§ 2. សមាសធាតុសំខាន់នៃកោសិកា eukaryotic
កោសិកា Eukaryotic (រូបភាពទី 8 និងទី 9) ត្រូវបានរៀបចំស្មុគស្មាញជាងកោសិកា prokaryotic ។ ពួកវាមានទំហំចម្រុះណាស់ (ពីមីក្រូម៉ែត្រជាច្រើនទៅច្រើនសង់ទីម៉ែត្រ) មានរូបរាង និងលក្ខណៈរចនាសម្ព័ន្ធ (រូបភាព 10)។
អង្ករ។ 8. រចនាសម្ព័ន្ធនៃកោសិកា eukaryotic ។ គ្រោងការណ៍ទូទៅ
អង្ករ។ 9. រចនាសម្ព័ន្ធកោសិកាយោងតាមមីក្រូទស្សន៍អេឡិចត្រុង
អង្ករ។ 10. កោសិកា eukaryotic ផ្សេងៗ៖ 1 - epithelial; 2 - ឈាម (e - erythroitis, / - leukitis); 3 - ឆ្អឹងខ្ចី; 4 - ឆ្អឹង; 5 - សាច់ដុំរលោង; 6 - ជាលិកាភ្ជាប់; 7 - កោសិកាសរសៃប្រសាទ; 8 - សរសៃសាច់ដុំ striated
ទោះជាយ៉ាងណាក៏ដោយ អង្គការទូទៅ និងវត្តមាននៃសមាសធាតុជាមូលដ្ឋានគឺដូចគ្នានៅក្នុងកោសិកា eukaryotic ទាំងអស់ (រូបភាព 11) ។
អង្ករ។ 11. កោសិកា Eukaryotic (ដ្យាក្រាម)
Plasmalemma (ភ្នាសកោសិកាខាងក្រៅ) ។មូលដ្ឋាននៃភ្នាសប្លាស្មាដូចជាភ្នាសផ្សេងទៀតនៅក្នុងកោសិកា (ឧទាហរណ៍ mitochondria, plastids ។ ចាប់តាំងពីម៉ូលេគុល lipid គឺប៉ូល (ប៉ូលមួយគឺ hydrophilic ពោលគឺ ទាក់ទាញដោយទឹក និងមួយទៀតគឺ hydrophobic ពោលគឺ repelled ដោយទឹក) ពួកវាត្រូវបានរៀបចំតាមលំដាប់ជាក់លាក់មួយ។ ចុងអ៊ីដ្រូហ្វីលីកនៃម៉ូលេគុលនៃស្រទាប់មួយត្រូវបានតម្រង់ឆ្ពោះទៅរកបរិយាកាស aqueous - ចូលទៅក្នុង cytoplasm នៃកោសិកា និងស្រទាប់ផ្សេងទៀត - នៅខាងក្រៅកោសិកា - ឆ្ពោះទៅរកសារធាតុអន្តរកោសិកា (នៅក្នុងសារពាង្គកាយពហុកោសិកា) ឬបរិយាកាស aqueous (នៅក្នុងសារពាង្គកាយឯកតា។ )
អង្ករ។ 12. រចនាសម្ព័ននៃភ្នាសកោសិកាយោងទៅតាមគំរូវត្ថុរាវ។ ប្រូតេអ៊ីន និង glycoproteins ត្រូវបានជ្រមុជនៅក្នុងស្រទាប់ពីរនៃម៉ូលេគុល lipid ដោយចុង hydrophilic របស់ពួកគេ (រង្វង់) បែរមុខទៅខាងក្រៅ ហើយចុង hydrophobic របស់ពួកគេ (បន្ទាត់រលក) បែរមុខទៅជ្រៅទៅក្នុងភ្នាស។
ម៉ូលេគុលប្រូតេអ៊ីនត្រូវបានបង្កប់ដោយ mosaically នៅក្នុងស្រទាប់ bimolecular នៃ lipids ។ នៅខាងក្រៅកោសិកាសត្វ ម៉ូលេគុល polysaccharide ត្រូវបានភ្ជាប់ទៅនឹង lipids និងម៉ូលេគុលប្រូតេអ៊ីននៃ plasmalemma បង្កើតជា glycolipids និង glycoproteins ។
ការប្រមូលផ្តុំនេះបង្កើតជាស្រទាប់ glycocalyx ។ភ្ជាប់ជាមួយគាត់ មុខងារទទួលភ្នាសប្លាស្មា (សូមមើលខាងក្រោម); វាក៏អាចប្រមូលផ្តុំសារធាតុផ្សេងៗដែលប្រើដោយកោសិកា។ លើសពីនេះទៀត glycocalyx បង្កើនស្ថេរភាពមេកានិចនៃប្លាស្មា។
កោសិការបស់រុក្ខជាតិ និងផ្សិតក៏មានជញ្ជាំងកោសិកាដែលដើរតួនាទីគាំទ្រ និងការពារ។ នៅក្នុងរុក្ខជាតិ វាមានសារជាតិសែលុយឡូស ហើយនៅក្នុងផ្សិត វាត្រូវបានបង្កើតឡើងដោយ chitin ។
ភ្នាសកោសិកាខាងក្រៅអនុវត្តមុខងារមួយចំនួន រួមមានៈ
♦ មេកានិច(ការគាំទ្រ, ការបង្កើតទម្រង់);
♦ របាំង - ការដឹកជញ្ជូន(ភាពជ្រាបចូលនៃសារធាតុផ្សេងៗគ្នា៖ ការចូលទៅក្នុងកោសិកានៃសារធាតុចាំបាច់ និងការដកយកចេញនូវសារធាតុដែលមិនចាំបាច់ និងបង្កគ្រោះថ្នាក់);
♦ អ្នកទទួល(ការកំណត់សារធាតុគីមីជាច្រើនដែលរកឃើញនៅជិតកោសិកា ការយល់ឃើញនៃសញ្ញាក្នុងទម្រង់អ័រម៉ូន ការទទួលស្គាល់ប្រូតេអ៊ីន "បរទេស" ដោយកោសិកានៃប្រព័ន្ធភាពស៊ាំ។ល។)។
ការផ្លាស់ប្តូរសារធាតុរវាងកោសិកា និងបរិស្ថានត្រូវបានអនុវត្តតាមវិធីផ្សេងៗគ្នា - អកម្ម និងសកម្ម។
ម៉ូលេគុលនៃទឹក និងអ៊ីយ៉ុងផ្សេងៗដោយអកម្ម (ដោយសារតែការសាយភាយ, osmosis) ដោយគ្មានកោសិកាចំណាយថាមពលចូលតាមរន្ធញើសពិសេស - ទាំងនេះគឺជា ការដឹកជញ្ជូនអកម្ម។ម៉ាក្រូម៉ូលេគុល ដូចជា ប្រូតេអ៊ីន ប៉ូលីស្យូស សូម្បីតែកោសិកាទាំងមូល ត្រូវបានផ្គត់ផ្គង់ដោយ phagocytosisនិង pinocytosisជាមួយនឹងការប្រើប្រាស់ថាមពល - ការដឹកជញ្ជូនសកម្ម។
ដោយ phagocytosis កោសិកាទាំងមូលឬភាគល្អិតធំត្រូវបានខ្ទាស់ (ឧទាហរណ៍គិតអំពីអាហារូបត្ថម្ភនៅក្នុងអាមីបាឬ phagocytosis ដោយកោសិកាឈាមការពារនៃបាក់តេរី) ។ ក្នុងអំឡុងពេល pinocytosis ភាគល្អិតតូចៗ ឬដំណក់ទឹកនៃសារធាតុរាវត្រូវបានស្រូបយក។ ជាទូទៅចំពោះដំណើរការទាំងពីរគឺថា សារធាតុដែលស្រូបចូលត្រូវបានហ៊ុំព័ទ្ធដោយភ្នាសខាងក្រៅដែលឈ្លានពានដើម្បីបង្កើតជា vacuole ដែលបន្ទាប់មកផ្លាស់ទីជ្រៅទៅក្នុង cytoplasm នៃកោសិកា។
Exocytosis គឺជាដំណើរការមួយ (ជាការដឹកជញ្ជូនសកម្មផងដែរ) ទល់មុខនឹង phagocytosis និង pinocytosis (រូបភាព 13) ។ ដោយមានជំនួយរបស់វា អាហារដែលមិនបានរំលាយនៅក្នុងប្រូតូហ្សូអា ឬសារធាតុសកម្មជីវសាស្រ្តដែលបង្កើតឡើងនៅក្នុងកោសិកាសំងាត់អាចត្រូវបានយកចេញ។
ស៊ីតូប្លាស្មា។ Cytoplasm គឺជាខ្លឹមសារនៃកោសិកា ដែលកំណត់ដោយប្លាស្មា លើកលែងតែស្នូល។ វាមាន សារធាតុដី (hyaloplasm), សរីរាង្គនិង ការរួមបញ្ចូល។
Hyaloplasma- វត្ថុរាវដែលមានជាតិ viscous អាចស្ថិតនៅក្នុងស្ថានភាពណាមួយ។ ហ្សូឡា(រាវ) ឬ ជែល(ដូចចាហួយ)។
បើចាំបាច់ cytoplasm មានសមត្ថភាពផ្លាស់ប្តូរបញ្ច្រាសពីរដ្ឋមួយទៅរដ្ឋមួយទៀត។ ឧទាហរណ៍ក្នុងអំឡុងពេលចលនា amoeboid (ចងចាំផ្នែក "Protozoa" ពីវគ្គសិក្សានៃសត្វវិទ្យា) ក្នុងអំឡុងពេលនៃការបង្កើត pseudopod ការផ្លាស់ប្តូរយ៉ាងឆាប់រហ័សនៃ cytoplasm ពីជែលទៅ sol និងច្រាសមកវិញកើតឡើង។ នេះគឺដោយសារតែវត្តមាននៅក្នុង cytoplasm នៃចំនួនដ៏ច្រើននៃម៉ូលេគុលប្រូតេអ៊ីន filamentous អាទីណា។នៅពេលដែលពួកវាភ្ជាប់គ្នាទៅវិញទៅមកដើម្បីបង្កើតជាបណ្តាញបីវិមាត្រ cytoplasm ស្ថិតនៅក្នុងស្ថានភាពជែលមួយ ហើយនៅពេលដែលបណ្តាញនេះបែកខ្ញែក វាស្ថិតនៅក្នុងស្ថានភាពសូល។
hyaloplasm មានសារធាតុផ្សេងៗ - អង់ស៊ីម ប្រូតេអ៊ីន កាបូអ៊ីដ្រាត ខ្លាញ់ និងសារធាតុសរីរាង្គ និងសារធាតុរ៉ែ។ ដំណើរការគីមីផ្សេងៗកើតឡើងនៅទីនេះ - ការបំបែកសារធាតុ ការសំយោគ និងការកែប្រែ (ការផ្លាស់ប្តូរ)។
សរីរាង្គ។ទាំងនេះគឺជាសមាសធាតុអចិន្ត្រៃយ៍នៃកោសិកាដែលមានរចនាសម្ព័ន្ធ និងមុខងារជាក់លាក់ ដែលមានទីតាំងនៅក្នុង cytoplasm របស់វា។ នៅក្នុងអ្វីដែលខាងក្រោមយើងនឹងនិយាយអំពី សរីរាង្គគោលបំណងទូទៅ,មាននៅក្នុងកោសិកាគ្រប់ប្រភេទនៃ eukaryotes ទាំងអស់។ ពួកវាត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការធានានូវមុខងារសំខាន់ៗនៃមុខងារចុងក្រោយ។ អង្គការគោលបំណងពិសេសត្រូវបានរកឃើញតែនៅក្នុងកោសិកានៃប្រភេទជាក់លាក់មួយ (ឯកទេសខ្ពស់) - ឧទាហរណ៍ myofibrils នៅក្នុងកោសិកាសាច់ដុំ។
សរីរាង្គនៃគោលបំណងទូទៅមានរចនាសម្ព័ន្ធដូចគ្នា ដោយមិនគិតពីកោសិកា និងសារពាង្គកាយណាមួយដែលពួកវាជាកម្មសិទ្ធិ។ ប៉ុន្តែក្នុងចំណោមពួកគេមានក្រុមដែលមានភ្នាស (endoplasmic reticulum, Golgi apparatus, mitochondria, plastids, lysosomes, vacuoles),ក៏ដូចជាមិនមែនភ្នាស ( ribosomes មជ្ឈមណ្ឌលកោសិកា)រចនាសម្ព័ន្ធ។
រ៉េទីកូឡាមអេនដូប្លាសមីម (ER) ។ EPS មានភ្នាស និងជាប្រព័ន្ធសាខាស្មុគស្មាញនៃ tubules និង cisterns ដែលជ្រាបចូលទៅក្នុង cytoplasm ទាំងមូលនៃកោសិកា (រូបភាព 14) ។ EPS មានពីរប្រភេទ - រដុបនិង រលោង។ Ribosomes ត្រូវបានភ្ជាប់ទៅនឹងភ្នាសរដុប (ពីផ្នែកខាង cytoplasmic) ប៉ុន្តែវាមិនមានវត្តមាននៅលើភ្នាសរលោងនោះទេ។
អង្ករ។ 14. Endoplasmic reticulum
reticulum endoplasmic អនុវត្តមុខងារសំខាន់ៗមួយចំនួននៅក្នុងកោសិកា eukaryotic:
♦ កំណត់ព្រំដែន(ការបែងចែកបរិមាណខាងក្នុងនៃកោសិកាទៅជាចន្លោះប្រតិកម្មផ្សេងៗ);
♦ ការចូលរួមក្នុងការសំយោគសារធាតុសរីរាង្គ(ribosomes មានទីតាំងនៅលើភ្នាសនៃ ER រដុប ហើយអង់ស៊ីមស្មុគ្រស្មាញមានទីតាំងនៅលើរលោង ធានាការសំយោគនៃ lipids កាបូអ៊ីដ្រាត ។ល។
♦ ការចូលរួមក្នុងការបង្កើតធាតុនៃបរិធាន Golgi, lysosomes;
♦ ការដឹកជញ្ជូនសារធាតុ។
ឧបករណ៍ Golgi ។ឧបករណ៍ Golgi (AG) គឺជាប្រព័ន្ធមួយ។ រថក្រោះ(vacuoles ផ្ទះល្វែង) និង ពពុះ(vesicles) ដែលមានទីតាំងនៅជិតស្នូលកោសិកាដែលត្រូវបានបង្កើតឡើងដោយសារតែ EPS ដែលជាលទ្ធផលនៃការបំបែកនៃបំណែកតូចៗរបស់វា (រូបភាព 15) ។ នៅពេលដែលបំណែកទាំងនេះបញ្ចូលគ្នា អាងស្តុកទឹកថ្មីនៃបរិធាន Golgi កើតឡើង ខណៈពេលដែលសារធាតុផ្សេងៗត្រូវបានដឹកជញ្ជូនពី EPS ដែលចូលរួមក្នុងការប្រមូលផ្តុំនៃសមាសធាតុសរីរាង្គស្មុគស្មាញ (ប្រូតេអ៊ីន + កាបូអ៊ីដ្រាត ប្រូតេអ៊ីន + lipid ជាដើម) ដែលត្រូវបានយកចេញជាមួយ ជំនួយរបស់ AG នៅខាងក្រៅកោសិកា។ សារធាតុសកម្មជីវសាស្រ្តទាំងនេះត្រូវបានដកចេញពីកោសិកា (ដោយប្រើ secretory vacuoles ដោយ exocytosis) ឬជាផ្នែកមួយនៃ lysosomes (សូមមើលខាងក្រោម) ដែលបង្កើតឡើងដោយ AG ។
អង្ករ។ 15. ឧបករណ៍ Golgi៖
ឧបករណ៍ Golgi អនុវត្តមុខងារដូចខាងក្រោមៈ
♦ ការសំយោគសារធាតុសកម្មជីវសាស្រ្តដែលផលិតដោយកោសិកា;
♦ ការសម្ងាត់ (ការយកចេញពីកោសិកា) នៃសារធាតុផ្សេងៗ(អ័រម៉ូន អង់ស៊ីម សារធាតុដែលជញ្ជាំងកោសិកាត្រូវបានសាងសង់។ល។);
♦ ការចូលរួមក្នុងការបង្កើត lysosomes ។
មីតូខន់ឌ្រី។គ្រប់ប្រភេទនៃកោសិកា eukaryotic មាន mitochondria (រូបភាព 16) ។ ពួកវាមើលទៅដូចជារាងមូល ឬកំណាត់ មិនសូវជាញឹកញាប់ទេ - ខ្សែស្រឡាយ។ ទំហំរបស់ពួកគេមានចាប់ពី 1 ដល់ 7 មីក្រូ។ ចំនួននៃ mitochondria នៅក្នុងកោសិកាមួយមានចាប់ពីរាប់រយទៅរាប់ម៉ឺននាក់ (នៅក្នុងប្រូតូហ្សូអាធំ)។
អង្ករ។ 16. Mitochondria ។ ខាងលើ - mitochondria (a) នៅក្នុងប្រឡាយទឹកនោមដែលអាចមើលឃើញនៅក្នុងមីក្រូទស្សន៍ពន្លឺ។ ខាងក្រោមនេះគឺជាគំរូបីវិមាត្រនៃអង្គការ mitochondrial: 1 - cristae; 2 - ភ្នាសខាងក្រៅ; 3 - ភ្នាសខាងក្នុង; ៤- ម៉ាទ្រីស
mitochondrion ត្រូវបានបង្កើតឡើងដោយភ្នាសពីរ - ខាងក្រៅនិង ខាងក្នុង,រវាងដែលមានទីតាំងនៅ ចន្លោះអន្តរភ្នាស។ភ្នាសខាងក្នុងបង្កើតបានជាអវយវៈជាច្រើន - cristae ដែលជាចានឬបំពង់។ អង្គការនេះផ្តល់នូវតំបន់ដ៏ធំនៃភ្នាសខាងក្នុង។ វាមានអង់ស៊ីមដែលធានាការបំប្លែងថាមពលដែលមាននៅក្នុងសារធាតុសរីរាង្គ (កាបូអ៊ីដ្រាត ខ្លាញ់) ទៅជាថាមពល ATP ដែលចាំបាច់សម្រាប់ជីវិតរបស់កោសិកា។ ដូច្នេះមុខងាររបស់ mitochondria គឺចូលរួម ថាមពលដំណើរការកោសិកា។ នោះហើយជាមូលហេតុដែលមួយចំនួនធំនៃ mitochondria មាននៅក្នុងកោសិកាសាច់ដុំដែលធ្វើការច្រើន។
ផ្លាស្ទិច។នៅក្នុងកោសិការុក្ខជាតិ សរីរាង្គពិសេសត្រូវបានរកឃើញ - plastids ដែលជារឿយៗមានរាងមូល ឬរាងមូល ជួនកាលស្មុគស្មាញជាង។ ផ្លាស្ទីតមានបីប្រភេទ - ក្លរ៉ូផ្លាស្ទីស (រូបភាពទី 17) ក្រូម៉ូប្លាស និងលីកូប្លាស។
Chloroplastពណ៌បៃតងខុសគ្នា ដែលបណ្តាលមកពីសារធាតុពណ៌ ក្លរ៉ូហ្វីលធានាដំណើរការ រស្មីសំយោគឧ. ការសំយោគសារធាតុសរីរាង្គពីទឹក (H 2 O) និងកាបូនឌីអុកស៊ីត (CO 2) ដោយប្រើថាមពលនៃពន្លឺព្រះអាទិត្យ។ Chloroplast ត្រូវបានរកឃើញជាចម្បងនៅក្នុងកោសិកាស្លឹក (នៅក្នុងរុក្ខជាតិខ្ពស់ជាង) ។ ពួកវាត្រូវបានបង្កើតឡើងដោយភ្នាសពីរដែលមានទីតាំងនៅស្របគ្នាទៅវិញទៅមកជុំវិញមាតិកានៃ chloroplasts - ស្ត្រូម៉ា។ភ្នាសខាងក្នុងបង្កើតជាថង់សំប៉ែតជាច្រើន - thylakoids,ដែលត្រូវបានជង់ (ដូចជាកាក់កាក់) - គ្រាប់ធញ្ញជាតិ -ហើយដេកនៅក្នុង stroma ។ វាគឺជា thylakoids ដែលមានក្លរ៉ូហ្វីល។
Chromoplastsកំណត់ពណ៌លឿង ពណ៌ទឹកក្រូច និងពណ៌ក្រហមនៃផ្កា និងផ្លែឈើជាច្រើននៅក្នុងកោសិកាដែលពួកវាមានវត្តមានក្នុងបរិមាណដ៏ច្រើន។ សារធាតុពណ៌សំខាន់ៗនៅក្នុងសមាសភាពរបស់ពួកគេគឺ carotene ។គោលបំណងមុខងាររបស់ chromoplasts គឺដើម្បីទាក់ទាញសត្វដែលមានពណ៌ ធានាការលំអងនៃផ្កា និងការបែកខ្ញែកនៃគ្រាប់ពូជ។
អង្ករ។ 17. Plastids: a – chloroplasts នៅក្នុងកោសិកានៃស្លឹក Elodea ដែលអាចមើលឃើញនៅក្នុងមីក្រូទស្សន៍ពន្លឺ។ ខ - ដ្យាក្រាមនៃរចនាសម្ព័ន្ធខាងក្នុងនៃ chloroplast ជាមួយ grana ដែលជាជង់នៃថង់សំប៉ែតដែលមានទីតាំងនៅកាត់កែងទៅនឹងផ្ទៃនៃ chloroplast; គ - ដ្យាក្រាមលម្អិតបន្ថែមទៀតដែលបង្ហាញពីបំពង់ anastomosing តភ្ជាប់បន្ទប់កង្ហារនីមួយៗ
លីកូប្លាសគឺជាផ្លាស្ទីតដែលគ្មានពណ៌ដែលមាននៅក្នុងកោសិកានៃផ្នែកក្រោមដីនៃរុក្ខជាតិ (ឧទាហរណ៍នៅក្នុងមើមដំឡូង) គ្រាប់ពូជ និងស្នូលនៃដើម។ នៅក្នុង leukoplasts ម្សៅត្រូវបានបង្កើតឡើងជាចម្បងពីគ្លុយកូសនិងកកកុញនៅក្នុងសរីរាង្គផ្ទុករបស់រុក្ខជាតិ។
Plastids នៃប្រភេទមួយអាចបំប្លែងទៅជាមួយផ្សេងទៀត។ ឧទាហរណ៍ នៅពេលដែលស្លឹកផ្លាស់ប្តូរពណ៌ក្នុងរដូវស្លឹកឈើជ្រុះ chloroplasts ផ្លាស់ប្តូរទៅជា chromoplasts ។
លីសូសូម។សរីរាង្គទាំងនេះមើលទៅដូចជា vesicles ព័ទ្ធជុំវិញដោយភ្នាសដែលមានអង្កត់ផ្ចិតរហូតដល់ 2 មីក្រូ។ ពួកវាមានអង់ស៊ីមរាប់សិបដែលបំបែកប្រូតេអ៊ីន អាស៊ីតនុយក្លេអ៊ីក ប៉ូលីសាខ័រ និងលីពីត។ មុខងាររបស់ lysosomes គឺចូលរួមនៅក្នុងដំណើរការនៃការបំបែក intracellular នៃសមាសធាតុសរីរាង្គស្មុគស្មាញ (ឧទាហរណ៍ សារធាតុចិញ្ចឹម ឬសារធាតុនៃសមាសធាតុកោសិកា "ចំណាយ")។ Lysosomes ផ្សំជាមួយ vacuoles phagocytic (ឬ pinocytic) ដើម្បីបង្កើតជា vacuole រំលាយអាហារ។
ការបង្កើត lysosomes កើតឡើងដោយសារតែការពន្លកចេញពី cisternae នៃឧបករណ៍ Golgi ។
រីបូសូម។ Ribosomes (Fig ។ 18) មានវត្តមាននៅក្នុងកោសិកានៃ eukaryotes និង prokaryotes ចាប់តាំងពីពួកវាបំពេញមុខងារសំខាន់នៅក្នុង ជីវសំយោគប្រូតេអ៊ីន(សូមមើលជំពូកទី 5) ។ កោសិកានីមួយៗមានរាប់សិបពាន់ (រហូតដល់រាប់លាន) នៃសរីរាង្គរាងមូលតូចៗទាំងនេះ។
អង្ករ។ 18. គ្រោងការណ៍នៃរចនាសម្ព័ន្ធនៃ ribosome មួយដែលអង្គុយនៅលើភ្នាសនៃ reticulum endoplasmic: 1 - subunit តូច; 2 - tRNA; 3 - aminoacyl-tRNA; 4 - អាស៊ីតអាមីណូ; 5 - ផ្នែករងធំ; 6 - ភ្នាស reticulum endoplasmic; 7 - ខ្សែសង្វាក់ polypeptide សំយោគ
ribosome មានផ្នែករងមិនស្មើគ្នាពីរ (ផ្នែក)។ ពួកវាត្រូវបានបង្កើតឡើងដោយឡែកពីគ្នា និងផ្សំ RNA នៃអ្នកនាំសារ "គ្របដណ្តប់" កំឡុងពេលសំយោគម៉ូលេគុលប្រូតេអ៊ីន។ Ribosomes មានប្រូតេអ៊ីន និង ribosomal RNAs ជាច្រើនប្រភេទ។
ការរួមបញ្ចូលកោសិកា។នេះគឺជាឈ្មោះដែលបានផ្តល់ឱ្យសមាសធាតុមិនអចិន្ត្រៃយ៍នៅក្នុងកោសិកាដែលមានវត្តមាននៅក្នុងសារធាតុសំខាន់នៃ cytoplasm ក្នុងទម្រង់ជាគ្រាប់ធញ្ញជាតិ granules ឬដំណក់ទឹក។ ការរួមបញ្ចូលអាចឬមិនអាចត្រូវបានហ៊ុំព័ទ្ធដោយភ្នាស។
តាមមុខងារ ការដាក់បញ្ចូលមានបីប្រភេទ៖ សារធាតុចិញ្ចឹមបម្រុង(ម្សៅ, glycogen, ខ្លាញ់, ប្រូតេអ៊ីន), ការរួមបញ្ចូលអាថ៌កំបាំង(សារធាតុលក្ខណៈនៃកោសិកាក្រពេញផលិតដោយពួកវា - អរម៉ូននៃក្រពេញ endocrine ។ល។) និង ការរួមបញ្ចូលគោលបំណងពិសេស(នៅក្នុងកោសិកាឯកទេសខ្ពស់ ឧទាហរណ៍ អេម៉ូក្លូប៊ីនក្នុងកោសិកាឈាមក្រហម)។
§ 3. ការរៀបចំនៃស្នូលកោសិកា។ ក្រូម៉ូសូម
ស្នូលកោសិកា (សូមមើលរូបភាពទី 8 និងទី 9) គឺមានសារៈសំខាន់បំផុតក្នុងជីវិតរបស់កោសិកា ដោយសារវាបម្រើជាឃ្លាំងនៃព័ត៌មានតំណពូជដែលមាននៅក្នុងក្រូម៉ូសូម (សូមមើលខាងក្រោម)។
ស្នូលត្រូវបានចងភ្ជាប់ដោយស្រោមសំបុត្រនុយក្លេអ៊ែរដែលបំបែកមាតិការបស់វា។ (ការ៉ាយ៉ូប្លាស្មា)ពី cytoplasm ។ សែលមានភ្នាសពីរដែលបំបែកដោយគម្លាតមួយ។ ពួកវាទាំងពីរត្រូវបាន permeated ជាមួយរន្ធញើសជាច្រើន, អរគុណដែលការផ្លាស់ប្តូរនៃសារធាតុរវាងស្នូលនិង cytoplasm គឺអាចធ្វើទៅបាន។ នៅក្នុងស្នូលកោសិកានៃ eukaryotes ភាគច្រើនមានពី 1 ទៅ 7 នុយក្លេអូលី។ដំណើរការនៃការសំយោគ RNA និង tRNA ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងពួកគេ។
សមាសធាតុខឺណែលចម្បង - ក្រូម៉ូសូមបង្កើតឡើងពីម៉ូលេគុល DNA និងប្រូតេអ៊ីនផ្សេងៗ។ នៅក្នុងមីក្រូទស្សន៍ពន្លឺពួកវាអាចមើលឃើញយ៉ាងច្បាស់តែក្នុងអំឡុងពេលនៃការបែងចែកកោសិកាប៉ុណ្ណោះ។ ( mitosis, meiosis) ។នៅក្នុងកោសិកាដែលមិនបែងចែក ក្រូម៉ូសូមមើលទៅដូចជាខ្សែស្តើងវែងដែលចែកចាយពាសពេញបរិមាណទាំងមូលនៃស្នូល។
ក្នុងអំឡុងពេលនៃការបែងចែកកោសិកា ខ្សែក្រូម៉ូសូមបង្កើតជាវង់ក្រាស់ ជាលទ្ធផលដែលពួកវាអាចមើលឃើញ (ដោយប្រើមីក្រូទស្សន៍ធម្មតា) ក្នុងទម្រង់ជាកំណាត់ "កន្សែងសក់"។ បរិមាណទាំងមូលនៃព័ត៌មានហ្សែនត្រូវបានចែកចាយរវាងក្រូម៉ូសូមនៃស្នូល។ នៅក្នុងដំណើរការនៃការសិក្សាពួកគេ គំរូខាងក្រោមត្រូវបានកំណត់៖
♦ នៅក្នុងស្នូលនៃកោសិកា somatic (ពោលគឺកោសិការាងកាយ មិនមែនផ្លូវភេទ) បុគ្គលទាំងអស់នៃប្រភេទដូចគ្នាមានចំនួនក្រូម៉ូសូមដូចគ្នាដែលបង្កើតបានជា សំណុំនៃក្រូម៉ូសូម(រូបភព 19);
អង្ករ។ 19. ក្រូម៉ូសូមនៃប្រភេទរុក្ខជាតិ និងសត្វផ្សេងៗគ្នា ដែលបង្ហាញនៅលើមាត្រដ្ឋានដូចគ្នា៖ 1,2 – អាមីបា; 3,4 - diatoms; 5-8, 18,19 - សារាយបៃតង; 9 - រុយ agaric; 10 - លីនដិន; ១១-១២ - ដ្រូសូហ្វីឡា; 13 - ត្រី salmon; 14 - skerda (គ្រួសារ Asteraceae); 15 - រុក្ខជាតិពីគ្រួសារ aroid; 16 - មេអំបៅ Corydalis; 17 - សត្វល្អិតពីគ្រួសារកណ្ដូប; 20 - មេរោគ stride ទឹក; 21 - សត្វល្អិតផ្កា; 22 - Amphibian Ambystoma; 23 - aloe (គ្រួសារ Lily)
♦ ប្រភេទសត្វនីមួយៗត្រូវបានកំណត់ដោយក្រូម៉ូសូមផ្ទាល់ខ្លួនរបស់វាយោងទៅតាមចំនួនរបស់វា (ឧទាហរណ៍មនុស្សម្នាក់មានក្រូម៉ូសូម 46 រុយ Drosophila មាន 8 ដង្កូវមូលមាន 4 ក្តាមមាន 196 សេះមាន 66 និងពោតមាន 104) ។
♦ ក្រូម៉ូសូមនៅក្នុងស្នូលនៃកោសិកា somatic អាចត្រូវបានដាក់ជាក្រុមជាគូហៅថា ក្រូម៉ូសូមដូចគ្នា។ដោយផ្អែកលើភាពស្រដៀងគ្នារបស់ពួកគេ (នៅក្នុងរចនាសម្ព័ន្ធនិងមុខងារ);
♦ នៅក្នុងស្នូលនៃកោសិកាមេរោគ (gametes) គូនៃក្រូម៉ូសូមដូចគ្នាមានតែមួយ ពោលគឺសំណុំក្រូម៉ូសូមសរុបគឺពាក់កណ្តាលនៃកោសិកា somatic ។
♦ សំណុំក្រូម៉ូសូមតែមួយនៅក្នុងកោសិកាមេរោគត្រូវបានគេហៅថា haploidនិងត្រូវបានតំណាងដោយអក្សរ n និងនៅក្នុង somatic - diploid(2 ន)
ពីខាងលើវាច្បាស់ណាស់ថាគូនៃក្រូម៉ូសូមដូចគ្នាត្រូវបានបង្កើតឡើងដោយការរួបរួមនៃក្រូម៉ូសូមមាតានិងមាតាអំឡុងពេលបង្កកំណើត ពោលគឺការលាយបញ្ចូលគ្នានៃកោសិកាផ្លូវភេទ (gametes)។ ផ្ទុយទៅវិញ ក្នុងអំឡុងពេលនៃការបង្កើតកោសិកាមេរោគ ពីក្រូម៉ូសូមដូចគ្នានីមួយៗ មានតែមួយប៉ុណ្ណោះដែលចូលទៅក្នុង gamete ។
ក្រូម៉ូសូម ខុសគ្នាគូ homologous ខុសគ្នាក្នុងទំហំ និងរូបរាង (រូបភាព 20 និង 21)។
អង្ករ។ 20. រចនាសម្ព័ន្ធ និងប្រភេទនៃក្រូម៉ូសូម៖ a – រូបរាង 1 – centromere; 2 - ស្មាខ្លី; 3 - ស្មាវែង); រចនាសម្ព័ន្ធខាងក្នុងនៃក្រូម៉ូសូមដូចគ្នា (1 - centromere; - ម៉ូលេគុល DNA); គ - ប្រភេទនៃក្រូម៉ូសូម (1 - ប្រដាប់អាវុធតែមួយ; អាវុធផ្សេងគ្នា; 3 - អាវុធស្មើគ្នា: X - ដៃ, V - កណ្តាល)
អង្ករ។ 21. ក្រូម៉ូសូមមួយមាន DNA និងប្រូតេអ៊ីន។ ម៉ូលេគុល DNA ត្រូវបានចម្លង។ ឧទ្ធម្ភាគចក្រពីរ DNA ដូចគ្នាបេះបិទនៅតែតភ្ជាប់គ្នានៅតំបន់កណ្តាល។ ច្បាប់ចម្លងទាំងនេះក្លាយជាក្រូម៉ូសូមដាច់ដោយឡែកនៅពេលក្រោយកំឡុងពេលបែងចែកកោសិកា
នៅក្នុងរាងកាយនៃក្រូម៉ូសូមមាន ការរឹតបន្តឹងបឋម (ហៅថាកណ្តាល)ដែលខ្សែស្រឡាយត្រូវបានភ្ជាប់ spindles ប្រសព្វ។វាបែងចែកក្រូម៉ូសូមជាពីរ ស្មាក្រូម៉ូសូមអាចមានអាវុធស្មើគ្នា អាវុធមិនស្មើគ្នា ឬអាវុធតែមួយ។
ជំពូកទី 5. ការរំលាយអាហារ
§ 1. Metabolism ជាការរួបរួមនៃ assimilation និង dissimilation
កោសិកា និងសារពាង្គកាយមានជីវិតទាំងអស់គឺជាប្រព័ន្ធបើកចំហ ពោលគឺពួកវាស្ថិតក្នុងស្ថានភាពនៃការផ្លាស់ប្តូរថាមពល និងសារធាតុជាបន្តបន្ទាប់ជាមួយបរិស្ថាន។ មានប្រព័ន្ធបើកចំហនៅក្នុងធម្មជាតិគ្មានជីវិត ប៉ុន្តែអត្ថិភាពរបស់ពួកវាមានលក្ខណៈគុណភាពខុសពីសារពាង្គកាយមានជីវិត។ សូមពិចារណាឧទាហរណ៍នេះ៖ ដុំស្ពាន់ធ័រដែលឆេះនៅក្នុងស្ថានភាពផ្លាស់ប្តូរជាមួយបរិស្ថាន។ នៅពេលដែលវាឆេះ O 2 ត្រូវបានស្រូបយកហើយ SO 2 និងថាមពល (ក្នុងទម្រង់ជាកំដៅ) ត្រូវបានបញ្ចេញ។ ទោះយ៉ាងណាក៏ដោយក្នុងករណីនេះដុំស្ពាន់ធ័រដែលជារូបរាងកាយត្រូវបានបំផ្លាញហើយបាត់បង់រចនាសម្ព័ន្ធចម្បងរបស់វា។
សម្រាប់សារពាង្គកាយមានជីវិត ការផ្លាស់ប្តូរជាមួយបរិស្ថានប្រែទៅជាលក្ខខណ្ឌមួយសម្រាប់ការថែរក្សា និងថែរក្សារចនាសម្ព័ន្ធរបស់ពួកវាតាមរយៈការបន្តឡើងវិញដោយខ្លួនឯងនូវសារធាតុ និងសមាសធាតុទាំងអស់ដែលពួកវាមាន។
មេតាបូលីស (Metabolism) គឺជាសំណុំនៃដំណើរការដែលកើតឡើងនៅក្នុងសារពាង្គកាយមានជីវិត (ការប្រើប្រាស់ ការផ្លាស់ប្តូរ ការប្រមូលផ្តុំ និងការបញ្ចេញសារធាតុ និងថាមពល) ដែលធានានូវសកម្មភាពសំខាន់ៗ ការអភិវឌ្ឍន៍ ការលូតលាស់ ការបន្តពូជ។ កំឡុងពេលដំណើរការមេតាបូលីស ម៉ូលេគុលដែលបង្កើតកោសិកាត្រូវបានបំបែក និងសំយោគ។ ការបន្តនៃរចនាសម្ព័ន្ធកោសិកា និងសារធាតុអន្តរកោសិកា។
ការរំលាយអាហារគឺផ្អែកលើដំណើរការដែលទាក់ទងគ្នាទៅវិញទៅមក assimilation(anabolism) និង ការបង្ខូចទ្រង់ទ្រាយ(catabolism) ។ ក្នុងអំឡុងពេល assimilation (ការផ្លាស់ប្តូរផ្លាស្ទិច) សារធាតុស្មុគស្មាញត្រូវបានសំយោគពីវត្ថុសាមញ្ញ។ វាគឺជាការអរគុណចំពោះបញ្ហានេះដែលសារធាតុសរីរាង្គទាំងអស់នៅក្នុងកោសិកាដែលចាំបាច់សម្រាប់ការសាងសង់សមាសធាតុរចនាសម្ព័ន្ធរបស់វាប្រព័ន្ធអង់ស៊ីមជាដើមត្រូវបានបង្កើតឡើង។ Assimilation តែងតែត្រូវបានអនុវត្តជាមួយនឹងការចំណាយថាមពល។
កំឡុងពេលបំបែក (ការបំប្លែងថាមពល) សារធាតុសរីរាង្គស្មុគ្រស្មាញត្រូវបានបំបែកទៅជាសារធាតុសាមញ្ញ ឬអសរីរាង្គ។ ក្នុងករណីនេះថាមពលត្រូវបានបញ្ចេញដែលត្រូវបានចំណាយដោយកោសិកាដើម្បីអនុវត្តដំណើរការផ្សេងៗដែលធានានូវសកម្មភាពសំខាន់របស់វា (ការសំយោគ និងការដឹកជញ្ជូនសារធាតុ ការងារមេកានិច។ល។)។
ភាវៈរស់ទាំងអស់អាចបែងចែកជាពីរក្រុម៖ autotrophsនិង heterotrophs,ដែលមានភាពខុសប្លែកគ្នានៅក្នុងប្រភពថាមពល និងសារធាតុចាំបាច់ ដើម្បីធានាបាននូវមុខងារសំខាន់ៗរបស់វា។
អូតូត្រូហ្វ- សារពាង្គកាយដែលសំយោគសមាសធាតុសរីរាង្គពីសារធាតុអសរីរាង្គដោយប្រើប្រាស់ថាមពលនៃពន្លឺព្រះអាទិត្យ (ដូច phototrophs- រុក្ខជាតិ cyanobacteria) ឬថាមពលដែលទទួលបានពីការកត់សុីនៃសារធាតុរ៉ែ (អសរីរាង្គ) (ដូចជា គីមីវិទ្យា- បាក់តេរីស្ពាន់ធ័រ បាក់តេរីដែក ។ល។) អាស្រ័យហេតុនេះ ពួកគេអាចបង្កើតសារធាតុចាំបាច់សម្រាប់សកម្មភាពជីវិតរបស់ពួកគេដោយឯករាជ្យ។
§ 2. Dissimilation នៅក្នុងសារពាង្គកាយ anaerobic និង aerobic
សារពាង្គកាយអាចចែកចេញជាពីរក្រុម និងយោងទៅតាមលក្ខណៈនៃការបែងចែក - អេរ៉ូប៊ីនិង anaerobes ។ Aerobes (មកពីភាសាក្រិក។ បិសាច- ខ្យល់) ត្រូវការអុកស៊ីសែនដោយឥតគិតថ្លៃសម្រាប់ជីវិត។ នៅក្នុង anaerobes (ភាសាក្រិច។ ac- ភាគល្អិតអវិជ្ជមាន) វាមិនចាំបាច់ទេ។ នៅក្នុងពួកគេការបំបែកត្រូវបានអនុវត្តដោយការ fermentation - គ្មានអុកស៊ីហ៊្សែនការបំបែកអង់ស៊ីមនៃសារធាតុសរីរាង្គជាមួយនឹងការបង្កើតសារធាតុសរីរាង្គសាមញ្ញនិងការបញ្ចេញថាមពល។ ឧទាហរណ៍:
♦ អាស៊ីតឡាក់ទិក fermentation:
C 6 H 12 O 6 + 2H 3 PO 4 + 2ADP → 2PH + 2ATP + 2H 2 O;
♦ ជាតិអាល់កុល fermentation:
C 6 H 12 O 6 + 2PH + 2ADP → 2C 2 H 5 OH + 2CO 2 + 2ATP + 2H 2 O ។
សារធាតុដែលបានបង្កើតឡើងកំឡុងពេល fermentation គឺសរីរាង្គ ហើយដូច្នេះនៅតែមានថាមពលច្រើន។
អង្ករ។ 22. ទំនាក់ទំនងរវាង assimilation និង dissimilation នៅក្នុង autotrophic និង heterotrophic organisms
នៅក្នុងសារពាង្គកាយ aerobic ក្នុងអំឡុងពេលដំណើរការនៃការដកដង្ហើមនៅក្នុង mitochondria ការបំបែកពេញលេញនៃសារធាតុសរីរាង្គកើតឡើង (ដោយប្រើ O 2) ទៅនឹងផលិតផលចុងថាមពលដែលខ្សោយ CO 2 និង H 2 O ហើយបរិមាណថាមពលកាន់តែច្រើនត្រូវបានបញ្ចេញ៖
C 6 H 12 0 6 (គ្លុយកូស) + 0 2 > 6C0 2 + 6H 2 0 + ថាមពល (ដោយសារម៉ូលេគុល ATP 38 ត្រូវបានសំយោគ)។
ចូរយើងពិចារណាក្នុងទម្រង់នៃគ្រោងការណ៍ទូទៅនៃការរំលាយអាហារនៃសារពាង្គកាយ aerobic autotrophic និង heterotrophic (រូបភាព 22) ។
ការរួមផ្សំ។ដំណើរការសំខាន់បំផុតរបស់វាគឺ រស្មីសំយោគនិង biosynthesis នៃប្រូតេអ៊ីន។
ការតភ្ជាប់នៃសារពាង្គកាយជាមួយបរិស្ថាន តាមទស្សនៈរូបវិទ្យា គឺជាប្រព័ន្ធបើកចំហ នោះគឺជាប្រព័ន្ធដែលដំណើរការជីវគីមីកំពុងដំណើរការ។ សារធាតុចាប់ផ្តើមចេញមកពីបរិស្ថាន ហើយសារធាតុដែលត្រូវបានបង្កើតឡើងជាបន្តបន្ទាប់ក៏ត្រូវបាននាំយកទៅខាងក្រៅ។ លំនឹងរវាងល្បឿន និងកំហាប់នៃផលិតផលនៃប្រតិកម្មពហុទិសដៅនៅក្នុងរាងកាយគឺតាមលក្ខខណ្ឌ ស្រមើស្រមៃ ចាប់តាំងពីការទទួលទាន និងការដកយកសារធាតុចេញមិនឈប់។ ការតភ្ជាប់ជាបន្តបន្ទាប់ជាមួយបរិស្ថានអនុញ្ញាតឱ្យយើងពិចារណាសារពាង្គកាយមានជីវិតជាប្រព័ន្ធបើកចំហ។
សម្រាប់កោសិការស់ទាំងអស់ ប្រភពថាមពលគឺព្រះអាទិត្យ។ កោសិការុក្ខជាតិចាប់យកថាមពលពីពន្លឺព្រះអាទិត្យដោយមានជំនួយពី chlorophyll ដោយប្រើវាសម្រាប់ប្រតិកម្ម assimilation ក្នុងអំឡុងពេលដំណើរការនៃរស្មីសំយោគ។ កោសិការបស់សត្វ ផ្សិត និងបាក់តេរីប្រើប្រាស់ថាមពលពន្លឺព្រះអាទិត្យដោយប្រយោល កំឡុងពេលបំបែកសារធាតុសរីរាង្គដែលសំយោគដោយរុក្ខជាតិនៅលើផែនដី។
សារធាតុចិញ្ចឹមមួយចំនួនរបស់កោសិកាត្រូវបានបំបែកកំឡុងពេលដកដង្ហើមកោសិកា ដូច្នេះការផ្គត់ផ្គង់ថាមពលចាំបាច់សម្រាប់ប្រភេទផ្សេងៗនៃសកម្មភាពកោសិកា។ ដំណើរការនេះកើតឡើងនៅក្នុង organelles ហៅថា mitochondria ។ Mitochondria មានភ្នាសពីរ៖ ខាងក្រៅបំបែកសរីរាង្គចេញពី cytoplasm និងផ្នែកខាងក្នុងបង្កើតជាផ្នត់ជាច្រើន។ ផលិតផលសំខាន់នៃការដកដង្ហើមគឺ ATP ។ វាចាកចេញពី mitochondria ហើយត្រូវបានគេប្រើជាប្រភពថាមពលសម្រាប់ប្រតិកម្មគីមីជាច្រើននៅក្នុង cytoplasm និងភ្នាសកោសិកា។ ប្រសិនបើអុកស៊ីសែនត្រូវបានទាមទារសម្រាប់ការដកដង្ហើមតាមកោសិកា នោះការដកដង្ហើមត្រូវបានគេហៅថា aerobic ប៉ុន្តែប្រសិនបើមានប្រតិកម្មកើតឡើងនៅពេលអវត្ដមាននៃអុកស៊ីហ្សែន នោះយើងនិយាយអំពីការដកដង្ហើម anaerobic ។
សម្រាប់ប្រភេទការងារណាមួយដែលបានអនុវត្តនៅក្នុងកោសិកា ថាមពលត្រូវបានប្រើប្រាស់ក្នុងទម្រង់តែមួយ និងតែមួយគត់ - ក្នុងទម្រង់ជាថាមពលពីចំណងផូស្វ័រនៃ ATP ។ ATP គឺជាសមាសធាតុចល័តយ៉ាងងាយស្រួល។ ការបង្កើត ATP កើតឡើងនៅលើភ្នាសខាងក្នុងនៃ mitochondria ។ ATP ត្រូវបានសំយោគនៅក្នុងកោសិកាទាំងអស់ក្នុងអំឡុងពេលដកដង្ហើមដោយសារតែថាមពលនៃការកត់សុីនៃកាបូអ៊ីដ្រាត ខ្លាញ់ និងសារធាតុសរីរាង្គផ្សេងទៀត។ នៅក្នុងកោសិការុក្ខជាតិពណ៌បៃតង បរិមាណសំខាន់នៃ ATP ត្រូវបានសំយោគនៅក្នុង chloroplasts ដោយសារតែថាមពលពន្លឺព្រះអាទិត្យ។ កំឡុងពេលធ្វើរស្មីសំយោគ ពួកវាផលិត ATP ច្រើនជាង mitochondria ច្រើនដង។ ATP រលាយជាមួយនឹងការដាច់នៃចំណងផូស្វ័រ - អុកស៊ីហ្សែន និងការបញ្ចេញថាមពល។ វាកើតឡើងនៅក្រោមសកម្មភាពនៃអង់ស៊ីម ATPase កំឡុងពេល hydrolysis នៃ ATP - ការបន្ថែមទឹកជាមួយនឹងការលុបបំបាត់ម៉ូលេគុលអាស៊ីតផូស្វ័រ។ ជាលទ្ធផល ATP ត្រូវបានបំប្លែងទៅជា ADP ហើយប្រសិនបើម៉ូលេគុលអាស៊ីតផូស្វ័រពីរត្រូវបានបំបែក នោះទៅជា AMP។ ប្រតិកម្មនៃការលុបបំបាត់អាស៊ីតម៉ូលេគុលក្រាមនីមួយៗត្រូវបានអមដោយការបញ្ចេញ 40 kJ ។ នេះគឺជាទិន្នផលថាមពលដ៏ធំមួយ ដែលនេះជាមូលហេតុដែលចំណងផូស្វ័រ-អុកស៊ីហ្សែននៃ ATP ត្រូវបានគេហៅថា macroergistic (ថាមពលខ្ពស់)។
ការប្រើប្រាស់ ATP ក្នុងប្រតិកម្មផ្លាស់ប្តូរផ្លាស្ទិចត្រូវបានអនុវត្តដោយការភ្ជាប់ពួកវាជាមួយ ATP hydrolysis ។ ម៉ូលេគុលនៃសារធាតុផ្សេងៗត្រូវបានចោទប្រកាន់ដោយថាមពលដោយការភ្ជាប់ក្រុមផូស្វ័រដែលបញ្ចេញក្នុងអំឡុងពេល hydrolysis ពីម៉ូលេគុល ATP ពោលគឺដោយ phosphorylation ។
ភាពប្លែកនៃដេរីវេនៃផូស្វ័រគឺថាពួកគេមិនអាចចាកចេញពីកោសិកាបានទេទោះបីជាទម្រង់ "បញ្ចេញ" របស់ពួកគេឆ្លងកាត់ដោយសេរីតាមរយៈភ្នាសក៏ដោយ។ សូមអរគុណចំពោះបញ្ហានេះ ម៉ូលេគុល phosphorylated នៅតែមាននៅក្នុងកោសិការហូតដល់ពួកវាត្រូវបានប្រើក្នុងប្រតិកម្មសមស្រប។
ដំណើរការបញ្ច្រាសនៃការបំលែង ADP ទៅជា ATP កើតឡើងដោយការបន្ថែមម៉ូលេគុលអាស៊ីតផូស្វ័រទៅ ADP បញ្ចេញទឹក និងស្រូបយកថាមពលយ៉ាងច្រើន។
ដូច្នេះ ATP គឺជាប្រភពថាមពលសកល និងផ្ទាល់សម្រាប់សកម្មភាពកោសិកា។ នេះបង្កើតមូលនិធិកោសិកាតែមួយនៃថាមពល និងធ្វើឱ្យវាអាចចែកចាយឡើងវិញ និងដឹកជញ្ជូនវាពីតំបន់មួយនៃកោសិកាទៅមួយទៀត។
ការផ្ទេរក្រុម phosphate ដើរតួនាទីយ៉ាងសំខាន់ក្នុងប្រតិកម្មគីមី ដូចជាការប្រមូលផ្តុំ macromolecules ពី monomers ។ ឧទាហរណ៍ អាស៊ីតអាមីណូអាចត្រូវបានបញ្ចូលគ្នាទៅជា peptides តែបន្ទាប់ពីត្រូវបាន phosphorylated ពីមុន។ ដំណើរការមេកានិកនៃការកន្ត្រាក់ ឬចលនា ការដឹកជញ្ជូនសារធាតុរំលាយប្រឆាំងនឹងជម្រាលកំហាប់ និងដំណើរការផ្សេងទៀតពាក់ព័ន្ធនឹងការប្រើប្រាស់ថាមពលដែលរក្សាទុកក្នុង ATP ។
ដំណើរការនៃការរំលាយអាហារថាមពលអាចត្រូវបានតំណាងដូចខាងក្រោម។ សារធាតុសរីរាង្គម៉ូលេគុលខ្ពស់នៅក្នុង cytoplasm ត្រូវបាន enzymatically ដោយ hydrolysis បំប្លែងទៅជាសារធាតុសាមញ្ញជាង ដែលវាមានៈ ប្រូតេអ៊ីន - ទៅជាអាស៊ីតអាមីណូ poly- និង disaccharides - ទៅជា monosaccharides (+ glucose) ខ្លាញ់ទៅជា glycerol និង fatty acids ។ មិនមានដំណើរការអុកស៊ីតកម្មទេថាមពលតិចតួចត្រូវបានបញ្ចេញដែលមិនត្រូវបានប្រើហើយចូលទៅក្នុងទម្រង់កម្ដៅ។ កោសិកាភាគច្រើនប្រើកាបូអ៊ីដ្រាតដំបូង។ Polysaccharides (ម្សៅនៅក្នុងរុក្ខជាតិ និង glycogen នៅក្នុងសត្វ) ត្រូវបាន hydrolyzed ទៅគ្លុយកូស។ ការកត់សុីគ្លុយកូសកើតឡើងជាបីដំណាក់កាលគឺ glycolysis, oxidative decarboxylation (វដ្ត Krebs - វដ្តនៃអាស៊ីតនៃក្រូចឆ្មា) និង phosphorylation អុកស៊ីតកម្ម (ខ្សែសង្វាក់ផ្លូវដង្ហើម) ។ Glycolysis ជាលទ្ធផលដែលម៉ូលេគុលមួយនៃជាតិស្ករត្រូវបានបំបែកជាពីរម៉ូលេគុលនៃអាស៊ីត pyruvic ជាមួយនឹងការបញ្ចេញម៉ូលេគុលពីរនៃ ATP កើតឡើងនៅក្នុង cytoplasm ។ អវត្ដមាននៃអុកស៊ីហ្សែន អាស៊ីត pyruvic ត្រូវបានបំប្លែងទៅជាអេតាណុល (ការបន្សាបជាតិអាស៊ីត) ឬអាស៊ីតឡាក់ទិក (ការដកដង្ហើម anaerobic) ។
នៅពេលដែល glycolysis កើតឡើងនៅក្នុងកោសិកាសត្វ ម៉ូលេគុលកាបូន 6 នៃគ្លុយកូសបំបែកទៅជាម៉ូលេគុលពីរនៃអាស៊ីតឡាក់ទិក។ ដំណើរការនេះមានច្រើនដំណាក់កាល។ វាត្រូវបានអនុវត្តជាបន្តបន្ទាប់ដោយ 13 អង់ស៊ីម។ ក្នុងអំឡុងពេល fermentation គ្រឿងស្រវឹង ម៉ូលេគុលពីរនៃអេតាណុល និងម៉ូលេគុលពីរនៃ CO2 ត្រូវបានបង្កើតឡើងពីម៉ូលេគុលគ្លុយកូស។
Glycolysis គឺជាដំណាក់កាលធម្មតានៃការដកដង្ហើមតាមបែប aerobic និង aerobic; ដំណើរការនៃអុកស៊ីតកម្មគ្មានអុកស៊ីហ្សែន ដែលក្នុងនោះមានតែផ្នែកមួយនៃថាមពលនៃសារធាតុរំលាយអាហារប៉ុណ្ណោះដែលត្រូវបានបញ្ចេញ និងប្រើប្រាស់ គឺចុងក្រោយសម្រាប់សារពាង្គកាយ anaerobic ។ នៅក្នុងវត្តមាននៃអុកស៊ីហ៊្សែនអាស៊ីត pyruvic ឆ្លងកាត់ចូលទៅក្នុង mitochondria ដែលជាលទ្ធផលនៃប្រតិកម្មបន្តបន្ទាប់គ្នាវាត្រូវបានកត់សុីទាំងស្រុងទៅជា H2O និង CO2 ជាមួយនឹង phosphorylation ដំណាលគ្នានៃ ADP ទៅ ATP ។ ក្នុងករណីនេះម៉ូលេគុល ATP ពីរត្រូវបានផលិតដោយ glycolysis ពីរដោយវដ្ត Krebs និង 34 ដោយខ្សែសង្វាក់ផ្លូវដង្ហើម។ ទិន្នផលសុទ្ធសម្រាប់ការកត់សុីពេញលេញនៃម៉ូលេគុលគ្លុយកូសមួយទៅ H2O និង CO2 គឺ 38 ម៉ូលេគុល។
ដូច្នេះនៅក្នុងសារពាង្គកាយ aerobic ការបំបែកចុងក្រោយនៃសារធាតុសរីរាង្គត្រូវបានអនុវត្តដោយការកត់សុីពួកវាជាមួយនឹងអុកស៊ីសែនបរិយាកាសទៅជាសារធាតុអសរីរាង្គសាមញ្ញ៖ CO2 និង H2O ។ ដំណើរការនេះកើតឡើងនៅលើ cristae នៃ mitochondria ។ ក្នុងករណីនេះចំនួនអតិបរមានៃថាមពលឥតគិតថ្លៃត្រូវបានបញ្ចេញដែលជាផ្នែកសំខាន់មួយដែលត្រូវបានបម្រុងទុកនៅក្នុងម៉ូលេគុល ATP ។ វាងាយមើលឃើញថា អុកស៊ីតកម្មតាមបែប aerobic ផ្តល់ឱ្យកោសិកានូវថាមពលដោយឥតគិតថ្លៃដល់កម្រិតដ៏អស្ចារ្យបំផុត។
ជាលទ្ធផលនៃ catabolism ម៉ូលេគុល ATP សម្បូរថាមពលប្រមូលផ្តុំនៅក្នុងកោសិកា ហើយ CO2 និងទឹកលើសត្រូវបានបញ្ចេញទៅក្នុងបរិយាកាសខាងក្រៅ។
ម៉ូលេគុលស្ករដែលមិនត្រូវការសម្រាប់ការដកដង្ហើមអាចត្រូវបានរក្សាទុកនៅក្នុងកោសិកា។ ជាតិខ្លាញ់លើសត្រូវបានបំបែកចេញ បន្ទាប់ពីនោះផលិតផលនៃការបំបែករបស់វាចូលទៅក្នុង mitochondria ជាស្រទាប់ខាងក្រោមសម្រាប់ការដកដង្ហើម ឬត្រូវបានទុកជាទុនបម្រុងនៅក្នុង cytoplasm ក្នុងទម្រង់ជាដំណក់ខ្លាញ់។ ប្រូតេអ៊ីនត្រូវបានបង្កើតឡើងពីអាស៊ីតអាមីណូចូលទៅក្នុងកោសិកា។ ការសំយោគប្រូតេអ៊ីនកើតឡើងនៅក្នុងសរីរាង្គដែលហៅថា ribosomes ។ ribosome នីមួយៗមានភាគល្អិតរងពីរ - ធំ និងតូច៖ ភាគល្អិតទាំងពីររួមមានម៉ូលេគុលប្រូតេអ៊ីន និងម៉ូលេគុល RNA ។
Ribosomes ជារឿយៗត្រូវបានភ្ជាប់ទៅប្រព័ន្ធភ្នាសពិសេសដែលមានអាងទឹក និង vesicles - ដែលគេហៅថា endoplasmic reticulum (ER); នៅក្នុងកោសិកាដែលផលិតប្រូតេអ៊ីនច្រើន កោសិកា endoplasmic ជារឿយៗត្រូវបានអភិវឌ្ឍយ៉ាងល្អ និងគ្របដណ្តប់ដោយ ribosomes ។ អង់ស៊ីមខ្លះមានប្រសិទ្ធភាពលុះត្រាតែពួកវាជាប់នឹងភ្នាស។ អង់ស៊ីមភាគច្រើនដែលពាក់ព័ន្ធនឹងការសំយោគ lipid មានទីតាំងនៅទីនេះ។ ដូច្នេះ កោសិកា endoplasmic reticulum គឺដូចជាប្រភេទនៃកោសិកាការងារ។
លើសពីនេះទៀត ER បែងចែក cytoplasm ទៅជាផ្នែកដាច់ដោយឡែក ពោលគឺវាបំបែកដំណើរការគីមីផ្សេងៗដែលកើតឡើងក្នុងពេលដំណាលគ្នានៅក្នុង cytoplasm ហើយដោយហេតុនេះកាត់បន្ថយលទ្ធភាពដែលដំណើរការទាំងនេះនឹងរំខានដល់គ្នាទៅវិញទៅមក។
ផលិតផលដែលផលិតដោយកោសិកាដែលបានផ្តល់ឱ្យជារឿយៗត្រូវបានគេប្រើនៅខាងក្រៅកោសិកា។ ក្នុងករណីបែបនេះ ប្រូតេអ៊ីនដែលត្រូវបានសំយោគនៅលើ ribosomes ឆ្លងកាត់ភ្នាសនៃ endoplasmic reticulum ហើយត្រូវបានខ្ចប់ចូលទៅក្នុងភ្នាសភ្នាសដែលបង្កើតនៅជុំវិញពួកវា ដែលបន្ទាប់មកត្រូវបានផ្តាច់ចេញពី ER ។ សរសៃទាំងនេះ មានរាងសំប៉ែត និងដាក់ជង់លើគ្នា ដូចជានំផេនខេកជង់គ្នា បង្កើតបានជារចនាសម្ព័ន្ធលក្ខណៈមួយហៅថា ស្មុគ្រស្មាញ Golgi ឬឧបករណ៍ Golgi ។ ក្នុងអំឡុងពេលស្នាក់នៅក្នុងបរិធាន Golgi ប្រូតេអ៊ីនឆ្លងកាត់ការផ្លាស់ប្តូរមួយចំនួន។ នៅពេលដែលពេលវេលាមកដល់សម្រាប់ពួកវាចេញពីកោសិកា ភ្នាសភ្នាសបានបញ្ចូលគ្នាជាមួយភ្នាសកោសិកា ហើយត្រូវបានបញ្ចេញចោល បង្ហូរមាតិការបស់ពួកគេចេញ ពោលគឺការសំងាត់កើតឡើងដោយ exocytosis ។
ឧបករណ៍ Golgi ក៏ផលិត lysosomes - ថង់ភ្នាសដែលមានអង់ស៊ីមរំលាយអាហារ។ ការស្វែងយល់ពីរបៀបដែលកោសិកាបង្កើត វេចខ្ចប់ និងនាំចេញប្រូតេអ៊ីនជាក់លាក់ និងរបៀបដែលវា "ដឹង" ប្រូតេអ៊ីនដែលវាគួររក្សាទុកសម្រាប់ខ្លួនវាគឺជាសាខាដ៏គួរឱ្យចាប់អារម្មណ៍បំផុតមួយនៃ cytology ទំនើប។
ភ្នាសនៃកោសិកាណាមួយមានចលនា និងផ្លាស់ប្តូរជានិច្ច។ ភ្នាស ER ផ្លាស់ទីយឺតៗពេញកោសិកា។ ផ្នែកនីមួយៗនៃភ្នាសទាំងនេះដាច់ដោយឡែក និងបង្កើតជា vesicles ដែលក្លាយជាផ្នែកមួយនៃឧបករណ៍ Golgi ជាបណ្ដោះអាសន្ន ហើយបន្ទាប់មកតាមរយៈដំណើរការនៃ exocytosis បញ្ចូលគ្នាជាមួយភ្នាសកោសិកា។
ក្រោយមកសម្ភារៈភ្នាសត្រូវបានត្រលប់ទៅ cytoplasm ដែលវាត្រូវបានគេប្រើម្តងទៀត។
ការផ្លាស់ប្តូរសារធាតុចូលទៅក្នុងកោសិកា ឬបញ្ចេញដោយវានៅខាងក្រៅ ក៏ដូចជាការផ្លាស់ប្តូរសញ្ញាផ្សេងៗជាមួយនឹងមីក្រូ និងម៉ាក្រូ កើតឡើងតាមរយៈភ្នាសខាងក្រៅនៃកោសិកា។ ដូចដែលត្រូវបានគេស្គាល់ ភ្នាសកោសិកាគឺជាស្រទាប់ខ្លាញ់ដែលចូលទៅក្នុងកោសិកាដែលម៉ូលេគុលប្រូតេអ៊ីនជាច្រើនត្រូវបានបង្កប់ដែលដើរតួជាអ្នកទទួលឯកទេស បណ្តាញអ៊ីយ៉ុង ឧបករណ៍ដែលដឹកជញ្ជូនយ៉ាងសកម្ម ឬដកសារធាតុគីមីផ្សេងៗ ទំនាក់ទំនងរវាងកោសិកា។ល។ ភ្នាស asymmetrically: ផ្ទៃខាងក្រៅមាន sphingomyelin និង phosphatidylcholine, ផ្ទៃខាងក្នុង - នៃ phosphatidylserine និង phosphatidylethanolamine ។ ការថែរក្សា asymmetry បែបនេះតម្រូវឱ្យមានការចំណាយថាមពល។ ដូច្នេះនៅក្នុងព្រឹត្តិការណ៍នៃការខូចខាតកោសិកា ការឆ្លងមេរោគ ឬការអត់ឃ្លានថាមពល ផ្ទៃខាងក្រៅនៃភ្នាសត្រូវបានសំបូរទៅដោយសារធាតុ phospholipids ដែលខុសពីធម្មតា ដែលវាក្លាយជាសញ្ញាសម្រាប់កោសិកា និងអង់ស៊ីមផ្សេងទៀតអំពីការខូចខាតកោសិកាជាមួយនឹងប្រតិកម្មដែលត្រូវគ្នាចំពោះបញ្ហានេះ។ តួនាទីដ៏សំខាន់បំផុតត្រូវបានលេងដោយទម្រង់រលាយនៃ phospholipase A2 ដែលបំបែកអាស៊ីត arachidonic និងបង្កើត lysoforms ពី phospholipids ដែលបានរៀបរាប់ខាងលើ។ អាស៊ីត Arachidonic គឺជាតំណភ្ជាប់កំណត់សម្រាប់ការបង្កើតអ្នកសម្រុះសម្រួលរលាកដូចជា eicosanoids និងម៉ូលេគុលការពារ - pentraxins (ប្រូតេអ៊ីន C-reactive (CRP) មុនគេនៃប្រូតេអ៊ីន amyloid) - ត្រូវបានភ្ជាប់ទៅនឹង lysoforms នៅក្នុងភ្នាសបន្ទាប់មកដោយការធ្វើឱ្យសកម្មនៃការបំពេញបន្ថែម។ ប្រព័ន្ធតាមបណ្តោយផ្លូវបុរាណ និងការបំផ្លាញកោសិកា។
រចនាសម្ព័ន្ធនៃភ្នាសជួយរក្សាលក្ខណៈនៃបរិយាកាសខាងក្នុងនៃកោសិកាភាពខុសគ្នារបស់វាពីបរិយាកាសខាងក្រៅ។ នេះត្រូវបានធានាដោយការជ្រាបចូលនៃភ្នាសកោសិកា និងអត្ថិភាពនៃយន្តការដឹកជញ្ជូនសកម្មនៅក្នុងវា។ ការរំខានរបស់ពួកគេជាលទ្ធផលនៃការខូចខាតដោយផ្ទាល់ឧទាហរណ៍ដោយ tetrodotoxin, ouabain, tetraethylammonium ឬក្នុងករណីនៃការផ្គត់ផ្គង់ថាមពលមិនគ្រប់គ្រាន់ដល់ "ស្នប់" ដែលត្រូវគ្នានាំឱ្យមានការរំខាននៃសមាសធាតុអេឡិចត្រូលីតនៃកោសិកាការផ្លាស់ប្តូរការរំលាយអាហាររបស់វាការរំខាន។ មុខងារជាក់លាក់ - ការកន្ត្រាក់ ដំណើរការនៃកម្លាំងរំញោច ជាដើម។ ការរំខាននៃបណ្តាញអ៊ីយ៉ុងកោសិកា (កាល់ស្យូម សូដ្យូម ប៉ូតាស្យូម និងក្លរួ) នៅក្នុងមនុស្សក៏អាចត្រូវបានកំណត់ដោយហ្សែនដោយការផ្លាស់ប្តូរហ្សែនដែលទទួលខុសត្រូវចំពោះរចនាសម្ព័ន្ធនៃបណ្តាញទាំងនេះ។ អ្វីដែលគេហៅថា channelopathies បណ្តាលឱ្យមានជំងឺតំណពូជនៃប្រព័ន្ធសរសៃប្រសាទសាច់ដុំនិងប្រព័ន្ធរំលាយអាហារ។ ការបញ្ចូលទឹកច្រើនពេកទៅក្នុងកោសិកាអាចនាំអោយមានការប្រេះស្រាំរបស់វា - ស៊ីតូលីស៊ីស - ដោយសារតែការជ្រាបចូលនៃភ្នាសនៅពេលដែលការបំពេញបន្ថែមត្រូវបានធ្វើឱ្យសកម្ម ឬការវាយប្រហារដោយកោសិកា lymphocytes cytotoxic និងកោសិកាឃាតករធម្មជាតិ។
ភ្នាសកោសិកាមានអ្នកទទួលជាច្រើនដែលបង្កើតឡើងនៅក្នុងវា - រចនាសម្ព័ន្ធដែលនៅពេលរួមបញ្ចូលគ្នាជាមួយម៉ូលេគុលសញ្ញាជាក់លាក់ដែលត្រូវគ្នា (ligands) បញ្ជូនសញ្ញាមួយនៅខាងក្នុងកោសិកា។ វាកើតឡើងតាមរយៈបណ្តុំបទប្បញ្ញត្តិផ្សេងៗដែលមានម៉ូលេគុលសកម្មអង់ស៊ីមដែលត្រូវបានធ្វើឱ្យសកម្មជាបន្តបន្ទាប់ ហើយចុងក្រោយរួមចំណែកដល់ការអនុវត្តកម្មវិធីកោសិកាផ្សេងៗ ដូចជាការលូតលាស់ និងការរីកសាយ ភាពខុសគ្នា ចលនា ភាពចាស់ និងការស្លាប់កោសិកា។ និយតកម្មមានច្រើនណាស់ ប៉ុន្តែចំនួនរបស់វាមិនទាន់ត្រូវបានកំណត់ពេញលេញនៅឡើយ។ ប្រព័ន្ធនៃ receptors និង cascades និយតកម្មដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងពួកវាក៏មាននៅក្នុងកោសិកាផងដែរ។ ពួកគេបង្កើតបណ្តាញបទប្បញ្ញត្តិជាក់លាក់មួយជាមួយនឹងចំណុចនៃការប្រមូលផ្តុំ ការចែកចាយ និងការជ្រើសរើសផ្លូវសញ្ញាបន្ថែមទៀត អាស្រ័យលើស្ថានភាពមុខងាររបស់កោសិកា ដំណាក់កាលនៃការអភិវឌ្ឍន៍របស់វា និងសកម្មភាពដំណាលគ្នានៃសញ្ញាពីអ្នកទទួលផ្សេងទៀត។ លទ្ធផលនៃការនេះអាចជាការរារាំង ឬការពង្រឹងសញ្ញា ដោយដឹកនាំវាតាមផ្លូវបទប្បញ្ញត្តិផ្សេងគ្នា។ ទាំងឧបករណ៍ទទួល និងផ្លូវបញ្ជូនសញ្ញាតាមរយៈបណ្តុំបទប្បញ្ញត្តិ ឧទាហរណ៍ទៅស្នូល អាចត្រូវបានរំខានជាលទ្ធផលនៃពិការភាពហ្សែនដែលកើតឡើងជាពិការភាពពីកំណើតនៅកម្រិតសារពាង្គកាយ ឬជាលទ្ធផលនៃការផ្លាស់ប្តូរ somatic នៅក្នុងកោសិកាជាក់លាក់មួយ។ ប្រភេទ។ យន្តការទាំងនេះអាចត្រូវបានបំផ្លាញដោយភ្នាក់ងារបង្ករោគ ជាតិពុល និងការផ្លាស់ប្តូរផងដែរក្នុងអំឡុងពេលដំណើរការចាស់។ ដំណាក់កាលចុងក្រោយនៃការនេះអាចជាការរំខានដល់មុខងាររបស់កោសិកា ដំណើរការនៃការរីកសាយ និងភាពខុសគ្នារបស់វា។
នៅលើផ្ទៃនៃកោសិកាក៏មានម៉ូលេគុលដែលដើរតួនាទីយ៉ាងសំខាន់ក្នុងដំណើរការនៃអន្តរកោសិកា។ ទាំងនេះអាចរាប់បញ្ចូលទាំងប្រូតេអ៊ីន adhesion កោសិកា អង់ទីករនៃភាពឆបគ្នា histocompatibility antigens ជាក់លាក់នៃជាលិកា ភាពខុសគ្នានៃ antigens ។ល។ ការផ្លាស់ប្តូរសមាសភាពនៃម៉ូលេគុលទាំងនេះបណ្តាលឱ្យមានការរំខាននៃអន្តរកម្មរវាងកោសិកា និងអាចបណ្តាលឱ្យមានការធ្វើឱ្យសកម្មនៃយន្តការសមស្របសម្រាប់ការលុបបំបាត់កោសិកាបែបនេះ ព្រោះវាបង្កឱ្យ គ្រោះថ្នាក់ជាក់លាក់ចំពោះភាពសុចរិតនៃរាងកាយ ជាអាងស្តុកទឹកនៃការឆ្លងមេរោគ ជាពិសេសមេរោគ ឬជាអ្នកផ្តើមសក្តានុពលនៃការលូតលាស់ដុំសាច់។
ការរំលោភលើការផ្គត់ផ្គង់ថាមពលនៃកោសិកា
ប្រភពនៃថាមពលនៅក្នុងកោសិកាគឺជាអាហារ បន្ទាប់ពីការបំបែកថាមពលត្រូវបានបញ្ចេញទៅជាសារធាតុចុងក្រោយ។ កន្លែងសំខាន់នៃការផលិតថាមពលគឺ mitochondria ដែលក្នុងនោះសារធាតុត្រូវបានកត់សុីដោយជំនួយពីអង់ស៊ីមនៃខ្សែសង្វាក់ផ្លូវដង្ហើម។ អុកស៊ីតកម្មគឺជាអ្នកផ្គត់ផ្គង់ថាមពលដ៏សំខាន់ព្រោះជាលទ្ធផលនៃ glycolysis ថាមពលមិនលើសពី 5% ត្រូវបានបញ្ចេញចេញពីបរិមាណដូចគ្នានៃស្រទាប់ខាងក្រោមអុកស៊ីតកម្ម (គ្លុយកូស) បើប្រៀបធៀបទៅនឹងអុកស៊ីតកម្ម។ ប្រហែល 60% នៃថាមពលដែលបានបញ្ចេញកំឡុងពេលអុកស៊ីតកម្មត្រូវបានប្រមូលផ្តុំដោយ phosphorylation អុកស៊ីតកម្មនៅក្នុងផូស្វាតដែលមានថាមពលខ្ពស់ (ATP, creatine phosphate) នៅសល់ត្រូវបានរលាយជាកំដៅ។ ក្រោយមក ផូស្វាតថាមពលខ្ពស់ត្រូវបានប្រើប្រាស់ដោយកោសិកាសម្រាប់ដំណើរការដូចជា ប្រតិបត្តិការបូម ការសំយោគ ការបែងចែក ចលនា ការសម្ងាត់។ល។មានយន្តការបីគឺការខូចខាតដែលអាចបណ្តាលឱ្យមានការរំខានដល់ការផ្គត់ផ្គង់ថាមពលរបស់កោសិកា៖ ទីមួយគឺ យន្តការនៃការសំយោគអង់ស៊ីមមេតាបូលីសថាមពល, ទីពីរគឺជាយន្តការនៃ phosphorylation អុកស៊ីតកម្ម, ទីបីគឺជាយន្តការនៃការប្រើប្រាស់ថាមពល។
ការរំខាននៃការដឹកជញ្ជូនអេឡិចត្រុងនៅក្នុងសង្វាក់ផ្លូវដង្ហើម mitochondrial ឬ uncoupling នៃអុកស៊ីតកម្ម ADP និង phosphorylation ជាមួយនឹងការបាត់បង់សក្តានុពលប្រូតុង ដែលជាកម្លាំងជំរុញសម្រាប់ការបង្កើត ATP នាំឱ្យចុះខ្សោយនៃ phosphorylation អុកស៊ីតកម្មតាមរបៀបដែលថាមពលភាគច្រើនត្រូវបានរលាយដូចជាកំដៅ និង ចំនួននៃសមាសធាតុថាមពលខ្ពស់ថយចុះ។ ការបំបែកអុកស៊ីតកម្ម និង phosphorylation ក្រោមឥទ្ធិពលនៃ adrenaline ត្រូវបានប្រើដោយកោសិកានៃសារពាង្គកាយកំដៅផ្ទះដើម្បីបង្កើនការផលិតកំដៅ ខណៈពេលដែលរក្សាសីតុណ្ហភាពរាងកាយថេរក្នុងអំឡុងពេលត្រជាក់ ឬបង្កើនវាអំឡុងពេលក្តៅខ្លួន។ ការផ្លាស់ប្តូរដ៏សំខាន់នៅក្នុងរចនាសម្ព័ន្ធ mitochondrial និងការរំលាយអាហារថាមពលត្រូវបានគេសង្កេតឃើញនៅក្នុង thyrotoxicosis ។ ការផ្លាស់ប្តូរទាំងនេះដំបូងគឺអាចត្រឡប់វិញបាន ប៉ុន្តែបន្ទាប់ពីចំណុចជាក់លាក់មួយ ពួកវាប្រែជាមិនអាចត្រឡប់វិញបាន៖ បំណែក mitochondria បែកខ្ញែក ឬហើម បាត់បង់ cristae ប្រែទៅជា vacuoles ហើយនៅទីបំផុតប្រមូលផ្តុំសារធាតុដូចជា hyaline ferritin កាល់ស្យូម lipofuscin ។ ចំពោះអ្នកជំងឺដែលមាន scurvy, mitochondria fuse ដើម្បីបង្កើត chondriospheres, ប្រហែលជាដោយសារតែការបំផ្លាញភ្នាសដោយសមាសធាតុ peroxide ។ ការខូចខាតយ៉ាងសំខាន់ចំពោះ mitochondria កើតឡើងក្រោមឥទិ្ធពលនៃវិទ្យុសកម្មអ៊ីយ៉ូដ កំឡុងពេលបំប្លែងកោសិកាធម្មតាទៅជាសាហាវ។
Mitochondria គឺជាឃ្លាំងដ៏មានឥទ្ធិពលនៃអ៊ីយ៉ុងកាល់ស្យូម ដែលកំហាប់របស់វាគឺមានលំដាប់លំដោយខ្ពស់ជាងនៅក្នុង cytoplasm ។ នៅពេលដែល mitochondria ត្រូវបានខូចខាត ជាតិកាល់ស្យូមចូលទៅក្នុង cytoplasm ដែលបណ្តាលឱ្យមានការធ្វើឱ្យសកម្មនៃប្រូតេអ៊ីនជាមួយនឹងការខូចខាតដល់រចនាសម្ព័ន្ធខាងក្នុងកោសិកា និងការរំខានដល់មុខងារនៃកោសិកាដែលត្រូវគ្នា ឧទាហរណ៍ ការចុះកិច្ចសន្យាកាល់ស្យូម ឬសូម្បីតែ "ការស្លាប់នៃជាតិកាល់ស្យូម" នៅក្នុងណឺរ៉ូន។ ជាលទ្ធផលនៃការរំខាននៃសមត្ថភាពមុខងាររបស់ mitochondria ការបង្កើតសមាសធាតុ peroxide រ៉ាឌីកាល់សេរីកើនឡើងយ៉ាងខ្លាំងដែលមានប្រតិកម្មខ្ពស់ហើយដូច្នេះបំផ្លាញសមាសធាតុសំខាន់ៗនៃកោសិកា - អាស៊ីត nucleic ប្រូតេអ៊ីននិង lipids ។ បាតុភូតនេះត្រូវបានគេសង្កេតឃើញក្នុងអំឡុងពេលដែលហៅថាភាពតានតឹងអុកស៊ីតកម្មហើយអាចមានផលវិបាកអវិជ្ជមានចំពោះអត្ថិភាពនៃកោសិកា។ ដូច្នេះការខូចខាតភ្នាសខាងក្រៅនៃ mitochondria ត្រូវបានអមដោយការបញ្ចេញចូលទៅក្នុង cytoplasm នៃសារធាតុដែលមាននៅក្នុងចន្លោះ interemembrane ជាចម្បង cytochrome C និងសារធាតុសកម្មជីវសាស្រ្តមួយចំនួនផ្សេងទៀត ដែលបង្កឱ្យមានប្រតិកម្មសង្វាក់ដែលបណ្តាលឱ្យស្លាប់កោសិកាតាមកម្មវិធី - apoptosis ។ ដោយការបំផ្លាញ DNA របស់ mitochondrial ប្រតិកម្មរ៉ាឌីកាល់សេរីបង្ខូចទ្រង់ទ្រាយព័ត៌មានហ្សែនដែលចាំបាច់សម្រាប់ការបង្កើតអង់ស៊ីមខ្សែសង្វាក់ផ្លូវដង្ហើមមួយចំនួនដែលត្រូវបានផលិតជាពិសេសនៅក្នុង mitochondria ។ នេះនាំឱ្យមានការរំខានកាន់តែច្រើននៃដំណើរការអុកស៊ីតកម្ម។ ជាទូទៅ ឧបករណ៍ហ្សែនផ្ទាល់ខ្លួនរបស់ mitochondria បើប្រៀបធៀបទៅនឹងឧបករណ៍ហ្សែននៃស្នូល គឺមិនសូវត្រូវបានការពារពីឥទ្ធិពលដែលបង្កគ្រោះថ្នាក់ដែលអាចផ្លាស់ប្តូរព័ត៌មានហ្សែនដែលបានអ៊ិនកូដនៅក្នុងវា។ ជាលទ្ធផល ភាពមិនដំណើរការនៃ mitochondria កើតឡើងពេញមួយជីវិត ឧទាហរណ៍ កំឡុងពេលដំណើរការចាស់ កំឡុងពេលផ្លាស់ប្តូរកោសិកាសាហាវ ក៏ដូចជាប្រឆាំងនឹងផ្ទៃខាងក្រោយនៃជំងឺ mitochondrial តំណពូជដែលទាក់ទងនឹងការផ្លាស់ប្តូរ DNA mitochondrial នៅក្នុងស៊ុត។ បច្ចុប្បន្ននេះការផ្លាស់ប្តូរ mitochondrial ជាង 50 ត្រូវបានពិពណ៌នាដែលបណ្តាលឱ្យមានជំងឺ degenerative តំណពូជនៃប្រព័ន្ធសរសៃប្រសាទនិងសាច់ដុំ។ ពួកវាត្រូវបានបញ្ជូនទៅកុមារទាំងស្រុងពីម្តាយ ចាប់តាំងពី mitochondria នៃមេជីវិតឈ្មោលមិនមែនជាផ្នែកនៃ zygote ហើយយោងទៅតាមសារពាង្គកាយថ្មី។
ការបំពានលើការរក្សាទុក និងការបញ្ជូនព័ត៌មានហ្សែន
ស្នូលកោសិកាផ្ទុកនូវព័ត៌មានហ្សែនភាគច្រើន ហើយដោយហេតុនេះធានានូវដំណើរការធម្មតារបស់វា។ តាមរយៈកន្សោមហ្សែនជ្រើសរើស វាសម្របសម្រួលសកម្មភាពកោសិកាកំឡុងពេលអន្តរដំណាក់កាល រក្សាទុកព័ត៌មានហ្សែន និងបង្កើតឡើងវិញ និងបញ្ជូនសម្ភារៈហ្សែនអំឡុងពេលបែងចែកកោសិកា។ ការចម្លង DNA និងការចម្លង RNA កើតឡើងនៅក្នុងស្នូល។ កត្តាបង្កជំងឺផ្សេងៗ ដូចជាកាំរស្មីអ៊ុលត្រាវីយូឡេ និងអ៊ីយ៉ូដ អុកស៊ីតកម្មរ៉ាឌីកាល់សេរី សារធាតុគីមី មេរោគអាចបំផ្លាញ DNA ។ វាត្រូវបានគេប៉ាន់ស្មានថាកោសិកានីមួយៗនៃសត្វដែលមានឈាមក្តៅត្រូវចំណាយពេល 1 ថ្ងៃ។ បាត់បង់មូលដ្ឋានច្រើនជាង 10,000 ។ នៅទីនេះយើងគួរតែបន្ថែមការបំពាននៅពេលចម្លងកំឡុងពេលបែងចែក។ ប្រសិនបើការខូចខាតទាំងនេះនៅតែបន្ត កោសិកានឹងមិនអាចរស់បានឡើយ។ ការការពារស្ថិតនៅក្នុងអត្ថិភាពនៃប្រព័ន្ធជួសជុលដ៏មានអានុភាពដូចជា អ៊ុលត្រាវីយូឡេស អង់ដូណូក្លូស ការជួសជុលការចម្លង និងប្រព័ន្ធជួសជុលឡើងវិញដែលជំនួសការខូចខាត DNA ។ ពិការភាពហ្សែននៅក្នុងប្រព័ន្ធជួសជុលបណ្តាលឱ្យមានការវិវត្តនៃជំងឺដែលបណ្តាលមកពីការបង្កើនភាពប្រែប្រួលទៅនឹងកត្តាដែលបំផ្លាញ DNA ។ នេះគឺជា xeroderma pigmentosum ក៏ដូចជារោគសញ្ញានៃភាពចាស់ដែលបង្កើនល្បឿនមួយចំនួន អមដោយទំនោរកើនឡើងក្នុងការវិវត្តទៅជាដុំសាច់សាហាវ។
ប្រព័ន្ធសម្រាប់ធ្វើនិយតកម្មដំណើរការនៃការចម្លង DNA ការចម្លងសារ RNA (mRNA) និងការបកប្រែព័ត៌មានហ្សែនពីអាស៊ីត nucleic ទៅក្នុងរចនាសម្ព័ន្ធនៃប្រូតេអ៊ីនគឺពិតជាស្មុគស្មាញ និងពហុកម្រិត។ បន្ថែមពីលើបទប្បញ្ញត្តិដែលបង្កឱ្យមានសកម្មភាពនៃកត្តាចម្លងដែលមានចំនួនសរុបជាង 3000 ដែលធ្វើឱ្យហ្សែនមួយចំនួនសកម្ម វាក៏មានប្រព័ន្ធគ្រប់គ្រងពហុកម្រិតដែលសម្របសម្រួលដោយម៉ូលេគុល RNA តូចៗ (រំខាន RNA; RNAi) ។ ហ្សែនរបស់មនុស្សដែលមានប្រហែល 3 ពាន់លាន purine និង pyrimidine bases មានត្រឹមតែ 2% នៃហ្សែនរចនាសម្ព័ន្ធដែលទទួលខុសត្រូវចំពោះការសំយោគប្រូតេអ៊ីន។ នៅសល់ផ្តល់នូវការសំយោគនៃ RNA និយតកម្មដែលក្នុងពេលដំណាលគ្នាជាមួយកត្តាចម្លងធ្វើឱ្យសកម្មឬរារាំងការងារនៃហ្សែនរចនាសម្ព័ន្ធនៅកម្រិត DNA ក្នុងក្រូម៉ូសូមឬមានឥទ្ធិពលលើដំណើរការនៃការបកប្រែ RNA (mRNA) កំឡុងពេលបង្កើតម៉ូលេគុល polypeptide នៅក្នុង cytoplasm ។ ការរំលោភលើព័ត៌មានហ្សែនអាចកើតឡើងទាំងនៅកម្រិតនៃហ្សែនរចនាសម្ព័ន្ធ និងផ្នែកនិយតកម្មនៃ DNA ជាមួយនឹងការបង្ហាញដែលត្រូវគ្នាក្នុងទម្រង់នៃជំងឺតំណពូជផ្សេងៗ។
ថ្មីៗនេះ ការយកចិត្តទុកដាក់ជាច្រើនត្រូវបានទាក់ទាញចំពោះការផ្លាស់ប្តូរនៃហ្សែនដែលកើតឡើងក្នុងអំឡុងពេលនៃការអភិវឌ្ឍន៍បុគ្គលនៃសារពាង្គកាយមួយ ហើយត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការរារាំង ឬការធ្វើឱ្យសកម្មនៃផ្នែកខ្លះនៃ DNA និងក្រូម៉ូសូមដោយសារតែ methylation, acetylation និង phosphorylation របស់វា។ ការផ្លាស់ប្តូរទាំងនេះនៅតែបន្តកើតមានក្នុងរយៈពេលយូរ ជួនកាលពេញមួយជីវិតនៃសារពាង្គកាយចាប់ពីអំប្រ៊ីយ៉ុងរហូតដល់វ័យចាស់ ហើយត្រូវបានគេហៅថាតំណពូជអេពីដេណូមិច។
ការរីកសាយនៃកោសិកាដែលមានព័ត៌មានហ្សែនប្រែប្រួលក៏ត្រូវបានរារាំងដោយប្រព័ន្ធ (កត្តា) ដែលគ្រប់គ្រងវដ្តមីតូស។ ពួកវាធ្វើអន្តរកម្មជាមួយប្រូតេអ៊ីន kinases ដែលពឹងផ្អែកលើ cyclin និងផ្នែករងកាតាលីកររបស់ពួកគេ - cyclins - ហើយរារាំងកោសិកាមិនឱ្យឆ្លងកាត់វដ្ត mitotic ពេញលេញ ដោយបញ្ឈប់ការបែងចែកនៅព្រំដែនរវាងដំណាក់កាល presynthetic និង synthetic (ប្លុក G1/S) រហូតដល់ការជួសជុល DNA ត្រូវបានបញ្ចប់។ ហើយប្រសិនបើវាមិនអាចទៅរួចនោះ ពួកគេចាប់ផ្តើមកោសិកាស្លាប់ដែលបានកំណត់កម្មវិធី។ កត្តាទាំងនេះរួមមានហ្សែន p53 ការផ្លាស់ប្តូរដែលបណ្តាលឱ្យបាត់បង់ការគ្រប់គ្រងលើការរីកសាយនៃកោសិកាផ្លាស់ប្តូរ។ វាត្រូវបានគេសង្កេតឃើញស្ទើរតែ 50% នៃជំងឺមហារីកមនុស្ស។ ចំណុចត្រួតពិនិត្យទីពីរនៃវដ្ត mitotic គឺនៅព្រំដែន G2/M ។ នៅទីនេះ ការចែកចាយត្រឹមត្រូវនៃសម្ភារៈក្រូម៉ូសូមរវាងកោសិកាកូនស្រីនៅក្នុង mitosis ឬ meiosis ត្រូវបានគ្រប់គ្រងដោយប្រើសំណុំនៃយន្តការដែលគ្រប់គ្រងកោសិកា spindle កណ្តាល និង centromeres (kinetochores) ។ ភាពគ្មានប្រសិទ្ធភាពនៃយន្តការទាំងនេះនាំឱ្យមានការរំខានដល់ការចែកចាយក្រូម៉ូសូម ឬផ្នែករបស់ពួកគេ ដែលត្រូវបានបង្ហាញដោយអវត្តមាននៃក្រូម៉ូសូមណាមួយនៅក្នុងកោសិកាកូនស្រី (aneuploidy) វត្តមាននៃក្រូម៉ូសូមបន្ថែម (polyploidy) ការបំបែកនៃ ផ្នែកនៃក្រូម៉ូសូម (ការលុប) និងការផ្ទេររបស់វាទៅក្រូម៉ូសូមមួយផ្សេងទៀត (ការផ្ទេរទីតាំង) ។ ដំណើរការបែបនេះត្រូវបានគេសង្កេតឃើញជាញឹកញាប់ក្នុងអំឡុងពេលការរីកសាយនៃកោសិកាដែលខូច និងបំប្លែងយ៉ាងសាហាវ។ ប្រសិនបើរឿងនេះកើតឡើងក្នុងអំឡុងពេល meiosis ជាមួយកោសិកាមេរោគ វានាំទៅដល់ការស្លាប់របស់ទារកនៅដំណាក់កាលដំបូងនៃការអភិវឌ្ឍន៍អំប្រ៊ីយ៉ុង ឬដល់កំណើតនៃសារពាង្គកាយដែលមានជំងឺក្រូម៉ូសូម។
ការរីកសាយកោសិកាដែលមិនអាចគ្រប់គ្រងបានក្នុងអំឡុងពេលលូតលាស់នៃដុំសាច់កើតឡើងជាលទ្ធផលនៃការផ្លាស់ប្តូរហ្សែនដែលគ្រប់គ្រងការរីកសាយកោសិកា ហើយត្រូវបានគេហៅថា oncogenes ។ ក្នុងចំណោមសារធាតុ oncogenes ដែលគេស្គាល់ច្រើនជាង 70 ដែលភាគច្រើនជាធាតុផ្សំនៃបទបញ្ជាការលូតលាស់កោសិកា ខ្លះត្រូវបានតំណាងដោយកត្តាចម្លងដែលគ្រប់គ្រងសកម្មភាពហ្សែន ក៏ដូចជាកត្តាដែលរារាំងការបែងចែក និងការលូតលាស់កោសិកា។ កត្តាមួយទៀតដែលកំណត់ការពង្រីកលើសលប់ (ការរីករាលដាល) នៃកោសិកាដែលរីកធំធាត់គឺការបង្រួមចុងនៃក្រូម៉ូសូម - telomeres ដែលមិនអាចចម្លងបានពេញលេញជាលទ្ធផលនៃអន្តរកម្មស្តេរ៉ូអ៊ីតសុទ្ធសាធ ដូច្នេះបន្ទាប់ពីការបែងចែកកោសិកានីមួយៗ telomeres ត្រូវបានខ្លីដោយ ផ្នែកជាក់លាក់នៃមូលដ្ឋាន។ ដូច្នេះ ការរីកសាយកោសិកានៃសារពាង្គកាយពេញវ័យបន្ទាប់ពីការបែងចែកមួយចំនួន (ជាធម្មតាពី 20 ទៅ 100 អាស្រ័យលើប្រភេទនៃសារពាង្គកាយ និងអាយុរបស់វា) អស់ប្រវែង telomere ហើយការចម្លងក្រូម៉ូសូមបន្ថែមទៀតឈប់។ បាតុភូតនេះមិនកើតឡើងនៅក្នុងកោសិកាមេជីវិតឈ្មោល កោសិកា enterocytes និងអំប្រ៊ីយ៉ុងនោះទេ ដោយសារតែវត្តមានរបស់អង់ស៊ីម telomerase ដែលស្ដារប្រវែង telomere ឡើងវិញ បន្ទាប់ពីការបែងចែកនីមួយៗ។ នៅក្នុងកោសិកាភាគច្រើននៃសារពាង្គកាយមនុស្សពេញវ័យ telomerase ត្រូវបានរារាំង ប៉ុន្តែជាអកុសលវាត្រូវបានធ្វើឱ្យសកម្មនៅក្នុងកោសិកាដុំសាច់។
ការតភ្ជាប់រវាងស្នូលនិង cytoplasm និងការដឹកជញ្ជូនសារធាតុក្នុងទិសដៅទាំងពីរត្រូវបានអនុវត្តតាមរយៈរន្ធញើសនៅក្នុងភ្នាសនុយក្លេអ៊ែរដោយមានការចូលរួមពីប្រព័ន្ធដឹកជញ្ជូនពិសេសដែលប្រើប្រាស់ថាមពល។ នៅក្នុងវិធីនេះ ថាមពល និងសារធាតុប្លាស្ទិក ម៉ូលេគុលសញ្ញា (កត្តាចម្លង) ត្រូវបានបញ្ជូនទៅស្នូល។ លំហូរបញ្ច្រាសចូលទៅក្នុងម៉ូលេគុល cytoplasm នៃ mRNA និងផ្ទេរ RNA (tRNA) ribosomes ចាំបាច់សម្រាប់ការសំយោគប្រូតេអ៊ីននៅក្នុងកោសិកា។ ផ្លូវដូចគ្នានៃការដឹកជញ្ជូនសារធាតុក៏មាននៅក្នុងមេរោគផងដែរ ជាពិសេសដូចជាមេរោគអេដស៍។ ពួកវាផ្ទេរសារធាតុហ្សែនរបស់ពួកគេទៅក្នុងស្នូលនៃកោសិកាមេ ជាមួយនឹងការបញ្ចូលបន្ថែមរបស់វាទៅក្នុងហ្សែនមេ និងការផ្ទេរ RNA មេរោគដែលបានបង្កើតថ្មីទៅក្នុង cytoplasm សម្រាប់ការសំយោគប្រូតេអ៊ីននៃភាគល្អិតមេរោគថ្មី។
ការរំលោភលើដំណើរការសំយោគ
ដំណើរការសំយោគប្រូតេអ៊ីនកើតឡើងនៅក្នុងអាងនៃ reticulum endoplasmic ដែលមានទំនាក់ទំនងយ៉ាងជិតស្និទ្ធជាមួយនឹងរន្ធញើសនៅក្នុងភ្នាសនុយក្លេអ៊ែរ ដែលតាមរយៈនោះ ribosomes, tRNA និង mRNA ចូលទៅក្នុង reticulum endoplasmic ។ នៅទីនេះការសំយោគនៃខ្សែសង្វាក់ polypeptide ត្រូវបានអនុវត្តដែលជាបន្តបន្ទាប់ទទួលបានទម្រង់ចុងក្រោយរបស់ពួកគេនៅក្នុង reticulum endoplasmic agranular និង lamellar complex (Golgi complex) ដែលពួកគេឆ្លងកាត់ការកែប្រែក្រោយការបកប្រែ និងផ្សំជាមួយកាបូអ៊ីដ្រាត និងម៉ូលេគុល lipid ។ ម៉ូលេគុលប្រូតេអ៊ីនដែលទើបបង្កើតថ្មីមិនស្ថិតនៅទីតាំងនៃការសំយោគទេ ប៉ុន្តែតាមរយៈដំណើរការគ្រប់គ្រងដ៏ស្មុគស្មាញហៅថា ប្រូតេអ៊ីនkinesisត្រូវបានផ្ទេរយ៉ាងសកម្មទៅផ្នែកដាច់ស្រយាលនៃក្រឡា ដែលពួកវានឹងអនុវត្តមុខងារដែលគេចង់បាន។ ក្នុងករណីនេះ ជំហានដ៏សំខាន់មួយគឺការរៀបចំរចនាសម្ព័ន្ធនៃម៉ូលេគុលដែលបានផ្ទេរទៅក្នុងការកំណត់រចនាសម្ព័ន្ធលំហដែលសមស្រប ដែលមានសមត្ថភាពអាចបំពេញមុខងាររបស់វាបាន។ រចនាសម្ព័ន្ធនេះកើតឡើងដោយជំនួយពីអង់ស៊ីមពិសេស ឬនៅលើម៉ាទ្រីសនៃម៉ូលេគុលប្រូតេអ៊ីនឯកទេស - chaperones ដែលជួយម៉ូលេគុលប្រូតេអ៊ីនដែលទើបបង្កើត ឬផ្លាស់ប្តូរដោយសារឥទ្ធិពលខាងក្រៅ ដើម្បីទទួលបានរចនាសម្ព័ន្ធបីវិមាត្រត្រឹមត្រូវ។ នៅក្នុងព្រឹត្តិការណ៍នៃផលប៉ះពាល់អវិជ្ជមានលើកោសិកានៅពេលដែលមានលទ្ធភាពនៃការរំខាននៃរចនាសម្ព័ន្ធនៃម៉ូលេគុលប្រូតេអ៊ីន (ឧទាហរណ៍ជាមួយនឹងការកើនឡើងនៃសីតុណ្ហភាពរាងកាយដំណើរការឆ្លងការស្រវឹង) ការប្រមូលផ្តុំនៃ chaperones នៅក្នុងកោសិកាកើនឡើង។ យ៉ាងខ្លាំង។ ដូច្នេះម៉ូលេគុលបែបនេះត្រូវបានគេហៅផងដែរ។ ប្រូតេអ៊ីនស្ត្រេស, ឬ ប្រូតេអ៊ីនឆក់កំដៅ. ការបំពានលើរចនាសម្ព័ន្ធនៃម៉ូលេគុលប្រូតេអ៊ីននាំទៅរកការបង្កើតនូវបណ្តុំអសកម្មគីមី ដែលត្រូវបានដាក់ក្នុងកោសិកា ឬនៅខាងក្រៅវាអំឡុងពេល amyloidosis ជំងឺ Alzheimer's ។ ករណី ប្រសិនបើរចនាសម្ព័ន្ធបឋមកើតឡើងមិនត្រឹមត្រូវ ម៉ូលេគុលជាបន្តបន្ទាប់ទាំងអស់ក៏នឹងខូចដែរ។ ស្ថានភាពនេះកើតឡើងនៅក្នុងអ្វីដែលគេហៅថាជំងឺ prion (scrapie ក្នុងចៀម, គោឆ្កួត, kuru, ជំងឺ Creutzfeldt-Jakob ចំពោះមនុស្ស) នៅពេលដែលពិការភាពនៃប្រូតេអ៊ីនភ្នាសមួយនៃកោសិកាសរសៃប្រសាទបណ្តាលឱ្យមានការប្រមូលផ្តុំជាបន្តបន្ទាប់នៃម៉ាស់អសកម្មនៅក្នុងកោសិកា។ និងការរំខានដល់មុខងារសំខាន់ៗរបស់វា។
ការរំខាននៃដំណើរការសំយោគនៅក្នុងកោសិកាអាចកើតឡើងនៅដំណាក់កាលផ្សេងៗរបស់វា៖ ការចម្លង RNA នៅក្នុងស្នូល ការបកប្រែនៃសារធាតុ polypeptides នៅក្នុង ribosomes ការកែប្រែក្រោយការបកប្រែ ការ hypermethylation និង glycosylation នៃម៉ូលេគុលបន៍ត្នោតខ្ចី ការដឹកជញ្ជូន និងការចែកចាយប្រូតេអ៊ីននៅក្នុងកោសិកា និងការយកចេញរបស់ពួកគេ។ ទៅខាងក្រៅ។ ក្នុងករណីនេះ គេអាចសង្កេតឃើញការកើនឡើង ឬថយចុះនៃចំនួន ribosomes ការបំបែកនៃ polyribosomes ការពង្រីកនៃ cisterns នៃ granular endoplasmic reticulum ការបាត់បង់ ribosomes និងការបង្កើត vesicles និង vacuoles ។ ដូច្នេះនៅពេលដែលពុលដោយ grebe ស្លេក អង់ស៊ីម RNA polymerase ត្រូវបានខូចខាត ដែលរំខានដល់ការចម្លង។ ជាតិពុលរោគខាន់ស្លាក់ ដោយការធ្វើឱ្យកត្តាពន្លូតសកម្ម រំខានដល់ដំណើរការបកប្រែ បណ្តាលឱ្យខូចខាតដល់សាច់ដុំបេះដូង។ មូលហេតុនៃការរំខាននៃការសំយោគនៃម៉ូលេគុលប្រូតេអ៊ីនជាក់លាក់មួយចំនួនអាចជាភ្នាក់ងារបង្ករោគ។ ឧទាហរណ៍វីរុស Herpes រារាំងការសំយោគនិងការបញ្ចេញម៉ូលេគុល MHC antigen ដែលអនុញ្ញាតឱ្យពួកគេជៀសវាងការគ្រប់គ្រងភាពស៊ាំដោយផ្នែក bacilli - ការសំយោគនៃអ្នកសម្របសម្រួលនៃការរលាកស្រួចស្រាវ។ រូបរាងនៃប្រូតេអ៊ីនមិនធម្មតាអាចបញ្ឈប់ការបំបែកបន្ថែមទៀតរបស់វាហើយនាំទៅដល់ការប្រមូលផ្តុំនៃសារធាតុ inert ឬសូម្បីតែសារធាតុពុល។ នេះអាចត្រូវបានសម្របសម្រួលដោយការរំខាននៃដំណើរការពុករលួយក្នុងកម្រិតជាក់លាក់មួយ។
ការរំខាននៃដំណើរការរលួយ
ក្នុងពេលដំណាលគ្នាជាមួយនឹងការសំយោគប្រូតេអ៊ីននៅក្នុងកោសិកាការបំបែករបស់វាកើតឡើងជាបន្តបន្ទាប់។ នៅក្រោមលក្ខខណ្ឌធម្មតា នេះមានសារៈសំខាន់ជាបទប្បញ្ញត្តិ និងទម្រង់ជាឧទាហរណ៍ ក្នុងអំឡុងពេលនៃការធ្វើឱ្យសកម្មនៃទម្រង់អសកម្មនៃអង់ស៊ីម អ័រម៉ូនប្រូតេអ៊ីន និងប្រូតេអ៊ីននៃវដ្ត mitotic ។ ការលូតលាស់ និងការអភិវឌ្ឍន៍កោសិកាធម្មតាទាមទារឱ្យមានតុល្យភាពដែលគ្រប់គ្រងយ៉ាងល្អរវាងការសំយោគ និងការរិចរិលនៃប្រូតេអ៊ីន និងសរីរាង្គ។ ទោះបីជាយ៉ាងណាក៏ដោយ នៅក្នុងដំណើរការនៃការសំយោគប្រូតេអ៊ីន ដោយសារតែមានកំហុសក្នុងប្រតិបត្តិការឧបករណ៍សំយោគ រចនាសម្ព័ន្ធខុសប្រក្រតីនៃម៉ូលេគុលប្រូតេអ៊ីន និងការខូចខាតរបស់វាដោយភ្នាក់ងារគីមី និងបាក់តេរី ម៉ូលេគុលខូចមួយចំនួនធំត្រូវបានបង្កើតឡើងឥតឈប់ឈរ។ យោងតាមការប៉ាន់ប្រមាណមួយចំនួនចំណែករបស់ពួកគេគឺប្រហែលមួយភាគបីនៃប្រូតេអ៊ីនសំយោគទាំងអស់។
កោសិកាថនិកសត្វមានមេមួយចំនួន វិធីបំផ្លាញប្រូតេអ៊ីន៖តាមរយៈ lysosomal proteases (pentide hydrolases), ប្រូតេអ៊ីនដែលពឹងផ្អែកលើកាល់ស្យូម (endopeptidases) និងប្រព័ន្ធ proteasome ។ លើសពីនេះទៀតក៏មានប្រូតេអ៊ីនពិសេសផងដែរដូចជា caspases ។ សរីរាង្គសំខាន់ដែលការរិចរិលនៃសារធាតុកើតឡើងនៅក្នុងកោសិកា eukaryotic គឺ lysosome ដែលមានអង់ស៊ីម hydrolytic ជាច្រើន។ ដោយសារតែដំណើរការនៃ endocytosis និងប្រភេទផ្សេងៗនៃ autophagy នៅក្នុង lysosomes និង phagolysosomes ទាំងម៉ូលេគុលប្រូតេអ៊ីនដែលខូច និងសរីរាង្គទាំងមូលត្រូវបានបំផ្លាញ: mitochondria ដែលខូចខាត ផ្នែកនៃភ្នាសប្លាស្មា ប្រូតេអ៊ីន extracellular មួយចំនួន និងមាតិកានៃ secretory granules ។
យន្តការសំខាន់មួយសម្រាប់ការរិចរិលប្រូតេអ៊ីនគឺ proteasome ដែលជារចនាសម្ព័ន្ធប្រូតេអ៊ីន multicatalytic នៃរចនាសម្ព័ន្ធស្មុគស្មាញដែលបានធ្វើមូលដ្ឋានីយកម្មនៅក្នុង cytosol, nucleus, endoplasmic reticulum និងនៅលើភ្នាសកោសិកា។ ប្រព័ន្ធអង់ស៊ីមនេះទទួលខុសត្រូវក្នុងការបំបែកប្រូតេអ៊ីនដែលខូចក៏ដូចជាប្រូតេអ៊ីនដែលមានសុខភាពល្អដែលត្រូវតែយកចេញសម្រាប់ដំណើរការធម្មតានៃកោសិកា។ ក្នុងករណីនេះ ប្រូតេអ៊ីនដែលត្រូវបំផ្លាញត្រូវបានផ្សំជាបឋមជាមួយនឹងសារធាតុ polypeptide ជាក់លាក់មួយ ubiquitin ។ ទោះជាយ៉ាងណាក៏ដោយ ប្រូតេអ៊ីនដែលមិនមាននៅគ្រប់ទីកន្លែងក៏អាចបំផ្លាញដោយផ្នែកនៅក្នុង proteasomes ផងដែរ។ ការបំបែកម៉ូលេគុលប្រូតេអ៊ីននៅក្នុង proteasomes ទៅជា polypeptides ខ្លីៗ (ដំណើរការ) ជាមួយនឹងការបង្ហាញជាបន្តបន្ទាប់របស់ពួកគេជាមួយនឹងប្រភេទ I MHC ម៉ូលេគុល គឺជាតំណភ្ជាប់ដ៏សំខាន់ក្នុងការគ្រប់គ្រងភាពស៊ាំនៃ antigenic homeostasis នៅក្នុងខ្លួន។ នៅពេលដែលមុខងារ proteasome ត្រូវបានចុះខ្សោយ ប្រូតេអ៊ីនដែលខូច និងមិនចាំបាច់កកកុញ ដែលអមនឹងភាពចាស់នៃកោសិកា។ ការរំលោភលើការរិចរិលនៃប្រូតេអ៊ីនដែលពឹងផ្អែកលើ cyclin នាំឱ្យមានការរំខាននៃការបែងចែកកោសិកាការរិចរិលនៃប្រូតេអ៊ីន secretory - ដល់ការវិវត្តនៃ cystofibrosis ។ ផ្ទុយទៅវិញ ការកើនឡើងនៃមុខងារ proteasome អមជាមួយនឹងការថយចុះនៃរាងកាយ (អេដស៍ មហារីក)។
ជាមួយនឹងជំងឺដែលបានកំណត់ហ្សែននៃការរិចរិលប្រូតេអ៊ីន សារពាង្គកាយមិនអាចឋិតឋេរបាន ហើយស្លាប់នៅដំណាក់កាលដំបូងនៃការបង្កើតអំប្រ៊ីយ៉ុង។ ប្រសិនបើការបំបែកខ្លាញ់ ឬកាបូអ៊ីដ្រាតត្រូវបានរំខាននោះ ជំងឺផ្ទុក (thesaurismosis) កើតឡើង។ ក្នុងករណីនេះបរិមាណច្រើនហួសប្រមាណនៃសារធាតុឬផលិតផលនៃការបំបែកមិនពេញលេញរបស់ពួកគេ - lipid, polysaccharides - កកកុញនៅខាងក្នុងកោសិកាដែលធ្វើឱ្យខូចមុខងាររបស់កោសិកាយ៉ាងខ្លាំង។ ភាគច្រើនវាត្រូវបានគេសង្កេតឃើញនៅក្នុងកោសិកា epithelial ថ្លើម (hepatocytes), ណឺរ៉ូន, fibroblasts និង macrophagocytes ។
ជំងឺដែលទទួលបាននៃដំណើរការនៃការបំបែកសារធាតុអាចកើតឡើងជាលទ្ធផលនៃដំណើរការរោគសាស្ត្រ (ឧទាហរណ៍ប្រូតេអ៊ីនខ្លាញ់កាបូអ៊ីដ្រាតនិងការថយចុះសារធាតុពណ៌) ហើយត្រូវបានអមដោយការបង្កើតសារធាតុមិនធម្មតា។ ការរំខាននៅក្នុងប្រព័ន្ធ proteolysis lysosomal នាំឱ្យមានការថយចុះនៃការសម្របខ្លួនក្នុងអំឡុងពេលតមអាហារឬភាពតានតឹងកើនឡើងនិងការកើតឡើងនៃមុខងារ endocrine មួយចំនួន - ការថយចុះកម្រិតនៃអាំងស៊ុយលីន thyroglobulin cytokines និងអ្នកទទួលរបស់ពួកគេ។ ការថយចុះនៃប្រូតេអ៊ីនធ្វើឱ្យថយចុះអត្រានៃការព្យាបាលមុខរបួស បណ្តាលឱ្យមានការវិវត្តនៃជំងឺក្រិនសរសៃឈាម និងប៉ះពាល់ដល់ការឆ្លើយតបនៃប្រព័ន្ធភាពស៊ាំ។ ជាមួយនឹង hypoxia ការផ្លាស់ប្តូរ pH intracellular ការរងរបួសវិទ្យុសកម្មត្រូវបានកំណត់ដោយការកើនឡើង peroxidation នៃ lipids ភ្នាសក៏ដូចជានៅក្រោមឥទ្ធិពលនៃសារធាតុ lysosomotropic - endotoxins បាក់តេរីការរំលាយអាហារនៃផ្សិតពុល (sporofusarin) គ្រីស្តាល់អុកស៊ីដស៊ីលីកុន - ស្ថេរភាពនៃភ្នាស lysosome ។ ការផ្លាស់ប្តូរ អង់ស៊ីម lysosomal សកម្មត្រូវបានបញ្ចេញទៅក្នុង cytoplasm ដែលបណ្តាលឱ្យមានការបំផ្លាញរចនាសម្ព័ន្ធកោសិកា និងការស្លាប់របស់វា។
ជំពូកទី 1
មូលដ្ឋានគ្រឹះនៃសរីរវិទ្យាកោសិកា
I. Dudel
ភ្នាសប្លាស្មា . កោសិកាសត្វត្រូវបានចងដោយភ្នាសប្លាស្មា (រូបភាព 1.1) ។ យើងនឹងរស់នៅលើរចនាសម្ព័ន្ធរបស់វា ដែលស្រដៀងនឹងរចនាសម្ព័ន្ធនៃភ្នាសខាងក្នុងជាច្រើន ដោយលម្អិតបន្តិចទៀត។ ម៉ាទ្រីសសំខាន់នៃភ្នាសមាន ជាតិខ្លាញ់,ជាចម្បង phosphatidylcholine ។ lipid ទាំងនេះមានក្រុមក្បាល hydrophilic ដែលខ្សែសង្វាក់អ៊ីដ្រូកាបូនអ៊ីដ្រូកាបូនវែងត្រូវបានភ្ជាប់។ នៅក្នុងទឹក សារធាតុ lipids បែបនេះកើតឡើងដោយឯកឯងបង្កើតជាខ្សែភាពយន្ត bilayer ក្រាស់ 4-5 nm ដែលក្នុងនោះក្រុម hydrophilic ប្រឈមមុខនឹងមជ្ឈដ្ឋាន aqueous ហើយខ្សែសង្វាក់ hydrophobic hydrocarbon ត្រូវបានរៀបចំជាពីរជួរ បង្កើតជាដំណាក់កាល lipid anhydrous ។ ភ្នាសកោសិកាគឺជាស្រទាប់ខ្លាញ់នៃប្រភេទនេះ និងមានផ្ទុក glycolipids កូលេស្តេរ៉ុល និង phospholipids (រូបភាព 1.2) ។ ផ្នែក hydrophilic នៃ glycolipids ត្រូវបានបង្កើតឡើងដោយ oligosaccharides ។ Glycolipids តែងតែស្ថិតនៅលើផ្ទៃខាងក្រៅនៃភ្នាសប្លាស្មា ដោយផ្នែក oligosaccharide នៃម៉ូលេគុលតម្រង់ទិសដូចជាសក់ដែលជ្រលក់ក្នុងបរិស្ថាន។ ខ្ចាត់ខ្ចាយក្នុងចំណោម phospholipids ក្នុងបរិមាណស្ទើរតែស្មើគ្នា ម៉ូលេគុលកូលេស្តេរ៉ុលធ្វើឱ្យភ្នាសមានស្ថេរភាព។ ការចែកចាយ lipid ផ្សេងៗនៅក្នុងស្រទាប់ខាងក្នុង និងខាងក្រៅនៃភ្នាសគឺមិនដូចគ្នាទេ ហើយសូម្បីតែនៅក្នុងស្រទាប់មួយក៏មានតំបន់ដែលប្រភេទ lipid មួយចំនួនត្រូវបានប្រមូលផ្តុំ។ ការចែកចាយមិនស្មើគ្នានេះ។
អង្ករ។ ១.១. គំនូសតាងនៃក្រឡាដែលបង្ហាញពីសរីរាង្គសំខាន់បំផុត
ប្រហែលជាមានមុខងារមួយចំនួន ដែលមិនទាន់ច្បាស់នៅឡើយ។
ធាតុមុខងារសំខាន់ៗដែលបង្កប់ក្នុងម៉ាទ្រីស lipid inert នៃភ្នាសគឺ កំប្រុក(រូបភាព 1.2) ។ ប្រូតេអ៊ីនតាមទម្ងន់មានពី 25 ទៅ 75% នៅក្នុងភ្នាសផ្សេងៗ ប៉ុន្តែដោយសារម៉ូលេគុលប្រូតេអ៊ីនមានទំហំធំជាងម៉ូលេគុល lipid 50% ដោយទម្ងន់គឺស្មើនឹងសមាមាត្រនៃម៉ូលេគុលប្រូតេអ៊ីន 1 ទៅ 50 ម៉ូលេគុល lipid ។ ប្រូតេអ៊ីនខ្លះជ្រាបចូលទៅក្នុងភ្នាសពីខាងក្រៅទៅខាងក្នុងរបស់វា ខណៈដែលខ្លះទៀតត្រូវបានជួសជុលក្នុងស្រទាប់តែមួយ។ ម៉ូលេគុលប្រូតេអ៊ីនជាធម្មតាត្រូវបានតម្រង់ទិស ដូច្នេះក្រុម hydrophobic របស់ពួកគេត្រូវបានបង្កប់នៅក្នុងភ្នាស lipid ហើយក្រុមប៉ូល hydrophilic នៅលើផ្ទៃនៃភ្នាសត្រូវបានបង្កប់ក្នុងដំណាក់កាល aqueous ។ ប្រូតេអ៊ីនជាច្រើននៅលើផ្ទៃខាងក្រៅនៃភ្នាសគឺ glycoproteins; ក្រុម hydrophilic saccharide របស់ពួកគេប្រឈមមុខនឹងបរិយាកាសក្រៅកោសិកា។
ប្រព័ន្ធ Membrane នៃសរីរាង្គខាងក្នុង .
ប្រហែលពាក់កណ្តាលនៃបរិមាណកោសិកាត្រូវបានកាន់កាប់ដោយ organelles ដាច់ដោយឡែកពី cytosol ដោយភ្នាស។ ផ្ទៃសរុបនៃភ្នាសនៃសរីរាង្គខាងក្នុងគឺយ៉ាងហោចណាស់ 10 ដងធំជាងផ្ទៃនៃភ្នាសប្លាស្មា។ ប្រព័ន្ធភ្នាសដែលប្រើយ៉ាងទូលំទូលាយបំផុតគឺ reticulum endoplasmic,តំណាងឱ្យបណ្តាញ

អង្ករ។ ១.២.ការបង្ហាញគ្រោងការណ៍នៃភ្នាសប្លាស្មា។ ប្រូតេអ៊ីនត្រូវបានបង្កប់នៅក្នុងស្រទាប់ phospholipid bilayer ដោយពួកវាខ្លះជ្រាបចូលទៅក្នុង bilayer ខណៈពេលដែលផ្សេងទៀតត្រូវបានបោះយុថ្កាទៅស្រទាប់ខាងក្រៅឬខាងក្នុងប៉ុណ្ណោះ។
បំពង់ដែលមានរាងមូលខ្ពស់ ឬរចនាសម្ព័ន្ធពន្លូតដូចថង់; តំបន់ធំនៃ reticulum endoplasmic ត្រូវបាន studd ជាមួយ ribosomes; reticulum បែបនេះត្រូវបានគេហៅថា granular ឬ rough (រូបភាព 1.1) ។ ឧបករណ៍ Golgiក៏មានភ្នាសដែលចងភ្ជាប់ lamellae ដែល vesicles ឬ vesicles បំបែកចេញ (រូបភាព 1.1) ។ Lysosomes និង peroxisomes- ទាំងនេះគឺជា vesicles ឯកទេសតូច។ នៅក្នុងសរីរាង្គផ្សេងៗទាំងនេះ ភ្នាស និងចន្លោះដែលវាហ៊ុមព័ទ្ធមានសំណុំអង់ស៊ីមជាក់លាក់។ នៅខាងក្នុងសរីរាង្គផលិតផលមេតាបូលីសពិសេសកកកុញ ប្រើដើម្បីបំពេញមុខងារផ្សេងៗនៃសរីរាង្គ។
ស្នូលនិង មីតូខនឌ្រីខុសគ្នាត្រង់ថាសរីរាង្គនីមួយៗទាំងនេះត្រូវបានហ៊ុំព័ទ្ធដោយភ្នាសពីរ។ ស្នូលគឺទទួលខុសត្រូវចំពោះការគ្រប់គ្រង kinetic នៃការរំលាយអាហារ; ភ្នាសខាងក្នុងបត់នៃ mitochondria គឺជាកន្លែងនៃការរំលាយអាហារអុកស៊ីតកម្ម; នៅទីនេះ ដោយសារតែការកត់សុីនៃសារធាតុ pyruvate ឬអាស៊ីតខ្លាញ់ សមាសធាតុថាមពលខ្ពស់ adenosine triphosphate (ATP ឬ ATP) ត្រូវបានសំយោគ។
Cytoskeleton . cytoplasm ជុំវិញសរីរាង្គមិនអាចនៅក្នុងវិធីណាមួយត្រូវបានចាត់ទុកថាជាអាម៉ូញាក់; វាត្រូវបានជ្រាបចូលដោយបណ្តាញ cytoskeletal ។ cytoskeleton រួមមាន microtubules, actin filaments និង intermediate filaments (រូបភាព 1.1) ។ មីក្រូបំពង់មានអង្កត់ផ្ចិតខាងក្រៅប្រហែល 25 nm; ពួកវាត្រូវបានបង្កើតឡើងដូចជាវត្ថុធាតុ polymer ធម្មតាដែលជាលទ្ធផលនៃការប្រមូលផ្តុំម៉ូលេគុលប្រូតេអ៊ីន tubulin ។ សរសៃ Actin -សរសៃ contractile ដែលមានទីតាំងនៅក្នុងស្រទាប់ជិតភ្នាស និងទូទាំងកោសិកា ភាគច្រើនចូលរួមនៅក្នុងដំណើរការដែលទាក់ទងនឹងចលនា។ filaments កម្រិតមធ្យមមានប្លុកនៃសមាសធាតុគីមីផ្សេងគ្នានៅក្នុងប្រភេទផ្សេងគ្នានៃកោសិកា; ពួកវាបង្កើតជាភាពខុសគ្នានៃទំនាក់ទំនងរវាងធាតុ cytoskeletal ពីរផ្សេងទៀតដែលបានរៀបរាប់ខាងលើ។ សរីរាង្គ និងភ្នាសប្លាស្មាក៏ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹង cytoskeleton ដែលមិនត្រឹមតែរក្សារូបរាងរបស់កោសិកា និងទីតាំងនៃសរីរាង្គនៅក្នុងវាប៉ុណ្ណោះទេ ប៉ុន្តែថែមទាំងកំណត់ការផ្លាស់ប្តូររូបរាងរបស់កោសិកា និងការចល័តរបស់វាផងដែរ។
ស៊ីតូសូល។ . ប្រហែលពាក់កណ្តាលនៃបរិមាណកោសិកាត្រូវបានកាន់កាប់ដោយ cytosol ។ ដោយសារវាមានប្រូតេអ៊ីនប្រហែល 20% ដោយទម្ងន់ វាគឺជាជែលច្រើនជាងដំណោះស្រាយទឹកមួយ។ ម៉ូលេគុលតូចៗ រួមទាំងសរីរាង្គ និងអសរីរាង្គ អ៊ីយ៉ុងរំលាយនៅក្នុងដំណាក់កាល aqueous ។ ការផ្លាស់ប្តូរអ៊ីយ៉ុងកើតឡើងរវាងកោសិកា និងបរិស្ថាន (ចន្លោះក្រៅកោសិកា); ដំណើរការមេតាបូលីសទាំងនេះនឹងត្រូវបានពិភាក្សានៅក្នុងផ្នែកបន្ទាប់។ ការផ្តោតអារម្មណ៍នៃអ៊ីយ៉ុងនៅក្នុងលំហ extracellular ត្រូវបានរក្សាជាមួយនឹងភាពត្រឹមត្រូវសន្ធឹកសន្ធាប់នៅកម្រិតថេរមួយ; កំហាប់ intracellular នៃអ៊ីយ៉ុងនីមួយៗក៏មានកម្រិតជាក់លាក់មួយ ខុសពីខាងក្រៅកោសិកា (តារាង 1.1)។ សារធាតុ cation ច្រើនក្រៃលែងនៅក្នុងបរិយាកាសក្រៅកោសិកាគឺណា+ នៅក្នុងកោសិកាកំហាប់របស់វាគឺទាបជាង 10 ដង។ ផ្ទុយទៅវិញ កំហាប់ K + គឺខ្ពស់បំផុតនៅខាងក្នុងកោសិកា នៅខាងក្រៅកោសិកា វាទាបជាងច្រើនជាងលំដាប់នៃរ៉ិចទ័រ។ ជម្រាលដ៏អស្ចារ្យបំផុតរវាងកំហាប់ extracellular និង intracellular មានសម្រាប់ Ca 2+ កំហាប់នៃអ៊ីយ៉ុងសេរីនៅខាងក្នុងកោសិកាគឺទាបជាង 10,000 ដងជាងនៅខាងក្រៅវា។ មិនមែនអ៊ីយ៉ុងទាំងអស់ត្រូវបានរំលាយនៅក្នុង cytosol ទេ; ឧទាហរណ៍ ក្នុងករណី Ca 2+ អ៊ីយ៉ុងចងមានច្រើនជាង អ៊ីយ៉ុងឥតគិតថ្លៃ។ ភាគច្រើននៃប្រូតេអ៊ីន cytosolic គឺជាអង់ស៊ីមដោយមានការចូលរួមដែលដំណើរការជាច្រើននៃការរំលាយអាហារកម្រិតមធ្យមត្រូវបានអនុវត្ត: glycolysis និង gluconeogenesis ការសំយោគឬការបំផ្លាញអាស៊ីតអាមីណូ ការសំយោគប្រូតេអ៊ីននៅលើ ribosomes (រូបភាព 1.1) ។ cytosol ក៏មានដំណក់ទឹក lipid និង glycogen granules ដែលបម្រើជាទុនបំរុងនៃម៉ូលេគុលសំខាន់ៗ។
តារាង 1.1 ។កំហាប់អ៊ីយ៉ុងខាងក្នុង និងក្រៅកោសិកានៅក្នុងកោសិកាសាច់ដុំនៃសត្វដែលមានកម្តៅ។ ក – - អ៊ីយ៉ុងកោសិកាទម្ងន់ម៉ូលេគុលខ្ពស់
|
ការផ្តោតអារម្មណ៍ក្នុងកោសិកា |
ការផ្តោតអារម្មណ៍ក្រៅកោសិកា |
||
|
ណា+ |
12 មីល្លីលីត្រ / លីត្រ |
ណា+ |
145 មីល្លីលីត្រ / លីត្រ |
|
១៥៥ មីល្លីលីត្រ / លីត្រ |
K+ |
៤ មីល្លីលីត្រ/លីត្រ |
|
|
Ca 2+ |
10-7 10-8 mmol / l |
Ca 2+ |
2 mmol / លីត្រ |
|
គ l - |
៤ មីល្លីលីត្រ/លីត្រ |
C1 – |
120 មីល្លីលីត្រ / លីត្រ |
|
HCO 3 – |
៨ មីល្លីលីត្រ / លីត្រ |
HCO 3 – |
២៧ មីល្លីលីត្រ / លីត្រ |
|
ក – |
១៥៥ មីល្លីលីត្រ / លីត្រ |
ផ្សេងៗ cations |
5 មីល្លីលីត្រ / លីត្រ |
|
សក្តានុពលសម្រាក - 90 mV |
|||
១.២. ការផ្លាស់ប្តូរសារធាតុរវាងកោសិកា និងបរិស្ថាន
យើងបានពិពណ៌នាយ៉ាងខ្លីអំពីរចនាសម្ព័ន្ធនៃកោសិកា ដូច្នេះយើងអាចប្រើការពិពណ៌នានេះដើម្បីពិនិត្យមើលមូលដ្ឋាននៃសរីរវិទ្យាកោសិកា។ គ្មានករណីណាមួយអាចចាត់ទុកកោសិកាថាជាធាតុឋិតិវន្តបានឡើយ ព្រោះមានការផ្លាស់ប្តូរសារធាតុជាប្រចាំរវាងបន្ទប់ក្នុងកោសិកាផ្សេងៗ ព្រមទាំងរវាងផ្នែកនិងបរិស្ថាន។ រចនាសម្ព័ន្ធកោសិកាស្ថិតនៅក្នុងលំនឹងថាមវន្ត ហើយអន្តរកម្មនៃកោសិកាគ្នាទៅវិញទៅមក និងជាមួយបរិយាកាសខាងក្រៅគឺជាលក្ខខណ្ឌចាំបាច់សម្រាប់រក្សាជីវិតរបស់សារពាង្គកាយដែលមានមុខងារ។ នៅក្នុងជំពូកនេះ យើងនឹងពិនិត្យមើលយន្តការជាមូលដ្ឋាននៃការផ្លាស់ប្តូរបែបនេះ។ នៅក្នុងជំពូកបន្តបន្ទាប់ យន្តការទាំងនេះនឹងត្រូវបានចាត់ទុកថាត្រូវបានអនុវត្តចំពោះកោសិកាប្រសាទ និងមុខងាររបស់វា។
ទោះយ៉ាងណាក៏ដោយ យន្តការដូចគ្នានេះ បញ្ជាក់ពីដំណើរការនៃសរីរាង្គផ្សេងទៀតទាំងអស់។
ការសាយភាយ។ដំណើរការសាមញ្ញបំផុតនៃការផ្លាស់ទីវត្ថុគឺការសាយភាយ។ នៅក្នុងដំណោះស្រាយ (ឬឧស្ម័ន) អាតូម និងម៉ូលេគុលផ្លាស់ទីដោយសេរី ហើយភាពខុសគ្នានៃការប្រមូលផ្តុំមានតុល្យភាពដោយសារតែការសាយភាយ។ ចូរយើងពិចារណាបរិមាណពីរដែលពោរពេញទៅដោយរាវ ឬឧស្ម័ន (រូបភាព 1.3) ដែលសារធាតុមានកំហាប់ c 1 និង c 2 និងបំបែកដោយស្រទាប់ដែលមានផ្ទៃ A និងកម្រាស់ឃ. លំហូរនៃរូបធាតុ m ក្នុងអំឡុងពេល t បានពិពណ៌នា ច្បាប់ដំបូងនៃការសាយភាយរបស់ Fick៖
dm/ dt= ឃក/ ឃ ( គ 1 -C 2) =ឃក/ ឃឃ គ(1)
ដែល D គឺជាមេគុណនៃការសាយភាយ ថេរសម្រាប់សារធាតុដែលបានផ្តល់ឱ្យ សារធាតុរំលាយ និងសីតុណ្ហភាព។ នៅក្នុងទម្រង់ទូទៅបន្ថែមទៀតសម្រាប់ភាពខុសគ្នានៃការប្រមូលផ្តុំ dc នៅចម្ងាយ dx
dm/dt= –D A dc/dx, (2)
លំហូរតាមរយៈផ្នែក A គឺសមាមាត្រទៅនឹងជម្រាលនៃការប្រមូលផ្តុំ dc/dx . សញ្ញាដកលេចឡើងក្នុងសមីការ ដោយសារតែការផ្លាស់ប្តូរការផ្តោតអារម្មណ៍ក្នុងទិសដៅ x គឺអវិជ្ជមាន។
ការសាយភាយ គឺជាដំណើរការដ៏សំខាន់បំផុត ដោយសារតែម៉ូលេគុលភាគច្រើននៅក្នុងដំណោះស្រាយ aqueous ផ្លាស់ទីក្នុងចម្ងាយខ្លី។ នេះក៏អនុវត្តចំពោះចលនារបស់ពួកគេនៅក្នុងកោសិកាផងដែរ ដែលការសាយភាយមិនត្រូវបានរារាំងដោយភ្នាស។ សារធាតុជាច្រើនអាចសាយភាយដោយសេរីនៅទូទាំងភ្នាសខ្លាញ់ ជាពិសេសទឹក និងឧស្ម័នដែលរលាយដូចជា O 2 និង CO 2 ។ រលាយជាតិខ្លាញ់

អង្ករ។ ១.៣.គ្រោងការណ៍នៃការសាយភាយបរិមាណ។ ចន្លោះទាំងពីរត្រូវបានបំបែកដោយស្រទាប់ក្រាស់ឃនិងតំបន់ ក.គ - កំហាប់ខ្ពស់នៃភាគល្អិតនៅផ្នែកខាងឆ្វេងនៃបរិមាណ C: - កំហាប់ភាគល្អិតទាបនៅខាងស្តាំ ផ្នែក, ផ្ទៃពណ៌ផ្កាឈូក- ជម្រាលនៃការផ្តោតអារម្មណ៍នៅក្នុងស្រទាប់សាយភាយ។ លំហូរនៃការសាយភាយ dm/dt - សូមមើល សមីការ (1)
សារធាតុក៏សាយភាយបានយ៉ាងល្អតាមរយៈភ្នាស; នេះក៏អនុវត្តចំពោះម៉ូលេគុលប៉ូលដែលមានទំហំតូចល្មម ដូចជាអេតាណុល និងអ៊ុយ ខណៈដែលជាតិស្ករឆ្លងកាត់ស្រទាប់ខ្លាញ់ដោយពិបាក។ ក្នុងពេលជាមួយគ្នានេះ ស្រទាប់ lipid គឺមិនអាចជ្រាបចូលបានចំពោះម៉ូលេគុលដែលមានបន្ទុក រួមទាំងសូម្បីតែអ៊ីយ៉ុងអសរីរាង្គ។ សម្រាប់មិនមែនអេឡិចត្រូលីត សមីការនៃការសាយភាយ (1) ជាធម្មតាត្រូវបានបំប្លែងដោយការរួមបញ្ចូលលក្ខណៈនៃភ្នាស និងសារធាតុដែលសាយភាយទៅជាមួយ។ ប៉ារ៉ាម៉ែត្រ permeability (P):
dm/dt=P កឃ គ.(3)
នៅក្នុងរូបភព។ 1.4 ត្រូវបានប្រៀបធៀប permeability (P) នៃភ្នាស lipid សម្រាប់ម៉ូលេគុលផ្សេងៗ។
ការសាយភាយតាមរយៈរន្ធញើសភ្នាស . ភ្នាសប្លាស្មា (និងភ្នាសកោសិកាផ្សេងទៀត) គឺអាចជ្រាបចូលបានមិនត្រឹមតែសារធាតុដែលសាយភាយតាមស្រទាប់ខ្លាញ់ប៉ុណ្ណោះទេ ថែមទាំងអ៊ីយ៉ុង ជាតិស្ករ អាស៊ីតអាមីណូ និងនុយក្លេអូទីតជាច្រើនផងដែរ។ សារធាតុទាំងនេះឆ្លងកាត់ភ្នាសតាមរយៈរន្ធញើសដែលបានបង្កើតឡើង ដឹកជញ្ជូនប្រូតេអ៊ីន,ជ្រមុជនៅក្នុងភ្នាស។ នៅខាងក្នុងប្រូតេអ៊ីនបែបនេះមានឆានែលដែលពោរពេញទៅដោយទឹកដែលមានអង្កត់ផ្ចិតតិចជាង 1 nm ដែលតាមរយៈនោះម៉ូលេគុលតូចៗអាចសាយភាយ។ ពួកវាផ្លាស់ទីតាមជម្រាលនៃការផ្តោតអារម្មណ៍ ហើយប្រសិនបើពួកគេផ្ទុកបន្ទុក នោះចលនារបស់ពួកគេតាមរយៈបណ្តាញក៏ត្រូវបានគ្រប់គ្រងដោយសក្តានុពលភ្នាសផងដែរ។ ឆានែល Membrane គឺជ្រើសរើសតិចតួច

អង្ករ។ ១.៤.ភាពជ្រាបចូលនៃស្រទាប់ lipid សិប្បនិម្មិតទៅនឹងសារធាតុផ្សេងៗ
ទាក់ទងទៅនឹងប្រភេទនៃម៉ូលេគុលដែលអាចឆ្លងកាត់ពួកវាបាន ឧទាហរណ៍ ប៉ូតាស្យូម សូដ្យូម និងកាល់ស្យូម ឆានែលដែលនីមួយៗមិនអាចជ្រាបចូលបានស្ទើរតែគ្រប់អ៊ីយ៉ុង លើកលែងតែជាក់លាក់មួយ។ បែប ការជ្រើសរើសត្រូវបានកំណត់ដោយបន្ទុកឬរចនាសម្ព័ន្ធនៃកន្លែងចងនៅក្នុងជញ្ជាំងឆានែលដែលជួយសម្រួលដល់ការដឹកជញ្ជូនម៉ូលេគុលជាក់លាក់មួយនិងការពារការជ្រៀតចូលនៃសារធាតុផ្សេងទៀតតាមរយៈឆានែល (រូបភាពទី 2) ។ 1.5, ក) .
នៅពីក្រោយអាកប្បកិរិយា ឆានែលអ៊ីយ៉ុងភ្នាសងាយស្រួលក្នុងការសង្កេតដោយសារតែចរន្តដែលកើតចេញពីចលនារបស់អ៊ីយ៉ុងអាចត្រូវបានវាស់សូម្បីតែសម្រាប់ឆានែលតែមួយក៏ដោយ។ វាត្រូវបានបង្ហាញថាឆានែលដោយឯកឯងនិងជាមួយនឹងប្រេកង់ខ្ពស់ផ្លាស់ប្តូរស្ថានភាពរបស់ពួកគេពីបើកចំហទៅបិទ។ ឆានែលប៉ូតាស្យូមត្រូវបានកំណត់លក្ខណៈដោយជីពចរបច្ចុប្បន្នជាមួយនឹងទំហំប្រហែល 2 pA (2 10 –12 A) និងរយៈពេលជាច្រើនមិល្លីវិនាទី (មើលរូបភាព 2.12 ទំព័រ 37) [3] ។ ក្នុងអំឡុងពេលនេះ អ៊ីយ៉ុងរាប់ម៉ឺន ឆ្លងកាត់វា។ ការផ្លាស់ប្តូរនៃប្រូតេអ៊ីនពីការអនុលោមមួយទៅការអនុលោមតាមមួយផ្សេងទៀតត្រូវបានសិក្សាដោយប្រើកាំរស្មី X, Mössbauer spectroscopy និងអនុភាពម៉ាញេទិកនុយក្លេអ៊ែរ (NMR) ។ ដូច្នេះ ប្រូតេអ៊ីនមានថាមពលខ្លាំង រចនាសម្ព័ន្ធចល័ត ហើយឆានែលដែលឆ្លងកាត់ប្រូតេអ៊ីនមិនគ្រាន់តែជាបំពង់រឹង និងពោរពេញដោយទឹកទេ (រូបភាព 1.5, A) ប៉ុន្តែជា labyrinth នៃក្រុមម៉ូលេគុលដែលផ្លាស់ទីយ៉ាងឆាប់រហ័ស និងបន្ទុក។ លក្ខណៈថាមវន្តនៃឆានែលនេះត្រូវបានឆ្លុះបញ្ចាំងនៅក្នុង ទម្រង់ថាមពលនៃឆានែល,បង្ហាញក្នុងរូប។ 1.5, B. នៅទីនេះ អ័ក្ស abscissa តំណាងឱ្យប្រវែងនៃឆានែលពីដំណោះស្រាយខាងក្រៅជាមួយនឹងកំហាប់អ៊ីយ៉ុង C 0 និងសក្តានុពល 0 ទៅកាន់ដំណោះស្រាយខាងក្នុងជាមួយកំហាប់ C 1 និងសក្តានុពល E. អ័ក្សតម្រៀប

អង្ករ។ ១.៥.A. គ្រោងការណ៍នៃប្រូតេអ៊ីនដែលបង្កើតជាឆានែលប៉ូតាស្យូមដែលបង្កប់នៅក្នុងស្រទាប់ខ្លាញ់នៃភ្នាសប្លាស្មា។ ការចោទប្រកាន់អវិជ្ជមានចំនួនបួនត្រូវបានជួសជុលនៅលើ "ជញ្ជាំង" នៃឆានែល។ ខ. ទម្រង់ថាមពលតាមគ្រោងការណ៍នៃឆានែលដែលបង្ហាញក្នុងរូប។ A. អ័ក្សតម្រៀបបង្ហាញពីតម្លៃនៃថាមពល kinetic ដែលត្រូវការសម្រាប់ការឆ្លងកាត់ឆានែល; តាមបណ្តោយអ័ក្ស abscissa - ចម្ងាយរវាងផ្ទៃខាងក្នុងនិងខាងក្រៅនៃភ្នាស។ មីនីម៉ាថាមពលត្រូវគ្នាទៅនឹងកន្លែងចងនៃអ៊ីយ៉ុងដែលមានបន្ទុកវិជ្ជមានជាមួយនឹងបន្ទុកអវិជ្ជមានថេរនៅក្នុងជញ្ជាំងឆានែល។ ថាមពលអតិបរមាត្រូវគ្នាទៅនឹងឧបសគ្គនៃការសាយភាយនៅក្នុងឆានែល។ ការអនុលោមតាមប្រូតេអ៊ីនឆានែលត្រូវបានសន្មត់ថាលំយោលដោយឯកឯង; ជម្រើសទម្រង់ថាមពលត្រូវបានបង្ហាញជាមួយនឹងបន្ទាត់រឹង និងបន្ទាត់ដាច់; លំយោលទាំងនេះជួយសម្រួលយ៉ាងខ្លាំងដល់ការភ្ជាប់អ៊ីយ៉ុងនៅពេលយកឈ្នះឧបសគ្គថាមពល (កែប្រែ)
កម្រិតថាមពលអ៊ីយ៉ុងនៅកន្លែងភ្ជាប់ឆានែលត្រូវបានបង្ហាញ; កំពូលនៅក្នុងក្រាហ្វតំណាងឱ្យរបាំងភាពជ្រាបចូលដែលថាមពលអ៊ីយ៉ុងត្រូវតែលើសពីដើម្បីជ្រាបចូលទៅក្នុងឆានែល ហើយ "ជ្រលក់" នៅក្នុងក្រាហ្វតំណាងឱ្យស្ថានភាពដែលមានស្ថេរភាព (ការចង) ។ ទោះបីជាមានឧបសគ្គនៃកំពូលថាមពលក៏ដោយ អ៊ីយ៉ុងអាចជ្រាបចូលទៅក្នុងបណ្តាញ ប្រសិនបើទម្រង់ថាមពលផ្លាស់ប្តូរដោយឯកឯង។ ដូច្នេះ អ៊ីយ៉ុងអាចរកឃើញខ្លួនវា "នៅម្ខាងទៀត" នៃកំពូលថាមពល ហើយអាចបន្តផ្លាស់ទីទៅក្នុងកោសិកា។ អាស្រ័យលើបន្ទុក ទំហំ និងកម្រិតនៃជាតិទឹកនៃអ៊ីយ៉ុង និងសមត្ថភាពរបស់វាក្នុងការភ្ជាប់ទៅនឹងរចនាសម្ព័ន្ធជញ្ជាំងឆានែល ទម្រង់ថាមពលនៃឆានែលប្រែប្រួលសម្រាប់អ៊ីយ៉ុងផ្សេងៗគ្នា ដែលអាចពន្យល់ពីជម្រើសនៃប្រភេទឆានែលនីមួយៗ។
ការសាយភាយលំនឹងនៃអ៊ីយ៉ុង . ការសាយភាយនៃអ៊ីយ៉ុងផ្សេងៗតាមរយៈបណ្តាញភ្នាសគួរតែនាំទៅដល់ការលុបបំបាត់ភាពខុសគ្នានៃការប្រមូលផ្តុំរវាងបរិយាកាសខាងក្រៅ និងខាងក្នុង។ ដូចដែលអ្នកអាចឃើញពីតារាង។ 1.1 ភាពខុសគ្នាបែបនេះនៅតែមាន ដូច្នេះត្រូវតែមានខ្លះ លំនឹងរវាងការសាយភាយ និងដំណើរការដឹកជញ្ជូនផ្សេងទៀតឆ្លងកាត់ភ្នាស។ ផ្នែកពីរបន្ទាប់និយាយអំពីវិធីដែលលំនឹងនេះត្រូវបានសម្រេច។ ក្នុងករណីអ៊ីយ៉ុង លំនឹងសាយភាយត្រូវបានប៉ះពាល់ដោយបន្ទុករបស់វា។ ការសាយភាយនៃម៉ូលេគុលដែលមិនបានបញ្ចូលត្រូវបានធានាដោយភាពខុសគ្នានៃការប្រមូលផ្តុំឌីស៊ី ហើយនៅពេលដែលការប្រមូលផ្តុំត្រូវបានស្មើគ្នា ការដឹកជញ្ជូនខ្លួនឯងឈប់។ ភាគល្អិតដែលត្រូវបានចោទប្រកាន់ត្រូវបានប៉ះពាល់បន្ថែមដោយវាលអគ្គិសនី។ ឧទាហរណ៍ នៅពេលដែលអ៊ីយ៉ុងប៉ូតាស្យូមទុកកោសិកាមួយតាមជម្រាលនៃការប្រមូលផ្តុំរបស់វា វាផ្ទុកបន្ទុកវិជ្ជមានតែមួយ។ ដូច្នេះ បរិយាកាសខាងក្នុងកោសិកាកាន់តែមានបន្ទុកអវិជ្ជមាន ដែលបណ្តាលឱ្យមានភាពខុសគ្នាដ៏មានសក្តានុពលនៅទូទាំងភ្នាស។ បន្ទុកអវិជ្ជមានក្នុងកោសិការារាំងអ៊ីយ៉ុងប៉ូតាស្យូមថ្មីពីការចាកចេញពីកោសិកា ហើយអ៊ីយ៉ុងទាំងនោះដែលចេញពីកោសិកានឹងបង្កើនបន្ទុកបន្ថែមទៀតនៅលើភ្នាស។ លំហូរនៃអ៊ីយ៉ុងប៉ូតាស្យូមឈប់នៅពេលដែលសកម្មភាពនៃវាលអគ្គីសនីទូទាត់សងសម្រាប់សម្ពាធនៃការសាយភាយដោយសារតែភាពខុសគ្នានៃការប្រមូលផ្តុំ។ អ៊ីយ៉ុងបន្តឆ្លងកាត់ភ្នាសប៉ុន្តែក្នុងបរិមាណស្មើគ្នាក្នុងទិសដៅទាំងពីរ។ ដូច្នេះសម្រាប់ភាពខុសគ្នាដែលបានផ្តល់ឱ្យនៅក្នុងការប្រមូលផ្តុំអ៊ីយ៉ុងនៅលើភ្នាសមាន សក្តានុពលលំនឹងអ៊ីអ៊ីយ៉ុង ដែលលំហូរនៃអ៊ីយ៉ុងតាមរយៈភ្នាសឈប់។ សក្តានុពលលំនឹងអាចត្រូវបានកំណត់យ៉ាងងាយស្រួលដោយប្រើ សមីការ Nernst៖
អ៊ីអ៊ីយ៉ុង= រធ/ zច* lnគចេញ/ C ក្នុង(4)
ដែលជាកន្លែងដែល R - ថេរឧស្ម័ន, T - សីតុណ្ហភាពដាច់ខាត, z - វ៉ាល់អ៊ីយ៉ុង (អវិជ្ជមានសម្រាប់អ៊ីយ៉ុង)គចេញ - កំហាប់អ៊ីយ៉ុងក្រៅកោសិកា C ក្នុង - ការប្រមូលផ្តុំអ៊ីយ៉ុងខាងក្នុង;ច លេខហ្វារ៉ាដេយ។ ប្រសិនបើយើងជំនួសថេរទៅក្នុងសមីការ នោះនៅសីតុណ្ហភាពរាងកាយ (T = 310 K) សក្ដានុពលលំនឹងសម្រាប់អ៊ីយ៉ុងប៉ូតាស្យូមអ៊ី។ K គឺស្មើនឹង៖
ឯក= –61 មខ/(5)
ប្រសិនបើ [K + ចេញ]/[ K + ចូល ] = 39 ដូចខាងក្រោមពីតារាង។ 1.1 បន្ទាប់មក
ឯក = –61 m B log 39= –97 mV ។
ជាការពិត វាត្រូវបានគេរកឃើញថាកោសិកាទាំងអស់មាន សក្តានុពលភ្នាស;នៅក្នុងកោសិកាសាច់ដុំថនិកសត្វកម្រិតរបស់វាគឺប្រហែល -90 mV ។ អាស្រ័យលើលក្ខខណ្ឌ និងការប្រមូលផ្តុំអ៊ីយ៉ុងដែលទាក់ទង កោសិកាអាចមានសក្តានុពលភ្នាសចាប់ពី -40 ដល់ -120 mV ។ សម្រាប់ក្រឡាក្នុងឧទាហរណ៍ខាងលើ (តារាង 1.1) សក្តានុពលសម្រាក,ស្មើនឹងប្រហែល -90 mV បង្ហាញថាលំហូរនៃអ៊ីយ៉ុងប៉ូតាស្យូមតាមរយៈបណ្តាញភ្នាសគឺប្រហែលនៅក្នុងលំនឹង។ នេះមិនមែនជារឿងគួរឱ្យភ្ញាក់ផ្អើលនោះទេព្រោះនៅក្នុងភ្នាសដែលសម្រាកស្ថានភាពបើកចំហនៃបណ្តាញប៉ូតាស្យូមគឺទំនងបំផុត i.e. ភ្នាសគឺអាចជ្រាបចូលបានច្រើនបំផុតទៅនឹងអ៊ីយ៉ុងប៉ូតាស្យូម។ ទោះជាយ៉ាងណាក៏ដោយសក្តានុពលនៃភ្នាសត្រូវបានកំណត់ដោយលំហូរនៃអ៊ីយ៉ុងផ្សេងទៀត។
ភាពងាយស្រួលដែលភាគល្អិតដែលមិនបានបញ្ចូលអាចសាយភាយតាមភ្នាសត្រូវបានគណនាក្នុងសមីការ (3)។ ភាពជ្រាបចូលនៃភាគល្អិតដែលមានបន្ទុក ត្រូវបានពិពណ៌នាដោយសមីការស្មុគស្មាញបន្តិច៖
ទំ= ម រធ/ dF(6)
កន្លែងណា ម- ការចល័តអ៊ីយ៉ុងនៅក្នុងភ្នាស;ឃ - កម្រាស់ភ្នាស a R, T និង F - ស្គាល់ថេរនៃទែរម៉ូឌីណាមិក។ តម្លៃ permeability សម្រាប់អ៊ីយ៉ុងផ្សេងៗដែលបានកំណត់តាមវិធីនេះអាចត្រូវបានប្រើដើម្បីគណនាសក្តានុពលភ្នាសអេម នៅពេលដែលប៉ូតាស្យូម សូដ្យូម និងក្លរីន អ៊ីយ៉ុងឆ្លងកាត់ភ្នាសក្នុងពេលដំណាលគ្នា (ជាមួយ permeability P K, P Na និង P Cl រៀងៗខ្លួន)។ វាត្រូវបានសន្មត់ថាសក្តានុពលធ្លាក់ចុះស្មើភាពគ្នានៅក្នុងភ្នាសដូច្នេះថាកម្លាំងវាលគឺថេរ។ ក្នុងករណីនេះវាត្រូវបានអនុវត្ត សមីការ Goldman ឬសមីការវាលថេរ :
អ៊ីm= R T/F * ln(P K + P Na + P Cl )/ (P K + P Na + P Cl )(7)
សម្រាប់ភ្នាសកោសិកាភាគច្រើន Pខេ ប្រហែល 30 ដងខ្ពស់ជាង Pណា (សូមមើលផ្នែក 1.3 ផងដែរ)។ តម្លៃដែលទាក់ទង P Cl ប្រែប្រួលយ៉ាងខ្លាំង; សម្រាប់ភ្នាសជាច្រើន។ P Cl តូចបើធៀបនឹង Rខេ ទោះយ៉ាងណាក៏ដោយសម្រាប់អ្នកដទៃ (ឧទាហរណ៍នៅក្នុងសាច់ដុំគ្រោងឆ្អឹង) P Cl ខ្ពស់ជាង Pខេ
ការដឹកជញ្ជូនសកម្ម បូមសូដ្យូម . ផ្នែកមុនពិពណ៌នាអំពីការសាយភាយអកម្មនៃអ៊ីយ៉ុង និងសក្តានុពលនៃភ្នាសលទ្ធផលនៅឯកំហាប់អ៊ីយ៉ុងខាងក្នុង និងក្រៅកោសិកាដែលបានផ្តល់ឱ្យ។ ទោះជាយ៉ាងណាក៏ដោយ ជាលទ្ធផលនៃដំណើរការនេះ ការប្រមូលផ្តុំអ៊ីយ៉ុងនៅខាងក្នុងកោសិកាមិនមានស្ថេរភាពដោយស្វ័យប្រវត្តិទេ ចាប់តាំងពីភ្នាស
សក្ដានុពលគឺមានលក្ខណៈអេឡិចត្រុងច្រើនជាងបន្តិចអ៊ីខេ និងច្រើនបើប្រៀបធៀបទៅនឹង Eណា (ប្រហែល +60 mV) ។ ដោយសារតែការសាយភាយ ការប្រមូលផ្តុំអ៊ីយ៉ុងក្នុងកោសិកា យ៉ាងហោចណាស់ប៉ូតាស្យូម និងសូដ្យូម ត្រូវតែស្មើគ្នាជាមួយនឹងកោសិកាក្រៅ។ ស្ថេរភាពនៃជម្រាលអ៊ីយ៉ុងត្រូវបានសម្រេចតាមរយៈការដឹកជញ្ជូនសកម្ម៖ ប្រូតេអ៊ីនភ្នាសដឹកជញ្ជូនអ៊ីយ៉ុងឆ្លងកាត់ភ្នាសប្រឆាំងនឹងជម្រាលអគ្គិសនី និង/ឬកំហាប់ ការប្រើប្រាស់ថាមពលមេតាប៉ូលីសសម្រាប់ការនេះ។ ដំណើរការដ៏សំខាន់បំផុតនៃការដឹកជញ្ជូនសកម្មគឺការងារណា/K - ស្នប់ដែលមាននៅក្នុងកោសិកាស្ទើរតែទាំងអស់;
ស្នប់បូមអ៊ីយ៉ុងសូដ្យូមចេញពីកោសិកា ក្នុងពេលដំណាលគ្នាបូមអ៊ីយ៉ុងប៉ូតាស្យូមទៅក្នុងកោសិកា។ នេះធានានូវកំហាប់អ៊ីយ៉ុងសូដ្យូមក្នុងកោសិកាទាប និងកំហាប់ខ្ពស់នៃអ៊ីយ៉ុងប៉ូតាស្យូម (តារាង 1.1)។ ជម្រាលកំហាប់នៃអ៊ីយ៉ុងសូដ្យូមនៅលើភ្នាសមានមុខងារជាក់លាក់ទាក់ទងនឹងការបញ្ជូនព័ត៌មានក្នុងទម្រង់ជាកម្លាំងជំរុញអគ្គិសនី (សូមមើលផ្នែកទី 2.2) ក៏ដូចជាការថែរក្សាយន្តការដឹកជញ្ជូនសកម្មផ្សេងទៀត និងបទប្បញ្ញត្តិនៃបរិមាណកោសិកា (សូមមើលខាងក្រោម)។ ដូច្នេះវាមិនគួរឱ្យភ្ញាក់ផ្អើលទេដែលថាមពលច្រើនជាង 1/3 នៃថាមពលប្រើប្រាស់ដោយកោសិកាមួយត្រូវបានចំណាយលើស្នប់ Na/K ហើយនៅក្នុងកោសិកាសកម្មបំផុតមួយចំនួនរហូតដល់ 70% នៃថាមពលត្រូវបានចំណាយលើប្រតិបត្តិការរបស់វា។
ប្រូតេអ៊ីនដឹកជញ្ជូន Na/K គឺជា ATPase ។ នៅលើផ្ទៃខាងក្នុងនៃភ្នាសវាបំបែក ATP ទៅជា ADP និង phosphate (រូបភាព 1.6) ។ ថាមពលនៃម៉ូលេគុល ATP មួយត្រូវបានប្រើដើម្បីដឹកជញ្ជូនអ៊ីយ៉ុងសូដ្យូមបីចេញពីកោសិកា ហើយក្នុងពេលដំណាលគ្នា អ៊ីយ៉ុងប៉ូតាស្យូមពីរចូលទៅក្នុងកោសិកា ពោលគឺសរុបមក បន្ទុកវិជ្ជមានមួយត្រូវបានយកចេញពីកោសិកាក្នុងវដ្តមួយ។ ដូច្នេះម៉ាស៊ីនបូម Na/K គឺ អេឡិចត្រុង(បង្កើតចរន្តអគ្គិសនីឆ្លងកាត់ភ្នាស) ដែលនាំទៅរកការកើនឡើងនៃ electronegativity នៃសក្តានុពលភ្នាសប្រហែល 10 mV ។ ប្រូតេអ៊ីនដឹកជញ្ជូនអនុវត្តប្រតិបត្តិការនេះក្នុងល្បឿនលឿន: ពី 150 ទៅ 600 អ៊ីយ៉ុងសូដ្យូមក្នុងមួយវិនាទី។ លំដាប់អាស៊ីតអាមីណូនៃប្រូតេអ៊ីនដឹកជញ្ជូនត្រូវបានគេស្គាល់ ប៉ុន្តែយន្តការនៃការដឹកជញ្ជូនមេតាបូលីសដ៏ស្មុគស្មាញនេះមិនទាន់ច្បាស់នៅឡើយទេ។ ដំណើរការនេះត្រូវបានពិពណ៌នាដោយប្រើទម្រង់ថាមពលនៃការផ្ទេរអ៊ីយ៉ុងសូដ្យូមឬប៉ូតាស្យូមដោយប្រូតេអ៊ីន (រូបភាព 1.5,5) ។ ដោយធម្មជាតិនៃការផ្លាស់ប្តូរទម្រង់ទាំងនេះដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការផ្លាស់ប្តូរថេរក្នុងការអនុលោមតាមប្រូតេអ៊ីនដឹកជញ្ជូន (ដំណើរការដែលត្រូវការថាមពល) មនុស្សម្នាក់អាចវិនិច្ឆ័យ stoichiometry នៃការផ្លាស់ប្តូរ: អ៊ីយ៉ុងប៉ូតាស្យូមពីរត្រូវបានផ្លាស់ប្តូរសម្រាប់អ៊ីយ៉ុងសូដ្យូមបី។
ម៉ាស៊ីនបូម Na/K ដូចគ្នាទៅនឹងការដាច់ឆ្ងាយណា+ /K + -dependent membrane ATPase, រារាំងជាពិសេសដោយ glycoside បេះដូង ouabain (strophanthin) ។ ដោយសារប្រតិបត្តិការរបស់ម៉ាស៊ីនបូម Na/K គឺជាប្រតិកម្មគីមីច្រើនដំណាក់កាល វាដូចជាប្រតិកម្មគីមីទាំងអស់ ពឹងផ្អែកយ៉ាងធំទៅលើសីតុណ្ហភាព ដែល

អង្ករ។ ១.៦.ដ្យាក្រាមនៃ Na/K pump-ATPase (ដាក់ចូលទៅក្នុងស្រទាប់ខ្លាញ់នៃភ្នាសប្លាស្មា) ដែលក្នុងវដ្តមួយដកអ៊ីយ៉ុង Na + ចំនួនបីចេញពីកោសិកាប្រឆាំងនឹងជម្រាលសក្តានុពល និងកំហាប់ ហើយនាំ K ions ពីរចូលទៅក្នុងកោសិកា។ + . ក្នុងអំឡុងពេលដំណើរការនេះ ម៉ូលេគុល ATP មួយត្រូវបានបំបែកទៅជា ADP និង phosphate ។ ដ្យាក្រាមបង្ហាញ ATPase ជាឌីម័រដែលមានអនុផ្នែកធំ (មុខងារ) និងតូច។ នៅក្នុងភ្នាសវាមានជា tetramer ដែលបង្កើតឡើងដោយអនុពីរធំ និងតូចពីរ
បង្ហាញក្នុងរូប។ ១.៧. នៅទីនេះលំហូរនៃអ៊ីយ៉ុងសូដ្យូមពីកោសិកាសាច់ដុំត្រូវបានបង្ហាញទាក់ទងទៅនឹងពេលវេលា; នេះគឺសមមូលទៅនឹងលំហូរនៃអ៊ីយ៉ុងសូដ្យូមដែលសម្របសម្រួលដោយប្រតិបត្តិការនៃស្នប់ Na/K ពីព្រោះលំហូរអកម្មនៃអ៊ីយ៉ុងសូដ្យូមប្រឆាំងនឹងការប្រមូលផ្តុំ និងជម្រាលសក្តានុពលគឺតូចខ្លាំងណាស់។ ប្រសិនបើថ្នាំត្រូវបានត្រជាក់ប្រហែល 18 អង្សាសេ លំហូរនៃអ៊ីយ៉ុងសូដ្យូមពីកោសិកានឹងថយចុះយ៉ាងឆាប់រហ័ស 15 ដង ហើយភ្លាមៗបន្ទាប់ពីកំដៅ វានឹងត្រលប់មកកម្រិតដើមវិញ។ ការថយចុះនេះនៃលំហូរនៃអ៊ីយ៉ុងសូដ្យូមពីកោសិកាគឺច្រើនដងច្រើនជាងអ្វីដែលត្រូវគ្នាទៅនឹងការពឹងផ្អែកសីតុណ្ហភាពនៃដំណើរការសាយភាយ ឬប្រតិកម្មគីមីសាមញ្ញ។ ឥទ្ធិពលស្រដៀងគ្នានេះត្រូវបានគេសង្កេតឃើញនៅពេលដែលថាមពលមេតាបូលីសត្រូវបានបាត់បង់ជាលទ្ធផលនៃការពុល dinitrophenol (DNP) (រូបភាព 1.7,5) ។ ជាលទ្ធផលលំហូរនៃអ៊ីយ៉ុងសូដ្យូមពីកោសិកាត្រូវបានធានាដោយប្រតិកម្មដែលពឹងផ្អែកលើថាមពល - ស្នប់សកម្ម។ លក្ខណៈមួយទៀតនៃស្នប់ រួមជាមួយនឹងសីតុណ្ហភាព និងថាមពលដ៏សំខាន់គឺវត្តមាននៃកម្រិតតិត្ថិភាព (ដូចជាប្រតិកម្មគីមីផ្សេងទៀតទាំងអស់); នេះមានន័យថាល្បឿននៃស្នប់មិនអាចកើនឡើងដោយគ្មានកំណត់ទេ ដោយសារកំហាប់នៃអ៊ីយ៉ុងដឹកជញ្ជូនកើនឡើង (រូបភាព 1.8)។ ផ្ទុយទៅវិញ លំហូរនៃសារធាតុដែលសាយភាយដោយអសកម្មកើនឡើងក្នុងសមាមាត្រទៅនឹងភាពខុសគ្នានៃកំហាប់ ស្របតាមច្បាប់នៃការសាយភាយ (សមីការទី 1 និងទី 2) ។

អង្ករ។ ១.៧. ក, ខ។ការដឹកជញ្ជូន Na សកម្ម +. អ័ក្ស Y៖លំហូរនៃវិទ្យុសកម្ម 24 Na + ពីកោសិកា (imp./min) ។ អ័ក្ស Abscissa៖ពេលវេលាចាប់តាំងពីការចាប់ផ្តើមនៃការពិសោធន៍។ ក.ក្រឡាត្រូវបានត្រជាក់ពី 18.3 ° C ទៅ 0.5 ° C; លំហូរណា+ ពីកោសិកាក្នុងអំឡុងពេលនេះត្រូវបានរារាំង។ ខ.ការបង្ក្រាបលំហូរនៃ Na + ពីកោសិកាជាមួយ dinitrophenol (DNP) នៅកំហាប់ 0.2 mmol / l (កែប្រែ)
បន្ថែមពីលើស្នប់ Na/K ភ្នាសប្លាស្មាមានយ៉ាងហោចណាស់ស្នប់មួយបន្ថែមទៀត - កាល់ស្យូម;ស្នប់នេះបូមជាតិកាល់ស្យូមអ៊ីយ៉ុង (Ca 2+) ចេញពីកោសិកា ហើយចូលរួមក្នុងការរក្សាកំហាប់ខាងក្នុងកោសិកានៅកម្រិតទាបបំផុត (តារាង 1.1)។ ម៉ាស៊ីនបូមកាល់ស្យូមមានវត្តមាននៅក្នុងដង់ស៊ីតេខ្ពស់នៅក្នុងកោសិកាសាច់ដុំ sarcoplasmic reticulum ដែលប្រមូលផ្តុំអ៊ីយ៉ុងកាល់ស្យូមដែលជាលទ្ធផលនៃការបំបែកម៉ូលេគុល ATP (សូមមើលជំពូកទី 4) ។
ឥទ្ធិពលនៃស្នប់ Na/K លើសក្តានុពលភ្នាស និងបរិមាណកោសិកា . នៅក្នុងរូបភព។ រូបភាពទី 1.9 បង្ហាញពីសមាសធាតុផ្សេងៗនៃចរន្តភ្នាស និងបង្ហាញពីការប្រមូលផ្តុំអ៊ីយ៉ុងក្នុងកោសិកាដែល

អង្ករ។ ១.៨.ទំនាក់ទំនងរវាងអត្រានៃការដឹកជញ្ជូនម៉ូលេគុលនិងការប្រមូលផ្តុំរបស់ពួកគេ (នៅច្រកចូលទៅឆានែលឬនៅកន្លែងភ្ជាប់នៃស្នប់) កំឡុងពេលសាយភាយតាមឆានែលឬកំឡុងពេលដឹកជញ្ជូនបូម។ ក្រោយមកទៀតក្លាយជាឆ្អែតនៅកំហាប់ខ្ពស់ (ល្បឿនអតិបរមា, Vmax ) តម្លៃនៅលើអ័ក្ស x ដែលត្រូវគ្នានឹងពាក់កណ្តាលល្បឿនបូមអតិបរមា ( Vmax /2) គឺជាកំហាប់លំនឹង TO ម

អង្ករ។ ១.៩.ដ្យាក្រាមបង្ហាញពីការប្រមូលផ្តុំ Na+ , K+ និង Cl - ខាងក្នុង និងខាងក្រៅកោសិកា និងផ្លូវសម្រាប់ការជ្រៀតចូលនៃអ៊ីយ៉ុងទាំងនេះតាមរយៈភ្នាសកោសិកា (តាមរយៈបណ្តាញអ៊ីយ៉ុងជាក់លាក់ ឬដោយប្រើស្នប់ Na/K ។ នៅកម្រិតកំហាប់ដែលបានផ្តល់ឱ្យ សក្តានុពលលំនឹង E Na, E K និង E С l – ស្មើនឹងអ្វីដែលបានបង្ហាញ សក្តានុពលភ្នាសអេម = – 90 mV
ធានានូវអត្ថិភាពរបស់ពួកគេ។ ចរន្តខាងក្រៅនៃអ៊ីយ៉ុងប៉ូតាស្យូមត្រូវបានគេសង្កេតឃើញតាមរយៈបណ្តាញប៉ូតាស្យូម ចាប់តាំងពីសក្តានុពលនៃភ្នាសគឺមានលក្ខណៈអេឡិចត្រូប៉ូតាស្យូមច្រើនជាងសក្តានុពលលំនឹងសម្រាប់អ៊ីយ៉ុងប៉ូតាស្យូម។ ចរន្តសរុបនៃបណ្តាញសូដ្យូមគឺទាបជាងបណ្តាញប៉ូតាស្យូម ពោលគឺឧ។ ឆានែលសូដ្យូមត្រូវបានបើកញឹកញាប់តិចជាងឆានែលប៉ូតាស្យូមនៅសក្តានុពលសម្រាក; ទោះយ៉ាងណាក៏ដោយ ប្រមាណចំនួនដូចគ្នានៃអ៊ីយ៉ុងសូដ្យូមចូលទៅក្នុងកោសិកា នៅពេលដែលអ៊ីយ៉ុងប៉ូតាស្យូមចេញពីកោសិកា ពីព្រោះការប្រមូលផ្តុំដ៏ធំ និងជម្រាលសក្តានុពលត្រូវបានទាមទារសម្រាប់អ៊ីយ៉ុងសូដ្យូមដើម្បីសាយភាយចូលទៅក្នុងកោសិកា។ ម៉ាស៊ីនបូម Na/K ផ្តល់សំណងដ៏ល្អសម្រាប់ចរន្តសាយភាយអកម្ម ព្រោះវាដឹកជញ្ជូនអ៊ីយ៉ុងសូដ្យូមចេញពីកោសិកា និងអ៊ីយ៉ុងប៉ូតាស្យូមចូលទៅក្នុងវា។ ដូច្នេះ ស្នប់មានអេឡិចត្រូហ្សែន ដោយសារភាពខុសគ្នានៃចំនួននៃការចោទប្រកាន់ដែលបានផ្ទេរចូល និងចេញពីកោសិកា ដែលក្នុងល្បឿនធម្មតានៃប្រតិបត្តិការរបស់វាបង្កើតសក្តានុពលភ្នាសប្រហែល 10 mV គឺមានលក្ខណៈអេឡិចត្រុងច្រើនជាងប្រសិនបើវាត្រូវបានផលិតដោយលំហូរអ៊ីយ៉ុងអកម្មតែម្នាក់ឯង (សូមមើលសមីការទី 7) ។ ជាលទ្ធផលសក្តានុពលភ្នាសចូលទៅជិតសក្តានុពលលំនឹងប៉ូតាស្យូមដែលកាត់បន្ថយការលេចធ្លាយនៃអ៊ីយ៉ុងប៉ូតាស្យូម។ សកម្មភាពម៉ាស៊ីនបូម / K ត្រូវបានគ្រប់គ្រងដោយកំហាប់ខាងក្នុងកោសិកានៃអ៊ីយ៉ុងសូដ្យូម។ ល្បឿននៃការបូមថយចុះនៅពេលដែលកំហាប់អ៊ីយ៉ុងសូដ្យូមដែលត្រូវយកចេញពីកោសិកាថយចុះ (រូបភាព 1.8) ដូច្នេះហើយការបូម និងលំហូរនៃអ៊ីយ៉ុងសូដ្យូមទៅក្នុងកោសិកាមានតុល្យភាពគ្នា រក្សាកំហាប់សូដ្យូមក្នុងកោសិកា។ អ៊ីយ៉ុងនៅកម្រិតប្រហែល 10 mmol / L ។
ដើម្បីរក្សាលំនឹងរវាងការបូម និងចរន្តភ្នាសអកម្ម ម៉ូលេគុលបូម Na/K ជាច្រើនទៀតត្រូវការជាចាំបាច់ជាងប្រូតេអ៊ីនឆានែលសម្រាប់អ៊ីយ៉ុងប៉ូតាស្យូម និងសូដ្យូម។ នៅពេលដែលឆានែលត្រូវបានបើក អ៊ីយ៉ុងរាប់សិបពាន់ឆ្លងកាត់វាក្នុងរយៈពេលពីរបីមីលីវិនាទី (សូមមើលខាងលើ) ហើយចាប់តាំងពីឆានែលនេះជាធម្មតាបើកច្រើនដងក្នុងមួយវិនាទី សរុបមានច្រើនជាង 10 5 អ៊ីយ៉ុងឆ្លងកាត់ក្នុងអំឡុងពេលនេះ។ ប្រូតេអ៊ីនបូមតែមួយផ្លាស់ទីអ៊ីយ៉ុងសូដ្យូមជាច្រើនរយក្នុងមួយវិនាទី ដូច្នេះភ្នាសប្លាស្មាត្រូវតែមានម៉ូលេគុលបូមប្រហែល 1000 ដងច្រើនជាងម៉ូលេគុលឆានែល។ ការវាស់វែងនៃចរន្តឆានែលនៅពេលសម្រាកបានបង្ហាញពីវត្តមានជាមធ្យមនៃប៉ូតាស្យូមមួយនិងឆានែលបើកចំហសូដ្យូមមួយក្នុងមួយភ្នាស 1 μm 2; វាកើតឡើងពីនេះដែលម៉ូលេគុលប្រហែល 1000 នៃស្នប់ Na/K គួរតែមានវត្តមាននៅក្នុងចន្លោះដូចគ្នាពោលគឺឧ។ ចម្ងាយរវាងពួកវាគឺជាមធ្យម 34 nm; អង្កត់ផ្ចិតនៃប្រូតេអ៊ីនបូមដែលជាប្រូតេអ៊ីនឆានែលគឺ 8-10 nm ។ ដូច្នេះ ភ្នាសមានដង់ស៊ីតេច្រើនជាមួយម៉ូលេគុលបូម
ការពិតដែលថាលំហូរនៃអ៊ីយ៉ុងសូដ្យូមចូលទៅក្នុងកោសិកានិងអ៊ីយ៉ុងប៉ូតាស្យូមចេញពីកោសិកាត្រូវបានផ្តល់សំណងដោយប្រតិបត្តិការនៃស្នប់នោះមានផលវិបាកមួយផ្សេងទៀតដែលមាននៅក្នុងការរក្សាសម្ពាធ osmotic មានស្ថេរភាពនិង កម្រិតសំឡេងថេរ។នៅខាងក្នុងកោសិកាមានកំហាប់ខ្ពស់នៃ anions ធំ ដែលភាគច្រើនជាប្រូតេអ៊ីន (A - ក្នុងតារាង 1.1) ដែលមិនអាចជ្រាបចូលទៅក្នុងភ្នាស (ឬជ្រាបចូលយឺតៗ) ដូច្នេះហើយគឺជាសមាសធាតុថេរនៅក្នុងកោសិកា។ ដើម្បីធ្វើឱ្យមានតុល្យភាពបន្ទុកនៃ anions ទាំងនេះ ចំនួនស្មើគ្នានៃ cations គឺត្រូវការ។ ដោយសារតែសកម្មភាពរបស់ម៉ាស៊ីនបូម Na/K សារធាតុ cations ទាំងនេះភាគច្រើនជាអ៊ីយ៉ុងប៉ូតាស្យូម។ ការកើនឡើងគួរឱ្យកត់សម្គាល់នៃកំហាប់អ៊ីយ៉ុងក្នុងកោសិកាអាចកើតឡើងតែជាមួយនឹងការកើនឡើងនៃកំហាប់អ៊ីយ៉ុងដោយសារតែលំហូរនៃ C1 - តាមបណ្តោយជម្រាលនៃការប្រមូលផ្តុំទៅក្នុងកោសិកា (តារាង 1.1) ប៉ុន្តែសក្តានុពលភ្នាសប្រឆាំងនឹងបញ្ហានេះ។ ចរន្តចូល Cl - ត្រូវបានគេសង្កេតឃើញរហូតដល់សក្តានុពលលំនឹងសម្រាប់អ៊ីយ៉ុងក្លរីនត្រូវបានឈានដល់។ នេះត្រូវបានសង្កេតឃើញនៅពេលដែលជម្រាលអ៊ីយ៉ុងក្លរីនគឺស្ទើរតែផ្ទុយទៅនឹងជម្រាលអ៊ីយ៉ុងប៉ូតាស្យូម ចាប់តាំងពីអ៊ីយ៉ុងក្លរីនត្រូវបានចោទប្រកាន់អវិជ្ជមាន (សមីការទី 4)។ ដូច្នេះកំហាប់ intracellular ទាបនៃអ៊ីយ៉ុងក្លរីនត្រូវបានបង្កើតឡើង ដែលត្រូវគ្នាទៅនឹងកំហាប់ក្រៅកោសិកាទាបនៃអ៊ីយ៉ុងប៉ូតាស្យូម។ លទ្ធផលគឺជាការកំណត់នៃចំនួនអ៊ីយ៉ុងសរុបនៅក្នុងក្រឡា។ ប្រសិនបើសក្តានុពលនៃភ្នាសធ្លាក់ចុះនៅពេលដែលស្នប់ Na/K ត្រូវបានរារាំង ឧទាហរណ៍ក្នុងអំឡុងពេល anoxia នោះសក្តានុពលលំនឹងសម្រាប់អ៊ីយ៉ុងក្លរីនថយចុះ ហើយកំហាប់ខាងក្នុងនៃអ៊ីយ៉ុងក្លរួកើនឡើងទៅតាមនោះ។ ស្ដារតុល្យភាពនៃការចោទប្រកាន់, អ៊ីយ៉ុងប៉ូតាស្យូមក៏ចូលទៅក្នុងកោសិកា; ការប្រមូលផ្តុំសរុបនៃអ៊ីយ៉ុងនៅក្នុងកោសិកាកើនឡើងដែលបង្កើនសម្ពាធ osmotic; នេះបង្ខំឱ្យទឹកចូលទៅក្នុងកោសិកា។ កោសិកាហើម។ ការហើមនេះត្រូវបានអង្កេតនៅក្នុង Vivo នៅក្នុងលក្ខខណ្ឌនៃការខ្វះថាមពល។
ជម្រាលការផ្តោតអារម្មណ៍ ណា + ជាកម្លាំងជំរុញសម្រាប់ការដឹកជញ្ជូនភ្នាស . សារៈសំខាន់នៃស្នប់ Na/K សម្រាប់កោសិកាមិនត្រូវបានកំណត់ចំពោះស្ថេរភាពនៃជម្រាល K + និង Na + ធម្មតានៅលើភ្នាសទេ។ ថាមពលត្រូវបានរក្សាទុកក្នុងជម្រាលភ្នាសណា+ ជាញឹកញាប់ត្រូវបានគេប្រើដើម្បីជួយសម្រួលដល់ការដឹកជញ្ជូនភ្នាសនៃសារធាតុផ្សេងទៀត។ ឧទាហរណ៍នៅក្នុងរូបភព។ 1.10 បង្ហាញ "ភាពសាមញ្ញ"ណា+ និងម៉ូលេគុលស្ករចូលទៅក្នុងកោសិកា។ ប្រូតេអ៊ីនដឹកជញ្ជូនភ្នាសដឹកជញ្ជូនម៉ូលេគុលស្ករចូលទៅក្នុងកោសិកាសូម្បីតែប្រឆាំងនឹងជម្រាលនៃការផ្តោតអារម្មណ៍ ខណៈពេលដែល Na + ផ្លាស់ទីតាមបណ្តោយការផ្តោតអារម្មណ៍ និងជម្រាលសក្តានុពល ផ្តល់ថាមពលសម្រាប់ ការដឹកជញ្ជូនស្ករ។ការដឹកជញ្ជូនជាតិស្ករនេះគឺពឹងផ្អែកទាំងស្រុងទៅលើអត្ថិភាពនៃជម្រាលខ្ពស់។ណា+ ; ប្រសិនបើការប្រមូលផ្តុំ intracellularណា+ កើនឡើងយ៉ាងខ្លាំងការដឹកជញ្ជូនជាតិស្ករឈប់។ សម្រាប់ផ្សេងៗគ akharov មានប្រព័ន្ធស៊ីមកាតផ្សេងៗគ្នា។ ការដឹកជញ្ជូនអាស៊ីតអាមីណូចូលទៅក្នុងកោសិកាដែលស្រដៀងនឹងការដឹកជញ្ជូនគ akharov បានបង្ហាញនៅក្នុងរូបភព។ ១.១០; វាក៏ត្រូវបានផ្តល់ដោយជម្រាលផងដែរ។ណា+ , មានយ៉ាងហោចណាស់ប្រព័ន្ធ symport ប្រាំផ្សេងគ្នា ដែលនីមួយៗមានឯកទេសសម្រាប់ក្រុមមួយនៃអាស៊ីតអាមីណូដែលពាក់ព័ន្ធ។
បន្ថែមពីលើប្រព័ន្ធស៊ីមកាតក៏មានផងដែរ។ "អ្នកប្រឆាំងច្រកទ្វារ" ។ឧទាហរណ៍ មួយក្នុងចំនោមពួកគេក្នុងវដ្តមួយផ្ទេរអ៊ីយ៉ុងកាល់ស្យូមមួយពីកោសិកាជាថ្នូរនឹងអ៊ីយ៉ុងសូដ្យូមចូលបី (រូបភាព 1.10) ។ ថាមពលសម្រាប់ ការដឹកជញ្ជូន Ca 2+ត្រូវបានបង្កើតឡើងដោយសារតែការបញ្ចូលអ៊ីយ៉ុងសូដ្យូមចំនួនបីតាមបណ្តោយការផ្តោតអារម្មណ៍ និងជម្រាលសក្តានុពល។ ថាមពលនេះគឺគ្រប់គ្រាន់ (នៅសក្តានុពលសម្រាក) ដើម្បីរក្សាជម្រាលអ៊ីយ៉ុងកាល់ស្យូមខ្ពស់ (ពីតិចជាង 10-7 mol/L នៅខាងក្នុងកោសិកាទៅប្រហែល 2 mmol/L នៅខាងក្រៅកោសិកា)។
Endo- និង exocytosis . ចំពោះសារធាតុមួយចំនួនដែលចូលទៅក្នុងកោសិកា ឬត្រូវតែបញ្ចេញចេញ

អង្ករ។ ១.១០.ប្រូតេអ៊ីនដែលបង្កប់ក្នុងស្រទាប់ lipid bilayer នៃភ្នាសសម្របសម្រួលភាពស៊ីសង្វាក់គ្នានៃជាតិគ្លុយកូស និង Na + ទៅក្នុងកោសិកា ក៏ដូចជា Ca2+/Na+ -antiport ដែលកម្លាំងជំរុញគឺ Na + gradient នៅលើភ្នាសកោសិកា
ពីវាមិនមានបណ្តាញដឹកជញ្ជូន; សារធាតុទាំងនេះរួមបញ្ចូលឧទាហរណ៍ប្រូតេអ៊ីននិងកូលេស្តេរ៉ុល។ ពួកគេអាចឆ្លងកាត់ភ្នាសប្លាស្មាចូលទៅក្នុង vesicles,ឬ ពពុះ,តាមរយៈ endo- និង exocytosis ។ នៅក្នុងរូបភព។ រូបភាព 1.11 បង្ហាញពីយន្តការសំខាន់ៗនៃដំណើរការទាំងនេះ។ នៅក្នុង exocytosis សរីរាង្គមួយចំនួន (សូមមើលខាងក្រោម) បង្កើតជា vesicles ដែលពោរពេញទៅដោយសារធាតុដែលត្រូវការដកចេញពីកោសិកា ដូចជាអរម៉ូន ឬអង់ស៊ីមក្រៅកោសិកា។ នៅពេលដែល vesicles បែបនេះទៅដល់ភ្នាសប្លាស្មា ភ្នាស lipid របស់ពួកវាប្រសព្វជាមួយវា ដូច្នេះហើយទើបអាចឱ្យមាតិកាទាំងនោះគេចចេញពីបរិយាកាសខាងក្រៅ។ នៅក្នុងដំណើរការផ្ទុយគ្នា endocytosis, ភ្នាសប្លាស្មា invaginates, បង្កើតជារណ្តៅមួយ, បន្ទាប់មកកាន់តែជ្រៅនិងបិទ, បង្កើត vesicle intracellular ពោរពេញទៅដោយសារធាតុរាវ extracellular និង macromolecules មួយចំនួន។ ដើម្បីធានាបាននូវការបញ្ចូលគ្នានៃភ្នាសនេះ និងការបិទនៃ vesicle ធាតុ contractile នៃ cytoskeleton ធ្វើសកម្មភាពដោយភ្ជាប់ជាមួយភ្នាសខ្លួនឯង (សូមមើលខាងក្រោម) ។ Endocytosis មិនតែងតែពាក់ព័ន្ធនឹងការចាប់យកបរិយាកាសខាងក្រៅចូលទៅក្នុងកោសិកានោះទេ។ ភ្នាសកោសិកាជារឿយៗផ្ទុកនូវអ្នកទទួលជាក់លាក់សម្រាប់ម៉ាក្រូម៉ូលេគុល ដូចជាអាំងស៊ុយលីន ឬអង់ទីហ្សែន ដែលជារឿយៗត្រូវបានរៀបចំជាក្រុមឯកទេស។ បន្ទាប់ពី macromolecules ទាំងនេះភ្ជាប់ទៅនឹង receptor របស់ពួកគេ endocytosis កើតឡើងនៅក្នុងតំបន់ភ្នាសជុំវិញ receptor ហើយ macromolecule ត្រូវបានជ្រើសរើសចូលទៅក្នុងកោសិកា (រូបភាព 1.12, B) ។
Endo- និង exocytosis កើតឡើងជាបន្តបន្ទាប់នៅក្នុងកោសិកា។ បរិមាណនៃការផ្លាស់ប្តូរសម្ភារៈភ្នាសមានសារៈសំខាន់; ក្នុងរយៈពេល 1 ម៉ោង macrophage ស្រូបយកពីរដងនៃផ្ទៃនៃភ្នាស cytoplasmic របស់វាក្នុងទម្រង់ជា vesicles ។ នៅក្នុងកោសិកាភាគច្រើន ការផ្លាស់ប្តូរនៃសម្ភារៈភ្នាសមិនកើតឡើងខ្លាំងនោះទេ ប៉ុន្តែវាគួរតែនៅតែមានសារៈសំខាន់។

អង្ករ។ ១.១១.Exocytosis និង endocytosis ។ ឡើង៖ vesicle intracellular fuses ជាមួយ lipid bilayer នៃភ្នាសប្លាស្មា ហើយបើកចូលទៅក្នុង extracellular space។ ដំណើរការនេះត្រូវបានគេហៅថា exocytosis ។ នៅខាងក្រោមនេះ:ភ្នាសប្លាស្មាចូលទៅក្នុងផ្ទៃតូចមួយ ហើយបញ្ចេញ vesicle ដែលពោរពេញទៅដោយសារធាតុ extracellular ។ ដំណើរការនេះត្រូវបានគេហៅថា endocytosis
១.៣. ការដឹកជញ្ជូនសារធាតុនៅក្នុងកោសិកា
Endo- និង exocytosis មិនត្រឹមតែជាដំណើរការនៃការដឹកជញ្ជូនសារធាតុឆ្លងកាត់ភ្នាសកោសិកាប៉ុណ្ណោះទេ ប៉ុន្តែវាក៏ជាដំណើរការនៃការផ្លាស់ប្តូរភ្នាសផងដែរ - សមាសធាតុរចនាសម្ព័ន្ធនៃកោសិកាខ្លួនឯង។ ប្រធានបទនៃការពិចារណានៅក្នុងផ្នែកនេះគឺជាដំណើរការដឹកជញ្ជូនស្រដៀងគ្នាផ្សេងទៀតនៅក្នុងកោសិកា និងសរីរាង្គរបស់វា។
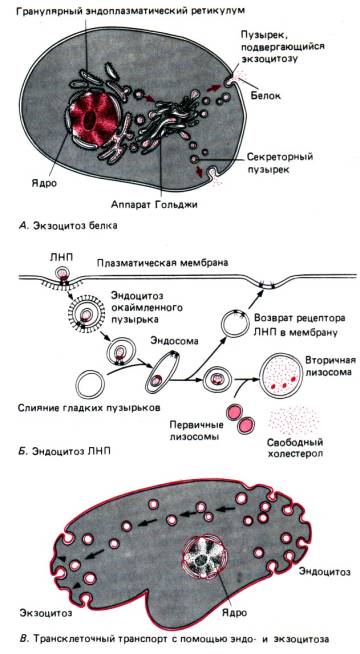
អង្ករ។ ១.១២. ក-ខ។គ្រោងការណ៍នៃដំណើរការរួមទាំង exo- និង endocytosis ។ ក.ប្រូតេអ៊ីនដែលត្រូវបានសំយោគនៅក្នុង reticulum endoplasmic granular ត្រូវបានដឹកជញ្ជូនតាមបរិធាន Golgi ទៅកាន់ភ្នាសប្លាស្មាដែលជាកន្លែងដែលវាត្រូវបានលាក់ដោយ exocytosis ។ ខ.កូលេស្តេរ៉ុលភ្ជាប់ទៅនឹង LDL (Low-density lipoprotein) ភាគល្អិតភ្ជាប់ទៅនឹងភ្នាសប្លាស្មា ជំរុញការបង្កើត vesicle endocytic នៅក្នុងតំបន់នៃភ្នាសនេះ ហើយត្រូវបានបញ្ជូនទៅ lysosomes ដែលវាត្រូវបានបញ្ចេញ។ INវត្ថុធាតុក្រៅកោសិកាដែលចាប់យកអំឡុងពេលជំងឺ endocytosis (ក្នុងរូប នៅខាងស្តាំ)ដឹកជញ្ជូនតាមរយៈកោសិកានៅក្នុង vesicles ឬ vesicles និងត្រូវបានបញ្ចេញតាមរយៈ exocytosis (ក្នុងរូបភាព ឆ្វេង)
ការសាយភាយ . ជាធម្មតានៅក្នុង cytosol ភាពខុសគ្នានៃការប្រមូលផ្តុំត្រូវបានលុបចោលដោយសារតែការសាយភាយ; ដូចគ្នានេះដែរគឺជាការពិតសម្រាប់សារធាតុរាវដែលមាននៅក្នុងសរីរាង្គ។ ដោយសារតែកំហាប់ខ្ពស់នៃប្រូតេអ៊ីនរលាយ ការសាយភាយនៅទីនេះគឺយឺតជាងក្នុងទឹក។ ភ្នាស lipid - នៅជុំវិញកោសិកានិងក្នុងសរីរាង្គ - គឺជាវត្ថុរាវពីរវិមាត្រដែលការសាយភាយកើតឡើង។ Lipids នៅក្នុងភ្នាស bilayer សាយភាយនៅក្នុងស្រទាប់របស់វាផ្ទាល់ ដោយកម្រផ្លាស់ទីពីមួយទៅមួយទៀត។ ប្រូតេអ៊ីនដែលបានជ្រមុជក្នុងពួកវាគឺជាការចល័តណាស់; ពួកវាបង្វិលជុំវិញអ័ក្សកាត់កែងទៅនឹងភ្នាស ឬសាយភាយនៅពេលក្រោយជាមួយនឹងថេរនៃការសាយភាយខុសគ្នាខ្លាំង យឺតជាង phospholipids 2-10,000 ដង។ ដូច្នេះប្រសិនបើប្រូតេអ៊ីនខ្លះផ្លាស់ទីដោយសេរីនៅក្នុងស្រទាប់ lipid និងក្នុងល្បឿនដូចគ្នាទៅនឹងម៉ូលេគុល lipid ខ្លួនវា នោះផ្សេងទៀតត្រូវបានបោះយុថ្កា ពោលគឺឧ។ ភ្ជាប់យ៉ាងរឹងមាំទៅនឹង cytoskeleton ។ មានការប្រមូលផ្តុំ "អចិន្រ្តៃយ៍" នៃប្រូតេអ៊ីនជាក់លាក់នៅក្នុងភ្នាស ឧទាហរណ៍ រចនាសម្ព័ន្ធមុន និងក្រោយសំយោគនៃកោសិកាប្រសាទ។ ប្រូតេអ៊ីនដែលផ្លាស់ទីដោយសេរីអាចត្រូវបានបង្ហាញដោយការចងពួកវាទៅនឹងថ្នាំជ្រលក់ fluorescent ដែលជាពន្លឺនៃពន្លឺដែលត្រូវបានបង្កឡើងដោយការបំភ្លឺខ្លីៗនៃតំបន់តូចមួយនៃភ្នាស។ ការពិសោធន៍បែបនេះបង្ហាញថាក្នុងរយៈពេលតិចជាង 1 នាទី ប្រូតេអ៊ីនដែលភ្ជាប់ទៅនឹងសារធាតុជ្រលក់ត្រូវបានចែកចាយស្មើៗគ្នាលើភ្នាសក្នុងចម្ងាយរហូតដល់ 10 μm។
ការដឹកជញ្ជូនសកម្មនៅក្នុងភ្នាសសរីរាង្គ .
ដំណើរការដឹកជញ្ជូនសកម្ម ដែលដើរតួនាទីយ៉ាងសំខាន់ក្នុងដំណើរការនៃភ្នាសប្លាស្មា ក៏កើតឡើងនៅក្នុងកោសិកា នៅក្នុងភ្នាសនៃសរីរាង្គ។ មាតិកាជាក់លាក់នៃសរីរាង្គផ្សេងៗត្រូវបានបង្កើតឡើងមួយផ្នែកដោយសារតែការសំយោគខាងក្នុង និងមួយផ្នែកដោយសារការដឹកជញ្ជូនសកម្មពី cytosol ។ ឧទាហរណ៍មួយនៃការបន្ទាប់គឺការបូម Ca 2+ ដែលបានរៀបរាប់ខាងលើនៅក្នុង sarcoplasmic reticulum នៃកោសិកាសាច់ដុំ។ វាគួរឱ្យចាប់អារម្មណ៍ជាពិសេសថានៅក្នុងករណីនៃការសំយោគ ATP នៅក្នុង mitochondria គោលការណ៍ផ្ទុយអនុវត្តចំពោះអ្វីដែលកើតឡើងនៅក្នុងស្នប់ ATPase នៃភ្នាសប្លាស្មា (រូបភាព 1.6) ។ កំឡុងពេលសំយោគ ATP ការរំលាយអាហារអុកស៊ីតកម្មនាំទៅរកការបង្កើតជម្រាលដ៏ចោត H+ នៅលើភ្នាសខាងក្នុង។ ជម្រាលនេះគឺជាកម្លាំងជំរុញសម្រាប់ដំណើរការបញ្ច្រាសនៃវដ្តបូមនៃការដឹកជញ្ជូនសកម្មនៃម៉ូលេគុល: អ៊ីយ៉ុង H + ផ្លាស់ទីឆ្លងកាត់ភ្នាសតាមបណ្តោយជម្រាល ហើយថាមពលដែលបានបញ្ចេញជាលទ្ធផលផ្តល់នូវការសំយោគ ATP ពី ADP និងផូស្វាត។ ជាលទ្ធផល ATP ផ្តល់ថាមពលដល់កោសិកា រួមទាំងការដឹកជញ្ជូនសកម្ម។
ការដឹកជញ្ជូននៅក្នុង vesicles . កោសិកានេះមានចំនួនច្រើននៃសរីរាង្គ និង vesicles ដែលពាក់ព័ន្ធ (រូបភាព 1.1) ។ សរីរាង្គទាំងនេះ និងជាពិសេស vesicles ស្ថិតនៅក្នុងចលនាថេរ ដឹកជញ្ជូនមាតិការបស់វាទៅសរីរាង្គផ្សេងទៀត ឬទៅភ្នាសប្លាស្មា។ vesicles ក៏អាចធ្វើចំណាកស្រុកពីភ្នាសកោសិកាទៅសរីរាង្គដូចជានៅក្នុង endocytosis ដែរ។
ដំណើរការ ការសម្ងាត់ប្រូតេអ៊ីនបង្ហាញក្នុងរូប។ 1.12, ក.ប្រូតេអ៊ីនត្រូវបានសំយោគនៅជិតស្នូលកោសិកានៅលើ ribosomes ដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹង reticulum endoplasmic (ដែលគេហៅថា granular ឬ rough, endoplasmic reticulum); នៅពេលដែលនៅក្នុង reticulum endoplasmic ប្រូតេអ៊ីនត្រូវបានខ្ចប់ចូលទៅក្នុង vesicles ដឹកជញ្ជូនដែលបំបែកចេញពីសរីរាង្គនិងធ្វើចំណាកស្រុកទៅបរិធាន Golgi ។ នៅទីនេះពួកវាបញ្ចូលគ្នាជាមួយ cisternae នៃបរិធាន Golgi ដែលប្រូតេអ៊ីនត្រូវបានកែប្រែ (ឧ. បំប្លែងទៅជា glycoprotein)។ នៅចុងបញ្ចប់នៃ cisternae, vesicles បំបែកម្តងទៀត។ សរសៃឈាមសម្ងាត់ដែលផ្ទុកប្រូតេអ៊ីនដែលបានកែប្រែផ្លាស់ទីទៅភ្នាសប្លាស្មាហើយបញ្ចេញមាតិការបស់វាដោយ exocytosis ។
ឧទាហរណ៍មួយទៀតនៃផ្លូវដឹកជញ្ជូនក្នុងក្រឡាមួយត្រូវបានបង្ហាញក្នុងរូប។ 1.12, ខ; នេះគឺជាការស្រូបយកកូលេស្តេរ៉ុលដោយកោសិកា។ កូលេស្តេរ៉ុលដែលដឹកជញ្ជូនក្នុងឈាមត្រូវបានចងភ្ជាប់ជាចម្បងទៅនឹងប្រូតេអ៊ីន ដូចជាភាគល្អិត " lipoprotein ដង់ស៊ីតេទាប"(LDL) ។ ភាគល្អិតបែបនេះភ្ជាប់ទៅនឹងតំបន់ភ្នាសជាក់លាក់ដែលមានអ្នកទទួល LDL ដែលជាកន្លែងដែល endocytosis កើតឡើងហើយ LDL ត្រូវបានបញ្ជូនទៅកោសិកានៅក្នុង vesicles "ព្រំដែន" ។ vesicles ទាំងនេះ fuses ដើម្បីបង្កើត endosomes និងបាត់បង់ "edging" របស់ពួកគេក្នុងអំឡុងពេលដំណើរការនេះ។ Endosomes បញ្ចូលគ្នាជាមួយ lysosomes បឋម ដែលមានអង់ស៊ីម hydrolytic លើសលុប ហើយបង្កើតជា lysosomes ធំជាងទីពីរ។ នៅក្នុងពួកវា កូលេស្តេរ៉ុលត្រូវបានបញ្ចេញចេញពីភាគល្អិត LDL ហើយសាយភាយចូលទៅក្នុង cytosol ដែលជាកន្លែងដែលវាអាចប្រើបាន ឧទាហរណ៍សម្រាប់ការសំយោគភ្នាស lipid ។ vesicles ដែលមិនមាន LDL ក៏ត្រូវបានបំបែកចេញពី endosomes ដែលផ្លាស់ទីក្នុងវិធីពិសេសមួយទៅកាន់ភ្នាសប្លាស្មា ហើយបញ្ចូលគ្នាជាមួយវា ត្រឡប់សម្ភារៈភ្នាស ហើយប្រហែលជាអ្នកទទួល LDL ។ ចាប់ពីពេលដែលភាគល្អិត LDL ភ្ជាប់ទៅនឹងភ្នាសរហូតដល់កូលេស្តេរ៉ុលត្រូវបានបញ្ចេញចេញពី lysosome បន្ទាប់បន្សំ 10-15 នាទីឆ្លងកាត់។ ការចុះខ្សោយនៃការផ្សារភ្ជាប់ និងការស្រូបយក LDL ពោលគឺការផ្គត់ផ្គង់កូលេស្តេរ៉ុលដល់កោសិកា ដើរតួយ៉ាងសំខាន់ក្នុងការវិវត្តនៃជំងឺធ្ងន់ធ្ងរ និងរីករាលដាល atherosclerosis ("ការឡើងរឹង" នៃសរសៃឈាម)។
មានផ្លូវដឹកជញ្ជូនជាច្រើនទៀតដែលស្រដៀងនឹងផ្លូវដែលបង្ហាញក្នុងរូប។ 1.11 និង 1.12, A ដោយមានជំនួយពី vesicles ជាក់លាក់ផ្លាស់ទីនៅក្នុងកោសិកា។ វាមិនត្រូវបានគេដឹងច្បាស់ពីរបៀបដែលពួកវាផ្លាស់ទីនោះទេ ប៉ុន្តែធាតុ cytoskeletal ប្រហែលជាពាក់ព័ន្ធនឹងដំណើរការនេះ។ vesicles អាចរុញតាម microtubules ក្នុងករណីនេះថាមពលសម្រាប់ចលនាត្រូវបានផ្តល់ឱ្យជាក់ស្តែងដោយប្រូតេអ៊ីនដែលមានទំនាក់ទំនងជាមួយ vesicles ATPase (សូមមើលខាងក្រោម) ។ វានៅតែមិនច្បាស់ថា តើមាន vesicles ផ្សេងគ្នាប៉ុន្មាន ដែលផ្លាស់ទីមួយទៅទិសមួយទៀត ទៅដល់គោលដៅរបស់ពួកគេ។ ពួកគេច្បាស់ជាត្រូវ "សម្គាល់" តាមរបៀបដែលត្រូវបានទទួលស្គាល់ដោយប្រព័ន្ធដឹកជញ្ជូន និងបំប្លែងទៅជាចលនាគោលដៅ។
ការដឹកជញ្ជូនដោយការបង្កើតនិងការបំផ្លាញសរីរាង្គ . រហូតមកដល់ពេលនេះយើងបានចាត់ទុក endo- និង exocytosis ជាដំណើរការនៃការដឹកជញ្ជូនមាតិកានៃ vesicles ។ មានទិដ្ឋភាពមួយផ្សេងទៀតនៃដំណើរការទាំងនេះដែលស្ថិតនៅក្នុងការពិតដែលថាការដកយកចេញនៃភ្នាសប្លាស្មានៅលើផ្នែកមួយនៃផ្ទៃកោសិកាដោយ endocytosis ហើយផ្ទុយទៅវិញការបន្ថែមរបស់វានៅលើមួយទៀតដោយ exocytosis ផ្លាស់ទីតំបន់ធំនៃភ្នាស (រូបភាព . 1.12.E) ផ្តល់ឱកាសដល់ក្រឡា ជាឧទាហរណ៍ បង្កើតជាការកើនឡើង ឬផ្លាស់ទី។
ការរៀបចំឡើងវិញស្រដៀងគ្នានេះក៏ជាតួយ៉ាងសម្រាប់ cytoskeleton ជាពិសេសសម្រាប់ microfilaments និង microtubules (រូបភាព 1.1)។ មីក្រូហ្វីលមានជាចម្បងនៃ ប្រូតេអ៊ីន F-actin,ដែលមានសមត្ថភាពប្រមូលផ្តុំទៅជាបណ្តុំសរសៃដែលជាលទ្ធផលនៃវត្ថុធាតុ polymerization នៃ monomer ពី cytosol ។ បណ្តុំត្រូវបានបែងចែកជារាងប៉ូល ពោលគឺពួកវាច្រើនតែដុះនៅចុងម្ខាង ដោយប្រមូលផ្តុំម៉ូលេគុល actin ថ្មី ចំណែកចុងម្ខាងទៀតគឺ inert ឬ disassembly កើតឡើងនៅទីនេះ។ ដោយសារតែការលូតលាស់រាងប៉ូលនេះ មីក្រូហ្វីលផ្លាស់ទីយ៉ាងមានប្រសិទ្ធភាព ហើយរចនាសម្ព័ន្ធនៃបណ្តាញរបស់វាអាចផ្លាស់ប្តូរបាន។ ការផ្លាស់ប្តូរនៃ actin ពីស្ថានភាព depolymerized (sol) ទៅរដ្ឋរៀបចំ (gel) អាចកើតឡើងយ៉ាងឆាប់រហ័សនៅក្រោមឥទ្ធិពលនៃប្រូតេអ៊ីនផ្សេងទៀតឬការផ្លាស់ប្តូរនៅក្នុងកំហាប់អ៊ីយ៉ុង (សូមមើលខាងក្រោម) ។ វាក៏មានប្រូតេអ៊ីនដែលបណ្តាលឱ្យសរសៃ actin បំបែកទៅជាបំណែកខ្លីៗ។ ការព្យាករណ៍ស្តើងនៃកោសិកាជាច្រើន - filopodia - មានបណ្តុំកណ្តាលនៃ actin (រូបភាព 1.1) ហើយចលនាផ្សេងៗនៃ filopodia ប្រហែលជាដោយសារការផ្លាស់ប្តូរ actin: polymerization - depolymerization ។
មីក្រូបំពង់ជារឿយៗក៏មានចលនាស្រដៀងគ្នាដែរ។ យន្តការនៃចលនាទាំងនេះគឺស្រដៀងគ្នា - ការធ្វើវត្ថុធាតុ polymerization នៃ tubulin ពី cytosol តាមរបៀបដែលចុងម្ខាងនៃ microtubule លូតលាស់ ខណៈពេលដែលម្ខាងទៀតមិនផ្លាស់ប្តូរ ឬ disassembly កើតឡើងនៅទីនោះ។ ដូច្នេះ microtubule ដោយការបន្ថែម ឬដកចេញសម្ភារៈសមស្រប អាចផ្លាស់ទីពេញ cytosol ។
ចលនាសកម្មនៃ cytoskeleton . ការផ្លាស់ប្តូររចនាសម្ព័ន្ធ cytoskeletal អាចកើតឡើងជាលទ្ធផលនៃចលនាសកម្មនិងការរៀបចំឡើងវិញដែលបានពិពណ៌នាខាងលើ។ ក្នុងករណីជាច្រើន ចលនារបស់ microtubules និង actin filaments ត្រូវបានជំរុញដោយប្រូតេអ៊ីន contractile ដែលភ្ជាប់សរសៃ ឬ tubules ហើយអាចផ្លាស់ទីពួកវាទាក់ទងគ្នាទៅវិញទៅមក។ កំប្រុក myosin និង dyneinមានវត្តមាននៅក្នុង cytosol នៃកោសិកាទាំងអស់នៅក្នុងការប្រមូលផ្តុំខ្ពស់ទាក់ទង; ពួកវាជាធាតុដែលបំប្លែងថាមពលទៅជាចលនានៅក្នុងកោសិកាឯកទេស (សាច់ដុំ) និងសរីរាង្គ (ស៊ីលីយ៉ា)។ នៅក្នុងកោសិកាសាច់ដុំ myosin បង្កើតជាសរសៃក្រាស់តម្រង់ទិសស្របទៅនឹងសរសៃ actin ។ ម៉ូលេគុល myosin ដែលមាន "ក្បាល" របស់វាភ្ជាប់ទៅនឹងសរសៃ actin ហើយដោយប្រើថាមពលរបស់ ATP ។ ផ្លាស់ទី myosin តាមបណ្តោយម៉ូលេគុល actin ។ Myosin បន្ទាប់មកបំបែកចេញពី actin ។ ការរួមបញ្ចូលគ្នានៃវដ្តនៃការតភ្ជាប់ - ផ្តាច់ទំនាក់ទំនងជាច្រើននាំទៅរកម៉ាក្រូស្កូប ការបង្រួមនៃសរសៃសាច់ដុំ(ជំពូកទី 4) ។ Dynein ដើរតួស្រដៀងគ្នាក្នុងចលនានៃ microtubules កំឡុងពេល cilia (រូបភាព 1.1) ។ នៅក្នុង cytoplasm នៃកោសិកាដែលមិនមានឯកទេស myosin និង dynein មិនបង្កើតជាសរសៃធម្មតាទេ ប៉ុន្តែក្នុងករណីភាគច្រើន ក្រុមតូចៗនៃម៉ូលេគុល។ សូម្បីតែនៅក្នុងការប្រមូលផ្តុំតូចបែបនេះក៏ដោយ ពួកវាមានសមត្ថភាពក្នុងការផ្លាស់ទី actin filaments ឬ microtubules ។ អង្ករ។ រូបភាពទី 1.13 បង្ហាញពីដំណើរការនេះ នៅពេលដែលម៉ូលេគុល myosin រាងប៉ូលផ្ទុយគ្នាក៏ត្រូវបានភ្ជាប់ទៅនឹងសរសៃ actin ពីរផងដែរ ដែលមានរាងប៉ូលក្នុងទិសដៅផ្សេងៗគ្នា។ ក្រុមក្បាល myosin ពត់ទៅកន្ទុយនៃម៉ូលេគុល ដោយប្រើប្រាស់ ATP នៅក្នុងដំណើរការ ហើយសរសៃ actin ទាំងពីរផ្លាស់ទីក្នុងទិសដៅផ្ទុយ បន្ទាប់មក myosin ផ្ដាច់ចេញពីពួកគេ។ ចលនានៃប្រភេទនេះ ក្នុងអំឡុងពេលដែលថាមពល ATP ត្រូវបានបំប្លែងទៅជាការងារមេកានិក អាចផ្លាស់ប្តូររូបរាងរបស់ cytoskeleton ហើយជាលទ្ធផលកោសិកា ហើយក៏ផ្តល់នូវការដឹកជញ្ជូនសរីរាង្គដែលទាក់ទងនឹង cytoskeleton ផងដែរ។
ដំណើរការនៃការដឹកជញ្ជូន intracellular អាចត្រូវបានបង្ហាញយ៉ាងច្បាស់បំផុតនៅលើ axon នៃកោសិកាសរសៃប្រសាទមួយ។ ការដឹកជញ្ជូន Axonត្រូវបានពិភាក្សានៅទីនេះយ៉ាងលម្អិត ដើម្បីបង្ហាញពីព្រឹត្តិការណ៍ដែលទំនងជាកើតឡើងតាមរបៀបស្រដៀងគ្នានៅក្នុងកោសិកាភាគច្រើន។ អ័ក្សដែលមានអង្កត់ផ្ចិតត្រឹមតែពីរបីមីក្រូអាចឈានដល់ប្រវែងមួយម៉ែត្រ ឬច្រើនជាងនេះ ហើយចលនានៃប្រូតេអ៊ីនដោយការសាយភាយចេញពីស្នូលទៅចុងចុងនៃអ័ក្សនឹងចំណាយពេលរាប់ឆ្នាំ។ វាត្រូវបានគេស្គាល់ជាយូរមកហើយថានៅពេលដែលផ្នែកណាមួយនៃ axon ឆ្លងកាត់ការរឹតបន្តឹងផ្នែកនៃ axon ដែលមានទីតាំងនៅជិតនឹងពង្រីក។ វាមើលទៅដូចជាលំហូរ centrifugal ត្រូវបានរារាំងនៅក្នុងអ័ក្ស។ បែប លំហូរ - ការដឹកជញ្ជូនអ័ក្សអាកាសលឿនត្រូវបានបង្ហាញដោយចលនានៃសញ្ញាសម្គាល់វិទ្យុសកម្ម ដូចនៅក្នុងការពិសោធន៍បង្ហាញក្នុងរូប។ ១.១៤. Radiolabeled leucine ត្រូវបានចាក់ចូលទៅក្នុង ganglion root dorsal ហើយបន្ទាប់មកវិទ្យុសកម្មត្រូវបានវាស់នៅក្នុងសរសៃប្រសាទ sciatic នៅចម្ងាយ 166 មីលីម៉ែត្រពីកោសិកាសរសៃប្រសាទពីម៉ោង 2 ដល់ទី 10 ។ លើសពី 10 ម៉ោង កម្រិតកំពូលនៃវិទ្យុសកម្មនៅកន្លែងចាក់ថ្នាំបានផ្លាស់ប្តូរបន្តិច។ ប៉ុន្តែរលកវិទ្យុសកម្មបានសាយភាយតាមអ័ក្សក្នុងល្បឿនថេរប្រហែល ៣៤ មីលីម៉ែត្រក្នុងរយៈពេល ២ ម៉ោង ឬ ៤១០ មីលីម៉ែត្រក្នុងមួយថ្ងៃ។ វាត្រូវបានបង្ហាញថានៅក្នុងណឺរ៉ូនទាំងអស់នៃសត្វដែលមានកំដៅក្នុងផ្ទះ ការដឹកជញ្ជូនអ័ក្សអាកាសលឿនកើតឡើងក្នុងល្បឿនដូចគ្នា ហើយមិនមានភាពខុសគ្នាគួរឱ្យកត់សម្គាល់ណាមួយត្រូវបានគេសង្កេតឃើញរវាងសរសៃស្តើង គ្មានមីអ៊ីលីន និង អ័ក្សក្រាស់បំផុត ក៏ដូចជារវាងសរសៃម៉ូទ័រ និងសរសៃប្រសាទ។ ប្រភេទនៃសញ្ញាសម្គាល់វិទ្យុសកម្មក៏មិនប៉ះពាល់ដល់អត្រានៃការដឹកជញ្ជូនតាមអ័ក្សលឿនដែរ។ សញ្ញាសម្គាល់អាចបម្រើជាភាពខុសគ្នានៃវិទ្យុសកម្ម

អង្ករ។ ១.១៣.ស្មុគ្រស្មាញ myosin ដែលមិនមែនជាសាច់ដុំ ជាមួយនឹងការតំរង់ទិសជាក់លាក់មួយ អាចភ្ជាប់ទៅនឹងសរសៃអំបោះនៃប៉ូលផ្សេងគ្នា ហើយដោយប្រើថាមពលនៃ ATP ផ្លាស់ប្តូរពួកវាទាក់ទងគ្នាទៅវិញទៅមក។
ម៉ូលេគុល ដូចជាអាស៊ីតអាមីណូផ្សេងៗ ដែលត្រូវបានរួមបញ្ចូលនៅក្នុងប្រូតេអ៊ីននៃរាងកាយកោសិកានៃណឺរ៉ូន។ ប្រសិនបើយើងវិភាគផ្នែកខាងចុងនៃសរសៃប្រសាទដើម្បីកំណត់ពីធម្មជាតិនៃអ្នកដឹកជញ្ជូនវិទ្យុសកម្មដែលដឹកជញ្ជូនមកទីនេះ នោះក្រុមហ៊ុនដឹកជញ្ជូនបែបនេះត្រូវបានរកឃើញជាចម្បងនៅក្នុងប្រភាគប្រូតេអ៊ីន ប៉ុន្តែក៏មាននៅក្នុងសមាសភាពនៃអ្នកសម្របសម្រួល និងអាស៊ីតអាមីណូឥតគិតថ្លៃផងដែរ។ ដោយដឹងថាលក្ខណៈសម្បត្តិនៃសារធាតុទាំងនេះមានភាពខុសប្លែកគ្នា ហើយទំហំនៃម៉ូលេគុលរបស់វាមានភាពខុសប្លែកគ្នាជាពិសេសនោះ យើងអាចពន្យល់ពីល្បឿនថេរនៃការដឹកជញ្ជូនដោយយន្តការដឹកជញ្ជូនធម្មតាសម្រាប់ពួកវាទាំងអស់។
បានពិពណ៌នាខាងលើ ការដឹកជញ្ជូនលឿន axonគឺ anterograde, i.e. ដឹកនាំឆ្ងាយពីរាងកាយកោសិកា។ វាត្រូវបានបង្ហាញថាសារធាតុមួយចំនួនផ្លាស់ទីពីបរិមាត្រទៅរាងកាយកោសិកាដោយប្រើ ការដឹកជញ្ជូន retrograde ។ឧទាហរណ៍ acetylcholinesterase ត្រូវបានដឹកជញ្ជូនក្នុងទិសដៅនេះក្នុងល្បឿន 2 ដងយឺតជាងល្បឿននៃការដឹកជញ្ជូនតាមអ័ក្សលឿន។ សញ្ញាសម្គាល់ដែលត្រូវបានប្រើជាញឹកញាប់នៅក្នុង neuroanatomy, horseradish peroxidase, ក៏ផ្លាស់ទីដោយការដឹកជញ្ជូន retrograde ។ ការដឹកជញ្ជូន Retrograde ប្រហែលជាដើរតួយ៉ាងសំខាន់ក្នុងការគ្រប់គ្រងការសំយោគប្រូតេអ៊ីននៅក្នុងរាងកាយកោសិកា។ ពីរបីថ្ងៃបន្ទាប់ពីការកាត់ axon, chromatolysis ត្រូវបានគេសង្កេតឃើញនៅក្នុងរាងកាយកោសិកាដែលបង្ហាញពីការរំខាននៃការសំយោគប្រូតេអ៊ីន។ ពេលវេលាដែលត្រូវការសម្រាប់ chromatolysis ទាក់ទងទៅនឹងរយៈពេលនៃការដឹកជញ្ជូន retrograde ពីទីតាំងនៃការផ្លាស់ប្តូរ axon ទៅរាងកាយកោសិកា។ លទ្ធផលនេះក៏បង្ហាញពីការពន្យល់សម្រាប់ជំងឺនេះផងដែរ - ការបញ្ជូនពីបរិមាត្រនៃ "សារធាតុសញ្ញា" ដែលគ្រប់គ្រងការសំយោគប្រូតេអ៊ីនត្រូវបានរំខាន។ វាច្បាស់ណាស់ថា "យានជំនិះ" សំខាន់ៗដែលប្រើសម្រាប់ axonal លឿន
អង្ករ។ ១.១៤.ការពិសោធន៍បង្ហាញពីការដឹកជញ្ជូនតាមអ័ក្សយ៉ាងលឿននៅក្នុងសរសៃវិញ្ញាណនៃសរសៃប្រសាទ sciatic របស់ឆ្មា។ Leucine ដែលមានស្លាក Tritium ត្រូវបានចាក់ចូលទៅក្នុង ganglion ឬស dorsal ហើយវិទ្យុសកម្មនៅក្នុង ganglion ហើយសរសៃវិញ្ញាណត្រូវបានវាស់នៅ 2, 4, 6, 8 និង 10 ម៉ោងបន្ទាប់ពីការចាក់។ (ខាងក្រោមរូបភាព)។ដោយ អ័ក្ស xចម្ងាយពី ganglion ទៅតំបន់នៃសរសៃប្រសាទ sciatic ដែលការវាស់វែងត្រូវបានធ្វើឡើងត្រូវបានគ្រោងទុក។ នៅលើអ័ក្សតម្រៀបសម្រាប់តែខ្សែកោងខាងលើ និងខាងក្រោមប៉ុណ្ណោះ វិទ្យុសកម្ម (ជីពចរ/នាទី) ត្រូវបានរៀបចំតាមមាត្រដ្ឋានលោការីត។ "រលក" នៃវិទ្យុសកម្មកើនឡើង (ព្រួញ)ផ្លាស់ទីក្នុងល្បឿន 410 មីលីម៉ែត្រ / ថ្ងៃ (តាមបណ្តោយ)
ការដឹកជញ្ជូនគឺ vesicles (vesicles) និង organelles,ដូចជា mitochondria ដែលមានសារធាតុដែលត្រូវដឹកជញ្ជូន។ ចលនានៃ vesicles ឬ mitochondria ធំបំផុតអាចត្រូវបានគេសង្កេតឃើញដោយប្រើមីក្រូទស្សន៍នៅក្នុង Vivo . ភាគល្អិតបែបនេះធ្វើចលនាខ្លី លឿនក្នុងទិសដៅមួយ ឈប់ ជារឿយៗផ្លាស់ទីថយក្រោយបន្តិច ឬទៅចំហៀង ឈប់ម្តងទៀត ហើយបន្ទាប់មកលោតក្នុងទិសដៅសំខាន់។ 410 មីលីម៉ែត្រ / ថ្ងៃត្រូវគ្នាទៅនឹងល្បឿនមធ្យមនៃចលនា anterograde ប្រហែល 5 μm / s; ដូច្នេះល្បឿននៃចលនានីមួយៗត្រូវតែខ្ពស់ជាង ហើយប្រសិនបើយើងគិតគូរពីទំហំនៃសរីរាង្គ សរសៃអំបោះ និង microtubules នោះចលនាទាំងនេះពិតជាលឿនណាស់។ ការដឹកជញ្ជូនតាមអ័ក្សលឿនតម្រូវឱ្យមានកំហាប់សំខាន់នៃ ATP ។ សារធាតុពុលដូចជា colchicine ដែលរំខានដល់ microtubules ក៏រារាំងការដឹកជញ្ជូនតាមអ័ក្សលឿនផងដែរ។ វាធ្វើតាមពីនេះថានៅក្នុងដំណើរការដឹកជញ្ជូនដែលយើងកំពុងពិចារណា vesicles និង organelles ផ្លាស់ទីតាមបណ្តោយ microtubules និង actin filaments; ចលនានេះត្រូវបានផ្តល់ដោយការប្រមូលផ្តុំតូចៗនៃម៉ូលេគុល dynein និង myosin ដែលដើរតួដូចបង្ហាញក្នុងរូប។ 1.13 ដោយប្រើថាមពល ATP ។
ការដឹកជញ្ជូន axon លឿនក៏អាចចូលរួមផងដែរ។ ដំណើរការរោគសាស្ត្រ។មេរោគ neurotropic មួយចំនួន (ឧទាហរណ៍ មេរោគ herpes ឬ polio virus) ចូលទៅក្នុង axon នៅតាមបរិវេណ និងផ្លាស់ទីតាមរយៈការដឹកជញ្ជូន retrograde ទៅកាន់រាងកាយ neuron ដែលពួកគេគុណ និងបញ្ចេញឥទ្ធិពលពុលរបស់វា។ ជាតិពុលតេតាណូស ដែលជាប្រូតេអ៊ីនដែលផលិតដោយបាក់តេរីដែលចូលទៅក្នុងខ្លួននៅពេលដែលស្បែកត្រូវបានខូចខាត ត្រូវបានចាប់យកដោយចុងសរសៃប្រសាទ និងដឹកជញ្ជូនទៅកាន់រាងកាយណឺរ៉ូន ដែលជាកន្លែងដែលវាបណ្តាលឱ្យមានការកន្ត្រាក់សាច់ដុំលក្ខណៈ។ មានករណីដែលគេដឹងថាមានឥទ្ធិពលពុលលើការដឹកជញ្ជូន axon ខ្លួនវាជាឧទាហរណ៍ ការប៉ះពាល់នឹងសារធាតុរំលាយអាគ្រីឡាមីដក្នុងឧស្សាហកម្ម។ លើសពីនេះទៀតវាត្រូវបានគេជឿថាការបង្ករោគនៃកង្វះវីតាមីន beriberi និង polyneuropathy គ្រឿងស្រវឹងពាក់ព័ន្ធនឹងការរំលោភលើការដឹកជញ្ជូន axonal លឿន។
បន្ថែមពីលើការដឹកជញ្ជូន axonal លឿននៅក្នុងកោសិកាក៏មានកម្លាំងខ្លាំងផងដែរ។ ការដឹកជញ្ជូន axon យឺត។ Tubulin ផ្លាស់ទីតាមអ័ក្សក្នុងល្បឿនប្រហែល 1 មម/ថ្ងៃ ហើយ actin ផ្លាស់ទីលឿនជាងមុន — រហូតដល់ 5 មម/ថ្ងៃ។ ប្រូតេអ៊ីនផ្សេងទៀតធ្វើចំណាកស្រុកជាមួយសមាសធាតុ cytoskeletal ទាំងនេះ; ឧទាហរណ៍ អង់ស៊ីមហាក់ដូចជាត្រូវបានផ្សារភ្ជាប់ជាមួយនឹង actin ឬ tubulin ។ អត្រានៃចលនារបស់ tubulin និង actin គឺប្រហាក់ប្រហែលនឹងអត្រាកំណើនដែលបានរកឃើញសម្រាប់យន្តការដែលបានពិពណ៌នាពីមុន ដែលម៉ូលេគុលត្រូវបានដាក់បញ្ចូលទៅក្នុងចុងសកម្មនៃ microtubule ឬ microfilament ។ ដូច្នេះយន្តការនេះអាចនឹងបង្ហាញពីការដឹកជញ្ជូនតាមអ័ក្សយឺត។ អត្រានៃការដឹកជញ្ជូនតាមអ័ក្សយឺតក៏ប្រហាក់ប្រហែលនឹងអត្រានៃការលូតលាស់ axon ដែលជាក់ស្តែងបង្ហាញពីការរឹតបន្តឹងដែលដាក់ដោយរចនាសម្ព័ន្ធនៃ cytoskeleton នៅលើដំណើរការទីពីរ។
សេចក្តីសន្និដ្ឋានផ្នែកនេះ វាគួរតែត្រូវបានសង្កត់ធ្ងន់ថាកោសិកាមិនមានរចនាសម្ព័ន្ធឋិតិវន្តទេ ដូចដែលវាលេចឡើង ឧទាហរណ៍នៅក្នុងរូបថតមីក្រូទស្សន៍អេឡិចត្រុង។ ភ្នាសប្លាស្មានិងជាពិសេស សរីរាង្គស្ថិតនៅក្នុងចលនាលឿនថេរនិងការរៀបចំឡើងវិញថេរ;នោះហើយជាមូលហេតុដែលពួកគេអាចដំណើរការបាន។ លើសពីនេះ ទាំងនេះមិនមែនជាអង្គជំនុំជម្រះសាមញ្ញ ដែលប្រតិកម្មគីមីកើតឡើងនោះទេ។ បណ្តុំនៃភ្នាស និងសរសៃដែលមានការរៀបចំខ្ពស់ដែលប្រតិកម្មកើតឡើងក្នុងលំដាប់ដែលរៀបចំយ៉ាងល្អប្រសើរ។
១.៤. បទប្បញ្ញត្តិនៃមុខងារកោសិកា
ការថែរក្សាកោសិកាបុគ្គលជាឯកតាមុខងារត្រូវបានគ្រប់គ្រងជាចម្បងដោយស្នូល; ការសិក្សាអំពីយន្តការនិយតកម្មបែបនេះគឺជាប្រធានបទនៃជីវវិទ្យាកោសិកា និងជីវគីមី។ ក្នុងពេលជាមួយគ្នានេះ កោសិកាត្រូវតែកែប្រែមុខងាររបស់ពួកគេស្របតាមលក្ខខណ្ឌបរិស្ថាន និងតម្រូវការរបស់កោសិកាផ្សេងទៀតនៃរាងកាយ ពោលគឺពួកវាបម្រើជាវត្ថុនៃបទបញ្ជាមុខងារ។ ខាងក្រោមនេះយើងនឹងពិចារណាដោយសង្ខេបអំពីរបៀបដែលឥទ្ធិពលបទប្បញ្ញត្តិទាំងនេះធ្វើសកម្មភាពលើភ្នាសប្លាស្មា និងរបៀបដែលពួកវាទៅដល់សរីរាង្គខាងក្នុង។
សក្តានុពលនៃភ្នាស . ក្នុងករណីជាច្រើនបទប្បញ្ញត្តិនៃមុខងារកោសិកាត្រូវបានអនុវត្តដោយការផ្លាស់ប្តូរសក្តានុពលភ្នាស។ ការផ្លាស់ប្តូរក្នុងស្រុកនៅក្នុងសក្តានុពលគឺអាចធ្វើទៅបាននៅពេលដែល: 1) ចរន្តពីតំបន់ជាប់គ្នានៃកោសិកាឬបង្កើតដោយកោសិកាមួយផ្សេងទៀតហូរតាមរយៈភ្នាស; 2) កំហាប់ extracellular នៃការផ្លាស់ប្តូរអ៊ីយ៉ុង (ជាញឹកញាប់ [K +]ចេញ ); 3) បណ្តាញអ៊ីយ៉ុងភ្នាសបើក។ ការផ្លាស់ប្តូរសក្ដានុពលនៃភ្នាសអាចប៉ះពាល់ដល់ការអនុលោមតាមប្រូតេអ៊ីនភ្នាស ដែលបណ្តាលឱ្យជាពិសេស បណ្តាញបើក ឬបិទ។ ដូចដែលបានពិពណ៌នាខាងលើ ដំណើរការនៃស្នប់ភ្នាសខ្លះអាស្រ័យលើសក្តានុពលនៃភ្នាស។ កោសិកាសរសៃប្រសាទមានឯកទេសក្នុងការយល់ឃើញការផ្លាស់ប្តូរសក្តានុពលភ្នាសជាព័ត៌មានដែលត្រូវតែដំណើរការ និងបញ្ជូន (សូមមើលជំពូកទី 2)។
សារធាតុនិយតកម្មក្រៅកោសិកា . យន្តការនិយតកម្មដ៏សំខាន់បំផុតដែលពាក់ព័ន្ធនឹងសារធាតុក្រៅកោសិកាគឺអន្តរកម្មរបស់ពួកគេជាមួយអ្នកទទួលជាក់លាក់នៅលើភ្នាសប្លាស្មា ឬនៅខាងក្នុងកោសិកា។ សារធាតុទាំងនេះរួមមាន synaptic transmitter ដែលបញ្ជូនព័ត៌មានរវាងកោសិកាប្រសាទ ភ្នាក់ងារក្នុងតំបន់ និងសារធាតុដែលចរាចរក្នុងឈាម និងទៅដល់កោសិកាទាំងអស់នៃរាងកាយ ដូចជាអរម៉ូន និងអង់ទីហ្សែន។ ឧបករណ៍បញ្ជូន Synapticគឺជាម៉ូលេគុលតូចៗដែលបញ្ចេញចេញពីចុងសរសៃប្រសាទនៅ synapse;
នៅពេលដែលពួកគេទៅដល់ភ្នាសប្លាស្មានៃកោសិកាជិតខាង កោសិកា postsynaptic ពួកវាបង្កឱ្យមានសញ្ញាអគ្គិសនី ឬយន្តការនិយតកម្មផ្សេងទៀត។ បញ្ហានេះត្រូវបានពិភាក្សាលម្អិតនៅក្នុងជំពូក។ ៣.
ភ្នាក់ងារគីមីក្នុងស្រុក ជារឿយៗត្រូវបានលាក់ដោយកោសិកាឯកទេស។ ពួកវាសាយភាយដោយសេរីនៅក្នុងចន្លោះក្រៅកោសិកា ប៉ុន្តែសកម្មភាពរបស់ពួកគេត្រូវបានកំណត់ចំពោះក្រុមតូចមួយនៃកោសិកាដោយសារតែការបំផ្លាញយ៉ាងឆាប់រហ័សនៃសារធាតុទាំងនេះ ទាំងដោយឯកឯង ឬក្រោមសកម្មភាពនៃអង់ស៊ីម។ ឧទាហរណ៍មួយនៃការបញ្ចេញភ្នាក់ងារបែបនេះគឺការចេញផ្សាយ អ៊ីស្តាមីនកោសិកា mast ដោយសារតែការខូចខាតឬការឆ្លើយតបនៃប្រព័ន្ធភាពស៊ាំ។ អ៊ីស្តាមីន បណ្តាលឱ្យមានការបន្ធូរបន្ថយកោសិកាសាច់ដុំរលោងនៃសរសៃឈាម បង្កើនភាពជ្រាបចូលនៃសរសៃឈាម endothelium និងរំញោចចុងសរសៃប្រសាទដែលសម្របសម្រួលអារម្មណ៍នៃការរមាស់។ ភ្នាក់ងារគីមីក្នុងស្រុកផ្សេងទៀតត្រូវបានបញ្ចេញដោយកោសិកាជាច្រើនទៀត។ ភ្នាក់ងារក្នុងស្រុកធម្មតាគឺ ថ្នាំ prostaglandins,បង្កើតជាក្រុមនៃដេរីវេនៃអាស៊ីតខ្លាញ់ប្រហែល 20 ។ ពួកវាត្រូវបានបញ្ចេញជាបន្តបន្ទាប់ពីកោសិកាដែលចែកចាយយ៉ាងទូលំទូលាយ ប៉ុន្តែធ្វើសកម្មភាពតែក្នុងមូលដ្ឋាន ព្រោះពួកវាត្រូវបានបំផ្លាញយ៉ាងឆាប់រហ័សដោយភ្នាស phospholipases ។ prostaglandins ជាច្រើនមានវិសាលគមធំទូលាយនៃសកម្មភាព: ពួកគេអាចបង្កឱ្យមានការកន្ត្រាក់នៃកោសិកាសាច់ដុំរលោងបណ្តាលឱ្យមានការប្រមូលផ្តុំប្លាកែតឈាម (ប្លាកែត) ឬទប់ស្កាត់ការវិវត្តនៃ corpus luteum នៅក្នុងអូវែ។
ភ្នាក់ងារក្នុងស្រុកផ្សេងទៀតបម្រើ កត្តាលូតលាស់។កត្តាលូតលាស់សរសៃប្រសាទ (NGF) សម្រាប់សរសៃប្រសាទដែលចេះអាណិតអាសូរ ដែលចាំបាច់សម្រាប់ការលូតលាស់ និងការរស់រានមានជីវិតនៃសរសៃប្រសាទទាំងនេះអំឡុងពេលអភិវឌ្ឍនៅក្នុង Vivo ឬនៅក្នុងវប្បធម៌កោសិកា។ ជាក់ស្តែង កោសិកាគោលដៅសម្រាប់ថ្នាក់នៃណឺរ៉ូននេះសម្ងាត់ NGF ហើយដោយហេតុនេះធានាឱ្យមានដំណើរការខាងក្នុងត្រឹមត្រូវ។ នៅពេលបង្កើតសរីរាង្គ កោសិកាជារឿយៗត្រូវ "ស្វែងរកផ្លូវរបស់ពួកគេ" ទៅកាន់កោសិកាគោលដៅ ដែលអាចស្ថិតនៅចម្ងាយដ៏ច្រើនសន្ធឹកសន្ធាប់។ ដូច្នោះហើយត្រូវតែមានកត្តាកំណើនឯកទេសជាច្រើនដែលស្រដៀងនឹង NGF ។
អរម៉ូននិងអង់ទីហ្សែន បញ្ជូនដោយឈាមទៅកាន់កោសិកាទាំងអស់។ អង់ទីហ្សែនបង្កឲ្យមានការឆ្លើយតបនៃប្រព័ន្ធភាពស៊ាំពីកោសិកាដែលមានអង្គបដិប្រាណជាក់លាក់។ ទោះជាយ៉ាងណាក៏ដោយ អង់ទីហ្សែន ជាក្បួនគឺជាសារធាតុបរទេសដែលមិនត្រូវបានផលិតនៅក្នុងសារពាង្គកាយដែលមានប្រតិកម្ម (សម្រាប់ព័ត៌មានលម្អិត សូមមើលជំពូកទី 18) ។ អ័រម៉ូនមួយចំនួនដូចជា អាំងស៊ុយលីន ឬ thyroxine ប៉ះពាល់ដល់ប្រភេទកោសិកាជាច្រើន ហើយអរម៉ូនផ្សេងទៀតដូចជា អរម៉ូនភេទ ប៉ះពាល់តែប្រភេទជាក់លាក់នៃកោសិកា។ អ័រម៉ូនគឺជា peptides ដែលសកម្មភាពរបស់វាត្រូវបានបង្កឡើងដោយការភ្ជាប់ទៅនឹង receptor នៅលើភ្នាសកោសិកា ឬ steroids និង thyroxine ដែលសាយភាយតាមរយៈភ្នាស lipid និងភ្ជាប់ទៅនឹងអ្នកទទួល intracellular ។ អរម៉ូនស្តេរ៉ូអ៊ីតភ្ជាប់ទៅនឹងក្រូម៉ាទីននុយក្លេអែរដែលបណ្តាលឱ្យមានការចម្លងនៃហ្សែនមួយចំនួន។ ប្រូតេអ៊ីនដែលជាលទ្ធផលបណ្តាលឱ្យមានការផ្លាស់ប្តូរមុខងារកោសិកាដែលជាឥទ្ធិពលជាក់លាក់នៃអរម៉ូន។ បញ្ហាដែលទាក់ទងនឹងការបញ្ចេញ និងសកម្មភាពរបស់អរម៉ូនត្រូវបានពិភាក្សាយ៉ាងលម្អិតនៅក្នុងជំពូក។ ១៧.
ការប្រាស្រ័យទាក់ទងក្នុងកោសិកាដែលពាក់ព័ន្ធនឹងអ្នកនាំសារទីពីរ
មុខងារនិយតកម្មដែលបានពិពណ៌នាខាងលើរួមមានឥទ្ធិពលលើភ្នាសកោសិកា។ ព័ត៌មានដែលទទួលបានដោយភ្នាសកោសិកាជារឿយៗត្រូវតែធ្វើឱ្យសរីរាង្គមានប្រតិកម្ម ហើយត្រូវបានផ្ទេរទៅឱ្យពួកវាដោយសារធាតុផ្សេងៗដែលគេស្គាល់ថាជាអ្នកនាំសារទីពីរ (ផ្ទុយពីអ្នកនាំសារទីមួយដែលចូលមកកោសិកាពីប្រភពខាងក្រៅ)។ ការសិក្សាអំពីអ្នកសម្របសម្រួលទីពីរកំពុងវិវឌ្ឍយ៉ាងឆាប់រហ័ស ហើយមិនមានការធានាថាកម្រិតនៃការយល់ដឹងអំពីបញ្ហាបច្ចុប្បន្ននឹងពេញលេញគ្រប់គ្រាន់នោះទេ។ នៅទីនេះយើងនឹងប៉ះអ្នកសម្របសម្រួលបីនាក់ដែលបានសិក្សាយ៉ាងល្អ: Ca 2+, cAMP និង inositol triphosphate ។
កាល់ស្យូម។អ្នកសម្រុះសម្រួលក្នុងកោសិកាដ៏សាមញ្ញបំផុតគឺ Ca 2+ ion ។ ការផ្តោតអារម្មណ៍ដោយឥតគិតថ្លៃរបស់វានៅក្នុងកោសិកាសម្រាកគឺទាបណាស់ ហើយបរិមាណដល់ 10_–8–10–7 mol/l ។ វាអាចចូលទៅក្នុងកោសិកាតាមរយៈបណ្តាញភ្នាសជាក់លាក់ នៅពេលដែលពួកវាស្ថិតនៅក្នុងស្ថានភាពបើកចំហ ជាឧទាហរណ៍ នៅពេលដែលសក្តានុពលភ្នាសផ្លាស់ប្តូរ (សូមមើលជំពូកទី 2)។ ការកើនឡើងជាលទ្ធផលនៃកំហាប់ Ca 2+ បង្កឱ្យមានប្រតិកម្មសំខាន់ៗនៅក្នុងកោសិកា ដូចជាការកន្ត្រាក់នៃ myofibrils ដែលជាមូលដ្ឋាននៃការកន្ត្រាក់សាច់ដុំ (សូមមើលជំពូកទី 4) ឬការបញ្ចេញ vesicles ដែលមានសារធាតុបញ្ជូនសរសៃប្រសាទពីចុងសរសៃប្រសាទ (សូមមើលជំពូកទី 3)។ . ប្រតិកម្មទាំងពីរទាមទារកំហាប់ Ca 2+ ប្រហែល 10-5 mol/L ។ Ca 2+ ដែលមានឥទ្ធិពលនិយតកម្ម ក៏អាចត្រូវបានបញ្ចេញចេញពីហាង intracellular ដូចជា endoplasmic reticulum ផងដែរ។ ការចេញផ្សាយ Ca 2+ ពីដេប៉ូទាមទារឱ្យមានការចូលរួមពីអន្តរការីផ្សេងទៀត (សូមមើលឧទាហរណ៍រូបភាព 1.16)។
ស៊ីក្លូ adenosine monophosphate, cAMP ។ ថ្មីៗនេះ វាត្រូវបានបង្ហាញថា cyclic adenosine monophosphate (cAMP) ដែលជាដេរីវេនៃប្រភពថាមពលសំខាន់របស់រាងកាយ ATP គឺជាអ្នកនាំសារទីពីរដ៏សំខាន់។ ខ្សែសង្វាក់ប្រតិកម្មស្មុគស្មាញដែលបង្ហាញក្នុងរូប។ 1.15 ចាប់ផ្តើមជាមួយនឹងការទទួល R s នៅលើផ្ទៃខាងក្រៅនៃភ្នាសប្លាស្មា ដែលអាចបម្រើជាកន្លែងភ្ជាប់ជាក់លាក់សម្រាប់អ្នកសម្រុះសម្រួល និងអរម៉ូនផ្សេងៗ។ បន្ទាប់ពីការភ្ជាប់ទៅនឹងម៉ូលេគុល "រំញោច" ជាក់លាក់មួយ។ R s ផ្លាស់ប្តូរការអនុលោមតាមរបស់វា; ការផ្លាស់ប្តូរទាំងនេះប៉ះពាល់ដល់ប្រូតេអ៊ីន G s នៅលើផ្ទៃខាងក្នុងនៃភ្នាសតាមរបៀបដែលការធ្វើឱ្យសកម្មនៃសារធាតុក្រោយដោយ guanosine triphosphate (GTP) ចូលក្នុងកោសិកាអាចធ្វើទៅបាន។ ប្រូតេអ៊ីនធ្វើឱ្យសកម្ម G s នៅក្នុងវេនជំរុញអង់ស៊ីមនៅលើផ្ទៃខាងក្នុងនៃភ្នាស adenylate cyclase (AC) ដែលជំរុញការបង្កើត cAMP ពី ATP ។ cAMP រលាយក្នុងទឹក គឺជាអ្នកសម្រុះសម្រួលដែលបញ្ជូនឥទ្ធិពល

អង្ករ។ ១.១៥.ខ្សែសង្វាក់នៃប្រតិកម្មពាក់ព័ន្ធនឹង cAMP (cyclic adenosine monophosphate) ។ សញ្ញាខាងក្រៅដែលគួរឱ្យរំភើបឬរារាំងធ្វើឱ្យអ្នកទទួលភ្នាសសកម្ម R s ឬ Ri . អ្នកទទួលទាំងនេះគ្រប់គ្រងដំណើរការចងជី - ប្រូតេអ៊ីនដែលមាន GTP intracellular (guanosine triphosphate) ដោយហេតុនេះរំញោចឬរារាំង intracellular adenylate cyclase (AC) ។ អង់ស៊ីម amplifying AC បំប្លែង adenosine triphosphate (ATP) ទៅជា cAMP ដែលបន្ទាប់មកត្រូវបានបំបែកទៅជា AMP ដោយ phosphodiesterase ។ cAMP ឥតគិតថ្លៃសាយភាយចូលទៅក្នុងកោសិកា និងធ្វើឱ្យសកម្ម adenylate kinase (A-kinase) បញ្ចេញនូវអនុunit C របស់វា ដែលជំរុញឱ្យមាន phosphorylation នៃប្រូតេអ៊ីន intracellular ពោលគឺឧ។ បង្កើតជាឥទ្ធិពលចុងក្រោយនៃការរំញោចក្រៅកោសិកា។ ដ្យាក្រាមក៏បង្ហាញពីឱសថសាស្ត្រ និងសារធាតុពុលដែលបង្កឱ្យមាន (+) ឬរារាំង (-) ប្រតិកម្មមួយចំនួន (កែប្រែ)
ការរំញោចអ្នកទទួលខាងក្រៅ R s ទៅរចនាសម្ព័ន្ធខាងក្នុងនៃកោសិកា។
ស្របជាមួយនឹងខ្សែសង្វាក់រំញោចនៃប្រតិកម្មដែលពាក់ព័ន្ធ R s ការផ្សារភ្ជាប់អ្នកសម្របសម្រួល inhibitory និងអរម៉ូនទៅនឹងអ្នកទទួលដែលត្រូវគ្នាគឺអាចធ្វើទៅបាន R i ដែលម្តងទៀតតាមរយៈប្រូតេអ៊ីន GTP-ធ្វើឱ្យសកម្មជី រារាំង AC ហើយដូច្នេះការផលិត cAMP ។ ការសាយភាយចូលទៅក្នុងកោសិកា cAMP មានប្រតិកម្មជាមួយ adenylate kinase (A-kinase); នេះចេញផ្សាយរង C ដែល ជំរុញ phosphorylation នៃប្រូតេអ៊ីន P. phosphorylation នេះបំប្លែងប្រូតេអ៊ីនទៅជាទម្រង់សកម្ម ហើយឥឡូវនេះពួកគេអាចបញ្ចេញឥទ្ធិពលបទប្បញ្ញត្តិជាក់លាក់របស់ពួកគេ (ឧទាហរណ៍ បណ្តាលឱ្យមានការរិចរិល glycogen)។ ប្រព័ន្ធនិយតកម្មដ៏ស្មុគស្មាញនេះមានប្រសិទ្ធភាពខ្លាំង ពីព្រោះលទ្ធផលចុងក្រោយគឺ phosphorylation នៃប្រូតេអ៊ីនជាច្រើន ពោលគឺសញ្ញានិយតកម្មធ្វើដំណើរឆ្លងកាត់សៀគ្វីជាមួយនឹងការកើនឡើងខ្ពស់។ អ្នកសម្របសម្រួលខាងក្រៅដែលភ្ជាប់ទៅនឹងអ្នកទទួល R s និង R i ជាក់លាក់ចំពោះពួកគេនីមួយៗ ចម្រុះណាស់។ Adrenaline, ភ្ជាប់ជាមួយ R s ឬ R i ចូលរួមក្នុងបទប្បញ្ញត្តិនៃការរំលាយអាហារ lipid និង glycogen ក៏ដូចជាក្នុងការបង្កើនការកន្ត្រាក់នៃសាច់ដុំបេះដូងនិងប្រតិកម្មផ្សេងទៀត (សូមមើលជំពូកទី 19) ។ អរម៉ូនរំញោចក្រពេញទីរ៉ូអ៊ីត, កំពុងធ្វើឱ្យសកម្ម R s ជំរុញការបញ្ចេញអរម៉ូន thyroxine ដោយក្រពេញទីរ៉ូអ៊ីត ហើយ prostaglandin I រារាំងការប្រមូលផ្តុំប្លាកែតក្នុងឈាម។ ឥទ្ធិពលរារាំង រួមទាំង adrenaline សម្របសម្រួលតាមរយៈ R i ត្រូវបានបង្ហាញក្នុងការថយចុះនៃ lipolysis ។ ដូច្នេះ ប្រព័ន្ធ cAMP គឺជាប្រព័ន្ធគ្រប់គ្រងកោសិកាខាងក្នុងពហុមុខងារ។ដែលអាចត្រូវបានគ្រប់គ្រងយ៉ាងជាក់លាក់ដោយសារធាតុ stimulatory extracellular និង inhibitory signaling ។
Inositol phosphate "IF" h " ប្រព័ន្ធ intracellular នៃអ្នកនាំសារទីពីរគឺ inositol phosphate ត្រូវបានគេរកឃើញថ្មីៗនេះ (រូបភាព 1.16) ។ ក្នុងករណីនេះមិនមានផ្លូវរារាំងទេប៉ុន្តែមានភាពស្រដៀងគ្នាទៅនឹងប្រព័ន្ធ cAMP ដែលឥទ្ធិពលនៃការរំញោចនៃអ្នកទទួល R ត្រូវបានផ្ទេរទៅប្រូតេអ៊ីន GTP-activated G នៅលើផ្ទៃខាងក្នុងនៃភ្នាស។ នៅដំណាក់កាលបន្ទាប់ភ្នាសរំអិលធម្មតា phosphatidylinositol (PI) ដែលពីមុនបានទទួលក្រុមផូស្វាតពីរបន្ថែមទៀតត្រូវបានបំប្លែងទៅជា PI-diphosphate (PIP 2) ដែលត្រូវបានបំបែកដោយ phosphodiesterase (PDE) សកម្មចូលទៅក្នុង inositol triphosphate(IFz) និង lipid diacylglycerol(DAG) ។ Inositol triphosphate គឺជាអ្នកនាំសារទីពីររលាយក្នុងទឹក ដែលសាយភាយចូលទៅក្នុង cytosol ។ វាដើរតួជាចម្បងដោយការបញ្ចេញ Ca 2+ ពីកោសិកា endoplasmic reticulum ។ Ca 2+ នៅក្នុងវេនដើរតួជាអ្នកសម្រុះសម្រួល ដូចដែលបានរៀបរាប់ខាងលើ។ ឧទាហរណ៍ វាធ្វើឱ្យសកម្ម Ca 2+ -dependent phosphokinase ដែល phosphorylates អង់ស៊ីម។ ផ្នែករង lipid នៃ DAG (រូបភាព 1.16) ក៏ផ្ទុកសញ្ញាផងដែរ ដោយបានសាយភាយក្នុងដំណាក់កាល lipid នៃភ្នាសប្លាស្មាទៅ C-kinase ដែលមានទីតាំងនៅលើផ្ទៃខាងក្នុងរបស់វា ដែលត្រូវបានធ្វើឱ្យសកម្មដោយមានការចូលរួមពី phosphatidylserine ជា cofactor ។ បន្ទាប់មក C-kinase បង្កឱ្យមាន phosphorylation នៃប្រូតេអ៊ីនដោយបំប្លែងពួកវាទៅជាទម្រង់សកម្ម។
ប្រព័ន្ធផ្ញើសារទីពីរ intracellular IF 3 ក៏អាចត្រូវបានគ្រប់គ្រងដោយភាពខុសគ្នានៃអ្នកសម្រុះសម្រួលខាងក្រៅ និងអរម៉ូន រួមទាំង acetylcholine, serotonin, vasopressin និងអរម៉ូនរំញោចក្រពេញទីរ៉ូអ៊ីត។ ដូចជាប្រព័ន្ធ cAMP វាត្រូវបានកំណត់ដោយភាពខុសគ្នានៃឥទ្ធិពល intracellular ។ វាអាចទៅរួចដែលថាប្រព័ន្ធនេះក៏ត្រូវបានធ្វើឱ្យសកម្មដោយពន្លឺនៅក្នុងឧបករណ៍ទទួលដែលមើលឃើញនៃភ្នែក និងដើរតួនាទីសំខាន់ក្នុងការបញ្ជូនពន្លឺ (សូមមើលជំពូកទី 11) ។ ជាលើកដំបូងនៅក្នុងការអភិវឌ្ឍបុគ្គលនៃសារពាង្គកាយអ្នកទទួលនៃប្រព័ន្ធ IG ត្រូវបានធ្វើឱ្យសកម្មដោយមេជីវិតឈ្មោលដែលជាលទ្ធផលដែល IG ចូលរួមក្នុងប្រតិកម្មបទប្បញ្ញត្តិដែលអមនឹងការបង្កកំណើតនៃស៊ុត។
ប្រព័ន្ធ cAMP និង IFz-DAG មានប្រសិទ្ធភាពខ្ពស់។ អ្នកបង្កើនជីវសាស្រ្ត។ពួកគេ

អង្ករ។ ១.១៦.ខ្សែសង្វាក់នៃប្រតិកម្មដែលពាក់ព័ន្ធនឹងអ្នកសម្រុះសម្រួលខាងក្នុងកោសិកា IPG (inositol triphosphate) ។ ដូចនៅក្នុងប្រព័ន្ធ cAMP សញ្ញា extracellular ត្រូវបានសម្របសម្រួលតាមរយៈប្រូតេអ៊ីនជី, ដែលក្នុងករណីនេះធ្វើឱ្យ phosphodiesterase (PDE) សកម្ម។ អង់ស៊ីមនេះបំបែក phosphatidylinosine diphosphate (PIP 2 ) នៅក្នុងភ្នាសប្លាស្មាមុន IF h និង diacylglycerol (DAG); IF h សាយភាយចូលទៅក្នុង cytoplasm ។ នៅទីនេះវាបណ្តាលឱ្យមានការចេញផ្សាយ Ca 2+ ពី reticulum endoplasmic; ការកើនឡើងកំហាប់ Ca 2+ នៅក្នុង cytoplasm ([Ca 2+ ]i ) ធ្វើឱ្យប្រូតេអ៊ីន kinase សកម្មដែល phosphorylates ហើយដូច្នេះធ្វើឱ្យអង់ស៊ីមសកម្ម។ ផលិតផលមួយផ្សេងទៀត DAG នៅតែមាននៅក្នុងភ្នាសនិងធ្វើឱ្យសកម្មប្រូតេអ៊ីន kinase C (phosphatidylserine cofactor, PS) ។ ប្រូតេអ៊ីន kinase C ក៏ phosphorylates អង់ស៊ីមដែលសម្របសម្រួលសកម្មភាពជាក់លាក់ដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការរំញោចខាងក្រៅ។រ . សាខានៃខ្សែសង្វាក់ប្រតិកម្មដែលពាក់ព័ន្ធនឹង IF h និង DAG អាចត្រូវបានធ្វើឱ្យសកម្មដោយឯករាជ្យដោយ ionomycin និង phorbol ester រៀងគ្នា (កែប្រែ)
បំប្លែងប្រតិកម្មរវាងឧបករណ៍បញ្ជូន និងឧបករណ៍ទទួលភ្នាសខាងក្រៅទៅជា phosphorylation នៃពពួកប្រូតេអ៊ីនខាងក្នុងកោសិកា ដែលបន្ទាប់មកអាចមានឥទ្ធិពលលើមុខងារកោសិកាផ្សេងៗ។ ទិដ្ឋភាពសំខាន់មួយនៃបញ្ហាគឺថា ដូចដែលបានដឹងនាពេលបច្ចុប្បន្ននេះ មានតែប្រព័ន្ធនិយតកម្មដែលទាក់ទងគ្នាយ៉ាងជិតស្និទ្ធទាំងពីរនៃប្រភេទនេះ ដែលត្រូវបានប្រើប្រាស់ដោយអ្នកសម្រុះសម្រួលខាងក្រៅជាច្រើន ដើម្បីគ្រប់គ្រងដំណើរការផ្សេងៗនៃកោសិកា។ ទន្ទឹមនឹងនេះ ប្រព័ន្ធនិយតកម្មទាំងនេះ រួមទាំង Ca 2+ មានទំនាក់ទំនងយ៉ាងជិតស្និទ្ធជាមួយគ្នាទៅវិញទៅមក ដែលអនុញ្ញាតឱ្យពួកគេអនុវត្តបទប្បញ្ញត្តិដ៏ល្អនៃមុខងារកោសិកា។
ការបង្រៀន និងការណែនាំ
1. អាល់ប៊ើត IN., Bray D., Lewis J., Raff M., Roberts TO, Watson J.D.ជីវវិទ្យាម៉ូលេគុលនៃកោសិកា, ញូវយ៉ក និងឡុងដ៍, Garland Publishing Inc., 1983 ។
2. Czihak G., Longer H., Ziegler H.(eds ។ ) ជីវវិទ្យា។ ទីក្រុងប៊ែកឡាំង ទីក្រុង Heidelberg ទីក្រុង New York ទីក្រុង Springer ឆ្នាំ 1983 ។
3. ហ៊ីល។ IN. ឆានែលអ៊ីយ៉ុងនៃភ្នាសគួរឱ្យរំភើប។ Sunderland, Mass., Sinauer Assoc., 1984 ។
4. Hoppe W., Lohmann W.. Marki H., Ziegler H.(eds ។ ) ជីវរូបវិទ្យា។ ទីក្រុងប៊ែកឡាំង, ហាយឌែលប៊ឺក, ញូវយ៉ក, ស្ព្រីងហ្គឺ, ឆ្នាំ ១៩៨៤។
5. Jungermann TO, Mahler H.ជីវគីមី។ ទីក្រុងប៊ែកឡាំង ទីក្រុង Heidelberg ទីក្រុង New York ទីក្រុង Springer ឆ្នាំ 1980 ។
6. Kandel E.R., Schwartz–J.H.,(eds ។ ) គោលការណ៍នៃវិទ្យាសាស្ត្រសរសៃប្រសាទ, ញូវយ៉ក, ទីក្រុង Amsterdam, Oxford, Elsevier, 1985 ។
7. Schiebler ធ. H., Schmidt W.កាយវិភាគសាស្ត្រ des Menschen ។ ទីក្រុងប៊ែកឡាំង, ហាយឌែលប៊ឺក, ញូវយ៉ក, តូក្យូ, ស្ព្រីងហ្គឺ, ឆ្នាំ ១៩៨៣។
អត្ថបទដើម និងការវាយតម្លៃ
8. Berridge M.J.មូលដ្ឋានម៉ូលេគុលនៃការទំនាក់ទំនងនៅក្នុងកោសិកា, Sci ។ Amer, 253, 124-134 (1985) ។
9. Berridge M.J., Irvine R.F. Inositol triphosphate ដែលជាអ្នកនាំសារទីពីរប្រលោមលោកក្នុងការបញ្ជូនសញ្ញាកោសិកា។ ធម្មជាតិ, 312, 315 321 (1984) ។
10. Bretscher M.S.ម៉ូលេគុលនៃភ្នាសកោសិកា Sci ។ Amer., 253, 124–134 (1985)។
11. Daut J.កោសិការស់ជាម៉ាស៊ីនបំលែងថាមពល។ គំរូតិចតួចនៃការរំលាយអាហារ myocardial, Biochem ។ និង Biophys ។ Acta, 895, 41–62 (1987) ។
12. Hodgkin A.L., Katz IN. ឥទ្ធិពលនៃអ៊ីយ៉ុងសូដ្យូមលើសកម្មភាពអគ្គិសនីនៃ axon យក្សនៃមឹក។ J. Physiol ។ ( ឡង។ ), 108, 37–77 (1949) ។
13. Hodgkin A.L., Keynes R.D.ការដឹកជញ្ជូនសកម្មនៃ cations នៅក្នុង axons យក្សពី ស៊ីភីយ៉ានិង ឡូលីហ្គោ J. Physiol ។ ( ឡង។ ), 128, 28–42 (1955) ។
14. យូរជាង P.ឆានែលអ៊ីយ៉ុងដែលមានអនុរដ្ឋអនុលោមភាព, ជីវវិទ្យា។ J., 47, 581–590 (1985) ។
15. Ochs S., Worth P.M.ការដឹកជញ្ជូន Exoplasmic នៅក្នុងប្រព័ន្ធធម្មតានិងរោគសាស្ត្រ។ នៅក្នុង: សរីរវិទ្យានិងរោគវិទ្យានៃ Axons, S.G. Waxmannអេដ។ ញូវយ៉ក, សារព័ត៌មាន Raven, 1978 ។
សេចក្តីសង្ខេបនៃបទបង្ហាញផ្សេងៗ"វិធីសាស្រ្តនៃការបង្រៀនជីវវិទ្យា" - សត្វវិទ្យាសាលា។ ណែនាំសិស្សឱ្យប្រើប្រាស់ទិន្នន័យសត្វវិទ្យាវិទ្យាសាស្ត្រ។ ការអប់រំសីលធម៌។ ពរជ័យបន្ថែមនៃទ្រុងមាន់។ ការជ្រើសរើសវិធីសាស្រ្ត។ ដំណើរការជីវិត។ ត្រីអាងចិញ្ចឹមត្រី។ អាហារូបត្ថម្ភ។ ការអប់រំបរិស្ថាន។ ខ្លឹមសារនៃដំណើរការជីវិត។ លទ្ធផលអវិជ្ជមាន។ សិស្សយកចិត្តទុកដាក់។ ទម្រង់ចាំបាច់។ មើលសត្វតូចៗ។ គោលបំណងនិងគោលបំណងនៃជីវវិទ្យា។ រឿង។
"ការរៀនផ្អែកលើបញ្ហាក្នុងមេរៀនជីវវិទ្យា" - ចំណេះដឹង។ សៀវភៅសិក្សាថ្មី។ ផ្លូវទៅកាន់ដំណោះស្រាយ។ បញ្ហា។ សិក្ខាសាលា។ តើអ្វីជាភារកិច្ច? លោក Albrecht Durer ។ ការរៀនផ្អែកលើបញ្ហាក្នុងមេរៀនជីវវិទ្យា។ មេរៀនមិនស្តង់ដារ។ អ្វីដែលមានន័យដោយការរៀនផ្អែកលើបញ្ហា។ គុណភាពនៃជីវិត។ ជីវវិទ្យាជាមុខវិជ្ជាសិក្សា។ សំណួរ។ មេរៀនដោះស្រាយបញ្ហា។ ការថយចុះចំណាប់អារម្មណ៍លើប្រធានបទ។ ថ្នាក់មន្ទីរពិសោធន៍ផ្អែកលើបញ្ហា។
"ការត្រិះរិះពិចារណាក្នុងមេរៀនជីវវិទ្យា" - បច្ចេកវិទ្យានៃ "ការគិតប្រកបដោយការរិះគន់" ។ ការប្រើប្រាស់បច្ចេកវិទ្យានៃ "ការអភិវឌ្ឍនៃការគិតរិះគន់" ។ តារាងសម្រាប់មេរៀន។ ការលើកទឹកចិត្តក្នុងការសិក្សា។ ប្រព័ន្ធអេកូឡូស៊ី។ អត្ថន័យនៃ "ការអភិវឌ្ឍការគិតពិចារណា" ។ សញ្ញានៃបច្ចេកវិទ្យា។ បច្ចេកវិទ្យា RKM ។ រចនាសម្ព័ន្ធមេរៀន។ ទិសដៅសំខាន់ៗ។ ប្រវត្តិនៃបច្ចេកវិទ្យា។ បច្ចេកវិទ្យាគរុកោសល្យ។ ច្បាប់នៃបច្ចេកវិទ្យា។ កិច្ចការជីវវិទ្យា។ រស្មីសំយោគ។ បច្ចេកទេសដែលបានប្រើនៅដំណាក់កាលផ្សេងៗគ្នានៃមេរៀន។
"មេរៀនជីវវិទ្យាជាមួយក្តារខៀនអន្តរកម្ម" - សៀវភៅសិក្សាអេឡិចត្រូនិច។ អត្ថប្រយោជន៍សម្រាប់សិស្ស។ ក្តារខៀនអន្តរកម្មជួយបញ្ជូនព័ត៌មានដល់សិស្សគ្រប់រូប។ កិច្ចការ Didactic ។ ការដោះស្រាយបញ្ហាជីវសាស្រ្ត។ គុណសម្បត្តិនៃការធ្វើការជាមួយក្តារខៀនអន្តរកម្ម។ ធ្វើការជាមួយបទបង្ហាញ។ ធ្វើការលើការប្រៀបធៀបវត្ថុ។ វត្ថុផ្លាស់ទី។ ការប្រើប្រាស់សៀវភៅបញ្ជី។ ការប្រើក្តារខៀនអន្តរកម្មក្នុងដំណើរការបង្រៀនសិស្សសាលា។ អត្ថប្រយោជន៍សម្រាប់គ្រូបង្រៀន។
"វិធីសាស្រ្តសកម្មភាពប្រព័ន្ធក្នុងជីវវិទ្យា" - សំណួរសិក្ខាសាលា។ វិធីសាស្រ្តសកម្មភាព។ Dryopithecus ។ ផ្លូវក្រៅភពនៃប្រភពដើមរបស់មនុស្ស។ លីសូសូម។ អង្គការគីមី។ កាយសម្ព័ន្ធ។ មេតាបូលីស។ អ្នកវិភាគ។ វិធីសាស្រ្តប្រព័ន្ធ-សកម្មភាពក្នុងការបង្រៀនជីវវិទ្យា។ ក្រូម៉ូសូម។ ស៊ីតូប្លាស្មា។ ពិការភ្នែក។ ប្រវែងត្រចៀក។ ចំណាត់ថ្នាក់នៃបុគ្គល។ គ្រោងឆ្អឹងនៃថនិកសត្វ។ ផ្លូវនៃការវិវត្តន៍របស់មនុស្ស។ មីតូស៊ីស។ ស្មុគ្រស្មាញ។ សំណួរដែលមានបញ្ហា។ នុយក្លេអូលូស។ ស្រោមសំបុត្រនុយក្លេអ៊ែរ។
"កុំព្យូទ័រក្នុងជីវវិទ្យា" - សកម្មភាពសហការរបស់សិស្ស។ គ្រួសារ Angiosperm ។ ការបណ្តុះបណ្តាលអន្តរកម្ម។ គំរូនៃការរៀនសូត្រ។ ឧទាហរណ៍នៃប្រព័ន្ធវាយតម្លៃ។ សំណួរកាតណែនាំ។ ឧទាហរណ៍នៃកាតណែនាំ។ អ្នកស្រាវជ្រាវ។ ក្រុមមីក្រូ។ បច្ចេកវិទ្យាសិក្សាអន្តរកម្ម។ វង់។ បច្ចេកវិទ្យាសិក្សាអន្តរកម្ម។ វិធីសាស្រ្តអន្តរកម្មក្នុងមេរៀនជីវវិទ្យា។ ទម្រង់ការងារជាក្រុម។ ភារកិច្ចសម្រាប់ក្រុម "អ្នកស្រាវជ្រាវ" ។
