जटिल सरल भाषा के बारे में।
यह विषय जटिल और जटिल है, जो हमारे शरीर में होने वाली जैव रासायनिक प्रक्रियाओं की एक बड़ी संख्या को तुरंत प्रभावित करता है। लेकिन आइए अभी भी यह पता लगाने की कोशिश करें कि माइटोकॉन्ड्रिया क्या हैं और वे कैसे काम करते हैं।
और इसलिए, माइटोकॉन्ड्रिया एक जीवित कोशिका के सबसे महत्वपूर्ण घटकों में से एक है। सरल शब्दों में हम कह सकते हैं कि यह सेल पावरहाउस. उनकी गतिविधि मांसपेशियों के संकुचन के कार्यान्वयन के लिए कार्बनिक यौगिकों के ऑक्सीकरण और विद्युत क्षमता (एटीपी अणु के टूटने के दौरान जारी ऊर्जा) के उत्पादन पर आधारित है।
हम सभी जानते हैं कि हमारा शरीर ऊष्मप्रवैगिकी के पहले नियम के अनुसार सख्ती से काम करता है। ऊर्जा हमारे शरीर में निर्मित नहीं होती, केवल रूपांतरित होती है। जीव केवल ऊर्जा परिवर्तन के रूप को चुनता है, इसे उत्पन्न किए बिना, रासायनिक से यांत्रिक और थर्मल तक। पृथ्वी ग्रह पर सभी ऊर्जा का मुख्य स्रोत सूर्य है। प्रकाश के रूप में हमारे पास आकर, ऊर्जा पौधों के क्लोरोफिल द्वारा अवशोषित की जाती है, जहां यह हाइड्रोजन परमाणु के इलेक्ट्रॉन को उत्तेजित करती है और इस प्रकार जीवित पदार्थ को ऊर्जा देती है।
हम अपने जीवन को एक छोटे इलेक्ट्रॉन की ऊर्जा के लिए देते हैं।
माइटोकॉन्ड्रियन का कार्य श्वसन श्रृंखला (प्रोटीन की इलेक्ट्रॉन परिवहन श्रृंखला) के प्रोटीन परिसरों के समूहों में मौजूद धातु परमाणुओं के बीच हाइड्रोजन इलेक्ट्रॉन ऊर्जा के चरणबद्ध हस्तांतरण में होता है, जहां प्रत्येक बाद के परिसर में एक इलेक्ट्रॉन के लिए एक उच्च आत्मीयता होती है, जो इसे आकर्षित करती है। पिछले एक, जब तक इलेक्ट्रॉन आणविक ऑक्सीजन के साथ गठबंधन नहीं करता है, जिसमें उच्चतम इलेक्ट्रॉन आत्मीयता होती है।
हर बार जब एक इलेक्ट्रॉन सर्किट से गुजरता है, तो ऊर्जा निकलती है, जो एक विद्युत रासायनिक ढाल के रूप में जमा होती है और फिर मांसपेशियों के संकुचन और गर्मी रिलीज के रूप में महसूस की जाती है।
माइटोकॉन्ड्रिया में ऑक्सीडेटिव प्रक्रियाओं की एक श्रृंखला जो आपको एक इलेक्ट्रॉन की ऊर्जा क्षमता को स्थानांतरित करने की अनुमति देती है, कहलाती है "इंट्रासेल्युलर श्वसन"या अक्सर "श्वसन श्रृंखला", चूंकि इलेक्ट्रॉन को परमाणु से परमाणु तक श्रृंखला के साथ पारित किया जाता है, जब तक कि वह ऑक्सीजन परमाणु के अपने अंतिम लक्ष्य तक नहीं पहुंच जाता।
ऑक्सीकरण प्रक्रिया के दौरान माइटोकॉन्ड्रिया को ऊर्जा ले जाने के लिए ऑक्सीजन की आवश्यकता होती है।
माइटोकॉन्ड्रिया हमारे द्वारा सांस लेने वाली ऑक्सीजन का 80% तक उपभोग करते हैं।
माइटोकॉन्ड्रिया एक स्थायी कोशिका संरचना है जो इसके साइटोप्लाज्म में स्थित होती है। माइटोकॉन्ड्रिया आमतौर पर 0.5 से 1 माइक्रोन व्यास के होते हैं। आकार में, इसकी एक दानेदार संरचना होती है और यह सेल की मात्रा का 20% तक कब्जा कर सकती है। कोशिका की इस स्थायी कार्बनिक संरचना को ऑर्गेनेल कहा जाता है। ऑर्गेनेल में मायोफिब्रिल्स भी शामिल हैं - मांसपेशी कोशिका की सिकुड़ा इकाइयाँ; और कोशिका केन्द्रक भी एक अंगक है। सामान्य तौर पर, कोई भी स्थायी कोशिका संरचना एक ऑर्गेनेल-ऑर्गेनेल है।
माइटोकॉन्ड्रिया की खोज की गई थी और पहली बार 1894 में जर्मन एनाटोमिस्ट और हिस्टोलॉजिस्ट रिचर्ड ऑल्टमैन द्वारा वर्णित किया गया था, और इस ऑर्गेनेल का नाम एक अन्य जर्मन हिस्टोलॉजिस्ट के. बेंड ने 1897 में दिया था। लेकिन केवल 1920 में, फिर से, जर्मन बायोकेमिस्ट ओटो वैगबर्ग ने साबित किया कि सेलुलर श्वसन की प्रक्रियाएं माइटोकॉन्ड्रिया से जुड़ी हैं।
एक सिद्धांत है कि माइटोकॉन्ड्रिया आदिम कोशिकाओं द्वारा कब्जा करने के परिणामस्वरूप प्रकट हुआ, कोशिकाएं जो स्वयं ऊर्जा उत्पन्न करने के लिए ऑक्सीजन का उपयोग नहीं कर सकती थीं, प्रोटोजोनोट बैक्टीरिया जो ऐसा कर सकते थे। यह ठीक है क्योंकि माइटोकॉन्ड्रियन पहले एक अलग जीवित जीव था कि आज भी इसका अपना डीएनए है।
माइटोकॉन्ड्रिया एक स्वतंत्र जीवित जीव हुआ करता था।
विकास के क्रम में, पूर्वजों ने अपने कई जीनों को नाभिक में धोखा दिया जो कि ऊर्जा दक्षता में वृद्धि के कारण बने थे और स्वतंत्र जीव नहीं रह गए थे। माइटोकॉन्ड्रिया सभी कोशिकाओं में मौजूद होते हैं। यहां तक कि शुक्राणु में भी माइटोकॉन्ड्रिया होता है। यह उनके लिए धन्यवाद है कि शुक्राणु की पूंछ गति में सेट होती है, जो इसके आंदोलन को करती है। लेकिन उन जगहों पर विशेष रूप से कई माइटाकॉन्ड्रिया हैं जहां किसी भी जीवन प्रक्रिया के लिए ऊर्जा की आवश्यकता होती है। और हां, यह मुख्य रूप से मांसपेशी कोशिकाएं हैं।
मांसपेशियों की कोशिकाओं में, माइटोकॉन्ड्रिया को विशाल शाखाओं वाले माइटोकॉन्ड्रिया के समूहों में जोड़ा जा सकता है, जो एक दूसरे से इंटरमिटोकॉन्ड्रियल संपर्कों का उपयोग करके जुड़े होते हैं, जिसमें वे एक समन्वित कार्य सहकारी प्रणाली बनाना. ऐसे क्षेत्र में स्थान में इलेक्ट्रॉन घनत्व में वृद्धि हुई है। नए माइटोकॉन्ड्रिया का निर्माण पिछले जीवों के सरल विभाजन से होता है। सबसे "सरल" और सभी कोशिकाओं के लिए ऊर्जा आपूर्ति के तंत्र को अक्सर ग्लाइकोलाइसिस की सामान्य अवधारणा कहा जाता है।
यह ग्लूकोज के पाइरुविक एसिड में क्रमिक अपघटन की प्रक्रिया है। अगर यह प्रक्रिया होती है आणविक ऑक्सीजन की भागीदारी के बिनाया अपर्याप्त उपस्थिति के साथ, इसे कहा जाता है अवायवीय ग्लाइकोलाइसिस. इस मामले में, ग्लूकोज को अंतिम उत्पादों तक नहीं, बल्कि लैक्टिक और पाइरुविक एसिड में तोड़ा जाता है, जो बाद में किण्वन के दौरान और परिवर्तनों से गुजरता है। इसलिए, जारी ऊर्जा कम है, लेकिन ऊर्जा उत्पादन की दर तेज है। एनारोबिक ग्लाइकोलाइसिस के परिणामस्वरूप, एक कोशिका को एक ग्लूकोज अणु से 2 एटीपी अणु और 2 लैक्टिक एसिड अणु प्राप्त होते हैं। ऐसी "बुनियादी" ऊर्जा प्रक्रिया किसी भी कोशिका के अंदर हो सकती है। माइटोकॉन्ड्रिया के बिना.
पर आणविक ऑक्सीजन की उपस्थितिमाइटोकॉन्ड्रिया के अंदर किया जाता है एरोबिक ग्लाइकोलाइसिसश्वसन श्रृंखला के भीतर। एरोबिक स्थितियों के तहत पाइरुविक एसिड ट्राइकारबॉक्सिलिक एसिड चक्र या क्रेब्स चक्र में शामिल होता है। इस बहु-चरणीय प्रक्रिया के परिणामस्वरूप, एक ग्लूकोज अणु से 36 एटीपी अणु बनते हैं। विकसित माइटोकॉन्ड्रिया और कोशिकाओं के साथ एक सेल के ऊर्जा संतुलन की तुलना जहां वे विकसित नहीं हैं शो(पर्याप्त ऑक्सीजन के साथ) कोशिका के अंदर ग्लूकोज ऊर्जा के उपयोग की पूर्णता में अंतर लगभग 20 गुना है!
मनुष्यों में, कंकाल की मांसपेशी फाइबर कर सकते हैं सशर्तयांत्रिक और उपापचयी गुणों के आधार पर तीन प्रकारों में विभाजित: - धीमा ऑक्सीकरण; - तेज ग्लाइकोलाइटिक; - तेजी से ऑक्सीकरण-ग्लाइकोलाइटिक।

तेजी से मांसपेशी फाइबरतेज और कड़ी मेहनत के लिए बनाया गया है। अपने संकुचन के लिए, वे मुख्य रूप से ऊर्जा के तेज स्रोतों का उपयोग करते हैं, जैसे कि क्रिएटिन फॉस्फेट और एनारोबिक ग्लाइकोलाइसिस। इस प्रकार के तंतुओं में माइटोकॉन्ड्रिया की सामग्री धीमी मांसपेशी फाइबर की तुलना में बहुत कम होती है।
धीमी मांसपेशी फाइबरधीमी गति से संकुचन करते हैं, लेकिन लंबे समय तक काम करने में सक्षम होते हैं। वे एरोबिक ग्लाइकोलाइसिस और वसा से ऊर्जा के संश्लेषण को ऊर्जा के रूप में उपयोग करते हैं। यह अवायवीय ग्लाइकोलाइसिस की तुलना में बहुत अधिक ऊर्जा प्रदान करता है, लेकिन बदले में अधिक समय की आवश्यकता होती है, क्योंकि ग्लूकोज क्षरण की श्रृंखला अधिक जटिल होती है और इसके लिए ऑक्सीजन की उपस्थिति की आवश्यकता होती है, जिसे ऊर्जा रूपांतरण के स्थान पर ले जाने में भी समय लगता है। मायोग्लोबिन के कारण धीमी मांसपेशी फाइबर को लाल कहा जाता है, फाइबर में ऑक्सीजन लाने के लिए जिम्मेदार प्रोटीन। धीमी मांसपेशी फाइबर में माइटोकॉन्ड्रिया की एक महत्वपूर्ण मात्रा होती है।
प्रश्न उठता है कि किस प्रकार और किन व्यायामों की सहायता से पेशीय कोशिकाओं में माइटोकॉन्ड्रिया का एक व्यापक नेटवर्क विकसित किया जा सकता है? सामग्री में विभिन्न सिद्धांत और प्रशिक्षण के तरीके और उनके बारे में हैं।
अधिकांश कोशिकाओं की विशेषता। मुख्य कार्य कार्बनिक यौगिकों का ऑक्सीकरण और जारी ऊर्जा से एटीपी अणुओं का उत्पादन है। एक छोटा माइटोकॉन्ड्रिया पूरे जीव का मुख्य ऊर्जा केंद्र है।
माइटोकॉन्ड्रिया की उत्पत्ति
आज वैज्ञानिकों के बीच, यह राय बहुत लोकप्रिय है कि विकास के दौरान माइटोकॉन्ड्रिया स्वतंत्र रूप से कोशिका में प्रकट नहीं हुआ। सबसे अधिक संभावना है, यह एक आदिम कोशिका द्वारा कब्जा करने के कारण हुआ, जो उस समय अपने दम पर ऑक्सीजन का उपयोग करने में सक्षम नहीं था, बैक्टीरिया, जो जानता था कि यह कैसे करना है और, तदनुसार, ऊर्जा का एक उत्कृष्ट स्रोत था। ऐसा सहजीवन सफल रहा और बाद की पीढ़ियों में इसकी जड़ें मजबूत हुईं। यह सिद्धांत माइटोकॉन्ड्रिया में अपने स्वयं के डीएनए की उपस्थिति द्वारा समर्थित है।
माइटोकॉन्ड्रिया कैसे व्यवस्थित होते हैं?
माइटोकॉन्ड्रिया में दो झिल्ली होती हैं: बाहरी और भीतरी। बाहरी झिल्ली का मुख्य कार्य कोशिका के कोशिका द्रव्य से ऑर्गेनॉइड को अलग करना है। इसमें एक बिलीपिड परत होती है और इसमें घुसने वाले प्रोटीन होते हैं, जिसके माध्यम से काम के लिए आवश्यक अणुओं और आयनों का परिवहन किया जाता है। चिकनी होने पर, आंतरिक कई तह बनाता है - क्राइस्ट, जो इसके क्षेत्र को काफी बढ़ाता है। आंतरिक झिल्ली ज्यादातर प्रोटीन से बनी होती है, जिनमें श्वसन श्रृंखला एंजाइम, परिवहन प्रोटीन और बड़े एटीपी सिंथेटेज़ कॉम्प्लेक्स होते हैं। यहीं पर एटीपी का संश्लेषण होता है। बाहरी और आंतरिक झिल्लियों के बीच में अपने निहित एंजाइमों के साथ इंटरमेम्ब्रेन स्पेस होता है।
माइटोकॉन्ड्रिया के आंतरिक भाग को मैट्रिक्स कहा जाता है। यहां फैटी एसिड और पाइरूवेट के ऑक्सीकरण के लिए एंजाइम सिस्टम, क्रेब्स चक्र के एंजाइम, साथ ही माइटोकॉन्ड्रिया की वंशानुगत सामग्री - डीएनए, आरएनए और प्रोटीन-संश्लेषण तंत्र हैं।
माइटोकॉन्ड्रिया किसके लिए हैं?
माइटोकॉन्ड्रिया का मुख्य कार्य रासायनिक ऊर्जा के एक सार्वभौमिक रूप का संश्लेषण है - एटीपी। वे ट्राइकारबॉक्सिलिक एसिड चक्र में भी भाग लेते हैं, पाइरूवेट और फैटी एसिड को एसिटाइल-सीओए में परिवर्तित करते हैं और फिर इसे ऑक्सीकरण करते हैं। इस अंग में, माइटोकॉन्ड्रिया के सामान्य कामकाज के लिए आवश्यक टीआरएनए, आरआरएनए और कुछ प्रोटीन के प्रजनन को एन्कोड करते हुए, माइटोकॉन्ड्रियल डीएनए संग्रहीत और विरासत में मिला है।
माइटोकॉन्ड्रियाएक सर्पिल, गोल, लम्बी या शाखित अंग है।
माइटोकॉन्ड्रिया की अवधारणा को पहली बार 1897 में बेंडा द्वारा प्रस्तावित किया गया था। माइटोकॉन्ड्रिया को जीवित कोशिकाओं में अनाज, कणिकाओं या फिलामेंट्स के रूप में चरण-विपरीत और हस्तक्षेप माइक्रोस्कोपी का उपयोग करके पता लगाया जा सकता है। ये बल्कि मोबाइल संरचनाएं हैं जो एक दूसरे के साथ जुड़ सकती हैं, विलय कर सकती हैं, साझा कर सकती हैं। जब प्रकाश माइक्रोस्कोपी के साथ मृत कोशिकाओं में विशेष विधियों द्वारा दाग दिया जाता है, तो माइटोकॉन्ड्रिया छोटे अनाज (ग्रेन्यूल्स) की तरह दिखते हैं जो साइटोप्लाज्म में वितरित होते हैं या इसके कुछ विशिष्ट क्षेत्रों में केंद्रित होते हैं।
ऑक्सीजन की उपस्थिति में ग्लूकोज और वसा के विनाश के परिणामस्वरूप, माइटोकॉन्ड्रिया में ऊर्जा उत्पन्न होती है, और कार्बनिक पदार्थ पानी और कार्बन डाइऑक्साइड में परिवर्तित हो जाते हैं। यह इस तरह है कि पशु जीवों को जीवन के लिए आवश्यक मुख्य ऊर्जा प्राप्त होती है। ऊर्जा एडेनोसाइन ट्राइफॉस्फेट (एटीपी) में संग्रहीत होती है, या बल्कि, इसके मैक्रोर्जिक बॉन्ड में। माइटोकॉन्ड्रिया का कार्य कार्बनिक यौगिकों के ऑक्सीकरण और एटीपी अणुओं के संश्लेषण के लिए उनके क्षय के दौरान जारी ऊर्जा के उपयोग से निकटता से संबंधित है। इसलिए, माइटोकॉन्ड्रिया को अक्सर कोशिका के ऊर्जा केंद्र या सेलुलर श्वसन के अंग कहा जाता है। एटीपी अपने ऊर्जा-समृद्ध अंत फॉस्फेट समूहों में से एक को दूसरे अणु में स्थानांतरित करके ऊर्जा आपूर्तिकर्ता के रूप में कार्य करता है और इस प्रक्रिया में एडीपी में परिवर्तित हो जाता है।
यह माना जाता है कि विकास में माइटोकॉन्ड्रिया प्रोकैरियोटिक सूक्ष्मजीव थे जो एक प्राचीन कोशिका के शरीर में सहजीवी बन गए। इसके बाद, वे महत्वपूर्ण हो गए, जो पृथ्वी के वायुमंडल में ऑक्सीजन की मात्रा में वृद्धि से जुड़ा था। एक ओर, माइटोकॉन्ड्रिया ने कोशिका के लिए विषाक्त अतिरिक्त ऑक्सीजन को हटा दिया, और दूसरी ओर, उन्होंने ऊर्जा प्रदान की।
माइटोकॉन्ड्रिया के बिना, कोशिका व्यावहारिक रूप से ऑक्सीजन को एक पदार्थ के रूप में उपयोग करने में असमर्थ है जो ऊर्जा आपूर्ति प्रदान करती है, और केवल अवायवीय प्रक्रियाओं के माध्यम से अपनी ऊर्जा जरूरतों को पूरा कर सकती है। इस प्रकार, ऑक्सीजन एक जहर है, लेकिन कोशिका के लिए महत्वपूर्ण जहर है, और ऑक्सीजन की अधिकता इसकी कमी के समान ही हानिकारक है।
माइटोकॉन्ड्रिया अपना आकार बदल सकते हैं और कोशिका के उन क्षेत्रों में जा सकते हैं जहाँ उनकी सबसे अधिक आवश्यकता होती है। तो, कार्डियोमायोसाइट्स में, माइटोकॉन्ड्रिया मायोफिब्रिल्स के पास, वृक्क नलिकाओं की कोशिकाओं में, बेसल प्रोट्रूशियंस आदि के पास स्थित होते हैं। एक कोशिका में एक हजार माइटोकॉन्ड्रिया होते हैं, और उनकी संख्या कोशिका की गतिविधि पर निर्भर करती है।
माइटोकॉन्ड्रिया का औसत अनुप्रस्थ आयाम 0.5…3 माइक्रोन है। आकार के आधार पर, छोटे, मध्यम, बड़े और विशाल माइटोकॉन्ड्रिया पृथक होते हैं (वे एक व्यापक नेटवर्क बनाते हैं - माइटोकॉन्ड्रियल रेटिकुलम)। माइटोकॉन्ड्रिया का आकार और संख्या कोशिका गतिविधि और ऊर्जा खपत से निकटता से संबंधित है। वे अत्यंत परिवर्तनशील हैं और, कोशिका गतिविधि, ऑक्सीजन सामग्री और हार्मोनल प्रभावों के आधार पर, वे सूज सकते हैं, क्राइस्ट की संख्या और संरचना को बदल सकते हैं, संख्या, आकार और आकार में भिन्न हो सकते हैं, साथ ही साथ एंजाइमी गतिविधि भी कर सकते हैं।
माइटोकॉन्ड्रिया का आयतन घनत्व, उनकी आंतरिक सतह के विकास की डिग्री और अन्य संकेतक कोशिका की ऊर्जा आवश्यकताओं पर निर्भर करते हैं। लिम्फोसाइटों में केवल कुछ माइटोकॉन्ड्रिया होते हैं, और यकृत कोशिकाओं में उनमें से 2-3 हजार होते हैं।
माइटोकॉन्ड्रिया एक मैट्रिक्स, एक आंतरिक झिल्ली, एक पेरिमिटोकॉन्ड्रियल स्पेस और एक बाहरी झिल्ली से बना होता है। बाहरी माइटोकॉन्ड्रियल झिल्ली ऑर्गेनेल को हाइलोप्लाज्म से अलग करती है। आमतौर पर इसमें समरूपता होती है और इसे बंद कर दिया जाता है ताकि यह एक झिल्ली बैग हो।
बाहरी झिल्ली को लगभग 10-20 एनएम चौड़ा एक पेरिमिटोकॉन्ड्रियल स्पेस द्वारा आंतरिक झिल्ली से अलग किया जाता है। आंतरिक माइटोकॉन्ड्रियल झिल्ली माइटोकॉन्ड्रिया की वास्तविक आंतरिक सामग्री को सीमित करता है - मैट्रिक्स। आंतरिक झिल्ली माइटोकॉन्ड्रिया के अंदर कई प्रोट्रूशियंस बनाती है, जो सपाट लकीरें या क्राइस्ट की तरह दिखती हैं।
क्राइस्ट का आकार प्लेटों (ट्रैबिकुलर) और नलिकाओं (कट पर बहुकोशिकीय) के रूप में हो सकता है, और उन्हें माइटोकॉन्ड्रिया के संबंध में अनुदैर्ध्य या अनुप्रस्थ रूप से निर्देशित किया जाता है।
प्रत्येक माइटोकॉन्ड्रियन एक मैट्रिक्स से भरा होता है जो आसपास के साइटोप्लाज्म की तुलना में इलेक्ट्रॉन माइक्रोग्राफ पर सघन दिखाई देता है। माइटोकॉन्ड्रियल मैट्रिक्स सजातीय (सजातीय) है, कभी-कभी विभिन्न इलेक्ट्रॉन घनत्वों के ठीक-ठीक होते हैं। यह पतले फिलामेंट्स को लगभग 2...3 एनएम की मोटाई और लगभग 15...20 एनएम के आकार के दानों को प्रकट करता है। मैट्रिक्स के धागे डीएनए अणु होते हैं, और छोटे दाने माइटोकॉन्ड्रियल राइबोसोम होते हैं। मैट्रिक्स में एंजाइम होते हैं, एक एकल-फंसे, चक्रीय डीएनए, माइटोकॉन्ड्रियल राइबोसोम, कई सीए 2+ आयन।
माइटोकॉन्ड्रिया के प्रोटीन संश्लेषण की स्वायत्त प्रणाली को हिस्टोन से मुक्त डीएनए अणुओं द्वारा दर्शाया जाता है। डीएनए छोटा, रिंग के आकार का (चक्रीय) होता है और इसमें 37 जीन होते हैं। परमाणु डीएनए के विपरीत, इसमें व्यावहारिक रूप से गैर-कोडिंग न्यूक्लियोटाइड अनुक्रम नहीं होते हैं। संरचना और संगठन की विशेषताएं बैक्टीरिया कोशिकाओं के डीएनए के साथ माइटोकॉन्ड्रिया के डीएनए को एक साथ लाती हैं। माइटोकॉन्ड्रिया के डीएनए पर, विभिन्न प्रकार के आरएनए अणुओं को संश्लेषित किया जाता है: सूचनात्मक, स्थानांतरण (परिवहन) और राइबोसोमल। माइटोकॉन्ड्रियल मैसेंजर आरएनए स्प्लिसिंग (सूचना भार नहीं रखने वाले क्षेत्रों को काटने) से नहीं गुजरता है। माइटोकॉन्ड्रियल डीएनए अणुओं का छोटा आकार सभी माइटोकॉन्ड्रियल प्रोटीन के संश्लेषण को निर्धारित नहीं कर सकता है। अधिकांश माइटोकॉन्ड्रियल प्रोटीन कोशिका नाभिक के आनुवंशिक नियंत्रण में होते हैं और साइटोप्लाज्म में संश्लेषित होते हैं, क्योंकि माइटोकॉन्ड्रियल डीएनए कमजोर रूप से व्यक्त किया जाता है और ऑक्सीडेटिव फास्फारिलीकरण श्रृंखला के एंजाइमों के केवल एक हिस्से का गठन प्रदान कर सकता है। माइटोकॉन्ड्रियल डीएनए दस से अधिक प्रोटीन को एनकोड नहीं करता है जो झिल्ली में स्थानीयकृत होते हैं और संरचनात्मक प्रोटीन होते हैं जो माइटोकॉन्ड्रियल झिल्ली के व्यक्तिगत कार्यात्मक प्रोटीन परिसरों के सही एकीकरण के लिए जिम्मेदार होते हैं। परिवहन कार्यों को करने वाले प्रोटीन भी संश्लेषित होते हैं। प्रोटीन संश्लेषण की ऐसी प्रणाली माइटोकॉन्ड्रिया के सभी कार्यों को प्रदान नहीं करती है, इसलिए माइटोकॉन्ड्रिया की स्वायत्तता सीमित और सापेक्ष है।
स्तनधारियों में, माइटोकॉन्ड्रिया को केवल अंडे के माध्यम से निषेचन के दौरान स्थानांतरित किया जाता है, और शुक्राणु नए जीव में परमाणु डीएनए लाता है।
माइटोकॉन्ड्रिया के मैट्रिक्स में, राइबोसोम बनते हैं जो साइटोप्लाज्म के राइबोसोम से भिन्न होते हैं। वे कई माइटोकॉन्ड्रियल प्रोटीन के संश्लेषण में शामिल होते हैं जो नाभिक द्वारा एन्कोडेड नहीं होते हैं। माइटोकॉन्ड्रियल राइबोसोम में 60 की अवसादन संख्या होती है (80 की अवसादन संख्या के साथ साइटोप्लाज्मिक राइबोसोम के विपरीत)। अवसादन संख्या सेंट्रीफ्यूजेशन और अल्ट्रासेंट्रीफ्यूजेशन के दौरान अवसादन दर है। संरचना के संदर्भ में, माइटोकॉन्ड्रियल राइबोसोम प्रोकैरियोटिक जीवों के समान होते हैं, लेकिन आकार में छोटे होते हैं और कुछ एंटीबायोटिक दवाओं (लेवोमाइसेटिन, टेट्रासाइक्लिन, आदि) के प्रति संवेदनशीलता में भिन्न होते हैं।
आंतरिक माइटोकॉन्ड्रियल झिल्ली में पदार्थों के परिवहन में उच्च स्तर की चयनात्मकता होती है। ऑक्सीडेटिव फास्फारिलीकरण श्रृंखला के निकटवर्ती एंजाइम, इलेक्ट्रॉन वाहक प्रोटीन, परिवहन प्रणाली एटीपी, एडीपी, पाइरूवेट, आदि इसकी आंतरिक सतह से जुड़े होते हैं। उत्प्रेरक प्रक्रियाओं की दक्षता।
इलेक्ट्रॉन माइक्रोस्कोपी ने मशरूम के आकार के कणों को मैट्रिक्स लुमेन में फैलाया। उनके पास एटीपी-सिंथेटिक (एडीपी से एटीपी रूप) गतिविधि है। इलेक्ट्रॉनों का परिवहन श्वसन श्रृंखला के साथ होता है, आंतरिक झिल्ली में स्थानीयकृत होता है, जिसमें चार बड़े एंजाइम कॉम्प्लेक्स (साइटोक्रोम) होते हैं। जब इलेक्ट्रॉन श्वसन श्रृंखला से गुजरते हैं, तो हाइड्रोजन आयनों को मैट्रिक्स से पेरिमिटोकॉन्ड्रियल स्पेस में पंप किया जाता है, जो एक प्रोटॉन ग्रेडिएंट (पंप) का निर्माण सुनिश्चित करता है। इस ढाल की ऊर्जा (पदार्थों की सांद्रता और एक झिल्ली क्षमता के निर्माण में अंतर) का उपयोग एटीपी के संश्लेषण और मेटाबोलाइट्स और अकार्बनिक आयनों के परिवहन के लिए किया जाता है। आंतरिक झिल्ली पर निहित वाहक प्रोटीन इसके माध्यम से कार्बनिक फॉस्फेट, एटीपी, एडीपी, अमीनो एसिड, फैटी एसिड, ट्राई- और डाइकारबॉक्सिलिक एसिड का परिवहन करते हैं।
माइटोकॉन्ड्रिया की बाहरी झिल्ली कम आणविक भार वाले पदार्थों के लिए अधिक पारगम्य है, क्योंकि इसमें कई हाइड्रोफिलिक प्रोटीन चैनल होते हैं। विशिष्ट रिसेप्टर कॉम्प्लेक्स बाहरी झिल्ली पर स्थित होते हैं, जिसके माध्यम से मैट्रिक्स से प्रोटीन को पेरिमिटोकॉन्ड्रियल स्पेस में ले जाया जाता है।
इसकी रासायनिक संरचना और गुणों के संदर्भ में, बाहरी झिल्ली अन्य इंट्रासेल्युलर झिल्ली और प्लास्मलेम्मा के करीब है। इसमें एंजाइम होते हैं जो वसा को चयापचय करते हैं, अमाइन, अमीनो ऑक्सीडेज के परिवर्तन को सक्रिय (उत्प्रेरित) करते हैं। यदि बाहरी झिल्ली के एंजाइम सक्रिय रहते हैं, तो यह माइटोकॉन्ड्रिया की कार्यात्मक सुरक्षा का सूचक है।
माइटोकॉन्ड्रिया में दो स्वायत्त उप-कम्पार्टमेंट होते हैं। जबकि पेरिमिटोकॉन्ड्रियल स्पेस, या माइटोकॉन्ड्रिया (बाहरी उपकम्पार्टमेंट) का बाहरी कक्ष, हाइलोप्लास्मिक प्रोटीन कॉम्प्लेक्स के प्रवेश के कारण बनता है, आंतरिक उपकम्पार्टमेंट (माइटोकॉन्ड्रियल मैट्रिक्स) आंशिक रूप से माइटोकॉन्ड्रियल डीएनए की सिंथेटिक गतिविधि के कारण बनता है। आंतरिक उपकम्पार्टमेंट (मैट्रिक्स) में डीएनए, आरएनए और राइबोसोम होते हैं। यह हाइलोप्लाज्म की तुलना में सीए 2+ आयनों के उच्च स्तर द्वारा प्रतिष्ठित है। बाहरी उपकम्पार्टमेंट में हाइड्रोजन आयन जमा होते हैं। बाहरी और आंतरिक उपखंडों की एंजाइमेटिक गतिविधि, प्रोटीन की संरचना बहुत भिन्न होती है। आंतरिक उपकम्पार्टमेंट में बाहरी की तुलना में अधिक इलेक्ट्रॉन घनत्व होता है।
माइटोकॉन्ड्रिया के विशिष्ट मार्कर एंजाइम साइटोक्रोम ऑक्सीडेज और सक्सेनेट डिहाइड्रोजनेज हैं, जिसका पता लगाने से माइटोकॉन्ड्रिया में ऊर्जा प्रक्रियाओं की मात्रा निर्धारित करना संभव हो जाता है।
माइटोकॉन्ड्रिया का मुख्य कार्य- एटीपी संश्लेषण। प्रारंभ में, शर्करा (ग्लूकोज) हाइलोप्लाज्म में लैक्टिक और पाइरुविक एसिड (पाइरूवेट) में एटीपी की एक छोटी मात्रा के एक साथ संश्लेषण के साथ नष्ट हो जाते हैं। एक ग्लूकोज अणु के ग्लाइकोलाइसिस के परिणामस्वरूप, दो एटीपी अणुओं का उपयोग किया जाता है, और चार बनते हैं। इस प्रकार, केवल दो एटीपी अणु एक सकारात्मक संतुलन बनाते हैं। ये प्रक्रियाएं बिना ऑक्सीजन (एनारोबिक ग्लाइकोलाइसिस) के होती हैं।
ऊर्जा उत्पादन के सभी बाद के चरण एरोबिक ऑक्सीकरण की प्रक्रिया में होते हैं, जो बड़ी मात्रा में एटीपी का संश्लेषण प्रदान करता है। इस मामले में, कार्बनिक पदार्थ सीओ 2 और पानी में नष्ट हो जाते हैं। ऑक्सीकरण के साथ प्रोटॉन अपने स्वीकर्ता को स्थानांतरित कर देते हैं। इन प्रतिक्रियाओं की मध्यस्थता माइटोकॉन्ड्रियल मैट्रिक्स में पाए जाने वाले कई ट्राइकारबॉक्सिलिक एसिड चक्र एंजाइमों द्वारा की जाती है।
इलेक्ट्रॉन ट्रांसफर सिस्टम और इससे जुड़े एडीपी फास्फारिलीकरण (ऑक्सीडेटिव फास्फारिलीकरण) क्राइस्ट की झिल्लियों में निर्मित होते हैं। इस मामले में, इलेक्ट्रॉनों को एक इलेक्ट्रॉन स्वीकर्ता प्रोटीन से दूसरे में स्थानांतरित किया जाता है और अंत में, वे ऑक्सीजन से बंधे होते हैं, जिसके परिणामस्वरूप पानी बनता है। इसी समय, इलेक्ट्रॉन परिवहन श्रृंखला में इस तरह के ऑक्सीकरण के दौरान जारी ऊर्जा का हिस्सा एडीपी फॉस्फोराइलेशन के दौरान एक मैक्रोर्जिक बंधन के रूप में संग्रहीत होता है, जिससे बड़ी संख्या में एटीपी अणुओं का निर्माण होता है - मुख्य इंट्रासेल्युलर ऊर्जा समकक्ष। माइटोकॉन्ड्रियल क्राइस्ट की झिल्लियों पर, यहां स्थित ऑक्सीकरण श्रृंखला प्रोटीन और एडीपी-एटीपी सिंथेटेस फॉस्फोराइलेशन एंजाइम की मदद से ऑक्सीडेटिव फास्फारिलीकरण की प्रक्रिया होती है। ऑक्सीडेटिव फास्फारिलीकरण के परिणामस्वरूप, एक ग्लूकोज अणु से 36 एटीपी अणु बनते हैं।
माइटोकॉन्ड्रिया में कुछ हार्मोन और पदार्थों के लिए विशिष्ट (आत्मीयता) रिसेप्टर्स होते हैं। ट्राईआयोडोथायरोनिन सामान्य रूप से माइटोकॉन्ड्रिया की सिंथेटिक गतिविधि को तेज करता है। इंटरल्यूकिन -1 और ट्राईआयोडोथायरोनिन की उच्च सांद्रता ऑक्सीडेटिव फास्फारिलीकरण की श्रृंखलाओं को अलग करती है, माइटोकॉन्ड्रियल सूजन का कारण बनती है, जो थर्मल ऊर्जा के उत्पादन में वृद्धि के साथ होती है।
न्यू माइटोकॉन्ड्रिया विभाजन, कसना या नवोदित द्वारा बनते हैं। बाद के मामले में, एक प्रोटोमाइटोकॉन्ड्रियन बनता है, धीरे-धीरे आकार में बढ़ रहा है।
प्रोटोमाइटोकॉन्ड्रियन बाहरी और आंतरिक झिल्लियों वाला एक छोटा अंग है। आंतरिक झिल्ली में अविकसित क्राइस्ट नहीं होता है या नहीं होता है। ऑर्गेनेल को एरोबिक फॉस्फोराइलेशन के निम्न स्तर की विशेषता है। जब कसना बनता है, तो माइटोकॉन्ड्रिया की सामग्री दो नए बल्कि बड़े जीवों के बीच वितरित की जाती है। प्रजनन की किसी भी विधि के साथ, नवगठित माइटोकॉन्ड्रिया में से प्रत्येक का अपना जीनोम होता है।
पुराने माइटोकॉन्ड्रिया ऑटोलिसोसोम के निर्माण के साथ ऑटोलिसिस (लाइसोसोम की मदद से कोशिका द्वारा आत्म-पाचन) द्वारा नष्ट हो जाते हैं। अवशिष्ट शरीर का निर्माण ऑटोलिसोसोम से होता है। पूर्ण पाचन के साथ, अवशिष्ट शरीर की सामग्री, जिसमें कम आणविक भार कार्बनिक पदार्थ होते हैं, एक्सोसाइटोसिस द्वारा उत्सर्जित होते हैं। अपूर्ण पाचन के साथ, माइटोकॉन्ड्रियल अवशेष कोशिका में निपोफसिन के साथ स्तरित निकायों या कणिकाओं के रूप में जमा हो सकते हैं। माइटोकॉन्ड्रिया के हिस्से में, अघुलनशील कैल्शियम लवण क्रिस्टल के निर्माण के साथ जमा होते हैं - कैल्सीफिकेशन। माइटोकॉन्ड्रियल अध: पतन उत्पादों के संचय से सेल डिस्ट्रोफी हो सकती है।
माइटोकॉन्ड्रिया (एमटी) मेरे लिए अनुसंधान के सबसे दिलचस्प क्षेत्रों में से एक है। लगभग 1.6 अरब साल पहले एंडोसिम्बायोसिस के दौरान एक अन्य कोशिका के साथ माइटोकॉन्ड्रिया का जुड़ाव एक जटिल संरचना वाले सभी बहुकोशिकीय यूकेरियोट्स का आधार बन गया। संभवतः, माइटोकॉन्ड्रिया की उत्पत्ति α-proteobacteria जैसी कोशिकाओं से हुई है।
माइटोकॉन्ड्रिया पर सबसे अच्छा हालिया समीक्षा अध्ययन वर्नर कुलब्रांट का "माइटोकॉन्ड्रियल झिल्ली प्रोटीन परिसरों की संरचना और कार्य" है। यदि आप अंग्रेजी जानते हैं और इन जीवों की संरचना में रुचि रखते हैं, तो मैं इसे पढ़ने की जोरदार सलाह देता हूं। यह लेख इतना अच्छा है कि यह आसानी से एक अच्छी आणविक जीव विज्ञान पाठ्यपुस्तक का प्रमुख बन सकता है। पहले तो मैं पूरे लेख का अनुवाद करना चाहता था, लेकिन इसमें अक्षम्य रूप से लंबा समय लगेगा और मुझे अन्य चीजों से दूर ले जाएगा। इसलिए, मैं खुद को थीसिस और तस्वीरों तक सीमित रखूंगा। समय-समय पर अपने विचारों से सब कुछ पतला करना।
कोशिका से अलग डीएनए (mtDNA) और प्रोटीन ट्रांसक्रिप्शन के लिए पूरे "उत्पादन" चक्र की उपस्थिति के बावजूद, माइटोकॉन्ड्रिया स्वयं केवल 13 प्रोटीनों को एनकोड करता है। एक पृथक माइटोकॉन्ड्रिया कुछ समय के लिए अपनी संरचना और कार्य को बनाए रख सकता है।
चित्र 1. माइटोकॉन्ड्रिया झिल्ली के अवयव। बाहरी झिल्ली माइटोकॉन्ड्रिया को साइटोप्लाज्म से अलग करती है। यह आंतरिक झिल्ली को घेर लेती है, जो प्रोटीन युक्त केंद्रीय मैट्रिक्स से इंटरमेम्ब्रेन स्पेस को अलग करती है। आंतरिक झिल्ली को आंतरिक सीमित झिल्ली और क्राइस्ट में विभाजित किया गया है। ये दो भाग क्राइस्ट के लगाव बिंदुओं पर निरंतर हैं (क्राइस्टे संगम) क्राइस्ट मैट्रिक्स में कम या ज्यादा गहराई तक फैली हुई है और माइटोकॉन्ड्रियल ऊर्जा रूपांतरण की मुख्य साइट है। इंटरमेम्ब्रेन स्पेस में एक छोटा प्रोटॉन ग्रेडिएंट (पीएच7.2-7.4) और मैट्रिक्स (पीएच7.9-8.0) क्राइस्ट की झिल्लियों में एटीपी सिंथेज़ द्वारा एटीपी के निर्माण की ओर ले जाते हैं।
बाहरी झिल्ली झरझरा होती है और साइटोप्लाज्म से पदार्थों को गुजरने देती है। आंतरिक झिल्ली घनी होती है, इसे पार करने के लिए परिवहन प्रोटीन की आवश्यकता होती है [गिल्बर्ट लिंग यथोचित असहमत], अवरोध की निरंतरता आंतरिक झिल्ली को -180 mV की विद्युत रासायनिक क्षमता रखने की अनुमति देती है। मैट्रिक्स में एक उच्च पीएच (7.9-8) है। एक बार फिर मैं लिंग में जाऊँगा। क्षारीय (7 से ऊपर) पीएच एक अधिक प्रकट प्रोटीन संरचना को बढ़ावा देता है। उच्च पीएच हाइड्रोजन और नमक बंधनों को तोड़ता है, जिससे ध्रुवीकृत सीओ और एनएच पानी के अणुओं के लिए उपलब्ध हो जाते हैं, जिससे सभी इंट्रासेल्युलर पानी के द्विध्रुवीय क्षण में वृद्धि होती है और इसे बांधता है। इस नस में, कोशिका के अंदर प्रोटोप्लाज्म को "पकड़ने" के लिए एक झिल्ली की उपस्थिति की आवश्यकता नहीं होती है (यह उच्च पीएच पर स्वयं प्रोटीन द्वारा किया जाता है), लेकिन एक क्षमता रखने के लिए।
एमटीडीएनए न्यूक्लियोटाइड्स में पाया जाता है, जो लगभग 1000 प्रति सेल होते हैं। मैट्रिक्स का प्रोटीन घनत्व काफी अधिक (500 मिलीग्राम / एमएल तक) है, जो क्रिस्टलीकृत प्रोटीन के करीब है।
आंतरिक झिल्ली क्राइस्टे नामक आक्रमण बनाती है जो मैट्रिक्स में गहराई से प्रवेश करती है। क्राइस्ट माइटोकॉन्ड्रिया के तीसरे "डिब्बे" को परिभाषित करता है - क्राइस्ट का लुमेन (क्राइस्ट लुमेन)। क्राइस्टा झिल्ली में सबसे अधिक, यदि सभी नहीं, तो इलेक्ट्रॉन परिवहन श्रृंखला और एटीपी सिंथेज़ के पूरी तरह से "इकट्ठे" परिसर होते हैं। क्राइस्टा के लुमेन में शामिल है एक बड़ी संख्या कीएक छोटा घुलनशील प्रोटीन इलेक्ट्रॉन वाहक (साइटोक्रोम सी)। इस प्रकार माइटोकॉन्ड्रियल क्राइस्ट सभी गैर-प्रकाश संश्लेषक यूकेरियोट्स में जैविक ऊर्जा रूपांतरण का मुख्य स्थल है।
क्रिस्टल के साथ भी कई दिलचस्प चीजें हैं। क्राइस्टा के ऑप्टिकल गुण ऊतकों में प्रकाश के प्रसार और उत्पादन को प्रभावित करते हैं। मुझे यह भी विचार आया कि क्राइस्टस की सतह टोपोलॉजिकल इंसुलेटर की सतहों के समान (धारणा) है (चार्ज अपव्यय के बिना सुपरकंडक्टिविटी निहित थी)।

चित्रा 2. श्वसन श्रृंखला के झिल्ली प्रोटीन परिसरों। जटिलमैं (नाधी/ubiquinone oxidoreductase, नीला), कॉम्प्लेक्सद्वितीय(सक्सेनेट डिहाइड्रोजनेज, गुलाबी), कॉम्प्लेक्सतृतीय(साइटोक्रोम सी रिडक्टेस, ऑरेंज), कॉम्प्लेक्सचतुर्थ(साइटोक्रोम सी ऑक्सीडेज, हरा) और माइटोकॉन्ड्रियल एटीपी सिंथेज़ (कॉम्प्लेक्स के रूप में जाना जाता है)वी, बेज) ऑक्सीडेटिव फास्फारिलीकरण के दौरान एक साथ काम करते हैं ताकि कोशिकाएं ऊर्जा का उपयोग कर सकें। परिसरमैं, तृतीय, चतुर्थक्राइस्टा झिल्ली के साथ प्रोटॉन को पंप करें, एक प्रोटॉन ढाल का निर्माण करें जो एटीपी संश्लेषण को उत्तेजित करता है।
अब कॉम्प्लेक्स II पर थोड़ा ध्यान दें। आपको याद है कि वसा (कीटो) चयापचय में FADH2 और जटिल II पर जोर देता है। वे CoQ जोड़ी को पुनर्स्थापित करते हैं, कुछ बिंदु पर ऑक्सीकृत CoQ इलेक्ट्रॉनों को जटिल III में ले जाने के लिए पर्याप्त नहीं है और सुपरऑक्साइड के गठन के साथ जटिल I में इलेक्ट्रॉनों का एक रिवर्स प्रवाह बनाता है। लंबे समय तक एचएफएलसी पोषण के साथ, जटिल I विपरीत रूप से नष्ट हो जाएगा, जबकि यह एक सामान्य शारीरिक अनुकूलन है।
मैं आपसे यह भी नोट करने के लिए कहता हूं कि कॉम्प्लेक्स II प्रोटॉन को पंप नहीं करता है। क्या हमारे अंदर प्रोटॉन प्रवणता को फैलाता है, फास्फारिलीकरण को बाधित करता है और गर्मी के लिए वसा के जलने को उत्तेजित करता है? यह सही है, ठंडा तनाव। थर्मोजेनेसिस एक जटिल के माध्यम से चयापचय से जुड़ा हुआ है जो प्रोटॉन को बंद नहीं करता है, जिससे एटीपी सिंथेज़ के लिए कोई अतिरिक्त प्रोटॉन नहीं मिलता है। कोई केवल आश्चर्य कर सकता है कि हमारे जीव को कितना अद्भुत माना जाता है।
क्रिस्टल माउंट औरमाइक्रोस
क्राइस्ट जंक्शन छोटे गोल छेद होते हैं जिनका व्यास लगभग 25 एनएम होता है। सभी जीवों के माइटोकॉन्ड्रिया में MICOS (माइटोकॉन्ड्रिया संपर्क साइट और बाहरी झिल्ली के लिए क्राइस्ट) प्रणाली होती है, जो पांच झिल्ली और एक घुलनशील प्रोटीन का एक संयोजन होता है जो क्राइस्ट को बाहरी झिल्ली से जोड़ता है।
बढ़ी हुई ऊर्जा आवश्यकता वाली कोशिकाओं में, जैसे कि कंकाल और हृदय की मांसपेशियां, क्राइस्ट अधिकांश माइटोकॉन्ड्रियल मात्रा को घनी रूप से भरते हैं। कम ऊर्जा आवश्यकताओं वाले ऊतकों में, जैसे कि यकृत और गुर्दे, क्राइस्ट एक साथ कसकर पैक नहीं होते हैं। मैट्रिक्स में बायोसिंथेटिक एंजाइमों के लिए अधिक जगह है।
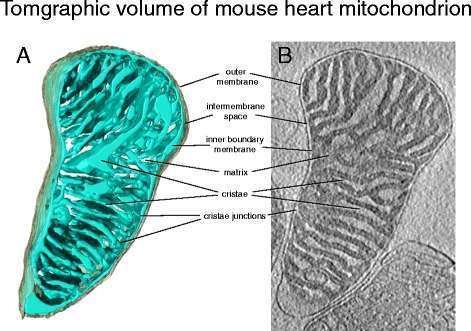
चित्रा 3. माउस दिल माइटोकॉन्ड्रिया की टोमोग्राफिक मात्रा। ए) क्रायो-ईटी द्वारा कैप्चर किए गए माउस हार्ट माइटोकॉन्ड्रिया की त्रि-आयामी मात्रा। बाहरी झिल्ली (ग्रे) आंतरिक झिल्ली (हल्का नीला) को ढक लेती है। आंतरिक झिल्ली घनी रूप से क्राइस्ट से भरी होती है b) आयतन का टोमोग्राफिक खंड। अधिकांश माइटोकॉन्ड्रियल प्रोटीन युक्त घनी भरी हुई मैट्रिक्स एक इलेक्ट्रॉन माइक्रोस्कोप पर अंधेरा दिखाई देती है। जबकि इंटरमेम्ब्रेन स्पेस और क्राइस्ट के लुमेन प्रोटीन की कम सांद्रता के कारण हल्के दिखते हैं।
एटीपी सिंथेज़ डिमर
माइटोकॉन्ड्रियल F1-F0 एटीपी सिंथेज़ क्राइस्टा का सबसे प्रमुख प्रोटीन कॉम्प्लेक्स है। एटीपी सिंथेज़ एक प्राचीन नैनोमशीन है जो घूर्णी कटैलिसीस के माध्यम से एएफटी बनाने के लिए आंतरिक झिल्ली के चारों ओर एक इलेक्ट्रोकेमिकल प्रोटॉन ग्रेडिएंट का उपयोग करता है। झिल्ली के F0 परिसर के माध्यम से घूमने वाले प्रोटॉन 8 (स्तनधारियों में) या 10 (खमीर में) सी-नोड्स के रोटर को घुमाते हैं। केंद्रीय तना सी-रोटर टॉर्क को एफ1 कैटेलिटिक हेड में स्थानांतरित करता है, जहां एटीपी का निर्माण एडीपी और फॉस्फेट से होता है, जो कि गठनात्मक परिवर्तनों के अनुक्रम के माध्यम से होता है। परिधीय तना F1 हेड के F0 कॉम्प्लेक्स के विरुद्ध अनुत्पादक रोटेशन को रोकता है।
कई वर्षों तक यह सोचा गया था कि एटीपी सिंथेज़ आंतरिक झिल्ली पर बेतरतीब ढंग से स्थित था। लेकिन यह पता चला कि डबल पंक्तियों में व्यवस्थित एटीपी सिंथेज़. इसके अलावा, एटीपी सिंथेज़ की रैखिक पंक्तियाँ सभी जीवित माइटोकॉन्ड्रिया का एक मूलभूत गुण हैं।

चित्रा 4. सात विभिन्न प्रजातियों में एटीपी सिंथेज़ की दोहरी पंक्तियाँ।
एटीपी सिंथेज़ की पंक्तियाँ मुख्य रूप से क्राइस्ट की लकीरों के साथ स्थित होती हैं। डिमर्स लिपिड बाईलेयर को मोड़ते हैं और परिणामस्वरूप पंक्तियों में स्व-व्यवस्थित हो जाते हैं। जब खमीर माइटोकॉन्ड्रिया में एपीटी सिंथेज़ के ई और जी नोड्स को खटखटाया गया, तो तनाव अपने जंगली समकक्षों की तुलना में 60% धीमा हो गया, और उनके माइटोकॉन्ड्रिया की झिल्ली क्षमता आधी हो गई। प्रोकैरियोटिक एपीटी सिंथेज़ में कई डिमर-जुड़े नोड्स का अभाव है, और बैक्टीरिया और आर्किया में डिमर पंक्तियाँ नहीं पाई गई हैं। एएफटी सिंथेज़ डिमर की क्राइस्ट और पंक्तियाँ इस प्रकार जीव की उच्च ऊर्जा आवश्यकताओं के लिए एक अनुकूलन हैं।

चित्रा 5. पॉलीमेला एसपी माइटोकॉन्ड्रिया के एटीपी सिंथेज़ डिमर की संरचना। एटीपी सिंथेज़ के वी-आकार के डिमर का साइड व्यू।
श्वसन श्रृंखला के परिसर और सुपरकंपलेक्स
आंतरिक झिल्ली के चारों ओर प्रोटॉन ढाल तीन बड़े झिल्ली परिसरों द्वारा बनाई गई है जिन्हें जटिल I, जटिल III और जटिल IV के रूप में जाना जाता है (चित्र 2 देखें)। कॉम्प्लेक्स I को एनएडीएच से इलेक्ट्रॉनों के साथ खिलाया जाता है, जब एक इलेक्ट्रॉन दान किया जाता है तो ऊर्जा चार प्रोटॉन को पंप करती है। कॉम्प्लेक्स III कम किए गए क्विनोल से एक इलेक्ट्रॉन प्राप्त करता है और इसे एक इलेक्ट्रॉन वाहक (साइटोक्रोम सी) में स्थानांतरित करता है, इस प्रक्रिया में एक प्रोटॉन को पंप करता है। कॉम्प्लेक्स IV साइटोक्रोम सी से एक इलेक्ट्रॉन प्राप्त करता है और इसे आणविक ऑक्सीजन को दान करता है, पानी में परिवर्तित प्रत्येक ऑक्सीजन अणु के लिए 4 प्रोटॉन पंप करता है। कॉम्प्लेक्स II प्रोटॉन को पंप नहीं करता है, सीधे क्विनोल को इलेक्ट्रॉनों का दान करता है। एनएडीएच से क्विनोल में इलेक्ट्रॉन स्थानांतरण प्रोटॉन स्थानान्तरण से कैसे संबंधित है, यह अभी तक स्पष्ट नहीं है। कॉम्प्लेक्स I, III और IV के संयुक्त से बड़ा है।

चित्रा 6. गोजातीय हृदय माइटोकॉन्ड्रिया का जटिल I। मैट्रिक्स भाग में आठ लौह-सल्फर (Fe-S) समूहों की एक पंक्ति होती है जो मैट्रिक्स-झिल्ली जंक्शन पर NADH से क्विनॉल तक इलेक्ट्रॉनों को निर्देशित करती है। झिल्ली वाले हिस्से में 78 ब्लेड होते हैं, जिनमें अणु भी शामिल हैं जो प्रोटॉन को पंप करते हैं।
परिसरमैंतृतीय औरIV सुपरकॉम्प्लेक्स में संयोजित होता हैया श्वसन संबंधी। बेकर के खमीर में ( Saccharomycescerevisiae) कोई जटिल I नहीं है, उनके सुपरकंपलेक्स में III और IV शामिल हैं। सुपरकॉम्प्लेक्स की भूमिका अभी स्पष्ट नहीं है। ऐसा माना जाता है कि यह इलेक्ट्रॉन परिवहन को अधिक कुशल बनाता है, लेकिन इसका अभी तक कोई प्रत्यक्ष प्रमाण नहीं है।

चित्रा 7. गोजातीय हृदय माइटोकॉन्ड्रियल सुपरकंपलेक्स। कॉम्प्लेक्स I और III के बीच की दूरी पर ध्यान दें, जो कि क्विनोल के साथ किया जाना चाहिए। तीर सुपरकंपलेक्स में एक इलेक्ट्रॉन की गति दिखाते हैं।
क्राइस्टा के लुमेन में मुख्य प्रोटीन साइटोक्रोम सी है, जो एक इलेक्ट्रॉन को जटिल III से जटिल IV में स्थानांतरित करता है। यदि साइटोक्रोम सी कोशिका के कोशिका द्रव्य में छोड़ा जाता है, तो यह एपोप्टोसिस का कारण बनता है.

चित्रा 8. एटीपी सिंथेज़ डिमर की पंक्तियाँ क्राइस्ट के आकार को निर्धारित करती हैं। क्राइस्टा के रिज पर, एएफटी सिंथेज़ (पीला) प्रोटॉन (लाल) के लिए एक संगम बनाता है, इलेक्ट्रॉनिक सर्किट (हरा) के प्रोटॉन पंप डिमर की पंक्तियों के दोनों किनारों पर स्थित होते हैं। प्रोटॉन को स्रोत से एटीपी सिंथेज़ तक निर्देशित करके, क्राइस्ट कुशल एटीपी उत्पादन की अनुमति देने के लिए प्रोटॉन गाइड के रूप में कार्य करता है। लाल तीर प्रोटॉन प्रवाह की दिशा दिखाते हैं।
उम्र बढ़ने के दौरान झिल्ली पुनर्गठन
सभी यूकेरियोट्स में बुढ़ापा एक मौलिक और खराब समझी जाने वाली प्रक्रिया है। कवक पर माइटोकॉन्ड्रियल उम्र बढ़ने का अध्ययन किया गया था पोडोस्पोरा अंसेरिनाजो सिर्फ 18 दिन जीते हैं। सामान्य माइटोकॉन्ड्रिया में, क्राइस्ट मैट्रिक्स में गहराई तक फैलती है। इसके लिए एटीपी सिंथेज़ डिमर की पंक्तियों और क्राइस्ट के अटैचमेंट साइट्स पर एक एमआईसीओएस कॉम्प्लेक्स की आवश्यकता होती है। उम्र के साथ, क्राइस्ट झिल्ली की सतह के करीब आने लगते हैं, एपीटी सिंथेज़ के डिमर मोनोमर्स में परिवर्तित हो जाते हैं, और सब कुछ साइटोक्रोम सी और कोशिका मृत्यु की रिहाई के साथ समाप्त होता है।
इलेक्ट्रॉन परिवहन परिसरों I और III में सुपरऑक्साइड बनाता है। यह चयापचय का उप-उत्पाद है। एक ही समय में आवश्यक और घातक। उम्र बढ़ने के दौरान, संलयन पर विखंडन पूर्वता लेने लगता है। यह क्षतिग्रस्त माइटोकॉन्ड्रिया को संलयन द्वारा "बचाने" से रोकता है और अपरिहार्य को तेज करता है।

चित्रा 9. माइटोकॉन्ड्रियल उम्र बढ़ने के दौरान आंतरिक झिल्ली और एटीपी सिंथेज़ डिमर के आकारिकी में परिवर्तन।
जैसा कि आप देख सकते हैं, भविष्य के शोध के लिए बहुत सारे क्षेत्र हैं। मैं भौतिकी और जीव विज्ञान के चौराहे पर अनुमान लगा रहा हूं, जहां भौतिक विज्ञानी यह समझाने की कोशिश करेंगे कि ऐसी संरचना अधिक ऊर्जा कुशल क्यों है। विशेष रूप से क्वांटम जीव विज्ञान पर प्रकाशन
माइटोकॉन्ड्रिया
माइटोकॉन्ड्रिया रॉड के आकार या अंडाकार आकार की संरचनाएं हैं (जीआर। मितोस- एक धागा, चोंड्रोस- दाना)। वे सभी पशु कोशिकाओं (परिपक्व एरिथ्रोसाइट्स को छोड़कर) में पाए जाते हैं: उच्च पौधों में, शैवाल और प्रोटोजोआ में। वे केवल बैक्टीरिया के प्रोकैरियोट्स में अनुपस्थित हैं।
इन जीवों को पहली बार पिछली शताब्दी के अंत में ऑल्टमैन द्वारा खोजा और वर्णित किया गया था। कुछ समय बाद, इन संरचनाओं को माइटोकॉन्ड्रिया कहा गया। 1948 में होगेबूम ने कोशिकीय श्वसन के केंद्र के रूप में माइटोकॉन्ड्रिया के महत्व को इंगित किया, और 1949 में कैनेडी और लेहिंगर ने स्थापित किया कि माइटोकॉन्ड्रिया में ऑक्सीडेटिव फास्फारिलीकरण का एक चक्र होता है। तो यह साबित हो गया कि माइटोकॉन्ड्रिया ऊर्जा उत्पादन के स्थान के रूप में कार्य करता है।
माइटोकॉन्ड्रिया एक पारंपरिक प्रकाश माइक्रोस्कोप में विशेष धुंधला विधियों के साथ दिखाई दे रहे हैं। एक चरण में - विपरीत सूक्ष्मदर्शी और "अंधेरे क्षेत्र" में उन्हें जीवित कोशिकाओं में देखा जा सकता है।
संरचना, आयाम, आकार माइटोकॉन्ड्रिया अत्यधिक परिवर्तनशील हैं। यह मुख्य रूप से कोशिकाओं की कार्यात्मक स्थिति पर निर्भर करता है। उदाहरण के लिए, यह स्थापित किया गया है कि 2 घंटे तक लगातार उड़ने वाली मक्खियों के मोटोकॉन्ड्रिया में बड़ी संख्या में गोलाकार माइटोकॉन्ड्रिया दिखाई देते हैं, जबकि चिपके पंखों वाली मक्खियों में माइटोकॉन्ड्रिया की संख्या बहुत कम होती है और उनका एक रॉड के आकार का आकार होता है ( एल बी लेविंसन)। आकार में, वे एक ही सेल के भीतर भी फिल्मी, छड़ के आकार, गोल और डम्बल के आकार के हो सकते हैं।
माइटोकॉन्ड्रिया कोशिका में स्थानीयकृत होते हैं, एक नियम के रूप में, या तो उन क्षेत्रों में जहां ऊर्जा की खपत होती है, या सब्सट्रेट के संचय के पास (उदाहरण के लिए, लिपिड बूंदें), यदि कोई हो।
माइटोकॉन्ड्रिया का एक सख्त अभिविन्यास शुक्राणु के फ्लैगेला के साथ धारीदार मांसपेशी ऊतक में पाया जाता है, जहां वे मायोफिब्रिल्स के साथ स्थित होते हैं, वृक्क नलिकाओं के उपकला में वे तहखाने की झिल्ली के आक्रमण में स्थानीयकृत होते हैं, आदि।
कोशिकाओं में माइटोकॉन्ड्रिया की संख्या में अंग की विशेषताएं होती हैं, उदाहरण के लिए, चूहे के जिगर की कोशिकाओं में 100 से 2500 माइटोकॉन्ड्रिया होते हैं, और गुर्दे की एकत्रित नलिकाओं की कोशिकाओं में - 300, विभिन्न जानवरों की प्रजातियों के शुक्राणुओं में 20 से 72 तक, एक में विशाल अमीबा अराजकता अराजकताउनकी संख्या 500,000 तक पहुँचती है। माइटोकॉन्ड्रिया का आकार 1 से 10 माइक्रोन तक होता है।
माइटोकॉन्ड्रिया की अल्ट्रामाइक्रोस्कोपिक संरचना उनके आकार और आकार की परवाह किए बिना समान होती है। वे दो लिपोप्रोटीन झिल्ली से ढके होते हैं: बाहरी और आंतरिक। उनके बीच इंटरमेम्ब्रेन स्पेस है।
माइटोकॉन्ड्रियल शरीर में निकलने वाली आंतरिक झिल्ली के आक्रमण कहलाते हैं क्राइस्टे. माइटोकॉन्ड्रिया में क्राइस्ट की व्यवस्था अनुप्रस्थ या अनुदैर्ध्य हो सकती है। क्राइस्ट का आकार सरल या शाखित हो सकता है। कभी-कभी वे एक जटिल नेटवर्क बनाते हैं। कुछ कोशिकाओं में, उदाहरण के लिए, अधिवृक्क ग्रंथि के ग्लोमेरुलर क्षेत्र की कोशिकाओं में, क्राइस्ट नलिकाओं की तरह दिखती हैं। क्राइस्ट की संख्या माइटोकॉन्ड्रिया में होने वाली ऑक्सीडेटिव प्रक्रियाओं की तीव्रता के सीधे आनुपातिक होती है। उदाहरण के लिए, कार्डियोमायोसाइट्स के माइटोकॉन्ड्रिया में हेपेटोसाइट्स के माइटोकॉन्ड्रिया की तुलना में कई गुना अधिक होते हैं। आंतरिक झिल्ली से घिरा हुआ स्थान माइटोकॉन्ड्रिया का आंतरिक कक्ष बनाता है। इसमें, क्राइस्ट के बीच, एक माइटोकॉन्ड्रियल मैट्रिक्स होता है - एक अपेक्षाकृत इलेक्ट्रॉन-घना पदार्थ।
आंतरिक झिल्ली प्रोटीन को माइटोरिबोसोम द्वारा संश्लेषित किया जाता है, जबकि बाहरी झिल्ली प्रोटीन को साइटोरिबोसोम द्वारा संश्लेषित किया जाता है।
"माइटोकॉन्ड्रिया की बाहरी झिल्ली कई मामलों में ईपीएस झिल्ली के समान है। यह ऑक्सीडेटिव एंजाइमों में खराब है। झिल्ली स्थान में उनमें से कुछ हैं। लेकिन आंतरिक झिल्ली और माइटोकॉन्ड्रियल मैट्रिक्स सचमुच उनके साथ संतृप्त हैं। इस प्रकार, क्रेब्स चक्र और फैटी एसिड ऑक्सीकरण के एंजाइम माइटोकॉन्ड्रियल मैट्रिक्स में केंद्रित होते हैं। आंतरिक में झिल्ली इलेक्ट्रॉन परिवहन श्रृंखला, फॉस्फोराइलेशन एंजाइम (एडीपी से एटीपी का गठन), कई परिवहन प्रणालियों द्वारा स्थानीयकृत होती है।
प्रोटीन और लिपिड के अलावा, माइटोकॉन्ड्रियल झिल्ली की संरचना में आरएनए, डीएनए शामिल हैं, बाद में एक आनुवंशिक विशिष्टता है, और इसके भौतिक रासायनिक गुणों में परमाणु डीएनए से भिन्न है।
इलेक्ट्रॉन सूक्ष्म अध्ययनों से पता चला है कि बाहरी झिल्ली की सतह छोटे गोलाकार प्राथमिक कणों से ढकी होती है। आंतरिक झिल्ली और क्राइस्ट में "पैर", तथाकथित मशरूम निकायों पर समान प्राथमिक कण होते हैं। इनमें तीन भाग होते हैं: एक गोलाकार सिर (व्यास 90–100 A°), बेलनाकार पैर, 5 एनएम लंबा और 3–4 एनएम चौड़ा, और आधार 4 बाय 11 एनएम। मशरूम के शरीर के सिर फॉस्फोराइलेशन से जुड़े होते हैं, फिर सिर में एटीपी-आईडी गतिविधि वाला एक एंजाइम पाया जाता है।
इंटरमेम्ब्रेन स्पेस में मैट्रिक्स की तुलना में कम इलेक्ट्रॉन घनत्व वाला पदार्थ होता है। यह झिल्लियों के बीच संचार प्रदान करता है और सहायक उत्प्रेरक-कोएंजाइम के साथ दोनों झिल्लियों में स्थित एंजाइमों की आपूर्ति करता है।
अब यह ज्ञात है कि माइटोकॉन्ड्रिया की बाहरी झिल्ली कम आणविक भार वाले पदार्थों, विशेष रूप से प्रोटीन यौगिकों के लिए अत्यधिक पारगम्य है। माइटोकॉन्ड्रिया की आंतरिक झिल्ली चुनिंदा पारगम्य है। यह आयनों के लिए व्यावहारिक रूप से अभेद्य है (Cl -1, Br -1, SO 4 -2, HCO 3 -1, Sn +2, Mg +2 उद्धरण, कई शर्करा और अधिकांश अमीनो एसिड, जबकि Ca 2+, Mn 2 +, फॉस्फेट, पॉलीकारबॉक्सिलिक एसिड आसानी से इसके माध्यम से प्रवेश करते हैं। कई वाहकों की आंतरिक झिल्ली में उपस्थिति पर डेटा होते हैं जो कि आयनों और उद्धरणों को भेदने के अलग-अलग समूहों के लिए विशिष्ट होते हैं। झिल्ली के माध्यम से पदार्थों का सक्रिय परिवहन के उपयोग के कारण किया जाता है ATPase प्रणाली की ऊर्जा या कार्य श्वसन श्रृंखला के परिणामस्वरूप झिल्ली पर उत्पन्न विद्युत क्षमता। यहां तक कि माइटोकॉन्ड्रिया में संश्लेषित एटीपी भी एक वाहक (युग्मित परिवहन) की मदद से बाहर निकल सकता है।
माइटोकॉन्ड्रियल मैट्रिक्स को एक महीन दाने वाले इलेक्ट्रॉन-घने पदार्थ द्वारा दर्शाया जाता है। इसमें माइटोरिबोसोम, तंतुमय संरचनाएं होती हैं जिनमें डीएनए अणु और दाने होते हैं जिनका व्यास 200A से अधिक होता है, जो लवण द्वारा निर्मित होता है: Ca 3 (PO 4), Ba 3 (PO 4) 2, Mg 3 (PO 4)। ऐसा माना जाता है कि दाने Ca +2 और Mg +2 आयनों के भंडार के रूप में काम करते हैं। माइटोकॉन्ड्रियल झिल्ली की पारगम्यता में परिवर्तन के साथ उनकी संख्या बढ़ जाती है।
माइटोकॉन्ड्रिया में डीएनए की उपस्थिति आरएनए और विशिष्ट प्रोटीन के संश्लेषण में माइटोकॉन्ड्रिया की भागीदारी सुनिश्चित करती है, और साइटोप्लाज्मिक आनुवंशिकता के अस्तित्व को भी इंगित करती है। प्रत्येक माइटोकॉन्ड्रियन में, इसके आकार के आधार पर, एक या अधिक डीएनए अणु (2 से 10 तक) होते हैं। प्रोटोजोआ, खमीर, कवक में माइटोकॉन्ड्रियल डीएनए का आणविक भार लगभग (30-40) * 10 6 होता है। उच्च जंतुओं में लगभग (9–10)*10 6.
खमीर में इसकी लंबाई लगभग 5 माइक्रोन, पौधों में - 30 माइक्रोन के बराबर होती है। माइटोकॉन्ड्रियल डीएनए में निहित आनुवंशिक जानकारी की मात्रा छोटी है: इसमें 15-75 हजार आधार जोड़े होते हैं, जो औसतन 25-125 प्रोटीन श्रृंखलाओं को लगभग 40,000 के आणविक भार के साथ एन्कोड कर सकते हैं।
माइटोकॉन्ड्रियल डीएनए कई मायनों में परमाणु डीएनए से भिन्न होता है: इसमें संश्लेषण की उच्च दर (5–7 गुना) होती है, यह DNase की क्रिया के लिए अधिक प्रतिरोधी होती है, यह दो-रिंग वाला अणु होता है, इसमें अधिक गुआनिन होता है और साइटोसिन, यह उच्च तापमान पर विकृत होता है और इसे ठीक करना आसान होता है। हालांकि, सभी माइटोकॉन्ड्रियल प्रोटीन माइटोकॉन्ड्रियल सिस्टम द्वारा संश्लेषित नहीं होते हैं। इस प्रकार, साइटोक्रोम सी और अन्य एंजाइमों का संश्लेषण नाभिक में निहित जानकारी द्वारा प्रदान किया जाता है। माइटोकॉन्ड्रिया के मैट्रिक्स में, विटामिन ए, बी 2, बी 12, के, ई, साथ ही ग्लाइकोजन स्थानीयकृत होते हैं।
माइटोकॉन्ड्रियल फ़ंक्शन इसमें कोशिकाओं की महत्वपूर्ण गतिविधि के लिए आवश्यक ऊर्जा का निर्माण होता है। विभिन्न यौगिक कोशिका में ऊर्जा के स्रोत के रूप में काम कर सकते हैं: प्रोटीन, वसा, कार्बोहाइड्रेट। हालांकि, ऊर्जा प्रक्रियाओं में तुरंत शामिल होने वाला एकमात्र सब्सट्रेट ग्लूकोज है।
जैविक प्रक्रियाएं, जिसके परिणामस्वरूप माइटोकॉन्ड्रिया में ऊर्जा उत्पन्न होती है, को 3 समूहों में विभाजित किया जा सकता है: समूह I - ऑक्सीडेटिव प्रतिक्रियाएं, जिसमें दो चरण शामिल हैं: अवायवीय (ग्लाइकोलिसिस) और एरोबिक। समूह II - डीफॉस्फोराइलेशन, एटीपी विभाजन और ऊर्जा रिलीज। समूह III - ऑक्सीकरण प्रक्रिया से जुड़ा फॉस्फोराइलेशन।
ग्लूकोज ऑक्सीकरण की प्रक्रिया पहले पाइरुविक या लैक्टिक एसिड में ऑक्सीजन (एनारोबिक या ग्लाइकोलाइटिक तरीके) की भागीदारी के बिना होती है।
हालांकि, केवल थोड़ी मात्रा में ऊर्जा जारी की जाती है। भविष्य में, ये एसिड ऑक्सीजन की भागीदारी के साथ होने वाली ऑक्सीकरण प्रक्रियाओं में शामिल होते हैं, अर्थात वे एरोबिक होते हैं। पाइरुविक और लैक्टिक एसिड के ऑक्सीकरण की प्रक्रिया के परिणामस्वरूप, जिसे क्रेब्स चक्र कहा जाता है, कार्बन डाइऑक्साइड, पानी और बड़ी मात्रा में ऊर्जा बनती है।
परिणामी ऊर्जा गर्मी के रूप में जारी नहीं होती है, जिससे कोशिकाओं का अधिक गर्म होना और पूरे जीव की मृत्यु हो जाती है, लेकिन एडेनोसिन ट्राइफॉस्फोरिक एसिड (एटीपी) के रूप में भंडारण और परिवहन के लिए सुविधाजनक रूप में जमा हो जाती है। एटीपी का संश्लेषण एडीपी और फॉस्फोरिक एसिड से होता है और इसलिए इसे कहा जाता है फास्फारिलीकरण.
स्वस्थ कोशिकाओं में, फास्फारिलीकरण ऑक्सीकरण से जुड़ा होता है। रोगों में, संयुग्मन अछूता हो सकता है, इसलिए सब्सट्रेट ऑक्सीकरण होता है, और फॉस्फोराइलेशन नहीं होता है, और ऑक्सीकरण गर्मी में बदल जाता है, और कोशिकाओं में एटीपी सामग्री कम हो जाती है। नतीजतन, तापमान बढ़ जाता है और कोशिकाओं की कार्यात्मक गतिविधि कम हो जाती है।
तो, माइटोकॉन्ड्रिया का मुख्य कार्य कोशिका की लगभग सभी ऊर्जा का उत्पादन करना है और स्वयं ऑर्गेनॉइड की गतिविधि के लिए आवश्यक घटकों का संश्लेषण, "श्वसन पहनावा", फॉस्फोलिपिड और प्रोटीन के एंजाइम होते हैं।
माइटोकॉन्ड्रिया की गतिविधि का दूसरा पक्ष विशिष्ट संश्लेषण में उनकी भागीदारी है, उदाहरण के लिए, स्टेरॉयड हार्मोन और व्यक्तिगत लिपिड के संश्लेषण में। विभिन्न जानवरों के oocytes में, माइटोकॉन्ड्रिया में जर्दी का संचय बनता है, जबकि वे अपनी मुख्य प्रणाली खो देते हैं। थका हुआ माइटोकॉन्ड्रिया भी उत्सर्जन उत्पादों को जमा कर सकता है।
कुछ मामलों में (यकृत, गुर्दे), माइटोकॉन्ड्रिया हानिकारक पदार्थों और जहरों को जमा करने में सक्षम होते हैं जो कोशिका में प्रवेश करते हैं, उन्हें मुख्य कोशिका द्रव्य से अलग करते हैं और इन पदार्थों के हानिकारक प्रभावों को आंशिक रूप से रोकते हैं। इस प्रकार, माइटोकॉन्ड्रिया अन्य सेल ऑर्गेनेल के कार्यों को संभालने में सक्षम होते हैं, जब सामान्य या चरम परिस्थितियों में किसी विशेष प्रक्रिया को पूरी तरह से समर्थन देने की आवश्यकता होती है।
माइटोकॉन्ड्रिया का जैवजनन। माइटोकॉन्ड्रिया एक छोटे जीवन चक्र के साथ संरचनाओं का नवीनीकरण कर रहे हैं (चूहे के जिगर की कोशिकाओं में, उदाहरण के लिए, माइटोकॉन्ड्रिया का आधा जीवन लगभग 10 दिन है)। माइटोकॉन्ड्रिया का निर्माण पूर्ववर्ती माइटोकॉन्ड्रिया की वृद्धि और विभाजन के परिणामस्वरूप होता है। उनका विभाजन तीन तरीकों से हो सकता है: कसना, छोटे क्षेत्रों का नवोदित होना और माँ के अंदर बेटी माइटोकॉन्ड्रिया का उभरना। माइटोकॉन्ड्रिया का विभाजन (प्रजनन) अपनी आनुवंशिक प्रणाली - माइटोकॉन्ड्रियल डीएनए के प्रजनन से पहले होता है।
इसलिए, अधिकांश शोधकर्ताओं के विचारों के अनुसार, माइटोकॉन्ड्रिया का निर्माण मुख्य रूप से उनके स्व-प्रजनन के माध्यम से होता है।
