលការវិវត្តនៃវិធីសាស្រ្តនៃការបន្តពូជដោយរបៀបណា?
16. តើអ្នកយល់យ៉ាងណាចំពោះការលូតលាស់ និងការអភិវឌ្ឍន៍នៃសារពាង្គកាយ? តើអ្វីជាទំនាក់ទំនងរវាងការលូតលាស់កោសិកា និងភាពខុសគ្នា?
17. តើអ្វីជាមូលដ្ឋានម៉ូលេគុលនៃការបែងចែកកោសិកា? តើអ្នកដឹងអ្វីខ្លះអំពីកោសិកាដើម?
18. បង្កើតគំនិតនៃ ontogenesis និងដាក់ឈ្មោះរយៈពេលនៃ ontogenesis ។
19. តើអ្វីជាភាពខុសគ្នារវាងការអភិវឌ្ឍន៍ដោយផ្ទាល់ និងដោយប្រយោល?
20. តើការបង្កកំណើតមានផលប៉ះពាល់អ្វីខ្លះទៅលើស៊ុត?
21. តើដំណាក់កាលណានៃការអនុវត្តព័ត៌មានហ្សែនគឺការគ្រប់គ្រងសកម្មភាពរបស់ហ្សែនត្រូវបានអនុវត្ត?
22. តើពងដែលបង្កកំណើតទៅជារចនាសម្ព័ន្ធពហុកោសិកាយ៉ាងដូចម្តេច?
23. តើកោសិកា និងជាលិកាដែលកំពុងតែវិវឌ្ឍមានភាពខុសប្លែកពីគ្នាក្នុងពេលកំពុងអភិវឌ្ឍដោយរបៀបណា?
24. តើផ្នែករាងកាយដែលបាត់បង់ ឬខូចអាចស្តារឡើងវិញបានទេ?
25. តើអ្វីជាទំនាក់ទំនងនៃ ontogenesis រវាងអាយុចាស់ និងអាយុសង្ឃឹមរស់?
26. បង្កើត និងកំណត់ភាពខុសគ្នារវាងធម្មជាតិ (ប្រហែល) និងអាយុសង្ឃឹមរស់ជាក់ស្តែង។
27. តើអ្នកដឹងទ្រឹស្ដីអ្វីខ្លះនៃភាពចាស់នៃសារពាង្គកាយ?
29. តើកត្តាអ្វីខ្លះមានឥទ្ធិពលលើអាយុសង្ឃឹមរស់?
30. តើការក្លូនសារពាង្គកាយផ្អែកលើអ្វី? ផ្តល់ឧទាហរណ៍នៃសារពាង្គកាយក្លូន។
31. តើមេជីវិតឈ្មោល ឬខ្លឹមសាររបស់វាជ្រាបចូលទៅក្នុងកោសិកាទេ?
32. តើទិន្នន័យក្លូនបង្ហាញអ្វីខ្លះ?
33. Preformationism and epigenesis in the history of ជីវវិទ្យា។ តើពួកគេមានសារៈសំខាន់ខាងវិទ្យាសាស្ត្រទេ?
34. តើអ្វីជាភាពខុសគ្នារវាងការស្លាប់តាមគ្លីនិក និងជីវសាស្រ្ត?
ការបន្តពូជគឺជាសមត្ថភាពរបស់សារពាង្គកាយដើម្បីបង្កើតប្រភេទរបស់វា។ ការបន្តពូជគឺជាលក្ខណៈសម្បត្តិដ៏សំខាន់បំផុតមួយនៃជីវិត ហើយអាចធ្វើទៅបានដោយសារតែសមត្ថភាពទូទៅនៃសារពាង្គកាយក្នុងការបង្កើតកូនចៅ។ ទោះជាយ៉ាងណាក៏ដោយកូនចៅភ្លាមៗមិនតែងតែស្រដៀងនឹងឪពុកម្តាយរបស់ពួកគេទេ។ ឧទាហរណ៍ពី Sern Spores កូនចៅជាច្រើនដុះឡើងជាកូនចៅដែលមានតំណាងដោយពន្លកដែលមិនស្រដៀងនឹងរុក្ខជាតិស្ពានរបស់ម្តាយ។ នៅលើការរីកដុះដាល, នៅក្នុងវេន, រុក្ខជាតិមិនដូចខ្លួនវាលេចឡើង - sporophyte មួយ។ បាតុភូតនេះត្រូវបានគេហៅថា ការផ្លាស់ប្តូរនៃជំនាន់។
ទម្រង់ពិសេសនៃការបន្តពូជផ្លូវភេទគឺ parthenogenesis ឬការបន្តពូជព្រហ្មចារី ការវិវត្តនៃសារពាង្គកាយពីស៊ុតដែលមិនទាន់បង្កកំណើត។ ទម្រង់នៃការបន្តពូជនេះគឺជាលក្ខណៈចម្បងនៃប្រភេទសត្វដែលមានវដ្ដជីវិតខ្លី ជាមួយនឹងការផ្លាស់ប្តូររដូវច្បាស់លាស់។ Parthenogenesis អាចជា haploid ឬ diploid ។ នៅក្នុង haploid (generative) parthenogenesis សារពាង្គកាយថ្មីមួយបានវិវត្តន៍ចេញពីស៊ុត haploid ។ បុគ្គលដែលទទួលបានលទ្ធផលអាចជាបុរសតែស្ត្រី ឬទាំងពីរនាក់ប៉ុណ្ណោះ។ នេះអាស្រ័យលើការកំណត់ភេទរបស់ក្រូម៉ូសូម។
ការអភិវឌ្ឍន៍បុគ្គលនៃសារពាង្គកាយមួយ ឬ ontogenesis គឺជាសំណុំនៃការផ្លាស់ប្តូរ morphological សរីរវិទ្យា និងជីវគីមីជាបន្តបន្ទាប់ដែលកើតឡើងដោយសារពាង្គកាយចាប់ពីពេលចាប់ផ្តើមរហូតដល់ស្លាប់។ ក្នុងអំឡុងពេល ontogenesis ការអនុវត្តព័ត៌មានតំណពូជដែលទទួលបានដោយរាងកាយពីឪពុកម្តាយរបស់វាកើតឡើង។ មានរយៈពេលសំខាន់ពីរនៅក្នុង ontogenesis - អំប្រ៊ីយ៉ុងនិង postembryonic ។ នៅដំណាក់កាលអំប្រ៊ីយ៉ុង អំប្រ៊ីយ៉ុងមួយត្រូវបានបង្កើតឡើងនៅក្នុងសត្វ ដែលក្នុងនោះប្រព័ន្ធសរីរាង្គសំខាន់ៗត្រូវបានបង្កើតឡើង។ នៅក្នុងអំឡុងពេល postembryonic ដំណើរការអភិវឌ្ឍន៍ត្រូវបានបញ្ចប់ភាពពេញវ័យការបន្តពូជភាពចាស់និងការស្លាប់កើតឡើង។
