წყალბადის ბმები წარმოიქმნება ერთი ბაზის ამინოჯგუფსა და მეორის კარბონილის ჯგუფს შორის, აგრეთვე ამიდისა და იმინის აზოტის ატომებს შორის. მაგალითად, ორი წყალბადის ბმა იქმნება ადენინსა და თიმინს შორის და ეს ჰეტეროციკლური ფუძეები ქმნიან დამატებით წყვილს. ეს ნიშნავს, რომ ერთი ჯაჭვის ადენინის ფუძე შეესაბამება მეორე ჯაჭვის თიმინურ ფუძეს. დამატებითი ბაზის კიდევ ერთი წყვილი არის გუანინი და ციტოზინი, რომელთა შორის არის სამი წყალბადის ბმა.
წყვილი დამატებითი ბაზის მიერ დაკავებული ტერიტორიები დაახლოებით ერთნაირია.
ბაზების კომპლემენტარულობა საფუძვლად უდევს E. Chargaff-ის მიერ ჩამოყალიბებულ შაბლონებს (ჩარგაფის წესები):
1. - პურინის ფუძეების რაოდენობა უდრის პირიმიდინის ფუძეების რაოდენობას;
2. - ადენინის რაოდენობა ტოლია თიმინის რაოდენობას; გუანინის რაოდენობა უდრის ციტოზინის რაოდენობას;
3. - ადენინის და ციტოზინის ჯამი უდრის გუანინისა და თიმინის ჯამს.
(A / T \u003d C / G \u003d 1).
ორჯაჭვიანი დნმ დამატებითი პოლინუკლეოტიდური ჯაჭვებით იძლევა მოლეკულის თვითგაორმაგების (რეპლიკაციის) შესაძლებლობას. ეს რთული პროცესი შეიძლება წარმოდგენილი იყოს შემდეგნაირად: გაორმაგებამდე წყალბადის ბმები იშლება და ორი ჯაჭვი იხსნება და განსხვავდება. შემდეგ თითოეული ღერი ემსახურება როგორც შაბლონს მასზე დამატებითი ღეროს ფორმირებისთვის. რეპლიკაციის შემდეგ წარმოიქმნება ორი ქალიშვილი დნმ-ის მოლეკულა, რომელთაგან თითოეულში ერთი სპირალი აღებულია მშობლის დნმ-დან, ხოლო მეორე (შემავსებელი) ხელახლა სინთეზირებულია.
დნმ-ის ორმაგ სპირალს აქვს რამდენიმე ფორმა, რაც დამოკიდებულია მოლეკულის ჰიდრატაციის ხარისხზე. ფორმები განსხვავდება ბაზის წყვილების სიბრტყის მდებარეობით სპირალის ღერძთან მიმართებაში, ერთმანეთთან, სპირალის გადახვევის ვარიანტის მიხედვით (მარჯვნივ, მარცხნივ) და სხვა მახასიათებლების მიხედვით.
ჯაჭვებს შორის არის ორი "ბეწვი" - დიდი და პატარა. ამ ღარებში ცილებს შეუძლიათ კონკრეტულად ურთიერთქმედება გარკვეულ ატომებთან. ნუკლეინის მჟავა, ამოიცნობს სპეციფიკურ ნუკლეოტიდურ თანმიმდევრობებს დარღვევის გარეშე დამატებითი ურთიერთქმედებებიორმაგი სპირალი. დადგენილია, რომ ასეთი ურთიერთქმედების გამო, მარეგულირებელ პროტეინებს შეუძლიათ გააკონტროლონ გენის ექსპრესია.


დნმ-ის მოლეკულის მეორადი სტრუქტურის ფორმები
რნმ-ის მეორადი სტრუქტურა
რნმ-ის მოლეკულა აგებულია ერთი პოლინუკლეოტიდური ჯაჭვიდან. ნუკლეოტიდების რაოდენობა შეიძლება იყოს 75-დან რამდენიმე ათასამდე და მოლეკულური მასაშეიცვლება 25 ათასიდან რამდენიმე მილიონამდე დიაპაზონში.
რნმ პოლიპეპტიდურ ჯაჭვს არ აქვს მკაცრად განსაზღვრული სტრუქტურა. მას შეუძლია თავის თავზე იკეცოს და შექმნას ცალკეული ორჯაჭვიანი სექციები წყალბადის ბმებიპურინისა და პირმიდინის ფუძეებს შორის. რნმ-ში წყალბადის ბმები არ ემორჩილება მკაცრი წესები vDNA-ს მსგავსად.
tRNA დეტალურად არის შესწავლილი. tRNA მოლეკულას აქვს მეორადი სტრუქტურა, რომელიც შედგება ოთხი სპირალური განყოფილებისგან, სამი, ოთხი ერთჯაჭვიანი მარყუჟისგან. ამ სტრუქტურას "სამყურას" უწოდებენ.
მარჯვნივ არის ყველაზე დიდი ადამიანის დნმ-ის სპირალი, რომელიც აშენებულია ადამიანებისგან ვარნას სანაპიროზე (ბულგარეთი), რომელიც გინესის რეკორდების წიგნში შევიდა 2016 წლის 23 აპრილს.
Დეზოქსირიბონუკლეინის მჟავა. Ზოგადი ინფორმაცია
დნმ (დეზოქსირიბონუკლეინის მჟავა) - ერთგვარი ცხოვრების გეგმა, რთული კოდი, რომელიც შეიცავს მონაცემებს მემკვიდრეობითი ინფორმაციის შესახებ. ამ რთულ მაკრომოლეკულას შეუძლია შეინახოს და გადასცეს მემკვიდრეობითი გენეტიკური ინფორმაცია თაობიდან თაობას. დნმ განსაზღვრავს ნებისმიერი ცოცხალი ორგანიზმის ისეთ თვისებებს, როგორიცაა მემკვიდრეობა და ცვალებადობა. მასში კოდირებული ინფორმაცია განსაზღვრავს ნებისმიერი ცოცხალი ორგანიზმის განვითარების მთელ პროგრამას. გენეტიკურად ჩადებული ფაქტორები წინასწარ განსაზღვრავს როგორც ადამიანის, ისე ნებისმიერი სხვა ორგანიზმის ცხოვრების მთელ კურსს. ხელოვნური ან ბუნებრივი გავლენა გარე გარემოშეუძლია მხოლოდ ოდნავ იმოქმედოს ცალკეული გენეტიკური მახასიათებლების საერთო სიმძიმეზე ან გავლენა მოახდინოს დაპროგრამებული პროცესების განვითარებაზე.
Დეზოქსირიბონუკლეინის მჟავა(დნმ) არის მაკრომოლეკულა (სამიდან ერთ-ერთი მთავარი, დანარჩენი ორი არის რნმ და ცილა), რომელიც უზრუნველყოფს შენახვას, გადაცემას თაობიდან თაობას და ცოცხალი ორგანიზმების განვითარებისა და ფუნქციონირების გენეტიკური პროგრამის განხორციელებას. დნმ შეიცავს ინფორმაციას სტრუქტურის შესახებ სხვადასხვა სახისრნმ და ცილები.
ევკარიოტულ უჯრედებში (ცხოველები, მცენარეები და სოკოები) დნმ გვხვდება უჯრედის ბირთვში, როგორც ქრომოსომების ნაწილი, ასევე ზოგიერთ უჯრედულ ორგანელებში (მიტოქონდრია და პლასტიდები). პროკარიოტული ორგანიზმების (ბაქტერიები და არქეები) უჯრედებში შიგნიდან მიმაგრებულია დნმ-ის წრიული ან ხაზოვანი მოლეკულა, ე.წ. უჯრედის მემბრანა. მათ და ქვედა ევკარიოტებს (მაგალითად, საფუარს) ასევე აქვთ მცირე ავტონომიური, ძირითადად წრიული დნმ-ის მოლეკულები, რომლებსაც პლაზმიდები ეწოდება.
თან ქიმიური წერტილიდნმ არის გრძელი პოლიმერული მოლეკულა, რომელიც შედგება განმეორებადი ერთეულებისგან, რომელსაც ეწოდება ნუკლეოტიდები. თითოეული ნუკლეოტიდი შედგება აზოტოვანი ფუძისგან, შაქრისგან (დეოქსირიბოზა) და ფოსფატის ჯგუფისგან. ჯაჭვში ნუკლეოტიდებს შორის კავშირები წარმოიქმნება დეზოქსირიბოზას მიერ ( თან) და ფოსფატი ( ფ) ჯგუფები (ფოსფოდიესტერული ბმები).

ბრინჯი. 2. ნუკლეტიდი შედგება აზოტოვანი ფუძისგან, შაქრისგან (დეზოქსირიბოზა) და ფოსფატის ჯგუფისგან.
შემთხვევების აბსოლუტურ უმრავლესობაში (გარდა ზოგიერთი ვირუსისა, რომელიც შეიცავს ერთჯაჭვიან დნმ-ს), დნმ მაკრომოლეკულა შედგება ორი ჯაჭვისგან, რომლებიც ორიენტირებულია აზოტოვანი ბაზებით ერთმანეთზე. ეს ორჯაჭვიანი მოლეკულა ხვეულია სპირალში.
დნმ-ში ნაპოვნია აზოტოვანი ფუძის ოთხი ტიპი (ადენინი, გუანინი, თიმინი და ციტოზინი). ერთ-ერთი ჯაჭვის აზოტოვანი ფუძეები დაკავშირებულია მეორე ჯაჭვის აზოტოვან ფუძებთან წყალბადური ბმებით კომპლემენტარობის პრინციპის მიხედვით: ადენინი ერწყმის მხოლოდ თიმინს ( A-T), გუანინი - მხოლოდ ციტოზინით ( G-C). სწორედ ეს წყვილები ქმნიან დნმ-ის ხვეული „კიბის“ „საფეხურებს“ (იხ.: სურ. 2, 3 და 4).

ბრინჯი. 2. აზოტოვანი ფუძეები
ნუკლეოტიდების თანმიმდევრობა საშუალებას გაძლევთ „დაშიფვროთ“ ინფორმაცია სხვადასხვა სახისრნმ, რომელთაგან ყველაზე მნიშვნელოვანია ინფორმაცია ან შაბლონი (mRNA), რიბოსომული (rRNA) და ტრანსპორტი (tRNA). ყველა ამ ტიპის რნმ სინთეზირდება დნმ-ის შაბლონზე დნმ-ის თანმიმდევრობის კოპირებით ტრანსკრიპციის დროს სინთეზირებულ რნმ-ის თანმიმდევრობაში და მონაწილეობს ცილების ბიოსინთეზში (თარგმანის პროცესი). გარდა კოდირების თანმიმდევრობისა, უჯრედის დნმ შეიცავს თანმიმდევრობებს, რომლებიც ასრულებენ მარეგულირებელ და სტრუქტურულ ფუნქციებს.

ბრინჯი. 3. დნმ-ის რეპლიკაცია
ძირითადი კომბინაციების ადგილმდებარეობა ქიმიური ნაერთებიდნმ და რაოდენობრივი კოეფიციენტებიამ კომბინაციებს შორის უზრუნველყოფს მემკვიდრეობითი ინფორმაციის დაშიფვრას.
Განათლება ახალი დნმ (რეპლიკაცია)
- რეპლიკაციის პროცესი: დნმ-ის ორმაგი სპირალის გახსნა - დამატებითი ჯაჭვების სინთეზი დნმ პოლიმერაზას მიერ - დნმ-ის ორი მოლეკულის წარმოქმნა ერთიდან.
- ორმაგი სპირალი "იხსნება" ორ ტოტად, როდესაც ფერმენტები არღვევს კავშირს ქიმიური ნაერთების ფუძე წყვილებს შორის.
- თითოეული ტოტი არის ახალი დნმ-ის ელემენტი. ახალი ბაზის წყვილები დაკავშირებულია იმავე თანმიმდევრობით, როგორც მშობლის ფილიალში.
დუბლირების დასრულების შემდეგ წარმოიქმნება ორი დამოუკიდებელი სპირალი, რომლებიც შექმნილია მშობელი დნმ-ის ქიმიური ნაერთებისგან და აქვთ იგივე გენეტიკური კოდი. ამ გზით, დნმ-ს შეუძლია ინფორმაციის ამოღება უჯრედიდან უჯრედში.
უფრო დეტალური ინფორმაცია:
ნუკლეინის მჟავების სტრუქტურა

ბრინჯი. 4 . აზოტოვანი ფუძეები: ადენინი, გუანინი, ციტოზინი, თიმინი
Დეზოქსირიბონუკლეინის მჟავა(დნმ) ეხება ნუკლეინის მჟავებს. Ნუკლეინის მჟავაარის არარეგულარული ბიოპოლიმერების კლასი, რომელთა მონომერები არის ნუკლეოტიდები.
ნუკლეოტიდებიშედგება აზოტოვანი ბაზადაკავშირებულია ხუთნახშირბადიან ნახშირწყალთან (პენტოზა) - დეზოქსირიბოზა(დნმ-ის შემთხვევაში) ან რიბოზა(რნმ-ის შემთხვევაში), რომელიც უკავშირდება დანარჩენს ფოსფორმჟავა(H 2 PO 3 -).
აზოტოვანი ფუძეებიარსებობს ორი ტიპი: პირიმიდინის ფუძეები - ურაცილი (მხოლოდ რნმ-ში), ციტოზინი და თიმინი, პურინის ფუძეები - ადენინი და გუანინი.

ბრინჯი. 5. აზოტოვანი ფუძეების სახეები: პირიმიდინი და პურინი
ნახშირბადის ატომები პენტოზას მოლეკულაში დანომრილია 1-დან 5-მდე. ფოსფატი ერწყმის მესამე და მეხუთე ნახშირბადის ატომებს. ასე უკავშირდებიან ნუკლეინის მჟავები და ქმნიან ნუკლეინის მჟავების ჯაჭვს. ამრიგად, ჩვენ შეგვიძლია გამოვყოთ დნმ-ის ჯაჭვის 3' და 5' ბოლოები:

ბრინჯი. 6. დნმ-ის ჯაჭვის 3' და 5' ბოლოების იზოლირება
დნმ-ის ორი ჯაჭვი იქმნება ორმაგი სპირალი. ეს ჯაჭვები სპირალურად არის ორიენტირებული საპირისპირო მიმართულებით. AT სხვადასხვა ჯაჭვებიდნმ-ის აზოტოვანი ფუძეები ერთმანეთთან არის დაკავშირებული წყალბადის ბმები. ადენინი ყოველთვის ერწყმის თიმინს, ხოლო ციტოზინი ყოველთვის აერთიანებს გუანინს. მას ეძახიან კომპლემენტარობის წესი.
კომპლემენტარობის წესი:
| A-T G-C |
მაგალითად, თუ გვეძლევა დნმ-ის ჯაჭვი, რომელსაც აქვს თანმიმდევრობა
3'-ATGTCCTAGCTGCTCG - 5',
მაშინ მეორე ჯაჭვი იქნება მისი შემავსებელი და მიმართული საწინააღმდეგო მიმართულება- 5'-დან 3'-ბოლომდე:
5'- TACAGGATCGACGAGC- 3'.

ბრინჯი. 7. დნმ-ის მოლეკულის ჯაჭვების მიმართულება და აზოტოვანი ფუძეების შეერთება წყალბადური ბმების გამოყენებით.
რეპლიკაცია
დნმ-ის რეპლიკაციაარის დნმ-ის მოლეკულის დუბლირების პროცესი მატრიცის სინთეზი. რეპლიკაცია ხდება ნახევრად კონსერვატიული მექანიზმი. ეს ნიშნავს, რომ დნმ-ის ორმაგი სპირალი იხსნება და მის თითოეულ ჯაჭვზე სრულდება ახალი ჯაჭვი კომპლემენტარობის პრინციპის მიხედვით. ამრიგად, ქალიშვილი დნმ-ის მოლეკულა შეიცავს ერთ ჯაჭვს მშობლის მოლეკულიდან და ერთ ახლად სინთეზირებულს. რეპლიკაცია ხდება მშობელი ჯაჭვის 3'-დან 5'-მდე მიმართულებით.

ბრინჯი. 8. დნმ-ის მოლეკულის რეპლიკაცია (გაორმაგება).
დნმ სინთეზი- ეს არც ისე რთული პროცესია, როგორც ერთი შეხედვით შეიძლება ჩანდეს. თუ დაფიქრდებით, ჯერ უნდა გაარკვიოთ რა არის სინთეზი. ეს არის რაღაცის გაერთიანების პროცესი. ახალი დნმ-ის მოლეკულის ფორმირება ხდება რამდენიმე ეტაპად:
- დნმ ტოპოიზომერაზა, რომელიც განლაგებულია რეპლიკაციის ჩანგლის წინ, ჭრის დნმ-ს, რათა ხელი შეუწყოს მის განტვირთვასა და განტვირთვას.
- დნმ-ის ჰელიკაზა, ტოპოიზომერაზას შემდეგ, გავლენას ახდენს დნმ-ის სპირალის „გახსნის“ პროცესზე.
- დნმ-ის დამაკავშირებელი პროტეინები ახორციელებენ დნმ-ის ჯაჭვების შეკავშირებას და ასევე ახორციელებენ მათ სტაბილიზაციას, რაც ხელს უშლის მათ ერთმანეთთან მიბმას.
- დნმ პოლიმერაზა ასინთეზებს ქალიშვილის დნმ-ის წამყვან ჯაჭვს.

ბრინჯი. 9. რეპლიკაციის პროცესის სქემატური წარმოდგენა, რიცხვები მიუთითებს: (1) ლაგ-ჯაჭვა, (2) წამყვანი ჯაჭვი, (3) დნმ პოლიმერაზა (Polα), (4) დნმ ლიგაზა, (5) რნმ პრაიმერი, (6) პრიმაზა , (7) ოკაზაკის ფრაგმენტი, (8) დნმ პოლიმერაზა (Polδ), (9) ჰელიკაზა, (10) ერთჯაჭვიანი დნმ-ის დამაკავშირებელი ცილები, (11) ტოპოიზომერაზა
რნმ სტრუქტურა
რიბონუკლეინის მჟავა(რნმ) არის სამი ძირითადი მაკრომოლეკულიდან ერთ-ერთი (დანარჩენი ორი არის დნმ და ცილა), რომლებიც გვხვდება ყველა ცოცხალი ორგანიზმის უჯრედებში.
ისევე, როგორც დნმ, რნმ შედგება გრძელი ჯაჭვისაგან, რომელშიც თითოეულ რგოლს ნუკლეოტიდი ეწოდება. თითოეული ნუკლეოტიდი შედგება აზოტოვანი ფუძისგან, რიბოზის შაქრისა და ფოსფატის ჯგუფისგან. თუმცა, დნმ-ისგან განსხვავებით, რნმ-ს ჩვეულებრივ აქვს ერთი და არა ორი ჯაჭვი. რნმ-ში პენტოზა წარმოდგენილია რიბოზათ და არა დეზოქსირიბოზით (რიბოზას აქვს დამატებითი ჰიდროქსილის ჯგუფინახშირწყლების მეორე ატომზე). დაბოლოს, დნმ განსხვავდება რნმ-ისგან აზოტოვანი ბაზების შემადგენლობით: თიმინის ნაცვლად ( თ) ურაცილი არის რნმ-ში ( U) , რომელიც ასევე ავსებს ადენინს.
ნუკლეოტიდების თანმიმდევრობა რნმ-ს გენეტიკური ინფორმაციის კოდირების საშუალებას აძლევს. ყველა ფიჭური ორგანიზმებიგამოიყენეთ რნმ (მრნმ) ცილების სინთეზის დასაპროგრამებლად.
უჯრედული რნმ წარმოიქმნება პროცესში ე.წ ტრანსკრიფცია , ანუ რნმ-ის სინთეზი დნმ-ის შაბლონზე, რომელსაც ახორციელებს სპეციალური ფერმენტები - რნმ პოლიმერაზები.
მესინჯერი რნმ-ები (mRNA) შემდეგ მონაწილეობენ პროცესში ე.წ გადაცემა, იმათ. ცილის სინთეზი mRNA შაბლონზე რიბოზომების მონაწილეობით. სხვა რნმ-ები განიცდიან ქიმიურ მოდიფიკაციას ტრანსკრიპციის შემდეგ და მეორადი და მესამეული სტრუქტურებიასრულებს ფუნქციებს რნმ-ის ტიპის მიხედვით.

ბრინჯი. 10. განსხვავება დნმ-სა და რნმ-ს შორის აზოტოვანი ფუძის მიხედვით: თიმინის (T) ნაცვლად რნმ შეიცავს ურაცილს (U), რომელიც ასევე ავსებს ადენინს.
ტრანსკრიპცია
ეს არის რნმ-ის სინთეზის პროცესი დნმ-ის შაბლონზე. დნმ იხსნება ერთ-ერთ ადგილზე. ერთ-ერთი ჯაჭვი შეიცავს ინფორმაციას, რომელიც საჭიროებს კოპირებას რნმ-ის მოლეკულაზე - ამ ჯაჭვს კოდირებას უწოდებენ. დნმ-ის მეორე ჯაჭვს, რომელიც ავსებს კოდირების ჯაჭვს, ეწოდება შაბლონი. შაბლონის ჯაჭვზე ტრანსკრიფციის პროცესში 3'-5' მიმართულებით (დნმ-ის ჯაჭვის გასწვრივ) სინთეზირდება მისი დამატებითი რნმ-ის ჯაჭვი. ამრიგად, იქმნება კოდირების ჯაჭვის რნმ-ის ასლი.
![]()
ბრინჯი. 11. ტრანსკრიფციის სქემატური წარმოდგენა
მაგალითად, თუ გვეძლევა კოდირების სტრიქონის თანმიმდევრობა
3'-ATGTCCTAGCTGCTCG - 5',
შემდეგ, კომპლემენტარობის წესის მიხედვით, მატრიცის ჯაჭვი ატარებს თანმიმდევრობას
5'- TACAGGATCGACGAGC- 3',
და მისგან სინთეზირებული რნმ არის თანმიმდევრობა
გადაცემა
განიხილეთ მექანიზმი ცილის სინთეზირნმ-ის მატრიცაზე, ასევე გენეტიკურ კოდსა და მის თვისებებზე. ასევე, სიცხადისთვის, ქვემოთ მოცემულ ბმულზე, გირჩევთ უყუროთ მოკლე ვიდეოს ცოცხალ უჯრედში მიმდინარე ტრანსკრიფციისა და თარგმნის პროცესების შესახებ:

ბრინჯი. 12. ცილის სინთეზის პროცესი: დნმ კოდირებს რნმ-ს, რნმ კოდირებს ცილას
გენეტიკური კოდი
გენეტიკური კოდი - ცილების ამინომჟავების თანმიმდევრობის კოდირების მეთოდი ნუკლეოტიდების თანმიმდევრობის გამოყენებით. თითოეული ამინომჟავა დაშიფრულია სამი ნუკლეოტიდის თანმიმდევრობით - კოდონი ან ტრიპლეტი.
გენეტიკური კოდი საერთოა პრო- და ევკარიოტების უმეტესობისთვის. ცხრილში მოცემულია 64-ვე კოდონი და ჩამოთვლილია შესაბამისი ამინომჟავები. საბაზისო რიგი არის mRNA-ს 5"-დან 3"-მდე ბოლომდე.
ცხრილი 1. სტანდარტული გენეტიკური კოდი
|
1-ლი არა |
მე-2 ბაზა |
მე-3 არა |
|||||||
|
U |
C |
ა |
გ |
||||||
|
U |
U U U |
(Phe/F) |
U C U |
(სერ/ს) |
U A U |
(წ/წ) |
U G U |
(Cys/C) |
U |
|
U U C |
U C C |
U A C |
U G C |
C |
|||||
|
U U A |
(ლეუ/ლ) |
U C A |
U A A |
კოდონის გაჩერება** |
U G A |
კოდონის გაჩერება** |
ა |
||
|
U U G |
U C G |
U A G |
კოდონის გაჩერება** |
U G G |
(Trp/W) |
გ |
|||
|
C |
C U U |
C C U |
(პრო/პ) |
C A U |
(მისი/ჰ) |
C G U |
(არგ/რ) |
U |
|
|
C U C |
C C C |
C A C |
C G C |
C |
|||||
|
C U A |
C C A |
C A A |
(Gln/Q) |
CGA |
ა |
||||
|
C U G |
C C G |
C A G |
C G G |
გ |
|||||
|
ა |
A U U |
(Ile/I) |
A C U |
(Thr/T) |
A A U |
(Asn/N) |
A G U |
(სერ/ს) |
U |
|
A U C |
A C C |
A A C |
A G C |
C |
|||||
|
A U A |
A C A |
A A A |
(Lys/K) |
A G A |
ა |
||||
|
A U G |
(მეტ/მ) |
A C G |
A A G |
A G G |
გ |
||||
|
გ |
G U U |
(Val/V) |
G C U |
(ალა/ა) |
G A U |
(Asp/D) |
გ გ უ |
(Gly/G) |
U |
|
G U C |
G C C |
G A C |
G G C |
C |
|||||
|
გ უ ა |
G C A |
G A A |
(გლუ/ე) |
G G A |
ა |
||||
|
გ უ გ |
G C G |
G A G |
G G G |
გ |
|||||
სამეულებს შორის არის 4 სპეციალური თანმიმდევრობა, რომლებიც მოქმედებს როგორც "პუნქტუაციის ნიშნები":
- *სამმაგი აუგ, ასევე მეთიონინის კოდირებას, ე.წ დაწყების კოდონი. ეს კოდონი იწყებს ცილის მოლეკულის სინთეზს. ამრიგად, ცილის სინთეზის დროს, პირველი ამინომჟავა თანმიმდევრობით ყოველთვის იქნება მეთიონინი.
- ** სამეული UAA, UAGდა UGAდაურეკა შეაჩერე კოდონებიდა არ კოდებს რაიმე ამინომჟავას. ამ თანმიმდევრობებზე ცილის სინთეზი ჩერდება.
გენეტიკური კოდის თვისებები
1. სამეული. თითოეული ამინომჟავა კოდირებულია სამი ნუკლეოტიდის თანმიმდევრობით - ტრიპლეტი ან კოდონი.
2. უწყვეტობა. სამეულებს შორის დამატებითი ნუკლეოტიდები არ არის, ინფორმაცია მუდმივად იკითხება.
3. გადახურვის გარეშე. ერთი ნუკლეოტიდი არ შეიძლება იყოს ერთდროულად ორი სამეულის ნაწილი.
4. უნიკალურობა. ერთ კოდონს შეუძლია მხოლოდ ერთი ამინომჟავის კოდირება.

5. დეგენერაცია. ერთი ამინომჟავა შეიძლება დაშიფრული იყოს რამდენიმე სხვადასხვა კოდონით.

6. მრავალმხრივობა. გენეტიკური კოდი ყველა ცოცხალი ორგანიზმისთვის ერთნაირია.
მაგალითი. ჩვენ მოცემულია კოდირების სტრიქონის თანმიმდევრობა:
3’- CCGATTGCACGTCGATCGTATA- 5’.
მატრიცის ჯაჭვს ექნება თანმიმდევრობა:
5’- GGCTAACGTGCAGCTAGCATAT- 3’.
ახლა ჩვენ „ასინთეზირებთ“ ინფორმაციულ რნმ-ს ამ ჯაჭვიდან:
3’- CCGAUUGCACGUCGAUCGUAUA- 5’.
პროტეინის სინთეზი მიდის 5' → 3' მიმართულებით, ამიტომ, გენეტიკური კოდის "წაკითხვისთვის" უნდა გადავატრიალოთ თანმიმდევრობა:
5’- AUAUGCUAGCUGCACGUUAGCC- 3’.
ახლა იპოვნეთ საწყისი კოდონი AUG:
5’- AU AUG CUAGCUGCACGUUAGCC- 3’.
დაყავით თანმიმდევრობა სამეულებად:
ასე ჟღერს: დნმ-დან ინფორმაცია გადადის რნმ-ში (ტრანსკრიფცია), რნმ-დან ცილაში (თარგმანი). ასევე შესაძლებელია დნმ-ის დუბლირება რეპლიკაციით და პროცესიც შესაძლებელია საპირისპირო ტრანსკრიფცია, როცა რნმ-ის შაბლონიდან დნმ სინთეზირდება, მაგრამ ასეთი პროცესი ძირითადად ვირუსებისთვისაა დამახასიათებელი.

ბრინჯი. 13. მოლეკულური ბიოლოგიის ცენტრალური დოგმა
გენომი: გენები და ქრომოსომა
(ზოგადი ცნებები)
გენომი - ორგანიზმის ყველა გენის მთლიანობა; მისი სრული ქრომოსომული ნაკრები.
ტერმინი „გენომი“ შემოგვთავაზა გ. ვინკლერმა 1920 წელს, რათა აღეწერა გენების მთლიანობა, რომელიც შეიცავს ერთი ორგანიზმის ქრომოსომების ჰაპლოიდურ კომპლექტს. სახეობა. ამ ტერმინის თავდაპირველი მნიშვნელობა მიუთითებდა, რომ გენომის კონცეფცია, გენოტიპისგან განსხვავებით, არის გენეტიკური მახასიათებელისახეობა მთლიანობაში და არა ინდივიდი. განვითარებასთან ერთად მოლეკულური გენეტიკაშეიცვალა ტერმინის მნიშვნელობა. ცნობილია, რომ დნმ, რომელიც არის გენეტიკური ინფორმაციის მატარებელი უმეტეს ორგანიზმებში და, შესაბამისად, ქმნის გენომის საფუძველს, მოიცავს არა მხოლოდ გენებს თანამედროვე გრძნობაეს სიტყვა. უმეტესობაევკარიოტული უჯრედების დნმ წარმოდგენილია არაკოდიციური ("ზედმეტი") ნუკლეოტიდური თანმიმდევრობებით, რომლებიც არ შეიცავს ინფორმაციას ცილებისა და ნუკლეინის მჟავების შესახებ. ამრიგად, ნებისმიერი ორგანიზმის გენომის ძირითადი ნაწილია მისი ქრომოსომების ჰაპლოიდური ნაკრების მთელი დნმ.
გენები არის დნმ-ის მოლეკულების სეგმენტები, რომლებიც კოდირებენ პოლიპეპტიდებს და რნმ-ის მოლეკულებს.
უკან ბოლო საუკუნეჩვენი გაგება გენების შესახებ მკვეთრად შეიცვალა. ადრე, გენომი იყო ქრომოსომის რეგიონი, რომელიც კოდირებს ან განსაზღვრავს ერთ მახასიათებელს ან ფენოტიპური(ხილული) თვისება, როგორიცაა თვალის ფერი.
1940 წელს ჯორჯ ბიდლმა და ედვარდ ტატამმა შესთავაზეს მოლეკულური განმარტებაგენი. მეცნიერებმა დაამუშავეს სოკოს სპორები ნეიროსპორა კრასა რენტგენიდა სხვა აგენტები, რომლებიც იწვევენ ცვლილებებს დნმ-ის თანმიმდევრობაში ( მუტაციები), და აღმოაჩინა სოკოს მუტანტური შტამები, რომლებმაც დაკარგეს გარკვეული სპეციფიკური ფერმენტები, რამაც ზოგიერთ შემთხვევაში გამოიწვია მთლიანობის დარღვევა. მეტაბოლური გზა. Beadle და Tatham მივიდნენ დასკვნამდე, რომ გენი არის გენეტიკური მასალის ნაწილი, რომელიც განსაზღვრავს ან კოდირებს ერთ ფერმენტს. ასეა ჰიპოთეზა "ერთი გენი, ერთი ფერმენტი". ეს კონცეფცია მოგვიანებით გაფართოვდა განსაზღვრებამდე "ერთი გენი - ერთი პოლიპეპტიდი"რადგან ბევრი გენი აკოდირებს ცილებს, რომლებიც არ არიან ფერმენტები და პოლიპეპტიდი შეიძლება იყოს რთული ცილის კომპლექსის ქვედანაყოფი.
ნახ. 14 გვიჩვენებს დიაგრამას, თუ როგორ განსაზღვრავს დნმ-ის სამეულები პოლიპეპტიდს, ცილის ამინომჟავის თანმიმდევრობას, შუამავლობით mRNA. დნმ-ის ერთ-ერთი ჯაჭვი ასრულებს mRNA-ს სინთეზის შაბლონის როლს, რომლის ნუკლეოტიდური ტრიპლეტები (კოდონები) ავსებენ დნმ-ის სამეულს. ზოგიერთ ბაქტერიაში და ბევრ ევკარიოტში, კოდირების თანმიმდევრობა წყდება არაკოდირების რეგიონებით (ე.წ. ინტრონები).
თანამედროვე ბიოქიმიური განმარტებაგენი კიდევ უფრო კონკრეტულად. გენები არის დნმ-ის ყველა განყოფილება, რომელიც კოდირებს საბოლოო პროდუქტების პირველადი თანმიმდევრობას, რომელიც მოიცავს პოლიპეპტიდებს ან რნმ-ს, რომლებსაც აქვთ სტრუქტურული ან კატალიზური ფუნქცია.
გენებთან ერთად, დნმ ასევე შეიცავს სხვა თანმიმდევრობებს, რომლებიც მოქმედებს ექსკლუზიურად მარეგულირებელი ფუნქცია. მარეგულირებელი თანმიმდევრობებიშეიძლება აღინიშნოს გენების დასაწყისი ან დასასრული, გავლენა მოახდინოს ტრანსკრიფციაზე ან მიუთითოს რეპლიკაციის ან რეკომბინაციის დაწყების ადგილი. ზოგიერთი გენი შეიძლება გამოიხატოს სხვადასხვა გზით, დნმ-ის ერთი და იგივე ნაწილი სხვადასხვა პროდუქტის ფორმირების შაბლონად ემსახურება.
ჩვენ შეგვიძლია უხეშად გამოვთვალოთ გენის მინიმალური ზომაშუალედური ცილის კოდირება. თითოეული ამინომჟავა პოლიპეპტიდურ ჯაჭვში კოდირებულია სამი ნუკლეოტიდის თანმიმდევრობით; ამ სამეულების (კოდონების) თანმიმდევრობა შეესაბამება მოცემული გენის მიერ კოდირებულ პოლიპეპტიდში ამინომჟავების ჯაჭვს. პოლიპეპტიდური ჯაჭვი 350 ამინომჟავის ნარჩენებისგან შუა სიგრძე) შეესაბამება 1050 b.p. ( ბპ). თუმცა, ბევრი ევკარიოტული გენი და ზოგიერთი პროკარიოტული გენი წყდება დნმ-ის სეგმენტებით, რომლებიც არ ატარებენ ინფორმაციას ცილის შესახებ და, შესაბამისად, ბევრად უფრო გრძელია, ვიდრე მარტივი გაანგარიშება აჩვენებს.
რამდენი გენია ერთ ქრომოსომაზე?
 ბრინჯი. 15. ქრომოსომების ხედი პროკარიოტულ (მარცხნივ) და ევკარიოტულ უჯრედებში. ჰისტონები არის ბირთვული ცილების ფართო კლასი, რომლებიც ასრულებენ ორ ძირითად ფუნქციას: ისინი მონაწილეობენ ბირთვში დნმ-ის ჯაჭვების შეფუთვაში და ბირთვული პროცესების ეპიგენეტიკურ რეგულირებაში, როგორიცაა ტრანსკრიფცია, რეპლიკაცია და შეკეთება.
ბრინჯი. 15. ქრომოსომების ხედი პროკარიოტულ (მარცხნივ) და ევკარიოტულ უჯრედებში. ჰისტონები არის ბირთვული ცილების ფართო კლასი, რომლებიც ასრულებენ ორ ძირითად ფუნქციას: ისინი მონაწილეობენ ბირთვში დნმ-ის ჯაჭვების შეფუთვაში და ბირთვული პროცესების ეპიგენეტიკურ რეგულირებაში, როგორიცაა ტრანსკრიფცია, რეპლიკაცია და შეკეთება.
მოგეხსენებათ, ბაქტერიულ უჯრედებს აქვთ ქრომოსომა დნმ-ის ჯაჭვის სახით, რომელიც შეფუთულია კომპაქტურ სტრუქტურაში - ნუკლეოიდში. პროკარიოტული ქრომოსომა ეშერიხია კოლი, რომლის გენომი მთლიანად გაშიფრულია, არის წრიული დნმ-ის მოლეკულა (ფაქტობრივად, ეს არ არის მარჯვენა წრე, არამედ მარყუჟი დასაწყისისა და დასასრულის გარეშე), რომელიც შედგება 4,639,675 b.p. ეს თანმიმდევრობა შეიცავს დაახლოებით 4300 ცილის გენს და კიდევ 157 გენს სტაბილური რნმ-ის მოლეკულებისთვის. AT ადამიანის გენომიდაახლოებით 3,1 მილიარდი ბაზის წყვილი, რომელიც შეესაბამება თითქმის 29000 გენს, რომლებიც განლაგებულია 24 სხვადასხვა ქრომოსომაზე.
პროკარიოტები (ბაქტერიები).
 ბაქტერია E. coliაქვს ერთი ორჯაჭვიანი წრიული დნმ-ის მოლეკულა. იგი შედგება 4,639,675 ბ.წ. და აღწევს სიგრძე დაახლოებით 1,7 მმ, რაც აღემატება თავად უჯრედის სიგრძეს E. coliდაახლოებით 850 ჯერ. გარდა დიდი წრიული ქრომოსომისა, როგორც ნუკლეოიდის ნაწილი, ბევრი ბაქტერია შეიცავს ციტოზოლში თავისუფლად განლაგებულ ერთ ან მეტ წრიულ დნმ-ის მოლეკულას. ამ ექსტრაქრომოსომულ ელემენტებს ე.წ პლაზმიდები(სურ. 16).
ბაქტერია E. coliაქვს ერთი ორჯაჭვიანი წრიული დნმ-ის მოლეკულა. იგი შედგება 4,639,675 ბ.წ. და აღწევს სიგრძე დაახლოებით 1,7 მმ, რაც აღემატება თავად უჯრედის სიგრძეს E. coliდაახლოებით 850 ჯერ. გარდა დიდი წრიული ქრომოსომისა, როგორც ნუკლეოიდის ნაწილი, ბევრი ბაქტერია შეიცავს ციტოზოლში თავისუფლად განლაგებულ ერთ ან მეტ წრიულ დნმ-ის მოლეკულას. ამ ექსტრაქრომოსომულ ელემენტებს ე.წ პლაზმიდები(სურ. 16).
პლაზმიდების უმეტესობა შედგება მხოლოდ რამდენიმე ათასი ბაზის წყვილისგან, ზოგი შეიცავს 10000 bp-ზე მეტს. ისინი ატარებენ გენეტიკურ ინფორმაციას და მრავლდებიან შვილობილი პლაზმიდების წარმოქმნით, რომლებიც ხვდებიან ქალიშვილი უჯრედებიმშობელი უჯრედის გაყოფის დროს. პლაზმიდები გვხვდება არა მხოლოდ ბაქტერიებში, არამედ საფუარში და სხვა სოკოებშიც. ხშირ შემთხვევაში, პლაზმიდები არ ანიჭებენ უპირატესობას მასპინძელ უჯრედებს და მათი ერთადერთი ამოცანაა დამოუკიდებლად გამრავლება. თუმცა, ზოგიერთი პლაზმიდი ატარებს მასპინძლისთვის სასარგებლო გენებს. მაგალითად, პლაზმიდებში შემავალ გენებს შეუძლიათ ბაქტერიული უჯრედების ანტიბაქტერიული აგენტებისადმი წინააღმდეგობის გაწევა. პლაზმიდები გენის მატარებელიβ-ლაქტამაზები ანიჭებენ წინააღმდეგობას β-ლაქტამური ანტიბიოტიკების მიმართ, როგორიცაა პენიცილინი და ამოქსიცილინი. პლაზმიდები შეიძლება გადავიდნენ ანტიბიოტიკებისადმი მდგრადი უჯრედებიდან იმავე ან სხვადასხვა ბაქტერიული სახეობის სხვა უჯრედებში, რის გამოც ეს უჯრედები ასევე რეზისტენტული ხდება. ანტიბიოტიკების ინტენსიური გამოყენება არის ძლიერი შერჩევითი ფაქტორი, რომელიც ხელს უწყობს ანტიბიოტიკების რეზისტენტობის მაკოდირებელი პლაზმიდების გავრცელებას (ისევე, როგორც ტრანსპოზონებს, რომლებიც აკოდირებენ მსგავს გენებს) შორის. პათოგენური ბაქტერიები, და იწვევს ბაქტერიული შტამების წარმოქმნას რამდენიმე ანტიბიოტიკის მიმართ რეზისტენტობით. ექიმები იწყებენ ანტიბიოტიკების ფართო გამოყენების საშიშროების გააზრებას და დანიშნავენ მათ მხოლოდ მაშინ, როცა აბსოლუტურად აუცილებელია. მსგავსი მიზეზების გამო, ფერმის ცხოველების სამკურნალოდ ანტიბიოტიკების ფართო გამოყენება შეზღუდულია.
Იხილეთ ასევე: რავინ ნ.ვ., შესტაკოვი ს.ვ. პროკარიოტების გენომი // Vavilov Journal of Genetics and Breeding, 2013. V. 17. No 4/2. გვ 972-984.
ევკარიოტები.
ცხრილი 2. ზოგიერთი ორგანიზმის დნმ, გენები და ქრომოსომა
|
საერთო დნმ, ბ.წ. |
ქრომოსომების რაოდენობა* |
გენების სავარაუდო რაოდენობა |
|
|
ეშერიხია კოლი(ბაქტერია) |
4 639 675 |
4 435 |
|
|
Saccharomyces cerevisiae(საფუარი) |
12 080 000 |
16** |
5 860 |
|
Caenorhabditis elegans(ნემატოდი) |
90 269 800 |
12*** |
23 000 |
|
Arabidopsis thaliana(მცენარე) |
119 186 200 |
33 000 |
|
|
დროზოფილა მელანოგასტერი(ხილის ბუზი) |
120 367 260 |
20 000 |
|
|
ორიზა სატივა(ბრინჯი) |
480 000 000 |
57 000 |
|
|
მუს კუნთი(მაუსი) |
2 634 266 500 |
27 000 |
|
|
ჰომო საპიენსი(ადამიანი) |
3 070 128 600 |
29 000 |
Შენიშვნა.ინფორმაცია მუდმივად განახლდება; დამატებითი უახლესი ინფორმაციისთვის ეწვიეთ ცალკეული გენომიური პროექტის ვებსაიტებს.
* ყველა ევკარიოტისთვის, საფუარის გარდა, მოცემულია ქრომოსომების დიპლოიდური ნაკრები. დიპლოიდურინაკრები ქრომოსომა (ბერძნულიდან diploos - ორმაგი და eidos - ხედი) - ქრომოსომების ორმაგი ნაკრები (2n), რომელთაგან თითოეულს აქვს ჰომოლოგიური.
**ჰაპლოიდური ნაკრები. საფუარის ველურ შტამებს, როგორც წესი, აქვთ ამ ქრომოსომების რვა (ოქტაპლოიდური) ან მეტი ნაკრები.
*** ორი X ქრომოსომის მქონე ქალებისთვის. მამაკაცებს აქვთ X ქრომოსომა, მაგრამ არა Y, ანუ მხოლოდ 11 ქრომოსომა.
საფუარის უჯრედს, ერთ-ერთ ყველაზე პატარა ევკარიოტს, აქვს 2,6-ჯერ მეტი დნმ, ვიდრე უჯრედს. E. coli(ცხრილი 2). ხილის ბუზის უჯრედები დროზოფილა, კლასიკური ობიექტი გენეტიკური კვლევა, შეიცავს 35-ჯერ მეტ დნმ-ს, ხოლო ადამიანის უჯრედები შეიცავს დაახლოებით 700-ჯერ მეტ დნმ-ს, ვიდრე უჯრედები E. coli.ბევრი მცენარე და ამფიბია კიდევ უფრო მეტ დნმ-ს შეიცავს. ევკარიოტული უჯრედების გენეტიკური მასალა ორგანიზებულია ქრომოსომების სახით. ქრომოსომების დიპლოიდური ნაკრები (2 ნ) დამოკიდებულია ორგანიზმის ტიპზე (ცხრილი 2).
მაგალითად, ადამიანის სომატურ უჯრედში არის 46 ქრომოსომა ( ბრინჯი. 17). თითოეული ქრომოსომა ევკარიოტულ უჯრედში, როგორც ნაჩვენებია ნახ. 17, ა, შეიცავს ერთ ძალიან დიდ ორჯაჭვიან დნმ-ის მოლეკულას. ადამიანის ოცდაოთხი ქრომოსომა (22 დაწყვილებული ქრომოსომა და ორი სქესის ქრომოსომა X და Y) განსხვავდება სიგრძით 25-ჯერ მეტით. თითოეული ევკარიოტული ქრომოსომა შეიცავს გენების სპეციფიკურ კომპლექტს.

ბრინჯი. 17. ევკარიოტული ქრომოსომა.ა- წყვილი დაკავშირებული და შედედებული დის ქრომატიდიდან ადამიანის ქრომოსომიდან. ამ ფორმით, ევკარიოტული ქრომოსომა რჩება რეპლიკაციის შემდეგ და მეტაფაზაში მიტოზის დროს. ბ- ქრომოსომების სრული ნაკრები წიგნის ერთ-ერთი ავტორის ლეიკოციტიდან. თითოეული ნორმალური ადამიანის სომატური უჯრედი შეიცავს 46 ქრომოსომას.
თუ თქვენ დააკავშირებთ ადამიანის გენომის დნმ-ის მოლეკულებს (22 ქრომოსომა და ქრომოსომა X და Y ან X და X) ერთმანეთთან, მიიღებთ დაახლოებით ერთი მეტრის სიგრძის თანმიმდევრობას. შენიშვნა: ყველა ძუძუმწოვარსა და სხვა ჰეტეროგამეტურ მამრობითი ორგანიზმში მდედრს აქვს ორი X ქრომოსომა (XX), ხოლო მამრს აქვს ერთი X ქრომოსომა და ერთი Y ქრომოსომა (XY).
ადამიანის უჯრედების უმეტესობა, ამიტომ ასეთი უჯრედების დნმ-ის მთლიანი სიგრძე დაახლოებით 2 მ-ია. ზრდასრულ ადამიანს აქვს დაახლოებით 10 14 უჯრედი, ამიტომ დნმ-ის ყველა მოლეკულის მთლიანი სიგრძეა 2-10 11 კმ. შედარებისთვის, დედამიწის გარშემოწერილობა არის 4~104 კმ, ხოლო დედამიწიდან მზემდე მანძილი 1,5~108 კმ. აი, საოცრად კომპაქტურად შეფუთული დნმ არის ჩვენს უჯრედებში!
ევკარიოტულ უჯრედებში არის დნმ-ის შემცველი სხვა ორგანელები - ეს არის მიტოქონდრია და ქლოროპლასტები. მრავალი ჰიპოთეზა წამოაყენეს მიტოქონდრიული და ქლოროპლასტის დნმ-ის წარმოშობასთან დაკავშირებით. დღეს საყოველთაოდ მიღებული თვალსაზრისი არის ის, რომ ისინი უძველესი ბაქტერიების ქრომოსომების საფუძვლებია, რომლებიც შეაღწიეს მასპინძელი უჯრედების ციტოპლაზმაში და გახდნენ ამ ორგანელების წინამორბედები. მიტოქონდრიული დნმკოდირებს მიტოქონდრიულ tRNA-ს და rRNA-ს, ასევე რამდენიმე მიტოქონდრიულ ცილას. მიტოქონდრიული ცილების 95%-ზე მეტი კოდირებულია ბირთვული დნმ-ით.
გენების სტრუქტურა
განვიხილოთ გენის სტრუქტურა პროკარიოტებში და ევკარიოტებში, მათი მსგავსება და განსხვავებები. იმისდა მიუხედავად, რომ გენი არის დნმ-ის განყოფილება, რომელიც კოდირებს მხოლოდ ერთ პროტეინს ან რნმ-ს, გარდა უშუალო კოდირების ნაწილისა, მასში შედის აგრეთვე მარეგულირებელი და სხვა სტრუქტურული ელემენტებიმქონე განსხვავებული სტრუქტურაპროკარიოტებში და ევკარიოტებში.
კოდირების თანმიმდევრობა- გენის მთავარი სტრუქტურული და ფუნქციური ერთეული, მასშია ნუკლეოტიდების სამეული, რომელიც კოდირებსამინომჟავების თანმიმდევრობა. ის იწყება საწყისი კოდონით და მთავრდება გაჩერების კოდონით.
კოდირების თანმიმდევრობამდე და მის შემდეგ არის გადაუთარგმნელი 5' და 3' თანმიმდევრობები. ისინი ასრულებენ მარეგულირებელ და დამხმარე ფუნქციებს, მაგალითად, უზრუნველყოფენ რიბოსომის დაშვებას mRNA-ზე.
გადაუთარგმნელი და კოდირებული თანმიმდევრობები ქმნიან ტრანსკრიფციის ერთეულს - ტრანსკრიბირებული დნმ-ის რეგიონს, ანუ დნმ-ის რეგიონს, საიდანაც სინთეზირებულია mRNA.
ტერმინატორიდნმ-ის არატრანსკრიბირებული რეგიონი გენის ბოლოს, სადაც რნმ-ის სინთეზი ჩერდება.
გენის დასაწყისში არის მარეგულირებელი ტერიტორია, რომელიც შეიცავს პრომოუტერიდა ოპერატორი.
პრომოუტერი- თანმიმდევრობა, რომლითაც პოლიმერაზა უკავშირდება ტრანსკრიფციის დაწყებისას. ოპერატორი- ეს ის უბანია, რომელსაც სპეციალური პროტეინები შეიძლება დაუკავშირდეს - რეპრესორები, რომელსაც შეუძლია შეამციროს რნმ-ის სინთეზის აქტივობა ამ გენიდან - სხვა სიტყვებით რომ ვთქვათ, შეამციროს იგი გამოხატულება.
გენების სტრუქტურა პროკარიოტებში
პროკარიოტებში და ევკარიოტებში გენების სტრუქტურის ზოგადი გეგმა არ განსხვავდება - ორივე მათგანი შეიცავს მარეგულირებელ რეგიონს პრომოტორთან და ოპერატორთან, ტრანსკრიპციის ერთეულს კოდირების და არათარგმნილი თანმიმდევრობით და ტერმინატორთან. თუმცა, გენების ორგანიზაცია პროკარიოტებსა და ევკარიოტებში განსხვავებულია.

ბრინჯი. 18. გენის სტრუქტურის სქემა პროკარიოტებში (ბაქტერიებში) -სურათი გადიდებულია
ოპერონის დასაწყისში და ბოლოს არის საერთო მარეგულირებელი რეგიონები რამდენიმე სტრუქტურული გენისთვის. ოპერონის ტრანსკრიბირებული რეგიონიდან იკითხება ერთი mRNA მოლეკულა, რომელიც შეიცავს რამდენიმე კოდირების თანმიმდევრობას, რომელთაგან თითოეულს აქვს საკუთარი საწყისი და გაჩერების კოდონი. თითოეული ამ სფეროდანსინთეზირდება ერთი ცილა. ამრიგად, რამდენიმე ცილის მოლეკულა სინთეზირდება ერთი i-RNA მოლეკულიდან.
პროკარიოტები აერთიანებს რამდენიმე გენს ერთში ფუნქციური ერთეული -ოპერონი. ოპერონის მუშაობა შეიძლება დარეგულირდეს სხვა გენებით, რომლებიც შესამჩნევად შეიძლება მოიხსნას თავად ოპერონიდან - რეგულატორები. ამ გენიდან ნათარგმნ ცილას ე.წ რეპრესორი. ის აკავშირებს ოპერონის ოპერატორს, არეგულირებს მასში შემავალი ყველა გენის ექსპრესიას ერთდროულად.
ფენომენი ახასიათებს პროკარიოტებსაც ტრანსკრიფცია და თარგმანის კონიუგაციები.
![]()
ბრინჯი. 19 პროკარიოტებში ტრანსკრიფციისა და თარგმანის კონიუგაციის ფენომენი - სურათი გადიდებულია
რევერსიული ტრანსკრიპტაზას მიერ კატალიზებული რეაქციაში.
cDNA ხშირად გამოიყენება პროკარიოტებში ევკარიოტული გენების კლონირებისთვის. დამატებითი დნმ ასევე იწარმოება რეტროვირუსებით (აივ-1, აივ-2, სიმიანური იმუნოდეფიციტის ვირუსი) და შემდეგ ინტეგრირდება მასპინძლის დნმ-ში პროვირუსის წარმოქმნით.
ხშირად, ევკარიოტული გენები შეიძლება გამოხატული იყოს პროკარიოტულ უჯრედებში. ყველაზე მეტად მარტივი შემთხვევა, მეთოდი გულისხმობს ევკარიოტული დნმ-ის პროკარიოტულ გენომში ჩასმას, შემდეგ დნმ-ის mRNA-ში ტრანსკრიფციას და შემდეგ mRNA-ს ცილებად გადაყვანას. პროკარიოტულ უჯრედებს არ აქვთ ინტრონის გამომჭრელი ფერმენტები და ამიტომ ინტრონები უნდა მოიჭრას ევკარიოტული დნმ-დან პროკარიოტულ გენომში ჩასმამდე. დნმ-ს, რომელიც ავსებს მომწიფებულ mRNA-ს, ამგვარად ეწოდება კომპლემენტარული დნმ - cDNA(cDNA). პროკარიოტებში ევკარიოტული cDNA-ში კოდირებული ცილების წარმატებული ექსპრესიისთვის საჭიროა აგრეთვე პროკარიოტული გენების მარეგულირებელი ელემენტები (მაგ. პრომოტორები).
აუცილებელი გენის (დნმ-ის მოლეკულა) მიღების ერთ-ერთი მეთოდი, რომელიც დაექვემდებარება რეპლიკაციას (კლონირებას) რეპლიკების მნიშვნელოვანი რაოდენობის გამოშვებით, არის mRNA-ზე დამატებითი დნმ-ის (cDNA) აგება. ეს მეთოდი მოითხოვს საპირისპირო ტრანსკრიპტაზას, ფერმენტის გამოყენებას, რომელიც იმყოფება რნმ-ის შემცველ ზოგიერთ ვირუსში და უზრუნველყოფს დნმ-ის სინთეზს რნმ-ის შაბლონზე.
მეთოდი ფართოდ გამოიყენება cDNA-ს მისაღებად და მოიცავს mRNA-ს იზოლაციას მთლიანი ქსოვილის mRNA-დან, რომელიც კოდირებს სპეციფიკური ცილის (მაგალითად, ინტერფერონის, ინსულინის) ტრანსლაციას ამ mRNA-ზე შემდგომი სინთეზით, როგორც აუცილებელი cDNA-ს შაბლონი, საპირისპირო გამოყენებით. ტრანსკრიპტაზა.
გენი, რომელიც მიღებული იქნა ზემოაღნიშნული პროცედურის გამოყენებით (cDNA) უნდა შეიყვანონ ბაქტერიულ უჯრედში ისე, რომ იგი ინტეგრირდეს მის გენომში. ამისათვის ისინი ქმნიან რეკომბინანტული დნმ, რომელიც შედგება cDNA-სგან და სპეციალური დნმ-ის მოლეკულისგან, რომელიც მართავს როგორც გამტარებელს, ანუ ვექტორს, რომელსაც შეუძლია შეაღწიოს მიმღებს უჯრედში. ვირუსები ან პლაზმიდები გამოიყენება როგორც ვექტორები cDNA-სთვის. პლაზმიდები არის მცირე წრიული დნმ-ის მოლეკულები, რომლებიც განცალკევებულია ნუკლეოიდისგან. ბაქტერიული უჯრედიშეიცავს რამდენიმე გენს, რომლებიც მნიშვნელოვანია მთელი უჯრედის ფუნქციონირებისთვის (მაგალითად, ანტიბიოტიკებისადმი რეზისტენტობის გენები და შეუძლიათ რეპლიკაცია უჯრედის მთავარი გენომის (დნმ) დამოუკიდებლად. ბიოლოგიურად მნიშვნელოვანია და პრაქტიკულად გამოსადეგი გენეტიკური ინჟინერიაპლაზმიდების თვისებებია მათი გადაცემის უნარი ერთი უჯრედიდან მეორეში ტრანსფორმაციის ან კონიუგაციის მექანიზმით, აგრეთვე ბაქტერიულ ქრომოსომაში ჩართვისა და მასთან ერთად რეპლიკაციის უნარი.
უოტსონმა და კრიკმა აჩვენეს, რომ წყალბადის ობლიგაციებისა და რეგულარული ორმაგი სპირალის ფორმირება შესაძლებელია მხოლოდ მაშინ, როდესაც ერთ ჯაჭვში უფრო დიდ პურინის ფუძე ადენინს (A) აქვს უფრო პატარა პირიმიდინის ფუძე თიმინი (T), როგორც პარტნიორი მეორე ჯაჭვში, და გუანინი (G) ასოცირებული ციტოზინთან (C). ეს ნიმუში შეიძლება წარმოდგენილი იყოს შემდეგნაირად:კორესპონდენცია A "T და G" C ეწოდება კომპლემენტარობის წესს, და თავად ჯაჭვები შემავსებელი. ამ წესის მიხედვით დნმ-ში ადენინის შემცველობა ყოველთვის თიმინის შემცველობის ტოლია, ხოლო გუანინის რაოდენობა ყოველთვის ციტოზინის ტოლია. უნდა აღინიშნოს, რომ დნმ-ის ორი ჯაჭვი, რომლებიც ქიმიურად განსხვავდება, ერთსა და იმავე ინფორმაციას ატარებს, რადგან კომპლემენტარობის გამო, ერთი ჯაჭვი ცალსახად განსაზღვრავს მეორეს.რნმ-ის სტრუქტურა ნაკლებად მოწესრიგებულია. ჩვეულებრივ, ეს არის ერთჯაჭვიანი მოლეკულა, თუმცა ზოგიერთი ვირუსის რნმ შედგება ორი ჯაჭვისგან. მაგრამ ასეთი რნმც უფრო მოქნილი ვიდრე დნმ. რნმ-ის მოლეკულის ზოგიერთი მონაკვეთი ერთმანეთს ავსებს და, როდესაც ჯაჭვი მოხრილია, წყვილდება, ქმნიან ორჯაჭვიან სტრუქტურებს (თმის სამაგრებს). უპირველეს ყოვლისა, ეს ეხება გადაცემის რნმ-ებს (tRNA). tRNA-ს ზოგიერთი ბაზა მოლეკულის სინთეზის შემდეგ განიცდის მოდიფიკაციას. მაგალითად, ზოგჯერ მეთილის ჯგუფები ერთვის მათ.
ნუკლეინის მჟავების ფუნქცია ნუკლეინის მჟავების ერთ-ერთი მთავარი ფუნქციაა ცილის სინთეზის განსაზღვრა. ინფორმაცია დნმ-ის ნუკლეოტიდურ თანმიმდევრობაში კოდირებული ცილების სტრუქტურის შესახებ უნდა გადაეცეს ერთი თაობიდან მეორეს და ამიტომ მისი უტყუარი კოპირება აუცილებელია, ე.ი. ზუსტად იგივე დნმ-ის მოლეკულის სინთეზი (რეპლიკაცია).რეპლიკაცია და ტრანსკრიფცია. ქიმიური თვალსაზრისით ნუკლეინის მჟავას სინთეზი არის პოლიმერიზაცია, ე.ი. სამშენებლო ბლოკების თანმიმდევრული კავშირი. ასეთი ბლოკებია ნუკლეოზიდის ტრიფოსფატები; რეაქცია შეიძლება წარმოდგენილი იყოს შემდეგნაირად: სინთეზისთვის საჭირო ენერგია გამოიყოფა პიროფოსფატის დაშლისას და სპეციალური ფერმენტები, დნმ პოლიმერაზები, ახდენენ რეაქციის კატალიზებას.
სინთეზისთვის საჭირო ენერგია გამოიყოფა პიროფოსფატის დაშლისას და სპეციალური ფერმენტები, დნმ პოლიმერაზები, ახდენენ რეაქციის კატალიზებას. ასეთი სინთეზური პროცესის შედეგად მივიღებდით შემთხვევითი ფუძის მიმდევრობის მქონე პოლიმერს. თუმცა, პოლიმერაზების უმეტესობა მუშაობს მხოლოდ წინასწარ არსებული ნუკლეინის მჟავის თანდასწრებით, შაბლონი, რომელიც კარნახობს რომელი ნუკლეოტიდი იქნება მიმაგრებული ჯაჭვის ბოლოს. ეს ნუკლეოტიდი უნდა იყოს შესაბამისი შაბლონის ნუკლეოტიდის შემავსებელი, ისე რომ ახალი ძაფები ავსებს ორიგინალს. შემდეგ დამატებითი სტრიქონის მატრიცის სახით გამოყენებით, მივიღებთ ზუსტი ასლიორიგინალური.
დნმ შედგება ორი ურთიერთშემავსებელი ჯაჭვისგან. რეპლიკაციის დროს ისინი განსხვავდებიან და თითოეული მათგანი ემსახურება როგორც შაბლონს ახალი ძაფის სინთეზისთვის:
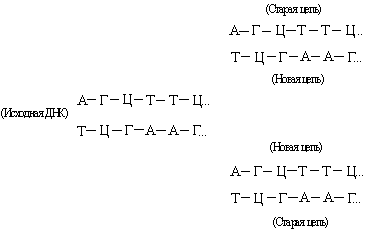 ეს ქმნის ორ ახალ ორმაგ სპირალს, იგივე ბაზის თანმიმდევრობით, როგორც ორიგინალური დნმ. ზოგჯერ რეპლიკაციის პროცესი „ჩაიშლება“ და ხდება მუტაციები (იხილეთ ასევე
მემკვიდრეობა).
დნმ-ის ტრანსკრიფციის შედეგად წარმოიქმნება უჯრედული რნმ (mRNA, rRNA და tRNA):
ეს ქმნის ორ ახალ ორმაგ სპირალს, იგივე ბაზის თანმიმდევრობით, როგორც ორიგინალური დნმ. ზოგჯერ რეპლიკაციის პროცესი „ჩაიშლება“ და ხდება მუტაციები (იხილეთ ასევე
მემკვიდრეობა).
დნმ-ის ტრანსკრიფციის შედეგად წარმოიქმნება უჯრედული რნმ (mRNA, rRNA და tRNA): ისინი ავსებენ დნმ-ის ერთ-ერთ ჯაჭვს და წარმოადგენენ მეორე ჯაჭვის ასლს, გარდა იმისა, რომ თიმინის ადგილს ურაცილი იკავებს. ამ გზით, დნმ-ის ერთ-ერთი ჯაჭვის მრავალი რნმ-ის ასლის მიღება შეიძლება.ნორმალურ უჯრედში ინფორმაცია გადაეცემა მხოლოდ დნმ-ის მიმართულებით.® დნმ და დნმ ® რნმ. თუმცა, ვირუსით ინფიცირებულ უჯრედებში შესაძლებელია სხვა პროცესებიც: რნმ® რნმ და რნმ ® დნმ. მრავალი ვირუსის გენეტიკური მასალა წარმოდგენილია რნმ-ის მოლეკულით, ჩვეულებრივ ერთჯაჭვიანი. მასპინძელ უჯრედში შეღწევის შემდეგ, ეს რნმ იმეორებს დამატებით მოლეკულას, რომელზედაც, თავის მხრივ, სინთეზირებულია ორიგინალური ვირუსული რნმ-ის მრავალი ასლი:
ისინი ავსებენ დნმ-ის ერთ-ერთ ჯაჭვს და წარმოადგენენ მეორე ჯაჭვის ასლს, გარდა იმისა, რომ თიმინის ადგილს ურაცილი იკავებს. ამ გზით, დნმ-ის ერთ-ერთი ჯაჭვის მრავალი რნმ-ის ასლის მიღება შეიძლება.ნორმალურ უჯრედში ინფორმაცია გადაეცემა მხოლოდ დნმ-ის მიმართულებით.® დნმ და დნმ ® რნმ. თუმცა, ვირუსით ინფიცირებულ უჯრედებში შესაძლებელია სხვა პროცესებიც: რნმ® რნმ და რნმ ® დნმ. მრავალი ვირუსის გენეტიკური მასალა წარმოდგენილია რნმ-ის მოლეკულით, ჩვეულებრივ ერთჯაჭვიანი. მასპინძელ უჯრედში შეღწევის შემდეგ, ეს რნმ იმეორებს დამატებით მოლეკულას, რომელზედაც, თავის მხრივ, სინთეზირებულია ორიგინალური ვირუსული რნმ-ის მრავალი ასლი: ვირუსული რნმ შეიძლება გადაიწეროს ფერმენტის მიერ-
საპირისპირო ტრანსკრიპტაზა-
დნმ-ში, რომელიც ზოგჯერ შედის მასპინძელი უჯრედის ქრომოსომულ დნმ-ში. ახლა ეს დნმ ატარებს ვირუსულ გენებს და ტრანსკრიფციის შემდეგ, ვირუსული რნმ შეიძლება გამოჩნდეს უჯრედში. ამრიგად, მას შემდეგ დიდი დრო, რომლის დროსაც უჯრედში ვირუსი არ არის ნაპოვნი, ის ხელახლა გამოჩნდება მასში ხელახალი ინფექციის გარეშე. ვირუსები, რომელთა გენეტიკური მასალა ჩართულია მასპინძელი უჯრედის ქრომოსომაში, ხშირად კიბოს გამომწვევი მიზეზია.
ვირუსული რნმ შეიძლება გადაიწეროს ფერმენტის მიერ-
საპირისპირო ტრანსკრიპტაზა-
დნმ-ში, რომელიც ზოგჯერ შედის მასპინძელი უჯრედის ქრომოსომულ დნმ-ში. ახლა ეს დნმ ატარებს ვირუსულ გენებს და ტრანსკრიფციის შემდეგ, ვირუსული რნმ შეიძლება გამოჩნდეს უჯრედში. ამრიგად, მას შემდეგ დიდი დრო, რომლის დროსაც უჯრედში ვირუსი არ არის ნაპოვნი, ის ხელახლა გამოჩნდება მასში ხელახალი ინფექციის გარეშე. ვირუსები, რომელთა გენეტიკური მასალა ჩართულია მასპინძელი უჯრედის ქრომოსომაში, ხშირად კიბოს გამომწვევი მიზეზია.
1953 წელს ისეთი ნივთიერების მოლეკულური ორგანიზაციის პრინციპის აღმოჩენის შემდეგ, როგორიცაა დნმ, დაიწყო განვითარება. მოლეკულური ბიოლოგია. გარდა ამისა, კვლევის პროცესში მეცნიერებმა გაარკვიეს, თუ როგორ ხდება დნმ-ის რეკომბინაცია, მისი შემადგენლობა და როგორ არის მოწყობილი ჩვენი ადამიანის გენომი.
ყოველ დღე მოლეკულური დონეხდება ყველაზე რთული პროცესები. როგორ არის მოწყობილი დნმ-ის მოლეკულა, რისგან შედგება? რა როლს ასრულებენ დნმ-ის მოლეკულები უჯრედში? მოდი დეტალურად ვისაუბროთ ორმაგი ჯაჭვის შიგნით მიმდინარე ყველა პროცესზე.
რა არის მემკვიდრეობითი ინფორმაცია?
მაშ, როგორ დაიწყო ეს ყველაფერი? ჯერ კიდევ 1868 წელს ნაპოვნი იქნა ბაქტერიების ბირთვებში. 1928 წელს კი ნ.კოლცოვმა წამოაყენა თეორია, რომ ცოცხალი ორგანიზმის შესახებ ყველა გენეტიკური ინფორმაცია დაშიფრულია დნმ-ში. შემდეგ ჯ. უოტსონმა და ფ. კრიკმა 1953 წელს იპოვეს ახლა უკვე კარგად ცნობილი დნმ-ის სპირალის მოდელი, რისთვისაც დამსახურებულად მიიღეს აღიარება და ჯილდო - ნობელის პრემია.
მაინც რა არის დნმ? ეს ნივთიერება შედგება 2 კომბინირებული ძაფისგან, უფრო ზუსტად სპირალებისგან. ასეთი ჯაჭვის მონაკვეთს გარკვეული ინფორმაციით ეწოდება გენი.

დნმ ინახავს ყველა ინფორმაციას იმის შესახებ, თუ რა სახის ცილები წარმოიქმნება და რა თანმიმდევრობით. დნმ მაკრომოლეკულა არის მასალის გადამზიდავიწარმოუდგენლად მოცულობითი ინფორმაცია, რომელიც ჩაწერილია ინდივიდუალური სამშენებლო ბლოკების - ნუკლეოტიდების მკაცრი თანმიმდევრობით. სულ 4 ნუკლეოტიდია, ისინი ერთმანეთს ავსებენ ქიმიურად და გეომეტრიულად. მეცნიერებაში შევსების, ანუ კომპლემენტარობის ეს პრინციპი მოგვიანებით იქნება აღწერილი. ეს წესი მოქმედებს საკვანძო როლიგენეტიკური ინფორმაციის კოდირებასა და გაშიფვრაში.
ვინაიდან დნმ-ის ჯაჭვი წარმოუდგენლად გრძელია, ამ თანმიმდევრობაში არ არის გამეორებები. ყველა ცოცხალ არსებას აქვს თავისი უნიკალური დნმ-ის ჯაჭვი.
დნმ-ის ფუნქციები
ფუნქციებში შედის მემკვიდრეობითი ინფორმაციის შენახვა და შთამომავლობაზე გადაცემა. ამ ფუნქციის გარეშე, სახეობის გენომის შენარჩუნება და განვითარება შეუძლებელია ათასწლეულების განმავლობაში. ორგანიზმები, რომლებმაც განიცადეს ძირითადი გენის მუტაციები, უფრო მეტად არ გადარჩებიან ან დაკარგავენ შთამომავლობის წარმოქმნის უნარს. ასე რომ, არსებობს ბუნებრივი დაცვა სახეობების გადაგვარებისგან.

კიდევ ერთი აუცილებელი მნიშვნელოვანი ფუნქცია— შენახული ინფორმაციის განხორციელება. უჯრედს არ შეუძლია რაიმე სასიცოცხლო პროტეინის შექმნა ინსტრუქციების გარეშე, რომლებიც ინახება ორმაგ ძაფში.
ნუკლეინის მჟავების შემადგენლობა
ახლა უკვე საიმედოდ ცნობილია რისგან შედგება თავად ნუკლეოტიდები, დნმ-ის სამშენებლო ბლოკები. ისინი მოიცავს 3 ნივთიერებას:
- ორთოფოსფორის მჟავა.
- აზოტოვანი ბაზა. პირიმიდინის ფუძეები – რომლებსაც აქვთ მხოლოდ ერთი რგოლი. მათ შორისაა თიმინი და ციტოზინი. პურინის ფუძეები, რომლებიც შეიცავს 2 რგოლს. ეს არის გუანინი და ადენინი.
- საქაროზა. დნმ შეიცავს დეზოქსირიბოზას, რნმ შეიცავს რიბოზას.
ნუკლეოტიდების რაოდენობა ყოველთვის უდრის აზოტოვანი ფუძეების რაოდენობას. სპეციალურ ლაბორატორიებში ხდება ნუკლეოტიდის გაყოფა და მისგან იზოლირება აზოტოვანი ბაზა. ასე რომ, ისინი სწავლობენ ამ ნუკლეოტიდების ინდივიდუალურ თვისებებს და მათში შესაძლო მუტაციებს.
მემკვიდრეობითი ინფორმაციის ორგანიზების დონეები
არსებობს ორგანიზაციის 3 დონე: გენი, ქრომოსომული და გენომი. ახალი ცილის სინთეზისთვის საჭირო ყველა ინფორმაცია ჯაჭვის მცირე მონაკვეთში – გენშია. ანუ გენი ითვლება ინფორმაციის კოდირების ყველაზე დაბალ და მარტივ დონედ.

გენები, თავის მხრივ, იკრიბებიან ქრომოსომებად. მემკვიდრეობითი მასალის მატარებლის ასეთი ორგანიზაციის წყალობით, თვისებების ჯგუფები მონაცვლეობენ გარკვეული კანონების მიხედვით და გადაეცემა ერთი თაობიდან მეორეს. უნდა აღინიშნოს, რომ ორგანიზმში წარმოუდგენლად ბევრი გენი არსებობს, მაგრამ ინფორმაცია არ იკარგება, მაშინაც კი, როდესაც ის ბევრჯერ არის შერწყმული.
არსებობს რამდენიმე სახის გენი:
- on ფუნქციური დანიშნულებაგამოიყოფა 2 ტიპი: სტრუქტურული და მარეგულირებელი თანმიმდევრობა;
- უჯრედში მიმდინარე პროცესებზე გავლენის მიხედვით განასხვავებენ: ზევიტალური, ლეტალური, პირობითად ლეტალური გენები, ასევე მუტაციური და ანტიმუტაციური გენები.
გენები განლაგებულია ქრომოსომის გასწვრივ ხაზოვანი წესრიგი. ქრომოსომებში ინფორმაცია შემთხვევით არ არის ფოკუსირებული, არის გარკვეული რიგი. არსებობს რუკაც კი, სადაც ნაჩვენებია პოზიციები, ან გენის ლოკაციები. მაგალითად, ცნობილია, რომ მონაცემები ბავშვის თვალების ფერის შესახებ დაშიფრულია ქრომოსომა 18-ში.
რა არის გენომი? ეს არის ნუკლეოტიდური თანმიმდევრობის მთელი ნაკრების სახელი სხეულის უჯრედში. გენომი ახასიათებს მთელი ხედი, არც ერთი ინდივიდი.
რა არის ადამიანის გენეტიკური კოდი?
ფაქტია, რომ მთელი უზარმაზარი პოტენციალი ადამიანის განვითარებაჩამოყალიბებულია ჩასახვის დროს. ყველა მემკვიდრეობითი ინფორმაცია, რომელიც აუცილებელია ზიგოტის განვითარებისა და ბავშვის ზრდისთვის დაბადების შემდეგ, დაშიფრულია გენებში. დნმ-ის სექციები მემკვიდრეობითი ინფორმაციის ყველაზე ძირითადი მატარებელია.

ადამიანებს აქვთ 46 ქრომოსომა, ანუ 22 სომატური წყვილი პლუს ერთი სქესის განმსაზღვრელი ქრომოსომა თითოეული მშობლისგან. ქრომოსომების ეს დიპლოიდური ნაკრები კოდირებს ადამიანის მთელ ფიზიკურ გარეგნობას, მის გონებრივ და ფიზიკურ შესაძლებლობებს და დაავადებებისადმი მიდრეკილებას. სომატური ქრომოსომა გარეგნულად არ განსხვავდება, მაგრამ ისინი სხვადასხვა ინფორმაციას ატარებენ, რადგან ერთი მათგანი მამისგანაა, მეორე კი დედისგან.
მამრობითი კოდი განსხვავდება ქალის კოდისგან ქრომოსომების ბოლო წყვილში - XY. ქალის დიპლოიდური ნაკრები არის ბოლო წყვილი, XX. მამრები იღებენ ერთ X ქრომოსომას ბიოლოგიური დედისგან, შემდეგ კი ის გადაეცემათ ქალიშვილებს. სქესის Y ქრომოსომა გადაეცემა ვაჟებს.
ადამიანის ქრომოსომა ზომით ძალიან განსხვავდება. მაგალითად, ყველაზე პატარა წყვილი ქრომოსომა არის #17. და ყველაზე დიდი წყვილი არის 1 და 3.
ორმაგი სპირალის დიამეტრი ადამიანებში მხოლოდ 2 ნმ-ია. დნმ იმდენად მჭიდროდ არის დახვეული, რომ უჯრედის პატარა ბირთვში ჯდება, თუმცა მისი ამოღების შემთხვევაში სიგრძე 2 მეტრამდე იქნება. სპირალის სიგრძე ასობით მილიონი ნუკლეოტიდია.
როგორ ხდება გენეტიკური კოდის გადაცემა?
მაშ, რა როლს ასრულებენ დნმ-ის მოლეკულები უჯრედში გაყოფის დროს? გენები - მემკვიდრეობითი ინფორმაციის მატარებლები - სხეულის ყველა უჯრედშია. იმისათვის, რომ მათი კოდი გადასცეს ქალიშვილ ორგანიზმს, ბევრი არსება თავის დნმ-ს ყოფს 2 იდენტურ სპირალში. ამას რეპლიკაცია ჰქვია. რეპლიკაციის პროცესში დნმ იხსნება და სპეციალური „მანქანები“ ავსებენ თითოეულ ჯაჭვს. გენეტიკური სპირალის გაყოფის შემდეგ ბირთვი და ყველა ორგანელა დაყოფას იწყებს, შემდეგ კი მთელი უჯრედი.
მაგრამ ადამიანს გენის გადაცემის განსხვავებული პროცესი აქვს – სექსუალური. მამისა და დედის ნიშნები შერეულია, ახალი გენეტიკური კოდი შეიცავს ინფორმაციას ორივე მშობლისგან.
მემკვიდრეობითი ინფორმაციის შენახვა და გადაცემა შესაძლებელია იმის გამო კომპლექსური ორგანიზაციადნმ-ის ჯაჭვები. ყოველივე ამის შემდეგ, როგორც ვთქვით, ცილების სტრუქტურა დაშიფრულია გენებში. ჩასახვის დროს შექმნის შემდეგ, ეს კოდი კოპირდება მთელი ცხოვრების განმავლობაში. ორგანოს უჯრედების განახლებისას კარიოტიპი (ქრომოსომების პირადი ნაკრები) არ იცვლება. ინფორმაციის გადაცემა ხდება სქესის გამეტების - მამრობითი და მდედრობითი სქესის დახმარებით.
მხოლოდ ვირუსებს, რომლებიც შეიცავს რნმ-ის ერთ ჯაჭვს, არ შეუძლიათ თავიანთი ინფორმაციის გადაცემა შთამომავლებისთვის. ამიტომ, გამრავლებისთვის მათ სჭირდებათ ადამიანის ან ცხოველის უჯრედები.
მემკვიდრეობითი ინფორმაციის დანერგვა
უჯრედის ბირთვში მუდმივია მნიშვნელოვანი პროცესები. ქრომოსომებში ჩაწერილი ყველა ინფორმაცია გამოიყენება ამინომჟავებისგან ცილების შესაქმნელად. მაგრამ დნმ-ის ჯაჭვი არასოდეს ტოვებს ბირთვს, ამიტომ აქ სხვა ადამიანის დახმარებაა საჭირო. მნიშვნელოვანი კავშირი= რნმ. მხოლოდ რნმ-ს შეუძლია შეაღწიოს ბირთვულ მემბრანაში და ურთიერთქმედება დნმ-ის ჯაჭვთან.
დნმ-ისა და რნმ-ის 3 ტიპის ურთიერთქმედების გზით ხდება ყველა კოდირებული ინფორმაციის რეალიზება. რა დონეზეა მემკვიდრეობითი ინფორმაციის დანერგვა? ყველა ურთიერთქმედება ხდება ნუკლეოტიდის დონეზე. მესინჯერი რნმ აკოპირებს დნმ-ის ჯაჭვის სეგმენტს და მოაქვს ეს ასლი რიბოსომამდე. აქ იწყება ახალი მოლეკულის ნუკლეოტიდების სინთეზი.
იმისათვის, რომ mRNA-მ დააკოპიროს ჯაჭვის აუცილებელი ნაწილი, სპირალი იშლება და შემდეგ, გადაკოდირების პროცესის დასრულების შემდეგ, კვლავ აღდგება. უფრო მეტიც, ეს პროცესი შეიძლება მოხდეს ერთდროულად 1 ქრომოსომის 2 მხარეს.
კომპლემენტარობის პრინციპი
ისინი შედგება 4 ნუკლეოტიდისგან - ეს არის ადენინი (A), გუანინი (G), ციტოზინი (C), თიმინი (T). ისინი დაკავშირებულია წყალბადის ბმებით კომპლემენტარობის წესის მიხედვით. ამ წესის დადგენაში დაეხმარა ე. ჩარგაფის ნაშრომებმა, ვინაიდან მეცნიერმა შეამჩნია გარკვეული ნიმუშები ამ ნივთიერებების ქცევაში. ე. ჩარგაფმა აღმოაჩინა, რომ ადენინის და თიმინის მოლური თანაფარდობა ერთის ტოლია. და ანალოგიურად, გუანინის და ციტოზინის თანაფარდობა ყოველთვის ერთის ტოლია.
მის ნაშრომზე დაყრდნობით გენეტიკოსებმა შექმნეს ნუკლეოტიდების ურთიერთქმედების წესი. კომპლემენტარობის წესი ამბობს, რომ ადენინი აერთიანებს მხოლოდ თიმინს, ხოლო გუანინი ციტოზინს. სპირალის გაშიფვრისა და რიბოსომაში ახალი ცილის სინთეზის დროს, მონაცვლეობის ეს წესი ეხმარება სწრაფად იპოვოთ საჭირო ამინომჟავა, რომელიც მიმაგრებულია გადამტან რნმ-ზე.
რნმ და მისი ტიპები
რა არის მემკვიდრეობითი ინფორმაცია? ნუკლეოტიდები დნმ-ის ორმაგ ჯაჭვში. რა არის რნმ? Რაარის მისი სამუშაო? რნმ ან რიბონუკლეინის მჟავა ხელს უწყობს დნმ-დან ინფორმაციის ამოღებას, მის გაშიფვრას და კომპლემენტარობის პრინციპის საფუძველზე შექმნას. უჯრედებს სჭირდებათცილები.
საერთო ჯამში იზოლირებულია რნმ-ის 3 ტიპი. თითოეული მათგანი მკაცრად ასრულებს თავის ფუნქციას.
- საინფორმაციო (mRNA), ან მას ასევე უწოდებენ მატრიცას. ის პირდაპირ მიდის უჯრედის ცენტრში, ბირთვში. ის ერთ-ერთ ქრომოსომაში პოულობს ცილის ასაშენებლად აუცილებელ გენეტიკურ მასალას და აკოპირებს ორმაგი ჯაჭვის ერთ-ერთ მხარეს. კოპირება კვლავ ხდება კომპლემენტარობის პრინციპის მიხედვით.
- ტრანსპორტი- ეს მცირე მოლეკულა, რომელსაც ერთ მხარეს აქვს ნუკლეოტიდური დეკოდერები, ხოლო მეორე მხარეს მთავარი კოდის შესაბამისი ამინომჟავები. tRNA-ს ამოცანაა მისი მიწოდება „საამქროში“, ანუ რიბოსომაში, სადაც ის ასინთეზებს საჭირო ამინომჟავას.
- rRNA არის რიბოსომული.ის აკონტროლებს წარმოებული ცილის რაოდენობას. შედგება 2 ნაწილისაგან - ამინომჟავისა და პეპტიდური ადგილისგან.
დეკოდირებისას ერთადერთი განსხვავება ისაა, რომ რნმ-ს არ აქვს თიმინი. თიმინის ნაცვლად აქ არის ურაცილი. მაგრამ შემდეგ, ცილის სინთეზის პროცესში, tRNA-სთან ერთად, ის მაინც სწორად აყალიბებს ყველა ამინომჟავას. თუ ინფორმაციის გაშიფვრაში რაიმე წარუმატებლობაა, მაშინ ხდება მუტაცია.
დაზიანებული დნმ-ის მოლეკულის აღდგენა
დაზიანებული ორმაგი ძაფის შეკეთების პროცესს რეპარაცია ეწოდება. აღდგენის პროცესში ხდება დაზიანებული გენების ამოღება.

შემდეგ ელემენტების საჭირო თანმიმდევრობა ზუსტად რეპროდუცირებულია და ჯდება იმავე ადგილას ჯაჭვზე, საიდანაც იგი იქნა ამოღებული. ეს ყველაფერი განსაკუთრებულის წყალობით ხდება ქიმიკატები- ფერმენტები.
რატომ ხდება მუტაციები?
რატომ იწყებს ზოგიერთი გენი მუტაციას და წყვეტს თავისი ფუნქციის შესრულებას - სასიცოცხლო მემკვიდრეობითი ინფორმაციის შენახვას? ეს გამოწვეულია დეკოდირების შეცდომით. მაგალითად, თუ ადენინი შემთხვევით შეიცვალა თიმინით.
ასევე არსებობს ქრომოსომული და გენომიური მუტაციები. ქრომოსომული მუტაციებიწარმოიქმნება, თუ მემკვიდრეობითი ინფორმაციის მონაკვეთები ამოვარდება, გაორმაგდება ან ზოგადად გადადის და ინტეგრირდება სხვა ქრომოსომაში.

გენომური მუტაციები ყველაზე სერიოზულია. მათი მიზეზი ქრომოსომების რაოდენობის ცვლილებაა. ანუ, როდესაც წყვილის ნაცვლად - დიპლოიდური ნაკრები, კარიოტიპში იმყოფება ტრიპლოიდური ნაკრები.
უმეტესობა ცნობილი მაგალითიტრიპლოიდური მუტაცია არის დაუნის სინდრომი, რომლის დროსაც ქრომოსომების პერსონალური ნაკრები არის 47. ასეთ ბავშვებში 21-ე წყვილის ნაცვლად 3 ქრომოსომა ყალიბდება.
ასევე არსებობს ისეთი მუტაცია, როგორიცაა პოლიპლოიდი. მაგრამ პოლიპლოიდი გვხვდება მხოლოდ მცენარეებში.
