Sanaysay
sa paksa: "Pagpaparami"
Panimula 3
1. Mga uri ng pagpaparami 4
1.1 Asexual reproduction 4
1.2 Sekswal na pagpaparami 6
2. Indibidwal na pag-unlad ng mga organismo 10
2.1 Embryonic na panahon ng pag-unlad 10
2.2 Postembryonic na panahon ng pag-unlad 13
2.3 Pangkalahatang mga pattern ng pag-unlad. Biogenetic Law 15
Konklusyon 18
Mga Sanggunian 18
Panimula
Ang kakayahang magparami, i.e. ang paggawa ng bagong henerasyon ng mga indibidwal ng parehong species ay isa sa mga pangunahing katangian ng mga buhay na organismo. Sa panahon ng proseso ng pagpaparami, ang genetic na materyal ay inililipat mula sa magulang na henerasyon hanggang sa susunod na henerasyon, na nagsisiguro sa pagpaparami ng mga katangian hindi lamang ng isang partikular na species, ngunit ng mga partikular na indibidwal na magulang. Para sa isang species, ang kahulugan ng pagpaparami ay upang palitan ang mga kinatawan nito na namatay, na nagsisiguro sa pagpapatuloy ng pagkakaroon ng species; bilang karagdagan, sa ilalim ng angkop na mga kondisyon, ang pagpaparami ay ginagawang posible upang madagdagan ang kabuuang bilang ng mga species.
Ang bawat bagong indibidwal, bago maabot ang yugto kung saan ito ay may kakayahang magparami, ay kailangang dumaan sa ilang yugto ng paglaki at pag-unlad. Ang ilang mga indibidwal ay namamatay bago umabot sa yugto ng reproduktibo (o sekswal na kapanahunan) bilang resulta ng pagkasira ng mga mandaragit, mga sakit at iba't ibang mga random na kaganapan; samakatuwid, ang mga species ay maaaring mabuhay lamang sa kondisyon na ang bawat henerasyon ay gumagawa ng mas maraming supling kaysa sa mga magulang na indibidwal na nakibahagi sa pagpaparami. Ang laki ng populasyon ay nagbabago depende sa balanse sa pagitan ng pagpaparami at pagkalipol ng mga indibidwal. Mayroong ilang iba't ibang mga diskarte sa pagpapalaganap, bawat isa ay may natatanging mga pakinabang at disadvantages; lahat ng mga ito ay ilalarawan sa abstract na ito.
1. Mga uri ng pagpaparami
Alam ang iba't ibang anyo ng pagpaparami, ngunit lahat sila ay maaaring pagsamahin sa dalawang uri: sekswal at asexual.
Ang sekswal na pagpaparami ay tumutukoy sa pagbabago ng mga henerasyon at pag-unlad ng mga organismo mula sa mga espesyal na selula ng kasarian na nabuo sa mga gonad. Sa kasong ito, ang isang bagong organismo ay bubuo bilang resulta ng pagsasanib ng dalawang selulang mikrobyo na nabuo ng magkaibang mga magulang. Gayunpaman, sa mga invertebrate na hayop, ang tamud at itlog ay madalas na nabuo sa katawan ng isang organismo. Ang phenomenon na ito, bisexuality, ay tinatawag na hermaphroditism. Ang mga namumulaklak na halaman ay bisexual din. Sa karamihan ng mga species ng angiosperms (namumulaklak) na mga halaman, ang isang bisexual na bulaklak ay kinabibilangan ng parehong mga stamen, na bumubuo ng mga male sex cell (sperm cell), at pistil, na naglalaman ng mga itlog. Sa humigit-kumulang isang-kapat ng mga species, ang lalaki (staminate) at babae (pistilate) na mga bulaklak ay bubuo nang nakapag-iisa, i.e. unisexual ang mga bulaklak nila. Ang isang halimbawa ay abaka. Sa ilang mga halaman (mais, birch), ang parehong lalaki at babaeng bulaklak ay lumilitaw sa parehong indibidwal.
Ang ilang uri ng hayop at halaman ay bubuo
hindi fertilized na itlog. Ang ganitong uri ng pagpaparami ay tinatawag na birhen o parthenogenetic.
Ang asexual reproduction ay nailalarawan sa katotohanan na ang isang bagong indibidwal ay bubuo mula sa mga non-sexual, somatic (katawan) na mga selula.
1.1 Asexual reproduction
Sa asexual reproduction, ang isang bagong organismo ay maaaring lumabas mula sa isang cell o mula sa ilang asexual (somatic) cells ng ina. Ang asexual reproduction ay nagsasangkot lamang ng isang magulang. Dahil ang mga selula na nagmumula sa mga organismo ng anak na babae ay lumitaw bilang isang resulta ng mitosis, lahat ng mga inapo ay magiging katulad sa namamana na mga katangian sa indibidwal na ina.
kanin. 1. Pagpaparami ng berdeng euglena
Maraming protozoa (amoeba, green euglena, atbp.), unicellular algae (Chlamydomonas) ang nagpaparami sa pamamagitan ng mitotic cell division (Fig. 1). Iba pang unicellular ilang mas mababang fungi, algae (chlorella), mga hayop, halimbawa ang causative agent ng malaria malarial plasmodium, ay nailalarawan sa pamamagitan ng sporulation. Sa kasong ito, ang cell ay nahahati sa isang malaking bilang ng mga indibidwal, katumbas ng bilang ng nuclei na dating nabuo sa parent cell bilang resulta ng paulit-ulit na paghahati ng nucleus nito. Ang mga multicellular na organismo ay may kakayahang sporulation: ito ay mga mosses, mas mataas na fungi, multicellular algae, pteridophytes at ilang iba pa.
Sa parehong unicellular at multicellular na organismo, ang budding ay isa ring paraan ng asexual reproduction. Halimbawa, sa yeast fungi at ilang ciliates (pagsuso ng ciliates), kapag namumuko sa mother cell, isang maliit na tubercle na naglalaman ng nucleus, isang bud, ay unang nabuo. Lumalaki ito, umabot sa sukat na malapit sa katawan ng ina, at pagkatapos ay naghihiwalay, nagpapatuloy sa malayang pag-iral. Sa mga multicellular na organismo (freshwater hydra), ang bato ay binubuo ng isang pangkat ng mga selula mula sa magkabilang patong ng dingding ng katawan. Ang usbong ay lumalaki, humahaba, at may lalabas na bukana ng bibig sa anterior dulo nito, na napapalibutan ng mga galamay. Ang budding ay nagtatapos sa pagbuo ng isang maliit na hydra, na pagkatapos ay naghihiwalay mula sa ina na organismo.
Sa mga multicellular na hayop, ang asexual reproduction ay nangyayari sa parehong paraan (jellyfish, annelids, flatworms, echinoderms). Mula sa bawat bahaging iyon ay bubuo ang isang ganap na indibidwal.
Ang vegetative propagation ay laganap sa mga halaman, i.e. mga bahagi ng katawan pinagputulan, tendrils, tubers. Kaya, ang mga patatas ay nagpaparami sa pamamagitan ng binagong mga bahagi sa ilalim ng lupa ng stem - tubers. Madaling nag-ugat ang mga sanga at pinagputulan ng jasmine at willow. Ang mga ubas, currant, at gooseberries ay pinalaganap gamit ang mga pinagputulan.
Ang mahahabang gumagapang na mga tangkay ng mga tendril ng strawberry ay bumubuo ng mga putot, na, na nag-ugat, ay nagbubunga ng isang bagong halaman. Ilang mga halaman, tulad ng begonia, ang maaaring palaganapin ng mga pinagputulan ng dahon (talim ng dahon at tangkay). Sa ilalim ng dahon, sa mga lugar kung saan ang mga malalaking ugat ay nagsasanga, lumilitaw ang mga ugat, sa itaas na bahagi ay may mga putot, at pagkatapos ay mga shoots.
Ginagamit din ang ugat para sa vegetative propagation. Sa paghahardin, ang mga raspberry, seresa, plum, at rosas ay pinalaganap gamit ang mga pinagputulan mula sa mga lateral na ugat. Ang Dahlias ay nagpaparami gamit ang root tubers. Ang pagbabago ng underground na bahagi ng stem rhizome ay bumubuo rin ng mga bagong halaman. Halimbawa, ang paghahasik ng tistle sa tulong ng mga rhizome ay maaaring makagawa ng higit sa isang libong mga bagong indibidwal bawat 1 m2 ng lupa.
1.2 Sekswal na pagpaparami
Ang sexual reproduction ay may napakahusay na evolutionary advantage kumpara sa asexual reproduction. Ito ay dahil sa ang katunayan na ang genotype ng mga supling ay lumitaw sa pamamagitan ng pagsasama-sama ng mga gene na kabilang sa parehong mga magulang. Bilang resulta, ang kakayahan ng mga organismo na umangkop sa mga kondisyon sa kapaligiran ay tumataas. Dahil ang mga bagong kumbinasyon ay isinasagawa sa bawat henerasyon, ang isang mas malaking bilang ng mga indibidwal ay maaaring iakma sa mga bagong kondisyon ng pag-iral kaysa sa panahon ng asexual reproduction. Ang paglitaw ng mga bagong kumbinasyon ng gene ay nagsisiguro ng mas matagumpay at mabilis na pagbagay ng mga species sa pagbabago ng mga kondisyon sa kapaligiran.
Kaya, ang kakanyahan ng sekswal na pagpaparami ay nakasalalay sa kumbinasyon sa namamana na materyal ng inapo ng genetic na impormasyon mula sa dalawang magkakaibang mga mapagkukunan - ang mga magulang.
Ang mga sex cell ay nabubuo sa gonad: male sperm, female ovule (o mga itlog). Sa unang kaso, ang kanilang pag-unlad ay tinatawag na spermatogenesis, sa pangalawa - oogenesis (mula sa Latin ovo - itlog).
Sa proseso ng pagbuo ng mga cell ng mikrobyo, ang isang bilang ng mga yugto ay nakikilala. Ang unang yugto ay ang panahon ng pagpaparami, kung saan ang mga primordial germ cells ay nahahati sa pamamagitan ng mitosis, na nagreresulta sa pagtaas ng kanilang bilang.
Ang ikalawang yugto ay ang panahon ng paglago. Sa immature male gametes hindi ito binibigkas. Ang kanilang mga sukat ay bahagyang tumaas. Sa kabaligtaran, ang mga hinaharap na itlog oocyte ay tumataas sa laki, minsan daan-daan, at mas madalas libu-libo at kahit milyon-milyong beses. Ang paglaki ng mga oocytes ay isinasagawa dahil sa mga sangkap na nabuo ng iba pang mga selula ng katawan. Kaya, sa isda, amphibian at, sa mas malaking lawak, sa mga reptilya at ibon, ang karamihan sa itlog ay ang pula ng itlog. Ito ay na-synthesize sa atay, dinadala sa isang espesyal na natutunaw na anyo ng dugo sa obaryo, tumagos sa lumalaking oocytes at idineposito doon sa anyo ng mga yolk plate. Bilang karagdagan, sa hinaharap na reproductive cell mismo, maraming mga protina at isang malaking bilang ng iba't ibang mga RNA ang na-synthesize: transportasyon, ribosomal at impormasyon. Ang yolk ay isang koleksyon ng mga sustansya (taba, protina, carbohydrates, bitamina, atbp.) na kinakailangan upang mapangalagaan ang pagbuo ng embryo, at tinitiyak ng RNA ang synthesis ng mga protina sa isang maagang yugto ng pag-unlad, kapag ang sarili nitong nakapipinsalang impormasyon ay hindi pa ginagamit.
Ang susunod na yugto, ang panahon ng pagkahinog, o meiosis, ay ipinakita sa Figure 2. Ang mga cell na papasok sa panahon ng pagkahinog ay naglalaman ng isang diploid na hanay ng mga chromosome at isang dobleng halaga ng DNA.

kanin. 2. Pagkahinog ng mga selulang mikrobyo (meiosis)
Ang kakanyahan ng meiosis ay ang bawat sex cell ay tumatanggap ng isang solong, haploid, set ng mga chromosome. Gayunpaman, sa parehong oras, ang meiosis ay isang yugto kung saan ang mga bagong kumbinasyon ng mga gene ay nilikha sa pamamagitan ng pagsasama-sama ng iba't ibang maternal at paternal chromosomes; ang recombination ng hereditary inclinations ay nangyayari rin bilang resulta ng pagtawid - ang pagpapalitan ng mga seksyon sa pagitan ng mga homologous chromosome sa panahon ng proseso. ng meiosis.
Ang Meiosis ay nagsasangkot ng dalawang magkakasunod na dibisyon. Tulad ng sa mitosis, ang bawat meiotic division ay may apat na yugto: prophase, metaphase, anaphase at telophase.
Unang (I) meiotic division. Nagsisimula ang prophase I sa spiralization ng mga chromosome. Tulad ng naaalala mo, ang bawat chromosome ay binubuo ng dalawang chromatids na konektado sa sentromere. Pagkatapos ang mga homologous chromosome ay magkakalapit, ang bawat punto ng bawat chromatid ng isang chromosome ay pinagsama sa kaukulang punto ng chromatid ng isa pa, homologous chromosome. Ang prosesong ito ng tumpak at malapit na pagsasama-sama ng mga homologous chromosome sa meiosis ay tinatawag na conjugation. Sa hinaharap, maaaring mangyari ang pagtawid sa pagitan ng mga naturang chromosome - isang palitan ng magkapareho, o homologous, ibig sabihin, naglalaman ng parehong mga gene, mga rehiyon. Sa pagtatapos ng prophase, ang mga salungat na puwersa ay lumitaw sa pagitan ng mga homologous chromosome. Una, lumilitaw ang mga ito sa rehiyon ng sentromere, at pagkatapos ay sa iba pang mga lugar.
Sa metaphase I, ang chromosome spiralization ay maximum. Ang mga conjugated chromosome ay matatagpuan sa kahabaan ng ekwador, kasama ang mga sentromer ng homologous chromosome na nakaharap sa iba't ibang pole ng cell. Ang mga spindle thread ay nakakabit sa kanila.
Sa anaphase I, ang mga braso ng homologous chromosome sa wakas ay naghihiwalay, at ang mga chromosome ay lumipat sa iba't ibang pole. Dahil dito, mula sa bawat pares ng homologous chromosome, isa lamang ang nakakapasok sa daughter cell. Ang bilang ng mga chromosome ay bumababa ng kalahati, ang chromosome set ay nagiging haploid. Gayunpaman, ang bawat chromosome ay binubuo ng dalawang chromatids, ibig sabihin, naglalaman pa rin ito ng dalawang beses sa dami ng DNA.
Sa telophase I, isang nuclear envelope ang nabuo sa maikling panahon. Sa panahon ng interphase sa pagitan ng una at pangalawang dibisyon ng meiosis, hindi nangyayari ang reduplication ng DNA. Ang mga cell na nabuo bilang isang resulta ng unang dibisyon ng pagkahinog ay naiiba sa komposisyon ng mga chromosome ng ama at ina at, dahil dito, sa hanay ng mga gene.
Halimbawa, ang lahat ng mga selula ng tao, kabilang ang mga primordial germ cell, ay naglalaman ng 46 na chromosome. Sa mga ito, 23 ang natanggap mula sa ama at 23 mula sa ina. Kapag nabuo ang mga cell ng mikrobyo pagkatapos ng unang meiotic division, ang mga spermatocytes at oocytes ay tumatanggap din ng 23 chromosome. Gayunpaman, dahil sa randomness ng divergence ng paternal at maternal chromosomes sa anaphase I, ang mga nagresultang cell ay tumatanggap ng isang malawak na iba't ibang mga kumbinasyon ng mga parental chromosome. Halimbawa, ang isa sa kanila ay maaaring may 3 paternal at 20 maternal chromosome, isa pang 10 paternal at 13 maternal, isang ikatlong 20 paternal at 3 maternal, atbp. Napakalaki ng bilang ng mga posibleng kumbinasyon. Kung isasaalang-alang din natin ang pagpapalitan ng mga homologous na seksyon ng mga chromosome sa prophase ng unang dibisyon ng meiosis, kung gayon ay malinaw na ang bawat resultang germ cell ay genetically unique, dahil nagdadala ito ng sarili nitong natatanging hanay ng mga gene.
Dahil dito, ang meiosis ay ang batayan ng combinative genotypic variability.
Pangalawang (II) meiotic division. Ang pangalawang dibisyon ng meiosis sa pangkalahatan ay nagpapatuloy sa parehong paraan tulad ng ordinaryong mitotic division, na may pagkakaiba lamang na ang naghahati na selula ay haploid. Sa anaphase II, ang mga centromeres na nagkokonekta sa mga kapatid na chromatids sa bawat chromosome ay nahahati, at ang mga chromatids, tulad ng sa mitosis, mula sa sandaling ito ay nagiging mga independiyenteng chromosome. Sa pagkumpleto ng telophase II, ang buong proseso ng meiosis ay nagtatapos: apat na haploid cell ang nabuo mula sa orihinal na pangunahing germ cell.
Sa mga lalaki, lahat ng mga ito ay na-convert sa gametes - tamud. Sa mga babae, dahil sa hindi pantay na meiosis, isang cell lamang ang gumagawa ng isang mabubuhay na itlog. Ang iba pang tatlong anak na selula ay mas maliit; sila ay nagiging tinatawag na paggabay, o pagbabawas, mga katawan, na malapit nang mamatay. Mula sa isang biological na pananaw, ang pagbuo ng isang itlog lamang at ang pagkamatay ng tatlong genetically complete guide bodies ay dahil sa pangangailangan na mapanatili sa isang cell ang lahat ng reserbang nutrients na kakailanganin para sa pagbuo ng hinaharap na embryo.
Ang panahon ng pagbuo ay binubuo ng mga cell na nakakakuha ng isang tiyak na hugis at sukat na naaayon sa kanilang pag-andar.
Sa panahon ng proseso ng pagkahinog, ang mga babaeng selula ng mikrobyo ay natatakpan ng mga lamad at handa na para sa pagpapabunga kaagad pagkatapos makumpleto ang meiosis. Sa maraming mga kaso, halimbawa sa mga reptilya, ibon at mammal, dahil sa aktibidad ng mga selula na nakapalibot sa itlog, lumilitaw ang isang bilang ng mga karagdagang lamad sa paligid nito. Ang kanilang tungkulin ay protektahan ang itlog at pagbuo ng embryo mula sa panlabas na masamang impluwensya. Ang tamud ay maaaring magkaroon ng iba't ibang laki at hugis.
Ang tungkulin ng tamud ay maghatid ng genetic na impormasyon sa itlog at pasiglahin ang pag-unlad nito. Ang nabuong tamud ay naglalaman ng mitochondria, ang Golgi apparatus, na nagtatago ng mga enzyme na natutunaw ang lamad ng itlog sa panahon ng pagpapabunga, iyon ay, sa panahon ng pagsasanib ng tamud at ng itlog. Ang nagresultang diploid cell ay tinatawag na zygote.
2. Indibidwal na pag-unlad ng mga organismo
Ang indibidwal na pag-unlad, o ontogenesis, ay tumutukoy sa buong panahon ng buhay ng isang indibidwal, mula sa sandaling ang tamud ay sumanib sa itlog at ang pagbuo ng isang zygote hanggang sa pagkamatay ng organismo. Ang ontogenesis ay nahahati sa dalawang panahon: 1) embryonic mula sa pagbuo ng zygote hanggang sa kapanganakan o paglabas mula sa mga lamad ng itlog; 2) postembryonic mula sa paglabas mula sa mga lamad ng itlog o pagsilang hanggang kamatayan ng organismo.
Ang agham na nag-aaral ng mga pattern ng indibidwal na pag-unlad ng mga organismo sa yugto ng embryonic ay tinatawag na embryology (mula sa Greek embryo embryo).
2.1 Embryonic na panahon ng pag-unlad
Sa karamihan ng mga multicellular na hayop, anuman ang pagiging kumplikado ng kanilang organisasyon, ang mga yugto ng pag-unlad ng embryonic na pinagdadaanan ng embryo ay pareho. Sa panahon ng embryonic, mayroong tatlong pangunahing yugto: cleavage, gastrulation at pangunahing organogenesis.
Naghihiwalay. Ang pag-unlad ng isang organismo ay nagsisimula sa solong yugto ng cell. Ang isang fertilized na itlog ay isang cell at sa parehong oras ay isang organismo sa pinakamaagang yugto ng pag-unlad nito. Bilang resulta ng paulit-ulit na paghahati, ang isang solong selulang organismo ay nagiging isang multicellular. Ang diploid nucleus, na lumilitaw sa panahon ng pagpapabunga sa pamamagitan ng pagsasanib ng isang tamud at isang itlog, ay nagsisimulang hatiin sa loob ng ilang minuto, at ang cytoplasm ay nahahati din dito. Ang mga resultang cell ay bumababa sa laki sa bawat dibisyon, kaya ang proseso ng paghahati ay tinatawag na cleavage. Sa panahon ng pagkapira-piraso, ang cellular na materyal ay nag-iipon para sa karagdagang pag-unlad. Ang fragmentation ay nagtatapos sa pagbuo ng isang multicellular embryo, ang blastula. Ang blastula ay may cavity na puno ng fluid, ang tinatawag na primary body cavity.
Sa mga kaso kung saan mayroong maliit na pula ng itlog sa cytoplasm ng itlog (tulad ng sa lancelet) o medyo maliit (tulad ng sa palaka), kumpleto ang fragmentation, ibig sabihin, ang itlog ay ganap na nahahati.
Kung hindi man, ang panahon ng pagkapira-piraso ay nangyayari sa mga ibon. Yolk-free cytoplasm ay bumubuo lamang ng 1% ng kabuuang dami ng isang itlog ng manok; ang buong natitirang cytoplasm ng itlog, at samakatuwid ang zygote, ay puno ng isang masa ng yolk. Kung titingnan mo nang mabuti ang isang itlog ng manok, sa isa sa mga pole nito nang direkta sa yolk maaari mong makita ang isang maliit na lugar - isang blastula, o germinal disc, na nabuo bilang isang resulta ng pagdurog sa yolk-free na seksyon ng cytoplasm na naglalaman ng nucleus. Sa ganitong mga kaso, ang pagdurog ay tinatawag na hindi kumpleto. Ang hindi kumpletong pagkapira-piraso ay katangian din ng ilang isda at reptilya.
Sa lahat ng mga kaso, pareho sa lancelet, at sa mga amphibian, at sa mga ibon, pati na rin sa iba pang mga hayop, ang kabuuang dami ng mga cell sa yugto ng blastula ay hindi lalampas sa dami ng zygote. Sa madaling salita, ang mitotic division ng zygote ay hindi sinamahan ng paglaki ng mga nagresultang anak na mga cell sa dami ng ina, at ang kanilang mga sukat bilang resulta ng isang serye ng mga sunud-sunod na dibisyon ay unti-unting bumababa. Ang tampok na ito ng mitotic cell division sa panahon ng cleavage ay sinusunod sa panahon ng pagbuo ng mga fertilized na itlog sa lahat ng mga hayop.
Ang ilang iba pang mga tampok ng pagdurog ay katangian din ng iba't ibang uri ng hayop. Halimbawa, ang lahat ng mga cell sa isang blastula ay may isang diploid na hanay ng mga chromosome, ay magkapareho sa istraktura at naiiba sa bawat isa pangunahin sa dami ng yolk na nilalaman nito. Ang ganitong mga cell, na walang mga palatandaan ng espesyalisasyon upang maisagawa ang ilang mga pag-andar, ay tinatawag na mga hindi espesyalisado (o hindi nakikilala) na mga selula. Ang isa pang tampok ng cleavage ay ang napakaikling mitotic cycle ng mga blastomeres kumpara sa mga selula ng isang pang-adultong organismo. Sa napakaikling interphase, tanging ang pagdoble ng DNA ang nangyayari.
Gastrulation. Ang blastula, bilang panuntunan, na binubuo ng isang malaking bilang ng mga blastomeres (halimbawa, sa lancelet mula sa 3000 na mga cell), sa panahon ng proseso ng pag-unlad ay pumasa sa isang bagong yugto, na tinatawag na gastrula (mula sa Greek gaster tiyan). Ang embryo sa yugtong ito ay binubuo ng malinaw na nakikilalang mga layer ng mga cell - ang tinatawag na mga layer ng mikrobyo: ang panlabas, o ectoderm (mula sa Greek ectos - matatagpuan sa labas), at ang panloob, o endoderm (mula sa Greek entos - matatagpuan sa loob) . Ang hanay ng mga proseso na humahantong sa pagbuo ng isang gastrula ay tinatawag na gastrulation.
Sa lancelet, ang gastrulation ay isinasagawa sa pamamagitan ng invagination ng isa sa mga pole ng blastula papasok, patungo sa isa pa; sa iba pang mga hayop, alinman sa pamamagitan ng delamination ng pader ng blastula, o sa pamamagitan ng paglaki ng napakalaking vegetative pole na may maliliit na selula ng poste ng hayop.
Sa mga multicellular na hayop, maliban sa mga coelenterates, kahanay ng gastrulation o, tulad ng sa lancelet, pagkatapos nito, ang ikatlong layer ng mikrobyo na mesoderm (mula sa Greek mesos na matatagpuan sa gitna) ay lilitaw, na isang hanay ng mga elemento ng cellular na matatagpuan sa pagitan ng ecto- at endoderm sa pangunahing cavity ng katawan blastocele. Sa paglitaw ng mesoderm, ang embryo ay nagiging tatlong-layered.
Kaya, ang kakanyahan ng proseso ng gastrulation ay ang paggalaw ng mga masa ng cell. Ang mga selula ng embryo ay halos nahati at hindi lumalaki. Gayunpaman, sa yugtong ito ang paggamit ng genetic na impormasyon ng mga embryonic cell ay nagsisimula, at ang mga unang palatandaan ng pagkita ng kaibahan ay lilitaw.
Ang differentiation, o differentiation, ay ang proseso ng paglitaw nito at ang pagtaas ng mga pagkakaiba sa istruktura at functional sa pagitan ng mga indibidwal na selula at mga bahagi ng embryo. Mula sa isang morphological point of view, ang pagkita ng kaibahan ay ipinahayag sa pagbuo ng ilang daang uri ng mga cell ng isang tiyak na istraktura na naiiba sa bawat isa. Mula sa mga hindi espesyal na selula ng blastula, ang mga epithelial cell ng balat, ang epithelium ng bituka, ang mga baga ay unti-unting lumalabas, ang mga selula ng nerve at kalamnan, atbp. Mula sa isang biochemical point of view, ang espesyalisasyon ng cell ay nakasalalay sa kakayahang mag-synthesize ng ilang mga protina na katangian lamang ng isang partikular na uri ng cell. Lymphocytes synthesize proteksiyon protina antibodies, kalamnan cell contractile protina myosin. Ang bawat uri ng cell ay gumagawa ng sarili nitong mga protina, na natatangi dito. Ang biochemical na pagdadalubhasa ng mga cell ay sinisiguro ng pumipili, pagkakaiba-iba ng aktibidad ng mga gene, ibig sabihin, sa mga selula ng iba't ibang mga layer ng mikrobyo, ang mga simulain ng ilang mga organo at sistema, ang iba't ibang mga grupo ng mga gene ay nagsisimulang gumana.
Sa iba't ibang uri ng hayop, ang parehong mga layer ng mikrobyo ay nagdudulot ng parehong mga organo at tisyu. Ibig sabihin homologous sila. Kaya, mula sa mga cell ng panlabas na layer ng mikrobyo - ectoderm - sa mga arthropod, chordates, kabilang ang mga isda, amphibian, reptile, ibon at mammal, ang balat at ang kanilang mga derivatives, pati na rin ang nervous system at sensory organ ay nabuo. Ang homology ng mga layer ng mikrobyo ng karamihan sa mga hayop ay isa sa mga patunay ng pagkakaisa ng mundo ng hayop.
Organogenesis. Matapos makumpleto ang gastrulation, ang embryo ay bumubuo ng isang complex ng axial organs: neural tube, notochord, at intestinal tube. Sa lancelet, ang mga axial organ ay nabuo tulad ng sumusunod: ang ectoderm sa dorsal side ng embryo ay yumuko sa kahabaan ng midline, nagiging isang uka, at ang ectoderm na matatagpuan sa kanan at kaliwa nito ay nagsisimulang tumubo sa mga gilid nito. Ang uka, ang rudiment ng nervous system, ay lumulubog sa ilalim ng ectoderm, at ang mga gilid nito ay malapit. Ang isang neural tube ay nabuo. Ang natitirang bahagi ng ectoderm ay ang rudiment ng epithelium ng balat.
Ang dorsal na bahagi ng endoderm, na matatagpuan mismo sa ilalim ng nerve rudiment, ay nahihiwalay mula sa natitirang bahagi ng endoderm at nakatiklop sa isang siksik na kurdon na tinatawag na notochord. Mula sa natitirang bahagi ng endoderm, nabuo ang mesoderm at bituka epithelium. Ang karagdagang pagkita ng kaibhan ng mga embryonic cell ay humahantong sa paglitaw ng maraming mga derivatives ng mga layer ng mikrobyo - mga organo at tisyu. Sa proseso ng pagdadalubhasa ng mga selula na bumubuo sa mga layer ng mikrobyo, ang sistema ng nerbiyos, mga organo ng pandama, epithelium ng balat, at enamel ng ngipin ay nabuo mula sa ectoderm; mula sa endoderm intestinal epithelium, digestive glands atay at pancreas, epithelium ng hasang at baga; mula sa mesoderm muscle tissue, connective tissue, kabilang ang maluwag na connective tissue, cartilage at bone tissue, dugo at lymph, pati na rin ang circulatory system, kidney, gonads.
2.2 Postembryonic na panahon ng pag-unlad
Sa sandali ng kapanganakan o paglabas ng organismo mula sa mga shell ng itlog, ang panahon ng embryonic ay nagtatapos at ang postembryonic na panahon ng pag-unlad ay nagsisimula. Ang postembryonic development ay maaaring direkta o sinamahan ng pagbabago (metamorphosis).
Sa panahon ng direktang pag-unlad (sa mga reptilya, ibon, mammal), isang organismo na may maliit na sukat ay lumalabas mula sa mga shell ng itlog o mula sa katawan ng ina, ngunit kasama ang lahat ng mga pangunahing organo na katangian ng isang may sapat na gulang na hayop na nabuo na. Ang postembryonic development sa kasong ito ay nababawasan pangunahin sa paglaki at pagdadalaga.
Sa panahon ng pag-unlad na may metamorphosis, isang larva ang lumalabas mula sa itlog, kadalasang mas simple sa istraktura kaysa sa isang pang-adultong hayop, na may mga espesyal na larva na organo na wala sa pang-adultong estado. Ang larva ay nagpapakain, lumalaki, at sa paglipas ng panahon ang mga larva na organo ay pinapalitan ng mga organo na katangian ng mga indibidwal na nasa hustong gulang. Dahil dito, sa panahon ng metamorphosis, ang mga larval organ ay nawasak at ang mga organo na katangian ng mga adult na hayop ay lumilitaw.
Tingnan natin ang ilang mga halimbawa ng hindi direktang pag-unlad ng postembryonic. Ang ascidian larva (phylum Chordata, subphylum Larval-Chordata) ay mayroong lahat ng pangunahing katangian ng mga chordates: isang notochord, isang neural tube, at gill slits sa pharynx. Malaya itong lumalangoy, pagkatapos ay nakakabit sa ilang solidong ibabaw sa ilalim ng dagat, kung saan nangyayari ang metamorphosis: ang buntot, notochord, at mga kalamnan nito ay nawawala, at ang neural tube ay nahahati sa mga indibidwal na selula, na karamihan ay phagocytosed. Ang lahat na natitira sa larval nervous system ay isang grupo ng mga selula na nagbubunga ng nerve ganglion. Ang istraktura ng katawan ng isang may sapat na gulang na ascidian, na humahantong sa isang nakalakip na pamumuhay, ay hindi katulad ng karaniwang mga tampok ng samahan ng mga chordates. Ang kaalaman lamang sa mga tampok ng ontogenesis ay posible upang matukoy ang sistematikong posisyon ng mga ascidian: ang istraktura ng larvae ay nagpapahiwatig ng kanilang pinagmulan mula sa mga chordates na humantong sa isang libreng pamumuhay. Sa panahon ng proseso ng metamorphosis, ang mga ascidian ay lumipat sa isang laging nakaupo na pamumuhay, at samakatuwid ang kanilang organisasyon ay pinasimple.
Ang larval form ng amphibians ay isang tadpole, na kung saan ay nailalarawan sa pamamagitan ng gill slits, isang lateral line, isang two-chambered heart, at isang bilog ng sirkulasyon ng dugo. Sa panahon ng proseso ng metamorphosis, na nangyayari sa ilalim ng impluwensya ng thyroid hormone, ang buntot ay nalulutas, lumilitaw ang mga paa, nawawala ang lateral line, ang mga baga at isang pangalawang bilog ng sirkulasyon ng dugo ay nabuo. Kapansin-pansin ang pagkakatulad ng isang bilang ng mga tampok na istruktura ng mga tadpoles at isda (lateral line, istraktura ng puso at sistema ng sirkulasyon, mga gill slits).
Ang pag-unlad ng mga insekto ay maaari ding magsilbi bilang isang halimbawa ng metamorphosis. Ang mga butterfly caterpillar o dragonfly larvae ay naiiba nang husto sa istraktura, pamumuhay at tirahan mula sa mga adult na hayop at kahawig ng kanilang mga ninuno, annelids.
Ang postembryonic period of development ay may iba't ibang tagal. Halimbawa, ang mga mayflies ay nabubuhay ng 2-3 taon sa larval state, at mula 2-3 oras hanggang 2-3 araw sa mature state, depende sa species. Sa karamihan ng mga kaso, ang postembryonic period ay mas mahaba. Sa mga tao, kabilang dito ang yugto ng pagdadalaga, ang yugto ng kapanahunan at ang yugto ng katandaan.
Sa mga mammal at tao, mayroong kilalang pag-asa sa pag-asa sa buhay sa tagal ng pagdadalaga at pagbubuntis. Karaniwang lumalampas ang pag-asa sa buhay
pre-reproductive period ng ontogenesis ng 5-8 beses.
Ang pag-unlad ng postembryonic ay sinamahan ng paglaki. Ang isang pagkakaiba ay ginawa sa pagitan ng hindi tiyak na paglago, na nagpapatuloy sa buong buhay, at tiyak na paglago, na limitado sa isang tiyak na panahon. Ang hindi tiyak na paglaki ay sinusunod sa makahoy na anyo ng mga halaman, ilang mollusk, vertebrates, isda, at daga.
Sa maraming mga hayop, humihinto ang paglaki sa ilang sandali pagkatapos maabot ang sekswal na kapanahunan. Sa mga tao, ang paglaki ay nagtatapos sa edad na 20-25.
2.3 Pangkalahatang mga pattern ng pag-unlad. Batas ng biogenetic
Lahat ng multicellular organism ay nabubuo mula sa isang fertilized na itlog. Ang pagbuo ng mga embryo sa mga hayop na kabilang sa parehong uri ay halos magkapareho. Sa lahat ng mga chordates, sa panahon ng embryonic, ang axial skeleton notochord ay nabuo, ang neural tube ay lilitaw, at ang mga gill slits ay nabuo sa anterior na bahagi ng pharynx. Ang structural plan ng chordates ay pareho din. Sa mga unang yugto ng pag-unlad, ang mga vertebrate embryo ay halos magkapareho (Larawan 3). Ang mga katotohanang ito ay nagpapatunay sa bisa ng batas ng embryonic na pagkakatulad na binuo ni K. Baer: "Ang mga embryo ay nagpapakita, na mula sa pinakamaagang yugto, ng isang tiyak na pangkalahatang pagkakatulad sa loob ng uri." Ang pagkakatulad ng mga embryo ng iba't ibang sistematikong grupo ay nagpapahiwatig ng kanilang karaniwang pinagmulan. Kasunod nito, ang istraktura ng mga embryo ay nagpapakita ng mga katangian ng klase, genus, species, at, sa wakas, mga katangian na katangian ng isang indibidwal. Ang pagkakaiba-iba ng mga katangian ng mga embryo sa panahon ng pag-unlad ay tinatawag na embryonic divergence at sumasalamin sa ebolusyon ng isang partikular na sistematikong pangkat ng mga hayop, ang kasaysayan ng pag-unlad ng isang partikular na species.

kanin. 3. Pagkakatulad ng germline sa mga vertebrates: 1 monotremes (echidna), 2 marsupial (kangaroos), 3 artiodactyls (deer), 4 carnivores (pusa), 5 primates (unggoy), 6 - tao
Malaking pagkakatulad sa pagitan ng mga embryo sa mga unang yugto ng pag-unlad at
ang kababalaghan ng mga pagkakaiba sa mga susunod na yugto ay may sariling paliwanag.
Ang katawan ay napapailalim sa pagkakaiba-iba sa buong pag-unlad.
Ang proseso ng mutation ay nakakaapekto sa mga gene na tumutukoy sa structural at metabolic features ng mga pinakabatang embryo. Ngunit ang mga istruktura na lumitaw sa kanila (mga sinaunang katangian na katangian ng malayong mga ninuno) ay may napakahalagang papel sa mga proseso ng karagdagang pag-unlad. Tulad ng ipinahiwatig, ang notochord primordium ay nagpapahiwatig ng pagbuo ng neural tube, at ang pagkawala nito ay humahantong sa pagtigil ng pag-unlad. Samakatuwid, ang mga pagbabago sa mga unang yugto ay kadalasang humahantong sa hindi pag-unlad at pagkamatay ng indibidwal. Sa kabaligtaran, ang mga pagbabago sa mga susunod na yugto, na nakakaapekto sa hindi gaanong makabuluhang mga katangian, ay maaaring maging kapaki-pakinabang para sa organismo at sa mga ganitong kaso ay nakuha ng natural na pagpili.
Ang hitsura sa embryonic na panahon ng pag-unlad ng mga modernong hayop ng mga katangian na katangian ng kanilang malayong mga ninuno ay sumasalamin sa ebolusyonaryong pagbabago sa istraktura ng mga organo.
Sa pag-unlad nito, ang organismo ay dumadaan sa isang single-celled stage (zygote stage), na maaaring ituring bilang isang pag-uulit ng phylogenetic stage ng primitive amoeba. Sa lahat ng mga vertebrates, kabilang ang kanilang pinakamataas na kinatawan, ang isang notochord ay nabuo, na pagkatapos ay pinalitan ng isang gulugod, at sa kanilang mga ninuno, na hinuhusgahan ng lancelet, ang notochord ay nanatili para sa buhay. Sa panahon ng embryonic development ng mga ibon at mammal, kabilang ang mga tao, ang mga gill slits at kaukulang septa ay lumilitaw sa pharynx. Ang katotohanan ng pagbuo ng mga bahagi ng gill apparatus sa mga embryo ng terrestrial vertebrates ay ipinaliwanag sa pamamagitan ng kanilang pinagmulan mula sa mga ninuno na parang isda na huminga gamit ang mga hasang. Ang istraktura ng puso ng isang embryo ng tao sa maagang yugto ng pagbuo ay kahawig ng istraktura ng organ na ito sa isda: mayroon itong isang atrium at isang ventricle. Ang mga balyena na walang ngipin ay nagkakaroon ng mga ngipin sa panahon ng embryonic. Ang mga ngipin na ito ay hindi pumuputok; sila ay nawasak at natutunaw.
Ang mga halimbawang ibinigay dito at marami pang iba ay tumutukoy sa isang malalim na koneksyon sa pagitan ng indibidwal na pag-unlad ng mga organismo at ng kanilang makasaysayang pag-unlad. Ang koneksyon na ito ay ipinahayag sa biogenetic na batas na binuo ni F. Müller at E. Haeckel noong ika-19 na siglo: ang ontogenesis (indibidwal na pag-unlad) ng bawat indibidwal ay isang maikli at mabilis na pag-uulit ng phylogeny (historical development) ng species kung saan kabilang ang indibidwal na ito. .
Konklusyon
Ang pagkumpleto ng gawain sa abstract, maaari tayong makarating sa konklusyon na ang kakayahang magparami, o pagpaparami sa sarili, ay isa sa pinakamahalagang katangian ng organikong kalikasan. Ang pagpaparami ay isang pag-aari na likas sa lahat ng nabubuhay na organismo nang walang pagbubukod, mula sa bakterya hanggang sa mga mammal.
Ang pagkakaroon ng anumang uri ng hayop at halaman, bakterya at fungi, ang pagpapatuloy sa pagitan ng mga indibidwal na magulang at kanilang mga supling ay pinananatili lamang sa pamamagitan ng pagpaparami. Ang malapit na nauugnay sa pagpaparami ng sarili ay isa pang pag-aari ng mga nabubuhay na bagay.pag-unlad ng mga organismo. ItoIto rin ay likas sa lahat ng buhay sa Earth: ang pinakamaliit na single-celled na organismo, at multicellular na mga halaman at hayop.
Bibliograpiya
Bogen G. Makabagong biyolohiya. - M.: Mir, 1970.
Green N., Stout W., Taylor D. Biology: sa 3 volume. T. 3: trans. mula sa English/ed. R. Soper. - M.: Mir, 1990.
Mamontov S.G. Biology. Pangkalahatang mga pattern. M.: Bustard, 2002.
Mula sa hayop hanggang sa tao. M.: Nauka, 1971.
Slyusarev A.A. Biology na may pangkalahatang genetika. - M.: Medisina, 1978.
Lecture No. 3 Ontogenesis
1. Gametogenesis
2. Embryonic period
3. Postembryonic period
Ontogenesis– indibidwal na pag-unlad ng isang organismo mula sa zygote hanggang biological na kamatayan. Proogenesis– ang panahon bago ang ontogenesis, kasama ang gametogenesis at fertilization. Phylogenesis– ebolusyonaryong pag-unlad ng mga species.
Ang mga tao ay nailalarawan sa pamamagitan ng sekswal na pagpaparami; ito ay nailalarawan sa pamamagitan ng: pagkakaroon ng mga gonad, gametes, proseso ng pagpapabunga, at sekswal na dimorphism.
Gametogenesis – proseso ng pagbuo ng mga selula ng mikrobyo ovogenesis - pagkahinog ng oocyte, spermatogenesis– spermatozoa. Ang gametogenesis ay nangyayari sa mga gonad ng katawan. Ang mga gametes ay nabuo mula sa mga embryonic epithelial cells, na nabuo sa panahon ng embryonic na pag-unlad ng organismo.
Sa proseso ng pagbuo, ang mga selula ng mikrobyo ay dumaan sa tatlong yugto:
1. Panahon ng pagpaparami (ang mga selula ng embryonic epithelium ay nahahati sa pamamagitan ng mitosis);
2. Panahon ng paglago;
3. Ang panahon ng pagkahinog, ang mga cell ay nahahati sa pamamagitan ng meiosis, na nagreresulta sa pagbuo ng mga gametes (Larawan 5).
kanin. 5. Dibisyon ng mga selulang mikrobyo
Ang tamud ay mature sa seminiferous tubules ng testes. Matatagpuan sa pagitan ng mga seminiferous tubules interstitial tissue, gumagawa ng male sex hormone - testosterone. Mga male sex hormones - androgens, kinokontrol ang reproductive function, gametogenesis, at pagbuo ng pangalawang sekswal na katangian. Ang tamud ay mature sa loob ng 70 araw. Ang mga seminiferous tubules ay naglalaman ng mga gametes sa iba't ibang yugto ng pagkahinog. Sa 5 ml. Ang seminal fluid ng tao ay naglalaman ng 12 milyong tamud. Ang mga ito ay negatibong sinisingil, pinipigilan sila ng singil na magkadikit. Ang isang malaking bilang ng mga male gametes ay biologically expedient; ang pagbaba sa kanilang bilang ng 40% ay ginagawang imposible ang proseso ng pagpapabunga. Ang Spermatozoa ay maliit, motile na mga selula na binubuo ng ulo, leeg, at buntot. Sa ulo ay akrosom, isang binagong lysosome, naglalaman ito ng mga enzyme na tumutunaw sa lamad ng itlog sa panahon ng pagpapabunga. Ang pag-asa sa buhay ay mula 6 hanggang 72 oras.
Ang oogenesis ay nagsisimula sa embryonic na panahon ng pag-unlad ng babaeng katawan. Sa yugto ng prophase ng unang dibisyon ng meiosis, kapag nangyari ang conjugation at crossing over, nangyayari ang isang pansamantalang paghinto. Ang karagdagang pag-unlad ng itlog ay nagpapatuloy sa panahon ng pagdadalaga ng batang babae. Sa ilalim ng impluwensya ng mga babaeng sex hormones - estrogens, isang follicle, isang cellular vesicle na nagpoprotekta at nagpapalusog sa cell, ay nabuo sa paligid ng first-order oocyte.
Habang tumatanda ang follicle, ito ay gumagalaw sa gilid ng obaryo at pagkatapos ay sasabog; ang yugto ng pag-unlad ng itlog na ito ay tinatawag na ovarian o follicular; ito ay tumatagal ng 12 araw. Ang pagkalagot ng follicle at ang paglabas ng itlog mula sa obaryo papunta sa lukab ng tiyan ay tinatawag na obulasyon.
Pagkatapos ng obulasyon, posible ang pagpapabunga. Ang babaeng gamete ay nakuha ng fimbriae ng fallopian tube. Sa tulong ng ciliated epithelium, lumilipat ito sa matris; ang yugto ng matris ay tumatagal ng 12-14 araw. Sa oras na ito, sa ilalim ng impluwensya ng mga babaeng sex hormones, ang uterine epithelium ay lumuwag. Kung hindi nangyari ang pagpapabunga, ang epithelium at mga itlog ay pinalabas mula sa katawan bilang resulta ng pag-urong ng makinis na mga kalamnan ng mga dingding ng matris. Ang prosesong ito ay tinatawag na mensis at tumatagal ng 3-4 na araw. Kaya, ang oras mula sa simula ng pagbuo ng follicle hanggang sa simula ng mensis ay 20-30 araw at tinatawag na ovarian-menstrual cycle. (Larawan 6).

kanin. 6. Obulasyon at pagtatanim ng itlog sa matris
Kasama dito ovarian stage, uterine at mensis. Sa panahon ng ovarian-menstrual cycle, ang buong reproductive system ng isang babae ay muling itinatayo, ang qualitative at quantitative na komposisyon ng mga hormone ay nagbabago, performance, lakas, flexibility, at endurance change. Ang mga pagbabago sa physiological sa babaeng katawan ay isinasaalang-alang kapag nag-aayos ng proseso ng pagsasanay.
Pagpapabunga- Ito ay ang pagsasanib ng isang itlog at isang tamud. Mayroong tatlong yugto ng pagpapabunga:
1) acrosomal reaction (20 segundo) – pagkalusaw ng egg shell sa pamamagitan ng acrosome enzymes at pagtagos ng sperm sa itlog. Ito ay pinaniniwalaan na ang tamud na lumilikha ng pinakamalaking potensyal na pagtagos (katuwaan) sa lamad ng itlog ay tumagos sa itlog;
2) pansamantalang paghinto - walang nakikitang pagbabago na nagaganap sa itlog, tumataas ang metabolismo;
3) pagsasanib ng nuclei ng itlog at tamud, pagpapanumbalik ng diploid na hanay ng mga chromosome.
Ang fertilization ay nangyayari sa itaas na ikatlong bahagi ng fallopian tube. Ang fertilized egg ay tinatawag zygote. Sa pagbuo ng isang zygote, nagsisimula ang ontogenesis ng tao.
2. Ontogenesis Ang tao ay may dalawang yugto: embryonic at postembryonic.
Embryonic na yugto ng pag-unlad nagpapatuloy mula sa zygote hanggang sa kapanganakan. Kasama ang mga yugto: zygote, blastula, gastrula, histogenesis, organogenesis.
Zygote- paulit-ulit na nahahati sa pamamagitan ng mitosis, na nagreresulta sa pagbuo ng isang multicellular single-layer na embryo - blastula. Ang blastula ay hindi tumataas sa laki, dahil sa oras na ito ito ay nasa fallopian tube at dapat na malayang lumipat sa matris. Sa ikaanim na araw, pumapasok ito sa matris at nakakabit sa dingding nito, bumulusok sa lumuwag na epithelium. Ang prosesong ito ay tinatawag na pagtatanim. Ang embryo ay patuloy na umuunlad at nagiging dalawa, pagkatapos ay tatlong-layered. Sa yugtong ito ng pag-unlad ito ay tinatawag gastrula. Bilang resulta, nabuo ang tatlong layer ng mikrobyo: ectoderm, endoderm, mesoderm. Ang mga tissue at organ ay nabuo mula sa mga embryonic system.
kaya: blastula- single-layer na embryo, gastrula– dalawa at tatlong-layer na embryo. Histogenesis– paglalatag ng mga tisyu mula sa mga layer ng mikrobyo. Organogenesis- paglalagay ng organ. Sa edad na 8 linggo, ang masa ng embryo ng tao ay 4 g, laki ng 5 mm. Sa oras na ito, siya ay sumailalim sa mga proseso ng pagbuo at nakuha ang mga contour ng isang katawan ng tao. Pagkatapos ng walong linggo ng pagbubuntis, ang embryo ng tao ay tinatawag na fetus.
Sa panahon ng pag-unlad ng embryo ng tao, pansamantalang awtoridad, na pagkatapos ng kapanganakan ay nawawala ang kanilang mga tungkulin. Ang mga pansamantalang awtoridad ay kinabibilangan ng: germinal shell, chorion, amnion at inunan, umbilical cord. Inunan Tinitiyak ang malapit na pakikipag-ugnayan sa pagitan ng mga capillary ng bata at ng ina. Ang umuunlad na organismo ay tumatanggap ng mga sustansya, oxygen, mga antibodies sa pamamagitan ng inunan, at nag-aalis ng mga produktong metabolic. Ang inunan ay gumaganap ng isang barrier function, na nagpoprotekta sa embryo mula sa mga microorganism, ilang mga gamot at mga lason (Larawan 7).

kanin. 7. Inunan
1 – matris; 2 – amnion; 3 – embryo; 4 – amniotic fluid; 5 – inunan; 6 - mga capillary ng inunan; 7 – ugat; 8 – mga arterya
Ang mga banayad na mekanismo ng pagbuo ng mga organo at tisyu ng bata ay maaaring maputol kapag nalantad sa nakakalason, narcotic substance, alkohol, nikotina na nasa dugo ng ina. Ang mga karamdaman sa pag-unlad ng embryonic ng tao ay tinatawag na congenital deformities at malformations. Kabilang dito ang: "cleft palate" (kawalan ng upper palate), "cleft lip" (non-fusion of maxillary bones), polydactyly (extra fingers), syndactyly (fused fingers). Ang mga hindi kanais-nais na kadahilanan sa kapaligiran na nagdudulot ng mga congenital malformations ay tinatawag teratogenic(nagdudulot ng mga deformidad) (Larawan 8).

kanin. 8. Mga salik sa kapaligiran na nakakaimpluwensya sa pag-unlad ng katawan ng tao
Ang embryonic na panahon ng pag-unlad ng tao ay tumatagal ng 36 na linggo.
3. Ang panahon pagkatapos ng kapanganakan at bago ang biyolohikal na kamatayan ay tinatawag postembryonic. Kasama dito kabataan, pagdadalaga mga panahon ng pag-unlad at pagtanda.
Juvenile – panahon bago ang pagdadalaga. Pagbibinata– panahon ng pagdadalaga.
Sa pagsasanay sa palakasan, pisyolohiya, pedagogy, sikolohiya, ginagamit ang sumusunod na periodization ng edad postembryonic ontogenesis ng mga tao.
Sa panahon mula 1 hanggang 10 araw ay tinawag ang bata bagong panganak;
Mula 10 araw hanggang 1 taon - dibdib;
mula 1 taon hanggang tatlo – panahon maagang pagkabata;
mula 4 hanggang 7 taon - unang pagkabata;
mula 8 taon hanggang 12 para sa mga babae, at hanggang 13 para sa mga lalaki ikalawang pagkabata;
mula 12 taon hanggang 15 para sa mga babae at mula 13 hanggang 16 para sa mga lalaki - mga teenager;
17-21 (lalaki), 16-20 taon (babae) - pagdadalaga;
20-55 para sa mga babae at 21-60 para sa mga lalaki pagdadalaga(pagbibinata);
mula 55 at 60 hanggang 70 - matatandang tao;
mula 70 hanggang 90 matatandang lalaki;
higit sa 90 mga sentenaryo.
Sa ontogenesis ng tao mayroong sensitibo at kritikal mga panahon.
Mga kritikal na panahon ay nailalarawan sa pamamagitan ng pagtaas ng aktibidad ng mga indibidwal na gene at ang kanilang mga complex na kumokontrol sa pagbuo ng anumang mga palatandaan ng katawan. Sa mga panahong ito, mayroong isang makabuluhang muling pagsasaayos ng mga proseso ng regulasyon, isang husay at dami ng paglukso sa pagbuo ng mga indibidwal na organo at mga functional na sistema. Sa mga panahong ito, ang katawan ay pinaka-sensitibo sa mga epekto ng mga salik sa kapaligiran. Ang mga kritikal na panahon ng ontogenesis ay itinuturing na implantation, placentation, pagbuo ng axial organs, neural at intestinal tubes, notochords, heart formation, at iba pa.
Kung ang isang babae ay nagkaroon ng rubella sa pagitan ng ika-3 at ika-9 na linggo ng pagbubuntis, may panganib na ang fetus ay magkaroon ng mga depekto tulad ng sakit sa puso, katarata, at pagkabingi. Sa ibang pagkakataon, ang rubella ay hindi nagiging sanhi ng mga malformasyon ng pangsanggol.
Mga sensitibong panahon– ito ay mga panahon ng pagbaba ng genetic control at pagtaas ng sensitivity ng mga indibidwal na katangian ng katawan sa mga impluwensya sa kapaligiran, kabilang ang pedagogical at coaching. Kaya, ang sensitibong panahon para sa pagpapakita ng iba't ibang mga tagapagpahiwatig ng kalidad ng bilis ay nangyayari sa edad na 11-14 taon at sa edad na 15 ang pinakamataas na antas nito ay naabot, kapag posible ang mataas na mga tagumpay sa palakasan. Ang isang katulad na larawan ay sinusunod sa ontogenesis para sa pagpapakita ng mga katangian ng dexterity at flexibility.
Para sa mga coach at guro na nagtatrabaho sa larangan ng pisikal na edukasyon at palakasan, ang kaalaman sa mga sensitibong panahon ay napakahalaga, dahil sa mga sensitibong panahon ang pinakadakilang epekto ng pagsasanay.
Maaari silang hatiin depende sa kung gaano karaming mga cell ang kasangkot sa proseso ng asexual reproduction: asexual reproduction kung saan ang isang anak na henerasyon ay nagmumula sa isang cell: cell division multiple cell division schizogony sporulation sporulation budding sa unicellular yeast...
Ibahagi ang iyong trabaho sa mga social network
Kung ang gawaing ito ay hindi angkop sa iyo, sa ibaba ng pahina ay may isang listahan ng mga katulad na gawa. Maaari mo ring gamitin ang pindutan ng paghahanap
Pagpaparami at pag-unlad ng mga organismo
Plano
- Ang konsepto at kahulugan ng pagpaparami.
- Mga anyo at uri ng pagpaparami.
- Ikot ng cell. Mitosis. Meiosis.
- Ang istraktura ng gametes. Gametogenesis.
1. Ang konsepto at kahulugan ng pagpaparami
Isa sa mga katangian ng mga bagay na may buhay ay discreteness, mga. sa anumang antas ng organisasyon, ang buhay na bagay ay kinakatawan ng elementarya na mga yunit ng istruktura. Ang bawat indibidwal ng isang partikular na species ay mortal, at ang pagkakaroon ng species ay pinananatili pagpaparami mga organismo. Kaya, ang discreteness ng buhay presupposes nito reproduction, i.e. proseso ng pagpaparami.
Pagpaparami ito ang kakayahan ng mga nabubuhay na nilalang na magparami ng kanilang sariling uri. Tinitiyak nito ang pagpapatuloy at pagpapatuloy ng buhay. Ang pagpapatuloy ay tinutukoy ng katotohanan na sa panahon ng proseso ng pagpaparami, ang genetic na materyal ay inilipat mula sa mga magulang hanggang sa mga supling, bilang isang resulta kung saan ang mga katangian ng magulang ay ipinakita sa isang antas o iba pa sa mga organismo ng anak na babae.
Halaga ng pagpaparami:
- Pagdaragdag o pagpapanatili ng bilang ng isang species. Dahil sa pagpaparami, hindi lamang pagpaparami ang nangyayari (i.e., pagpaparami ng sariling uri), kundi pati na rin ang pagtaas ng bilang ng mga nabubuhay na organismo.
- Pagpapatuloy ng buhay. Salamat sa pagpaparami, higit pa o hindi gaanong malalaking grupo ng mga indibidwal (halimbawa, mga populasyon at species) ay maaaring umiral nang mahabang panahon, dahil ang pagbaba sa kanilang mga bilang dahil sa natural na pagkamatay ng mga indibidwal ay binabayaran ng patuloy na pagpaparami ng mga organismo at ang pagpapalit ng mga patay ng mga bagong silang.
- Pagpapatuloy sa pagitan ng mga henerasyon. Sa panahon ng proseso ng pagpaparami, ang genetic na impormasyon ay ipinapadala mula sa mga indibidwal ng henerasyon ng magulang, na tinitiyak ang pagpaparami ng mga katangian ng mga partikular na magulang at ang buong species kung saan kabilang ang organismo.
- Ang mga katangian ng mga nabubuhay na bagay tulad ng pagmamana at pagkakaiba-iba ay natanto.
2. Mga anyo at uri ng pagpaparami
Mayroong dalawang pangunahing anyo ng pagpaparami: sekswal at asexual.
Asexual reproduction
Sa panahon ng asexual reproduction, lumilitaw ang isang bagong indibidwal mula sa hindi espesyalisadong non-reproductive somatic cells ng katawan. Samakatuwid, ang asexual reproduction ay nagsasangkot ng isang indibidwal. Tinitiyak ng asexual reproduction ang pagpaparami ng malaking bilang ng magkakaparehong indibidwal, na kapaki-pakinabang para sa mga species na naninirahan sa pare-parehong mga kondisyon. Ang mabilis at maraming pagpaparami ng mga supling na kapareho ng sa ina ay ang biological na kahulugan ng asexual reproduction.
Sa kalikasan, mayroong iba't ibang mga opsyon para sa asexual reproduction:division, sporulation, fragmentation, budding, vegetative propagation, cloning.
Maaari silang hatiin depende sa kung gaano karaming mga cell ang kasangkot sa proseso ng asexual reproduction:
- asexual reproduction, kung saan ang henerasyon ng anak na babae ay nagmumula sa isang cell:
- paghahati ng selula
- maramihang dibisyon ng cell (schizogony)
- sporulation (sporulation)
- namumuko sa mga unicellular na organismo (yeast)
2) asexual reproduction, na batay sa paghahati ng isang pangkat ng mga cell:
- vegetative
- pagkakapira-piraso
- namumuko sa mga multicellular na organismo (hydra)
May isa pang pag-uuri, ayon sa kung saan ang mga opsyon para sa asexual reproduction ay:
- vegetative propagation (ibig sabihin, paghahati ayon sa mga bahagi ng katawan ng ina):
Dibisyon
Maramihang fission (schizogony)
Namumuko
Pagkapira-piraso
Pagpapalaganap ng halaman sa pamamagitan ng mga pinagputulan, bombilya, dahon, rhizome
- pagpaparami sa pamamagitan ng spores, i.e. sporulation
Tingnan natin ang mga uri ng asexual reproduction nang mas detalyado.
Dibisyon. Ang pinakasimpleng anyo ng asexual reproduction. Mayroong iba't ibang mga pagpipilian sa dibisyon:
- Dibisyon sa pamamagitan ng simpleng paghihigpit sa pagbuo ng dalawang anak na organismo mula sa isang magulang na organismo. Katangian ng bacteria at cyanobacteria.
- Dibisyon sa pamamagitan ng mitotic division ng nucleus na sinusundan ng paghihiwalay ng cytoplasm. Katangian ng mga unicellular organism (maraming protozoa - amoeba, green euglena, atbp.; unicellular algae - chlamydomonas, atbp.).
Sa parehong mga kaso ito ay nangyayari binary dibisyon, i.e. sa dalawang cell. Gayunpaman, posible rin ang isa pang pagpipilian:
- Maramihang fission (schizogony). Una, ang nucleus ay paulit-ulit na nahahati at pagkatapos ay ang cytoplasm ay nahahati sa mga bahagi. Halimbawa, ang pagpaparami ng Plasmodium falciparum (ang causative agent ng malaria) sa mga erythrocytes ng tao. Sa kasong ito, ang plasmodia ay sumasailalim sa paulit-ulit na dibisyon ng nucleus nang maraming beses, pagkatapos ay nahahati ang cytoplasm. Bilang resulta, ang 1 plasmodium ay nagbibigay ng 12-24 na mga organismo ng anak na babae.
Pagbubuo ng spore (sporulation). Isang napakalawak na paraan ng asexual reproduction sa mga nabubuhay na organismo at matatagpuan sa halos lahat ng halaman, fungi at ilang protozoa (halimbawa, ang uri ng sporozoan), pati na rin ang mga prokaryotic na organismo (maraming bacteria, blue-green algae).
Spore Ito ay isang cell na natatakpan ng isang binuo na proteksiyon na takip - isang spore membrane, na nagbibigay-daan dito upang mapaglabanan ang mga epekto ng iba't ibang hindi kanais-nais na mga kadahilanan sa kapaligiran.Dahil sa maliit na sukat nito, ang spore ay karaniwang naglalaman lamang ng kaunting mga reserbang nutrisyon. Sa maraming mga organismo, ito ang pangunahing yunit ng dispersal, dahil ang malalaking dami ng mga light spores na nabuo ay malayang dinadala sa mga makabuluhang distansya sa pamamagitan ng paggalaw ng mga masa ng hangin at mga daloy ng tubig.Kadalasan, ang mga spores ay nabuo sa maraming dami at may hindi gaanong timbang, na ginagawang mas madali silang kumalat sa pamamagitan ng hangin, pati na rin ng mga hayop, pangunahin ang mga insekto.
Bilang karagdagan, sa ilang mga anyo (protozoa, fungi), ang mga spores ay maaaring kumilos bilang isang espesyal na yugto ng siklo ng buhay, na nagpapahintulot sa kanila na "makaligtas" sa mga epekto ng hindi kanais-nais na mga kadahilanan sa kapaligiran.Ang mga bacterial spores, mahigpit na nagsasalita, ay hindi nagsisilbi para sa pagpaparami, ngunit upang mabuhay sa ilalim ng hindi kanais-nais na mga kondisyon, dahil ang bawat bacterium ay gumagawa lamang ng isang spore. Ang mga bacterial spores ay kabilang sa mga pinaka-lumalaban: halimbawa, madalas silang makatiis ng paggamot na may malalakas na disinfectant at kumukulo sa tubig.
Sa maraming mga halaman, ang proseso ng pagbuo ng spore(sporogenesis) isinasagawa sa mga espesyal na sac-like structure na tinatawag na sporangia. Ang mga spores ay maaaring maging motile dahil sa pagkakaroon ng isang flagellar apparatus (sa kasong ito ay tinatawag sila zoospores), at hindi gumagalaw, pinagkaitan ng kakayahang aktibong gumalaw.
Ang mga zoospores ay sinusunod sa ilang berdeng algae. Sa mga hayop, ang sporulation ay sinusunod sa malarial plasmodium at isang buong grupo ng mga sporozoans (unicellular organisms).
Namumuko. Ito ay matatagpuan kapwa sa mga unicellular na organismo, halimbawa, sa lebadura at ilang mga uri ng ciliates, at sa mga multicellular na organismo, halimbawa, sa mga kinatawan ng uri ng coelenterate (hydra), pati na rin sa mga tunicates (class ascidians).
Para sa mga unicellular na organismo, ang pamamaraang ito ay binubuo sa pagbuo ng isang tubercle (outgrowth) na may nucleus sa mother cell, na pagkatapos ay naghihiwalay at nagiging isang malayang organismo.
Para sa mga multicellular na organismo, ang pamamaraang ito ay binubuo sa katotohanan na unang lumilitaw ang isang maliit na tubercle sa katawan ng indibidwal na ina, na tumataas sa laki, pagkatapos ay lumilitaw ang mga rudiment ng lahat ng mga istraktura at organo na katangian ng organismo ng ina. Pagkatapos ay ang paghihiwalay (budding) ng indibidwal na anak na babae ay nangyayari. Pagkatapos nito, ang bata, bagong hiwalay na organismo ay lumalaki at umabot sa laki ng orihinal na ispesimen.
kanin. Namumuko sa mga coelenterates (hydra), 1 organismong nasa hustong gulang, 2 namumuong organismong anak na babae.
Ang isang hindi pangkaraniwang anyo ng namumuko ay inilarawan sa isang makatas na halaman Bryophyllum - xerophyte, madalas na lumaki bilang isang ornamental houseplant: ang mga maliliit na halaman na nilagyan ng maliliit na ugat ay bubuo sa mga gilid ng mga dahon nito; ang mga "buds" na ito ay tuluyang nalalagas at nagsimulang umiral bilang mga independiyenteng halaman.
Pagkapira-piraso paghahati ng isang indibidwal sa dalawa o higit pang mga bahagi, na ang bawat isa ay bubuo sa isang bagong organismo. Ang pagkapira-piraso ay nangyayari, halimbawa, sa filamentous algae tulad ng Spirogyra. Ang Spirogyra filament ay maaaring masira sa dalawang bahagi sa anumang lugar, kung saan ang dalawang organismo ay kasunod na nabuo.
Ang pagkapira-piraso ay sinusunod din sa ilang mas mababang mga hayop, na, hindi tulad ng mas mataas na organisadong mga anyo, ay nagpapanatili ng isang makabuluhang kakayahan upang muling buuin mula sa medyo mahinang pagkakaiba-iba ng mga selula. Halimbawa, ang katawan ng mga nemertean (isang grupo ng mga primitive worm, pangunahin sa dagat) ay lalong madaling mapunit sa maraming bahagi, na ang bawat isa ay maaaring magbunga ng isang bagong indibidwal bilang resulta ng pagbabagong-buhay. Sa kasong ito, ang pagbabagong-buhay ay isang normal at regulated na proseso; gayunpaman, sa ilang mga hayop (halimbawa, starfish), ang pagpapanumbalik mula sa mga indibidwal na bahagi ay nangyayari lamang pagkatapos ng aksidenteng pagkakapira-piraso. Ang pagkapira-piraso ay sinusunod sa mga espongha, coelenterates (hydras), dikya, echinoderms, annelids at flatworms. Minsan ang kakayahang ito ay napakahusay na binuo na ang isang buong indibidwal ay naibalik mula sa isang hiwalay na fragment.
kanin. . Pagbabagong-buhay ng isang starfish mula sa isang sinag. A, B, C sunud-sunod na yugto ng pagbabagong-buhay
Pagpaparami ng halamanito ay pagpaparami, kung saan ang isang bagong organismo ng anak na babae ay nabubuo mula sa isang bahagi na hiwalay sa organismo ng ina. Sa kasong ito, ang isang medyo mahusay na pagkakaiba-iba na bahagi ay nahiwalay mula sa ina na ispesimen, na higit na bubuo sa isang malayang halaman. O ang halaman ay bumubuo ng mga espesyal na istruktura na partikular na idinisenyo para sa vegetative propagation. Halimbawa, mga bombilya, tubers, corms, rhizomes, tendrils, buds. Ang ilan sa mga istrukturang ito ay nagsisilbi ring mag-imbak ng mga sustansya, na nagpapahintulot sa halaman na mabuhay sa mga panahon ng hindi kanais-nais na mga kondisyon tulad ng malamig o tagtuyot. Ang vegetative propagation ay katangian ng maraming grupo ng mga halaman, mula sa algae hanggang sa mga namumulaklak na halaman.
Sa prinsipyo, ang vegetative propagation ay halos hindi naiiba sa fragmentation o budding, ngunit ayon sa kaugalian, ang terminong ito ay inilalapat sa mga organismo ng halaman at minsan lamang sa mga hayop, kumpara sa fragmentation at budding.
Pag-clone. Ito ay isang artipisyal na paraan ng pagpaparami na hindi natural na nangyayari. Ito ay naging laganap lamang sa nakalipas na 30-40 taon at lalong ginagamit para sa mga layuning pang-ekonomiya. Mayroong ilang mga espesyal na diskarte na nagbibigay-daan sa iyo upang i-clone ang ilang mga halaman at hayop. I-clone ay tumutukoy sa genetically identical na supling na nakuha mula sa isang indibidwal bilang resulta ng isa o ibang paraan ng asexual reproduction. Dito nagmula ang pangalan ng pamamaraang ito.
Ang mga eksperimento sa pag-clone ay lalo na malawak na isinasagawa sa mga halaman, na dahil sa kanilang mataas na kakayahang muling makabuo. Ang mga indibidwal na selula ay inilalagay sa nutrient media kung saan sila ay nahahati at, gamit ang mga espesyal na pamamaraan, ang isang di-organisadong masa ng mga selula ay nakuha, o kalyo. Pagkatapos ay nagiging sanhi sila ng pagkakaiba-iba ng pangunahing homogenous calli at ang pagbuo ng iba't ibang mga tisyu at organo at, sa huli, isang buong organismo ng halaman na may lahat ng parehong mga katangian tulad ng orihinal kung saan kinuha ang mga selula.
Gamit ang cloning method, maaaring makuha ang iba't ibang hybrid form. Kaya, sa tulong ng mga enzyme o ultrasound, ang mga cell wall ng mga cell ng halaman ay tinanggal, pagkatapos nito ang mga nagresultang "hubad" na mga protoplast ay maaaring pagsamahin, na nagreresulta sa mga hybrid na selula (halimbawa, tomato-potato o tobacco-petunia hybrids). Pagkatapos nito, ang mga pader ng cell ay naibalik, nabuo ang callus, at pagkatapos ay isang buong hybrid na halaman.
Sa kaso ng mga hayop, ang sumusunod na pamamaraan ay ginagamit: ang nucleus ng itlog ay tinanggal o nawasak, at ang nucleus ng isang somatic cell (halimbawa, isang epithelial cell) ay inilalagay sa lugar nito. Sa hinaharap, mula sa naturang itlog ay maaaring makuha ang isang organismo na magkapareho sa mga katangian ng donor ng nucleus ng hayop. Sa ganitong paraan, maaari kang makakuha ng mga clone ng ilang mga hayop na clawed palaka(Xenopus), newts (Triturus). Sa kasalukuyan, kahit na ang mga clone ng mga mammal ay nakuha, tulad ng kilalang tupa na si Dolly. Ang pagbuo ng mga diskarte sa pag-clone ay isinasagawa ng isang espesyal na sangay ng biology - biotechnology, ang mga gawain na hindi limitado dito.
Sekswal na pagpaparami
Sekswal na pagpaparamitinatawag na pagbabago ng mga henerasyon at pag-unlad ng mga organismo batay sa pagsasanib ng mga dalubhasang selula ng mikrobyo ( gametes ) at ang pagbuo ng isang zygote. Ang pagbuo ng mga gametes (babae at lalaki, itlog at tamud) ay nangyayari sa mga gonad.
Ang sexual reproduction ay tipikal para sa karamihan ng mga nabubuhay na nilalang, ay mas progresibo kumpara sa asexual reproduction at may napakalaking genetic advantages. Pinakamainam na tinitiyak ng sexual reproduction ang genetic diversity ng supling, dahil mayroong kumbinasyon ng mga gene na dating pag-aari ng parehong mga magulang. Ang pagkakaiba-iba ng mga genotype ng mga indibidwal na bumubuo sa mga species ay nagbibigay ng posibilidad ng mas matagumpay at mabilis na pagbagay ng mga species sa pagbabago ng mga kondisyon sa kapaligiran.
Sa panahon ng pagpapabunga, ang mga gametes ay nagsasama upang bumuo ng isang diploid zygote mula sa kung saan, sa proseso ng pag-unlad, ang isang mature na organismo ay nakuha. Ang mga gametes ay haploid - naglalaman sila ng isang hanay ng mga kromosom, ang zygote ay diploid, na naglalaman ng isang dobleng hanay ng mga kromosom. Ito ang unang cell ng hinaharap na organismo.
Ang mga gametes ay karaniwang may dalawang uri - lalaki at babae. Ang mga ito ay ginawa ng mga magulang na lalaki at babae kung ang species ay dioecious; ( tulad ng ilang namumulaklak na halaman, karamihan sa mga hayop at tao) o ng parehong indibidwal (harmaphroditism).
Katangian para sa lalaki at babaeng hayopsekswal na dimorphism- ibig sabihin. mga pagkakaiba sa sekswal sa istraktura, hitsura (laki, kulay at iba pang mga katangian), pati na rin sa pag-uugali. Sa mga hayop, nangyayari na ito sa mas mababang mga yugto ng pag-unlad ng ebolusyon, halimbawa, sa mga bilog na helminth at arthropod, at naabot ang pinakadakilang pagpapahayag nito sa mga vertebrates, kung saan ang mga panlabas na pagkakaiba sa pagitan ng mga lalaki at babae ay napaka nagpapahayag. Sa mga halaman ng mga species na nailalarawan sa pagkakaroon ng mga lalaki at babae na indibidwal, nangyayari din ang sekswal na dimorphism, ngunit ito ay napakaliit na ipinahayag.
Ang paghahambing ng sekswal at asexual na pagpaparami ay ibinigay sa talahanayan. 5
mesa 5
Paghahambing ng asexual at sekswal na pagpaparami
|
Asexual reproduction |
Sekswal na pagpaparami (hindi kasama ang bacteria) |
|
|
Isang magulang |
Kadalasan dalawang magulang |
|
|
Ang mga gametes ay hindi nabuo |
Ang mga haploid gametes ay nabuo, ang nuclei kung saan nagsasama (fertilization) upang bumuo ng isang diploid zygote |
|
|
Walang meiosis |
Sa ilang yugto ng siklo ng buhay, nangyayari ang meiosis, na pumipigil sa pagdoble ng mga kromosom sa bawat henerasyon. |
|
|
Ang mga supling ay magkapareho sa kanilang mga magulang. Ang tanging pinagmumulan ng genetic variation ay random mutations |
Ang mga inapo ay hindi magkapareho mga indibidwal ng magulang. Nagpapakita sila ng genetic variation na nagreresulta mula sa genetic recombination. Itinataguyod nito ang natural na pagpili ng pinakamalakas at pinakamatibay na indibidwal, at, dahil dito, ebolusyon. |
|
|
Katangian ng mga halaman, ilang mas mababang hayop at mikroorganismo. Hindi matatagpuan sa matataas na hayop |
Katangian ng karamihan sa mga halaman at hayop |
|
|
Kadalasan ay nagreresulta sa mabilis na paglikha ng isang malaking bilang ng mga inapo |
Mas mabilis na pagtaas ng bilang |
Ang sekswal na pagpaparami, depende sa ratio ng mga selula ng mikrobyo sa laki at paggana, ay maaaring nasa tatlong opsyon:
Isogamy (algae, protozoa). Ang mga reproductive cell ng lalaki at babae ay magkapareho sa laki, istraktura at kadaliang kumilos.
Heterogamy. Ang mga cell ay naiiba sa laki at istraktura.
Oogamy. Ito ay isa sa mga variant ng heterogamy, kapag ang itlog ay malaki at hindi kumikibo, at ang tamud ay mas maliit sa laki, may isang organ ng paggalaw at may kadaliang kumilos.
Mayroong mga espesyal na anyo ng sekswal na pagpaparami, tulad ng hermaphroditism at parthenogenesis.
Hermaphroditism. Ang terminong "hermaphroditism" ay kumbinasyon ng mga pangalang Griyego na Hermes (diyos ng kagandahan ng lalaki) at Aphrodite (diyosa ng kagandahang babae).
Gayunpaman, sa karamihan ng mga hermaphroditic species, ang pagpapabunga ay kinabibilangan ng mga gametes na nagmula sa iba't ibang indibidwal, at mayroon silang maraming genetic, morphological at physiological adaptations na pumipigil sa self-fertilization at pinapaboran ang cross-fertilization. Halimbawa, sa maraming protozoa ang self-fertilization ay pinipigilan ng genetic incompatibility, sa maraming namumulaklak na halaman sa pamamagitan ng istraktura ng androecium at gynoecium, at sa maraming hayop sa pamamagitan ng katotohanan na ang mga itlog at tamud ay nabuo sa parehong indibidwal sa iba't ibang oras.
Ang kababalaghan ng tunay na hermaphroditism ay matatagpuan din sa mas organisadong mga nilalang. Sa partikular, ito ay matatagpuan sa mga mammal. Halimbawa, sa mga baboy, ang pag-unlad ng mga ovary ay minsan ay sinusunod sa isang bahagi ng katawan, at ang pagbuo ng mga testes (testes) sa kabilang banda, o ang pagbuo ng pinagsamang mga istraktura (ovotestis), at sa parehong mga kaso ang synthesis ng functionally ang mga aktibong itlog at tamud ay nagaganap. Ang mga naturang hayop ay inuri bilang isang "intermediate" na kasarian, na ang karamihan ng mga indibidwal ng intermediate na uri ng sekswal ay mga babae na may dalawang XX chromosome. Ang isang katulad na kababalaghan ay nabanggit sa mga kambing.
Ang tunay na hermaphroditism ay nangyayari rin sa mga tao, na nagreresulta mula sa mga karamdaman sa pag-unlad. Ang mga genotype ng hermaphrodites ay 46XX o 46 XY , na karamihan sa mga kaso ay tumutukoy sa XX (mga 60%). Ang maling hermaphroditism ay kilala rin, kapag ang mga indibidwal ay may panlabas na ari at pangalawang sekswal na katangian na katangian ng parehong kasarian, ngunit gumagawa ng mga selulang mikrobyo ng isang uri lamang - lalaki o babae.
Karamihan sa mga namumulaklak na halaman ay may mga hermaphroditic na bulaklak, na karaniwang tinatawag na bisexual dahil ang bawat bulaklak ay naglalaman ng pistil at stamens. Para sa kadahilanang ito, ang mga prutas ay nabuo mula sa lahat ng mga bulaklak. Ang trigo, cherry, mansanas at marami pang ibang uri ng halaman ay bisexual. Bilang karagdagan sa mga bisexual, sa panahon ng ebolusyon, ang mga halaman na may paghihiwalay ng mga kasarian sa loob ng parehong species ay nabuo, ibig sabihin, ang monoecious at dioecy ng mga halaman ay lumitaw. Ang mga halaman na naglalaman ng parehong pistillate (babae) at staminate (lalaki) na mga bulaklak ay tinatawag na monoecious. Sa mga monoecious na halaman, ang mga prutas ay bubuo lamang mula sa mga bulaklak ng pistillate. Ang mais ay monoeciouspipino, kalabasa at iba pa. Sa kaibahan, ang mga dioecious na halaman ay mga halaman na naglalaman ng alinman sa pistillate o staminate na mga bulaklak (sa loob ng parehong species). Sa mga dioecious na halaman, ang mga may pistillate na bulaklak (mga babae) lamang ang namumunga. Ang poplar, strawberry at iba pang uri ng makahoy at mala-damo na halaman ay dioecious.
Parthenogenesis (mula sa Greek. parthenos birhen at genesis kapanganakan) ay isa sa mga pagbabago ng sekswal na pagpaparami kung saan ang babaeng gamete ay nabubuo sa isang bagong indibidwal na walang pagpapabunga ng male gamete. Ang bentahe ng parthenogenesis ay na sa ilang mga kaso pinatataas nito ang rate ng pagpaparami, at sa mga social na insekto pinapayagan ka nitong ayusin ang bilang ng mga inapo ng bawat uri. Ang parthenogenesis ay maaaring obligado (obligado) at facultative (opsyonal).
Halimbawa, sa honey bee(Apis mellifera) ang reyna ay nangingitlog ng fertilized(2п = 32), na, kapag umuunlad, ay nagbubunga ng mga babae (mga reyna o manggagawa), at mga hindi pa nabubuong itlog(P = 16), na gumagawa ng mga lalaki (drone) na gumagawa ng spermine sa pamamagitan ng mitosis sa halip na meiosis.
kanin. Scheme ng mga uri ng parthenogenesis.
- Ikot ng cell. Mitosis. Meiosis
Ang paghahati ng cell ay ang batayan ng pagpaparami at indibidwal na pag-unlad ng mga organismo.
Ang lahat ng nabubuhay na organismo ay binubuo ng mga selula. Ang pag-unlad, paglaki, at pagbuo ng tipikal na istraktura ng katawan ay isinasagawa sa pamamagitan ng pagpaparami ng isa o isang grupo ng mga orihinal na selula. Sa proseso ng buhay, ang ilan sa mga selula ng katawan ay nawawala, tumatanda at namamatay. Upang mapanatili ang istraktura at normal na paggana, ang katawan ay dapat gumawa ng mga bagong selula upang palitan ang mga luma. Ang tanging paraan upang bumuo ng mga cell ay sa pamamagitan ng paghahati sa mga nauna.
Cell division isang mahalagang proseso para sa lahat ng mga organismo. Sa katawan ng tao, na binubuo ng humigit-kumulang 10 13 mga cell, ilang milyon sa kanila ang dapat hatiin bawat segundo.
Tatlong paraan ng paghahati ng mga eukaryotic cell ay inilarawan: amitosis (direktang paghahati), mitosis (hindi direktang paghahati) at meiosis (pagbawas dibisyon).
Amitosis isang medyo bihira at hindi gaanong pinag-aralan na paraan ng paghahati ng cell. Ito ay inilarawan para sa pagtanda at pathologically altered na mga cell. Sa amitosis, ang interphase nucleus ay nahahati sa pamamagitan ng constriction, at ang pare-parehong pamamahagi ng namamana na materyal ay hindi natiyak. Kadalasan ang nucleus ay nahahati nang walang kasunod na paghihiwalay ng cytoplasm at ang mga selulang binucleate ay nabuo. Ang isang cell na sumailalim sa amitosis ay pagkatapos ay hindi makapasok sa normal na mitotic cycle. Samakatuwid, ang amitosis ay nangyayari, bilang isang panuntunan, sa mga selula at mga tisyu na napapahamak sa kamatayan, halimbawa, sa mga selula ng mga embryonic lamad ng mga mammal at sa mga selula ng tumor.
Mitosis isang unibersal na paraan ng paghahati ng mga eukaryotic cell. Ang tagal nito sa mga selula ng hayop ay humigit-kumulang 1 oras. Ang mitosis ay isang tuluy-tuloy na proseso, na karaniwang nahahati sa apat na yugto: prophase, metaphase, anaphase at telophase.
Ang pagkakasunud-sunod ng mga kaganapan na nangyayari sa pagitan ng pagbuo ng isang naibigay na cell at ang paghahati nito sa mga anak na selula ay tinatawagsiklo ng cell. Ang siklo na ito ay binubuo ng tatlong pangunahing yugto:
1. Interphase. Isang panahon ng matinding synthesis atpaglago. Ang cell ay nag-synthesize ng maraming mga sangkap na kinakailangan para sa paglaki nito at ang pagpapatupad ng lahat ng mga likas na pag-andar nito. Sa panahon ng interphase, nangyayari ang pagtitiklop ng DNA.
2. Mitosis. Ito ang proseso ng nuclear division (karyokinesis), kung saan ang mga chromatid ay pinaghihiwalay sa isa't isa at muling ipinamahagi bilang mga chromosome sa pagitan ng mga daughter cell.
3. Ang cytokinesis ay ang proseso ng paghahati ng cytoplasm (cytokinesis) sa pagitan ng dalawang anak na selula.
Ang haba ng cell cycle ay depende sa uri ng cell at sa panlabas na mga kadahilanan tulad ng temperatura, nutrients at oxygen. Ang mga bacterial cell ay maaaring hatiin tuwing 20 minuto, bituka epithelial cell - bawat 8-10, mga cell sa dulo ng isang ugat ng sibuyas - bawat 20 oras, at maraming mga cell ng nervous system ay hindi kailanman nahahati.
Ang ugnayan sa pagitan ng mga proseso sa paglipas ng panahon ay ipinapakita sa kanin.

kanin . Mga yugto ng cell cycle
Interphase ay binubuo ng ilang mga panahon: G 1, S, G 2.
Panahon G 1 tinatawag na presynthetic. Karamihan sa variable sa tagal. Sa oras na ito, ang mga proseso ng biological synthesis ay isinaaktibo sa cell, pangunahin ang istruktura at functional na mga protina. Ang cell ay lumalaki at naghahanda para sa susunod na panahon. Sa panahong ito, nagaganap ang masinsinang mga proseso ng biosynthesis. Ang pagbuo ng mitochondria, chloroplast (sa mga halaman), endoplasmic reticulum, lysosomes, Golgi apparatus, vacuoles at vesicle. Ang nucleolus ay gumagawa ng rRNA, mRNA at tRNA; nabuo ang mga ribosom; ang cell ay synthesizes structural at functional na mga protina. Matinding cellular metabolism na kinokontrol ng mga enzyme. Paglago ng cell. Ang pagbuo ng mga sangkap na pumipigil o nagpapasigla sa pagsisimula ng susunod na yugto.
Panahon S tinatawag na synthetic. Ito ang pangunahing panahon sa mitotic cycle. Sa paghahati ng mga selula ng mammalian, tumatagal ito ng mga 6 x 10 oras. Dito nangyayari ang pagtitiklop ng DNA. Synthesis ng mga molekula ng protina na tinatawag na histones, na nagbubuklod sa bawat strand ng DNA. Ang bawat chromosome ay nagiging dalawang chromatid.
Ang G2 period ay tinatawag postsynthetic.Ito ay medyo maikli, sa mga selula ng mammalian ito ay mga 2 x 5 na oras. Sa oras na ito, ang bilang ng mga centrioles, mitochondria at plastids ay doble, ang mga aktibong proseso ng metabolic ay nagaganap, ang mga protina at enerhiya ay naipon para sa paparating na dibisyon. Nagsisimulang hatiin ang selula. Nangyayari ang masinsinang proseso ng biosynthesis. Dibisyon ng mitochondria at chloroplast. Nadagdagang reserbang enerhiya. Pagtitiklop ng centrioles (sa mga cell kung saan sila naroroon) at ang simula ng pagbuo ng spindle
Mitosis kondisyon na nahahati sa apat na yugto: prophase, metaphase, anaphase at telophase
Prophase . Nagsisimula ang spiralization ng DNA at unti-unting tumataas sa nucleus. Ang mga chromosome ay umiikli, lumalapot, nagiging nakikita, at nakakakuha ng isang tipikal na istraktura ng bichromatid. Unti-unting nawawala ang nucleolus. Sa cytoplasm, ang mga microtubule ay nakatuon sa paligid ng bawat pares ng centrioles, na bumubuo ng mga sentro ng spindle. Ang mga centriole ay lumipat sa iba't ibang mga pole, ang mga microtubule ay umaabot sa kahabaan ng axis ng cell, at nagsisimula ang pagbuo ng isang achromatin spindle. Ang nuklear na sobre ay nahati sa magkakahiwalay na maliliit na fragment. Ang mga chromosome ay lumilipat patungo sa gitna ng cell
Metaphase . Ang mga chromosome ay maximally spiralized at nakaayos sa paraang ang kanilang mga centromeres ay nasa parehong eroplano - ang eroplano ng cell equator. Ang isang metaphase plate ay nabuo. Ang pagbuo ng mitotic spindle ay nakumpleto. Ang mga centriole ay matatagpuan sa magkapares sa magkasalungat na mga pole, at ang mga spindle thread mula sa iba't ibang mga pole ay nakakabit sa centromere ng bawat chromosome.
Anaphase . Ito ang pinakamaikling yugto ng mitosis. Dito, nangyayari ang longitudinal splitting ng bawat chromosome, ang pagbabawas ng mga threadspindles at divergence ng chromatids (daughter chromosomes) patungo sa mga pole ng cell.
Telofase . Ang mga chromosome ng anak na babae, na binubuo ng isang chromatid, ay umaabot sa mga pole ng cell. Ang DNA na bumubuo sa mga ito ay nagsisimula sa despiral, lumilitaw ang isang nucleolus, isang nuclear membrane ay nabuo sa paligid ng bawat pangkat ng mga anak na chromosome, at ang mga thread ng achromatin spindle ay unti-unting nawasak. Nakumpleto ang nuclear fission.
Nagsisimula ang cytoplasmic division(cytotomy) at ang pagbuo ng isang septum sa pagitan ng mga cell ng anak na babae. Ang mga selula ng hayop ay nagsasagawa ng cytotomy sa pamamagitan ng paghihigpit ng cytoplasmic membrane. Sa mga halaman, ang isang membrane septum ay nabuo sa eroplano ng cell equator, na lumalaki sa gilid, na umaabot sa cell wall. Bilang isang resulta, dalawang ganap na pinaghiwalay na mga cell ng anak na babae ay nabuo.
Subaybayan natin pagbabago sa namamana na materyalsa panahon ng mitotic cycle. Ang mga pangunahing kaganapan ng mitotic cycle ay Pagtitiklop ng DNA nagaganap sa interphase at humahantong sa pagdodoble ng dami ng namamana na impormasyon, atchromatid segregation,nagaganap sa anaphase ng mitosis at tinitiyak ang pare-parehong pamamahagi ng namamana na impormasyon sa pagitan ng mga cell ng anak na babae. Isinasagawa ng namamanang materyal ang mga prosesong ito habang nasa iba't ibang anyo ng istruktura. Sumasailalim ang replicative synthesisinterphase chromatin, inkung saan ang molekula ng DNA ay nasa medyo despiralized na estado. Ang pamamahagi ng genetic na impormasyon ay isinasagawamitotic chromosome,kung saan ang DNA ay maximally spiralized.
Sa mitotic cycle, nagbabago rin ang dami ng hereditary material. Kung ang bilang ng mga chromosome sa isang haploid set ay tinutukoy ng titik p (sa isang diploid set, ayon sa pagkakabanggit, 2n), at ang bilang ng mga molekula ng DNA ay tinutukoy ng titik kasama, pagkatapos ay posible na masubaybayan ang pagbabago sa formula ng nucleus ng isang somatic cell sa iba't ibang yugto ng mitotic cycle. dati S -panahon kung kailan ang bawat chromosome ay binubuo ng isang molekula ng DNA, ang kabuuang dami ng DNA sa nucleus ay tumutugma sa bilang ng mga chromosome sa loob nito, at ang formula ng isang diploid cell ay may anyo 2p2s. Pagkatapos ng pagtitiklop, kapag ang DNA ng bawat chromosome ay nagdodoble sa sarili nito, ang kabuuang dami ng DNA sa nucleus ay nagdodoble at ang cell formula ay nasa anyo na 2n4c. Bilang resulta ng paghihiwalay ng chromatid sa anaphase ng mitosis, ang nuclei ng anak na babae ay tumatanggap ng isang diploid na hanay ng mga single-chromatid chromosome. Nagiging muli ang formula ng mga daughter cell 2p2s.
Biological na kahalagahan ng mitosisay bilang isang resulta ng pamamaraang ito ng paghahati, ang mga cell ay nabuo na may namamana na impormasyon na qualitatively at quantitatively magkapareho sa impormasyon ng mother cell. Ang pare-parehong pamamahagi ng namamana na materyal ay tinitiyak ng mga proseso ng pagtitiklop ng DNA at pagdodoble ng chromosome sa interphase ng mitotic cycle, pati na rin sa pamamagitan ng spiralization at pare-parehong pamamahagi ng mga chromatids sa pagitan ng mga cell ng anak na babae sa panahon ng mitosis. Tinitiyak ng Mitosis ang pagpapanatili ng constancy ng karyotype sa isang bilang ng mga henerasyon ng cell at nagsisilbing isang mekanismo ng cellular para sa mga proseso ng paglaki at pag-unlad ng katawan, pati na rin ang pagbabagong-buhay at asexual reproduction.
Ang pagkilos ng isang bilang ng mga kadahilanan sa kapaligiran ay maaaring makagambala sa normal na kurso ng mitosis at humantong sa pinsala sa mga chromosome, gayundin sa mga pagbabago sa bilang ng mga indibidwal na chromosome o buong set ng chromosome sa mga somatic cells ng katawan. Ang mga pathological mitoses ay maaaring maging sanhi ng isang bilang ng mga chromosomal na sakit. Ang mga pathological mitoses ay sinusunod lalo na madalas sa mga selula ng tumor.
Diagram ng mitosis:
MITOSIS
|
1. Interphase Madalas itong maling tinatawag na yugto ng pahinga. Ang tagal ng interphase ay nag-iiba at depende sa paggana ng isang naibigay na cell. Ito ang panahon kung saan ang cell ay karaniwang nag-synthesize ng mga organelles at lumalaki ang laki. Ang nucleoli ay malinaw na nakikita at aktibong synthesize ang ribosomal na materyal. Bago ang paghahati ng cell, ang DNA at mga histone ng bawat chromosome ay ginagaya. Ang bawat chromosome ay kinakatawan na ngayon ng isang pares ng chromatids na konektado sa isa't isa ng isang centromere. Ang sangkap ng mga chromosome ay may mantsa at tinatawag na chromatin, ngunit ang mga istrukturang ito mismo ay mahirap makita. |
|
|
2. Prophase Karaniwan ang pinakamahabang yugto ng paghahati ng cell. Ang mga Chromatid ay umiikli (hanggang 4% ng kanilang orihinal na haba) at lumapot bilang resulta ng kanilang spiralization at condensation. Kapag nabahiran, ang mga chromatid ay malinaw na nakikita, ngunit ang mga sentromere ay hindi nakikita. Sa iba't ibang mga pares ng chromatids, ang sentromere ay matatagpuan sa ibang paraan. Sa mga selula ng hayop at sa mas mababang mga halaman, ang mga centriole ay naghihiwalay sa magkasalungat na mga poste ng selula. Mula sa bawat centriole, ang mga maikling microtubule ay nagmumula sa anyo ng mga sinag, na sama-samang bumubuo ng isang bituin. Ang nucleoli ay nagiging mas maliit dahil ang kanilang nucleic acid ay bahagyang inilipat sa ilang mga pares ng chromatids. Sa pagtatapos ng prophase, ang nuclear membrane ay naghiwa-hiwalay at isang fission spindle ay nabuo. |
|
|
3. Metaphase Ang mga pares ng chromatids ay ikinakabit ng kanilang mga sentromer sa mga filament ng spindle (microtubules) at gumagalaw pataas at pababa sa spindle hanggang sa pumila ang kanilang mga centromeres sa kahabaan ng ekwador ng spindle na patayo sa axis nito. |
|
|
4. Anaphase Ito ay isang napakaikling yugto. Ang bawat centromere ay nahahati sa dalawa, at ang mga filament ng spindle ay hinihila ang anak na babae na sentromere sa magkabilang poste. Hinihila ng mga centromeres sa likod nila ang mga chromatid na humiwalay sa isa't isa, na tinatawag na ngayong mga chromosome. |
|
|
5. Telofase Ang mga kromosom ay umaabot sa mga cell pole, despiral, humahaba, at hindi na sila malinaw na makilala. Ang mga filament ng spindle ay nawasak at ang mga centriole ay ginagaya. Ang isang nuclear membrane ay nabuo sa paligid ng mga chromosome sa bawat poste. Muling lumitaw ang nucleolus. Ang Telophase ay maaaring agad na sundan ng cytokinesis (dibisyon ng buong cell sa dalawa). |
|





Meiosis (mula sa Greek meiosis pagbabawas) isang kakaibang paraan ng paghahati ng cell, na humahantong sa isang paghahati ng bilang ng mga chromosome sa kanila. Ang Meiosis ay ang gitnang link gametogenesis sa mga hayop at sporogenesis sa mga halaman. Binubuo ang Meiosis ng dalawang magkakasunod na dibisyon na pinangungunahan ng isang solong reduplication ng DNA. Ang lahat ng mga sangkap at enerhiya na kinakailangan para sa parehong mga dibisyon ay naka-imbak sa panahon ng interphase bago ang meiosis ako. Interphase II halos wala, at ang mga dibisyon ay mabilis na sumusunod sa isa't isa. Sa bawat isa sa mga meiotic division, ang parehong apat na yugto ay nakikilala: prophase, metaphase, anaphase at telophase, na katangian ng mitosis, ngunit naiiba sa isang bilang ng mga tampok.
Unang meiotic division (meiosis ako ) ay humahantong sa paghati ng bilang ng mga kromosom at tinatawag na pagbabawas. Bilang resulta, mula sa isang diploid cell(2p 4c) dalawang haploid cells ang nabuo(p 2c) mga cell.
Prophase I Ang meiosis ang pinakamahaba at pinakamasalimuot. Bilang karagdagan sa mga proseso ng DNA helixation at spindle formation na tipikal ng prophase ng mitosis, ako Dalawang napakahalagang biological na kaganapan ang nagaganap: banghay o synapsis homologous chromosome at tumatawid.
Ang conjugation ay ang proseso ng pagkakalapit ng mga homologous chromosome. Nabubuo ang mga magkapares na chromosome na ito bivalent at pinananatili sa komposisyon nito sa tulong ng mga espesyal na protina. Dahil ang bawat chromosome ay binubuo ng dalawang chromatids, ang isang bivalent ay kinabibilangan ng apat na chromatids at tinatawag ding kuwaderno. Gumagawa ang isang diploid cell P bivalents. Pagkatapos ng conjugation, ang cell formula ay kumukuha ng form pumasa.
Sa ilang mga lugar ng bivalent, ang mga chromatids ng conjugated chromosome ay nagsalubong, nasira, at nagpapalitan ng kaukulang mga seksyon. Ang prosesong ito ng pagpapalitan ng mga fragment ng homologous chromosome ay tinatawag na crossing over. Tinitiyak nito ang pagbuo ng mga bagong kumbinasyon ng paternal at maternal genes sa mga chromosome ng hinaharap na gametes. Maaaring mangyari ang pagtawid sa ilang lugar (multiple crossing over), na nagbibigay ng mas mataas na antas ng recombination ng namamana na impormasyon sa mga gametes. Sa pagtatapos ng prophase ako ang antas ng chromosome spiralization ay tumataas, ang mga chromatids ay nagiging malinaw na nakikilala, ang mga spindle thread mula sa bawat poste ay nakakabit sa centromere ng isa sa mga bivalent chromosome. Ang nuclear envelope ay nawasak, at ang bivalents ay nakadirekta patungo sa equatorial plane ng cell.
Sa metaphase I Kinukumpleto ng Meiosis ang pagbuo ng spindle, ang mga bivalents ay naka-install sa equatorial plane ng cell. Ang mga spindle strands mula sa isang poste ay nakakabit sa centromere ng bawat chromosome.
Sa anaphase I Sa meiosis, sa ilalim ng pagkilos ng mga spindle thread, ang mga homologous chromosome ay lumayo sa isa't isa, patungo sa magkasalungat na pole ng cell. Bilang resulta, sa bawat poste ng cell ahaploid setchromosome, na naglalaman ng isang bichromatid chromosome mula sa bawat pares ng homologous chromosome. Sa anaphase ako chromosome ng iba't ibang pares, i.e. Ang mga di-homologous na chromosome ay ganap na kumikilos nang hiwalay sa isa't isa, na tinitiyak ang pagbuo ng isang malawak na iba't ibang uri ng mga kumbinasyon paternal at maternal chromosome sa haploid set ng hinaharap na gametes. Ang bilang ng mga naturang kumbinasyon ay tumutugma sa formula 2 P, kung saan p bilang ng mga pares ng homologous chromosome. Sa mga tao, ang halagang ito ay katumbas ng 2, i.e. 8.4 10 mga variant ng mga kumbinasyon ng paternal at maternal chromosome ay posible sa mga gametes ng tao.
Kaya, ang pagkakaiba-iba ng mga homologous chromosome sa anaphase ako Tinitiyak ng meiosis hindi lamang ang pagbawas sa bilang ng mga chromosome sa hinaharap na mga selula ng mikrobyo, kundi pati na rin ang malaking pagkakaiba-iba ng huli dahil sa random na kumbinasyon ng mga chromosome ng ama at ina ng iba't ibang mga pares.
Sa telophase I Ang Meiosis ay nangyayari sa pagbuo ng mga selula na ang nuclei ay may haploid na hanay ng mga chromosome at doble ang dami ng DNA, dahil ang bawat chromosome ay binubuo ng dalawang chromatids. Ang mga cell na nagreresulta mula sa unang meiotic division ay may formula p2s at pagkatapos ng isang maikling interphase ay sinimulan nila ang susunod na dibisyon.
Pangalawang meiotic division (meiosis II ) nagpapatuloy bilang isang tipikal na mitosis (Larawan 5.3), ngunit naiiba dahil ang mga selulang pumapasok dito ay naglalaman ng isang haploid na hanay ng mga kromosom. Bilang resulta ng dibisyong ito P bichromatid chromosomes (r2c), paghahati, anyo P iisang chromatid chromosomes(ps). Ang dibisyong ito ay tinatawag na equational (o pagpantay-pantay).
Kaya, pagkatapos ng dalawang sunud-sunod na meiotic division, mula sa isang cell na may diploid set ng two-chromatid chromosome (2x4c), apat na cell na may haploid set ng single-chromatid chromosome ay nabuo.(ps).
Biological na kahalagahan ng meiosisBinubuo ang pagbuo ng mga cell na may pinababang hanay ng mga chromosome at pagpapanatili ng constancy ng karyotype sa isang bilang ng mga henerasyon ng mga organismo na nagpaparami. sexually transmitted. Ang Meiosis ay nagsisilbing batayan para sa combinative variability, na nagbibigay ng genetic diversity ng gametes sa pamamagitan ng mga proseso ng crossing over, divergence at combinatorics ng paternal at maternal chromosomes. Mga pagbabago sa istruktura ng chromosome dahil sa hindi pantay na pagtawid, pagkagambala sa pagkakaiba-iba ng lahat o indibidwal na chromosome sa anaphase I at II Ang mga meiotic division ay humahantong sa pagbuo ng mga abnormal na gametes at maaaring magsilbing batayan para sa pagkamatay ng organismo o pagbuo ng isang bilang ng mga chromosomal syndrome sa mga inapo.
Diagram at maikling paglalarawan ng sunud-sunod na yugto ng meiosis sa isang selula ng hayop.
|
1. Interphase Ang tagal ay nag-iiba sa iba't ibang species. Nagaganap ang pagtitiklop ng organelle at lumalaki ang laki ng cell. Ang pagtitiklop ng DNA at mga histone ay pangunahing nagtatapos sa premeiotic interphase, ngunit bahagyang nagsasangkot din ng simula ng prophase. Ang bawat chromosome ay kinakatawan na ngayon ng isang pares ng chromatids na konektado ng isang centromere. Ang materyal na chromosomal ay nabahiran, ngunit sa lahat ng mga istruktura ay ang nucleoli lamang ang malinaw na nakikita. |
A. Maagang prophase ako |
|
2. Prophase Ang pinakamahabang yugto. Ito ay madalas na nahahati sa limang yugto (leptotema, zygonema, pachynema, diplonema at diakinesis), ngunit dito ito ay ituturing bilang isang tuluy-tuloy na pagkakasunud-sunod ng mga pagbabago sa chromosome. A . Ang mga chromosome ay umiikli at nagiging nakikita bilang magkahiwalay na mga istraktura. Sa ilang mga organismo, ang mga ito ay mukhang mga string ng mga kuwintas: mga lugar ng matinding mantsang materyal - chromomeres - kahalili ng mga hindi nabahiran na lugar. Ang mga Chromomere ay ang mga lugar kung saan ang materyal na chromosomal ay lubos na nakapulupot. |
|
|
B. Ang mga homologous chromosome, na nagmula sa nuclei ng maternal at paternal gametes, ay lumalapit sa isa't isa at conjugate. Ang mga chromosome na ito ay magkapareho ang haba, ang kanilang mga sentromere ay sumasakop sa parehong posisyon, at sila ay karaniwang naglalaman ng parehong bilang ng mga gene na nakaayos sa parehong linear sequence. Ang chromomeres ng homologous chromosome ay magkatabi. Ang proseso ng conjugation ay maaaring magsimula sa ilang mga punto sa mga chromosome, na pagkatapos ay konektado sa buong haba (na parang naka-zip na magkasama). Ang mga pares ng conjugated homologous chromosome ay madalas na tinatawag na bivalents. Ang bivalents ay umiikli at lumapot. Sa kasong ito, ang parehong mas siksik na pag-iimpake ay nangyayari sa antas ng molekular at panlabas na kapansin-pansin na pag-twist (spiralization). Ngayon ang bawat chromosome na may sentromere nito ay malinaw na nakikita. |
B. Prophase I |
|
SA. Ang mga homologous chromosome na bumubuo sa bivalent ay bahagyang pinaghihiwalay, na parang nagtutulak palayo sa isa't isa. Ngayon ay makikita mo na ang bawat chromosome ay binubuo ng dalawang chromatids. Ang mga chromosome ay konektado pa rin sa isa't isa sa ilang mga punto. Ang mga puntong ito ay tinatawag na chiasmata (mula sa Griyego. chiasma - krus). Sa bawat chiasma, ang mga seksyon ng chromatids ay ipinagpapalit bilang resulta ng mga break at reunion, kung saan ang dalawa sa apat na thread na nasa bawat chiasm ay kasangkot. Bilang resulta, ang mga gene mula sa isang chromosome (halimbawa, paternal - A, B, C) ay nauugnay sa mga gene mula sa isa pang chromosome (maternal a, b , c), na humahantong sa mga bagong kumbinasyon ng gene sa mga nagresultang chromatids. Ang prosesong ito ay tinatawag na crossing over. Ang mga homologous chromosome ay hindi naghihiwalay pagkatapos tumawid, dahil ang mga kapatid na chromatids (ng magkabilang chromosome) ay nananatiling matatag na konektado hanggang sa anaphase. |
B. Pagtawid sa panahon ng prophase ako |
|
G . Ang mga chromatid ng mga homologous chromosome ay patuloy na nagtataboy sa isa't isa, at ang mga bivalents ay nakakakuha ng isang tiyak na pagsasaayos depende sa bilang ng chiasmata. Ang mga bivalents na may isang chiasmata ay may hugis cruciform, na may dalawang chiasmata na hugis singsing, at may tatlo o higit pang mga loop na nakahiga patayo sa isa't isa. Sa pagtatapos ng prophase, ang lahat ng mga chromosome ay ganap na condensed at matinding mantsa. Ang iba pang mga pagbabago ay nangyayari sa cell: paglipat ng mga centrioles (kung mayroon man) sa mga pole, pagkasira ng nucleoli at nuclear membrane, at pagkatapos ay ang pagbuo ng mga spindle filament. |
|
|
2. Metaphase Ang mga bivalents ay pumila sa equatorial plane, na bumubuo ng isang metaphase plate. Ang kanilang mga centromeres ay kumikilos bilang mga solong istruktura (bagaman madalas silang lumilitaw na doble) at nag-aayos ng mga spindle filament na nakakabit sa kanila, na ang bawat isa ay nakadirekta sa isa lamang sa mga pole. Bilang resulta ng mahinang puwersa ng paghila ng mga sinulid na ito, ang bawat bivalent ay matatagpuan sa rehiyon ng ekwador, at ang parehong mga sentromere nito ay nasa parehong distansya mula dito, ang isa sa ibaba at ang isa sa itaas. |
D. Late metaphase ako |
|
3. Anaphase Ang dalawang sentromer na nasa bawat bivalent ay hindi pa nahahati, ngunit ang mga kapatid na chromatids ay hindi na magkatabi. Hinihila ng mga filament ng spindle ang mga sentromere, na ang bawat isa ay nauugnay sa dalawang chromatids, patungo sa magkabilang poste ng spindle. Bilang resulta, ang mga chromosome ay nahahati sa dalawang haploid set na napupunta sa mga anak na selula. |
E. Anaphase I |
|
4. Telofase Ang divergence ng mga homologous centromeres at mga nauugnay na chromatids sa magkasalungat na pole ay nangangahulugan ng pagkumpleto ng unang meiotic division. Ang bilang ng mga chromosome sa isang set ay naging kalahati ng laki, ngunit ang mga chromosome sa bawat poste ay binubuo ng dalawang chromatids. Dahil sa pagtawid sa panahon ng pagbuo ng chiasmata, ang mga chromatid na ito ay genetically non-identical, at sa ikalawang meiotic division ay maghihiwalay sila. Ang mga spindle at ang kanilang mga sinulid ay karaniwang nawawala. Sa mga hayop at ilang halaman, ang mga chromatids ay despiral, isang nuclear membrane ang bumubuo sa kanilang paligid sa bawat poste, at ang resultang nucleus ay pumapasok sa interphase. Pagkatapos ang paghahati ng cytoplasm ay nagsisimula (sa mga hayop) o ang pagbuo ng isang naghahati na pader ng cell (sa mga halaman), tulad ng sa mitosis. Sa maraming halaman, walang telophase, o cell wall formation, o interphase na sinusunod, at ang cell ay direktang lumilipat mula sa anaphase. I para manghula II. |
G. Telofase I sa isang selula ng hayop |







|
Interphase II Ang yugtong ito ay karaniwang sinusunod lamang sa mga selula ng hayop: ang tagal nito ay nag-iiba. Phase S ay wala, at walang karagdagang pagtitiklop ng DNA na nagaganap. Ang mga prosesong kasangkot sa ikalawang dibisyon ng meiosis ay katulad sa kanilang mekanismo sa mga nagaganap sa mitosis. Kasama sa mga ito ang paghihiwalay ng mga chromatids sa parehong mga cell ng anak na babae na nagreresulta mula sa unang meiotic division. Ang pangalawang dibisyon ng meiosis ay naiiba sa mitosis pangunahin sa dalawang paraan: 1) sa metaphase II meiosis, ang mga kapatid na chromatid ay madalas na mahigpit na nakahiwalay sa isa't isa; 2) ang bilang ng mga chromosome ay haploid. |
|
|
Prophase II Sa mga cell na nawawalan ng interphase II , nawawala rin ang yugtong ito. Tagal ng prophase II inversely proportional sa tagal ng telophase ako . Ang nucleoli at nuclear membrane ay nawasak, at ang mga chromatid ay pinaikli at pinalapot. Ang mga centriole, kung naroroon, ay lumipat sa magkabilang poste ng mga selula; Lumilitaw ang mga filament ng spindle. Ang mga chromatid ay nakaayos sa paraang ang kanilang mahahabang axes ay patayo sa spindle axis ng unang meiotic division. |
Z. Prophase P |
|
Metaphase II Sa panahon ng ikalawang dibisyon, ang mga sentromere ay kumikilos tulad ng dobleng istruktura. Inayos nila ang mga thread ng spindle na nakadirekta patungo sa parehong mga poste, at sa gayon ay ihanay ang spindle sa ekwador. |
Metaphase II |
|
Anaphase II Gitna Nahati ang omer, at hinihila sila ng mga thread ng spindle sa magkabilang poste. Ang mga centromeres ay humihila sa magkakahiwalay na chromatids, na tinatawag na ngayong mga chromosome. Telofase II Ang yugtong ito ay halos kapareho sa telophase ng mitosis. Ang mga kromosom ay despiral, nababanat at pagkatapos ay mahirap na makilala. Ang mga filament ng spindle ay nawawala at ang mga centriole ay gumagaya. Sa paligid ng bawat nucleus, na ngayon ay naglalaman ng kalahati ng (haploid) na bilang ng mga chromosome ng orihinal na parent cell, muling nabuo ang isang nuclear membrane. Bilang resulta ng kasunod na paghahati ng cytoplasm (sa mga hayop) o pagbuo ng isang cell wall (sa mga halaman), apat na anak na cell ang nakuha mula sa isang orihinal na parent cell. |
|


Mga pagkakaiba sa pagitan ng mga yugto ng mitosis at meiosis
|
Yugto |
Mitosis |
Meiosis |
|
Prophase |
Ang mga chromomere ay hindi nakikita Ang mga homologous chromosome ay pinaghihiwalay Ang Chiasmata ay hindi nabuo Hindi nangyayari ang mga tawiran |
Nakikita ang mga chromomere Ang mga homologous chromosome ay conjugated. Nabuo ang chiasmata. Maaaring mangyari ang pagtawid |
|
Metaphase |
Ang mga pares ng Chromatid ay matatagpuan sa spindle equator Pumila ang mga centromeres sa parehong eroplano sa spindle equator |
Ang mga pares ng chromatids ay matatagpuan sa spindle equator lamang sa ikalawang dibisyon ng meiosis Ang mga centromeres sa unang dibisyon ng meiosis ay matatagpuan sa itaas at ibaba ng ekwador sa pantay na distansya mula dito |
|
Anaphase |
Naghahati ang mga sentromer. Ang mga chromatid ay naghihiwalay. Magkapareho ang divergent chromatid |
Ang mga centromeres ay nahahati lamang sa ikalawang yugto ng meiosis. Ang mga chromatid ay naghihiwalay sa panahon ng ikalawang dibisyon ng meiosis. Sa unang dibisyon, ang buong chromosome ay naghihiwalay. Maaaring hindi magkapareho ang mga diverging chromosome bilang resulta ng pagtawid |
|
Telolophase |
Ang bilang ng mga chromosome sa mga anak na selula ay pareho sa mga selula ng magulang Ang mga cell ng anak na babae ay naglalaman ng parehong homologous chromosome (sa diploids) |
Ang bilang ng mga chromosome sa mga anak na selula ay kalahati kaysa sa mga selula ng magulang Ang mga selulang anak na babae ay naglalaman lamang ng isa sa bawat pares ng mga homologous chromosome |
|
Saan nangyayari ang ganitong uri ng dibisyon? |
Posible sa haploid, diploid at polyploid cells Nangyayari sa panahon ng pagbuo ng mga somatic cell at ilang mga spores, gayundin sa panahon ng pagbuo ng mga gametes sa mga halaman kung saan nagaganap ang paghahalili ng mga henerasyon |
Sa diploid at polyploid cells lamang Sa panahon ng gameto- o sporogenesis |
- Ang istraktura ng gametes. Gametogenesis
Ang sekswal na pagpaparami ay isinasagawa gamit ang mga espesyal na selula ng mikrobyo na tinatawag na gametes. Ang mga babaeng gametes ay tinatawag itlog, lalaki spermatozoa.Ang mga gametes ay naiiba sa mga somatic cell pangunahin sa pagkakaroon ng kalahati ng bilang ng mga chromosome, pati na rin ang isang mababang antas ng mga metabolic na proseso.
Mga ovule medyo malalaking non-motile cells, kadalasang bilog ang hugis; Bilang karagdagan sa mga tipikal na organelles, ang cytoplasm ay naglalaman ng mga pagsasama ng mga reserbang nutrients sa anyo ng yolk ( kanin .). Sa nuclei ng mga itlog, maraming mga kopya ng ribosomal genes at mRNA ang nabuo, na tinitiyak ang synthesis ng mga mahahalagang protina ng hinaharap na embryo. Ang mga itlog ng iba't ibang mga organismo ay naiiba sa dami at likas na katangian ng pamamahagi ng yolk sa kanila. Mayroong ilang mga uri ng mga itlog.

Isolecithalay tinatawag na medyo maliit na mga itlog na may isang maliit na halaga ng pantay na distributed yolk. Ang core sa kanila ay matatagpuan mas malapit sa gitna. Ang ganitong mga itlog ay matatagpuan sa mga bulate, bivalve at gastropod, echinoderms, at lancelet.Katamtamang telolecithalAng mga itlog ng sturgeon at amphibian ay may diameter na humigit-kumulang 1.5 × 2 mm at naglalaman ng isang average na halaga ng yolk, ang karamihan nito ay puro sa isa sa mga pole (vegetative). Sa kabaligtaran na poste (hayop), kung saan may maliit na pula ng itlog, mayroong nucleus ng itlog.
Malakas na telolecithalAng mga itlog ng ilang isda, reptilya, ibon at oviparous na mammal ay naglalaman ng maraming pula ng itlog, na sumasakop sa halos buong dami ng cytoplasm ng itlog. Sa poste ng hayop mayroong isang germinal disc na may aktibong cytoplasm na walang yolk. Ang mga sukat ng mga itlog na ito ay malaki, 10 15 mm o higit pa.
Alecital ang mga itlog ay halos walang pula ng itlog, may maliit na maliit na sukat (0.1 × 0.3 mm) at katangian ng mga placental mammal, kabilang ang mga tao.
Ang tamud o ang tamud ay napakaliit na mobile male gametes (halimbawa, ang tamud ng tao ay 50 × 70 µm ang haba, at ang buwaya ay 20 µm) na nabuo ng mga male gonad - testes; ang kanilang bilang ay nasa milyun-milyon. Ang hugis ng tamud ay nag-iiba sa iba't ibang mga hayop, ngunit ang kanilang istraktura ay pareho, karamihan sa kanila ay may ulo at leeg. Sa ulo ng tamud mayroong isang nucleus na naglalaman ng haploid na bilang ng mga chromosome at sakop akrosom. Ang acrosome ay isang espesyal na istraktura, isang binagong Golgi complex, na naglalaman ng mga enzyme para sa pagtunaw ng lamad ng itlog sa panahon ng pagpapabunga, na napapalibutan ng isang lamad. Ang leeg ay naglalaman ng maraming mitochondria at dalawang centrioles. Ang isang buntot na nabuo sa pamamagitan ng microtubule ay lumalaki mula sa leeg at tinitiyak ang sperm motility. Ang gitnang bahagi ay pinalawak dahil sa maraming mitochondria na nilalaman nito, na binuo sa isang spiral sa paligid ng flagellum. Ang mga mitochondria na ito ay nagbibigay ng enerhiya para sa mga mekanismo ng contractile na nagbibigay ng paggalaw ng flagellum. Ang pangunahing at buntot na bahagi ng tamud ay may istrakturang katangian ng flagella.

Kung titingnan mo ang ulo ng isang tamud ng tao mula sa itaas, ito ay lumilitaw na bilog, at kapag tiningnan mula sa gilid, ito ay lumilitaw na patag. Ang paggalaw ng flagellar lamang ay hindi sapat para ang tamud ay maglakbay sa distansya mula sa puki patungo sa lugar kung saan nangyayari ang pagpapabunga. Ang pangunahing gawain ng lokomotor ng tamud ay ang kulubot sa paligid ng oocyte at i-orient ang kanilang mga sarili sa isang tiyak na paraan bago tumagos sa mga lamad ng oocyte.
Pagbuo ng mga gametes (gametogenesis).Mga cell ng germinal epithelium sa mga lalaki At Ang mga babaeng gonad ay sumasailalim sa isang serye ng mga sequential mitotic at meiotic division, na pinagsama-samang tinatawag na gametogenesis, na nagreresulta sa pagbuo ng mga mature male gametes (spermatogenesis) at female gametes (oogenesis). Sa parehong mga kaso, ang proseso ay nahahati sa tatlong yugto - ang yugto ng pagpaparami, ang yugto ng paglago at ang yugto ng pagkahinog. Ang reproductive phase ay nagsasangkot ng paulit-ulit na mitotic division na humahantong sa pagbuo ng maraming spermatogonia o oogonia. Ang bawat isa sa kanila ay sumasailalim sa isang panahon ng paglago bilang paghahanda para sa unang meiotic division at kasunod na cytokinesis. Pagkatapos ay magsisimula ang yugto ng pagkahinog, kung saan nangyayari ang una at pangalawang meiotic division Sa kasunod na pagkita ng kaibhan ng mga haploid cells At pagbuo ng mga mature gametes.
Pag-unlad ng tamud (spermatogenesis).Ang spermatozoa (spermine) ay nabuo bilang resulta ng sunud-sunod na paghahati ng cell, na pinagsama-samang tinatawagspermatogenesis,sinusundan ng isang kumplikadong proseso ng pagkita ng kaibhan na tinatawag spermiogenesis (Larawan 20.31 ). Ang proseso ng pagbuo ng tamud ay tumatagal ng humigit-kumulang 70 araw; bawat 1 g ng testicle weight 10 ay nabuo 7 tamud bawat araw. Ang epithelium ng seminiferous tubule ay binubuo ng isang panlabas na layer ng mga cellgerminal epitheliumat humigit-kumulang anim na layer ng mga cell ang nabuo bilang resulta ng paulit-ulit na paghahati ng cell ng layer na ito (Larawan 20.32 at 20.33); ang mga layer na ito ay tumutugma sa sunud-sunod na mga yugto ng pag-unlad ng tamud. Una, ang dibisyon ng germinal epithelial cells ay nagdudulot ng marami spermatogonia na lumalaki sa laki at nagigingunang order ng spermatocytes.Bilang resulta ng unang meiotic division, ang mga spermatocytes na ito ay bumubuo ng haploidpangalawang order spermatocytes,pagkatapos nito ay sumasailalim sila sa pangalawang meiotic division at nagiging spermatids. Sa pagitan ng mga "strands" ng pagbuo ng mga cell ay may malaki Sertoli cells, o mga trophic na selula,matatagpuan sa buong espasyo mula sa panlabas na layer ng tubule hanggang sa lumen nito.

Ang mga spermatocytes ay matatagpuan sa maraming invaginations sa mga lateral surface ng Sertoli cells; dito sila nagiging spermatids, at pagkatapos ay lumipat sa gilid ng Sertoli cell, na nakaharap sa lumen ng seminiferous tubule, kung saan sila ay nag-mature, na bumubuo. spermine (Larawan 20.33 ). Tila, ang mga selulang Sertoli ay nagbibigay ng mekanikal na suporta, proteksyon at nutrisyon sa pagkahinog ng tamud. Ang lahat ng nutrients at oxygen na inihatid sa pagbuo ng mga gametes sa pamamagitan ng mga daluyan ng dugo na nakapalibot sa mga seminiferous tubules, at metabolic waste na inilabas sa dugo, ay dumadaan sa mga selula ng Sertoli. Ang mga cell na ito ay naglalabas din ng likido kung saan ang tamud ay dumadaan sa mga tubule.
Pag-unlad ng itlog sa mga tao (oogenesis).Hindi tulad ng pagbuo ng tamud, na nagsisimula sa mga lalaki lamang sa pagdadalaga, ang pagbuo ng mga itlog sa mga kababaihan ay nagsisimula kahit bago ang kanilang kapanganakan at nakumpleto para sa bawat ibinigay na itlog pagkatapos lamang ng pagpapabunga nito. Ang mga yugto ng oogenesis ay ipinapakita sa Fig. 20.36. Sa panahon ng pag-unlad ng pangsanggol, ang mga primordial germ cells ay paulit-ulit na nahahati sa pamamagitan ng mitosis, na gumagawa ng maraming malalaking selula na tinatawag na oogonia. Ang Oogonia ay muling sumasailalim sa mitosis at anyounang order oocygs,na nananatili sa yugto ng prophase halos hanggang sa obulasyon. Ang mga first order oocytes ay napapalibutan ng isang solong layer ng mga cell- granulosa lamad-at bumuo ng tinatawag naprimordial follicles.Ang isang babaeng fetus kaagad bago ipanganak ay naglalaman ng mga 2 10 6 ang mga follicle na ito, ngunit halos 450 lamang sa kanila ang umabot sa yugto ng second-order oocytes at umalis sa obaryo (ovulation). Bago ang obulasyon, ang first-order oocyte ay sumasailalim sa unang meiotic division, na bumubuo ng isang haploidpangalawang order oocyte At unang polar body.Ang ikalawang meiotic division ay umabot sa metaphase stage, ngunit hindi nagpapatuloy hanggang ang oocyte ay nagsasama sa sperm. Sa panahon ng pagpapabunga, ang pangalawang-order na oocyte ay sumasailalim sa pangalawang meiotic division, na bumubuo ng isang malaking cell - itlog at gayundin pangalawang polar body.Ang lahat ng polar body ay maliliit na selula; wala silang ginagampanan sa oogenesis at tuluyang nawasak.

parthenogenesis
obligasyon
opsyonal
Ang pag-unlad ng mga organismo lamang mula sa isang hindi fertilized na itlog.
Mga babae lang ang nabuo.
Halimbawa, Caucasian rock lizard
Ang mga letlet ay maaaring bumuo ng parehong walang pagpapabunga at may pagpapabunga
Uri ng babae
Ang mga lalaki ay nabubuo mula sa hindi na-fertilized na mga itlog
Uri ng lalaki
Sa ilang isogamous algae
Iba pang katulad na mga gawa na maaaring interesante sa iyo.vshm> |
|||
| 6644. | Indibidwal na pag-unlad ng mga organismo (ontogenesis) | 78.69 KB | |
| Ito ay pinaniniwalaan na ang mga nuklear na nilalaman ng isang tamud lamang ay tumagos sa itlog ng mga hayop. Sa kaso ng mga tao at kung minsan ay mas mataas na mga hayop, ang panahon ng pag-unlad bago ang kapanganakan ay madalas na tinatawag na prenatal pagkatapos ng kapanganakan at postnatal. Sa karamihan ng mga multicellular na hayop, anuman ang pagiging kumplikado ng kanilang organisasyon, ang mga yugto ng pag-unlad ng embryonic na pinagdadaanan ng embryo ay pareho. Ang likas na katangian ng pagdurog at mga uri ng blastula sa iba't ibang vertebrates... | |||
| 13714. | Indibidwal na pag-unlad ng mga organismo at ang kanilang pag-uugali. Ontogenesis. Siklo ng buhay sa mga halaman at hayop | 9.96 KB | |
| Ang pinagmulan ng personalidad ng ontogenesis ay ang indibidwal na pag-unlad ng isang organismo mula sa sandali ng pagbuo ng zygote hanggang sa kamatayan nito. Ang mga gametes ay mga sex cell na nagdadala ng namamana na impormasyon at mayroong haploid set. Ang fertilization ay ang proseso ng pagsasanib ng lalaki at babaeng reproductive cell ng mga halaman o hayop at ito ang batayan ng proseso ng sekswal. Ang embryonic germinal period ay ang panahon ng ontogenesis mula sa pagbuo ng zygote hanggang sa pagsilang o paglitaw mula sa mga lamad ng itlog o pagtubo. | |||
| 10427. | Pagpaparami | 6.75 KB | |
| Sa isang malaking pagkakaiba-iba ng mga anyo ng pagpaparami ng mga organismo, lahat sila ay maaaring mabawasan sa dalawang pangunahing uri: asexual at sekswal. Sa asexual reproduction, ang pagpaparami ng mga supling ay nangyayari mula sa isang magulang sa pamamagitan ng pagbuo ng mga spores o vegetatively. Sa panahon ng vegetative propagation, ang mga supling ay nagmumula sa mga bahagi ng katawan na hiwalay sa ina. Sa panahon ng vegetative propagation, ang mga halaman ay nagpapanatili ng heterozygosity sa maraming henerasyon. | |||
| 21332. | Naglilimita sa mga kadahilanan. Pag-angkop ng mga organismo sa mga salik | 303.8 KB | |
| Ang ilang mga hayop ay mahilig sa matinding init, ang iba ay mas pinahihintulutan ang katamtamang temperatura sa kapaligiran, atbp. Bilang karagdagan, ang mga nabubuhay na organismo ay nahahati sa mga may kakayahang umiiral sa isang malawak o makitid na hanay ng mga pagbabago sa anumang kadahilanan sa kapaligiran. Kung ang impluwensya ng mga kondisyon sa kapaligiran ay hindi umabot sa matinding halaga, ang mga nabubuhay na organismo ay tumutugon dito na may ilang mga aksyon o pagbabago sa kanilang estado, na sa huli ay humahantong sa kaligtasan ng mga species. Ang layunin ng pag-aaral sa gawaing ito ay mga salik sa kapaligiran; ang paksa ay nililimitahan ang mga salik at pagbagay ng mga organismo sa... | |||
| 8875. | Pangkalahatang mga pattern ng pakikipag-ugnayan sa pagitan ng mga organismo at mga salik sa kapaligiran | 193.58 KB | |
| Biotic factor ng terrestrial at aquatic na kapaligiran ng mga lupa Biologically active substances of living organisms Anthropogenic factors Pangkalahatang pattern ng interaksyon sa pagitan ng mga organismo at environmental factors Ang konsepto ng isang limiting factor. Ang pinakamababang batas ng Liebig Batas ni Shelford Mga detalye ng epekto ng mga anthropogenic na kadahilanan sa isang organismo Pag-uuri ng mga organismo na may kaugnayan sa mga kadahilanan sa kapaligiran 1. Ang mga kondisyon ng feather grass steppes ay kumakatawan sa ganap na magkakaibang mga rehimen ng mga abiotic na kadahilanan. | |||
| 12700. | Mga biyolohikal na katangian ng mga peste at mga hakbang upang labanan ang mga ito | 62.79 KB | |
| Ang mga partikular na makabuluhang pagkalugi sa pananim ay nangyayari bilang resulta ng pagkakaroon ng mga damo na nag-aalis ng mga sustansya at kahalumigmigan mula sa lupa, lilim ng mga halaman na nilinang, at sa maraming mga kaso ay nakontamina ang mga produkto na may mga nakakalason na sangkap at mga buto na nagdudulot ng pagkalason sa mga tao at hayop. Ang mga pangunahing direksyon ng chemicalization ng agrikultura: ang paggamit ng mga pataba, kemikal, proteksyon ng halaman mula sa mga peste, sakit at mga damo, ang paggamit ng mga produktong kemikal sa mga hayop, pag-canning ng mga produktong agrikultura at... | |||
| 13403. | Ang mga virus bilang non-cellular life forms. Istraktura, pag-uuri, pakikipag-ugnayan ng virus sa mga selula ng iba't ibang mga organismo | 12.75 KB | |
| Ito ay mga biological na bagay, mga genome na binubuo ng mga nucleic acid na DNA o RNA, na ginawa sa mga buhay na selula gamit ang kanilang biosynthetic apparatus. Mga pagkakaiba sa pagitan ng mga virus at iba pang anyo ng buhay: wala silang cellular na istraktura; 1 uri ng mga nucleic acid; DNA o RNA lamang; wala silang sariling metabolismo. Hypothesis ng pinagmulan ng mga virus: ang mga virus ay lumitaw mula sa mga bahagi ng isang normal na cell na nakatakas sa kontrol ng mga mekanismo ng regulasyon at naging isang independiyenteng yunit; marahil, isang serye ng mga pagbabago sa genetic ang naganap sa isang seksyon ng DNA na... | |||
| 18798. | Mga istruktura at functional na tagapagpahiwatig ng mga organismo ng mesofauna ng lupa sa kagubatan at bukas na biotopes ng lambak ng Pozim River | 61.54 KB | |
| Gayunpaman, ngayon ang pag-unlad ng teritoryong ito ay bunga ng maraming problema sa kapaligiran. Sa partikular, malawakang isinasagawa ang trabaho sa lambak upang maubos ang mga parang, na direktang nakakaapekto sa mga natural na biotop. | |||
| 19386. | Pag-unlad ng liberalismo sa CIS | 35.55 KB | |
| Ang kumplikadong sistema ng kalikasan ng mga pagbabagong isinasaalang-alang sa mga proseso ng ebolusyon ng mga dayuhang relasyon sa ekonomiya ay tumutukoy sa rebisyon at pagbabago ng buong axiomatics ng mga tradisyonal na ideya tungkol sa pagkakakilanlan, mga hangganan at soberanya ng pambansang ekonomiya... | |||
| 8867. | EDUKASYON AT PAG-UNLAD | 162.32 KB | |
| Ang kakanyahan ng problema ng relasyon sa pagitan ng pagsasanay at pag-unlad. Mga pangunahing diskarte sa paglutas ng problema ng relasyon sa pagitan ng pagsasanay at pag-unlad. Ang konsepto ng zone ng proximal development L. Ang kakanyahan ng problema ng relasyon sa pagitan ng pagsasanay at pag-unlad. | |||
Ang pagpaparami ay ang pag-aari ng mga organismo upang makagawa ng mga supling o ang kakayahan ng mga organismo na magparami ng kanilang mga sarili. Bilang pinakamahalagang pag-aari ng mga nabubuhay na bagay, tinitiyak ng pagpaparami ang pagpapatuloy ng buhay at ang pagpapatuloy ng mga species.
Ang proseso ng pagpaparami ay lubhang kumplikado at nauugnay hindi lamang sa paglipat ng genetic na impormasyon mula sa mga magulang patungo sa mga supling, kundi pati na rin sa anatomical at physiological na mga katangian ng mga organismo, kasama ang kanilang pag-uugali, at hormonal control. Ang pagpaparami ng mga organismo ay sinamahan ng mga proseso ng kanilang paglaki at pag-unlad.
Ang mga buhay na nilalang ay nailalarawan sa pamamagitan ng matinding pagkakaiba-iba sa kanilang mga pamamaraan ng pagpaparami. Gayunpaman, mayroong dalawang pangunahing paraan ng pagpaparami - asexual at sekswal (Larawan 16). Asexual reproduction, o apomixis (mula sa Greek. aro - walang, mixis - paghahalo) ay isang proseso kung saan isang magulang lamang (cell o multicellular organism) ang nakikilahok. Sa kabaligtaran, ang sekswal na pagpaparami ay nagsasangkot ng dalawang magulang, na ang bawat isa ay may sariling reproductive system at gumagawa ng mga sex cell (gametes), na, pagkatapos ng pagsasanib, ay bumubuo ng isang zygote (fertilized egg), na pagkatapos ay nag-iiba sa isang embryo. Dahil dito, sa panahon ng sekswal na pagpaparami ay may pinaghalong namamana na mga kadahilanan, ibig sabihin, isang proseso na tinatawag na amphimixis (mula sa Griyego. amphi - sa magkabilang panig, mixis - paghahalo).
Asexual Reproduction
Ang asexual reproduction ay katangian ng mga organismo ng maraming species, parehong halaman at hayop. Ito ay matatagpuan sa mga virus, bacteria, algae, fungi, vascular plants, protozoa, sponges, coelenterates, bryozoans at tunicates.
Ang pinakasimpleng anyo ng asexual reproduction ay katangian ng mga virus. Ang kanilang proseso ng pag-aanak ay nauugnay sa mga molekula ng nucleic acid, na may kakayahan ng mga molekulang ito na mag-self-duplicate
at batay sa pagtitiyak ng medyo mahina na mga bono ng hydrogen sa pagitan ng mga nucleotide.
kanin. 16.Mga paraan ng pagpaparami ng mga organismo
Kaugnay ng iba pang mga organismo na nagpaparami nang walang seks, ginagawa ang pagkakaiba sa pagitan ng vegetative reproduction at reproduction sa pamamagitan ng sporulation.
Ang vegetative propagation ay pagpaparami kung saan ang isang bagong organismo ay bubuo mula sa isang bahagi na hiwalay sa maternal na organismo. Ang ganitong uri ng pagpaparami ay katangian ng parehong unicellular at multicellular na mga organismo, ngunit may iba't ibang mga pagpapakita sa kanila.
Sa mga unicellular na organismo, ang vegetative reproduction ay kinakatawan ng mga form tulad ng division, multiple fission at budding. Ang paghahati sa pamamagitan ng simpleng paghihigpit sa pagbuo ng dalawang anak na organismo mula sa isang magulang na organismo ay katangian ng bakterya at asul-berdeng algae (cyanobacteria). Sa kabaligtaran, ang pagpaparami sa pamamagitan ng dibisyon ng kayumanggi at berdeng algae, pati na rin ang mga unicellular na hayop (sarcodes, flagellates at ciliates) ay nangyayari sa pamamagitan ng mitotic division ng nucleus na sinusundan ng constriction ng cytoplasm.
Ang pagpaparami sa pamamagitan ng maraming fission (schizogony) ay nagsasangkot ng paghahati ng nucleus na sinusundan ng paghahati ng cytoplasm sa mga bahagi. Bilang resulta ng paghahati na ito, maraming anak na organismo ang nabuo mula sa isang cell. Ang isang halimbawa ng maramihang dibisyon ay ang pagpaparami ng falciparum plasmodium (P. vivax) sa mga erythrocyte ng tao. Sa kasong ito, sa plasmodia, ang paulit-ulit na dibisyon ng nuklear ay nangyayari nang maraming beses nang walang cytokinesis, na sinusundan ng cytokinesis. Bilang isang resulta, ang isang plasmodium ay nagbibigay ng 12-24 na mga organismo ng anak na babae.
Sa multicellular na mga organismo ng halaman, ang vegetative propagation sa pamamagitan ng dibisyon ay isinasagawa sa pamamagitan ng mga pinagputulan, bombilya, dahon, at rhizomes. Ngunit ito ay mahalagang artipisyal na pagpapalaganap na ginagamit sa pagsasanay sa agrikultura. Ang pagpaparami ng mas matataas na halaman sa ilalim ng mga artipisyal na kondisyon ay posible rin mula sa isang cell. Ang mga organismo na nabubuo mula sa isang cell (clone) ay mayroong lahat ng mga katangian ng orihinal na multicellular na organismo. Ang pagpapalaganap na ito ay tinatawag na clonal micropropagation. Ang isa sa mga anyo ng vegetative propagation ay maaaring paghugpong o paglipat ng maraming mga nilinang halaman, na binubuo ng paglipat ng usbong o bahagi ng isang shoot mula sa isang halaman patungo sa isa pa. Siyempre, ito rin ay isang paraan ng pagpaparami, na hindi nangyayari sa kalikasan, ngunit malawak na ginagamit sa agrikultura.
Sa mga multicellular na hayop, ang vegetative reproduction ay nangyayari sa pamamagitan ng paghahati-hati ng kanilang katawan sa mga bahagi, pagkatapos nito ang bawat isa
bahagi ay bubuo sa isang bagong hayop. Ang ganitong pagpaparami ay tipikal para sa mga espongha, coelenterates (hydras), nemerteans, flatworms, echinoderms (starfish) at ilang iba pang mga organismo. Ang isang malapit na anyo ng fragmentation sa vegetative reproduction ng mga hayop ay polyembryony ng hayop, na binubuo sa katotohanan na sa isang tiyak na yugto ng pag-unlad ang embryo ay nahahati sa ilang bahagi, na ang bawat isa ay bubuo sa isang independiyenteng organismo. Ang polyembryony ay nangyayari sa armadillos. Gayunpaman, ang huli ay nagpaparami nang sekswal. Samakatuwid, ang polyembryony ay sa halip ay isang natatanging yugto sa sekswal na pagpaparami, at ang mga supling na nagreresulta mula sa polyembryony ay kinakatawan ng monozygotic twins.
Ang budding ay binubuo ng pagbuo ng isang tubercle (outgrowth) na may nucleus sa mother cell, na pagkatapos ay naghihiwalay at nagiging isang malayang organismo. Ang budding ay nangyayari kapwa sa mga unicellular na halaman, halimbawa sa yeast, at sa unicellular na hayop, halimbawa sa ilang mga species ng ciliates.
Ang pagpaparami sa pamamagitan ng sporulation ay nauugnay sa pagbuo ng mga dalubhasang mga cell - spores, na naglalaman ng isang nucleus, cytoplasm, ay natatakpan ng isang siksik na lamad at may kakayahang pangmatagalang pag-iral sa hindi kanais-nais na mga kondisyon, na, bilang karagdagan, ay nag-aambag sa kanilang dispersal. Kadalasan, ang ganitong pagpaparami ay nangyayari sa bakterya, algae, fungi, mosses, at ferns. Sa ilang berdeng algae, ang tinatawag na zoospores ay maaaring mabuo mula sa mga indibidwal na selula.
Sa mga hayop, ang pagpaparami sa pamamagitan ng sporulation ay sinusunod sa mga sporozoan, lalo na sa falciparum plasmodium.
Sa mga organismo ng maraming species, ang asexual reproduction ay maaaring kahalili ng sexual reproduction.
SEKSWAL NA REPRODUKSI
Ang sexual reproduction ay nangyayari sa parehong unicellular at multicellular na halaman at hayop.
Tulad ng nabanggit sa Kabanata V at XIII, ang sekswal na pagpaparami sa bakterya ay isinasagawa sa pamamagitan ng conjugation, na nagsisilbing isang analogue ng sekswal na proseso at isang sistema ng recombination ng mga organismo na ito, habang sa protozoa, ang sekswal na pagpaparami ay nangyayari din sa pamamagitan ng conjugation o sa pamamagitan ng syngamy. at autogamy.
Sa mga multicellular organism (halaman at hayop), ang sekswal na pagpaparami ay nauugnay sa pagbuo ng mikrobyo o sex cell (gametes), pagpapabunga at pagbuo ng mga zygotes.
Ang sexual reproduction ay isang makabuluhang evolutionary acquisition ng mga organismo. Sa kabilang banda, nag-aambag ito sa reassortment ng mga gene, ang paglitaw ng pagkakaiba-iba ng mga organismo at pagtaas ng kanilang pagiging mapagkumpitensya sa patuloy na pagbabago ng mga kondisyon sa kapaligiran.
Sa mga single-celled na organismo, ang sekswal na pagpaparami ay nangyayari sa iba't ibang anyo. Sa bakterya, ang sekswal na pagpaparami ay maaaring maitulad sa conjugation na nangyayari sa kanila, na binubuo sa paglipat ng plasmid o chromosomal DNA mula sa mga donor cell (naglalaman ng plasmids) sa mga cell ng tatanggap (hindi naglalaman ng mga plasmids), pati na rin sa transduction ng bakterya , na binubuo sa paglipat ng genetic na materyal mula sa ilang bacterial cell patungo sa iba pang mga phage. Ang conjugation ay matatagpuan din sa mga ciliates, kung saan sa prosesong ito ang paglipat ng nuclei ay nangyayari mula sa isang indibidwal patungo sa isa pa, na sinusundan ng paghahati ng huli.
Sa multicellular na mga halaman at hayop, ang sekswal na pagpaparami ng mga selula ay nangyayari sa pamamagitan ng pagbuo ng mga babae at lalaki na mga selula ng mikrobyo (mga itlog at tamud), kasunod na pagpapabunga ng itlog sa pamamagitan ng isang tamud at ang pagbuo ng isang zygote. Sa mga halaman, ang mga sex cell ay ginawa sa mga espesyal na istruktura ng reproduktibo; sa mga hayop, ang mga ito ay ginawa sa mga gonad na tinatawag na gonads (mula sa Greek. wala na - buto).
Mayroong mahalagang pagkakaiba sa pagitan ng somatic at reproductive cells ng mga hayop. Ito ay nakasalalay sa katotohanan na ang mga somatic cell ay may kakayahang dibisyon, iyon ay, nagpaparami sila sa kanilang sarili, at, bilang karagdagan, ang mga selula ng mikrobyo ay nabuo mula sa kanila. Sa kabaligtaran, ang mga sex cell ay hindi nahahati, ngunit sila ay "nagsisimula" sa pagpaparami ng buong organismo.
Ang diploid somatic cells kung saan nabuo ang mga male germ cell ay tinatawag na spermatogonia, at kung saan nabuo ang mga babaeng germ cell - oogonia. Ang proseso ng pagbuo (paglaki at pagkita ng kaibhan) ng mga selulang mikrobyo ng lalaki at babae ay tinatawag na gametogenesis.
Ang gametogenesis ay batay sa meiosis (mula sa Greek. meiosis - reduce), na isang pagbawas na dibisyon ng cell nuclei, na sinamahan ng pagbawas sa bilang ng mga chromosome bawat nucleus. Ang Meiosis ay nangyayari sa mga espesyal na selula ng reproductive organ ng mga buhay na nilalang na sekswal na nagpaparami (Larawan 17). Halimbawa, sa mga pteridophytes, ang meiosis ay nangyayari sa mga espesyal na sporangial cell na matatagpuan sa ibabang ibabaw ng mga dahon ng mga halaman na ito at nagiging mga spore at pagkatapos ay sa mga gametophytes. Ang huli ay umiiral nang hiwalay, sa huli ay gumagawa ng lalaki at babaeng gametes. Sa mga namumulaklak na halaman, ang meiosis ay nangyayari sa mga espesyal na selula ng mga ovule, na nagiging mga spore. Ang huli ay gumagawa ng isang gametophyte na may isang itlog. Bilang karagdagan, sa mga halaman na ito, ang meiosis ay nangyayari rin sa mga espesyal na selula ng anter, na nabubuo din sa mga spores na sa huli ay gumagawa ng pollen na may dalawang male gametes. Sa mga earthworm, na mga hermaphrodite at naglalaman ng mga male reproductive organ sa isang bahagi ng katawan at babae sa isa pa at nailalarawan sa pamamagitan ng
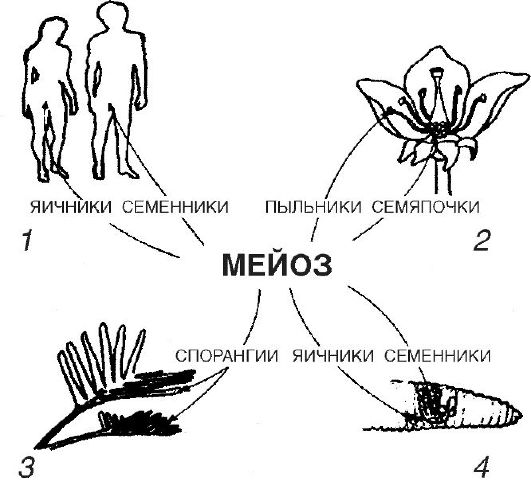
kanin. 17. Meiosis sa iba't ibang organismo: 1 - Tao; 2 - namumulaklak na mga halaman; 3 - pako; 4 - bulate
Mayroon silang kakayahang mag-cross-fertilize sa pagitan ng iba't ibang indibidwal; mayroon silang kakayahang sabay na magsagawa ng spermatogenesis at oogenesis.
Ang Meiosis ay nangyayari sa mga espesyal na selula ng testes at ovaries, na gumagawa ng mga male at female gametes, ayon sa pagkakabanggit. Ang mga protina na inducers ng meiosis ay natukoy.
Sa panahon ng proseso ng meiosis, ang diploid na bilang ng mga chromosome (2n), na katangian ng mga somatic cell (cell nuclei) at mga immature cell germs, ay nagbabago sa haploid number (n), na katangian ng mga mature na germ cell. Kaya, bilang isang resulta ng gametogenesis, ang mga cell ng mikrobyo ay tumatanggap lamang ng kalahati ng mga chromosome ng mga somatic cells (Larawan 18).

kanin. 18.
Ang pag-uugali ng mga chromosome sa panahon ng gametogenesis sa mga hayop ay pareho sa parehong lalaki at babae. Gayunpaman, ang mga kasarian ay naiiba sa oras ng pinagmulan ng iba't ibang yugto ng meiosis, na lalo na
lalo na kapansin-pansin sa mga tao. Sa postpubertal na mga lalaki, ang buong proseso ng meiosis ay nakumpleto sa loob ng halos dalawang buwan, habang sa mga babae, ang unang meiotic division ay nagsisimula sa fetal ovary at hindi natatapos hanggang sa magsimula ang obulasyon, na nangyayari sa mga 15 taong gulang.
Sa mas mataas na mga hayop, sa kaso ng mga lalaki, ang meiosis ay sinamahan ng pagbuo ng apat na functionally active gametes.
Sa kabaligtaran, sa mga babae, ang bawat pangalawang-order na oocyte ay gumagawa lamang ng isang itlog. Ang iba pang mga produkto ng nuklear ng babaeng meiosis ay tatlong katawan ng pagbabawas na hindi nakikilahok sa pagpaparami at pagkabulok.
Ang Meiosis ay binubuo ng dalawang dibisyong nuklear. Ang unang meiotic nuclear division ay naghihiwalay sa mga miyembro ng bawat pares ng homologous chromosome pagkatapos nilang ipares sa isa't isa (synapsis) at magpalitan ng genetic material (crossing over). Bilang resulta ng dibisyong ito, nabuo ang dalawang haploid nuclei. Ang pangalawang meiotic division ay naghihiwalay sa dalawang longitudinal na halves ng chromosome (chromatids) sa bawat isa sa mga nuclei na ito, na gumagawa ng apat na haploid nuclei.
Sa panahon ng proseso ng gametogenesis, nangyayari rin ang pagkakaiba-iba ng mga itlog (ovogenesis) at spermatozoa (spermatogenesis), na isang kinakailangan para sa kanilang mga pag-andar. Ang mga itlog ng hayop ay mas malaki kaysa sa tamud, kadalasang hindi kumikibo at naglalaman ng nutritional material na nagsisiguro sa pag-unlad ng embryo sa unang panahon pagkatapos ng fertilization. Ang tamud ng karamihan sa mga hayop ay may flagellum, na nagsisiguro sa kanilang kalayaan sa paggalaw.
Ang Meiosis ay may natitirang biological na kahalagahan. Salamat sa meiosis, ang isang pare-parehong bilang ng mga chromosome ay pinananatili sa mga selula ng mga organismo, anuman ang bilang ng mga henerasyon. Samakatuwid, pinapanatili ng meiosis ang katatagan ng mga species. Sa wakas, sa meiosis, bilang resulta ng pagtawid, nangyayari ang recombination ng gene, na isa sa mga salik ng ebolusyon.
SPERMATOGENESIS AT OVOGENESIS
Ang spermatogenesis ay ang proseso ng pagbuo ng mga mature na male germ cells. Ang spermatozoa ay nabubuo sa mga male gonad (testes, o testes) mula sa mga espesyal na somatic cells (Larawan 19). Ang mga espesyal na selula ay nagsisilbing sumusunod:
tinatawag na primordial germ cells, na lumilipat sa testis sa maagang yugto ng embryogenesis ng indibidwal na lalaki. Dahil dito, ang mga primordial cell ay ang mga ninuno (precursors) ng mga mature na germ cell.
Ang mga testes ng tao ay binubuo ng maraming tubule, ang mga dingding nito ay nabuo sa pamamagitan ng mga patong ng mga selula na nasa iba't ibang yugto ng pag-unlad ng tamud. Ang panlabas na layer ng tubules ay nabuo ng malalaking selula na tinatawag na spermatogonia. Ang mga cell na ito ay naglalaman ng isang diploid na hanay ng mga chromosome at mga inapo ng primordial germ cells sa testis. Sa panahon ng pagbibinata ng isang indibidwal, ang bahagi ng spermatogonia ay gumagalaw sa panloob na layer ng mga tubule, kung saan, bilang resulta ng meiosis, sila ay bubuo sa mga cell na tinatawag na first-order spermatocytes (spermatocytes I), pagkatapos ay sa second-order spermatocytes. (spermatocytes II) at, sa wakas,

kanin. 19.Spermatogenesis at oogenesis
sa spermatids, na mga haploid germ cells na sa huli ay naiba sa mature sperm. Kaya, sa pangkalahatang mga termino, maaari nating sabihin na ang spermatogenesis ay pinasimulan sa diploid somatic cells (spermatogonia), na sinusundan ng isang panahon ng pagkahinog ng cell ng mikrobyo, kung saan ang dalawang nuklear na dibisyon ay nangyayari sa pamamagitan ng meiosis, na humahantong sa pagbuo ng mga spermatids. Ito ang larawan ng isang tao.
Ang Meiosis (Larawan 20) sa spermatogenesis ay nangyayari sa ilang yugto (mga yugto). Sa pagitan ng mga dibisyon ay may dalawang interphases. Kaya, ang meiotic division ay maaaring kinakatawan bilang isang serye ng mga kaganapan na sumusunod sa isa't isa, katulad: interphase I - ang unang meiotic division (maagang prophase I, late prophase I, metaphase I, anaphase I, telophase I) - interphase II (interokinesis) - pangalawang meiotic division (prophase I, metaphase II, anaphase II, telophase II). Ang proseso ng meiosis ay napaka-dynamic, kaya ang mga mikroskopikong pagkakaiba sa pagitan ng iba't ibang yugto ay hindi sumasalamin sa likas na katangian ng mga yugto mismo, ngunit sa halip ang mga katangian ng mga kromosom sa iba't ibang yugto. Interphase I

kanin. 20.Mga yugto ng Meiosis: 1 - unang meiotic division; 2 - pangalawang meiotic division
nailalarawan sa pamamagitan ng katotohanan na ang pagtitiklop ng chromosome (pagdodoble ng DNA) ay nangyayari dito, na halos ganap na nakumpleto sa simula ng maagang prophase I.
Ang unang meiotic division ay nagsisimula sa pangunahing spermatocyte at nailalarawan sa pamamagitan ng isang mahabang prophase, na binubuo ng prophase I at prophase II na lumilipat sa isa't isa. Sa prophase I, mayroong limang pangunahing yugto - leptonema, zygonema, pachynema, diplonema at diakinesis.
Sa yugto ng leptonema, ang mga chromosome sa nucleus ay ipinakita sa anyo ng mga manipis na spiral thread na naglalaman ng maraming madilim na kulay na mga butil (chromomeres). Ang paghahati ng mga chromomere at mga thread ay hindi nabanggit, ngunit pinaniniwalaan na ang mga chromosome sa yugtong ito ay doble, ibig sabihin, diploid. Ang mga homologue ng bawat pares ng chromosome ay pinagsama ng mga chromomere sa haba ng mga ito ayon sa prinsipyo ng clasp.
Ang yugto ng zygonema ay nailalarawan sa pamamagitan ng pagtatatag ng mga synapses sa pagitan ng mga homologous chromosome, na nagreresulta sa pagbuo ng mga ipinares na chromosome (bivalents). Ang mga Chromosome X at Y ay medyo naiiba kumpara sa mga autosome. Nag-condense sila sa madilim na kulay na mga heterochromatic na katawan, na ipinares bilang resulta ng mga homologous na rehiyon sa kanilang mga dulo.
Sa yugto ng pachynema, na siyang pinakamahabang yugto sa meiotic prophase, nangyayari ang condensation ng bivalents at ang paghahati ng bawat chromatid sa dalawa, bilang isang resulta kung saan ang bawat bivalent ay isang kumplikadong helical na istraktura na binubuo ng apat na kapatid na chromatids (tetrads). Sa pagtatapos ng yugtong ito, magsisimula ang paghihiwalay ng mga ipinares na bivalent chromosome. Ngayon ang mga homologous chromosome ay maaaring obserbahan magkatabi. Samakatuwid, sa ilang mga paghahanda ay makikita ng isa ang apat na chromosome, na nabuo bilang isang resulta ng pagdoble ng bawat homolog, na bumubuo ng mga kapatid na chromatids. Sa yugtong ito, nagaganap ang mga palitan sa pagitan ng mga homologue at pagbuo ng chiasmata.
Sa yugto ng diplonema, ang pagpapaikli, pampalapot at mutual repulsion ng mga kapatid na chromatids ay nangyayari, bilang isang resulta kung saan ang mga chromatids sa bivalent ay halos magkahiwalay. Ang paghihiwalay ay itinuturing na hindi kumpleto dahil ang centromere ay hindi pa nahahati sa bawat pares ng mga kromosom. Tulad ng para sa mga bivalents, ang mga ito ay gaganapin sa iba't ibang mga lugar kasama ang kanilang haba sa pamamagitan ng chiasmata, na mga istruktura na nabuo sa pagitan ng mga homologous chromatid bilang isang resulta.
nakaraang pagtawid sa pagitan ng synaptically connected homologs. Sa mahusay na paghahanda, ang isa hanggang ilang chiasmata ay maaaring maobserbahan, depende sa haba ng bivalent. Ang bawat chiasma na naobserbahan sa yugtong ito ay kumakatawan sa resulta ng isang palitan na naganap sa pagitan ng mga hindi magkapatid na chromatid sa yugto ng pachynema. Habang tumitindi ang compression at repulsion ng bivalents, gumagalaw ang chiasmata patungo sa dulo ng mga chromosome, ibig sabihin, nangyayari ang terminalization ng mga chromosome. Sa pagtatapos ng diplonema, ang despiralization ng mga chromosome ay nangyayari; ang mga homologue ay patuloy na nagtataboy sa isa't isa.
Sa yugto ng diakinesis, na katulad ng diplonema, ang pagpapaikli ng bivalents ay nagpapatuloy at ang chiasmata ay humina (pagbaba), bilang isang resulta kung saan ang mga discrete unit sa anyo ng mga chromatids (apat) ay nabuo. Kaagad pagkatapos makumpleto ang yugtong ito, ang paglusaw ng nuclear membrane ay nangyayari.
Sa metaphase I, ang mga bivalents ay umabot sa kanilang pinakamataas na konsentrasyon. Nagiging hugis-itlog, sila ay matatagpuan sa ekwador na bahagi ng nucleus, kung saan sila ay bumubuo ng mga ekwador na plato ng meiotic metaphase I. Ang hugis ng bawat bivalent ay tinutukoy ng bilang at lokasyon ng chiasmata. Sa mga lalaki, ang bilang ng chiasmata bawat bivalent sa metaphase I ay karaniwang 1-5. Ang XY bivalent ay nagiging rod-shaped bilang resulta ng isang chiasm na matatagpuan sa terminal.
Sa anaphase I, ang paggalaw ng magkasalungat na sentromere sa magkasalungat na pole ng cell ay nagsisimula. Bilang resulta, ang mga homologous chromosome ay pinaghihiwalay. Ang bawat chromosome ngayon ay binubuo ng dalawang chromatids na pinagsasama-sama ng isang centromere, na hindi naghahati at nananatiling buo. Tinutukoy nito ang anaphase I ng meiosis mula sa anaphase ng mitosis, kung saan ang centromere ay sumasailalim sa dibisyon. Mahalagang tandaan na ang pagtawid ay gumagawa ng bawat chromatid na genetically naiiba.
Sa yugto ng telophase I, ang mga chromosome ay umaabot sa mga pole, na nagtatapos sa unang meiotic division. Pagkatapos ng telophase I, isang maikling interphase (interkinesis) ang nagaganap, kung saan ang mga chromosome ay despiral at nagiging diffuse, o ang telophase I ay direktang dumadaan sa prophase II ng pangalawang meiotic division. Sa alinmang kaso ay hindi sinusunod ang pagtitiklop ng DNA. Pagkatapos ng unang meiotic division, ang mga cell ay tinatawag na second-order spermatocytes. Ang bilang ng mga chromosome sa bawat naturang cell ay bumababa mula sa 2n hanggang 1n, ngunit hindi pa nagbabago ang nilalaman ng DNA.
Ang pangalawang meiotic division ay nangyayari sa ilang yugto (prophase II, metaphase II, anaphase II, telophase II) at katulad ng mitotic division. Sa prophase II, ang mga chromosome ng pangalawang spermatocytes ay nananatili sa mga pole. Sa metaphase II, ang centromere ng bawat isa sa mga double chromosome ay nahahati, na nagbibigay sa bawat bagong chromosome ng sarili nitong centromere. Nagsisimula ang pagbuo ng spindle, patungo sa poste kung saan gumagalaw ang mga bagong chromosome. Sa telophase II, ang pangalawang meiotic division ay nagtatapos, bilang isang resulta kung saan ang bawat pangalawang-order na spermatocyte ay gumagawa ng dalawang spermatids, kung saan ang spermatozoa ay nag-iiba. Tulad ng sa pangalawang spermatocyte, ang bilang ng mga chromosome sa spermatid ay haploid (1n). Gayunpaman, ang mga chromosome ng spermatids ay iisa, habang ang mga chromosome ng pangalawang spermatocytes II ay doble, na binuo mula sa dalawang chromatids. Dahil dito, ang nucleus ng bawat spermatid ay may isang solong hanay ng mga non-homologous chromosome. Ang pangalawang meiotic division ay isang dibisyon ng mitotic type (equatorial division). Pinaghihiwalay nito ang mga double sister chromatids at iba ito sa reduction division, kung saan pinaghihiwalay ang mga homologous chromosome. Ang tanging makabuluhang pagkakaiba mula sa klasikal na mitosis ay mayroong isang haploid na hanay ng mga kromosom.
Kaya, ang unang meiotic division ng first-order spermatocytes ay humahantong sa pagbuo ng dalawang pangalawang spermatocytes (pangalawang order). Ang parehong mga chromatids ng mga istruktura na nabuo bilang isang resulta ng pagbawas ng paghahati ay mga kapatid na chromatids. Ang huli ay lumitaw bilang isang resulta ng pagtitiklop bago ang unang meiotic division. Ang pangalawang meiotic division ng bawat pangalawang spermatocyte ay nagreresulta sa pagbuo ng apat na spermatids. Kaya, sa tipikal na meiosis, ang mga cell ay nahahati nang dalawang beses, habang ang mga chromosome ay nahahati lamang ng isang beses (Larawan 21).
Ang huling yugto sa spermatogenesis ay nauugnay sa pagkita ng kaibhan, na nagtatapos sa bawat isa sa medyo malaki, spherically immobile spermatids na nagiging isang maliit na pinahabang motile sperm.
Sa karamihan ng mga hayop na may sapat na gulang (sexually mature) na lalaki, ang spermatogenesis ay nangyayari sa mga testes nang palagian o pana-panahon (pana-panahon). Halimbawa, sa mga insekto ay tumatagal lamang ng ilang araw upang makumpleto ang cycle ng spermatogenesis, habang sa mga mammal ang siklo na ito ay tumatagal ng ilang linggo o kahit na.

kanin. 21.Pamamahagi ng mga chromosome sa panahon ng gametogenesis
buwan. Sa isang may sapat na gulang, ang spermatogenesis ay nangyayari sa buong taon. Ang oras ng pagbuo ng primitive spermatogonia sa mature sperm ay humigit-kumulang 74 araw.
Ang mga male reproductive cell, na ginawa ng mga organismo ng iba't ibang species, ay nailalarawan sa pamamagitan ng kadaliang kumilos at matinding pagkakaiba-iba sa laki at istraktura (Larawan 22). Ang bawat tamud ng tao ay binubuo ng tatlong seksyon - ang ulo, gitnang bahagi at buntot (Larawan 23). Ang ulo ng tamud ay naglalaman ng isang nucleus na naglalaman ng isang haploid na hanay ng mga chromosome.
Ang ulo ay nilagyan ng isang acrosome, na naglalaman ng mga lytic enzymes na kinakailangan para sa mga nilalaman ng tamud na makapasok sa itlog. Dalawang centrioles ay naisalokal din sa ulo - proc-

kanin. 22. Mga hugis ng tamud

kanin. 23. Istraktura ng tamud: A - light microscopic na imahe sa iba't ibang eroplano: 1 - ulo, 2 - gitnang bahagi, 3 - buntot; B - schematic reconstruction ng isang electron microscopic image: 1 - nucleus, 2 - acrosome, 3 - centrosome (proximal centriole), 4 - central ring, 5 - mitochondrial helix,
6 - axial thread
ang simal, na nagpapasigla sa paghahati ng itlog na pinataba ng tamud, at ang distal, na nagbubunga ng aximal shaft ng buntot. Ang gitnang bahagi ng tamud ay naglalaman ng basal na katawan ng buntot at mitochondria. Ang buntot (proseso) ng tamud ay nabuo sa pamamagitan ng isang panloob na aximal rod at isang panlabas na kaluban, na kung saan ay cytoplasmic pinagmulan. Ang spermatozoa ng tao ay nailalarawan sa pamamagitan ng makabuluhang motility.
OVOGENESIS AT OGGLE CELLS
Ang proseso ng pagbuo ng itlog ay tinatawag na oogenesis. Ang mga tungkulin nito ay upang matiyak ang haploid na hanay ng mga kromosom sa nucleus ng itlog at ang mga pangangailangan sa nutrisyon ng zygote. Ang oogenesis sa pagpapakita nito ay karaniwang maihahambing sa spermatogenesis.
Sa mga mammal at tao, ang oogenesis ay nagsisimula sa prenatal period (bago ipanganak). Ang Oogonia, na mga maliliit na selula na may medyo malaking nucleus at naisalokal sa mga ovarian follicle, ay nagsisimulang mag-iba sa mga pangunahing oocytes. Ang huli ay nabuo na sa ikatlong buwan ng pag-unlad ng intrauterine, pagkatapos ay pumasok sila sa prophase ng unang meiotic division. Sa oras na ipinanganak ang isang batang babae, ang lahat ng mga pangunahing oocytes ay nasa prophase na ng unang meiotic division. Ang mga pangunahing oocyte ay nananatili sa prophase hanggang sa pagsisimula ng pagdadalaga sa babaeng indibidwal. Ang isang oocyte ay naglalaman ng hanggang 100,000 mitochondria. Kapag ang mga ovarian follicle ay nag-mature sa simula ng pagbibinata, ang meiotic prophase ay nagpapatuloy sa mga pangunahing oocytes. Ang unang meiotic division para sa bawat umuunlad na itlog ay nakumpleto ilang sandali bago ang oras ng obulasyon ng itlog na iyon. Bilang resulta ng unang meiotic division at hindi pantay na pamamahagi ng cytoplasm, ang isang resultang cell ay nagiging pangalawang oocyte, ang isa pa - isang polar (pagbawas) na katawan.
Ang pangalawang meiotic division ay nangyayari kapag ang pangalawang oocyte (nabubuong itlog) ay pumasa mula sa obaryo patungo sa fallopian tube. Gayunpaman, ang paghahati na ito ay hindi nakumpleto hanggang ang tamud ay tumagos sa pangalawang oocyte, na kadalasang nangyayari sa fallopian tube. Kapag ang tamud ay tumagos sa pangalawang oocyte, ang huli ay nahahati, na nagreresulta sa pagbuo ng isang ovote (mature na itlog) na may pronucleous na naglalaman ng isang set.
ng 23 maternal chromosome. Ang iba pang cell na nagreresulta mula sa dibisyong ito ay isang pangalawang polar body, na walang kakayahang higit pang pag-unlad. Sa oras na ito, ang polar (reduction) na katawan ay sumasailalim din sa paghahati sa dalawa. Kaya, ang pagbuo ng isang first-order oocyte ay sinamahan ng pagbuo ng isang ovotide at tatlong reduction body. Sa mga ovary, 300-400 oocytes ang karaniwang nag-mature sa ganitong paraan sa buong buhay, ngunit isang oocyte lamang ang nag-mature bawat buwan. Sa panahon ng pagkakaiba-iba ng mga itlog, ang mga lamad ay nabuo at ang kanilang nucleus ay bumababa sa laki.
Sa ilang mga species ng hayop, ang oogenesis ay nangyayari nang mabilis at tuluy-tuloy at nagreresulta sa paggawa ng malaking bilang ng mga itlog.
Sa kabila ng mga pagkakatulad sa spermatogenesis, ang oogenesis ay nailalarawan sa pamamagitan ng ilang mga tiyak na tampok. Ang nutritional material (yolk) ng pangunahing oocyte ay hindi pantay na ipinamamahagi sa apat na mga cell na nabuo bilang resulta ng meiotic divisions. Ang pangunahing halaga ng yolk ay nakaimbak sa isang malaking cell, habang ang mga polar body ay naglalaman ng napakakaunting sangkap na ito. Bilang resulta ng mga dibisyon, ang una at pangalawang polar na katawan ay tumatanggap ng parehong mga set ng chromosome bilang pangalawang oocytes, ngunit hindi sila nagiging mga cell ng mikrobyo. Samakatuwid, ang mga itlog ay mas mayaman sa nutritional material kumpara sa tamud. Ang pagkakaiba na ito ay lalo na binibigkas sa kaso ng mga hayop na nangingitlog.
Ang mga mammal na itlog ay may isang hugis-itlog o medyo pinahabang hugis (Larawan 24) at nailalarawan sa pamamagitan ng mga tipikal na katangian ng cellular na istraktura. Naglalaman ang mga ito ng lahat ng mga istraktura na katangian ng mga somatic cell, gayunpaman, ang intracellular na organisasyon ng itlog ay napaka tiyak at tinutukoy ng katotohanan na ang itlog ay din ang kapaligiran na nagsisiguro sa pag-unlad ng zygote. Ang isa sa mga katangian ng mga itlog ay ang pagiging kumplikado ng istraktura ng kanilang mga lamad. Sa maraming mga hayop, ang pangunahin, pangalawa at tertiary na lamad ng mga itlog ay nakikilala. Ang pangunahing shell (panloob) ay nabuo sa yugto ng oocyte. Kumakatawan sa ibabaw na layer ng oocyte, mayroon itong isang kumplikadong istraktura, dahil ito ay natagos ng mga outgrowth ng mga follicular cell na katabi nito. Ang pangalawang (gitnang) shell ay ganap na nabuo ng mga follicular cell, at ang tertiary (panlabas) na shell ay nabuo ng mga sangkap na mga produkto ng pagtatago ng mga glandula ng oviduct kung saan dumadaan ang mga itlog. Sa mga ibon, halimbawa,
Ang tertiary membranes ng mga itlog ay ang albumen, subshell at shell membranes. Ang mga itlog ng mammalian ay nailalarawan sa pagkakaroon ng dalawang lamad. Ang istraktura ng mga intracellular na bahagi ng mga itlog ay tiyak sa mga tuntunin ng mga species, at kung minsan ay may mga indibidwal na katangian.

kanin. 24. Istraktura ng itlog: 1 - follicular cell; 2 - shell; 3 - cytoplasm; 4- core
PAGPAPATABO
Ang pagpapabunga ay ang proseso ng pagsasama-sama ng lalaki at babaeng gametes, na humahantong sa pagbuo ng isang zygote at ang kasunod na pag-unlad ng isang bagong organismo. Sa panahon ng proseso ng pagpapabunga, ang isang diploid na hanay ng mga chromosome ay itinatag sa zygote, na tumutukoy sa natitirang biological na kahalagahan ng prosesong ito.
Depende sa mga species ng mga organismo na nagpaparami nang sekswal, ang panlabas at panloob na pagpapabunga ay nakikilala. Ang panlabas na pagpapabunga ay nangyayari sa kapaligiran kung saan pumapasok ang mga reproductive cell ng lalaki at babae. Halimbawa, ang pagpapabunga sa isda ay panlabas. Ang lalaki (gatas) at babae (caviar) na mga reproductive cell na inilabas ng mga ito ay pumapasok sa tubig, kung saan sila nagtagpo at nagkakaisa.
Ang panloob na pagpapabunga ay tinitiyak ng paglipat ng tamud mula sa katawan ng lalaki patungo sa katawan ng babae. Ang ganitong pagpapabunga ay nangyayari sa mga mammal, ang gitnang punto kung saan ay ang pagsasanib ng nuclei ng mga selulang mikrobyo. Ito ay pinaniniwalaan na ang mga nilalaman ng isang tamud ay tumagos sa itlog. Sa mga mekanismo ng pagpapabunga
pagkumpleto, marami pa rin ang hindi malinaw. Ang data sa pagpapabunga sa mga sea urchin ay nagpapahiwatig na 2 segundo pagkatapos ng pakikipag-ugnay ng tamud at itlog, ang mga pagbabago ay nangyayari sa mga de-koryenteng katangian ng lamad ng plasma ng huli. Ang pagsasanib ng mga gametes ay nangyayari sa loob ng 7 segundo. Ipinapalagay na ang pagtagos ng mga nilalaman ng isa lamang sa maraming tamud sa itlog ay ipinaliwanag sa pamamagitan ng mga pagbabago sa mga electrical properties ng plasma membrane nito. Ang isang fertilized na itlog ay nagbibigay ng isang zygote. Mayroong dalawang mga opinyon sa mga dahilan para sa pag-activate ng metabolismo ng itlog sa pamamagitan ng tamud. Ang ilan ay naniniwala na ang pagbubuklod ng tamud sa mga panlabas na receptor sa ibabaw ng mga selula ay isang senyas na pumapasok sa itlog sa pamamagitan ng lamad at nagpapagana ng inositol triphosphate at mga calcium ions doon. Ang iba ay naniniwala na ang tamud ay naglalaman ng isang espesyal na kadahilanan sa pagsisimula.
Ang mga eksperimentong pagpapaunlad na isinagawa sa mga nakaraang taon ay nagpakita na ang pagpapabunga ng mga itlog ng mga mammal, kabilang ang mga tao, ay posible sa vitro. Bukod dito, ang mga embryo na binuo sa vitro ay maaaring itanim sa matris ng isang babae, kung saan sila ay dumaranas ng karagdagang normal na pag-unlad. Maraming mga kaso ng pagsilang ng mga "test tube" na bata.
Hindi tulad ng mga hayop na nagpaparami sa pamamagitan ng zygogenesis, maraming mga organismo ang may kakayahang magparami sa pamamagitan ng parthenogenesis (mula sa Greek. parthenos - birhen at genos - kapanganakan), na tumutukoy sa pagpaparami ng mga organismo mula sa isang hindi pa fertilized na itlog. Mayroong obligado at facultative parthenogenesis. Ang obligadong parthenogenesis ay naging pangunahing paraan ng pagpaparami ng mga organismo ng ilang mga species, halimbawa, ang Caucasian rock lizard. Ang mga hayop ng species na ito ay babae lamang. Sa kabaligtaran, ang facultative parthenogenesis ay nangangahulugan na ang mga itlog ay maaaring bumuo ng parehong walang pagpapabunga at pagkatapos ng pagpapabunga. Ang facultative parthenogenesis, naman, ay babae at lalaki. Ang babaeng parthenogenesis ay madalas na naobserbahan sa mga bubuyog, langgam, at rotifers, kung saan ang mga lalaki ay nabubuo mula sa hindi na-fertilized na mga itlog. Ang male parthenogenesis ay nangyayari sa ilang isogamous algae.
Ang parthenogenesis ay maaaring maging natural at artipisyal (sapilitan). Ang mekanismo ng artificial parthenogenesis ay ang pangangati ng mga itlog gamit ang pisikal o kemikal
mga kadahilanan, na humahantong sa pag-activate ng mga itlog at, bilang isang resulta, sa pag-unlad ng mga hindi na-fertilized na itlog. Ang artipisyal na parthenogenesis ay naobserbahan sa mga hayop ng maraming mga sistematikong grupo - echinoderms, worm, mollusks at kahit na mga mammal.
Ang isang kilalang anyo ng parthenogenesis ay tinatawag na androgenesis (mula sa Griyego. andros - lalaki, genesis - pinagmulan). Kung ang nucleus sa isang itlog ay hindi aktibo at pagkatapos ay maraming tamud ang tumagos dito, kung gayon ang isang lalaki na organismo ay bubuo mula sa naturang itlog bilang resulta ng pagsasanib ng lalaki (sperm) nuclei.
May mga kaso kung saan ang parthenogenesis ay nangyayari sa paikot, depende sa panahon. Halimbawa, ang rotifers, daphnia at aphids ay nagpaparami sa tag-araw sa pamamagitan ng parthenogenesis, at sa taglagas sa pamamagitan ng pagpapabunga ng mga itlog at pagbuo ng mga zygotes, ibig sabihin, sa pamamagitan ng zygogenesis.
Ang papel ng parthenogenesis at ang mga anyo nito sa kalikasan ay maliit, dahil hindi ito nagbibigay ng malawak na kakayahang umangkop ng mga organismo.
Sa kaibahan sa zygogenesis at parthenogenesis, mayroong gynogenesis, na pseudogamy, kapag ang tamud ay pumasok sa itlog at na-activate ito, ngunit ang sperm nucleus ay hindi nagsasama sa nucleus ng itlog. Sa kasong ito, ang mga nagresultang supling ay binubuo lamang ng mga babae. Ang Gynogenesis ay natural na nangyayari sa mga nematode at isda, ngunit maaari rin itong ma-induce nang artipisyal. Sa partikular, ang mga kaso ng artipisyal na gynogenesis sa mga silkworm, isda at amphibian ay inilarawan.
ALTERNATING HAPLOIDY AT DIPLOIDY.
PAGSALIT NG MGA HENERASYON
Ang mga organismo na nagpaparami nang sekswal ay nailalarawan sa pamamagitan ng mga alternating haploid at diploid phase sa kanilang pag-unlad. Sa maraming mga organismo, kabilang ang mga mammal, ang paghahalili na ito ay regular, at ang pangangalaga ng mga katangian ng mga species ng mga organismo ay batay dito.
Maraming mga organismo ang nailalarawan din sa pamamagitan ng pagbabago ng mga henerasyon, kapag ang mga henerasyon ng mga indibidwal na nagpaparami ng asexual ay pinalitan ng mga henerasyon ng mga indibidwal na nagpaparami nang sekswal na may pagbuo ng mga gametes. Ito ay tinatawag na pangunahing pagbabago ng mga henerasyon. Ito ay matatagpuan sa mga sporozoan, flagellates at maraming halaman. Ang pangunahing pagbabago ng mga henerasyon ay regular, at ito ay naroroon
Ang matatagpuan sa kalikasan ay nagpapahiwatig ng pangangalaga sa phylogeny ng maraming mga organismo ng parehong asexual at sekswal na pagpaparami. Sa ibang mga organismo, nangyayari ang paghalili ng sekswal na pagpaparami sa parthenogenesis. Ito ay tinatawag na pangalawang henerasyong pagbabago. Halimbawa, sa trematodes, ang sekswal na pagpaparami ay regular na pinapalitan ng parthenogenesis. Ang pangalawang paghahalili ng mga henerasyon ay tinatawag na heterogony. Sa mga coelenterates, sa ilang mga yugto ng pag-unlad, nangyayari ang isang paglipat mula sa sekswal na pagpaparami hanggang sa asexual (vegetative). Ang anyo ng pangalawang paghahalili ng mga henerasyon ay tinatawag na metagenesis.
SEKSUAL DIMORPHISMO. HERMAPHRODITISM
Ang mga lalaki at babae ay nailalarawan sa pamamagitan ng mga tiyak na phenotypic na katangian. Ang mga pagkakaiba sa pagitan ng babae at lalaki sa kanilang mga katangian ay tinatawag na sexual dimorphism. Sa mga hayop, nangyayari na ito sa mas mababang yugto ng pag-unlad ng ebolusyon, halimbawa sa mga bilog na helminth at arthropod, at umabot sa pinakadakilang ekspresyon nito sa mga vertebrates.
Kung ang mga reproductive cell ng lalaki at babae ay ginawa ng parehong indibidwal, na mayroong parehong lalaki at babaeng gonad, kung gayon ang hindi pangkaraniwang bagay na ito ay tinatawag na totoong hermaphroditism. Ito ay matatagpuan sa mga flatworm, annelids, at mollusks. Sa mga flatworm, ang lalaki at babae na gonad ay gumagana sa buong buhay ng indibidwal. Sa kabaligtaran, sa mga mollusk ang gonad ay gumagawa ng mga itlog at tamud nang salit-salit.
Ang tunay na hermaphroditism ay nangyayari rin sa mga tao, na nagreresulta mula sa mga karamdaman sa pag-unlad. Ang mga genotype ng hermaphrodites ay 46 XX o 46 XY, na ang karamihan sa mga kaso ay XX (mga 60%). Ang genotype XX ay madalas na matatagpuan sa mga hermaphrodites ng mga populasyon ng Negroid African, habang ang XY ay kadalasang matatagpuan sa mga Japanese. Sa mga hermaphrodites ng parehong uri, ang isang pagkahilig sa bilateral gonadal asymmetry ay nabanggit. Kabilang sa mga tunay na hermaphrodites, mayroon ding mga chromosomal mosaic, na sa ilang mga somatic cells ay may isang pares ng XX chromosomes, sa iba pa - isang pares ng XY.
Ang maling hermaphroditism ay kilala rin, kapag ang mga indibidwal ay may panlabas na ari at pangalawang katangiang sekswal na katangian ng parehong kasarian, ngunit gumagawa lamang ng isang uri ng mga selulang mikrobyo - lalaki o babae.
PINAGMULAN NG MGA PARAAN NG REPRODUCTION
Ito ay pinaniniwalaan na ang pinaka sinaunang ay asexual reproduction, sa partikular na vegetative reproduction. Mula sa huli, ang pagpaparami sa pamamagitan ng pagbuo ng spore ay nabuo, ang walang alinlangan na bentahe nito ay nagbibigay ito ng mas mahusay na mga pagkakataon para sa pangangalaga ng mga species at lalo na para sa kanilang dispersal.
Ang sekswal na pagpaparami ay ang pinakamabisang paraan ng pagpaparami ng mga organismo. Ito ay pinaniniwalaan na ito ay nabuo mula sa asexual, umuusbong mga 1 bilyong taon na ang nakalilipas, at ang mga unang yugto ay nauugnay sa komplikasyon sa pagbuo ng mga gametes. Ang mga primitive gametes ay nailalarawan sa pamamagitan ng hindi sapat na morphological differentiation, bilang isang resulta kung saan ang isogamy (mula sa Greek. iso- pantay, gamos - kasal), kapag ang mga sex cell ay mga motile isogametes, hindi naiba sa anyo ng lalaki at babae.
Kasunod nito, nabuo ang anisogamy (mula sa Greek. anisos- hindi pantay, gamos - kasal), na nailalarawan sa pagkakaroon ng magkakaibang mga gametes na naiiba lamang sa laki. Sa mga huling yugto ng ebolusyon, lumitaw ang matalim na pagkakaiba sa kadaliang kumilos, hugis at laki ng mga gametes. Sa proseso ng ebolusyon, ang mga vertebrates ay nakabuo din ng ilang karagdagang mga aparato na nagpapadali sa paglipat ng male sperm sa babaeng reproductive tract at lumikha ng mga kondisyon para sa pagbuo ng isang fertilized na itlog. Sa panahon ng ebolusyon, ang mga aparatong ito ay nabuo mula sa excretory system, na humantong sa pagbuo ng genitourinary system.
Ang diploid state ay nagbibigay ng matinding benepisyo sa mga organismo dahil ang akumulasyon ng iba't ibang alleles ay nangyayari sa estadong ito. Samakatuwid, ang sekswal na pagpaparami ay mayroon ding kalamangan na ibinibigay nito sa mga organismoOmas malaking posibilidad ng pagkakaiba-iba kumpara sa asexual, at ito ay gumaganap ng isang mahalagang papel sa ebolusyon.
Ang parthenogenesis ay may malinaw na reproductive advantage, dahil ito ay gumagawa lamang ng mga babaeng supling. Gayunpaman, ito ay bihira. Dalawang hypotheses ang ginagamit upang ipaliwanag ang mababang dalas ng pagpapalit ng sekswal na pagpaparami ng parthenogenesis sa mga natural na populasyon ng mga organismo. Ayon sa isa sa kanila (mutation-accumulative), ang sex ay isang adaptive adaptation, dahil "nilinis" nito ang genome mula sa mga mutation na umuulit sa paglipas ng panahon, habang ayon sa isa pang hypothesis (ecological), ang sex ay
Ang mga unang ideya tungkol sa paglago at pag-unlad ay nagmula sa sinaunang mundo. Kahit na si Hippocrates (460-377 BC) ay ipinapalagay na ang mga itlog ay naglalaman na ng isang ganap na nabuong organismo, ngunit sa isang napakababang anyo. Ang ideyang ito ay binuo sa doktrina ng preformationism (mula sa lat. preformatio - preformation), na naging tanyag lalo na noong ika-17-18 siglo. Ang mga tagasuporta ng preformationism ay sina Harvey, Malpighi at marami pang ibang kilalang biologist at manggagamot noong panahong iyon. Para sa mga preformationist, ang kontrobersyal na isyu ay kung saan ang mga sex cell ang organismo ay preformed - babae o lalaki. Ang mga nagbigay ng kagustuhan sa mga itlog ay tinatawag na mga ovist, at ang mga nagbigay ng higit na kahalagahan sa mga reproductive cell ng lalaki ay tinatawag na mga animalculist. Ang preformationism ay isang metapisiko na doktrina mula simula hanggang wakas, dahil tinanggihan nito ang pag-unlad. Ang mapagpasyang suntok sa preformationism ay hinarap ni C. Bonnet (1720-1793), na nakatuklas ng parthenogenesis noong 1745 gamit ang halimbawa ng pag-unlad ng mga aphids mula sa hindi na-fertilized na mga itlog. Pagkatapos nito, ang preformationism ay hindi na makabawi at nagsimulang mawala ang kahalagahan nito.
Sa sinaunang mundo, lumitaw ang isa pang doktrina na kabaligtaran ng preformationism at kalaunan ay natanggap ang pangalang epigenesis (mula sa Greek. epi- pagkatapos, genesis - pag-unlad). Tulad ng preformationism, naging laganap din ang epigenesis noong ika-17-18 siglo. Sa paglaganap ng epigenesis, ang mga pananaw ni K.F. ay napakahalaga. Wolf (1733-1794), na buod sa kanyang aklat na "The Theory of Development" (1759). K.F. Naniniwala si Wolf na ang itlog ay hindi naglalaman ng isang preformed na organismo o mga bahagi nito at na ito ay binubuo ng isang homogenous na masa sa una. Sa kaibahan sa mga preformist, ang mga pananaw ni K.F. Ang lobo at iba pang mga tagasuporta ng epigenesis ay progresibo para sa kanilang panahon, dahil naglalaman sila ng ideya ng pag-unlad. Gayunpaman, nang maglaon ay lumitaw ang mga bagong sandali. Sa partikular, noong 1828, inilathala ni K. Baer ang kanyang akdang "The History of Animal Development," kung saan ipinakita niya na ang mga nilalaman ng itlog ay heterogenous, iyon ay, nakabalangkas, at ang antas ng istraktura ay tumataas habang ang embryo ay bubuo. Kaya, ipinakita ni K. Baer ang hindi pagkakapare-pareho ng parehong preformationism at epigenesis.
Ang paglaki at pag-unlad ay ang pinakamahalagang katangian ng mga bagay na may buhay. Ang paglaki ay isang pagtaas sa masa ng buong organismo bilang isang resulta ng isang pagtaas sa bilang ng mga selula, habang ang pag-unlad ay ang mga pagbabago sa husay sa katawan na tinutukoy ng pagkita ng kaibahan ng cell at morphogenesis, na tinitiyak ang mga progresibong pagbabago sa mga indibidwal, simula sa mga itlog. at nagtatapos sa kanilang pang-adultong estado.
Ontogenesis (mula sa Greek. ontos - nilalang, genesis - pag-unlad) ay ang kasaysayan (cycle) ng pag-unlad ng isang indibidwal, na nagsisimula sa pagbuo ng mga selulang mikrobyo na nagbunga nito at nagtatapos sa pagkamatay nito. Ang mga ideya tungkol sa ontogeny ay batay sa data sa paglago, pag-unlad at pagkakaiba-iba. Ang pag-aaral ng mga pangunahing prinsipyo ng ontogenesis ay mahalaga para sa pag-unawa sa biology ng tao.
Ang modernong data sa pag-unlad ng mga organismo ay tinatanggihan ang parehong preformationism at epigenesis. Sa loob ng balangkas ng mga modernong konsepto, ang pag-unlad ng isang organismo ay nauunawaan bilang isang proseso kung saan ang mga istrukturang nabuo nang mas maaga ay nagpapasigla sa pagbuo ng mga kasunod na istruktura. Ang proseso ng pag-unlad ay tinutukoy ng genetic at malapit na nauugnay sa kapaligiran. Dahil dito, ang pag-unlad ay tinutukoy ng pagkakaisa ng panloob at panlabas na mga kadahilanan.
PAGKAKAISA NG PAGLAGO AT PAG-UNLAD
Ang paglaki ng isang organismo ay isang unti-unting pagtaas ng masa nito at pagbabago sa hugis bilang resulta ng pagtaas ng bilang ng mga selula at ang kanilang pagkakaiba-iba, ang pagbuo ng mga tisyu at organo, at mga pagbabagong biochemical sa mga selula at tisyu. Kaya, ang paglago ay resulta ng dami ng mga pagbabago sa anyo ng isang pagtaas sa bilang ng mga cell (timbang ng katawan) at mga pagbabago sa husay sa anyo ng pagkita ng kaibahan ng cell at morphogenesis. Ang cell differentiation ay ang proseso kung saan ang ilang mga cell ay nagiging morphologically, biochemically, at functionally na naiiba sa ibang mga cell. Ang pagpaparami at pagkita ng kaibhan ng ilang mga selula ay palaging naaayon sa paglaki at pagkakaiba ng iba. Ang parehong mga prosesong ito ay nangyayari sa buong ikot ng buhay ng organismo. Dahil ang pagkakaiba-iba ng mga cell ay nagbabago ng kanilang hugis, at ang mga grupo ng mga cell ay kasangkot sa mga pagbabago sa hugis, ito ay sinamahan ng morphogenesis, na isang hanay ng mga proseso na tumutukoy sa istrukturang organisasyon ng mga cell at tisyu, pati na rin ang pangkalahatang morpolohiya ng mga organismo.
Maaaring masukat ang paglaki sa pamamagitan ng pagbuo ng mga kurba ng laki ng katawan, timbang, tuyong masa, numero ng cell, nilalaman ng nitrogen at iba pang mga tagapagpahiwatig batay sa mga resulta ng pagsukat.
ONTOGENESIS AT MGA URI NITO. PERIODISATION NG ONTOGENESIS
Ang ontogenesis, depende sa likas na katangian ng pag-unlad ng mga organismo, ay inuri sa direkta at hindi direkta, at samakatuwid ang isang pagkakaiba ay ginawa sa pagitan ng direkta at hindi direktang pag-unlad. Ang direktang pag-unlad ng mga organismo sa kalikasan ay nangyayari sa anyo ng non-larval at intrauterine development, habang ang hindi direktang pag-unlad ay sinusunod sa anyo ng larval development. Sa kaibahan sa ontogeny, ang kategorya ng species ay phylogeny.
Pag-unlad ng larva. Ang pag-unlad na ito ay nauunawaan bilang hindi direktang pag-unlad, dahil ang mga organismo ay may isa o higit pang mga yugto ng larval sa kanilang pag-unlad. Ang pag-unlad ng larva ay tipikal para sa mga insekto, amphibian, at echinoderms. Ang larvae ng mga hayop na ito ay humantong sa isang malayang pamumuhay, pagkatapos ay sumasailalim sa mga pagbabago. Samakatuwid, ang pag-unlad na ito ay tinatawag ding pag-unlad na may metamorphosis (tingnan sa ibaba).
Pag-unlad na hindi larva. Ang anyo ng pag-unlad na ito ay katangian ng mga organismo na direktang umuunlad, halimbawa, isda, reptilya at ibon, na ang mga itlog ay mayaman sa pula ng itlog (nutrient material). Dahil dito, ang isang makabuluhang bahagi ng ontogeny ay nagaganap sa mga itlog na inilatag sa panlabas na kapaligiran. Ang metabolismo ng mga embryo ay sinisiguro sa pamamagitan ng pagbuo ng mga pansamantalang organo, na mga embryonic membranes (yolk sac, amnion, allantois).
Pag-unlad ng intrauterine. Ang pag-unlad na ito ay katangian din ng mga organismo na direktang umuunlad, tulad ng mga mammal, kabilang ang mga tao. Dahil ang mga itlog ng mga organismo na ito ay napakahirap sa mga sustansya, ang lahat ng mahahalagang pag-andar ng mga embryo ay ibinibigay ng katawan ng ina sa pamamagitan ng pagbuo ng mga pansamantalang organo mula sa mga tisyu ng ina at ang embryo, kung saan ang pangunahing isa ay ang inunan. Sa ebolusyon, ang intrauterine development ay ang pinakabagong anyo, ngunit ito ay pinaka-kapaki-pakinabang para sa mga embryo, dahil epektibo nitong tinitiyak ang kanilang kaligtasan.
Ang ontogenesis ay nahahati sa proembryonic, embryonic at postembryonic period. Sa kaso ng mga tao, ang panahon ng pag-unlad bago ang kapanganakan ay tinatawag na prenatal o antenatal, pagkatapos ng kapanganakan - postnatal. Ang pagbuo ng embryo bago ang pagbuo ng mga organ rudiment ay tinatawag na isang embryo, pagkatapos ng pagbuo ng mga organ rudiment - isang fetus.
PROEMBRYONAL DEVELOPMENT
Ang panahong ito sa indibidwal na pag-unlad ng mga organismo ay nauugnay sa pagbuo ng mga gametes sa proseso ng gametogenesis. Ang mga male reproductive cell ay walang makabuluhang pagkakaiba mula sa iba pang mga cell, habang ang mga itlog ay naiiba dahil naglalaman sila ng maraming yolk. Isinasaalang-alang ang dami ng yolk at ang pamamahagi nito sa mga itlog, ang huli ay inuri sa tatlong uri:
1) isolecithal na itlog, naglalaman ng isang maliit na pula ng itlog, na kung saan ay naisalokal nang pantay-pantay sa buong cell. Ang mga itlog na ito ay ginawa ng mga echinoderms (sea urchins), lower chordates (lancelets), at mammals;
2) mga itlog ng telolecithal naglalaman ng isang malaking halaga ng pula ng itlog, na kung saan ay puro sa isa sa mga pole - ang vegetative. Ang ganitong mga itlog ay ginawa ng mga mollusk, amphibian, reptilya, at ibon. Halimbawa, ang mga itlog ng palaka ay binubuo ng 50% pula ng itlog, mga itlog ng manok (karaniwang mga itlog ng manok) - 95%. Sa kabilang poste (hayop) ng telolecithal na mga itlog, ang cytoplasm at nucleus ay puro;
3) centrolecithal oocytes, kung saan mayroong maliit na pula ng itlog at ito ay sumasakop sa isang sentral na posisyon. Ang cytoplasm ay matatagpuan sa periphery ng naturang mga itlog. Ang mga centrolecithal na itlog ay ginawa ng mga arthropod.
Ang panahon ng proembryonic ay nailalarawan din ng katotohanan na sa panahong ito ang mga proseso ng metabolic na nauugnay sa akumulasyon ng mga molekula ng DNA ay nangyayari sa mga gametes.
EMBRYONAL DEVELOPMENT
Embryogenesis (mula sa Greek. etbryop - mikrobyo), o panahon ng embryonic, ay nagsisimula sa pagsasanib ng mga selulang mikrobyo ng lalaki at babae, na kumakatawan sa proseso ng pagpapabunga ng mga itlog.
Sa mga organismo na nailalarawan sa pamamagitan ng intrauterine development, ang embryonic period ay nagtatapos sa kapanganakan, at sa mga organismo na nailalarawan sa pamamagitan ng larval at non-larval na uri ng pag-unlad, ang embryonic period ay nagtatapos sa paglabas ng organismo mula sa itlog o embryonic membranes, ayon sa pagkakabanggit. Sa loob ng panahon ng embryonic, ang mga yugto ng zygote, cleavage, blastula, pagbuo ng mga layer ng mikrobyo, histogenesis at organogenesis ay nakikilala.
Zygote.Ang pagpapabunga ay nagsasangkot ng isang serye ng mga proseso kung saan ang male reproductive cell ay nagpasimula ng pagbuo ng isang itlog. Sa itlog na na-activate ng male gamete, maraming pisikal at kemikal na proseso ang nagaganap, kabilang ang pagtaas ng synthesis ng protina. Ang paggalaw ng protoplasm ay humahantong sa pagtatatag ng bilateral symmetry ng itlog. Ang nuclei fuse, at ang diploid set ng mga chromosome ay naibalik. Lumilikha ito ng isang solong selulang organismo.
Naghihiwalay.Ito ay kumakatawan sa unang panahon ng pag-unlad ng zygote (fertilized egg), na binubuo ng paghahati ng zygote sa pamamagitan ng mitosis. Ang dibisyon ay nagsisimula sa hitsura ng isang tudling sa ibabaw ng itlog. Ang unang furrow ay humahantong sa pagbuo ng dalawang cell - dalawang blastomeres, ang pangalawa - apat na blastomeres, ang pangatlo - walong blastomeres (Fig. 25). Ang isang pangkat ng mga selula na nabuo bilang resulta ng sunud-sunod na pagkapira-piraso ay tinatawag na morula (mula sa Latin. morum- mulberry).
Ang biological na kahalagahan ng yugtong ito ay nakasalalay sa katotohanan na mula sa isang malaking cell, na kung saan ay isang itlog, mas maliit na mga cell ay nabuo kung saan ang ratio ng cytoplasm sa nucleus ay nabawasan.
Ang fragmentation ng zygote ay nagtatapos sa pagbuo ng isang multicellular na istraktura na tinatawag na blastula (mula sa Greek. blastos - usbong). Ang istraktura na ito ay hugis tulad ng isang vesicle na tinatawag na blastoderm, na binubuo ng isang solong layer ng mga cell. Ngayon ang mga cell na ito ay tinatawag na embryonic. Ang blastula ay katulad ng laki sa itlog. Sa panahon ng fragmentation, ang bilang ng nuclei at ang kabuuang bilang ng DNA ay tumataas. Ang maliit na halaga ng mRNA at tRNA ay na-synthesize din, habang ang ribosomal RNA ay hindi pa nakikita.
Ang lahat ng mga hayop ay dumaan sa yugto ng blastula, ngunit ang bawat kaso ay may sariling mga katangian. Sa mga mammal, ang dibisyon ay hindi pantay, kaya ang morulae ay binubuo ng iba't ibang bilang ng mga selula. Bilang karagdagan, ang isang bahagi ng mga selula ay bumubuo ng isang istraktura na tinatawag na trophoblast,

Blastocoel Larva Tadpole
anyo
kanin. 25.Fragmentation ng zygotes at pagbuo ng blastulas sa iba't ibang mga organismo: 1 - orihinal na itlog; 2 - dalawang blastomeres; 3 - apat na blastomeres; 4 - walong blastomeres; 5 - blastula; 6 - pang-adultong anyo
ang mga selula kung saan nagpapalusog sa embryo at, salamat sa mga enzyme, tinitiyak ang pagtagos ng huli sa dingding ng matris. Nang maglaon, ang mga selula ng trophoblast ay nag-alis mula sa embryo at bumubuo ng isang vesicle, na puno ng likido mula sa tisyu ng matris.
Ang biological na kahalagahan ng yugtong ito ay nakasalalay sa katotohanan na mula sa isang malaking selula, tulad ng itlog, ang mga maliliit na selula ay nabuo kung saan ang ratio ng cytoplasm sa nucleus ay nabawasan at ang nucleus ay may bagong cytoplasmic na kapaligiran.
Gastrulation(mula sa Greek gastre - cavity ng sisidlan). Ito ang proseso ng paggalaw ng mga embryonic cell kasunod ng pagbuo ng blastula, na sinamahan ng pagbuo ng dalawa o tatlong (depende sa uri ng hayop) na layer ng embryo, o ang tinatawag na germ layers (Fig. 26). ).
Ang pag-unlad (gastrulation) ng isolecithal na mga itlog ay nangyayari sa pamamagitan ng invagination (invagination) ng vegetative pole sa loob ng blastula, bilang isang resulta kung saan ang mga kabaligtaran na pole ay halos sumanib, at ang blastocoel (blastula cavity) ay halos o ganap na nawawala. Panlabas
kanin. 26.Gastrulation sa iba't ibang mga organismo: 1 - blastocoel; 2 - pangunahing mesenchyme; 3, 4, 5 - pangunahing gat
Ang layer ng mga embryonic cell ay tinatawag na ectoderm (mula sa Greek. ectos - sa labas, derma - balat), o ang panlabas na layer ng mikrobyo, habang ang panloob ay ang endoderm (mula sa Griyego. entos - sa loob), o panloob na layer ng mikrobyo. Ang lukab na nabuo sa kasong ito ay tinatawag na gastrocoel, o pangunahing bituka, ang pasukan kung saan ay tinatawag na blastopore (pangunahing bibig).
Ang pagbuo ng dalawang layer ng mikrobyo ay katangian ng mga espongha at coelenterates. Gayunpaman, ang mga chordates sa panahon ng gastrulation ay nailalarawan sa pamamagitan ng pag-unlad ng ikatlong layer ng mikrobyo - mesoderm (mula sa Greek. mesos - gitna), nabuo sa pagitan ng ectoderm at endoderm.
Ang gastrulation ay isang kinakailangang paunang kinakailangan para sa mga susunod na yugto ng pag-unlad, dahil dinadala nito ang mga selula sa isang posisyon na ginagawang posible upang bumuo ng mga organo. Ang materyal na embryonic na naiba sa tatlong embryonic anlages ay nagbibigay ng lahat ng mga tisyu at organo ng pagbuo ng embryo.
Ang pag-unlad (pagkita ng kaibhan) ng mga layer ng mikrobyo ay sinamahan ng pagbuo ng iba't ibang mga tisyu at organo mula sa kanila. Sa partikular, ang epidermis ng balat, kuko at buhok, sebaceous at sweat glands, ang nervous system (utak, spinal cord, ganglia, nerves), receptor cells ng sensory organs, lens ng mata, epithelium ng bibig. , lukab ng ilong at anus, ngipin -
naya enamel. Mula sa endoderm ang epithelium ng esophagus, tiyan, bituka, gallbladder, trachea, bronchi, baga, urethra, pati na rin ang atay, pancreas, thyroid, parathyroid at thymus glands ay bubuo. Mula sa mesoderm bumuo ng makinis na mga kalamnan, kalansay at mga kalamnan ng puso, dermis, connective tissue, buto at kartilago, dental dentin, dugo at mga daluyan ng dugo, mesentery, bato, testes at ovaries. Sa mga tao, ang utak at spinal cord ang unang naghihiwalay. Pagkatapos ng 2 buwan, halos lahat ng mga istraktura ng katawan ay lilitaw. Ang koneksyon ng embryo sa kapaligiran ay isinasagawa sa pamamagitan ng mga pansamantalang organo. Ang organogenesis ay nagtatapos sa pagtatapos ng panahon ng embryonic. Kung ang tiyak na pagbubukas ng bibig ay nabuo sa site ng pangunahing bibig (blastopore), kung gayon ang mga hayop na ito ay tinatawag na protostomes (worm, mollusks, arthropods).
Kung ang tiyak na bibig ay nabuo sa kabaligtaran na lugar, kung gayon ang mga hayop na ito ay tinatawag na deuterostomes (echinoderms, chordates).
Upang matiyak ang koneksyon ng embryo sa kapaligiran, ginagamit ang tinatawag na mga provisional organ, na pansamantalang umiiral. Depende sa uri ng mga itlog, ang mga pansamantalang organo ay iba't ibang mga istraktura. Sa isda, reptilya at ibon, kasama sa mga pansamantalang organo ang yolk sac. Sa mga mammal, ang yolk sac ay nabuo sa simula ng embryogenesis, ngunit hindi nabubuo. Mamaya nabawasan. Ang panlabas na lamad ng embryo ay tinatawag na chorion. Lumalaki ito sa matris. Ang lugar ng pinakamalaking ingrowth sa matris ay tinatawag na inunan. Ang embryo ay konektado sa inunan sa pamamagitan ng umbilical cord, o umbilical cord, na naglalaman ng mga daluyan ng dugo na nagbibigay ng sirkulasyon ng dugo ng inunan. Ang metabolismo ng fetus ay sinisiguro sa pamamagitan ng inunan.
Ang formative interaction ng mga bahagi ng embryo ay batay sa ilang mga coordinated metabolic process. Ang pattern ng pag-unlad ay heterochrony, na nauunawaan bilang ang iba't ibang pagbuo ng mga pagbabago sa organ sa paglipas ng panahon at ang iba't ibang intensity ng kanilang pag-unlad. Ang mga organ at sistemang iyon na dapat magsimulang gumana nang mas maaga ay mas mabilis na umuunlad. Halimbawa, sa mga tao, ang mga pangunahing bahagi ng itaas na mga paa ay umuunlad nang mas mabilis kaysa sa mas mababang mga paa.
Ang mga embryo ay lubhang sensitibo sa iba't ibang impluwensya. Samakatuwid, ang mga kritikal na panahon ay nakikilala, ibig sabihin, mga panahon kung kailan ang mga embryo ay pinaka-sensitibo sa mga nakakapinsalang kadahilanan. Sa kaso ng mga tao, ang mga kritikal na panahon ng embryonic ontogenesis
ay ang mga unang araw pagkatapos ng pagpapabunga, ang oras ng pagbuo ng inunan at panganganak.
Ang nuclei ng somatic cells ay may kakayahang tiyakin ang normal na pag-unlad ng mga itlog, na nilinaw sa mga eksperimento sa paglipat ng somatic cell nuclei sa mga itlog na kulang sa nuclei.
Ipinakita din ng mga eksperimento na ang pagbabago ng isang blastomere mula sa 8- at 16-cell na embryo ng tupa ng isang lahi patungo sa anucleate na kalahati ng itlog (pagkatapos putulin ang huli sa kalahati) ng isa pang lahi ay sinamahan ng pagbuo ng mga mabubuhay na embryo. at ang pagsilang ng mga tupa.
Mula noong panahon ni Hippocrates (ika-5 siglo BC), ang tanong ng mga dahilan na nagpasimula ng pagsilang ng isang fetus ay tinalakay. Sa partikular, iminungkahi mismo ni Hippocrates na ang pag-unlad ng pangsanggol ay nagsisimula ng sarili nitong kapanganakan. Ang pinakabagong eksperimentong gawain ng mga mananaliksik sa Ingles, na isinagawa sa mga tupa, ay nagpakita na sa mga tupa ang pagsisimula ng tupa ay kinokontrol ng isang kumplikadong hypothalamus + pituitary gland + adrenal glands ng fetus. Ang pinsala sa hypothalamic nuclei, ang pag-alis ng anterior lobe ng pituitary gland o adrenal gland ay nagpapatagal sa pagbubuntis ng tupa. Sa kabaligtaran, ang pagbibigay ng adenocorticotropic hormone (isang pagtatago mula sa anterior pituitary gland) o cortisol (isang pagtatago mula sa adrenal glands) sa mga tupa ay nagpapaikli sa tagal ng kanilang pagbubuntis.
Kaya, sa proseso ng pag-unlad ng mas mataas na eukaryotes, ang isang solong fertilized zygote cell, sa panahon ng karagdagang pag-unlad bilang isang resulta ng mitosis, ay nagbibigay ng mga cell ng iba't ibang uri - epithelial, nervous, bone, blood cells at iba pa, na kung saan ay nailalarawan sa pamamagitan ng isang iba't ibang morphology at macromolecular composition. Gayunpaman, katangian din ng mga cell ng iba't ibang uri na naglalaman ang mga ito ng parehong hanay ng mga gene, ngunit lubos na dalubhasa, na gumaganap lamang ng isa o ilang mga partikular na function, iyon ay, ang ilang mga gene ay "gumagana" sa mga cell, habang ang iba ay hindi aktibo. Halimbawa,

kanin. 27. Pag-unlad ng monozygotic twins
ang mga pulang selula ng dugo lamang ang tiyak sa synthesis at imbakan ng hemoglobin. Sa parehong paraan, ang mga epidermal cell lamang ang nag-synthesize ng keratin. Samakatuwid, ang mga tanong ay matagal nang lumitaw tungkol sa genetic na pagkakakilanlan ng nuclei ng mga somatic cells at tungkol sa mga mekanismo ng kontrol ng pagbuo ng mga fertilized na itlog bilang isang kinakailangan para sa pag-unawa sa mga mekanismo na pinagbabatayan ng pagkakaiba-iba ng cell. Ang mga stem cell ay may malaking kahalagahan sa cell differentiation (tingnan sa ibaba). Ipinakita ng mga eksperimento sa mga daga na ang mga cell ng isang uri ay may kakayahang mag-convert sa mga cell ng ibang uri. Sa partikular, ang pagkakaiba-iba o pagkakaiba-iba ng mga selula ng atay ay ipinakita na nagko-convert sa mga pancreatic cell.
Mula noong 50s. XX siglo Sa maraming mga laboratoryo, ang mga eksperimento ay isinagawa sa matagumpay na paglipat ng somatic cell nuclei sa mga itlog na artipisyal na pinagkaitan ng kanilang sariling nuclei. Ang isang pag-aaral ng DNA mula sa nuclei ng iba't ibang magkakaibang mga cell ay nagpakita na sa halos lahat ng mga kaso ang mga genome ay naglalaman ng parehong mga hanay ng mga sequence ng mga pares ng nucleotide. May mga kilalang eksepsiyon kung saan ang mga mammalian red blood cell ay nawawala ang kanilang nuclei sa huling yugto ng differentiation. Ngunit sa oras na ito ang mga pool
Ang ilang mga hemoglobin mRNA ay na-synthesize na, kaya ang nuclei ay hindi na kailangan ng mga pulang selula ng dugo. Kasama sa iba pang mga halimbawa ang immunoglobulin at T-cell na mga gene na binago sa panahon ng pag-unlad.
Ang isa sa mga pangunahing yugto sa direksyon ng pag-unawa sa mga mekanismo ng kontrol ng embryonic ontogenesis ay ang mga resulta ng mga eksperimento na isinagawa noong 1960-1970s. Ang English researcher na si D. Gurdon upang malaman kung ang nuclei ng mga somatic cells ay may kakayahang tiyakin ang karagdagang pag-unlad ng mga itlog kung ang mga nuclei na ito ay ipinapasok sa mga itlog kung saan ang kanilang sariling nuclei ay dati nang inalis. Sa Fig. Ipinapakita ng Figure 28 ang isang diagram ng isa sa mga eksperimentong ito, kung saan ang nuclei ng tadpole somatic cells ay inilipat sa mga itlog ng palaka na ang nuclei ay naalis na dati. Ipinakita ng mga eksperimentong ito na ang nuclei ng mga somatic cell ay talagang masisiguro ang karagdagang pag-unlad ng mga itlog, dahil nagawa nilang lagyan ng pataba ang mga itlog at "puwersa" ang mga ito na umunlad pa. Pinatunayan nito ang posibilidad ng pag-clone ng mga hayop.

kanin. 28.Scheme ng isang eksperimento sa paglipat ng somatic cell nuclei sa mga anucleated na itlog (D. Gurdon, 1968)
Nang maglaon, nagsagawa ng mga eksperimento ang iba pang mga mananaliksik kung saan ipinakita nila na ang paglipat ng mga indibidwal na blastomeres mula sa 8- at 16 na araw na mga embryo ng tupa ng isang lahi sa anucleated na kalahati ng itlog (pagkatapos putulin ang huli sa kalahati) ng isa pang lahi ay sinamahan ng ang pagbuo ng mga mabubuhay na embryo na may kasunod na kapanganakan ng mga tupa.
Sa simula ng 1997, itinatag ng mga may-akda ng Ingles na ang pagpapakilala ng somatic cell nuclei (mga selula ng mga embryo, fetus o udders ng mga adultong tupa) sa artipisyal na denucleated na mga itlog ng tupa, at pagkatapos ay ang pagtatanim ng mga itlog sa gayon ay napataba sa matris ng tupa ay sinamahan ng ang paglitaw ng pagbubuntis na sinusundan ng pagsilang ng mga tupa (Larawan 29). Ang isa sa mga tupa ay pinangalanang Dolly. Noong 2003, namatay si Dolly. Sa panahong ito, nakuha ang mga embryo ng mga daga, baka, kuneho, kabayo, daga at iba pang hayop.

Pagpapasok ng kultura sa katawan ng ina
kanin. 29.Paglipat ng somatic cell nuclei sa mga anucleated na itlog
Ang pagsusuri sa mga resultang ito ay nagpapakita na ang mga mammal ay maaaring magparami nang walang seks, na gumagawa ng mga supling ng mga hayop na ang mga selula ay naglalaman ng nuclear material na pinagmulan ng ama o ina, depende sa kasarian ng donor na tupa. Sa gayong mga cell
tanging ang cytoplasm at mitochondria ang pinanggalingan ng ina. Ang konklusyon na ito ay may napakahalagang pangkalahatang biological na kahalagahan at nagpapalawak ng aming mga pananaw sa potensyal na reproduktibo ng mga hayop. Ngunit mahalagang idagdag din na pinag-uusapan natin ang mga genetic manipulations na hindi umiiral sa kalikasan. Sa kabilang banda, sa mga praktikal na termino, ang mga genetic manipulations na ito ay kumakatawan sa isang direktang paraan ng pag-clone ng mataas na organisadong mga hayop na may ninanais na mga katangian, na may malaking kahalagahan sa ekonomiya. Sa mga terminong medikal, ang rutang ito ay maaaring gamitin sa hinaharap upang madaig ang pagkabaog ng lalaki.
Kaya, ang genetic na impormasyon na kinakailangan para sa normal na pag-unlad ng embryo ay hindi nawawala sa panahon ng pagkita ng kaibahan ng cell dahil sa tinatawag na mga stem cell, na may potensyal na umunlad sa iba't ibang uri ng mga selula ng katawan. Kapag nahati ang mga stem cell, ang bawat bagong cell ay may potensyal na manatiling isang stem cell o maging isang cell na may mas espesyal na function (mga selula ng kalamnan, mga selula ng dugo, o mga selula ng utak). Ang isang fertilized na itlog ay totipotent dahil ito ay nagbibigay ng iba't ibang uri ng mga selula sa katawan. Ang mga totipotent stem cell ay maaaring magbunga ng anumang uri ng cell maliban sa mga kinakailangan para sa pagbuo ng pangsanggol. Ang mga stem cell, na maaaring magbunga ng iba't ibang uri ng cell, ay karaniwang tinatawag multitotipotent mga selula. Ito ay itinatag na ang mga adult stem cell ay maaaring makabuo ng mga pagkakaiba-iba ng mga cell mula sa mga embryonic na hindi nauugnay na mga tisyu. Ang mga somatic cell ay mayroon ding isang ari-arian na tinatawag na totipotensi, ibig sabihin, ang kanilang genome ay naglalaman ng lahat ng impormasyong natanggap nila mula sa fertilized na itlog na nagbigay sa kanila ng pagtaas bilang resulta ng pagkakaiba-iba. Ang pagkakaroon ng mga data na ito ay walang alinlangan na nangangahulugan na ang pagkakaiba-iba ng cell ay napapailalim sa genetic control. Ang pag-aaral ng mga stem cell ay may implikasyon para sa medisina.
Ito ay itinatag na ang matinding synthesis ng protina kasunod ng pagpapabunga sa karamihan ng mga eukaryote ay hindi sinamahan ng mRNA synthesis. Ang pag-aaral ng oogenesis sa mga vertebrates, lalo na sa mga amphibian, ay nagpakita na ang masinsinang transkripsyon ay nangyayari kahit sa panahon ng prophase I (lalo na ang diplonema) ng meiosis. Samakatuwid, ang mga transcript ng gene sa anyo ng mga molekula ng mRNA o pro-mRNA ay naka-imbak sa mga itlog sa isang dormant na estado. Ito ay itinatag na sa mga embryonic cell ay may tinatawag na asymmetric
ternary division, na binubuo sa katotohanan na ang paghahati ng isang embryonic cell ay nagbubunga ng dalawang cell, kung saan isa lamang ang nagmamana ng mga protina na kasangkot sa transkripsyon. Kaya, ang hindi pantay na pamamahagi ng mga salik ng transkripsyon sa pagitan ng mga cell ng anak na babae ay humahantong sa pagpapahayag ng iba't ibang mga hanay ng mga gene sa kanila pagkatapos ng paghahati, ibig sabihin, sa pagkakaiba-iba ng cell.
Sa mga amphibian at marahil karamihan sa mga vertebrates, ang mga genetic na programa na kumokontrol sa maagang pag-unlad (bago ang yugto ng blastula) ay itinatag sa panahon ng oogenesis. Ang mga susunod na yugto ng pag-unlad, kapag nagsimula ang cellular differentiation (humigit-kumulang mula sa yugto ng gastrula), ay nangangailangan ng mga bagong programa para sa pagpapahayag ng gene. Kaya, ang pagkakaiba-iba ng cell ay nauugnay sa reprogramming ng genetic na impormasyon sa isang direksyon o iba pa.
Ang isang katangian ng pagkita ng kaibhan ng cell ay na ito ay hindi maibabalik na humahantong sa isa o ibang uri ng cell. Ang prosesong ito ay tinatawag na pagpapasiya at nasa ilalim din ng genetic control, at gaya ng ipinapalagay ngayon, ang pagkita ng kaibahan at pagpapasiya ng cell ay kinokontrol ng pakikipag-ugnayan ng mga cell batay sa mga senyas na isinasagawa ng mga kadahilanan ng paglago ng peptide sa pamamagitan ng mga receptor ng tyrosine kinase. Marahil ay maraming mga ganoong sistema. Ang isa sa mga ito ay ang pagkakaiba-iba ng mga selula ng kalamnan at nerbiyos ay kinokontrol ng mga neuroregulin, na mga protina ng lamad na kumikilos sa pamamagitan ng isa o higit pang mga receptor ng tyrosine kinase.
Ang genetic na kontrol sa pagpapasiya ay ipinapakita din sa pamamagitan ng pagkakaroon ng tinatawag na homeiotropic o homeotic mutations, na ipinakita sa mga insekto upang magdulot ng mga pagbabago sa determinasyon sa mga partikular na imaginal disc. Bilang resulta, ang ilang bahagi ng katawan ay nabubuo nang wala sa lugar. Halimbawa, sa Drosophila, binabago ng mga mutasyon ang pagpapasiya ng antennal disc sa isang disc na nabubuo sa apendiks ng isang paa na umaabot mula sa ulo. Sa mga insekto ng genus Ophthalmoptera ang mga istruktura ng pakpak ay maaaring bumuo mula sa disc ng mata. Sa mga daga, ipinakita ang pagkakaroon ng isang kumpol ng gene (komplikadong) Hox, na binubuo ng 38 mga gene at kinokontrol ang pag-unlad ng mga paa.
Ang isyu ng regulasyon ng aktibidad ng gene sa panahon ng pag-unlad ng embryonic ay may independiyenteng kahalagahan. Ito ay pinaniniwalaan na sa panahon ng pagkita ng kaibhan, ang mga gene ay kumikilos sa iba't ibang oras, na ipinahayag
sa transkripsyon sa iba't ibang magkakaibang mga selula ng iba't ibang mRNA, ibig sabihin, ang panunupil at derepression ng mga gene ay nagaganap. Halimbawa, ang bilang ng mga gene na na-transcribe sa RNA sa sea urchin blastocytes ay 10%, sa mga rat liver cells ito ay 10% din, at sa mga cattle thymus cells ito ay 15%. Ipinapalagay na ang mga non-histone na protina ay kasangkot sa kontrol ng transkripsyon na katayuan ng mga gene. Sinusuportahan ng sumusunod na data ang pagpapalagay na ito. Kapag nasa phase ang cell chromatin S na-transcribe sa system sa vitro, pagkatapos ay ang histone mRNA lamang ang na-synthesize, na sinusundan ng mga histones. Sa kabaligtaran, kapag ginamit ang chromatin ng mga cell mula sa β-phase, walang histone mRNA ang na-synthesize. Kapag ang mga non-pistone na protina ay inalis mula sa chromatin ng β-phase at pinalitan ng mga non-histone chromosomal na protina na na-synthesize sa yugto S, pagkatapos ay pagkatapos ng transkripsyon ng naturang chromatin sa vitro Ang histone mRNA ay na-synthesize. Bukod dito, kapag ang mga non-histone na protina ay nagmula sa 1-phase na mga cell, at ang DNA at mga histone ay nagmula sa S-phase na mga cell, walang histone mRNA ang na-synthesize. Ang mga resultang ito ay nagpapahiwatig na ang mga non-histone na protina na nasa chromatin ay tumutukoy sa kakayahang mag-transcribe ng mga gene na nag-encode ng mga histone Samakatuwid naniniwala na ang mga non-histone chromosomal na protina ay maaaring may mahalagang papel sa regulasyon at pagpapahayag ng mga gene sa mga eukaryote.
Iminumungkahi ng magagamit na data na ang mga protina at steroid na hormone ay kasangkot sa regulasyon ng transkripsyon sa mga hayop. Ang mga hormone ng protina (insulin) at steroid (estrogon at testosterone) ay dalawang sistema ng pagbibigay ng senyas na ginagamit sa mga intercellular na komunikasyon. Sa mas mataas na mga hayop, ang mga hormone ay na-synthesize sa mga espesyal na secretory cell. Inilabas sa daluyan ng dugo, pumapasok sila sa mga tisyu. Dahil ang mga molekula ng mga hormone ng protina ay medyo malaki, hindi sila tumagos sa mga selula, kaya ang kanilang mga epekto ay ibinibigay ng mga protina ng receptor na naisalokal sa mga lamad ng mga target na selula at intracellular na antas ng cyclic AMP (cAMP). Sa kabaligtaran, ang mga steroid hormone ay maliliit na molekula, bilang isang resulta kung saan madali silang tumagos sa mga selula sa pamamagitan ng mga lamad. Sa sandaling nasa loob ng mga selula, nagbubuklod sila sa mga partikular na protina ng receptor na matatagpuan sa cytoplasm lamang ng mga target na selula. Ang mga kumplikado ay isinasaalang-alang hormone - receptor ng protina, Nakatuon sa nuclei ng mga target na selula, ina-activate nila ang transkripsyon ng mga partikular na gene sa pamamagitan ng pakikipag-ugnayan sa ilang
mi non-histone na mga protina na nagbubuklod sa mga rehiyon ng promoter ng mga partikular na gene. Dahil dito, ang pagbubuklod ng complex hormone + protina (receptor ng protina) na may mga non-histone na protina ay nagpapalaya sa mga rehiyon ng promoter para sa paggalaw ng RNA polymerase. Ang pagbubuod ng data sa genetic control ng embryonic period sa ontogenesis ng mga organismo, maaari nating tapusin na ang kurso nito ay kinokontrol ng differential switching on at off ng pagkilos ng mga gene sa iba't ibang mga cell (tissue) sa pamamagitan ng kanilang derepression at repression.
PAG-UNLAD NG POSTEMBRYONAL
Pagkatapos ng kapanganakan ng isang organismo, ang postembryonic development nito ay nagsisimula (postnatal para sa mga tao), na sa iba't ibang mga organismo ay tumatagal mula sa ilang araw hanggang daan-daang taon, depende sa kanilang mga species. Dahil dito, ang pag-asa sa buhay ay isang uri ng hayop na katangian ng mga organismo na hindi nakadepende sa antas ng kanilang organisasyon (tingnan sa ibaba).
Sa postembryonic ontogenesis, ang pagkakaiba ay ginawa sa pagitan ng juvenile at pubertal period, pati na rin ang panahon ng katandaan, na nagtatapos sa kamatayan.
Juvenile period. Ang panahong ito (mula sa lat. juvenilis- bata) ay tinutukoy ng oras mula sa pagsilang ng organismo hanggang sa pagdadalaga. Ito ay nangyayari sa iba't ibang paraan at depende sa uri ng ontogenesis ng mga organismo. Ang panahong ito ay nailalarawan sa pamamagitan ng alinman sa direkta o hindi direktang pag-unlad.
Sa kaso ng mga organismo na nailalarawan sa pamamagitan ng direktang pag-unlad (maraming invertebrates, isda, reptilya, ibon, mammal, tao), ang mga napisa mula sa mga shell ng itlog o bagong panganak ay katulad ng mga pang-adultong anyo, na naiiba mula sa huli sa mas maliliit na sukat, pati na rin. bilang hindi pag-unlad ng mga indibidwal na organo at hindi perpektong proporsyon ng katawan (Larawan 30).
Ang isang tampok na katangian ng paglaki sa juvenile period ng mga organismo na napapailalim sa direktang pag-unlad ay ang pagkakaroon ng pagtaas sa bilang at laki ng mga selula, at ang mga proporsyon ng pagbabago ng katawan. Ang paglaki ng tao sa iba't ibang panahon ng kanyang ontogenesis ay ipinapakita sa Fig. 31. Ang paglaki ng iba't ibang organo ng tao ay hindi pantay. Halimbawa, ang paglaki ng ulo ay nagtatapos sa pagkabata, ang mga binti ay umabot sa proporsyonal na laki ng mga 10 taon. Ang panlabas na genitalia ay napakabilis na lumalaki sa pagitan ng edad na 12 at 14 na taon. Ang isang pagkakaiba ay ginawa sa pagitan ng tiyak at hindi tiyak na paglago. Ang isang tiyak na paglaki ay katangian ng mga organismo na humihinto sa paglaki sa isang tiyak na edad,
halimbawa mga insekto, mammal, tao. Ang hindi tiyak na paglaki ay katangian ng mga organismo na lumalaki sa buong buhay nila, halimbawa, mga mollusk, isda, amphibian, reptilya, at maraming uri ng halaman.

kanin. tatlumpu.Direkta at hindi direktang pag-unlad ng mga organismo ng iba't ibang species
Sa kaso ng hindi direktang pag-unlad, ang mga organismo ay sumasailalim sa mga pagbabagong tinatawag na metamorphoses (mula sa lat. metamorphosis - pagbabagong-anyo).

kanin. 31.Paglago at pag-unlad sa iba't ibang panahon ng ontogenesis ng tao
Kinakatawan nila ang mga pagbabago ng mga organismo sa panahon ng pag-unlad. Ang mga metamorphoses ay malawakang matatagpuan sa coelenterates (hydra, jellyfish, coral polyps), flatworms (fasciola), roundworms (roundworms), mollusks (oysters, mussels, octopuses), arthropods (crayfish, river crab, lobsters, shrimp, scorpions, spiders, mites , insekto) at maging sa ilang chordates (tunicates at amphibians). Sa kasong ito, ang kumpleto at hindi kumpletong metamorphoses ay nakikilala. Ang pinaka-nagpapahayag na mga anyo ng metamorphosis ay sinusunod sa mga insekto na sumasailalim sa parehong hindi kumpleto at kumpletong metamorphosis.
Ang hindi kumpletong pagbabago ay isang pag-unlad kung saan ang isang organismo ay lumalabas mula sa mga shell ng itlog, ang istraktura nito ay katulad ng istraktura ng isang pang-adultong organismo, ngunit ang laki nito ay mas maliit. Ang ganitong organismo ay tinatawag na larva. Sa panahon ng proseso ng paglaki at pag-unlad, ang laki ng larvae ay tumataas, ngunit ang umiiral na chitinized na takip ay pumipigil sa isang karagdagang pagtaas sa laki ng katawan, na humahantong sa molting, ibig sabihin, ang pagbubuhos ng chitinized na takip, kung saan mayroong isang malambot na cuticle. Ang huli ay tumuwid, at ito ay sinamahan ng pagtaas ng laki ng hayop. Pagkatapos ng ilang mga moults, ang hayop ay umabot sa kapanahunan. Ang hindi kumpletong pagbabago ay karaniwan, halimbawa, sa kaso ng pag-unlad ng mga surot.
Ang kumpletong metamorphosis ay isang pag-unlad kung saan ang isang larva ay inilabas mula sa mga shell ng itlog, na makabuluhang naiiba
sa istraktura mula sa mga indibidwal na may sapat na gulang. Halimbawa, sa mga butterflies at maraming insekto ang larvae ay mga uod. Ang mga uod ay napapailalim sa molting, at maaaring molt ng ilang beses, pagkatapos ay nagiging pupae. Mula sa huli, nabuo ang mga porma ng pang-adulto (imago), na hindi naiiba sa mga orihinal.
Sa vertebrates, ang metamorphosis ay nangyayari sa mga amphibian at bony fish. Ang yugto ng larval ay nailalarawan sa pagkakaroon ng mga pansamantalang organo, na maaaring umuulit sa mga katangian ng mga ninuno o may malinaw na adaptive na kahalagahan. Halimbawa, ang isang tadpole, na kung saan ay ang larval form ng isang palaka at inuulit ang mga katangian ng orihinal na anyo, ay nailalarawan sa pamamagitan ng isang isda-tulad ng hugis, ang pagkakaroon ng hasang paghinga at isang bilog ng sirkulasyon ng dugo. Ang mga tampok na adaptive ng tadpoles ay ang kanilang mga suckers at mahabang bituka. Ito rin ay katangian ng mga larval form na, kumpara sa mga adult na form, sila ay inangkop sa buhay sa ganap na magkakaibang mga kondisyon, na sumasakop sa ibang ecological niche at ibang lugar sa food chain. Halimbawa, ang larvae ng palaka ay may gill respiration, habang ang mga adult form ay may pulmonary respiration. Hindi tulad ng mga pang-adultong anyo, na mga carnivore, ang mga larvae ng palaka ay kumakain ng mga pagkaing halaman.
Ang pagkakasunud-sunod ng mga kaganapan sa pag-unlad ng mga organismo ay madalas na tinatawag na mga siklo ng buhay, na maaaring maging simple o kumplikado. Ang pinakasimpleng mga siklo ng pag-unlad ay katangian, halimbawa, ng mga mammal, kapag ang isang organismo ay bubuo mula sa isang fertilized na itlog, na muling gumagawa ng mga itlog, atbp. Ang mga kumplikadong biological cycle ay mga cycle sa mga hayop, na nailalarawan sa pamamagitan ng pag-unlad na may metamorphosis. Ang kaalaman tungkol sa mga biological cycle ay praktikal na kahalagahan, lalo na sa mga kaso kung saan ang mga organismo ay mga pathogen o mga carrier ng mga pathogen sa mga hayop at halaman.
Ang pag-unlad at pagkakaiba-iba na nauugnay sa metamorphosis ay ang resulta ng natural na seleksyon, dahil sa kung saan maraming mga larval form, tulad ng mga insect caterpillar at frog tadpoles, ay mas mahusay na iniangkop sa kapaligiran kaysa sa mga adult na sexually mature form.
Pagbibinata. Ang panahong ito ay tinatawag ding mature, at ito ay nauugnay sa sekswal na kapanahunan ng mga organismo. Ang pag-unlad ng mga organismo sa panahong ito ay umabot sa pinakamataas nito.
Ang paglaki at pag-unlad sa postembryonic period ay lubos na naiimpluwensyahan ng mga salik sa kapaligiran. Para sa mga halaman, ang mga mapagpasyang kadahilanan ay liwanag, halumigmig, temperatura, dami at kalidad ng mga sustansya sa lupa. Para sa mga hayop, ang tamang pagpapakain ay pinakamahalaga (ang pagkakaroon ng mga protina, carbohydrates, lipids, mineral salts, bitamina, microelements sa feed). Mahalaga rin ang oxygen, temperatura, liwanag (vitamin D synthesis).
Ang paglaki at indibidwal na pag-unlad ng mga organismo ng hayop ay napapailalim sa neurohumoral na regulasyon ng humoral at nervous regulatory mechanisms. Ang mga aktibong sangkap na tulad ng hormone na tinatawag na phytohormones ay natuklasan sa mga halaman. Ang huli ay nakakaapekto sa mahahalagang pag-andar ng mga organismo ng halaman.
Sa mga selula ng hayop, sa panahon ng kanilang mga proseso sa buhay, ang mga chemically active substance ay na-synthesize na nakakaapekto sa mga proseso ng buhay. Ang mga nerve cell ng invertebrates at vertebrates ay gumagawa ng mga sangkap na tinatawag na neurosecretes. Ang endocrine, o panloob, na mga glandula ng pagtatago ay naglalabas din ng mga sangkap na tinatawag na mga hormone. Ang mga glandula ng endocrine, lalo na ang mga nauugnay sa paglaki at pag-unlad, ay kinokontrol ng mga neurosecretion. Sa mga arthropod, ang regulasyon ng paglaki at pag-unlad ay napakahusay na inilalarawan ng epekto ng mga hormone sa pag-molting. Ang synthesis ng larval secretions ng mga cell ay kinokontrol ng mga hormone na naipon sa utak. Ang isang espesyal na glandula sa mga crustacean ay gumagawa ng isang hormone na pumipigil sa molting. Tinutukoy ng mga antas ng mga hormone na ito ang dalas ng molting. Sa mga insekto, ang hormonal regulation ng egg maturation at diapause ay naitatag.
Sa vertebrates, ang mga endocrine gland ay ang pituitary gland, pineal gland, thyroid, parathyroid, pancreas, adrenal glands at gonads, na malapit na konektado sa isa't isa. Ang pituitary gland sa mga vertebrates ay gumagawa ng gonadotropic hormone, na nagpapasigla sa aktibidad ng mga gonad. Sa mga tao, ang pituitary hormone ay nakakaapekto sa paglaki. Sa isang kakulangan, bubuo ang dwarfism; sa labis, bubuo ang gigantismo. Ang pineal gland ay gumagawa ng isang hormone na nakakaapekto sa pana-panahong pagbabagu-bago sa sekswal na aktibidad ng mga hayop. Ang thyroid hormone ay nakakaimpluwensya sa metamorphosis ng mga insekto at amphibian. Sa mga mammal, ang underdevelopment ng thyroid gland ay humahantong sa growth retardation at underdevelopment ng genital organs. Sa mga tao, dahil sa isang depekto sa thyroid gland, ang ossification at paglaki ay naantala.
(dwarfism), hindi nangyayari ang pagdadalaga, huminto ang pag-unlad ng kaisipan (cretinism). Ang adrenal glands ay gumagawa ng mga hormone na nakakaimpluwensya sa metabolismo, paglaki at pagkita ng kaibahan ng mga selula. Ang mga gonad ay gumagawa ng mga sex hormone na tumutukoy sa pangalawang sekswal na katangian. Ang pag-alis ng mga gonad ay humahantong sa hindi maibabalik na mga pagbabago sa isang bilang ng mga katangian. Halimbawa, sa castrated roosters, humihinto ang paglaki ng suklay at nawawala ang sexual instinct. Ang isang castrated na lalaki ay nakakakuha ng isang panlabas na pagkakahawig sa isang babae (isang balbas at buhok ay hindi lumalaki sa balat, ang taba ay idineposito sa dibdib at pelvic area, ang timbre ng boses ay napanatili, atbp.).
Ang mga phytohormones ng halaman ay mga auxin, cytokinin at gibberellins. Kinokontrol nila ang paglaki at paghahati ng cell, ang pagbuo ng mga bagong ugat, pag-unlad ng bulaklak at iba pang mga katangian ng halaman.
Sa lahat ng panahon ng ontogenesis, ang mga organismo ay may kakayahang ibalik ang nawala o nasira na mga bahagi ng katawan. Ang pag-aari na ito ng mga organismo ay tinatawag pagbabagong-buhay, na maaaring pisyolohikal at reparative.
Physiological regeneration - Ito ang pagpapalit ng mga nawawalang bahagi ng katawan sa panahon ng buhay ng katawan. Ang mga pagbabagong-buhay ng ganitong uri ay karaniwan sa mundo ng hayop. Halimbawa, sa mga arthropod ito ay kinakatawan ng molting, na nauugnay sa paglago. Sa mga reptilya, ang pagbabagong-buhay ay ipinahayag sa pagpapalit ng buntot at kaliskis, sa mga ibon - mga balahibo, kuko at spurs. Sa mga mammal, isang halimbawa ng physiological regeneration ay ang taunang pagpapadanak ng mga sungay ng usa.
Reparative regeneration - Ito ay ang pagpapanumbalik ng isang bahagi ng katawan ng isang organismo na marahas na napunit. Ang pagbabagong-buhay ng ganitong uri ay posible sa maraming mga hayop, ngunit ang mga pagpapakita nito ay nag-iiba. Halimbawa, karaniwan ito sa mga hydras at nauugnay sa pagpaparami ng huli, dahil ang buong organismo ay muling nabuo mula sa isang bahagi. Sa iba pang mga organismo, ang pagbabagong-buhay ay nagpapakita ng sarili sa anyo ng kakayahan ng mga indibidwal na organo na mabawi pagkatapos ng pagkawala ng anumang bahagi. Sa mga tao, ang epithelial, connective, muscle at bone tissues ay may medyo mataas na regenerative na kakayahan.
Ang mga halaman ng maraming mga species ay may kakayahang muling pagbuo.
Ang data sa pagbabagong-buhay ay may malaking kahalagahan hindi lamang sa biology. Malawakang ginagamit ang mga ito sa agrikultura, gamot, lalo na sa operasyon.
Ang katandaan bilang isang yugto ng ontogenesis. Ang katandaan ay ang huling yugto ng ontogenesis ng hayop, at ang tagal nito ay tinutukoy
ang kabuuang haba ng buhay, na nagsisilbing isang katangian ng species at nag-iiba-iba sa iba't ibang mga hayop. Ang katandaan ay pinakatumpak na pinag-aralan sa mga tao.
Mayroong iba't ibang mga kahulugan ng katandaan ng tao. Sa partikular, ang isa sa mga pinakasikat na kahulugan ay ang katandaan ay ang akumulasyon ng sunud-sunod na pagbabago na kaakibat ng pagtaas ng edad ng isang organismo at pagtaas ng posibilidad ng pagkakasakit o kamatayan nito. Ang agham ng pagtanda ng tao ay tinatawag na gerontology (mula sa Greek. geron - matanda, matanda, mga logo - ang agham). Ang gawain nito ay pag-aralan ang mga pattern ng paglipat ng edad sa pagitan ng kapanahunan at kamatayan.
Ang siyentipikong pananaliksik sa gerontology ay umaabot sa iba't ibang lugar, mula sa mga pag-aaral ng mga pagbabago sa aktibidad ng mga cellular enzymes hanggang sa pagpapaliwanag ng impluwensya ng psychological at sociological mitigations sa environmental stress sa pag-uugali ng mga matatanda.
Sa kaso ng mga tao, ang physiological old age ay nakikilala; katandaan na nauugnay sa edad ng kalendaryo; at maagang pagtanda na dulot ng panlipunang mga salik at sakit. Alinsunod sa mga rekomendasyon ng WHO, ang isang matanda ay dapat ituring na mga 60-75 taong gulang, at gulang - 75 taon o higit pa.
Ang katandaan ng tao ay nailalarawan sa pamamagitan ng isang bilang ng mga panlabas at panloob na mga palatandaan.
Kabilang sa mga panlabas na palatandaan ng katandaan, ang pinaka-kapansin-pansin ay ang pagbaba sa kinis ng mga paggalaw, mga pagbabago sa pustura, pagbaba ng pagkalastiko ng balat, bigat ng katawan, katatagan at pagkalastiko ng mga kalamnan, ang paglitaw ng mga wrinkles sa mukha at iba pang bahagi ng katawan, at pagkawala ng ngipin. Kaya, halimbawa, ayon sa pangkalahatang data, ang isang tao sa edad na 30 ay nawalan ng 2 ngipin (bilang resulta ng pagkawala), sa 40 taong gulang - 4 na ngipin, sa 50 taong gulang - 8 ngipin, at sa 60 taong gulang - 11 ngipin na. Ang unang sistema ng pagbibigay ng senyas ay sumasailalim sa mga kapansin-pansing pagbabago (ang katalinuhan ng mga pandama na organo ay napurol). Halimbawa, ang maximum na distansya kung saan nakikilala ng malulusog na tao ang ilang magkatulad na tunog ay 12 m sa 20-30 taong gulang, 10 m sa 50 taong gulang, 7 m sa 60 taong gulang, at 4 m lamang sa 70 taong gulang. Kapansin-pansin din ang mga pagbabago pangalawang sistema ng pagbibigay ng senyas (nagbabago ang intonasyon ng pagsasalita, nagiging mapurol ang boses).
Kabilang sa mga panloob na palatandaan, una sa lahat, dapat nating banggitin ang mga palatandaan tulad ng reverse development (involution) ng mga organo. Mayroong pagbaba sa laki ng atay at bato, pati na rin sa bilang
ang bilang ng mga nephron sa mga bato (sa edad na 80, halos kalahati), na binabawasan ang pag-andar ng mga bato at nakakaapekto sa metabolismo ng tubig-electrolyte. Bumababa ang pagkalastiko ng mga daluyan ng dugo, bumababa ang perfusion ng dugo ng mga tisyu at organo, at tumataas ang resistensya ng peripheral vascular. Ang mga di-organikong asing-gamot ay naiipon sa mga buto, nagbabago ang kartilago (nagpapa-calcify), at bumababa ang kakayahan ng mga organo na muling buuin. Ang mga makabuluhang pagbabago ay nangyayari sa mga cell, ang paghahati at pagpapanumbalik ng kanilang functional tone ay bumabagal, ang nilalaman ng tubig ay bumababa, ang aktibidad ng cellular enzymes ay bumababa, ang koordinasyon sa pagitan ng asimilasyon at dissimilation ay nagambala. Sa utak, ang synthesis ng protina ay nagambala, na nagreresulta sa pagbuo ng mga abnormal na protina. Ang lagkit ng mga lamad ng cell ay tumataas, ang synthesis at paggamit ng mga sex hormone ay nagambala, at ang mga pagbabago ay nangyayari sa istruktura ng mga neuron. Ang mga pagbabago sa istruktura ay nangyayari sa mga protina ng nag-uugnay na tisyu at ang pagkalastiko ng tisyu na ito. Ang mga reaksyon ng immunological ay humina, at ang posibilidad ng mga reaksyon ng autoimmune ay tumataas. Ang mga pag-andar ng mga endocrine system, lalo na ang mga gonad, ay nabawasan. Ang pag-uugali ng iba pang mga palatandaan sa katandaan ay ipinapakita sa Fig. 32.

kanin. 32.Mga pagbabago sa ilang katangian ng tao sa edad: 1 - bilis ng mga impulses ng nerve; 2 - antas ng basal metabolismo; 3 - index ng puso; 4 - antas ng pagsasala ng bato para sa insulin; 5 - tidal volume ng mga baga; 6 - antas ng daloy ng plasma sa mga bato
Ang pagnanais na maunawaan ang likas na katangian ng pagtanda ng katawan ay nasa loob ng mahabang panahon. Sa Sinaunang Greece, naniniwala si Hippocrates na ang pagtanda ay nauugnay sa labis na pagkain at hindi sapat na pagkakalantad sa sariwang hangin. Ayon kay Aristotle, ang pagtanda ay sanhi ng pagkonsumo ng thermal energy ng katawan. Napansin din ni Galen ang kahalagahan ng pagkain bilang salik sa pagtanda. Ngunit sa loob ng mahabang panahon ay walang sapat na pang-agham na data upang lubos na maunawaan ang problemang ito. Noong ika-19 na siglo lamang. Nagkaroon ng ilang pag-unlad sa pag-aaral ng pagtanda, at nagsimulang lumitaw ang mga teorya ng pagtanda.
Ang isa sa mga unang pinakatanyag na teorya ng pagtanda ng katawan ng tao ay ang teorya ng Aleman na doktor na si H. Hufeland (1762-1836), na nagbigay ng kahalagahan sa aktibidad ng paggawa sa mahabang buhay. Narinig natin ang kanyang pahayag na walang ni isang tamad na nabuhay hanggang sa pagtanda. Mas sikat pa endocrine theory ng pagtanda, na nagmula sa mga eksperimento na isinagawa kahit sa kalagitnaan ng siglo bago ang huli ni Berthold (1849), na nagpakita na ang paglipat ng mga testes mula sa isang hayop patungo sa isa pa ay sinamahan ng pag-unlad ng pangalawang sekswal na katangian. Nang maglaon, ang Pranses na physiologist na si C. Brown-Séquard (1818-1894), batay sa mga resulta ng pag-iniksyon sa kanyang sarili ng mga extract mula sa testes, ay nagtalo na ang mga iniksyon na ito ay gumagawa ng isang kapaki-pakinabang at nakapagpapasiglang epekto. Sa simula ng ika-20 siglo. Mayroon nang paniniwala na ang pagsisimula ng katandaan ay nauugnay sa pagkalipol ng aktibidad ng mga glandula ng endocrine, lalo na ang mga gonad. Noong 20-30s. XX siglo Batay sa paniniwalang ito, maraming operasyon ang ginawa sa iba't ibang bansa upang pabatain ang mga matatanda o matatanda. Halimbawa, itinali ni G. Steinach sa Austria ang mga spermatic cord ng mga lalaki, na humantong sa pagtigil ng panlabas na pagtatago ng mga gonad at diumano sa ilang pagpapabata. S.A. Inilipat ni Voronov sa France ang mga testes mula sa mga batang hayop hanggang sa mga matanda at mula sa mga unggoy sa mga lalaki, at si Tushnov sa USSR ay nagpabata ng mga tandang sa pamamagitan ng pag-iniksyon sa kanila ng mga histolysates ng mga gonad. Ang lahat ng mga operasyong ito ay humantong sa ilang mga epekto, ngunit pansamantala lamang. Pagkatapos ng mga epektong ito, nagpatuloy ang proseso ng pagtanda, at mas matindi pa.
Sa simula ng ika-20 siglo. bumangon teorya ng microbiological ng pagtanda, ang lumikha nito ay si I.I. Mechnikov, na nakikilala sa pagitan ng physiological at pathological na katandaan. Naniniwala siya na ang katandaan ng tao ay pathological, i.e. napaaga. Ang batayan ng mga ideya ni I.I Ang doktrina ni Mechnikov ng orthobiosis (orthos -
tama, bios - buhay), ayon sa kung saan ang pangunahing sanhi ng pagtanda ay pinsala sa mga selula ng nerbiyos sa pamamagitan ng mga produktong pagkalasing na nabuo bilang resulta ng pagkabulok sa malaking bituka. Pagbuo ng doktrina ng isang normal na pamumuhay (pagsunod sa mga panuntunan sa kalinisan, regular na trabaho, pag-iwas sa masamang gawi), I.I. Iminungkahi din ni Mechnikov ang isang paraan upang sugpuin ang putrefactive na bituka na bakterya sa pamamagitan ng pagkonsumo ng fermented milk products.
Noong 30s XX siglo naging laganap teorya tungkol sa papel ng central nervous system (CNS) sa pagtanda. Ang lumikha ng teoryang ito ay si I.P. Pavlov, na nagtatag ng integrating role ng central nervous system sa normal na paggana ng mga organismo. Mga tagasunod ng I.P. Ang mga eksperimento ni Pavlov sa mga hayop ay nagpakita na ang napaaga na pagtanda ay sanhi ng nervous shocks at matagal na nervous overstrain.
Worth mentioning teorya ng mga pagbabago na nauugnay sa edad sa connective tissue, nabuo noong 30s. XX siglo A.A. Bogomolets (1881-1946). Naniniwala siya na ang physiological activity ng katawan ay sinisiguro ng connective tissue (bone tissue, cartilage, tendons, ligaments at fibrous connective tissue) at ang mga pagbabago sa colloidal state ng mga cell, pagkawala ng kanilang turgor, atbp. ay tumutukoy sa mga pagbabagong nauugnay sa edad. sa mga organismo. Ipinapahiwatig ng modernong data ang kahalagahan ng akumulasyon ng calcium sa mga nag-uugnay na tisyu, dahil nag-aambag ito sa pagkawala ng pagkalastiko nito, pati na rin ang pagpapatigas ng mga daluyan ng dugo.
Ang mga modernong diskarte sa pag-unawa sa kakanyahan at mga mekanismo ng pag-iipon ay nailalarawan sa pamamagitan ng malawakang paggamit ng data mula sa physicochemical biology, at lalo na ang mga tagumpay ng molecular genetics. Ang pinaka-karaniwang modernong mga ideya tungkol sa mekanismo ng pagtanda ay kumukulo sa katotohanan na sa panahon ng buhay, ang mga somatic mutations ay naipon sa mga selula ng katawan, bilang isang resulta kung saan ang synthesis ng mga may sira na protina o hindi naayos na mga cross-link ng DNA na may mga protina ay nangyayari. Dahil ang mga may sira na protina ay gumaganap ng isang disintegrating na papel sa cellular metabolism, ito ay humahantong sa pagtanda. Sa kaso ng mga kulturang fibroblast, ang mga protina at mRNA na nauugnay sa mga lumang selula ay ipinakita upang sugpuin ang synthesis ng DNA sa mga batang fibroblast.
Mayroon ding kilalang hypothesis ayon sa kung saan ang pagtanda ay itinuturing na resulta ng mga pagbabago sa mitochondrial metabolites na may kasunod na dysfunction ng enzymes.
Sa mga tao, ang pagkakaroon ng mga gene ay ipinakita na tumutukoy sa tiyempo ng pag-unlad ng namamana na mga degenerative na proseso na nauugnay sa pagtanda. Ang ilang mga mananaliksik ay naniniwala na ang sanhi ng pagtanda ay ang mga pagbabago sa immunological defense system ng katawan, lalo na ang mga autoimmune na reaksyon sa mga istruktura ng katawan na napakahalaga. Sa wakas, kapag ipinapaliwanag ang mga mekanismo ng pagtanda, binibigyang pansin ng mga eksperto ang pinsala sa protina na nauugnay sa pagbuo ng mga libreng radikal. Sa wakas, ang kahalagahan ay minsan ay nakakabit sa mga hydrolase na inilabas pagkatapos ng pagkasira ng mga lysosome, na sumisira sa mga selula.
Gayunpaman, ang isang komprehensibong teorya ng pagtanda ay hindi pa nagagawa, dahil malinaw na wala sa mga teoryang ito ang makapag-iisa na ipaliwanag ang mga mekanismo ng pagtanda.
Kamatayan.Ang kamatayan ay ang huling yugto ng ontogenesis. Ang tanong ng kamatayan sa biology ay sumasakop sa isang espesyal na lugar, dahil ang pakiramdam ng kamatayan "... ay ganap na likas sa kalikasan ng tao at palaging isa sa mga pinakadakilang alalahanin ng tao" (I.I. Mechnikov, 1913). Bukod dito, ang tanong ng kamatayan ay nasa sentro ng atensyon ng lahat ng pilosopikal at relihiyosong mga turo, bagaman ang pilosopiya ng kamatayan ay ipinakita sa iba't ibang mga makasaysayang panahon. Sa sinaunang mundo, pinagtatalunan nina Socrates at Plato ang imortalidad ng kaluluwa, habang tinanggihan ni Aristotle ang ideya ni Plato tungkol sa imortalidad ng kaluluwa at naniniwala sa imortalidad ng espiritu ng tao, na patuloy na nabubuhay pagkatapos ng kamatayan ng isang tao. Kinilala din nina Cicero at Seneca ang buhay sa hinaharap, ngunit itinuturing ni Marcus Aurelius ang kamatayan bilang isang natural na kababalaghan na dapat tanggapin nang walang reklamo. Noong ika-18 siglo Naniniwala rin sina I. Kant at I. Fichte (1762-1814) sa isang buhay sa hinaharap, at si G. Hegel ay sumunod sa paniniwala na ang kaluluwa ay hinihigop ng isang "ganap na nilalang," bagaman ang likas na katangian ng "pagiging" na ito ay hindi nahayag. .
Alinsunod sa lahat ng kilalang turo ng relihiyon, ang buhay sa lupa ng isang tao ay nagpapatuloy pagkatapos ng kanyang kamatayan, at ang isang tao ay dapat na walang pagod na maghanda para sa hinaharap na kamatayan. Gayunpaman, ang mga natural na siyentipiko at pilosopo na hindi kumikilala sa imortalidad ay naniniwala at naniniwala pa rin na ang kamatayan ay, gaya ng paulit-ulit na idiniin ng I.I. Mechnikov, ang natural na kinalabasan ng buhay ng isang organismo. Ang isang mas matalinghagang kahulugan ng kamatayan ay na ito ay "...ay isang malinaw na tagumpay ng kawalang-kabuluhan laban sa kahulugan, kaguluhan sa kalawakan" (V. Solovyov, 1894).
Iminumungkahi ng siyentipikong ebidensya na sa mga single-celled na organismo (halaman at hayop), ang kamatayan ay dapat na makilala mula sa pagtigil.
kaalaman sa kanilang pag-iral. Kamatayan ang kanilang pagkasira, habang ang pagtigil ng pag-iral ay nauugnay sa kanilang pagkakahati. Dahil dito, ang kahinaan ng mga single-celled na organismo ay binabayaran ng kanilang pagpaparami. Sa mga multicellular na halaman at hayop, ang kamatayan ay nasa buong kahulugan ng salitang katapusan ng buhay ng organismo.
Sa mga tao, ang posibilidad ng kamatayan ay tumataas sa panahon ng pagdadalaga. Sa mga umuunlad na bansa sa partikular, ang posibilidad ng kamatayan ay tumataas nang halos exponentially pagkatapos ng edad na 28.
Mayroong klinikal at biyolohikal na pagkamatay ng isang tao. Ang klinikal na kamatayan ay ipinahayag sa pagkawala ng malay, pagtigil ng tibok ng puso at paghinga, ngunit karamihan sa mga selula at organo ay nananatiling buhay. Nagaganap ang self-renewal ng mga cell at nagpapatuloy ang peristalsis ng bituka. Ang klinikal na kamatayan ay hindi "umaabot" sa biological na kamatayan, dahil ito ay nababaligtad, dahil sa isang estado ng klinikal na kamatayan ang isa ay maaaring "bumalik" sa buhay. Halimbawa, ang mga aso ay "ibinalik" sa buhay pagkatapos ng 5-6 minuto, ang mga tao - pagkatapos ng 6-7 minuto mula sa simula ng klinikal na kamatayan. Ang biological na kamatayan ay nailalarawan sa pamamagitan ng katotohanan na ito ay hindi maibabalik. Ang pagtigil sa tibok ng puso at paghinga ay sinamahan ng pagtigil ng mga proseso ng pag-renew ng sarili, pagkamatay at pagkabulok ng mga selula. Gayunpaman, ang pagkamatay ng cell ay hindi nagsisimula sa lahat ng mga organo sa parehong oras. Una, ang cerebral cortex ay namatay, pagkatapos ay ang epithelial cells ng bituka, baga, atay, kalamnan cell, at puso ay namatay.
Ang mga hakbang para sa resuscitation (revival) ng mga organismo ay batay sa mga ideya tungkol sa klinikal na kamatayan, na may pambihirang kahalagahan sa modernong medisina.
HABA NG BUHAY
Ang paghahambing ng data sa pag-asa sa buhay ng mga kinatawan ng flora at fauna ay nagpapakita na sa mga halaman at hayop, ang iba't ibang mga organismo ay nabubuhay sa iba't ibang panahon. Halimbawa, ang mga mala-damo na halaman (ligaw at nilinang) ay nabubuhay nang isang panahon. Sa kabaligtaran, ang kanilang makahoy na mga halaman ay nailalarawan sa pamamagitan ng mas mahabang buhay. Halimbawa, ang cherry ay nabubuhay ng 100 taon, spruce - 1000 taon, oak -
2000 taon, pine - hanggang 2000-4000 taon.
Ang mga isda ng maraming mga species ay nabubuhay ng 55-80 taon, mga palaka - 16 na taon, mga buwaya - 50-60 taon, mga ibon ng ilang mga species - hanggang sa 100 taon. Ang habang-buhay ng mga mammal ay mas maikli.
Halimbawa, ang maliliit na hayop ay nabubuhay ng 20-25 taon, baka - 30 taon o higit pa, kabayo - hanggang 30 taon, aso - 20 taon o higit pa, lobo - 15 taon, oso - 50 taon, elepante - 100 taon. Sa mga mammal, ang mga tao ang pinakamahabang buhay. Maraming tao ang nabuhay hanggang 115-120 taong gulang o higit pa.
Alinsunod sa umiiral na mga ideya, ang pag-asa sa buhay ay isang tiyak na uri ng dami ng katangiang napapailalim sa kontrol ng genotype. Ito ay pinaniniwalaan na ang genetically na tinutukoy na pag-asa sa buhay ay nauugnay sa mga panahon ng ontogenesis (Talahanayan 5).
Talahanayan 5.Depende sa antas ng fibroblast sa panahon ng paglilinang sa edad ng donor
Ipinapalagay na ang haba ng buhay ng mga species ay isang evolutionary acquisition ng species. Tulad ng para sa mahabang buhay ng mga indibidwal, maaari itong ipaliwanag sa pamamagitan ng pagpapalagay ng pagkakaroon ng mga kumbinasyon ng ilang mga gene sa kanilang mga genotype, o isang maliit na bilang o kumpletong kawalan ng mga mutasyon sa kanilang mga selula.
A.A. Bogomolets at I.I. Kinakalkula ni Schmalhausen na ang natural na pag-asa sa buhay ng isang tao ay dapat na 120-150 taon. Gayunpaman, iilan lamang ang nabubuhay hanggang sa edad na ito. Samakatuwid, ang aktwal na pag-asa sa buhay ay hindi tumutugma sa natural na pag-asa sa buhay.
Ang average na pag-asa sa buhay ay naiimpluwensyahan ng ilang salik (pagbaba ng dami ng namamatay sa sanggol, pagiging epektibo ng pagkontrol sa impeksyon, pagsulong sa operasyon, pagpapabuti sa nutrisyon at pangkalahatang kondisyon ng pamumuhay). Ang mga pangunahing dahilan ng pagbaba ng pag-asa sa buhay ay ang pagkamatay mula sa gutom, sakit, at hindi sapat na pangangalagang medikal. Sa Russia, sa mga nakaraang taon, isang negatibong populasyon ang naobserbahan sa lahat ng mga lungsod at kanayunan ng Russia dahil sa pagbaba sa rate ng kapanganakan at mataas na dami ng namamatay.
Sa mga terminong medikal, ang pag-asa sa buhay ay isang tagapagpahiwatig ng kalusugan ng isang bansa. Ang USSR ay unang niraranggo sa mundo sa mga tuntunin ng bilang ng mga matatanda. Halimbawa, bawat 1 milyong naninirahan ay mayroong 104 katao na higit sa 90 taong gulang, habang sa England - 6, France - 7
at ang USA - 15 tao. Sa kasalukuyan, mayroong pagbabago sa mga hangganan ng populasyon ng nagtatrabaho kumpara, halimbawa, sa 30s. XX siglo Ang agwat sa pagitan ng edad ng pagreretiro at mga antas ng aktibidad ng mga tao ay magkakaiba din. Noong 1982, ang World Assembly on the Problems of the Aging of the World Population ay ginanap sa Vienna, kung saan ang mga pagtataya ay nabuo sa isyung ito hanggang 2025. Ipinapalagay na ang bilang ng mga taong may edad na 60 taong gulang at mas matanda ay tataas ng 5 beses kumpara hanggang 1950, mga taong higit sa 80 taong gulang - 7 beses. Ayon sa internasyonal na forum na ito, may mga pagkakaiba sa rate ng pagtanda ng populasyon sa iba't ibang mga tao, bansa at rehiyon.
Ang Geriatrics ay isang agham na ang gawain ay bumuo ng mga paraan upang gawing normal ang pagbabago ng mga function ng isang tumatanda na katawan. Gayunpaman, ang modernong gamot ay wala pang mga pamamaraan at paraan ng pag-impluwensya sa mga normal na proseso ng physiological. Samakatuwid, ang papel ng mga geriatric ay limitado sa paggamot ng mga sakit na lumitaw sa katandaan at ang pag-aalis (kung maaari) ng mga kadahilanan ng panganib na nagdudulot ng maagang pagtanda.
MGA ISYU PARA SA TALAKAYAN
1. Ano ang naiintindihan mo sa sekswal na pagpaparami ng mga organismo at ano ang biological na papel nito?
2. Ilarawan ang asexual reproduction at pangalanan ang mga anyo nito.
3. Ilarawan ang mga tampok ng prosesong sekswal sa unicellular at multicellular na organismo.
4. Ano ang gametogenesis?
5. Ano ang tungkulin ng bawat uri ng gamete?
6. Anong mga yugto ang alam mo sa pagbuo ng mga gametes?
7. Ano ang mga pagkakatulad at pagkakaiba sa pagitan ng spermatogenesis at oogenesis?
8. Ano ang meiosis at ano ang biological significance nito?
9. Ilarawan ang mga yugto ng meiosis.
10. Ang pagtawid ba ay nakakaapekto sa mga resulta ng meiosis at paano?
11. Ilarawan ang kakanyahan ng pagpapabunga.
12. Ano ang pagkakaiba sa pagitan ng zygogenesis at parthenogenesis?
13. Ano ang biological na papel ng alternating haploidy at diploidy?
14. Ano ang sexual dimorphism? Ano ang naiintindihan mo sa hermaphroditism? Ang mga kaso ba ng hermaphroditism ay sinusunod sa mga tao at gaano kadalas?
15. Paano mo maiisip ang ebolusyon ng mga paraan ng pagpaparami?
16. Ano ang naiintindihan mo sa paglaki at pag-unlad ng mga organismo? Ano ang kaugnayan sa pagitan ng paglaki ng cell at pagkita ng kaibhan?
17. Ano ang mga molecular na batayan ng cell differentiation? Ano ang alam mo tungkol sa mga stem cell?
18. Bumuo ng konsepto ng ontogenesis at pangalanan ang mga panahon ng ontogenesis.
19. Ano ang mga pagkakaiba sa pagitan ng direkta at hindi direktang pag-unlad?
20. Ano ang epekto ng pagpapabunga sa mga itlog?
21. Sa anong yugto ng pagpapatupad ng genetic information isinasagawa ang kontrol sa pagkilos ng mga gene?
22. Paano nagiging multicellular structure ang fertilized egg?
23. Paano nagiging iba ang pagbuo ng mga selula at tisyu sa bawat isa sa panahon ng pag-unlad?
24. Maaari bang maibalik ang nawala o nasirang bahagi ng katawan?
25. Ano ang koneksyon sa ontogenesis sa pagitan ng pagtanda at pag-asa sa buhay?
26. Bumalangkas at tukuyin ang pagkakaiba sa pagitan ng natural (malamang) at aktwal na pag-asa sa buhay.
27. Anong mga teorya ng pagtanda ng mga organismo ang alam mo?
29. Anong mga salik ang nakakaimpluwensya sa pag-asa sa buhay?
30. Ano ang batayan ng pag-clone ng mga organismo? Magbigay ng mga halimbawa ng pag-clone ng mga organismo.
31. Ang tamud ba o ang mga nilalaman nito ay tumagos sa selula?
32. Ano ang ipinahihiwatig ng cloning data?
33. Preformationism at epigenesis sa kasaysayan ng biology. Mayroon ba silang pang-agham na kahalagahan?
34. Ano ang mga pagkakaiba sa pagitan ng klinikal at biyolohikal na kamatayan?
Ang pagpaparami ay ang kakayahan ng mga organismo na bumuo ng kanilang sariling uri. Ang pagpaparami ay isa sa pinakamahalagang katangian ng buhay at posible dahil sa pangkalahatang kakayahan ng mga organismo na makagawa ng mga supling. Gayunpaman, ang mga agarang inapo ay hindi palaging katulad ng kanilang mga magulang. Halimbawa, mula sa mga spore ng pako, maraming mga supling ang lumalaki, na kinakatawan ng mga shoots na hindi katulad ng halaman na nagdadala ng spore ng ina. Sa paglaki, sa turn, lumilitaw ang isang halaman na hindi katulad nito - isang sporophyte. Ang kababalaghang ito ay tinatawag na alternation of generations.
Ang isang espesyal na anyo ng sekswal na pagpaparami ay parthenogenesis, o virgin reproduction, ang pagbuo ng isang organismo mula sa isang hindi pa nataba na itlog. Ang anyo ng pagpaparami na ito ay pangunahing katangian ng mga species na may maikling siklo ng buhay na may binibigkas na mga pagbabago sa panahon. Ang parthenogenesis ay maaaring haploid o diploid. Sa haploid (generative) parthenogenesis, isang bagong organismo ang bubuo mula sa isang haploid na itlog. Ang mga resultang indibidwal ay maaaring lalaki lamang, babae lamang, o pareho. Depende ito sa chromosomal sex determination.
Ang indibidwal na pag-unlad ng isang organismo, o ontogenesis, ay isang hanay ng mga sunud-sunod na pagbabagong morphological, physiological at biochemical na pinagdaanan ng organismo mula sa sandali ng pagsisimula nito hanggang sa kamatayan. Sa panahon ng ontogenesis, ang pagpapatupad ng namamana na impormasyon na natanggap ng katawan mula sa mga magulang nito ay nangyayari. Mayroong dalawang pangunahing panahon sa ontogenesis - embryonic at postembryonic. Sa yugto ng embryonic, ang isang embryo ay nabuo sa mga hayop, kung saan ang mga pangunahing sistema ng organ ay nabuo. Sa postembryonic period, ang mga proseso ng pag-unlad ay nakumpleto, ang pagdadalaga, pagpaparami, pagtanda at kamatayan ay nangyayari.
